
Integrated biostratigraphy and palaeoenvironments of an upper Santonian – upper Campanian...
16
Integrated biostratigraphy and palaeoenvironments of an upper Santonian e upper Campanian succession from the southern part of the Eastern Carpathians, Romania Claudia G. Cetean a, * , Ramona B alc b , Michael A. Kaminski c , Sorin Filipescu a a Babes ¸-Bolyai University, Department of Geology, M. Kog alniceanu 1, 400084 Cluj Napoca, Romania b Babes ¸-Bolyai University, Faculty of Environmental Sciences, Fântânele Street 30, 400294 Cluj Napoca, Romania c Earth Sciences Department, King Fahd University of Petroleum and Minerals, Dhahran 31261, Saudi Arabia article info Article history: Received 16 June 2010 Accepted in revised form 10 November 2010 Available online 19 November 2010 Keywords: Foraminifera Calcareous nannofossils SantonianeCampanian Romania Agglutinated foraminifera morphogroups abstract Study of an upper Santonian to upper Campanian hemipelagic succession from the southern part of the Romanian Eastern Carpathians enables us to establish an integrated biostratigraphy based on planktonic foraminifera and calcareous nannofossils and to compare this record with the agglutinated foraminiferal biozonation used for the Carpathians. Benthic foraminiferal assemblages were investigated using several methods, such as agglutinated and calcareous benthic foraminiferal morphogroups, and the benthic foraminiferal oxygen index in order to determine their response to environmental parameters in the basin (correlated with sea-level maxima documented by regional sea-level curves for the Tethys). A pattern of changes in benthic foraminiferal communities associated with increased organic carbon flux and rising sea-levels can be summarized as follows in the studied succession. As sea-level begins to rise there is an increase in the proportion of calcareous benthic foraminifera at the expense of agglutinated foraminifera within the benthic assem- blages (earliest Campanian, mid-late Campanian). Once sea-level rises, an increase in the elongate keeled morphotype of agglutinated foraminifera (shallower water forms) can be observed, and if sea-level remains high for an extended period (as in the early Campanian) then an invasion of both agglutinated and benthic calcareous foraminifera characteristic of outer shelf-upper slope environments take place in the basin. The variations in tubular and deep infaunal morphotypes of agglutinated foraminifera are ascribed to varying levels of organic carbon flux. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction The late Santonian to late Campanian interval represents a period of major sea-level fluctuations in the Tethyan area (Lüning et al., 1998; Li et al., 2000). Among these changes the earliest Campanian sea-level maximum appears to be the most pronounced one with documented changes in benthic foraminiferal assem- blages worldwide. Low diversity foraminiferal assemblages and high numbers of radiolaria are reported in the Polish and Romanian Carpathians in the lower Campanian (Neagu, 1970; Ba ˛ k, 2000), in the Boreal seaway between Greenland and Norway (Gradstein et al., 1999), including a peak in pyritized diatoms, and faunal changes are even reported in the southern Atlantic by Basov and Krasheninnikov (1983). These most probably reflect a period with high organic carbon flux a result of mostly increased primary productivity during periods of sea-level rise (Erbacher et al., 1996). Other important sea-level maxima and minima are documented in the Tethys in the late Santonian to late Campanian interval in regional sea-level curves from Egypt and Tunisia (Hancock, 1993; Lüning et al., 1998; Li et al., 2000) such as late Santonian and mid-late Campanian maxima or a late Campanian minimum. Continuous successions of Upper Cretaceous middle bathyal deposits are rarely exposed in the Carpathians; usually outcrops of this age in the Romanian, Polish, and Czech Carpathians expose turbidite deposits representing a deeper (basinal) environment (e.g., Neagu, 1970). In central-eastern Europe the chalk deposits from Poland and Romania are better studied for this time interval (e.g., Neagu, 1992; Peryt, 1988; Gawor-Biedowa, 1992). Lithologi- cally similar deposits to those exposed in the southern Eastern Carpathians of Romania were studied in the Subsilesian Unit of the Polish Carpathians already in the late 19th century by Grzybowski (1896) revealing well preserved and diverse benthic foraminiferal * Corresponding author. Present address: Institute of Geological Sciences, Polish Academy of Sciences, ul. Senacka 1, 31-002 Kraków, Poland. E-mail address: [email protected] (C.G. Cetean). Contents lists available at ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/CretRes 0195-6671/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.cretres.2010.11.001 Cretaceous Research 32 (2011) 575e590
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Integrated biostratigraphy and palaeoenvironments of an upper Santonian – upper Campanian...

lable at ScienceDirect
Cretaceous Research 32 (2011) 575e590
Contents lists avai
Cretaceous Research
journal homepage: www.elsevier .com/locate/CretRes
Integrated biostratigraphy and palaeoenvironments of an upperSantonian e upper Campanian succession from the southern partof the Eastern Carpathians, Romania
Claudia G. Cetean a,*, Ramona B�alc b, Michael A. Kaminski c, Sorin Filipescu a
aBabes-Bolyai University, Department of Geology, M. Kog�alniceanu 1, 400084 Cluj Napoca, RomaniabBabes-Bolyai University, Faculty of Environmental Sciences, Fântânele Street 30, 400294 Cluj Napoca, Romaniac Earth Sciences Department, King Fahd University of Petroleum and Minerals, Dhahran 31261, Saudi Arabia
a r t i c l e i n f o
Article history:Received 16 June 2010Accepted in revised form 10 November 2010Available online 19 November 2010
Keywords:ForaminiferaCalcareous nannofossilsSantonianeCampanianRomaniaAgglutinated foraminifera morphogroups
* Corresponding author. Present address: InstituteAcademy of Sciences, ul. Senacka 1, 31-002 Kraków, P
E-mail address: [email protected] (C.G. Cete
0195-6671/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.cretres.2010.11.001
a b s t r a c t
Study of an upper Santonian to upper Campanian hemipelagic succession from the southern part of theRomanian Eastern Carpathians enables us to establish an integrated biostratigraphy based on planktonicforaminifera and calcareous nannofossils and to compare this record with the agglutinated foraminiferalbiozonation used for the Carpathians.
Benthic foraminiferal assemblages were investigated using several methods, such as agglutinated andcalcareous benthic foraminiferal morphogroups, and the benthic foraminiferal oxygen index in order todetermine their response to environmental parameters in the basin (correlated with sea-level maximadocumented by regional sea-level curves for the Tethys). A pattern of changes in benthic foraminiferalcommunities associated with increased organic carbon flux and rising sea-levels can be summarized asfollows in the studied succession. As sea-level begins to rise there is an increase in the proportion ofcalcareous benthic foraminifera at the expense of agglutinated foraminifera within the benthic assem-blages (earliest Campanian, mid-late Campanian). Once sea-level rises, an increase in the elongate keeledmorphotype of agglutinated foraminifera (shallower water forms) can be observed, and if sea-levelremains high for an extended period (as in the early Campanian) then an invasion of both agglutinatedand benthic calcareous foraminifera characteristic of outer shelf-upper slope environments take place inthe basin. The variations in tubular and deep infaunal morphotypes of agglutinated foraminifera areascribed to varying levels of organic carbon flux.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
The late Santonian to late Campanian interval representsa period of major sea-level fluctuations in the Tethyan area (Lüninget al., 1998; Li et al., 2000). Among these changes the earliestCampanian sea-level maximum appears to be themost pronouncedone with documented changes in benthic foraminiferal assem-blages worldwide. Low diversity foraminiferal assemblages andhigh numbers of radiolaria are reported in the Polish and RomanianCarpathians in the lower Campanian (Neagu, 1970; Bak, 2000), inthe Boreal seaway between Greenland and Norway (Gradsteinet al., 1999), including a peak in pyritized diatoms, and faunalchanges are even reported in the southern Atlantic by Basov andKrasheninnikov (1983). These most probably reflect a period with
of Geological Sciences, Polisholand.an).
All rights reserved.
high organic carbon flux a result of mostly increased primaryproductivity during periods of sea-level rise (Erbacher et al., 1996).Other important sea-level maxima and minima are documented inthe Tethys in the late Santonian to late Campanian interval inregional sea-level curves from Egypt and Tunisia (Hancock, 1993;Lüning et al., 1998; Li et al., 2000) such as late Santonian andmid-late Campanian maxima or a late Campanian minimum.
Continuous successions of Upper Cretaceous middle bathyaldeposits are rarely exposed in the Carpathians; usually outcrops ofthis age in the Romanian, Polish, and Czech Carpathians exposeturbidite deposits representing a deeper (basinal) environment(e.g., Neagu, 1970). In central-eastern Europe the chalk depositsfrom Poland and Romania are better studied for this time interval(e.g., Neagu, 1992; Peryt, 1988; Gawor-Biedowa, 1992). Lithologi-cally similar deposits to those exposed in the southern EasternCarpathians of Romania were studied in the Subsilesian Unit of thePolish Carpathians already in the late 19th century by Grzybowski(1896) revealing well preserved and diverse benthic foraminiferal

Fig. 1. Location map of the Izlaz Valley section, Eastern Carpathians.
C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590576
assemblages. Unfortunately, the present-day outcrops of the Sub-silesian Unit are often tectonised and represented by small andpatchy exposures. Therefore, the Izlaz Valley section analyzed inthis study, is an ideal setting to document the biostratigraphy ofbenthic foraminifera and interpret their palaeoecology in relationto global changes, since it occupied an approximately centralposition in Tethys and the Upper Cretaceous deposits are wellexposed in the studied area.
The objectives of this study are: (1) to establish an integratedbiostratigraphy of the upper Santonian to upper Campanianinterval by means of planktonic foraminifera, agglutinated fora-minifera and calcareous nannofossils (2) to calibrate the biostrati-graphical schemes based on agglutinated foraminifera against theplanktonic microfossil record (3) to document the changes in thebenthic foraminifera assemblages as a result of changes in envi-ronmental conditions in the basin (associated with sea-levelmaxima) by using quantitative methods such as agglutinatedforaminiferal morphogroups, calcareous benthic morphogroups,and the benthic foraminiferal oxygen index (4) to investigate theidea that variations in the agglutinated foraminiferal morphotypescan be correlated with the organic carbon flux and therefore serveas a proxy for sea-level change in the mentioned period.
2. Material and Methods
The deposits investigated in this study represent a part of theUpper Cretaceous post-tectonic cover (the westernmostextremity) of the Ceahl�au Nappe, Outer Dacides, Eastern Carpa-thians (S�andulescu et al., 1981). The upper Santonian to upperCampanian succession is exposed on a small right-side tributary(Izlaz Valley; 45�23’N, 25�18’E) of the Dâmbovita River, in theCotenesti locality, in the southernmost part of the Eastern Car-pathians (Fig. 1). The lower part of the exposures consists of
alternating red glauconitic sandstones, grey siltstones andmudstones of the Plaiu Formation (Stef�anescu, 1970; 1995). Redsiltstones are also present near the base of the outcrop (1e3 m).Based on the present integrated biostratigraphy we assign thisformation to the upper Santonian e upper Campanian (Fig. 2),while Stef�anescu (1995) considered it to represent only the upperCampanian. However, this interpretation was arbitrarily based onthe fact that it is overlain by the Gura Beliei Formation, which is oflate Campanian age at its base. The three glauconitic sand/sand-stone levels, two in the upper Santonian (Fig. 2) and one in theupper part of the lower Campanian, consist of only quartz grains,sponge spicules, glauconite, and foraminifera with very rarehematite, and are here interpreted to represent importantregressions. The upper part of the succession consists of varie-gated but predominately red siltstones of the lower Member of theGura Beliei Formation.
We investigated 72 samples for foraminifera and 58 samplesfor calcareous nannoplankton from the Izlaz Valley exposure.Samples investigated for calcareous nannofossils were preparedusing the standard smear slide technique. No special techniquewas used to clean and concentrate the biotic fraction, the mainpurpose being preservation of the initial assemblage composition.Observations were carried out under a light microscope ata magnification of 1000�. The preservation of the calcareousnannofossils is moderate, samples yielding a medium diversity (ca.50 species/sample) while abundance is low (1e5 specimens/fieldof view).
Foraminiferal samples were prepared using standard micropa-leontological methods, with more than 300 specimens/samplepicked. Preservation of the foraminiferal assemblages varies frommoderate to good with the exception of the uppermost Santonianto lower Campanian when dissolution of calcareous (both benthicand planktonic) foraminifera is visible. Examples include the loss of
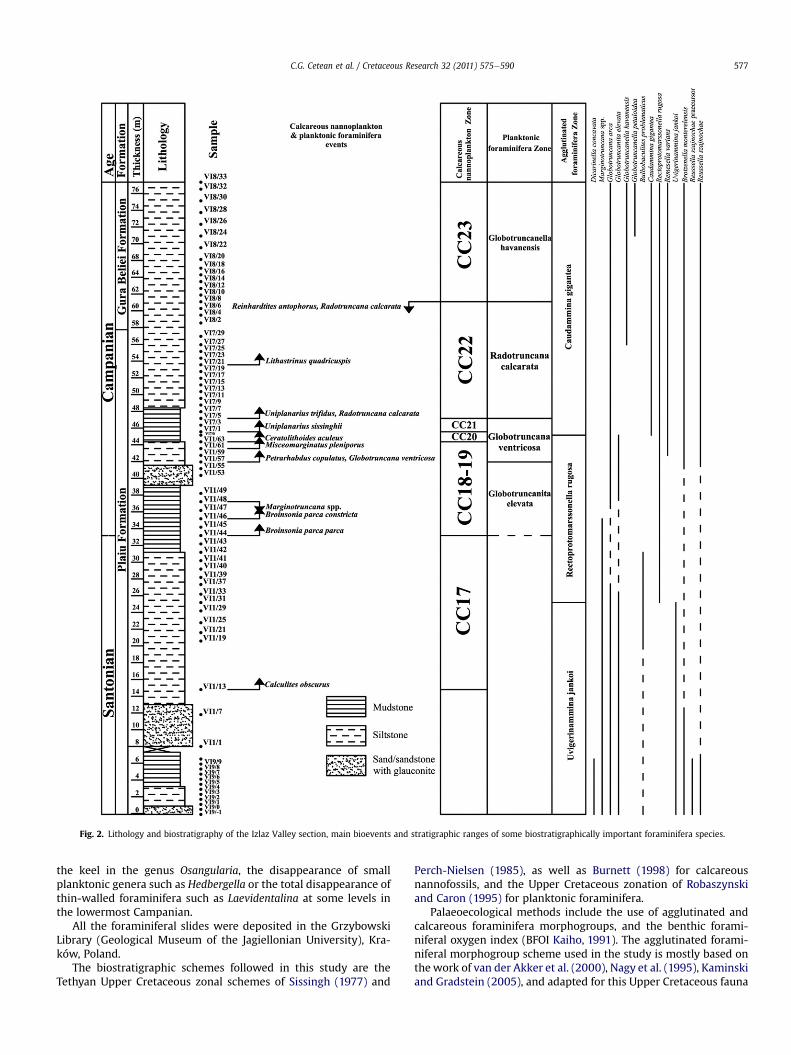
Fig. 2. Lithology and biostratigraphy of the Izlaz Valley section, main bioevents and stratigraphic ranges of some biostratigraphically important foraminifera species.
C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590 577
the keel in the genus Osangularia, the disappearance of smallplanktonic genera such as Hedbergella or the total disappearance ofthin-walled foraminifera such as Laevidentalina at some levels inthe lowermost Campanian.
All the foraminiferal slides were deposited in the GrzybowskiLibrary (Geological Museum of the Jagiellonian University), Kra-ków, Poland.
The biostratigraphic schemes followed in this study are theTethyan Upper Cretaceous zonal schemes of Sissingh (1977) and
Perch-Nielsen (1985), as well as Burnett (1998) for calcareousnannofossils, and the Upper Cretaceous zonation of Robaszynskiand Caron (1995) for planktonic foraminifera.
Palaeoecological methods include the use of agglutinated andcalcareous foraminifera morphogroups, and the benthic forami-niferal oxygen index (BFOI Kaiho, 1991). The agglutinated forami-niferal morphogroup scheme used in the study is mostly based onthe work of van der Akker et al. (2000), Nagy et al. (1995), Kaminskiand Gradstein (2005), and adapted for this Upper Cretaceous fauna

Table 1Agglutinated foraminifera morphotypes used in this study (modified after Cetean, 2009).
Morphogroup Morphotype Test form Life position Feeding habit Environment Main genera
M1 Tubular Erect epifauna Suspension feedingTranquil bathyal and abyssalwith low organic flux
NothiaRhizamminaKalamopsisBathysiphonArthrodendronDendrophrya
M2
M2a Globular Shallow infaunaSuspension feeding and/or Passive deposit feeding
Common inbathyal and abyssal
SaccamminaPlacentamminaCaudamminaHyperamminadilatata
M2b
Rounded trochospiraland streptospiral
Surficial epifauna Active deposit feeding Shelf to deep marine
RccurvoidesCribrostomoidesAtaxophragmium
Planoconvextrochospiral
Trochammina
M2c Elongate keeled Surficial epifauna Active deposit feeding Shelf to marginal marineSpiroplectinellaSpiroplectammina
M3
M3a
Flattened trochospiral
Surficial epifaunaActive and passivedeposit feeding
Lagoonal to abyssal
Flattened planispiraland streptospiral
AmmodiscusGlomospiraRepmaninaRzehakinaLituotuba
M3b Flattened irregular Surficial epifauna Suspension feeding Upper bathyal to abyssal Ammolagena
M3c Flattened streptospiral Surficial epifaunaActive and passivedeposit feeding
Upper bathyal to abyssalParatrochamminoidesTrochamminoidesAmmosphaeroidina
C.G.Cetean
etal./
CretaceousResearch
32(2011)
575e590
578

M4
M4a
Rou
nded
planispiral
Surfi
cial
epifau
naan
d/
orsh
allow
infauna
Activedep
ositfeed
ing
Inner
shelfto
upper
bathya
lHap
lophragm
oides
M4b
Elon
gate
subc
ylindrical
Dee
pinfauna
Activedep
ositfeed
ing
Inner
shelfto
upper
bathya
lwith
increa
sedorga
nic
matterflux
Rem
esella
Gau
dryina
Dorothia
Trftax
iaMarsson
ella
Arenob
ulim
ina
Geroc
ham
mina
Hormosina
Elon
gate
tapered
Reo
phax
Subreo
phax
Bulbob
aculites
Ammob
aculites
C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590 579
by Cetean (2009, Table 1). Calcareous foraminiferal morphogroupsused in the study are based on the work of Koutsoukos and Hart(1990) and Frenzel (2000, Table 2). It should be mentioned thatalthough Koutsoukos and Hart (1990) considered the genus Osan-gularia to be an epifaunal form, this study follows Frenzel (2000) inregarding it as an infaunal form (probably shallow infaunal) due toits thin wall and almost flattened morphology (high surface/volume ratio interpreted as characteristic for infaunal forms, Corlissand Chen, 1988).
3. Results
3.1. Biostratigraphy
3.1.1. Planktonic foraminiferaUpper Santonian. Unfortunately, the index species Dicarinella
asymetrica is rare in coeval investigated foraminiferal assemblagesin Romania (Ion and Szasz, 1994). This taxon is absent in the IzlazValley section e only Dicarinella concavata was found in thesuccession, but this species is also very rare and therefore itsobserved LO may be doubtful (Fig. 2). The planktonic foraminiferarecovered in the lower part of the section consist of Con-tusotruncana fornicata, Marginotruncana spp., Globotruncana arca,G. lapparenti, Globotruncanita elevata, G. stuartiformis and D. con-cavata. This assemblage permits, albeit tentatively, the assignmentof these deposits to the upper Santonian.
Campanian. Uppermost Santonian to lowermost Campanianplanktonic foraminiferal assemblages from the Izlaz Valley sectionare poor owing to dissolution. Small genera such as Heterohelix andHedbergella are almost absent from the assemblages, as previouslyshown, small planktonic foraminiferawere the first ones to dissolve(e.g., Nguyen et al., 2009). Among the larger planktonic genera,Marginotruncana appears to have been more resistant to dissolu-tion than Globotruncanita and Globotruncana probably due to itsthicker wall.
As mentioned above, because Dicarinella asymetrica is missing,the lower limit of the Globotruncanita elevata Zone cannot beestablished precisely in this section. The Santonian/Campanianboundary is approximated here by the FO of the calcareous nan-nofossil Broinsonia parca parca. The disappearance of the genusMarginotruncana takes place in the middle part of the Globo-truncanita elevata Zone, above the FOs of the calcareous nannofossilBroinsonia parca parca as observed in the Bottacione section ofcentral Italy by Premoli Silva and Sliter (1995). Other planktonicforaminiferal bioevents observed in the Izlaz Valley section include:the LO of Globotruncanita elevata in the Globotruncana ventricosaZone and the FO of Globotruncanella havanensis in the Radotruncanacalcarata Zone (Plate 2).
3.1.2. Calcareous nannoplanktonCalcareous nannofossils also yield a late Santonian to late
Campanian age for the deposits exposed in Izlaz Valley. Based onseveral bioevents, seven biozones were recognized, starting withthe upper Santonian (Plate 4).
The FO of Calculites obscurus marks the base of Zone CC17; theupper part of this biozone broadly coincides with the upper part ofthe Dicarinella asymetrica Zone in Tethys (e.g., in the Bottacionesection of Italy, reported by Premoli Silva and Sliter, 1995). ZoneCC17was subdivided byWagreich (1992) into three subzones basedon bioevents that were not observed in the Izlaz Valley section.
The CC18 biozone is defined by the FO of Broinsonia parca parcaand corresponds with the lower part of the Globotruncanita elevataZone. The absence in the studied section of the planktonic fora-minifer Dicarinella asymetrica leads to a questionable lower limit ofthe Globotruncanita elevata Zone, but this zonal boundary is usually

C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590580
placed above the base of Zone CC18 (Erba et al., 1995; Premoli Silvaand Sliter, 1995; Gardin et al., 2001; Romero De et al., 2003; Huet al., 2005) or at the same level (Erba et al., 1995 adapted to the2008 time scale).
The CC19 biozone cannot be recognized in the Izlaz Valleysection due to the absence of the index species Marthasterites fur-catus, a species that was not encountered anywhere in the section.The absence of this species was also noted in Romania in thenorthwestern part of the Hateg Basin (Grigorescu and Melinte,2002) and in the Apuseni Mountains (B�alc et al., 2007). In thisinterval (CC18-CC19) a series of bioevents were recorded: the FO ofBroinsonia parca constricta, FO of Petrarhabdus copulatus and the FOof Misceomarginatus pleniporus. The FO of Petrarhabdus copulatuswas observed below the FOs of the Tethyan species Ceratolithoidesaculeus, Uniplanarius sissinghii and Uniplanarius trifidus and ata lower stratigraphic level than observed in northwestern Australiaby Campbell et al. (2004).
The FO of C. aculeus defines the base of Zone CC20 and takesplace in the Globotruncana ventricosa Zone as previously recordedat Gubbio by Premoli Silva and Sliter (1995) and Gardin et al. (2001).
Zones CC21 and CC22 are separated based on the followingbioevents: the FO of Uniplanarius sissinghii and respectively the FOof Uniplanarius trifidus. The latest event occurs in the Izlaz Valleysection at the same level as the FO of the planktonic foraminiferRadotruncana calcarata. In Austria, Neuhuber and Wagreich (2009)also placed these two bioevents at the same level, as well as Gardinet al. (2001) in the Bottacione section, Italy, but usually the FOs ofthis species are recorded at different stratigraphic levels (Erba et al.,1995; Cunha et al., 1997; Yildiz and Özdemir, 1999). The CC21biozone was subdivided by Sissingh (1977) into three subzonesbased on the range of Ceratolithoides arcuatus, but this marker wasnot observed in the studied section. The FO of Prediscosphaerastoveri was observed in this biozone, although this species isrecorded from lowerMaastrichtian (Catanzariti et al., 2007) or fromthe base of the UC15d Zone (Burnett, 1998).
The upper limit of Zone CC22, as defined by the LO of Rein-hardtites antophorus, coincides with the upper limit of the Rado-truncana calcarata Zone, as both species disappear at the samelevel, thus these two biozones are coeval in the Izlaz Valley section.The FO of Lithastrinus quadricuspis was observed in the middle ofZone CC22. Sissingh (1977) suggested subdividing this zone basedon the FO of Reinhardtites levis, but this is difficult to accomplish inthe studied section because of the presence of intermediary formsbetween this species and Reinhardtites antophorus.
Table 2Calcareous benthic foraminiferal morphogroups used in this study (modified after Frenz
Morphogroup Sub-Morphogroup Morphotype Life posi
CH-A 1 Trochospiral, conical Epifauna
2 Planoconvex trochospiral Epifauna
3 Biconvex trochospiral, lenticular Epifauna4 not present in this study5 Planispiral to Liniserial Epifauna
shallowCH-B 1 Cylindrical Infauna
2 Planispiral with many pores Infauna3 Spherical Infauna4 Pyramidal lo conical Infauna
5 Flattened Infauna6 Ovoidal Infauna7 Low trochospiral,
symmetric bilateralInfauna
CH-C not present in this study
The exposed upper part of the Gura Beliei siltstones is assignedto the CC23 Zone and corresponds with the Globotruncanellahavanensis Zone of planktonic foraminifera.
3.1.3. Agglutinated foraminiferaThe biozonation based on agglutinated foraminifera realized in
this study follows the Upper Cretaceous scheme of Neagu et al.(1992) for the Eastern Carpathians, Romania, and Geroch andNowak (1984) for the Polish Carpathians. There are, however,small differences between the two schemes, which are mentionedbelow.
Top of the Uvigerinammina jankoi Zone. The U. jankoi Zone hasbeen used in the Carpathians in the upper Turonian to Santonianinterval by Neagu et al. (1992) and Geroch and Nowak (1984); itsupper limit is given by the FO of Rectoprotomarssonella rugosa andusually coincides with the Santonian/Campanian boundary. Onlythe upper part of this biozone is exposed in the Izlaz Valley sectionand the FO of R. rugosa was observed here at the same level as theLO of U. jankoi in the uppermost Santonian.
Rectoprotomarsonella rugosa Zone. Neagu et al. (1992) in theRomanian Eastern Carpathians and Geroch and Nowak (1984) inthe Polish Carpathians defined a “Goessella rugosa” (reassigned tothe new genus Rectoprotomarssonella by Kaminski et al., 2008)biozone for the lower Campanian to cover the interval up to the firstoccurrence of Caudammina gigantea, which usually appears in theupper part of the lower Campanian in the Carpathians. In the IzlazValley section, the FO of Rectoprotomarssonella rugosawas observedin the upper Santonian, and coincides with the LO of Uviger-inammina jankoi as mentioned above.
Caudammina gigantea Zone. In the Izlaz Valley section the FO ofCaudammina gigantea was observed in the middle Globotruncanaventricosa Zone (upper Campanian, Fig. 2). It is possible that the FOand distribution of this taxon is depth related as suggested byKuhnt et al. (1998) in the Atlantic Ocean Hole 959D where the FOwas recorded as high as upper Maastrichtian. This anomalouslylate FO was ascribed to the shallow setting of the site before thelate Maastrichtian. If Caudammina gigantea is a depth-restrictedspecies (restricted to the lower bathyal zone according to Kuhntet al., 1998) this will explain the large difference between itsfirst occurrences within the Carpathians, Tethys, and Atlanticbasins. However, as estimated by Cetean (2009), the deposition inthe Izlaz Valley section took place around and above 1000 mpaleodepth, and therefore the upper depth limit of this taxon maybe considered as middle bathyal. This hypothesis was confirmed
el, 2000).
tion Feeding habit Main genera
Active deposit feeding Gyroidinoides Eponides,Globorotalites, Nuttalinella
Active deposit feeding Gavelinella, Brotzenella,Cibicidoides,Alabamina, Stensioeina
Active deposit feeding Lenticulina Angulogavelinella, Planularia
/infauna
Deposit feeding Neoflabellina, Vagimdina, Frondicularia,Astacolus, Citharinella
Deposit feeding Nodosarids, Hemirobulina, Marginiilina,Ramulina, Ellipsoidella grcicilima, E. kugleri
Active deposit feeding AnomalinoidesActive deposit feeding Pullenia, LagenidsActive deposit feeding Reussella, Pleurostomella, Ellipsodimorphina,
Lagenids (elongated farms), PraebuliminaActive deposit feeding CoryphostomellaActive deposit feeding Fissurina, Guttulina, AllomorphinaActive deposit feeding Osangularia

late 1. Agglutinated foraminifera from the Izlaz Valley section. 1. Dendrophrya dichotomica Neagu, (VI7/11) 2. Rhizammina sp., (VI8/26) 3. Psammosphaera irregularis (Grzybowski),I7/11) 4. Ammodiscus cretaceus (Reuss), (VI7/11) 5. Dolgenia pennyi (Cushman and Jarvis), (VI7/15) 6. Glomospira irregularis (Grzybowski), (VI7/27) 7. Rzehakina epigona (Rzehak),I8/24) 8. Caudammina crassa (Geroch), (VI7/19) 9. Caudammina ovuloides (Grzybowski), (VI7/13) 10. Arthrodendron diffusum Ulrich, (VI8/14) 11. Arthrodendron moniliformis Neagu,I8/14) 12. Arthrodendron moniliformis Neagu, detail of the wall showing a Contusotruncana used to build the test, (VI8/14) 13. Reophax pilulifer (Brady), (VI7/11) 14. Hormosina
elascoensis (Cushman), (VI7/27) 15. Ammosphaeroidina pseudopauciloculata (Mjatliuk), (VI8/30) 16. Haplophragmoides aff. horridus (Grzybowski), (VI8/33) 17. Spiroplectamminaavarroana Cushman, (VI7/21) 18. Spiroplectinella israelskyi (Hillebrandt), (VI8/14) 19. Spiroplectinella dentata (Alth), (VI8/20) 20. Tritaxia subparisiensis (Grzybowski), (VI7/11)1. Spiroplectinata compressiuscula (Chapman), (VI1/37) 22. Gaudryina carinata Franke, Izlaz (VI8/24) 23. Remesella varians (Glaessner), (VI8/28) 24. Rectoprotomarssonella rugosaanzlíková), (VI8/2) 25. Marssonella trochus (d’Orbigny), (VI8/2)
P(V(V(Vvn2(H

late 2. Planktonic and calcareous benthic foraminifera from the Izlaz Valley section. 1. Chrysalogonium elongatum, Cushman and Jarvis, (VI7/11) 2. Laevidentalina gracilis (d’Or-igny), (VI7/15) 3. Laevidentalina oligostegia (Reuss), (VI8/24) 4. Laevidentalina pseudonana (Ten Dam), (VI8/14) 5. Nodosaria limbata (d’Orbigny), (VI7/0) 6. Pseudonodosaria manifestaeuss), (VI8/33) 7. Neoflabellina jarvisi (Cushman), (VI8/16) 8. Hemirobulina sp., (Cushman), (VI8/16) 9. Lagena paucicosta Franke, (VI8/32) 10. Fissurina bicornis Neagu, (VI7/19) 11a-c.lobotruncana ventricosa White, a e spiral view, b e umbilical view, c e lateral view, (VI7/0) 12a-c. Globotruncanita elevata (Brotzen), a e spiral view, b e umbilical view, c e lateraliew, (VI7/0) 13a-b. Different specimens of Radotruncana calcarata (Cushman), a e spiral view (VI7/23), b e umbilical view (VI7/27) 14. Aragonia ouezzanensis (Rey), (VI8/24) 15.eussella szajnochae (Grzybowski), (VI7/5) 16. Ellipsoidella kugleri (Cushman and Renz), (VI7/11) 17. Ellipsopolymorphina velascoensis (Cushman), (VI8/16) 18. Pleurostomella dacicaeagu, (VI7/5) 19. Pleurostomella pseudocurta Neagu, (VI7/9) 20a-b. Cibicidoides voltzianus (d’Orbigny), a e spiral view, b e oblique-lateral view (VI8/16) 21a-b. Cibicidoideslascoensis (Cushman), a e spiral view, b e oblique-lateral view, (VI8/30)
Pb(RGvRNve

Plate 3. Calcareous benthic foraminifera from the Izlaz Valley section. 1a-b. Quadrimorphina allomorphinoides (Reuss), a e spiral view, b e umbilical view (VI7/3) 2a-b. Anomalinoidesnobilis (Brotzen), a e spiral view, be umbilical view, (VI7/27) 3. Gyroidinoides nitidus (Reuss), lateral view, (VI7/15) 4a-b. Gyroidinoides globosus (Hagenow), a e spiral view, b e lateralview, (VI 8/32) 5a-c. Stensioeina pommerana (Brotzen), ae spiral view, bedorsal view, ce lateral view, (VI7/0). 6a-b.Gavelinella sandidgei (Brotzen), ae spiral view, bedorsal view, (VI7/5) 7a-b.Brotzenellamonterelensis (Marie), ae spiral view, bedorsal view, (VI7/3) 8.Brotzenellamonterelensis (Marie), spiral view, (VI7/19) 9.Angulogavelinella avnimelechi (Reuss), spiralview, (VI8/30) 10-11a-b. Eponides praemegastoma (Mjatliuk),10e spiral view (VI7/27),11ae spiral viewandbeumbilical view, (VI8/30) 12a-b.Gyroidina anomaloides (White), ae spiralview, b e umbilical view, (VI8/26)

C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590584

Fig. 3. Diversity (Fisher a), agglutinated foraminifera morphotypes (morphotypes under 5% were not plotted) and relative abundance (% of the assemblage) of two agglutinatedgeneraMarssonella and Gaudryina, plotted against sea-level curves for Tethys (1 e Hancock, 1993; 2 e Lüning et al., 1998; 3 eLi et al., 2000), arrows pointing at the highest values ofM4b morphotype and the lowest values of M1 morphotype during the early Campanian and mid-late Campanian sea-level maxima.
C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590 585
by Bak (2000) in the Pieniny Klippen Belt of Poland where he alsoestimated an upper depth limit of upper-middle bathyal for thisspecies. The FO of Caudammina gigantea differs greatly within theCarpathian basins. Neagu et al. (1992) observed it in the RomanianEastern Carpathians in the lower Campanian as Geroch and Nowak(1984) did in the Polish Outer Carpathians. Olszewska (1997) alsoestablished an acme zone with Caudammina gigantea in the upperSantonian e lower Campanian in the Polish Outer Carpathians.
In this zone, the FO of Remesella varians is the earliest observedoccurrence (Fig. 2), which takes place in the Globotruncana ven-tricosa Zone in the Izlaz Valley section. Usually this taxon appears inthe Maastrichtian, either in the lower Maastrichtian in Poland andRomania (Olszewska, 1997; Ion et al., 1998) or more commonly inthe middle-upper Maastrichtian in the rest of the Tethys and NorthAtlantic where it is used in several stratigraphical schemes basedon agglutinated foraminifera (Kuhnt et al., 1992; Kuhnt andKaminski, 1997).
Plate 4. Calcareous nannofossils from the Izlaz Valley section. All photographs taken under cmagnum Wind and Wise in Wise and Wind, 1977 (VI7/5) 3. Broinsonia parca constricta (HattnNoël, 1969 (VI1/44) 5. Ceratolithoides aculeus (Stradner, 1961) Prins and Sissingh in Sissingh, 1(Deflandre, 1959) Prins and Sissingh, in Sissingh, 1977 (VI1/29) 8. Cyclagelosphaera rein(Arkhangelsky, 1912) Deflandre, in Piveteau, 1952 (VI7/5) 10. Eiffelithus eximius (Stover, 1966)1954) Reinhardt, 1965 (VI1/19) 12. Kamptnerius magnificus Deflandre, 1959 (VI1/55) 13. Loxodecoratus Deflandre, 1959 (VI8/33) Eiffelithus turriseiffelii (Deflandre, in Deflandre and Fert, 191977 (VI 1/61) 16. Micula staurophora Vekshina, 1959 (VI 1/19) 17. Petrarhabdus copulatus(Arkhangelsky, 1912) Gartner, 1968 (VI1/19) 19. Placozygus fibuliformis (Reinhardt, 1964) Hoff(VI1/41) 21. Reinhardtites levis (Deflandre, 1959) Perch-Nielsen, 1968 (VI1/63) 22. RetecapsaRucinolithus? magnus Bukry, 1975 (VI7/5) 24. Tranolithus orionatus Stover, 1966 (VI1/19) 25.(VI1/55) 26-27. Uniplanarius ssissinghii Perch-Nielsen, 1984 (VI7/1) 28.Uniplanarius trifidus ((VI7/5) 29. Watznaueria barnesae (Black, 1959) Perch-Nielsen, 1984 (VI1/29) 30. Zeugrhabdo
3.1.4. Calcareous benthic foraminiferaSome calcareous benthic taxa used in biostratigraphic schemes
in the Santonian to lower Campanian such as Reusella szajnochaepraecursor and Brotzenella monterelensis (Gawor-Biedowa 1992;Robaszynski et al., 2005) yielded a different stratigraphic range inthe studied section. Both R. szajnochae and R. szajnochae praecursorwere found to range from the base of the outcrop (upper Santonian)in the Izlaz Valley section. While R. szajnochae praecursor disap-pears in the upper Santonian together with R. szajnochae, the laterreappears in the lower part of the G. ventricosa Zone and hasa continuous stratigraphic range in the upper Campanian. Itstemporarily disappearance can be linked with the presence ofa different type of fauna in the uppermost Santonian to lowerCampanian as documented in this study (Plate 3).
Stensioeina pommerana is present in the foraminiferal assem-blages starting with the upper Santonian, which is consistent withthe findings of Olszewska (1997) from the Polish Outer Carpathians
ross-polarised light. 1. Arkhangelskiella cymbiformis Vekshina, 1959 (VI1/44) 2. Biscutumer et al., 1980) Perch-Nielsen, 1984a (VI1/46) 4. Broinsonia parca parca (Stradner, 1963)977 (VI1/63) 6. Ceratolithoides sesquipedalis Burnett, 1998 (VI7/25) 7. Calculites obscurushardtii (Perch-Nielsen, 1968) Romein, 1977 (VI1/39) 9. Cribrosphaerella ehrenbergiiPerch-Nielsen, 1968 (VI1/19) 11. Eiffelithus turriseiffelii (Deflandre, in Deflandre and Fert,lithus armilla (Black in Black and Barnes, 1959) Noel, 1965 (VI1/29) 14. Microrhabdulus54) Reinhardt, 1965 15.Misceomarginatus pleniporus Wind and Wise in Wise and Wind,(Deflandre, 1959) Wind and Wise in Wise, 1983 (VI1/57) 18. Prediscosphaera cretaceamann, 1970(VI1/39) 20. Reinahardtites antophorus (Deflandre, 1959) Perch-Nielsen, 1968crenulata (Bramlette and Martini, 1964) Grün in Grün and Allemann, 1975 (VI1/19) 23.Uniplanarius gothicus (Deflandre, 1959) Prins and Perch-Nielsen in Manivit et al. (1977)Stradner in Stradner and Papp, 1961) Prins and Perch-Nielsen in Manivit et al. (1977)tus embergeri (Noel, 1959) Perch-Nielsen, 1984 (VI1/40)

Fig. 4. Agglutinated/calcareous benthic foraminifera ratio, relative abundance (% of the assemblage) of the genera Globorotalites, Osangularia and Stensioeina, calcareous benthicmorphogroups and BFOI in the Izlaz Valley section, plotted against sea-level curves for Tethys (1 e Hancock, 1993; 2 e Lüning et al., 1998; 3 e Li et al., 2000), arrows pointing at thehighest values of the calcareous benthic foraminifera and the highest values of the benthic foraminifera infauna morphogroups during the early Campanian and mid-late Cam-panian sea-level maxima.
C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590586
and those of Gawor-Biedowa (1992) from epicontinental depositsin SE Poland. Usually this species is recorded for the first time in thelower Campanian in the chalk facies as reported by Neagu (1992) inDobrogea, Hampton et al. (2007) in the English chalk at SeafordHead, Sussex; and Peryt (1988) from the Lublin Upland, Poland.
Brotzenella monterelensis is also present in the benthic forami-niferal assemblages beginning with the upper Santonian, while theFO of this species is usually recorded in the lower or upper Cam-panian in chalk deposits (Neagu, 1992; Robaszynski et al., 2005;Gawor-Biedowa, 1992). This species displays the same behavior asReussella szajnochae and temporarily disappears in the uppermostSantonian to lower Campanian only to return in the lower part ofG. ventricosa Zone.
3.2. Benthic foraminiferal assemblages
3.2.1. Diversity and type of assemblagesDiversity varies greatlywithin the studied section; values are high
in the upper Santonian but decrease significantly in the lowermostCampanian. Low values are also observed in the mid-upper Campa-nian (upper part of theG. ventricosa Zone to lowermostRadotruncanacalcarata Zone). The low values probably reflect impoverishedassemblages dominated by both agglutinated and calcareous benthicspecies which prefer more eutrophic conditions as a result of theincreased organic flux into the basin associated with the two sea-levelmaxima that occur at these levels in the Tethys (Hancock,1993;Lüning et al., 1998; Li et al., 2000). Diversity gradually increasestowards the upper Campanian and attains a maximum in the red(oxic) deposits of the Gura Beliei Formation (Fig. 3).
The benthic assemblages are dominated by agglutinated fora-minifera with values around 70-80%, however, calcareous benthicforaminifera appear to increase in number during the two sea-levelmaxima mentioned above (Fig. 4). The same decrease in theproportion of agglutinated foraminifera within benthic assem-blages was observed in lowermost Campanian chalk deposits inPoland (Peryt, 1988) and in the upper Cenomanian of the Basco-Cantabrian Basin in Spain (Gräfe andWendler, 2003), both intervalswith raised sea-levels. It is possible that coarse agglutinated fora-miniferal species may have difficulties in building a test duringthese intervals, as finer sedimentation prevails and the availabilityof larger quartz grains becomes more limited.
Agglutinated foraminifera are dominated by tubular coarselyagglutinated forms, which have mean values of 50% of the wholebenthic assemblage. However, during the sea-level maxima calcar-eous-cemented species such as Clavulinoides, Dorothia,Marssonella,Arenobulimina, andGaudryina increase in number, becoming almostequal in numbers with the tubular forms.
In general, the mixture of agglutinated, calcareous benthic andplanktonic foraminifera fits verywell within the low latitude “Slope-marl” biofacies of Kaminski and Gradstein (2005). However, thepresence of Rzehakina, a genus restricted to flysch-type assemblages(Kaminski and Gradstein, 2005) in the upper Santonian and upperCampanian, together with other species characteristic for this biof-aciespoint toa transition fromthe “Slope-marl”biofacies toa “Flysch-type” one, especially in the upper Campanian where the FOs ofseveral other flysch-type agglutinated species such as Arthrodendrondiffusum, A. moniliformis, and Nothia latissima are observed.
As mentioned above, the proportion of agglutinated calcareous-cemented species such as Clavulinoides, Marssonella (Fig. 3),

C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590 587
increased during the earliest Campanian and the mid-late Cam-panian sea-level maxima, and the assemblages can be consideredto represent more the concept of the “Slope-marl” biofacies. This isparticularly true for the earliest Campanian sea-level maximum,which due to its longer extent, allowed bathymetrically shallowtaxa to invade the basin, an interpretation sustained both bycalcareous benthic and agglutinated foraminifera. Agglutinatedspecies such as Tritaxia dubia, T. tripleura, Spiroplectammina bau-douiniana, and S. elongata known from the chalk deposits inDobrogea, France and England (e.g., Neagu,1989) seem to appear inthe deeper part of the basin and disappear as soon as the earlyCampanian regression took place. During the two sea-level maximawe also observe an increase of several species of Gaudryina (Fig. 3).These are associated with an increased organic flux and dysaerobicconditions, as mentioned by Nyong and Olsson (1983) andHolbourn et al. (1999).
The hypothesis of a shallow water fauna (outer shelf-upperslope) invasion into the basin during the earliest Campanian is alsosupported by changes in the calcareous benthic assemblages. Thereare few species that particularly increase in number during thistime such as Globorotalites michelinianus, a species characteristic toan outer shelf - upper slope environments (e.g., Peryt, 1988), whichusually has a patchy distribution in the studied section but attains8% of the foraminiferal assemblage in the lowermost Campanian.Others species such as Osangularia cordieriana show increasedvalues also during the mid-late Campanian sea-level maximum,even higher than in the earliest Campanian. However, this can alsorepresent an effect of dissolution since Osangularia is a thin-walledform that can be easily dissolved (Fig. 4). The invasion of shallowwater taxa during the early Campanian is also proved by thepresence of typical outer shelf - upper bathyal species such asLingulogavelinella stelligera and L. lenticula (Neagu, 1992; Peryt,1988; Ion et al., 1998, etc.) which were not present in the lateSantonian and disappeared during the regression that followed theearly Campanian sea-level maximum.
Sedimentological studies of the upper Cretaceous successions inthe area do not show any evidence for a shallowing trend whichmay explain this change of the foraminifera assemblages, the depthof deposition increases through the Late Cretaceous towards theEocene flysch deposits (Stef�anescu, 1970; 1995). Also regionalfluctuations of the CCD are improbable since the this basin wasa part of Tethys and the presence of similar benthic assemblageswith Contessa Highway section, Italy (Cetean, 2009) or Poland (Bak,2000) do not support any restrictions, as well as other microfossils(Melinte and Jipa, 2005). Finally, while redeposition cannot becompletely excluded, it is highly unlikely, foraminifera are notabraded or broken and the type of sediment (mudstones) does notindicate a high-energy environment.
3.2.2. Paleoenvironments as documented by agglutinated andcalcareous morphogroups and BFOI
In the agglutinated foraminifera morphogroup model presentedby Kaminski and Gradstein (2005), organic matter arriving at thesea floor will be first scavenged by the tubular (M1 morphotype)and globular (M2a morphotype) agglutinated species that extendtheir pseudopodia to capture food from the water column. Theorganic matter that arrives on the sediment surface will be thenconsumed by epifaunal or shallow infaunal (M2b, M2c, M3a, M3b,M4c morphotypes) that utilize various feeding strategies, eitheractive or passive. The remaining organic matter will be incorpo-rated by bioturbators into the sediment, and made available to thedeep infaunal taxa (M4a, M4b morphotypes).
We expect that during periods of rapid sea-level rise associatedwith higher primary productivity and therefore higher organicmatter influx into the basin (Erbacher et al., 1996), more organic
matter will be available to sustain high populations of the deepinfaunal morphogroup (M4b), which is situated at the end of thefood chain. The increase in the infaunal morphogroup can also beaugmented by the fact that oxidation of organic matter can result indysoxic pore waters within the sediment. This favours the infaunalforms, as they are already able to tolerate low oxygen conditions.This hypothesis seems to be accurate in the Izlaz Valley aggluti-nated foraminiferal morphotype analysis (Fig. 3). During theearliest Campanian and mid-late Campanian (upper G. ventricosa-lowermost Radotruncana calcarata) sea-level maxima the values ofthe infaunal agglutinated morphotype M4b increase to 50% and38% respectively.
By contrast, higher values of morphotype M1 of tubular agglu-tinated foraminifera (due to its feeding strategy) and the lowestvalues of the deep infaunal morphogroups are expected duringperiods of low organic matter flux to the sea floor, for example thiscan be observed in the late Campanian (Globotruncanella hava-nensis Zone) and can be associated with an important regression,documented by Li et al. (2000) in Tunisia.
In the early Campanian an increase in morphotype M2c repre-sented by Spiroplectinella and Spiroplectammina can be alsoobserved. These are generally regarded as shallow water taxa (shelfto marginal marine). Their increase together with the deep infaunamorphotype M4b may further lead to the idea of an invasion ofshallower taxa in this middle bathyal setting, as these formsprobably found more favourable conditions at greater depthsduring periods of high sea-level. Similar high values in this mor-photypewere documented by Cetean (2009) in the same area in theuppermost Cenomanian, suggesting that variations in this mor-photype can be used as a proxy for high sea-level.
As is the case with agglutinated foraminifera, the infaunalmorphogroup (CH-B) of calcareous benthic foraminifera showshigher values during the mid-late Campanian sea-level maxima(Fig. 4). However, the epifauna morphogroup seem to dominate thelatest Santonian to early Campanian interval, but we assume thatthis can be caused by dissolution of thin-walled infaunal taxa suchas Laevidentalina and Osangularia during this interval.
The benthic foraminiferal dissolved oxygen index confirms ourinterpretation regarding a higher organic flux into the basin (asshown by the increase in infaunal morphogroups both in calcar-eous and agglutinated foraminifera) and dysaerobic conditionsduring the two sea-level maxima. BFOI values fall in the uppermostSantonian to lowermost Campanian and mid-upper Campanian,and indicate dysoxic bottom waters. Though expected to showlower values during the early Campanian sea-level maximum, theBFOI shows that bottom waters were still dysoxic, but with loweroxygen values than in the mid-late Campanian. As mentionedabove, it is possible that due to the dissolution recorded in thisperiod, thin-walled infaunal forms will have lower numberspreserved in the sediment, therefore any analysis based on ratios ofcalcareous benthics will be compromised by dissolution.
4. Discussion
4.1. Integrated biostratigraphy
Low sedimentation rates in the lowermost upper Campanianmay account for the closely spaced FO’s of some species of calcar-eous nannofossils such as Ceratolithoides aculeus, Uniplanarius sis-singhii and U. trifidus. High sea-level during this interval is inferredfrom changes in the benthic foraminiferal assemblages and also bythe deposition of finer sediments, but the relative order of thenannofossil first occurrences is correct. Also, high sea-level isdocumented by several regional sea-level curves during thisinterval (Lüning et al., 1998; Li et al., 2000).

C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590588
The integrated planktonic foraminiferal e calcareous nanno-fossil biostratigraphy from the Izlaz Valley section shows signifi-cantly differences with respect to the 2004 and 2008 GeologicalTime Scale (Ogg et al., 2008, calcareous nannofossils and planktonicforaminifera modified from Erba et al., 1995) but it seems to matchvery well with other integrated biostratigraphic schemes such asthe one published by Yildiz and Özdemir (1999) from SE Turkey,Campbell et al. (2004) from the northwestern Australian marginand partially to the Premoli Silva and Sliter (1995) biostratigraphicscheme from the Bottacione section, central Italy, and the�Svábenická et al. (1997) scheme from the Magura Nappes of theCzech Republic.
The presence of the agglutinated species Spiroplectinata com-pressiuscula during the lower Campanian and of the nannofossilPetrarhabdus copulatus in the uppermost lower Campanian, pointout to a southern latitude influence in the early Campanian.P. copulatus is a species frequently mentioned from latitudesbetween 30�-40� in the southern hemisphere (Wind, 1975) and israrely encountered in the northern hemisphere (Varol, 1992;�Svábenická, 1995; Gasi�nski et al., 1999; �Svábenická et al., 2002;Kedzierski, 2008). The species Spiroplectinata compressiuscula(Chapman) was described from the Santonian of WesternAustralia (Chapman, 1917) and mentioned also in Santonian -Campanian deposits of western South Atlantic Leg 39 by Sliter(1977).
4.2. Utility of foraminifera morphogroups
Morphogroups of agglutinated foraminifera are used in a varietyof studies as indicators of paleobathymetry and/or organic carbonflux (Kaminski et al., 2006; Kender et al., 2008). This method has itslimitations as discussed by Kender et al. (2008), but overall it seemsmore reliable than calcareous benthic foraminifera morphogroupsespecially in settings affected temporarily by dissolution, as in thisstudy. In general, in the Izlaz Valley section the agglutinated mor-photypes seem to represent reliable proxies for the increasedcarbon flux during the earliest Campanian andmid-late Campaniansea-level maxima widely recognized in Europe (Hancock, 1993;Lüning et al., 1998; Li et al., 2000).
High-resolution sampling of this section raises the questionwhether or not additional variations of organic carbon fluxassociated with sea-level fluctuations can be recognized in thestudied interval. For example in the upper Santonian, high valuesof morphotype M4b and low values of the M1 tubular forms mayindicate another period of high sea-level followed by an impor-tant regression (represented by the second level of glauconitesand/sandstone), after which sea-level started to rise againtowards the earliest Campanian as indicated again by aggluti-nated morphotypes, with another increase in the M4b infaunamorphotype. This observation fits well with the sea-level curvefor NW Europe (Hancock, 1993), with the same succession oftransgresive e regressive cycles.
In the upper Campanian Radotruncana calcarata Zone, thegeneral trend of increase in the M1 morphotype and decrease ofmorphotype M4b values suggest low organic flux associated withdecreasing sea-levels in the Tethys (e.g., Lüning et al., 1998; Li et al.,2000). The high percentage of the M1 tubular morphotype and lowM4b deep infaunal numbers in the uppermost part of Globo-truncanella havanensis Zone may also indicate another period oflow organic carbon flux associated with the late Campanianregression, as seen in the Tethyan sea-level curves (e.g., Li et al.,2000). Unfortunately, this event is impossible to accurately corre-late owing to the absence of Gansserina gansseri in the studiedsection.
5. Conclusions
Based on calcareous nannofossils, planktonic and agglutinatedforaminifera from the Izlaz Valley section, an integrated biostra-tigraphy was established for upper Santonian to upper Campanianof the southern Eastern Carpathians, Romania (Plate 1).
Study of the benthic foraminiferal assemblages in the IzlazValley section document the earliest Campanian invasion ofbathymetrically shallower taxa (outer shelf-upper bathyal) into thismiddle bathyal setting, which points out that caution is necessarywhen interpreting paleobathymetry based on benthic foraminifera.The late Santonian deep-water type of benthic foraminiferalassemblage is almost completely replaced in the latest Santonian e
earliest Campanian by assemblages that bear strong affinities withthose reported from chalk deposits in the nearby Dobrogea shelfand other European chalk deposits.
We consider the use of the planktonic/ benthonic ratio (P/Bratio) as indicator of paleobathymetry to be doubtful during sea-level maxima in this type of setting due to several factors that caninfluence it (as documented by Zwaan van der et al., 1990) such asdissolution of planktonic foraminifera and higher numbers ofbenthic foraminifera (especially infauna) in dysoxic environments(to be expected during sea-level maxima). In the Izlaz Valleysection Cetean (2009) had shown that the P/B ratio suddenlydecreases from 90% in the late Santonian to 50% in the earliestCampanian only to return to values over 90% in themid Campanian.Another drop to 78% corresponding to themid-late Campanian sea-level maximum was observed after which the values remainedaround 95% for the whole late Campanian. While the appliedcorrection suggested by Zwaan van der et al. (1990) of not takinginto account the infaunal forms during dysoxic intervals pushed themid-late Campanian P/B values to similar values in the data set(around 90%), the early Campanian values of the P/B ratio stillremained around 65e70%.
Therefore, the use of the P/B ratio during these intervalstogether with the composition of the benthic foraminiferalassemblages will lead to the false conclusion that the depositionalsetting was bathymetrically shallower.
As this study shows in the Izlaz Valley section, agglutinatedforaminiferal morphotypes can be considered good indicators oforganic carbon flux. The high values of the deep infauna morpho-type M4b (and the decrease of M1 morphotype of tubular forms)points to an increased organic carbon flux, which correlates withsea-level maxima in the Tethys during the earliest Campanian andmid-late Campanian. Also, morphotypeM2c represented usually byshallow water taxa (shelf to marginal marine) increases in valueswhen sea-level remains high for a longer period, as was the case inthe early Campanian (e.g., Lüning et al., 1998). Agglutinated fora-miniferal morphogroups are considered here to be more reliablethan calcareous benthic foraminiferal morphogroups, which areaffected by dissolution during sea-level maxima.
The benthic foraminifera dissolved oxygen index (BFOI) appearsalso correlates with the sea-level changes indicating dysoxicbottom waters during the two sea-level maxima, which can beexpected owing to sluggish deep-water circulation.
A middle bathyal setting with hemipelagic deposition seemsideal for using the agglutinated foraminiferal morphotypes, butfurther studies of sedimentological and geochemical studies andagglutinated foraminiferal morphogroups will be needed to assesstheir paleoecological importance.
Acknowledgements
CGC wishes to thank the NURC (CNCSIS grant 475/2007) forfinancial support obtained during PhD studies, and the Cushman

C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590 589
Foundation for a grant to support fieldwork (theW.V. Sliter Award).We thank Theodor Neagu (University of Bucharest) for help withsome of the taxonomical problems encountered in this study, EiichiSetoyama (PAN Kraków) for redrawing the agglutinated morpho-types table and two anonymous reviewers for suggestion thatimproved the present manuscript.
References
Akker, van der, T.J.H.A., Kaminski, M.A., Gradstein, F.M., Wood, J., 2000. Campanianto Palaeocene biostratigraphy and palaeoenvironments in the Foula Sub-basin,west of the Shetland Islands, UK. Journal of Micropalaeontology 19, 23e43.
Basov, I.A., Krasheninnikov, V.A., 1983. Benthic foraminifers in Mesozoic andCenozoic sediments of the southwestern Atlantic as an indicator of paleo-environment, Deep Sea Drilling Project Leg 71. Initial Reports of the Deep SeaDrilling Project 71, 739e787.
B�alc, R., Suciu-Krausz, E., Borbei, F., 2007. Biostratigraphy of the Cretaceous depositsin the Western Transylvanides from Ampoi Valley. Studia Universitatis Babes-Bolyai, Geologia 52, 37e43.
Bak, K., 2000. Biostratigraphy of deep-water agglutinated Foraminifera in ScagliaRossa-type deposits of the Pieniny Klippen Belt, Carpathians, Poland. In:Hart, M.B., Kaminski, M.A., Smart, C.W. (Eds.), Proceedings of the Fifth Inter-national Workshop on Agglutinated Foraminifera. Grzybowski FoundationSpecial Publication 7, 15e40.
Burnett, J.A., 1998. Upper Cretaceous. In: Bown, P.R. (Ed.), Calcareous nannofossilsbiostratigraphy. British Micropaleontology Society Series. Chapman and Hall,London, pp. 132e199.
Campbell, R.J., Howe, R.W., Rexilius, J.P., 2004. Middle CampanianelowermostMaastrichtian nannofossil and foraminiferal biostratigraphy of the north-western Australian margin. Cretaceous Research 25, 827e864.
Catanzariti, R., Ellero, A., Levi, N., Ottria, G., Pandolfi, L., 2007. Calcareous nannofossilbiostratigraphy of the Antola Unit succession (Northern Apennines, Italy): newage constraints for the Late Cretaceous Helminthoid Flysch. CretaceousResearch 28, 841e860.
Chapman, F., 1917. Monograph of the foraminifera and ostracoda of the GinginChalk. Western Australia Geological Survey Bulletin 72, 1e43.
Cetean, C.G., 2009. Cretaceous Foraminifera from the southern part of the EasternCarpathians, between Stoenesti and Cet�ateni. Paleocology and Biostratigraphy.Unpublished Ph.D. thesis. Babes – Bolyai University, Cluj Napoca, 214 pp.
Corliss, B.H., Chen, C., 1988. Morphotype patterns of Nowegian deep-sea benthicforaminifera and ecological implications. Geology 16, 716e719.
Cunha, A.S., Antunes, R.L., Burnett, J.A., 1997. Calcareous nannofossils and theSantonian/Campanian and Campanian/Maastrichtian boundaries on the Bra-zilian Continental Margin: historical overview and state of the art. CretaceousResearch 18, 823e832.
Erba, E., Watkins, D., Mutterlose, J., 1995. Campanian Dwarf calcareous nannofossilsfrom Wodejebato Guyot. Proceedings of the Ocean Drilling Program. ScientificResults 144, 141e156.
Erbacher, J., Thurow, J., Littke, R., 1996. Evolution patterns of radiolaria and organicmatter variations; a new approach to identify sea-level changes in Mid-Creta-ceous pelagic environments. Geology 24, 499e502.
Frenzel, P., 2000. Die benthischen Foraminiferen der Rügener Schreibkreide (unter-Maastricht, NE Deutchland). þ46 taf.. In: Neue Paläontologischen Abhand-lungen. CPress, Dresden, 3, 361 pp.
Gardin, S., Del Panta, F., Monechi, S., Pozzi, M., 2001. A Tethyan reference record forthe Campanian and Maastrichtian stages: the Bottacione section (central Italy);review of data and new calcareous nannofossils results). In: Odin, G.S. (Ed.), TheCampanian-Maastrichtian Stage Boundary, Characterisation at Tercis les Bains(France) and correlation with Europe and other continents. Developments inPalaeontology and Stratigraphy 19, 746e757.
Gasi�nski, M.D., Jugowiec, M., �Slaczka, A., 1999. Late Cretaceous foraminiferids andcalcareous nannoplankton from the Weglówka Marls (Subsilesian Unit, OuterCarpathians, Poland). Geologica Carpathica 50, 63e73.
Gawor-Biedowa, E., 1992. Campanian and Maastrichtian foraminifera from theLublin Upland, Eastern Poland. Palaeontologia Polonica 52, 1e187. þ 39 pls.
Geroch, S., Nowak, W., 1984. Proposal of Zonation for the late Tithonian - lateEocene, based upon arenaceous foraminifera from the outer Carpathians,Poland. April 11-15, 1983. In: Oertli, H. (Ed.), Benthos ’83; 2nd InternationalSymposium on Benthic Foraminifera Pau (France). Elf Aquitaine, Esso Rep andTotal cfp, Pau and Bourdeaux, pp. 225e239.
Gradstein, F.M., Kaminski, M.A., Agterberg, F.P., 1999. Biostratigraphy and paleo-ceanography of the Cretaceous seaway between Norway and Greenland. EarthScience Reviews 46, 27e98.
Gräfe, K., Wendler, J., 2003. Foraminifers and calcareous dinoflagellate cysts asproxies for deciphering sequence stratigraphy, sea-level change, and paleo-ceanography of Cenomanian-Turonian hemipelagic sediments in WesternEurope. Micropaleontological proxies for sea-level change and stratigraphicdiscontinuities. SEPM Special Publications. 75, 229e262.
Grigorescu, D., Melinte, M.C., 2002. The stratigraphy of the Upper Cretaceousmarine sediments from the NW Hateg Area (South Carpathians, Romania). ActaPalaeontologica Romaniae 3, 153e159.
Grzybowski, J., 1896. Otwornice czerwonych i1ów z Wadowic. RozprawyWydzia1u Matematyczno-Przyrodniczego. Akademia Umiejetno�sci w Krako-wie seryja 2 (30), 261e308.
Hampton, M.J., Bailey, H.W., Gallagher, L.T., Mortimore, R.N., Wood, C.J., 2007. Thebiostratigraphy of Seaford Head, Sussex, southern England; an internationalreference section for the basal boundaries for the Santonian and CampanianStages in chalk facies. Cretaceous Research 28, 46e60.
Holbourn, A., Kuhnt, W., Albani, A.L., Gomez, A.L.R., Herbin, J.P., 1999. Paleoenvir-onments and paleobiogeography of the Late Cretaceous Casamance transect(Senegal, NW Africa): distribution patterns of benthic foraminifera, organiccarbon and terrigenous flux. Neues Jahrbuch für Geologie und Paläontologie,Abhandlungen 212, 335e377.
Hancock, J.M., 1993. Transatlantic correlations in the Campanian- Maastrichtianstages by eustatic changes of sea-level. In: Hailwood, E.A., Kidd, R.B. (Eds.), HighResolution Stratigraphy. Geological Society of London, Special Publication 70,241e256.
Hu, X., Jansa, L., Wang, C., Sarti, M., Bak, K., Wagreich, M., Michalik, J., Sotak, J., 2005.Upper Cretaceous oceanic red beds (CORBs) in the Tethys: occurrences, lith-ofacies, age, and environments. Cretaceous Research 26, 3e20.
Ion, J., Szasz, L., 1994. Biostratigraphy of the Upper Cretaceous of Romania. Creta-ceous Research 15, 59e87.
Ion, J., Iva, M., Melinte, M.C., Florescu, C., 1998. Subsurface Santonian e Maas-trichtian deposits in North-Eastern South Dobrogea (Romania). RomanianJournal of Stratigraphy 77, 47e63.
Kaiho, K., 1991. Global changes of Paleogene aerobic/anaerobic benthic foraminiferaand deep-sea circulation. Palaeogeography Palaeoclimatology Palaeoecology83, 65e85.
Kaminski, M.A., Gradstein, F.M., 2005. Atlas of Paleogene cosmopolitan deep-water agglutinated foraminifera. Grzybowski Foundation Special Publication,10, 574þvii pp.
Kaminski, M.A., Bubík, M., Cetean, C.G., 2008. Rectoprotomarssonella n.gen. a newagglutinated foraminiferal genus from the Upper Cretaceous of the CarpathianFlysch. Micropaleontology 53, 517e521.
Kaminski, M.A., Silye, L., Kender, S., 2006. Miocene deep-water agglutinated fora-minifera from ODP Hole 909C: Implications for the paleoceanography of theFram Strait Area, Greenland Sea. Micropaleontology 51, 373e403.
Kender, S., Kaminski, M.A., Jones, R.W., 2008. Early to middle Miocene foraminiferafrom the deep-sea Congo Fan, offshore Angola. Micropaleontology 54, 477e568.
Kedzierski, M., 2008. Calcareous nannofossil and inoceramid biostratigraphies ofa Middle Turonian to Middle Coniacian section from the Opole Trough of SWPoland. Cretaceous Research 29, 451e467.
Koutsoukos, A.M., Hart, M.B., 1990. Cretaceous foraminiferal morphogroup distri-bution patterns, paleocommunities and trophic structures: a case of study fromthe Sergipe Basin, Brazil. Transactions of the Royal Society of Edinburgh. EarthSciences 81, 221e246.
Kuhnt, W., Kaminski, M.A., 1997. Cenomanian to lower Eocene deep-water agglu-tinated foraminifera from the Zumaya section, northern Spain. Annales Soci-etatis Geologorum Poloniae 67, 257e270.
Kuhnt, W., Moullade, M., Kaminski, M.A., 1998. Upper Cretaceous, K/T boundary,and Paleocene agglutinated foraminifers from Hole 959D (Côte d"Ivoire eGhana transform Margin). Proceedings of the Ocean Drilling Program. ScientificResults 159, 389e411.
Kuhnt, W., Geroch, S., Kaminski, M.A., Moullade, M., Neagu, T., 1992. Upper Creta-ceous Abyssal Claystones in the North Atlantic and the Western Tethys - CurrentStatus of Biostratigraphical Correlation using Agglutinated Foraminifers.Cretaceous Research 13, 467e478.
Li, L.Q., Keller, G., Adatte, T., Stinnesbeck, W., 2000. Late Cretaceous sea-levelchanges in Tunisia: a multi-disciplinary approach. Journal of the GeologicalSociety of London 157, 447e458.
Lüning, S., Marzouk, A.M., Morsi, A.M., Kuss, J., 1998. Sequence stratigraphy of theUpper Cretaceous of south-east Sinai, Egypt. Cretaceous Research 19, 153e196.
Melinte, M.C., Jipa, D., 2005. Campanian-Maastrichtian marine red beds in Romania:biostratigraphic and genetic significance. Cretaceous Research 26, 49e56.
Nagy, J., Gradstein, F.M., Kaminski, M.A., Holbourn, A.E.L., 1995. Late Jurassic to EarlyCretaceous foraminifera of Thakkhola, Nepal: Paleoenvironments and descrip-tion of new taxa. In: Kaminski, M.A., Geroch, S., Gasi�nski, M.A. (Eds.),Proceedings of the Fourth International Workshop on Agglutinated Forami-nifera. Grzybowski Foundation Special Publication 3, 181e209.
Neagu, T., 1970. Micropaleontological and stratigraphical study of the upperCretaceous deposits between the upper valleys of the Buzau and Riul NegruRivers (Eastern Carpathians). Memorii, Institutul Geologic 12, 7e109.
Neagu, T., 1989. Benthonic foraminiferal fauna from the white chalk of southernDobrogea, Romania. Part II: Agglutinated foraminifera. Revista Española deMicropaleontologia 3, 331e354.
Neagu, T., 1992. White chalk foraminiferal fauna from southern Dobrogea,Romania. III. Calcareous benthic foraminifera. Revista Española de Micro-paleontologia 24, 59e115.
Neagu, T., Platon, E., Dumitrescu, G., Selea, A., 1992. The biostratigraphical signifi-cance of agglutinated foraminifera in the Eastern Carpathians (Upper Creta-ceous). Analele Universitatii Bucuresti 15-16, 45e49.
Neuhuber, S., Wagreich, M., 2009. The Late Campanian Globotruncanita calcarataZone (Ultrahelvetics, Austria): implications from nannofossil stratigraphy,stable isotopes and geochemistry. In: Hart, M.B. (Ed.), 8th InternationalSymposium on the Cretaceous System, 6th - 12th September, 2009, Plymouth,Abstract Volume, p. 91.

C.G. Cetean et al. / Cretaceous Research 32 (2011) 575e590590
Nguyen, T.M.P., Petrizzo, M.R., Speijer, R.P., 2009. Experimental dissolution of a fossilforaminiferal assemblage (PaleoceneeEocene Thermal Maximum, Dababiya,Egypt): Implications for paleoenvironmental reconstructions. Marine Micro-paleontology 73, 241e258.
Nyong, E.E., Olsson, R.K., 1983. A paleoslope model of Campanian to lower Maas-trichtian foraminifera in the North Atlantic Basin and adjacent continentalmargin. Marine Micropaleontology 8, 437e477.
Ogg, J.G., Ogg, G., Gradstein, F.M., 2008. The Concise Geologic Time Scale. CambridgeUniversity Press, Cambridge, 184 pp.
Olszewska, B., 1997. Foraminiferal biostratigraphy of the Polish Outer Carpathians:a recordofbasingeohistory.AnnalesSocietatisGeologorumPoloniae67, 325e337.
Perch-Nielsen, K., 1985. Mesozoic calcareous nannofossils. In: Bolli, H.M.,Saunders, J.B., Perch-Nielsen, K. (Eds.), Plankton stratigraphy. CambridgeUniversity Press, Cambridge, pp. 329e426.
Peryt, D., 1988. Paleoecology of Middle and Late Cretaceous foraminifers from theLublin Upland (SE Poland). Revue de Paleobiologie 2, 311e321.
Premoli Silva, I., Sliter, W.V., 1995. Cretaceous planktonic biostratigraphy andevolutionary trends from Bottaccione section, Gubbio, Italy. PaleontographiaItalica 82, 1e89.
Robaszynski, F., Caron, M., 1995. Foraminifères planctoniques du Crètacé: com-mentaire de la zonation Europe-Méditerraneé. Bulletin de la Société Géologiquede France 166, 681e692.
Robaszynski, F., Pomerol, B., Masure, E., Bellier, J.P., Deconinck, J.F., 2005. Stratig-raphy and stage boundaries in reference sections of the Upper Cretaceous Chalkin the east of the Paris Basin: the ‘‘Craie 700’’ Provins boreholes. CretaceousResearch 26, 157e169.
Romero De, L.M., Truskowski, I.M., Bralower, T.J., Berge, J.A., Odreman, O.,Zachos, J.C., Galea-Alvarez, F.A., 2003. An Integrated Calcareous MicrofossilBiostratigraphic and Carbon-Isotope Stratigraphic Framework for the La LunaFormation, Western Venezuela. Palaios 18, 349e366.
S�andulescu, M., Stef�anescu, M., Butac, A., Patrut, I., Zaharescu, P., 1981. Genetical andstructural relationship between the Flysch and Molasse (the East Carpathiansmodel). Volume of the Carpatho-Balkan Geological Association. In: GuidebookSeries of the Geological Institute of Romania. 12th Congress, Bucharest. 19, 1e94.
Sissingh, W., 1977. Biostratigraphy of Cretaceous calcareous nannoplankton. Geo-logie en Mijnbouw 56, 37e65.
Sliter, W.V., 1977. Cretaceous benthic foraminifers from the western South AtlanticLeg 39, DSDP. Initial Reports of the Deep Sea Drilling Project 29, 657e697.
Stef�anescu, M., 1970. Structura geologica a regiunii dintre Valea Talea si ValeaIalomitei. Dari de seama ale Instutului de Geologie si Geofizica 57, 191e219.
Stef�anescu, M., 1995. Stratigraphy and structure of Cretaceous and Paleogene flyshdeposits between Prahova and Ialomita Valleys. Romanian Journal of Tectonicsand Regional Geology 76 suppl., 1e41.
�Svábenická, L., 1995. The stratigraphical correlation of the Campanian low- andhigh-latitude calcareous nannofossils in Southern Moravia (Western Carpa-thians). Geologica Carpathica 46, 297e302.
�Svábenická, L., Bubík, M., Krejcí, O., Stráník, Z., 1997. Stratigraphy of the Cretaceoussediments of the Magura group of nappes in Moravia (Czech Republic). Geo-logica Carpathica 48 (3), 179e191.
�Svábenická, L., Wagreich, M., Egger, H., 2002. Upper Cretaceous calcareous nanno-fossils assemblages at a transect from the northernTethys to the temperate realmin Central Europe. In: Michalik, J. (Ed.), Tethyan/Boreal Cretaceous Correlation.VEDA, Pub. House of the Slovak Academy of Sciences, Bratislava, pp. 187e212.
Varol, O., 1992. Taxonomic revision of the Polycyclolithaceae and its contribution toCretaceous biostratigraphy. Newsletters on Stratigraphy, Berlin-Stuttgart. 27,93e127.
Wagreich, M., 1992. Correlation of Late Cretaceous calcareous nannofossil zoneswith ammonite zones and planktonic Foraminifera: the Austrian Gosausections. Cretaceous Research 13, 505e516.
Wind, F.H., 1975. Tetralithus copulatus Deflandre (Coccolithophyceae) from TheIndian Ocean: a possible paleoecological indicator. Antarctic Journal of theUnited States 10, 265e268.
Yildiz, A., Özdemir, Z., 1999. Biostratigraphic and isotopic data on the ÇöreklikMember of the Hekimhan Formation (Campanian-Maastrichtian) of SE Turkeyand their palaeoenvironmental significance. Cretaceous Research 20, 107e117.
Zwaan van der, G.J., Jorissen, F.J., Stigter, H.C., 1990. The dept dependency ofplanktonic/benthic foraminiferal ratios: Constraints and applications. MarineGeology 95, 1e16.




















