
Initial diversity in sheep and goat management in Neolithic south-western Asia
17
Initial diversity in sheep and goat management in Neolithic south-western Asia Benjamin S. Arbuckle and Levent Atici In this paper we survey a large body of faunal data for the practice of young male culling in Neolithic south-western Asia. Although the young male kill-off model is one of the most widely used models for identifying animal domestication in Neolithic south-western Asia, its ubiquity has never been addressed on a regional scale. By focusing on a combination of kill-off age and the shape of the distributions of biometric data, we are able to address the emergence and ubiquity of young male culling amongst Neolithic sheep and goat herders. Although the intensive culling of young males has been presented as a ‘leading edge marker’ for the initiation of sheep and goat herding, we find that clear evidence for young male kill-off appears in the faunal record only in the early 8th millennium cal BC — considerably later than the origins of caprine management. Instead, Neolithic caprine management practices appear to have been characterized by a high degree of ‘initial diversity’, especially in the 9th and early 8th millennia, suggesting that early management strategies may have been much more varied than previously realized. However, after c. 7500 cal BC young male kill-off was widely practised across south-western Asia, suggesting this efficient and effective management technology quickly replaced the diversity of local management strategies prevalent earlier. Keywords: Neolithic, sheep, goats, animal domestication, young male culling, management Introduction The origins of domesticated plants and animals represents one of the most consequential technologi- cal transformations in human history and has there- fore been a major focus of archaeological research for more than a century (Binford 1968; Childe 1936; Pumpelly 1908). Beginning with the observation that domesticates exhibit a distinctive set of phenotypes, often including smaller size, compared to their wild ancestors, early researchers focused on using skeletal measurements to identify the origins of domestic animals (Ba ¨umler 1921; Hammer 1984; Uerpmann 1979; Winge 1900). By the 1970s, a series of more comprehensive and varied lines of evidence for identifying the process of animal domestication was developed, including the use of demographic profiles to interpret culling practices associated with intentional human management (Bo ¨ko ¨ nyi 1969; Ducos 1978; Hesse 1978; Meadow 1989). Recently, increasingly sophisticated methods have been added to the arsenal that archaeologists have at their disposal to address the origins of animal husbandry, including isotopic and phytolith evidence for foddering and manipulation of herd mobility, geochemical evidence for penning and the use of animal dung, morphometric approaches to population dispersals, as well as the results of paleogenetic studies (Ludwig et al. 2009; Makarewicz and Tuross 2012; Meiggs 2010; Naderi et al. 2008; Ottoni et al. 2012; Pearson et al. 2007). Together, the application of these methods has resulted in tremendous progress mapping out the broad chronological and spatial patterns for the origins and spread of domestic animals (Peters et al. 2013; Vigne et al. 2011; Zeder 2008b). Despite this progress, however, the early stages of the develop- ment and spread of herding economies are still poorly understood, and basic questions such as ‘what husbandry methods did the earliest herders use’ and ‘how did herd management technologies spread’ are Benjamin S. Arbuckle (corresponding author), Department of Anthropology, University of North Carolina at Chapel Hill, CB#3115, 301 Alumni Building, Chapel Hill, NC 27599-3115, USA; email: bsarbu@ email.unc.edu. Levent Atici, Department of Anthropology, University of Nevada, Las Vegas, NV 89154-5003, USA; email: levent.atici@unlv. ß Council for British Research in the Levant 2013 Published by Maney DOI 10.1179/0075891413Z.00000000026 Levant 2013 VOL 45 NO 2 219
Transcript of Initial diversity in sheep and goat management in Neolithic south-western Asia

Initial diversity in sheep and goatmanagement in Neolithic south-western Asia
Benjamin S. Arbuckle and Levent Atici
In this paper we survey a large body of faunal data for the practice of young male culling in
Neolithic south-western Asia. Although the young male kill-off model is one of the most widely
used models for identifying animal domestication in Neolithic south-western Asia, its ubiquity has
never been addressed on a regional scale. By focusing on a combination of kill-off age and the
shape of the distributions of biometric data, we are able to address the emergence and ubiquity
of young male culling amongst Neolithic sheep and goat herders. Although the intensive culling of
young males has been presented as a ‘leading edge marker’ for the initiation of sheep and goat
herding, we find that clear evidence for young male kill-off appears in the faunal record only in the
early 8th millennium cal BC — considerably later than the origins of caprine management.
Instead, Neolithic caprine management practices appear to have been characterized by a high
degree of ‘initial diversity’, especially in the 9th and early 8th millennia, suggesting that early
management strategies may have been much more varied than previously realized. However,
after c. 7500 cal BC young male kill-off was widely practised across south-western Asia,
suggesting this efficient and effective management technology quickly replaced the diversity of
local management strategies prevalent earlier.
Keywords: Neolithic, sheep, goats, animal domestication, young male culling, management
Introduction
The origins of domesticated plants and animals
represents one of the most consequential technologi-
cal transformations in human history and has there-
fore been a major focus of archaeological research for
more than a century (Binford 1968; Childe 1936;
Pumpelly 1908). Beginning with the observation that
domesticates exhibit a distinctive set of phenotypes,
often including smaller size, compared to their wild
ancestors, early researchers focused on using skeletal
measurements to identify the origins of domestic
animals (Baumler 1921; Hammer 1984; Uerpmann
1979; Winge 1900). By the 1970s, a series of more
comprehensive and varied lines of evidence for
identifying the process of animal domestication
was developed, including the use of demographic
profiles to interpret culling practices associated with
intentional human management (Bokonyi 1969;
Ducos 1978; Hesse 1978; Meadow 1989). Recently,
increasingly sophisticated methods have been added
to the arsenal that archaeologists have at their disposal
to address the origins of animal husbandry, including
isotopic and phytolith evidence for foddering and
manipulation of herd mobility, geochemical evidence
for penning and the use of animal dung, morphometric
approaches to population dispersals, as well as the
results of paleogenetic studies (Ludwig et al. 2009;
Makarewicz and Tuross 2012; Meiggs 2010; Naderi et al.
2008; Ottoni et al. 2012; Pearson et al. 2007).
Together, the application of these methods has
resulted in tremendous progress mapping out the
broad chronological and spatial patterns for the
origins and spread of domestic animals (Peters et al.
2013; Vigne et al. 2011; Zeder 2008b). Despite this
progress, however, the early stages of the develop-
ment and spread of herding economies are still poorly
understood, and basic questions such as ‘what
husbandry methods did the earliest herders use’ and
‘how did herd management technologies spread’ are
Benjamin S. Arbuckle (corresponding author), Department ofAnthropology, University of North Carolina at Chapel Hill, CB#3115, 301Alumni Building, Chapel Hill, NC 27599-3115, USA; email: [email protected]. Levent Atici, Department of Anthropology, University ofNevada, Las Vegas, NV 89154-5003, USA; email: levent.atici@unlv.
� Council for British Research in the Levant 2013Published by ManeyDOI 10.1179/0075891413Z.00000000026 Levant 2013 VOL 45 NO 2 219

yet to be answered. This is partly the result of
limitations in the application of faunal data and
traditional lines of evidence to address the nature of
the earliest animal management strategies. For
example, researchers have shown that changes in
phenotype, including decrease in size and transfor-
mation of horn morphology in domestic bovids,
likely occurred centuries after the initiation of
intensive management regimes (i.e. herding), thus
making it difficult to use these phenotypic markers to
identify the earliest phases in the establishment of
animal husbandry practices (Zeder 2006). In addi-
tion, interpretations of demographic evidence for
herd management are often hindered by widespread
variations in the methods, localities and seasonality
of hunting practices, which may target a range of
demographic cohorts especially in wild bovids
characterized by dramatic seasonal changes in the
territoriality and demographic composition of social
groups (Schaller 1977; Simmons and Ilany 1975–77;
Watson 1978). The common, and often necessary,
analytical practice of combining faunal data into
broad multi-component assemblages, often creates
unintelligible palimpsests of evidence for animal
exploitation, thus making interpretation difficult
(Halstead 1998). Compounding these problems is
the fact that faunal researchers have been slow to
apply new methods which target evidence for specific
husbandry practices including foddering, stalling and
herd mobility, in favour of approaches seeking to
broadly characterize the ‘status’ of animal populations
as wild or domestic (although see Henton 2012;
Makarewicz and Tuross 2012; Vanpoucke et al. 2009).
Some of the most productive recent research
documenting the origins of animal management has
focused on sheep and goats, which may have been the
earliest domesticated food animals (Peters et al. 2005).
Current research suggests that these animals were
brought under human control in a process that began
sometime in the 9th millennium cal BC in a region
extending from south-eastern Turkey to north-western
Iran (Zeder 2011). One of the most effective methods
for documenting this initiation of animal husbandry has
focused on identifying a combination of sex and age
specific culling practices known as young male kill-off.
Payne (1973) has defined the most widely used
model describing young male kill-off as a typical herd
management strategy used by sheep and goat herders.
In this model, surplus immature males are the
primary target of slaughter, while females are
generally culled as adults when their reproductive
potential begins to decline. As a result of this
management system, faunal assemblages exhibit clear
biometric and demographic patterns. Because goats
and, to a lesser extent, sheep are characterized by
sexual dimorphism, and because males achieve most of
their larger body size within the first year (Davis 2000;
Zeder and Hesse 2000), young male kill-off produces
diagnostic patterns in which most immature specimens
represent large males, while most adult specimens
represent smaller females. These patterns can be
identified through the analysis of breadth and depth
measurements of postcranial skeletal elements which
bear evidence for epiphyseal fusion thus providing
evidence for both the age and sex of culled individuals.
Zeder and Hesse (2000; Zeder 2008b) have
effectively used this combined biometric-demo-
graphic approach to identify the earliest management
of morphologically wild goats in western Iran in the
early 8th millennium BC at the site of Ganj Dareh.
Expanding the use of this approach, Zeder has
succeeded in mapping out the spread of goat and
also sheep management practices across the Zagros
region. As a result of this innovative and detailed
work, it is argued that young male kill-off, identified
through this combined biometric-demographic
approach, is the best ‘leading edge marker’ for the
origins of animal management (Zeder 2006).
Curiously, and despite the effectiveness of this
approach, it has rarely been applied outside the
Zagros region (although see Arbuckle et al. 2009;
Makarewicz 2009), and there has never been a
comprehensive test for the timing and ubiquity of
evidence for young male kill-off in the archaeofaunal
record of Neolithic south-western Asia. As a result, it
is currently not clear how ubiquitous young male kill-
off was as a method for managing Neolithic herds of
sheep and goat or when and where it first emerged as
a management strategy.
In this paper, we provide the first regional scale test
for evidence for the practice of young male kill-off in
Neolithic south-western Asia. We focus on a survey
of published biometric and demographic data from
78 assemblages from across the region, particularly
those dating to the 10th through 8th millennia BC
(PPNA and PPNB periods in the Levantine chron-
ological framework), with the goal of identifying
when and where the practice of young male kill-off is
evident and characterizing its ubiquity as an early
strategy of herd management (Fig. 1; see Tables 1
and 2 for lists of assemblages).
Methods: identifying young male kill-off in thearchaeofaunal record
In order to identify evidence for the management
strategy of young male kill-off, we focus on published
Arbuckle and Atici Diversity in sheep and goat management
220 Levant 2013 VOL 45 NO 2

biometric data and, secondarily, age data for both
sheep and goats. Biometric data are ideal for identify-
ing evidence for young male kill-off in caprines for
several reasons. First, they have the advantage that
they are published for a large number of Neolithic
assemblages using a reliable and standardized format
either as raw measurements (following von den
Driesch 1976) or transformed using the Log Size
Index (LSI) method (Meadow 1999).
Secondly, in sexually dimorphic taxa such as goats
and, to a lesser extent, sheep, males and females can
be distinguished based on the breadth and depth
measurements of long bones (Davis 2000; Zeder and
Hesse 2000). For goats (Capra aegagrus/hircus),
which exhibit a relatively high degree of dimorphism,
it is often possible to attribute most measurements of
long bone epiphyses to a particular sex, and
researchers have used several statistical methods
including Mixture analysis (Monchot 1999;
Monchot and Lechelle 2002) and the Mahalanobis
distance between known male and female samples
(Zeder 2001) to achieve this result with high
confidence intervals.
Sheep (Ovis orientalis/aries) exhibit a lesser degree
of sexual dimorphism than do goats (Davis 2000;
Vigne 2011b). There is a greater degree of overlap in
the size of males and females in bone measurements
and it is therefore more difficult to attribute
individual measurements to a particular sex.
However, dimorphism in some measurements includ-
ing the distal breadth of the metacarpus (also scapula
SLC, tibia Bd, metatarsal Bd, radius Bp, humerus
BT, calcaneum GL) is significant within both
domestic (t-test; p,0?001; N526) (Davis 2000) and
wild populations (t-test; p50?004; N526; based on
recent specimens from Iran curated in the Field
Museum, Chicago) (Fig. 2). Although the degree of
dimorphism in sheep is limited, it is possible to
generate general estimates of the proportions of
males and females in archaeofaunal populations by
examining the shape (skewness and kurtosis) of the
distribution produced by biometric data utilizing
Figure 1 Map showing the location of sites mentioned in the text
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 221
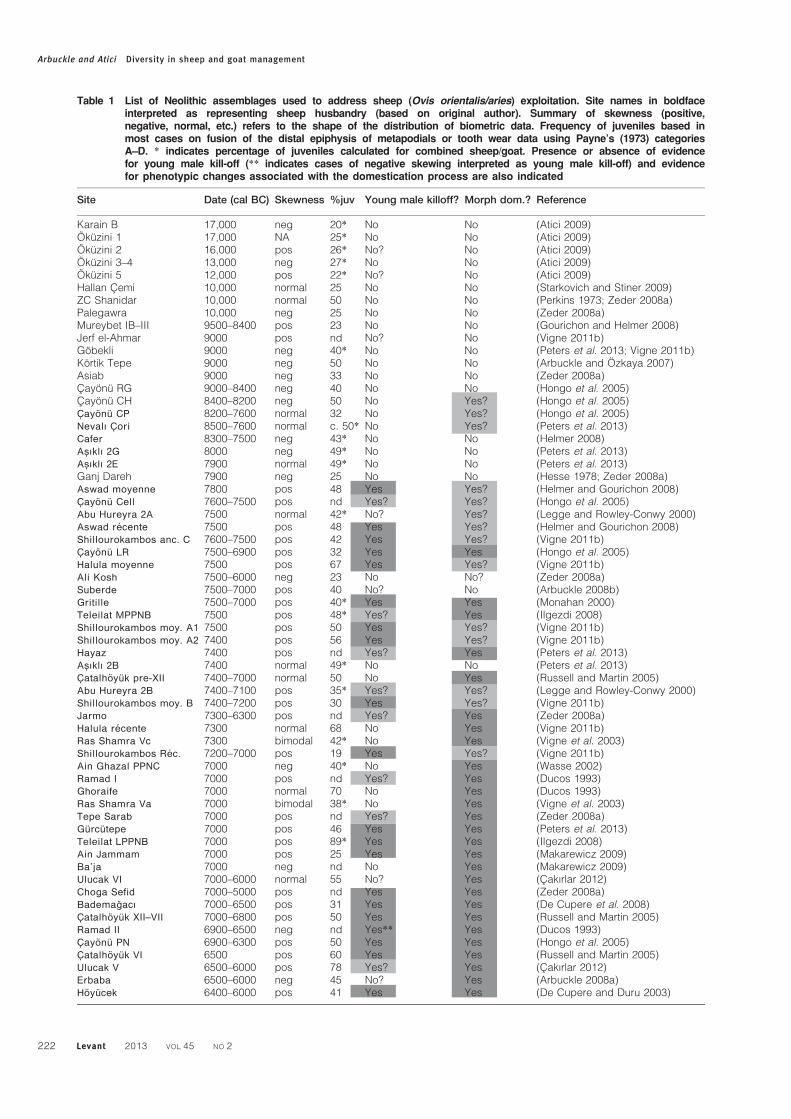
Table 1 List of Neolithic assemblages used to address sheep (Ovis orientalis/aries) exploitation. Site names in boldfaceinterpreted as representing sheep husbandry (based on original author). Summary of skewness (positive,negative, normal, etc.) refers to the shape of the distribution of biometric data. Frequency of juveniles based inmost cases on fusion of the distal epiphysis of metapodials or tooth wear data using Payne’s (1973) categoriesA–D. * indicates percentage of juveniles calculated for combined sheep/goat. Presence or absence of evidencefor young male kill-off (** indicates cases of negative skewing interpreted as young male kill-off) and evidencefor phenotypic changes associated with the domestication process are also indicated
Site Date (cal BC) Skewness %juv Young male killoff? Morph dom.? Reference
Karain B 17,000 neg 20* No No (Atici 2009)Okuzini 1 17,000 NA 25* No No (Atici 2009)Okuzini 2 16,000 pos 26* No? No (Atici 2009)Okuzini 3–4 13,000 neg 27* No No (Atici 2009)Okuzini 5 12,000 pos 22* No? No (Atici 2009)Hallan Cemi 10,000 normal 25 No No (Starkovich and Stiner 2009)ZC Shanidar 10,000 normal 50 No No (Perkins 1973; Zeder 2008a)Palegawra 10,000 neg 25 No No (Zeder 2008a)Mureybet IB–III 9500–8400 pos 23 No No (Gourichon and Helmer 2008)Jerf el-Ahmar 9000 pos nd No? No (Vigne 2011b)Gobekli 9000 neg 40* No No (Peters et al. 2013; Vigne 2011b)Kortik Tepe 9000 neg 50 No No (Arbuckle and Ozkaya 2007)Asiab 9000 neg 33 No No (Zeder 2008a)Cayonu RG 9000–8400 neg 40 No No (Hongo et al. 2005)Cayonu CH 8400–8200 neg 50 No Yes? (Hongo et al. 2005)Cayonu CP 8200–7600 normal 32 No Yes? (Hongo et al. 2005)Nevalı Cori 8500–7600 normal c. 50* No Yes? (Peters et al. 2013)Cafer 8300–7500 neg 43* No No (Helmer 2008)Asıklı 2G 8000 neg 49* No No (Peters et al. 2013)Asıklı 2E 7900 normal 49* No No (Peters et al. 2013)Ganj Dareh 7900 neg 25 No No (Hesse 1978; Zeder 2008a)Aswad moyenne 7800 pos 48 Yes Yes? (Helmer and Gourichon 2008)Cayonu Cell 7600–7500 pos nd Yes? Yes? (Hongo et al. 2005)Abu Hureyra 2A 7500 normal 42* No? Yes? (Legge and Rowley-Conwy 2000)Aswad recente 7500 pos 48 Yes Yes? (Helmer and Gourichon 2008)Shillourokambos anc. C 7600–7500 pos 42 Yes Yes? (Vigne 2011b)Cayonu LR 7500–6900 pos 32 Yes Yes (Hongo et al. 2005)Halula moyenne 7500 pos 67 Yes Yes? (Vigne 2011b)Ali Kosh 7500–6000 neg 23 No No? (Zeder 2008a)Suberde 7500–7000 pos 40 No? No (Arbuckle 2008b)Gritille 7500–7000 pos 40* Yes Yes (Monahan 2000)Teleilat MPPNB 7500 pos 48* Yes? Yes (Ilgezdi 2008)Shillourokambos moy. A1 7500 pos 50 Yes Yes? (Vigne 2011b)Shillourokambos moy. A2 7400 pos 56 Yes Yes? (Vigne 2011b)Hayaz 7400 pos nd Yes? Yes (Peters et al. 2013)Asıklı 2B 7400 normal 49* No No (Peters et al. 2013)Catalhoyuk pre-XII 7400–7000 normal 50 No Yes (Russell and Martin 2005)Abu Hureyra 2B 7400–7100 pos 35* Yes? Yes? (Legge and Rowley-Conwy 2000)Shillourokambos moy. B 7400–7200 pos 30 Yes Yes? (Vigne 2011b)Jarmo 7300–6300 pos nd Yes? Yes (Zeder 2008a)Halula recente 7300 normal 68 No Yes (Vigne 2011b)Ras Shamra Vc 7300 bimodal 42* No Yes (Vigne et al. 2003)Shillourokambos Rec. 7200–7000 pos 19 Yes Yes? (Vigne 2011b)Ain Ghazal PPNC 7000 neg 40* No Yes (Wasse 2002)Ramad I 7000 pos nd Yes? Yes (Ducos 1993)Ghoraife 7000 normal 70 No Yes (Ducos 1993)Ras Shamra Va 7000 bimodal 38* No Yes (Vigne et al. 2003)Tepe Sarab 7000 pos nd Yes? Yes (Zeder 2008a)Gurcutepe 7000 pos 46 Yes Yes (Peters et al. 2013)Teleilat LPPNB 7000 pos 89* Yes Yes (Ilgezdi 2008)Ain Jammam 7000 pos 25 Yes Yes (Makarewicz 2009)Ba’ja 7000 neg nd No Yes (Makarewicz 2009)Ulucak VI 7000–6000 normal 55 No? Yes (Cakırlar 2012)Choga Sefid 7000–5000 pos nd Yes Yes (Zeder 2008a)Bademagacı 7000–6500 pos 31 Yes Yes (De Cupere et al. 2008)Catalhoyuk XII–VII 7000–6800 pos 50 Yes Yes (Russell and Martin 2005)Ramad II 6900–6500 neg nd Yes** Yes (Ducos 1993)Cayonu PN 6900–6300 pos 50 Yes Yes (Hongo et al. 2005)Catalhoyuk VI 6500 pos 60 Yes Yes (Russell and Martin 2005)Ulucak V 6500–6000 pos 78 Yes? Yes (Cakırlar 2012)Erbaba 6500–6000 neg 45 No? Yes (Arbuckle 2008a)Hoyucek 6400–6000 pos 41 Yes Yes (De Cupere and Duru 2003)
Arbuckle and Atici Diversity in sheep and goat management
222 Levant 2013 VOL 45 NO 2
relatively large samples of measurements (Monchot
1999; Vigne 2011b). Although it is preferable to use
measurements from a single skeletal part (e.g.
metacarpal Bd) for this purpose, log transformed
LSI data are also effective in cases where sample sizes
are limited. Recently, both Vigne (2011b) and Zeder
(2008a) have successfully used this method to
interpret Neolithic sheep exploitation practices in
the Neolithic Near East (also see Marom and Bar-Oz
2013; Wolverton 2008).
Finally, postcranial depth and breadth measure-
ments from long bones with epiphyses record
evidence of both the size (and therefore sex) and
age of culled individuals. Since the ages of fusion for
sheep and goat epiphyses are known (e.g. Habermehl
1985; Silver 1963), the fusion status of skeletal
elements can be used to identify individuals as either
older or younger than the age range at which fusion
takes place for a particular skeletal part. Since
caprines achieve most of their skeletal growth by c.
1–2 years, even unfused specimens (especially for
later fusion skeletal parts) can effectively be used to
distinguish between males and females (Davis 2000;
Zeder 2001). Thus, by combining evidence for
epiphyseal fusion with biometric evidence for sex
ratios, we can attempt to identify assemblages that
exhibit the characteristic pattern of large-sized,
unfused specimens and, especially, the small-sized
fused specimens typically produced by the practice of
young male kill-off.
Several strategies were utilized to accommodate the
fact that this study focuses on identifying young male
kill-off through published biometric data. First,
because the samples of measurements from individual
sites identified as either sheep or goat are often
limited, we use the LSI method to combine and
standardize measurements from multiple skeletal
elements (Meadow 1999). Although allometric differ-
ences between archaeological populations and the
standard animal used to generate LSI values can
create variation which potentially blurs differences
between the sexes (Arbuckle and Makarewicz 2009;
Russell et al. 2005), previous work suggests that for
both sheep and goats LSI transformed data still
exhibit clear signals of sexual dimorphism (Vigne
2011b). Second, since specimens with unfused epi-
physes were rarely measured before Zeder’s work, we
focus on biometric data from fused specimens, which
nonetheless provide ample evidence for young male
kill-off (fused specimens will be dominated by small
females when young male kill-off is practised). In
addition, whenever possible, later fusing skeletal parts
which exhibit the highest degree of sexual dimorphism
(e.g. metacarpal Bd, metatarsal Bd, calcaneum GL,
tibia Bd) (based on Davis 2000 for sheep, and Zeder
2001 for goats) were preferentially selected for
analysis, although in some cases (due to small sample
sizes) all available measurements were utilized.
In order to identify young male kill-off, histograms
of LSI values were generated for sheep and for goats
for each of 42 archaeofaunal assemblages. The shape
of the distribution of each of these histograms was
then described in terms of its skewness and kurtosis.
Skewness describes the asymmetry of the distribution
of a variable, with positive skewing representing a
curve with a tail extending further to the right of the
mean, and negative skewing a curve with a tail
extending further to the left of the mean (Moore and
McCabe 2001). A skewness value of ‘0’ represents a
relatively symmetrical distribution. Kurtosis, on the
other hand, refers to the height of a distribution with
positive values representing a tall, peaked curve while
negative values indicate a low peak in the distribution
(Moore and McCabe 2001).
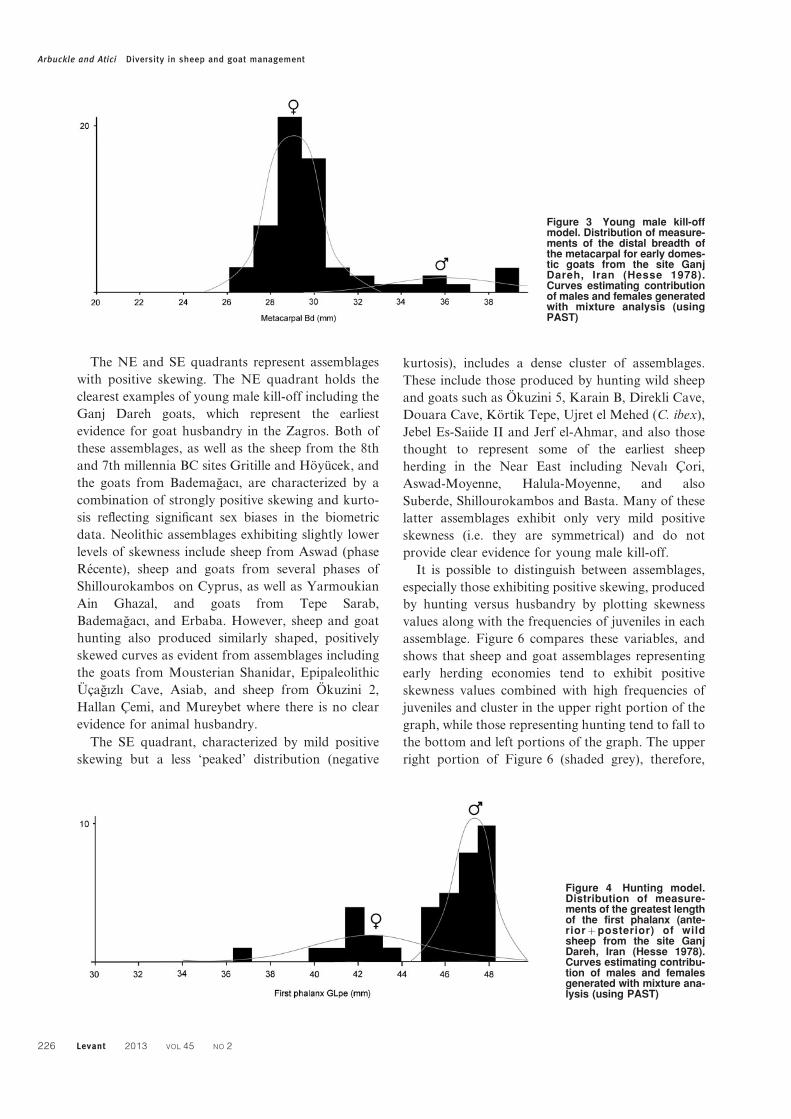
The practice of young male kill-off is expected to
result in diagnostic patterns in biometric data. These
patterns are represented in Figure 3 based on distal
breadth measurements for fused metacarpals for
goats from Ganj Dareh — the site with which this
model has been most clearly associated. This
characteristic distribution exhibits strong positive
skewing and positive kurtosis. The peak on the left
Site Date (cal BC) Skewness %juv Young male killoff? Morph dom.? Reference
Qdeir 6500 normal 49* No Yes (Helmer 1992)Umm El Tlel 6500 neg 60* No Yes (Helmer 1992)El Kowm 2 6500 pos 40* Yes Yes (Helmer 1992)Ain Ghazal Yarmoukian 6500 pos 46* Yes Yes (Wasse 2002)Bouqras 6500 neg nd No Yes (Wasse 2002)Khirokitia III–I 6000 normal nd No No? (Vigne 2011b)Kosk Hoyuk V 6000 pos 61* Yes Yes (Arbuckle 2006)Banahilk 6000–5000 pos nd Yes Yes (Zeder 2008a)Ilipinar X–IX 6000–5800 pos 35 Yes Yes (Buitenhuis 2008)
Table 1 Continued
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 223
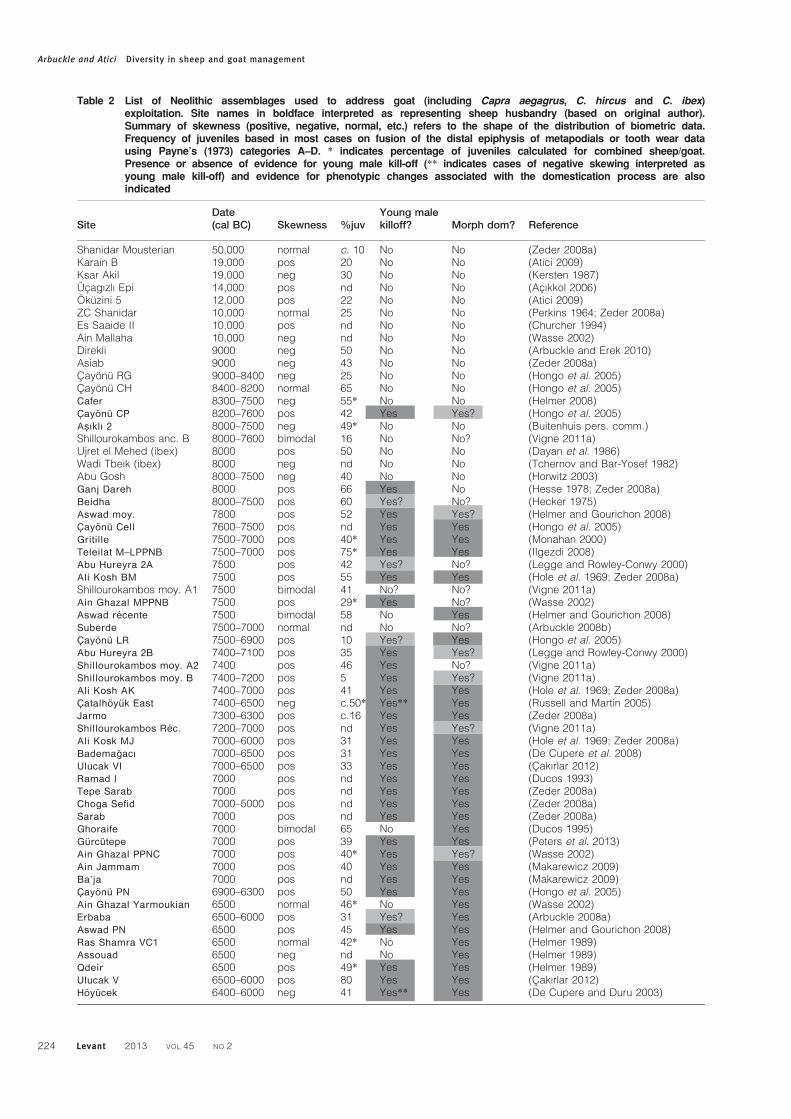
Table 2 List of Neolithic assemblages used to address goat (including Capra aegagrus, C. hircus and C. ibex)exploitation. Site names in boldface interpreted as representing sheep husbandry (based on original author).Summary of skewness (positive, negative, normal, etc.) refers to the shape of the distribution of biometric data.Frequency of juveniles based in most cases on fusion of the distal epiphysis of metapodials or tooth wear datausing Payne’s (1973) categories A–D. * indicates percentage of juveniles calculated for combined sheep/goat.Presence or absence of evidence for young male kill-off (** indicates cases of negative skewing interpreted asyoung male kill-off) and evidence for phenotypic changes associated with the domestication process are alsoindicated
SiteDate(cal BC) Skewness %juv
Young malekilloff? Morph dom? Reference
Shanidar Mousterian 50,000 normal c. 10 No No (Zeder 2008a)Karain B 19,000 pos 20 No No (Atici 2009)Ksar Akil 19,000 neg 30 No No (Kersten 1987)Ucagızlı Epi 14,000 pos nd No No (Acıkkol 2006)Okuzini 5 12,000 pos 22 No No (Atici 2009)ZC Shanidar 10,000 normal 25 No No (Perkins 1964; Zeder 2008a)Es Saaide II 10,000 pos nd No No (Churcher 1994)Ain Mallaha 10,000 neg nd No No (Wasse 2002)Direkli 9000 neg 50 No No (Arbuckle and Erek 2010)Asiab 9000 neg 43 No No (Zeder 2008a)Cayonu RG 9000–8400 neg 25 No No (Hongo et al. 2005)Cayonu CH 8400–8200 normal 65 No No (Hongo et al. 2005)Cafer 8300–7500 neg 55* No No (Helmer 2008)Cayonu CP 8200–7600 pos 42 Yes Yes? (Hongo et al. 2005)Asıklı 2 8000–7500 neg 49* No No (Buitenhuis pers. comm.)Shillourokambos anc. B 8000–7600 bimodal 16 No No? (Vigne 2011a)Ujret el Mehed (ibex) 8000 pos 50 No No (Dayan et al. 1986)Wadi Tbeik (ibex) 8000 neg nd No No (Tchernov and Bar-Yosef 1982)Abu Gosh 8000–7500 neg 40 No No (Horwitz 2003)Ganj Dareh 8000 pos 66 Yes No (Hesse 1978; Zeder 2008a)Beidha 8000–7500 pos 60 Yes? No? (Hecker 1975)Aswad moy. 7800 pos 52 Yes Yes? (Helmer and Gourichon 2008)Cayonu Cell 7600–7500 pos nd Yes Yes (Hongo et al. 2005)Gritille 7500–7000 pos 40* Yes Yes (Monahan 2000)Teleilat M–LPPNB 7500–7000 pos 75* Yes Yes (Ilgezdi 2008)Abu Hureyra 2A 7500 pos 42 Yes? No? (Legge and Rowley-Conwy 2000)Ali Kosh BM 7500 pos 55 Yes Yes (Hole et al. 1969; Zeder 2008a)Shillourokambos moy. A1 7500 bimodal 41 No? No? (Vigne 2011a)Ain Ghazal MPPNB 7500 pos 29* Yes No? (Wasse 2002)Aswad recente 7500 bimodal 58 No Yes (Helmer and Gourichon 2008)Suberde 7500–7000 normal nd No No? (Arbuckle 2008b)Cayonu LR 7500–6900 pos 10 Yes? Yes (Hongo et al. 2005)Abu Hureyra 2B 7400–7100 pos 35 Yes Yes? (Legge and Rowley-Conwy 2000)Shillourokambos moy. A2 7400 pos 46 Yes No? (Vigne 2011a)Shillourokambos moy. B 7400–7200 pos 5 Yes Yes? (Vigne 2011a)Ali Kosh AK 7400–7000 pos 41 Yes Yes (Hole et al. 1969; Zeder 2008a)Catalhoyuk East 7400–6500 neg c.50* Yes** Yes (Russell and Martin 2005)Jarmo 7300–6300 pos c.16 Yes Yes (Zeder 2008a)Shillourokambos Rec. 7200–7000 pos nd Yes Yes? (Vigne 2011a)Ali Kosk MJ 7000–6000 pos 31 Yes Yes (Hole et al. 1969; Zeder 2008a)Bademagacı 7000–6500 pos 31 Yes Yes (De Cupere et al. 2008)Ulucak VI 7000–6500 pos 33 Yes Yes (Cakırlar 2012)Ramad I 7000 pos nd Yes Yes (Ducos 1993)Tepe Sarab 7000 pos nd Yes Yes (Zeder 2008a)Choga Sefid 7000–5000 pos nd Yes Yes (Zeder 2008a)Sarab 7000 pos nd Yes Yes (Zeder 2008a)Ghoraife 7000 bimodal 65 No Yes (Ducos 1995)Gurcutepe 7000 pos 39 Yes Yes (Peters et al. 2013)Ain Ghazal PPNC 7000 pos 40* Yes Yes? (Wasse 2002)Ain Jammam 7000 pos 40 Yes Yes (Makarewicz 2009)Ba’ja 7000 pos nd Yes Yes (Makarewicz 2009)Cayonu PN 6900–6300 pos 50 Yes Yes (Hongo et al. 2005)Ain Ghazal Yarmoukian 6500 normal 46* No Yes (Wasse 2002)Erbaba 6500–6000 pos 31 Yes? Yes (Arbuckle 2008a)Aswad PN 6500 pos 45 Yes Yes (Helmer and Gourichon 2008)Ras Shamra VC1 6500 normal 42* No Yes (Helmer 1989)Assouad 6500 neg nd No Yes (Helmer 1989)Qdeir 6500 pos 49* Yes Yes (Helmer 1989)Ulucak V 6500–6000 pos 80 Yes Yes (Cakırlar 2012)Hoyucek 6400–6000 neg 41 Yes** Yes (De Cupere and Duru 2003)
Arbuckle and Atici Diversity in sheep and goat management
224 Levant 2013 VOL 45 NO 2
represents small adult females, which dominate the
assemblage, while the tail on the right represents a
small number of males allowed to survive past the age
of metacarpal fusion (c. 18–24 months). The esti-
mated distributions of females and males in the
population are further indicated in Figure 3 based on
the results of mixture analysis (PAST software),
emphasizing the highly biased sex ratio among
skeletally mature individuals reflected in this assem-
blage. This pattern, combined with age data indicat-
ing a young kill-off in which most goats (66%) were
culled by 24 months, clearly reflects a management
strategy in which young surplus males are targeted
for slaughter and females are allowed to survive into
adulthood (Zeder 2008a). This pattern is thought to
represent the archetypal management system used by
sheep and goat pastoralists (Payne 1973) and is
unlikely to be produced by hunting strategies.
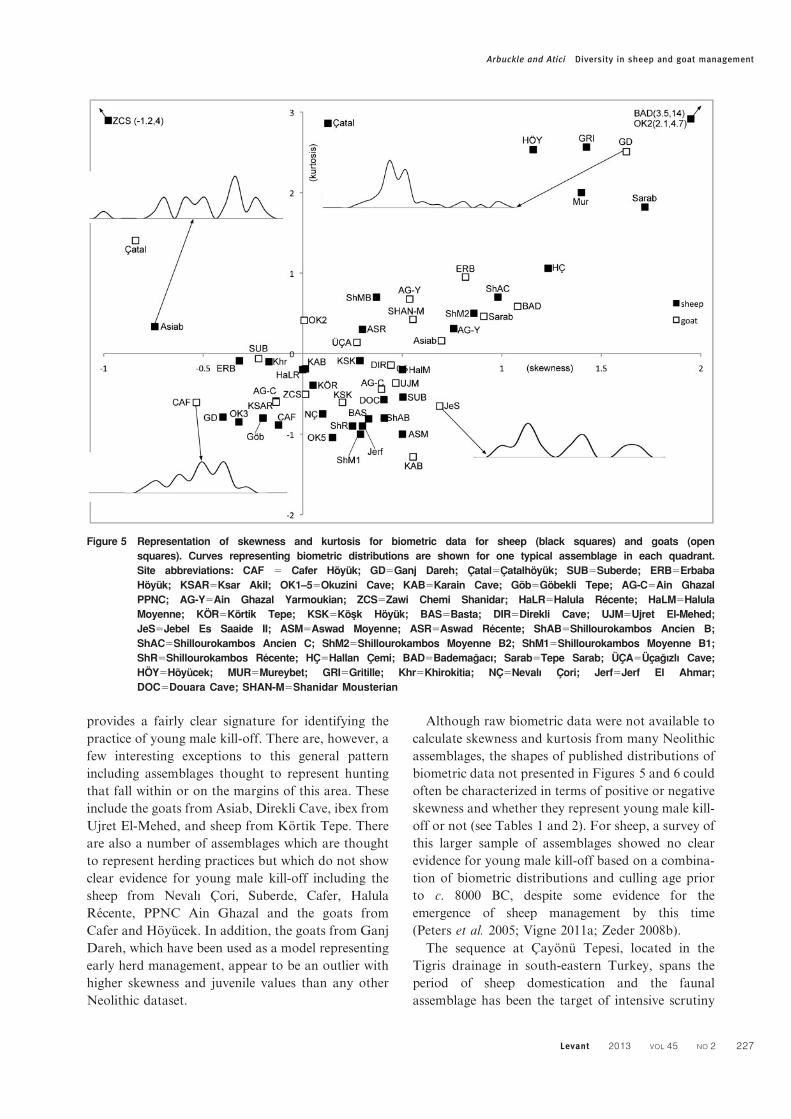
Although hunting wild sheep and goats may
potentially result in a wide variety of demographic
patterns, a frequently reoccurring example of the
distribution of biometric data is presented in Figure 4.
This classic hunting model is based on measurements
of wild sheep from Ganj Dareh and exhibits strongly
negative skewing and negative kurtosis. The peak on
the right represents large adult males, which dominate
the assemblage, while the tail to the left of the peak
represents a smaller numbers of adult females. Mixture
analysis suggests that this pattern reflects a significant
sex bias with males dominating the population
(Fig. 4). In addition, the slaughter age for this
assemblage is quite high, with only 25% of sheep
slaughtered before 24 months, which is a typical,
although not universal, feature of demographic
profiles produced by hunting (Zeder 2008a).
Results
With these two models in mind, skewness and
kurtosis values were plotted for 42 Neolithic assem-
blages for sheep and goats in Figure 5. In this figure,
the NW and SW quadrants represent assemblages
exhibiting negative skewing. In accordance with the
model presented in Figure 5, many of the assem-
blages in these quadrants represent caprine exploita-
tion strategies interpreted as hunting. The majority of
these assemblages exhibit a combination of mild
negative skewing and negative kurtosis as exemplified
by the sheep data from Ganj Dareh, Gobekli Tepe,
Okuzini 3, and goats from Ksar Akil. Only three
examples fall in the NW quadrant including the
assemblages from Asiab and Zawi Chemi Shanidar
which are thought to represent sheep hunting. The
goats from Catalhoyuk, which represent morpholo-
gically domesticated animals, also fall in this quad-
rant but this is likely the result of the near absence of
large males represented in the biometric sample used
in this study (i.e. the tail to the right is missing).
Interestingly, in the SW quadrant, in addition to
assemblages representing wild caprine hunting, there
are also several Neolithic assemblages thought to
represent herding economies including the sheep and
goats from Cafer, goats from Suberde and
Khirokitia, and sheep from PPNC Ain Ghazal
(Arbuckle 2008b; Helmer 2008; Wasse 2002).
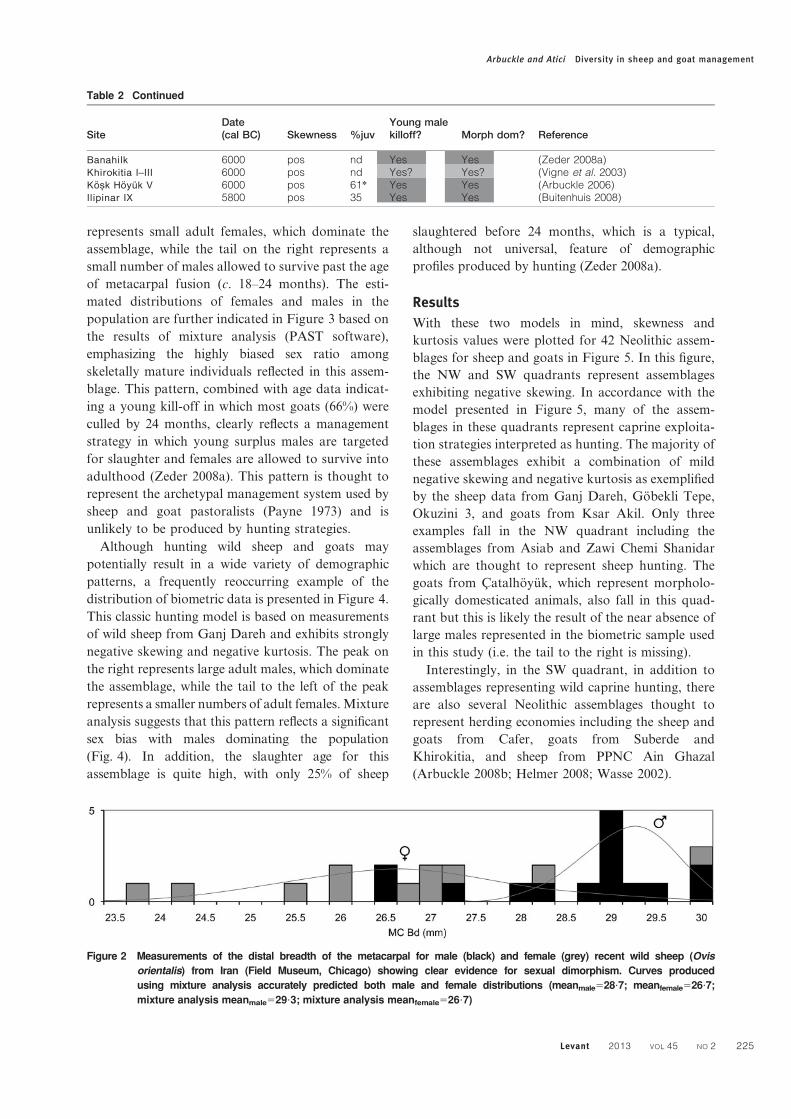
Figure 2 Measurements of the distal breadth of the metacarpal for male (black) and female (grey) recent wild sheep (Ovis
orientalis) from Iran (Field Museum, Chicago) showing clear evidence for sexual dimorphism. Curves produced
using mixture analysis accurately predicted both male and female distributions (meanmale528?7; meanfemale526?7;
mixture analysis meanmale529?3; mixture analysis meanfemale526?7)
SiteDate(cal BC) Skewness %juv
Young malekilloff? Morph dom? Reference
Banahilk 6000 pos nd Yes Yes (Zeder 2008a)Khirokitia I–III 6000 pos nd Yes? Yes? (Vigne et al. 2003)Kosk Hoyuk V 6000 pos 61* Yes Yes (Arbuckle 2006)Ilipinar IX 5800 pos 35 Yes Yes (Buitenhuis 2008)
Table 2 Continued
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 225
The NE and SE quadrants represent assemblages
with positive skewing. The NE quadrant holds the
clearest examples of young male kill-off including the
Ganj Dareh goats, which represent the earliest
evidence for goat husbandry in the Zagros. Both of
these assemblages, as well as the sheep from the 8th
and 7th millennia BC sites Gritille and Hoyucek, and
the goats from Bademagacı, are characterized by a
combination of strongly positive skewing and kurto-
sis reflecting significant sex biases in the biometric
data. Neolithic assemblages exhibiting slightly lower
levels of skewness include sheep from Aswad (phase
Recente), sheep and goats from several phases of
Shillourokambos on Cyprus, as well as Yarmoukian
Ain Ghazal, and goats from Tepe Sarab,
Bademagacı, and Erbaba. However, sheep and goat
hunting also produced similarly shaped, positively
skewed curves as evident from assemblages including
the goats from Mousterian Shanidar, Epipaleolithic
Ucagızlı Cave, Asiab, and sheep from Okuzini 2,
Hallan Cemi, and Mureybet where there is no clear
evidence for animal husbandry.
The SE quadrant, characterized by mild positive
skewing but a less ‘peaked’ distribution (negative
kurtosis), includes a dense cluster of assemblages.
These include those produced by hunting wild sheep
and goats such as Okuzini 5, Karain B, Direkli Cave,
Douara Cave, Kortik Tepe, Ujret el Mehed (C. ibex),
Jebel Es-Saiide II and Jerf el-Ahmar, and also those
thought to represent some of the earliest sheep
herding in the Near East including Nevalı Cori,
Aswad-Moyenne, Halula-Moyenne, and also
Suberde, Shillourokambos and Basta. Many of these
latter assemblages exhibit only very mild positive
skewness (i.e. they are symmetrical) and do not
provide clear evidence for young male kill-off.
It is possible to distinguish between assemblages,
especially those exhibiting positive skewing, produced
by hunting versus husbandry by plotting skewness
values along with the frequencies of juveniles in each
assemblage. Figure 6 compares these variables, and
shows that sheep and goat assemblages representing
early herding economies tend to exhibit positive
skewness values combined with high frequencies of
juveniles and cluster in the upper right portion of the
graph, while those representing hunting tend to fall to
the bottom and left portions of the graph. The upper
right portion of Figure 6 (shaded grey), therefore,
Figure 3 Young male kill-offmodel. Distribution of measure-ments of the distal breadth ofthe metacarpal for early domes-tic goats from the site GanjDareh, Iran (Hesse 1978).Curves estimating contributionof males and females generatedwith mixture analysis (usingPAST)
Figure 4 Hunting model.Distribution of measure-ments of the greatest lengthof the first phalanx (ante-riorzposterior) of wildsheep from the site GanjDareh, Iran (Hesse 1978).Curves estimating contribu-tion of males and femalesgenerated with mixture ana-lysis (using PAST)
Arbuckle and Atici Diversity in sheep and goat management
226 Levant 2013 VOL 45 NO 2
provides a fairly clear signature for identifying the
practice of young male kill-off. There are, however, a
few interesting exceptions to this general pattern
including assemblages thought to represent hunting
that fall within or on the margins of this area. These
include the goats from Asiab, Direkli Cave, ibex from
Ujret El-Mehed, and sheep from Kortik Tepe. There
are also a number of assemblages which are thought
to represent herding practices but which do not show
clear evidence for young male kill-off including the
sheep from Nevalı Cori, Suberde, Cafer, Halula
Recente, PPNC Ain Ghazal and the goats from
Cafer and Hoyucek. In addition, the goats from Ganj
Dareh, which have been used as a model representing
early herd management, appear to be an outlier with
higher skewness and juvenile values than any other
Neolithic dataset.
Although raw biometric data were not available to
calculate skewness and kurtosis from many Neolithic
assemblages, the shapes of published distributions of
biometric data not presented in Figures 5 and 6 could
often be characterized in terms of positive or negative
skewness and whether they represent young male kill-
off or not (see Tables 1 and 2). For sheep, a survey of
this larger sample of assemblages showed no clear
evidence for young male kill-off based on a combina-
tion of biometric distributions and culling age prior
to c. 8000 BC, despite some evidence for the
emergence of sheep management by this time
(Peters et al. 2005; Vigne 2011a; Zeder 2008b).
The sequence at Cayonu Tepesi, located in the
Tigris drainage in south-eastern Turkey, spans the
period of sheep domestication and the faunal
assemblage has been the target of intensive scrutiny
Figure 5 Representation of skewness and kurtosis for biometric data for sheep (black squares) and goats (open
squares). Curves representing biometric distributions are shown for one typical assemblage in each quadrant.
Site abbreviations: CAF 5 Cafer Hoyuk; GD5Ganj Dareh; Catal5Catalhoyuk; SUB5Suberde; ERB5Erbaba
Hoyuk; KSAR5Ksar Akil; OK1–55Okuzini Cave; KAB5Karain Cave; Gob5Gobekli Tepe; AG-C5Ain Ghazal
PPNC; AG-Y5Ain Ghazal Yarmoukian; ZCS5Zawi Chemi Shanidar; HaLR5Halula Recente; HaLM5Halula
Moyenne; KOR5Kortik Tepe; KSK5Kosk Hoyuk; BAS5Basta; DIR5Direkli Cave; UJM5Ujret El-Mehed;
JeS5Jebel Es Saaide II; ASM5Aswad Moyenne; ASR5Aswad Recente; ShAB5Shillourokambos Ancien B;
ShAC5Shillourokambos Ancien C; ShM25Shillourokambos Moyenne B2; ShM15Shillourokambos Moyenne B1;
ShR5Shillourokambos Recente; HC5Hallan Cemi; BAD5Bademagacı; Sarab5Tepe Sarab; UCA5Ucagızlı Cave;
HOY5Hoyucek; MUR5Mureybet; GRI5Gritille; Khr5Khirokitia; NC5Nevalı Cori; Jerf5Jerf El Ahmar;
DOC5Douara Cave; SHAN-M5Shanidar Mousterian
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 227

(Hongo et al. 2005). Here, morphologically wild
sheep were hunted in small numbers in the early and
mid-9th millennium BC, producing biometric dis-
tributions exhibiting negative skewing suggesting an
emphasis on hunting large adult males. By the late
9th millennium, smaller-sized individuals, probably
representing domesticates, begin to appear at Cayonu
but the biometric distribution is symmetrical with no
indication of young male kill-off. It is not until the
Cell Room phase, dating from c. 7500 BC, that
positively skewed biometric data along with evidence
for changes in phenotype suggest that domestic sheep
were subject to young male kill-off at Cayonu.
In the upper Euphrates basin at Nevalı Cori, Peters
(Losch et al. 2006; Peters et al. 2005) has argued for
the earliest appearance of sheep husbandry, including
evidence for phenotypic changes associated with
domestication and isotopic evidence for foddering.
However, in this case the distribution of sheep
measurements is notably symmetrical with negative
kurtosis providing no clear evidence for young male
kill-off in the late 9th and early 8th millennia BC
(Peters et al. 2013). At Cafer Hoyuk, in the highlands
of eastern Anatolia, Helmer (2008) has argued for a
system of sheep husbandry combined with mouflon
hunting in the late 9th and early 8th millennia BC.
However, the sheep at Cafer seem to retain a largely
wild phenotype and biometric data exhibit negative
skewing, suggesting that large adult males were
frequently exploited providing no evidence for
the culling of young males. At Asıklı Hoyuk, in
central Anatolia, where sheep are the dominant
taxon, biometric data from levels 2G to 2B,
representing the early to late 8th millennium BC,
show either negative skewing or no skewing at all.
Despite convincing arguments for ‘proto-domestica-
tion’ management of sheep at Asıklı (Buitenhuis
1997; Matthews 1998) the biometric patterns do not
fit with expectations for young male kill-off. The
earliest available evidence for young male kill-off for
sheep comes from the northern margin of the
southern Levant at Tell Aswad, where by the early
8th millennium BC both morphological domesticates
and positively skewed biometric data suggest sheep
herds were subject to young male kill-off (Helmer and
Gourichon 2008).
Figure 6 Plot showing skewness and frequency of juveniles for Neolithic sheep (black squares) and goat (grey squares)
assemblages. Frequency of juveniles based in most cases on either fusion of metapodials or tooth wear data
using Payne’s (1973) categories A–D. Shaded grey area represents typical pattern for young male kill-off. Site
codes are the same as in Figure 5
Arbuckle and Atici Diversity in sheep and goat management
228 Levant 2013 VOL 45 NO 2

Biometric evidence for young male kill-off becomes
much more common after 7500 BC across south-
western Asia. Out of the assemblages surveyed, 65%
(31/48) reflect fairly clear evidence for young male
kill-off compared to only 13% from earlier Neolithic
assemblages (2/15). In south-eastern Turkey, assem-
blages from Gritille and Teleilet both show evidence
for young male kill-off in the mid- to late 8th
millennium, and this management strategy is also
evident from contemporary levels at Halula and Abu
Hureyra 2B on the Middle Euphrates. At the site of
Shillourokambos, on the island of Cyprus, Vigne
(2011a; Vigne et al. 2011) has argued that domestic
sheep were introduced in small numbers from the
mainland by c. 8000 BC and positive skewing
combined with the culling of juveniles suggests that
young male kill-off was practised by 7500 BC.
Finally, in the Zagros region, central/western
Anatolia, and the southern Levant positive skewing
and young male kill-off are evident at Tepe Sarab,
Pottery Neolithic levels of Catalhoyuk, Bademagacı,
Ulucak V and Ain Jammam in the early 7th
millennium cal BC.
However, young male kill-off is not ubiquitous
even among later Neolithic sites with strong evidence
for sheep herding. For example, at Abu Hureyra on
the Middle Euphrates, domestic sheep are thought to
have appeared in level 2A (c. 7500 BC) but biometric
evidence for young male kill-off is only evident in
level 2B dating to the late 8th millennium (Legge and
Rowley-Conwy 2000). In central Anatolia, domestic
sheep are present at Catalhoyuk from the earliest
levels of the site, c. 7400 BC. However, the distribu-
tion of biometric data from these early levels is
symmetrical (Russell and Martin 2005); it is only in
the early-7th-millennium levels that the shape of the
biometric distribution suggests young male culling at
Catalhoyuk. Moreover, in western and south-western
Anatolia, neither Suberde, Erbaba, nor the earliest
levels of Ulucak, exhibit clear evidence for young
male kill-off. At Suberde skewing of fused measure-
ments is only weakly positive and measurements of
unfused specimens do not suggest that young males
were preferentially culled. At Erbaba, although sheep
with domestic phenotypes are present in this late-7th-
millennium assemblage, biometric data exhibit nega-
tive skewing and large adult males are well represented
suggesting a mixed exploitation system perhaps similar
to that described for Cafer. In the southern Levant,
where domestic sheep were imported in the late 8th
millennium (Horwitz et al. 1999), several assemblages
show, either bimodality, no skewing, or negative
skewing indicating a surprising degree of variability
in evidence for sheep management at sites including
Ras Shamra, Ghoraife, PPNC Ain Ghazal and on the
island of Cyprus at Khirokitia.
As was the case with sheep, evidence for young
male kill-off of goats is limited before c. 8000 BC
(Table 2). Biometric and survivorship evidence from
Cayonu suggest that wild males were targeted by
hunting parties in the 9th millennium BC, with a shift
towards positive skewing and morphological domes-
ticates suggesting the initiation of young male culling
in the Cobble Paved phase dating to the late 9th and
early 8th millennia BC (Hongo et al. 2005). These
patterns become clearer in the following Cell phase
dating to the mid-8th millennium. In addition, at
Cafer, Helmer (2008) has argued that goats were both
herded and hunted in the late 9th and early 8th
millennia, although morphological changes are not
evident and the biometric data exhibit strongly
negative skewing. The same pattern is evident in
central Anatolia at Asıklı Hoyuk despite arguments
for management in the early 8th millennium.
Although goats were imported to the island of
Cyprus by c. 8300 BC, Vigne (2011a) has argued
that they were fundamentally a wild resource and
were hunted. This is confirmed by the strongly
bimodal shape of biometric data from the early levels
of Shillourokambos (Ancienne B–C) and low fre-
quencies of juveniles.
The earliest and most precisely dated evidence for
the practice of culling young male goats comes from
Ganj Dareh in western Iran where it can be closely
dated to the first centuries of the 8th millennium BC
(Zeder and Hesse 2000). In the far southern Levant,
the assemblage from Beidha shows some potentially
early evidence for young male culling including a high
frequency of juveniles and positive skewing of
biometric data, although the chronology of the
assemblage is not well understood and the presence
of both goats and ibex complicate the interpretation
of biometric patterns (Hecker 1975). Roughly con-
temporaneous with Ganj Dareh, the Middle PPNB
levels of Tell Aswad provide the earliest evidence for
young male culling combined with morphological
changes in the goat population (Helmer and
Gourichon 2008). This is significant in that it
indicates that this management strategy was utilized
on opposite sides of the Fertile Crescent by the early
8th millennium.
As was the case with sheep, evidence for culling
young male goats is common in Neolithic assem-
blages across south-western Asia after 7500 BC,
where it is identified in 84% (37/44) of the assem-
blages examined in this study compared to only 35%
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 229

(5/14) of those dating to the earlier Neolithic. In the
late 8th millennium, positive skewing, high frequen-
cies of juveniles and morphological domesticates
indicate young male kill-off was practised along the
Middle Euphrates at Abu Hureyra 2A, at Gritille and
Teleilat in south-eastern Turkey, at Ali Kosh in
western Iran, at Catalhoyuk in central Anatolia,
Ulucak in western Anatolia, in southern Jordan at
Ain Jammam and Ba’ja, and it makes its appearance
on Cyprus at Shillourokambos (Moyenne A.2) by
7400 BC. However, culling young male goats is not
evident in all later Neolithic assemblages, including
Suberde in west central Anatolia; Ghoraife, Aswad
Recente and Tell Assouadin, Syria; and Yarmoukian
Ain Ghazal and Ras Shamra in the southern Levant.
Discussion
The results of this broad survey of faunal evidence for
early sheep and goat management in south-western
Asia emphasize several important features of Neolithic
animal economies. First, we are able to address the
timing of the appearance of young male kill-off as a
major herd management strategy. This is important
both for understanding the evolution of animal
husbandry, but also because Zeder’s detailed analysis
of goat exploitation in the Zagros region has shown
that young male kill-off preceded the appearance of
morphological changes and is, therefore, presented as
the best ‘leading edge marker’ for the earliest stages of
the domestication process (Zeder 2006).
The data examined here suggest that the practice of
young male kill-off was applied to goat herds at
Cayonu in the upper Tigris basin and at Ganj Dareh
in western Iran by c. 8000 BC, and to sheep herds a
few centuries later at Cayonu, Tell Aswad in the
Damascus Basin, and Shillourokambos on Cyprus.
However, these early-8th-millennium communities,
which are spread over a wide geographic area, seem
to be isolated outposts in their choice of herd
management strategies since young male culling is
not evident at contemporaneous sites in the region.
In addition, although young male culling clearly
precedes the appearance of phenotypically domestic
goats in the Zagros region, in the upper Euphrates
region the opposite pattern is evident. Here, at Nevalı
Cori, Peters (Peters et al. 2013) has argued that
phenotypic changes (size decrease) and isotopic
evidence for foddering suggest that sheep and,
perhaps, goat herding was well underway as early
as the late 9th millennium BC. Further upstream,
Helmer (2008) suggests that caprine management
may have been practised at Cafer Hoyuk from the
late 9th and early 8th millennia. However, there is no
evidence to suggest that young males were targeted
for preferential culling at either site. In fact, evidence
for young male culling is not present in the Euphrates
basin until the mid- to late 8th millennium at Teleilat,
Gritille and Gurcutepe.
Thus in the Euphrates basin, young male kill-off is
evident well after the appearance of evidence for herd
management, including phenotypic changes, and
seems not to be the leading edge marker for the
initiation of caprine management as it was for the
Zagros region. The reasons for this reversal are
unclear. It is possible that it is the result of biases in
the archaeofaunal record. For example, it is possible
that the delay in the appearance of young male
culling is the result of small sample sizes from the
earliest phases of important sites, including Aswad
(ancien), Cafer (Early phase), and Shillourokambos
(ancien A), or the common practice of ‘lumping’
faunal data from multiple archaeological phases into
one analytical unit which might obscure patterns in
the data.
However, it is also possible that this pattern
represents a different reality in the Euphrates basin,
where populations of managed caprines in the 9th
and early 8th millennia were very small, and
represented only a minor component of the animal
economy compared to the Zagros where they were
the dominant taxa (Arbuckle 2012; Legge 1996;
Vigne and Helmer 2007). In these cases where local
populations were small, access to wild caprines was
limited, and husbandry likely played as much a social
as subsistence role (Russell 2012), it may not have
made economic (or social) sense for herders to cull
large numbers of young animals. However, these
small, isolated populations of managed caprines may
have developed phenotypic changes associated with
the domestication process more rapidly than their
managed neighbours in uplands regions, which likely
maintained gene flow with wild populations.
This apparent delay in the application of young
male culling in the Euphrates basin may, therefore,
reflect differences in the historical trajectories of
caprine management systems in lowland regions
where gazelle and equids were long the focus of
animal economies, versus upland regions within the
natural habitat of the wild sheep and goats where
caprine management likely first emerged. However,
even in the upland regions of Anatolia where caprines
were heavily exploited (e.g. Asıklı, Cafer), the data
still do not suggest a pattern like that seen in the
Zagros. Here, neither evidence for young male culling
nor phenotypic changes are evident in the late 9th or
early 8th millennia BC.
Arbuckle and Atici Diversity in sheep and goat management
230 Levant 2013 VOL 45 NO 2

The second significant finding of this study relates
to the ubiquity of young male culling as a Neolithic
pastoral management strategy. In the 9th and early
8th millennia BC, sheep and goat management
strategies seem to have been very diverse with only
a minority of communities intensively culling young
males. Young male kill-off is only evident in 30% (7/
23) of the sheep and goat assemblages dating to the
9th and early 8th millennia — it was, therefore, far
from ubiquitous as a management strategy in the
early Neolithic.
However, a major change took place in the mid-7th
millennium BC, after which young male kill-off
became the dominant pastoral management strategy
across south-western Asia. Among sheep and goat
assemblages dating between 7500–6000 BC, clear
evidence for young male kill-off was recognized in
almost 75% (62/84) of cases. That young male kill-off
shifted from an experimental management strategy
used in only a handful of communities to become the
dominant mode of herd management in the mid-8th
millennium BC, is no accident. This period represents
the major inflection point in the evolution of
Neolithic animal economies across south-western
Asia. The frequencies of sheep and goats — which
now exhibit domestic phenotypes — increase drama-
tically at this time, becoming the mainstay of animal
economies across the region, with domesticates
appearing outside of their natural habitat including
goats on the middle Euphrates and sheep in the
southern Levant (Arbuckle 2012; Legge 1996; Vigne
and Helmer 2007; Zeder 2011). This new emphasis on
herding domestic caprines seems to represent the
widespread adoption of intensive and large-scale
mixed-caprine pastoralism throughout much of the
Near East. It was in these new economies of large-
scale sheep and goat pastoralism, combined with
mature agriculture systems (Asouti and Fuller 2012),
that young male kill-off emerged from the cacophony
of diverse local practices as the preferred strategy for
herd management in the late 8th and 7th millennia.
However, even though the majority of later
Neolithic assemblages exhibit clear evidence for
young male kill-off there is still a surprising number
(c. 25%) that do not. This degree of variation in
evidence for herd management, although greatly
reduced compared to the early Neolithic, is interest-
ing. If culling young males is the most efficient and
productive method of herd management, what, then,
is the source of this continued variation?
There are several possibilities. First, with the
development of large-scale pastoral systems seasonal
mobility may have increased in the later Neolithic
(Meiggs 2010). In Neolithic France, Helmer et al.
(2005) have shown that seasonal mobility can result
in truncated demographic profiles, with faunal data
from village settlements reflecting only a portion of a
larger, spatially diverse, exploitation system, thus
blurring evidence for young male culling. This may
explain the lack of evidence for young male culling in
the later Neolithic at sites including Ain Ghazal,
Umm el-Tlel and Tell Assouad.
Second, variation in evidence for management may
be the result of the application of multiple exploita-
tion strategies or types of management systems with
no recent parallels. This is perhaps especially the case
within the natural habitat zone of caprines in upland
regions of north-western Iran and central and eastern
Anatolia, where management may have been com-
bined with hunting to produce biometric and demo-
graphic palimpsests which are difficult to interpret.
For example, at 7th-millennium Erbaba Hoyuk,
negative skewing in sheep biometrics combined with
the presence of phenotypically wild sheep suggests
that local mouflon were the target of hunting parties.
However, the presence of small-sized domesticates
and the fact that the mean LSI value for unfused
specimens is larger than that for fused specimens
suggests that management, including young male kill-
off, may have been practised as well (Arbuckle 2008a).
In addition, continued inter-breeding between mana-
ged and wild populations likely resulted in the long
delay in the appearance of morphological changes
evident at upland sites including Cafer, Ganj Dareh,
Asıklı and Suberde.
In other cases variation may simply reflect the
application of management systems with no modern
analogues. This variation may reflect a period of
early experimentation in caprine management
regimes, a common stage in the development of
technological systems (Skibo and Schiffer 2008). The
evidence for variation in herd management strategies,
especially in the early Neolithic, may represent a
period of ‘initial diversity’ in management technolo-
gies, in which individual communities were in the
process of developing husbandry systems that fit into
their unique local environmental, social, historical
and economic contexts. It was only after c. 7500 BC
that this diverse array of strategies, reflecting funda-
mentally local answers to the problems associated with
caprine husbandry, began to be replaced as herders
settled on young male culling as the most effective
answer to their needs — a shift perhaps fuelled by the
development of large, regional populations of produc-
tive domesticates and increased mobility and inter-
community interaction. Even so, pockets of variation
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 231

continued into the late Neolithic with some commu-
nities still exhibiting evidence for local adaptations to
their subsistence needs (Asouti and Fuller 2012).
Finally, variations in herd management may have
been affected by the biological limitations of herding
phenotypically wild sheep and goats, especially in the
9th and 8th millennia prior to the widespread
availability of caprines with domestic phenotypes.
Domestic livestock exhibit a series of developmental,
morphological, and behavioural changes, known as
the ‘domestication syndrome’ (Hammer 1984), which
include rapid development, earlier sexual maturity,
and, in males, larger testes and greater sperm
production (Kenagy and Trombulak 1986; Lincoln
et al. 1990; Preston et al. 2003). In recent pastoral
societies practicing young male culling, adult males
often represent only 1–10% of the adult population,
requiring males to inseminate a large number of
females (Redding 1981). In the earliest herding
economies this may have posed a problem for
phenotypically wild male caprines in the early stages
of the domestication process, thus potentially creat-
ing a biological limitation to the effectiveness of
young male kill-off. In addition, Ryder (1960) has
suggested that wild sheep, and presumably also goats,
exhibit significantly slower weight gain compared to
domesticates, which may have also made slaughtering
young animals less attractive to early herders.
Although the impact of these and other issues
associated with herding phenotypically wild animals
are not well understood, they must have affected
Neolithic management regimes. In this context, the
rapid increase in phenotypically domesticated sheep
and goats across south-western Asia in the mid- to
late 8th millennium, may represent a response to the
increased productivity of these new and ‘improved’
domesticates, which spurred the widespread adoption
of intensive caprine pastoralism as well as systems of
young male culling.
Conclusion
In this paper we have surveyed a large body of faunal
data focusing on evidence for the practice of young
male culling in Neolithic south-western Asia. By
focusing on a combination of kill-off age and the
shape of the distributions of biometric data, espe-
cially those from skeletal parts that exhibit the
greatest levels of sexual dimorphism, we are able to
address the emergence and ubiquity of young male
culling amongst Neolithic sheep and goat herders.
Although the intensive culling of young males has
been presented as the best ‘leading edge marker’ for
the initiation of sheep and goat herding, this survey
of available data suggests that young male kill-off
appears in the faunal record only in the early 8th
millennium BC — considerably later than the origins
of caprine management. Instead, Neolithic caprine
management practices appear to have been charac-
terized by a high degree of ‘initial diversity’,
especially in the 9th and early 8th millennia,
suggesting that early management strategies may
have been much more varied than previously realized.
However, after c. 7500 BC young male kill-off was
widely practised across south-western Asia, suggest-
ing this efficient and effective management technol-
ogy quickly replaced the diversity of local
management strategies prevalent earlier — although
some local management systems continued into the
7th millennium BC. This mid- 8th-millennium transi-
tion represents an important shift in the evolution of
animal economies in the Near East, characterized by
the widespread appearance of phenotypically domes-
tic caprines and the spread of intensive and large-
scale sheep and goat pastoralism.
Finally, by highlighting diversity in early caprine
management strategies, this survey also suggests that
local-scale variability was also present in the Late
Pleistocene and earliest Holocene animal economies
that preceded herding economies. This suggests that
10th- and early-9th-millennia assemblages within the
natural habitat zone of wild caprines, including
Hallan Cemi and Kortik Tepe in south-eastern
Turkey, Asiab and Zawi Chemi Shanidar in the
Zagros, and sites such as Jeftelik and Qarassa in Syria
and Nachcharini in Lebanon (Garrard et al. 2003;
Ibanez et al. 2010; Rodrıguez Rodrıguez et al. 2010),
may warrant closer scrutiny in order to identify the
origins of local histories of caprine exploitation that
likely led to the management regimes of the later 9th
millennium (Zeder 2012).
Moreover, new data indicating the complexity and
time depth of human manipulation of wild animals
(e.g. Gifford-Gonzalez and Hanotte 2011; Vigne et al.
2003), suggests that to further clarify the picture of
early animal management, future work needs to focus
on a combination of high-resolution methods and
locally oriented models that seek to identify specific
management practices and define their histories of
use, perhaps extending back to the Pleistocene-
Holocene boundary in particular regions. Although
this includes addressing evidence for young male
culling, it is also necessary to explore evidence for
other management practices, including foddering,
control over mobility, penning and manipulation of
weaning age (Balasse and Tresset 2002; Brochier
1993; Makarewicz and Tuross 2012; Meiggs 2010;
Arbuckle and Atici Diversity in sheep and goat management
232 Levant 2013 VOL 45 NO 2

Upex et al. 2012). Only with a better understanding
of these specific management practices will we be able
to develop a detailed understanding of the local
histories from which the earliest systems of animal
husbandry emerged.
Acknowledgements
The authors would like to thank Bill Finlayson and
Cheryl Makarewicz for the invitation to present an
earlier draft of this paper at a session of the Society
for American Archaeology annual meeting in 2012
and to participate in this volume. Financial support
has been provided to BA by Baylor University.
BibliographyAcıkkol, A. (2006) Ucagızlı Magarası Faunasının Zooarkeolojik
Acıdan Incelenmesi: Capra, Capreolus, Dama ve Cervus ların
Morfometrik Acıdan Analizi. Unpublished PhD thesis, Ankara
University.
Arbuckle, B. S. (2006) The evolution of sheep and goat pastoralism and
social complexity in Central Anatolia. Unpublished PhD thesis,
Department of Anthropology, Harvard University.
— (2008a) Caprine exploitation at Erbaba Hoyuk: a Pottery Neolithic
village in Central Anatolia. Pp. 345–65 in L. Gourichon and E.
Vila (eds), Archaeozoology of Southwestern Asia and Adjacent
Areas VIII. Paris: Travaux de la Maison de l’Orient.
— (2008b) Revisiting Neolithic caprine exploitation at Suberde,
Turkey. Journal of Field Archaeology 32, 219–36.
— (2012) Animals in the ancient world. Pp. 201–19 in D. T. Potts (ed.),
A Companion to the Archaeology of the Ancient Near East. Oxford:
Wiley-Blackwell.
— and Erek, C. M. (2010) Late Epipaleolithic hunters of the central
Taurus: faunal remains from Direkli cave, Kahramanmaras,
Turkey. International Journal of Osteoarchaeology 22, 694–707.
— and Makarewicz, C. A. (2009) The early management of cattle (Bos
taurus) in Neolithic central Anatolia. Antiquity 83, 669–86.
— and Ozkaya, V. (2007) Animal exploitation at Kortik Tepe: an early
Aceramic Neolithic site in southeastern Turkey. Paleorient 32,
198–211.
—, Oztan, A. and Gulcur, S. (2009) The evolution of sheep and goat
husbandry in central Anatolia. Anthropozoologica 44, 129–57.
Asouti, E. and Fuller, D. Q. (2012) From foraging to farming in the
southern Levant: the development of Epipaleolithic and Pre-
pottery Neolithic plant management strategies. Vegetation History
and Archaeobotany 21, 149–62.
Atici, L. (2009) Implications of age structures for Epipaleolithic
hunting strategies in the western Taurus mountains, Southwest
Turkey. Anthropozoologica 44, 13–40.
Balasse, M. and Tresset, A. (2002) Early weaning of Neolithic domestic
cattle (Bercy, France) revealed by Intra-tooth Variation in
Nitrogen Isotope Ratios. Journal of Archaeological Science 29,
853–59.
Baumler, H. (1921) Die morphologischen Veranderungen des
Schweineschadels unter dem EInfluss der Domestikation. Archiv
fur Naturgeschichte 87, 140–78.
Binford, L. R. (1968) Post-pleistocene adaptations. Pp. 313–41 in S.
Binford and L. R. Binford (eds), New Perspectives in Archaeology.
Chicago: Aldine.
Bokonyi, S. (1969) Archaeological problems and methods of recogniz-
ing animal domestication. Pp. 219–29 in P. J. Ucko and G. W.
Dimbleby (eds), The Domestication and Exploitation of Plants and
Animals. Chicago: Aldine.
Brochier, J. E. (1993) Cayonu Tepese. Domestication, rythmes et
environnment au PPNB. Paleorient 19, 39–49.
Buitenhuis, H. (1997) Asıklı Hoyuk: a ‘protodomestication’ site.
Anthropozoologica 25–26, 655–62.
— (2008) Ilipinar: The faunal remains from the late Neolithic and early
Chalcolithic periods. Pp. 299–322 in E. Vila, L. Gourichon, A.
Choyke and H. Buitenhuis (eds), Archaeozoology of the Near East
VIII. Lyon: Maison de l’Orient et de la Mediterranee.
Cakırlar, C. (2012) The evolution of animal husbandry in Neolithic central-
west Anatolia: the zooarchaeological record from Ulucak Hoyuk
(c. 7040–5660 cal BC, Izmir, Turkey). Anatolian Studies 62, 1–33.
Childe, V. G. (1936) Man Makes Himself. London: Watts and
Company.
Churcher, C. S. (1994) The vertebrate fauna from the Natufian level at
Jebel es-Saaıde (Saaıde II), Lebanon. Paleorient 20, 35–58.
Davis, S. J. (2000) The effect of castration and age on the development
of the Shetland sheep skeleton and a metric comparison between
bones of males, females and castrates. Journal of Archaeological
Science 27, 373–90.
Dayan, T., Tchernov, E., Bar-Yosef, O. and Yom-Tov, Y. (1986)
Animal exploitation in Ujrat El-Mehed, a Neolithic site in
southern Sinai. Paleorient 12, 105–16.
De Cupere, B. and Duru, R. (2003) Faunal remains from Neolithic
Hoyucek (SW-Turkey) and the presence of early domestic cattle in
Anatolia. Paleorient 29, 107–20.
De Cupere, B., Duru, R. and Umurtak, G. (2008) Animal husbandry at
the Early Neolithic to Early Bronze Age site of Bademagacı
(Antalya province, SW Turkey): evidence from the faunal remains.
Pp. 367–406 in E. Vila, L. Gourichon, A. Choyke and H.
Buitenhuis (eds), Archaeozoology of the Near East VIII. Lyon:
Maison de l’Orient et de la Mediterranee.
Ducos, P. (1978) ‘Domestication’ defined and methodological
approaches to its recognition in faunal assemblages. Pp. 53–56 in
R. H. Meadow and M. Zeder (eds), Approaches to faunal analysis in
the Middle East. Cambridge: Peabody Museum Bulletin 2.
— (1993) Proto-elevage et elevage au Levant Sud au VIIe millenaire BC
les donnees de la Damascene. Paleorient 19, 153–73.
— (1995) Note preliminaire dur les faunes d’Aswad et Ghoraife. Pp.
339–49 in H. de Contenson (ed.), Aswad et Ghoraife. Sites
Neolithiques en Damascene (Syrie), aux IXeme et VIIIeme
millenaires avant l’ere Chretienne. Beyrouth: Bibliotheque
Archeologique et Historique.
Garrard, A., Pirie, A., Schroeder, B. and Wasse, A. (2003) Survey of
Nachcharini Cave and prehistoric settlement in the northern Anti-
Lebanon highlands. Bulletin d’archeologie et d’architecture liba-
naises 7, 15–48.
Gifford-Gonzalez, D. and Hanotte, O. (2011) Domesticating animals in
Africa: implications of genetic and archaeological findings.
Journal of World Prehistory 24, 1–23.
Gourichon, L. and Helmer, D. (2008) Etude archeozoologique de
Mureybet. Pp. 115–228 in J. J. Ibanez (ed.), Le site neolithique de
Tell Mureybet (Syrie du Nord). Oxford: BAR International Series
1843.
Habermehl, K.-H. (1985) Altersbestiummung bei Wildund Pelztieren.
Hamburg: Verlag Paul Parey.
Halstead, P. (1998) Mortality models and milking: Problems of
uniformitarianism, optimality and equifinality reconsidered.
Anthropozoologica 27, 3–20.
Hammer, K. (1984) Das Domestikationssyndrom. Die Kulturpflanze
32, 11–34.
Hecker, H. (1975) The faunal analysis of the primary food animals
from the Pre-Pottery Neolithic Beidha (Jordan). Unpublished
PhD thesis, Columbia University.
Helmer, D. (1989) Le developpement de la domestication au Proche-
Orient de 8500 a 7500 BP: les nouvelles donnees d’El Kowm et de
Ras Shamra. Paleorient 15, 111–21.
— (1992) La domestication des animaux par les homes prehistoriques.
Paris: Masson.
— (2008) Revision de la faune de Cafer Hoyuk (Malatya, Turquie):
apports des methodes de l’analyse des melanges et de l’analyse de
Kernel a la mise en evidence de la domestication. Pp. 169–96 in E.
Vila, L. Gourichon, A. Choyke and H. Buitenhuis (eds),
Archaeozoology of the Near East VIII. Lyon: Maison de l’Orient
et de la Mediterranee.
— and Gourichon, L. (2008) Premieres donnees sur les modalites de
subsistance a Tell Aswad (Syrie, PPNB Moyen et Recent,
Neolithique Ceramique Ancien) - Fouilles 2001–2005. Pp. 119–
51 in E. Vila, L. Gourichon, A. Choyke and H. Buitenhuis (eds),
Archaeozoology of the Near East VIII. Lyon: Maison de l’Orient et
de la Mediterranee.
—, Gourichon, L., Sidi Maamar, H. and Vigne, J.-D. (2005) L’elevage
des caprines neolithiques dans le Sud-Est de la France: saisonna-
lite des abattages, relations entre grottes bergeries et sites de plein
air. Anthropozoologica 40, 167–90.
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 233

Henton, E. (2012) The combined use of oxygen isotopes and microwear
in sheep teeth to elucidate seasonal management of domestic
herds: the case study of Catalhoyuk, central Anatolia. Journal of
Archaeological Science 39, 3264–76.
Hesse, B. (1978) Evidence for husbandry from the early Neolithic site of
Ganj Dareh in western Iran. Unpublished PhD thesis, Columbia
University.
Hole, F., Flannery, K. V. and Neely, J. A. (1969) Prehistory and human
ecology on the Deh Luran Plain. Museum of Anthropology,
University of Michigan, Memoir No.1. Ann Arbor: Museum of
Anthropology, University of Michigan.
Hongo, H., Meadow, R. H., Oksuz, B. and Ilgezdi, G. (2005) Sheep
and goat remains from Cayonu Tepesi, Southeastern Anatolia. Pp.
112–23 in H. Buitenhuis, A. Choyke, L. Martin, L. Bartosiewicz
and M. Mashkour (eds), Archaeozoology of the Near East VI.
Proceedings of the Sixth International Symposium on the
Archaeozoology of Southwestern Asia and Adjacent Areas.
Groningen: ARC Publication 123.
Horwitz, L. K. (2003) The Neolithic fauna. Pp. 87–101 in H. Khalaily
and O. Marder (eds), The Neolithic Site of Abu Gosh. The 1995
Excavations. Jerusalem Israeli Antiquities Authority Reports vol.
19.
—, Tchernov, E., Ducos, P., Becker, C., Driesch, A. v. d., Martin, L.
and Garrard, A. (1999) Animal domestication in the southern
Levant. Paleorient 25, 63–80.
Ibanez, J. J., Balbo, A., Braemer, F., Gourichon, L., Iriarte, E.,
Santana, J. and Zapata, L. (2010) The early PPNB levels of Tell
Qarassa North (Sweida, southern Syria). Antiquity 84.
Ilgezdi, G. (2008) The domestication process in Southeastern Turkey:
the evidence of Mezraa-Teleilat. Unpublished PhD thesis,
Geowissenschaftlichen Fakultat der Eberhard-Karls-Universitat
Tubingen.
Kenagy, G. J. and Trombulak, S. C. (1986) Size and function of
mammalian testes in relation to body size. Journal of Mammalogy
67, 1–22.
Kersten, A. M. P. (1987) Age and sex composition of Epipaleolithic
fallow deer and wild goat from Ksar ‘Akil. Palaeohistoria 29, 119–31.
Legge, A. J. (1996) The beginning of caprine domestication in
southwest Asia. Pp. 238–62 in D. Harris (ed.), Origins and
Spread of Agriculture and Pastoralism in Eurasia. London: UCL
Press.
— and Rowley-Conwy, P. A. (2000) The exploitation of animals. Pp.
423–71 in A. M. T. Moore, G. C. Hillman and A. J. Legge (eds),
Village on the Euphrates. Oxford: Oxford University Press.
Lincoln, G. A., Lincoln, C. E. and McNeilly, A. S. (1990) Seasonal
cycles in the blood plasma concentration of FSH, inhibin and
testosterone, and testicular size in rams of wild, feral and
domestication breeds. Journal of Reproduction and Fertility 88,
623–33.
Losch, S., Grupe, G. and Peters, J. (2006) Stable isotopes and dietary
adaptations in humans and animals at Pre-Pottery Neolithic
Nevalı Cori, Southeast Turkey. American Journal of Physical
Anthropology 131, 181–93.
Ludwig, A., Pruvost, M., Reissmann, M., Benecke, N., Brockmann,
G. A., Castanos, P., Cieslak, M., Lippold, S., Llorente, L.,
Malaspinas, A.-S., Slatkin, M. and Hofreiter, M. (2009) Coat
color variation at the beginning of horse domestication. Science
324, 485.
Makarewicz, C. (2009) Complex caprine harvesting practices and
diversified hunting strategies: Integrated animal exploitation
systems at Late Pre-Pottery Neolithic B ‘Ain Jammam.
Anthropozoologica 44, 79–102.
— and Tuross, N. (2012) Finding fodder and tracking transhumance:
Isotopic detection of goat domestication processes in the Near
East. Current Anthropology 53, 495–505.
Marom, N. and Bar-Oz, G. (2013) The prey pathway: a regional history
of cattle (Bos taurus) and pig (Sus scrofa) domestication in the
northern Jordan Valley, Israel. PLOS One 8, e55958.
Matthews, W. (1998) Appendix 2. Micromorphological analysis of
occupation sequences at the Aceramic Neolithic settlement of
Asikli Hoyuk: an assessment, and comparison to depositional
sequences at Catalhoyuk. Catalhoyuk 1998 Archive Report.
Meadow, R. H. (1989) Osteological evidence for the process of animal
domestication. Pp. 80–96 in J. Clutton-Brock (ed.), The Walking
Larder: Patterns of Domestication, Pastoralism, and Predation.
London: Unwin Hyman.
— (1999) The use of size index scaling techniques for research on
archaeozoological collections from the Middle East. Pp. 285–300
in C. Becker, H. Manhart, J. Peters and J. Schibler (eds), Historia
animalium ex ossibus: Beitrage zur palaoanatomie, archaologie,
agyptologie, ethnologie, and geschichte der tiermedizin. Rahden:
Verlag Marie Leidorf.
Meiggs, D. (2010) Investigation of Neolithic ovicaprine herding
practices by multiple isotope analysis: a case study at PPNB
Gritlle, Southeastern Turkey. Unpublished PhD thesis,
Department of Anthropology, University of Wisconsin.
Monahan, B. H. (2000) The organization of domestication at Gritille, a
Pre-Pottery Neolithic B site in southeastern Turkey. Unpublished
PhD thesis, Northwestern University.
Monchot, H. (1999) Mixture analysis and mammalian sex ratio among
middle Pleistocene mouflon of Arago Cave, France. Quaternary
Research 52, 259–68.
— and Lechelle, J. (2002) Statistical nonparametric methods for the
study of fossil populations. Paleobiology 28, 55–69.
Moore, D. S. and McCabe, G. P. (2001) Introduction to the Practice of
Statistics. New York: W.H. Freeman and Company.
Naderi, S., Rezaei, H.-R., Pompanon, F., Blum, M. G. B., Negrini, R.,
Naghash, H.-R., Balkiz, O., Mashkour, M., Gaggiotti, O. E.,
Ajmone-Marsan, P., Kence, A., Vigne, J.-D. and Taberlet, P.
(2008) The goat domestication process inferred from large-scale
mitochondrial DNA analysis of wild and domestic individuals.
PNAS 105, 17659–64.
Ottoni, C., Flink, L. G., Evin, A., Georg, C., De Cupere, B., Van Neer,
W., Bartosiewicz, L. S., Linderholm, A., Barnett, R., Peters, J.,
Decorte, R., Waelkens, M., Vanderheyden, N., Ricaut, F. O.-X.,
Rus Hoelzel, A., Mashkour, M., Karimlu, A. F. M., Seno, S. S.,
Daujat, J., Brock, F., Pinhasi, R., Hongo, H., Perez-Enciso, M.,
Rasmussen, M., Frantz, L., Megens, H.-J., Crooijmans, R.,
Groenen, M., Arbuckle, B., Benecke, N., Vidarsdottir, U. S.,
Burger, J., Cucchi, T., Dobney, K. and Larson, G. (2012) Pig
domestication and human-mediated dispersal in western Eurasia
revealed through ancient DNA and geometric morphometrics.
Molecular Biology and Evolution 30(4), 824–832.
Payne, S. (1973) Kill-off patterns in sheep and goats: the Mandibles
from Asvan-kale. Anatolian Studies 23, 281–303.
Pearson, J. A., Buitenhuis, H., Hedges, R. E. M., Martin, L., Russell,
N. and Twiss, K. (2007) New light on early caprine herding
strategies from isotope analysis: a case study from Neolithic
Anatolia. Journal of Archaeological Science 34, 2170–79.
Perkins, D. (1964) Prehistoric fauna from Shanidar, Iraq. Science 144,
1565–66.
— (1973) The beginnings of animal domestication in the Near East.
American Journal of Archaeology 77, 279–82.
Peters, J., Buitenhuis, H., Grupe, G., Schmidt, K. and Pollath,
N. (2013) The long and winding road. Ungulate exploitation
and domestication in early Neolithic Anatolia (10,000–7,000
cal BC). In S. Colledge, J. Connolly, K. Dobney, K. Manning
and Shennan, S. (eds), Origins and Spread of Domestic Animals
in Southwest Asia and Europe. Walnut Creek: CA Left Coast
Press, 83–114.
Peters, J., von den Driesch, A. and Helmer, D. (2005) The upper
Ephrates-Tigris basin: cradle of agro-pastoralism? Pp. 96–124 in J.-
D. Vigne, J. Peters and D. Helmer (eds), The First Steps of Animal
Domestication: New Archaeological Approaches. Proceedings of the
9th ICAZ Conference, Durham 2002. Oxford: Oxbow.
Preston, B. T., Stevenson, I. R., Pemberton, J. M., Coltman, D. W. and
Wilson, K. (2003) Overt and covert competition in a promiscuous
mammal: the importance of weaponry and testes size to male
reproductive success. Proceedings of the Royal Society B 270, 633–40.
Pumpelly, R. (1908) Explorations in Turkestan; Expedition of 1904.
Prehistoric Civilizations of Anau; Origins, Growth and Influence of
Environment. Washington DC: Carnegie Institution of
Washington Publication No. 73.
Redding, R. (1981) Decision making in subsistence herding of sheep and
goats in the Middle East. Unpublished PhD thesis, Departments of
Anthropology and Biology, University of Michigan.
Rodrıguez Rodrıguez, A., Haıdar-Boustani, M. G. U., Ibanez, J. J., Al-
Maqdissi, M., Terradas, X. and Zapata, L. (2010) Jeftelik: a new
Early Natufian site in the Levant (Homs Gap, Syria). Antiquity 84.
Russell, N. (2012) Social Zooarchaeology: Humans and Animals in
Prehistory. Cambridge: Cambridge University Press.
Arbuckle and Atici Diversity in sheep and goat management
234 Levant 2013 VOL 45 NO 2

— and Martin, L. (2005) The Catalhoyuk mammal remains. Pp. 33–98
in I. Hodder (ed.), Inhabiting Catalhoyuk: Reports from the 1995–
1999 seasons. Cambridge: McDonald Institute for Archaeological
Research.
—, Martin, L. and Buitenhuis, H. (2005) Cattle domestication at
Catalhoyuk revisited. Current Anthropology 46 Supplement, S101–8.
Ryder, M. L. (1960) A study of the coat of the mouflon Ovis musimon
with special reference to seasonal change. Proc. Zool. Lond.135,
387–408.
Schaller, G. B. (1977) Mountain Monarchs: Wild Sheep and Goats of the
Himalyah. Chicago: University of Chicago Press.
Silver, I. A. (1963) The aging of domestic animals. Pp. 250–68 in D.
Brothwell, E. Higgs and G. Clark (eds), Science in Archaeology.
London: Thames and Hudson.
Simmons, A. H. and Ilany, G. (1975–77) What mean these bones?
Paleorient 3, 269–74.
Skibo, J. M. and Schiffer, M. B. (2008) People and Things: A Behavioral
Approach to Material Culture. New York: Springer.
Starkovich, B. M. and Stiner, M. C. (2009) Hallan Cemi Tepesi: high-
ranked game exploitation alongside intensive seed processing at
the Epipaleolithic-Neolithic transition in southeastern Turkey.
Anthropozoologica 44, 41–62.
Tchernov, E. and Bar-Yosef, O. (1982) Animal exploitation in the Pre-
Pottery Neolithic B Period at Wadi Tbeik, Southern Sinai.
Paleorient 8, 17–37.
Uerpmann, H.-P. (1979) Probleme der Neolithisierung des
Mittelmeeraums. Wiesbaden: Verlag.
Upex, B., Balasse, M., Tresset, A., Arbuckle, B. and Dobney, K. (2012)
Protocol for recording enamel hypoplasia in modern and
archaeological caprine populations. International Journal of
Osteoarchaeology, DOI: 10.1002/oa.2227.
Vanpoucke, S., Mainland, I., De Cupere, B. and Waelkens, M. (2009)
Dental microwear study of pigs from the classical site of
Sagalassos (SW Turkey) as an aid for the reconstruction of
husbandry practices in ancient times. Environmental Archaeology
14, 137–54.
Vigne, J.-D. (2011a) Histoire de la gestion des chevres et des moutons.
Pp. 1039–57 in J. Guilaine, F. Briois and V. Jean-Denis (eds),
Shillourokambos: Un etablissement neolithique pre-ceramique a
Chypre. Les fouilles du secteur 1. Paris: Editions Errance.
— (2011b) Le mouton (Ovis aries). Pp. 1021–38 in J. Guilaine, F. Briois
and V. Jean-Denis (eds), Shillourokambos: Un etablissement
neolithique pre-ceramique a Chypre. Les fouilles du secteur 1.
Paris: Editions Errance.
—, Carrere, I., Briois, F. and Guilaine, J. (2011) The early process of
mammal domestication in the Near East. Current Anthropology
52, S255–71.
—, Carrere, I. and Guilaine, J. (2003) Unstable status of early domestic
ungulates in the Near East: the example of Shillourokambos
(Cyprus, IX–VIIIth millennia cal BC). Pp. 239–51 in J. Guilaine
and A. Le Brun (eds), Le Neolithique de Chypre. Actes du colloque
international organise par le Department des Antiquites de Chypre
et l’Ecole Francaise d’Athenes, Nicosie 17–19, Mai 2001. Athens:
Ecole francaise d’Athenes.
— and Helmer, D. (2007) Was milk a ‘secondary product’ in the Old
World Neolithisation process? Its role in the domestication of
cattle, sheep, and goats. Anthropozoologica 42, 9–40.
von den Driesch, A. (1976) A guide to the measurements of animals
bones from archaeological sites. Peabody Museum Bulletin 1.
Cambridge, MA: Peabody Museum of Archaeology and
Ethnology, 1–137.
Wasse, A. (2002) Final results of an analysis of the sheep and goat
bones from Ain Ghazal, Jordan. Levant 34, 59–82.
Watson, J. P. N. (1978) The interpretation of epiphyseal fusion data.
Pp. 97–101 in D. R. Brothwell, K. D. Thomas and J. Clutton-
Brock (eds), Research Problems in Zooarchaeology. London:
Institute of Archaeology Occasional Publication No. 3.
Winge, H. (1900) Bløddyr-skaller. Knogler af dyr. Plante Levninger. Pp.
158–63.
Wolverton, S. (2008) Harvest pressure and environmental carrying
capacity: an ordinal-scale model of effects on ungulate prey.
American Antiquity 73, 179–99.
Zeder, M. (2001) A metrical analysis of a collection of modern goats
(Capra hircus aegagrus and C. h. hircus) from Iran and Iraq:
Implications for the study of caprine domestication. Journal of
Archaeological Science 28, 61–79.
— (2006) Archaeological approaches to documenting animal domestica-
tion. Pp. 171–80 in M. Zeder, D. Bradley, E. Emshwiller and B. D.
Smith (eds), Documenting Domestication: New Genetic and
Archaeological Paradigms. California: University of California Press.
— (2008a) Animal domestication in the Zagros: an update and
directions for future research. Pp. 243–78 in E. Vila,
L. Gourichon, A. Choyke and H. Buitenhuis (eds),
Archaeozoology of the Near East VIII. Lyon: Maison de l’Orient
et de la Mediterranee.
— (2008b) Domestication and early agriculture in the Mediterranean
Basin: Origins, diffusion, and impact. Proceedings of the National
Academy of Sciences 105, 11597–604.
— (2011) The origins of agriculture in the Near East. Current
Anthropology 52, 221–35.
— (2012) The Broad Sprectrum Revolution at 40: resource diversity,
intensification, and an alternative to optimal foraging explana-
tions. Journal of Anthropological Archaeology 31, 241–64.
— and Hesse, B. (2000) The initial domestication of goats (Capra hircus)
in the Zagros mountains 10,000 years ago. Science 287, 2254–57.
Arbuckle and Atici Diversity in sheep and goat management
Levant 2013 VOL 45 NO 2 235




















