Digestive enzymes of two brachyuran and two anomuran land ...

Inhibitory effect of Pistia tannin on digestive enzymesof Indian major carps: an in vitro study
Sudipta Mandal • Koushik Ghosh
Received: 26 June 2009 / Accepted: 22 March 2010 / Published online: 6 April 2010
� Springer Science+Business Media B.V. 2010
Abstract Aquatic weeds are one of the major
unconventional feed ingredients tested for aquafeed
formulation. Tannin content in the water lettuce,
Pistia, has been quantified (26.67 mg g-1; dry
weight) and graded levels of which (12.5–200 lg)
have been incorporated in the reaction mixtures to
evaluate any change in the in vitro activity of the
principal digestive enzymes from the three Indian
major carps (IMC), namely rohu (Labeo rohita), catla
(Catla catla) and mrigala (Cirrhinus mrigala). Result
of the experiment revealed that the Pistia tannin (PT)
significantly inhibit/lower the activities of the diges-
tive enzymes from three IMCs in a dose-dependant
manner, even at very low concentration. Significant
variation in the reduction of the enzyme activities
was noticed between the three fish species, as well as
between the three enzymes studied. Among the three
species studied, digestive enzymes from L. rohita
were found to be the most sensitive to the PT,
whereas enzymes from C. catla were found to be
comparatively least affected. On the other hand,
protease and lipase activities were comparatively
more affected than the amylase activity. The results
of the study suggest that more stress should be given
on the elimination of tannin while incorporating feed
ingredients of plant origin in fish diets.
Keywords Indian major carps � Tannin �Protease � Amylase � Lipase � Pistia
Introduction
The growing demand to substitute fish meal in
aquafeed has compelled to search for alternative less
expensive and protein-rich sources. Plant proteins are
considered to be the most viable alternative in this
respect for economic fish production in most of the
developing countries (Wee and Wang 1987; Mukho-
padhyay and Ray 1996; Becker and Makkar 1999).
The use of plant-derived materials (e.g., aquatic
weeds, legume seeds, different types of oilseed cake,
leaf meals, leaf protein concentrates and root tuber
meals) as fish feed ingredients is limited by the
presence of a wide variety of antinutritional sub-
stances (Francis et al. 2001). Even though possibilities
for uses of plant ingredients have been investigated
and achieved experimental success (Hasan et al. 1990;
Patra et al. 2002; Kalita et al. 2007). The water lettuce,
Pistia, is an aquatic weed; infestation of which causes
great problem in the aquaculture ponds. In compar-
ison with the terrestrial conventional vegetal meals,
the use of aquatic weeds like Pistia involves minimum
S. Mandal � K. Ghosh (&)
Aquaculture Laboratory, Department of Zoology,
The University of Burdwan, Golapbag, Burdwan 713104,
West Bengal, India
e-mail: [email protected]; [email protected]
123
Fish Physiol Biochem (2010) 36:1171–1180
DOI 10.1007/s10695-010-9395-6

expenditure as it requires only the cost of labor for
collection. Ray and Das (1995) have shown that Pistia
has the potential to serve as one of such alternate
ingredient in fish feed. However, higher levels of
incorporation of Pistia meal in aquafeeds were
associated with poor growth, feed conversion and
protein utilization, which may be due to the interfer-
ence caused by the antinutrients therein. Tannins are
one such naturally occurring plant polyphenols that
may interfere with the digestive processes (Liener
1989). These are secondary compounds of various
chemical structures which are generally divided into
hydrolysable and condensed tannins (Francis et al.
2001). Experimental evidence has shown to induce
feed rejection by hydrolysable tannin when included
in the feed of the common carp, Cyprinus carpio
(Becker and Makkar 1999). Poor growth performance
has also been recorded in the same species as a result
of feeding tannic acid incorporated diet (Hossain and
Jauncey 1989). However, studies on the interaction of
tannin with fish intestinal enzymes are scanty. Maitra
and Ray (2003) demonstrated the inhibition of the
intestinal enzymes in Labeo rohita fingerlings to a
significant level by the tannin extracted from Acacia,
though the effect of the hydrolysable tannin on the
digestive enzymes of the other carp species has not
been worked out. Indian major carps might not share
same pattern of inhibition with regard to the digestive
enzymes as they vary in feeding regime specificity
and also in the enzyme activities (Jhingran 1997).
Therefore, the present study was undertaken to
demonstrate the comparative effect of the hydrolysa-
ble tannin extracted from Pistia on the digestive
enzymes of three Indian major carps, rohu, Labeo
rohita; catla, Catla catla and mrigal, Cirrhinus
mrigala.
Materials and methods
Extraction of crude tannin
Tannin was extracted from Pistia leaves following
the method described by Schanderi (1970). Pistia
leaves were collected from a local water body, oven
dried (at 55 ± 5�C) for 48 h and finely powdered in a
mixer grinder (Remi Laboratory Blender). The pow-
dered material (5 g) was mixed with distilled water
(200 ml) and kept at room temperature for overnight.
After soaking, the mixture was boiled for 30 min,
cooled and centrifuged at 2,000 rpm for 20 min. The
supernatant was collected and used as tannin extract.
The extracted tannin was not purified further.
Estimation of extracted tannin
Concentration of tannin in the extract was measured
by Folin–Denis method (Schanderi 1970) with minor
modifications. The crude extract (0.2 ml) was diluted
with 8.3 ml of distilled water and then mixed with
0.5 ml of Folin–Denis reagent. The reaction mixture
was alkalinized by the addition of 1 ml of 15% (w/v)
sodium carbonate solution and kept in dark for
30 min at room temperature. The absorbance of the
solution was read at 700 nm using spectrophotometer
(Shimadzu UV/VIS-1700), and the concentration of
tannin in the extract was determined using pure
tannic acid (MERCK, India) as standard.
Experimental fish
Fingerlings of Labeo rohita, Catla catla and Cirrhi-
nus mrigala were collected from three local compos-
ite carp culture farms and acclimatized separately
according to their source in glass aquaria (75 L) for
10 days, during which fish were fed ad libitum with a
diet containing approximately 40% crude protein
having fish meal as the chief protein source. Average
weight of the fishes examined and their feeding habits
are presented in Table 1.
Preparation of crude enzyme
The experimental fish were weighed to the nearest
gram on a single-pan top-loading balance, and the
anterior and middle intestinal parts were dissected out
on cooled plates and placed in prechilled Petri dishes.
Blood and other debris were washed out with chilled
phosphate buffer (0.1 mol L-1, pH 7.4) containing
0.89% sodium chloride (phosphate buffered saline,
PBS). The cleaned intestinal parts were minced with
scissors, and a 10% homogenate was prepared in the
same buffer using tissue homogenizer (REMI, Model
RQ-127A2). The homogenate was centrifuged
(REMI, Model No.C24) at 10,000 rpm for 30 min
at 4�C. The supernatant was separated and used as
enzyme extract. Preparation of enzyme extract and
inhibition assay was done on the same day, and the
1172 Fish Physiol Biochem (2010) 36:1171–1180
123

extract was kept at 4�C in between. Protein content of
the extract was determined following the method
described by Lowry et al. (1951) using BSA as
standard. Three specimens of each fish species from
each collection site were used for preparation of
enzyme extract for every replicate, and there were
three replicates for each fish species.
Inhibition assay
Inhibitory effect of tannin extracted from Pistia leaf
meal on the activity of principal digestive enzymes
were studied in vitro by adding graded levels of
extracted tannin (12.5–200 lg of tannin) to the test
tubes containing enzyme extracts. Then, substrate and
PBS were added to the tubes and incubated at 37�C
for the optimal time period. Any minor change in pH
and volume of the reaction mixture due to the
addition of tannin was corrected by adding PBS. A
concurrent control set without any tannin was main-
tained for each experimental set. Decrease in the
enzyme activity due to the addition of tannin was
expressed as percent reduction when compared to the
control set (100% activity). There were three repli-
cates for each of the experimental set.
Protease assay
Protease activity was measured according to Anson
(1938) using bovine serum albumin (BSA) as
substrate. Graded levels of extracted tannin were
added to 0.1 ml of enzyme extract followed by
0.1 ml of substrate (10 mg ml-1 BSA solution),
0.5 ml of phosphate buffer (pH 7.4, 0.1 M) and
0.3 ml of distilled water. The reaction mixture was
incubated for 1 h at 37�C. One milliliter of 10%
trichloroacetic acid (TCA) was then added to stop the
reaction and precipitate any remaining substrate. It
was then centrifuged at 3,000 rpm for 10 min. To
2 ml of the supernatant, 1 ml of distilled water and
1 ml of ninhydrin reagent were added in a test tube
and heated in a boiling water bath for 20 min, placing
a marble on top of each tube. After cooling to room
temperature, the developed color was read at 570 nm
in a spectrophotometer (Shimadzu UV/VIS-1700).
Blanks were obtained by adding TCA to the substrate
prior to incubation. The amino acid liberated was
measured following Moore and Stein (1948). Prote-
ase activity was expressed as lg of glycine liberated
h-1 mg protein-1.
a-Amylase assay
a-Amylase activity was determined following the
method described by Bernfeld (1955). Graded levels
of extracted tannin were added to 1 ml of enzyme
extract followed by 1 ml of substrate (1% soluble
starch). The mixture was incubated at 37�C for
20 min. Then, 2 ml of dinitrosalicylic acid reagent
(DNSA reagent, containing 1% dinitrosalicylic acid,
30% sodium potassium tartrate in 0.4 N NaOH
solution) was added to each tube and kept in a
boiling water bath for 5 min. Then, the tubes were
cooled and intensity of the color developed was read
at 540 nm. Blanks were obtained by adding DNSA
reagent prior to incubation. Amylase activity was
expressed as mg maltose liberated h-1 mg protein-1.
Lipase assay
Lipase activity was measured following the method
described by Colowick and Kaplan (1955) using olive
oil as substrate. Briefly, graded levels of extracted
tannin (12.5–200 lg of tannin) were added to 0.5 ml
of enzyme extract followed by 2.5 ml of substrate
(olive oil emulsion in 2% polyvinyl alcohol) and
0.5 ml of calcium chloride solution (110 mM). The
reaction mixture was taken in 100-ml conical flask
and incubated at 37�C for 1 h in a shaker incubator
(Lab. companion, SL-300R) with continuous shaking
Table 1 Food habit and average weight of the fishes examined
Fish species Food habit Average weight (g) (±SD)
Catla catla Zooplanktophagous 17.67 ± 1.8
Labeo rohita Omnivorous, mostly plant matter 12.44 ± 1.67
Cirrhinus mrigala Detritivorous 10.11 ± 1.45
Values are mean ± standard deviation (SD) of nine specimens
Fish Physiol Biochem (2010) 36:1171–1180 1173
123

(120 rpm). Then, 10 ml of acetone–ethanol mixture
(1:1) was added to stop the reaction. Few drops of
phenolphthalein indicator were added, and fatty acid
liberated as a result of enzymatic action was titrated
with 0.02 N NaOH solutions till the appearance of
faint pink color. One milliliter of 0.02 N NaOH is
equivalent to 100 lM of free fatty acid. Blanks were
obtained using boiled enzyme. Lipase activity was
expressed as l mole of fatty acid liberated h-1 mg
protein-1.
Statistical analysis
The data obtained on the enzyme activity in presence
of the tannin were subjected to regression analysis,
using the concentration of the inhibitor as the
independent variable. Further, the data on the prote-
ase, amylase and lipase enzymes were applied for
three-way ANOVA followed by Tukey’s test to infer
about the differences in the enzyme inhibition
between the three fish species. All the statistical
analyses were carried out following Zar (1999) using
SPSS Ver10 (Kinnear and Gray 2000) software.
Results
The tannin content in powdered Pistia was
26.67 mg g-1 of leaf meal. The concentration of
tannin in the tannin extract was 0.89 mg mL-1. The
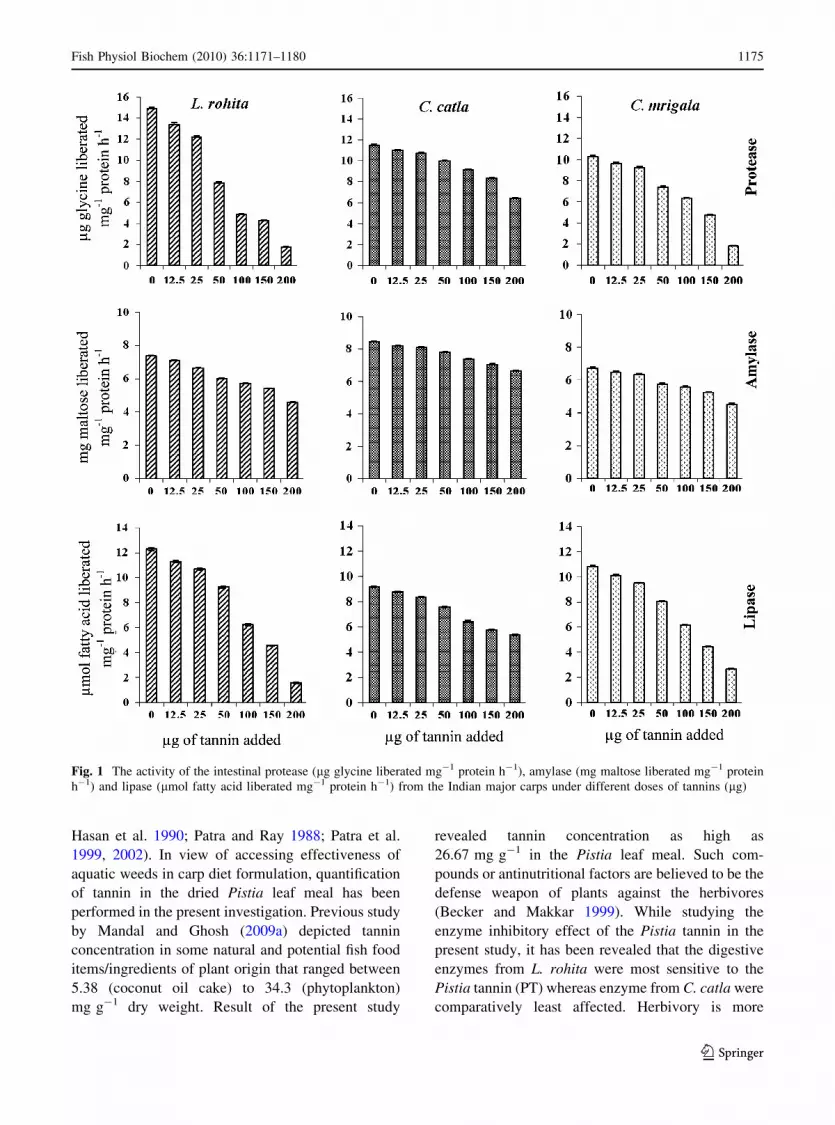
results of the experiment revealed that the Pistia
tannin (PT) significantly inhibit/lower the activities
of the digestive enzymes from three IMCs in a dose
dependant manner (Fig. 1) as evident from the
regression equations (Table 2). In all the instances
irrespective of the carp species, the enzyme activity
was noted to decrease with the increase in the tannin
concentrations. However, variation in the reduction
of enzyme activities was noticed between the three
fish species, as well as the enzymes studied. Among
the fish species, digestive enzymes from L. rohita
were found to be the most sensitive to the PT
followed by C. mrigala, whereas enzymes from
C. catla were comparatively least affected. The
activities of protease and lipase were more affected
compared to the amylase.
Percent inhibition of the enzyme activities are
presented in Table 3. Protease activities in the
control sets (without tannin) in L. rohita, C. catla
and C. mrigala were 14.85 (±0.15), 11.52 (±0.07)
and 10.32 (±0.13) lg of glycine liberated h-1 mg
protein-1, respectively. There was a reduction of 9.65
(±0.13)% to 88.28 (±0.68)% enzyme activity in
L. rohita when 12.5–200 lg of tannin was added.
With similar doses of tannin, reduction in the
protease activities ranged between 4.37 (±0.06)%
to 43.96 (±0.09)% and 6.43 (±0.17)% to 82.53
(±0.99)% in C. catla and C. mrigala, respectively.
Amylase activities in the control sets were 7.34
(±0.04), 8.43 (±0.06) and 6.73 (±0.07) mg maltose
liberated h-1 mg protein-1 in L. rohita, C. catla and
C. mrigala, respectively. Reduction in the enzyme
activity due to the addition of tannin was reduced as
3.22 (±0.07)% to 37.88 (±0.77)% in L. rohita, 3.12
(±0.03)% to 21.63 (±0.33)% in C. catla and 3.67
(±0.14)% to 32.74 (±0.39)% in C. mrigala.
Activities of lipase in the control sets in L. rohita,
C. catla and C. mrigala were recorded as 12.27
(±0.11), 9.17 (±0.05) and 10.82 (±0.06) lmol of
fatty acid liberated h-1 mg protein-1, respectively.
Reduction in the enzyme activities varied between
7.93 (±0.2)% to 87.07 (±0.59)% in L. rohita, 4.43
(±0.04)% to 41.35 (±0.4)% in C. catla and 6.41
(±0.03)% to 75.35 (±0.62)% in C. mrigala with the
addition of tannin.
The results of the three-way ANOVA (Table 4)
revealed significant differences in enzyme activity
due to tannin in terms of dose, type of enzyme and
the species. The post hoc Tukey test revealed
significant differences in protease, amylase and lipase
activity between the three fish species (Between
L. rohita and C. catla: 1.09; between L. rohita and
C. mrigala: 0.51; between C. catla and C. mrigala:
1.61; for all values P \ 0.001).
Discussions
Chemical compounds that render plant tissues unpal-
atable are known to be widely distributed in the plant
kingdom (Liener 1989; Makkar 1993; Hagerman et al.
1997). Tannins are one such group of phenolic
compounds which have received a lot of attention
with respect to their possible nutritional and physio-
logical interference. The nutritional value and possi-
bilities of utilization of aquatic weeds have been
evaluated in some of the literatures in the last decade
(Edwards et al. 1985; Ray and Das 1992, 1994, 1995;
1174 Fish Physiol Biochem (2010) 36:1171–1180
123
Hasan et al. 1990; Patra and Ray 1988; Patra et al.
1999, 2002). In view of accessing effectiveness of
aquatic weeds in carp diet formulation, quantification
of tannin in the dried Pistia leaf meal has been
performed in the present investigation. Previous study
by Mandal and Ghosh (2009a) depicted tannin
concentration in some natural and potential fish food
items/ingredients of plant origin that ranged between
5.38 (coconut oil cake) to 34.3 (phytoplankton)
mg g-1 dry weight. Result of the present study
revealed tannin concentration as high as
26.67 mg g-1 in the Pistia leaf meal. Such com-
pounds or antinutritional factors are believed to be the
defense weapon of plants against the herbivores
(Becker and Makkar 1999). While studying the
enzyme inhibitory effect of the Pistia tannin in the
present study, it has been revealed that the digestive
enzymes from L. rohita were most sensitive to the
Pistia tannin (PT) whereas enzyme from C. catla were
comparatively least affected. Herbivory is more
Fig. 1 The activity of the intestinal protease (lg glycine liberated mg-1 protein h-1), amylase (mg maltose liberated mg-1 protein
h-1) and lipase (lmol fatty acid liberated mg-1 protein h-1) from the Indian major carps under different doses of tannins (lg)
Fish Physiol Biochem (2010) 36:1171–1180 1175
123

common in the L. rohita than C. mrigala; on the other
hand, C. catla feeds on zooplanktons (Jhingran 1997;
Mohanty 2003). Most of the previous studies reported
L. rohita to be herbivore (Alikunhi 1958; Khan and
Siddiqui 1973); however, some of the recent inves-
tigations have focused on omnivorous feeding apti-
tude of this species by analyzing feed intake/
preference (Mohanty 2003; Rahman et al. 2006) or
symbiotic gut microbial community (Ghosh et al.
2002). Therefore, maximum inhibition of digestive
enzymes from L. rohita by plant tannin and appear-
ance of omnivorous feeding aptitude may be viewed
as co-evolution where tannin in the leaf meal appears
to play as a defense compound. To reduce damage
caused by herbivory, plants have evolved a broad
array of defenses, majority of which are chemicals.
Herbivores prefer to avoid ingesting tannin if alternate
foods are available (Wynne-Edwards 2001). Thus,
defenses and counter defenses lay down the step for
the co-evolution of plants and herbivores.
Hydrolysable tannins are easily degraded in bio-
logical systems by non-specific esterase, and the
hydrolyzed products entering into the blood may
cause organ toxicity (particularly in liver and kidney)
once the level in blood rises beyond the detoxification
capability of these organs (Garg et al. 1992; Muller-
Harvey and McAllan Mueller-Harvey and McAllan
1992). In contrast, the condensed tannins are complex
and larger molecules which are not hydrolyzed in the
biological system, indicating their least effect in fish
(Makkar et al. 1995). Condensed tannin did not
appear to bind to proteins and other nutrients and
decrease their bioavailability in the carp intestine
(Becker and Makkar 1999). In the present study,
inhibition of digestive enzymes in carps must be from
their exposure to the hydrolysable tannin (or, tannic
acid), indicating its potential to interfere with the
digestive process.
Commonly used ingredients in formulated fish
feed like mustard oil cake and sun flower oil cake
contain tannin as high as 21.25 mg and
26.25 mg g-1 dry weight, respectively (Mandal and
Ghosh 2009a). It may be assumed that if fish of 100 g
biomass is fed at the rate of 3% of the body weight
daily with a formulated diet having only 10% of these
ingredients, the fish will be exposed to more than
6 mg of tannin daily. Natural feeding on phytoplank-
ton tends to increase this value. In the present study,
concentration of tannin used (12.5–200 lg) are much
lower than this presumptive value. The results of the
present study clearly indicated that tannin, even in
very low concentration, can inhibit or lower the
activities of protease, amylase and lipase in all the
three carp species studied, which is in agreement with
the study made by Maitra and Ray (2003) in L. rohita
fingerlings. In a study based on growth and hemato-
immunological parameters, Prusty et al. (2007)
opined that tannic acid in feed (up to 2%) may not
be detrimental for Labeo rohita fingerlings. However,
they did not address enzyme activities in the short-
term feeding trial of 60 days. Similarly, Becker and
Makkar (1999) assumed that tannic acid may not
affect nutrient availability and enzyme activity in
gastrointestinal tract of carp. In contradiction to these
observations, in vitro studies made in the present
experiment clearly indicated inhibition of the diges-
tive enzyme activities in carps by the tannic acid
extracted from the Pistia. Although experiment
Table 2 Regression equations of the activities of enzymes from the Indian major carps against graded levels of tannin
Enzymes Fish species Regression equation
(y = enzyme activity, x = dose of tannin)
Protease L. rohita y = 13.36 - 0.064x; F = 181.98, df = 1,19; p \ 0.001; R2 = 0.905
C. catla y = 11.37 - 0.023x; F = 834.63, df = 1,19; p \ 0.001; R2 = 0.978
C. mrigala y = 10.12 - 0.04x; F = 848.1, df = 1,19; p \ 0.001; R2 = 0.978
Amylase L. rohita y = 7.08 - 0.013x; F = 254.35, df = 1,19; p \ 0.001; R2 = 0.93
C. catla y = 8.3 - 0.009x; F = 614.45, df = 1,19; p \ 0.001; R2 = 0.97
C. mrigala y = 6.58 - 0.01x; F = 304.04, df = 1,19; p \ 0.001; R2 = 0.941
Lipase L. rohita y = 11.98 - 0.05x; F = 2687.66, df = 1,19; p \ 0.001; R2 = 0.993
C. catla y = 8.84 - 0.02x; F = 348.86, df = 1,19; p \ 0.001; R2 = 0.948
C. mrigala y = 10.49 - 0.04x; F = 2099.89, df = 1,19; p \ 0.001; R2 = 0.991
1176 Fish Physiol Biochem (2010) 36:1171–1180
123

conducted with commercial tannin may not corre-
spond with the tannins of the feed components
(Becker and Makkar 1999). Tannins may reduce
macro nutrient utilization by forming tannin–protein
complexes with various digestive enzymes preclud-
ing the formation of the product absorbable by the
small intestine (Carmona et al. 1996). In addition, it
may inhibit transport systems concerning carbohy-
drate assimilation such as glucosidase/maltase
(Bjorck and Nyman 1987), sucrase (Welsch et al.
1989) and the intestinal sodium-dependent glucose
uptake system (Karasov et al. 1992). Bean tannins
were shown to strongly inhibit pancreatic trypsin,
chymotrypsin and a-amylase (Sing 1984; Carmona
et al. 1991).
Tannins are considered as plant secondary metab-
olites which are distinguished from other polypheno-
lic compounds by their ability to precipitate proteins
(Silanikove et al. 2001). It has been postulated that
tannins interfere with protein and dry matter digest-
ibility by inhibiting protease and also forming
indigestible complexes with dietary protein (Krog-
dahl 1989). However, little is known about the effect
of tannin on fish (Makkar and Becker 1998). Vohra
et al. (1966) reported that tannins caused growth
depression in chickens at levels as low as 0.5% of the
diet. Similarly, Hossain and Jauncey (1989) observed
poor growth response in common carp (Cyprinus
carpio) fed diets containing 0.57 and 1.14% tannins.
In addition, high levels of tannins in feed have been
shown to have adverse effect on herbivorous and
omnivorous fish (Al-Owafeir 1999; Becker and
Makkar 1999; Olvera et al. 1988). The extents of
these growth inhibitory effects varied in different fishTa
ble
3P
erce
nt
red
uct
ion
of
inv
itro
acti
vit
ies
of
the
inte
stin
alen
zym
esfr
om
the
thre
eIn
dia
nm
ajo
rca
rps
by
gra
ded
lev
els
of
tan
nin
extr
acte
dfr
om
Pis
tia
leaf
mea
l
Tan
nin
add
ed(l
g)
%R
edu
ctio
nin
enzy
me
acti
vit
y(c
on
tro
l,w
ith
ou
tta
nn
inat
10
0%
acti
vit
y)
Pro
teas
eA
my
lase
Lip
ase
L.
roh
ita
C.
catl
aC
.m
rig
ala
L.
roh
ita
C.
catl
aC
.m
rig
ala
L.
roh
ita
C.
catl
aC
.m
rig
ala
12
.59
.65
±0
.13
4.3
7±
0.0
66
.43
±0
.17
3.2
2±
0.0
73
.12
±0
.03
3.6
7±
0.1
47
.93
±0
.24
.43
±0
.04
6.4
1±
0.0
3
25
17
.87
±0
.35
6.7
4±
0.1
41
0.6
6±
0.1
99
.31
±0
.23
3.6
8±
0.0
85
.75
±0
.14
12
.95
±0
.12
8.9
±0
.11
2.1
7±
0.1
6
50
47
.17
±0
.51
13
.23
±0
.21
28
.2±
0.1
91
8.5
3±
0.3
17
.79
±0
.16
14
.42
±0
.29
24
.69
±0
.13
17
.62
±0
.36
25
.79
±0
.16
10
06
7.3
5±
0.5
62
0.7
5±
0.1
13
8.6
1±
0.4
82
2.3
5±
0.3
71
2.6
5±
0.2
31
7.2
9±
0.1
54
9.2
8±
0.6
72
9.8
7±
1.0
44
3.1
±0
.67
15
07
1.4
2±
0.6
27
.47
±0
.27
54
.12
±0
.42
26
.52
±0
.28
16
.81
±0
.32
1.8
4±
0.5
36
2.8
1±
0.4
13
7.2
8±
0.7
55
9.0
9±
0.1
5
20
08
8.2
8±
0.6
84
3.9
6±
0.0
98
2.5
3±
0.9
93
7.8
8±
0.7
72
1.6
3±
0.3
33
2.7
4±
0.3
98
7.0
7±
0.5
94
1.3
5±
0.4
75
.35
±0
.62
Val
ues
are
mea
ns
±S
Eo
fth
ree
det
erm
inat
ion
s
Table 4 Results of the three-way factorial ANOVA for the
effect of tannin on the enzyme activities in the Indian major
carps (F-values are significant at P \ 0.001)
Source Sum of squares df Mean square F-value
DOSE (D) 631.45 5 126.29 9,222.37
ENZYME (E) 51.07 2 25.54 1,864.87
FISH SP. (F) 72.76 2 36.38 2,656.61
D 9 E 142.28 10 14.23 1,038.99
D 9 F 81.24 10 8.12 593.26
E 9 F 36.78 4 9.19 671.55
D 9 E 9 F 43.79 20 2.19 159.91
Error 1.48 108 0.01
Total 1,060.86 161
Fish Physiol Biochem (2010) 36:1171–1180 1177
123

species studied and also plant material used in the fish
feed formulation. Present study also revealed varied
degrees of tannin-induced inhibition of digestive
enzymes from different carp species studied. In
agreement with the reports discussed above, it may
be assumed from the results of the present study that
tannin may affect feed utilization efficiency and
digestibility in the major carps if used in feed
formulation. Although results obtained from such in
vitro studies should not be extrapolated directly to in
vivo conditions as several other factors, such as
presence of food, pH, microbiota, intestinal secre-
tions, etc. may influence the adversity of dietary
tannin. However, result of the present study empha-
sizes the need for eliminating tannin from aquatic
macrophytes or other feed stuffs of plant origin to
replace fish meal successfully. Low-cost physical
methods like heat inactivation and water soaking
(Egounlety and Aworh 2003; Ramachandran and Ray
2008) or bioinactivation processes using tannase
producing microbiota may be practiced as indicated
in some of the recent literatures (for review see
Ghosh and Mukhopadhyay 2006; Mandal and Ghosh
2009b).
Although decrease in enzyme activity has been
assumed as the result of formation of insoluble
complexes with the dietary protein in most of the
observations (Makkar 1993), in the present study,
tannin at such low concentration in the in vitro
reaction mixture may form no or negligible amount
of complex which may rule out the possibility of non-
availability of substrate for enzymatic degradation
(Maitra and Ray 2003). Moreover, tannin not only
affected the protease activity, but also inhibited
comparatively lipase activities to a greater extent
and amylase activities to a lesser extent. Lee and Pan
(2003) indicated a non-competitive mixed-type inhi-
bition of in vitro trypsin activity from grey mullet
(Mugil cephalus) by tannin which may throw some
light on possible inhibitory mechanism of such
compound. Limited literature on this particular issue
suggests that more effort should be given to gain a
better insight to find out the mechanism of tannin-
mediated inhibition of digestive enzymes of fish and
its probable effect at the physiological level.
Acknowledgments The authors are grateful to the Head,
Department of Zoology, The University of Burdwan, West
Bengal, India and The Department of Science and Technology
(FIST programme), New Delhi, India for providing research
facilities. The authors are obliged to Dr. G. Aditya, Department
of Zoology, The University of Burdwan for rendering help in
statistical analyses of data. The first author is grateful to The
University of Burdwan, Burdwan for awarding the university
fellowship.
References
Alikunhi KH (1958) Observations on the feeding habits of
young carp fry. Indian J Fish 5(1):95–106
Al-Owafeir M (1999) The effects of dietary saponin and tannin
on growth performance and digestion in Oreochromisniloticus and Clarias gariepinus. Ph.D. Thesis, Institute of
Aquaculture, University of Stirling, UK, pp 220
Anson ML (1938) The estimation of pepsin, trypsin, papain,
and cathepsin with Hemoglobin. J Gen Physiol 22:79–89
Becker K, Makkar HPS (1999) Effects of dietary tannic acid
and quebracho tannin on growth performance and meta-
bolic rates of common carp (Cyprinus carpio L.). Aqua-
culture 175:327–335
Bernfeld P (1955) Amylase (alpha) and (beta). In: Colowick
SP, Kaplan NO (eds) Methods in enzymology, vol I,
Academic press, New York, pp 149–150
Bjorck IM, Nyman ME (1987) In vitro effects of phytic acid
and polyphenols on starch digestion and fiber degradation.
J Food Sci 52:1588–1594
Carmona A, Seild DS, Jaffe WG (1991) Comparison of
extraction methods and assay procedures for the deter-
mination of the apparent tannin Content of common
beans. J Sci Food Agric 56:291–301
Carmona A, Borgudd L, Borges G, Levy-Benshimol A (1996)
Effect of black bean tannins on in vitro carbohydrate
digestion and absorption. Nutr Biochem 7:445–450
Colowick SP, Kaplan NO (1955) Lipases. In: Methods in
enzymology, vol I. Academic press, New York, pp 627
Edwards P, Kamal M, Wee KL (1985) Incorporation of com-
posted and dried water hyacinth in pelleted feed for the
tilapia Oreochromis niloticus (Peters). Aquacult Fish
Manage 16:233–248
Egounlety M, Aworh OC (2003) Effect of soaking, dehulling,
cooking and fermentation with Rhizopus oligosporus on
the oligosaccharides, trypsin inhibitor, phytic acid and
tannins of soybean (Glycine max Merr.), cowpea (Vignaunguiculata L. Walp) and groundbean (Macrotyloma
geocarpa Harms). J Food Eng 56:249–254
Francis G, Makkar HPS, Becker K (2001) Antinutritional
factors present in plant-derived alternate fish feed ingre-
dients and their effects in fish. Aquaculture 199(3–4):197–
227
Garg SK, Makkar HPS, Nagal KB, Sharma SK, Wadhwa DR,
Singh B (1992) Toxicological investigations into oak
(Quercus incana) leaf poisoning in cattle. Vet Human
Toxicol 34:161–164
Ghosh K, Mukhopadhyay PK (2006) Application of enzymes
in aqua feeds. Aqua Feeds Formulation Beyond 3(4):7–10
Ghosh K, Sen SK, Ray AK (2002) Characterization of bacilli
isolated from gut of rohu, Labeo rohita, fingerlings and its
significance in digestion. J Appl Aquaculture 12:33–42
1178 Fish Physiol Biochem (2010) 36:1171–1180
123

Hagerman AE, Zhao Y, Johnson S (1997) Methods for deter-
mination of condensed and hydrolysable tannins. In:
Shahidi F (ed) Antinutrients and phytochemicals in foods.
American Chemical Society. ACS Symposium Series,
662, pp 209–222
Hasan MR, Moniruzzaman M, Farooque AMO (1990) Evalu-
ation of Leucaena and water hyacinth leaf meal as dietary
protein sources for the fry of Indian major carp, Labeo
rohita (Hamilton). In: Hirano R, Hanyu I (eds) Proceed-
ings of the second Asian fisheries forum. Asian Fisheries
Society, Manila, pp 275–278
Hossain MA, Jauncey K (1989) Nutritional evaluation of some
Bangladeshi oilseed meals as partial substitutes for fish
meal in the diet of common carp., Cyprinus carpio L.
Aquacult Fish Manage 20:255–268
Jhingran VG (1997) Fish and fisheries of India, 3rd edn edn.
Hindustan Publishing Corporation, Delhi, pp 335–337
Kalita P, Mukhopadhyay PK, Mukherjee AK (2007) Evalua-
tion of the nutritional quality of four unexplored aquatic
weeds from northeast India for the formulation of cost-
effective fish feeds. Food Chem 103:204–209
Karasov WH, Meyer MW, Darker BW (1992) Tannic acid
inhibition of amino acid and sugar absorption by mouse
and vole intestine: test following acute and subchronic
exposure. J Chem Ecol 18:719–736
Khan RA, Siddiqui AQ (1973) Food selection by Labeo rohita(Ham.) and its feeding relationship with other major
carps. Hydrobiologia 43:429–442
Kinnear PR, Gray CD (2000) SPSS for Windows made simple.
Release 10. Psychology Press, Sussex
Krogdahl A (1989) Alternative protein sources from plants
contain anti-nutrients affecting digestion in salmonids. In:
Takeda M, Watanabe T (ed) The current status of fish
nutrition in aqua-culture. Proceedings of the third inter-
national symposium on feeding and nutrition in fish.
August 28–September 1. Tokyo University of Fisheries,
Tokyo, pp 253-261
Lee SM, Pan BS (2003) Effect of dietary sorghum distillery
residue on hematological characteristics of cultured grey
mullet (Mugil cephalus)—an animal model for presc-
reening antioxidant and blood thinning activities. J Food
Biochem 27(1):1–18
Liener IE (1989) Antinutritional factors in legume seeds:
state of the art. In: Huisman J, van der Poel AFB,
Liener IE (eds) Recent advances in research in anti-
nutritional factors in legume seeds. Pudoc, Wageningen,
pp 6–14
Lowry OH, Ronebrough NJ, Farr AL, Randell RJ (1951)
Protein measurement with Folin phenol reagent. J Biol
Chem 193:265–276
Maitra S, Ray AK (2003) Inhibition of digestive enzymes in
rohu, Labeo rohita (Hamilton) fingerlings by tannin, an in
vitro study. Aquacult Res 34:93–95
Makkar HPS (1993) Antinutritional factors in foods for live-
stock. In: Gill M, Owen E, Pollot GE, Lawrence TLJ (eds)
Animal production in developing countries. Occasional
publication no. 16. British Society of Animal Production,
pp 6–14
Makkar HPS, Blummel M, Becker K (1995) In vitro effects
and interactions of tannins and saponins and fate of tan-
nins in rumen. J Sci Food Agric 69:481–493
Mandal S, Ghosh K (2009a) Accumulation of tannin in dif-
ferent tissues of Indian major carps and exotic carps.
Aquacult Res 1–4 (Early view) doi:10.1111/j.1365-
2109.2009.02371.x
Mandal S, Ghosh K (2009b) Development of plant-derived
low-cost fish feed through overcoming adverse effects of
plant anti-nutrients. Fishing Chimes 29(1):156–161
Mohanty RK (2003) Feed intake pattern and growth perfor-
mance of Indian Major Carps and freshwater prawn in a
rice-fish integration system. J Asian Fish Sci 16:307–316
Moore S, Stein WW (1948) Photometric ninhydrin method for
use in the chromatography of amino acids. J Biol Chem
176:367–388
Mueller-Harvey I, McAllan AB (1992) Tannins: their bio-
chemistry and nutritional properties. Adv Plant Cell Bio-
chem Biotechnol 1:151–217
Mukhopadhyay N, Ray AK (1996) The potential of deoiled sal
(Shorea robusta) seed meal as a feedstuff in pelleted feed
for Indian major carp, rohu, Labeo rohita (Hamilton)
fingerlings. Aquacult Nutr 2:221–227
Olvera NMA, Martinez P, Galvan CR, Chavez SC (1988) The
use of seed of the leguminous plant Sesbania grandijloraas a partial replacement for fish meal in the diets for
tilapia (Oreochromis mossambicus). Aquaculture 71:
51–60
Patra BC, Ray AK (1988) A preliminary study on the utiliza-
tion of the aquatic weed. Hydrilla verticillata (L.f.) Rayle
as feed by the carp, Labeo rohita (Hamilton): growth and
certain biochemical composition of flesh. Ind Biol
20(1):44–50
Patra BC, Maity J, Banerjee S, Patra S (1999) Making aquatic
weeds useful III: nutritive value of Nechamandraalternifolia (Roxb.ex Weight) Thw. meal as feed for the
Indian major carps Labeo rohita (Hamilton). J Biol Sci
5:35–45
Patra BC, Maity J, Debnath J, Patra S (2002) Making aquatic
weeds useful II: Nymphoides cristatum (Roxb.) O. Kuntze
as feed for an Indian major carp Labeo rohita (Hamilton).
Aquacult Nutr 8:33–42
Prusty AK, Sahu NP, Pal AK, Reddy AK, Kumar S (2007)
Effect of dietary tannin on growth and haemato-immu-
nological parameters of Labeo rohita (Hamilton) finger-
lings. Anim Feed Sci Technol 136:96–108
Rahman MM, Verdegem MCJ, Nagelkerke LAJ, Wahab MA,
Milstein A, Verreth JAJ (2006) Growth production and
food preference of rohu Labeo rohita (H.) in monoculture
and in polyculture with common carp Cyprinus carpio(L.) under fed and non-fed ponds. Aquaculture 257:359–
372
Ramachandran S, Ray AK (2008) Effect of different process-
ing techniques on the nutritive value of grass pea Lathyrussativus L., seed meal in compound diets for indian major
carp rohu, Labeo rohita (Hamilton), fingerlings. Arch Pol
Fish 16:189–202
Ray AK, Das I (1992) Utilization of diets containing com-
posted aquatic weed (Salvinia cuculata) by the Indian
major carp, rohu (Labeo rohita Ham.) fingerlings. Biore-
sour Technol 40:67–72
Ray AK, Das I (1994) Apparent digestibility of some aquatic
macrophytes in rohu Labeo rohita (Ham.) fingerlings.
J Aquacult Tropics 9:335–342
Fish Physiol Biochem (2010) 36:1171–1180 1179
123

Ray AK, Das I (1995) Evaluation of dried aquatic weed, Pistiastratiotes meal as a feedstuff in pelleted feed for rohu,
Labeo rohita fingerlings. J Appl Aquacult 5:35–44
Schanderi SH (1970) Methods in food analysis. Academic
Press, New York, p 709
Silanikove N, Perevolotsky A, Provenza FD (2001) Use of
tannin-binding chemicals to assay for tannins and their
negative post ingestive effects in ruminants. Anim Feed
Sci Technol 91:69–81
Singh U (1984) The inhibition of digestive enzymes by poly-
phenols of chick pea (Cieer arrierinum L) and pigeon pea
(Cajamus cajan (L.) Millsp.). Nutr Rep Intern 29:745–753
Vohra P, Kratzer FH, Joslyn MA (1966) The growth depressing
and toxic effects of tannins to chicks. Poultry Sci 46:135–142
Wee KL, Wang SS (1987) Nutritive value of Leucaena leaf
meal in pelleted feed for Nile Tilapia. Aquaculture 62:97–
108
Welsch CA, Lachance PA, Wasserman BP (1989) Effects of
native and oxidized phenolic compounds on sucrase
activity in rat brush border membrane vesicles. J Nutr
119:1737–1740
Wynne-Edwards KE (2001) Evolutionary biology of plant
defenses against herbivory and their predictive implica-
tions for endocrine disruptor susceptibility in vertebrates.
Environ Health Perspect 109(5):443–448
Zar JH (1999) Biostatistical analysis, IV ed. Pearson Education
Singapore Pte. Ltd (Indian Branch). New Delhi, India, p
663
1180 Fish Physiol Biochem (2010) 36:1171–1180
123
Copyright © 2022 FDOKUMEN