
Interaction of the monoxenic trypanosomatid Blastocrithidia culicis with the Aedes aegypti salivary...
10
Acta Tropica 113 (2010) 269–278 Contents lists available at ScienceDirect Acta Tropica journal homepage: www.elsevier.com/locate/actatropica Interaction of the monoxenic trypanosomatid Blastocrithidia culicis with the Aedes aegypti salivary gland Michelle Tanny C. Nascimento a , Maria Carolina F. Garcia a , Karen Pereira da Silva a , Lucia Helena Pinto-da-Silva b , Georgia C. Atella c , Maria Cristina M. Motta d,∗∗ , Elvira M. Saraiva a,∗ a Laboratório de Imunobiologia das Leishmanioses, Departamento de Imunologia, Instituto de Microbiologia Prof. Paulo de Góes, Universidade Federal do Rio de Janeiro, RJ, Brazil b Dept. Microbiologia e Imunologia Veterinária, Instituto de Veterinária, Universidade, Federal Rural do Rio de Janeiro, RJ, Brazil c Instituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, RJ, Brazil d Laboratório de Ultraestrutura Celular Hertha Meyer – Instituto de Biofísica Carlos Chagas, Filho, Universidade Federal do Rio de Janeiro, RJ, Brazil article info Article history: Received 21 July 2009 Received in revised form 18 November 2009 Accepted 27 November 2009 Available online 3 December 2009 Keywords: Blastocrithidia Aedes aegypti Mosquito salivary gland carbohydrates Mosquito salivary gland lectin Protozoa–insect interaction Monoxenic trypanosomatid abstract Blastocrithidia culicis is a monoxenous trypanosomatid that inhabits mosquitoes. Although its life cycle in these insects has not been described, we recently demonstrated that B. culicis colonizes the Aedes aegypti digestive tract and is also able to reach the mosquito’s haemocoel. In this work, we describe the interaction of B. culicis with the A. aegypti salivary gland in vitro and in vivo. Ultrastructural analysis reveals different steps of the invasion process, beginning with the insertion of the B. culicis anterior flagellum into the basal lamina of the gland and ending with the protozoan inside the salivary gland acini compartment. Carbohydrates are involved in the initial adhesion of B. culicis cells to the salivary glands, as demonstrated by protozoan binding inhibition assays and fluorescent lectin labeling of the trypanosomatid–salivary gland interaction. B. culicis is able to survive after incubation in vitro in the mosquito haemolymph, and trypanosomatid binding to salivary glands was confirmed by the injection of radioactively labeled protozoa into the mosquito haemocoel. These results suggest that salivary gland invasion could be part of the B. culicis life cycle in A. aegypti, raising the possibility that B. culicis can be transmitted by these mosquitoes. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Trypanosomatid protozoa develop within different insect organs, including the midgut, hindgut, Malphigian tubules, sali- vary glands and haemocoel (Wallace, 1966). Although in some cases these trypanosomatids reach a secondary host, such as vertebrates via blood sucking insects or plants via phytophagous bugs, part of their life cycle takes place in an insect. There are few studies of the complex interactions between trypanosomatids and insects in the literature, especially in the case of monoxenic trypanosomatids, whose life cycle occurs only in insects (Podlipaev, 2000). The endosymbiont-bearing monoxenic trypanosomatid Blast- ocrithidia culicis and the Dipteran mosquito Aedes aegypti constitute an attractive model system for studying protozoan–insect interac- tions. A. aegypti is a globally distributed vector of the dengue and yellow fever viruses, and one of the most studied mosquito species due to its importance as a vector and amenability to laboratory ∗ Corresponding author. Tel.: +55 21 2562 6747; fax: +55 21 2560 8344. ∗∗ Corresponding author. Tel.: +55 21 2562 6580; fax: +55 21 2260 2364. E-mail addresses: [email protected] (M.C.M. Motta), [email protected] (E.M. Saraiva). rearing. B. culicis is a promiscuous trypanosomatid isolated from mosquitoes such as Aedes vexans, Culex pipens, Mansonia richardii and Anopheles maculipennis, though its life cycle in invertebrates remains unknown (Wallace, 1966). Although considered nonpathogenic for mammals, monoxenic trypanosomatids have been reported to infect different vertebrate hosts. For example, McGhee (1957) and Jansen et al. (1988) demon- strated that these trypanosomatids can infect chicken embryos and opossum stink glands. Importantly, the monoxenic trypanosomatid Leptomonas pulexsimulantis was identified as the causative agent of a syndrome similar to visceral leishmaniasis, while a protozoan closely related to Herpetomonas samuelpessoai, also a monoxenic trypanosomatid, was found in the blood of an HIV-positive patient (Pacheco et al., 1998; Morio et al., 2008). Moreover, we demon- strated that B. culicis could survive in human macrophages whose microbicidal activity had been deactivated by HIV-1 infection (Barreto-de-Souza et al., 2008). Using the A. aegypti–B. culicis interaction model, we demon- strated that this trypanosomatid is capable of colonizing the mosquito’s digestive tract and crossing the intestinal epithelia, reaching the haemocoel (Fampa et al., 2003; Corrêa-da-Silva et al., 2006). This finding prompted the present study, in which we asked whether B. culicis could survive in the mosquito haemolymph. We 0001-706X/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.actatropica.2009.11.010
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Interaction of the monoxenic trypanosomatid Blastocrithidia culicis with the Aedes aegypti salivary...

IA
MLa
b
c
d
a
ARR1AA
KBAMMPM
1
ovtvtttw
oatyd
(
0d
Acta Tropica 113 (2010) 269–278
Contents lists available at ScienceDirect
Acta Tropica
journa l homepage: www.e lsev ier .com/ locate /ac ta t ropica
nteraction of the monoxenic trypanosomatid Blastocrithidia culicis with theedes aegypti salivary gland
ichelle Tanny C. Nascimentoa, Maria Carolina F. Garciaa, Karen Pereira da Silvaa,ucia Helena Pinto-da-Silvab, Georgia C. Atellac, Maria Cristina M. Mottad,∗∗, Elvira M. Saraivaa,∗
Laboratório de Imunobiologia das Leishmanioses, Departamento de Imunologia, Instituto de Microbiologia Prof. Paulo de Góes, Universidade Federal do Rio de Janeiro, RJ, BrazilDept. Microbiologia e Imunologia Veterinária, Instituto de Veterinária, Universidade, Federal Rural do Rio de Janeiro, RJ, BrazilInstituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, RJ, BrazilLaboratório de Ultraestrutura Celular Hertha Meyer – Instituto de Biofísica Carlos Chagas, Filho, Universidade Federal do Rio de Janeiro, RJ, Brazil
r t i c l e i n f o
rticle history:eceived 21 July 2009eceived in revised form8 November 2009ccepted 27 November 2009vailable online 3 December 2009
a b s t r a c t
Blastocrithidia culicis is a monoxenous trypanosomatid that inhabits mosquitoes. Although its life cyclein these insects has not been described, we recently demonstrated that B. culicis colonizes the Aedesaegypti digestive tract and is also able to reach the mosquito’s haemocoel. In this work, we describethe interaction of B. culicis with the A. aegypti salivary gland in vitro and in vivo. Ultrastructural analysisreveals different steps of the invasion process, beginning with the insertion of the B. culicis anteriorflagellum into the basal lamina of the gland and ending with the protozoan inside the salivary gland
eywords:lastocrithidiaedes aegyptiosquito salivary gland carbohydratesosquito salivary gland lectin
rotozoa–insect interaction
acini compartment. Carbohydrates are involved in the initial adhesion of B. culicis cells to the salivaryglands, as demonstrated by protozoan binding inhibition assays and fluorescent lectin labeling of thetrypanosomatid–salivary gland interaction. B. culicis is able to survive after incubation in vitro in themosquito haemolymph, and trypanosomatid binding to salivary glands was confirmed by the injectionof radioactively labeled protozoa into the mosquito haemocoel. These results suggest that salivary glandinvasion could be part of the B. culicis life cycle in A. aegypti, raising the possibility that B. culicis can be
quito
onoxenic trypanosomatid transmitted by these mos. Introduction
Trypanosomatid protozoa develop within different insectrgans, including the midgut, hindgut, Malphigian tubules, sali-ary glands and haemocoel (Wallace, 1966). Although in some caseshese trypanosomatids reach a secondary host, such as vertebratesia blood sucking insects or plants via phytophagous bugs, part ofheir life cycle takes place in an insect. There are few studies ofhe complex interactions between trypanosomatids and insects inhe literature, especially in the case of monoxenic trypanosomatids,hose life cycle occurs only in insects (Podlipaev, 2000).
The endosymbiont-bearing monoxenic trypanosomatid Blast-crithidia culicis and the Dipteran mosquito Aedes aegypti constitute
n attractive model system for studying protozoan–insect interac-ions. A. aegypti is a globally distributed vector of the dengue andellow fever viruses, and one of the most studied mosquito speciesue to its importance as a vector and amenability to laboratory∗ Corresponding author. Tel.: +55 21 2562 6747; fax: +55 21 2560 8344.∗∗ Corresponding author. Tel.: +55 21 2562 6580; fax: +55 21 2260 2364.
E-mail addresses: [email protected] (M.C.M. Motta), [email protected]. Saraiva).
001-706X/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.actatropica.2009.11.010
es.© 2009 Elsevier B.V. All rights reserved.
rearing. B. culicis is a promiscuous trypanosomatid isolated frommosquitoes such as Aedes vexans, Culex pipens, Mansonia richardiiand Anopheles maculipennis, though its life cycle in invertebratesremains unknown (Wallace, 1966).
Although considered nonpathogenic for mammals, monoxenictrypanosomatids have been reported to infect different vertebratehosts. For example, McGhee (1957) and Jansen et al. (1988) demon-strated that these trypanosomatids can infect chicken embryos andopossum stink glands. Importantly, the monoxenic trypanosomatidLeptomonas pulexsimulantis was identified as the causative agentof a syndrome similar to visceral leishmaniasis, while a protozoanclosely related to Herpetomonas samuelpessoai, also a monoxenictrypanosomatid, was found in the blood of an HIV-positive patient(Pacheco et al., 1998; Morio et al., 2008). Moreover, we demon-strated that B. culicis could survive in human macrophages whosemicrobicidal activity had been deactivated by HIV-1 infection(Barreto-de-Souza et al., 2008).
Using the A. aegypti–B. culicis interaction model, we demon-
strated that this trypanosomatid is capable of colonizing themosquito’s digestive tract and crossing the intestinal epithelia,reaching the haemocoel (Fampa et al., 2003; Corrêa-da-Silva et al.,2006). This finding prompted the present study, in which we askedwhether B. culicis could survive in the mosquito haemolymph. We
2 cta Tr
awpwoictonbbrBah
2
2
f(gw7
2
rDmsweai
2
MwpCOtnb1w((
2
fNgAtts
70 M.T.C. Nascimento et al. / A
lso wanted to determine whether B. culicis is capable of interactingith the A. aegypti salivary glands, likewise Plasmodium sp. and Try-
anosoma rangeli (Pimenta et al., 1994; Schaub, 1994). Additionally,e examined the role of carbohydrates and ultrastructural aspects
f the adhesion of B. culicis to salivary glands, and investigated thisnteraction in vivo by injecting B. culicis into the mosquito haemo-oel. Our results demonstrate that B. culicis is not only able to adhereo A. aegypti salivary glands, but is also capable of invading thisrgan, reaching the cytoplasm of the epithelial cells and the aci-ar space, where saliva is stored. This protozoan–salivary glandinding process is mediated by carbohydrates, as demonstratedy inhibition assays using different carbohydrates and polysaccha-ides, as well as by fluorescent lectin binding analyses. In addition,. culicis is able to survive in the mosquito haemolymph anddhere to the salivary glands after inoculation into the mosquitoaemocoel.
. Materials and methods
.1. Protozoan culture
The endosymbiont-harboring B. culicis (ATCC 30257), isolatedrom Aedes vexans, was maintained at 26 ◦C in Warren’s mediumWarren, 1960) supplemented with 10% fetal bovine serum. Afterrowth for 24 h (logarithmic phase) or 72 h (stationary phase), cellsere washed 3 times in 0.01 M phosphate buffered saline (PBS), pH
.2 and used for all assays.
.2. Mosquito
A. aegypti (Rockfeller strain) was obtained from the Labo-atório de Fisiologia e Controle de Vetores (LAFICAVE–Entomologyept.–Fundacão Oswaldo Cruz, Rio de Janeiro, Brazil). Adult femaleosquitoes, 5–7 days post-emergence, were maintained on 30%
ugar solution ad libitum until use. For sterile assays, mosquitoesere washed for 2 min in hypochlorite solution (3%), followed by
xtensive washings in PBS supplemented with penicillin (50 U/mL)nd streptomycin (50 �g/mL). Salivary glands were then dissectednside the laminar flow.
.3. Protozoan–insect organ interaction assay
Groups of 10–15 midguts, salivary glands, ovaries, crops andalpighian tubules from female mosquitoes were incubated for 1 hith 1 × 106 B. culicis in Hank’s balanced salt solution (Sigma) sup-lemented with 1% bovine serum albumin (BSA, Sigma), 0.15 mMaCl2 and 1 mM MgCl2 (complete Hank’s) at room temperature.rgans were gently washed in PBS after binding assays, individually
ransferred to microtubes containing PBS, homogenized and theumber of attached B. culicis was determined in a Neubauer cham-er. Longer B. culicis–salivary gland binding assays were done forh in sterile conditions as described above. The organs were thenashed and re-incubated for 4 or 18 h in Schneider insect medium
Sigma) supplemented with penicillin (50 U/mL) and streptomycin50 �g/mL) at 26 ◦C.
.4. Carbohydrate inhibition assays
Salivary glands were dissected and immediately incubatedor 30 min in the presence of 100 mM �-methyl-d-mannoside,-acetyl-d-glucosamine (GlucNAc), l-fucose, 200 mM N-acetyl-d-
alactoside (GalNAc), 100 �g of mannan or succinyl Concanavalin(S-Con A) in complete Hank’s. B. culicis (1 × 106) was added tohese salivary gland preparations and incubated for 1 h at roomemperature. Pre-treatment of B. culicis with mannan (100 �g) oralivary glands with S-Con A (100 �g) was performed for 30 min,
opica 113 (2010) 269–278
followed by washings in PBS and then processing as describedabove.
2.5. Fluorescence microscopy
Salivary gland–B. culicis interactions, salivary glands or B. culi-cis alone were fixed in 4% paraformaldehyde diluted in PBS. Afterwashing, samples were blocked with 1% PBS–BSA and incubatedfor 1 h in the presence of 100 �g/mL of one of the followingFITC-conjugated lectins: Concanavalin A (Con A), wheat germagglutinin (WGA), peanut agglutinin (PNA) or soybean agglutinin(SBA) (Sigma). Topro-3 (1 �M, Molecular Probes, USA) or DAPI(10 �g/ml, Sigma) were used for nuclear and kinetoplast staining,and Evans blue (0.05%) was used to quench nonspecific fluores-cence. After labeling, the samples were washed with PBS andmounted with Vectashield (Vector Lab Inc, USA). Fluorescenceimages were obtained using a Zeiss Axioplan and Confocal imageswere taken in a Zeiss LSM 510 META.
2.6. Transmission electron microscopy (TEM)
B. culicis–salivary gland preparations, done as described above,were washed and fixed in Karnovsky solution containing 2.5% glu-taraldehyde and 4% paraformaldehyde plus 4% sucrose and 5 mMCaCl2, diluted in 0.1 M cacodylate buffer, pH 7.2. All samples werepost-fixed in 1% osmium tetroxide containing 0.8% potassium fer-ricyanide diluted in the same buffer. After washing, cells weredehydrated in graded series of acetone solutions and embeddedin Epoxy resin. Ultrathin sections were stained with uranyl acetateand lead citrate and observed in a Zeiss 900 transmission electronmicroscope.
2.7. Scanning electron microscopy (SEM)
B. culicis–salivary gland preparations done as described abovewere fixed and post-fixed as for TEM. Dehydration was done ingraded series of ethanol solutions, and then samples were dried bythe critical point method, coated with gold and observed in a JeolJSM-5310 Scanning Electron Microscope.
2.8. Flow cytometry
B. culicis was fixed in 4% paraformaldehyde diluted in PBS.After washing, cells were blocked with 1% PBS–BSA, incubated for1 h with FITC-ConA (10 �g/mL) and analyzed in a fluorescence-activated cell sorter (FACSCalibur, BD Bioscience, USA).
2.9. Haemolymph survival assays
Mosquitoes were anesthetized at −20 ◦C and maintained at 4 ◦Cduring injection of 2–3 �l of Aedes saline (Hagedorn et al., 1977)into the haemocoel using a PLI-100 microinjector (Harvard Appa-ratus). Subsequently, haemolymph was collected by capillary aftergentle pressing of the mosquito body and dispensed in Terasakimicroplates. Pooled haemolymph from 12 to 15 mosquitoes dilutedwith the same volume of Aedes saline was incubated with 106 B.culicis for 1 h at room temperature. Viable, motile protozoa werecounted in a Neubauer chamber.
2.10. In vivo binding assay
B. culicis cells were labeled with 125Iodine (300 �Ci) in RPMImedium for 1 h and washed three times in PBS. Then 1 �L contain-ing 1 × 105 radiolabeled protozoa was injected into the haemocoelof adult female mosquitoes using a PLI-100 microinjector. One daylater, the salivary glands, midguts, crops, ovaries and Malpighian

M.T.C. Nascimento et al. / Acta Tropica 113 (2010) 269–278 271
Fig. 1. Blastocrithidia culicis–A. aegypti organ interactions. Protozoa in the logarith-mic growth phase were incubated for 1 h in complete Hank’s with the indicatedoBea
twWb
2
pti
3
3
ec2gcnmws
3i
tmseg(estatiba
Fig. 2. Scanning electron microscopy of Blastocrithidia culicis–salivary gland interac-tions. (A) Whole A. aegypti salivary gland illustrating protozoa (P) attached to lateral(LL) and median (ML) lobules. The inset shows a higher magnification of B. culicis(P) interacting by its flagellum with the medium lobule (bar = 50 �m; inset = 5 �m).(B) Protozoa (P) attached to the salivary gland surface by the flagellum after 1 h of
rgans explanted from A. aegypti females. Results are shown as mean number of. culicis cells attached to the different organs and were not statistically differ-nt between the organs analyzed. Each symbol represents the number of B. culicisttached per organ.
ubules of inoculated mosquitoes were dissected and radioactivityas determined using an automatic gamma counter (Wallac 1470izard). Radioactivity associated to carcasses (the whole mosquito
ody minus the listed organs) was also quantified.
.11. Statistical analysis
Data were analyzed using the GraphPad Prism 4.0 softwareackage (GraphPad Software Inc., San Diego, CA, USA). Unpaired-tests were performed, and P values ≤0.05 were considered signif-cant.
. Results
.1. In vitro interaction of Blastocrithidia with Aedes organs
To study the parasite–mosquito organ interaction in vitro,xplanted organs were incubated in the presence of 1 × 106 B. culi-is for 1 h. Binding assays with different organs showed a mean of.2, 3.1 and 2.3 × 103 protozoa bound to the basal lamina of salivarylands, midguts and ovaries, respectively (Fig. 1). In these assays, B.ulicis was unable to adhere to crops and Malpighian tubules (dataot shown), demonstrating that the adhesion to salivary glands,idguts and ovaries was specific. The binding to salivary glandsas not influenced by the growth phase of the protozoa (data not
hown).
.2. Ultrastructural aspects of the B. culicis–salivary glandnteraction
Ultrastructural aspects of the B. culicis–salivary gland interac-ion were analyzed by scanning (SEM) and transmission electron
icroscopies (TEM). B. culicis were usually seen attached to thealivary gland by the anterior flagellum (Fig. 2A and B). No prefer-ntial adhesion of B. culicis to the different regions of the salivaryland was observed in any of the protozoan–gland pairs analyzedFig. 2A). Protozoa at different entrance stages and partially cov-red by the basal lamina were observed. Lesions in the basal laminaeemed to be formed by the insertion of the flagellum, which allowshe protozoan to enter a space formed between the basal lamina
nd the epithelial cell membrane (Fig. 2B and C). To strengthenhis pattern of interaction, we used TEM to analyze B. culicis bind-ng to the A. aegypti salivary glands. Similarly to what we foundy SEM, B. culicis flagellum interacts with the basal lamina (Fig. 3And B) by a close contact between both structures (Fig. 3C). Afterinteraction (arrow; bar = 1 �m). The asterisk indicates damage of the basal lamina.(C) Protozoa are visible under the basal lamina, which seems to have been dam-aged by these organisms after incubation overnight. Arrowhead points to protozoainvading the gland and asterisks indicate scars in the basal lamina (bar = 5 �m).
4 h of interaction, the trypanosomatid flagellum was observed in
the space between the basal lamina and the plasma membraneof the glandular cells (Fig. 3D and E). As revealed by TEM, proto-zoa could be observed in the cell cytoplasm of all salivary glandlobules after overnight interaction (Fig. 4). In the medium lob-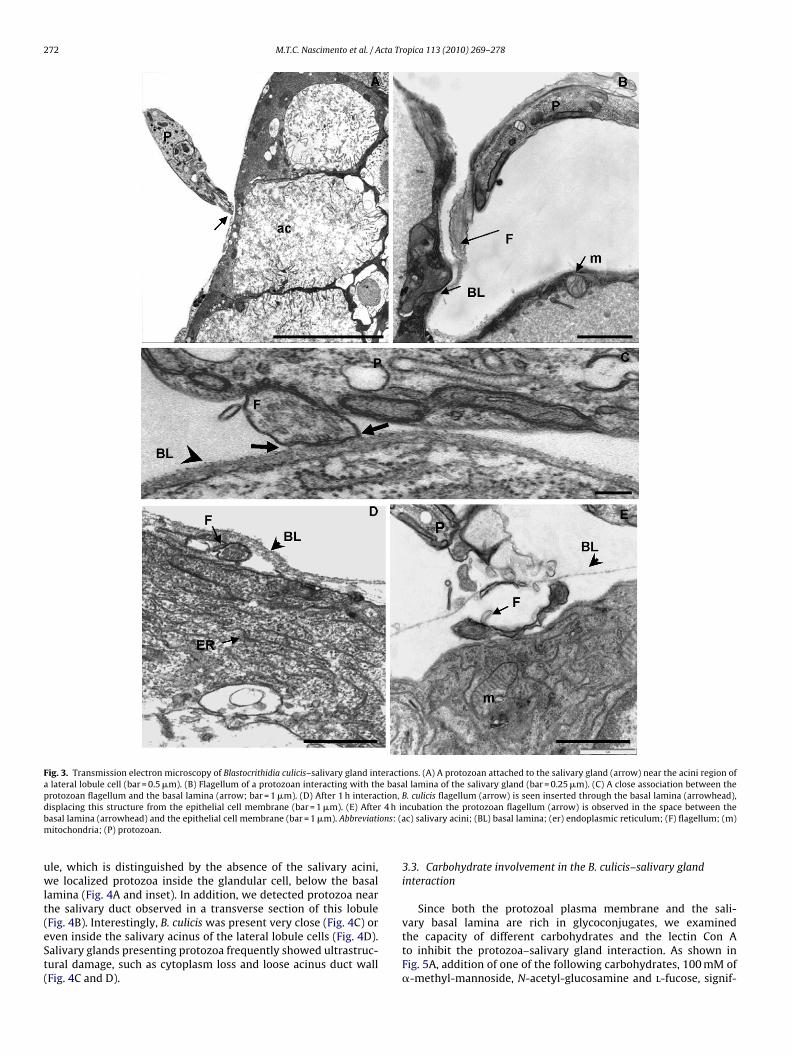
272 M.T.C. Nascimento et al. / Acta Tropica 113 (2010) 269–278
Fig. 3. Transmission electron microscopy of Blastocrithidia culicis–salivary gland interactions. (A) A protozoan attached to the salivary gland (arrow) near the acini region ofa lateral lobule cell (bar = 0.5 �m). (B) Flagellum of a protozoan interacting with the basal lamina of the salivary gland (bar = 0.25 �m). (C) A close association between theprotozoan flagellum and the basal lamina (arrow; bar = 1 �m). (D) After 1 h interaction, B. culicis flagellum (arrow) is seen inserted through the basal lamina (arrowhead),d 4 h ib ons: (am
uwlt(eSt(
isplacing this structure from the epithelial cell membrane (bar = 1 �m). (E) Afterasal lamina (arrowhead) and the epithelial cell membrane (bar = 1 �m). Abbreviatiitochondria; (P) protozoan.
le, which is distinguished by the absence of the salivary acini,e localized protozoa inside the glandular cell, below the basal
amina (Fig. 4A and inset). In addition, we detected protozoa nearhe salivary duct observed in a transverse section of this lobule
Fig. 4B). Interestingly, B. culicis was present very close (Fig. 4C) orven inside the salivary acinus of the lateral lobule cells (Fig. 4D).alivary glands presenting protozoa frequently showed ultrastruc-ural damage, such as cytoplasm loss and loose acinus duct wallFig. 4C and D).ncubation the protozoan flagellum (arrow) is observed in the space between thec) salivary acini; (BL) basal lamina; (er) endoplasmic reticulum; (F) flagellum; (m)
3.3. Carbohydrate involvement in the B. culicis–salivary glandinteraction
Since both the protozoal plasma membrane and the sali-
vary basal lamina are rich in glycoconjugates, we examinedthe capacity of different carbohydrates and the lectin Con Ato inhibit the protozoa–salivary gland interaction. As shown inFig. 5A, addition of one of the following carbohydrates, 100 mM of�-methyl-mannoside, N-acetyl-glucosamine and l-fucose, signif-
M.T.C. Nascimento et al. / Acta Tropica 113 (2010) 269–278 273
Fig. 4. Ultrastructure of the Blastocrithidia culicis invasion of the salivary gland. (A) After overnight interaction, a protozoan (left upper square) is identified by its endosymbiontinside a cell of the median lobule, close to the transversal section of the salivary duct (asterisk; bar = 1 �m). Inset: detail of an endosymbiont-bearing protozoan under the basallamina (arrow; bar = 1 �m). (B) Longitudinal section of the salivary duct showing a protozoan (enclosed in a rectangle) whose flagellum is evidenced by the axonema. Notethe gland damage close to the protozoan (bar = 1 �m). Inset: higher magnification of the flagellum; arrow indicates the axonema (bar = 0.25 �m). (C) After gland invasion, B.culicis (P) is seen between the basal lamina and the acini of the salivary gland and also attached to the duct wall (asterisk). Note membrane profiles present in the same space( ry glaa d conda ; (SD)
i(pSiciwg
wmiwT
arrowhead; bar = 1 �m). (D) Protozoa were observed inside the acini of the salivalterations, including membrane shedding (arrow), a loose duct wall (arrowhead) ancini; (BL) basal lamina; (E) endosymbiont; (F) flagellum; (N) nucleus; (P) protozoa
cantly inhibited B. culicis adhesion to the salivary gland surfaceP < 0.0001). N-Acetyl-galactosamine was unable to affect therotozoan–salivary gland attachment. Addition of mannan and-Con A to the B. culicis–salivary gland interaction significantlynhibited binding (Fig. 5B). Interestingly, the pre-treatment of B.ulicis with mannan significantly decreased the protozoan bind-ng; however, pre-treatment of the salivary gland with S-Con A
as unable to affect the binding of the protozoa to the salivaryland.
To further explore carbohydrate expression in the protozoa,
e analyzed Con A-FITC binding to B. culicis by fluorescenceicroscopy and flow cytometry (Fig. 6). Con A strongly and specif-cally labeled B. culicis, as evidenced by the lectin binding to thehole protozoan cell membrane, including the flagellum (Fig. 6A).
his result was confirmed by flow cytometry analysis and by
nd cells. Salivary gland epithelial cells invaded by B. culicis display ultrastructuralensed chromatin on the nuclear periphery (bar = 1 �m). Abbreviations: (ac) salivarysalivary duct. Bars = 1 �m.
the specific inhibition of Con A labeling by �-methyl-mannoside(Fig. 6B).
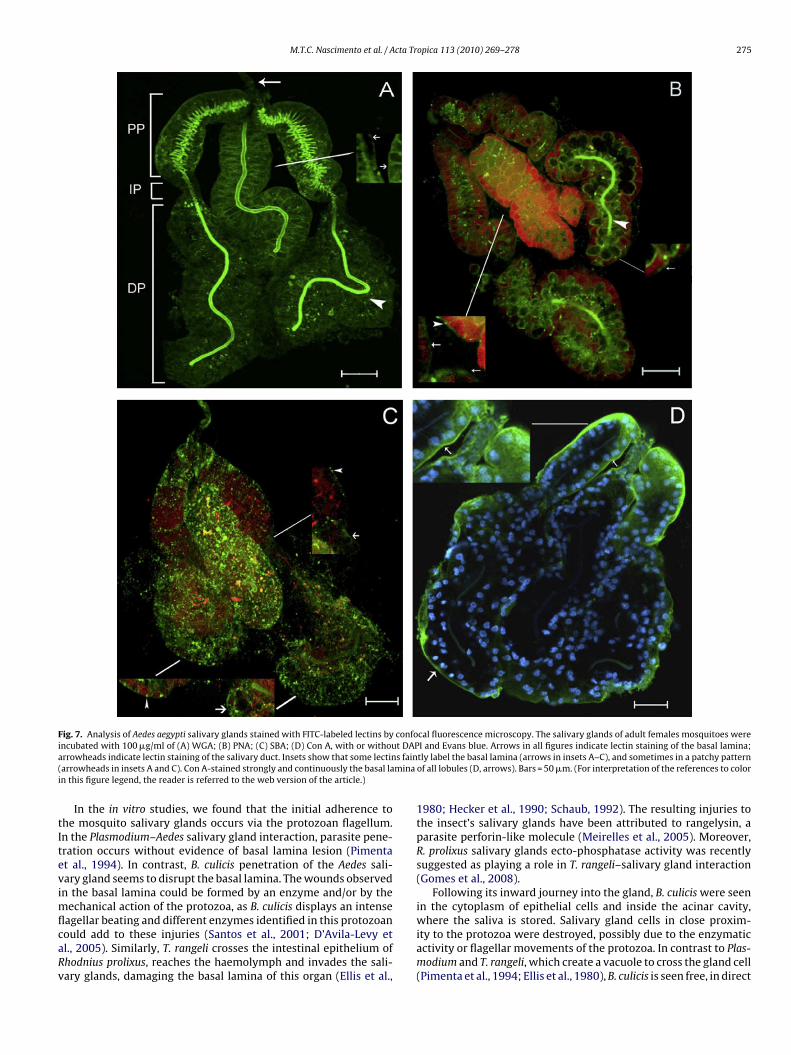
Additionally, carbohydrates present in the salivary glandswere analyzed using FITC-labeled lectins and confocal microscopy(Fig. 7). The results of salivary gland staining showed that allfour lectins tested bound to this organ. The different labeling pat-terns of particular lectins reveal the diversity of carbohydratesexpressed in the salivary glands. Wheat germ agglutinin (WGA)labeled the basal lamina, mainly at its proximal portion (Fig. 7A),while peanut lectin (PNA) uniformly labeled the basal lamina in
scattered patches (Fig. 7B) and soybean agglutinin (SBA) labeledthe basal lamina in patches (Fig. 7C). The entire extension of thesalivary duct was strongly labeled by WGA (Fig. 7A), but only itsdistal portion was labeled by PNA (Fig. 7B). SBA was unable to bindto the salivary duct (Fig. 7C). Con A labeled the basal lamina of all
274 M.T.C. Nascimento et al. / Acta Tr
Fig. 5. Carbohydrate inhibition of Blastocrithidia culicis adhesion to salivary glands.(A) Salivary glands were pre-incubated with 100 mM of �-methyl-mannoside(�-Man), N-acetyl-glucosamine (GluNAc) or l-fucose (l-Fuc) and 200 mM of N-acetyl-galactosamine (GalNAc), and then incubated with the protozoa. After 1 h ofinteraction, binding of B. culicis to the salivary glands was significantly inhibited bythe carbohydrates (P < 0.0001), except GalNAc. (B) Mannan or succinyl ConcanavalinA (S-Con A) at 100 �g/ml significantly inhibited the interaction (P = 0.0028 and 0.028,respectively). Additionally B. culicis was pre-treated with mannan (Bc pre-mannan)or the salivary glands were pre-treated with S-Con A (Sg pre-S-Con A) before theinteraction. Binding of B. culicis to the salivary gland was significantly inhibited onlyafter pre-treatment of the protozoa with mannan (P = 0.008). Each point representsthe number of B. culicis cells attached per salivary gland.
Fig. 6. Analysis of Concanavalin A binding to Blastocrithidia culicis by fluorescence microscbrightly labeled on the whole surface, as observed by confocal microscopy (bar = 10 �m).Con A (10 �g/mL) was confirmed by flow cytometry analysis (green line). Con A binding tocontrol protozoa (red line). (For interpretation of the references to color in this figure leg
opica 113 (2010) 269–278
salivary gland lobules uniformly but did not stain the salivary duct(Fig. 7D).
To confirm the involvement of mannose in the binding betweenthe protozoa and the salivary gland, we stained the B. culicisadhered to salivary gland with Con A-FITC (Fig. 8). Con A-FITC-labeled the cell surface of the protozoa, as well as the salivarygland basal lamina and the plasma membrane of the epithelial cells(Fig. 8A–D).
3.4. Survival of B. culicis in A. aegypti haemolymph
In order to adhere to and penetrate the mosquito salivary glands,B. culicis must survive the mosquito haemolymph defense mech-anisms. Thus, we tested for survival of the protozoa in dilutedmosquito haemolymph (Fig. 9). As determined by its motility after1-h incubation with mosquito haemolymph, B. culicis is indeed ableto survive the haemolymph defense mechanisms.
3.5. In vivo interaction of B. culicis with A. aegypti organs
To investigate whether B. culicis is able to bind to mosquitoorgans immersed in the haemolymph, we inoculated radiolabeledprotozoa into the mosquito thorax and measured the number ofB. culicis cells attached per organ after overnight incubation. Ourfindings show that, as in the in vitro assays, B. culicis is capable toadhere to the salivary glands (Fig. 10). Radioactive protozoa boundto the midgut and a small number bound to the ovaries were alsoobserved. A larger number of B. culicis cells was detected in thecarcasses of mosquitoes whose other organs were explanted; how-ever, we could not discriminate if they were adhered to the carcassitself or to the fat body that is normally attached to the carcass.
4. Discussion
The colonization of insects by trypanosomatids depends directlyon the interaction of the protozoa with the organs of these hosts. Bystudying the B. culicis–A. aegypti interaction, we previously demon-
strated that this protozoan is capable to colonize the intestine andcross the intestinal epithelium, reaching the mosquito haemocoel(Corrêa-da-Silva et al., 2006). Here we have demonstrated that B.culicis can survive in Aedes haemolymph and is able to adhere tothe A. aegypti salivary glands, both in vitro and in vivo.opy and flow cytometry. (A) Protozoa incubated with FITC-Con A (100 �g/mL) wereNonspecific fluorescence was quenched by Evans blue. (B) Labeling of B. culicis byB. culicis was specifically inhibited by 250 mM �-mannoside (blue line). Unlabeled
end, the reader is referred to the web version of the article.)
M.T.C. Nascimento et al. / Acta Tropica 113 (2010) 269–278 275
Fig. 7. Analysis of Aedes aegypti salivary glands stained with FITC-labeled lectins by confocal fluorescence microscopy. The salivary glands of adult females mosquitoes werei t DAPa s fain( minai
tItevimflcaRv
ncubated with 100 �g/ml of (A) WGA; (B) PNA; (C) SBA; (D) Con A, with or withourrowheads indicate lectin staining of the salivary duct. Insets show that some lectinarrowheads in insets A and C). Con A-stained strongly and continuously the basal lan this figure legend, the reader is referred to the web version of the article.)
In the in vitro studies, we found that the initial adherence tohe mosquito salivary glands occurs via the protozoan flagellum.n the Plasmodium–Aedes salivary gland interaction, parasite pene-ration occurs without evidence of basal lamina lesion (Pimentat al., 1994). In contrast, B. culicis penetration of the Aedes sali-ary gland seems to disrupt the basal lamina. The wounds observedn the basal lamina could be formed by an enzyme and/or by the
echanical action of the protozoa, as B. culicis displays an intense
agellar beating and different enzymes identified in this protozoanould add to these injuries (Santos et al., 2001; D’Avila-Levy etl., 2005). Similarly, T. rangeli crosses the intestinal epithelium ofhodnius prolixus, reaches the haemolymph and invades the sali-ary glands, damaging the basal lamina of this organ (Ellis et al.,I and Evans blue. Arrows in all figures indicate lectin staining of the basal lamina;tly label the basal lamina (arrows in insets A–C), and sometimes in a patchy patternof all lobules (D, arrows). Bars = 50 �m. (For interpretation of the references to color
1980; Hecker et al., 1990; Schaub, 1992). The resulting injuries tothe insect’s salivary glands have been attributed to rangelysin, aparasite perforin-like molecule (Meirelles et al., 2005). Moreover,R. prolixus salivary glands ecto-phosphatase activity was recentlysuggested as playing a role in T. rangeli–salivary gland interaction(Gomes et al., 2008).
Following its inward journey into the gland, B. culicis were seenin the cytoplasm of epithelial cells and inside the acinar cavity,
where the saliva is stored. Salivary gland cells in close proxim-ity to the protozoa were destroyed, possibly due to the enzymaticactivity or flagellar movements of the protozoa. In contrast to Plas-modium and T. rangeli, which create a vacuole to cross the gland cell(Pimenta et al., 1994; Ellis et al., 1980), B. culicis is seen free, in direct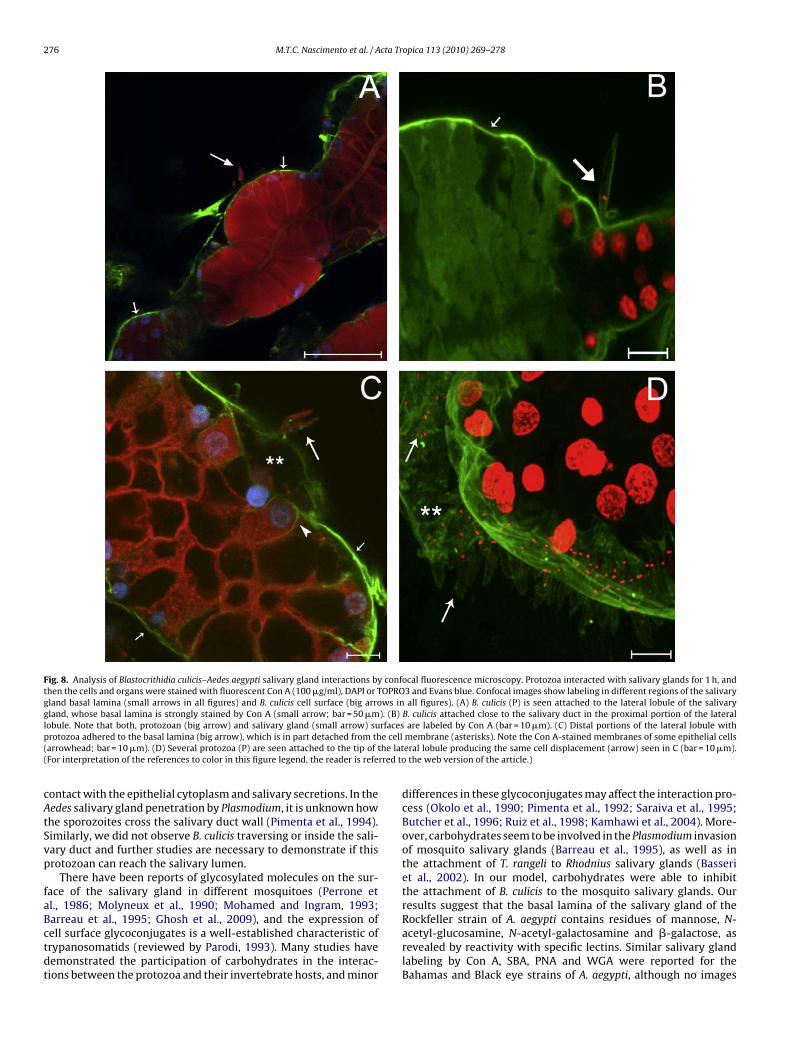
276 M.T.C. Nascimento et al. / Acta Tropica 113 (2010) 269–278
Fig. 8. Analysis of Blastocrithidia culicis–Aedes aegypti salivary gland interactions by confocal fluorescence microscopy. Protozoa interacted with salivary glands for 1 h, andthen the cells and organs were stained with fluorescent Con A (100 �g/ml), DAPI or TOPRO3 and Evans blue. Confocal images show labeling in different regions of the salivarygland basal lamina (small arrows in all figures) and B. culicis cell surface (big arrows in all figures). (A) B. culicis (P) is seen attached to the lateral lobule of the salivarygland, whose basal lamina is strongly stained by Con A (small arrow; bar = 50 �m). (B) B. culicis attached close to the salivary duct in the proximal portion of the laterall rfacesp e cell( he lat( rred to
cAtSvp
faBctdt
obule. Note that both, protozoan (big arrow) and salivary gland (small arrow) surotozoa adhered to the basal lamina (big arrow), which is in part detached from tharrowhead; bar = 10 �m). (D) Several protozoa (P) are seen attached to the tip of tFor interpretation of the references to color in this figure legend, the reader is refe
ontact with the epithelial cytoplasm and salivary secretions. In theedes salivary gland penetration by Plasmodium, it is unknown howhe sporozoites cross the salivary duct wall (Pimenta et al., 1994).imilarly, we did not observe B. culicis traversing or inside the sali-ary duct and further studies are necessary to demonstrate if thisrotozoan can reach the salivary lumen.
There have been reports of glycosylated molecules on the sur-ace of the salivary gland in different mosquitoes (Perrone etl., 1986; Molyneux et al., 1990; Mohamed and Ingram, 1993;arreau et al., 1995; Ghosh et al., 2009), and the expression of
ell surface glycoconjugates is a well-established characteristic ofrypanosomatids (reviewed by Parodi, 1993). Many studies haveemonstrated the participation of carbohydrates in the interac-ions between the protozoa and their invertebrate hosts, and minorare labeled by Con A (bar = 10 �m). (C) Distal portions of the lateral lobule withmembrane (asterisks). Note the Con A-stained membranes of some epithelial cellseral lobule producing the same cell displacement (arrow) seen in C (bar = 10 �m).the web version of the article.)
differences in these glycoconjugates may affect the interaction pro-cess (Okolo et al., 1990; Pimenta et al., 1992; Saraiva et al., 1995;Butcher et al., 1996; Ruiz et al., 1998; Kamhawi et al., 2004). More-over, carbohydrates seem to be involved in the Plasmodium invasionof mosquito salivary glands (Barreau et al., 1995), as well as inthe attachment of T. rangeli to Rhodnius salivary glands (Basseriet al., 2002). In our model, carbohydrates were able to inhibitthe attachment of B. culicis to the mosquito salivary glands. Ourresults suggest that the basal lamina of the salivary gland of theRockfeller strain of A. aegypti contains residues of mannose, N-
acetyl-glucosamine, N-acetyl-galactosamine and �-galactose, asrevealed by reactivity with specific lectins. Similar salivary glandlabeling by Con A, SBA, PNA and WGA were reported for theBahamas and Black eye strains of A. aegypti, although no images
M.T.C. Nascimento et al. / Acta Tr
Fig. 9. Blastocrithidia culicis survival in Aedes aegypti haemolymph. Protozoawere incubated with Aedes haemolymph or saline (ctrl) and motile viable try-panosomatids were counted in a Neubauer chamber. Results from at least threeindependent experiments are expressed as number of protozoa/mL. No signifi-cant difference was observed between the two groups, indicating that the insecthaemolymph does not promote protozoal lysis.
Fig. 10. In vivo binding of Blastocrithidia culicis to Aedes aegypti organs. (A) 125I-labeled-protozoa (105) were injected into a mosquito’s thorax, and indicated organswere explanted after incubation overnight. Results are expressed as the mean organ-acte
wwettcfrmlTgiiNtc
ailM
ssociated cpm (black bars) or mean number of protozoa/organ (gray bars), afteronverting organ-associated cpm to trypanosomatid numbers. These data representhe mean of two independent experiments, with 50 mosquitoes organs analyzed inach experiment.
ere shown (Perrone et al., 1986; Barreau et al., 1995). In addition,e have revealed labeling of internal salivary gland structures. For
xample, the labeling of the whole duct extension by WGA and onlyhe distal portion by PNA suggests that the carbohydrate composi-ion of this structure varies. Moreover, the presumed presence ofhitin in the A. aegypti salivary duct (Pimenta et al., 1994) was rein-orced by the bright labeling of this structure by WGA, a lectin thatecognizes chitin. The images presented here demonstrate that theannose-specific lectin Con A displayed a continuous and strong
abeling of the basal lamina, as well as of the B. culicis cell surface.o further confirm the role of this sugar in the B. culicis–salivaryland interaction, we found that mannan and S-Con A significantlynhibited protozoan adhesion. Moreover, mannose strongly inhib-ted B. culicis interaction with the salivary gland, as did fucose and-acetyl-glucosamine residues. Together, these results suggest that
he attachment of B. culicis to A. aegypti salivary glands involvesarbohydrates.
Although able to adhere to mosquito salivary glands, B. culicislso bound to midguts and ovaries, as demonstrated by in vitro bind-ng and confirmed by the in vivo binding assays using radioactivelyabeled protozoa. Contrary to B. triatomae, which interacts with
alpighian tubules of Triatoma infestans (Schaub and Schnitker,
opica 113 (2010) 269–278 277
1988; Schaub et al., 1990), B. culicis did not adhere to Malpighiantubules, or crop of A. aegypti (data not shown), suggesting somespecificity in organ adhesion.
A cascade of humoral and cellular responses is triggered byorganisms that gain access to the haemocoel of insects (Schaub,1994; Müller et al., 2008). T. rangeli survives in R. prolixushaemolymph and infects the insect salivary glands; however, thissame parasite never infects T. infestans salivary glands, becauseit is eliminated when it reaches the insect haemolymph (Heckeret al., 1990). Moreover, trypanolytic activity was described in theRhodnius haemolymph (Gregório and Ratcliffe, 1991). Thus, B. culi-cis should survive in the insect haemolymph in order to adhere toand penetrate mosquito salivary glands. As we demonstrated hereB. culicis survived in the A. aegypti haemolymph, and adhered todifferent organs after in vivo inoculation. However, the number ofB. culicis adhered to the different organs was smaller in vivo thanin vitro. Interestingly, it was recently reported that malaria sporo-zoites have a narrow window for salivary gland invasion, due toan efficient immune response in the mosquito haemocoel, whichlimits the number of parasites able to infect the gland (Hillyer etal., 2007).
In this report we demonstrate for the first time that B. culicis isable to adhere to A. aegypti salivary glands in vitro and in vivo, andsurvive after in vitro incubation with mosquito haemolymph. How-ever, further studies are needed to elucidate whether the protozoareaching the haemocoel are able to infect the salivary gland. Theseresults suggest that salivary gland invasion by B. culicis could be partof the life cycle of this monoxenic trypanosomatid in mosquitoes.
Acknowledgements
We thank the Laboratório de Fisiologia e Controle de Vetores,LAFICAVE for the Aedes aegypti supply and the Program for Techno-logical Development in Tools for Health-PDTIS/FIOCRUZ (FundacãoOswaldo Cruz, Rio de Janeiro) for use of confocal facilities. Wealso thank Bernardo Pascarelli for the confocal analysis. The exper-iments comply with the current laws of Brazil. This work wassupported by CNPq and FAPERJ.
References
Barreto-de-Souza, V., Medeiros, T.X., Motta, M.C.M., Bou-Habib, D.C., Saraiva, E.M.,2008. HIV-1 infection and HIV-1 Tat protein permit the survival and replicationof a non-pathogenic trypanosomatid in macrophages through TGF-�1 produc-tion. Microbes Infect. 10, 642–649.
Basseri, H.R., Tew, I.F., Ratcliffe, N.A., 2002. Identification and distribution of carbo-hydrate moieties on the salivary glands of Rhodnius prolixus and their possibleinvolvement in attachment/invasion by Trypanosoma rangeli. Exp. Parasitol. 100,226–234.
Barreau, C., Touray, M., Pimenta, P.F., Miller, L.H., Vernick, K.D., 1995. Plasmodiumgallinaceum: sporozoite invasion of Aedes aegypti salivary glands is inhibited byanti-gland antibodies and by lectins. Exp. Parasitol. 81, 332–343.
Butcher, B.A., Turco, S.J., Hilty, B.A., Pimenta, P.F., Panunzio, M., Sacks, D.L., 1996.Deficiency in beta 1,3-galactosyltransferase of a Leishmania major lipophos-phoglycan mutant adversely influences the Leishmania-sand fly interaction. J.Biol Chem. 271, 20573–20579.
Corrêa-da-Silva, M.S., Fampa, P., Lessa, L.P., Silva, E.R., Santos, J.R.M., Saraiva, E.M.,Motta, M.C.M., 2006. Colonization of Aedes aegypti midgut by the endosymbiont-bearing trypanosomatid Blastocrithidia culicis. Parasitol. Res. 99, 384–391.
D’Avila-Levy, C.M., Araújo, F.M., Vermelho, A.B., Soares, R.M., Santos, A.L., Bran-quinha, M.H., 2005. Proteolytic expression in Blastocrithidia culicis: influenceof the endosymbiont and similarities with virulence factors of pathogenic try-panosomatids. Parasitology 130, 413–420.
Ellis, D.S., Evans, D.A., Stamford, S., 1980. The penetration of the salivary glands ofRhodnius prolixus by Trypanosoma rangeli. Z. Parasitenkd. 62, 63–74.
Fampa, P., Corrêa-da-Silva, M.S., Lima, D.C., Oliveira, S.M., Motta, M.C., Saraiva, E.M.,
2003. Interaction of insect trypanosomatids with mosquitoes, sand fly and therespective insect cell lines. Int. J. Parasitol. 33, 1019–1026.Ghosh, A.K., Devenport, M., Jethwaney, D., Kalume, D.E., Pandey, A., Anderson, V.E.,Sultan, A.A., Kumar, N., Jacobs-Lorena, M., 2009. Malaria parasite invasion of themosquito salivary gland requires interaction between the Plasmodium TRAP andthe Anopheles saglin proteins. PLoS Pathog. 5, e1000265.

2 cta Tr
G
G
H
H
H
J
K
M
M
M
M
M
M
O
78 M.T.C. Nascimento et al. / A
omes, S.A., Fonseca de Souza, A.L., Kiffer-Moreira, T., Dick, C.F., Santos, A.L.,Meyer-Fernandes, J.R., 2008. Ecto-phosphatase activity on the external sur-face of Rhodnius prolixus salivary glands: modulation by carbohydrates andTrypanosoma rangeli. Acta Trop. 106, 137–142.
regório, E.A., Ratcliffe, N.A., 1991. The distribution of agglutinins and lytic activityagainst Trypanosoma rangeli and erythrocytes in Rhodnius prolixus and Tri-atoma infestans tissue extracts and haemolymph. Mem. Inst. Oswaldo Cruz 86,181–186.
agedorn, H.H., Turner, S., Hagedorn, E.A., Pontecorvo, D., Greenbaum, P., Pfeiffer,D., Wheelock, G., Flanagan, T.R., 1977. Postemergence growth of the ovarianfollicles of Aedes aegypti. J. Insect Physiol. 23, 203–206.
ecker, H., Schwarzenbach, M., Rudin, W., 1990. Development and interactions ofTrypanosoma rangeli in and with the reduviid bug Rhodnius prolixus. Parasitol.Res. 76, 311–318.
illyer, J.F., Barreau, C., Vernick, K.D., 2007. Efficiency of salivary gland invasionby malaria sporozoites is controlled by rapid sporozoite destruction in themosquito haemocoele. Int. J. Parasitol. 37, 673–681.
ansen, A.M., Carreira, J.C., Deane, M.P., 1988. Infection of a mammal by mono-genetic insect trypanosomatids (Kinetoplastida, Trypanosomatidae). Mem. Inst.Oswaldo Cruz 83, 271–272.
amhawi, S., Ramalho-Ortigão, M., Pham, V.M., Kumar, S., Lawyer, P.G., Turco, S.J.,Barillas-Mury, C., Sacks, D.L., Valenzuela, J.G., 2004. A role for insect galectins inparasite survival. Cell 119, 329–341.
eirelles, R.M., Henriques-Pons, A., Soares, M.J., Steindel, M., 2005. Penetration of thesalivary glands of Rhodnius domesticus Neiva & Pinto, 1923 (Hemiptera: Reduvi-idae) by Trypanosoma rangeli Tejera, 1920 (Protozoa: Kinetoplastida). Parasitol.Res. 97, 259–269.
cGhee, R.B., 1957. Infection of chick embryos by a Crithidia from a phytophagoushemipteron. Science 125, 157–158.
ohamed, H.A., Ingram, G.A., 1993. Salivary gland surface carbohydrate variationsin three species of the Anopheles gambiae complex. Ann. Soc. Belg. Med. Trop.73, 197–207.
olyneux, D.H., Okolo, C.J., Lines, J.D., Kamhawi, M., 1990. Variation in fluorescein-labelled lectin staining of salivary glands in the Anopheles gambiae complex.Med. Vet. Entomol. 4, 459–462.
orio, F., Reynes, J., Dollet, M., Pratlong, F., Dedet, J.P., Ravel, C., 2008. Isola-tion of a protozoan parasite genetically related to the insect trypanosomatid
Herpetomonas samuelpessoai from a human immunodeficiency virus-positivepatient. J. Clin. Microbiol. 46, 3845–3847.üller, U., Vogel, P., Alber, G., Schaub, G.A., 2008. The innate immune system ofmammals and insects. Contrib. Microbiol. 15, 21–44.
kolo, C.J., Jenni, l., Molineaux, D.H., Wallbanks, K.R., 1990. Surface car-bohydrate differences of Glossina salivary glands and infectivity of Try-
opica 113 (2010) 269–278
panosoma brucei gambiense to Glossina. Ann. Soc. Belg. Med. Trop. 70,39–47.
Pacheco, R.S., Marzochi, M.C.A., Pires, M.Q., Brito, C.M.M., Madeira, M.F.,Barbosa-Santos, E.G.O., 1998. Parasite genotypically related to a monoxenoustrypanosomatid of dog’s flea causing opportunistic infection in an HIV positivepatient. Mem. Inst. Oswaldo Cruz 93, 531–537.
Parodi, A.J., 1993. N-glycosylation in trypanosomatid protozoa. Glycobiology 3,193–199.
Perrone, J.B., DeMaio, J., Spielman, A., 1986. Regions of mosquito salivary glandsdistinguished by surface lectin-binding characteristics. Insect Biochem. 16,313–318.
Pimenta, P.F., Turco, S.J., McConville, M.J., Lawyer, P.G., Perkins, P.V., Sacks, D.L., 1992.Stage-specific adhesion of Leishmania promastigotes to the sand fly midgut.Science 256, 1812–1815.
Pimenta, P.F., Touray, M., Miller, L., 1994. The journey of malaria sporozoites in themosquito salivary gland. J. Eukaryot. Microbiol. 41, 608–624.
Podlipaev, S.A., 2000. Insect trypanosomatids: the need to know more. Mem. Inst.Oswaldo Cruz 95, 517–522.
Ruiz, R.C., Favoreto, S., Dorta, M.L., Oshiro, M.E.M., Ferreira, A.T., Manke, P.M.,Yoshida, N., 1998. Infectivity of Trypanosoma cruzi strains is associated withdifferential expression of surface glycoprotein with differential Ca+2 signalingactivity. Biochem. J. 330, 505–511.
Santos, A.L., Abreu, C.M., Batista, L.M., Alviano, C.S., de Araújo Soares, R.M., 2001.Cell-associated and extracellular proteinases in Blastocrithidia culicis: influenceof growth conditions. Curr. Microbiol. 43, 100–106.
Saraiva, E.M., Pimenta, P.F., Brodin, T.N., Rowton, E., Modi, G.B., Sacks, D.L., 1995.Changes in lipophosphoglycan and gene expression associated with the devel-opment of Leishmania major in Phlebotomus papatasi. Parasitolology 111,275–287.
Schaub, G.A., 1992. The effects of trypanosomatids on insects. Adv. Parasitol. 31,255–319.
Schaub, G.A., 1994. Pathogenicity of trypanosomatids on insects. Parasitol. Today 10,463–468.
Schaub, G.A., Reduth, D., Pudney, M., 1990. The peculiarities of Blastocrithidia tri-atomae. Parasitol. Today 6, 361–363.
Schaub, G.A., Schnitker, A., 1988. Influence of Blastocrithidia triatomae (Trypanoso-matidae) on the reduviid bug Triatoma infestans: alterations in the Malpighian
tubules. Parasitol. Res. 75, 88–97.Wallace, F.G., 1966. The trypanosomatids parasites of insects and arachnids. Exp.Parasitol. 18, 124–193.
Warren, L.G., 1960. Metabolism of Schizotrypanum cruzi, Chagas. 1. Effect of cul-ture age and substrate concentration on respiratory rate. J. Parasitol. 46,529–539.




















