
A preliminary assessment of cisco (Coregonus spp.) diversity in Yellowknife Bay, Great Slave Lake,...
24
A preliminary assessment of cisco (Coregonus spp.) diversity in Yellowknife Bay, Great Slave Lake, Northwest Territories Paul Vecsei 1 , Craig T. Blackie 1,2 , Andrew M. Muir 1,3 *, Hilary M. Machtans 1 , J.D. Reist 4 with 8 figures and 5 tables Abstract: Northern freshwater fishes, particularly members of the genus Coregonus, have been used as a model for studying microevolution due to their well known colonization history coupled with parallel patterns of phenotypic differentiation. We assessed and described the phenotypic diversity among ciscoes in the Yellowknife Bay region of Great Slave Lake using 1) traditional taxonomic methods for cisco species and 2) contemporary linear and geometric morphometric methods. Four morphs were identified using a qualitative visual assessment of gross morphology and/or location of capture in comparison to published taxonomy for the genus Coregonus. The morphs identified were assigned the following colloquial names: A) ‘adfluvial’, B) ‘lacustrine’, C) ‘shortjaw’, and D) ‘least’. Univariate analyses on taxonomic measures indicated that cisco varied in terms of age, number of lateral line scales, number of anal fin rays, middle gill raker length, and lower gill arch length. By contrast, five morphs were identified using cluster analysis of linear and geometric morphometric data combined with age and gill raker number. The additional phenotypic cluster entailed a splitting of large and small lacustrine morphs into two groups. Multivariate analyses indicated a strong relationship between body shape and gill raker number, eye size, age and mouth morphology. These results suggested that the segregation among morphs was based on differential usage of trophic and/or habitat resources as well as life history variation such as age at maturity. Our results indicated that the level of phenotypic diversity was greater than previously documented for Great Slave Lake and that this diversity was similar to that reported for ciscoes in the Laurentian Great Lakes. Keywords: Coregonus spp., cisco, Great Lakes, morphometrics, divergence, polymorphism. Authors’ addresses: 1 Golder Associates Ltd. 9-4905 48th Street, Yellowknife, Northwest Territories, X1A 3S3, Canada. 2 Biology Department, Dalhousie University, 1355 Oxford St., Halifax, Nova Scotia, B3H 4J1, Canada. 3 Current address: Great Lakes Fishery Commission, 2100 Commonwealth Blvd., Suite 100, Ann Arbor, MI, 48105, USA. 4 Arctic Fish Ecology and Assessment Research Section, Fisheries and Oceans Canada, Winnipeg, Manitoba, R3T 2N6, Canada. * Corresponding author, e-mail: [email protected] 1612-166X/12/0063-0299 $ 6.00 © 2012 E. Schweizerbart’sche Verlagsbuchhandlung, 70176 Stuttgart, Germany Advanc. Limnol. 63, p. 299–322 Biology and Management of Coregonid Fishes – 2008
Transcript of A preliminary assessment of cisco (Coregonus spp.) diversity in Yellowknife Bay, Great Slave Lake,...

A preliminary assessment of cisco (Coregonus spp.) diversity in Yellowknife Bay, Great Slave Lake, Northwest Territories
Paul Vecsei1, Craig T. Blackie1,2, Andrew M. Muir1,3*, Hilary M. Machtans1, J.D. Reist4
with 8 fi gures and 5 tables
Abstract: Northern freshwater fi shes, particularly members of the genus Coregonus, have been used as a model for studying microevolution due to their well known colonization history coupled with parallel patterns of phenotypic differentiation. We assessed and described the phenotypic diversity among ciscoes in the Yellowknife Bay region of Great Slave Lake using 1) traditional taxonomic methods for cisco species and 2) contemporary linear and geometric morphometric methods. Four morphs were identifi ed using a qualitative visual assessment of gross morphology and/or location of capture in comparison to published taxonomy for the genus Coregonus. The morphs identifi ed were assigned the following colloquial names: A) ‘adfl uvial’, B) ‘lacustrine’, C) ‘shortjaw’, and D) ‘least’. Univariate analyses on taxonomic measures indicated that cisco varied in terms of age, number of lateral line scales, number of anal fi n rays, middle gill raker length, and lower gill arch length. By contrast, fi ve morphs were identifi ed using cluster analysis of linear and geometric morphometric data combined with age and gill raker number. The additional phenotypic cluster entailed a splitting of large and small lacustrine morphs into two groups. Multivariate analyses indicated a strong relationship between body shape and gill raker number, eye size, age and mouth morphology. These results suggested that the segregation among morphs was based on differential usage of trophic and/or habitat resources as well as life history variation such as age at maturity. Our results indicated that the level of phenotypic diversity was greater than previously documented for Great Slave Lake and that this diversity was similar to that reported for ciscoes in the Laurentian Great Lakes.
Keywords: Coregonus spp., cisco, Great Lakes, morphometrics, divergence, polymorphism.
Authors’ addresses:1 Golder Associates Ltd. 9-4905 48th Street, Yellowknife, Northwest Territories, X1A 3S3, Canada.2 Biology Department, Dalhousie University, 1355 Oxford St., Halifax, Nova Scotia, B3H 4J1,
Canada.3 Current address: Great Lakes Fishery Commission, 2100 Commonwealth Blvd., Suite 100, Ann
Arbor, MI, 48105, USA.4 Arctic Fish Ecology and Assessment Research Section, Fisheries and Oceans Canada, Winnipeg,
Manitoba, R3T 2N6, Canada.* Corresponding author, e-mail: [email protected]
1612-166X/12/0063-0299 $ 6.00© 2012 E. Schweizerbart’sche Verlagsbuchhandlung, 70176 Stuttgart, Germany
Advanc. Limnol. 63, p. 299–322Biology and Management of Coregonid Fishes – 2008

300 P. Vecsei et al.
Introduction
Discerning the patterns and processes that drive ecological divergence is crucial for under-standing microevolution. Northern freshwater fi shes are prominently featured in the scien-tifi c literature of microevolution because they are evolutionarily young (i.e., <10,000 y), gen-erally have a well understood colonization history (TAYLOR 1999), and occur in systems that have low fi sh species richness (ROBINSON & PARSONS 2002). Therefore, sympatric morphs are believed to be the result of divergent natural selection, largely facilitated by increased eco-logical opportunity (ROBINSON & SCHLUTER 2000). Interestingly, striking phenotypic conver-gence occurs among a broad range of northern genera in both form and function relationships (often referred to as resource polymorphisms; SKULASON & SMITH 1994). Despite a complex evolutionary history involving colonization from multiple glacial refugia (BENATCHEZ & WIL-SON 1998), much of the phenotypic diversity in northern fi shes appears to have arisen in situ via common selection pressures (ROGERS & BERNATCHEZ 2005). To understand the processes that promote adaptive divergence, it is important to consider the patterns of morphological differentiation among geographically distinct systems.
Diversifi cation among North American coregonid fi shes (Coregonus spp.) is one example that appears to be the result of recent adaptive radiation (BERNATCHEZ 2004). Cisco (Coregon-us artedi), in particular, exhibit considerable life history, ecological, and morphological vari-ation throughout their range (MCPHAIL 2007, TODD et al. 1981, TODD & SMITH 1980, CLARKE 1973, DYMOND 1943, KOELZ 1929). Previous studies of cisco morphological diversifi cation have been confi ned to the Laurentian Great Lakes basin (e.g., KOELZ 1929, TURGEON et al. 1999 & TODD & SMITH 1980), where parallel patterns of morphological diversifi cation have been suggested. By contrast, the taxonomy and evolutionary history of the cisco complex in Great Slave Lake, Northwest Territories has received relatively less attention (however, see: HARPER & NICHOLS 1919, RAWSON 1951, CLARKE 1973, STEINHILBER et al. 2002, TODD and STEINHILBER 2002, MURRAY & REIST 2003, MURRAY 2006). In comparison to the Laurentian Great Lakes, inter- and intra-specifi c taxonomic distinctions have not been made for the cisco in Great Slave Lake. Uncertainty has raised conservation concerns over the management of current commercial and recreational fi sheries. Thus, it is important to inventory the amount of phenotypic diversity within Great Slave Lake ciscoes and to discern how this variation is partitioned into particular forms (i.e., morphs or species). Moreover, Great Slave Lake is relatively unperturbed. Although a substantial commercial fi shery has existed on the lake since 1945, exploitation has been primarily concentrated in the main basin of the lake. In addition, Great Slave Lake has been minimally affected by anthropogenic development and contains an intact assemblage of native species. As such, Great Slave Lake provides a rare opportunity to study ecological divergence in a more ‘natural’ setting in comparison to the Laurentian Great Lakes, where much of the native diversity has been lost.
The objectives of our study were twofold: 1) provide a preliminary assessment of cisco diversity in the Yellowknife Bay region of Great Slave Lake, and 2) provide a preliminary description of the variation among cisco morphs within the lake. To achieve these objectives we employed two complementary approaches. First, we grouped cisco into taxonomic units using traditional methods such as visual assessment and basic statistical assessment of lin-ear measures and meristic counts of key characteristics. Second, we partitioned phenotypic variation into groups using geometric morphometric methods. We compared traditional and

Cisco diversity in Yellowknife Bay, Great Slave Lake 301
morphometric grouping methods and discussed aspects differentiating morphs within the context of trophic specialization and parallel divergence in the Laurentian Great Lakes.
Methods
Study site
Great Slave Lake covers an area of about 28,000 km2 and is the tenth largest lake in the world and the third largest lake in Canada (EVANS et al. 2000). It is located within the Mackenzie River Basin, approxi-mately 230 km south of the Arctic Circle (Fig. 1) and can be subdivided into a central main basin, and northern and eastern arms. The central main basin is relatively shallow, turbid and nutrient rich, while the northern and eastern arms are deep and highly oligotrophic (RAWSON 1950). Yellowknife Bay, situ-ated in the north arm, is 18 km long and 3 km at its widest point. The Yellowknife River is the largest inlet draining into the northernmost portion of Yellowknife Bay (Fig. 1).
Ciscoes were sampled from three sites in Yellowknife Bay: 1) Kam Point (62°23’32.42’’ – 114°22’08.05’’); 2) Sub Islands north (62°22’20.73’’ – 114°21’30.25’’); and 3) Sub Islands south (62°21’30.54’’ – 114°22’40.48’’) and one site in the Yellowknife River: 1) Tartan Rapids (62°33’32.12’’ – 114°13’05.67’’; Fig. 1). Sample sites were selected using local fi shers’ knowledge of bathymetric features and cisco distribution in combination with the Canadian Hydrographic Service chart of Yel-lowknife Bay (chart number 6369).
Fig. 1. Cisco sampling sites on Great Slave Lake, NWT; Kam Point (latitude 62°23ʹ32.42ʺ; longitude 114°22ʹ08.05ʺ); Sub Islands north (latitude 62°22ʹ20.73ʺ; longitude 114°21ʹ30.25ʺ) and Sub Islands south (latitude 62°21ʹ30.54ʺ; longitude 114°22ʹ40.48ʺ); and Tartan Rapids (latitude 62°33ʹ32.12ʺ; longitude 114°13ʹ05.67ʺ).

302 P. Vecsei et al.
Fish collections
Fifty-two cisco were dip-netted from Tartan Rapids during October 2006 (Fig. 1). This mass movement of fi sh into the river is apparently a spawning migration, but specifi c spawning locations are currently unknown. Water depth at the collection site was approximately 1 m. One hundred and nineteen cisco were collected from three sites in Yellowknife Bay between April and July 2008 using gillnets (100 mlong by 2 m deep) ranging from 5-cm to 10-cm stretch mesh. Nets were deployed on bottom for a maxi-mum of 24 h at depths ranging from 20 to 50 m.
Field processing of fi sh
A calibrated digital image of the left side of each individual was captured using a Nikon D200 camera (Nikon Inc, Toronto, Canada). A long focal-length lens (i.e., Nikor 17-55 ED set at 55-mm focal length [equivalent in SLR to 1.5 x 55 mm]) was used to eliminate potential parallax distortion. Macro images of key morphological and meristic features, such as the fi ns and head were also captured for illustrative purposes. Fork length (FL±1mm) and whole body weight (±0.5g) were measured for each individual, with the exception of the Tartan Rapids fi sh, which were not weighed. Cisco were individually labelled and frozen at -20 °C prior to laboratory analysis.
Laboratory processing of fi sh
Frozen fi sh were thawed. Three meristic variables were enumerated (i.e., lateral line scales, anal rays, and gillrakers). Scales in the lateral line were enumerated between the posterior edge of the opercle and the terminus of the hypural complex; when scales were missing due to handling or net damage, scale pockets were counted. To enumerate the number of rays in the anal fi n, the anterior rays were excluded unless they were two-thirds the length of the longest ray. Where the shortest ray was split at its base, it was counted as a single ray. The fi rst left gill arch was extracted and rakers were enumerated under a Leica ES2 dissection scope (Leica Microsystems Inc., Richmond Hill, Ontario) using refl ected light at a magnifi cation of either 10X or 30X, depending on gill raker size. All rakers were enumerated including rudimentary rakers at either end of the arch. Two linear morphometric measurements were recorded—longest gill raker and lower gill arch. The longest gill raker, which we defi ned as the fi rst raker of the lower arch where it joined the upper and lower arches, was measured point-to-point using a digital caliper (± 0.01 mm). After external measurements were made, sex was recorded and sagittal otoliths were extracted for age estimation. Ninety-eight percent of the individuals in the sample were sexually mature.
Age estimation
Ages were estimated by John Tost, North Shore Environmental Services, Thunder Bay, Ontario using the otolith crack-and-burn technique. Otoliths were cracked through the transverse plane to expose growth increments and the cracked surface was charred using an alcohol burner to highlight growth zones. Otoliths were analyzed under a Wild Leitz M3Z dissection microscope (Wild Leitz Instruments, Heidelberg, Germany) using refl ected light at 6.5X–40X magnifi cation; mineral oil was used as a clear-ing agent to accentuate annuli. Annual increments were defi ned as an opaque zone (i.e., summer grow-ing season) followed by a narrow translucent zone (i.e., winter growing season; HOWLAND et al. 2004).

Cisco diversity in Yellowknife Bay, Great Slave Lake 303
Taxonomy
Both traditional (i.e., visual and basic statistical assessment of key characters) and quantitative (i.e., geometric morphometric) methods were used to assign individuals to morphs. We used the ‘morph’ designation, which we defi ned as a ‘form’ or ‘variant’ (COAD & MCALLISTER 2009), to group cisco into distinguishable units due to the taxonomic uncertainty of this particular group. For the traditional method, individuals were grouped into morphs based on habitat of capture (i.e., adfl uvial versus lacus-trine) and a suite of six external characters (i.e., dorsal colouration, dorsal head shape, mouth position, mandible length, eye size, and fi n length; Table 1) that were visually assessed from digital images. Ini-tial classifi cations were subsequently verifi ed using meristic data (i.e., gill raker number). Colouration was visually assessed and designated using the standardized hex-triplet system (http://en.wikipedia.org/wiki/List_of_colors).
To describe variation in external gross morphology and variation in appearance among morphs, a series of colour illustrations were created from enlarged digital images (all illustrations by P. Vecsei). Colour images were printed, placed on a light table, and the outline was transferred to cotton archival two-ply vellum (Strathmore Artist Papers, Westfi eld, Massachusetts). Illustrations were drawn using a combination of graphite and polychromos colour pencils (Faber-Castell, Cleveland, Ohio) and fi nished with multiple wash layers of water colour pencil.
Geometric morphometrics
To quantify differences in shape among morphs and to validate traditional assessments, variation in body shape was analyzed using geometric methods (ZELDITCH et al. 2004, ROHLF AND BOOKSTEIN 2003, BOOKSTEIN 1989). Shape data were collected from 16 homologous and four semi-landmarks using the Thin Plate Spline suite of software (TPS; State University of New York at Stony Brook; http://life.bio.sunysb.edu/morp; Fig. 2). Landmarks were placed on digital images using the program TPSdig. Semi-landmarks (17–20 on Fig. 2) were non-homologous points that were placed at points situated at 0.20, 0.30, 0.40, and 0.50*standard length to quantify body depth. To facilitate semi-landmark placement, a grid was layered over the image. Landmark data were then used to obtain relative warp scores using TPSrelw. To complement the data set, eye diameter, pectoral fi n length, maxillary length, and mouth orientation were quantifi ed from digital images using TPSdig (see TURGEON et al. 1999). Eye diameter was measured from the anterior to the posterior edge of the orbit. Pectoral fi n length was measured from
Table 1. Suite of characters used to visually assign cisco to one of four morphs: A) adfl uvial; B) lacustrine; C) shortjaw; and D) least.
MorphCharacter A B C DHabitat of capture adfl uvial lacustrine lacustrine lacustrineDorsal colouration blue-black or
greenolive-brown or
greencinnamon brown cobalt blue
Dorsal head shape fl at fl at convex fl atEye size moderate small moderate largeFin length short short moderate longMouth position terminal terminal subterminal or
terminalsuperior
Mandible length even with premaxilla
extended past premaxilla
does not extend past premaxilla
extended well past premaxilla

304 P. Vecsei et al.
the origin to the end of the fi rst ray. Maxillary length was measured from the tip of the snout to the pos-terior margin of the maxillary. Mouth orientation was measured as the angle of the outer premaxillary relative to the lateral plane (i.e., parallel to the lateral line) using the compass tool in TPSdig. Prior to analysis, all linear measurements were size corrected following REIST (1985).
Statistical methods
To support traditional taxonomic methods, a series of univariate analyses of covariance (ANCOVA) were conducted. These analyses tested for differences in the number of anal fi n rays, lateral line scales, gill rakers, lengths of longest raker and lower gill raker arch, and age among morphs. Fork length was used as a covariate. The signifi cance criterion for multiple comparisons was corrected using the Bonfer-roni method. To examine general trends in size-at-age among morphs, mean length-at-age was plotted and visually assessed.
Two separate statistical analyses were undertaken to examine taxonomic structure using morphomet-rics. The number of unique cisco morphs in Yellowknife Bay was fi rst assessed with a K-means cluster analysis using the 36 partial warp scores for body shape, select linear measurements (i.e., fork length, eye diameter, pectoral fi n length, and maxillary length), gill raker number, and mouth orientation as independent variables. For this analysis, the number of unique morphs present was not assumed. A range of K values (i.e., number of potential clusters) from 2 to 10 were assessed using three replicates. Euclidean distances were measured between group centroids and 100 fi tting iterations were run. The resulting within and among cluster sum of squares (SS) were averaged among replicates and the respec-tive curves were plotted and assessed visually to identify the most parsimonious number of groups. This clustering approach minimized the within cluster SS while maximizing among cluster SS. As such, the most suitable number of clusters was expected to correspond to a minimum rate of change in slope of a curve fi t to the regression of among (and within) cluster SS and number of clusters present. Asymptotes were located visually and used to estimate the number of clusters (i.e., unique morphs). A discriminant function analysis (DFA) tested how well phenotypic traits clustered individuals into groups. Only body shape data (i.e., 36 partial warps) were used for this analysis and the validity of the resulting group structure was assessed using jackknife classifi cation where successful individual assignment back to groups was quantifi ed.
Fig. 2. Sixteen homologous and four semi-landmarks used for geometric morphometric analyses. Note that semi-landmarks represent a proportion of standard length (i.e., 20, 30, 40 & 50%).

Cisco diversity in Yellowknife Bay, Great Slave Lake 305
A correlation analysis assessed whether particular phenotypic traits were associated with body shape variation. For this analysis, Pearson correlation coeffi cients were computed for the associations between body shape as represented by DFA scores along the fi rst four axes and linear measures of phe-notype described by fork length, eye diameter, pectoral fi n length, maxillary length, gill raker number, and mouth orientation. To visualize how body shape varied in relation to other morphometric and meristic variables, a PCA was conducted on select linear measures and meristic counts and partial warps from the shape analysis were subsequently regressed on the resulting PCA scores using TPS regress. To further investigate how body shape varied in relation to particular phenotypes, partial warps were regressed onto eye diameter, pectoral fi n length, maxillary length, gill raker number, and mouth orientation in independent analyses using TPS regress. This analysis measured how body shape var-ied in response to all linear measures and meristic counts combined. Deformation grids were visually assessed. All statistical analyses (signifi cance level was set at α = 0.05) were conducted using SPSS 14.0 (SPSS Inc., Chicago, Illinois), SigmaStat 3.5 (Systat Software Inc., San Jose, California), and SYSTAT 12 (Systat Software, a subsidiary of Cranes Software International Ltd., Chicago, Illinois).
Results
Cisco in the Yellowknife Bay region of Great Slave Lake varied in terms of habitat use (i.e., riverine versus lacustrine), external gross morphology, body shape, and colouration (Fig. 3 a-d). On the basis of a qualitative assessment of digital images, ciscoes were categorized into four morphs. For descriptive purposes, the following colloquial names were given to each morph: A) ‘adfl uvial’, B) ‘lacustrine’, C) ‘shortjaw’, and D) ‘least’.
Age (ANCOVA; F3,143 = 11.34; P < 0.001), number of scales in the lateral line (ANCOVA; F3,159 = 6.22; P = 0.001), number of anal fi n rays (ANCOVA; F3,168 = 17.76; P < 0.001), number of gill rakers (ANCOVA; F3,169 = 62.04; P < 0.001), middle gill raker length (ANCOVA; F3,168
= 35.69; P < 0.001), and lower gill arch length (ANCOVA; F3,168 = 57.31; P < 0.001) differed signifi cantly among the four morphs. The nature of these differences is presented below in more detail under the subheading for each morph.
Fig. 3a. Morph A (adfl uvial) from Tartan Rapids, Yellowknife River, 215 mm FL. Illustration P. Vecsei Golder-DFO © 2008.

306 P. Vecsei et al.
Fig. 3b. Morph B (lacustrine) from Sub Islands, Yellowknife Bay, 398 mm FL. Illustration P. Vecsei Golder-INAC-DFO © 2008. Note: the specimen illustrated was one of the large forms identifi ed by geometric morphometrics as a separate group (Morph E – see text).
Fig. 3c (i). Morph C (shortjaw) from Sub Islands, Yellowknife Bay, 285 mm FL. Illustration P. Vecsei Golder-INAC-DFO © 2008. The specimen illustrated was the more typical form of this morph.
Fig. 3c (ii). Morph C (shortjaw) from Sub Islands, Yellowknife Bay, 236 mm FL. Illustration P. Vecsei Golder-DFO © 2008. The specimen illustrated was an atypical form of this morph; note differences in head and snout arrangement.

Cisco diversity in Yellowknife Bay, Great Slave Lake 307
Fig. 3d (i). Morph D (least) from Sub Islands, Yellowknife Bay, 298 mm FL. Illustration P. Vecsei Golder-INAC © 2008.
Fig. 3 d (ii). Morph D (least) from Sub Islands, Yellowknife Bay, 280 mm FL. Illustration P. Vecsei Golder-DFO © 2008.
Traditional taxonomic assessments
Morph A (adfl uvial)
Fifty-two reproductively mature individuals (n = 38 males and 14 females) were captured in the Yellowknife River and classifi ed as Morph A (Fig. 3a). Adults of this morph were consistently small in comparison to mature individuals of the other morphs (Table 2). Cisco classifi ed as Morph A tended to be more elongate in body shape, and typically had terminal mouths with the lower mandible even with or extending slightly beyond the premaxilla when the mouth was closed. Morph A had a less sloped head profi le (i.e., fl at) and an anteriorly pigmented mandible and maxilla (i.e., dark chin and dark snout). Typically, the paired fi ns were immaculate (i.e., lacking pigmentation), the dorsal fi n had dark black pigment along the
308 P. Vecsei et al.
membrane between the rays, and the caudal fi n was darkly pigmented, but lacked the black outer margin that was typical of other morphs. The anal fi n was either immaculate or had light pigmentation towards the posterior margin. Dorsal colouration of Morph A consisted of a combination of myrtle (21421E), dark green (013220) and army green (4B5320) or blue (pigment; 333399), cobalt (0047AB), dark slate grey (2F4F4F), indigo blue (00416A), midnight blue (191970), Persian indigo (32127A), Prussian blue (003153), and sapphire (082567). Atmospheric conditions and the iridescent nature of the scales greatly infl uenced how colour was perceived.
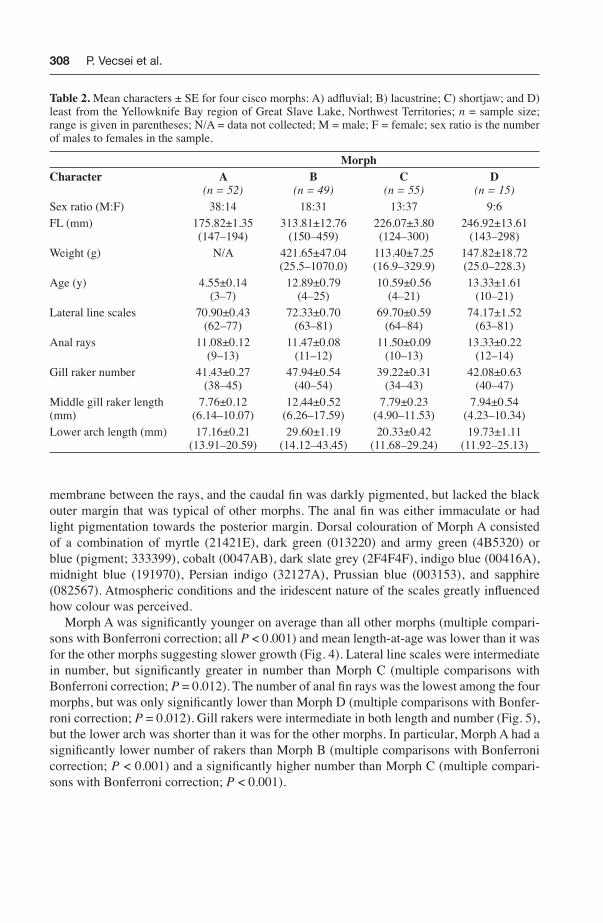
Morph A was signifi cantly younger on average than all other morphs (multiple compari-sons with Bonferroni correction; all P < 0.001) and mean length-at-age was lower than it was for the other morphs suggesting slower growth (Fig. 4). Lateral line scales were intermediate in number, but signifi cantly greater in number than Morph C (multiple comparisons with Bonferroni correction; P = 0.012). The number of anal fi n rays was the lowest among the four morphs, but was only signifi cantly lower than Morph D (multiple comparisons with Bonfer-roni correction; P = 0.012). Gill rakers were intermediate in both length and number (Fig. 5), but the lower arch was shorter than it was for the other morphs. In particular, Morph A had a signifi cantly lower number of rakers than Morph B (multiple comparisons with Bonferroni correction; P < 0.001) and a signifi cantly higher number than Morph C (multiple compari-sons with Bonferroni correction; P < 0.001).
Table 2. Mean characters ± SE for four cisco morphs: A) adfl uvial; B) lacustrine; C) shortjaw; and D) least from the Yellowknife Bay region of Great Slave Lake, Northwest Territories; n = sample size; range is given in parentheses; N/A = data not collected; M = male; F = female; sex ratio is the number of males to females in the sample.
MorphCharacter A
(n = 52)B
(n = 49)C
(n = 55)D
(n = 15)
Sex ratio (M:F) 38:14 18:31 13:37 9:6FL (mm) 175.82±1.35
(147–194)313.81±12.76
(150–459)226.07±3.80(124–300)
246.92±13.61(143–298)
Weight (g) N/A 421.65±47.04(25.5–1070.0)
113.40±7.25(16.9–329.9)
147.82±18.72(25.0–228.3)
Age (y) 4.55±0.14(3–7)
12.89±0.79(4–25)
10.59±0.56(4–21)
13.33±1.61(10–21)
Lateral line scales 70.90±0.43(62–77)
72.33±0.70(63–81)
69.70±0.59(64–84)
74.17±1.52(63–81)
Anal rays 11.08±0.12(9–13)
11.47±0.08(11–12)
11.50±0.09(10–13)
13.33±0.22(12–14)
Gill raker number 41.43±0.27(38–45)
47.94±0.54(40–54)
39.22±0.31(34–43)
42.08±0.63(40–47)
Middle gill raker length (mm)
7.76±0.12(6.14–10.07)
12.44±0.52(6.26–17.59)
7.79±0.23(4.90–11.53)
7.94±0.54(4.23–10.34)
Lower arch length (mm) 17.16±0.21(13.91–20.59)
29.60±1.19(14.12–43.45)
20.33±0.42(11.68–29.24)
19.73±1.11(11.92–25.13)

Cisco diversity in Yellowknife Bay, Great Slave Lake 309
Fig. 4. Mean fork length (mm) at age (y) ± SE for adfl uvial and lacustrine morphs (panel A) and shortjaw and least morphs (panel B); morphs were identifi ed using traditional taxonomic methods from the Yellowknife Bay region of Great Slave Lake.

310 P. Vecsei et al.
Morph B (lacustrine)
Forty-nine individuals (n = 18 males, 27 females, and 4 unknown) were captured in Yel-lowknife Bay and classifi ed as Morph B (Fig. 3b). Morph B adults attained the largest body size and the greatest mean size-at-age (Fig. 4) among the four morphs, but both length and weight were highly variable (Table 2). Cisco classifi ed as Morph B tended to be more ellipti-cal in body shape, typically had terminal mouths with the lower mandible extending beyond the premaxilla, had a less sloped head, and lightly pigmented mandible and maxilla. The paired fi ns were immaculate or lightly pigmented and had a prominent black margin along the distal edge of the dorsal and caudal fi ns, especially on larger specimens. Light pigmenta-tion was often observed on the membrane of the anal fi n. In general, dorsal colouration of Morph B was a combination of army green (4B5320) and viridian (40826D). Minor colours associated with iridescence and pigmentation included olive (80800), olive drab (6B8E23), olivine (9AB973), sea green (2E8B57), Emerald (50C878), dark spring green (177245), and myrtle (21421E).
Mean age and the number of lateral line scales were intermediate and did not differ statisti-cally from other morphs. Anal fi n-ray number was the second lowest among the four morphs, but was only statistically lower than Morph D (multiple comparisons with Bonferroni cor-rection; P < 0.001). Morph B had a signifi cantly greater number of gill rakers than all other
Fig. 5. Gill raker frequency for four cisco morphs in the Yellowknife Bay region of Great Slave Lake.

Cisco diversity in Yellowknife Bay, Great Slave Lake 311
morphs (multiple comparisons with Bonferroni correction; all P < 0.001; Fig. 5). Neither gill raker length nor lower arch length differed between Morphs A and B; however, Morph B had signifi cantly longer rakers (multiple comparisons with Bonferroni correction; all P < 0.001) and a signifi cantly longer lower arch (multiple comparisons with Bonferroni correction; all P < 0.01) than Morphs C and D.
Morph C (shortjaw)
Fifty-fi ve individuals (n = 16 males, 32 females, and 7 unknown) were captured in Yellow-knife Bay and classifi ed as Morph C (Fig. 3c i-ii). Adults of this morph were intermediate in mean size-at-age (Fig. 3) among the four morphs (Table 2). Morph C was the deepest anteriorly and more ovate laterally in comparison to the other morphs. The mouth was either sub-terminal or terminal, the head was more convex, and there was light pigmentation on the mandible and maxilla. Paired and anal fi ns were immaculate, but the distal edge of the dorsal and caudal fi ns ranged from being lightly shaded to exhibiting a conspicuous black margin. In general, the dorsum of Morph C was a combination of tan (D2B48C), raw umber (734A12) and cinnamon (D2691E). Minor colours included dark chestnut (986960), dark goldenrod (B8860B), dark tan (918151), Ecru (C2B280), khaki (C3B091), lavender grey (C4C3D0), and mauve taupe (915F6D).
On average, Morph C was signifi cantly older than Morph A (multiple comparisons with Bonferroni correction; P < 0.001), but did not differ in mean age from the other morphs. The number of lateral line scales was signifi cantly lower than it was for Morphs A and D (multiple comparisons with Bonferroni correction; all P < 0.01). Anal fi n-ray number did not differ from Morphs A and B, but was statistically lower than for Morph D (multiple comparisons with Bonferroni correction; P < 0.001). Gill raker number and length did not differ between morphs C and D, but morph C had signifi cantly fewer and shorter rakers that Morphs A and B (multiple comparisons with Bonferroni correction; all P < 0.001). The lower arch length was intermediate in comparison to the other morphs, and was signifi cantly shorter than Morphs A and B (multiple comparisons with Bonferroni correction; P = 0.21 and P = 0.18, respectively) and signifi cantly longer than Morph D (multiple comparison with Bonferroni correction; P = 0.001).
Morph D (least)
Fifteen individuals (n = 9 males and 6 females) were captured in Yellowknife Bay and clas-sifi ed as Morph D (Fig. 3d i-ii). Adults of this morph were intermediate in mean size-at-age (Fig. 4) among the four morphs (Table 2). The eyes were set forward on the head, large, and often extended beyond the dorsal outline of the head when viewed laterally. The interorbital distance was strikingly short, giving the appearance of the eyes being close together when viewed from the dorsal aspect. Multiple prominent morphological features distinguished this morph from the others. Morph D had a terminal mouth, but because of the lower mandible angle and that the tip of the dentary extended well past the premaxilla, the mouth appeared superior. Morph D appeared slender overall when viewed laterally, but was noticeably robust

312 P. Vecsei et al.
in the lumbar region compared to the more elliptical or anteriorly ovate sympatric cisco. The dorsal aspect of the head appeared fl atter in profi le, exhibiting a straight line from the pre-maxilla to the occipital, when compared to the other morphs (Fig. 3d).
The mandible and premaxilla of Morph D were darkly pigmented in comparison to the other morphs. The dorsal border and outer margin of the pectorals were lined with black pigment. The pelvic fi ns were conspicuously pigmented with a distinct black lateral band along the mid to distal portion of the structure. Anal fi n pigmentation was moderate to heavy, particularly on the membrane and often along the outer margin of the fi n. In general, the dorsum of Morph D consisted of a combination of slate grey (2F4F4F), charcoal (464646), dark green (013220), indigo (dye; 00416A), Prussian blue (003153), and sapphire (082567).
Fin length was a key feature that distinguished Morph D. For instance, all paired fi ns appeared large relative to other sympatric cisco. The dorsal fi n appeared shorter at its base, but the fi rst and second rays were long, giving it a ‘tall’, ‘sail-like’ appearance. The anal fi n was longer at its base when compared with sympatric cisco, likely a result of the greater number of rays. The caudal fi n appeared slightly more lunate and lacked the distinct paddle like or ‘lobate’ shape in the upper and lower lobe associated with Morph C.
Mean age was signifi cantly higher for Morph D than that for Morph A (multiple compari-son with Bonferroni correction; all P = 0.038), but not statistically different from the other two morphs. Anal ray number was signifi cantly higher than it was for all other morphs (mul-tiple comparisons with Bonferroni correction; all P < 0.001). Morph D had a signifi cantly greater number of gill rakers than Morph C and signifi cantly lower number of rakers than Morph B (multiple comparisons with Bonferroni correction; all P < 0.001). Morph D had the shortest gill rakers of the morphs and they differed signifi cantly from Morphs A and B (multiple comparisons with Bonferroni correction; all P < 0.001). The lower gill raker arch was signifi cantly lower than all other morphs (multiple comparisons with Bonferroni correc-tion; all P < 0.01).
Morphometric taxonomic assessment
In contrast to the traditional assessment, in which four morphs were identifi ed, the morpho-metric analyses supported the existence of fi ve morphs. Large and small lacustrine forms of Morph B were distinguished from one another using this analytical method. For the pur-pose of reporting, we refer to the large lacustrine form as Morph E. Averaged among cluster K-means SS values ranged from 8000 for K = 2 to 22,000 for K = 10 (Fig. 6). The averaged within cluster SS ranged from 18,000 for K = 2 to 4,000 for K = 10. For both curves, the asymptote corresponded to K = 5, indicating that fi ve morphs were most likely present in the sample. Individuals were assigned to one of the fi ve morphs using a hierarchical cluster analysis. These clusters were tentatively named on the basis of results from the descriptive portion of the study using information such as location of capture, body size, and gill raker number. These morph assignments formed the basis for testing how well particular traits defi ned the groups.
The resulting clusters were well supported by the DFA model (F164, 530 = 9.75; P < 0.001; Tables 3, 4, and Fig. 7). Overall, 79% of the variation in body shape was explained along the fi rst two DF axes. Morph B (small lacustrine) had the lowest jackknife classifi cation success

Cisco diversity in Yellowknife Bay, Great Slave Lake 313
Fig. 6. The number of cisco morphs present in the Yellowknife Bay region of Great Slave Lake, NT was determined based on sum of squares values from K-means cluster analyses; (a) illustrates among cluster sum of squares while (b) illustrates within cluster sum of squares values.
Table 3. Discriminant function analysis results of phenotypic variation for Yellowknife Bay (Great Slave Lake) and Yellowknife River cisco (Coregonus sp.) by predetermined clusters.
Variable Eigenvalue Canonical correlation
Wilk’sLambda
F Jackknifed classifi cation success
Assigned cluster
7.2184.1732.3030.594
0.9370.8980.8350.594
0.005 9.750df (164,530)P < 0.001
Clusters:Lacustrine – 52%Large lacustrine – 87%Least – 93%Shortjaw – 84%Adfl uvial – 96%_______________Total: 84%
Table 4. Jackknifed classifi cation matrix for individual cisco from Yellowknife Bay region of Great Slave Lake based assigned groups from K-means cluster analysis; for Morph A) adfl uvial; B) small lacustrine; C) shortjaw; D) least; and E) large lacustrine; %correct = the number of individuals correctly assigned to each of the four morphs.
Morph A B C D E % CorrectA 48 1 1 0 0 96B 1 12 8 0 2 52C 3 5 47 0 1 84D 1 0 0 14 0 93E 0 5 0 0 33 87
(52%) whereas 96% and 93% of Morph A (adfl uvial) and Morph D (least) were correctly classifi ed to their respective groups (Table 4). The majority of the misclassifi ed Morph B were assigned to the Morph C (shortjaw) cluster. Also, all misclassifi ed Morph E (large lacus-trine) individuals were assigned to Morph B (small lacustrine; Table 4).
Traditional linear measures of phenotypic variation were correlated to variation in body morphology along DF axis-1. In particular, variation in body shape along DF axis-1 was

314 P. Vecsei et al.
Table 5. Pearson correlation coeffi cients between traditional measures of phenotype and geometric shape indices (DF scores) for ciscoes (Coregonus sp.) in Yellowknife Bay region, Great Slave Lake. Aspects of body shape separating morphs along DF1 are correlated with age, gill raker number, and eyes size. Along DF2, mouth orientation is correlated with differences in body shape.
FunctionVariable 1 2 3 4Age 0.629 0.155 0.015 -0.107Gillraker number 0.537 0.264 -0.408 -0.116Pectoral fi n length -0.036 0.090 0.100 -0.038Eye diameter -0.655 0.214 0.080 0.230Maxillary length 0.020 -0.006 -0.262 0.070Mouth orientation 0.188 0.337 0.048 0.019
Fig. 7. Discriminant analysis of body shape among Great Slave Lake cisco morphs grouped by K-means clustering on body morphology and traditional taxonomic characters. Elipses were hand-drawn to depict grouping structure.
associated with gill raker number, eye size, and age (all r > 0.5; Table 5). Variation in body shape along DF axis-2 was primarily associated with differences in mouth morphology. Gill raker number and eye diameter were the two variables that separated most clearly along PC1 and PC2 with highly negative scores associated with lower gill raker number and smaller eyes and highly positive scores associated with higher gill raker number and larger eyes. When body shape variation was superimposed on PC axes for the linear measures and counts, the traditional taxonomic descriptions that we assigned in the fi rst part of the study were

Cisco diversity in Yellowknife Bay, Great Slave Lake 315
generally reinforced (Fig. 8). Differences in the positioning of the 20 landmarks were obvious as one moved from morph to morph along a continuum of body shape variation. For example, shape landmarks for Morph A (adfl uvial; left panel in Fig. 8a) were generally compressed along all axes of the body in comparison to the more robust Morph E (large lacustrine; right panel in Fig. 8a) as one moves along a gradient of phenotypic variation.
Discussion
The presence of multiple morphs in the Great Slave Lake drainage
Both traditional and geometric morphometric analyses of phenotypic traits supported the hypothesis that multiple cisco forms exist in the Yellowknife Bay region of Great Slave Lake. Previous researchers (HARPER & NICHOLS 1919, CLARK 1973, RAWSON 1951, ROBERGE et al. 1985, STEINHILBER et al. 2002) who have examined the taxonomy of Great Slave Lake ciscoes have found inconclusive and in some cases confl icting results. For example, RAWSON (1951) reported that at least three cisco species existed in Great Slave Lake—these collections were confi rmed by J.R. Dymond to be lake cisco (Coregonus artedi), a morph that resembled blackfi n cisco (C. nigripinnis), and a large morph that could not be identifi ed at that time. These specimens were deposited in the Royal Ontario Museum and subsequently assessed by T. Todd (USGS Great Lakes Science Center, personal communication), R. Eshenroeder (Great Lakes Fishery Commission), and N. Mandrak (Fisheries and Oceans Canada), who suggested that the collection contained C. artedi, C. nigripinnis, C. zenithicus, and possibly C. sardinella. By contrast, MURRAY and REIST (2003) were unable to confi rm the presence of C. zenithicus in Great Slave Lake using a suite of over 30 morphometric and meristic charac-ters on cisco obtained from (1) Hay River in the western main basin, (2) Lutsel’ke in the east arm, and (3) Simpson Islands in the east arm.
Fig. 8. Aspects of geometric morphometric shape variation among cisco in Yellowknife Bay, Great Slave Lake. Panel (a) illustrates differences in body depth observed between Morph E (large lacustrine; right illustration) and other cisco (left illustration), while panel (b) illustrates differences in mouth orientation which was a key feature discriminating Morphs C (left illustration) and D (right illustration).

316 P. Vecsei et al.
The preliminary analyses undertaken herein support the idea that the degree of cisco diver-sity in Yellowknife Bay, Great Slave Lake, is greater than previously described. Visual assign-ment to groups using traditional taxonomic criteria, such as gross morphology support the existence of four morphs, whereas geometric morphometric analyses support the existence of fi ve morphs. The additional morph identifi ed using the latter approach separated Morph B into a small lacustrine form that exhibited a lower number of gill rakers (i.e., Morph B) and a larger lacustrine form with a higher number of gill rakers (Morph E), despite controlling for the effect of body size in the analysis. This was also evidenced by a bimodal gill raker dis-tribution (Fig. 5) with modes around 44 and 50. However, the existence of Morph E was not well supported due to the high misclassifi cation rate. Individuals in Morph E were most com-monly misclassifi ed as Morph B, suggesting that the phenotypic distinction between these two morphs may be questionable. Given that Morph B was represented by small individu-als, while Morph E consisted primarily of large individuals, phenotypic differentiation may develop through ontogeny. Therefore, Morph B may represent earlier life stages of Morph E. The latter possibility is supported by the observation that high gill raker counts were always attributed to larger and older members of the group. If small, low-raker and large, high-raker forms of the lacustrine morph do indeed exist, then this raises some interesting questions. For instance, why did we not capture any small, high-raker forms of this morph in either the bay or the river? The possibility that cisco develop additional gill rakers through ontogeny would contradict previous heritability estimates for this trait (TODD et al. 1981), but would be consistent with the coexistence of small and large cisco morphs in Lake Cormorant and Lake Ryan, Alberta (TURGEON & BERNATCHEZ 2003). Phenotypic shifts during ontogeny are believed to be common among morphs of northern freshwater fi shes (PARSONS & ROBINSON 2007), although there are few quantitative examples in the literature (ZIMMERMAN et al. 2009).
Life history variation
Yellowknife River cisco exhibit a combination of life history characteristics and morphological attributes that differ from Yellowknife Bay cisco. On the basis of our preliminary evaluation, the Yellowknife River spawning-run consisted of specimens 3–6 years of age, which suggests an early-maturing, short-lived phenotypic variant. Aside from the obvious difference in their migrating into rivers or streams apparently for spawning, these adfl uvial forms differed from larger-bodied lacustrine forms (Morphs B & E) in gross morphology, growth, longevity, and possibly age of maturation. TURGEON & BERNATCHEZ (2003) mentioned anadromy as a north-ern phenomenon, but did not cite any literature on the topic. Moreover, it is uncertain if these authors were referring to anadromy (i.e., migrations from salt to fresh water) or potadromy (i.e., migrations entirely within freshwater). We did not capture any obvious adfl uvial forms (i.e., Morph B) in the greater Yellowknife Bay. This begs the question, what habitat does this morph occupy during non-spawning times of the year? By contrast, Yellowknife Bay cisco appear to be long lived and likely mature at an older age. Mean length-at-age data for the lacustrine morphs suggest that maximum size is achieved within 10–15 years followed by a pronounced reduction in somatic growth (Fig. 4). This life history strategy of possible delayed maturation with rapid initial growth may be a response to predation from burbot (Lota lota) and lake trout (Salvelinus namaycush) in Yellowknife Bay (HJORT 1914).

Cisco diversity in Yellowknife Bay, Great Slave Lake 317
Phenotypic form and function relationships and trophic specialization
Although gut contents of cisco in this study were not analyzed, hypotheses about the trophic ecology of the morphs can be generated from classical form-function relationships (WEBB 1984)—these hypotheses will be tested in a follow-on study. Correlations among trophic-related phenotypic traits suggest that cisco morphs in Great Slave Lake are specialized for different foraging strategies and for exploiting specifi c trophic resources. For example, Morph A has a fusiform body shape, intermediate gill raker number, and a terminal mouth. These traits are typically associated with specialized foraging on relatively large prey items within the water column.
The greatest phenotypic differentiation among Great Slave Lake ciscoes was between Morphs C and D, where mouth orientation, eye size, caudal peduncle depth, and lateral posi-tion of paired fi ns differed. The nature of differences in these characters, suggest that Morph C may be adapted to benthic foraging via prolonged fast swimming, while morph D may be more adapted to utilizing a burst-and-glide foraging strategy within the pelagic zone. Morph D had proportionally large eyes; therefore, it may feed during low light, either at night, or in deep water. The anterior positioning of paired fi ns further supports the hypothesis of a burst-and-glide foraging strategy.
Despite the correlations among phenotypic traits and the hypothesized links to trophic specializations, there is also evidence that morphs may be partitioning particular trophic niches. While Morphs C and D exhibited very different phenotypic traits, their gill raker numbers overlapped. One hypothesis for the existence of two competing forms is that it could refl ect sharing common trophic resources but spatial partitioning of physical habitat. One hypothesis for the origin of multiple cisco morphs involves adaptive radiation into deepwater habitats to prey on vertically migrating opossum shrimp, Mysis relicta (ESHENRODER et al. 1999). It is possible that Morphs C and D both exploit Mysis, but in different habitats. For instance, Morph C may prey on Mysis during the day when they are adjacent to the lake-bottom sediments, while morph D may target them during the night after they have ascended into the water column. Investigating food web structure, cisco stomach contents, prey selec-tion, and stable isotope and fatty acid signatures of predators and prey may provide insights into how trophic niches are being partitioned among the cisco morphs in Great Slave Lake.
Taxonomy: phenotypic parallelism with the Laurentian Great Lakes
In several respects, the cisco morphs identifi ed in Yellowknife Bay are similar to those described from the Laurentian Great Lakes. Body shape, gill raker number, and mouth ori-entation suggest that Morphs A (adfl uvial), B (small lacustrine), and E (large lacustrine) are most similar to C. artedi, while Morph C (shortjaw) appears to be most closely affi liated with C. zenithicus. Morph D (least) is an exception in that it does not have Laurentian Great Lakes analog and likely colonized the lake from a northwestern source. Morph D appears to share affi nities with both C. nigripinnis and C. sardinella.
The small cisco from the Yellowknife River (Morph A) is also unusual in that it pos-sesses characteristics of both C. artedi and C. sardinella. In some gross morphological fea-tures, such as overall body shape, head length, and eye diameter, Morph A overlapped in

318 P. Vecsei et al.
characters with C. sardinella. However, snout length and pelvic fi n position in relation to body and meristic characters, such as dorsal fi n ray number, differed considerably from C. sardinella. The life history trait of spawning in fast-fl owing shallow water has been docu-mented for some northern populations of C. artedi (TURGEON & BERNATCHEZ 2003). It is pos-sible that the Yellowknife River cisco is a small-bodied morph of C. artedi (i.e., a ‘dwarf’) with an adfl uvial life history component. However, lower numbers of gill rakers and general gross morphological differences such as larger eyes set along the medial body axis, short-er pectoral fi ns, and a less elliptical body shape distinguish it from the lacustrine morphs (morphs B & E).
The small lacustrine Morph B was most similar to C. artedi in terms of morphometric characters and assumed habitat. Both appear to inhabit mid to deep-water lacustrine habitats. Gillraker number for Morph B was within the lower range typically observed for C. artedi (KOELZ 1929, MCPHAIL & LINDSEY 1970, MCPHAIL 2007). Morph B had relatively long rakers, which is also characteristic of C. artedi. Morph B also exhibited a moderate eye diameter (21–26% of head length for C. artedi), snout length (i.e., premaxilla to origin of eye) that was in excess of the eye diameter, and a terminal mouth, which are all characteristic of C. artedi. In general, all meristic counts for Morph B fell within the range reported for C. artedi (SCOTT & CROSSMAN 1973).
Morph C was most similar to C. zenithicus described from the Laurentian Great Lakes. Gill raker number was within the range documented for C. zenithicus, albeit on the higher end of the spectrum (SCOTT & CROSSMAN 1973, STEINHILBER 2002, MURRAY & REIST 2003). While Morph C did not exhibit life-history variants as observed in other forms, a range of interesting morphologies were observed, suggesting that differentiation may be occurring within this group (Fig. 3c[ii]). One example was a long-snouted variant which lacked the pronounced down-curved premaxilla associated with C. zenithicus from the Great Lakes. The long-snouted variant had a fl eshy, rounded, protruding snout more typical of long snouted forms of C. williamsoni and may represent a unique form of local endemism rather than a more widespread morphotype.
Morph D appears to be unique among the ciscoes, but shares some traits in common with C. nigrippinnis. For example, both forms exhibit dark pigmentation on the fi ns and posterior portion of dentary and maxilla, a long mandible extending beyond the premaxilla, a rela-tively large eye, and overlap in some meristic characters. However, Morph D differs from C. nigripinnis in that it has a pointed, upturned snout with a superior mouth. This mouth arrangement differs markedly from C. nigripinnnis, where the mouth opening forms a 45° to 60° angle with the horizontal axis of the head (SCOTT 1960). Morph D also had a less ellipti-cal body shape, large paired fi ns comparable to C. kiyi, and a more ‘pointed’ or ‘sail-like’ dorsal fi n. Morph D also differed from C. nigripinnis in that gill raker number was far lower and outside the range of what is known for that species (T. Todd, United States Geological Survey, personal communication). However, gill raker number and length for Morph D were in the middle of the range observed and overlapped with all other sympatric cisco morphs. Morph D also shares several morphological characteristics with C. sardinella, a member of the Arctic cisco assemblage. Indeed, multiple specimens of this morph exist in the Royal Ontario Museum (e.g., ROM catalogue # 17107; collector Kennedy 1946) and were subse-quently identifi ed as C. sardinella by ROM ichthyologists. Further evidence to support the C. sardinella designation comes from mtDNA sequences (TURGEON & BERNATCHEZ 2003).

Cisco diversity in Yellowknife Bay, Great Slave Lake 319
If Morph E is C. sardinella, then it is of interest because it may represent a southern, land-locked lacustrine population that differs greatly in morphology and habitat from its Alaskan counterpart.
Future considerations
There were two potential caveats that may have biased the results of our study. The fi rst is that because this was a preliminary investigation, small sample sizes limited the power with which we could resolve taxonomic associations. To increase the understanding of cisco diversity in Great Slave Lake, we initiated a collaborative research program during 2008. The second issue was that we did not conduct a quantitative assessment of maturity or diet. An understanding of how body shape varies throughout ontogeny and how ontogenetic shifts in diet relate to those changes will provide a better understanding of the processes driving phenotypic divergence within the cisco complex. Another issue that we identifi ed was that although colour is typically used as a key distinguishing feature between C. zenithicus and other cisco, we found that body colouration was of limited value for distinguishing among cisco morphs in Great Slave Lake. When live specimens were caught in good condition, digital images were captured immediately to more accurately document body colouration. However, photography is an imperfect medium because colour rendition and ‘temperature’ are affected by environmental conditions. In the case of iridescent fi shes, such as cisco, per-ceived colour is also affected by the clothing of the individual handling the fi sh. The colours that we described for each morphotype may appear in varying combinations or hues, which are often an artefact of the variable atmospheric conditions during observation rather than a wide range of colour gamut on a particular individual. To obtain enough accuracy to use the internationally accepted Munsell Colour Code System was deemed impossible as it goes into descriptive levels such as hue, value, and chroma that are simply impossible to discern with the naked eye.
Conclusions
Discerning the patterns of phenotypic differentiation is a critical fi rst step in understanding the processes driving species divergence and ultimately evolution. The cisco complex in Great Slave Lake is composed of several morphs and the pattern of phenotypic differentiation may parallel that seen in other large systems such as the Laurentian Great Lakes. We found several levels of variation within the diverse Yellowknife Bay cisco taxonomic assemblage. Morph A (adfl uvial) may represent a life history variant of C. artedi. Morph B (small lacus-trine) may be a biological variant of C. artedi differing in size, gill raker number, and body proportions from the sympatric large lacustrine form (Morph E). Alternatively, Morph B may be an earlier life stage of Morph E with gill raker number being a function of ontogenetic changes in size, habitat, or food habits. Morph C (shortjaw) may represent a phenotypic vari-ant of C. artedi, with local morphological adaptations to better exploit resources less utilized by the common C. artedi. Alternatively, Morph C may indeed be a closely related form of C. zenithicus. Morph D likely represents a form or morph of C. sardinella. Morph E (large

320 P. Vecsei et al.
lacustrine) like Morph B, may represent a biological variant of C. artedi exhibiting an unusu-ally large adult size.
At this point, it is unknown how cisco are partitioning their resources (i.e., habitat versus trophic) and how that resource partitioning is driving diversifi cation; however, these studies are currently underway. The relative infl uences of allopatric versus sympatric divergence will need to be addressed as both have likely played a role in shaping contemporary population structure (TURGEON & BERNATCHEZ 2003). Rigorous comparisons between Great Slave Lake and other systems will be necessary to address the question of parallel evolution. Moreover, estimates of genetic differentiation will aid in the assignment of conservation units to respec-tive morphs. The high degree of phenotypic diversity in Great Slave Lake appears to adhere to the common paradigm of ecological speciation and this complex system will undoubtedly offer new insights into how ecological factors drive evolutionary divergence.
Acknowledgements
We thank D. Bourassa, K. Bourassa, and P. Smith for their integral assistance with fi sh sampling and processing and R. Bourke and C. McLeod for comments on a previous version of the manuscript. This research was originally presented at the 2008 Coregonid Symposium in Winnipeg Manitoba. Funding was provided by Indian and Northern Affairs Canada (Contract 50A 00-06-6014-2 to IMG Golder) and by Fisheries and Oceans Canada through Public Works Canada (Contract EW699-090962/001/PWZ 99 to Golder Associates Ltd).
Literature cited
BERNATCHEZ, L. (2004): Ecological theory of adaptive radiation: an empirical assessment from Coregonine fi shes (Salmoniformes). – In: HENDRY, A.P. & STEARNS, S.C. (eds.): Evolution illuminated: salmonids and their relatives. Oxford Univ. Press, New York. pp. 175–207.
BERNATCHEZ, L. & WILSON, C.C. (1998): Comparative phylogeography of Nearctic and Palearctic fi shes. – Mol. Ecol. 7:431–452.
BOOKSTEIN, F.L. (1989): Principal warps, thin plate splines and the decomposition of deformations. – Ieee Transactions on Pattern Analysis and Machine Intelligence. 11(6): 567–585.
COAD, B.W. & MCALLISTER, D.E. (2009): Dictionary of Ichthyology. http://www.briancoad.com/Dictionary/M.htm. Accessed 03 March 2009.
CLARKE, R.M. (1973): The systematics of cisco (Coregonidae) in central Canada. Doctoral dissertation, University of Manitoba, Winnipeg.
DYMOND, J.R. (1943): The coregonine fi shes of northwestern Canada. – Transactions of the Royal Canadian Institute. 24: 171–232.
ESHENRODER, R.L., SIDELEVA, V.G. & TODD, T.N. (1999): Functional convergence among pelagic sculpins of Lake Baikal and deepwater cisco of the Great Lakes. – J. Great Lakes Res. 25: 847–855.
EVANS, M.S. (2000): The large lake ecosystems of northern Canada. – Aquatic Ecosyst. Health Management. 3: 65–79.
KOELZ, W.N. (1929): Coregonid fi shes of the Great Lakes. – Bulletin U.S. Bureau of Fisheries. 43: 279–643.
HARPER, G. & NICHOLS, J.T. (1919): Six new fi shes from northwestern Canada. – Bull. Am. Mus. Nat. Hist. XLI: 263–271.
HJORT, J. (1914): Fluctuations in the great fi sheries of northern Europe viewed in light of biological research. – Rapports et Procès-Verbaux des Reunions Conseil Permanent International pour L’Exploration de la Mer. 20: 1–228.

Cisco diversity in Yellowknife Bay, Great Slave Lake 321
HOWLAND, K.L., GENDRON, M., TONN, W.M. & TALLMAN, R.F. (2004): Age determination of a long lived coregonid from the Canadian North: comparison of otoliths, fi n rays and scales in inconnu (Stenodus leucichthys). – Ann. Zool. Fenn. 41: 205–214.
MCPHAIL, J.D. (2007): The Freshwater Fishes of British Columbia. The University of Alberta Press. 620 pp.
MCPHAIL, J.D. & Lindsey, C.C. (1970): Freshwater Fishes of Northwestern Canada and Alaska. Fisheries Research Board of Canada Bulletin 173, Ottawa, Ontario.
MURRAY, L. (2006): A morphological examination of sympatric cisco forms in four lakes with specifi c reference to the occurrence of shortjaw cisco (coregonus zenithicus) in Manitoba. Master’s thesis, Zoology, University of Manitoba, Winnipeg.
MURRAY, L. & REIST, J.D. (2003): Status report on the shortjaw cisco Coregonus zenithicus in central and western Canada. – Can. Manuscr. Rep. Fish Aquat. Sci. 2638: vii + 56 pp.
PARSONS, K.J. & ROBINSON, B.W. (2007): Foraging performance of diet-induced morphotypes in pumpkinseed sunfi sh (Lepomis gibbosus) favours resource polymorphism. – European Society for Evolutionary Biology. 20: 673–684.
RAWSON, D.S. (1950): The physical limnology of Great Slave Lake. – J. Fish. Res. Board Can. 8: 1–166.RAWSON, D.S. (1951): Studies of the fi sh of Great Slave Lake. – J. Fish. Res. Board Can. 8: 207–240.REIST, J.D. (1985): An empirical evaluation of several univariate methods that adjust for size variation
in morphometric data. – Can. J. Zool. 63: 1429–1439.ROBERGE, M.M., LOW, G. & READ, C.J. (1985): Data from an experimental gillnetting program on
Great Slave Lake, Northwest Territories, 1980–81. Department of Fisheries and Oceans, Winnipeg, Manitoba.
ROBINSON, B.W. & PARSONS, K.J. (2002): Changing times, spaces and faces: tests and implications of adaptive morphological plasticity in the fi shes of northern postglacial lakes. – Can. J. Fish. Aquat. Sci. 59: 1819–1833.
ROBINSON, B.W. & SCHLUTER, D. (2000): Natural selection and the evolution of adaptive genetic variation in northern freshwater fi shes. – In: MOUSSEAU, T.A., SINERVO, B. & ENDLER, J.A. (Eds.) Adaptive genetic variation in the wild. Oxford Univ. Press, NY. pp. 65–94.
ROGERS S.M. & BERNATCHEZ, L. (2005): Integrating QTL mapping and genomic scans towards the characterization of candidate loci under parallel directional selection in these lake whitefi sh (Coregonus clupeaformis). – Mol. Ecol. 14: 351–361.
ROHLF, F.J. & BOOKSTEIN, F.L. (2003): Computing the uniform component of shape variation. – Syst. Biol. 52(1): 66–69.
SCOTT, B.W. (1960): Summaries of current information on shortjaw cisco, shortnose cisco, and blackfi n cisco. Research Branch Ontario Department of Lands and Forests. Research Information Paper (Fisheries) No. 9, Maple, Ontario.
SCOTT, B.W. & CROSSMAN, E.J. (1973): Freshwater Fishes of Canada. Bulletin 184. 966 pp.SKULASON, S. & SMITH, T.B. (1994): Resource polymorphisms in vertebrates. – Trends Ecol. Evol.
10:366–370.STEINHILBER, M., NELSON, J.S. & REIST, J.D. (2002): A morphological and genetic re-examination
of sympatric shortjaw cisco (Coregonus zenithicus) and lake herring (C. artedi) in Barrow Lake, Alberta, Canada. – Archives fur Hydrobiologie. 57: 463–478.
TAYLOR. E.B. (1999): Species pairs of north temperate freshwater fi shes: taxonomy evolution, and conservation. – Reviews in Fish Biology and Fisheries. 9:299–324.
TODD, T.N., SMITH, G.R. & CABLE, L. (1981): Environmental and genetic contributions to morphological differentiation in cisco (Coregoninae) of the Great Lakes. – Can. J. Fish. Aquat. Sci. 38 (1): 59–67.
TODD T.N. & SMITH, G.R. (1980): Differentiation in Coregonus zenithicus in Lake Superior. – Can. J. Fish. Aquat. Sci. 37: 2228–2235.
TODD, T.N. & STEINHILBER, M.A. (2002): Diversity in shortjaw cisco (Coregonus zenithicus) in North America. – In: TODD, T.N. & FLEISCHER, G. (Eds.) Biology and Management of Coregonid Fishes, 1999. – Spec. Issues Advanc. Limnol. Archiv für Hydrobiologie. 57: 517–525.
TURGEON, J. & BERNATCHEZ, L. (2003): Reticulate evolution and phenotypic diversity in North American cisco, Coregonus spp. (Teleostei: Salmonidae): implications for the conservation of an evolutionary legacy. – Conserv. Genet. 4:67–81.

322 P. Vecsei et al.
TURGEON, J., ESTOUP, A. & BERNATCHEZ, L. (1999): Species fl ock in the North American Great Lakes: molecular ecology of Lake Nipigon cisco (Teleostei: Coregonidae: Coregonus). – Evolution. 53: 1857–1871.
WEBB, P. (1984): Body form, locomotion and foraging in aquatic vertebrates. – Am. Zool. 24: 107–120.ZELDITCH, M.L., SWIDERSKI, D.L., SHEETS, D.H. & FINK, W.L. (2004): Geometric morphometrics for
biologists: a primer. Elsevier Academic Press, New York.ZIMMERMAN, M.S, STEPHANIE N., SCHMIDT S.N., KRUEGER C.C., VANDER ZANDEN M.J. & ESHENRODER R.L.
(2009): Ontogenetic niche shifts and resource partitioning of lake trout morphs. – Can. J. Fish. Aquat. Sci. 66: 1007–1018.




















