
Instituto do Cérebro - UFRN
102
Universidade Federal do Rio Grande do Norte Programa de Pós-Graduação em Neurociências – Instituto do Cérebro DISSERTAÇÃO DE MESTRADO IMPLANTAÇÃO DA TÉCNICA DE REGISTRO DE CÉLULAS DE LUGAR UTILIZANDO MICRODRIVES DE TETRODOS MÓVEIS EM RATOS IMPLEMENTATION OF PLACE CELL RECORDINGS USING MOVABLE TETRODES IN RATS Estudante: Rafael Hugo de Andrade Pedrosa Orientador: Prof. Dr. Adriano Bretanha Lopes Tort Co-orientador: Prof. Dr. Hindiael Aeraf Belchior Natal / RN 2018
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Instituto do Cérebro - UFRN

Universidade Federal do Rio Grande do Norte
Programa de Pós-Graduação em Neurociências – Instituto do Cérebro
DISSERTAÇÃO DE MESTRADO
IMPLANTAÇÃO DA TÉCNICA DE REGISTRO DE CÉLULAS DE LUGAR
UTILIZANDO MICRODRIVES DE TETRODOS MÓVEIS EM RATOS
IMPLEMENTATION OF PLACE CELL RECORDINGS USING
MOVABLE TETRODES IN RATS
Estudante: Rafael Hugo de Andrade Pedrosa
Orientador: Prof. Dr. Adriano Bretanha Lopes Tort
Co-orientador: Prof. Dr. Hindiael Aeraf Belchior
Natal / RN
2018

IMPLANTAÇÃO DA TÉCNICA DE REGISTRO DE CÉLULAS DE LUGAR
UTILIZANDO MICRODRIVES DE TETRODOS MÓVEIS EM RATOS
por
Rafael Hugo de Andrade Pedrosa
Dissertação apresentada ao Programa de Pós-Graduação em Neurociências da Universidade Federal do Rio Grande do Norte como requisito parcial para a obtenção do título de Mestre em Neurociências.
Orientador: Prof. Dr. Adriano Bretanha Lopes Tort Co-orientador: Prof. Dr. Hindiael Aeraf Belchior
Natal / RN
2018
2

Universidade Federal do Rio Grande do Norte - UFRN
Sistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial
Árvore do Conhecimento - Instituto do Cérebro - ICE
Pedrosa, Rafael Hugo de Andrade.
Implantação da técnica de registro de células de lugar utilizando
microdrives de tetrodos móveis em ratos / Rafael Hugo de Andrade
Pedrosa. - Natal, 2018.
102f.: il.
Universidade Federal do Rio Grande do Norte. Instituto do
Cérebro. Programa de Pós-Graduação em Neurociências.
Orientador: Adriano Bretanha Lopes Tort.
Coorientador: Hindiael Aeraf Belchior.
1. Célula de lugar. 2. Microdrive. 3. Tetrodo. 4. Oscilações. 5.
Hipocampo. 6. Labirinto linear. I. Tort, Adriano Bretanha Lopes. II.
Belchior, Hindiael Aeraf. III. Título.
RN/UF/Biblioteca Setorial Árvore do Conhecimento, Instituto do
Cérebro. CDU 612.82
Elaborado por ISMAEL SOARES PEREIRA - CRB-15/741
3

Agradecimentos
Agradeço antes de tudo a minha mãe e meu pai por todo o apoio e
paciência ao longo desses 2 anos. Foram meus financiadores e budas em todo o
processo. Também agradeço a meu irmão pelo apoio quase filosófico que me fez
ser um humano menos ignorante.
Agradeço imensamente ao professor Adriano Tort por ter me aceitado e
confiado no meu trabalho. Antes mesmo de trabalharmos em conjunto, você já era
uma inspiração para mim. Hoje, mais que tudo, é um exemplo do cientista que
almejo ser. É uma referência que com certeza vou levar pro resto da minha vida.
Enfim, no mais, eu espero que tenha correspondido minimamente suas
expectativas.
Ao meu pai científico, Hindiael Belchior. Seu respeito, paciência e confiança
só me fizeram querer crescer. Além disso, nunca se inibiu de me ensinar qualquer
coisa de seu conhecimento, é uma virtude que poucos conseguem ter. Foi muito
gratificante trabalhar esse tempo desde a iniciação científica com você, levo seu
nome no meu currículo com muito orgulho.
A Richardson pela ajuda material para que eu tivesse condições de fazer os
experimentos, e Katarina por todas as dicas e conselhos ao longo do meu tempo
no ICe. Além disso, ambos, juntamente com Diego Laplagne se disponibilizaram
em me acompanhar durante o mestrado.
Ao meu Robin científico, Alan Michel, que sem sua proatividade e
cooperação esse trabalho não poderia ter sido feito.
Aos meus colegas e ex de laboratório Lockmann, Bryan, Robinho
maravilha, Vitor, Zé, Rodrigo, Arthur, Pavão, César e Izabela por me ajudar sempre
que precisei.
Aos iceanos Annie, João Patriota, Renzo, Ana Raquel, Daniel Filho, Bruna,
Ingrid, Daiane, Davi, Jéssica, Lyvia, Martin, Janine, Andressa, Carolina e que me
ajudaram de alguma forma ou foram efetivos na manutenção da minha saúde
mental. Também agradeço ao corpo de funcionários do Instituto do Cérebro.
A algumas companhias que facilitaram o trabalho: Sci-hub, Monster, Open
Ephys e Google.
4

“Hamlet” - William Shakespeare
5

Resumo
A formação de mapas espaciais depende do hipocampo e de estruturas
associadas. A atividade eletrofisiológica na região CA1 do hipocampo codifica
representações espaciais através de aumentos da taxa de disparos de neurônios
piramidais, conhecidos por células de lugar. O presente trabalho visou a
implementação da técnica de registro eletrofisiológico hipocampal através da
utilização de microdrives de múltiplos tetrodos móveis. Para isso, desenvolvemos
um protótipo de microdrive e fizemos cirurgias estereotáxicas em ratos para o
implante crônico bilateral. O novo protótipo de microdrive conteve 16 tetrodos
móveis e permitiu o posicionamento progressivo individual dos tetrodos na
camada piramidal da região CA1 do hipocampo dorsal. Após a recuperação
cirúrgica dos animais, realizamos o registro da atividade eletrofisiológica
extracelular da região CA1 enquanto ratos buscavam por recompensa de água
nas extremidades de um labirinto linear. As formas de onda dos potenciais de
ação registrados foram então classificadas como unidades neuronais individuais
por algoritmos de classificação semi-automáticos. Cada disparo de um dado
neurônio foi então associado à posição instantânea do rato no labirinto linear, e
assim detectamos campos receptivos das células de lugar. Dessa forma,
pudemos validar o protótipo de microdrive desenvolvido e, com isso, fornecer uma
base metodológica importante para futuros estudos almejando entender a
codificação espacial do ambiente e a formação de memórias espaciais.
Palavras chave: Células de lugar, Microdrive, Tetrodos, Hipocampo, Labirinto
linear.
6

Abstract
The formation of spatial maps depends on the hippocampus and associated
structures. Electrophysiological activity in the CA1 region of the hippocampus
encodes spatial representations through increases in the firing rate of pyramidal
neurons, known as place cells. The present work aimed the implementation of
hippocampal electrophysiological recording technique through the use of
microdrives of multiple movable tetrodes. For this, we developed a microdrive
prototype and performed stereotaxic surgeries in rats for bilateral chronic implant.
The new prototype microdrive contained 16 movable tetrodes and allowed the
individual progressive positioning of the tetrodes in the pyramidal layer of CA1
region of the dorsal hippocampus. After the surgical recovery of the animals, we
recorded the extracellular electrophysiological activity of the CA1 region while rats
searched for water reward at the ends of a linear track. The waveforms of recorded
action potentials were then classified as individual neural units by semi-automatic
classification algorithms. Each firing of a given neuron was then associated with
the instantaneous position of the rat on the linear track, and thus we detected
place fields of the place cells. Thus, we validated the microdrive prototype
developed and, thereby, provide an important methodological basis for future
studies aiming to understand the spatial encoding of the environment and the
formation of spatial memories.
Keywords: Place cell, Microdrive, Tetrodes, Hippocampus, Linear track.
7

Lista de abreviaturas
MRO - Movimento rápido dos olhos (Rapid eye movement).
SOL - Sono de ondas lentas (Slow wave sleep).
CA1 - Cornu Ammonis area 1.
CA2 - Cornu Ammonis area 2.
CA3 - Cornu Ammonis area 3.
CE - Córtex entorrinal.
GD - Giro denteado.
PRE - Sessão de sono anterior à tarefa espacial.
POS - Sessão de sono posterior à tarefa espacial.
PCL - Potencial de campo local.
V1 - Velocidade 1.
V2 - Velocidade 2.
V3 - Velocidade 3.
A1 - Aceleração 1.
A2 - Aceleração 2.
A3 - Aceleração 3.
IM - Índice de modulação.
CNC - Comando numérico computadorizado.
PSD - Power spectral density (densidade de potência espectral).
8

Lista de figuras
Figura 1 | Esquema das subdivisões hipocampais e suas projeções.
Figura 2 | Esquemático das principais projeções hipocampais.
Figura 3 | Ritmos oscilatórios do cérebro de roedores.
Figura 4 | Ilustração de 4 células de lugar que apresentam potenciais de ação
associados às posições em que o rato se encontra no espaço.
Figura 5 | Esquema ilustrativo da classificação de unidades neuronais a partir do
registro eletrofisiológico feito com tetrodo.
Figura 6 | Ilustração de um microdrive implantado em um rato.
Figura 7 | Comparação entre os diferentes tipos de microdrives que existem
atualmente.
Figura 8 | Processo de manufatura do tetrodo e microdrive.
Figura 9 | Microdrive de 8 tetrodos móveis.
Figura 10 | Microdrive de 16 tetrodos móveis.
Figura 11 | Testes de deslocamento e tensão na base de proteção.
Figura 12 | Reposicionamento dos tetrodos ao longo dos dias.
Figura 13 | Ilustração do protocolo de labirinto linear utilizado na tarefa
comportamental.
Figura 14 | Exemplo de classificação autônoma utilizando o Klustakwik.
Figura 15 | Exemplo de classificação autônoma utilizando o modelo de misturas
de gaussianas desenvolvido em nosso laboratório.
Figura 16 | Posições do animal no labirinto linear em diferentes sessões.
Figura 17 | Atividade neuronal registrada por um tetrodo.
Figura 18 | Seleção de candidatos a células de lugar.
Figura 19 | Seletividade espacial de neurônio de CA1 dorsal.
Figura 20 | Células de lugar ao longo da trajetória percorrida no labirinto linear.
Figura 21 | Neurônio de CA1 dorsal seletivo e acoplado em teta.
Figura 22 | Potencial de campo local de CA1 durante uma travessia no labirinto
linear.
Figura 23 | Potenciais de campo local de CA1 em alguns estados.
9

Figura 24 | Comodulograma de estados do animal.
Figura 25 | Lesão eletrolítica e histologia.
Figura 26 | Ilustração do protocolo de velocidades e acelerações da tarefa.
Figura 27 | Ilustração da tarefa de locomoção na esteira.
Figura 28 | Exemplo de oscilações delta (1-4 Hz) durante a exploração do animal
no labirinto linear.
Figura 29 | Análise espectral durante três protocolos de velocidades constantes
(20, 30 e 40 cm/s).
Figura 30 | Análise espectral durante a aceleração (2,5 cm/s²) no protocolo da
esteira.
Figura 31 | Análise espectral do PCL hipocampal em três protocolos de
aceleração (2, 2,5 e 3 cm/s²).
Figura 32 | Pico da frequência e amplitude normalizado em delta e teta através
dos 20 segundos binados.
Figura 33 | Frequência respiratória durante a corrida de velocidade constante na
esteira.
Figura 33 | Comodulograma da fase-amplitude e a energia por fase dos três
protocolos de velocidade constante.
10

Sumário
Resumo 5
Abstract 6
Lista de abreviaturas 7
Lista de figuras 8
Apresentação 11
I - Introdução geral 12 A formação hipocampal 13 Eletrofisiologia hipocampal 16 Microdrive de tetrodos móveis 23
II - Capítulo 1: Desenvolvimento dos microdrives de tetrodos móveis 27 1. Objetivos 28 2. Métodos 28 3. Resultados 31
III - Capítulo 2: Implantação da técnica de registro de células de lugar utilizando microdrives de tetrodos móveis em ratos 35
1. Objetivos 37 2. Métodos 38 3. Resultados 45 4. Conclusão 58
IV - Capítulo 3: Velocidade de corrida modula a oscilação delta no hipocampo de ratos 60
1. Objetivos 61 2. Métodos 62 3. Resultados 64 4. Conclusão 76
V. Discussão geral 77
VI. Animais implantados 81
VII. Referências 82
VIII. Anexos 96
11

Apresentação
O presente trabalho constitui uma síntese das atividades de pesquisa
desenvolvidas durante o curso de mestrado. A dissertação tem como tema central
a atividade eletrofisiológica do hipocampo de roedores durante a realização de
comportamentos exploratórios, protocolos de exercícios físicos, e ciclos de
sono/vigília. Para integrar os assuntos, o texto foi dividido em uma introdução
geral seguida de três capítulos que abordam principalmente (1) a implementação
de técnicas de registro neurofisiológico intracerebral utilizando microdrives de
tetrodos móveis; (2) a realização de registros da atividade de disparo de células de
lugar e do potencial de campo hipocampal durante a exploração de um labirinto
linear; e (3) a realização de registros eletrofisiológicos do hipocampo de roedores
durante corridas de velocidades crescentes e constantes em uma esteira elétrica
controlada por computador. Além disso, os capítulos apresentam as respectivas
análises dos sinais neurais, comportamentais, e interpretações dos resultados
obtidos de modo a trazer contribuições originais a cada um desses campos
individualmente.
12

I - Introdução geral _________________________________________________________________________________
13

A formação hipocampal
O hipocampo é uma estrutura cerebral localizada no lobo temporal medial
que exerce papel fundamental na codificação de memórias declarativas ou
explícitas (Scoville e Milner, 1954; Buzsáki e Moser, 2013), além de codificar
representações espaciais durante a navegação juntamente com o córtex entorrinal
(CE) (Hafting et al, 2005; Buzsáki e Moser, 2013). A anatomia hipocampal começou
a ser conhecida no final do século XIX a partir dos trabalhos iniciais de Camilo
Golgi e Santiago Ramón y Cajal. Eles desenvolveram uma técnica de coloração
por nitrato de prata capaz de marcar o meio intracelular dos neurônios, sendo
assim possível visualizar as unidades neuronais através do microscópio óptico
(Bentivoglio e Swanson, 2001). A partir dessa técnica, foi possível observar que o
hipocampo possui subdivisões com diferentes características e projeções
(Bentivoglio e Swanson, 2001; Insausti, 1997; Freund e Buzsáki, 1996).
A anatomia hipocampal hoje é separada em três subáreas chamadas de
Cornu Ammonis: CA1, CA2 e CA3 (Figura 1-A). Uma das principais projeções
neuronais para o hipocampo se dá pelo CE a partir de duas vias (Insausti, 1997).
(1) A via perfurante, que tem início na camada II do CE e que cruza o subiculum
até chegar no giro denteado (GD) e em CA3. O GD se projeta para o CA3 a partir
das fibras musgosas (mossy fibers), e CA3 se conecta com CA1 pelas colaterais
de Schaffer . (2) A via temporoamônica, que se origina na camada III do CE e 1
projeta diretamente para CA1 (Figura 1-B). Tanto a via perfurante (CE II - DG)
quanto a temporoamônica (CE III - CA1) enviam projeções unidirecionais para o
hipocampo, as camadas IV e V do CE por sua vez recebem projeções
hipocampais originadas em CA1 (Insausti, 1997; van Groen, 2003). No geral, a
atividade excitatória e inibitória do CE é composta de neurônios piramidais
glutamatérgicos e interneurônios GABAérgicos, respectivamente. Nas duas vias
de projeção para o hipocampo (perfurante e temporoamônica), os neurônios de
projeção do CE são excitatórios.
1 Essa conexão que envolve o CE até CA1 a partir do GD e CA3 é conhecida como via trisináptica (Amaral e Witter, 1989; Insausti, 1993). É a maior comunicação entre o hipocampo e o córtex.
14

Figura 1: Esquema das subdivisões hipocampais e suas projeções. (A) A imagem mostra uma
visão do hipocampo a partir de um corte coronal com coloração Nissl à esquerda e à direita um
esquemático com as subdivisões de CA1, CA2, CA3 e giro denteado. Em (B) uma ilustração das
projeções hipocampais. Modificado de Andersen et al, 2007.
A citoarquitetura do hipocampo é formada por uma complexa rede de
neurônios piramidais e diferentes classes de interneurônios. Em CA1, CA2, CA3 e
GD é possível observar uma alta densidade neuronal, composta de um
agrupamento de neurônios piramidais. Modulando a atividade dos neurônios
piramidais, interneurônios GABAérgicos projetam seus axônios em diferentes
níveis do corpo celular piramidal (Freund e Buzsáki, 1996; Klausberger e Somogyi,
2008). Os interneurônios são classificados de acordo com suas características de
15

projeção sobre o neurônio piramidal, sendo eles: axo-axônicas (chandelier cells) , 2
basket cells , oriens lacunosum-moleculare (OLM) cells e bistratified cells . 3 4 5
A organização morfológica dos interneurônios e das células piramidais
separa as regiões de CA1, CA2, CA3 e GD em 5 diferentes lâminas ou estratos:
stratum oriens, stratum pyramidale (onde se localiza o corpo celular dos neurônios
piramidais), stratum lucidum, stratum radiatum e stratum lacunosum-moleculare
(Figura 2). Exclusivamente em CA3 o perfil laminar se dá pelas 5 camadas, sendo
os stratum lucidum, pyramidale e oriens alvos de projeções do DG. CA2 por sua
vez é composto de apenas 4 camadas, todas com exceção do stratum lucidum
(Amaral e Witter, 1989). Sua principal via de entrada se dá pelo stratum
lacunosum-moleculare com inputs do CE II, e oriens que recebe projeções das
colaterais associativas. Similar à CA2, CA1 apresenta as mesmas 4 camadas,
onde sua principal vía de entrada é o stratum lacunosum-moleculare que recebe
projeção do CE III. CA1 tem como principal camada de saída o stratum oriens que
tem projeções de feedback para as camadas V e VI do EC. Além disso, CA1
recebe inputs de CA3 no stratum radiatum pelas colaterais de Schaffer (Amaral et
al, 2006).
2 As chandelier cells são interneurônios que possuem seu corpo celular junto ao do neurônio piramidal no stratum pyramidale e têm conexão axo-axonal (Freund e Buzsáki, 1996). 3 Interneurônio de alta taxa de disparo que inibem a região peri-somática dos neurônios piramidais (Freund e Buzsáki, 1996). 4 As células OLM regulam a atividade dos neurônios piramidais e possuem corpo no stratum oriens com projeções axonais no stratum lacunosum-moleculare. 5 Essas células apresentam seus somas no stratum pyramidale e atividade inibitória nos dendritos das células piramidais no stratum oriens (Klausberger e Somogyi, 2008; Wheeler et al, 2015).
16

Figura 2: Esquemático das principais projeções hipocampais. Na figura pode-se ver as principais
projeções de entrada e saída do hipocampo nas diferentes camadas celulares. O CE e GD
fornecem projeções de entrada (em verde musgo e rosa). CA1 gera projeções de saída (verde).
Eletrofisiologia hipocampal
As trocas iônicas entre os meios intracelular e extracelular para que ocorra
o potencial de ação neuronal desencadeiam uma diferença de potencial capaz de
ser identificada por meio de eletrodos, como foi descrito por Luigi Galvani no
século XVIII (Verkhratsky et al, 2006). O conjunto dessas ativações neuronais gera
um sinal elétrico extracelular chamado de potencial de campo local (PCL) (da
Silva, 2009). O PCL representa uma propagação espacial em torno do eletrodo de
aproximadamente 400 µm na horizontal e 1 mm na vertical . Quando medido no 6
hipocampo, o PCL apresenta diferentes bandas de frequências que se relacionam
6 Essa dispersão é discutida atualmente por influência da condição de volume local x global. Trabalhos atualmente descrevem sua influência como sendo de uma área entorno de 6 mm (Kajikawa e Schroeder, 2011).
17

com diferentes estados comportamentais (Buzsaki, 2006; O'Keefe e Reece, 1993;
Csicsvari et al, 2003; Senior et al, 2008; Sirota e Buzsaki, 2008). As principais
bandas podem ser resumidamente separadas em: delta (1-4 Hz), teta (5-10 Hz),
beta (10-30 Hz ), low gamma (LG) (30-50 Hz) e high gamma (HG) (50-80 Hz) (Figura
3). Em estudos mais recentes, foi possível observar que esses ritmos oscilatórios
podem coexistir e estar acoplados. É possível encontrar uma relação de
fase-amplitude-frequência entre dois diferentes ritmos, em geral esse
acoplamento se dá entre uma oscilação de alta frequência e uma de baixa
frequência (Scheffer-Teixeira e Tort, 2016; Scheffer-Teixeira e Tort, 2017; Colgin et
al, 2009).
Figura 3: Ritmos oscilatórios do cérebro de roedores. (A) Espectro de potência de diferentes
ritmos hipocampais durante o sono. (B) Diferentes classes de ritmos oscilatórios do cérebro por
bandas de frequências. Modificado de Buzsáki e Draguhn, 2004.
18

No geral, o estado global do cérebro é definido como “alerta” e “quieto” a
partir das oscilações predominantes no hipocampo (Battaglia et al, 2011; Kirov et
al, 2009): oscilações delta (1-4 Hz) caracterizam o estado quieto, e as oscilações
teta (5-10 Hz) o estado alerta. Teta ocorre fortemente associado a oscilações
gama (30-80 Hz), que estão relacionadas a processos cognitivos (Sheffzuk et al,
2011; Tort et al , 2009; Kirov et al, 2009). Delta, teta e gama são atualmente os
ritmos hipocampais mais estudados, e podem estar envolvidos no processo de
transmissão de informações entre as áreas cerebrais. Por isso, introduziremos
abaixo seus principais aspectos individuais:
Delta. A oscilação delta é descrita como uma oscilação lenta entre 1 e 4
Hz, predominante no hipocampo de roedores durante estados de anestesia geral,
imobilidade na vigília e em períodos de sono descrito como sono de ondas lentas
(do inglês, slow wave sleep ou “SOL”). O ritmo delta reflete períodos tônicos de
despolarização e hiperpolarização cortical referidos como estados “up” e “down”
(Steriade et al, 1993). O estado up é descrito como o aumento do balanço entre a
atividade excitatória e a inibitória dos potenciais pós-sinápticos na comunicação
do hipocampo com o córtex, que gera um aumento do potencial de membrana
nessas áreas (Haider et al, 2006; Headley e Paré, 2017). Além disso, durante as
oscilações delta, um evento de alta frequência chamado de complexo sharp wave
ripple (SWR) (150-250 Hz) ocorre periodicamente durante o início do estado up
(Isomura et al, 2006; Battaglia et al, 2004). O SWR é caracterizado pela
sincronização de neurônios piramidais de CA1 com interneurônios de CA3, e
exerce um papel fundamental no processo de consolidação de memória (Ylinen et
al, 1995; Girardeau e Zugaro, 2011).
Teta. A oscilação teta no hipocampo dorsal de roedores pode variar entre 5
e 10 hz, e é presente tanto em atividades voluntárias, como andar, correr, pular, e
nadar dentre outras, quanto atividades involuntárias como durante o alerta e o
sono de movimento rápido dos olhos (MRO) (Vanderwolf, 1969; Buzsáki, 2002).
Além disso, foi mostrado que a oscilação teta apresenta funções cognitivas,
como tomada de decisão, aprendizagem, memória espacial e associação
contextual (Belchior et al, 2014; O’Keefe e Recce, 1993; Tort et al, 2009;
Benchenane et al, 2010). Sua atividade é dependente da banda medial/diagonal
19

do septo, que possui projeções GABAérgicas, colinérgicas e glutamatérgicas para
o hipocampo (Buzsáki, 2002; Lee et al, 1994). Trabalhos anteriores mostraram que
lesões no septo e via septohipocampal abolem a atividade do ritmo teta tanto em
GD quanto em CA1 (Sainbury e Bland, 1981; Andersen et al, 1979). Além disso,
Quilichini et al, 2010 mostraram que lesões no CE também influenciam a atividade
do ritmo teta em todo o hipocampo, o que indica uma função do CE como um
modulador externo do teta hipocampal.
A atividade de teta ao longo do tempo foi extremamente explorada por ser
o principal ritmo durante o estado alerta do cérebro de roedores. Dentre esses
estudos, McFarland et al em 1975 observaram a correlação desse ritmo com a
atividade locomotora de roedores. Ao mostrar que há um aumento da amplitude e
da frequência de teta durante a locomoção, seu trabalho se tornou precursor de
uma série de estudos seguintes que relaciona esse ritmo à dedicação física do
indivíduo. Em 1998, Slawińska e Kasicki mostraram que o aumento da frequência
teta durante a locomoção tem um fator motivacional inserido que modifica sua
dinâmica a partir de influências emocionais. Mais recentemente, Kuo e
colaboradores (2011) descobriram que essa dinâmica de atividade de teta é
variável com o tempo. Eles mostraram que no início da corrida há um aumento de
amplitude e de frequência dessa banda, mas que com o passar do tempo a
frequência retorna a um nível basal. Em outro trabalho publicado em 2014, Kuo e
Li também mostraram que há diferença na ritmicidade de teta entre corridas
voluntárias e involuntárias (Li et al, 2014). Os resultados mostram que nas 7
corridas involuntárias os animais apresentaram uma maior potência na frequência
de teta quando comparado com as corridas voluntárias. Nas corridas involuntárias
também foram encontrados aumentos nos batimentos cardíacos. Os autores
sugerem que interações emocionais e sensoriais durante a corrida podem estar
relacionadas com essas diferenças de atividade cerebral e cardíaca.
Em suma, a oscilação teta é fundamental para a aprendizagem e
consolidação de memória, além de estar envolvida nas principais ações do
indivíduo durante seu estado de vigília. Sua atividade modula diversos processos
7 Experimentos executados em uma roda de correr e em uma esteira controlada por computador.
20

cognitivos, dentre eles a formação de mapas espaciais (Buzsáki e Moser, 2013),
que será discutido mais à frente.
Gama. As oscilações gama possuem frequência entre 30 e 80 Hz. Sua
atividade ocorre ao longo do córtex, hipocampo e estriado tanto durante o sono
MRO quanto na vigília. A geração do ritmo gama é bastante estudada, sendo
atualmente dividida em dois modelos: (1) Modelo baseado na comunicação
inibitória de interneurônios, que se dá através de drives excitatórios que agem em
uma rede de interneurônios inibitórios. Quando esses drives atingem um
determinado nível de atividade, os interneurônios acabam silenciando
temporariamente sua própria rede, causando uma sincronia de disparos quando
saem da inibição. Essa atividade se torna cíclica e contínua (Wang e Buzsáki,
1996). (2) Em outro modelo, os neurônios piramidais ativam os interneurônios que
causam um feedback de inibição. Com o diminuir dessa inibição, as células
piramidais voltam a disparar, excitando novamente as pequenas redes de
interneurônios, que por sua vez voltam a prover feedback de inibição (Borges e
Kopell, 2003). Essa atividade também se torna cíclica e gera uma oscilação na
banda gama.
Atualmente, o ritmo gama observado em CA1 é dividido em dois
componentes que apresentam frequência, origem e funções diferentes. O low
gamma (LG) (30-50 Hz) é oriundo de CA3 e apresenta relação com a codificação
de memórias hipocampo-dependentes (Colgin e Moser, 2010). Enquanto o high
gamma (HG) (50-80 Hz) possui origem no CE e participa de processos de
associações espaciais (Colgin e Moser, 2010; Quilichini et al, 2010). Trabalhos
mostram que há um maior acoplamento entre teta e gama em tarefas que exigem
demandas associativa e cognitiva (Igarashi et al, 2014; Tort et al, 2009; Bott et al,
2016).
Dessa forma, durante a navegação espacial no ambiente, o hipocampo
possui atividade oscilatória de teta (5-10 Hz) e gama (30-80 Hz). Essas oscilações
modulam a atividade de disparos de potenciais de ação de diversas classes de
neurônios, dentre elas os neurônios piramidais (células principais). Foi descoberto
que alguns desses neurônios piramidais modulados por teta e gama apresentam
21

preferência de disparo em um lugar específico no ambiente (O’Keefe e Dostrovsky,
1971) e participam portanto da codificação espacial, tópico da próxima seção.
Células de Lugar
Em 1971, O’Keefe e Dostrovsky observaram que neurônios piramidais no
hipocampo de ratos disparam potenciais de ação em locais específicos do
ambiente (O’Keefe e Dostrovsky, 1971); estes neurônios são hoje em dia
chamados de “células de lugar” (do inglês, place cells). Essa descoberta deu
suporte à teoria do Mapas Cognitivos, a qual postula que no hipocampo são
integradas informações sensoriais recebidas de áreas corticais primárias, criando
assim uma representação espacial do ambiente (O’Keefe e Nadel, 1978). Por outro
lado, desde a descrição do caso do paciente H.M., já havia sido proposto que o
hipocampo e estruturas parahipocampais desempenham uma função primordial
na formação de novas memórias declarativas (Scoville e Milner, 1954). Atualmente,
de acordo com essa teoria, as células de lugar forneceriam o substrato neural
fundamental para a construção de representações dos contextos envolvidos na
memória episódica (Eichenbaum, 2000b; Wood et al., 2000). Contudo, a
interligação entre essas duas teorias ainda permanece por ser esclarecida.
Mais recentemente, em um labirinto linear com recompensas de água nas
extremidades, foi possível observar o disparo sequencial de populações de células
de lugar em função da posição e da direção do animal no espaço (Figura 4)
(Wilson e McNaughton, 1994; Jensen e Lisman, 2005; Dragoi e Buzsaki, 2006;
Foster and Wilson, 2006). Foi visto também que a sequência de células de lugar
ativas ao longo do percurso era frequentemente reativada durante os momentos
de vigília quieta e durante os episódios de sono pós-experimento (Louie e Wilson,
2001; Foster e Wilson, 2006). Essas reativações acontecem acompanhadas de
sharp-wave/ripples (150-250 Hz) (Buzsáki, 1998; Battaglia et al., 2011; Carr et al.,
2011). Uma série de estudos demonstrou que a interrupção das sharp-wave ripple
afeta o processo de aprendizagem e a consolidação de novas memórias
(Girardeau et al., 2009; Ego-Stengel e Wilson, 2010; Girardeau e Zugaro, 2011).
22

Figura 4: Ilustração de 4 células de lugar que apresentam potenciais de ação associados às
posições em que o rato se encontra no espaço (o exemplo mostra a plataforma de um labirinto
linear). À direita tem-se a representação espacial do campo de lugar em escala de cor. Retirado de
Nakazawa e colaboradores (2004).
Os desenvolvimentos técnicos e a evolução das ferramentas
computacionais para análise de dados permitiram avanços na compreensão da
atividade eletrofisiológica do hipocampo. Por exemplo, (1) foi observado que a
ativação das células de lugar está acoplada a determinadas frequências do
potencial de campo local (PCL), como as oscilações teta (5-10 Hz) e gama (30-80
Hz) (O'Keefe e Reece, 1993; Csicsvari et al., 2003; Senior et al., 2008).
Observou-se ainda que (2) as oscilações teta e gama acontecem no hipocampo de
roedores não somente durante os estados de alerta e de exploração ativa da
vigília, mas também surgem durante o sono MRO (Buzsaki, 2002;
Scheffer-Teixeira et al., 2012). Além disso, as análises da relação entre a fase das
oscilações e os disparos de potenciais de ação mostraram que (3) os disparos das
células de lugar ao longo do seu campo receptivo espacial variam em função da
fase do ciclo da oscilação teta subjacente (O'Keefe e Reece, 1993). Essa variação
de fases acontece de maneira ordenada, dando origem a uma precessão das
fases da onda onde os disparos acontecem à medida que o animal atravessa o
campo espacial dessa célula (Jensen e Lisman, 2000; O'Keefe e Reece, 1993).
Além da relação com as oscilações teta, acredita-se que (4) a coordenação
23

temporal exercida pelo PCL sobre os disparos de grandes populações de
neurônios permita a formação de assembleias neuronais (Harris et al., 2003),
descrita por Donald Hebb em 1949 como agrupamentos de neurônios que se
ativariam juntos para codificar uma representação mental (Hebb, 1949).
Mais recentemente, Hafting e colaboradores (2005) registraram no córtex
entorrinal medial, uma região que envia projeções ao hipocampo, e descobriram
neurônios que disparam potenciais de ação periodicamente no espaço, com
campos espaciais em formato de grades, chamando-lhes assim de células de
grade (do inglês, grid cells). Esse achado, acompanhado da descoberta das
células de lugar ocorrida no anos 70, fez com que John O’Keefe, Edvard Moser e
May-Brit Moser fossem agraciados com o Prêmio Nobel de Medicina e Fisiologia
em 2014 (Moser, Moser e O’Keefe, 2014).
Microdrive de tetrodos móveis
Uma dificuldade técnica para a obtenção de registros extracelulares da
atividade de disparos de unidades neuronais individuais no hipocampo de
roedores decorre do fato de as camadas celulares CA1, CA2 e CA3 apresentarem
os corpos celulares altamente compactadas em densas camadas. Para sobrepor
essa dificuldade, foram desenvolvidas duas técnicas para discriminar potenciais
de ação provenientes de diferentes neurônios a partir de registros extracelulares.
Inicialmente, foram usados estereotrodos (McNaughton et al., 1983), que se
caracterizam por dois eletrodos de mesmo material e espessura, agrupados
paralelamente e girados sobre o próprio eixo (O'Keefe e Reece, 1993; Wilson e
McNaughton, 1993). Posteriormente, foram usados tetrodos, caracterizados como
quatro eletrodos agrupados da mesma forma (Figura 5-A). Esses eletrodos
registram os mesmos disparos provenientes de uma pequena população neuronal,
e análises computacionais posteriores permitem então a caracterização de
unidades neurais individuais de acordo com os parâmetros do formato de onda
dos potenciais de ação registrados simultaneamente nos diferentes canais do
tetrodo (Figura 5-B). Esses desenvolvimentos metodológicos tornaram o registro
24

utilizando tetrodos a principal técnica eletrofisiológica para o estudo de
populações de neurônios individuais no hipocampo de roedores, e assim
permitiram a caracterização detalhada da atividade de disparos das células de
lugar.
Figura 5: Esquema ilustrativo da classificação de unidades neuronais a partir do registro
eletrofisiológico feito com tetrodo. A figura à esquerda (A) representa um esquemático de um
tetrodo localizado nas proximidades de 3 neurônios representados pelas cores vermelho, verde e
azul. A figura à direita (B) mostra um exemplo do processo de registro e classificação neuronal. O
sinal eletrofisiológico bruto é filtrado em baixas (1-1000 Hz) e altas (500-6000 Hz) frequências para
se obter o potencial de campo local e os disparos neuronais, respectivamente. A classificação dos
disparos é feita a partir da discrepância do formato de onda do disparo de cada neurônio,
mostrado na figura pelas cores seguindo os neurônios da figura (A). Figura ilustrada por Rodrigo
Pavão e modificada por Rafael Pedrosa.
Junto das técnicas de estereotrodo e tetrodo, foi desenvolvido também um
sistema para a movimentação progressiva dos eletrodos após a cirurgia de
implante, o microdrive. Esse aparelho permite a execução de ajustes finos no
posicionamento de modo que a ponta dos eletrodos esteja o mais próximo
possível dos neurônios registrados. Dependendo da necessidade, os eletrodos
podem se mover juntos ou individualmente, múltiplas vezes, e até mesmo com os
animais acordados. O uso dos microdrives também permite que o
experimentalista atinja o alvo no cérebro em diferentes camadas a partir de um
sistema móvel de parafuso que controla a altura do eletrodo no cérebro após a
cirurgia. Um exemplo de microdrive é mostrado na Figura 6.
25

Figura 6: Ilustração de um microdrive implantado em um rato. Em (A) e (B) temos um exemplo de
um animal implantado com um microdrive revestido de um papel laminado. (C) mostra o microdrive
pós implantado. Figura retirada de J. Yamamoto e Matthew Wilson (2008).
Os primeiros microdrives eram construídos usando componentes de metal
e plástico líquido, mas hoje em dia sua principal base é feita de componentes
plásticos produzidos por impressoras 3D. A principal vantagem desses
componentes plásticos é o baixo peso, alta precisão, baixo custo e a velocidade
da produção, consumindo muito menos tempo que o modelo anterior. Além disso,
as versões mais recentes se aprimoraram também em utilizar um maior número de
canais para registro, juntamente com técnicas de manufatura mais rápida.
Atualmente, o foco de melhorias nessa técnica ainda está voltado para o
aumento de número de tetrodos para controle individual juntamente com redução
do peso. Dos microdrives disponíveis no mercado atualmente, o flexDrive® é o
que apresenta uma melhor versão da relação peso/tetrodos, sendo o mais
26

utilizado nos laboratórios que trabalham com registro de unidades neuronais em
larga escala (Figura 7).
Figura 7: Comparação entre os diferentes tipos de microdrives que existem atualmente. O eixo X é
o número individual de agrupamento de eletrodos que se podem mover individualmente. Em Y o
peso de cada microdrive. A linha cinza é o peso limite que um camundongo pode sustentar.
Retirado de Voigts et al (2013).
O assunto abordado nesse tópico da introdução será necessário para o
entendimento do capítulo 1 desta dissertação. A saber, o trabalho desenvolvido
visou a implementação da técnica de registro eletrofisiológico através da utilização
de microdrives de múltiplos tetrodos móveis, uma tecnologia ainda incipiente no
Instituto do Cérebro da UFRN e no Brasil. Conforme visto nos outros tópicos
abordados nesta introdução, o desenvolvimento de tal tecnologia permitirá a
realização de pesquisas sobre importantes temas atuais da eletrofisiologia
hipocampal, como o estudo da atividade das células de lugar e de suas relações
com as oscilações neuronais.
27

II - Capítulo 1: Desenvolvimento de microdrives de
tetrodos móveis
_________________________________________________________________________________
28

A técnica de microdrive de tetrodos móveis, além de ser a mais eficaz
quanto ao quesito de número de neurônios registrados, vem sendo extremamente
utilizada em diversos laboratórios ao redor do mundo. Apesar de haver avanços
em sua eficiência, esse sistema ainda é dependente de peças e ferramentas
específicas produzidas por empresas ou grupos que monopolizam o mercado, e,
com isso, o valor individual de compra de cada microdrive ainda é inviável para
muitos laboratórios de países em desenvolvimento. Pensando nisso, e com
interesse em implementar a técnica no Brasil, desenvolvemos dois diferentes
protótipos de microdrives utilizando uma impressora 3D e uma máquina de
comando numérico computadorizado (CNC).
1. Objetivos
1.1 Objetivo geral
Desenvolver um novo protótipo de microdrives de tetrodos móveis e
implantar sua técnica no Laboratório de Neurofisiologia Computacional do
Instituto do Cérebro da UFRN.
1.2 Objetivos Específicos
1 – Desenvolver um protótipo de microdrive móvel para implantes em ratos.
2 – Utilizar o método de montagem em blocos para facilitar sua manufatura.
2. Métodos
2.1 Microdrive de tetrodos móveis
Desenvolvemos um protótipo próprio de microdrive inspirado em modelos
já existentes na literatura. Foram testadas tanto estratégias de confecção dos
protótipos via CNC quanto através de impressão 3D, e utilizados polylactic acid
(PLA) e acrylonitrile butadiene styrene (ABS). Foram empregados conectores
29

Omnetics® para a conexão do microdrive ao sistema de registro. Além disso,
parafusos e cânulas mais baratos foram adaptados aos microdrives
desenvolvidos.
2.2 Tetrodos
Os tetrodos foram formados por quatro microeletrodos de nicromo ou
tungstênio de 12,5 µm de diâmetro, revestidos de Teflón® (Politetrafluoretileno), e
emaranhados a partir de giros em relação ao seu eixo principal. Antes da cirurgia
de implante, os microeletrodos tiveram sua impedância corrigida para
aproximadamente 70 KΩ a 100 KΩ a partir da eletrólise em solução de ouro
contendo nano tubos de carbono, e de acordo com Redish e colaboradores
(2009).
2.3 Manufatura do microdrive de tetrodos móveis
Para a manufatura do microdrive é necessário imprimir e construir as partes
(usando a impressora 3D): 1) Support board - placa central em que o microdrive
será montado; 2) Protection base – estrutura tipo um cone que prevê sua
proteção; 3) Guide platforms – componentes para guiar o tetrodo, controlados por
parafusos; 4) Guiding base – peça encaixada embaixo da protection base e usada
para a distribuição espacial dos tetrodos no eixo XY; 5) Microdrive cover - tampa
para a proteção do microdrive quando não estiver em uso. A Figura 11 mostra um
passo a passo do processo de manufatura. As peças podem ser encontradas para
impressão no link: https://github.com/tortlab/Open-Source-Microdrive.
30

31

Figura 8: Processo de manufatura do tetrodo e microdrive. (A) Para a manufatura do tetrodo, cortamos dois pedaços de fio revestido de 20 cm de comprimento (níquel-cromo ou tungstênio, 12,5 µm diâmetro), dobramo-os juntos em forma de U e prendemos o laço na presilha pelas 4 pontas. B) & C) Abaixamos o suporte para suavemente prender a presilha no imã magnético para começar o protocolo de giro. D) depois do giro, usamos a heatgun para grudar os fios. Em seguida, cortamos o tetrodo perto da presilha e removemos cuidadosamente. E) Materiais usado para a confecção dos tetrodos/microdrive com o sistema de aquisição. Soldamos o conector Omnetics na printed circuit board (PCB) usando solda em pasta. Parafusos e porcas foram usados para prender a PCB na Support board. O terra (fio de prata ou aço inoxidável) também foi soldado na PCB e preso aos parafusos na ponta. F) Partes 3D do Microdrive: Microdrive cover, Support board, Guide platforms, Protection base e Guiding base. Cada uma das 16 guiding units é composta de barras de 19Ga, parafuso, porcas e uma cânula de 22Ga. Para cada guiding unit, tem-se 3 furos radialmente alinhados com a Support board para a barra, parafuso e cânula de 22Ga. Um pequeno tubo de sílica (que vai carregar o tetrodo) foi inserido dentro da cânula de 22Ga, e colado nele junto da Guide platform. A Guide platform desce pelo apoio da barra de 19Ga a partir do giro do parafuso, onde carrega a cânula de 22Ga e o pequeno tubo de sílica para a Support board. Dentro da Support board, o pequeno tubo de sílica desliza dentro de um tubo de sílica maior de 27G. A Support board encaixa na Protection base e essa por sua vez na Guiding base. G) Um passo adicional no conjunto do microdrive é o ancoramento das guiding units e das sílicas maiores de 27Ga (azul) dentro da Support board. Note que o pequeno tubo de sílica (verde) vai dentro do tubo da sílica de 27Ga (azul). Em outra etapa, as guiding units são baixadas a partir do giro dos parafusos, sendo ambas as sílicas cortadas na parte superior da Guiding base. Depois, eles são movidos de volta para cima e os tetrodos são carregados e colados nos pequenos tubos de sílica. H) Vista superior do microdrive sem as guiding units. A Support board possui 3 filas circulares de furos, através das quais são inseridas a cânula de 22Ga (fileira interna), o parafuso (fileira do meio, porca marcando em verde) e a barra de 19Ga (fileira externa). I) Vista frontal/lateral da Protection base, Guiding base (fixada dentro por resina acrílica) e o tubo de sílica de 27Ga ainda não cortada. Figura retirada de capítulo submetido.
3. Resultados
3.1 Microdrives
3.1.1 Microdrive de 8 tetrodos móveis
O primeiro protótipo de microdrive desenvolvido promove o movimento
simultâneo de 8 tetrodos que convergem na mesma região. O microdrive (5,0 cm
de altura; 1,4 cm de raio) apresenta uma parte móvel associada a 8 tetrodos
guiados por um parafuso de bronze com um passo de 0,28 mm a cada volta
(Figura 9). Uma placa de circuito impresso de fenolite foi desenvolvida para
conexão dos tetrodos e fio terra com o conector Omnetics®. As placas base
foram feitas com cortes precisos em uma CNC de modelo 3020. Os tetrodos se
encontram presos à sílica guia que se movimenta proporcionalmente ao passo
dado pelo parafuso guia. Em paralelo, as sílicas base estão fixadas à cânula presa
à placa base para estabilizar e direcionar a sílica guia com os tetrodos.
32

Figura 9: Microdrive de 8 tetrodos móveis. Na figura vemos o esquemático do microdrive, sendo a
primeira figura à esquerda, a representação de perspectiva do protótipo, seguido da vista lateral e
por último da vista superior. São indicadas por letras as peças: a - tetrodos; b - parafuso guia; c -
Base de proteção; d - Conector Omnetics®; e - Sílica guia; f - Sílica base; g - Placas base.
Ao todo 5 ratos foram implantados com esse modelo de microdrive, dos
quais foram obtidos registros de apenas 3. Os tetrodos foram progressivamente
aprofundados até a região do hipocampo. Os registros foram iniciados um dia
após observadas características eletrofisiológicas típicas do hipocampo, como a
ocorrência de SWR, oscilações teta de alta amplitude, rajadas (“bursts”) de
disparos de neurônios (típicos de neurônios piramidais). Ao fim de cada sessão de
registro, os tetrodos foram levemente aprofundados para o registro de novos
neurônios na sessão seguinte com intervalo mínimo de 20 horas entre sessões.
3.1.2 Microdrive de 16 tetrodos móveis
Com o objetivo de ampliar o número de tetrodos por implante, de se obter
movimentação individualizada, posicionamento preciso dos tetrodos de registro, e
de aumentar a quantidade de neurônios registrados, um segundo protótipo de
microdrive foi desenvolvido para o implante e registro bilateral da atividade
eletrofisiológica dos hipocampi de ratos. O microdrive (3,6 cm de altura; 2,3 cm de
raio e 14 gramas de peso) possui um sistema móvel de 16 tetrodos guiados
individualmente por 16 parafusos de bronze com um passo de 0,32 mm a cada
volta (Figura 10). Uma placa de circuito impresso de fenolite (modelo disponível
online pelo projeto Open ephys) foi utilizada para conexão dos 64 canais trilhados
33

com conectores Omnetics® (2 conectores com 32 canais cada). As peças
plásticas para a montagem do corpo do microdrive foram impressas em PLA por
uma impressora 3D com bico extrusor de 0,4 mm de precisão aquecido a 220 °C,
a partir de um modelo desenvolvido 3D por nós.
No total, 5 ratos foram implantados com o microdrive de 16 tetrodos
móveis, dos quais foram obtidos registros de apenas 3 animais. Por enquanto,
nem todos os registros colhidos foram analisados (ver na seção “VI - Animais
implantados”).
Figura 10: Microdrive de 16 tetrodos móveis. Na figura vemos um protótipo 3D do microdrive,
sendo apresentado na sequência as vistas de perspectiva, frontal e superior. As peças são
indicadas por letras: a - Tetrodos; b - Base de proteção; c - Plataforma guia; d - Conector
Omnetics®; e - Base guia.
3.1.2.1 Testes de resistência da Base de proteção
Testes de deslocamento e tensão (von Mises Stress) foram feitos para o
desenvolvimento de uma melhor estrutura de proteção do Microdrive (Figura 11).
Em simulação, foram definidos parâmetros como: material Abs, base fixada na
região inferior da Base de proteção e uma força de 2 N na região superior/lateral
para uma simulação de uma eventual batida forte do animal após implantado.
A simulação mostra que a peça apresenta maior deslocamento na região
superior, onde não há contato direto com nenhuma outra parte do microdrive
(Figura 11-A), podendo assim se movimentar sem que haja qualquer dano na
34

estrutura. A parte inferior, que é fixada na cabeça do animal com acrílico, não
apresentou qualquer movimentação no teste, proporcionando assim uma baixa
força de cisalhamento entre o implante e o acrílico.
Figura 11: Testes de deslocamento e tensão na base de proteção. Em (A) vemos um mapa de
calor do deslocamento da estrutura quando aplicada uma força de 2 N (representada pela seta
branca). Note que a região inferior da Base de proteção, a qual será fixada no animal, segue intacta
a qualquer movimento. (B) mostra o von Mises Stress, um tipo de medida de tensão da peça.
Perceba que existe pouco estresse no geral e que o vazamento ajuda na dissipação da tensão.
O von Mises Stress é uma medida de tensão baseada na relação da energia
de cisalhamento com o limite máximo de tensão do objeto. Foi observado que
houve pouco estresse na estrutura no geral (Figura 11-B). Os vazamentos da peça
ajudam na dissipação da tensão sofrida na base e na diminuição do peso,
influenciando assim na durabilidade do implante.
3.2 Posicionamento dos tetrodos
Como já dito anteriormente, uma das principais vantagens do uso de
microdrives é o reposicionamento dos tetrodos para registro de diferentes
neurônios ao longo do tempo. Como forma de validar o funcionamento do passo
dos parafusos, fizemos implantes crônicos dos microdrives em ratos com alvo em
CA1 e registramos sua atividade ao longo de diferentes camadas do tecido
cerebral (mais informações no capítulo 2). Sabendo que cada volta completa no
parafuso representa 0,32 mm, descemos os tetrodos progressivamente no
decorrer dos dias. Inicialmente o dado de cada tetrodo colhido foi filtrado entre
35

500-6000 Hz para detecção dos disparos e seus formatos de onda foram
armazenados. Em seguida analisamos a diferença de pico/pico de dois canais em
diferentes tetrodos ao longo de seu deslocamento no tecido cerebral (Figura 12).
Com propósito de registrar a camada piramidal de CA1 dorsal (2,3 mm de
profundidade a partir do início do córtex), espera-se que aproximadamente 7,2
voltas no parafuso seja suficiente para atingir o alvo. Nos registros da figura 12, a
camada piramidal foi atingida com 7,5 e 7,2 voltas no parafuso.
Nossos resultados mostram também que no decorrer da descida dos
tetrodos, particularidades da camada piramidal puderam ser observadas, como
maiores agrupamentos de disparos (Figura 12- A e B). Vimos também que, quando
no alvo, um leve torque no parafuso é suficiente para avistar novos agrupamentos
de disparos (Figura 12-C). Esse resultado se assemelha com o do Flexdrive® já
mostrado por Voigts e colaboradores, 2013.
Figura 12: Reposicionamento do tetrodo ao longo dos dias. Em (A) e (B) vemos dois tetrodos em
diferentes alturas no tecido cerebral, onde comparamos o pico/pico de 2 canais. Em cinza temos
36

um provável alvo da camada piramidal. (C) Representa um tetrodo possivelmente na camada
piramidal que apresenta diferença de agrupamentos no decorrer de pequenas descidas.
III - Capítulo 2: Implantação da técnica de registro de células
de lugar utilizando microdrives de tetrodos móveis em ratos
________________________________________________________________________
37

Embora o estudo dos correlatos neurais da codificação espacial constitua
uma área extremamente ativa de pesquisa atual, até o presente momento nenhum
laboratório no Brasil publicou estudos envolvendo registro de células de lugar, o
que portanto caracteriza um atraso científico de mais de 40 anos em relação aos
laboratórios de países desenvolvidos. O Laboratório de Neurofisiologia
Computacional do Instituto do Cérebro da UFRN, coordenado pelo orientador do
aluno de mestrado da presente dissertação, possui grande interesse nesta área de
pesquisa, e, de fato, já vem contribuindo para tal através da análise computacional
de registros de células de lugar colhidos por terceiros e que estão disponíveis na
internet em sites de compartilhamento de dados científicos (por exemplo, ver
www.crcns.org). Contudo, por já estarem colhidos, as perguntas que podem ser
respondidas por estes dados são restritas. Neste sentido, a fim de ganhar mais
autonomia científica, torna-se necessário que o laboratório, além de sua expertise
na análise computacional dos registros, também desenvolva expertise
experimental na coleta de seus próprios dados de atividade de células de lugar.
Para tanto, uma primeira etapa crucial é a implementação da tecnologia de
registros eletrofisiológicos do hipocampo através do uso de microdrives de
tetrodos móveis. Portanto, almejamos aqui uma validação da técnica, que servirá
de importante base para estudos futuros visando compreender a formação de
novas memórias espaciais dependentes do hipocampo.
38

Assim, o presente estudo visou implementar um sistema de registro
eletrofisiológico utilizando o novo protótipo de microdrive contendo 16 tetrodos
móveis desenvolvido no presente projeto (ver capítulo 1). Para validar este
protótipo, registramos a atividade de populações de células de lugar na região
CA1 do hipocampo dorsal de ratos durante uma tarefa de exploração de um
labirinto linear associado a recompensas de água nas extremidades.
1. Objetivos
1.1 Objetivo geral
Implementar no Laboratório de Neurofisiologia Computacional do Instituto
do Cérebro da UFRN a técnica de registro de potenciais de ação de células de
lugar no hipocampo de ratos utilizando um novo protótipo de um microdrive de
tetrodos móveis.
1.2 Objetivos Específicos
1 – Utilizar o novo protótipo de microdrive para o implante crônico de 16 tetrodos
móveis que possam ser precisamente posicionados após a cirurgia de implante.
2 – Utilizar tetrodos para registro eletrofisiológico do potencial de campo local e
da atividade de neurônios hipocampais individuais.
3 – Realizar o registro eletrofisiológico de potenciais de ação e de potenciais de
campo local na subárea CA1 do hipocampo dorsal de ratos enquanto os animais
39

buscam recompensa de água nas extremidades de um labirinto linear e durante o
sono pré e pós tarefa para futuras análises.
4 – Aplicar ferramentas de classificação de disparos (“spike sorting”) para
identificação de unidades neuronais individuais a partir das propriedades das
formas de onda do potencial de ação.
5– Fazer a análise do registro eletrofisiológico para quantificação da taxa de
disparos neuronal em relação à posição do animal ao longo do labirinto linear, a
fim de detectar células de lugar.
Métodos
2.1 Sujeitos experimentais
Utilizamos 10 ratos wistar machos adultos (Rattus Norvegicus) entre 2 e 4
meses de vida. Os animais foram mantidos em gaiolas coletivas sob ciclo
claro-escuro de 12 horas com água e ração ad libitum. Os experimentos foram
realizados em sua maioria na fase clara do ciclo. Todos os experimentos seguiram
as normas éticas estabelecidas pelo comitê de ética animal (CEUA 058/2016).
2.2 Protocolo de aprendizado espacial
Os animais foram submetidos a uma tarefa de aprendizado espacial para
obtenção de recompensa de água em labirinto linear. Alguns animais também
foram submetidos a sessões de sono de uma hora de duração antes e após a
execução da tarefa (Figura 13). Os animais foram privados de água durante as 18
horas que antecederam os experimentos. Ao final das sessões diárias de
experimento, os animais receberam água ad libitum durante 6 horas. A cada dia
40

do protocolo da tarefa, o animal foi submetido a uma sessão de exploração do
labirinto linear (180 cm de comprimento por 21 cm de largura) com duração de 30
minutos. A tarefa consistiu do deslocamento espacial do animal a partir de uma
extremidade do labirinto até a outra para a obtenção de uma recompensa de
água. A presença dos animais foi detectada por sensores de infravermelho
instalados nas paredes do labirinto e localizados a 25 cm do local de recompensa.
Ao passar pelos sensores, a presença do animal ativa um sistema automatizado
que aciona a liberação de 60 µL de água. A próxima recompensa liberada por esse
sensor só pode ser acionada após o animal visitar a outra extremidade do
labirinto. Foram registrados automaticamente os instantes de tempo em que o
animal atravessa os sensores. O comportamento dos animais foi registrado por
meio de uma câmera de vídeo digital (Logitech® c920) localizada
aproximadamente 200 cm acima do labirinto linear. Os arquivos foram adquiridos
à taxa de 30 imagens por segundo e armazenados em disco rígido para análise
posterior.
Figura 13: Ilustração do protocolo de labirinto linear utilizado na tarefa comportamental. O labirinto
é feito de madeira em suas extremidades e revestido de acrílico em suas paredes, sendo ele
conectado a sensores de infravermelho próximos à extremidade para detectar a presença do
animal e liberar a recompensa de água. Um LED vermelho é acoplado com o sensor infravermelho
para a sincronização do registro eletrofisiológico por vídeo.
2.3 Neurocirurgia para implante de microdrive
Cada animal foi submetido a uma cirurgia estereotáxica para o implante
permanente de um microdrive contendo oito ou dezesseis tetrodos móveis. Os
animais foram anestesiados através da administração intramuscular de sulfato de
41

atropina (0,04 mg/kg), seguido de cloridrato de cetamina (100 mg/kg) e cloridrato
de xilazina (8 mg/kg). Durante a cirurgia, o nível da anestesia era periodicamente
verificado através de alterações das frequências cardíaca e respiratória, e de
outros sinais fisiológicos como o reflexo da cauda. Doses suplementares de
cetamina (metade da dose inicial) eram administradas quando necessário. Todos
os esforços foram empregados para minimizar o sofrimento animal durante os
procedimentos experimentais. Após tricotomia, o animal foi posicionado em um
aparelho estereotáxico (Kopf). Lidocaína foi injetada no tecido subcutâneo
craniano onde foi realizada uma incisão longitudinal, expondo o crânio. O centro e
os quatro pontos marginais da craniotomia foram marcados segundo as
dimensões do microdrive de tetrodos e segundo as coordenadas estereotáxicas
da subárea CA1 do hipocampo dorsal (AP: -4,16 mm; ML: +2,4 mm; DV: 2,40 mm).
Brocas dentais de diâmetro apropriado foram utilizadas para a realização da
craniotomia de uma janela quadrada, e de seis pequenas perfurações para
inserção de parafusos de sustentação e de dois parafusos de
sustentação/aterramento. Os dois parafusos de sustentação/aterramento foram
implantados na região occipital, e entraram em contato com o líquido
cefalorraquidiano cerebelar, servindo como aterramento elétrico dos eletrodos de
registro. Os parafusos de sustentação/aterramento foram soldados a fios de aço e
conectados ao canal de referência da placa de circuito acoplada ao microdrive.
Após craniotomia da janela, as meninges cerebrais foram cuidadosamente
retiradas, expondo assim o córtex cerebral e permitindo a penetração dos
tetrodos. Durante a cirurgia de implante, os tetrodos foram inseridos apenas
superficialmente no tecido cerebral, aproximadamente 1 mm (equivalente a 3
voltas no parafuso-guia). O posicionamento final dos tetrodos na camada
piramidal de CA1 foi realizado conjuntamente à visualização dos sinais captados,
após a recuperação pós-cirúrgica dos animais. Acrílico dental polimerizável foi
usado para confeccionar um capacete sobre o crânio, dando sustentação e
estabilizando a posição final do microdrive. Após a cirurgia, os animais receberam
antibiótico Fluotril (i.m.) na dose de 2,5 mg/kg. Os animais tiveram acesso livre
diário a 200 ml de água contendo 20 gotas do analgésico paracetamol (200
42

mg/kg, sendo que 15 gotas contém 1 ml, que contém 200 mg de paracetamol), e
permaneceram em repouso por uma semana para recuperação do peso corporal.
2.4 Aquisição de dados eletrofisiológicos e comportamentais
Os sinais eletrofisiológicos foram adquiridos a uma taxa de 30 KHz, filtrados
entre 0,1 Hz e 7,5 KHz, amplificados em 20X e digitalizados através do uso de
headstages Intan® e do sistema de registro Open Ephys. Os movimentos da
cabeça do animal nos três eixos de deslocamento foram detectados por
acelerômetros localizados nos headstages Intan®. O comportamento foi filmado e
gravado em vídeo através de uma câmera Logitech c920 na frequência de 30
imagens por segundo.
2.5 Eutanásia por perfusão transcardíaca
Depois de injetarmos uma sobredose de Tiopental (100 mg/kg), o rato foi
posicionado sobre uma plataforma plana e seus membros foram presos por fitas à
mesa. Foi feita, então, uma marcação tecidual do posicionamento de cada
eletrodo por passagem de corrente elétrica contínua de 0,2 µA por 30 segundos.
Em seguida, foi feita uma incisão no abdome e estendida até o tórax de maneira a
expor o coração. Uma ponteira foi introduzida no ventrículo esquerdo do coração
e guiada até a aorta. Em seguida, foi feita uma incisão no ventrículo direito para a
circulação do sangue. Um sistema de bombeamento acoplado à ponteira injetou
500 ml de salina (preparada a 38°C) e 500 ml de paraformaldeído (preparado a
20°C). Após esse processo, o cérebro foi cuidadosamente retirado, mantido em
solução de paraformaldeído por 24 horas, seguido de solução de sacarose por 48
horas, e foi posteriormente congelado em gel para inclusão (Tissue Tek®) e
conservado em refrigerador à temperatura de -80°C para processamento
histológico posterior.
2.6 Análise histológica
O tecido cerebral foi seccionado em fatias coronais de 50 micrômetros de
espessura em um criostato (Micron®), em seguida montado em lâminas
gelatinizadas e corado com solução cresil violeta para registro fotográfico e
43

inspeção visual do posicionamento final dos eletrodos de registro através de
microscopia luminosa com lente amplificada em 5x.
2.7 Sincronização de registros eletrofisiológicos e de vídeo
Os vídeos e os registros eletrofisiológicos foram sincronizados para que a
posição instantânea do animal fosse alinhada ao tempo de disparo dos neurônios.
A sincronização se deu a partir da normalização dos mesmos pelo tempo de
registro do primeiro ao último evento identificados pelo infravermelho.
Inicialmente, foram detectados os frames em que o LED vermelho acoplado
ao infravermelho acende, para identificação do primeiro e último frame em que o
animal cruzou o labirinto, chamando-os assim de firstledon e lastledon (primeiro e
último frame em que o LED acende). Sabendo que a taxa de aquisição da câmera
é de 30 frames/s, cada frame será associado a um período de amostragem de
1/30 segundos, gerando assim um vetor de tempo de cada posição do animal na
tarefa (Frametime). Durante o registro, o infravermelho quando acionado envia
TTL’s para a Open Ephys, salvando assim o momento exato do registro
eletrofisiológico em que o animal cruza o labirinto naquela posição. O primeiro e
último eventos marcados (ftimeevent e ltimeevent) foram utilizados para identificar
o início e fim da tarefa nos registros eletrofisiológicos.
2.8 Análise de dados neurais
Todos os registros eletrofisiológicos e de vídeo foram analisados usando
rotinas programadas em MATLAB®. O registro eletrofisiológico foi filtrado entre
500-6000 Hz para a detecção dos disparos e armazenamento do formato de onda.
A detecção e a classificação dos potenciais de ação foram feitas a partir de
características das formas de onda. A classificação foi feita tanto pelo algoritmo
MClust, desenvolvido por David Redish, como por um algoritmo desenvolvido em
nosso laboratório (Souza et al, 2018) que utiliza modelos de misturas de
gaussianas (GMMs, do inglês, Gaussian mixture models) para a caracterização de
atributos da onda. A classificação neuronal foi feita utilizando tanto os sistemas
autônomos Klustakwik (Figura 14) e GMMs (Figura 15), juntamente de correções
44

manuais. Essas correções manuais se dão pela verificação da energia e amplitude
dos formatos de onda dos neurônios em relação à sua classificação.
Figura 14: Exemplo de classificação autônoma utilizando o Klustakwik a partir da análise dos
componentes principais. A figura apresenta as diferenças de cluster de 2 neurônios de um mesmo
eletrodo (A e B). Em (C e D) temos o formato de onda nos neurônios classificados (figura retirada
de Keopsell et al, 2009).
45

Figura 15: Exemplo de classificação autônoma utilizando o modelo de misturas de gaussianas
desenvolvido em nosso laboratório. A figura apresenta as diferenças de cluster de 5 neurônios de
um mesmo tetrodo em 4 diferentes pares de atributos do formato de onda.
Após a atribuição dos disparos a unidades neuronais individuais, iniciamos
a detecção das células de lugar. Como descrição clássica, a célula de lugar
apresenta uma maior taxa de disparo em um (ou mais) determinado lugar no
espaço. Para medir esse tipo de informação, a métrica que leva em consideração
ae taxa de disparo por taxa de ocupação definida por Skaggs e colaboradores
(1993) foi utilizada. Essa métrica calcula a informação espacial por disparos
neuronais em unidade de bits/spikes, como descrito na equação abaixo:
sendo o N o número total de bins, pi a probabilidade do animal estar no bin i, λi a
taxa de disparo no bin i e λ a taxa de disparo total da célula. No nosso caso, o
46

labirinto foi binado em bins de 4 cm², sendo que 25 cm foram excluídos em cada
extremidade.
3. Resultados
3.2 Comportamento espacial
O comportamento do animal durante a execução da tarefa espacial no
labirinto linear foi gravado em vídeo em todas as sessões de registros. Um LED
acoplado ao headstage foi utilizado para detectar a posição instantânea do animal
ao longo do labirinto linear. E então um algoritmo foi desenvolvido para rastrear a
posição do animal automaticamente.
Observamos que, durante as explorações espaciais, todos os animais
apresentaram um maior tempo de permanência nas extremidades do labirinto
linear, na maioria das vezes em uma das extremidades. Como exemplo, o rato 04
executou mais de 18 travessias em 4 das 8 sessões registradas (critério mínimo de
18 e máximo de 36 travessias), sendo as mesmas selecionadas para análises
adicionais. A trajetória do animal foi plotada e o tempo de permanência em cada
um dos 210 quadrantes do labirinto linear foi calculado. A Figura 16 mostra que o
animal explorou toda a área do labirinto e apresentou maior tempo de
permanência nas extremidades do labirinto (região próxima à área de recompensa
e que apresenta paredes escuras).
47

Figura 16: Posições do animal no labirinto linear em diferentes sessões. A área da plataforma foi
dividida em 42x5 bins de aproximadamente 4 cm². O tempo em cada bin foi somado para calcular
a matriz de ocupância (Occupation Matrix). Como mostram as matrizes de ocupância das 4
sessões, o rato tem maior tempo de permanência nas extremidades do labirinto, perto da área de
recompensa.
3.3 Potencias de Ação e Células de lugar
Nesse subtópico, 4 sessões de registros de 3 ratos diferentes foram
analisadas. Algumas outras sessões foram pré-analisadas e descartadas devido à
má qualidade do dado.
3.3.1 Classificação de unidades neuronais individuais
Inicialmente, os potenciais de ação registrados em cada tetrodo foram
atribuídos a unidades neuronais individuais através da técnica de spike sorting
usando o programa MClust®, desenvolvido por David Redish (Universidade de
Minnesota), ou pelo classificador desenvolvido em nosso laboratório por Bryan
Souza (Souza et al, 2018). Em ambos os programas, a classificação é feita por
algoritmos semi-automáticos seguido de ajustes manuais quando necessário.
48

Como exemplo de classificação, podemos ver na Figura 17 a distribuição dos
potenciais de ação obtidos de um único tetrodo em um registro de 5 minutos na
sleep box. Foram encontradas 5 unidades neuronais individuais a partir da
diferença de pico-vale do formato de onda e do autocorrelograma dos próprios
clusters.
49

Figura 17: Atividade neuronal registrada por um tetrodo. Em (A) pode-se ver a clusterização
neuronal de um tetrodo a partir da diferença pico-vale dos formatos de onda. (B) mostra o formato
50

de onda e o desvio padrão de 5 neurônios clusterizados em (A) (separados pelas cores) e seus
autocorrelogramas.
3.3.2 Neurônios candidatos a células de lugar
Com o objetivo de selecionar apenas os neurônios candidatos a células de lugar, analisamos a atividade de neurônios individuais de 2 registros e consideramos apenas aqueles que apresentaram uma taxa de disparo média acima de 0,2 Hz e um índice de informação (I) acima de 0,5. Com esses critérios, foram obtidas 18 células de lugar, de um total de 71 neurônios selecionados (25,3% do total) (Figura 18-A). Como exemplo ilustrativo, na Figura 18-B mostramos 3 exemplos de células: (1) um neurônio excluído de análises adicionais por apresentar baixa taxa de disparos, (2) um neurônio candidato a célula de lugar, e (3) um neurônio com alta taxa de disparo, mas pouca informação espacial e por isso excluído de análises adicionais.
Figura 18: Seleção de candidatos a células de lugar. Em (A) plotamos a taxa de disparo em função
da quantidade de informação espacial (I, bits/spike) dos neurônios analisados. As linhas tracejadas
em vermelho ilustram os critérios definidos para a detecção dos candidatos a células de lugar. Em
(B) é mostrada e taxa de disparo de 3 neurônios ao longo do labirinto linear binado. Note que
nesses exemplos, o turquesa e o azul não foram selecionados como candidatos a célula de lugar
por não atenderem aos critérios (taxa de disparo > 0,2 Hz e I > 0,5). Em vermelho um exemplo de
célula candidata a ser célula de lugar.
3.3.3 Células de lugar
Após selecionar as células de lugar, os disparos provenientes de cada
neurônio foram plotados em função das respectivas posições do animal no
51

labirinto (Figura 18-A). A somatória dos disparos em cada bin dividida pelo tempo
permanecido no mesmo foi usada para comparar a taxa de disparo do neurônio
em cada posição ocupada pelo animal ao longo do labirinto (Figura 18-B). Apenas
as sessões em que o animal executou um número de travessias acima do critério
foram analisadas.
Os resultados mostram que os neurônios selecionados como candidatos a
células de lugar apresentam um ou mais campos de lugar e taxas de disparos
aumentadas em locais específicos do labirinto linear (Figura 19).
52

Figura 19: Seletividade espacial de neurônio de CA1 dorsal. Os potenciais de ação desta célula
piramidal putativa (pontos vermelhos) foram plotados em função de 24 passagens (linha cinza)
executadas pelo rato ao longo do labirinto linear (superior). Taxas de disparos do mesmo neurônio
foram divididas e linearizadas em 42 bins ao longo do labirinto (as bordas foram descartadas)
(inferior). Note que há um aumento na taxa de disparo no lado direito do labirinto linear (limiar
acima de 7 Hz). Apenas períodos onde o animal se encontrava em movimento foram considerados
nesta análise.
Na Figura 20, os campos de lugar dessas células também podem ser
observados nos eixos X e Y da trajetória do animal. Por ser na extremidade do
labirinto o lugar onde o animal se encontra mais parado, 15 cm de cada
extremidade do aparato foram excluídos das análises. As células de lugar foram
binadas em 5 cm para que fosse possível encontrar o seu campo de lugar. A taxa
de disparo foi computada pela divisão de cada bin de contagem de disparos pela
ocupância do animal no bin.
Figura 20: Células de lugar ao longo da trajetória percorrida no labirinto linear. A figura mostra 4
células de lugar durante a corrida no labirinto linear. Em cima temos a trajetória percorrida pelo
animal em preto e em vermelho os momentos em que as células de lugar dispararam. Abaixo
vemos o percurso binado e também “suavizado”.
53
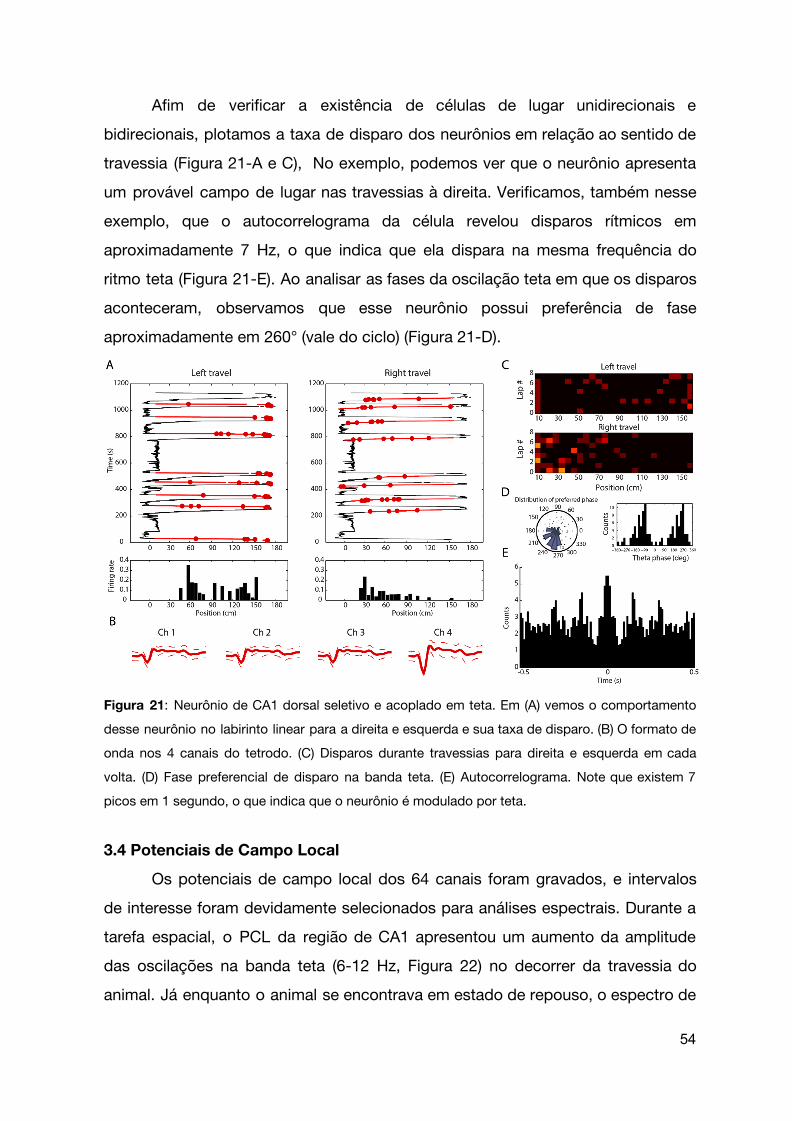
Afim de verificar a existência de células de lugar unidirecionais e
bidirecionais, plotamos a taxa de disparo dos neurônios em relação ao sentido de
travessia (Figura 21-A e C), No exemplo, podemos ver que o neurônio apresenta
um provável campo de lugar nas travessias à direita. Verificamos, também nesse
exemplo, que o autocorrelograma da célula revelou disparos rítmicos em
aproximadamente 7 Hz, o que indica que ela dispara na mesma frequência do
ritmo teta (Figura 21-E). Ao analisar as fases da oscilação teta em que os disparos
aconteceram, observamos que esse neurônio possui preferência de fase
aproximadamente em 260° (vale do ciclo) (Figura 21-D).
Figura 21: Neurônio de CA1 dorsal seletivo e acoplado em teta. Em (A) vemos o comportamento
desse neurônio no labirinto linear para a direita e esquerda e sua taxa de disparo. (B) O formato de
onda nos 4 canais do tetrodo. (C) Disparos durante travessias para direita e esquerda em cada
volta. (D) Fase preferencial de disparo na banda teta. (E) Autocorrelograma. Note que existem 7
picos em 1 segundo, o que indica que o neurônio é modulado por teta.
3.4 Potenciais de Campo Local
Os potenciais de campo local dos 64 canais foram gravados, e intervalos
de interesse foram devidamente selecionados para análises espectrais. Durante a
tarefa espacial, o PCL da região de CA1 apresentou um aumento da amplitude
das oscilações na banda teta (6-12 Hz, Figura 22) no decorrer da travessia do
animal. Já enquanto o animal se encontrava em estado de repouso, o espectro de
54

potências mostra o predomínio de frequências mais lentas, na banda delta (1-4
Hz). Nesse momento, também é possível observar a ocorrência de episódios
pontuais de oscilações de alta frequência, denominadas ripples (150-250 Hz).
Figura 22: Potencial de campo local de CA1 durante uma travessia no labirinto linear. Sinais brutos
de PCL obtido de cada canal (ch 1-4 de um tetrodo) mostram duas diferentes frequências
oscilatórias a partir da trajetória do animal (acima). Espectrograma do canal 4 (verde) mostra um
aumento da potência oscilatória em teta (5-12 Hz). A barra cinza indica quando o animal chegou ao
fim do labirinto linear (recompensa entregue).
Para uma melhor visualização do PCL, dividimos o registro em 3 estados:
os momentos de travessia no labirinto linear, episódios de SOL e de sono MRO
durante os períodos pré tarefa (Figura 23-A e B). Concatenamos momentos
provenientes de registros obtidos de 3 animais (Rato 04, Rato 05, e Rato 06) para
um vetor de cada condição.
Inicialmente foram feitas análises da potência espectral de cada animal em
SOL, MRO e durante a travessia (exemplo do Rato 04 na Figura 23-C). Os estágios
de sono foram definidos por observações manuais do PCL e vídeo devidamente
55

sincronizados. Com isso, integramos a área do PSD nos segmentos de teta (5-12
Hz), delta (1-5 Hz), LG (30-55 Hz) e HG (65-100 Hz) para análise da potência
(Figura 23-D). Podemos observar que o sinal dos animais corroboram achados
publicados na literatura, como o aumento de teta durante MRO e travessia, e o
aumento do ritmo delta durante o SOL (Buzsáki, 2002; Mole et al., 2006). O HG
também apresentou um aumento de amplitude na travessia comparado com SOL
e MRO (Sheffzuk et al., 2011).
Figura 23: Potenciais de campo local de CA1 em alguns estados. Em (A) temos exemplos de 2
segundos de um sinal bruto, filtrado em teta (5-12 Hz), LG (30-55 Hz) e HG (65-100 Hz) em SOL,
MRO e durante a travessia no labirinto linear. Em (B) vemos o espectrograma de todo o sinal
filtrado e concatenado do registro do mesmo rato (Rato 04). O colorbar foi modificado para “hot”
para facilitar a visualização da imagem, já que a escala foi ancorada para os três espectrogramas.
No painel (C), temos o PSD do sinal em (B). (D) Integral da PSD nos intervalos de teta, delta, LG e
HG para os 3 animais.
3.4.1 Comodulogramas
Fizemos análises de acoplamento fase-amplitude (Rato 04) durante os
momentos de sono pré tarefa (SOL e MOL) e de caminhada no labirinto linear
56

(Figura 24-A). Os períodos representam uma concatenação de 123 segundos de
SOL, 43 segundos de MRO e 200 segundos de caminhada (25 corridas).
Podemos observar que durante os episódios de SOL houve acoplamento
da fase de delta com LG e a amplitude de HG, e, em menor escala, entre a fase de
teta e a amplitude de HG. Durante os episódios de sono REM, podemos observar
um acoplamento claro entre a fase de teta e a amplitude de HG. Na caminhada, foi
observado um acoplamento tanto das fases de teta com a amplitude de LG, como
entre as fases de teta e a amplitude de HG, corroborando como o que já existe
relatado na literatura (Figura 24-B).
Ao que já sabemos, existe um acoplamento maior entre as fases de teta e a
amplitude de HG durante o sono MRO (Sheffzuk et al, 2011; Scheffer-Teixeira e
Tort, 2017) do que quando o animal está acordado, o que também foi observado
em nossos resultados. Vale ressaltar que os tempos analisados para cada
comodulograma podem ter sido curtos, o que pode influenciar em quão robustas
são as modulações.
57

Figura 24: Comodulograma fase-amplitude para os diferentes estados comportamentais. Em (A)
temos um exemplo de comodulograma de um registro de SOL, MRO e “Running”, este último
definido como a travessia do animal no labirinto linear. (B) mostra as distribuições fase-amplitude
dos exemplos de (A), computados para: delta (1-5 Hz) / LG (30-60 Hz), delta (1-5 Hz) / HG (65-100
Hz), teta (5-10 Hz) / LG (30-60 Hz) e teta (5-10 Hz) / HG (65-100 Hz). Os números em cima de cada
barra significa o valor de acoplamento (índice de modulação ,IM).
58
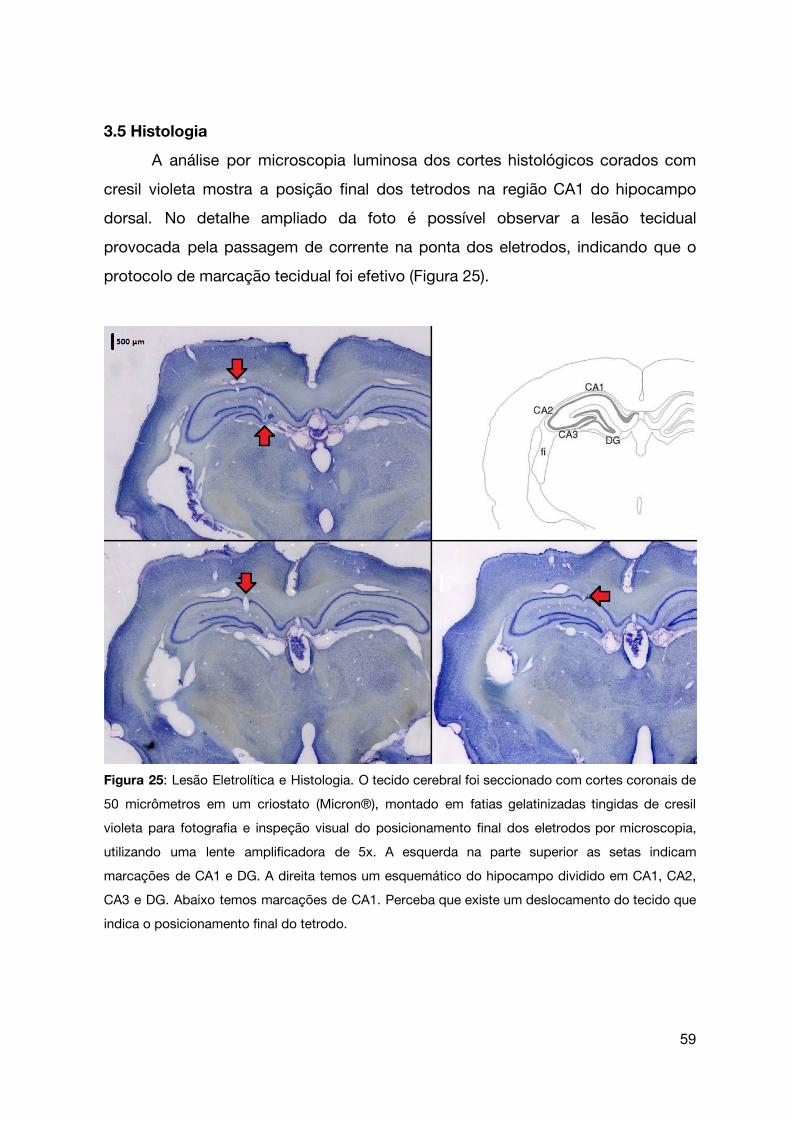
3.5 Histologia
A análise por microscopia luminosa dos cortes histológicos corados com
cresil violeta mostra a posição final dos tetrodos na região CA1 do hipocampo
dorsal. No detalhe ampliado da foto é possível observar a lesão tecidual
provocada pela passagem de corrente na ponta dos eletrodos, indicando que o
protocolo de marcação tecidual foi efetivo (Figura 25).
Figura 25: Lesão Eletrolítica e Histologia. O tecido cerebral foi seccionado com cortes coronais de
50 micrômetros em um criostato (Micron®), montado em fatias gelatinizadas tingidas de cresil
violeta para fotografia e inspeção visual do posicionamento final dos eletrodos por microscopia,
utilizando uma lente amplificadora de 5x. A esquerda na parte superior as setas indicam
marcações de CA1 e DG. A direita temos um esquemático do hipocampo dividido em CA1, CA2,
CA3 e DG. Abaixo temos marcações de CA1. Perceba que existe um deslocamento do tecido que
indica o posicionamento final do tetrodo.
59

4. Conclusão
Utilizando os 2 protótipos de microdrives desenvolvidos (ver capítulo 1),
registramos a atividade eletrofisiológica de neurônios do hipocampo de ratos.
Abaixo estão sumarizados os objetivos e suas devidas conclusões.
Objetivo 1:
- O protótipo foi desenvolvido e implantado com sucesso, e os tetrodos
foram precisamente posicionados na região de CA1.
Objetivo 2:
- Os tetrodos foram confeccionados e utilizados para registro da atividade de
disparos e das oscilações de populações de neurônios. No todo, os últimos
animais apresentaram uma grande qualidade nos registros de LFP e um
bom desempenho quanto à atividade neuronal.
Objetivo 3:
- Os animais foram devidamente treinados e executaram bem a tarefa no
labirinto linear, onde correram por recompensa de água, cruzando o
labirinto de 8 a 37 vezes. Foi percebido que no geral o animal preferia uma
das extremidades do labirinto para explorar mais.
Objetivo 4:
- Inicialmente houve um retardo nas análises de sorting dos dados por um
problema de conversão. Foram utilizados no total 4 classificadores
diferentes para finalmente padronizar as análises com o Klustakwik e o
GMM. Nem todos os dados colhidos foram analisados devido à quantidade
e tempo até o momento (9 registros foram analisados).
60

Objetivo 5:
- Os dados que foram selecionados para análise apresentaram em
aproximadamente metade de seus tetrodos a atividade neuronal
característica de CA1.
Em suma, a técnica de registro neuronal com microdrive de tetrodos
móveis foi implantada no laboratório. As análises dos dados mostraram atividade
de células de lugar no labirinto linear. Nosso trabalho, além de tudo, apresentou
grande importância metodológica para o Instituto do Cérebro em geral, que ganha
maiores opções de pesquisa com a padronização de uma técnica desenvolvida
com tecnologia própria.
61

IV - Capítulo 3: Velocidade de corrida modula a oscilação
delta no hipocampo de ratos
________________________________________________________________________
62

Durante os registros ocorridos no labirinto linear, foi possível notar que,
além da presença da oscilação teta (5-10 Hz), uma oscilação delta (1-4 Hz)
apresentava picos de coatividade durante períodos de aumento de velocidade. A
fim de investigar isso mais profundamente, fizemos a análise da atividade do PCL
durante a locomoção do animal. Para controlar a velocidade de locomoção dos
animais, utilizamos uma esteira elétrica controlada por computador enquanto os
ratos (wistar) corriam em protocolos de velocidades constantes e crescentes.
Observamos o surgimento de oscilações na banda delta no hipocampo enquanto
os ratos aumentam sua velocidade de locomoção. A amplitude e a frequência das
oscilações delta foram fortemente associadas à velocidade de corrida dos
animais, e a ritmicidade da banda delta se tornava notavelmente periódica acima
das velocidades de 30 cm/s. Nossos resultados demonstram que a velocidade de
corrida modula fortemente as oscilações delta no hipocampo de ratos, e que esse
ritmo e a oscilação teta podem coexistir.
1. Objetivos
1.1 Objetivo geral
Registrar a atividade eletrofisiológica do hipocampo de ratos enquanto os
animais correm em uma esteira elétrica controlada por computador em protocolos
de velocidades constantes e crescentes.
1.2 Objetivos específicos
1 – Desenvolver um protocolo experimental para a tarefa com fim de analisar a
relação entre a velocidade do animal e as frequências lentas do PCL hipocampal.
2 – Registrar os sinais eletrofisiológicos na subárea CA1 do hipocampo dorsal de
ratos enquanto correm em uma tarefa de locomoção em esteira.
63

3 – Analisar a relação entre oscilações neurais e a velocidade de locomoção dos
animais.
2. Métodos
2.1 Sujeitos experimentais
Utilizamos 4 ratos machos adultos (Rattus Norvegicus) entre 2 e 4 meses de
vida. Os animais foram mantidos em gaiolas individuais sob ciclo claro-escuro de
12 horas com água e ração ad libitum. Todos os experimentos foram realizados
durante a fase clara do ciclo. Nesses experimentos, foram utilizados os mesmos
animais implantados com o microdrive de 16 tetrodos móveis.
2.2 Protocolo da tarefa na esteira
Inicialmente os animais foram treinados em protocolos de baixas
velocidades contínuas por 15 minutos por 2 vezes ao dia para que aprendessem a
andar na esteira . Durante os treinamentos, uma barra de choque de 0,1 a 0,7 mA 8
foi ligada para que o animal evitasse descansar fora da cinta, na parte posterior da
esteira (Figura 26). Uma vez que os animais aprenderam a tarefa, era iniciado o
protocolo de velocidades constantes e velocidades crescentes (Figura 27). O início
do protocolo é sinalizado com 3 tons neutros de 100 ms intercalados por 150 ms,
seguido de um intervalo de 5 s com a esteira parada. Os protocolos de locomoção
(Aceleração1 (A1) = 0-20 cm/s, Aceleração2 (A2)= 0-30 cm/s, Aceleração3 (A3)=
0-40 cm/s, Velocidade1 (V1) = 20 cm/s, Velocidade2 (V2) = 30 cm/s, Velocidade3
(V3) = 40 cm/s) são iniciados de acordo com uma ordem previamente definida,
com duração de 20 s por protocolo e intervalo de 5 s entre protocolos. Essa
sequência de protocolos foi repetida 7 vezes, gerando um total de 42 episódios de
corrida, equivalente a 17 min e 30 s de tarefa. Nossos protocolos de corrida e de
choque foram baseados em estudos anteriores (Albeck et al, 2006; O’Callaghan et
8 A esteira possui 40 cm de comprimento e tem uma velocidade máxima de 120 cm/s. Além de possuir regulagem angular de inclinação, possui uma barra de choque na parte de trás e uma ventoinha na parte da frente adaptada.
64

al, 2007; Liu et al, 2009). Um dos animais preliminares (Rato 05) foi registrado com
velocidades de 40, 50 e 60 cm/s.
Figura 26: Ilustração da tarefa de locomoção na esteira. A esteira é revestida de acrílico e
apresenta um sistema gradeado de choque na parte posterior e uma ventoinha de refrigeração na
parte frontal. Um LED vermelho foi colocado em um quadrante da imagem capturada pelo vídeo
para a sincronização de registros eletrofisiológicos e de vídeo.
Figura 27: Ilustração da sequência de protocolos de velocidades na esteira. A tarefa inicia com 3
tons de 100 ms intercalados por 150 ms, seguido de 20 s de 6 diferentes condições de velocidade,
intercalados por 5 segundos de intervalo.
A etapa de treinos durou 1 mês, visto que os animais apresentaram
dificuldade em correr em altas velocidades. Nos últimos treinos antes do
procedimento cirúrgico e ao longo de todos os registros, o choque foi mantido
desligado. Após recuperação cirúrgica, o animal foi re-treinado por mais duas
semanas antes que houvesse o primeiro registro. Uma câmera Logitech c920 foi
utilizada para filmar o experimento com amostragem de 30 quadros/s. Um LED
vermelho foi acoplado em um dos quadrantes da imagem capturada pelo vídeo
para posterior sincronização entre vídeo e registro.
65

3. Resultados
Registramos 4 animais, sendo que um deles foi registrado com A1 = 0-40
cm/s, A2 = 0-50 cm/s, A3 = 0-60 cm/s, V1 = 40 cm/s, V2 = 50 cm/s e V3 = 60
cm/s e os demais seguindo o protocolo normal. Um trial é descrito como a corrida
individual de cada velocidade de locomoção (A1, A2, A3, V1, V2 e V3). Corridas
em que os animais apresentaram taxa de execução menor 18s dos 20s de cada
trial foram excluídas de qualquer análise (animal parar na barra de choque por 2
segundos por exemplo). A taxa de execução é dada pelo momento em que o
animal corre adequadamente durante a tarefa. Observamos que em todos os
registros e em todos os trials o animal desempenhou a tarefa em marcha de
caminhada, como esperado pela definição de Whishaw e Kolb (2005).
3.1 Picos de oscilações delta no labirinto linear
Nos experimentos descritos no capítulo 2, foi presenciada uma
proeminente oscilação teta (6-12 Hz) durante a movimentação do animal no
labirinto linear. Além disso, foram observados picos de potência na frequência
delta (1-4 Hz) durante períodos de maior velocidade (Figura 28-A). Isto é,
constatamos que além dos períodos de delta sem teta que ocorrem quando o
animal encontra-se parado, e de teta sem delta durante locomoção, também havia
períodos de teta associado a delta relacionados ao aumento de velocidade (Figura
28-B).
66

Figura 28: Exemplo de oscilações delta (1-4 Hz) durante a exploração do animal no labirinto linear.
(A) vemos o espectrograma de um canal durante uma tarefa de exploração espacial. A linha preta
representa a velocidade do rato. Note que durante momentos de maior velocidade aparece uma
maior frequência de delta independente do teta. (B) 3 momentos de 2 segundos mostrando a:
trajetória, sinal bruto, sinal filtrado em teta (6-12 Hz) e delta (1-4 Hz). No primeiro momento o
animal encontra-se parado, como pode ser observado na trajetória. O segundo é um período de
movimentação em que se tem alta amplitude das oscilações teta mas não de delta. O terceiro é um
momento de aceleração do animal em que se tem uma alta amplitude tanto de teta como de delta.
A partir dessas observações procuramos conduzir experimentos de forma
controlada em uma esteira, visando encontrar uma maior estabilidade da atividade
delta em relação à velocidade do animal.
3.2 Velocidades constantes
Inicialmente analisamos os três protocolos de velocidades constantes na
esteira. Vimos que tanto a oscilação teta (6-12 Hz) quanto delta (1-4 Hz)
coexistiam nas três velocidades (Figura 29), o que, em primeira instância, parece
67

contradizer a literatura de ritmos hipocampais que separa os estados “alerta” e
“quieto” a partir da oscilação predominante presente (Battaglia et al, 2011; Kirov et
al, 2009).
Em análises iniciais, pensamos que essa oscilação delta poderia ser
artefato de movimento da cabeça, o que rapidamente descartamos ao observar
que o movimento da cabeça do animal oscilava em torno de 6 Hz, como mostrado
na Figura 29. A fim de ver se se trata de um artefato elétrico oriundo do
funcionamento do motor da esteira, fizemos testes com o animal livre na esteira,
onde o motor estava ligado mas sem correia. Observamos que o motor não causa
interferência elétrica nos sinais registrados.
Figura 29: Análise espectral durante três protocolos de velocidades constantes (20, 30 e 40 cm/s).
Espectrograma hipocampal (esquerda) e PSD (meio) dos três protocolos de velocidades
constantes. À direita, o PSD do acelerômetro localizado na cabeça do animal.
Além de tudo, pudemos observar também que a frequência do delta é
variável de acordo com a diferença de velocidade, o que nos fez olhar mais
atenciosamente para os protocolos de velocidades crescentes, como será
mostrado abaixo.
68

3.3 Protocolo de aceleração na esteira
Após observar a atividade delta durante o protocolo de velocidades
constantes, decidimos investigar sua dinâmica durante a variação de velocidade.
Analisando o protocolo de velocidades crescentes na esteira, vimos que a
oscilação do PCL bruto varia de acordo com a velocidade de corrida (Figura 30),
onde o delta em certos momentos pode chegar a ter amplitude maior do que teta.
Figura 30: Análise espectral durante protocolo de aceleração (2.5 cm/s²) na esteira. Protocolo
“ramp up” de 0 a 50 cm/s em 20 s (acima). PCL hipocampal antes e durante a corrida (meio).
Espectrograma e PSD em blocos de 5s mostram o aumento da oscilação delta (abaixo).
Em seguida, analisamos a variação do delta nos diferentes protocolos de
velocidades crescentes (aceleração A1, A2 e A3, Figura 31). Na figura, é possível
observar que a variação da frequência de delta se relaciona com a velocidade de
locomoção instantânea, não pela aceleração. Além disso, é possível notar que o
aumento da potência do delta é mais robusto em acelerações maiores.
69

Figura 31: Análise espectral do PCL hipocampal em três protocolos de aceleração (2, 2.5 e 3
cm/s²). A amplitude e frequência da oscilação delta aumenta de acordo com a velocidade de
locomoção.
Por último, fizemos uma correlação entre o pico do PSD das frequências e
potências normalizadas de delta e teta com a velocidade de locomoção do animal
(Figura 32). Utilizando trials como entrada e analisando em bins de 1 segundo,
nossos resultados mostram que há uma correlação linear positiva (r = 0,29) com a
potência de delta e negativa com teta (r = -0,39), o que sugere inicialmente que
essas bandas se correlacionam de forma inversamente proporcional à velocidade.
Quanto à frequências, observamos que um aumento da velocidade se
correlaciona com a frequência apenas de delta, sugerindo assim que a banda
delta tenha uma relação agregada com a velocidade de locomoção.
70

Figura 32: Pico da frequência e amplitude normalizado em delta (acima) e teta (abaixo) através dos
20 segundos binados. A frequência e amplitude da oscilação delta aumenta com a velocidade,
enquanto a frequência de teta aumenta e a amplitude diminui.
3.4 Frequência respiratória na esteira
Atualmente, é sabido na literatura que aspectos fisiológicos estão
correlacionados com ritmos oscilatórios do cérebro (Fuhrmann et al, 2015;
Ledberg e Robbe, 2011; Lockmann et al, 2016). Dentre esses aspectos, o ritmo
respiratório está associado ao ritmo oscilatório no hipocampo de roedores
(Lockmann et al, 2016; Vivan Chi et al, 2016). Essa particularidade levanta a
hipótese de que o delta visto no hipocampo durante os protocolos de velocidades
constantes e crescentes esteja associado ao ritmo respiratório. Para testar isso,
fizemos um piloto, onde implantamos um thermocouple na cavidade nasal de um
rato Long Evans macho adulto enquanto corria em uma esteira nas velocidades
constantes de 20, 30 e 40 cm/s (Figura 33). O sinal foi registrado a uma taxa de
amostragem de 2030 e filtrado entre 0-15 Hz. Nosso resultado mostra que a
banda de frequência de respiração durante a tarefa nas 3 velocidades constantes
71

é entre 4-6 Hz (Figura 33-B), o que diverge da banda delta observada no
hipocampo durante a corrida em esteira (1-4 Hz). Ainda que esse achado não
tenha sido obtido dos mesmos animais, ele sugere que a oscilação delta no
hipocampo de roedores observada durante corrida em diferentes velocidades não
está associada à atividade do ritmo respiratório.
Figura 33: Frequência respiratória durante a corrida de velocidade constante na esteira. Em (A e B)
temos um exemplo da frequência respiratória do animal durante a corrida nas velocidades
constantes de 20, 30 e 40 cm/s. Em (C) o PSD mostra uma oscilação nas três velocidades,
variando entre 4 e 6 Hz.
3.5 Fase de delta modula a oscilação Low gamma
Em análises iniciais de acoplamento fase-amplitude, também encontramos
um alto índice de modulação entre as fases de delta (1-4 Hz) e a amplitude de LG
(20-55 Hz) em velocidades constantes, o que sugere a priori que o delta tenha
relação com a atividade de CA3. Vimos também que quanto maior é a velocidade,
maior a modulação de delta e LG (Figura 34), o que nos fez olhar para a energia
72
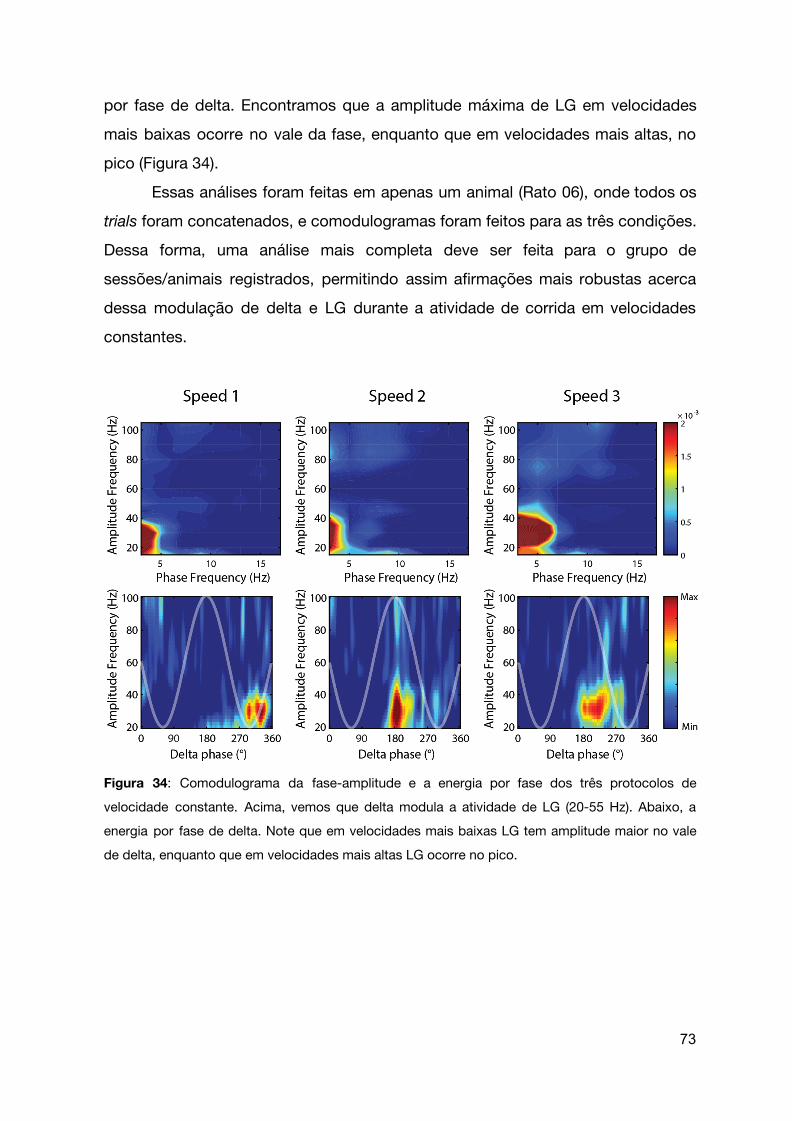
por fase de delta. Encontramos que a amplitude máxima de LG em velocidades
mais baixas ocorre no vale da fase, enquanto que em velocidades mais altas, no
pico (Figura 34).
Essas análises foram feitas em apenas um animal (Rato 06), onde todos os
trials foram concatenados, e comodulogramas foram feitos para as três condições.
Dessa forma, uma análise mais completa deve ser feita para o grupo de
sessões/animais registrados, permitindo assim afirmações mais robustas acerca
dessa modulação de delta e LG durante a atividade de corrida em velocidades
constantes.
Figura 34: Comodulograma da fase-amplitude e a energia por fase dos três protocolos de
velocidade constante. Acima, vemos que delta modula a atividade de LG (20-55 Hz). Abaixo, a
energia por fase de delta. Note que em velocidades mais baixas LG tem amplitude maior no vale
de delta, enquanto que em velocidades mais altas LG ocorre no pico.
73

4. Conclusão
A oscilação delta (proeminente em estados offlines do cérebro) pode
coexistir com a oscilação teta (proeminente em estados onlines) no hipocampo de
ratos em função da velocidade de locomoção. A oscilação delta apresenta
amplitude e frequência relacionadas à velocidade. Também existe modulação da
amplitude de LG pela fase de delta, o que indica que delta tenha relação com a
atividade de CA3. No mais, a origem e função da oscilação delta na locomoção
necessita de mais estudos para um melhor entendimento.
74

V. Discussão _________________________________________________________________________________
Apesar de distintos, os três capítulos dessa dissertação possuem uma
relação dependente que os une de forma sequencial. Observando passo a passo,
essa relação tem início com a intenção do Laboratório de Neurofisiologia
Computacional em implantar a técnica de registro de tetrodos utilizando
microdrives. Visando esse objetivo, nós desenvolvemos um novo modelo de
microdrive barato e eficiente que vem sendo não apenas utilizado por nosso
grupo, mas por outros laboratórios do Instituto do Cérebro da UFRN.
No capítulo 1 apresentamos dois diferentes modelos de microdrives de
tetrodos móveis. Pensando em facilidade e praticidade para futuras produções em
larga escala, seu desenvolvimento se deu a partir da utilização apenas de
ferramentas e máquinas disponíveis no Instituto do Cérebro, tais quais CNC e
impressora 3D, além de parafusos hexagonais e porcas específicas, diferente de
outros modelos da literatura que utilizam materiais exclusivos e comerciais (Voigts
et al, 2013; Battaglia et al, 2009). Além disso, os microdrives disponíveis
atualmente não possuem uma proteção apropriada, no geral improvisados com
algum plástico e papel alumínio. No microdrive elaborado, a arquitetura de
montagem por blocos conta com uma proteção leve e resistente que dá corpo ao
dispositivo. Essa montagem por blocos também ajuda em uma fácil adaptação
para o uso de fibras ópticas e eletrodos de estimulação em conjunto dos tetrodos,
caso seja necessário. Apesar de tudo, nosso modelo ainda se limita à utilização do
conector Omnetics, que é a componente mais cara do sistema hoje em dia. Com
isso, futuras adaptações podem ser feitas para baratear ainda mais sua produção.
Como forma de validar o funcionamento adequado do microdrive,
registramos células de lugar no hipocampo de ratos. Isso faz desse trabalho, o
primeiro projeto voltado para o registro de células de lugar no Brasil . Em 9
adicional, esse dado será utilizado para futuras análises em nosso laboratório para
9 As células de lugar foram descobertas por O’Keefe e Dostrovsky em 1971. Até o momento não há nenhum trabalho publicado que indique registros dessas células no Brasil.
75

um melhor entendimento da formação de mapas espaciais no hipocampo de
ratos.
Apesar de apenas parcialmente analisado, nossos registros reforçam a
existência de células de lugar no hipocampo dorsal de ratos. Alguns trabalhos têm
apontado uma média de 1/3 das células que são registradas no hipocampo como
sendo células de lugar (O’Keefe, 1976; Souza, 2015; Thompson e Best, 1989).
Nosso trabalho, nesse sentido, apresentou uma porcentagem de 25,3% de células
de lugar, o que corrobora com o esperado . Na literatura, a atividade das células 10
de lugar durante a corrida no labirinto linear é separada como sendo unidirecional
ou bidirecional, isso significa que podem apresentar formação do campo de lugar
apenas durante a travessia em um determinado sentido ou em ambos (Battaglia et
al, 2009; Foster e Wilson, 2006; Dragoi e Buzsáki, 2006). Em nossas análises,
também encontramos seletividade de direção, reforçando assim que as células de
lugar apresentam padrões complexos em relação à sua preferência espacial.
Apesar de não ter apresentado um resultado do grupo de animais, vimos também
que as células de lugar em sua maioria apresentam disparos acoplados às fases
do ritmo teta. Esse acoplamento já era esperado, e segundo alguns trabalhos,
pode ter relação com a precessão de fase durante a travessia no campo de lugar,
como mostra Jensen e Lisman (2000), O'Keefe e Reece (1993) e Skaggs e
McNaughton (1996).
Durante os registros, também observamos uma maior ocupância do rato
nas extremidades do labirinto linear. Esse comportamento também foi visto em
outros trabalhos, que acrescentam que durante esse período há reativação das
células de lugar (Foster e Wilson, 2006; Dragoi e Tonegawa, 2011).
Como análises adicionais e também como forma de validação dos registros
pelo modelo de microdrive desenvolvido nesta dissertação, analisamos o PCL de
ratos em diferentes estados comportamentais. Nossos resultados são
semelhantes ao que está descrito na literatura, como: (1) encontramos um
aumento de teta durante o MRO e durante travessia no labirinto linear, além de um
aumento do ritmo delta durante o SOL (Buzsáki, 2002; Mole et al, 2006); (2) houve
10 Vale ainda salientar que, durante a classificação pela métrica de Skaggs, não separamos as corridas por unidirecionalidade, o que implica que essa porcentagem poderia aumentar.
76

um aumento do HG durante a travessia, diferentemente de SOL e MRO (Sheffzuk
et al., 2011); (3) vimos também uma comodulação maior entre teta e HG durante o
sono MRO comparado com os momentos de travessia (Sheffzuk et al, 2011;
Scheffer-Teixeira e Tort, 2017). Durante as análises do PCL nos períodos de
travessia no labirinto linear, vimos que além da esperada oscilação teta no
hipocampo, houveram picos de atividade de delta que se correlacionaram com o
aumento de velocidade do rato. Com isso, dedicamos o capítulo 3 para uma
análise controlada desse ritmo.
Mesmo que sob análises iniciais, os resultados do capítulo 3 apresentam
achados interessantes quanto ao funcionamento oscilatório do hipocampo. Alguns
trabalhos têm definido (Hirase et al, 2001; Scheffer-Teixeira e Tort, 2016) a
atividade eletrofisiológica hipocampal em estados “alerta” e “quieto”, onde, por
natureza, esses estados são supressivos e excludentes entre si. Os estados são
definidos pelas oscilações presentes no hipocampo, sendo o alerta caracterizado
pela atividade de teta e gama, e o quieto pela atividade de delta acompanhada de
eventos de alta frequência chamados de sharp-wave ripple. Nosso achado neste
trabalho indica que essa separação é incongruente quando os animais estão em
uma atividade de locomoção, visto que mostramos que tanto as oscilações teta
quanto delta podem coexistir no hipocampo. Além disso, nossos resultados
indicam que as duas bandas de frequência são afetadas pela velocidade de
locomoção do animal, tanto na amplitude como frequência.
Até hoje, não existe nenhum estudo que mostra correlação entre a
amplitude e frequência de delta em relação à velocidade de locomoção. Ao invés
disso, vários estudos correlacionam locomoção apenas a banda teta e gama
(Fuhrmann et al, 2015; Andersen et al, 2007; McFarland et al, 1975), o que também
é reforçado pelo nosso trabalho, adicionando que encontramos correlações
inversas da potência de teta em relação a velocidade.
Como é sabido, aspectos fisiológicos estão associados a ritmos
oscilatórios no cérebro de roedores, como movimentos da cabeça, frequência
respiratória e outros (Fuhrmann et al, 2015; Ledberg e Robbe, 2011; Lockmann et
al, 2016), dessa maneira, a oscilação delta não apresenta ação específica em
atividade motora. Pesquisas recentes têm reportado uma atividade oscilatória
77

entre 2-4 Hz no giro denteado de animais anestesiados (Yanovsky et al, 2014; Ito
et al, 2014; Lockman et al, 2016). Outros estudos ainda apontam que a frequência
respiratória de um roedor pode variar de 2-10 Hz (Kabir et al, 2010; Nalivaiko et al,
2010). Das possíveis razões da origem do delta encontrado no nosso trabalho,
ainda não podemos afirmar com convicção se ele é oriundo ou não do ritmo
respiratório; para isso, experimentos adicionais devem ser feitos com registros do
PCL de CA1 e da atividade respiratória durante uma tarefa de corrida. Por outro
lado, Alves et al (2016) mostram que a taxa respiratória média das velocidades que
estamos registrando é de 8 Hz, e, juntamente com nosso resultado piloto
mostrando respiração acima de 4 Hz na esteira, supomos que o delta encontrado
na esteira em nossos protocolos de velocidades não seja uma influência
fisiológica direta do ritmo respiratório.
Um outro ponto a se observar é de que achados recentes apontam para um
acoplamento entre o córtex prefrontal e o hipocampo de 2-5 Hz em ratos
anestesiados (Roy et al, 2017). Nesse trabalho, os autores reportam que essa
banda de frequência é oriunda do córtex prefrontal e expressa no hipocampo via o
tálamo. Ao que se conhece na literatura (Giustino et al, 2015; Likhtik et al, 2014), o
córtex prefrontal possui papel crucial codificação de memória de medo. Essa
codificação é reportada por uma oscilação que gira em torno de 4 Hz, sendo
também encontrada na amígdala (Fujisawa, 2017; Karalis et al, 2016 ). O protocolo
de condicionamento aversivo na nossa tarefa com as barras de choque faz pensar
que a oscilação delta encontrada no hipocampo durante a locomoção possa ser
proveniente da oscilação lenta que o prefrontal projeta indiretamente ao
hipocampo. Para comprovar essa hipótese, outros experimentos devem ser feitos
a partir de registros do hipocampo e do córtex prefrontal.
Apesar de já termos feito algumas análises, o presente trabalho ainda
apresenta limitações, como o número de animais analisados até aqui e a falta de
uma análise de grupo. Contudo, os resultados prévios corroboram com uma ideia
nova de definição da atividade de delta e teta no hipocampo de ratos durante a
locomoção.
Em suma, durante o presente trabalho de mestrado foi desenvolvido e
implementado no Instituto do Cérebro da UFRN uma tecnologia ainda incipiente
78

no país de registro de atividade eletrofisiológica através de microdrives de
tetrodos móveis. A efetividade de tal tecnologia foi validada através do registro e
análise da atividade neuronal e de PCL do hipocampo de ratos se movimentando
em um labirinto linear e numa esteira motorizada. Além da caracterização das
células de lugar, já bem estabelecidas na literatura, como co-produto deste
trabalho mostramos também resultados preliminares inéditos acerca da atividade
delta durante locomoção em esteira.
79
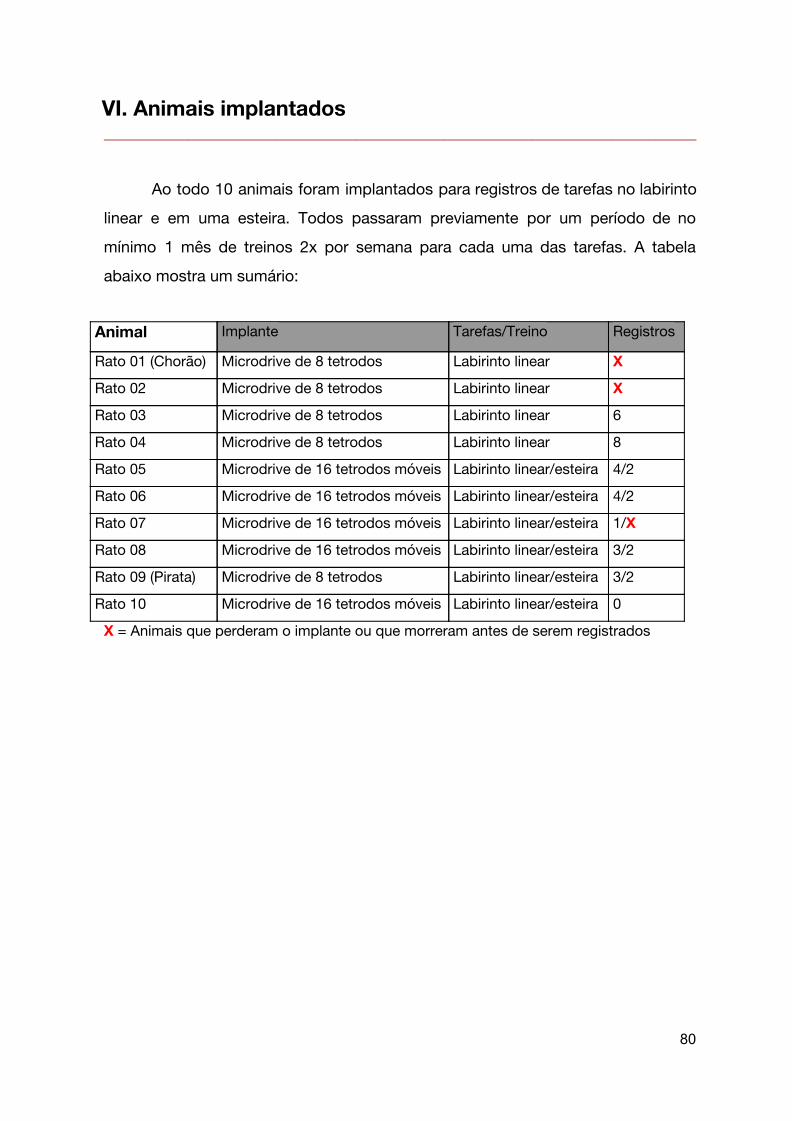
VI. Animais implantados _________________________________________________________________________________
Ao todo 10 animais foram implantados para registros de tarefas no labirinto
linear e em uma esteira. Todos passaram previamente por um período de no
mínimo 1 mês de treinos 2x por semana para cada uma das tarefas. A tabela
abaixo mostra um sumário:
Animal Implante Tarefas/Treino Registros
Rato 01 (Chorão) Microdrive de 8 tetrodos Labirinto linear X
Rato 02 Microdrive de 8 tetrodos Labirinto linear X
Rato 03 Microdrive de 8 tetrodos Labirinto linear 6
Rato 04 Microdrive de 8 tetrodos Labirinto linear 8
Rato 05 Microdrive de 16 tetrodos móveis Labirinto linear/esteira 4/2
Rato 06 Microdrive de 16 tetrodos móveis Labirinto linear/esteira 4/2
Rato 07 Microdrive de 16 tetrodos móveis Labirinto linear/esteira 1/X
Rato 08 Microdrive de 16 tetrodos móveis Labirinto linear/esteira 3/2
Rato 09 (Pirata) Microdrive de 8 tetrodos Labirinto linear/esteira 3/2
Rato 10 Microdrive de 16 tetrodos móveis Labirinto linear/esteira 0
X = Animais que perderam o implante ou que morreram antes de serem registrados
80

VII. Referências _________________________________________________________________________________
Ahmed, Omar J., and Mayank R. Mehta. "Running speed alters the frequency of
hippocampal gamma oscillations." Journal of Neuroscience 32.21 (2012):
7373-7383.
Albeck, D. S., Sano, K., Prewitt, G. E., & Dalton, L. (2006). Mild forced
treadmill exercise enhances spatial learning in the aged rat. Behavioural brain
research, 168(2), 345-348.
Alves, Joseph Andrews, Barbara Ciralli Boerner, and Diego Andrés Laplagne.
"Flexible coupling of respiration and vocalizations with locomotion and head
movements in the freely behaving rat." Neural plasticity 2016 (2016).
Amaral, D. G., and M. P. Witter. "The three-dimensional organization of the
hippocampal formation: a review of anatomical data." Neuroscience 31.3 (1989):
571-591.
Andersen, P., et al. "Septo-hippocampal pathway necessary for dentate theta
production." Brain research 165.1 (1979): 13-22.
Andersen P., Morris R., Amaral D., Bliss T., O’Keefe J. The Hippocampus Book.
New York: Oxford University Press, 2007. 832p.
Battaglia, F. P., Sutherland, G. R. & McNaughton, B. L. Hippocampal sharp wave
bursts coincide with neocortical “up-state” transitions. Learn. Mem. 11, 697–704
(2004).
Battaglia, Francesco P., et al. "The hippocampus: hub of brain network
communication for memory." Trends in cognitive sciences 15.7 (2011): 310-318.
81

Battaglia, Francesco P., et al. "The Lantern: an ultra-light micro-drive for
multi-tetrode recordings in mice and other small animals." Journal of neuroscience
methods 178.2 (2009): 291-300.
Battaglia, Francesco P., Gary R. Sutherland, and Bruce L. McNaughton. "Local
sensory cues and place cell directionality: additional evidence of prospective
coding in the hippocampus." Journal of Neuroscience 24.19 (2004): 4541-4550.
Belchior H, Lopes-Dos- Santos V, Tort AB, Ribeiro S (2014). Increase in
hippocampal theta oscillations during spatial decision making. Hippocampus,
24(6), 693-702.
Benchenane, K., Peyrache, A., Khamassi, M., Tierney, P. L., Gioanni, Y., Battaglia,
F. P., & Wiener, S. I. (2010). Coherent theta oscillations and reorganization of
spike timing in the hippocampal-prefrontal network upon learning. Neuron, 66(6),
921-936.
Börgers, C. & Kopell, N. Synchronization in networks of excitatory and inhibitory
neurons with sparse, random connectivity. Neural. Comput. 15, 509–538 (2003).
Bott, Jean-Bastien, et al. "Spatial Reference Memory is Associated with
Modulation of Theta–Gamma Coupling in the Dentate Gyrus." Cerebral Cortex 26.9
(2016): 3744-3753.
Buzsáki, György, and Andreas Draguhn. "Neuronal oscillations in cortical
networks." science 304.5679 (2004): 1926-1929.
Buzsáki, György, and Edvard I. Moser. "Memory, navigation and theta rhythm in
the hippocampal-entorhinal system." Nature neuroscience 16.2 (2013): 130-138.
82

Buzsáki, György. "Theta oscillations in the hippocampus." Neuron 33.3 (2002):
325-340.
Buzsaki, Gyorgy. Rhythms of the Brain. Oxford University Press, 2006.
Carr, Margaret F., Shantanu P. Jadhav, and Loren M. Frank. "Hippocampal replay
in the awake state: a potential substrate for memory consolidation and retrieval."
Nature neuroscience 14.2 (2011): 147-153.
Chi, Vivan Nguyen, et al. "Hippocampal respiration-driven rhythm distinct from
theta oscillations in awake mice." Journal of Neuroscience 36.1 (2016): 162-177.
Colgin, Laura Lee, and Edvard I. Moser. "Gamma oscillations in the hippocampus."
Physiology 25.5 (2010): 319-329.
Colgin, Laura Lee, et al. "Frequency of gamma oscillations routes flow of
information in the hippocampus." Nature 462.7271 (2009): 353.
Csicsvari, Jozsef, et al. "Mechanisms of gamma oscillations in the hippocampus of
the behaving rat." Neuron 37.2 (2003): 311-322.
Csicsvari, Jozsef, et al. "Reliability and state dependence of pyramidal
cell–interneuron synapses in the hippocampus: an ensemble approach in the
behaving rat." Neuron 21.1 (1998): 179-189.
Da Silva, Fernando Lopes. "EEG: origin and measurement." EEg-fMRI. Springer
Berlin Heidelberg, 2009. 19-38.
Dragoi, George, and György Buzsáki. "Temporal encoding of place sequences by
hippocampal cell assemblies." Neuron 50.1 (2006): 145-157.
83

Dragoi, George, and Susumu Tonegawa. "Preplay of future place cell sequences
by hippocampal cellular assemblies." Nature 469.7330 (2011): 397.
Ego Stengel, Valérie, and Matthew A. Wilson. "Disruption of ripple‐associated
hippocampal activity during rest impairs spatial learning in the rat."Hippocampus
20.1 (2010): 1-10.
Eichenbaum, Howard. "A cortical–hippocampal system for declarative memory."
Nature Reviews Neuroscience 1.1 (2000): 41-50.
Ferguson, John E., Chris Boldt, and A. David Redish. "Creating low-impedance
tetrodes by electroplating with additives." Sensors and Actuators A: Physical 156.2
(2009): 388-393.
Foster, David J., and Matthew A. Wilson. "Reverse replay of behavioural
sequences in hippocampal place cells during the awake state." Nature 440.7084
(2006): 680-683.
Freund, Tamas F., and Gyorgi Buzsáki. "Interneurons of the hippocampus."
Hippocampus 6.4 (1996): 347-470.
Fuhrmann, F., Justus, D., Sosulina, L., Kaneko, H., Beutel, T., Friedrichs, D.,
Schoch S., Schwarz M.K., Fuhrmann M. & Remy, S. (2015). Locomotion, theta
oscillations, and the speed-correlated firing of hippocampal neurons are controlled
by a medial septal glutamatergic circuit. Neuron, 86(5), 1253-1264.
Fujisawa, Shigeyoshi. "Slow Oscillation in Prefrontal Cortex Underlying Local
Computations and Large-Scale Interactions." The Physics of the Mind and Brain
Disorders. Springer, Cham, 2017. 233-245.
Girardeau, Gabrielle, and Michaël Zugaro. "Hippocampal ripples and memory
consolidation." Current opinion in neurobiology 21.3 (2011): 452-459.
84

Girardeau, Gabrielle, et al. "Selective suppression of hippocampal ripples impairs
spatial memory." Nature neuroscience 12.10 (2009): 1222-1223.
Giustino, Thomas F., and Stephen Maren. "The role of the medial prefrontal cortex
in the conditioning and extinction of fear." Frontiers in behavioral neuroscience 9
(2015).
Golgi, Camillo, Marina Bentivoglio, and L. Swanson. "On the fine structure of the
pes Hippocampi major (with plates XIII-XXIII). 1886." Brain research bulletin 54.5
(2001): 461-483.
Hafting, Torkel, et al. "Microstructure of a spatial map in the entorhinal
cortex." Nature 436.7052 (2005): 801-806.
Harris, Kenneth D., et al. "Organization of cell assemblies in the hippocampus."
Nature 424.6948 (2003): 552-556.
Headley, Drew B., and Denis Paré. "Common oscillatory mechanisms across
multiple memory systems." npj Science of Learning 2.1 (2017): 1.
Hebb, Donald O. "The organization of behavior." (1949).
Hirase, Hajime, et al. "Behavior-dependent states of the hippocampal network
affect functional clustering of neurons." Journal of Neuroscience 21.10 (2001):
RC145-RC145.
Igarashi, Kei M., et al. "Coordination of entorhinal–hippocampal ensemble activity
during associative learning." Nature 510.7503 (2014): 143.
85

Insausti, Ricardo, M. Herrero, and Menno P. Witter. "Entorhinal cortex of the rat:
cytoarchitectonic subdivisions and the origin and distribution of cortical efferents."
Hippocampus 7.2 (1997): 146-183.
Insausti, Ricardo. "Comparative anatomy of the entorhinal cortex and
hippocampus in mammals." Hippocampus 3.S1 (1993): 19-26.
Isomura, Y. et al. Integration and segregation of activity in entorhinal-hippocampal
subregions by neocortical slow oscillations. Neuron 52, 871–882 (2006).
Ito J., Roy S., Liu Y., Cao Y., Fletcher M., Lu L., Boughter J.D., Grün S. ; Heck D.H.
(2014). Whisker barrel cortex delta oscillations and gamma power in the awake
mouse are linked to respiration. Nature communications, 5:3572.
Jensen, Ole, and John E. Lisman. "Position reconstruction from an ensemble of
hippocampal place cells: contribution of theta phase coding." Journal of
neurophysiology 83.5 (2000): 2602-2609.
Jensen O, Lisman JE (2005) Hippocampal sequence-encoding driven by a cortical
multi-item working memory buffer. Trends in neurosciences 28:67-72.
Kabir, Muammar M., et al. "Respiratory pattern in awake rats: effects of motor
activity and of alerting stimuli." Physiology & behavior 101.1 (2010): 22-31.
Kajikawa, Yoshinao, and Charles E. Schroeder. "How local is the local field
potential?." Neuron 72.5 (2011): 847-858.
Karalis, Nikolaos, et al. "4-Hz oscillations synchronize prefrontal-amygdala circuits
during fear behavior." Nature neuroscience 19.4 (2016): 605-612.
86

Kirov, Roumen, et al. "Slow oscillation electrical brain stimulation during waking
promotes EEG theta activity and memory encoding." Proceedings of the National
Academy of Sciences 106.36 (2009): 15460-15465.
Klausberger, Thomas, and Peter Somogyi. "Neuronal diversity and temporal
dynamics: the unity of hippocampal circuit operations." Science 321.5885 (2008):
53-57.
Koepsell, Kilian, et al. "Retinal oscillations carry visual information to cortex."
Frontiers in systems neuroscience 3 (2009): 4.
Kuo, T. B. J., et al. "Changes in hippocampal theta activity during initiation and
maintenance of running in the rat." Neuroscience 194 (2011): 27-35.
Ledberg, Anders, and David Robbe. "Locomotion-related oscillatory body
movements at 6–12 Hz modulate the hippocampal theta rhythm." PloS one 6.11
(2011): e27575.
Lee, M. G., Chrobak, J. J., Sik, A., Wiley, R. G. & Buzsáki, G. Hippocampal theta
activity following selective lesion of the septal cholinergic system. Neuroscience
62, 1033–1047 (1994).
Li, Jia-Yi, et al. "Voluntary and involuntary running in the rat show different patterns
of theta rhythm, physical activity, and heart rate." Journal of neurophysiology
111.10 (2014): 2061-2070.
Likhtik, Ekaterina, et al. "Prefrontal entrainment of amygdala activity signals safety
in learned fear and innate anxiety." Nature neuroscience 17.1 (2014): 106-113.
Liu, Yu‐Fan, et al. "Differential effects of treadmill running and wheel running on
spatial or aversive learning and memory: roles of amygdalar brain‐derived
87

neurotrophic factor and synaptotagmin I." The Journal of physiology 587.13 (2009):
3221-3231.
Lockmann, André LV, et al. "A respiration-coupled rhythm in the rat hippocampus
independent of theta and slow oscillations." Journal of Neuroscience 36.19 (2016):
5338-5352.
Louie, Kenway, and Matthew A. Wilson. "Temporally structured replay of awake
hippocampal ensemble activity during rapid eye movement sleep."Neuron 29.1
(2001): 145-156.
McFarland, Willard L., Herman Teitelbaum, and Elizabeth K. Hedges. "Relationship
between hippocampal theta activity and running speed in the rat." Journal of
comparative and physiological psychology 88.1 (1975): 324.
McNaughton, Bruce L., John O'Keefe, and Carol A. Barnes. "The stereotrode: a
new technique for simultaneous isolation of several single units in the central
nervous system from multiple unit records." Journal of neuroscience methods 8.4
(1983): 391-397.
Molle, Matthias, et al. "Hippocampal sharp wave-ripples linked to slow oscillations
in rat slow-wave sleep." Journal of neurophysiology 96.1 (2006): 62-70
Montgomery, Sean M., Anton Sirota, and György Buzsáki. "Theta and gamma
coordination of hippocampal networks during waking and rapid eye movement
sleep." Journal of Neuroscience 28.26 (2008): 6731-6741.
Moser E, Moser M, O’Keefe J. “The Nobel Prize in Physiology or Medicine 2014".
Nobelprize.org. Nobel Media AB 2014. Web. 13 Apr 2016.
<http://www.nobelprize.org/nobel_prizes/medicine/laureates/2014/>”.
88

Nalivaiko, Eugene, et al. "Dorsomedial hypothalamus and medullary raphe mediate
respiratory arousal responses in rats." The FASEB Journal 24.1 Supplement (2010):
1019-16.
O'Keefe, John, and Lynn Nadel. The hippocampus as a cognitive map. Vol. 3.
Oxford: Clarendon Press, 1978.
O'Keefe, John, and Michael L. Recce. "Phase relationship between hippocampal
place units and the EEG theta rhythm." Hippocampus 3.3 (1993): 317-330.
O'Keefe, John. "Place units in the hippocampus of the freely moving rat."
Experimental neurology 51.1 (1976): 78-109.
O'Keefe J, Dostrovsky J (1971) The hippocampus as a spatial map: Preliminary
evidence from unit activity in the freely-moving rat. Brain research.
O’Callaghan, R. M., Ohle, R., & Kelly, A. M. (2007). The effects of forced
exercise on hippocampal plasticity in the rat: A comparison of LTP, spatial-and
non-spatial learning. Behavioural brain research, 176(2), 362-366.
Paxinos, G., and C. Watson. "Compact, 3rd edn: The Rat Brain in Stereotaxic
Coordinates." (1997).
Quilichini, P., Sirota, A. & Buzsáki, G. Intrinsic circuit organization and
theta-gamma oscillation dynamics in the entorhinal cortex of the rat. J. Neurosci.
30, 11128–11142 (2010).
Roy, Alexis, et al. "Prefrontal-hippocampal coupling by theta rhythm and by 2–5 Hz
oscillation in the delta band: The role of the nucleus reuniens of the thalamus."
Brain Structure and Function (2017): 1-12.
89

Sainsbury, Robert S., and Brian H. Bland. "The effects of selective septal lesions
on theta production in CA1 and the dentate gyrus of the hippocampus."
Physiology & behavior 26.6 (1981): 1097-1101.
Scheffer-Teixeira, Robson, and Adriano BL Tort. "On cross-frequency
phase-phase coupling between theta and gamma oscillations in the
hippocampus." eLife 5 (2016): e20515.
Scheffer-Teixeira, Robson, and Adriano BL Tort. "Unveiling fast field oscillations
through comodulation." eNeuro 4.4 (2017): ENEURO-0079.
Scheffer-Teixeira, Robson, et al. "On high-frequency field oscillations (> 100 Hz)
and the spectral leakage of spiking activity." The Journal of Neuroscience 33.4
(2013): 1535-1539.
Scheffer-Teixeira, Robson, et al. "Theta phase modulates multiple layer-specific
oscillations in the CA1 region." Cerebral Cortex (2011): bhr319.
Scheffzük, Claudia, et al. "Selective coupling between theta phase and neocortical
fast gamma oscillations during REM-sleep in mice." PLoS One 6.12 (2011):
e28489.
Scoville, William Beecher, and Brenda Milner. "Loss of recent memory after
bilateral hippocampal lesions." Journal of Neurology, Neurosurgery &
Psychiatry 20.1 (1957): 11-21.
Senior, Timothy J., et al. "Gamma oscillatory firing reveals distinct populations of
pyramidal cells in the CA1 region of the hippocampus." The Journal of
Neuroscience 28.9 (2008): 2274-2286.
Skaggs, William E., and Bruce L. McNaughton. "Theta phase precession in
hippocampal." Hippocampus 6 (1996): 149-172.
90

Skaggs, William E., Bruce L. McNaughton, and Katalin M. Gothard. "An
information-theoretic approach to deciphering the hippocampal code." Advances
in neural information processing systems. 1993.
Souza, Bryan C., et al. "Spike sorting with Gaussian mixture models." bioRxiv
(2018): 248864.
Souza, Bryan da Costa. Caracterização de células de lugar no hipocampo e de
suas relações com oscilações do potencial de campo local. MS thesis.
Universidade Federal do Rio Grande do Norte, 2015.
Steriade, M., Nuñez, A. & Amzica, F. A novel slow (< 1 Hz) oscillation of neocortical
neurons in vivo: depolarizing and hyperpolarizing components. J. Neurosci. 13,
3252–3265 (1993).
Sławińska, Urszula, and Stefan Kasicki. "The frequency of rat's hippocampal theta
rhythm is related to the speed of locomotion." Brain research 796.1-2 (1998):
327-331.
Thompson, L. T., and P. J. Best. "Place cells and silent cells in the hippocampus of
freely-behaving rats." Journal of Neuroscience 9.7 (1989): 2382-2390.
Tort, Adriano BL, et al. "Theta–gamma coupling increases during the learning of
item–context associations." Proceedings of the National Academy of Sciences
106.49 (2009): 20942-20947.
van Groen, Thomas, Pasi Miettinen, and Inga Kadish. "The entorhinal cortex of the
mouse: organization of the projection to the hippocampal formation."
Hippocampus 13.1 (2003): 133-149.
91

Verkhratsky, Alexei, Oleg A. Krishtal, and Ole H. Petersen. "From Galvani to patch
clamp: the development of electrophysiology." Pflügers Archiv 453.3 (2006):
233-247.
Voigts, Jakob, et al. "The flexDrive: an ultra-light implant for optical control and
highly parallel chronic recording of neuronal ensembles in freely moving mice."
Frontiers in systems neuroscience 7 (2013): 8.
Wang, X. J. & Buzsáki, G. Gamma oscillation by synaptic inhibition in a
hippocampal interneuronal network model. J. Neurosci. 16, 6402–6413 (1996).
Wheeler, Diek W., et al. "Hippocampome. org: a knowledge base of neuron types
in the rodent hippocampus." Elife 4 (2015).
Whishaw, Ian Q., and Bryan Kolb, eds. The behavior of the laboratory rat: a
handbook with tests. Oxford University Press, 2004.
Wilson, Matthew A., and Bruce L. McNaughton. "Reactivation of hippocampal
ensemble memories during sleep." Science 265.5172 (1994): 676-679.
Wilson MA, McNaughton BL (1994) Reactivation of hippocampal ensemble
ssociations." Proceedings of the National Academy of Sciences 106.49 (2009):
20942-20947.
Wood, Emma R., et al. "Hippocampal neurons encode information about different
types of memory episodes occurring in the same location." Neuron 27.3 (2000):
623-633.
Yamamoto, Jun, and Matthew A. Wilson. "Large-scale chronically implantable
precision motorized microdrive array for freely behaving animals." Journal of
neurophysiology 100.4 (2008): 2430-2440.
92

Yanovsky, Y., Ciatipis, M., Draguhn, A., Tort, A. B., & Brankačk, J. (2014).
Slow oscillations in the mouse hippocampus entrained by nasal respiration.
Journal of Neuroscience, 34(17), 5949-5964.
Ylinen, A. et al. Sharp wave-associated high-frequency oscillation (200 Hz) in the
intact hippocampus: network and intracellular mechanisms. J. Neurosci. 15, 30–46
(1995).
93

VIII. Anexos: _________________________________________________________________________________
1. Anexo 1:
94

2. Anexo 2: Poster apresentado no House Symposium do Instituto do cérebro
em dezembro de 2016
95

3. Anexo 3: Colaboração em um capítulo de livro da editora Elsevier
96

4. Anexo 4: Desenvolvimento de um tutorial para a manufatura de microdrives
97

5. Anexo 5: Desenvolvimento de um sistema de realidade virtual (VR) para
roedores.
Aparte de tudo, comecei a desenvolver um projeto de realidade virtual (VR)
para experimento com roedores. Inicialmente a ideia era desenvolver um aparato
experimental de registro head fixed com roedores em uma superfície esférica,
onde um projetor preso na superfície reproduz a imagem para uma plataforma
côncava de pano. Um sistema de recompensa foi pensado para ficar preso
próximo da boca do animal. O ambiente virtual pensado foi um labirinto linear
rodeado de dicas espaciais para facilitar a localização do animal.
O labirinto linear foi desenvolvido em Unity/C#, que é uma linguagem muito
utilizada no desenvolvimento de jogos, o que proporciona uma boa realidade e
uma programação fácil de Inputs e outputs em hardwares (Figura abaixo).
98

6. Anexo 6: Poster apresentado no House Symposium do Instituto do Cérebro
em dezembro de 2017.
99

7. Anexo 7: Aparelho de privação de sono.
Em colaboração com Daniel Filho, aluno de doutorado do Laboratório de
Memória e Sono, ajudei a desenvolver um aparelho de privação de sono
programado manualmente pelo hardware. O sistema possui um algoritmo de
escolha de protocolo que permite selecionar tanto uma rotina já preexistente de
privação de horas de sono como uma escolha manual de horas de privação
(Figura abaixo). O sistema também possui um input para o MATLAB® que permite
a escolha do tipo de sono (ex: SOL, MRO e fase de transição) a ser privado.
O aparelho é composto por duas partes, a primeira é uma caixa revestida
por acrílico com dois motores na superfície. A segunda parte é onde se pode
programar o aparelho manualmente para algum protocolo e é composta por um
Arduino due, fonte 24 V, 2 relés, LCD, keyboard, 1 transistor, 2 conversores de
voltagem e um RTC (Real Time Clock) (Figura abaixo). A privação de sono do
animal ocorre a partir do acionamento dos motores que movimentam a caixa em 2
cm no intervalo de 100 ms. Os intervalos desses acionamentos ocorre a partir de
um dos protocolos escolhidos manualmente ou por meio do MATLAB®.
100

Algoritmo de programação: O algoritmo foi programado em um Arduino para que o pesquisador
escolha o protocolo de privação.
101

Aparelho de privação de sono: O aparelho é composto pela caixa de onde o animal vai dormir e
pelo hardware de programação.
102




















