
INFORMATION TO USERS - University of Victoria
294
INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, vriiile others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e g., maps, drawings, charts) are reproduced by sectioning the original, b%inning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. Each original is also photographed in one exposure and is included in reduced form at the back of the book. Photographs included in the original manuscript have been reproduced xerographically in this copy. Higher quality 6” x 9” black and white photographic prints are available for any photographs or illustrations appearing in this copy for an additional charge. Contact UMI directly to order. UMI ABell &Hbwell Infonnation Compaiy 300 North Zeeb Road, Ann Arbor MI 48106-1346 USA 313/761^700 800/521-0600
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of INFORMATION TO USERS - University of Victoria

INFORMATION TO USERS
This manuscript has been reproduced from the microfilm master. UMI
films the text directly from the original or copy submitted. Thus, some
thesis and dissertation copies are in typewriter face, vriiile others may be
from any type o f computer printer.
The quality of this reproduction is dependent upon the quality of the
copy submitted. Broken or indistinct print, colored or poor quality
illustrations and photographs, print bleedthrough, substandard margins,
and improper alignment can adversely affect reproduction.
In the unlikely event that the author did not send UMI a complete
manuscript and there are missing pages, these will be noted. Also, if
unauthorized copyright material had to be removed, a note will indicate
the deletion.
Oversize materials (e g., maps, drawings, charts) are reproduced by
sectioning the original, b%inning at the upper left-hand comer and
continuing from left to right in equal sections with small overlaps. Each
original is also photographed in one exposure and is included in reduced
form at the back of the book.
Photographs included in the original manuscript have been reproduced
xerographically in this copy. Higher quality 6” x 9” black and white
photographic prints are available for any photographs or illustrations
appearing in this copy for an additional charge. Contact UMI directly to
order.
UMIABell &Hbwell Infonnation Compaiy
300 North Zeeb Road, Ann Arbor MI 48106-1346 USA 313/761^700 800/521-0600


Pro tein-N ucleic A d d In teractions of V\^Ims* T um or and TFIIIA ZincFinger Proteins.
byTATYANA HAMILTON
B. Sc., Novosibirsk State University, Russia, 1990
A D issertation Subm itted in Partial Fulfillm ent of the Requirem ents for the Degree of
DOCTOR OF PHILOSOPHY in the D epartm ent of Biochemistry a n d M icrobiology
We accept this thesis as conform ing to ih e i^buired standard
rvisor (Dept, of Biochemistry & Ibiology)
Olafson, D epartm ental Member 'o f B io ch e n jis^ & N/p:Sob^ology)
Dr. F rands E. N ano, Deps(Dept, of Bi
Member logy)
chael J. A shm (Dept, of Biology)
TJenry Pearson, Departm ental Member (Dept, of BigdaggUjtry & Microbiology)
Dr. D avid Setzer, E ^ m a l Exam iner Case W estern Reserve U niversity)
© TATYANA B. HAMILTON, 1997 U niversity of Victoria
All rights reserved. This thesis m ay not be reproduced in w hole or in part, by m im eograph or o ther m eans, w ithout the perm ission of the au th o r.

Supervisor: Dr. Paul J. Romaniuk
Abstract
This Fh.D . w ork represents the study of nucleic a d d interactions
o f tw o zinc finger proteins: m am m alian W ilm s' tum or su p p resso r
(W Tl) and Xenopus transcription factor UIA proteins (TFIIIA).
W Tl is a pu tative transcriptional regu latory p ro te in w h ich is
inactiva ted in a sub type of W ilm s' tum ours. Using se lec tion and
am plification b ind ing assay (SAAB) we determ ined that the h ighest
affinity b inding sites for WT1[-KTS] consist of a 12 base pa ir sequence
GCG-TGG-GCG-(T/G)(G/A/T)(T/G). These sequences have a four-fold
h igher affinity for the protein than the nonselected sequence GCG-
T G G -G C G -C C C , as m easured by a quantita tive nitrocellulose filter
binding assay.
The effects of Denys-Drash syndrom e (DOS) point m utations on
th e DNA b in d in g activity of W Tl were determ ined. SAAB assay
revealed th a t none of the DOS m utan t p ro te ins give rise to a new
sequence specifid ty . One m utation, R394W abolishes specific b ind ing
o f the pro tein . The rem ain ing m utations resu lt in red u ced DNA-
b ind ing activity, ranging from 1.4 to 14-fold, w hich suggests th a t even
sm all changes in D N A -binding activity m ay p red p ita te the clinical
phenotype of Denys-Drash syndrom e.
C om parative analysis of the DNA b in d in g characteristics of
W ilm s' tum our and Early grow th response pro teins was conducted .
T he stoichiom etry of the D N A -protein com plexes, their s tab ility to
d issodation , and the effects of pH , tem perature and salt concentration
o n the equilib rium binding of these proteins to their cognate DNA

n i
sequences have been determ ined. U nder the conditions of 0.1 M sa lt,
pH 7.5, a n d 22 * C W Tl-ZF has an apparen t dissociation constant (Kd)
of 1.14± 0.2 X 1 0 ^ M, and EGR-1 protein has a Kd of 3.55 ± 0.4 x 10"^ M.
In ad d itio n , w e tested rela tive contribution of each base pair in th e
consensus binding site to the high affinity b inding by po in t m utational
analysis, an d identified im portan t differences that exist in the b ind ing
m odes o f the tw o proteins.
T ran sc rip tio n factor UIA controls the ex p ress io n of the 5S
ribosom al RNA genes d u rin g developm ent of Xenopus laevis., a n d
specifically interacts w ith bo th 5S DNA an d 5S rRNA m olecules. T he
presen t s tu d y assesses contributions of the central z inc fingers fo u r
th ro u g h seven to specific DNA and RNA b ind ing activ ities of th e
protein. The results dem onstrate that each zinc finger in the zf 4-7
reg io n c o n trib u te s to b o th the h ig h affin ity D N A and R N A
in teractions: the largest effect on TFlllA-DNA b ind ing (10-fold) w as
p roduced w hen zinc finger 5 of TFIHA w as replaced w ith the do n o r
sequences of either p43 or W Tl. However, while all the zinc fingers 4-7
contribute to the high affinity 5S rRNA binding, substitu tion of an a -
helical p o rtio n of zinc finger 6 w ith the equivalen t sequences from
W Tl abo lished RNA -binding activity of TFIILA, suggesting that z inc
finger 6 p lays a particularly im portant role in binding to RNA.

IV
Exam iners:
Dr. P au l J. Rbmani rvisor (Dept, of B iochem istry & o o lo g y )
)lafson. D epartm ental M em ber (D ept^f Biochemistryj)^;^crobiology)
Dr. Francis E. Nano, D epartm ental M em ber (D ept, oT ^ochem istry & Microbiology)
Dr. 4rerry|
SnuthDr. M ichael J. Ashwi M em W r (Dept, of Biology)
Dr. D avid Setzer, External Examiner (Case W estern ReserveU niversity)

Table of Contents
Abstract.................................................................................................................................. ii
Table of C onten ts............................................................................................................... v
List of Tables..........................................................................................................................x
List of Figures.....................................................................................................................xii
List of A bbreviations....................................................................................................xv ii
Acknowledgm ents............................................................................................................. xx
C h ap te r 1.0 Early grow th response p ro te in (EGR-1): a p ro to typical
m em ber of a C2H 2 zinc finger family of transcription factors
1.1 Introduction............................................................................................................. 1
1.1.1 Overview of the EGR-1 family of transcription factors............................... 1
1.1.2 Gene targets of EGR-1 regulation .......................................................................6
1.1.3 Identification of EGR-1 cDNA, and characterization o f its protein
p ro d u c t.................................................................................................................................7
1.1.4 DNA-binding function of EGR-1.....................................................................14
1.1.5 Structures of other zinc finger-DNA complexes.......................................... 32
1.1.6 Studies tow ard a zinc finger recognition code..............................................43
C h ap te r 2.0 The W ilms' tum our gene product: a tum our suppressor
involved in regulation of k idney developm ent
2.1 Introduction.......................................................................................................... 47
2.1.1 The concept of tum or suppressor g e n es ....................................................... 47
2.1.2 Biology of Wilms' tum or and associated syndrom es................................ 47
2.1.3 Identification and characterization of the W Tl gene an d its
protein product................................................................................................................. 50
2.1.4 DNA-binding activity of Wilms’ tum or protein..........................................56

VI
2.1.5 Transcriptional regulatory functions o f W T l............................................. 65
C hap ter 3.0 Identification of h igh affinity b ind ing sites for the W ilms'
tum our suppressor protein W Tl
3.1 Introduction.......................................................................................................... 72
3.2 Materials and M ethods.......................................................................................75
3.2.1 Bacterial strains and DNA vectors.................................................................. 75
3.2.2 Other M aterials.....................................................................................................75
3.2.3 C onstruction and Purification of R ecom binant W T l-Z FP and
W TIAFI-ZFP P ro te in s ...................................................................................................76
3.2.4 Selection Amplification and Binding (SAAB) A ssay ................................80
3.2.5 Construction of plasm ids containing sequence elem ents o f the
insulin-like grow th factor II fetal prom oter, and the non-selected
sequence w ith the CCC subsite for finger 1 b ind ing ............................................ 86
3.2.6 Purification of oligonucleotides..................................................................... 87
3.2.7 End-labeling of DNA...........................................................................................88
3.2.8 Nitrocellulose Filter Binding A ssay ...............................................................88
3.2.9 Transient Transfection A ssays.........................................................................89
3.3 Results........................................................................................................................ 90
3.3.1 Isolation o f DNA bind ing subsite for finger 1 of W T l-Z F by
SA A B ..................................................................................................................................90
3.3.2 Q uantitative Binding of W Tl-ZFP to Various DNA Sequences................93
3.3.3 A n In vitro Selected W Tl Binding Site Acts as a Strong
Transcriptional Regulator...............................................................................................97
3.4 Discussion.................................................................................................................101
C hap ter 4.0 W Tl m utations and the Denys-Drash syndrom e

vu
4.1 In troduction ..........................................................................................................108
4.2 M aterials an d M ethods.....................................................................................113
4.2.1 Construction and expression of Denys-Drash m utant p ro te in s .............113
4.2.2 Selection amplification and binding (SAAB) assay................................... 114
4.2.3 Nitrocellulose Filter Binding A ssay............................................................ 117
4.3 R e su lts .................................................................................................................. 118
4.3.1 Selection of DNA binding sites for D enys-D rash m u tan t p ro te in s 118
4.3.2 M easuring the binding affinities of the m u tan t pep tid es for the
selected DNA sequences.............................................................................................. 121
4.4 D iscussion............................................................................................................ 125
C h ap te r 5.0. Com parative analysis of the DNA binding characteristics
o f W ilm s' tum our and Early G row th Response Proteins
5.1 In troduction .........................................................................................................129
5.2 M aterials an d M ethods..................................................................................... 130
5.2.1 Construction and purification of recom binant W Tl-ZF
and EGRl-ZF proteins....................................................................................................130
5.2.2 Construction of m utant W Tl-ZF and EGRl-ZF DNA b ind ing
seq u en ces ......................................................................................................................... 130
5.2.3 End-labeling of DNA......................................................................................... 132
5.2.4 N itrocellulose Filter Binding A ssay............................................................. 132
5.3 R esults.................................................................................................................. 132
5.3.1 Equilibrium binding constan ts ...................................................................... 132
5.3.2 M onovalent salt dependence of the Ka for DNA bind ing .......................139
5.3.3 p H dependence of K a.......................................................................................141
5.3.4 Effect of d ivalent metal ion concentration o n DNA b in d in g ................144
5.3.5 Tem perature dependence o f the Ka v a lu e ................................................. 144

vm
5.3.6 C ation and anion effects on binding ..............................................................147
5.3.7 Identification o f relative contributions of each base pair in the
consensus site to the b inding specificities of W Tl-ZF and EGR-1
p ro te in s ............................................................................................................................ 149
5.4 Discussion............................................................................................................. 155
C hap ter 6.0 TFIHA - a representative of the C2H 2 fam ily of zinc finger pro teins6.1 The C2H 2 zinc finger dom ain ....................................................................... 163
6.2 Structure / function analysis of TFIHA......................................................... 173
6.2.1 Purification and characterization of TFIHA gene p roduct....................173
6.2.2 TFIIIA gene expression.....................................................................................179
6.2.3 Roles of TFIHA in transcription and storage of 55 rRNA..................... 180
6.2.4 Structure and function of 5S rRNA and its gene.......................................182
6.2.5 Role of TFIIIA zinc fingers in RNA recognition .................................... 183
6.2.6 Role of TFIIIA zinc fingers in DNA recognition .................................... 194
C hapter 7.0 The study of the nucleic acid interactions of zinc finger 4-7
region of TFIHA
7.1 In tro d u c tio n ...................................................................................................... 206
7.2 M aterials and M ethods.................................................................................. 207
7.2.1 Construction of TFIIIA finger sw ap m u tan ts .......................................... 207
7.2.2 Expression and purification of TFIHA p ro te in s ........................................213
7.2.3 Radiolabeling o f 5S DN A................................................................................ 218
7.2.4 Synthesis and Radiolabeling of 5S rRNA.................................................... 218
7.2.5 Nitrocellulose filter binding assays............................................................ 221
7.3 R e su lts ................................................................................................................ 221

DC
7.3.1 Effects o f zinc finger su b stitu tio n m utagenesis on the D N A
binding activity of TFIIIA.............................................................................................221
7.3.2 Effects of zinc finger substitu tion m utagenesis on the RN A
binding activity of TFIIIA.............................................................................................228
7.4 Discussion............................................................................................................230
8.0 C onclusions....................................................................................................... 236
9.0 L iterature cited................................................................................................... 239

List of Tables
Table 3.1 Frequencies of Nucleotides Selected by WTl-ZFP and
WTIAFI-ZFP......................................................................................................................92
T able 3.2 Relative Affinities of W Tl B inding Sites for W Tl-ZFP and
WTIAFI-ZFP......................................................................................................................96T able 4.1 Finger 2 subsite sequences selected from a random ized template by w ü d type W Tl-ZFP and the finger 2 point m utan ts.....................119
Table 4.2 Frequencies of each nucleotide selected at the finger 2 subsite
positions by W Tl-ZFP and the finger 2 po in t m utants.......................................120
Table 4.3 Frequencies of each nucleotide selected at the finger 3 subsite
positions by W Tl-ZFP and the finger 3 po in t m utants.......................................120
T ab le 4.4 Finger 3 subsite sequences selected from a random ized
template by w ild type W Tl-ZFP and the finger 3 point m utan ts.....................122
Table 4.5 Binding affiiüties of wild type and finger 2 m utan t W Tl-ZFP
for selected DNA sequences........................................................................................ 124
Table 4.6 Binding affinities of wild type and finger 3 m utan t W Tl-ZFP
for selected DNA sequences........................................................................................ 124
T able 5.1 Sequences of m utan t oligonucleotides harbouring indiv idual
base pair substitutions in the EGRl-ZF and W Tl-ZF consensus DNA
binding site...................................................................................................................... 133
T ab le 5.2 Effect of the different m onovalent salts on the b ind ing of
W Tl-ZF and EGRl-ZF to DNA consensus sequences.........................................148
T able 5.3 Dissociation constants for W Tl-ZF and EGRl-ZF b inding to
w ild-type and m utant D N A consensus sequences.............................................. 150
T able 7.1 Sequences of oligonucleotides used in construction of TFIIIA
substitution m utants......................................................................................................208

XI
T ab le 7.2 Sequences of se lec tion p rim ers u sed in tran sfo rm e r
m utagenesis protocol to construct substitu tion m utan ts of TFIHA...............216
T able 7.3 The effects of TFIHA zinc finger substitu tion m utations on
the DNA and RNA b inding of the factor................................................................ 223
T able 7.4 The effects of TFIIIA zinc finger substitu tion m utations on
the DNA and RNA b inding of the factor................................................................ 227

XU
List of Figures
Figure 1.1 Biological processes m ediated by EGR-1 in response to
diverse stim uli (Cashier & Sukhatm e, 1995)............................................................ 2
Figure 1.2 C om parison of DNA-binding dom ains of the EGR-1 fam ily
of proteins............................................................................................................................4
Figure 1.3 Schem atic structure and amino a d d sequence of EGR-1
p ro te in ................................................................................................................................ 10
Figure 1.4 Sum m ary of EGR-1 functional dom ains............................................. 13
Figure 1.5 Sequence of the EGR-1 zinc finger peptide and of the DNA
binding site used in cocrystallization........................................................................ 16
Figure 1.6 The overall arrangem ent of the three zinc fingers of EGR-1
in the major groove of DNA (Pavletich & Pabo, 1991).........................................17
Figure 1.7 Sketch sum m arizing the principal am ino add-base contacts
as seen in the original EGR-1 X-ray structure (Pavletich & Pabo, 1991)...........19
Figure 1.8 Sum m ary of direct base and phosphate contacts in EGR-1-
DNA complex...................................................................................................................20
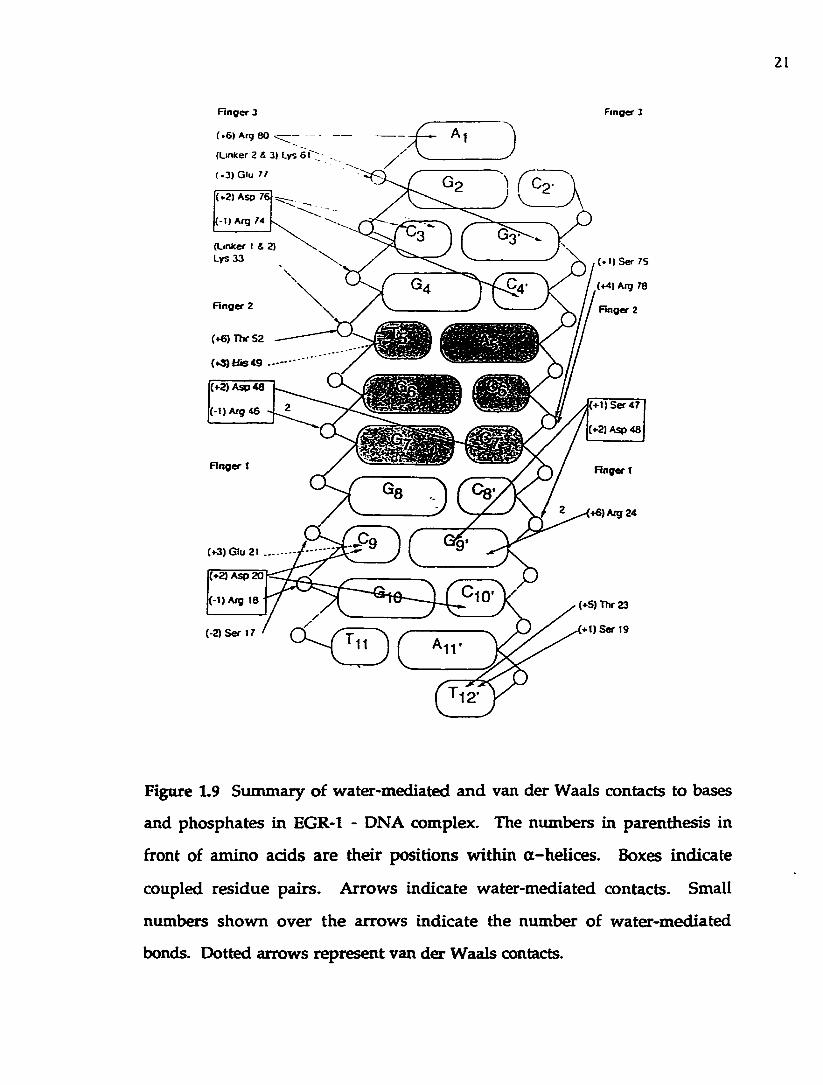
Figure 1.9 Sum m ary of w ater-m ediated and van der Waals contacts to
bases and phosphates in EGR-l-DNA complex...................................................... 21
Figure 1.10 Drawings of amino add-base pair contacts of the EGR-l-
DNA complex...................................................................................................................22
Figure 1.11 Schem atic diagram of EGR-1 zinc fingers interacting w ith
the proposed overlapping, 4-bp DNA subsites (Isalan et al., 1997)..................... 27
Figure 1.12 Schematic representation of hydrogen bonding betw een
Zn-coordinated histidine and DNA backbone........................................................29
Figure 1.13 Sequences of the GLI zinc finger dom ain and the DNA-
binding site used for cocrystallization.......................................................................33

x m
Figure 1.14 Sketch sum m arizing base and phosphate contacts m ade by
the GLI peptide................................................................................................................ 34
Figure 1.15 The sum m ary of Tramtrack-DNA contacts...................................... 36
Figure 1.16 Schematic summary of the principal protein-DNA contacts
observed in the cocrystal structures of the EGR-1 and Tram track.....................37
Figure 1.17 Schematic representation of the YYl-DNA interactions.............. 40
Figure 1.18 Com parison of YYl w ith other zinc finger structures...................41
Figure 2.1 The am ino acid sequence of W Tl pro tein .......................................... 52
Figure 2.2 Schematic diagram of the structure of the WTl m RN A and
p ro te in s ............................................................................................................................. 54
Figure 2.3 DNA sequences which bind W Tl proteins
(Reddy & Licht, 1996)..................................................................................................... 58
Figure 2.4 Protein-DNA contacts m ade by EGR-1 protein, and p roposed
DNA contacts for W Tl zinc fingers........................................................................... 62
Figure 2.5 Proposed W Tl zinc finger-DNA contacts (Reddy & Licht,
1996)....................................................................................................................................64
Figure 3.1 DNA sequence of the peptide encoding insert in p lasm id
pET-WTZFP, w ith the amino a d d sequence of the peptide................................73
Figure 3.2 Schematic representation of the recom binant W T l-Z F
plasm id, pUC18/W Tl-ZF. Restriction sites used in subsequent c lon ing
steps are show n............................................................................................................... 78
Figure 3.3 Plasmid m ap of pET-16b expression vector........................................ 79
Figure 3.4 Coomassie blue-stained 15% SDS-polyacrylamide gel
show ing purifiied W Tl-ZFP and W TIAFI-ZFP proteins....................................81
Figure 3.5 Protocol for the SAAB (Selection and Am plification of
B inding site assay)...........................................................................................................82

XIV
Figure 3.6 T he o ligonuc leo tide c o n ta in in g a ran d o m ized ta rg e t
sequence for SAAB analysis of WT1[-KTS] finger 1 subsite................................83
Figure 3.7 A n autoradiogram m of a SAAB ro u n d ............................................... 85
Figure 3.8 Sequences of bases 10-14 arising from the random SAAB
tem plate after four rounds of selection w ith W Tl-ZFP.......................................91
Figure 3.9 R esults of an nitrocellulose filter b ind ing assay m easuring
the equ ilib rium b ind ing of W Tl-ZFP an d W TIAFI-ZFP to tw o W Tl
elements...............................................................................................................................95
F igure 3.10 (A) Sequences of W Tl elem ents identified in the fetal
prom oter of the IGF-11 gene. (B) Results of an n itrocellu lose filter
b ind ing a ssay m easu ring the equ ilib rium b in d in g of W T l-Z F P to
different D N A elem ents..................................................................................................98
F ig u re 3 .11 T ra n s ie n t tra n s fe c tio n a ssay s m e a su r in g W Tl
responsiveness................................................................................................................100
Figure 3.12 The hydrogen-bond donors an d acceptors p resen ted by
W atson-Crick base pairs to the major groove and the m inor g roove........... 103
Figure 4.1 W T l m utations associated w ith Denys-Drash syndrom e.............109
Figure 4.2 C o m parison of the sequences of W T l and EGR-1 zinc
fingers.................................................................................................................................112
Figure 4.3 C oom assie b lue-sta ined 15% SD S-polyacrylam ide gel
show ing pu rifiied W Tl-ZFP Denys-Drash m u tan ts........................................... 115
F ig u re 4.4 T he SAAB tem p la te o lig o n u c leo tid e s c o n ta in in g
random ized sequences for the WT1[-KTS] finger 2 (A) or finger 3 (B)
recognition........................................................................................................................116
F igure 4.5 T he equilib rium b ind ing of the W Tl elem ent GCG TGG
GAG TGT to W Tl-ZFP, R366H and R366C.............................................................123

XV
Figure 5.1 Coomassie blue-stained 15% SDS-polyacrylamide gel
show ing EGRl-ZF pro tein purified by affinity chrom atography......................131
Figure 5.2 Equilibrium binding curves of W Tl-ZF and EG Rl-ZF
pro teins to their target DNA sequence.................................................................... 135
Figure 5.3 Scatchard analysis of W T1-2T (A) and EGRl-ZF..............................137
F igure 5.4 Time dependence of the binding of W Tl-ZF and EG Rl-ZF to
DNA consensus sequences..........................................................................................140
Figure 5.5 KCl concentration dependence of the binding of W T l-Z F
and EG R l-ZF to DNA consensus sequences.......................................................... 142
Figure 5.6 pH dependence of the binding of W Tl-ZF and EGRl-ZF to
DNA consensus sequences..........................................................................................143
F igure 5.7 Effect of the m agnesium ion concentration on the b ind ing of
W Tl-ZF (closed circles) and EGRl-ZF (open circles) to DNA consensus
sequences.......................................................................................................................... 145
Figure 5.8 T em perature dependence of the binding of W Tl-ZF an d
EG Rl-ZF to DNA consensus sequences.................................................................. 146
Figure 6.1 Schematic representation of a C2H 2 zinc finger...............................165
F igure 6.2 Representative structures from various zinc finger fam ilies........170
Figure 6.3 Schematic representation of the interfinger o rientations of
MBP-1 zinc fingers and EGR-1 fingers 1 and 2.......................................................174
Figure 6.4 Diagrams of the functional domains of eukaryotic zinc-
con tainng transcription factors. 175
F igure 6.5 Amino a d d sequence of the nucleic ad d binding do m ain of
TFIIIA .................................................................................................................................177
Figure 6.6 Schematic diagram illustrating alignm ent of TFIIIA zinc
finger region along ICR................................................................................................ 178
F igure 6.7 O rganization of the Xenopus 5S rRNA genes...................................184

XVI
Figure 6.8 The secondary structure of 5S rRNA. 185
Figure 6.9 M odel for the TFIIIA - 5S rRNA interaction.....................................188
Figure 6.10 A lignm ent of zinc finger sequences of TFIIIA and p43
p ro te in s.............................................................................................................................189
Figure 6.11 A schem atic representation of the Xenopus laevis 5S rR N A
gene in ternal control region......................................................................................195
Figure 6.12 M odels for TFIIIA - 5S DNA complex.............................................. 201
Figure 7.1 Schem atic diagram illustrating construction of (A) pUC19-
zf4-7; (B) pUC19-W Tl-BX...........................................................................................210
Figure 72. Schematic chart of the PCR-mediated site-directed
m utagenesis protocol used to create TFIIIA substitution m utants................. 212
Figure 7.3 Schematic diagram show ing the strategy for in troducing
m utations using the Transform er site-directed m utagenesis k it
(Clontech Inc., 1994)..................................................................................................... 214
Figure 7.4 Coom assie blue-stained 15% SDS-polyacrylamide gel
show ing TFIIIA w ild type and m utant proteins................................................. 219
Figure 7.5 Coom assie blue-stained 15% SDS-polyacrylamide gel
show ing TFIIIA w ild type and m utant proteins................................................. 220
Figure 7.6 C om parison of the amino acid sequences of zinc fingers 4-6
of the dono r pro tein p43 w ith the zinc fingers 4-6 of TFIIIA............................222
Figure 7.7 Sam ple nitrocellulose filter binding curves of TFIIIA w ild
type and m u ta n t proteins w ith the 58 rRNA gene..............................................225
Figure 7.8 C om parison of the amino a d d sequences of zinc fingers 1-4
of the dono r W Tl-ZF w ith the zinc fingers 4-7 of TFIIIA................................. 226
Figure 7.9 C om petition assay of TFIIIA and TF(4-7)WT for 5SrR N A
binding using tRNA^*^® as a competitor.................................................................229

xvu
List of Abbreviations
bp: base pair
BSA: bovine serum albumin
BWS: Beckwith-W iedem ann syndrom e
CAT: Chloram phenicolA cetylTransferase
cDNA: com plem entary deoxyribonucleic acid
cpm : counts per m inute
DDS: D enys-D rash syndrome
deoxynucleotide triphosphates:
dA TP, deoxyadenosine triphosphate
dCTP, deoxycytidine triphosphate
dG T P, deoxyguanine triphosphate
dTTP, deoxythym idine triphosphate
DEPC: diethy pyrocarbonate
DNA: deoxyribonucleic acid
DTT: dithiotreitol
E.coli: Escherichia coli
EDTA: ethylenediam ine-tetraacetic acid
EGRl: Early G row th Response 1
F.U.P.: Forw ard Universal Prim er
HEPES: N-2-hydroxyethylpiperazine-N '-2-ethanesulphonic acid
ICR: internal control region
IE: in term ediate elem ent
IGFII: Insulin-like G row th Factor II
IPTG: isopropyl-P-D-thiogalactopyranoside
KTS: lysine, threonine, serine

xvm
LB: Luria-Benton b ro th
LOH: Loss O f Heterozygosity
m RN A : m essenger ribonucleic acid
M W : m olecular w eight
N M R: nuclear m agnetic resonance
nt: nucleotide
N ucleotide triphosphates:
GTP, guanine triphosphate
CTP, cytidine triphosphate
ATP, adenosine triphosphate
UTP, u rid ine triphosphate
nucleotide bases:
G , guanine
C, cytosine
A, adenine
T, thym idine
N , either A, A, G, or T
PAGE: polyacrylam ide gel electrophoresis
PDGF-A: Platelet-D erived G row th Factor A
PEG: polyethylene glycol
PM SF: phenylm ethylsulfonyl fluoride
PPG: 2,5-diphenyloxazole
R.U.P.: Reverse U niversal P rim er
RB: retinoblastom a
RNA: ribonucleic acid
RN ase: ribonuclease
R N asin : ribonuclease inhibitor

XIX
RN P: ribonucleoprotein
rRN A : ribosom al nucleic acid
S: Svedberg unit
SAAB: selected amplification and binding
SAAB: Selection A nd Amplification Binding assay
SDS: sodium dodecyl sulphate
TBE: Tris base, borate, EDTA
TFIIIA: transcription factor UIA
TFIIIB: transcription factor UIB
TFIIIC: transcription factor HIC
T ris-H C l: tris-(hydroxym ethyl) am inom ethane hydrochloride
tRN A : transfer ribonucleic acid
TTK: T ram TracK protein
W A G R : W ilm s' tum our. A nirid ia , uroG enital m alform ations, m ental
R e ta rd a tio n
W T l: W ilm s' T um our 1
Xbo: Xenopus borealis oocyte
Xbs: Xenopus borealis somatic
Xlo: Xenopus laevis oocyte
Xls: Xenopus laevis somatic
XIt: Xenopus laevis trace-oocyte

XX
A cknow ledgm ents
I w ould like to thank m y supervisor. Dr. Paul J. Rom aniuk for
g iv ing m e the w onderfu l o p p o rtu n ity to join h is lab, and fo r h is
continuous support, encouragem ent and guidance over the years.
V aluable assistance was p ro v id ed by all the m em bers o f m y
supervisory committee: Dr. Robert W. Olafson, Dr. Francis E. N ano, Dr.
Terry W . Pearson and Dr. Michael J. Ashwood-Sm ith, for w hich I am
grateful. I would also like to m ention Dr. A1 T. M atheson, Dr. Ed E.
Ishiguro and Dr. Juan Ausio for their assistance.
M y sincere thanks to all the mem bers of Dr. Rom aniuk's lab -
Kathy Barilla, Frank Borel, John Ferris, Steve H endy , Maya Iskandar,
Colleen Nelson, N ik Veldhoen, Judy Wise, Q im in You, and W ei-Q ing
Z ang - for their help and friendship . My thanks also to the o ther
g raduate students a t the departm ent w ho d idn 't hesitate to help: G erry
Bearon, Siobhan C ow ley, Ben Forw ard , Lee A n n H ow e, A rm an d o
Jardim , Kizzy Mdluli, Dmitrii Rodionov, Caroline Stebeck.
I am grateful to Albert Labossiere and Scott Scholz, w ho w ere
always there to help w ith all m anner of technical assistance from fixing
a com puter to setting up a photo room.
I w ould very m uch like to acknowledge the help of the office:
Rosanne Poulson, M aree Roome, an d Claire Tugwell.
I w ou ld also like to thank M arion and A rth u r Fontaine for
connecting me to Dr. Paul Rom aniuk, for their infin ite kindness, and
for being like second parents to me.

XXI
Finally, I w ould like to thank A rthu r H am ilton, m y husb an d
and best friend , for his love, patience and support. W ithout him m y
life w ould never be complete.
Dedication
This w ork is dedicated to my mother, Lina Kletskova.

CHAPTER 1.0 EARLY GROWTH RESPONSE PROTEIN 1 (EGR-1); A
PROTOTYPICAL MEMBER OF A C2 H 2 ZINC FINGER FAMILY OF
TRANSCRIPTION FACTORS
1.1 Introduction
1.1.1 Overview o f the EGR-1 fam ily o f transcription factors
The proliferation and differentiation of eukaryotic cells is influenced by
a m u ltitu d e of stim uli, including grow th factors, adhesion m olecules and
o th er extracellular ligands. These complex long-term cellu lar responses are
m ed iated by changes in gene expression, an d are coupled to the initial signal
tran sd u c tio n even ts occurring a t the level of the plasm a m em brane. The
im m ediate-early genes are the earliest dow nstream nuclear ta rge ts for these
events. The activation of these genes is generally very rap id , transient, and
independen t of de novo protein synthesis. A subclass of these genes encodes
tran sc rip tio n factors, w hich fo rm the firs t step in in itia tio n o f genetic
p rogram s that w ould lead to an appropriate cellular response.
The best characterized m em bers of this g roup of im m ediate-early genes
in c lu d e c-jun, c-fos, and Egr-1. Each of these genes, in tu rn , represents a
p ro to ty p e for a fam ily of closely related proteins. N um erous stud ies have
dem onstra ted th a t EGR-1 induction is universal. Various s tim u li th a t induce
EGR-1 expression, as well as diverse cell types in w hich EGR-1 expression has
b een described, are sum m arized in Figure 1.1. E xtracellu lar stim uli th a t
induce EGR-1 can be grouped in to the follow ing categories: (1) m itogens; (2)
deve lopm en ta l o r d ifferentiation cues; (3) tissue or ra d ia tio n injury; (4)
signals that cause neuronal excitation. In practically every cell type examined,
EGR-1 expression is rapidly induced by such m itogens as ho rm ones, grow th
fac to rs, and the tum or prom oter TP A (phorbol) (Sukhatm e e t al., 1987;
Sukhatm e et al., 1988; Lau and N athans, 1987; Lemaire et al., 1988). Various

K cjp tfp jt
v iitocrn ic
S t i t n o l u s
POGF EGF rr.i lasulinpttyfohem aggluiininandjdenoune diphofptuic PDCF. »»sopcesiin tenimbFGF. PDCF BB p in ijl hcpafectomir GM-CSF. LPS endochelin angiotensin (I
Cell troc
ribroblascsntvobiasuhqsalocylcspenptieral blood lymphocytes B lymphocytes kidney epithelial cells kidney ittesjngial cells skeletal muscle Sol* cells skeletal muscle Sol* cells liverpentooeal macrophages astrocytesvascular smooth muscle cells
H ypertrophic
O ille ren tia tive
endothelin. angiotensin II
NGFretinoic acid. DSISO retinoacacid TP A. DMSO
myocyte
pheochrofflocytoma PCI 2 (neunll embryonal carcinoma PI9 embryonal calvarial cells. RCT-1 (osteoblast) myeloid leukemia HL60 and U 9)7
Tissue!radiation injury ischemia
ioniang tadialionkidney 29). SQ-20B
Neuronal ecciCatioo potassium ions metrazole NMDA visual stimuli efccnooonvTilstve shock therapy, dopattiine receptor activation, opiate withdrawalperipheral nervous system
PCI 2 (depolaricatioo) seizures in vivo hippocampus visual ctxtci CNS
sciativc nerve transecirao
Other MOCK. IXC-PKi leital epithelial celli
Figure 1.1 Biological processes m ediated b y EGR-1 in response to diverse
stim uli (Cashier & Sukhatme, 1995).

differentiative stim uli such as retinoic acid, DMSO (dim ethyl sulfoxide), and
pho rbo l give rise to EGR-1 expression, w hich can be co rre la ted w ith
differentiative processes of the kidney, spleen, brain, and m ost o ther tissues
(Sukhatme e t al., 1988; Lemaire e t al., 1988; Christy e t al., 1988). In the context
of tissue injury such as ischem ia or ionizing radiation, EGR-1 has been show n
to be strikingly induced (Bonventre e t al., 1991; H allahan e t al., 1991). It is
likely that EGR-1 is no t induced by the injury itself, bu t rather acts in response
to post-injury events, m ediating subsequent processes of cellular proliferation
a n d d ifferen tia tion . N u m ero u s lines of ev idence ex is t in d ica tin g an
im portan t role of EGR-1 in neuronal signaling. For exam ple, EGR-1 levels
increase dram atically in the brain following seizure activity (Sukhatm e e t al.,
1988). A d ram atic increase in EGR-1 levels is o b se rv ed fo llow ing
electroconvulsive shock therapy , dopam ine receptor activation, an d op iate
w ithdraw al (Bhat et al., 1992). Finally, the expression of EGR-1 in developing
and adu lt brain em phasizes the importance of EGR-1 in neurophysiological
processes (W atson and M ilbrandt, 1990). Thus, EGR-1 m ediates responses of
enorm ous com plexity. I t acts in different cellular contexts and is able to
respond to a m ultitude of extracellular signals.
All m em bers of the EGR family share highly sim ilar, C 2H 2 zinc finger
D N A -binding motifs. A t p resent, the closest m em bers of the EGR fam ily of
transcriptional regulators are: EGR-2/Krox20 (Chavrier e t al., 1988; Joseph et
al., 1988), EGR-3 (P a tw ardhan e t al., 1991), and EG R -4/N G FI-C /pA T133
(Patw ardhan et al., 1991; Crosby et al., 1991; M uller et al., 1991). All of the
above proteins have zinc finger dom ains that are virtually identical to th a t of
EGR-1 (Figure 1.2). The EGR-1 zinc-finger dom ain is over 95% identical to
tha t of EGR-2 (Joseph e t al., 1988) and 91% identical to th a t of EGR-3 a t the
am ino a d d level (Patw ardhan et al., 1991). The residues im portan t for specific

E f f l Ep 2 d g r i WTI Spl
- .K P 5 p K R r. r p N R P S K T P P H E R P r /kp r L R P R K r p N R P s K T P V H E R P r P
K p r R P R K r p H R P s K T P L H E R P H Ar. R f F K 1, S H L 0 K H s R K H T C E K P r 0T c p Y c K 0 s E c R C s C D P C K K K 0 H (
- - I S L« 2 1 . -Egr-1 CD c p V E S C o R R F S R S D E L T R K I R I H T C Q K P F OE ff-2 CD c p A E C C o R R F S R S 0 E L T R K I R I H T C H K P F OEgr-Î CD c p A E C C 0 R R F S R 5 0 E C, T R H E R I H T C H K P F OWTI C2) c 0 F K 0 C E R R F S R S 0 Q L K R H Q R R H T C V K P F QSpl CD c K C Q C C c K V r c £] T S K L R A H E R W H T C E R P F H
- . I S LS 21 •Egr-1 C2) c R t - - c K R N F S R 5 0 H L T T H I R T H T C E K P F AEgf-2 (2) c R c - - c K R N F S R S D K L T T H I R T K T C E K P F AEgr-3 (2) c R I - - c K R S F S R S 0 K L T T K I R T K T C E R P F AWTI O) c K T - - c Q R K F S R S 0 H L K T K T R T H T C Q R P F SSpl C2) c T W S V C C K R F T R S 0 E L Q R K R R T H T C E R R F AM IG l (1 ) c P I - - c K R A F H R L E K 0 T R K K R I H T C E R P H A
. . LS 1 1 2 1 * . • . . . .£gr-l C3) c - - o r e C R K F A K S O e R R R H T R I H E R Q K O K R A O R s V VEgf-2 (3) c - - o r e C R K F A R S 0 E R R R K T R I H E R Q R E R R S S A p S AEgr-3 (3) c - - E F c C R K F A R S D E R R R K A R I H E R Q R E R R A E R c c AWTI W c R w e s c Q K K F A R S 0 E t V R H H H K H Q R N K T K L Q L A L •Spl (3) c - - P E C p K R F K R S 0 K L S R H X R T H Q H R R C C P C V A L s VMIGl (2) c 0 F p c c V K R F S R S 0 E L T R K R R I H T M S K P R C R R C R R K
a - h d i x
Figure 1.2 Com parison of D N A -binding dom ains of the EGR-1 family of
proteins. 25nc fingers and adjacent sequences of the EGR-1 fam ily of proteins
are aligned. Conserved cysteine and histidine residues are m arked (•); the a~
helical region is underlined; conserved residues im portant fo r determ ining
b inding speciHdty are enclosed; basic residues are denoted (+) (Cashier &
Sukhatm e, 1995).

DNA recognition are conserved, and m ost of the changes are conservative
substitu tions (Figure 1.2). The hom ology betw een these proteins ex tends to
short stre tches of adjacent basic sequences bu t sharp ly drops o u ts id e this
region. This suggests tha t these proteins m ay recognize the same D N A target
sequences th ro u g h th e ir zinc fingers, b u t tha t in te rac tions w ith o ther
transcrip tional regulatory proteins are d istinct for each family m em ber. This
m ultigene fam ily offers a g reat system to study the re la tionsh ip betw een
signal tran sd u ctio n and gene expression in both norm al and tran sfo rm ed
cells.
The W ilm s' tum or gene p roduct has four zinc fingers, three o f w hich
possess re la te d bu t less hom ologous zinc finger D N A -binding dom ains
(approxim ately 65% identity to the EGR-1 zinc finger dom ain) (G essler et al.,
1990; Call e t al., 1990). The m am m alian ubiquitous transcrip tional activator
S p l has three related zinc fingers (K adonaga e t al., 1987). S p l finger 2 is m ost
similar to EGR-1 fingers 1 and 3 (Kadonaga e t al., 1987). Another d is tan t
m em ber of the EGR fam ily of proteins is a yeast p ro te in M IGl invo lved in
responses to glucose repression (N ehlin and Ronne, 1990). It con ta ins two
zinc fingers th a t are m ost sim ilar to th e second and th ird fingers o f EGR-1
protein, w ith 60% identical residues (N ehlin and Ronne, 1990). O u ts id e the
zinc fingers, MIG-1 has no obvious sim ilarity to other proteins. This absence
of sequence conservation outside the zinc finger motifs is a com m on finding
am ong C 2 H 2 zinc finger proteins. Therefore, a com parison of the fam ily
m em bers m u s t rely on a n alignm ent o f finger sequences. S tud ies o f the
im m ediate-early proteins a n d dow nstream prom oter elem ents th ey targe t
will con tinue to enhance o u r know ledge of protein-D N A in terac tions and
general m echanism s of transcriptional activation and repression.

1.1.2 G ene targets o f EGR-1 regulation
C onsisten t w ith its role in cellu lar p ro liferation a n d differentiation,
EGR-1 b in d s and reg u la te s genes invo lved in these functions. Some
discussion follows o n the best docum ented instances th a t involve regulation
by EGR-1.
Platelet-derived grow th factor A (PDGF-A) is a p o ten t mitogen w hich is
present a t elevated levels in response to grow th factors o r cytokines (W ang
and D euel, 1992). A n EGR-1 binding site has been defined in the prom oter
region o f the PDGF-A gene (Wang an d Deuel, 1992). The DNA fragm ent
bound by EGR-1 has the follow ing sequence: GAG-GAG-GAG-GAGGA.
A lthough th is site dev ia tes from the consensus EGR-1 b ind ing sequence,
w hich is G C G -G /TG G -G G G , it com petes equally w ell in the gel-sh ift
com petition experim ents. Similar m otifs have been iden tified in prom oters
of other g row th-related genes, such as the insulin receptor, the tum or grow th
factor P, the epiderm al grow th factor receptor, c-myc an d c-Ki-ras (W ang and
Deuel, 1992).
A no ther gene w hose expression is regulated by EGR-1 is thym idine
kinase (tk), an im portan t player in DNA biosynthesis. U sing m onoclonal anti
- EGR-1 antibodies, it w as shown that EGR-1 was one o f the com ponents of
the tk p ro m o te r com plex obtained from serum -stim ulated nuclear extract
(M olnar e t a l., 1994). F u rther tra n s ie n t tran s fe c tio n e x p erim en ts
dem onstrated that EGR-1 activates a reporter driven by a tk prom oter elem ent
(Molnar e t al., 1994).
EGR-1 has been show n to be required for differentiation of m yeloblasts
along the m acrophage lineage (N guen e t al., 1993). T he use o f EGR-1
an tisense o ligom ers in the cell c u ltu re m ed ium re su lte d in b locked
m acrophage d iffe ren tia tio n in norm al m yeloblasts as w ell as m yelo id

leukem ia cells. Thus, the authors clearly dem onstrated that expression of
EGR-1 is essential for m acrophage differentiation.
The m yosin heavy chain a gene (a-M HC) was show n to be expressed in
concordance w ith EGR-1 in stim ulated cultures of cardiac m yocytes (G upta et
al., 1991). Transient transfection assays using a CAT reporter contain ing a -
m yosin heavy chain p rom oter show ed 1 0 -fold induction of the prom oter
activity by an EGR-1 expression vector (Gupta et al., 1991). The reg ion of the
ra t a-M H C prom oter th a t is responsive to EGR-1 has been defined , and
show n to contain a potential EGR-1 b ind ing site (Gupta e t al., 1991). The a -
M HC induction appears to be tissue-specific since it w as observed in the
myogenic Sol8 cell line, b u t not in NIH3T3 fibroblasts (G upta et al., 1991).
EGR-1 is expressed a t high level in the rat adrenal g land an d in PC12
cells as dem onstrated by Elbert et al., (1994). A proposed function for EGR-1 in
adrenergic d ifferentiation m ay be via regulation of phenylethanolam ine N-
m ethy ltransferase (PNM T) (PNMT is an adrenal enzym e th a t converts
norep inephrine to ep inephrine) (E lbert e t al., 1994). The analysis o f the
PNMT prom oter revealed that it contains tw o potential EGR-1 b ind ing sites,
one of them differing only by one nucleotide from the EGR-1 consensus DNA
binding sequence. T ransient transfections show ed tha t EGR-1 can stim ulate a
PNMT reporter by fourfold (Elbert et al., 1994).
D espite the abundance of d a ta on EGR-1 induction by m itogenic
signals, additional EGR-1 gene targets aw ait elucidation. Finally, in vitro
studies of the EGR-1 involvem ent in cellular proliferation and differentiation
need to be correlated w ith the in vivo role of EGR-1.

1.13 Identification o f EGR-1 cDNA, and characterization of its protein p roduct
U sing d ifferen tia l screen ing techniques, several g ro u p s se t o u t to
iden tify novel genes which have a low level of expression in nond iv id ing
cells b u t which are rapidly up-regulated in cells stim ulated by m itogen. The
follow ing criteria w ere used to isolate such novel genes: (1 ) transcripts should
be rap id ly and transiently induced by serum stim ulation of quiescent cells; (2 )
these genes should be induced w ithou t intervening protein synthesis, i.e. the
induction should no t be affected by inhibitors of protein synthesis, such as
cyclohexim ide; (3) expression sh o u ld be induced by a w ide spectrum of
m itogens, such as g row th factors, horm ones, and o ther ligands; (4) expression
should be induced in a broad array of cell types; an d (5) the genes should be
highly conserved in evolution.
The novel im m ediate-early gene has been cloned by a num ber of
research groups. Sukhatm e e t al. (1987, 1988) identified a gene designated Egr-
1 . The authors used a differential screening technique to screen a library
from BALB/c 3T3 cells s tim u la te d w ith se ru m in the p resence of
cycloheximide. C lones which preferentially hybridized to cDNA from serum
and cyclohexim ide-treated fibroblasts were identified by com paring them to
cD N A from quiescent cells. A 3.4-kb transcrip t appeared u p o n m itogenic
stim ula tion of a varie ty of cell types, and was designated Egr-1. U sing a
s im ila r d ifferen tia l screening s tra tegy , M ilb rand t (1987) in d ep e n d en tly
iso la te d a tra n s c r ip t nam ed N G F I - A , w h ich was ac tiv a ted in r a t
pheochrom ocytom a PC12 cells by nerve grow th factor, and is a ra t analog of
the m ouse Egr-1. The same gene has been independently cloned by o ther
groups, using a sim ilar approach: zif268 was cloned from serum -stim ulated
3T3 fibroblasts from BALB/c m ice (Lau and N athans, 1987); tis8 w as
iden tified as a phorbol-inducible gene in 3T3 cells (Lim e t al., 1987); the

chicken hom olog , ce/5, was cloned as a v-src - inducib le gene from chicken
em bryo fibroblasts (Simmons et al., 1989); gene 225 w as identified as a T-cell-
activated transcrip t (W right et al., 1990), and Krox24 w as isolated from serum -
stim ulated 3T3 cells by hybridization to a highly conserved dom ain o f the
Drosophila factor K ruppel (Lemaire e t al., 1988).
Sequence analysis of EGR-1 revealed that the p ro te in contains three
tandem ly repeated zinc finger m otifs of C2 H 2 type th a t govern the DNA-
b ind ing function o f th is protein (Sukhatm e e t al., 1988; Lau an d N athans,
1987; Lem aire e t al., 1988; Christy e t al., 1988) (F igure 1.3A). The coding
sequence o f the p ro te in is highly conserved across species, ind ica ting the
im portance of the Egr-1 gene product. C om plem entary DNAs firom hum an
(Suggs e t al., 1990), ra t (M ilbrandt, 1987), m ouse (Sukhatm e e t al., 1988;
Lemaire e t al., 1988; C hristy et al., 1988), chicken (Sim m ons et al., 1989), and
zebrafish (D rum m ond e t al., 1994) are highly similar. The p roduct o f the Egr-
1 gene is a pro tein o f 80-82 kOa (Cao e t al., 1990). Cell firactionation studies
and im m unocy tochem istry dem onstra ted th a t EGR-1 is localized to the
nucleus, w hich is consistent with its pu tative D N A -binding function (Cao et
al., 1990; Day e t al., 1990). Further exam ination of the am ino acid sequence of
the EGR-1 p ro te in show ed it to contain basic residues clustered in the zinc
fingers and the adjacent sequence (Figure 1.3B). The am ino-term inal am ino
a d d s are rich in proline and serine-threonine residues, o rganized in stretches
of tree to five consecu tive am ino acids. There is o ne series o f seven
consecutive serine-th reonine residues, w hich is fo llow ed by seven g lydnes
(Figure 1.3B). The carboxyl term inus of the EGR-1 p ro te in is also rich in
proline and se rin e /th reo n in e residues. The regions o f the pro tein containing
proline am ino a d d s are predicted to lack a-helical secondary structure,
whereas the h igh con ten t of serine an d threonine residues indicates th a t

10
100
Egr-1 residues ZOO 300 400 SOO $33
1 1
pisn1------------ r
Sci/Thr-nch
T
K«C Al« Al« 4 ia Ly* K l^ Clu H«C Clci L«u K«C Sfi£ ^ro Leu
Cli* t i« s^f Kmp Pro Mm Cty Ser ptM fro « i* Sey Pro LLc M*c Amq AMt DgC Pro L^s
Leu Clu Clu Mec Not. Lou Lou S#r 4sn c ty Ato Pro Cln ft»« Leu Cty Ale Ala Cly Qlc
Pro Clu Cly Ser C ly C ly Asa Ser ter t# f tmf TTtf ter Cly Cly Cly c l y Cly C j
Cly S e e Asa ^or C ly t o f S ^ r Alo Ptte Asm Pro Clm Cty Clu Pro Scjc Clu Glm Pro_;Da2
Clu Hi* Leu Ttir Ttir c lu Ser Pbo te r Asp l i e Al» Leu Asm Asm Clu ty* Ale Net VmI
Clu TSr Kmr- ism- pyQ SoT C la Thr THr Afg Lou Pro Pro l ie Ttir Tvr tn r c l y Axg Ptie
Sec Leu Clu Pro A le Pro Asa Ser c ly Asm Ttir Lou Trp Pro Clu Pro Leu Pbe SeC Lw
v» l Sec c t y Lou Vol t o r Met Zhg Asa Pro Pro Thr t e r t e r te r Ser Ale Pro SfiC Pro
Al« At» t e r g e r t o r t e r to r Ala t e r Clm t e r Pro Pro Leu SfiC Cys AI» V»I Pro Sec19Asa Asp t e r t e r pro t l o Tyr t e r Al» Al» Pro f i ic Pro ZbC ^ro Asa %hC Asp t i e
Pho Pro Clu Pro O la ScC C la A la Pbe Pro C ly SfiC Ala Cly The. Ala Leu C la XXL. g o
Pro Pro Ala i v y Pro A la fh r Lys C ly C ly Pbo C la Pal Pro Met ( to Pro Asp Z3S tcu
Pbe Pro Cla C la C la C ly Asp Lou Scc Lou C ly Thr Pro Asp Cla Lus Pro A a Cla C ^
Lou Clu Asa Arg %bC C la C la Pro t f^ Lau tb r pro Lou t e r Thr t i e Ly* Ala Pbe Ala
Z2lC Cla Sec C ly t o r C la Asp Lou Lys Ala Low Asa Tbr Tbr Tvr C la t e r C la Lau
Lyt Pro t e r Arc Kac Arc Lys Tyg Pro Asa Arg Pro t e r L n XSiL Pro Prof C is Clu Argi
Pro Tyr Ala^ ÿ ^ Pro V al Clu t o r A s p Arg Arg Pbo te r Arg to r Asp Clu Lou
Argi l s ) H e Arg t l a ^ S ^ t b r C ly Cla Lys Pro Pbo C l a A r g t l o 0 y o ) wec Arg Asaj
Pbe te r Acg t e r Asp > £s Lau Tbr t h r ^ Z ^ t lo Arg Tbc(g £ i)Tbr C ly Clu Lys Pro g o
Ala y s ) Asp I l a 6 ^ ) c l y Arg Lys Pbo Ala Acg to r Asp c lu Arg Lys Arg ^ i ^ Tbr Lys
Lou Arc C la Lys Asp Lys Lys Ala As p Lys t e r V I v»l Ala te r Pro Ala ^ a
te r te r Lew t o r t a r Tyr Pro to r Pro Pal Ala Tbr t e r Tyr Pro te r Pro Ala tb r tbr
t e r Pbe Pro t o r Pro v a l Pro Tbr ter Tyr to r t e r Pro CLy ter tec Tbr Tyr Pro
Pro Ala His ta r C ly Pba Pro to r Pro to r Val Ala Tbr Tbr fbe Ala te r v » l pro Pro
Ala Pbe Pro Tbr C la Val ta r to r Pbo Pro to r Ala C ly Val ter tor tor Pbo te r TbrSN
te c Tbr Cly Lau t a r Asp Mac Tbr Ala Tbr Pbo to r Pro Arg Tbr l i e Clu t l o Cys$31
Figure 1.3 Schematic struc tu re and amino a d d sequence of EGR-1 protein.

11
F igu re 1.3 (A) A diagram of the functional dom ains o f EGR-1 protein: the
serine-threonine N -term inal dom ain is show n in the hatched box on the left;
b lack bars re p re se n t zinc finger m otifs; the C -term inal p ro line-serine-
th reonine dom ain is designated by a hatched box on the right. (B) Coding
sequence of m urine EGR-1. The three zinc finger motifs are enclosed in a box;
zinc-coordinating cysteine and histidine residues are circled; serine, threonine
a n d ty rosine re s id u es in th e N -term inal p o rtio n of th e sequence are
underlined (G ashler & Sukhatm e, 1995).

1 2
EGR-1 may be subject to phosphorylation. Indeed, alkaline phosphatase was
show n to convert slow m igrating form of EGR-1 to the faster m igrating form
as seen on SDS-PAGE (Cao et al., 1990; Day et al., 1990).
F u rther stru c tu re -fu n c tio n analysis of EGR-1 p ro te in defined its
ac tiva tion , rep re ss io n , DNA b in d in g and nuc lear lo ca liza tio n dom ains
(G ashler et al., 1993), sum m arized in a d iagram in F igure 1.4. Thus, the
organization of EGR-1 is m odular in nature, w hich is characteristic of many
classical transcription factors. A po ten t activational dom ain w as m apped to
the serine- a n d th reon ine-rich am ino term inal dom ain , u s in g d e le tio n
analysis of EGR-1 (Gashler e t al., 1993). These EGR-1 ac tiva tion sequences
w ere show n to be independent dom ains by p lacing them in the context of
o ther proteins. W hen residues 3-281, or 3-138, o r 138-281 w ere fused to the
D N A -binding do m ain of the yeast factor GAL4, they activated transcrip tion
100-fold as GAL4 fusions (Gashler e t al., 1993). These findings w ere confirm ed
by studying NGFI-A protein, the ra t homolog of EGR-1 (Russo e t al., 1993). A
w eak transactivation dom ain was m apped by several laboratories to the C-
term inus of EGR-1: am ino acids 420-533 (Russo e t al., 1993; G ash ler et al.,
1993).
A repression dom ain of EGR-1 was iden tified w hen a sm all in ternal
deletion 5' to the zinc-finger dom ain (amino acids 284-330) re su lted in alm ost
5-fold increased transactivation in HeLa cells (Gashler et al., 1993). This w as
an unexpected finding. Further experim ents have show n th a t th is repression
dom ain is also m odular in nature, since it could be fused to the D N A -binding
dom ain of GAL4, and represses transcription 7- to 10-fold (G ashler e t al., 1993).
T his se rin e / th reo n in e -rich d o m a in is h igh ly co n serv ed in e v o lu tio n
(D rum m ond e t al., 1994), and is d istinct from repression dom ains of other
transcriptional regulators, such as the alanine- and g lydne-rich repression

Il
sQ P 8 C . T 7 C . f f T l I I K X r A . T Q « a * Q
D L K & L 8 T T Y Q f f Q L CH S ) 3 tK p g R 8 R K T 7 8 R P 8 K T P
}fftr O Q K r r Q
IfftT o K K p r ^
4 1 7 *1»L K Q K D K K K O K f f V V X f f
ff ft ff O 8
ft ff O 8
ft ft ff O 8 C ft
I ft I
T CI
too
Egr*I amino acids
200 300 S M S3) 1 1
PWT"T r
Scr/Thr-richrZTE
A c t ir a d o a
Repression
DNA blading
N u d esr localization
s tro n g
m
U l 314
331 413
313 33# 331 413
Figure 1.4 Sum m ary of EGR-1 functional dom ains.
(A) Amino a d d sequence o f EGR-1 repression dom ain an d zinc fingers. (B)
Schematic m ap o f EGR-1 dom ains (Gashler & Sukhatme, 1995).

14
dom ain in K ruppel (Licht e t al., 1990), hydrophobic- an d proline-rich Even-
sk ip p e d rep resso r (H an a n d M anley, 1993), or p ro lin e - and glycine-rich
rep resso r o f W TI (M adden e t al., 1993). The fact that 7 o u t of 24 am ino a d d
residues of the EGR-1 repression dom ain are serine o r threonine suggests that
the rep ression function m ay be regulated by phosphorylation.
C onsisten t w ith its transcrip tional regulatory function, EGR-1 w as
show n to localize to the n u d e u s (Cao e t al., 1990; Day e t al., 1990). Generally,
n u d e a r localization signals are short stretches of 8 -1 0 am ino adds rich in basic
an d p ro line residues (Silver e t al., 1984). In EGR-1, the only regions rich in
basic residues are the three zinc finger region and the adjacent stretch of basic
am ino a d d s , suggesting th a t the n u d e a r localization signal resides in those
sequences. U sing a series o f deletion m utants of EGR-1 along w ith subcellular
fractionation and W estern b lo t analysis, am ino adds 315 to 429 were show n to
be im portan t for proper targeting to the nudeus (Day e t al., 1990). H ow ever,
the zinc finger region a lone (amino a d d s 331 to 419) w as not sufficient for
n u d e a r localization of the protein. The basic sequence 5' to the zinc finger
dom ain (am ino a d d s 315 to 330) was also required for nuclear targeting (Day
e t al., 1990). Thus, EGR-1 has a b ipartate nuclear localization signal, the
largest portion of which coinddes w ith the DNA-binding domain.
1.1.4 D N A -b ind ing function of EGR-1
The DNA sequence to which EGR-1 binds was initially identified based
on th e assu m p tio n th a t EGR-1 regulates its ow n expression a n d m ight,
therefore, b ind to the 5' upstream flanking sequence of the Egr-1 gene (Christy
an d N a th an s , 1989). U sin g bacterially expressed EGR-1 protein , sp ed fic
b in d in g to one prom oter fragm ent was observed in gel mobility sh ift assays
(C hristy and N athans, 1989). This b inding site was located w ithin 650 base

15
pa irs 5’ to the transcrip tion s ta rt site of Egr-1. D N ase-I fo o tp r in tin g
experim ents identified the sites of contact in the Egr-1 prom oter b ind ing
sequence, as well as a num ber of sites 5' to other genes (Christy and N athans,
1989). These sequences, taken together, were u sed to establish a consensus
high-affinity binding site sequence GCG-G/TGG-GCG. Further m éthylation
interference experim ents indicated tha t EGR-1 m akes extensive contacts along
this sequence (Christy and Nathans, 1989).
EGR-1 is the first C2H 2 zinc finger protein for w hich a high resolution
structu re w as obtained. The structure of the complex consisting of the cloned
three zinc finger pep tide and the 12-bp DNA con tain ing the specific 9-bp
sequence has been so lved at 2.1-Â resolution (Pavletich and Pabo, 1991)
(Figure 1.5). The structural inform ation obtained in this study served as a
topological blueprint for other m em bers of the zinc finger family. This EGR-1
- DN A complex was recently refined a t 1.6 Â (Elrod-Erickson et al., 1996). The
new structure confirms all the basic features of the 2 .1 Â model, and reveals
additional critical details of the complex.
The crystal struc tu res show th a t the three fingers w rap a ro u n d the
double helix, describing a C shape (Figure 1.6). The overall alignm ent of the
fingers on the DNA is antiparallel (finger 1 binds near the 3' end, and finger 3
b inds near the 5’ end of the 5 -GCGTGGGCG-3' consensus binding site). [The
m ode of TFIUA - DNA interaction is also an tipara lle l, w ith m ost of the
contacts involving the guanine-rich strand of the DNA (Smith e t al., 1984;
V rana e t al., 1988)]. The a-helices o f the zinc fingers are tipped a t ca.45°
relative to the plane of the base pairs; therefore, they are only approxim ately
a ligned w ith the m ajor groove. The N-term inal e n d of each a-helix m akes
specific contacts w ith the base pairs, each helix in teracting prim arily w ith the
3-bp subset of the 9-bp consensus binding site.

16
- 1 1 2 3 6M E R P Y A C P V E S C D R R F S R IS D E L T R H I RI HTIg Q K 1 5 10 15 20 25 30
P F Q C R I - - C M R N F S R Is D H L T T H I R T H t I G E K35 40 45 50 55 60
P F A C D I - - C G R K F A R IS D E R K R H T K I H L k o K D65 70 75 80 85
P-sheets a-helix
B1 2 3 4 5 6 7 8 9 10 11
A G C G T G G G C G T C G C A C C C G C A T
Figure 1.5 Sequence of the EGR-1 zinc finger peptide and of the DNA binding
site used in cocrystallization. (A) Sequences of the three zinc fingers are
aligned by conserved residues an d secondary structure elements, a-helices
are enclosed in boxes, and P-sheets are indicated by arrows. The conserved
cysteine and histidine residues are highlighted in bold. (B) Sequence of the
duplex oligonucleotide used in cocrystallization (Pavletich & Pabo, 1991).

17
F in g er J COOH
Hnger 1
Figure L 6 The overall arrangem ent of the three zinc fingers o f EGR-1 in the
m ajor groove of DN A (Pavletich & Pabo, 1991).

18
The N -term inal s trand of each P-sheet makes no contact w ith DNA,
w hile the m ore C -term inal s trand contacts the sugar-phosphate backbone
a long one DNA strand . The am ino acid side chains directly involved in
contacts w ith the DNA bases lie at a-helical positions -1, 2 ,3 , and 6 (num bered
relative to the first residue in each a-helix). Base specific contacts identified
in the original crystal structure are prim arily hydrogen bonds w ith the G-rich
strand of the DNA (Figure 1.7). The higher resolution structure provides a
m ore detailed view of the EGR-l-DNA interface w ith m ore direct and water-
m ed ia ted contacts. The sum m ary of direct base and phosphate contacts is
sh o w n in Figure 1.8. W ater-m ediated and van der W aals contacts to bases
and phosphates are sum m arized in Figure 1.9.
M any features of this complex were correctly predicted by sequence
ana ly ses an d m u ta tio n a l s tud ies (N ardelli et al., 1991). S ite-d irected
m utagenesis experim ents w ith DN A-binding dom ains of Krox-20 and Spl
p ro teins substitu ted the nonhom ologous amino acids in the recognition a -
helices and resulted in interconversion of the specificity of the m u tan t finger
(N ardelli e t al., 1991). Thus, even prior to the solution of the EGR-1 crystal
structure, some of the base-contacting positions had already been identified.
Fingers 1 and 3 have identical residues at positions -1, 2, 3 and 6 of the
a-helix: R, D, E, R, w hereas finger 2 has R, D, H, and T at the corresponding
positions. Fingers 1 and 3 m ake two prim ary contacts to the guanines of the
GCG subsite using the guanidinium group of the arginines to hydrogen bond
w ith the N 7 and 0 6 of the guanines in each subsite (Figure l.lO A). The
in teraction of arginine w ith guanine was predicted to p lay an im portan t role
in sequence-spec ific reco g n itio n based on its u n iq u e stereochem ical
com plem entarity (Seeman e t al., 1976). Indeed, these contacts are the m ost
p rom inen t ones in the EGR-1 - DNA complex. These arginines also interact

19
Ffnaer 3
Arg 80
Finger 2
His 49
Arg 46Finger 1
Arg 24
Figure 1.7 Sketch sum m arizing the principal amino add-base contacts as seen
in the original EGR-1 X-ray structure (Pavletich & Pabo, 1991).

20
Finger 3
( ,61 Arg 8 0
His 53
|( ,2 ) Asp 76 N ^ r g T i
A n g er 2
Arg 42 -— ■His 2 5 .
S a r4 5
(+3) His 49
|( * 2 ) A s p ^ - ( - l )A f g
R n g e r l
(♦6) Arg 24
) (|(» 2 )A sp 2 0 -( -l)A /g 1g
(Finger 3)
Figure 1.8 Sum m ary o f direct base a n d phosphate contacts in EGR-l-DNA
complex. The num bers in paren thesis in front of am ino acids a re their
positions w ith in a-helices. Boxes ind icate coupled res idue pairs. A rrow s
indicate hydrogen bonds; do tted a rro w s represent b o n d s w ith m arg inal
geom etry (derived from Elrod-Erickson e t al., 1996).
21
H n g cr 3
f *6) Arg 80 — --
(Linker 2 6 3) Lys 61
( .3 ) Glu 7f
Finger 3
(-2 )S er 17
(»2) Asp 76
(-1) Arg 74
(Linker I & 2) Lys 33 (♦I) Ser 75
M l Arg 78
Finger 2 R oger 2
(♦€) T hr52
( ♦ » « i s 4 9 -—
(♦^2) Asp 48
(-1) Arg 46 M l Ser 47
(r-2) Asp 48
R o g e r 1 R oger t
48) Arg 24
(*3) Glu 21
(♦2) Asp 20
(-1) Arg 18
c(♦S) Thr 23
(4 l)S e r 19
Figure 1.9 Sum m ary o f w ater-m ediated an d vsm der Waals contacts to bases
and phosphates in EGR-1 - DNA complex. The num bers in parenthesis in
front of am ino a d d s a re their positions w ithin a -h e lices . Boxes indicate
coupled residue pairs. A rrow s indicate w ater-m ediated contacts. Small
num bers show n over the arrow s indicate the num ber of w ater-m ediated
bonds. Dotted arrow s represen t van der W aals contacts.

22Basa pair 10 b r FinQor i
( Basa pair T for Finger 2 .
Base pair 4 b r Finger 3 )
020 in Finger I (048 InFingefZ:
RI8 In Fingar 1 076 in Finger 3 )(R46 InFIngerZ:
R74 In F in g ers)
B Besepeir'Sbr Finger I
( Base pair2 b r Bnger 3 )
n H
8eeepelr6
N.—
•H----
/------
(R80 lnRna«r3)
Figure 1.10 D raw ings of am ino add-base pair contacts o f the EGR-1-DNA
complex: (A) D rawing of the Asp-Arg-guanine interaction that occurs in all
three zinc fingers; (B) D raw ing of the Arg-guanine interaction that is present
in fingers 1 and 3; (C) D raw ing of the His-guanine interaction seen in finger 2
(Pavletich & Pabo, 1991).

23
w ith an aspartic ad d , w hich is the second residue w ith in each a-helix . The
carboxylate oxygens of the aspartic a d d make hydrogen bond-salt b ridges with
the Ne and N t] of the guanid in ium group of the arginines th a t a re already
bound to the first G of each subsite, thus stabilizing the interaction of the long
arginine side chain w ith the base (Figure l.lOB). Finger two uses a n arginine
and a histidine to contact the guanines of the m iddle base pair subsite TGG. A
histidine-guanine interaction that occurs in finger 2 is show n in Figure I.IOC.
The new 1.6 Â structu re helps explain the roles of the aspartic a d d
residues at position 2 of the recognition helices. The crystal struc tu re shows
that the A rg -1 /A sp2 residue pairs make w ater-m ediated contacts w ith the
cytosine w hich is base pa ired to the guanine contacted by Arg -1 (as well as
w ith the phosphate on the 5' side of this guanine). These contacts are seen in
all three zinc fingers. In fingers one and three, these aspartates also make
w ater-m ediated contacts w ith the neighboring base 5' of the critical guanine.
For instance. Asp +2 in finger 1 makes a w ater-m ediated contact w ith cytosine
9, and an analogous contact is made by Asp +2 of finger 3 to the N4 o f cytosine
3 (Figure 1.9). The crystal structure shows tha t Asp +2 of finger 2 clearly
contacts cytosine 8 ', and tentatively suggests that Asp +2 residues firom fingers
1 and 3 form s weak interactions w ith a base positioned outside the canonical
triplet on the secondary, C-rich strand of the DNA (Figure 1.8). In the original
X-ray structure one of the oxygens of the aspartic a d d w as show n to be w ithin
hydrogen bonding distance to a neighboring base on the parallel, C-rich strand
of the DNA. However, since the H -bonding geom etry was no t ideal, these
contacts w ere presum ed un im portan t for recognition (Pavletich an d Pabo,
1991). M ore favorable geom etries w ere observed in the refined structure
(Elrod-Erickson e t al., 1996). For example, w hen A rg -1 of finger 3 contacts G4 ,
this in teraction is stabilized by Asp +2 of the sam e finger, w h ich in tu rn

24
contacts As* (the com plem entary base to T5 on the opposite strand ) (Figure
1.8). Such contact was clearly dem onstrated for finger 2 (A sp48 in teracting
w ith Cg ). The co rrespond ing interactions for fingers 1 a n d 3 had less
favorable stereochem istry.
There are several p ieces of evidence suggesting th a t th is con tact
con tribu tes to recognition in o ther zinc finger p ro te ins. In the cocrystal
s truc tu re of the Drosophila regulatory pro tein Tram track com plexed w ith its
D N A the hom ologous aspartic acid in the second zinc finger w as show n to
m ake direct hydrogen bonds w ith the secondary DNA s tra n d (Fairall e t al.,
1993). Replacem ent of the aspartic acid by alanine in the second zinc finger of
the S. cerevisiae ADRl p ro te in resulted in significant re d u c tio n o f p ro te in
b ind ing to its cognate site (Thukral e t al., 1991; Thukral et al., 1992).
Several biochemical stud ies also propose the in teraction of A sp +2 w ith
the parallel s tran d of the DNA. Zinc finger phage d isp lay selection studies
p rov ided original support for this interaction. The technique o f phage display
m akes use o f the expression of zinc finger regions th a t carry partia lly
ran d o m ized recognition helices as part of a phage coat p ro te in . By th en
allow ing the phage carrying zinc finger peptides on its su rface to equilibrate
w ith a target DNA sequence, i t is possible to isolate and am plify the specific
zinc finger proteins that recognize desired DNA target sites. In the study by
C hoo and K lug (1994a) the sequence of the m iddle f in g er of EGR-1 w as
random ized to test new b ind ing specificities. Strikingly, in a lm ost all of the
selected fingers in w hich arg in ine interacts w ith the 3' gu an in e , aspartic acid
w as selected a t position +2. W hen position -1 w as not arg in ine, aspartic a d d
a t position +2 w as practically never selected. This suggests th a t the aspartic
a d d residue + 2 plays an im portan t role in contributing to the sp e d f id ty of the
p ro te in -D N A com plex. T his resu lt also em phasizes th e im portance o f

25
a rg in in e in conferring D N A -binding ability. In ou r study o f D enys-D rash
W TI p ro te in m u tan ts the substitu tion of a rg in ine for a try p to p h a n w as
de trim en ta l to DNA-binding activity (Borel e t al-, 1996). In a com plem entary
s tu d y by Choo and Klug (1994b), a bias toward G or T at the 3 rd position of the
recognition subsites w as dem onstrated, and it w as concluded th a t the am ino
a d d a t position + 2 w as able to m odulate the sp e d fid ty of the am ino a d d at
o th e r positions. Thus, it was proposed th a t A sp +2 m akes a cross-strand
contact. Sw irnoff and M ilbrandt (1995) carried o u t b inding s ite selections to
derive the high-affinity consensus binding site for EGR-1 fam ily o f proteins.
T h e ir find ings su p p o rt the im portance of a sp artic ac id fo r sequence
recogn ition by EGR-1 protein fam ily m em bers. The au th o rs no ted a very
s tro n g selection for thym ine at position 5 and less frequently for guanine,
w h ile ad en in e or cytosine w ere never se lec ted . Both A an d C, the
com plem entary bases to T or G a t position 5 have hydrogen bond donor
g ro u p s (N 6 and N4) that can interact w ith the carboxylate oxygen of Asp +2.
T hese resu lts su p p o rt the idea th a t the h y d ro g en bond p o ten tia l on the
secondary DNA stran d contributes to recognition. A nother im plication of
these findings is tha t a single zinc finger can actually specify no t 3 bp, as
o rig inally proposed, bu t 4 bp DNA sites, and these binding sites overlap by
one base pair. Recently published w ork by Isalan, e t al. (1997) dem onstrates
that EGR-1-hke zinc fingers can, in fact, specify overlapping, 4-bp subsites. In
this s tu d y , the th ird zinc finger of EGR-1 protein w as m odified to rem ove the
fin g er 's po ten tia l for DNA interactions. N orm ally , the th ird zinc finger
in teracts w ith the first triplet of the EGR-1 consensus sequence. How ever, as a
resu lt o f finger three alterations, changes were observed in th e 5’ position of
the m id d le trip let of the DNA b ind ing site. T hus, deleting th e contact from
finger 3 affects the specificity for the 5' base of the binding site of finger 2.

26
Further selection an d gel m obility shift experim ents po in ted to the sam e
conclusion (Isalan e t al., 1997). Thus, the potential of zinc fingers to specify
overlapping, 4-bp subsites was dem onstrated, and the role of aspartic acid at
helical position 2 w as em phasized. The specificity of each recognition subsite
is, therefore, m ediated by tw o adjacent fingers th a t are synergistic in their
m ode o f binding. This has led the authors to redefine the b ind ing subsites for
each zinc finger, and the new schem atic d iagram of recognition is show n in
Figure 1.11.
The roles of the third a-helical residues are also refined in the 1 .6 Â
structure (Elrod-Erickson et al., 1996). The role of His49 (the th ird residue in
finger 2 a-helix) w as previously discussed in the original EGR-1 crystal
structure (Pavletich and Pabo, 1991). This residue w as previously show n to
make a direct hydrogen bond to N 7 of guanine 6 . In the new structure, this
histidine m ay be contacting 0 6 of the guanine instead. In addition , His49 also
m akes a series of van der W aals interactions w ith the m ethy l g roup of
thym ine 5. The significance o f the h istid ine-thym ine in te rac tio n w as
dem onstrated by Sw irnoff and M ilbrandt (1995) in their b inding site selection
experim ents. Thus, histidine contributes to the recognition of tw o bases on
the G -rich DNA strand.
G lutam ic acid residues w hich occur a t a -h e lica l p o sitio n three of
fingers 1 and 3 were suggested to contribute to specificity because biochemical
site selection and b ind ing studies all show a preference for cytosine at the
center o f triplets recognized by fingers one an d th ree (Christy an d N athans,
1989; Jam ieson e t al., 1994; Swirnoff & M ilbrandt, 1995). As revealed in the
refined crystal structure, the carboxylate groups of Glu21 and Glu77 do not
make any base-specific contacts. However, the side chains of these glutam ates
do hydrogen bond to the backbone amides of the arginines im m ediately

27
Figure 1 .1 1 Schematic diagram of EGR-I zinc fingers in teracting with the
proposed overlapping, 4-bp DNA subsites (Isalan e t al., 1997).

28
p reced ing the a helix (ArglS in finger 1, a n d Arg74 in finger 3). Glu21 can
also hydrogen b o n d to the backbone am ide of S erl7 , w hile G lu77 m akes a
co rrespond ing contact to Ala73. In ad d ition , these glutam ic acid residues
m ake van der W aals contacts to the edge o f the co rrespond ing cytosines.
These contributions, albeit m odest, may p lay a role in favoring cytosines over
o ther bases. A nother good observation w as m ade by N ardelli e t al. (1992) and
Sw im off & M ilbrandt (1995): the reason for cytosines and not guanines in the
m id d le of trip le t 1 and 3 m ig h t be an electrostatic repu lsion betw een the
carboxylate oxygens of the glutam ates and the 0 6 and N7 g roups of guanines.
C y tosines do n o t have e ither of these g ro u p s, an d , therefo re , are better
to lerated at these positions.
A num ber of w ater-m ediated contacts w ere detected a t the protein-
D N A interface th a t were not seen in the o rig inal X-ray structu re . They are
su m m arized in F igure 1.9. A set of contacts w ith the su g ar-p h o sp h a te
backbone of the DNA is also detected in the X-ray structure, an d m ost of these
are m ade to the G -rich strand of the consensus site (Figure 1.8). Each finger
u ses an arginine in the second P-strand a n d the first z in c -co o rd in a tin g
h is tid in e to contact phosphodiester oxygens. These contacts include one
u n u su a l a rrangem en t in w h ich the im idazo le r in g of the f irs t h istid ine,
in v o lv e d in z in c c o o rd in a tio n th ro u g h N £ , m akes co n tac ts to the
p hosphod ieste r oxygens th ro u g h Ny (Figure 1.12). Since zinc coordinating
h istid ines are conserved in all zinc fingers, it is possible that they all use these
types of contacts. The 1.6 Â structure revealed additional, n u m erous w ater-
m ed ia ted phosphate contacts. They are sum m arized in F igure 1.9. Serine
res id u es that occur a t position 1 of the a helices all p ro v id e phosphate
contacts: Ser75 (in finger 3) m akes a h yd rogen bond to p h ospha te 7', w hile
Ser47 (in finger 2) m akes a w ater-m ediated contact w ith phosphate 9'.

29
DNAH
Figure 1 .1 2 Schematic rep resen tation of hydrogen bonding betw een Zn-
coordinated h istid ine and DNA backbone (O 'Halloran, 1993).

30
Sim ilarly, Serl9 (in finger 1) m akes a w ater-m ediated contact w ith 0 5 ’ of
thym ine in position 1 2 ’. Other residues also contribute to the netw ork of
w ater-m ediated phosphate contacts, nam ely Serl7 m akes a w ater-m ediated
contact to the 5’ phosphate of base 9; Arg78 (the fou rth a-helical residue in
finger 3) m akes a w ater-m ediated contact w ith phosphate 7’; a n d Thr23
(residue 5 in the helix of finger 1) contacts the 0 5 ’ of base 12’. In addition, the
conserved lysines th a t occur in the linker region betw een the zinc fingers also
make several w ater-m ediated contacts: Lys33 (located betw een fingers 1 and 2)
makes a pa ir of water-m ediated contacts to the 5’ phosphate of base 5; Lys61
(located betw een fingers 2 and 3) m akes an analogous w ater-m ediated contact
w ith the 5’ phosphate of base 2. A n analogous phosphate contact involving
lysine residue w as described for D N A -binding peptide of I'b lilA (Chop and
Klug, 1993). M utation of this residue in TFIIIA reduces its affinity for DNA by
about sevenfold.
The im portan t role of the linker sequences w as pointed o u t in the
study by W ilson e t al. (1992). The authors used a reversion-based analysis of
D N A -binding dom ains: they constructed a chimeric p ro tein containing the
D N A -binding dom ain of EGR-1 inserted betw een the LexA D N A -binding
dom ain and the GAL4 transcriptional activating dom ain (LAG). This dim eric
protein w hen expressed in yeast resulted in retardation o f their grow th, while
a dim eric pro tein containing only the LexA and GAL4 dom ains acted as an
activator. Some yeast cells harbouring LAG eventually reverted to w ild-type
g row th by inac tiva tion of LAG. Those rev ertan ts w ere analy sed for
m utations in LAG, w hich all m apped to the EGR-1 zinc finger D N A -binding
dom ain. The m ajority of the m utations w ere found to comprise the DNA-
contacting am ino a d d s identified in the crystal structure, while several of the
m utations lay in the linker regions o f the zinc fingers. The m utation of the

31
two lysines in the linker regions w as disruptive, a n d the au thors proposed
that the rem oval o f the positive charge im p a rted by the ly s in es may
destabilize the tertiary structure of the protein. Secondly, it was no ted that the
two linker regions w ere no t equally sensitive to m utation : the second linker
was m u ta ted seventeen tim es w hereas the first w as a ltered only th ree times,
suggesting that the linkers may n o t p lay identical ro les in o rienting the zinc
fingers.
Finally, the role o f D N A conform ation was ad d ressed in the recent X-
ray s tu d y (E lrod-E rickson et a l., 1996). It is w e ll know n th a t D N A
conform ation either before or subsequent to protein b ind ing is an im portan t
determ inant of sequence-specific recognition (Steitz, 1993). Thus, stretches of
A /T residues in the b ind ing sites of the zinc finger p ro te ins T ram track and
M IGl are required for recognition (Fairall et al., 1993; L undin e t al., 1994).
While there was no bend ing detected in the crystal struc tu res of EGR-1-D N A
complex, circular d ichroism studies show a striking difference occurring in
the EGR-1 binding site depending o n the presence o r absence of the protein
(E lrod-Erickson e t al., 1996). T his suggests th a t EGR-1 D N A changes
conform ation upon com plex form ation . The D N A also adop ts a novel
conform ation, both w ide and deep, tha t has been described as B e n ia r g e d g r o o v e
-D N A . The binding sites in Tram track and GLI have been show n to ad o p t the
same conform ation (Fairall e t al., 1993; N ekludova & Pabo, 1994). The
significance of this Beniarged groove"DNA conform ation w as not clear un til the
m odeling studies show ed that the observed in te rfinger contacts a n d the
linker leng th w ould n o t allow bind ing to canonical B -D N A (Elrod-Erickson et
al., 1996).
T he original crystal struc tu re of EGR-1 com plex (Pavletich & Pabo,
1991) gave an initial v iew of zinc finger-DN A interactions. The new , refined

32
EGR-1 - DNA stru c tu re (Elrod-Erickson et ai., 1996) revealed considerable
add itiona l com plexity, and changed the w idely he ld percep tion tha t EGR-1
uses a simple recognition schem e in its interaction w ith DNA.
1.1.5 Structures o f o th e r zinc finger-D N A complexes
The elucidation of high resolution structures of less closely related zinc
finger proteins - the hum an oncogene GLI (Pavletich & Pabo, 1993) and the
D rosophila reg u la to ry p ro tein T ram track (Fairall et al., 1993) p ro v id e a
b roader perspective o n the issue of specificity of zinc finger-DNA interactions.
The hum an oncogene protein GLI contains five zinc fingers, a n d has been
cocrystallized w ith the sequence of DNA d eriv ed from in vitro genom ic
selection studies (since the physiologically relevant GLI-binding sites are no t
ye t know n) (Figure 1.13). The com parison of GLI-DNA com plex w ith the
o r ig in a l EGR-1-DNA s tru c tu re h ad revea led m ore d iffe ren ces th a n
sim ilarities. H ow ever, w ith the elucidation of the refined, 1.6Â structu re of
the EGR-l-DNA com plex, m ore sim ilarities be tw een the tw o pro tein-D N A
com plexes became apparent. The overall structure of the GLI-DNA com plex
show s tha t fingers 2 to 5 are arranged similar to the EGR-1 fingers, w rapp ing
a ro u n d the DNA w ith the N -term inal portion of their a -helices fitting in to
the D N A major groove. The difference betw een the two com plexes is that,
surprisingly, finger 1 of GLI does n o t make any contacts w ith the DNA a t all
(F igure 1.14). F ingers 2 and 3 b ind a section o f the DNA largely non-
specifically, contacting the DNA backbone, w ith finger 2 p rov id ing the only
base contact in th is region. O n the other hand , fingers 4 an d 5 do b ind a
conserved 9-bp reg ion of the site in a sequence-specific m anner. These fingers
a re m ost im portan t for recognition. In EGR-1, all fingers m ake generally
sim ilar contributions to sequence-specific recognition. A lignm ent of the zinc

33
s ' ' . . i \ I* <rî I .
V f E r O C R W D G C S O E F Ü S O E O L V H H I N S E H I @ G E R k
K J9 M SI S / 50 6* S8
E F V C H W G G C S 0 6 lR P F @ A Q L V V @ M R R H F G E K
59 r2 n ai ar ao 9< 98
P H K C r F E G C R K S ® S @ L E N L 0 F H L R S H F G E K
99 102 10/ III 1 1 / 120 I2S 129
P 0 M C E H E G C S K A F S n | a | s j o 0 a [ k 0 Q N R © H S N E ®
IM 11 ] I j a 142 148_______ i s i ISS ISO
P Y V C K L P G C T K R ® ( 5 f D ] p | s J s | L | g ( K | H V K T V H G P O A
W V W V I_________________________ 1
B' s 10 I s 20
A C G T G G A C C A C C C A A G A C G A A G C A C C T G G T G G G T T C T G C T T T
Figure 1.13 Sequences of the GLI zinc finger domain and the DNA-binding site used for cocrystallization. (A) The five zinc fingers of GLI are aligned to show the conserved am ino a d d residues and secondary structure elements. The position of a-helices is underlined, an d ^-sheets are indicated by zig-zag lines. O pen boxes highlight residues that make base contacts in the crystal structure , and open circles indicate residues that make phosphate contacts. Sym bols below the GLI sequence indicate the corresponding positions o f side chain-base contacts (filled boxes) and side chain-phosphate contacts (filled d rd e s ) that were observed in EGR-1 complex. (B) DNA duplex used in cocrystallization (Pavletich and Pabo, 1993).

34
Finaec
Fmaer
QaasL
Saser
y FinaecJ
Figure 1.14 Soetch sum m arizing base and phosphate contacts made by the GLI peptide. Solid arrow s indicate base contacts and dotted arrows indicate phosphate contacts (Pavletich & Pabo, 1991).

35
finger sequences of GLI and EGR-1 show ed a clear co rre la tion betw een the
positions o f the base contacting residues in the two p ro teins. H ow ever, the
m ode of dock ing zinc fingers against D N A show s d ifferences, w hich are
reflected in the d ifferent p a tte rn of am ino acid-base in te rac tions a t the
p ro te in -D N A interface. D espite the fact th a t GLI a n d EGR-1 belong to
d ifferent subfam ilies, w ith very little am ino a d d sim ilarity in the zinc finger
region, a n d d ifferen t DNA sequences to w hich they b in d , the tw o pro teins
have the fo llow ing features of the protein-D N A com plex in com m on: (1)
each pro tein contacts prim arily four bases in each subsite, an d these subsites
can overlap; (2) four amino a d d s in the a helix are usually involved in these
contacts; (3) b o th proteins contact bases on bo th DNA stran d s, except that the
m ajority of EGR-1 contacts involve prim ary, G-rich strand o f the DNA, w hile
GLI contacts are more evenly distributed betw een the tw o strands; (4) in bo th
complexes the DNA assumes a conform ation that is in term ediate betw een A-
and B-form; (5) the DNA-contacting residues of EGR-1 fall a t positions -1, 2, 3,
a n d 6 of th e a -h e lix , GLI a lso uses th e D N A -co n tac tin g re s id u e s
co rresp o n d in g to positions -1, 2, 3, and 6. In GLI, h o w ev er, add itional
residues of the a-helix make spedfic contacts, in d u d in g positions 1 and 5.
W hile GLI fingers m ake some use of the p rim ary contacts identified in EGR-1,
several of these interactions are unique in term s of the subsite position of the
base contacted, the strand contacted, and the nature of the sidechain-base
in teraction .
The s truc tu re of a tw o zinc finger p ro te in T ram track com plexed w ith
its recognition DNA sequence shows m any sim ilarities w ith the EGR-l-DNA
complex. The contacts betw een Tram track and its DNA are sum m arized in
Figure 1.15. A com parison o f base-specific contacts fo rm ed by EGR-1 and
Tram track pro teins is diagram m ed in Figure 1.16. The follow ing features of

36
; 10 i 5s - ;
C T A A T A A G G A T A A C C r C C C A T T A T T C C T A T r G C \ C G C r
r - • • ;35 30 25
F igure 1.15 The sum m ary of Tramtrack-DNA contacts (Fairall e t ai., 1993).
The sequence of the DNA used for crystallization is shown in the top right of
the figure. The details of the protein-DNA contacts are discussed in the text.

37
l-:CK-l F-l & F3ECR-I F2
*2
TTKF2TTKFl
T
Figure 1.16 Schematic sum m ary of the principal protein-D N A contacts
observed in the cocrystal structures of the EGR-1 and Tramtrack. Residues are
num bered relative to their position in the a helix. A rrow s rep resen t
hydrogen bonds observed in the respective crystal structure (Choo & Klug,
1997).

38
the Tram track-DNA structure are similar to those of EGR-1: (1) bo th fingers of
T ram track use the N -term inus of their a-helices to contact specific bases in
the DNA m ajor groove; (2) the majority of contacts are m ade to one DNA
strand ; (3) the b ind ing sites for the adjoining fingers overlap; (4) a -h e l ic a l
positions -I , 2 ,3 , and 6 are used to make base-specific contacts, w hich coincide
w ith the sam e residue positions in EGR-1 complex; (5) finger 2 m akes very
sim ilar base and phosphate contacts to those of EGR-1 fingers. F inger 2 in
bo th proteins uses an arginine a t position -1 for contacting guanine. The use
of position 2 in finger 2 observed in the Tram track cocrystal s tru c tu re is
equ iva len t to the contact seen in finger 2 of EGR-1. In both p ro te in s, an
aspartic acid residue at this position accepts two hydrogen bonds from an
arginine and m akes an optim al hydrogen bond to a cytosine on the opposite
D N A strand . The crystal struc tu ra l data of the T ram track com plex also
illustra ted novel features. Thus, finger 1 o f Tram track has an add itional 3
strand , w hich doesn 't make contacts to DNA, bu t is probably required for the
overall stability of finger 1. The pattern of amino ac id /b ase contacts in finger
1 differs from those seen in finger 2 and in all three fingers of EGR-1: the
h istid ine a t position -1 does no t make a base-specific contact; in stead , it
contacts a phosphate; a short serine sidechain a t position 2 contacts a thym ine
(this is facilitated by the DNA bending), whereas in zinc fingers 1 an d 3 of
E G R -lanalogous contacts involve aspartates hydrogen bonding to adenine
bases on the opposite strand of the DNA. Curiously, about 50 % of zinc finger
sequences have a serine residue conserved a t this position (Fairall e t al., 1993).
Overall, although differing in some of the exact contacts, there is a rem arkable
concordance betw een the two structures.
Recently, the struc tu re of a designed zinc finger p ro te in b o u n d to
consensus DNA sequence has been solved (Kim & Berg, 1996). The overall

39
m ode of binding resem bles tha t o f EGR-1, w ith m ost o f the am ino add-base
contacts occurring as predicted. Finally, the cocrystal structure of the four zinc
finger protein YYl (Ying-Yang 1) com plexed w ith a 20-bp adeno-assodated
v irus P5 initiator has been reported (Houbaviy e t al., 1996). YYl is a hum an
G L I-related p ro te in capable o f su p p o rtin g specific in itia tio n o f m RNA
p ro d u c tio n (H oubaviy et al., 1996). A d iag ra m sum m ariz ing YYl-D NA
interactions is show n in Figure 1.17. All four fingers o f YYl bind in th e major
groove, w ith fingers 2 to 4 m aking m ultiple contacts w ith DNA. F inger 1, in
contrast, contacts only one base. YYl uses sim ilar a-he lical positions w ith in
the fingers as those used by EGR-1, GLI, an d T ram track for m ak ing base-
specific contacts: -1, 2, 3, and 6 (Figure 1.18 A). A schem atic com parison of the
DNA contacts m ade by EGR-1, GLI, and YYl is show n in Figure 1.18 B. An
u n u su a l feature of the YYl-DNA complex is the a b ru p t strand sw itching,
w hich is p roposed to play an essential role in estab lish ing the p o la rity of
transcrip tion from the initiator elem ent. The d is trib u tio n of DNA contacts is
a lm ost even betw een the two stran d s of DN A (tem plate and non-tem plate)
until the strand sw itch occurs in the m iddle o f the b ind ing site, a t AlO - T31
an d T i l - A30 base pairs (Figure 1.17). F rom AlO to A17, the rem aining
interactions are m ade to the tem plate strand w ith only one contact involving
the nontem plate s tran d (Figure 1.17). The m echan ism of strand sw itching
w as observed in o ther zinc finger - DNA complexes, b u t no t to the sam e
ex ten t as seen in YYl protein - DN A complex. In co n d u sio n , th is crystal
structure reveals EGR-1 - like b ind ing by fingers 3 and 4, w hile finger 1 mainly
m akes phosphate contacts, bu t on ly one contact to a DN A base, an d finger 2
uses an unusual lysine - g u an in e contact fro m p o sitio n 3 (F igure 1.17).
O verall, the YYl illustrates new versatility o f the zinc finger D N A binding
m otif.

40
io > - [ T I^ N 3 6 ^ )^ H 3 4 3
V346
•<-K351
Figure 1.17 Schem atic represen tation of the YYl-DNA interactions. The
tem plate strand is labeled T . Protein-DNA contacts are denoted w ith arrow s
(Houbaviy et al., 1996). The details of the complex are discussed in the tex t

41
A i*« .«»I I <
M E PR T I ACPHKCXrrKMFRDNSAMRKHLHTHC I P - R
VHVCAE- -CCKA FVESSKLKRH Qt,VH T ICEK
I
PFQCTFEGCG KRPSLD FX LRTHVR [ HT | GOR
*$#I I I
PYVCPFOGOnCKFAQSTNLKSHrt.THA ( KAJCKNQ
r iY2Y3Tf4Z1Z2Z3G2G3C4G5T1T2
: (x x x
S «
.4* 4*4- =4- • • ♦♦ ♦
B
GLI Axn
Figure 1.18 Com parison of YYl w ith other zinc finger structures.

42
Figure 1.18 C om parison of YYl w ith other zinc finger structures. (A) Amino
acid sequence of YYl. The zinc fingers are aligned by their zinc-coordinating
residues. The YYl contacts are indicated as follows: • , backbone contact; +,
base contact, =, touches both base and backbone. These contacts are also
presented w ith those seen in the structures of EGR-1 (Z1-Z3), GLI (G2-G5), and
Tram track (T l, T2). (B) Schematic comparisons of the DNA contacts m ade by
EGR-1, GLI, and YYl. DNA bases and backbone are represented by rectangles
and circles, respectively. Hatched rectangles and solid circles denote sites of
protein contact (H oubaviy et al., 1996).

43
1.1.6 Studies toward a zinc finger recognition code
The recognition principals deduced from the o rig ina l EGR-1 crystal
structure p rom p ted researchers to apply the recognition ru les o f EGR-1 to
other C2H 2 proteins, such as S p l transcription factor, the m am m alian W ilms'
tum or p ro te in (H aber and Backler, 1992), Xenopus tran scrip tion factor UIA,
the yeast M IG l repressor (Nehlin and Ronne, 1990) an d others. C om parison
of the sequences of m ore than 100 0 zinc finger m otifs has show n th a t am ino
ad d s in positions involved in direct base pair contacts in EGR-1 com plex are
particu larly variable in the other zinc fingers (Jacobs, 1992). O bservations
such as these seem to indicate that EGR-1 and a t least its close relatives
recognize DNA in a sim ilar m anner. This raised an e x d tin g possibility of
p red ic ting DNA b in d in g sites for p ro te ins w ith u n k n o w n sp ed fic ity , or
creating new proteins designed to recognize specific DNA sequences. This, in
turn, cou ld open new opportunities for bo th the study of gene regu la tion and
de novo design of p ro te ins w ith preprogram m ed sp e d f id tie s for therapeu tic
use.
In d e e d , such ideas have been developed in m u ta tio n a l s tu d ies
involv ing the con tact residues, some o f w hich re su lte d in p ro te in s w ith
changed D N A b ind ing sped fid ties (N ardelli et al., 1991; Desjarlais an d Berg,
1992). Desjarlais an d Berg designed several po lypep tides o f the S p l DNA-
binding dom ain tha t w ere m ade up of fingers of know n sp e d fid ty , an d every
po lypep tide d isp layed essentially the expected sequence p reference. The
observed "recognition ru les " were analyzed and com piled in to databases
(Nardelli e t al., 1991; Desjarlais and Berg, 1992). Subsequent stud ies show ed
that application of som e of these rules successfully enables zinc fingers to bind
to predeterm ined D N A -binding sites, and that those ru les can be transferred
from one p ro tein to ano ther (Desjarlais & Berg, 1993). H offm an e t al. (1993)

44
in troduced am ino a d d changes a t one of the three critical positions o f the
yeast transcrip tion factor A D R l, which reduced DNA b in d in g affinity w hile
n o t d isrup ting the structure of the finger. This suggests th a t the m u ta ted
am ino a d d s w ere those involved in direct contacts w ith DNA.
The w ork of Kriwaki et al. (1992) and Kuwahara e t al. (1993) p resen ted
evidence that fingers 2 and 3 of the transcription factor S p l b ind w ith h igher
affinity than does finger 1. This finding is also consistent w ith the u se of
EGR-1 contact positions, w here fingers 2 and 3 provide m ore hydrogen bonds
th an finger 1 . Furtherm ore, in one of the earlier stud ies derivatives o f the
S p l protein w ere generated, and bound to a predicted site w ith an affinity o f 2
nM (Nardelli e t al., 1991).
A differen t approach to designing a pro tein w ith the p rede term ined
b ind ing specificity involves a selection rather than a rational design strategy .
This technique is known as phage display, and is now being extensively used
by different laboratories (review ed in Choo & Klug, 1995) (a brief descrip tion
o f the technique can be found in Chapter 1). This has p roved to be a pow erfu l
selection m ethod used in m any studies of D N A -binding proteins. Som e of
the exam ples of using phage display to study zinc fingers include the stud ies
b y Rebar and Pabo (1994), Choo and Klug (1994a, 1994b), an d Jam ieson e t al.
(1994; 1996).
Rebar and Pabo (1994) used a phage display library expressing EGR-1-
p hage coat p ro tein fusions w hich contained random ized critical am ino a d d s
in the first finger. R ounds of affinity selections u sin g o ligonucleo tides
random ized in the region recognized by the first finger y ie lded v a rian ts of
EGR-1 carrying different am ino a d d s in zinc finger 1 , th a t b o u n d specifically
to the new DNA sites.

45
C hoo & Klug (1994a) in a very com prehensive s tu d y constructed a
phage d isp lay library containing over 2 m illion variants o f EGR-1 in w hich
the am ino acid sequence of finger 2 was random ized a t e igh t positions. This
lib rary w as th en screened against different DNA triplets rep resen ting DNA
bind ing subsites. As a result, 16 successful triplets were isolated and m atched
to a subpopu la tion of fingers able to bind them. A strong bias was observed
tow ards the base contacting positions revealed in the crystal structure of EGR-
1 (Choo & K lug, 1994a). O th e r stud ies used sim ila r stra teg ies, and
d em onstra ted th a t the phage d isp lay system is a pow erful tool for selecting
zinc fingers w ith novel DNA b ind ing specificities, w hich could be u sed in
research or ev en therapy.
A nother interesting study reports in znvo repression o f transcrip tion by
a C2H 2 zinc finger protein designed to specifically recognize an oncogenic
sequence (C hoo e t al., 1994c). A protein containing three zinc fingers w as
designed u sing a rationale based on phage d isp lay selection as well as the
available s tru c tu ra l data. A nuclear localization signal w as attached to the
p ro te in , w hich d irected it to the nucleus w here it bound its chrom osom al
DN A targe t sequence, and caused a specific inhibition of transcrip tion in
v iv o .
Several research groups have m ade a ttem pts to describe a DN A
recognition code for transcription factors. The group led by M asashi Suzuki is
one of the m ost determ ined ones, trying to reconcile the available struc tu ra l
d a ta to d ed u ce general rules for protein-D N A recognition (Suzuki, 1993;
Suzuki e t al. 1995). A recent publication by C hoo and Klug (1997) discusses
the fram ew ork of the zinc finger-DNA recognition code. H ow ever, a t the
p resen t tim e there isn 't sufficient data to draw any far-reaching conclusions.
The rem ark by H arrison and Aggarwal that "the variety of protein-base pair

46
contacts is m ore evident than their uniform ity" (A ggarw al e t al. 1988) still
holds today.
The new h igh resolution structures of zinc finger p ro te ins (discussed in
the previous sections of th is thesis) reveal very com plex in teractions a t
protein-D N A interfaces, and em phasize the difficulties in h eren t in try ing to
develop sim ple rules of protein-D N A recognition. M ore struc tu ra l da ta is
necessary for elucidation of a m ore com prehensive set o f ru les w hich cou ld
be used to p red ic t DNA b ind ing sites from the am ino acid sequence of a
pro tein .

47
CHA PTER 2.0 T H E W T l W ILM S’ TU M O R GENE PR O D U CT: A TUMOR
SUPPRESSOR INVOLVED IN REGULATION OF KIDNEY DEVELOPMENT.
2 .1 In troduction
2 .1 .1 T he concept of tum or suppressor genes.
The concept of tum or suppressor genes w hich play a role in negatively
regu la ting cell g ro w th was in troduced in th e late 1960s (H arris et al., 1969).
U ntil then, the concept that DNA and R N A tum or viruses in troduced new
oncogenes that acted in a genetically dom inan t fashion w as the only accepted
m echan ism le a d in g to neop lastic tran sfo rm a tio n . T he iso la tion an d
identification of tum or suppressor genes took m any years o f exploring using
various cloning a n d som atic cell hybrid ization techniques. These extensive
s tud ies have y ie lded a num ber of hum an tum or suppressor genes including
retinoblastom a (RB) (reviewed by W itm an, 1993; Hamel, 1993), p53 (reviewed
by Zam betti an d Levine, 1993), neuroblastom a (Brodeur, 1990), and Wilms'
tu m o r gene (H aber and H ousm an, 1992). It is w ith o u t d o u b t that the
d iscovery of m any new tum or suppressor genes is forthcom ing, and that the
understand ing o f these genes and their p roducts will even tually lead to the
developm ent of new clinical strategies and preventative therapies.
2.1.2 Biology of W ilm s' tum or and associated syndrom es.
W ilms' tu m o r is a pediatric k idney m alignancy th a t arises through
a b e rran t d ifferen tia tion of the k idney s tem cells. It rep re sen ts the m ost
com m on solid tu m o r of ch ildhood, affecting app rox im ate ly 1 in 1 0 , 0 0 0
c h ild re n , an d acco u n tin g for 85% o f a ll p e d ia tric n ep h ro b la s to m as
(Stanbridge, 1990; H aber and H ousm an, 1992). The average age of children
affected by W ilms' tum or is 3.5 years old, b u t m any of them d ie before the age
o f 2. In rare cases, th is tum or m ay develop a t an older age, typically in the

48
m id-20s . The tre a tm e n t of th is m alignancy in c lu d es su rg e ry and
chem otherapy, and , in the case o f advanced disease, radiation therapy. These
trea tm en ts usually resu lt in good long-term su rv iv a l (5 years follow-up):
a b o u t 80% of W ilm s tum or patien ts are successfully cured (Lem erle e t al.,
1983; D'Angio et al., 1989).
W ilm s’ tu m o r can occur in either fam ilial o r sporadic fo rm s, w ith
a lm ost 99% of the tum ors arising sporadically. M ost of the tum ors develop
unilaterally , and on ly 5-10% of children are affected w ith the bilateral form of
th e d isease, w h ich is usually inherited . H isto log ically , W ilm s tum ors
orig inate from residual islands of im m ature kidney, and consist of blastem al
cells as well as epithelial cells and muscle cells o f the supporting strom a. In
the m ajority of the cases W ilms' tum or is n o t associated w ith any other
system ic disease. H ow ever, it can be part of the follow ing syndrom es: the
W AGR syndrom e, w hich accounts for 1-2% of cases of VV ilms’ tu m or, and
in c lu d e s A .niridia (m alfo rm ation o r absence o f the iris), u ro G e n i ta l
m alform ations, and m ental R etardation (Baird e t al., 1992a); the Denys-Drash
syndrom e (2% of all W ilm s’ tum ors), which also includes various anomalies
of th e genitourinary tract and nephropathy (Coppes e t al., 1992b; Baird e t al.,
1992b); Beckw ith-W iedem ann syndrom e (BWS) w hich is characterized by
g ro w th abnorm alities an d a predisposition to W ilm s’ tum or (Koufos et al.,
1989; H enry et al., 1989). All these syndrom es have in com m on the more
frequen t occurrence of bilateral tum ors which appear a t an earlier age than in
spo rad ic cases no t associated w ith syndrom es. Based on the sim ilarities of
tu m o r p resen ta tion betw een the W ilm s’ tu m o r an d re tinob lastom a (RB)
(nam ely, an association betw een fam ilial inheritance an d bilateral onset of
tum ors), a "two-hit" hypothesis w as proposed by K nudson e t al. (Knudson,
1971; K nudson a n d S trong, 1972). A ccording to th is hypo thesis , two

49
in d e p e n d e n t even ts leading to inactivation o f both a lle les o f a tum or
su p p re sso r gene are requ ired for the tum or to develop. W hereas som e
W ilm s' tum ors (the familial cases in particular) m ay conform to the tw o-hit
hypothesis, the genetics of the majority of W ilm s' tum ors are likely to be
m uch m ore complex. There are several lines of evidence to suggest a more
com plex developm ent of W ilm s' tum or. F irst, the incidence of fam ilial
W ilm s' tum or cases is only 1 -2 %, which is very low com pared to the 35-40%
for re tin o b la s to m a (B rodeur, 1990; P e lle tie r e t al., 1991c). Second,
retinoblastom a results from inactivation of a single tum or susceptibility gene
located a t chrom osom e 13ql4 (Hamel et al., 1993). In the case of W ilms'
tum or, three separate genetic loci have been linked to p red isposition to the
disease.
The association of W ilms' tum or w ith W AGR syndrom e has been of
p a rticu la r im portance in iden tify ing a W ilm s' tum or su scep tib ility gene.
W AGR p a tien ts carry cy togenetically v is ib le ch rom osom al de le tions
invo lv ing band p l3 of the sh o rt arm of one o f the two chrom osom es 11
(Riccardi e t al., 1978; Francke e t al., 1979; H astie , 1994). T his observation
initially generated interest in the chrom osom al region l lp l3 . A dditionally,
abou t 20% of sporadic Wilms' tum ors show loss of heterozygosity (LOH) for
l l p l 3 m arkers, as indicated by analysis of restric tion frag m en t - length
polym orphism s (RFLPs) (Lewis et al., 1988; T on e t al., 1991). The WAGR-
asso d a ted deletion was later resolved into tw o separate loci: an anirid ia gene
(A N 2/Pax-6: sm all eye in chrom osom e 2 in m ouse), and a pu ta tive Wilms'
tu m o r supp resso r gene W Tl (Gessler et al. 1990). F u rther physiological
ev idence for th e involvem ent of chrom osom e 11 in W ilm s' tum ors w as
p ro v id ed by the observation th a t transfer of a norm al chrom osom e 11 (but

50
n o t chrom osom es 13 or X) in to cultured W ilm s' tu m o r cells can suppress
the ir tum origenic phenotype (W eissman e t al., 1987).
A second locus was m apped to chrom osom al reg io n l l p l 5 which has
b e en im p lica ted in B eckw ith-W iedem ann syndrom e a n d W ilm s' tu m o r
(C oppes e t al., 1992; H en ry e t al., 1989; Koufos e t a l., 1989). Loss of
heterozygosity for chrom osom e l lp l5 m arkers occurs in abou t 40% of W ilms'
tum ors (Coppes et al., 1992). This locus contains genes for IGF-n (insulin-like
g ro w th factor) and H19 mRNA. Changes in im prin ting of these genes have
been linked to tum origenesis (Zhang and Tycko, 1992; O gaw a e t al., 1993;
Rainier e t al., 1993).
A th ird locus, a lbeit n o t involved in h e red ita ry p red isposition to
W ilm s' tum or, is frequently lost d u rin g tum or p ro g ress io n , an d is thus
considered im portan t in W ilm s' tum or developm en t (C oppes et al., 1992;
M aw e t al., 1992). Tumor LOH for m arkers a t chrom osom e 16q is found in
a lm ost 20% of W ilms' tum or, and is alw ays accom panied by LOH at l l p l 3
an d l l p l 5 (Coppes et al., 1992).
2.1.3 Id en tifica tio n and characterization o f the W T l g e n e an d its pro tein
p ro d u c t
Three independen t g roups have rep o rted clon ing of the W Tl gene.
Genom ic probe WiT-13 was isolated by Call e t al. from a cosm id library of the
sh o rt a rm of chrom osome 11 (Call et al., 1990), which w as dele ted in sporadic
W ilm s' tum or. This deletion w as initially identified by loss of DNA m arkers
a t l l p l 3 (Lewis e t al., 1988). The WiT-13 DNA fragm ent w as then used to
p ro b e several cDNA libraries, and a cD N A clone d e s ig n a ted WT33 w as
isolated. By conducting N orthern blot analysis, this cD N A clone w as further
show n to hybridize to 3 kb-long mRNA species in baboon k idney and spleen.

51
w hile no detectab le h y b rid iza tio n w as observed in RNA d e riv e d from
m uscle, live r, o r brain (Call e t al., 1990). Sim ilarly, the WIT-2 gene was
iso lated by Bonetta e t al. (1990), using yeast artificial chrom osom es, and
show n to be identical to the WT33 sequence. Gessler e t al. (1990) u sed an
alternative strategy of chrom osom e jum ping to isolate the W Tl gene. First, a
genom ic p ro b e w as ob ta ined using rare-cutting enzym es, w hich p ro d u ced
sequences ind icating th e 5 '-end of a gene. This p ro b e m ap p ed w ith in
c h ro m o so m a l reg io n l l p l 3 w hich is d e le ted in W AGR p a tie n ts .
Subsequently , using this p robe, a cDNA clone term ed LK15 was iso lated from
a hum an fetal kidney cDNA library.
A n a ly s is of th e W T l gene sequence re v e a le d th a t i t sp a n s
approx im ate ly 50 kbp of DNA, and contains 10 exons. The W Tl m R N A is
abou t 3.5 kilobases long, a n d encodes a protein w ith a m olecular w eigh t of 47
to 49 kOa, depending on the presence or absence of tw o alternatively spliced
exons (H aber e t al., 1991). The W Tl m RNA is expressed in a lim ited num ber
of cell types, prim arily k idney cells and som e hem atopoetic cells (Gessler et
al., 1990; C all e t al., 1990). The predicted protein sequence has functional
dom ains considered to be hallm arks o f transcrip tional regu latory p ro te in s
(Figure 2.1). The amino term inus of W T l is rich in p ro line and g lu tam ine
residues (Lewis et al., 1988), which has been found in transactivation dom ains
o f a num ber o f transcription factors, such as EGR-1, SPl, C T F /N F l, an d others
(review ed in Pabo and Sauer, 1992; M itchell and Tijian, 1989). The carboxyl
term inus o f W T l contains four zinc fingers of C 2H 2 type, w hich rep resen ts a
pu tative D N A binding m o tif characteristic of a fam ily of sequence-specific
D N A -binding proteins (Pabo and Sauer, 1992). Fingers 2-4 of W Tl possess a
significant sim ilarity to the three zinc fingers of the im m ediate-early g row th
response genes, EGR.

52
W T lH G S D V R D L M A . L L P A . V P S L G G G C C C A L P V S
G A A Q W A P V L O P A P P G A S A Y G S L G G P A P P P
A p p p p p p P P P H S P r K Q E P S M C C A E P H E E O
C L S A P T V H P S G Q F T G T A G A C R l f G P F G P P P
F s q a s s g q a r m f p n a p y l p s c l e s q p a i r
n q g y s t v t f d g t p s y g h t p s h h a a q f p n r
S F K H E D P M G Q Q G S L G E Q Q Y S V P P P V Y G C Ht *
t p t d s c t g s q a l l l r t p y s s d m l y q m t s q
yAAGSSSSVKWTEGQSN
l e c m t w n q k n l g a t l k g ^h s t g y b s o n h t t
P r L C G A Q Y R I H T H G V F R G I Q ^ O V R R V P G V A
P T L V R S A S E T S E K R P P K © A Y P G © N K R Y P K
L S R L Q K © S R K © T G E K P Y Q © D F K D © E R R F F
R S D Q L K R © Q R R © T G V K P F Q © K T © Q R K F S ®
KTSS @ B L K T 0 T R T 0 T G^E K P F S © R W P S © Q K K F A
R S D E L V R a © R K 0 Q R N K T K E . Q L A L *
Figure 2.1 The am ino a d d sequence o f W Tl protein: the amino term inus is
rich in proline, g lu tam ine and g ly d n e residues, com m only occuring in
dusters; the DMA-binding zinc finger region is in the carbooxyl terminus; the
zinc-coordinating cysteines and h istid ines are d rd ed ; serine residues that are
potential targets for the D N A -activated protein kinase a re m arked by an
asterisk; the 17 am ino a d d , and th e three amino a d d insertions resulting
from alternative splicing are indicated above; the arginine and aspartic a d d
residues of finger three found m u ta ted in Denys-Drash patients are boxed
(Rauscher, 1993).

53
Four distinct isoforms of W T l can be generated by differential splicing
of th e prim ary transcrip t, reflecting the presence or absence o f tw o splice
insertions. The 17 amino a d d insertion is located in the am ino term inus,
and contains a num ber of contiguous serine residues. The th ree am ino a d d
insertion (lysine-threonine-serine, o r KTS) occurs betw een fingers 3 and 4 of
the h igh ly conserved DNA b ind ing dom ain (Figure 2.2). The m ost abundan t
W T l m RNA species is the one containing bo th insertions, an d the least
com m on is the one w ithout both insertions. The ratio of the fou r alternative
splice variants is conserved in h u m an and m ouse, and does n o t seem to be
developm entally regulated (H aber e t al., 1991). A m echanism of alternative
splicing to generate different b ind ing specifidties has been docum ented for a
num ber o f other zinc finger proteins. The chorion transcrip tion factor CF2
and the Tram track proteins, both from Drosophila, use differential splicing to
alter DN A-binding specifidties (Gogos e t ai., 1992; H su et al., 1992; Read et al.,
1992). A nother pro tein , proto-oncogene Evi-1 also undergoes a lternative
splicing in its zinc finger region (Bordereaux et al., 1990). H ow ever, it is n o t
yet k n o w n w hat effect this has o n DNA binding. Further heterogeneity of
W Tl isoform s m ay be generated by a non-AUG translational in itiation event,
as recently reported by Bruening a n d Pelletier (1996).
The presence of serine res id u es, especially in the 17 am ino a d d
alternative splice insertion, suggests potential targets for phosphory lation .
H ow ever, despite the abundance o f serine residues, the W Tl p ro te in appears
to be poorly phophorylated in COS-1 cells (Morris et al., 1991). M ore recent
in form ation suggests that the +KTS splice varian t of W Tl m ay be a better
candidate for phosphorylation (A nant e t al., 1994). Recently, Ye e t al. (1996)
dem onstra ted th a t bo th +KTS a n d -KTS isoform s of W Tl a re subject to
phosphorylation by both protein kinase A (PKA) and protein kinase C. These

SI
mRNA II7aa KTS
2 1 3
Proteins
A
B
C
a s ^ o c w d o n d o m a in ^
182re p re u io n domain activation domainminimal minimal
o n e finqer*
84 124 180 181 250 234 3 30 449
H B HI H i
Figure 2 .2 Schematic diagram o f the structure of the WTl m RN A and
proteins. Stippled boxes represent nucleotides or am ino adds th a t m ake up
the alternative splice insertions. H atched boxes represent proline-rich regions
of the protein. The exons of the m RNA and the zinc fingers of the protein
are num bered (Reddy & Licht, 1996).

55
phosphory la tion events inhibited the D N A -binding ability of b o th proteins,
as m ea su re d in gel m obility sh ift assays. M oreover, activa tion o f PKA
resulted in the reversal o f W Tl suppression of a repo rte r construct, as well as
in a lte ra tion in nuclear translocation of the p ro teins (a change from nuclear
to cy to p lasm ic re ten tio n ). T hus, p h o sp h o ry la tio n of W T l can have
significant an d pleiotropic effects on the function of the protein.
W T l protein also contains potential sites for N -linked glycosylation in
the 17 am ino acid insertion , as w ell as a num ber of potential recognition
m otifs fo r a D N A -activated pro te in kinase (Lees-M iller, et al., 1991). The
functional significance o f these features needs to be investigated further.
C onsisten t w ith its proposed function as a transcrip tional regulator,
W Tl has been show n to localize to the nucleus (Pelletier et al., 1991; M orris,
et al., 1991). Using im m unostaining an d confocal m icroscopy, it w as recently
o b served th a t WT1[+KTS] isoform co-localized p red o m in an tly w ith the
sp licing factors, w hile the WT1[-KTS] varian t p rim arily a ssoc ia ted w ith
transcrip tion factors (Larsson et al., 1995). It rem ains to be seen w hether
WT1[+KTS] or other isoform s of the pro tein play a role in mRNA splicing.
W ith respect to the subnuclear localization o f W T l isoforms, Englert et
al. (1995) have recently dem onstra ted that W Tl truncation m u ta n ts have
d is t in c t s u b n u c le a r c o m p a r tm e n ta l iz a t io n p ro p e r t ie s . U sin g
im m unofluorescence stud ies it w as dem onstra ted th a t WT1[-KTS] had a
d iffuse p a tte rn of exp ression th ro u g h o u t the e n tire nuc leus, w hereas
WT1(+KTS] p roduced a speckled appearance in subnuclear s truc tu res .
S im ila rly , tru n ca tio n m u tan ts o f W T l ex h ib ited a sp eck led nuc lear
d istribu tion . These speckled s truc tu res are sim ilar to the in te rch rom atin
granules w hich contain spliceosom al com ponents, a n d are though t to play a
role in sp liceosom al assem bly. M oreover, the d o m in an t-n eg a tiv e W Tl

56
m utants w ere show n to physically associate w ith the w ild-type WT1[-KTS] in
v ivo , a n d thus a lte r the nuclear localization o f the w ild -ty p e p ro te in ,
changing the d istribu tion of WT1[-KTS] form from a d iffuse to a speckled
pa tte rn and its subsequent functional inactivation. It is o f in te rest to po in t
ou t th a t m any of the truncation m utations iso lated occur in a heterozygous
state, w ith the w ild W T l allele still present (H aber et al., 1990; Pelletier et al.,
1991). Pelletier e t al. (1991) proposed that these isoforms, deficient for DNA
b inding , can act as dom inant negative m utants. Thus, d o m in an t m utations
of W Tl m ay lead to tumorigenesis w ithout requiring the loss of the wild type
allele (H aber et al., 1990). The m echanism by w hich m u tan t W T l proteins
can sequester w ild -type W Tl is analogous to th a t proposed for dom inant-
negative p53 m utan ts (Michalivitz e t al., 1990; M artinez e t al., 1991). The
difference betw een the two is that m utant p53 m ay associate w ith the w ild-
type p ro te in in the cytoplasm , while W Tl changes its sublocalization in the
nucleus. The m olecu lar m echanism for the dom in an t-n eg a tiv e effect
observed for W Tl truncation m utants was investigated by several groups,
and found to be due to protein self-association (Moffett e t al., 1995; Reddy e t
al., 1995). W Tl p ro te in was found to self-associate in vitro a n d in the yeast
tw o-hybrid assay (R eddy et al., 1995). It was found , in fact, th a t some W Tl
trunca tion m utan ts can oligom erize m ore efficiently th an w ild-type W Tl
(Moffett e t al., 1995), thus antagonizing the function of the w ild-type allele. In
addition, the dom ain of W Tl necessary for self-association w as m apped to the
N -term inal 182 am ino a d d s (Reddy et al., 1995).
2.1.4 D N A -binding activ ity of W ilm s' tum or p ro tein .
Once it becam e clear that the W Tl gene encoded a transcrip tion factor
containing four zinc fingers of the C2H 2 dass, the search began to identify the

57
DNA sequence the W T l protein recognizes. The first DN A-binding sequence
was identified by Rauscher et al. (1990). They used a strategy of selecting a
D N A -b in d in g se q u en c e o u t o f a pool o f ra n d o m iz e d sy n th e tic
oligonucleotides. The authors u sed a bacterially expressed recom binant
p ro te in encoding the four zinc fingers of the WT1[-KTS] isoform . The
resu lting selected sequence w as very GC-rich, a n d sim ilar (a lthough not
identical) to the EGR-1 consensus binding site 5 -GCGGGGGCG-3' (Gao et al.,
1990). Further analysis of the D N A -binding p roperties of W Tl an d EGR-1
proteins, involving com petition an d binding site m u ta tion experim ents have
confirm ed that EGR-1 and W Tl recognize a com m on DNA core elem ent
(Rauscher et al., 1990) (Figure 2.3 row A). This resu lt w as perhaps n o t so
su rp ris in g since bo th proteins sh a re a m odera te deg ree of am ino acid
sequence sim ilarity (about 60%) in the zinc finger region, w ith 6 o u t of 6
residues proposed to be im portant for DNA recognition conserved betw een
the tw o proteins (Pavletich and Pabo, 1991). Subsequently , using transien t
transfection assays, W Tl protein w as show n to regu la te transcrip tion from
prom oters containing an EGR-1 consensus binding site (M adden et al., 1991).
For quite some time the EGR-1 consensus site w as presum ed to be a canonical
WT1(-KTS] binding sequence. H ow ever, the results of o ther studies aim ed at
identification of W Tl DNA binding sites have revealed extended and
additional DNA binding sequences. These results are sum m arized in Figure
2.3.
A novel, TC-rich binding site for WT1[-KTS] w as iden tified from
DNAse footprinting analysis of the PDGF A-chain prom oter by W ang et al.
(1993a) (Figure 2.3 row C). This second W Tl binding m otif functioned equally
well in co-transfection experim ents as the EGR-1 consensus sequence (W ang
et al., 1993). The TCC repeat sequences were found w ith in a num ber of

58
S E Q U E N C E S W H IC H B IN D W T I f - K T S ) :
A RauscMf ef J/.. 1990 (random oigocnen)nc. (X'louuLAc. cfcaxcic. iu.iJir.icc . c x a x r z
(Ae auO ors d eo d ed on C n C T f T m r 'consensus* ta x e d on nocnalogy wott Egr-i
B. BckmoreetaJL. l992(genocraciiagfiwncx)ICTCTdCTdCTCTCTGTCTCTGrdCIGTOni^^iv>uuv>iv>iv>iuiviiviiuiuiClC(dUli;iUU>ICldC>ldC>lClUJn>il'llCCK2XCriAAAAAMCNiiccNXAiiriciiauLii'i'i-uuacMSiosrrciciUjTtuicictagiciciuiaJir-G'iG'i&'ni;ic n jio ta ;L M C .ic n ^ 'K;tuiu ic ium t;ic ;ux ;i'n ;r
C WangeraCI993(POGf^|itocro(e(); l O r rCCTOLUXTClOC
0. Nalagasnaefa(.l99S(gerainc sequence): CCS3SCCHSX
e. Haneton etaL I99S (random oigcrnen}: nOGTOOOOG(T/GI (C/A/T) (T/C)
F. RvaneraC l99S(Pax.2p<omoce«j: in r i-^-rrrfYTrraTrYTrrryrTTyr^(nrjkjuiur arrrxrcnmTrrarrnrTrir
SEQUENCES WHICH BIND BOTH VyTtMCTSl AND WTUa-KTSl:
G. Biciano(ae(a(.l992 (genomic sequences):a a rT ita a 'i i iix v ic m sic tj 'i i. 'ii ix ’iiM i.ni.’ita rtam n ry i'A i'i 'iT rjiiA t'l 'r-iiiw fa ran ram —A icx n iG K Z ccx cK T n cM C N X x zx aazzau a tsc rA3xncnazacixncnazac3xiamiaaxx2aGsaaGaz2R>Ma3iK3CMcicicr—ocrmxBicnGiGriGmAMcraximmGMaicnumacncicmcitxïDcriCDGr—ooœ cicnooT
H. Oiummond efaL I9M OGFJ: pmmoKer): if ^ iinrrrrrrrnnnnrrfgoggsÛOOOOQOCQZnoga g g a t f Z X m Z m z r ç gmis group also identKedGTC bom random oCgocnets as taxes best reoognûed by linclinger l
L ApptecMefaLI994(WTlpromoreq: OCHOCX30CWOOC. (UOGMOCTCTC
J. Wang et aL 1995 (POtff'A promoter):çyxRX nG yjffT T m T T rm am L 1 u m x lY M rm nnnrw i m m i m c m r c
K. RyanetaL 1995(Pax.2promoter):(NndsXTS«re#.binds «KTSalhigboonoenbalions) frTratTrrTrrrrrrrTTrt-ar-j-rj-i-i i «rr arrrrarr-
Figure 2.3 DNA sequences which bind W Tl proteins (Reddy & Licht, 1996).

59
p rom oters o f o ther g row th-related genes, such as Ki-ras, epithelial g row th
factor receptor, insulin receptor, c-myc, and tum or grow th factor p3 (W ang et
al., 1993). Thus, as ev iden t from the transient-transfection assays, W Tl can
function on a varie ty of prom oter sequences. How ever, the relevance of the
in vitro determ ined sequences to prom oter regulation in the chrom osom al
context has no t been determ ined.
The m o st a b u n d a n t sp lice varian t, WT1[+KTS], co n ta in in g the
trip ep tid e lysine, threonine and serine in the H-C linker reg ion betw een the
th ird and forth W T l zinc fingers, does not b in d to the EGR-1 core sequence
appreciab ly (Rauscher e t al., 1990). Since WT1[+KTS] rep resen ts the m ost
a b u n d an t isoform of the W T l protein, subsequent attem pts w ere aim ed at
identification of a b inding site for the WT1[+KTS].
Bickmore e t al. (1992) used a whole-genom e PCR approach to isolate
genom ic DNA sequences th a t b ind to either -KTS or +KTS splice variants of
W T l. The tw o W Tl isoform s w ere show n to have d ifferent DNA b inding
specificities. Sequences iso la ted w ith WT1[-KTS] form h a d runs o f GT
nucleotides (Figure 2.3 row B). In contrast, WT1[+KTS] d id no t select any GT-
rich sequences. Instead, tw o extended sequences (+P2 and +P5) of about 100 bp
w ere identified, each of w hich containing an EGR-l-Iike consensus b ind ing
site (Figure 2.3 row G). Interestingly, DNAse I footprin ting or m éthylation
interference analysis failed to identify the precise nucleotide sequences bound
b y the WT1[+KTS] zinc fingers. In addition, the analysis perform ed in this
s tu d y was only qualitative, w ithou t quantitative assessm ent of the b ind ing
affinities.
R upprecht e t al. (1994) h ad carried o u t footprinting analysis of the W Tl
p rom oter, and found m ultip le W Tl binding sites, on w hich W Tl acted as a
transcriptional repressor. Two of the WTl b ind ing sites could b ind both -KTS

60
and +KTS isofonns, and w ere not related to any o ther, previously identified
W T l b in d in g sequences (F igure 2.3 ro w I). H o w ev e r, m é th y la tio n
interference analysis again failed to reveal a distinct p a tte rn of protection due
to the WT1[+KTS] binding.
D rum m ond et al. (1994) analyzed binding of -KTS an d +KTS isoform s
of W Tl to the fetal IGF-II P3 prom oter, w hich is an active hum an k idney
prom oter (D rum m ond et al., 1992). Footprinting experim ents identified a 12-
bp sequence w hich bound bo th splice varian ts. T his site represents an
extended sequence containing features com m on to the EGR-1 consensus site.
A study of PDGF-A prom oter also identified a GC-rich sequence capable of
b ind ing both -KTS and +KTS variants (W ang et al., 1995) (Figure 2.3 row J).
This GC-rich sequence represents an extension of the EGR-1 binding motif.
H ow ever, the precise location of binding of WT1[+KTS] to this sequence is
no t clear. Recently, EGR-l-like sequences w ere also found to be contained in
the Pax-2 p rom oter (Ryan e t al., 1995) (Figure 2.3 row s F and K). Three
sequences w ere identified by footprinting assays; tw o of them bound WT1[-
KTS], and the th ird site could also b ind WT1[+KTS] , a lthough only w ith a
low affinity.
Thus, it rem ains unclear w hat constitu tes a WT1[+KTS] binding site.
Therefore, a t this point, it w ou ldn 't be p ru d en t to assign a specific DNA-
bind ing function to the WT1[+KTS]. Since it was dem onstra ted that the tw o
pro tein isoform s have distinct nuclear localizations, nam ely, that WT1[-KTS]
is found associated predom inantly w ith the transcription factors, w hereas the
WT1[+KTS] w as found w ith the splicing factors, it is reasonable to suggest that
the latter isoform m ay have a cellular function other th an binding DNA.
Since the three zinc fingers of EGR-1 and fingers 2-4 of W Tl have
structural sim ilarity as well as recognize a comm on D N A sequence, it w as

61
assum ed that W Tl zinc fingers 2-4 will in teract w ith the EGR-1 binding site in
a fundam entally similar m aim er. N am ely, that the struc tu ra l basis o f DNA
recognition for W Tl will be the sam e as found in the EGR-1 X-ray crystal
struc tu re , w hich is discussed in detail in C hapter 1 (Figure 2.4). H ow ever,
W T l p ro te in has an additional, first finger which suggests that W T l may
recognize extended DNA sequences.
To date , several g roups, including ours, have u ndertaken s tu d ies to
identify h igher affinity b ind ing sites for the WT[-KTS] p ro tein (Figure 2.3).
D ru m m ond e t al. (1994) perfo rm ed foo tp rin ting an a ly sis of the IGF-II
(insulin-like g row th factor) prom oter, a n d identified an ex tended , 1 2 bp
sequence containing three additional base pairs im m ediately dow nstream of
the EGR-1 consensus sequence. The au tho rs had show n tha t both WT1[-KTS]
and WT1[+KTS] isoforms could recognize the sam e 12-bp DNA sequence,
a lbeit w ith relatively low binding affinities (1.1 X lO'^S to 1.4 X lO'^^ M for
WT1[-KTS] and 2 - 7 X 10*06 m for W T l [+KTS]) (D rum m ond et al., 1994).
A dditionally , the authors also conducted binding site selection experim ents
a im ed a t identification of the W Tl finger 1 recognition subsite. They used a
p ep tide contain ing only zinc fingers 1-3 o f W Tl, an d fo u n d an add itional,
three-base pair sequence 5 -GTG-3' im m ediately dow nstream of the EGR-1
consensus sequence.
Recently, a whole-genome PCR w as used to identify physiological high-
affinity sites for the WT1[-KTS] (Nakagam a e t al., 1995). The identified 10-bp
sequence contained an EGR-1 - like site (except for an aden ine at the eighth
position) and an additional thym ine flanking the 3' end of the sequence: GCG
TGG GAGT (com pare to the EGR-1 consensus sequence GCG GGG GCG)
(Figure 2.3 row D). This sequence was show n to bind WT1[-KTS] w ith a 20 to
30-fold h igher affinity than the EGR-1 consensus sequence, w hich suggests

62
ONA-Protein Contacts Made By WTl and EGR-1
EGR-1
3 - G G A G C G G G G G C G -5 ' / \ / I I \/tSOELTK RSOHITT KSOERM
W Tl NH]
3 ‘-G G A G C G G G G G C G - 5' ? / \ / I / \
KI5HLQM JISOQUqi RSOHUa KOELVK
Figure 2.4 Protein-DNA contacts m ade by EGR-1 protein, and proposed DNA
contacts for WTl zinc fingers. The im portant recognition helix am ino ad d s
are show n. The aste risk s on finger 3 of the W Tl protein are residues
frequently m utated in Denys-Drash patients (Rauscher, 1993).

63
th a t W Tl and EGR-1 p ro te in share related, b u t n o t iden tical recognition
sequences.
In parallel w ith these studies, we conducted a binding site selection and
am plification assay in an a ttem p t to identify higher-affinity b in d in g sites for
WT1[-KTS] (H am ilton e t al., 1995). We determ ined th a t the h ig h es t affinity
binding sites for WT1[-KTS] consist of a 12-bp sequence, includ ing an EGR-1
consensus an d a 3-bp subsite recognized by zinc finger 1 (Figure 2.3 row E).
These experim ents are described in detail in C hap te r 3. O u r resu lts have
recently been once again confirm ed in the study by H ew itt e t al. (1996). It was
show n that zinc finger 1 recognizes an extended D N A binding m otif, where
guanines constitu te the d inucleo tide dow nstream o f the 9-bp EGR-1 - like
sequence.
On the basis of the above studies, as well as the EG R-l-D N A crystal
structural data , it is possible to propose contacts th a t are form ed in the W Tl-
DNA complex (Figure 2.5). H ow ever, more m utagenesis and structu ra l data
are necessary to unam biguously assign specific contacts to ind iv idua l amino
acids and DNA base pairs.
In conclusion, despite extensive studies, there is little ag reem ent as to
the DN A-binding specificity of WT1[+KTS] protein. The reported dissociation
constants (K js) for the WT1[+KTS] - DNA com plexes are typ ically several
orders of m agnitude h igher th a t those for the WT1[-KTS]. T he Kqs for the
WT1[-KTS] also vary from s tu d y to study, perhaps reflecting d ifferen t sources
of proteins, as well as different b inding assays and conditions used by different
researchers. Furtherm ore, m any groups do not perform quan titative analysis
of affinity constants, thus m ak ing it difficult to m ake d irec t com parisons
betw een different studies.
Recent studies described a specific RNA binding activity for W Tl

64
ZF<
H E R
ZF3
r H R
ZF2
R Q R
ZFI
M H K
P ro te in ,S o u r c e
IQ
s G C G T G G G A G r •> » - r WTU-KTS)(uO-tengtti
*1 v im translated
S X lO-'o Nakagama et al.. 1995
s G C G r G G G C G G a g 3 r G r
r
ZF(4CrS)bacterial
8.4 r 104 Hamilton et al.. 1995
s G C G G G G G C G G G C -3 ZF(-KTS)ZF{*KTS)bacterial
1.4 K 10^ 2.0X 10«
Drummond et ai ., 19945 - G C G G G G G C G G T G 3 Z F I Z 3
bacterialWD
G C G G ff' tS G A GC
G A G r o t
r cconsensus
Figure 2.5 Proposed W Tl zinc finger-DNA contacts (Reddy & Licht, 1996).

65
pro teins (Caricasoie e t al., 1996; Ye et al., 1996). T hese find ings fu rth e r
complicate the question o f the biological function of W Tl proteins and , a t the
sam e tim e, in troduce a new exciting possibility that one o f the im p o rtan t
roles of W T l proteins m ay be to bind to RNA. It w as first show n th a t the
n o rm al n u c le a r lo ca liz a tio n of W T l p ro te in s is n o t s e n s itiv e to
deoxyribonuclease, bu t ra ther to ribonuclease (Caricasoie e t al., 1996). Next,
the authors repo rt that W T l, bu t not EGR-1 proteins can b ind to specific IGF-II
(insulin-like g row th factor II) RNA sequences, em ploying zinc fingers for this
sequence-specific interaction. M utational analysis revealed tha t zinc finger 1
o f W T l, w hich has no coun terpart in EGR-1 protein, is m ore im p o rtan t for
th is RNA b in d in g activ ity . Therefore, a p o sttran sc rip tio n a l reg u la to ry
function for W T l was proposed. A study by Ye et al. (1996) dem onstra ted tha t
W T l can a lso b ind a n an tisense RNA, d esig n a ted W IT-1, w h ic h is
com plem entary to W Tl mRNA. Thus, W Tl m ay be ano ther p ro tein (besides
TFIIIA) to regu la te gene expression by b in d in g to b o th DNA a n d RNA.
Id en tif ic a tio n of o ther RN A b ind ing ta rg e ts for W T l aw aits fu r th e r
investigation .
2.1.5 T ranscriptional regulatory functions o f WTL
Since iden tifica tion of the W Tl D N A b ind ing site as a n E G R l-
consensus sequence, stud ies in to transcrip tional functions of W T l began ,
u sin g tran s ien t transfection assays in w h ich W Tl b o u n d to EGR-1 sites.
M adden e t al. (1991) first dem onstrated th a t W Tl expressed from a vector
u n d e r contro l o f a cytom egalovirus prom oter functioned as a rep resso r of
tran sc rip tio n w hen b o u n d to a CAT rep o rte r construct con ta in ing th ree
contiguous EGR-1 binding sites. It was also show n th a t D N A b in d in g w as
necessary, b u t no t sufficient for transcrip tional repression. U sing a se t of

66
tru n c a te d m u ta n t p ro te in s, the rep ress io n d o m ain w as m a p p e d to the
pro line- and g lutam ine-rich N -term inus of W Tl. In addition , w hen the W Tl
repression dom ain w as fused to the EGR-1 zinc finger dom ain , th is converted
EGR-1 from a transcrip tional activator to a repressor (M adden e t al., 1991).
Thus, a discrete, m odu lar character of the transcrip tional effector dom ain of
W Tl w as dem onstra ted , and a transcrip tional rep ression function w as first
p roposed for W Tl.
Subsequent studies further defined the transcrip tional effector dom ain
(M adden e t al., 1993; W ang e t a l., 1993b). M a d d e n e t al. (1993) had
dem onstra ted th a t W Tl contains a functionally transferable repressor dom ain
by fu sin g am ino acids 84-180 o f W T l to the GAL4 1-147 D N A -binding
dom ain . W ang e t al. (1993b) confirm ed that am ino acid region 84-189 was
su ffic ien t to co n fe r rep ression , w h ile am ino ac id s 180-294 en co d ed a
transcriptional activation dom ain th a t activated transcrip tion from the PDGF-
A p rom oter sites (Figure 2.2). T hus, W Tl p ro te in w as show n to contain
active rep ression an d activation dom ains. O ther tran scrip tio n factors that
have m odu lar transcrip tional effector dom ains are the Drosophila proteins
K ruppel, even-skipped, and engrailed (reviewed in Levine & M anley, 1989).
C uriously , M adden et al. (1993) found th a t the m inim al rep ressor
dom ain of W Tl (am ino acids 84-180) functioned in N IH 3T3 m ouse fibroblast
cells, b u t not in h u m an embryonic k idney 293 cells. To achieve repression in
293 cells, a dom ain consisting of am ino acids 1-298 w as required. Therefore,
in som e c ircum stances the tran scrip tio n a l ac tiv a tio n do m ain 180-294 is
needed to help the repression function of the 84-180 repressor dom ain . This
observation also suggests that the transcriptional rep ression function of W Tl
is co n tex t-d ep en d en t, a n d p robab ly involves in te rac tio n s w ith cell-type
specific pro tein co-factors. A poin t m utation a t position 201 of the repressor

67
dom ain had been described (Park e t al., 1993). This m utation resu lts in a
substitu tion of aspartic acid fo r glycine, and converts the function of W Tl
pro tein from a transcriptional repressor to an activator.
To date, a variety of n a tu ra l prom oters regu la ted by W T l have been
identified and analyzed. They include the prom oters of the follow ing genes:
insulin-like g ro w th factor II (D rum m ond et al., 1992), insulin-like grow th
factor I receptor (W erner e t a l., 1993; W erner e t al., 1994), p latelet-derived
g row th factor-A chain (W ang e t al., 1992; C ashier e t al., 1992; W ang et al.,
1993b; W ang e t al., 1995), transfo rm ing grow th fac to r-p i (Dey e t al., 1994),
colony-stim ulating factor-1 (H arring ton e t al., 1993), retinoic acid receptor-a
(Goodyer et al., 1995), PAX-2 (Ryan e t al., 1995), as well as the W T l prom oter
(R upprecht e t al., 1994). Ail o f these prom oters contain W Tl b ind ing sites,
a n d are rep re ssed by W T l w h e n assayed in tran s ien t tran sfec tio n
experim ents.
The find ing tha t IGF-II and PDGF-A are overexpressed in W ilms'
tum ors (Reeve e t al., 1985; Scott e t al., 1985; Fraizer e t al., 1987) suggested that
deregulated expression of g row th factor-encoding genes such as IGF-II a t a
critical time could contribute to W ilm s’ tum or developm ent. F u rther studies
of PDGF-A prom oter by W ang e t al. (1993b) had show n that W Tl can activate
or suppress transcrip tion , an d th a t separate dom ains of the p ro te in w ere
required for either transcriptional activation or suppression. The au thors also
fo u n d tha t W T l b ind ing s ites b o th upstream a n d dow nstream of the
transcrip tional s ta r t site are req u ired for the suppression function of W Tl,
w hereas activation only requ ires e ither an upstream or a dow nstream site
(W ang e t al., 1993b).
Prom oters such as IGF-II and IGF-I-R (D rum m ond et al., 1992; W em er
et al., 1994) have also been show n to require both upstream and dow nstream

68
b ind ing sites for rep ression by W Tl. H ow ever, o th e r p rom oters can be
rep re ssed from e ith er upstream or dow nstream sites, com plica ting the
m echanism of transcrip tional regulation by W Tl even further ( M ad d en et
al., 1991; Rackley et al., 1993; Madden e t al., 1993; R upprecht et al., 1994; H ew itt
et al., 1996). Furtherm ore, Hewitt and coworkers (1996) propose tha t m ultip le
W Tl b ind ing motifs are required for transcriptional repression by W T l, and
that the m ultip le b ind ing sites can be upstream of the transcrip tional start
site.
F u rther studies o f the W Tl repressor activity h ad defined the roles of
a lternative splice varian ts of W Tl. The study by R upprecht e t a l. (1994)
exam ined the repression abilities of all four W Tl splice isoforms, w h ich were
all capable o f repressing the W Tl prom oter. H ow ever, the splice varian t
contain ing b o th the 17-amino acid and KTS insertions (the m ost ab u n d an t
splice variant) was the strongest repressor, capable of 58.8-fold rep ression of
W Tl p rom oter, w hereas the isoform carrying the KTS insertion o n ly could
confer a w eak, 2.2-fold repression of the W Tl prom oter construct. In contrast,
the 17-am ino acid insertion had no ad d ed repressor function o n th e W Tl
protein lacking the KTS insertion. It appears that the 17-amino acid insertion
converts the W Tl into a powerful repressor. This find ing was confirm ed in
the s tu d y of PDGF-A prom oter by W ang et al. (1995). Furtherm ore, it was
show n th a t the 17 am ino acid insertion could confer repressor activ ity w hen
fused to e ither the zinc finger dom ain of W Tl or GAL-4. This rep ress ion
ability w as abolished w hen four consecutive serine residues w ere deleted
from the 17 am ino acid peptide. This indicates th a t the serine res id u es are
im portan t for the repressor activity of the isoforms carry ing the 17 am ino acid
insertion . To confer th e transcrip tional rep ress io n function , th e splice
variants w ith the 17 am ino acid peptide were bound upstream , dow nstream .

69
o r on bo th sides relative to the transcriptional start site. In contrast, the splice
variants w ithou t the 17 am ino a d d insertion could only repress transcrip tion
w hen they w ere b o u n d to both 5' a n d 3' sites; b ind ing o n either side alone
resulted in transcrip tional activation.
The m echanism of transcriptional repression by W T l is no t clear. If
W Tl is b o u n d upstream of the transcrip tiona l s ta rt s ite , perhaps it may
interfere w ith recru itm ent a n d /o r stab ility of the in itia tio n com plex. One
m ight env is ion th a t b in d in g of W T l p ro te in to the 3 ’ sites cou ld block
elongation of newly synthesized RN A molecules. H ow ever, it is no t know n
w hether W T l represses transcrip tion a t the level of e longation or initiation.
O n the o th er hand, the requ irem ent of sites both 3' a n d 5' relative to the
transcriptional start site could m ean th a t functional in teraction betw een W Tl
m olecules bound on bo th sites of the transcrip tional s ta r t is necessary for
repression. A sim ilar m echanism has been p roposed for the Drosophila
ev en -sk ip p ed p ro te in , acting o n th e Ultrabithorax p ro m o te r , w h ere
repression proceeds via a looping m echanism (TenHarm sel et al., 1993).
Recently, W Tl p ro te in was found to physically asso d a te w ith another
tum or-suppressor protein , p53 (M aherw aran e t al., 1993). This was show n by
c o im m u n o p re c ip i ta t io n an d c h e m ic a l c ro s s - l in k in g e x p e r im e n ts .
Transfection assays using EGR-1 prom oter and W Tl expression vector, b u t no
p53, dem onstra ted th a t W T l acted as a transcrip tional activator. H ow ever,
cotransfection of a p53 expression vector along w ith the W Tl, resu lted in
repression o f the sam e prom oter. T hus, the a ssoda tion of W Tl w ith other
proteins such as p53 can m odulate its regulatory activity. The first tw o zinc
fingers of W Tl were recently show n to be the site o f in teraction w ith p53
(M ahesw aran et al., 1995).

70
It sh o u ld be kep t in m ind , how ever, th a t p53 is know n to have
pleio tropic effects o n transcription, activating or repressing transcription from
a v arie ty o f p rom o te rs in e ither b ind ing s ite -d ep en d en t o r in d ep en d en t
fash ion (Z am betti an d Levine, 1993). The fact that transgenic mice w ith a
d is ru p te d p53 gene undergo norm al developm ent, a lb e it being p rone to
tu m o r fo rm ation (D onehow er e t al., 1992; Jacks e t al., 1994), w hile mice
lacking the in tact W T l gene (Kreidberg e t al., 1993) show prenatal lethality
an d o th er developm ental abnorm alities suggests that n o t all developm ental
functions o f W T l requ ire the presence of p53. It rem ains to be seen w hat
p ro te in factors in d ifferent cells determ ine w hether W T l w ill activate or
rep ress transcription.
Recently, severa l groups have p ro v id ed evidence th a t W Tl p ro te in
m ay associate w ith itself (Wang et al., 1995; R eddy et al., 1995; Moffett et al.,
1995). W ang e t al. first suggested that m u tan t W Tl pro teins m ay antagonize
the activ ity of the w ild-type protein. R eddy e t al. h ad m apped the self
association dom ain o f W Tl to the first 182 am ino acids of the protein. W hen
this dom ain w as expressed, or w hen tw o o ther proteins deficient for DNA-
b ind ing w ere expressed, they resulted in inhibition of transcrip tional activity
o f W T l (R eddy e t al., 1995). U sing an in vitro biochem ical assay and the in
vivo yeast tw o-hybrid assay W Tl w as found to self-associate. It w as thus
p roposed th a t W Tl m utants act in a dom inant-negative fashion, contributing
to the tum origenic developm ent. A sim ilar s tu d y by M offett e t al. (1995)
confirm ed the above findings, dem onstra ting bo th in vitro and in vivo that
W T l can self-associate. The authors used a D enys-Drash syndrom e allele to
d em o n stra te th a t it behaves in a dom inant-negative fash ion , and therefore
p ro p o sed a m echanism via w hich W Tl m uta tions m ay act in deregulating
cellu lar p ro liferation and differentiation.

71
D esp ite the accum ulated w ealth of stud ies on the m echan ism s of
transcrip tional regu la tion by W Tl proteins, there are still m any im resolved
problem s. A n u m b er of issues need to be ad d ressed in fu tu re studies,
including the identification of cellular genes that are actual targets for W Tl
regulation in bo th norm al and cancerous cells. It is n o t clear w h a t p ro tein co
factors m odu la te W Tl activity in particular cell types. It will be necessary to
identify m o re prom oters tha t are controlled by W T l proteins in o rd er to
understand the in vivo gene targets. This is com plicated by the heterogeneity
of the W T l b ind ing sites identified in vitro . M oreover, prom oters reported
to be regu la ted by W Tl proteins in transient cotransfection assays m ay not be
re levan t w h e n an a ly zed in vivo. C ellular p rom oters tha t req u ire W Tl
binding sites for tissue-specific expression in cell cu lture or in transgenic mice
need to be identified and analyzed. Finally, transcriptional regulation by WTl
proteins sh o u ld be stud ied in the context of native chrom atin struc tu re to
gain an in sigh t into gene regulatory events as they occur w ithin the nucleus.
The question rem ains as to the transcriptional effect of W T l: is it a t the
level of transcrip tional initiation, elongation, or both? Does W T l p lay a role
in p o s ttra n sc r ip tio n a l regu la tion , includ ing sp lic ing of n a sc e n t RNA
m olecules? W hat proteins of the basal transcriptional m achinery does W Tl
interact w ith? H ow is the expression of W Tl itself differentially regulated to
account fo r the tissue-specific and developm ent-specific expression of the
W Tl gene? A ll these questions need to be answ ered to p ro v id e an
u n d e rs ta n d in g o f the role of W Tl in both n o rm al d ev e lo p m en t and
neoplastic transform ation .

72
CHAPTER 3.0 IDENTIFICATION OF HIGH AFHNITY BINDING SITES FOR
THE WILMS’ TUMOUR SUPPRESSOR PROTEIN W Tl
3.1 Introduction
D espite the fact that bo th WT1[-KTS] and EGR-1 can bind to the same
DNA sequence, it is not clear w hat the preferred ta rg e t sequence for WT1[-
KTS] is, an d how the affinity and specificity of WT1[-KTS] for such sites w ould
com pare w ith com peting factors like EGR-1. Recent crystallographic studies
of a com plex form ed betw een the zinc finger dom ain of the EGR-1 protein
and a consensus DNA recognition sequence (Pavletich & Pabo, 1991; Elrod-
Erickson e t al., 1996) provided a structural basis for the observations that EGR-
1 and WT1[-KTS] recognize the same DNA sequence. These studies identified
those critical am ino acids w hich contact specific base pa irs in the consensus
EGR-1 b ind ing site GCG-TGG-GCG. These contact am ino acids are completely
conserved in the WT1[-KTS] protein, thus im plying a sim ilar role in the
binding of the last three zinc fingers of WT1[-KTS] to th e nine base pair EGR-1
recognition sequence (Figure 3.1 A ) .
The crystallographic data indicate that each of the three fingers of EGR-
1 interact w ith a specific 4 bp subsite w ithin the 9 base p a ir consensus binding
site (Figure 3.1B). The yeast M IGl repressor pro tein has tw o zinc fingers
co rrespond ing in sequence to fingers 2 and 3 of EGR-1. This p ro te in
recognizes a 6 base pair G rich sequence (Nehlin & R onne, 1990). This trend
predicts th a t WT1[-KTS] w ill recognize a specific 12 base pa ir sequence, the
first 9 base pairs of which w ill correspond to the EGR-1 consensus sequence.
The effects on W Tl-ZFP b ind ing of m utated sequences w ith in and flanking
an EGR-1 consensus site suggested that this zinc finger pep tide derived from
WT1[-KTS] recognizes a longer sequence (Rauscher et al., 1990; Morris e t al.,
1991).

» r c c c £ c * c c * r c a t c a t c * r c a t c a t c a t c * r c a c *c c « c e c c c c a t
73
• rc CAA CAT c c r r * r * r c ccA c c r c r c c c r c £ a c t a x c c c c & cr e r r s t a c c c r c c c c a r c r c a c a C C a c t c a c a a a c c c c c c r r c A rc11 * f l w t* r •r ^ • r t C * f • • • » f » t A r I Q • • f * 1 * w f C%w « A f « « r l y # « r ^
F I N G E R 1 CC 7 TA c CCA c c c r c c * A r a a c a c a t^ r x T T 'A A C ^ T c I r c c % C A C % r r A C A C ,A C C .C A C a c c a c c a a c c a c a c t c c t c a c a a a c c a t a c c a Ce rr* « U t y r A f * « t f c y # »#m l y * « c « c y r y « « * * l y i T i m J a « r > t a Z i v w # * r * r « t y * W c a r « l y t y * > r » t y r « 1 a
ANGER 2
ANGER:
ANGER 4
r c r CAC r r c a a c c a c r c r c a a o c a a c c r r r r r r j o c T j r c A c y * l y # A # y c y « « tw # r « # r « ##*# p * # | # y g l # # f
C A C jC A c CTC a a a H Z a I c a c c a a a c c ACA O A T ACA CC T c r c AAA OCA r r c c a c- - - » « I m t y | A f « | m # « I n « r « a c « A t * CAC # * y v # t t y t p c * p A * « 1 a
r c r AAA A C T — — rcT c a c c c a a a c c y « l f « t n r — c y # « t n « r « l y
c r r c r o c |c ë e |x c c |c A c |C A c |c r c • # A # • A r j A C l l — C | A # » | A t # i lp ««
AAC A C C CA C A C C ACC A C T A T ACA CC T CAA aaC C C C T T C A C C l y a C A C A t # CA T c u e ml* C A r « l y « t « i y « p c « p n « . * y
r e t c o s r c c o c a a c t r e t c a c « a a a a c r r r o o ec y # # r « c r p # e # # * r c y # « i n l y # l y # ptt* m l#
C A T CAA T TA C r C l Ô c C jC A X C A C AAC A T C O I T C A C ACA AAC A T S ACC AAA C T C «« A « I » l e w » # l | « c y A l « A l # ##m « m e A l # « 1 # # r « « a n m ac c n y l y # i# w
CAC CTC CCC e r r t c a t a c a c c t o b c a t c c « ! • t « « # 1 # I t t O T A A #0
BC C R -l
f l a g # »3 2 1I I I
n r iClnqecs
< 3 2 1 I I I IS ' -C O S s s e C O S -3 ' 5 -C O C TOG COG
Figure 3.1 DNA sequence of the peptide encoding insert in plasm id pET-
WTZFF, w ith the amino a d d sequence of the peptide. The region of sequence
underlined is deleted in p lasm id pET-W TlAFl, the pep tide product lacking
the first zinc finger of the W Tl protein. Am m o ad d s in fingers 2-4 that are
boxed correspond to the am ino ad d s of EGR-1 that are involved in specific
DNA recognition, and that in W Tl are proposed to play an analogous role.
A m no adds in finger 1 that are endosed w ith dashed boxes occupy positions
in the alpha helix that co u ld be invo lved in DNA interactions. (B)
D iagram m atic rep re sen ta tio n oof the z inc finger:3 bp DNA su b s ite
interactions in the EGR-l-DNA complex, and the proposed interaction of the
four W Tl zinc fingers with DNA.

74
To de term ine the role of the first zinc finger of WT1[-KTS] in D N A
binding, I u sed an in vitro b inding site selection assay (SAAB) (Blackwell &
W eintraub, 1990). From the crystal s tru c tu re data availab le for the EGR-
1:DNA com plex, it was reasonable to hypothesize that the first finger of WT1[-
KTS] should contact the 3-5 base pairs im m ediately dow nstream of the EGR-1
consensus s ite (Figure 3.IB). Therefore, an o ligonucleotide w ith flank ing
prim er sites a n d including the target sequence -GCGTGGGCGNNNNN- w as
synthesized an d used as a tem plate in the SAAB assay. A fter four cycles of
se le c tio n /a m p lif ic a tio n w ith e ither W T l-Z F P , a rec o m b in a n t p e p tid e
c o n ta in in g th e four zinc finger m otifs o f W T l, o r W T IA F I-Z F P , a
recom binant pep tide containing fingers 2-4 of W Tl, h igh affinity b inding sites
w ere cloned an d sequenced. The selected h igh affinity b ind ing sites for W T l-
ZFP had the sequence: GCG -TGG-GCG-(T/G)(A/T/G)(T/G)NN, w hereas the
W T IA F I-Z F P d id not d em o n stra te an y sequence p re fe ren ce a t th e
corresponding DNA binding subsite. These results indicate th a t the first zinc
finger of the WT1[-KTS] p ro tein is indeed required for the recognition of
additional specific DNA sequences contiguous w ith an EGR-1 binding site. A
q u an tita tiv e b in d in g assay w as used to dem onstra te th a t W T1-2TP has
app rox im ate ly a four fold h igher affin ity for the selec ted sites versus a
nonselected sequence. The ability of a consensus high affinity b inding site for
WT1[-KTS] to act as a regulatory sequence in m am m alian cells w as confirm ed
u sing a rep o rte r gene assay. DNA sequences in the feta l prom oter o f the
insu lin -like g ro w th factor U gene th a t confer W Tl responsiveness in a
transien t transfection assay b ind to the W Tl-ZFP w ith affin ities tha t v a ry
according to the num ber of consensus bases each sequence possesses in the
finger 1 subsite. All the experim ents in th is study were perfo rm ed by m yself,
w ith the exception of the oligonucleotide synthesis, w hich w as accom plished

75
by Dr. Paul J. Rom aniuk, and the transient transfection assays, w hich were
carried out by Kathleen C. Barilla.
3.2 M aterials and M ethods
3.2.1 Bacterial strains and DNA vectors
Plasm ids pET-W TlZFP and pET-WTlAFl contain the coding sequences
for the four zinc finger dom ain of W Tl and the last th ree zinc fingers of the
dom ain , respectively . The zinc finger cassettes w ere cloned in to the
N del/B am H l restriction sites of the T7 expression vector pET-16b (Studier et
al., 1990). The selected DNA sequences were c loned in to the B am H l and
EcoRI restriction sites of pUC19. All DNA vectors w ere m aintained in E.coli
s tra in JM109. Protein expression was carried out in E.coli strain BL21(DE3) (F-
ompT rb’mb")-
The m am m alian expression vector pCB6 + a n d its derivative pW Tl,
w h ich contains a complete cDNA encoding the W T l tum our suppresso r
p ro te in lacking either the 17 am ino acid insertion in the transcrip tional
regu la to ry d o m ain or the three am ino acid in se rtio n in the zinc finger
dom ain (W ang e t al., 1993), were generously p ro v id ed by Dr. T.F. Deuel.
P la s m id AMTV-CAT, w hich encodes a cDNA fo r ch lo ram phen ico l
acetyltransferase (CAT) under control of a m ouse m am m ary tu m o u r virus
prom oter (Thompson & Evans, 1989) was a kind gift from Dr. R. Evans.
3.2.2 O ther M aterials
Bacterial cell culture: yeast extract, tryptone and agar w ere purchased from
BDH Inc.
Chem icals: com m only used chemicals w ere purchased from Sigma Chemical
Inc., Fisher Scientific Com pany or BDH Inc.. Urea w as purchased from ICN

76
Pharm aceutical Inc.. Toluene w as purchased from VWR Scientific Inc.. 2, 5-
d iphenyl oxazole (PPO) w as purchased from Sigma Chemical Inc.
E nzym es: Restriction enzym es and T4 DNA ligase w ere purchased from
either New England Biolabs or Pharmacia. Taq DNA polym erase used in PCR
reactions was purchased from Promega.
N u c l e o t id e s : NTPs a n d dN T Ps w ere p u rch ased fro m P harm ac ia .
O ligonucleotides were syn thesized by Dr. R om aniuk u sin g an A pp lied
Biosystems Inc. 391 DNA synthesizer.
Iso topes: [a-32p] dATP (specific activity SOOOCi/mmol) w as purchased from
N ew England Nuclear (Du Pont).
O th e r: Agarose and low m elting tem perature agarose w ere purchased from
FMC Bioproducts. Gelatin w as purchased from Sigma. Poly (dl-dC) w as
pu rchased from Pharm acia. XAR-5 film w as pu rchased from K odak.
O ligonucleo tide purification cartridges w ere p u rch ased from A p p lied
Biosystems Inc.. Qiagen DNA purification colum ns w ith pre-packed anion-
exchange resin w ere purchased from Q iagen Inc.. M erm aid kit used for
recovering DNA from agarose gels was purchased from Bio 101 Inc.. His-Bind
resin and colum ns for p ro te in purification w ere purchased from N ovagen.
N itrocellulose filters were purchased from Millipore.
3.2.3 C onstruction and Purification of R ecom binant W Tl-ZFP and W TIAFI-
ZFP Proteins.
The synthetic genes encoding the four zinc finger dom ain (W Tl-ZFP)
an d the last th ree zinc fingers of the dom ain (W TIAFI-ZFP) o f WT1[-KTS]
w ere created by PCR, using cDNA prepared from hum an fetal k idney tissue
(Clontech) as a tem plate. For the construction of W Tl-ZFP, app rop ria te
p rim ers w ere syn thesized to allow for the am plification of a cassette

77
incorpora ting the WT1[-KTS] coding sequence from the un ique Aflin site to
the term ination codon (Figure 3.1A). PCR products w ere initially cloned by
b lun t-end ligation in to Sma I site of pUClS (Figure 3.2). The desired clones
w ere iden tified by DNA sequencing using a n ABI 373A au tom ated D N A
sequencer, and designated pUC-W TlZFP. A PCR cassette encoding only the
last three zinc fingers of WT1[-KTS] (WTIAFI-ZFP) w as generated using the
pU C -W TlZFP construct as a tem plate, the original dow nstream PCR prim er
a n d a n u p s t r e a m p r i m e r w i t h t h e s e q u e n c e
GCGCGCGGCCGAATTCATATGGAGAAA CCATACCAGTGT (Figure 3.1 A).
This PCR p ro d u c t w as cloned into Smal site of pU C lS an d the correct clone
identified by DNA sequencing using an ABI 373A autom ated DNA sequencer.
O nce the sequences w ere verified, the zinc finger cassettes w ere cloned in to
the N d e l/B am H I restriction sites of the T7 expression vector pET-16b (Studier
e t al., 1990) (Figure 3.3) to yield the plasmids pET-W TlZFP and pET-W TlAFl.
The resulting synthetic genes encoded zinc finger peptides w ith a histidine tag
and factor Xa cleavage site fused to the amino term inus (Figure 3.1 A).
Recom binant proteins were expressed by inducing log phase E.coli
(stra in BL21(DE3)) containing the pET-16b constructs w ith isopropyl-p-D -
th iogalac topyranoside (1 mM). The cells w ere harvested 3 hours la ter by
cen trifugation a t 8630 x g in a Beckman JA-14 rotor. The cell pellets w ere
w ashed w ith buffer A (10 mM TrisHCl, pH 7.5 a t 4 °C, 5 m M M gCli, 250 m M
N aC l, 10 m M PMSF, 10% glycerol, 5 mM DTT, 5 m M im idazole), th en
resu sp en d ed in the sam e buffer and lysed by u ltrasonication (8 X 15 second
pu lses on ice w ith 1.5 m inute intervals betw een pulses). Cell debris w as
pelleted by centrifugation in 15 ml corex tubes at 16,000 rp m for 15 m inutes in
a Beckman JA-21 rotor, and the supernatant w as discarded. The proteins w ere
extracted from the inclusion bodies by incubation overnight a t 4 °C in 10 m l

78
Ide I
WT1-ZF
p U C / W T
3152 bp
on
Figure 3.2 Schematic representation of the recom binant WTl-ZF plasmid,
pUC18/Wn-ZF. Restriction enzym e sites are indicated.

79
Aac IlcSâJg), Ssp f(SS20) ,
Sea 1(5196) Pvu 1(5086)
Pst 1(4961)
Bsa 1(4777)
EamllOS I(47i6)
EcoR 1(5709) ■Cla [(24) LHind“
Esp I(2S7)
A iw N 1(4239:
Sap 1(3707)
B stl 1071(3S94) Acc I(3S93) B îiA I(3S7S),
T th lll 1(3568)'
T7 transcription/ expression region
pET-16b(S T llbp )
-Bam H ((319). Xho 1(324) •Nde 1(331)
-Nco 1(397) -Xba K43I)
Bgl 11(497) SgrA 1(538)
Sph[(694) EcoN 1(754)
B pulO 1(2929)'
D id 11(942}
M lu 1(1219)
r Be! 1(1233)
B stE IKI400) -A pa 1(1430)
BssH 0(1630)
H paK i725)
PshAK2064)ra ilO S 1(2106)
X m a 01(2287) _ N cuI(2322) B%pM 10402)
'B o n 1(2707) >Bai 1(2794)
Figure 3.3 Plasmid m ap o f pET-16b expression vector (Novagen technical
bulletin). Protein coding sequences were cloned into Bam H l/Ndel sites.

80
of buffer B (buffer A containing 5M urea). Recom binant zinc finger peptides
were purified to hom ogeneity using a 1 m l nickel-chelate affinity colum n.
After w ashing the colum n w ith 2 ml o f buffer B containing 50 mM im idazole,
the recom binant p ro te in was eluted w ith 1 ml of buffer B containing 150 mM
im idazole follow ed by 1 ml of buffer B w ith 250 m M im idazole. Protein
concentrations w ere determ ined by the Bradford m ethod , using BSA as the
standard (Bradford, 1976), Protein yield averaged 10 m g per litre of bacterial
culture. O n SDS-PAGE (15%), the purified W Tl-ZFP m igrated w ith an
apparent m olecular m ass of 22 kilodaltons (kDa), and the W TIAFI-ZFP w ith
an apparen t m olecular mass of 17 kDa (Figure 3.4).
3.2.4 Selection A m plification and B inding (SAAB) Assay.
The SAAB assay was used according to the pub lished p rocedu re
(Blackwell & W eintraub, 1990) w ith several m inor m odifications (Figure 3.5).
A 69 bp oligonucleotide was synthesized that included the target sequence
-GCGTGGGCGNNNNN- flanked by Bam H l and EcoRl restriction sites and
by b ind ing sites for M13 universal forw ard and reverse sequencing prim ers
(Figure 3.6).
P rim er ex tension was used to convert the tem pla te oligonucleotide
into a m ixture of 1024 double stranded DNA sequences. The labeling reaction
for the initial round o f selection had a final volume of 40 pi, and consisted of:
10 pm ol o f SAAB tem plate, 80 pm ol R.U.P. (Reverse U niversal Erim er), NTB
buffer (50mM Tris-HCl, pH 7.2, 10 m M MgClz, 1 m M DTT, 50 m g /m l BSA).
First, the p rim er w as annealed by incubating the m ix ture at 65 °C for five
m inutes, then slow ly cooling to am bien t tem perature . The reaction w as
started by adding 8 nm ol each of dCTP, dGTP, d i IF , 20 pCi [a-32p]_^jATP and
10 units of Klenow firagment of E.coli DNA polym erase I. The reaction was

81
Figure 3.4 C oom assie b lue-sta ined 15% SD S-polyacrylam ide gel show ing
p u rifiied W T l-Z F P a n d W TIA FI-ZFP pro teins. Lane d e s ig n a ted M W
rep resen ts p ro te in m olecular w e ig h t m arker; L ane 1: W Tl-ZFP; Lane 2:
W TIAFI-ZFP.

82
Synthesize tem plate w ith random sequences in b inding site Synthesize opposite strand
NNN5' --------------------------------------------------------------------------- 3'
Incubate labeled double-stranded tem plate w ith protein
NNN5" --------------------------------------------------------------------------- S'S' --------------------------------------------------------------------------- 5'
N'N'N"+
ISeparate bound from free DNA by gel-shift
IIsolate b ound DNA and PCR-amplify
Label am plified selected b inding sites and re-select for binding as in steps 2-4
IBlunt en d clone into pUC,
Transform into E.coli
ISequence individual clones
Figure 3.5 Protocol for the SAAB (Selection and Am plification of Binding
site assay).

83
F.U.P. ^
5' GTTTTCCCAGTCACGACGAATTCTAATGCGTGGGCGE coR l
CCTTAGGATCCGTCATAGCTGTTTCCTG 3'B am H l ^
R.U.P.
Figure 3.6 The oligonucleotide containing a random ized target sequence for
SAAB analysis of WT1[-KTS] finger 1 subsite. Arrows represent the fo rw ard
(R.U.P.) and reverse (F.U.P.) universal prim ers. Endonuclease restriction sites
are underlined , the EGR-1 consensus sequence is show n in bold, an d the
random ized sequence is outlined.

84
allow ed to proceed for 45 m inutes, then the enzym e w as inactiva ted by
incubation at 65°C for five minutes. Following second strand synthesis, the
DNA w as labeled using [a-32p]-dATF, in the presence o f M13 universal
p rim ers éind the Klenow fragm ent of DNA polym erase I. The 40 p i labeling
reaction consisted o f the following: NTB buffer, 40 pm ol each of F.U.P.
( fo rw a rd U niversal Erim er) and R.U.P., lOpmol DNA tem plate, 40 pCi of [a-
32p]-dATP, 8 nmol each of dCTP, dTTP, dGTP, and 10 units K lenow fragm ent
of D N A polym erase I. The reaction was incubated for 30 m inu tes, and
stopped w ith the add ition of 8 nm ol dATP. The labeled DNA p roduct was
purified o n a 9% non-denaturing polyacrylam ide gel (29:1 crosslinking ratio
of ac ry lam id e to b is; 16 x 16 cm x 0.75 m m ), an d v isu a lize d by
autoradiography. Gel slices containing labeled DN A were excised, DNA was
elu ted overnight in 250 pi elution buffer (0.6 M NH^Ac, 1.0 m M EDTA, 0.1%
SDS) (Sam brook et al., 1989). The e lu ted DNA w as recovered by ethanol
precipitation, and resuspended in deionized water.
Labeled DNA (10-20,000 cpm, 0.005 pM) w as incubated w ith either
WT1-2IFP o r W TIAFI-ZFP for 20 m inutes at room tem perature in a buffer
containing 20 mM HEPES, pH 7.5, 70 m M NH4CI, 7 mM MgClz, 0.1% NP-40,
10 pM ZnCl2 , 2.5 m M DTT, 100 m g /m l BSA, 30 p g /m l poly dl-dC , 6 % glycerol.
B rom ophenol blue an d xylene cyanol w ere ad d ed and the reactions were
loaded im m ediately onto a non-denaturing 6% polyacrylam ide gel in a buffer
c o n ta in in g 89 m M T ris, 89 m M b o ra te , 2 m M EDTA. Follow ing
e lec trophoresis a t 200 V for 3 h a t 4 °C, the gels w ere sub jected to
au to rad iography (Figure 3.7). The band corresponding to the DN A -protein
complex w as cut ou t the gel, and the bound DNA in the gel slice w as eluted by
incubation overnight in 250 pi of 0.6 M am m onium acetate, 1 m M EDTA,
0.1% SDS a t 37 °C. The DNA was recovered by ethanol precipitation, and

85
A) 1 2 3 4 5 6 7 B) 1 2 3 4 5 6 7
I I I I I I I I I I I I I I
Bound
Loading wells
Figure 3.7 An autoradiogram m of round 3 of SAAB. (A) W Tl-ZF+DNA
template; (B) W TIAFI-ZFP+DNA tem plate. Lanes 1-7 represent p ro tein
concentrations 0; 1.56; 3.1; 6.25; 125; 25 and 50 nM, respectively.

8 6
resuspended in 40 pi of deionized water. Labeled DNA w as then p repared for
the nex t selection round using the e lu ted DNA as a tem p la te in a PCR
reaction (Blackwell & W eintraub, 1990). A standard PCR reaction h a d a
vo lum e of 100 p i, and contained the following: 10 mM Tris-H C l, p H 8.3 at
20°C, 1.5 m M MgCl2 , 25 mM KCl, 50 p g /m l gelatin, 10 pi selected tem plate
DNA, 20 nm ol each of dTTP, dCTP, dGTP, and dATP, 50 pm ol each F.U.P. and
R.U.P. p rim ers, and 2.5 un its Taq DNA polym erase. T he reaction w as
overla id w ith an equal volume of mineral oil, and carried o u t using a Techne
PCR therm ocycler. The reaction was carried ou t th ro u g h 25 ro u n d s of
th e rm a l cycling, each cycle consisting o f 1.5 m inu tes a t d é n a tu ra tio n
tem perature of 94°C, 1.5 m inutes a t an annealing tem perature of 55°C, a n d 1.5
m inu tes a t an extension tem perature of 72°C. The am plified DNA p ro d u c t
w as pu rified by phenolzCHClg extraction, an d ethanol p rec ip ita ted w ith the
add ition of glycogen as a carrier. The purified DNA was resuspended in 20 pi
o f deionized water.
In the first round of selection, the p ro te in concentrations were 25-100
nM ; in the next three rounds the concentrations were reduced to 6 , 3, an d 0.75
nM respectively. DNA isolated from the last round was am plified an d the
p ro d u c t d igested w ith Bam H l and EcoRl for cloning in to pUC19. A fter
tran sfo rm a tio n of E. coli JM109, th irty six to fifty clones w ere random ly
se lec ted , an d p lasm id DNA prepared . The base sequences of the EcoR
I/B a m H I inserts in these constructs were determ ined u sin g an ABI 373A
au tom ated DNA sequencer.
3.2.5 C onstruction of p lasm ids containing sequence e lem en ts o f the in su lin
lik e g row th factor II fetal p rom oter, and the non-selected sequence w ith the
CGC subsite for finger 1 b inding

87
The th ree W T l recognition sequences identified in th e insulin-like
grow th factor II (IGF-II)gene (D rum m ond et al., 1992), as well as a non-selected
sequence, GCG TGG GCG CCC were created de novo by Dr. R om aniuk using
a n A pp lied Biosystem s Inc. 391 DNA synthesizer. The to p s tra n d s of
oligonucleotides containing prom oter elements h ad the following sequences:
IGF-A: GAT GCG GTT GCG CGG GGG CGA CG
IGF-B: GAT GCG GTT GGG GGG GGG GGG GG
IGF-E: GAT GGA GGA GGG GGG GGG GGG GGG AAG G,
and incorporated a BamHL site for cloning into pUG19.
The top strand of the oligonucleotide w ith the non-selected sequence
had the sequence: AAT TGT AAT GGG TGG GGG CCC AGG GTT AG, and
in co rp o ra te d EcoRI and BamHl restriction sites for cloning in to pUG19
plasm id. The base sequences of the cloned inserts in these constructs were
determ ined using an ABI 373A autom ated DNA sequencer.
3.2.6 Purification o f oligonucleotides
Synthesized oligonucleotides were deprotected by incubation in 1.0 ml
of concentrated am m onium hydroxide for two hours at am bient tem perature.
The supernatan t was separated from the resin by centrifugation for 30 seconds
in a m icrocentrifuge, and incubated at 50®G overnight. The nex t day , the
so lu tion w as cooled prior to purification, and half of it was pu rified as per
protocol p ro v id ed by A pplied Biosystems Inc. The resulting e lu tan t was
evaporated in a RH40-11 Speed Vac concentrator, and the pellet resuspended
in 100 |il deionized water. O ligonucleotide concentrations w ere determ ined
by abso rbance a t 260 nm m ea su re d using DU-65 S pec tro p h o to m eter
(Beckman) (taking into account the OD^^O of a 10 M oligonucleotide solution).

88
3 ^ 7 End-labeling of DNA
A series of DNA sequences containing W Tl b in d in g sites were released
from p lasm id pUC19 using EcoRI an d H indU l a n d end-labeled w ith [a -
32p]dATP an d the Klenow fragm ent of E.coli DNA polym erase I (Sambrook et
al., 1989). The fill-in labeling reaction had a final vo lum e of 40 |i l , an d
included the following: 50mM Tris-HCl, p H 7.2, lOmM MgS0 4 , 0.1 m M DTT,
50 p g /m l BSA, 0.1 mM each dA TP, dTTP, dC TP an d dGTP, 25 pC i [a -
32P]dATP, an d 2.5 units of Klenow fragm ent of E.coli polym erase I. The
labeling reaction w as incubated a t am bient tem p era tu re for 30 m inu tes;
inactivated by incubation at 65® C for five m inutes, th en loaded onto a 9%
non-denatu ring polyacrylam ide gel (29:1 crosslinking ratio o f acrylam ide to
bis; 16 X 16 cm x 0.75 mm). Labeled DNA p ro d u c t w as v isualized by
autoradiography, the gel band excised, and the DNA w as e lu ted by incubation
overnight in 250 pi of 0.6 M am m onium acetate, 1 m M EDTA, 0.1% SDS. The
DNA w as recovered by ethanol precipitation, a n d resu sp en d ed in 40 p i of
deionized w ater.
3.2.8 N itrocellulose Filter B inding Assay
The binding affinities of a series of DNA sequences for the WT1[-KTS]
zinc finger peptides w ere quantified using a nitrocellulose filter binding assay
developed to study zinc finger protein-D N A in terac tions (Rom aniuk, 1985;
R om aniuk, 1990). The TMK binding reaction buffer contained 20 mM Tris-
HCl, 5m M MgClz, 100 m M K Q , 100 p g /m l BSA, Im M DTT, 10 pM ZnClz, and
was ad justed to pH 7.5 20°C. The proteins were serially d ilu ted in TMK buffer
to give fin a l concentrations ran g in g from 60 nM to 0.2 nM. P rio r to
incubation w ith DNA, the proteins w ere allowed to equilibrate in the b ind ing
buffer for 15 minutes. The binding reactions were in itia ted by the add ition of

89
approxim ately 10,000 cpm (approxim ately 0.005 pmol) of labeled DNA along
w ith the nonspecific com petitor poly (dl-dC) (Pharmacia) a t 30 ng per b inding
reaction. After incubation for an appropria te am ount o f tim e (m inim um 30
m inutes), a 180 p i a liquot w as rem oved, a n d vacuum filte red th rough pre
soaked 0.45 pm nitrocellulose filters (M illipore). The filters w ere d ried and
counted in toluene-based scintillant using a liquid scintillation counter (LKB).
Non-specific retention of labeled DNA on the filters was typically in the range
of 10-15 % of in p u t, and w as taken into account to norm alize m easurem ents
o f com plex fo rm atio n . E xperim en ta l d a ta (at le a s t 3 in d e p e n d e n t
determ inations per da ta po in t) w ere fitted using a non-linear least squares
function: m2 x m l x m O /(l + m l x mO); w here mO = varied ligand
concentration; m l = 1 0^, initially; and m 2 = 1 , initially (K aleidagraph version
3.0 Synergy Softw are, R ead ing , PA) d esig n ed for an A pp le M acin tosh
com pu ter. The d isso c ia tio n constan t (Kp) for a s im p le b im olecu lar
equilibrium (single com ponent pseudo first-order reaction) w as determ ined
as a concentration o f protein required to give 50 % saturation.
3.2.9 Transient T ransfection A ssays
The CAT re p o r te r p la sm id s w e re c o n s tru c te d by c lo n in g
oligonucleotides containing e ither one or th ree repeats o f the indicated W Tl
b ind ing motif into the un ique H indlQ site upstream of the transcrip tion start
o f the CAT gene in AMTV-CAT. HepG2 cells (ATCC HB8065) were cu ltu red
in m inim al essential m edia w ith Earle's salts, L-glutam ine and non-essential
am ino acids (Gibco BRL) w ith 1 mM sodium pyruvate a n d 10% fetal bovine
serum . Cotransfection of H epG 2 w ith 8 p g of either pW T l o r pCB6 +, 2 pg of
CAT reporter p lasm id and 3 p g of pSVP-gal (Prom ega) in 60 m m plates
u tilized a calcium phosphate protocol (Chen & Okayam a, 1987). Cell extracts

90
w ere p repared in IX R eporter Lysis Buffer (Promega) 48 h after transfection,
p-gaiactosidase activity w as m easured (Sam brook et al-, 1989) and w as then
used to norm alize the cell extracts for transfection efficiency before the
extracts w ere assayed for CAT activity. CAT activity was m easured by a non
chrom atograph ic techn ique u tilizing so lv en t p a rtitio n in g of the labeled
su b stra te ([^^C]-acetyl coenzym eA ) from the p roduct ([ l^ C ]-a c e ty la te d
chloram phenicol) (Sleigh, 1986).
2.3 Results
2.3.1 Isolation o f DNA b in d in g subsite for finger 1 of W Tl-ZF by SAAB.
The se lection /am plification assay (Blackwell & W eintraub, 1990) was
u sed to identify the p refe rred target sequences for the W Tl-ZFP pep tide ,
w h ich lacks the KTS in se rtio n betw een fingers 3 an d 4 in tro d u ced by
alternative splicing. Four cycles of SAAB analysis were carried ou t to select
h igh affinity binding sites from a low -degeneracy pool containing the target
sequence -G CG TG G G CG N N N N N - flanked by PCR p rim e r sites. This
oligonucleotide was syn thesized assum ing, based upon the crystal structure
d a ta available for the EGR-1: DNA com plex (Pavletich & Pabo, 1991), that
fingers 2-4 o f W Tl w o u ld in terac t w ith the EGR consensus sequence
GCGTGGGCG. By extension of the analogy to the EGR-1 protein, the first
finger of W Tl should contact the 3-4 bases im m ediately dow nstream of the
EGR consensus site, w h ich is the reg io n random ized in the tem pla te
oligonucleotide. The W TIAFI-ZFP p ro tein w hich lacks th e first zinc finger
w as used in a parallel selection procedure to serve as a negative control. After
the final round of selection and amplification, high affinity b inding sites from
the W Tl-ZFP selection, as w ell as sequences firom the W TIAFI-ZFP selection,
w ere cloned into pUC-19 an d 36 or more clones of each w ere sequenced. The

91
1 4 7 10 14 I I I I I
SAAB T e m p la t e : . .GCGTGGGCGNNNNN. .
o f b a s e s 1 0 - 1 4 o fGAGCG TAACA
TAACCGANCT
TACACGCACC TACAT
TACCCGCTCT
TAGCAGGGCA TAGGGGGGCG TAGTAGGGGT TAGTGGGGTGGGGTG TATCT
TATGCGGTAG TATGTGGTAT TATNNGGTGAGGTGC TCCAAGGTGG TCCCAGGTTG
TGACCGTAAA TGACCGTACC
TGCTAGTTGA
TGGACTGTAC TGGGATGTAC TGGGGTGTCA TGGGGTGTCATGTCT TTTAC
TTTCCTTGACTTGGATTGGC
Figure 3.8 Sequences of bases 10-14 arising from the random SAAB tem plate after four rounds o f selection w ith W Tl-ZFP. Fifty clones were isolated after lig a tio n into p lasm id pU C lS an d sequenced by s tan d ard m ethods. The sequences have been so rted accord ing to their iden tity at bases 1 0 -1 2 . N ucleo tides sh o w n as "N" cou ld no t be determ ined unam biguously , and these w ere no t used in calculating the d istribu tion frequencies show n in Table 3.1.

92
T ab le 3.1 Frequencies of N ucleotides Selected by W Tl-ZFP an d W TIAFI-ZFP
W Tl-ZFPa
Position; 1 0 11 12 13 14
A 0 .0 (0 .0 ) 30.0 (13.9) 14.0 (11.1) 24.0 (9.8) 30.0 (43.0)
C 0 .0 (0 .0 ) 8 .0 (0 .0 ) 12.0 (13.9) 38.0 (16.1) 34.0 (21.4)
G 36.0 (55.6) 46.0 (55.6) 34.0 (38.9) 26.0 (41.9) 22.0 (17.9)
T 64.0 (44.4) 16.0 (30.5) 38.0 (36.1) 12.0 (32.2) 14.0 (17.9)
W TlAFl-ZFPb
Position: 1 0 11 12 13 14
A 15.2 24.2 6.5 19.3 32.3
C 27.3 33.3 22.5 25.9 19.4
G 27.3 24.2 38.7 38.7 25.8
T 30.3 18.2 32.2 16.1 2 2 .6
^Com piled from the sequences of 50 clones chosen random ly after the final
selection round . N um bers in brackets com piled from the sequences of 36
clones ob tained from the fina l selection ro u n d of a second , in d ep en d en t
SAAB experim ent.
^C om piled from the sequences of 36 clones chosen random ly after the final
selection round.

93
sequences of 50 clones chosen from the final pool of DNAs selected by WTL-
ZFP are show n in F igure 3.8. The percentage frequencies of the four bases at
each o f the five random ized positions are show n in Table 3.1 for bo th W Tl-
ZFP a n d W TIAFI-ZFP. A chi-square test o f the base d istribu tion a t each
position indicated tha t only the distributions a t positions 1 0 , 11 and 1 2 in the
W Tl-ZTP selection experim ents were non-random (a<0.025), while all o ther
d istribu tions can be considered random (a>0.05). The h igh affinity b ind ing
site for W Tl-ZFP had th e sequence: GCG-TGG-GCG-(T/G)(A/G)(T/G)NN. In
contrast, W TIAFI-ZFP, th a t had a deletion o f the first zinc finger, show ed no
p reference a t all for D N A sequences d eriv ed from the five d eg en era te
positions. The reproduciblity of the SAAB results was tested by repeating the
se lec tion o f sequences w ith the W Tl-ZFP. The resu lts ob ta ined w ere
essen tia lly the sam e (Table 3.1), w ith a s ligh t difference in the preference
dem onstra ted for position 11 (G /T rather than A /G ). It is interesting to note
tha t only a three base p a ir sequence was selected for the W Tl-ZFP finger 1
b ind ing subsite , w hile 4 bp recognition subsites are observed for EGR-1
pro tein .
3.3.2 Q uan tita tive B ind ing of W Tl-ZFP to V arious DNA Sequences
A nitrocellu lose filter b ind ing assay (Rom aniuk, 1990) w as u sed to
determ ine the affinities of the W Tl-ZFP for several high affinity b ind ing sites
selected in the SAAB assay. A non-selected sequence, GCGTGGGCGCCC, was
also tested for b inding to W Tl-ZFP. Figure 3.9A shows typical b inding curves
m easured for W Tl recognition sequences containing TGT and CCC finger 1
subsites, a n d for an unrela ted sequence, a thyroid horm one receptor response
elem ent (TRE). The values of the apparent association constants (Kg) for the
b ind ing o f W Tl-ZFP to these DNA sequences can be derived from the data

94
assu m in g a s im p le b im olecular in te rac tion . T he resu lts o f severa l
Independen t experim ents indicate that the W Tl elem ent w ith the consensus
TGT finger 1 subsite binds WTl-ZFP w ith an apparen t Ka of 8.40±1.21 X 10®
w hile the nonconsensus CCC elem ent has an apparen t Ka of 2.10±0.41 X
10® M"^. In co m p ariso n , the TRE sequence b inds W T l-Z F P w ith
approxim ately tw o orders of magnitude less affinity (Ka estim ated to be ca. 3 X
1 0 ® M'^). Ka values for a num ber of the selected h igh affinity b in d in g sites
were m easured using the same assay, and these sequences have affinities for
W Tl-ZFP th a t are equal to that of the TGT sequence (Table 3.2). In
co m p ariso n , the W T l elem ent con ta in ing the CCC finger 1 subsite
significantly reduced the binding of the W Tl-ZFP, decreasing the affinity
four-fold com pared to the selected, h igh affinity b ind ing sites. The affinities
of the TGT elem ent (Ka = 1.67+0.05 X 10® M'^) and the CCC elem ent (Ka =
0.92±0.09 X 10® M‘1) for the m utant peptide W TIDFI-ZFP, w hich lacks the
first zinc finger of W T l, are almost equivalent (Figure 3.9B). It is apparen t
that the difference in affinities of selected vs. non-selected sequences for W Tl-
ZFP is a lm ost com pletely attributable to the interaction of finger 1 w ith the
base 10-12 subsite defined by the SAAB experiments (Table 3.2). These data
give a clear indication of the contribution m ade by finger 1 of the W Tl-ZFP to
the overall free energy of DNA binding. These results also indicate th a t the
SAAB assay easily discrim inates between elements tha t differ in their binding
affinity for a specific protein by as little as four-fold.
W T l binding sites have been identified in a num ber of eukaryotic
prom oters (D rum m ond e t al., 1990; Gashler et al., 1992; W ang et al., 1992;
H arrington e t al., 1993; Rupprecht et al., 1994; Dey et al., 1994). To determ ine
the biological significance of the WTl recognition sequences selected in the

95
T 3C3O
n
<zoco
0.0lO**
/(oc3oÛ<Zoco
0.6
0 .4
uCB
0.0 10'10*10[WT1AF1-ZFP1, M
1 0 "
Figure 3.9 (A) Results of an nitrocellulose filter b inding assay m easuring the
equilibrium b ind ing of WTl-ZFP to the WTl elem ent w ith a TGT subsite for
finger 1 (closed circles), the WTl elem ent w ith a CCC subsite for finger 1
(open squares) or a thyroid hormone receptor elem ent (open circles). Curves
rep resen t the best fit of the data in each case to a sim ple b im olecular
equilibrium . (B) Results of an nitrocellulose filter b ind ing assay m easuring
the equilib rium b ind ing of WTIAFI-ZFF to the WTl elem en t w ith a TGT
subsite for finger 1 (closed circles) and the WTl elem ent w ith a CCC subsite
for finger 1 (open squares).

96
T able 3.2 Relative Affinities of W T l Binding Sites for W Tl-ZFP and
W TIAFI-ZFP
D N A Sequence^ Relative Affinity for
W Tl-ZFPb
Relative A ffinity for
W TlA Fl-ZFPb
TGT 1.00 1 .00
CCC 0.22±0.04 0.56±0.07
GGG 0.98±0.26 n.d.
TAG 0.98+0.37 n.d.
GGT 0.81±0.30 n.d.
^Sequence of bases 10-12 (finger 1 subsite) are show n
^Determ ined as th e ratio of the apparen t Ka m easured for each b ind ing site to
tha t m easured for the TGT binding site in parallel. M ean ± s tan d ard deviation
reported for tw o o r m ore independent determ inations, n.d. = n o t determ ined.

97
SAAB assay, w e com pared the b inding affin ity of W T l-Z FP for three sites
id e n tif ie d in th e p rom oter o f the in su lin -lik e g ro w th factor II g ene
(D rum m ond e t al., 1992), w ith the affinity of the protein for the high affinity
TGT target sequence identified by SAAB analysis. As the results in Figure 3.10
sh o w , the three W T l sites isolated from the IGF-I I p ro m o te r act as h ig h
affin ity target sequences for the W Tl-ZFP, w ith affinities roughly equal to th a t
of the idealized TGT element. In relative term s, the th ree elem ents have
affin ities for W Tl-ZFP in the o rder (from h ighest to low est) of IGF-B^GF-
E>IGF-A (relative affinités being 1.00, 0.77±0.22 and 0.28±0.08 respectively).
T his correlates w ell w ith the SAAB results; IGF-B has bases 10 and 11 th a t
conform to the SAAB consensus, IGF-E has only 1 consensus base but it is in
the critical 10 position, and IGF-A has only 1 consensus base b u t it is in the
less critical 11 position. In sum m ary, our da ta dem onstrated th a t WT1[-KTS]
b in d s to a 12 bp DNA sequence w ith h igh affinity, a n d th a t the relative
affin ities of W Tl binding sites in genomic DNA can be understood based
u p o n this consensus binding sequence.
3.3.3 A n In v itro Selected W Tl B inding Site Acts as a S trong T ranscriptional
R egulator.
It has been dem onstrated that W Tl can activate, or repress,
transcrip tion from a prom oter depending up o n the num ber and location of
W T l binding sites (M adden et al., 1991; W ang e t al., 1993; W erner e t al., 1993).
To dem onstrate tha t a high affinity W Tl b ind ing site selected in vitro can act
as a transcriptional regulator in vivo, we tested a series of reporte r constructs
in w h ich the chloramphenicol acetyltransferase (CAT) gene w as fused to the
m ouse m am m ary tum or virus LTR prom oter containing a variable num ber
of th e TGT sites cloned upstream of the transcription start site. This reporter

98
c o n s e n s u s
r e f e r e n c e : IGF-A s i c e ; IGF-B s i c e ; IGF-E s i c e ;
GAGGCG-TGG-GCG-TGT
T
GCG-TGG-GCG-TGT GCG-CGG-GGG-cGa GCG-GGG-GCG-GGc GCG-GGG-GCG-Gcc
o3Oa
<ZOcoo«9
0 010* 10 '10
[WTl-ZFl. M
Figure 3.10 (A) Sequences of WTl elem ents identified in the fetal prom oter
of the IGF-II gene. (B) Results of a n nitrocellu lose filte r b ind ing assay
m easuring the equilibrium binding of W Tl-ZFP to the W T l elem ent w ith a
TGT subsite for finger 1 (closed cirdes), the IGF-A elem ent (open squares), the
IGF-B elem ent (closed squares) or the IGF-E elem ent (open circles). C urves
rep resen t the best fit of the data in each case to a sim p le b im olecular
equilibrium .

99
gene construct has been used to dem onstrate the gene regulatory function of
thyro id horm one reponsive elements cloned into the unique H ind HI site
(T hom pson & Evans, 1989). Similar tests of the regulatory activ ity of other
W T l b in d in g sites have utilized an analogous tk-CA T re p o rte r system
(M adden e t al., 1991). The MTV-CAT reporter constructs were co-transfected
in to H epG 2 cells w ith a eukaryotic expression vector containing the hum an
W T l cDNA, or the expression vector w ithout an insert. After 48 hours, cell
extracts w ere p repared an d after norm alization for transfection efficiency,
w ere assayed for CAT activity. As the results in Figure 3.11 show , insertion of
a single TGT site in the MTV prom oter gives rise to a significant stim ulation
of transcrip tion by W Tl com pared to basal expression. Insertion of three
tandem TGT sites gives a m uch larger responsiveness to W Tl. This increased
responsiveness is p rim arily the result of a sign ifican t decrease in basal
p rom oter activity w ith the insertion of three tandem sites, com pared to the
basal prom oter activity w ith the insertion of one site.
A num ber of the putative W Tl binding sites iden tified in several
prom oters do not conform to the consensus site we have identified by the
SAAB assay. To confirm that such sites also act as transcriptional regulators
responsive to W Tl in this system, we cloned either one or three copies of the
low er affinity CCC site in to the MTV LTR prom oter. As the results in Figure
3.11 show , a single CCC site has approxim ately the sam e effect o n prom oter
activity as a single TGT site has. In contrast, three tandem CCC sites provide
approxim ately twice the responsiveness to W Tl than tha t observed for three
tandem TGT sites. In th is case, the increased responsiveness observed for
three vs. one CCC site is directly the result of a decrease in the basal activity of
the prom oter: the level of CAT activity expressed as the result of W T l

100
20000,%u:vLlîi
jccc
15000
Q. 10000
4MTV-CAT CCC TOT 3CCC 3TGT
(I.7JC) (2.SXJ (2.«X) (171 X) ( 7 J Z X )
Reporter (fold activation)
Figure 3.11 (A) CAT reporter gene constructs used in transien t transfection
assays to m easure W Tl responsiveness. Boxes represent the num ber o f W Tl
elem ents w ith the indicated finger 1 subsite sequence inserted into the unique
H ind m site of the DMTV-CAT p lasm id . (B) A ssay of ch loram phenicol
acetyltransferase activity in cell extracts p repared from transient transfections.
Each transfection of HepG2 cells included the indicated reporter p lasm id , and
either the eukaryotic expression p lasm id pCB6 + which does not express W Tl
(open bars), or pW Tl (closed bars), a pCB6 + derivative th a t does express W Tl.
C pm va lues are reported as the m ean ± standard deviation from 5 o r more
in d ep e n d en t experim ents. Fold activa tion is reported as the ra tio o f cpm
ob tained in transfections w ith pW T l to the cpm ob tained in transfections
w ith pCB6 +.

101
activation is the sam e w hether the MTV prom oter contains one or th ree CCC
sites.
3.4 D iscussion
The DN A b ind ing dom ain of WT1[-KTS] consists of four zinc fingers,
the last th ree o f w hich have significant hom ology to the three zinc fingers of
EGR-1. M olecular details of the interaction of the zinc fingers of EGR-1 w ith a
consensus b inding site have been resolved by X-ray crystallography (Pavletich
& Pabo, 1991; E lrod-Erickson et al., 1996), and are discussed in detail in
C hapter 1 of this thesis. The three hom ologous zinc fingers found in WT1 [-
KTS] re ta in the specific am ino acids of EGR-1 tha t are involved in DNA
binding. The first, un ique zinc finger of WT1[-KTS] m ight be expected by
analogy to in teract w ith a subsite contiguous to the EGR-1 consensus site,
im plying tha t WT1[-KTS] recognizes a 1 2 (or longer) base pair sequence in
target prom oters.
In add ition , the structures of tw o other zinc fingerzDNA com plexes
have been solved by X-ray crystallography: the SWIzDNA complex (Fairall et
al., 1993) an d the GLIzDNA complex (Pavletich & Pabo, 1993). The results of
these investigations dem onstra ted th a t zinc finger interactions w ith double
stranded D N A are n o t restricted to one specific s tran d , nor to contiguous
three base p a ir subsites. Interactions can span bo th strands of the DNA,
subsites can be larger than 3 base pairs and can be overlapping, and som e zinc
fingers m ight not in teract w ith the D N A at all.
G iven this variability in the w ays by which zinc fingers can interact
w ith DNA, w e designed an experim ent to determ ine the full b ind ing site for
WT1[-KTS] th a t w o u ld favour the m o st likely hypothesis (m echanism of
interaction identical to EGR-1) w ithou t preventing the identification of other

102
m odes of b ind ing . The tem plate for the selection of high affin ity sites w as
d esig n ed to hav e a ran d o m sequence of five base p a irs im m ed ia te ly
dow nstream of the nine base pair EGR-1 consensus site. Based upon the
antiparallel b in d in g of EGR-1 to its nine base p a ir consensus site, the first
finger of WT1[-KTS] w ould be expected to interact w ith a contiguous subsite
im m ediately dow nstream . To verify that any sequences selected from this
tem plate by the complete WT1[-KTS] zinc finger dom ain w ere the result of
the specific in teraction o f finger 1 w ith the DNA, the selection assay w as
carried out in parallel w ith a zinc finger peptide containing only fingers 2 to 4
o f WT1[-KTS].
The re su lts of this experim ent indicate th a t the first z inc finger of
WT1[-KTS], like the three fingers homologous to the EGR-1 p ro tein , binds to
a three base p a ir subsite w ith in a 12 base pair consensus binding site for WT1[-
KTS]. This subsite has the consensus sequence (T o r G),(G or A or T), (G or T).
If the first zinc finger of WT1[-KTS] interacts specifically w ith the sam e strand
o f DNA as the o ther zinc fingers, it w ould appear firom the selected consensus
sequence that th is finger interacts preferentially w ith keto substituen ts in the
m ajor groove of the DNA (Figure 3.12). The absolute d iscrim ination for G o r
T in the first position (base 10) suggests that this nucleotide m ay form the
strongest in teraction w ith an amino acid side chain w ith in the a-helix of the
firs t zinc finger. Selection at the o ther two positions was som ew hat less
stringent (and varied from experim ent to experim ent for base 1 1 ), and so it is
no t clear exactly w hat role these two positions m igh t play in p ro te in binding:
they could p rov ide specific DN A -protein contacts, o r they m ig h t provide a
D N A sequence context th a t optim izes the in teraction of finger 1 w ith the
D N A .
While this work w as in progress, a report w as published describing the

103
♦ ♦ IM j | i ) r iircKivc
N406M7
NÿNI NIN9
N3 02N2
♦ I ♦ ON6 04
C5N7
N3NI NIN9
02
♦ ♦Minor groove
Figure 3.12 The hydrogen-bond donors and acceptors p resen ted by W atson-
Crick base pairs to the major groove and the m inor groove (Steitz, 1990).
Two inverted triang les represen t hydrogen bo n d donors; d iam ond-shaped
sym bols represent hydrogen bond acceptors; a circle - a m ethyl g roup.

104
re su lts of a SAAB experim en t co n d u c ted w ith a reco m b in an t p e p tid e
encom passing fingers 1-3 of WT1[-KTS] (WT1AF4-ZFP) (D rum m ond e t al.,
1994). A pparen tly selection experim ents conducted by this g ro u p w ith the
fu ll fou r finger d o m ain d id not re su lt in the se lection of a n y specific
sequences, m ost probably because of the h igh protein concentrations (>5 pM)
u se d in the assay. H ow ever, w hen the experim ent w as repeated w ith the
W T1AF4-ZFP p ro te in , a final selection round at 10 pM p ep tid e y ie ld ed a
s tro n g selection o f a three base pair subsite w ith a GT(C or T) consensus
sequence. My d a ta show s a som ew hat less stringent sequence w as selected at
approxim ately 1 0 ,0 0 0 -fold lower concentrations of a full four finger peptide.
N evertheless, th e sequence selected w ith the th ree finger W T 1A F4-Z FP
p e p tid e is a m em ber of the consensus sequence w e identified w ith th e full
zinc finger dom ain of WT1[-KTS].
W hile the SAAB experim ent is useful for iden tify ing h ig h affinity
b in d in g sites for a DN A binding pro tein , it provides little in sig h t in to the
re la tiv e affin ities th a t consensus sequence sites have for the p ro te in in
com parison to nonconsensus sequences. We used a n itrocellu lose filter
b in d in g assay to quantify the affinities th a t W Tl-ZFP and W TIA FI-ZFP had
for a num ber o f the selected sequences, an d the non-selected sequence GCG-
TGG-GCG-CCC. Four of the selected sequences chosen at random h a d equal
affinities for W Tl-ZFP, while the non-selected CCC elem ent h ad a four-fold
w eaker affinity. In com parison, the consensus TGT sequence a n d th e non-
se lec ted CCC e lem en t had roughly equal affinities for the W T IA F I-Z F P .
T herefore the ta rg e t sites identified by the SAAB experim ent fo r W T l-Z FP
h av e 4 fold h igher affinities for the p ro te in than a non-selected site as the
resu lt of specific interaction of the first zinc finger w ith DNA.

105
In a d d itio n to p rovid ing in form ation o n the re la tive affin ities of
se lec ted vs. non-selected sequences for W Tl-ZFP, the n itrocellu lose filter
b in d in g assay also provides a m easure of the apparen t association constant for
the equilibrium b in d in g of this pep tide to DNA. A previous s tu d y reported
the equ ilib rium constan ts m easured by a gel m obility sh ift assay for the
b in d in g of a W Tl zinc finger peptide to a 12 base pair W Tl selected sequence
to be 7.1 X 10^ M ‘i vs. a value of 4.5 X 10^ M"^ for a non-selected sequence
includ ing the 9 base pair EGR-1 consensus site (D rum m ond e t al., 1994). The
zinc finger pep tide W Tl-ZFP that w e have constructed b inds to a selected
D N A sequence w ith a Ka value m easured by a nitrocellulose filter binding
assay o f 8.40±1.21 X 10^ M'^ vs. 2.10±0.41 X 10* M 'l for a non-selected sequence.
In the gel m obility shift assay that w as used as p a rt of the SAAB experim ent,
w e observed strong binding of 0.75 nM W Tl-ZFP to selected DN A sequences
in ag reem ent w ith the affinities m easured by the filter b ind ing assay. The
va lues for equ ilib rium constants th a t we m easure are about tw o o rders of
m ag n itu d e h ig h er th an those rep o rted earlie r, an d consisten t w ith the
rep o rted equilibrium binding constant of 1.7 X 10* M‘l for the in teraction of
the zinc finger dom ain of EGR- Iw ith its consensus DNA sequence (Pavletich
& Pabo , 1991). T hus the zinc finger dom ain of W T l binds w ith h ig h affinity
to a tw elve base p a ir consensus sequence.
Putative W T l binding sites have been identified in a num ber of native
prom oters, and it has been dem onstrated by transien t transfection assays that
these p rom oters can be regulated by WT1[-KTS] (D rum m ond e t al., 1990;
G ashler et al., 1992; W ang et al., 1992; H arrington et al., 1993; R upprech t et al.,
1994; Dey et al., 1994). However, m any of these putative W T l recognition
sites d o not conform completely to the finger 1 subsite that w e have identified
by the in vitro SAAB assay. To determ ine w hether these n a tu ra lly occuring

106
sites b in d W T l w ith h igh affinity, w e m easured th e affinities o f three W Tl
sites from the fetal prom oter of the IGF-II gene fo r W Tl-ZFP. These three
sites h ad affinities for W Tl-ZFP th a t were s im ila r to the affin ity of the
consensus TGT selected site. H ow ever, the th ree IGF sites w ere not
e q u iv a le n t in the ir affin ities for W Tl-ZFP, b u t varied in affin ity in
relationship to their divergence from the finger 1 subsite consensus sequence.
These results link the in vitro SAAB experim ent to biologically re levan t sites
in genom ic DNA, and again dem onstrate the con tribu tion of the first zinc
finger of W T l to the interaction of the protein w ith DNA.
W T l acts as a transcriptional regulator in vivo, and n a tu ra l selection
will have designed W Tl recognition sites based bo th o n DNA b ind ing affinity
and th is regulatory function in a w ay that the in vitro SAAB assay is unable
to. T o p robe this relationship, we com pared the regu la to ry action of the
selected, h igh affinity TGT site w ith the regulatory action of the non-selected,
low er affinity CCC site in the MTV-CAT reporter gene system. In a single
copy, bo th sites confer a sim ilar positive regulatory response of the MTV LTR
prom oter in this reporter to W Tl. This result is consistent w ith reports that
the W T l p ro tein acts as a transcriptional activator o n prom oters th a t contain
b ind ing sites located exclusively upstream or dow nstream of the transcription
start site (W ang e t al., 1993). H ow ever, w hen the sites are p resen t in three
copies, it becom es apparen t that the low er affinity CCC site confers greater
responsiveness to W Tl than does the h igh affinity TGT site. This resu lt is
in trigu ing because it suggests that the strength of th e interaction of finger 1
w ith D N A m ay in some w ay m odulate the ability o f the p ro tein to regulate
transcription. In addition, the m ultiple copies of e ither W Tl site significantly
d am p en the basal level o f transcrip tion from the MTV prom oter, another

107
effect th a t m ay be active in natural selection. F u rther investigation w ill be
requ ired to completely understand these phenom ena.
In sum m ary, our da ta have dem onstrated th a t WT1[-KTS] binds to a 12
base pair DNA sequence w ith high affinity, and th a t the relative affinities of
W T l b ind ing sites in genom ic DNA can be u n d ers to o d based u p o n th is
consensus binding sequence. Investigation of the specific na ture o f am ino
ac id -D N A in terac tions of finger 1 o f W Tl w ill en h an ce fu r th e r o u r
understand ing of the structure and function of this tum or suppressor protein.

108
CHAPTER 4.0 W Tl MUTATIONS A N D THE DENYS-DRASH SYNDROME
4.1 Introduction
D enys-D rash syndrom e (DDS) is a rare hum an disease. Its sym ptom s
include W ilm s' tum or, genital anom alies an d n e p h ro p a th y (D rash e t ai.,
1970). H eterozygous W Tl po in t m utations have been identified in over 95 %
of DDS pa tien ts , pointing to a crucial ro le W Tl p lays in the genesis of the
D enys-D rash syndrom e (Pelletier et al., 1991). The W T l m utations observed
in DDS pa tien ts can be classified into four categories: (1) m issense m utations
of D N A -bind ing amino acids - the m ost com m on g ro u p of m utations; (2)
substitu tion o f zinc binding amino a d d s ; (3) nonsense m utations lead ing to a
fram eshift a n d rem oval of entire zinc fingers, a n d (4) early term ination .
These m utations are sum m arized in Figure 4.1 A.
The m ost common m utations affect the key am ino a d d s p roposed by
analogy to EGR-1 to contact DNA. A rginine 394 appears to be a hot spo t of the
missense m utations reported in 19 of 38 cases (50 %) of DDS patients (Pelletier
e t al., 1991; C oppes et al., 1992; Bruening e t al., 1992; Baird e t al., 1992; Sakai et
al., 1993; A kasaka et al., 1993; Baird & Cow ell, 1993; N ordenskjo ld et al., 1994).
This m u ta tio n is located in exon 9, w hich encodes zinc finger 3 of the W Tl
pro tein (Figure 4.IB). O ther m utations identified in exon 9 involve asparta te
396 substitu tions (reported in 6 individuals) (Bruening e t al., 1992; Baird e t a.,
1992b; N ordenckjold et al., 1994; Pelletier e t éd., 1991; Little e t al., 1993), as well
as fram e sh ift (at position 386; Ogawa e t al., 1993), an d truncation m utations
(at position 390; Little et al., 1992; Baird e t al., 1992a; Q uek e t al., 1993; Varanasi
e t al., 1994). In addition, a m utation in terfering w ith sp licing at exon 9 was
iso lated (H uff e t al., 1991). M utations m app ing to exon 8 (zinc finger 2)
involve arg in ine 366 (Henry e t al., 1989; Bruening e t al., 1992), h istid ine 373
(Little et al., 1993; Sharma et al., 1994), and histidine 377 (Pelletier et al., 1991).

109
B
M ut4Iion
I «3 C > Y
I tX % , Y
l '. î r . Y
lOO C , Y
< « )C > G
161 fratnethifi
162 R- > Mop
165 S > F
166 R - > H
166 R >C
121 H > Q
171 H > Y
177 H > Y
177 H > R
184 rnmcthift
186 fnincshift
190 R- > Mop
194 R. > W
1 9 4 R -> P
196 0 - > N
1 96D -> G
432 fnmeduTi
N o
(™)
(rail
Y rt n J2S C*YPCCKICRrFK6SU6QlllISRICHTCEICPrQ
c c r RX r £ • ?K Q Y f
Z F 2 JSS COFItOCERRrSRSDQLICRilQRRaTCVICPFO
P Cf W NZ F 3 J 8 S CRT C O R R F S aS D H L K tH T R T H T C K T eE K P F S
Z F 4 4 1 6 C R W P S C Q K R F R aS O eC V aaR N K U Q R K K T R I^ l^L
Figure 4.1 (A) W T l m utations associated w ith D enys-D rash syndrom e. (B)
P o in t m u ta tions an d fram eshifts in the z inc finger reg ion of W T l. *,
m utation to stop codon; f = fram eshift after the ind icated am ino acid (Reddy
et al., 1996).

110
C ysteine 330 to tyrosine transition was observed in zinc finger 1 (Huff e t al.,
1991).
Thus, m ost m utations described are single am ino acid substitu tions
c lu ste red in the zinc finger region of W Tl protein. They were therefore
postu la ted to interfere w ith DNA binding of W Tl by e ith er d isrup ting the
D N A -binding activity, o r low ering the affinity of the W T l for DNA. The
DDS phenotype elicited by the loss of DNA binding can also be explained by
a ltered W Tl dosage. G iven th a t the levels of W Tl p ro te in are very tightly
reg u la ted du ring developm ent, the d isrup tion of the dosage of functional
W T l protein m ay precipitate the observed DDS symptoms.
It is in trigu ing that the clinical m anifestations of DDS syndrom e in
d iffe ren t patien ts do not clearly correlate w ith any specific type of W Tl
m u ta tio n detected. Ind iv idua ls w ith identical m utations in W Tl pro tein
exh ib it the entire spectrum o f clinical sym ptom s. The sim plest explanation
m ay be that o ther genes influence the phenotypic expression of various W Tl
m utations. Finally, there is a case reported of an ind iv idual who carries a
W T l m utation, and yet is phenotypically norm al (Coppes e t al., 1992). This
ind iv idual's son has DDS syndrom e, and carries the sam e m utation as the
fa ther: exon 9 arg in ine 394 to tryp tophan m issense m u ta tion , the m ost
com m on m u ta tio n found in DDS patien ts (Coppes e t al., 1992). This
observation suggests that even though m ost DDS patients have constitutional
W T l m utations, no t all of them result in the disease. A possible explanation
for th is observation is tha t perhaps the m u tan t W Tl allele is not expressed
d u rin g the crucial stages in development. The genetic background and other
ep igenetic factors of an in d iv id u a l m ay also influence the effects of a
reduc tion in W Tl gene dosage. In addition, W Tl was suggested to play a
m ore im portan t role in m ale gen itourinary developm ent, since genetic

I ll
females w ith W T l m utations associated w ith DDS have a significantly low er
frequency of urogenital abnorm alities than m ales (Coppes e t al., 1993).
The m ajority of m u ta tions in DDS pa tien ts have b een found in a
heterozygous state, suggesting tha t the DN A-binding - deficient isoforms m ay
act in a dom inant-negative fashion. This hypothesis w as firs t p roposed by
Pelletier et al. (1991). Later, Bradeesy et al. (1994) dem onstrated that a p ro te in
containing am ino ad d s 1 - 2 2 2 represents a m inim al dom ain required for the
do m in an t negative effect. R eddy e t al. (1995) confirm ed tha t p ro te in
containing am ino acids 1 - 182 interferes w ith transactivation by w ild -type
W Tl, acting in a dom inant-negative fashion. Thus, dom inan t negative W T l
alleles could inh ib it the W T l transcriptional activation function , even tually
leading to progressive renal failure. A nother m echanism by w hich m u ta n t
W Tl isoform s m ay alter app rop ria te gene expression is th ro u g h conferring
new D N A -binding specificity, thus resulting in a change of gene netw orks
regulated by W T l protein.
To test th is hypothesis, we conducted selection a n d am plification
binding assays (SAAB) for the Denys-Drash m u tan t proteins. Five d ifferen t
m utations w ere in troduced into the zinc finger dom ain of W T l (Figure 4.2).
Two m issense m utations in the second zinc finger substitu te arginine 366
w ith a histidine o r a cysteine. This substitu tion was p roposed to d is ru p t an
a rg in ine-guan ine in teraction , based on analogy w ith the EGR-1 - D N A
complex (Pavletich and Pabo, 1991). The rem aining three m utations are in
finger 3 of W T l, and substitu te arginine 394 w ith a tryp tophan , and aspartic
acid 396 w ith e ither an asparagine or a glycine residue. DNA tem plates w ere
random ized for finger 2 and 3 binding subsites. DNA binding affinities of the
Denys-Drash m u tan t proteins for the selected sequences w ere m easured using
a nitrocellulose filter b inding assay. This work is a collaborative study w ith

112
UOWTJ-ZFI - K R P F M
ISOW T I-Z R - È K P Y Q
EGR-I-ZFl - E R ? Y A
n;VYTI-ZF3 - V 1C P F Q
6GR-1-ZF2 - Q K P F Q
WT1-ZF4 - E O P F S
EGR-1-ZF3 - E K P F A
R Y P G
D F K O
P V E S
K T - -
R I - -
R W P S
3 K - -
C H K P. Y F K L S K L 0 M
C. H
ERR 0 R ?. F S
»\
Q P. K F S
M R K F S
0 K K F R
G R K F A
E L V
E F. K
M H-k
R‘Aft-
R H
>■'
T HÎ
T H■Û
R K
3 H
H S R K
Q R R
Q R R
T 3 T
I R T
H N M
H K r
T C
T G
T G
T G
T G
O % L 3
F igure 4.2 C om parison of the sequences of W T l and EGR-1 zinc fingers.
R esidues in v o lv e d in DNA b in d in g and the ir co u n te rp arts in W Tl are
enclosed in clear boxes. Shaded boxes indicate residues that coordinate to zinc.
F inger 2 and 3 Denys-Drash m issense m utations are also indicated.

113
Frank Borel, K a th leen Barilla and M ay Iskandar from D r. R om aniuk 's
research group. I perfo rm ed site-directed m utagenesis and expression and
purification of the W T l-Z F Denys-Drash pro tein m utan ts, as well as the
initial D N A -binding assays with the m utant proteins.
4.2 M aterials and M ethods
4.2.1 C onstruction an d expression of D enys-Drash m utan t pro teins
The W Tl-ZFP D enys-D rash m utants (R366H, R366C, R394W, D396G, D396N)
were constructed by site-directed m utagenesis (Nelson & Long, 1989). The
plasm id pUC W TIZFP w hich contains the zinc finger dom ain of W Tl cloned
into pUC19 (construction described in Chapter 3), was used as a
template and the follow ing mutagenic primers w ere used. The base
alteration that in troduces each m utation is underlined:
R366C: AGG TXT TGT IG T TCA GAG GAG
R366H: AGG IT T TGT CAT TCA GAG CAG
R394W: AGG TCC T C C IG G TCC GAC CAC
D396G: TCC CGG TCC GQC CAC CTG AAG
D396N: TCC CGG TCC AAC CAC CTG AAG
DNA sequencing using an ABI 373A autom ated DNA sequencer w as
used to verify tha t on ly the desired m utation was in troduced in to the W Tl-
ZFP cDNA. M utan t cDNAs were cloned into the Nde I and Bam H I sites of
the T7 expression vector pET16b (Studier & Moffatt, 1986, S tudier e t al., 1990).
The resulting constructs encode zinc finger peptides w ith a histidine tag and a
factor Xa cleavage s ite fused to the am ino term inus and w ere used to
transform Escherichia coli BL21 (DE3) pLysS. Expression and purification of
the zinc finger p ro teins w ere carried o u t as described previously (Chapter 3).
Protein concentrations w ere determ ined by the Bradford m ethod, using BSA

114
as the standard (Bradford, 1976). SDS-poIyacrylamide gel analysis w as used to
determ ine pro tein purity (Figure 4.3). P rotein yield averaged 10 m g per litre
of bacterial culture. Fractional activity of each p ro te in p rep ara tio n (75%-
100%) w as determ ined by a saturation DNA binding assay.
4.2.2 Selection am plification and b ind ing (SAAB) assay
The SAAB assay w as carried o u t according to a pub lished m ethod
(Blackwell & W eintraub, 1990), with m inor modifications (Figure 3.5). The 67
base pair tem plate oligonucleotides contained a central region includ ing the
W Tl consensus sequence random ized in either the finger 2 o r finger 3
recognition sites (5' TAAT GCG TGG N N N TGT CCTAA 3' an d 5' TAAT
GCG N N N GCG TGT CCTAA 3' respectively) flanked by Eco RI (5') and
BamH I (3 ) recognition sequences and the M13 fo rw ard universal p rim er
(F.U.P.) sequence (5') and a sequence com plem entary to the M13 reverse
universal prim er (R.U.P., 3’) (Figure 4.4).
The tem plate was labeled for the initial round of selection by prim er
extension using 10 pm ol of the oligonucleotide annealed to 80 pm ol R.U.P. in
IX Klenow buffer (New England Biolabs), and extended in 0.2 mM dGTP, 0.2
mM dCTP, 0.2 mM dTTP, 20 pCi [a-32p]-dATP and 9 un its K lenow (New
England Biolabs) w ith incubation at room tem perature for 45 m inutes. The
resulting double-stranded DNA containing a mixture of 64 possible sequences
in the ran d o m ize d reg io n was p u rif ie d on a 1 2 % n o n -d e n a tu r in g
polyacrylam ide gel, and elu ted overnight a t room tem perature in 250 pi of 0.6
M am m onium acetate, 1 m M EDTA, 0.1% SDS.
Labeled DNA (200,000 cpm) was incubated with the proteins as described
previously (Chapter 3), except the incubation mixture u sed Tris ra th e r than
HEPES to buffer the solution. The reactions were loaded onto a 5% non-

115
MW
L I I I
Figure 4.3 C oom assie b lue-stained 15% SD S-polyacrylam ide gel show ing
pu rifiied W Tl-ZFP D enys-D rash m utants. Lane designated MW represen ts
protein m olecular w eigh t m arker; Lane 1: w ild-type W Tl-ZFP; Lane 2: R366H;
Lane 3: R366C; Lane 4: R394W; Lane 5: D396G; Lane 6 : D396N.

116
F.U.P. ►
5' GTTTTCCCAGTCACGACeAATTCTAATGCGTGGMM MEco RI
TGTCCTTAGGATCCGTCATAGCTGTTTœTG 3' B am H l ^
R.U.P.
(A) SAAB tem plate for WTl finger 2
F.U.P
5' GTTTTCCCAGTCACGACGAATTCTAATGCGMINl MGCGEco RI
TGTCCTTAGGATCCGTCATAGCTGTTTCCTG 3'B am H l ^ ............ "
RU .P
(B) SAAB tem plate for WTl finger 3
F igure 4.4 The SAAB tem plate oligonucleotides contain ing ran d o m ized
sequences for the WT1[-KTS] finger 2 (A) or finger 3 (B) recognition.

117
d enatu ring polyacrylam ide gel which had been p re-e lectrophoresed for 2 0
m inutes a t 250 V at 4°C. M igration on the gel was m onitored by load ing one
lane w ith 5 |jJ of 0.1% brom ophenol blue, 0.1% xylene cyanol, 25% glycerol.
The gel w as ru n in 0.3 X TBE at 250 V a t 4°C until the brom ophenol b lue had
m igrated 12 cm. The gel w as then subjected to au to rad iog raphy fo r 2 to 4
hours at 4°C, and the band corresponding to the low est protein concentration
a t which a protein-DNA complex was visible was excised and the DN A eluted
as described above. The elu ted DNA was amplified using F.U.P. and R.U.P. as
prim ers, radioactively labeled, and a new round of selection was begun using
low er p ro te in concentrations.
Four ro u n d s of selection were sufficient to isolate the highest affinity
binding sites for each protein from the pool of random DNA sequences. The
enrichm ent of high affinity sites for the W Tl proteins w as follow ed by the
proportion o f DNA bound to the proteins after each ro u n d of selection. The
low est p ro te in concentrations at w hich a protein-D N A com plex cou ld be
observed in round 4 were: WTl-ZFP, 0.5 nM; R366H, 1.25 nM; R366C, 7.5 nM;
R394W, 100 nM; D396N, 12.5 nM; D396G, 12.5 nM. The final PCR products of
ro u n d 4 w ere digested w ith Eco KL/Bam HI and cloned in to pUC19. The
original random ized oligonucleotides w ere converted to a doub le-s tranded
form w ith co ld dNTPs, and also subject to PCR am plification, restric tion
endonuclease digestion and ligation into pUC19. A sam ple of each ligation
w as used to transform E. coli strain JM109. Plasmid DN A w as isolated from at
least 50 colonies of each transform ation and sequenced using e ith e r the
Sequenase k it (Amersham) o r an ABI 373A autom ated DNA sequencer and
dye prim er chem istry (Applied Biosystems).

118
4.2.3 N itrocellulose Filter B inding Assay
The b ind ing affinities of a series of DNA sequences fo r the W Tl-ZFP
a n d the D enys-D rash m utants w ere quantified using a n itrocellu lose filter
b in d in g assay developed to s tu d y zinc finger p ro tein-D N A in teractions
(Romaniuk, 1990). The details of the assay are described in C hap ter 3.
4.3 Results
4.3.1 Selection of DNA binding sites for Denys-Drash m utant p ro te ins.
To determ ine w hether the Denys-Drash m u tan t p ro teins could confer
new D N A -binding specificities, fo u r rounds of selection, am plification, and
b inding assay (SAAB) were carried ou t for each m utant. A w ild-type W Tl-ZF
pep tide served as a negative control. The SAAB tem plates were: 5 -GCG TGG
N N N TGT-3' for finger 2 m utants, and 5 -GCG N N N GCG TGT-3’ for finger 3
m utants. PCR products of the final rounds of selections w ere cloned, and
about 50 random ly picked clones w ere sequenced for each SAAB experim ent.
The results of the finger 2 SAAB experim ents are show n in Table 4.1, w ith the
frequencies of the selected nucleotides sum m arized in Table 4.2. A pparently,
the m utation o f the recognition arginine at position -1 of the finger 2 a -helix
d id no t resu lt in a new specificity for either m utan t, as all the proteins
selected the w ild-type guanine w ith an almost 100% frequency. Interestingly,
adenine was selected w ith a h igh frequency a t position 8 o f the consensus
b ind ing site b y all finger 2 m utan ts. This is a definite dev ia tio n from the
w ild-type EGR-1 consensus site, an d , perhaps, reflects the real differences that
exist in the b ind ing specificities o f the two proteins. C uriously, a preference
for a thym ine w as revealed at position 9 for the R366H m utan t, w hereas the
w ild-type guanine was predom inantly selected by both the w ild-type W Tl-ZF
and the R366C proteins.

119
T able 4.1 Finger 2 subsite sequences selected from a random ized tem plate by
w ild type W Tl-ZFP and the finger 2 poin t m utants
Subsite Protein
Sequence W Tl-ZFP R366H R366C
CAC - 1 —
GAA — 1 —
GAC - 2 1
GAG 38 13 19GAT 3 23 3GCA - 2 7GCC 1 - 1
GCG 2 1 2
GCT - - 4GGA - - 2
GGC - 2 —
GGG 3 - 1
GGT - - 1
GTA - - 2
GTG 1 3 5GTT 1 1 2
TA A - - 1
TAG - 1 —
TCC 1 - —

120
hase 7 base K
prtiiL-in none WTl R366H R366C nunc WTl R366H R156CA II (I 0 25 82 82 17f IV 0 2 0 25 8 6 28G 21 Vf! 96 98 19 6 4 8r U 2 2 2 17 4 8 IX
b a se 9
none W Tl R366H R366C
•\ 17 0 6 24c 17 4 10 4c 29 88 36 53r 37 8 48 20
T able 4.2 Frequencies of each nucleotide selected a t the finger 2 subsite
positions by W Tl-ZFP and the fînger 2 point m utants
pnxcin:
base 4 base5
none W Tl D396G D396N R394W none W Tl 0396G 0396N R394W
A 19 0 0 0 0 20 6 0 0 22C 22 0 0 2 0 27 0 0 0 0c 30 34 46 10 IS 27 92 100 100 76r 29 66 54 gg 85 26 2 0 0 2
base 6none W T l D396C D396N R394W
•\ 1 0 0 0 0C 18 0 0 0 0G U (00 (00 100 44r 40 0 0 0 56
T able 4.3 Frequencies of each nucleotide selected a t the finger 3 subsite
positions by W Tl-ZFP and the finger 3 point m utants

121
The data for the finger 3 m utan ts are sh o w n in Table 4.3, w ith the
selection frequencies indicated in Table 4.4. Even though th ere w ere three
finger 3 m utants tested vs. two in finger 2, the num ber of subsites selected for
finger 3 m utants is alm ost half that selected for finger 2. T his indicates a
h igher stringency selection, perhaps pointing to a m ore im portan t role played
by finger 3 in DNA recognition. C uriously, the aspartic acid substitu tion
m u tan ts (D396G and D396N) along w ith the w ild-type W Tl-ZF selected the
w ild -type site (T/G)GG. Apparently, the loss of the bu ttressing interaction
betw een this aspartic a d d residue and an arginine does not resu lt in alteration
of b ind ing selectivity: a guanine base was selected w ith a 1 0 0 % frequency.
H ow ever, a substitu tion of an arginine 394 for a tryp tophan resu lted in a
change of specifidty: a thymine w as preferentially selected over a guanine at
position 6 of the binding site.
4.3.2 M easuring the b in d in g affinities of the m u tan t peptides fo r the selected
DN A sequences
The binding affinities of the m utan t proteins for the selected sequences
w ere m easured using a nitrocellulose filter b ind ing assay (Rom aniuk, 1990),
and com pared to those of the w ild-type protein. The representative binding
curves are show n in Figure 4.5. The values of the d issodation constants are
show n in Tables 4.5 and 4.6. The affinities correlate well w ith the firequendes
o f the selected n u d eo tid es at the g iven positions. Thus, a n adenine at
position 8 w as preferentially selected by the w ild type and the finger 2 m utant
proteins. Indeed, the affinity of the w ild-type p ro te in for the GAG triplet is
tw o -fo ld h igher th an for the GCG sequence d eriv ed from the EGR-1
consensus site. A thym ine (position 4) in the finger 3 recognition trip let was
also preferentially selected over a guanine, w hich is reflected in the two-fold

122
T able 4.4 Finger 3 subsite sequences selected from a random ized tem plate by
w ild ty p e WTl-ZFF and the finger 3 point m utants
Subsite Proteins
Sequence W Tl-ZFP D396G D396N R394W
CGG - 1 -
GAG - — — 1
GGG 17 23 5 6
GGT - — — 1
GTG 1 — — -
TAG 3 — — 5
TAT - — — 6
TGG 32 27 45 12
TGT - — — 2 2
TTT - — — 1

123
•OC3O
<zQCO58
1
0.8
0.8
0.4
0.2
01 0 -11 1 0 10 1 0 ® 1 0 ^
[Protein], M1 0 -7 1 0 "
Figure 4.5 The equilibrium binding of the W Tl elem ent GCG TGG GAG TGT
to W Tl-ZFP (open circles), R366H (closed d iam onds) a n d R366C (open
squares). Data points rep resen t the m ean of six o r more determ inations and
erro r bars indicate the s tan d ard deviations. Curves rep resen t the best fit of
the data to a sim ple bim olecular equilibrium .

124
G C G TGG CAC TGT
WTl -Zi'T* protein relative affinity*
wild type 0.77 ± 0.07 IK166K 2.99 ± 0 .1 9 0 2 6R )66C 10.8 ± 0 .5 7 0.07
GCG -TG G-CAT-TG T
W TI ZFP protein f . ( n M r relative a/Iiniiy* sclecuvity'
wild type 1.93 ± 0.08 1 2.50R366H 2 62 ± 0 .1 7 0.71 0 8 8RJ66C 14 1 ± 0 .9 1 0.14 1.30
Table 4.5 Binding affinities of w ild type and finger 2 m utan t W Tl-ZFP for
selected DNA sequences
cRatio of KrifG A G subsite)/fC<(GAT subsite)
WTl-ZFP proccinCCG-TGC-GCG-TGT
K t( a M r telative a f lin i^wild type 1.54 ± 0 .1 2 ID396G 4.11 ± 0 .2 9 0 J 7D396N 124 ± 0 .8 0 0.12
CCO-CCG-GCG-TGTWTl-ZFP protein K tlaM r relative aflituty* selectivity'
wild type 3.56 ± 0 3 4 1 233D396G 531 ± 0 3 9 0.63 1330396N 263 ± 2 4 4 0.14 213
Table 4.6 Binding affinities of w ild type and finger 3 m utant WTl-ZFP for
selected DNA sequences
^Dissociation constants a re expressed as the m ean with standard deviations of
a m inim um of six independen t determ inations
bRatio of BCd(mutant W T1-ZFP)/Kd(wild type WTl-ZFP)
cRatio of KdCEQG subsite)/K h(GG G subsite)

125
higher affinity for the TGG vs. GGG triplet. In contrast, a thymine a t position
6 of the finger 3 recognition triplet was never selected by the w ild type pro te in
o r the aspartic acid m utants (D396G and D396N). Accordingly, these p ro teins
show ed no detectable b ind ing affinity for the finger 3 subsite TGT. In
addition, the R394W m utant show ed no specific DN A-binding affinity fo r any
of the selected sequences. The fact that this protein selected some sequences
in the SAAB experim ent probably reflects nonspecific b ind ing due to the h igh
pro tein concentrations (100 nM or more) used in the assay. This is the only
protein m u tan t that lacks the specific DN A-binding activity. The o th e r four
D enys-D rash m utan ts show ed varying degrees of red u ced D N A -binding
affinities w hich range from 1.4 - to 14 - fold. It is of in terest to no te the
differences in severity o f each ind iv idual am ino acid m uta tion . T hus,
substitu ting a n arginine in zinc finger 2 for a cysteine p roduced a g rea ter
reduction in b ind ing affinity: 14-fold for the GAG subsite, and 7.2-fold for the
GAT subsite. Replacement of an arginine w ith histidine, o n the o ther hand ,
resulted in a 3.9-fold and a 1.4-fold reduction in affinity for the GAG an d
GAT subsites, respectively.
4.4 D iscussion
A lm ost all (95%) of the Denys-Drash patients have constitu tional W Tl
m utations (C oppes et al., 1993). These m utations involve key am ino acids
w ithin the zinc finger dom ain of W Tl, affecting the D N A -binding function of
the protein . This study w as designed to determ ine th e effects o f these
m uta tions o n the D N A -binding ability o f W T l, as w ell as to a d d re ss a
question of w hether these m utations confer new DN A-binding specificities.
The b in d in g site selection and am plification experim ents w ith the 5
zinc finger m u ta n t proteins d id not resu lt in a selection of a new , high-

126
affinity DNA sequence for any of the m utant proteins. In fact, the pro tein
w ith the m ost com m on m utation found in DDS patients, R394W, show s no
detectable D N A binding . The rem ain ing four p ro te in m u tan ts d isp layed
a ltered DNA b ind ing selectivities. Thus, both finger 2 m u tan ts R366H and
R366C had reduced sequence selectivity, w ith the largest effect observed for
the R366H m u ta n t (Table 4.5). H ow ever, this m u tan t had a four-fold lower
affinity for the selected GAG relative to the w ild-type p ro tein , w hereas the
R366C m utan t resulted in a 14-fold reduction in binding affinity. Presum ably,
h istid ine substitu tion at this position is less d isrup tive for the W T l DNA-
b ind ing function than the cysteine substitu tion due to the capacity of the
histidine to in teract w ith guanines. This histidine could also form a num ber
of van der W aals interactions w ith the thym ine in position 9 of the triplet,
thus explaining w hy bo th thymine and guanine w ere selected a t th is position.
A uniform selection of an adenine residue at position 8 o f the finger 2
b ind ing subsite by the w ild-type and the m utan t proteins, as w ell as the
higher affinity of the w ild type pro tein for the adenine-containing sequence
ind icates th a t th is aden ine base is an im portan t d e te rm in a n t in W Tl
sequence specificity . The results of the hum an genom ic D N A selection
stud ies also iden tified adenine at position 8 (N akagam a e t al., 1995). The
au thors p roposed an interaction betw een a glutam ine (Q369) of the W Tl and
an adenine. In addition , the results of the m utagenesis s tu d y described in
C hap ter 5 of th is thesis confirm the im portance of this aden ine for high-
affinity binding. Strikingly, when the guanine at position 9 w as replaced by a
thym ine, the affinity of the wild-type W Tl-ZF protein was reduced only by a
factor of 2.5. Position 9 w as identified as one of the key determ inan ts for
recognition in the EGR-1 X-ray study (Pavletich & Pabo, 1991). M utagenesis
s tu d ies w ith EGR-1 an d W Tl-ZF show that su b s titu tio n o f guan ine a t

127
p o sitio n 9 w ith a cytosine com pletely abolished binding o f b o th p ro te in s
(C hapter 5 of th is thesis). It rem ains to be seen w hy th ym ine is so w ell
to lerated a t this position.
In a recently refined crystal structure of EGR-1 - DNA com plex, aspartic
acid residues found a t the second positions of the a-helices w ere show n to
p lay an im p o rtan t role in recognition by no t on ly stabilizing the arg in ine-
guan ine interaction, bu t by establishing direct hydrogen bond contacts to the
secondary strand o f the DNA (Elrod-Erickson e t al., 1996). T hus, it is n o t
su rp rising that m uta tions of this am ino acid wUl have deleterious effects on
the p ro te in function. Indeed, bo th glycine an d asparagine su b stitu tio n s of
asparta te in finger 3 result in a three-fold an d 8 -fold reduction in b ind ing
affinity, respectively. In terms of selectivity, on ly a glycine m u ta n t had an
appreciably reduced sequence selectivity. Interestingly, all the pro te ins used
in the finger 3 selection experim ents preferred a thym ine over a guanine a t
position 4 of the b ind ing sequence, w hile adenine or a cytosine w ere never
selected. This ag rees well w ith the results o f the refined EGR-1 crystal
struc tu re (Elrod-Erickson et al., 1996), which show ed that both thym ine 4 and
its com plem entary base on the opposite strand of the DNA (aden ine 4’) are
invo lved in contacts w ith the p ro tein : thym ine 4 is used by a finger 2
h istid ine to establish a num ber of v an der W aals contacts, w hereas adenine
4' is contacted by an aspartic a d d residue of the first finger of EGR-1. Finally,
rep lacem ent of a guan ine at position 6 abolished W Tl-ZF b ind ing . This is
consistent w ith the role this base p lays in the EGR-1 - DNA recognition , as
identified in the X-ray structure (Pavletich & Pabo, 1991), as w ell as w ith a
n um ber of m utagenesis studies w hich observed the loss of b in d in g u p o n
substitu tion of the guanine at this position (Rauscher et al., 1990; N akagam a
e t al., 1995, H am ilton e t al., 1997). H ow ever, w hile guanines a t positions 6

128
and 9 play an equally im portant role in the binding o f EGR-1, there m u st be a
functional nonequivalence betw een these positions for the W Tl b ind ing as
thym ine is to lerated at position 9, but not at position 6 .
In su m m ary , the resu lts of this s tu d y p o in t to several in te res tin g
conclusions. T he SAAB experim ents d id not y ie ld new DNA sequence
specificities, th u s suggesting th a t the Denys-Drash syndrom e likely does not
arise due to a change in the gene networks regulated by the W Tl protein. The
m ost com m on DOS m utation , R394W, abolished specific b ind ing o f the
protein. O ther m utations resulted in reduced binding activities, ranging from
1.4 to 14-fold. This suggests that even sm all changes in the D N A -binding
activity of W T l p ro te in can contribute to the deve lopm en t of the D enys-
D rash syndrom e, suggesting th a t the cellular levels o f W Tl m ust be very
tightly reg u la ted . It is still no t clear w hether the red u ced D N A -binding
affinity of the DDS m utants is directly attributable to the phenotype seen in
DDS patients, o r w hether the dysfunctional W Tl p ro te in m ay interfere w ith
the function o f o ther cellular proteins. This study also provides insights into
the roles of som e of the key am ino acids to the W Tl recognition.

129
CHAPTER 5.0. COMPARATIVE ANALYSIS OF THE D N A BINDING
CHARACTERISTICS OF WILMS' TUMOUR A N D EARLY GROWTH
RESPONSE PROTEINS
5.1 Introduction
The dem onstra tion that the WTL protein is able to b in d to the EGR-1-
like sequences suggests that there may be a regulatory link betw een these tw o
proteins: the W T l protein m ay act as an antagonist of EGR-1 transcription
factors, or m ay be a tissue-specific factor th a t is involved in m aintaining a
particular differentiated phenotype. The balance in the levels of EGR-1 and
W Tl proteins m ay be critical: inactivation of W Tl could resu lt in the onset of
neoplasia.
The X-ray crystallographic studies of the EGR-1 zinc finger region
bound to a 9 base pair fragm ent of DNA containing an EGR-1 consensus site
have been serv ing as a topological blueprint to understand the m echanism of
DNA b inding by W Tl (Pavletich & Pabo, 1991; Elrod-Erickson, 1996). The
details of the high-resolution analysis are discussed in detail in Chapter 1. All
of the residues of the EGR-1 zinc fingers critical for specific b inding to DNA
are conserved in fingers 2-4 of W Tl. H ow ever, despite the h igh degree of
sim ilarity shared betw een the W Tl and EGR-1 D N A -binding dom ains, there
are a num ber of structural differences that suggest that the m olecular details
of DNA b ind ing are not identical between the tw o proteins. These structural
differences include approxim ately 50% dissim ilarity be tw een the W Tl and
EGR-1, as well as an additional zinc finger present in W Tl protein .
To p rov ide a clear quantitative picture of the in terac tion of W Tl and
EGR-1 p ro te ins w ith their DNA binding sites, we co n stru c ted pep tides
encom passing the zinc finger regions of these proteins (W Tl-ZF and EGRl-
ZF), and u se d a nitrocellu lose filter b ind ing assay to m easu re various

130
param eters of a bim olecular equilibrium . The stoichiom etry o f the DNA-
p ro te in com plexes, their stability to d issociation, and the effects of pH ,
tem p era tu re and sa lt concentration on the equ ilib rium b in d in g o f these
proteins to their cognate DNA sequences have been determ ined. In addition,
the relative contribution of each base pair in the consensus b ind ing site to the
high affin ity b ind ing w as determ ined by po in t m utational analysis. This
study is a collaborative effort of several m em bers of Dr. R om aniuk 's lab:
Cathy Ju rid c and Kathy Barilla m ade the DNA constructs incorporating single
m utations encom passing the W Tl and EGRl consensus binding site; Franck
Borel assay ed EG Rl-ZF b ind ing to the m u ta n t DNA sequences. The
rem aining experim ents I perform ed myself.
5.2 M aterials and M ethods
5.2.1 C onstruc tion an d purification o f recom binant W Tl-ZF a n d EGRl-ZF
pro teins
The construction of a synthetic gene encoding the zinc finger region of
EGR-l-ZF protein w as conducted by analogy w ith th a t of W T l-Z F (please,
refer to C hap ter 3). The purification of the recom binan t p ro te in s was
perfo rm ed as described previously (C hapter 3). Typical p ro te in yields
averaged 10 m g per litre of bacterial culture. O n SDS-PAGE (15%), the
purified EGRl-ZF m igrated with an apparent m olecular weight o f 17,000 kDa
(Figure 5.1). The purified protein products were aliquoted and sto red a t -80'C.
Protein aliquots were used only once w hen thaw ed.
5.2.2 C onstruction of m u tan t WTL-ZF and EGRl-ZF DNA b in d in g sequences
O ligonucleotides incorporating point m uta tions a t each one of the 1 2
positions of the W Tl-ZF and EGRl-ZF consensus b ind ing sequence w ere

131
m
Figure 5.1 Coom assie b lue-stained 15% SD S-poIyacrylam ide gel show ing
EG Rl-ZF protein purified by affinity chrom atography. Lane designated MW
rep re sen ts p ro te in m olecular w e ig h t m arker; la n e 1 : total co lu m n pass
through; lane 2: purified EGRl-ZF.

132
synthesized by Dr. Paul J. Rom aniuk using a n A pplied B iosystem s Inc. 391
DNA synthesizer. The oligonucleotide w ith a m utation at position 1 (1C) had
the follow ing sequence: GATCCA C C G T G G GCG T G T AG, w here a
h ig h lig h ted sequence rep resen ts the co n sen su s b in d in g site , and an
underlined position denotes a m utated base. Similarly, o th e r m utations w ere
introduced: 2(T, A, G), 3C, 4(A, C, G), 5C, 6C, 7C, 8 (A, G, T), 9C, IOC, IIC , 12C
(Table 5.1). The oligonucleotides were cloned in to BamHI site of pUC19, and
their sequences verified by autom ated DNA sequencing u sin g an ABI 373A
autom ated DNA sequencer and fluorescent d ye prim ers.
5.2.3 End-labeling o f DNA
Individual m u tan t DNA sequences w ere released from plasm id pUC19
using EcoRI and H in d m restric tion enzym es, an d e n d -la b e led w ith [a -
32p]dATP and the Klenow fragm ent of E.coli DNA polym erase I (Sambrook et
al., 1989). The details of the labeling protocol are provided in C hap ter 3.
5.2.4 N itrocellulose Filter B inding Assay
The binding affinities of a series of DNA sequences for the W Tl-ZF and
EGRl-ZF peptides w ere quantified using a nitrocellulose filter b inding assay
developed to study zinc finger protein-D N A interactions (Rom aniuk, 1990).
The details of the assay are described in C hapter 3.
5.3 Results
5.3.1 E quilibrium b ind ing constants
We have m ade a detailed investigation of the m echan ism s of DNA
b ind ing by both W Tl-ZF and EGRl-ZF p ro te ins, using m e th o d s that have
been applied successfully to the study of the D N A and RNA b ind ing

133
Table 5.1 Sequences of mutant oligonucleotides harbouring
individual base pair substitutions in the EGRl-ZF and W Tl-
ZF consensus DNA binding site
G IC G ATCCAÊCG TGG GCG TGT AG
C2A GATCCA g a g TGG GCG TGT AG
C2G GATCCA OfiG TGG GCG TGT AG
C2T GATCCA G IG TGG GCG TGT AG
G3C GATCCA GCG_TGG GCG TGT AG
T4A GATCCA GpG AGG GCG TGT AG
T4C GATCCAGCG GGG GCG TGT AG
T4G GATCCA GCG GGG GCG TGT AG
G5C GATCCAGCG TCG GCG TGT AG
G6 C GATCCAGCG TGÇ GCG TGT AG
G7C GATCCAGCG TGG gC G TGT AG
C8A GATCCA GCG TGG GAG TGT AG
C8 G GATCCAGCG TGG GfiG TGT AG
C8T GATCCAGCG TGG G IG TGT AGG9C GATCCAGCG TGG GCC.TGT AG
HOC GATCCAGCG TGG GCG f î? T AG
G llC GATCCAGCG TGG GCG T C I AG
T12C GATCCAGCG TGG GCG TGC AG

134
p ro p erties of the zinc finger p ro te in Xenopus tran scrip tio n fac to r TFiHA.
(Rom aniuk, P J. 1985; Rom aniuk, P J . 1990). Purified recom binant zinc finger
pep tides of W Tl-ZF and EGRl-ZF w ere used to determ ine the param eters of
the equ ilib rium b ind ing of the pro teins to th e ir p referred D N A b ind ing
sequences. DNA binding w as m easured by titra ting radiolabeled DNA w ith
increasing am ounts of purified pro tein and m easu ring the frac tion of DNA
bound by gel shift and filter b ind ing assays (Rom aniuk, 1990). Each data point
sh o w n in F igure 5.2 rep resen ts the m ean o f a t least 3 in d e p e n d e n t
determ inations. The apparent dissociation constant (Kd) m easu red for W Tl-
ZF - DN A complex is 1.14± 0.2 x 1 0 ^ M under conditions of 0.1 M KCl, pH 7.5
and incubation a t 22°C. The respective value for the EGRl-ZF - DNA binding
is 3.55 ± 0.4 X 10*^ M under the sam e conditions.
To determ ine the nature of the equilibrium , the dissociation constants
a n d the fraction o f active p ro te in , we p e rfo rm ed a S ca tchard analysis
(Scatchard, 1949) of the interaction betw een the D N A consensus site and both
W Tl-ZF and EGRl-ZF proteins. In this analysis the pro tein concentrations
w ere held constant (2 nM for W Tl-ZF, and 5 nM for EGRl-ZF), an d the DNA
concentrations w ere varied from 0.31 to 30 nM. The results o f the Scatchard
analysis are show n in Figure 5.3. The lines indicate the theoretical curves
ca lcu la ted for the form ation o f a sim ple b im olecu lar equ ilib rium . The
dissociation constants m easured from the slopes of the lines are 9.7 x 10"^ M
for W Tl-ZF - DNA complex, an d 2.05 x 1 0 ^ M for EGRl-ZF - D N A complex.
Both values are in close agreem ent w ith those m easured using the standard
filter b inding assay. The results of the Scatchard analysis also indicate that the
p ro tein preparations are 100% active in the DNA binding assays.
To determ ine the time requ ired for the tw o system s to reach a true
equilibrium , we conducted a tim e-dependence assay, in which the

(A)
135
0 .8 -■ Oc3OCD
0.6 -<Za
o 0 .4 -wu(Qi _U.
0.2 -
,-10 10
[W Tl-ZF], M
F igure 5.2 E qu ilib rium b in d in g curves of W T l-Z F (A) and E G R l-Z F (B)
p ro te in s to the ir target D N A sequence. The affinities of W Tl-ZF a n d EGRl-
ZF p ro te ins for their consensus D N A sequence w ere m easu red using a
n itrocellu lose filter b in d in g assay. Each line rep resen ts the best fit for the
form ation of a simple bim olecular com plex w ith K d values of 1.14 x 10-09 for
W Tl-ZF (R = 0.994), an d 3.55 x 10-09 for EGRl-ZF (R = 0.997). Each da ta point
is th e m ean of th ree o r m ore in d e p e n d e n t d e te rm in a tio n s , w ith the
associated standard deviations indicated w ith the e rro r bars.

136
0 .8 -• oc3Om
0.6 -
<ZÛ
o 0 .4 -4-*UCQk .
u _0 . 2 -
-10
[EGRl-ZF], M
F igure 5.2 (B) E qu ilib rium b in d in g curve of EG R l-ZF to its ta rge t D N A
sequence.
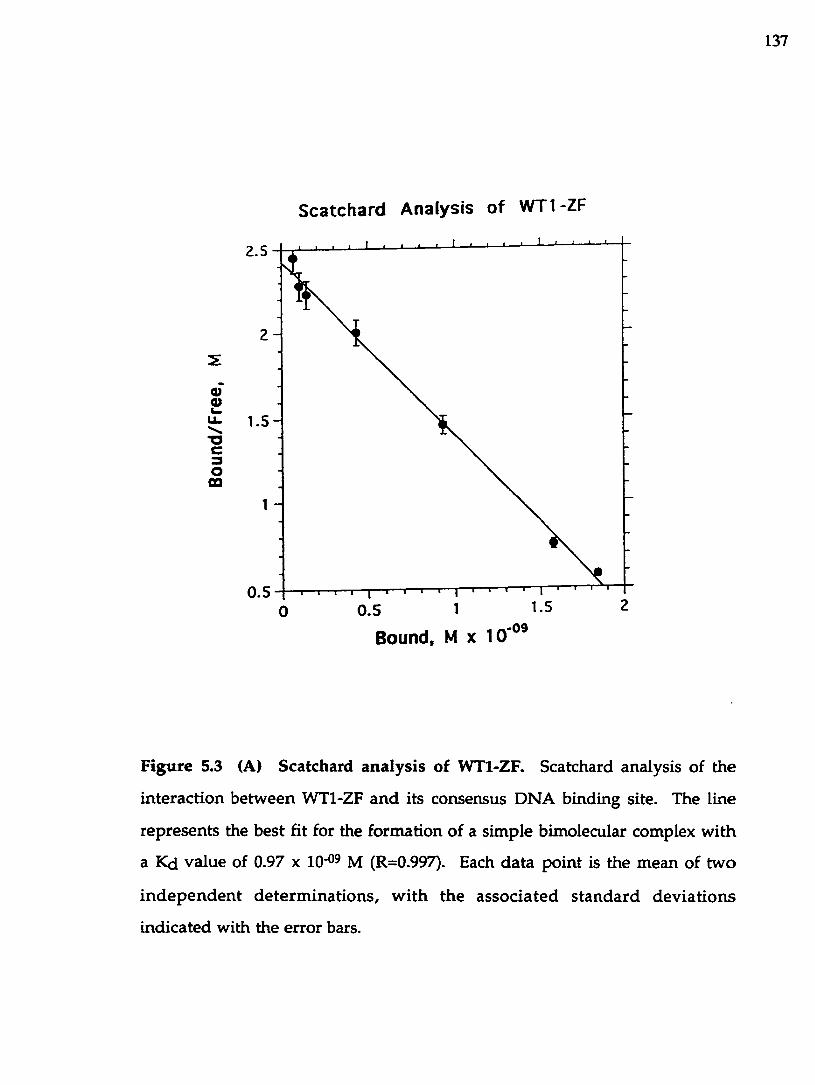
137
Scatchard Analysis of WTl-ZF
2.5
2 -
S
2u.—*oc3Om
1 .5 -
0.50.5
Bound, M X 10'°®
F igure 5.3 (A) Scatchard analysis of W Tl-ZF. Scatchard analysis of the
in teraction betw een W Tl-ZF and its consensus DN A binding site. The line
represents the best fit for the form ation of a sim ple bimolecular complex w ith
a Kd value of 0.97 x 10*09 M (R=0.997). Each da ta point is the m ean of tw o
in d e p e n d e n t d e te rm in a tio n s, w ith the a ssoc ia ted s tan d ard d ev ia tio n s
indicated with the error bars.

138
Scatchard analysis of EGR-1
2 .5 -
2
2u_"Oc3o
CQ
0.554320 1
Bound, MxlO-09
Figure 5.3 (B) Scatchard an a ly sis o f EGRl-ZF. Scatchard analysis of the
equilibrium binding of EGRl-ZF to its DNA sequence. The line represents
the theoretical curve calculated for a sim ple bim olecular equ ilib rium w ith a
K d value of 2.05 x 10'°^ M (R=0.999). Each data point is the m ean of tw o
in d e p e n d e n t de te rm in a tio n s, w ith the a sso c ia ted s ta n d a rd dev ia tions
indicated with the erro r bars.

139
d e p en d e n ce of th e a p p a re n t Ka values w ere m easured as a function of
in cu b a tio n times. F igure 5.4 show s th a t in a reaction m ixture containing a
ta rg e t D N A and a com petitor DNA (poly dl-dC) bo th pro teins reach the full
eq u ilib riu m after 60 m inutes of incubation. The difference in the shapes of
the tim e-dependence curves m ay reflect distinct m odes of b ind ing em ployed
by each protein. T he differences in the binding m odes m ay include different
k inetic rates of d issociation (not determ ined in this study), as w ell as o ther
u n id e n tif ie d co m p o n e n ts o f the eq u ilib riu m , su ch as in fluences of
co n fo rm atio n a l changes occu rring u p o n com plex fo rm ation , sequen tial
versus sim ultaneous b ind ing by indiv idual zinc fingers, and others. Because
the tim e-dependence experim ents w ere conducted last, incubation times used
in the experim ents described in this paper, were 45 m in (w ith the exception of
the tem p era tu re -d ep en d en ce assay w hich w as allow ed to proceed for 60
m inutes). Therefore, the apparen t association constants w ere m easured and
used in the com parative analysis.
5.3.2 M onovalent sa lt dependence of the Ka for DNA b ind ing
The affinity constants w ere m easured at potassium concentrations that
v a ried from 0 to 400 mM. The results given in Figure 5.5 show that the
b in d in g interaction has an op tim um betw een 100 m M and 250 mM for the
W T l-Z F protein, w ith affinity sharply declining at higher salt concentrations.
The o p tim u m b ind ing for the EGRl-ZF p ro tein is betw een 0 an d 100 m M
p o tass iu m salt w ith a decrease in b ind ing affinity observed a t higher sa lt
concentrations. Both proteins exhibit dram atic dependencies of their b inding
co n s tan ts on the m o n o v a len t sa lt concentra tion . A nalysis o f the sa lt
dep en d en ce of the p ro te in - DNA b ind ing p rov ides in form ation about the
n um ber of cationic g roups on the pro tein that interact w ith DNA phosphates

140
8 -
7 -
m
4 -Q.Q.
20 40 60 80 100 120 1400
Time (min)
Figure 5.4 Time dependence of the b ind ing of W Tl-ZF (closed circles) and
EG Rl-ZF (open circles) to DNA consensus sequences. A pparen t Ka s w ere
determ ined using the nitrocellulose filter b inding assay in TMK buffer. Each
data p o in t is the m ean of 3 or more independen t determ inations, w ith the
associated standard deviations indicated w ith the error bars.

141
in each com plex. The num ber of ion pa irs form ed in a protein-D N A
interaction can be determ ined using an analysis of the sa lt dependence of Ka
based u p o n ion displacem ent (von H ippel & Schleich, 1969; Record e t al.,
1976). The num ber of ion pairs can be obtained using the equation: In Kobsd =
In K° - Z y ln[M+] - Z ln(0.5 (l+ (l+4 KMgobsd [Mg2+])0.5)) w here K° is the
apparen t Ka a t IM salt, Z is the number of ion pairs form ed, y is the fractional
counterion b ound per phosphate in the DNA (assum ed to be 0.88 for double
stranded DNA), and K^Sobsd is the observed binding constant for the Mg2+ -
DNA interaction. From the results presented in Figure 5.5 w e calculated that
there are 7±1 ion pairs form ed in the W Tl-ZF-DNA com plex, and 6±1 ion
pairs form ed in the EGRl-ZF-DNA complex. This com parison indicates that
W Tl-ZF m akes m ore ionic bonds to DNA than does EGRl-ZF.
The values of electrostatic and nonelectrostatic contributions to the
observed free energy of binding can be determ ined using an analysis of the
salt dependence of Ka based upon ion displacem ent (von H ippel, P.H . and
Schleich, T. 1969; Record, M.T. e t al. 1976). How ever, the determ ination of
these values w as not possible since both proteins dem onstrate the presence of
anionic b ind ing sites.
5.3.3 pH dependence of Ka
The p H dependence of the ap p a ren t Ka for D N A b ind ing was
determ ined using appropriate buffers ranging from p H 5.5 to 9.0. As show n
in Figure 5.6, bo th the W Tl-ZF and EGRl-ZF-DNA interactions have a b road
p H op tim um from pH 5.5 to 8 . Both p ro teins d em o n stra te a strik ing
sim ilarity in the pH dependence of the binding affinity. The affinity decreases
a t pH values above 8 , which suggests that a t low proton concentrations some
of the functional groups on either protein (such as e-NHz groups of arginines)

142
22
2 0 -
m
C
32.521.50.5 1
-In [K+]
Figure 5.5. KCl concentration dependence of the b inding of W Tl-ZF (closed
circles) a n d EGRl-ZF (open circles) to consensus sequences. The effect of
increasing the KCl concentra tion in th e b ind ing bu ffer on the Ka w as
m easured using the nitrocellulose filter b ind ing assay. Each data point is the
m ean of 3 o r m ore independent determ inations, w ith the associated s tan d ard
dev ia tions indicated w ith the error bars. The experim ental da ta w ere fit to
equation 5 of Lohm an e t al (1980) by a regression m ethod w hich varied the
num ber o f ion pairs and the apparent Ka a t IM salt.

143
9.5
9 -
m8 .5 -O)
o
8 -
7.51 05 96 a7
pH
F ig u re 5.6. pH dependence of the b inding o f W Tl-ZF (closed circles) and
EG R l-ZF (open circles) to DNA consensus sequences. The effect of changing
the p H of the binding buffer from 5.5 to 9.0 on the Ka was m easured using the
n itrocellu lose filter b inding assay. Each data p o in t is the m ean o f 3 or more
in d e p e n d e n t de te rm ina tions, w ith the a sso c ia ted s ta n d a rd dev ia tions
ind ica ted w ith the erro r bars.

144
or D N A get depro tonated . This reduces the am oun t of ion release th a t
accom panies com plex form ation, and therefore the observed KaS decrease
w ith increasing pH.
5.3.4 Effect of d ivalen t metal ion concentration o n DNA b ind ing
The effect of the divalent m etal ion concentration on the efficiency of
b ind ing was m easured by varying the Mg^+ concentrations over a b road range:
from 0 mM to 20 mM. The data in Figure 5.7 illustrates that W Tl-ZF and
EGRl-ZF binding have an optim um at 4 to 5 m M Mg^+, gradually decreasing
at e ither lower or higher Mg^+ concentrations.
5.3.5 Tem perature dependence o f the Ka value
The effects o f tem perature on the b in d in g affinities of W Tl-ZF and
EGRl-ZF proteins have been determ ined using a range of tem peratures from
4®C to 37 °C. The data is shown in Figure 5.8, an d dem onstrates the opposing
effects of tem perature on the W Tl-ZF - DNA a n d EGRl-ZF - DN A complex
form ation . For W Tl-ZF, the affinity of the in te rac tion is h ighest a t 37°C,
w hile EGRl-ZF - DNA binding favors the low er values of the tem perature
range: the binding affinity is highest at 4°C.
A num ber of therm odynam ic param eters can be calculated from these
data: the free energy of the interactions can be calculated using the equation:
AG“= - RTlnKa (Lohm an, T.M. & M askotti, D.P. 1992), assum ing that the
a p p aren t Ka value derived from the filter b ind ing experim ents is equivalent
to th e association constant for the W Tl-ZF-D N A and EG Rl-ZF-D N A
equilibrium . The values of AH" can be calculated from van’t H off analysis,
and th e values of AS* can be derived using the equation: AG’=AH*-TAS".
A t 22"C, the AG" of the W Tl-DNA interaction has a value o f -12.1 kcal

145
9.5
m
O iO
8 -
7.52520151 050
[MgCIJ. mM
F igure 5.7. Effect of the m agnesium ion concentration on the b in d in g of
W T l-Z F (closed circles) and EGRl-ZF (open circles) to DNA consensus
sequences. The Ka v a lu es w ere determ ined using the nitrocellu lose filter
b ind ing assay in TMK buffer supplem ented w ith Mg2+ concentrations show n.
Each da ta poin t is the m ean of 3 or m ore independent determ inations, w ith
the associated standard deviations indicated w ith the error bars.

146
20.5 -
2 0 -c
1 9 .5 -
3.73.63.53.43.23 »1/T (xio’ ) "1C
Figure 5.8. Tem perature dependence of the binding of W Tl-ZF (closed circles) a n d EGRl-ZF (open circles) to DNA consensus sequences. The Ka v a lu e s w ere de te rm ined a t tem pera tu res rang ing from 4°C to 37°C, u s in g the nitrocellulose filter binding assay. The v an 't H off plot w as used to calculate the en thalpy of these interactions by fitting each da ta se t by linear least- squares analysis. The two correlation coefficients are g rea ter than 0.99. Each d a ta po in t is the m ean of 3 o r m ore in d ep en d en t determ inations, w ith the associated s tan d ard deviations indicated w ith the error bars. The en tha lpy values were determ ined from the slope of each line using the equation:
dlnKa -AH° d ( l / T ) “ R

147
m o l'i; AH* has a value o f +6 .6 kcal mol-^, and AS" has a va lue of +63.3 cal
mol'^ deg-i. This indicates that the W Tl-ZF-DNA com plex form ation is an
entropy d riv en process. The corresponding param eters fo r the EGRl-ZF-
DNA interaction are: AG°= -11.4 kcal m oH ; AH°= -6.9 kcal mol'^; and AS*=
15.3 cal m ol'i deg-i, and indicate that, in this case, the com plex form ation is an
enthalpy d riven process. The entropie b inding process m ay reflect the release
o f co u n te r ions and o rd e re d w ater m olecu les as w e ll as p o ss ib le
conform ational changes th a t accom pany protein-D N A com plex form ation.
The enthalpic contribution to the free energy change indicates the am ount of
heat transferred in the course of the reaction.
5.3.6 Cation and anion effects on binding
We m easured the effects of substitu ting potassium chloride by various
anions and cations in the b ind ing buffers. The varied cations and anions
w ere kep t a t 0.1 M concentration. R eplacem ent of p o tassiu m by sod ium ,
lithium , or am m onium d id n ’t have a significant effect on th e m easured KaS
for either protein. The substitu tion of chloride by anions like acetate, n itrate,
and iodide resu lted in the decrease of Ka observed in the follow ing o rder:
acetate > chloride > n itra te > iodide (Table 5.2). While acetate and n itra te
produced only m odest changes in the KaS, the largest effect w as observed by
substitu ting chloride for iod ide (about 1 0 0 - fold reduction of KaS for bo th
proteins). The dependence o f the b ind ing on the id en tity of the anionic
species indicates the presence of anionic b ind ing sites on the proteins.

148
T ab le 5.2 Effect of the different m onovalent salts on the b ind ing of W Tl-ZF
an d EGRl-ZF to DNA consensus sequences
W T l-Z F E G R l -ZF
SaltRelativeaffinity Kd {nM)
Relativeaffinity
KCl 1.14 ± 0.09 1.00 335 ± 0.02 1.00NaCl 0.76 ± 0.01 1.5 ± 0.75b 2.76 ± 0.08 1.29 ± 0.06UCl 0.86 ± 0.16 133 ± 020 2.47 ±0.1 1.44 ± 0.03NH4CI 0.58 ± 0.14 1.96 ± 035 321 ±0.11 1.11 ± 0.04NaNOg 1.47 ± 0.03 0.78 ± 0.06 4.99 ±0.13 0.71 ± 0.03NaCHaCOO 12 ±0.14 0.95 ± 0.04 2.71 ± 0.03 1.31 ± 0.11Nal 68.50 ± 6.61 0.017 ± 0.002 -c >0.005
=A pparen t dissociation constants determ ined by nitrocellulose filter binding
assay. The salt concentrations were kept constant at O.IM. Each value reported
rep resen ts the m ean of 3 or m ore independen t de te rm ina tions w ith the
associated standard deviations.
t>The errors for relative affinities are given by the expression s = {(si/M i)2 +
( s 2 / M 2 ) 2 } 1 / 2 X M 1/ M 2, w here Ml and M2 a re the respective d issocia tion
co n stan ts for w ild -type a n d m u tan t DN A and the s v a lu es are the
corresponding standard deviations for these determ inations.
^Binding affinity too low to be determ ined accurately.

149
5.3.7 Identification of relative contributions of each base pair in the consensus
site to the binding specificities of WTL-ZF and EGR-1 proteins
The relative contribution of each base pair in the W Tl-ZF consensus
sequence 5' GCG TGG GCG TGT 3' to the b inding affinity has been tested by
po in t m utational analysis. The binding affinity of each m u tan t DNA w as
determ ined using the nitrocellulose filter b inding assay. A n identical analysis
has been carried out for the EGRl-ZF pro tein interacting w ith the same
consensus DNA sequence. These experiments have yielded a detailed picture
o f the in teraction of each base pair in the DNA w ith the proteins u n d e r
investigation. The results are presented in Table 5.3, and provide valuable
inform ation in assessing the relative im portance of pu tative W Tl and EGR-1
binding sites as they are identified in genomic DNA.
Our analysis revealed that the guanine to cytosine substitutions at
positions 3, 5, 6 , 7, and 9 resulted in a drastically reduced (over 500-fold)
éiffinity of binding for both W Tl-ZF and EGRl-ZT proteins. This corresponds
to the pattern of guanines show n to be contacted in the EGR-1 - DNA co
crystal structure (Pavletich & Pabo, 1991; Elrod-Erickson et al., 1996). The
dissociation constants for these substitutions are indicated as "undetectable"
in Table 5.3, since we could not accurately m easure the equilibrium constants
for those DNA m utants. The first guanine of the consensus binding site has
also been show n to be contacted in the X-ray structure. H ow ever, this
position appears to have a less drastic effect on binding. W hen the guanine is
substituted by a cytosine, the binding affinity is reduced by approxim ately 1 0 -
fold for W Tl-ZF, and about 5-fold for EGRl-ZF. Replacement of guanine
residue w ith cytosine at position 1 removes the hydrogen-bonding potential
for arginines, since cytosine has no hydrogen-bond-accepting groups in the
m ajor groove. This results in diminished affinity for both proteins, how ever
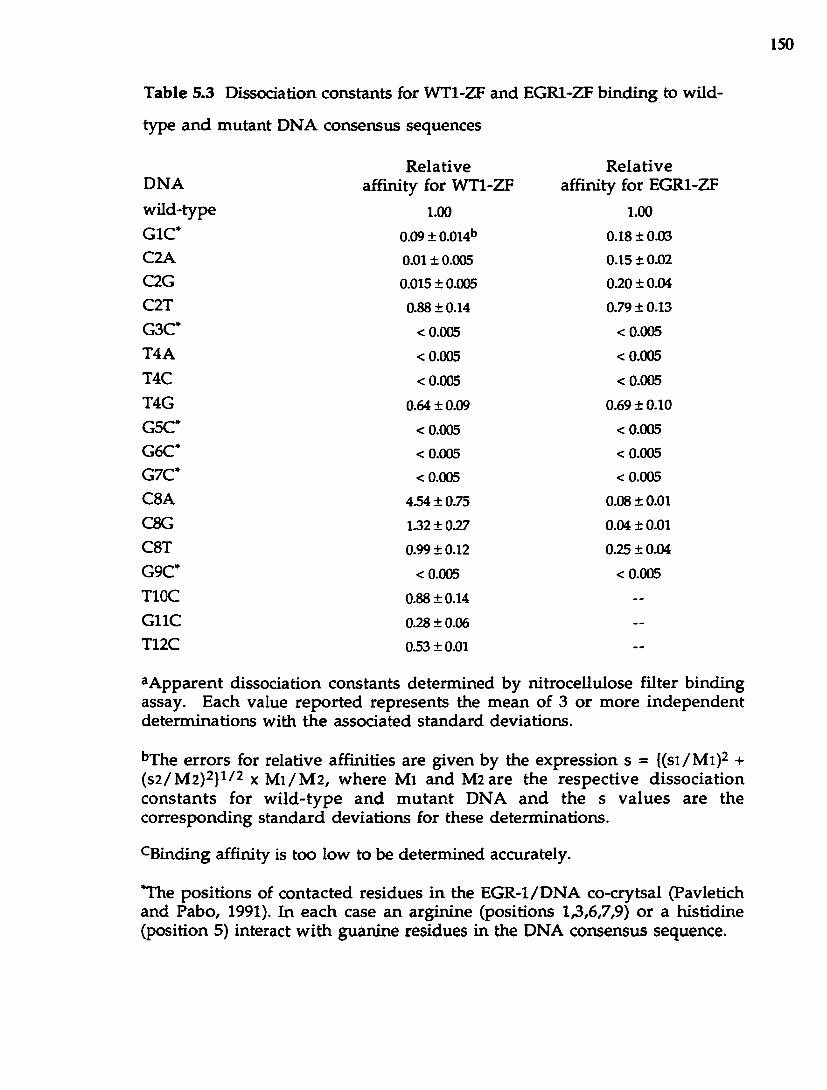
150
T able 5.3 Dissociation constants for W Tl-ZF and EGRl-ZF b ind ing to wild-
type a n d m utan t DNA consensus sequences
DNAw ild-typeQIC*C2AC2GC2TG3C*T4AT4CT4GG5C*G6C*G7C*C8 AC8GC8 TG9C*TIOCG llCT12C
R elative affinity for W Tl-ZF
1.00
0.09 ± 0.014b 0.01 ± 0.005 0.015 ± 0.005 0.88 ±0.14
<0.005
<0.005
<0.005
0.64 ± 0.09
<0.005 <0.005 <0.005
434 ±0.75 132 ±027
0.99 ±0.12
<0.005
0.88 ±0.14
0.28 ±0.06 033 ± 0.01
R ela tive affinity for EGRl-ZF
1.00
0.18 ± 0.03 0.15 ±0.02
0.20 ± 0.04 0.79 ±0.13
<0.005
<0.005
<0.005
0.69 ±0.10
<0.005 <0.005 <0.005
0.08 ± 0.01 0.04 ±0.01 0.25 ±0.04
<0.005
^A pparen t dissociation constants determ ined by nitrocellulose filter b inding assay. Each value rep o rted represents the m ean of 3 o r m ore independen t determ inations w ith th e associated standard deviations.
^The erro rs for relative affinities are given by the expression s = {(si/M i)2 + (s2 / M 2 )2 }1 /2 X M1 / M 2 , w here Ml and M2 are the respec tive d issoc ia tion co n stan ts for w ild -ty p e a n d m u ta n t D N A an d the s v a lu es are the corresponding standard deviations for these determ inations.
^Binding affinity is too low to be determ ined accurately.
*The positions of contacted residues in the EG R-1/D N A co-crytsal (Pavletich and Pabo, 1991). In each case an arginine (positions 1,3,6,7,9) o r a histidine (position 5) interact w ith guanine residues in the DNA consensus sequence.

151
still p rov id ing a significant level of binding affinity. W hile nucleotides 3, 6 ,
and 9 are contacted by an arg in ine residue im m ediately preceding the a -h e lix
(position -1 ), the first and seven th bases in the binding site are contacted by
fingers 1 and 3 w hich use arg in ines at a -helical position + 6 as identified in
the EGR-1 - D N A crystal s truc tu re . The reduction of the b inding s tren g th
observed for thhe m utan t a t position one m ay reflect nonequivalence o f the
contributions to the b ind ing affinity by the different nucleotides of the first
trip le t, as w ell as d ifferen t roles played by arginines d ep en d in g o n their
position in the recognition a-helix . H ow ever, we observed a m uch g rea ter
d rop in affirüty due to the substitu tion at position 7 than a t position 1. This is
ev iden t from the fact that bo th guanine at position 7 and its com plem entary
m ate (C7‘) o n the opposite DNA strand are involved in hydrogen b o n d
contacts from an Arg 24 an d an Asp 48, respectively, whereas there is only one
residue con tacted at position 1, as revealed in the crystal struc tu re . In
add ition , A rg + 6 (finger 3) in teraction w ith the first guanine is not stabilized
by a bu ttressing contact w ith an aspartic acid residue as observed in all three
zinc fingers for the arginines a t position -1. It m ay also be th a t individual zinc
fingers p rovide nonequivalent contributions to the binding affinity. O verall,
there is an excellent correspondence with the X-ray structural data in the w ay
b o th p ro teins respond to changes in positions identified as direct base-pair
contacts.
Base pairs 2, 4 and 8 are no t involved in d irect contacts w ith the EGR-1
zinc fingers. H ow ever, substitu tions at any one of them dem onstrated s trong
preferences a t each of these positions. Thus, substitu tion of a cytosine residue
a t position 2 w ith adenine o r guanine residues results in approxim ately 1 0 0 -
fo ld decrease in b inding affin ity for W Tl-ZF, b u t only a 5-6-fold d ro p in
affin ity for the EGRl-ZF p ro te in , suggesting th a t this in terac tion is m ore

152
critical for the W Tl-ZF p ro tein than for the EGRl-ZF. O n the other hand ,
in troduction of another pyrim idine, thym ine, does not appreciab ly affect the
binding s tren g th of either protein, resulting only in a sligh t (less than 15%)
decrease in the binding affinity. There m ay be several w ays of rationalizing
the observed effects: the identity of the base on the G -rich DN A strand , the
stereochem ical nature of the base, the contacts to the secondary , C-rich DNA
strand or a com bination of these features contribute to th e strong selective
pressure a t this position. Indeed, w ater-m ediated and van d e r W aals contacts
are m ade to the second cytosine as well as to the com plem entary guanine a t
position 2' of the secondary strand as detected in the EGR-1 - DNA crystal
structure. It is not readily apparent, how ever, why the m agn itude of the
affinity change is different betw een the tw o proteins. It is possible that W T l-
ZF contacts are m ore stereochem ically constricted that those of EGRl-ZF.
More m utagenesis and structural data will be necessary to determ ine w hy the
presence o f guanine or adenine at position 2 is more d isrup tive for the W T l-
DNA interaction.
The thym ine su b stitu tio n for e ither an adenine o r a cytosine a t
position 4 abolished the b inding ability of b o th W Tl-ZF a n d EGRl-ZF (over
500-fold red u c tio n in affinity), while bo th pro teins show ed ju s t a slightly
reduced (abou t 30 %) preference for a guanine residue in th is position. As
identified from the X-ray analysis, there are no d irect hydrogen -b ond ing
contacts m ad e a t th is position. The th reon ine w hich co u ld p ro v id e a
hydrogen from its hydroxyl g roup to the m ajor-groove hyd rogen -bond
acceptors o f thym ine (04) o r guanine (N7 and 06 ) is too far aw ay to m ake
such a contact (Pavletiich & Pabo, 1991). H ow ever, as described in C hapter 4,
His 49 m akes van der W aals contacts to the T4 as de tec ted in the refined
structure. A dditionally , Asp +2 of finger 3, w hich forms a residue pair w ith

153
Arg -1 th ro u g h a salt bridge, was tentatively suggested to accept a hydrogen
bond from the N 6 am ine group of the adenine A4’ located on the opposite
strand o f the DNA. In the X-ray structure , this in terac tion d id n o t have a
perfect geom etry, and therefore its im portance for DN A binding w as not clear.
In this s tudy , we show that this contact likely occurs, as it is very im portan t
for the overall affinity o f binding of bo th proteins. This in teraction w ould
also explain a preference for a cytosine a t position 4' (com plem entary to the
guanine o n the prim ary strand): a cytosine, analogous to an adenine, also has
a hydrogen-bond donating amine group - N4 - available for in teracting w ith
aspartic acid . In a recen t study, S w irnoff & M ilb ran d t (1995) also
dem onstrated a strong selection for a thym ine at position 4, and less strongly -
for a guanine, whereas adenine or cysteine were never selected. The authors
also show ed that a m ethyl group of thym ine was im p o rtan t for the optim um
bind ing affin ity by in troducing a uracil substitu tion a t position 4, w hich
reduced b ind ing strength of EGR-1.
F u rthe r differences in the b ind ing preferences for the W T l-Z F and
EGRl-ZF pro teins w ere revealed w hen the cytosine a t position 8 o f the
consensus site had been substituted by an adenine. The binding affinity o f the
EGRl-ZF pro tein d ropped by a factor of abou t 12, w hereas the b ind ing ability
of the W T l-Z F protein w as actually im proved by a factor of 4.5. The tw o
other substitu tions at position 8 - 8G and 8T gave nearly a w ild-type affinity
for the W Tl-ZF protein. However, in the case of EGRl-ZF protein , the 8 G
and 8 T m utations had resu lted in a 25-fold and a 4-fold reduction in b inding
affinity, respectively. Cytosines at positions 2 and 8 are sym m etrical in term s
of their location in the EGR-1 nonam er b inding site. H ow ever, there are
different degrees of tolerance to substitutions at these positions. In the case of
EGRl-ZF b ind ing , som e of the observed effects are im plicit in th e crystal

154
structure. Thus, cytosine 8 and its com plem entary base guanine 8 ' a re both
involved in w ater-m ediated and v an d e r Waals in teractions w ith a num ber
o f amino a d d s : Asp -1 of finger 1 m akes a w ater-m ediated contact to C 8 ; a t the
sam e tim e, G lu +3 of finger 1 makes van der Waals contacts to the sam e base;
while three am ino a d d s - Ser 47 and Asp +2 of finger 2 and Arg + 6 o f finger 1
make a series of w ater-m ediated interactions to G8 '. As m entioned earlier, at
position 2 there are tw o am ino a d d s m aking w ater-m ediated bonds to C2 and
G2’, and one am ino a d d m aking van der Waals contacts to C2. C learly , there
is nonequivalence betw een positions 2 and 8 w hich is reflected in vary ing
degrees o f tolerance to substitutions a t these positions.
In the case of W T l-Z F p ro tein , position 8 o f the DNA recogn ition
sequence d ea rly plays a different role than it does in EGRl-ZF binding. Finger
2 of W Tl-ZF has a glutam ine at a-helical position +3, w hich EGRl-ZF doesn 't
have in its corresponding finger 1. This glutam ine has the po ten tia l to be
either a hydrogen bond donor or acceptor, which w o u ld help explain the base
preferences seen in ou r d a ta (A > G > T or C). Therefore, W Tl-ZF pro tein
uses a d is tinc t mode of DNA b ind ing from that of EGRl-ZF. In add ition ,
com parison of the b in d in g preferences betw een fingers 4 and 2 o f W Tl
suggests th a t different fingers of W Tl provide nonequivalent contributions to
the binding.
The p rev iously iden tified positions 10-12 fo r the W Tl-ZF finger 1
b inding (D rum m ond e t al., 1994; H am ilton et al., 1995) h ad a relatively high
tolerance for sequence alteration (Table 5.1). The largest reduction (four-fold)
in affinity w as observed w hen a guan ine was su b stitu ted for a cy tosine at
position 11 of the binding site. Finger 1 of W Tl has a h istid ine a t position +3
of the a -he lix , which cou ld be invo lved in contacting guanine 11. M ore

155
stru c tu ra l a n d m utagenesis d a ta w ill be requ ired to unam biguously assign
specific am ino add-base interaction to finger 1 o f W Tl.
5.4 D iscussion
The dem onstra tion th a t W T l pro tein b inds to the D N A sequences
sim ilar to a consensus site for the EGR-1 protein , and can regulate prom oters
(M adden et al., 1991; R auscher e t al., 1990) contain ing such sequences,
su g g e s te d th a t W Tl and EGR-1 p ro te in s m ay act as a n ta g o n is ts in
developm ental regulation. W e conducted a de ta iled quan tita tive analysis to
define how the affinity and sp e d fid ty of W Tl-ZF for com m on regulatory sites
w o u ld com pare w ith com peting factors such as EGRl-ZF. W e investigated
the m olecular details of the interaction by m easuring the preferences of W T l-
ZF and EGRl-ZF proteins for various buffer com ponents, as well as effects of
tem perature an d incubation tim es on the b inding effidency.
The equilibrium dissodation constants w ere determ ined to be 1.14 ± 0.2
X 10“® M for W Tl-ZF, and 3.55 ± 0.4 x 1 0 ^ M for EGRl-ZF protein , assum ing
th a t the com plex formation is the result of a sim ple bim olecular equilibrium .
Scatchard analysis had confirm ed th a t both proteins form a 1 ; 1 com plex w ith
DNA. The association constants m easured by the slope of the Scatchard p lo t
w ere in close agreem ent w ith those m easu red using the s ta n d a rd filter
b in d in g assay . The Scatchard analysis a lso confirm s th a t the p ro te in
preparations w ere 1 0 0% active.
M easuring the m onovalent salt dependence of the K& for DN A binding
ind icated tha t W Tl-ZF interaction has an op tim um betw een 0 m M an d 200
m M potassium , whereas EGRl-ZF apparently requires low er m onovalent sa lt
concentrations (from 0 to 100 mM ) for the op tim um binding . Both p ro teins
d e m o n s tra te a d ram atic d ecrease in b in d in g a ffin ity a t h ig h e r sa lt

156
concentrations. In add ition , we estim ated the num ber of cationic residues
and phosphates that in teract in each complex: W Tl-ZF pro tein forms ab o u t 7
such in teractions, w hereas EGRl-ZF m akes 6 . This num ber of phosphate
contacts exactly corresponds to the num ber of phosphates found to be directly
contacted in the EGR-1 - DNA co-crystal structure (Pavletich & Pabo, 1991).
A dditional, w eaker w ater-m ediated phosphate contacts w ere also observed in
the refined EGR-1 structure (Elrod-Erickson, 1996), w hich were not detected in
ou r analysis.
D eterm ination of p H dependence o f the DNA b ind ing activity show ed
that bo th WT1-2T and EGRl-ZF proteins can achieve op tim um binding a t a
very b ro ad p H range, declining at h igher pH. The s tu d y of the effects of
d ivalen t m etal ion concentration on D N A binding h ad show n that W Tl-ZF
achieves its optim al b ind ing at 5 mM Mg^+, while EG Rl-ZF prefers a m ore
m odest concentration of about 1 mM Mg^+ for its op tim um binding.
A num ber of therm odynam ic param eters were obtained and com pared
for the W Tl-ZF - DNA and EGRl-ZF - D N A interactions. A t 22’ C, the W T l-
ZF - DNA complex had the following therm odynam ic values: AG°= -12.1 kcal
m o l ' i ; A H ’= +6 .6 kcal mol*^, and A S ’= +63.3 cal m ol'^ deg-i. The
corresponding param eters for EGRl-ZF - DNA interaction are: AG’= -11.4 kcal
m o l 'i ; A H ’= -6.9 kcal m ol'^, and AS’= +15.3 cal m ol'^ deg The large
positive value of AS’ for the W Tl-ZF - D N A interaction indicates that it is an
en tropy d riven process, w here ordered w ater molecules and counter ions m ay
becom e d isp laced from the binding interface upon com plex form ation. In
contrast, the EGRl-ZF - DNA interaction appears to be an enthalpy d riv en
p ro cess in the te m p e ra tu re ran g e s tu d ie d . T hese d ifferences in
therm odynam ic param eters provide us w ith im p o rtan t insights in to the
m echanism s of protein-D N A complex form ation in each case. As no ted by

157
Beaudette & Langerm an (1980), entropy-driven processes are often a reflection
of h y d ro p h o b ic in te rac tions tha t occur in the sy stem , w hereas a large
enthalpic contribution usually results from the form ation of hydrogen-bond
and van d e r Waals contacts. Thus, W Tl-ZF protein contains 7 hydrophobic
am ino acids in its recognition a-helices, com pared to 2 hydrophobic am ino
acids fo u n d in the corresponding regions of EGR1-2T, which is consistent
w ith ou r find ing of a large entropie contribution to the overall free energy of
b ind ing in the W Tl-ZF - DNA com plex. Refined EGR-1 structure , o n the
other h an d , revealed a n extensive netw ork of van d e r W aals a n d w ater-
m ediated contacts, w hich occur to bo th the DNA bases and the backbone
phosphates and provide additional contributions to the binding energy. This
is also consisten t w ith the enthalpic driv ing force lead ing to the EGRl-ZF -
DNA com plex formation.
W e used a nitrocellulose filter b inding assay to determ ine the affinities
of the W T l-Z F and EGRl-ZF proteins for a num ber of DNA po in t m utants
designed to test the relative contributions of each base pa ir to the h igh affinity
binding. The crystallographic data obtained for the EGR-1 - DNA com plex has
provided a structural fram ew ork for understanding the m olecular basis of the
interaction. Since the critical amino acids in EGR-l-ZF w hich contact specific
base pairs in the EGR-1 consensus site are completely conserved in the W Tl-
ZF protein , it was reasonable to predict that the last th ree zinc fingers of W Tl-
ZF will p lay a similar role in their b inding to the 9 b p EGR-1 recognition
sequence. However, this prediction h ad to be tested considering th a t W Tl-ZF
p ro tein con ta ins a n u m b er of s tru c tu ra l differences. These in c lu d e an
additional zinc finger (the first zinc finger of W Tl) w hich does n o t possess
significant hom ology w ith any of the three EGRl-ZF zinc fingers a t the am ino
acid level (Call, K.M. et al. 1990), and the approxim ately 50 % dissim ilarity in

158
the am ino acid sequences betw een the EGRl-ZF p ro te in and fingers 2-4 of the
W T l-Z F .
In addition, crystal structural da ta available for a num ber o f other zinc
finger proteins, includ ing Tram track (Fairall e t al., 1993), SWI (Fairall et al.,
1993), and GLI (Pavletich & Pabo , 1993) reveal com plex a n d nonun ifo rm
recognition m echanism s taking p lace in different p ro tein - D N A system s.
The details of these structures are discussed in C hap ter 4. In the view of this
d iversity in the b ind ing modes observed for various zinc finger proteins, and
the s tru c tu ra l d ifferences ex is tin g betw een the W Tl-ZF a n d EGRl-ZF
p ro te in s, we m ade a detailed investigation into th e roles of in d iv idua l base
pairs in the b ind ing of W Tl-ZF and EGRl-ZF proteins.
Overall, o u r results confirm the m ain recognition them es established
in EGR-1 crystal struc tu ra l d a ta , w hile revealing add itiona l de ta ils an d
p rov id ing a quan tita tive view of th e im portance o f indiv idual bases in high-
affin ity recognition. In agreem ent w ith the crystal struc tu re , the critical
guan ines of the p rim ary strand (positions 3, 5, 6 , 7, and 9) p ro v id e the m ost
im portan t energetic contributions to the high affinity b inding for b o th W Tl-
ZF an d EGRl-ZF proteins. Substitutions at any of these positions resulted in
the loss of b inding by bo th proteins. Position 1, a lbe it show n to be involved
in co n tac t in th e EGR-1 - D N A co-crystal, p ro v id es a less im p o rtan t
contribution to the overall binding: a guanine to cytosine substitu tion at this
position resulted in a 10-fold decrease in b inding for W Tl-ZF, a n d a 5-fold
decrease for EGRl-ZF. Previous m éthylation interference studies b y Lamaire
et al. (1990) indicated a weaker in teraction made by EGR-1 at position 1 . This
sm aller contribution from position 1 to the overall b ind ing e n erg y can be
underscored by the fact that there is only one contact m ade a t th is position,
and th is arginine-guanine contact is no t stabilized by the aspartic acid side

159
chain. A sim ilar Arg-G contact made at the third position in the b in d in g site
resu lted in the loss of b in d in g by bo th protein . The ad d itio n a l w ater-
m ediated contact formed betw een Asp+2 of finger 3 of EGR-1 (or, by analogy,
finger 4 o f W Tl) com bined w ith the stab ilized A rg-A sp-G co n tac t likely
contribute to the higher energetic cost of sequence d isrup tions at th is position.
Position 2 o f the recognition sequence w as found to be im p o rtan t fo r the
binding of bo th proteins. The sequence preferences w ere observed to be in the
follow ing order: C > T » A » G. However, substitution of cytosine w ith either
purine had a m ore deleterious effect on W Tl-ZF binding, reducing its affinity
by about 100-fold, whereas the corresponding drop in EGRl-ZF affin ity was
abou t 5-fold. It is likely th a t the stereochemical na ture o f the base p lays an
im portan t role in recognition. It is not clear, how ever, w hy the a ffin ity for
W Tl-ZF w as reduced to a greater extent than that for EGRl-ZF. T he ro le of
position 2 is discussed in m ore detail in the results section.
The refined EGR-1 crystal structures dem onstrated a direct ro le for the
aspartic acid residue in finger 2. H ow ever, it w as n o t clear w h e th e r the
co rrespond ing aspartic acid residues in fingers 1 a n d 3 also co n trib u te to
specific recognition, since they did no t have a perfec t geom etry (E lrod-
Erickson, 1996). O ur m utagenesis data supports the im portance o f these
aspartic acid residues for sequence-specific recognition o f the fo u r th and
seven th bases of the b ind ing site, w hich are in the th ird and seco n d zinc
finger b ind ing subsites, respectively. Both W Tl-ZF and EGRl-ZF p ro te ins lost
their DNA b ind ing activity w hen substitutions were m ade a t these positions.
H ow ever, w e d id n 't test the im portance of position 10 for EGRl-ZF binding .
This w ould have given us an insight as to the con tribu tion of a sp artic acid
res id u e in finger 1 to E G R l-ZF recognition, and , possib ly , w o u ld have
extended the EGRl-ZF recognition sequence to 10 base pairs. W Tl-ZF show ed

160
ju st a slightly decreased binding upon introduction of a cytosine at position
10. T here is evidence from the w ork of others w hich corroborates our
findings. Thus, a sim ilar contact was seen in the TTK /D N A crystal structure:
an aspartic a d d located at the tip of finger 2 makes a good hydrogen bond to a
cytosine o n the secondary strand (Fairall e t al., 1993). Replacem ent of the
hom ologous aspartic ac id in the second zinc finger o f A D Rl p ro te in
sign ifican tly reduced p ro te in b ind ing to its DNA recognition sequence
(Thukral e t al., 1991; T hukral et al., 1992). Binding site selection stud ies
support the sam e view: only thym ine o r guanine were selected a t position 4
and 10 of the DNA sequence (Swimoff & M ilbrandt, 1995). The latter authors
also p roposed that single zinc fingers of EGR-1 as w ell as rela ted fam ily
proteins m ay specify overlapping, 4-base pair subsites of DNA. This no tion is
supported in the refined EGR-1 crystal structure (Elrod-Erickson, 1996), in the
study by Isalan et al. (1997), as well as in our study. W hile both thym ine and
guanine a t position 4 could have satisfied a contact from aspartic acid to the
secondary D N A strand (providing am ine groups of A and C), we observed a
preference for a thym ine over a guanine for the b inding of both proteins.
This preference for a thym ine at position 4 can be explained by the v an der
W aals contacts m ade by histidine +3 o f the second finger of EGRl-ZF o r the
th ird finger of W Tl-ZF. A sim ilar observation was m ade by Sw irnoff &
M ilbrandt (1995), w ho suggested that histidine +3 of EGRl-ZF contributes to
the recognition of tw o adjacent bases a t positions 4 and 5. More discussion is
p rovided in the results section of this chapter.
As d iscussed earlie r, adenine a t position 8 p ro v id es a sign ifican t
con tribu tion to the h igh affinity b ind ing of the W Tl-ZF (4.5-fold increase
com pared to cytosine at this position). In contrast, this substitu tion reduced
the b ind ing strength of EGRl-ZF by 12-fold. This dem onstrates that the two

161
pro te ins share re la ted , but n o t identical DNA recognition specificities. The
im portance of a n adenine a t position 8 o f the DNA site has been previously
dem onstra ted for W Tl protein by N akagam a et al. (N akagam a e t al., 1995).
A n add itional in teraction p ro v id ed by a g lu tam ine residue (Q352) of the
W Tl-ZF, which EGRl-ZF does no t possess, was p roposed by N akagam a et al.
(1995). The re su lts of the p rev io u s h igh affinity b ind ing site selection
experim ents ob ta ined in our lab identified the sam e phenom ena: the w ild
type W Tl-ZF p ro te in selected an adenine at position 8 w ith h igh frequency
(Borel e t al., 1996). In addition, our data dem onstrate that EGRl-ZF has a
d istinct preference (1 0 -fold higher affinity) for a cytosine versus an adenine a t
position 8 .
In conclusion , the da ta p resen ted in this s tu d y p rovides accurate
m easurem ents of equilibrium b ind ing param eters for W Tl-ZF and EGRl-ZF
proteins. Q uantitative analysis of the contribution of each base p a ir in the
DNA consensus b inding sequence to the overall b ind ing affinity w as defined
using DNA p o in t m utants. Every base pair in the b ind ing sequence was
found to contribute, w ith vary ing degrees, to the h ig h affinity b ind ing for
bo th p ro teins. T he analysis dem onstra ted im p o rtan t d ifferences in the
b ind ing m echanism s that exist betw een the W Tl-ZF an d EGRl-ZF proteins,
w ith respect to b o th the therm odynam ics, and the sequence preferences. This,
in tu rn , suggests tha t the W T l and EGR-1 pro teins m ay act o n d istinct
p rom o te r sequences, while be ing able to com pete fo r com m on regu la to ry
sites. The inform ation obtained from these m utagenesis studies ex tends our
know ledge of the mechanism s o f b inding by the zinc finger proteins. Further
stud ies into the physiological roles of these tw o regu la to ry pro teins should
prov ide valuable inform ation in to the origins of specificity, as w ell as shed

162
ligh t o n the invo lvem en t of bo th p ro te in s in n o rm al d ev e lo p m en t and
neoplastic processes.

163
CHAPTER 6.0 TH IIA IS A PROTOTYPE O F THE C2H 2 FAMILY O F ZIN C FINGER PROTEINS
6 .1 The C2H 2 zinc finger dom ain
The m ajority o f b iological processes involve specific in te rac tio n s
betw een m acrom olecular species. The specific interactions are m ediated by a
m u ltitu d e o f d istinct s tru c tu ra l dom ains th a t m ain ta in an a p p ro p ria te
stereochem ical fold. T ypically , in o rd e r for the p o ly p ep tid es to fo ld
autonom ously, they need to be a t least 50 am ino acids long.
A bout a decade ago, a novel class of dom ains consisting on average of
30 am ino acids was d iscovered . These dom ains are too sm all to fo ld
in d ep en d en tly , bu t fo ld stab ly w hen coordinated zinc ions stab ilize the
conform ation. The ability of zinc to be tetrahedrally coordinated w ith in such
dom ains, as w ell as th e lack of redox activ ity of the zinc ion, led to the
evolution o f a wide range of zinc-stabilized structural dom ains now know n
to exist.
Zinc has been know n to be an essential trace elem ent for eukaryotes for
over a cen tury (Coleman, 1992). An average adu lt hum an body contains 2.3 g
of zinc (com pared w ith 4 g of iron) (Colem an, 1992). This m akes zinc the
second m ost abundant trace m etal in eukaryotes. As new families o f zinc
containing proteins continue to emerge, specific functions can be assigned to
zinc to justify its abundance.
The first discovery of a zinc m etalloprotein involved in transcrip tion
was m ade in 1983 by H anas e t al., who dem onstrated that transcription factor
n iA from Xenopus oocytes w as a zinc p ro te in , and th a t zinc coord ination
correlated w ith its ability to b ind DNA (H anas et al., 1983). Of all the C 2H 2
zinc fingers stud ied to d a te , this unique polyfinger DNA- and R N A -binding
protein is, perhaps, the m ost extensively stud ied of all.

164
From the nucleotide sequence of the gene, K lug and cow orkers (Miller
e t al., 1985) d ed u ced tha t the am ino acid sequence of the transla ted product
could be arranged so that a pair of conserved cysteines and a pa ir of conserved
h istid ines separated by a 12 - to 13 - residue spacer defined 9 repea t series of
am ino acid sequences of 28 residues w ithin the IF lllA protein as follows: - C
-X 2-5 - C -X12-13 - H -X3-4 - H - , w here X - any am ino acid. There was an
add itional pair o f conserved amino acids found w ith in the X12-13 spacers: an
arom atic am ino acid, F or Y, and a branched aliphatic amino a d d , commonly
L. The pairs o f C and H residues w ere proposed to tetrahedrally coordinate
Z n 2 + ions.
The term " zinc finger" was coined by M iller e t al. (1985) w ho proposed
th a t the 12 - 13 -residue loop or "finger" form ed the DNA b in d in g surface
(Figure 6.1 A). The m odel of tetrahedral Zn^+ coordination w as confirm ed by
ex tended X-ray absorption fine structure (EXAFS) w hich show ed that the zinc
abso rp tion edge in TFULA was fit to a tetrahedral coordination sphere w ith
tw o sulfur and tw o nitrogen atoms as ligands (D iakun et al., 1986). The zinc is
bo u n d to conserved histidine atom s at 2 À distance, and to cysteine SY
atom s a t 2.3 Â distance, w ith an affinity varying from 10^^ M to 10*^2 ^
depend ing on a particular zinc finger sequence (D iakun et al., 1986).
The sto ichiom etry of zinc m etal calculated for TFULA show ed it to
con tain 7 to 11 zinc atom s, which w as more consistent w ith the 9 zinc fingers
o f TFULA than the originally reported 2 zinc ions per IFlllA m olecule (Miller
e t al., 1985). Proteolysis studies, revealing periodic interm ediates of a limited
d ig est coupled w ith the analysis o f cDNA sequence of TFULA (M iller et al.,
1985) supported the general idea of a repeated structural unit, each of which
b o u n d a single zinc ion. Model structures for the zinc finger m o tif have been
p u t forw ard by Berg (1988) and Gibson et al. (1988). The first N M R structure of

165
C y s H is
ZnHisCys
8 Sx
F ig u re 6.1 Schem atic rep resen ta tion of a C z H z z in c f in g e r (Evans &
H ollenberg, 1988).

166
a zinc finger w as determ ined by Parraga et al. (1988) an d Lee e t al. (1989) and
resem b led m ore closely the m odel o f Berg. S ince th en several NM R
structures for zinc fingers have been published (Klevit e t al., 1990; Om ichinski
e t al., 1990, 1992; K ochoyan et al., 1991a, b; N euhaus, e t al., 1992). M odel
bu ild ing , em ploying a n insightful u se of the s tru c tu ra l da ta bases, two-
d im ensional nuclear m agnetic resonance analysis and X-ray crystal structures
have separately suggested an approxim ate structure for these zinc fingers.
T he struc tu re o f each of the zinc finger d o m ain s consists of two
an tiparalle l 3 -strands an d an a-helix surrounding a centrally bound zinc ion
(Figure 6.1 B). Three-dim ensional structures now availab le for a n um ber of
zinc fin g er p ro te ins v e ry nicely accom m odate th e conserved sequence
features. T he cysteine an d h is tid in e side chains are involved in zinc
c o o rd in a tio n , and the th ree o ther conserved re s id u e s pack to fo rm a
h y d ro p h o b ic core ad jacen t to the m etal coord ination un it. The use of
hy d ro p h o b ic am ino ac id s by zinc fingers to s tab ilize local zinc p ro te in
struc tu re d iffers from the folding stra tegy of m etallo thioneines w hich lack
arom atic side chains.
As w as p roposed earlier (Berg, 1988), the a -h e lix in a zinc finger is
directly invo lved in sequence-specific recognition, in te rac ting in the m ajor
groove o f the DNA. T hus, the zinc finger p rov ides yet another w ay of
inserting an a-helix - the m ost com m on m ode of D N A recognition - in to the
DNA m ajor groove.
The overall fold of an ind iv idual zinc finger has been show n to be
m ain ta ined entirely by the zinc coord ination as w ell as the packing o f the
hydrophobic firamework residues. M ichael et al. (1992) h ad used a m inim alist
finger pep tide consisting of 26 amino a d d s , 16 of w hich w ere alanine residues.

167
a n d th u s prov ided d irect evidence for the im portance of zinc coordination
a n d the central hydrophobic core for m aintaining the tertiary fold.
O nce folded, zinc fingers are very stab le structures, d isp lay ing little
in te rn a l m otion (Palm er et al., 1991), and resistance to p ro tease d igestion
(M iller e t al., 1985) o r thermal dénaturation (Frankel et al., 1987). Zinc fingers
iso la ted under m etal chelating conditions d isp lay no sequence specific DN A
b ind ing ability (Frankel et al., 1987; Lee e t al., 1991a). Therefore, the tertiary
structu re of a protein is inseparable from its function.
Significant variation exists w ithin the zinc requiring p ro te in fam ilies,
particu larly w ithin the Cys2H is2 class of zinc finger proteins. The differences
include such structural features as the natu re and positioning of hydrophobic
core residues, the spacing betw een the coord inating cysteine an d h istid ine
residues, the sequence of the interfinger linker regions, or th e num ber of 13-
strands in the zinc fingers. Two proteins, SWI5 and Tram track, deviate from
the classic PPa zinc finger fold. The am ino term inal finger of SWI5 has an
irregu lar P-sheet consisting of three strands as opposed to the canonical tw o
(N akaseko, 1992; N euhaus, 1992). Interestingly, folding of finger 1 appears to
be dependen t on the presence of the additional strand. A nother example of a
th ree -s tran d ed P-sheet was discovered in the am ino-term inal finger o f the
Drosophila transcrip tion repressor pro tein T ram track (Fairall e t al., 1993).
The th ird strand, w hile essential for D N A -binding activity, m akes no d irect
con tac t w ith DNA in a co-crystal s tru c tu re , suggesting it is som ehow
im p o rtan t for m aintaining finger structure. It rem ains to be seen why these
SWI5 and Tram track fingers differ from other know n C2H 2 fingers.
The extent o f zinc finger occurrence in eukaryotes is astonishing. It
w as estim ated that as much as 1% of the D N A in hum an cells specifies zinc
fingers (Rhodes and Klug, 1993). In chrom osom e 19 the figure is as h igh as

168
8 % (Rhodes and Klug, 1993). The zinc-finger containing proteins th a t have
been identified so far carry firom as few as two zinc fingers (M IGl), found in
yeast, to as m any as 37 tandem fingers (Xfin) in Xenopus. Well over 1300
zinc finger cDNA sequences have now been reported , suggesting tha t the
C2H 2 zinc finger dom ains represent the m ost abundant DNA b ind ing motif
in eukaryotic transcription factors.
T hese tran scrip tiona l regu la to rs are invo lved in a m u ltitu d e of
functions, such as differential gene expression [MIGl (yeast), MBP-1 (human)],
developm ental gene regulation [K ruppel and T ram track from Drosophila ,
EGR-1, m Kr2, Krox-20 from m ouse, Xfin, p43 and TFIIIA from Xenopus,
tum or su p p re ss io n [W T l (iden tified in hum an a n d m ouse)], hum an
oncogenesis [GLI (hum an)], ubiquitous transcriptional regulation [Spl], to
name b u t a few.
The term "zinc finger" is som etim es loosely app lied to other classes of
zinc containing proteins, and has been used to refer to to almost any sequence
that has a set of cysteines a n d /o r h istid ines w ith in a rela tively short
polypeptide chain. These other classes of proteins include large fam ilies such
as the nuclear horm one receptor superfam ily (Evans, 1988), the RING finger
protein fam ily (Freemont, 1993), w hich includes the breast and ovarian cancer
susceptibility gene BRCA 1 (Miki et al., 1994) and the retroviral CCHC motifs
rep resen ted by HIV-1 nucleocapsid p ro te in (Blake a n d Sum m ers, 1994).
Recently, an NMR structure of hum an transcriptional elongation factor iFilS
has been reported (Q uian e t al. 1993a). This small dom ain binds nucleic add
and consists of a three-stranded P-sheet. The zinc is coordinated tetrahedrally
by four cysteine residues. This type of structure is novel am ong the zinc-
b inding an d nudeic ad d -b in d in g dom ains and thus represents a further
striking exam ple of d iversity of zinc-containing motifs. A dditionally, other

169
proteins have been show n to contain structural zinc ions, such as th e tum or
suppressor p53 (Cho e t al., 1994) and the hum an g row th horm one-prolactin
receptor complex in w hich the zinc form s a bridge betw een the horm one and
the receptor (Somers e t al., 1994).
To give an apprecia tion for the diversity of zinc-b ind ing m otifs, the
schem atic rep resen ta tion in Figure 6.2 show s zinc m odu les from various
p ro te in fam ilies th a t d iffe r in m eta l coord ination s tra teg y , secondary
structure, and m odularity , giving rise to distinctly different global folds. Each
of these m olecules exhibits nucleic acid recognition strategies that a re unique
am ong the zinc m odules.
From an evolu tionary point of view , it could be a rgued tha t the C2H 2
class evolved from a single prim ordial ancestor, and tha t m ultifinger proteins
are the resu lt of in ternal gene duplication. This hypothesis is su p p o rted by
the conserva tion of the z inc-coord inating , h y d rophob ic and H / C link
residues, as well as the fact that m ost zinc fingers are encoded by separate
exons (six o f nine IF lllA fingers are localized to separate exons (Tso e t al.,
1986)).
To rationalize the evolution o f zinc fingers from the point o f view of
added d iversity , one m ig h t argue th a t the zinc fingers have o rig inally
evolved as sim ple repeating motifs th a t could interact w ith RNA m olecules
(w hich inhab ited the prebio tic soup before the D N A ), and w h ic h later
evolved to interact w ith DNA. Thus, the ability of IF lllA to in te rac t w ith
both DNA and RNA suggests that perhaps it is an exam ple of a n original
nucleic acid binding protein.
The reason for link ing several zinc finger un its together to create a
functional p ro tein can be underscored by the fact that iso lated single fingers
do not recognize DNA sequences specifically (Parraga et al., 1988). For

(i) (h) 170
(cl
(d)
Figure 6 .2 R epresentative structu res from various zinc finger fam ilies,
illustrating structural d iversity am ong zinc-binding and nucleic acid-binding
dom ains (Schm eideskam p & Klevit, 1994).

171
Figure 6.2 (a) EGR-1 finger 2 of C2H 2 type; (b) the GAL4 Z n2Cô b inuclear
cluster; (c) the glucocorticoid receptor D N A -binding dom ain; (d) the am ino-
term inal retroviral-type CCHC finger of HIV-1 nucleocapsid protein; (e) the
carboxy-term inal m etal-binding dom ain of GATA-1 .

172
exam ple, in a three-finger protein SWITCH 5 (SWI5) tw o linked fingers h ad
to be p resen t for the protein to be able to interact w ith DNA w ith a reasonable
affinity (N akaseko et al., 1992). By then applying NMR to th e first two zinc
finger m otifs of SWI5, it was confirmed that adjacent zinc fingers do not m eld
w ith each o ther, th a t they are tru ly independen t "reading heads" joined
together by flexible linkers.
T h is m o d u la r nature of zinc finger p ro te ins is u n iq u e am ong the
classes o f DNA b ind ing proteins. From an evo lu tionary p o in t of view ,
p roducing a pro tein w ith m ultiple fingers could be advantageous for several
reasons: such a protein could recognize longer stretches of D N A (im portant
for an o rgan ism w ith a larger genom e size); such a transcrip tional regulator
could b in d DNA w ith higher specificity, and thus p ro d u ce m ore stable
interaction; finally, the specificities of individual fingers cou ld be com bined,
giving rise to new binding specificities.
W h en sp e ak in g of "m odularity" o f z in c fin g er p ro te in s , the
independence of each zinc finger structure doesn 't always im ply that finger-
finger interactions do not exist. For m ost zinc finger proteins exam ined, there
are a few interfinger interactions taking place, and in general they are no t
conserved betw een different proteins.
F or exam ple, in the EGR-1 crystal s tru c tu re there a re conserved
hydrogen bonds observed betw een zinc fingers 1 and 2 w here an arginine
located in the C-term inal portion of an a-helix o f the first finger hydrogen
bonds to the backbone carbonyl of a serine located in the tu rn betw een the P~
sheet an d the a-helix of the next finger. A sim ilar in teraction is seen for
fingers 2 and 3, w here arginine hydrogen bonds to the backbone carbonyl of
an alanine residue (Pavletich & Pabo, 1991). A sim ilar p a tte rn o f interfinger

173
in teractions is observed in an N M R structure o f MBP-1 p ro te in (Om ichinski
e t aL, 1992): the tw o zinc fingers interact through hydrogen b o n d in g of amino
acids located in the same regions of the zinc fingers as se en in EGR-1 (C-
term inal region of an a-helix of a first zinc finger and the "tip" reg ion of the
nex t finger) (Figure 6.3). The GLI - DNA co-crystal structure has revealed that
b o th hydrogen bon d and hydrophobic interactions occur, especially betw een
zinc fingers 1 an d 2 (Fairall e t al. ,1993). As m entioned earlie r, an NMR
stru c tu re of SWI5 (Nakaseko e t al., 1992) revealed no in te rfin g er contacts.
H ow ever, a possibility exists th a t the adjacent zinc fingers m ay com e into
contact w hen com plexed w ith DNA.
A lthough the focus of th is overview is the structure a n d function of
zinc dom ains, it should be kept in m ind that DNA binding is on ly the initial
step in gene activation/repression . The m ajority of the p o ly p ep tid e chain in
th e n a tiv e z in c -co n ta in in g tran sc rip tio n fac to rs is ta k e n u p by the
tran sac tiv a tin g dom ains w hich are involved in in te rac tio n s w ith other
p ro te in s , m ost no tab ly the com ponen ts o f the basa l tra n sc rip tio n a l
m achinery. Figure 6.4 shows the schematic representations o f com plete zinc-
contain ing transcrip tion factors.
6.2 S tructure/function analysis of TFIIIA
6.2.1 Purification and characterization of TFIIIA gene product
The fact th a t TFIIIA represen ts one of the best s tu d ie d transcrip tion
fac to rs is, p e rhaps, best exp lained by its g rea t abundance in a natu ra lly
occurring system : Xenopus oocytes, which contain approxim ately 0.7-1 x 10^2
m olecules of TFULA per cell (Shastry et al., 1984). In the cy top lasm , TFULA
p red o m in an tly ex ists as a com ponen t of a 7S rib o n u cleo p ro te in particle,
w hich is com posed of one m olecule of TFULA and one m olecule of 5S rRNA

174
n:,
Figure 6.3 Schematic representation o f the in terfinger orientations o f (A)
MBP-1 zinc fingers and (B) EGR-1 fingers 1 and 2 (Omichinski et al., 1992).

175
Spl
TFIIIA
N [
1
Q-rich
344H i~ tn tn m ïm c
DNA Binding K-rich 9 Zinc-fingers
696 Z] C
DNA Binding 3 Zinc-fingers
1 421 486 7771 CGlucocorticoid N I i f
Receptor Modulates Transcription f Hormone BindingDNA Binding
1 53 120Thyroid Hormone Receptor
4101 C
GAL4
I Hormone Binding DNA Binding
1 62N
881H e
t Transactivating Domain (Negatively charged) DNA Binding ZnaCe Cluster
Figure 6.4 Diagram s of the functional dom ains of eukaryotic zinc-containing
transcription factors (Coleman, 1992).

176
(Picard & W egnez, 1979). U sing conventional chrom atography, TFIIIA was
purified to homogeneity (Picard & W egnez, 1979). O nly later, w hen it was
show n tha t TFIIIA could specifically b ind DNA (Engelke e t al., 1980) was it
realized tha t one of the roles o f TFIIIA is that o f transcrip tional regulation
(Engelke et al., 1980; Pelham & Brown, 1980; H onda & Reeder, 1980). O ther
m ethods for TFIIIA purification have been developed , and include density-
gradient centrifugation followed by DEAE cellulose chrom atography (Smith
et al., 1984), ion exchange chrom atography (R om aniuk, 1985), as well as
expression of recom binant TFIIIA in E.coli (Del Rio & Setzer, 1991).
The Xenopus laevis TFIIIA is a 344-am ino acid, 38.5 kD a protein ,
con ta in ing nine C 2 H 2 zinc finger motifs located a t the am ino term inus
(Brown e t al., 1985; M iller et al., 1985). TFIIIA p ro te in is p ro p o sed to be an
extended rather than a globular p ro te in (Reynolds & Gottesfeld, 1983; Hanas
et al., 1984) based on its m ode of DNA binding. First, TFIIIA b in d s a long
stre tch of DNA w ith o u t com pacting it in to a tigh ter con fo rm ation or
significantly changing the DNA linking num ber (Reynolds & G ottesfeld, 1983;
Hanas e t al., 1984). Second, proteolytic fragments o f TFIIIA reta in their ability
to b in d specific DNA regions, w hich suggests th a t TFIIIA m u st be in an
extended conform ation w hen in teracting w ith DNA, to be able to cover the
long internal control region (Ginsberg et al., 1984; M iller et al., 1985).
The amino acid sequence o f the D N A -binding dom ain o f TFIIIA is
show n in Figure 6.5, and a schem atic diagram show ing TFIIIA alignm ent
along the ICR is given in Figure 6 .6 , where the 30 kDa N -term inal DNA-
b inding dom ain, and a lOkDa C-term inal transcrip tional activa tion dom ain
are indicated (Smith e t al., 1984). Am ino acids 294 to 313 (Vrana e t al., 1988),
and 284 to 297 (Mao & Darby, 1993) w ithin the C-term inus have been show n
to be im portant for the transactivational function o f TFIIIA. This reg ion is

177
I :i I V :(i
f U C F I K A L P V V Y K R )12
^ ® r r f ! > s F * n(F>r. A a ( y ) K K K y K ^ Q
r n ^ fC • pm pfciK e E p ( c U - K - £ ( ^ ^T c p K ' NrmTfr)P î> p g(ç)p f p ^ T T !W ( K [ c F F K ( ^ g K A 0 K K P N 9CÇU -
o n f • ° fr)F ® P « ^A C - - - 0 P ® K K D_D S © S © V G K T y t Q Y
0 0 - - • L A Y © - - D v©N R K © p H K P Y © RE K E p T v t f r . r ^ P R n 0 © p P ? © T T A F
E E
A ® L C
R @ S . . L -
K ® 37
Z ® 67
K®F_N_ V ® S _ L
R F ® 98
• ? ® 129_a® £ K
L K ® V A
• 0 ® Q K ' S @ L g
277( D P E K R K L - K E K C P R P K R S L A S R L T C Y I P
S V S C T E K T O S L Y K N K P S C T E T f f i l G S L V L D
P K S K E
K L T [ Q
• Y ® 159
E Q ® 188
• T ® 214
S F ® 246
. Y ® 276
K1H)A 311
) 344
Figure 6.5 Amino acid sequence of the nucleic acid binding dom ain of TFULA
(Miller e t al., 1985).

178
Herna/ Control
Carboxyl Amino
Figure 6 .6 Schem atic diagram illustrating a lignm ent of TFULA. zinc finger
region along ICR (Miller et al., 1985).

179
rich in lysine residues w hich are proposed to m ediate interactions w ith other
com ponents of the transcriptional m achinery (Smith e t al., 1984). Potential
N -linked glycosylation sites were found in the C -term inal reg ion of TFluA
(M iller e t al., 1985). H ow ever, the role of this post-translational m odification
is no t ye t clearly determ ined. The elucidation of the zinc finger structure of
the TFIQA D N A -binding dom ain is described in the previous section of the
chapter.
6 .2 .2 TFIIIA gene expression
Xenopus laevis contains a single copy of the gene encoding TFIIIA per
hap lo id genom e (G insberg e t al., 1984; Taylor et al., 1986, Tso e t al., 1986;
Scotto e t al., 1989). The TFUIA gene is approxim ately 11 kilobasepairs in
length an d contains n ine exons separated by eight introns (Tso e t al., 1986).
The relative position of exons and introns, as well as the fact th a t 6 ou t of 9
zinc fingers of TFIIIA are encoded by separate exons suggest th a t TFIIIA
evolved from a prim ordial ancestor specifying a zinc finger m otif (Tso et al.,
1986). The TFIIIA prom oter was found located 5' to the transcrip tional start
site, an d has features characteristic of m any polymerase II transcribed genes
(Tso e t al., 1986). A consensus TATA box sequence is a t position -32, and a
CAAT box sequence is a t position -96 (Tso et al., 1986). Further regulatory
sequences include seven positive an d four negative cis-acting elem ents,
identified by deletion studies (M atsum oto & Korn, 1988; Hall & Taylor, 1989;
Scotto e t al., 1989; Pfaff e t al., 1991; Pfaff & Taylor, 1992).
The levels of TFIIIA mRNA and protein reach their m ax im um in the
early stages of oogenesis (approxim ately 10^2 m olecules/cell), a n d drop to
about 4x105 m olecules per cell in later stages of a ta ilbud em bryo (Shastry et
al., 1984; Andrews & Brown, 1989). Since the expression of TFULA is tightly

180
developm entally regulated (Andrews & Brown, 1987), bo th housekeeping and
stage-specific transcrip tion factors are expected to in te rac t w ith the TFULA
prom oter (Pfaff e t al., 1991). A protein th a t binds to one of the positive
regulatory sequences w as identified and show n to recognize the core sequence
CACGTG located at positions -269 to -264 (Scotto et al., 1989). This p ro te in ,
term ed th e d istal e lem ent factor (TDEF), is hom ologous to the ad en o v iru s
major late transcrip tion factor, which belongs to the helix-loop-helix class of
transcrip tional regulators (Sawadogo & Roeder, 1985; H all & T aylor, 1989;
Scotto e t al., 1989). The TDEF protein occurs in both oocyte and som atic cells.
H ow ever, it w as show n to regulate transcrip tion only in oocytes, suggesting
that it acts in a cell-specific fashion (Hall & Taylor, 1989).
A dditional level o f complexity in TFIIIA gene expression is in troduced
by the dem onstra tion th a t the oocyte a n d som atic form s of TFIIIA are
expressed from two d ifferen t, overlapp ing p rom oters, u tiliz ing d is tin c t
transcriptional start sites (Kim et al., 1990; M artinez et al., 1994). M oreover, it
was recently dem onstrated that both RNA polymerases II and IE are involved
in TFIIIA gene expression in somatic cells (Martinez e t al., 1994). H ow ever,
the significance of the involvem ent of the tw o RNA polym erases is n o t yet
u nders tood . Even th o u g h rem arkable progress has been m ade to w ard
understand ing the regulatory mechanisms of TFIIIA gene expression, several
questions still remain, such as the roles of other transcription factors, the fine
details of the differential regulation of som atic versus cxDcyte TFIIIA genes, as
well as the role of chrom atin structure.
6.2.3 Roles o f TFIIIA in transcrip tion and storage of 5S rRN A
T ranscrip tion factor HLA has a d u a l role as a positive tran sc rip tio n
factor requ ired for transcription of 5S rRNA genes (Honda & Roeder, 1980), as

181
well as a sto rage pro tein for 5S rRNA (Engelke e t aL, 1980). A ssociation of
TFIIIA w ith the 5S rRNA also serves to transport 5S rR N A from the nucleus
in to the cy to p lasm w here it is stored until it is re q u ire d for ribosom al
assem bly in im m atu re oocytes (G uddat et al., 1990). The b ind ing of TFIIIA to
the 5S rR N A gene nucleates an o rdered assem bly o f a tra n sc rip tio n
p rein itia tion com plex (Segall e t al., 1980; Shastry e t al., 1982; Bieker e t al.,
1985). First, TFUIA binds to the internal control region o f 55 rRNA gene, an d
form s a m etastab le complex (Engelke et al., 1980; Bieker & Roeder, 1984).
N ex t, TFIIIC b in d s and stab ilizes this com plex (L assa r e t al., 1983).
Subsequently, iF lilB binds (Bieker e t al., 1985; Bieker & R oeder, 1986), a n d is
responsib le fo r m axim al transcrip tion rates (K assavetis e t al., 1990). The
p re in itia tion com plex is very stable, and persists th ro u g h m any cycles of
transcrip tion by RNA polymerase HI (pol HI) (Bogenhagen e t al., 1982; Setzer
& Brown, 1985). Transcription of the 55 rRNAs from Xenopus has becom e a
m odel system for the study of the m echanisms that con tro l eukaryotic RNA
synthesis.
In e u k a ry o tic cells, RNA polym erase lU is re sp o n s ib le fo r the
transcrip tion o f genes encoding 55 rRNA, tRNA, severa l cytoplasm ic and
nuclear RNAs (75 K and L, 4.5 5 RNAs) and som e v irus-associated RN A s
(adenovirus-associated VAI and VAU, EBERI an d EBERII) (Ciliberto e t al.,
1983a). The m ost striking comm on feature shared by the genes transcribed by
pol m (w ith several exceptions) is that prom oters are con ta ined in the coding
sequence (5akonju e t al., 1980; Bogenhagen e t al., 1980). O ther sim ilarities
include the fact th a t Pol m genes are quite sho rt co m p ared to the average
leng th of pol I o r pol U genes (Ciliberto e t al., 1983); the cod ing sequence is
defined at the 3' end by at least one cluster of T residues, w h ich are requ ired
for efficient transcrip tional term ination (Bogenhagen & B row n, 1981).

182
The binding of TFIIIA to 5S rRNA forms a 7S ribonucleoprotein (RNP)
particle w hich serves to stabilize an d transport the 5S RNA m olecule (G uddat
et al., 1990). Upon expo rt from the nucleus to the cytoplasm , th e ribosom al
p ro tein L5, which also binds 5S rRNA, displaces TFULA from its interaction
w ith 5S rRNA, and the complex of L5 and 5S rR N A is inco rpo ra ted into a
ribosom e in the nucleolus (W orm ington, 1989).
6.2.4 S tructure and function of 55 rRNA and its gene
The 5S rRNA is an essential com ponent in all ribosom es, prokaryotic
and eukaryotic , except those from m am m alian m itochondria (Kjem s e t al.,
1984). The first discovery of ribosom al 55 rRNA w as m ade in 1963 (Erdm ann
et al., 1987). Since then , their s tru c tu re and function have b e e n stud ied
extensively. The ribosom al RNA com ponents are am ong the m o st highly
conserved m acrom olecules in all liv ing systems. Sequence com parisons of a
large num ber of 5S rRN A s isolated from a wide range of organism s revealed
a h igh deg ree of s tru c tu ra l conservation th ro u g h o u t ev o lu tio n (H ori &
O saw a, 1987; Erdm ann e t al., 1987; W olters & E rdm ann, 1988). D ue to this
high conservation, the sequences of 5S rRNA genes an d their transcrip ts were
used to m easure phylogenetic distances between d istan tly related organism s.
T here are three classes of 5S rRNA genes: m ajor-oocyte (Xlo), trace-
oocyte (Xlt), and som atic (Xls) that are transcribed (Fedoroff & Brow n, 1978;
Peterson e t al., 1980). The haploid Xenopus genom e contains approx im ate ly
2 0 ,0 0 0 copies of the major-oocyte type genes w h ich are transcribed only in
oocytes , and about 400 copies of som atic 5S rRNA genes that a re transcribed
in b o th oocyte and som atic cells (Peterson e t a l., 1980). T h ere are six
nucleotide differences betw een the som atic and th e oocyte g en e sequences
(Xing & W orcel, 1989). Trace oocyte genes are represen ted by approxim ately

183
2 ,0 0 0 copies per haploid genome, and combine som e features of the oocyte-
type genes, as well as som atic type (Brown et al., 1977). In addition, there are
approxim ately 24,000 copies of 5S rRNA pseudogenes per haploid genom e
that have no know n function since they do not p roduce a transcript (Fedoroff
& Brow n, 1978). The organization of the Xenopus 5S rRNA genes is show n
in F igure 6.7. The main components of each gene copy are the GC-rich region
w hich includes the gene and the pseudo-gene sequences, and a variable AT-
rich block, called the spacer DNA (Miller e t al., 1978; Fedoroff & Brown, 1978).
Based on the extensive sequence com parisons, a consensus m odel for a
5S rR N A m olecule has been proposed, an d is show n in Figure 6 .8 (Delihas &
A ndersen , 1982). These molecules are 120 nucleotides long, have a m olecular
w e ig h t of abou t 40 kDa, and assum e a Y-shape com posed of five helical
reg ions (I th ro u g h V), an d five loop dom ains (designated A th ro u g h E)
(Delihas & Andersen, 1982; Kiintzel et al., 1983; Delihas et al., 1984).
6.2.5 Role of TFIIIA zinc fingers in RNA recognition
The dua l function of TFIIIA as a positive transcrip tion factor an d a
storage p ro te in involved in regulation o f 5S rRNA gene expression requires it
to b in d to tw o different nucleic acids - DNA and RN A (Engelke e t al., 1980;
Pelham & Brown, 1980). TFIIIA interacts w ith DNA and RNA in a m utually
exclusive m anner (Pelham & Brown, 1980). Except for the W ilm s' tum or
p ro te in (Caricasole et al., 1996), TFIIIA is the only o ther C2H 2 class pro tein
capable of interacting w ith both DNA an d RNA. In parallel to the s tu d y of
TFUIA-5S DN A interaction, the RNA b ind ing function of the factor has been
the subject of intense investigations.
TFIIIA - 5S rRNA equilibrium b ind ing was characterized by R om aniuk
(1985). The factor binds 5S rRNA with an apparent association constant of 1.4

Xlo"J
184
— ' — ($00 ir» inOO bp
Xls -a-4TOb;.
Xlt ■ a— a — a310 bp
Xbo1000 to 2000 bp
X b s 1 I-850 bp
Figure 6.7 O rganization of the Xenopus 5S rRNA genes. Pseudogenes are
designated by a Y' (Kom, 1982).

185
rUUU c GG AU G c u G ^ GG U G G G A C CG G A C U (- u -.» ... UOG C « * U a G a a U a *
G» C “G» C
V A» U C» G »C» G A» U U» U A o A
- s:f®G«C G*C
U U “C»GC*GI AG» U
IV c » G C'G
" A G ®G A
Figure 6 .8 The secondary structure o f 5S rRNA (Delihas & A ndersen , 1982).
5S rR N A complex exhib its a b road p H op tim um , a n d is fav o red by both
en tro p y and enthalpy (Romaniuk, 1985).

186
X 10^ under the conditions of 0.1 M salt, p H 7.5 and 20®C. The affinity of
the nonspecific in teraction w ith tRNA^*^®, on the other hand , is at least two
o rd e rs o f m agn itude low er th an for the 5S rRNA. M utagenesis stud ies
covering the entire 5S rRNA m olecule com plem ented by nuclease digestion
and hydroxyl radical footprinting cum ulatively suggest the following features
of TFIIIA - 5S rRN A interaction (A nderson & Delihas, 1986; Baudin et al.,
1989; C hristiansen e t al., 1987; Pieler et al., 1984; Rom aniuk, 1985; Romaniuk
e t al., 1988; Rom aniuk, 1989; B audin et al., 1989; You & Rom aniuk, 1990; de
Stevenson et al., 1991; You et al., 1991; McBryant e t al., 1995; Z hang et al., 1995;
R aw lings et al., 1996): alm ost all of the 55 rRN A m olecule is involved in
TFIIIA interaction, especially helices H, IV & V and loops A & E; the binding
of TFIIIA is m ost notably a ltered by changes in the h igher o rder structure,
ra ther than base-specific contacts. The im portan t role of RN A conformation
for TFIIIA recognition is also ev iden t from the experim ents using 55 rRNAs
iso la ted from d iffe ren t species (H anas e t a l., 1984; P ie le r e t al., 1984b;
R om aniuk, 1985; A ndersen & Delihas, 1986). TFIIIA has com parable affinities
for various eukaryotic and bacterial 55 rRNAs, supporting the notion that the
o v e ra ll th ree -d im ensiona l fo ld of 55 rR N A m olecu les is the m ajor
de term inan t for specific interactions.
5ince TFIIIA interacts w ith tw o different nucleic acids, it w asn't clear
w he ther DNA and RNA interactions are m ed iated by d ifferen t zinc fingers,
and w h a t features o f ind iv idua l zinc fingers w ere responsib le for th is
d ifferen tia l activity. Hence, considerable effo rt w ent in to defining high-
affin ity RN A -binding fingers of TFIIIA. It w as soon found that while the
am ino-term inal three zinc fingers p redom inate in DNA bind ing , the central
zinc finger trip let - fingers 4 to 7 - p rim arily m ediate high-affinity RNA
b ind ing (Theunissen et al., 1992; Clemens e t al., 1993). In fact, a polypeptide

187
consisting of fingers 4 to 7 has been show n to have h igher affinity for 5S
rRNA than the full length pro tein (McBryant e t al., 1995). Sim ilarly, Setzer et
al. (1996) reported tha t polypeptide containing zinc fingers 4 to 9 bound 5S
rR N A w ith a tw o-fo ld higher affinity than the fu ll-length p ro tein . This
suggests that fingers 4 to 7 con ta in the necessary in fo rm ation for RNA
recognition, as w ell as that there is a certain degree of interference between
the zinc fingers. Com bining the available RNA m utagenesis d a ta (Sands &
Bogenhagen, 1987; You et al., 1991; Theunissen e t al., 1992) w ith the results
using truncated TFIIIA m utants Clemens et al. (1993) p roposed a m odel in
w hich ind iv idual fingers are assigned to d istinct structural e lem ents of 5S
rRN A (Figure 6.9). A peptide containing zinc fingers 5 to 7 w as show n to
have a 30-fold low er 5S rRNA affinity than the one w ith zinc fingers 4 to 7,
ind ica ting an im p o rtan t con tribu tion for finger 4 (Clem ens e t al., 1993).
M utagenesis of finger 4 resulted in a four-fold reduction in affinity relative to
the w ild-type protein , while m utations in finger 6 show ed a 30-fold drop in
b ind ing affinity (Clemens et al., 1993). Perhaps, the m utation in troduced in
finger 4 involved a residue (G lnl21) which m akes nonspecific backbone
contacts, whereas a threonine (Thrl76) in finger 6 m ay partic ipate in a base-
specific contact. This w ould exp lain the difference in the m agn itude of
affinity change for the two m utations. It is in teresting to p o in t ou t that aU
three m utations in troduced in finger 6 p roduced significant reduction in
b ind ing affinity. These m utations were made in the am ino ac id trip let TWT
w hich are the surface residues o f finger 6 (Clemens e t al., 1993). This triplet
sequence is conserved in the sam e position in finger 6 of an o th er 5S rRNA-
b ind ing protein, p43 (Joho, 1990). Like TFIIIA, p43 protein contains nine zinc
fingers, seven of w hich are the sam e size as their counterparts in TFIIIA (Joho
et al., 1990), (Figure 6.10). In addition, unlike TFULA, p43 only b inds to 55

188
10S ' G C C U A C O G C
r u u u c a o AUQCu a
rfS zf720
C
Q O
a c c c u g
U O O Q A C120
100
A UC - G C G A U
60
G C - C U G A CC O G A C U 40
zf8-9
70zfS
U*U A*A A - GG* U
G C U U 8 0 C - G C - G 1 A G* U C - G C - G
9 0 A G G A
r f 4
zf1-3
Figure 6.9 Model for the TFIIIA - 5S rRNA interaction. P roposed zinc finger
positions are indicated. N ucleotides enclosed in boxes ind icate th a t m ost
im portan t regions for recognition (Clem ens e t ai., 1993).

189
MM' TFUIA
nCrtUCf r CPC,nCECHL?UUVCHV
r c o t L^gpmmcRKAFYACEGCLOOGnACg U V C A V l U $ f A U Q G A A Ÿ M C M U C L O f Æ - C c f l
LcffluCHfflrRQSLrfl
S E O C P U c f l o i C C
r c E C P F P f i c f E C
ÎCUFAACAOILq [CCFrSLHHL
L A L C C L S f iP r f lG ^ n rF S rC C S L S f^ L V C j j j
rcECMF r g o s o c S o L R F r rc flH n c cQ F H R F f l
CEflUPLCgFUPCQCRSFHCCRRLRRÆSUffl
« I Cl CUVUgKFEf^CRFCCHMOLCuflof S 0
SMEPLSufiDUPCQsUCSSSURCLUfl^CRffl
r O O L P Ÿ E ^ K E C & C R F S L P S R L C R œ t l Æ
RC Ÿ ^ Y E C ÿ l TUSP r U fflLQ rffluccfl
RC Y Pg^O O SQ SFU C C T urL Y LC flu f lEC Q
PLELOQRR&KPFCCRSALRR&ATg 0 OL Ru§3 o Q h rc f RHCO VL Rofluc r f l
RCCPLOLPRPAQOROCrFSSUFMLTHgbACLg
E CERruYLgPROCgORSY T f RFKLRsfll OSFfl
LCLO rHRflPHSCflTRSFRnAESLLRflLUufl
EEORPFugEHRCQGCCFRnCCSLERQsUuQ
OPERCCLCLCOPECRCLCEC
Elofler
Figure 6.10 A lignm ent of zinc finger sequences of TFIIIA and p43 proteins.
The sh a d e d am ino acids re p re se n t the z inc-coord inating cyste ine and
h istid ine residues (Joho et al., 1990).

190
rRNA, a n d has no DN A-binding ability. There is, how ever, only a lim ited
am ino acid sequence iden tity (33%) shared by the tw o proteins. This am ino
acid id en tity extends to the consensus zinc ligands (C & H), hydrophob ic
am ino acids (Miller e t al., 1985; Brown et al., 1985) an d the TWT m otif in
finger 6 Qoho et al., 1990). The fact that only structure-determ ining am ino
acids are conserved betw een the two proteins suggests tha t it is not the overall
three-dim ensional fold o f a zinc finger that confers specificity for either DNA
or RNA.
T he im portance o f fingers 4 and 6 for 55 rRN A binding is fu rth er
dem onstra ted by the com plem entary 55 rRN A m utagenesis data. F ingers 4
and 6 w ere show n to b in d to loop E an d loop A regions of 55 rRN A ,
respectively (Clemens e t al., 1993; McBryant et al., 1995). Loop A represen ts
the cen tra l hinge reg ion of the 55 rRNA, and is p ro p o sed to o rien t the
adjacent arm s of the m olecule (Westhof e t al., 1989). Likewise, loop E was
also sh o w n to provide im portan t secondary and tertiary features necessary
for specific TFIIIA in te rac tio n (Rom aniuk, 1985; H u b e r & W ool, 1986;
C hristiansen e t al., 1987; W imberly et al., 1993). Thus, protection and m issing
nucleoside experim ents em phasize the role of loop E as a major s tru c tu ra l
de te rm inan t for TFIIIA - 55 rRNA recognition. 5 im ilarly , substitu tions in
loop A resu lt in reduction of TFIIIA binding affinity (Sands & Bogenhagen,
1987; R om aniuk , 1989; You et al., 1991). A lthough secondary and tertia ry
structure of 55 rRNA has been implicated as the major determ inant of TFIIIA
bind ing , it is still no t clear w hether loop A residues prov ide d irec t base-
specific contacts or serve to preserve the overall in teg rity of the 55 rRNA
m olecule.
In a recently published study RNA binding by a zinc finger 4-7 pep tide
of TFIIIA w as assayed using alanine substitutions m ade in the four positions

191
predicted to be a-helical in each of the RNA-binding fingers 4-7, as well as by
phage display of zinc finger 4 (Friesen & Darby, 1997). Substitutions in fingers
4 and 6 resulted in the m ost significant reduction in RNA affinity: 77- and 30-
fo ld , respectively . In contrast, substitu tions in fingers 5 o r 7 d id no t
appreciab ly affect the RN A -binding ability. T he effect o f h istid ine to
asparagine substitu tion in zinc fingers 4, 5 and 6 stud ied in the context of the
full-length IF lilA show ed only a m odest reduction in RNA binding: 2.6 - 4.1-
fold (Setzer et al., 1996). This effect could probably be explained by supposing
th a t the rem aining fingers assum e an alternative m ode o f b ind ing , and
com pensate for the m utated ones. O ther researchers observed a reduced
affinity of fingers 5-9 compared to fingers 6-9, and a 60-fold decrease in affinity
fo r a peptide containing fingers 1-6 relative to the 1-7 finger fragm ent (Darby
& Joho, 1992; Clem ens et al., 1993), suggesting that fingers 5 a n d 7 do provide
a n im portant contribution to the binding affinity. It is possible that am ino
acid determ inants providing RNA specificity in fingers 5 an d 7 are different
from those m u ta ted in the "alanine substitution" study (Friesen & Darby,
1997), and lie elsew here in the zinc fingers. RNAse protection assays show ed
a ltera tions visible only for m utan t fingers 4, 5 an d 6 (Setzer e t al., 1996).
F inger 5 and 6 m u tan ts showed a loss of protection over helices II, IV & V
(Setzer et al., 1996). In contrast, no protection cou ld be de tected for the 3-
finger amino term inal fragment, even though it w as show n to b ind 5S rRNA
w ith affinity th a t w as only two-fold lower relative to the w ild -type pro tein
(Setzer et al., 1996). An explanation offered by the authors is th a t perhaps the
finger 1-3 fragm ent - 5S RNA complex undergoes a very ra p id dissociation,
a n d the 5S RNA is cleaved by the CV l RNAse shortly thereafter, w hich
results in a lack o f an RNAse footprint.

192
In an attem pt to define am ino adds in zinc finger 4 im portan t for RN A
b ind ing , a phage-display m ethod was used to screen a collection of m u ta n t
finger 4 sequences in which am ino ad d s at positions -1, +2, +3 and + 6 w ere
ra n d o m iz e d to genera te the m u tan ts w ith every possib le am ino ac id
substitu tion a t these four positions (Friesen & Darby, 1997). These positions
are u sed by EGR-1 pro tein for m aking direct hydrogen bo n d contacts to the
D N A bases (Pavletich & Pabo, 1991), an d w ere, by analogy , p resu m ed
im portan t for TFIIIA - RNA recognition. Lysine at position -1 w as selected in
all th e clones sequenced, and conserved in the wild-type p ro tein , suggesting
an im p o rtan t RNA contact or zinc finger structural role for this residue. O ver
half o f the d o n es selected serine (versus asparagine in the w ild-type) a t + 2
position . Serine w as found a t the +2 position in a num ber of other z inc-
finger proteins (Pavletich & Pabo, 1991; Pavletich & Pabo, 1993; Kim & Berg,
1996). O nly five o ther dones revealed identity w ith the w ild-type: g lutam ine
at position +3, and valine at position +6 . Subsequent b ind ing assays show ed
that L y s 'i —> Ala'^ substitu tion reduced zf 4-7 binding affinity 37-fold, w hile
a lan ine substitu tions a t o ther positions resu lted in a 7-fold low er affinity .
Thus, som e aspects of zinc finger - RNA interaction appear to be sim ilar to
DNA recognition, w ith some of the a-helical positions used for recognition of
both n u d e ic adds.
Because different subsets of TFIIIA zinc fingers in teract w ith d ifferen t
nucleic acids, a view em erged th a t TFIIIA is a fusion o f a spedfic D N A -
b ind ing and an RNA -binding m odules. H ow ever, this is a n oversim plified
view w hich does no t acknow ledge the contributions to the b ind ing m ade by
the rem ain ing zinc fingers in each complex (Theunissen e t al., 1992; D arby &
Joho, 1992; Del Rio e t al., 1993; Setzer et al., 1996). In add ition , in form ation
o b ta in ed from stud ies using isolated sets o f zinc fingers o r zinc finger

193
m utan ts, w h ile p rov id ing usefu l insights, m ay not necessarily reflect the
b ind ing m echanism of the full-length protein. Setzer e t al. (1996) quantified
therm odynam ic contributions of different "broken finger” m u tan ts of TFIIIA
as well as N - and C-terminal TFIIIA fragm ents to RNA b ind ing , and revealed
tha t, s im ila r to DNA bind ing , there exists a therm odynam ic interference
betw een zinc fingers at opposite ends of the protein. The sum of the free-
energy change (AG°) observed for the N- and C-term inal fragm ents is greater
th an the AG° of the full-length TFIIIA. If the two fragm ents w ere binding
independen tly , the sum of AG° for the tw o fragm ents shou ld be less than or
equal to the AG° of the full-length protein.
O nly tw o proteins, TFIIIA and W ilm s' tum or p ro te in have been
dem onstra ted to bind both DNA and RNA. A nother Xenopus protein, p43,
(Joho et al., 1990) is able to b ind RNA. H ow ever, p43 does no t appear to bind
DNA. A possibility exists that, in addition to these pro te ins, the function of
o ther C2H 2 type zinc finger proteins may also be th rough specific interactions
w ith RNA. This possibility needs to be fu rther investigated. Furtherm ore, it
w as recently reported that som e zinc finger proteins m ay function th rough
sequence-specific recognition of DNA-RNA hybrids (Shi a n d Berg, 1995). The
basal tran scrip tion factor S p l and a designed finger p ro te in ZF-QQR w ere
show n to recognize DNA-RNA hybrids in a sequence-dependen t m anner.
Strikingly, the binding affinities of these p ro teins for the DNA-RNA hybrids
w ere equal to or even higher than those for the consensus DNA binding sites.
This DNA-RNA binding activity may be a general p roperty com m on to m any
o ther m em bers of the C2H 2 protein class. A lthough a t p resen t there are no
know n physiologically relevan t examples of such in te rac tions, DNA-RNA
heteroduplexes are formed in such biological processes as gene transcription,
DN A rep lication , and the reverse transcrip tion of re trov iru ses. Therefore,

194
the po ten tial of zinc finger proteins to specifically interact w ith DNA-RNA
heteroduplexes, and , perhaps, m acrom olecules o ther than nucleic acids,
exists, an d requires further investigation.
6.2.7 Role o f TFIIIA zinc fingers in DNA recognition
TFIIIA interacts specifically w ith the intragenic control region (ICR) of
the 5S rRNA gene (Engelke et al., 1980) which overlaps the p rom oter region
req u ire d fo r accu ra te tran sc rip tio n in itia tio n (Sakonju e t a l., 1980;
B ogenhagen et al., 1980). The stoichiom etry o f the in terac tion is 1 : 1
m easured in titration experiments (Smith et al., 1984; Bieker & Roeder, 1984;
Rom aniuk, 1990). From DNA m utagenesis and in vitro transcrip tion studies,
the sequence comprising the ICR w as defined an d includes a region from base
pairs +45 to +97 (Pieler e t al., 1985a; McConkey & Bogenhagen, 1987). The
functional dom ains of the Xenopus 55 rRNA gene prom oter consist of three
elements: box A (bp +50 to +64), box C (bp +80 to +97), and an in term ediate
spacer elem ent (IE) (bp +67 to +72) (Pieler et al., 1985b; Pieler e t al., 1987; You et
al., 1991; Hayes & Tullius, 1992) (Figure 6.11). The box A com ponent is
com m on to all pol IH genes th a t are regu la ted via in tragen ic p rom oter
elem ents (Pieler & Theunissen, 1993). Box C an d the in term ediate elem ent,
on the o ther hand, are specific to 5S rRNA genes (Sakonju & Brown, 1982;
Smith et al., 1984; Pieler et al., 1987). The 5S D N A ICR appears to assum e a
^enlarged groove-DNA conform ation (Fairall e t al., 1989) w hich is found in a
num ber of DNA binding sites of other zinc finger proteins (Fairall e t al., 1993;
N ekludova & Pabo, 1994). Studies using metal p robes which recognize DNA
on the basis of its shape (Huber et al., 1991) indicate that the structure of the
ICR is no t uniform throughout the sequence, b u t rather contains distinct
segm ents o f predom inantly A-like, or B-like conform ation, reflecting the

195
ICR
+ 1 +120
box A IE box C
50 64 67 72 79 97
Figure 6.11 A schematic representation of the Xenopus laevis 5S rRN A gene
internal control region.

196
periodicity w ith which TFIIIA interacts w ith the ICR. In addition, the binding
o f TFIIIA induces b en d in g of the 5S gene prom oter (Fairall e t al., 1989;
Schroth et al., 1989; Bazett-Jones & Brown, 1989).
Extensive studies of the interaction o f TFIIIA w ith the 5S p rom oter
DNA sequence indicate th a t three elements o f the ICR provide
n o n eq u iv a len t co n trib u tio n to the o v e ra ll b ind ing affin ity . C hem ical
m o d ifica tio n experim en ts revealed th a t m éthy la tion o f e ig h t guan ine
residues clustered at the 3' end of the ICR (box C) abolished TFIIIA binding
(Sakonju & Brown, 1982). The same dram atic drop in TFIIIA b inding affinity
w as observed upon éthylation of the eight phosphates in the sam e reg ion of
the TFIIIA binding site (Sakonju & Brown, 1982). The im portance o f the box
C e lem en t fo r TFIIIA in te rac tion w as d e m o n s tra te d in m u tag en esis
experim ents (Sakonju e t al., 1981; Pieler e t al., 1985a; Pieler e t al., 1987; You et
al., 1991) an d in m éthylation protection stud ies (Fairall e t al., 1986). The latter
show ed a g radual increase in TFIIIA protection of the D N A binding site in the
S' to 3' direction (toward the C box) (Fairall e t al., 1986). In contrast, box A and
th e in te rm ed ia te e lem en t p rov ide m o d es t con tribu tions to the overall
strength of IF lilA binding (Sakonju et al., 1981; Pieler e t al., 1985a; McConkey
& Bogenhagen, 1987; You et al., 1991). S tudies using substitu tion m utagenesis
w ith in the box C and IE (Veldhoen e t a l., 1994) have fu rther refined the
regions as w ell as ind iv idual base pairs th a t contribute to the h igh affinity
binding. Thus, base pairs 78-79 and 92-96 are not essential for the high-affinity
interaction. Substitution a t position 70 has a large effect, w hile substitu tion at
position 71 has a m odera te effect, and positions 67 to 69 have n o effect.
Substitu tions a t positions 81, 85, 89 and 91 have a d ram atic effect, w hereas
positions 80, 82, 83, 8 6 -8 8 , and 90 h ave vary ing deg rees of in fluence.

197
Therefore, alm ost every base pair w ithin the box C region plays an im portant
role in b ind ing of TFIIIA.
A quantitative m easurem ent of the TFII1A-5S D N A equ ilib rium has
been carried ou t by Rom aniuk (1990). IF lilA binds 5S DN A with an apparen t
association constant of 1.90 x 10^ in 0.1 M salt, pH 7.5, a t 22°C (Rom aniuk,
1990). A nalysis of the therm odynam ic param eters ob tained from m easuring
tem perature dependence of the binding o f TFULA to 55 DNA indicates that
TFIIIA - 55 DNA interaction is largely enthalpy driven a t tem peratures above
19®C, and is entropy d riv en a t lower tem peratures (Rom aniuk, 1990). The
DNA b ind ing activity of TFULA has a b road pH optim um betw een 6 and 8
units of pH . The op tim al d ivalent ion concentration (MgClz) is 5mM. The
binding is insensitive to the identity of the monovalent cation in the binding
buffer. The analysis o f the m onovalent sa lt dependence of the b ind ing
allowed the determ ination of the num ber of ion pairs th a t form a t the TFULA
- 55 DNA interface. There are as many as eight ion pairs form ed (Rom aniuk,
1990), w h ich is co n sis ten t w ith the p rev ious chem ical m o d ifica tio n
experim ents which d e te rm ined that e igh t phosphates a re essential fo r the
binding of TFULA (Sakonju & Brown, 1982). The b in d in g show s dram atic
dependence on the id en tity o f the an ion ic species, decreasing 1 0 0 -fold
following the lyotropic series (Romaniuk, 1990). This indicates the presence
of several anion-binding sites on TFULA.
N u m e ro u s s tu d ie s u s in g m u ta n t TFIIIA p ro te in s p ro v id e
com plem entary inform ation about TFULA - 55 RNA gene complex form ation.
The results of footprin ting experim ents using truncated TFULA dem onstrate
that the am ino-term inal zinc fingers in teract primarily w ith the C box o f the
ICR, w hile the carboxy-term inal fingers interact w ith th e A box reg ion
(Vrana e t al., 1988). DNase I and hydroxyl radical footprinting studies suggest

198
that TFUIA makes contacts p rim arily along one side of the doub le helix
(Engelke e t al., 1980; C hurchill e t al., 1990; V rana e t al., 1988). The
arrangem ent of the ind iv idual zinc fingers over the 5S RNA gene prom oter
has been dissected using a series o f recom binant polypeptides. The studies
revealed tha t the con tribu tions ftrom the zinc finger dom ains are no t
functionally equivalent. The three am ino-term inal zinc fingers (zf 1-3) are
necessary and sufficient for high-affinity DNA binding, providing abou t 95%
of the b ind ing ftree energy (Christensen et al., 1991; Liao et 1., 1992; Clemens et
al., 1992; Choo & Klug, 1993; H ansen et al., 1993; Z ang et al., 1995). TFIIIA
variants w ith altered fingers 2 or 3 have significantly reduced DNA binding
affinities (Theunissen e t al., 1992; D arby & Joho, 1992; Del Rio et al., 1993; Zang
et al., 1995). Finger sw ap and amino a d d substitution studies conducted in Dr.
Rom aniuk's laboratory had identified that TFIIIA zinc fingers 2 and 3 provide
the m ajority of free energy of TFIIIA - DNA com plex, w hereas finger 1
p ro v id es energetica lly less im p o rta n t contacts (Zang e t a l., 1995).
Furtherm ore, a num ber of amino a d d s in the a-helical regions of the zinc
fingers w ere identified as the m ost im portan t residues for estab lish ing high
affinity interaction. These residues are predicted to occur at positions w ithin
the a-helix involved in DNA base recognition, by analogy w ith the EGR-1 -
DNA com plex (Pavletich & Pabo, 1991). It was recently show n th a t alanine
substitutions at four critical a-helical positions -1, +2, +3, and + 6 in any one of
the first three fingers of zf 1-3 of TFIIIA eliminated D N A binding (Friesen &
Darby, 1997). These four a-helical positions were chosen by analogy w ith the
contact am ino adds identified in the available crystal structures o f C2H 2 zinc
finger pro teins. The resu lts of these studies ind icate that am in o acids
im portan t for TFIIIA DN A -binding affinity may be a t the same positions of
the recognition a-helix as those of EGR-1 and other dosely rela ted proteins.

199
Interestingly, w hereas alanine substitutions in fingers 1 and 2 abolished DN A
binding, the sam e substitutions in finger 3 produced less dram atic reduction
in DNA binding. This result is different from the "broken finger" m utants of
Th'illA, w hich d isp layed a relatively small change in affinity for DNA: 2.5-
and 7-fold for fingers 1 and 2, respectively (Del Rio et al., 1993). It also differs
from the data ob tained in our lab (Zang et al., 1995). The discrepancy m ay be
a ttribu tab le to the fact that the "broken finger" m utants w ere created in the
context o f the fu ll-length TFIIIA protein (Del Rio et al., 1993), w hereas the
a lan in e su b s titu tio n s w ere m ade in the th ree -finger am in o -te rm in a l
fragm ent (Friesen & Darby, 1997), where other zinc fingers are not p resen t to
com pensate for the loss of DNA contacts. In addition, different sources of the
proteins, as well as the m ethods employed could all have contributed to these
differring sets of data.
T he p ep tid e contain ing fingers 1-3 of TFIIIA b inds to the box C
prom oter elem ent (Liao e t al., 1992), covering the region o f approxim ately
nucleotide positions 79 to 92 (Hayes & Tullius, 1992). Judg ing from the
pa tte rn o f the hydroxyl radical and DNase I footprinting experim ents w ith
full len g th TFIIIA, o r truncation m utants, a change in the o rien tation of
TFIIIA relative to the DNA occurs at positions w here fingers 4 and 6 are
show n to bind (Clem ens e t al., 1992; Hayes & Clemens, 1992; Fairall & Rodes,
1992). Therefore, fingers 4 and 6 are proposed to cross the m inor groove.
Finger 5 binds in the major groove of the interm ediate elem ent, and the C-
term inal trip let - fingers 7-9 - interacts w ith the box A sequence (H ayes &
Tullius, 1992; C lem ens et al., 1994). The model in which fingers 1-3 and 7-9
w rap around the ends of the TFIIIA binding site, while fingers 4-6 span the
center, crossing the m inor groove twice was originally proposed by Hayes an d
Tullius based on the results of the missing-nucleoside and hydroxyl rad ical

200
foo tp rin ting experim ents (Hayes an d Tullius, 1992) (Figure 6.12A). A very
sim ilar model was proposed by Clem ens and cow orkers (1994). Interestingly,
it w as found tha t polypeptide containing zinc fingers 1 to 5 b in d s the DNA
w ith h igher affinity than the w ild-type TFULA, b u t the ad d itio n of finger 6
w eakens the b in d in g to DNA. These effects m ay be d u e to a n altered
conform ation tha t the peptides adop t, resulting in the change in the strength
of DN A-binding activity (Clemens et al., 1994). Fingers 7, 8 and 9 w ere show n
to be dispensable for DNA binding. It should be noted, how ever, that fingers
8 and 9 are absolutely required for transcriptional activity (Rollins e t al., 1993;
Del Rio & Setzer, 1993). Thus, it appears that d ifferen t subsets o f TFULA zinc
fingers provide energeticaUy d ifferen t contributions to the D N A interaction,
contact DNA reg ions of d ifferent size, and b in d in both m ajor and m inor
grooves of the DNA.
The previous m odels for the TFULA - 55 D N A interaction w ere based
on the assum ption th a t the nine fingers of TFULA w ould in terac t w ith DNA
in essentially the sam e way as does EGR-1 p ro te in (Fairall e t al., 1986;
C hurchül et al., 1990), w ith all the zinc fingers p rov id ing approxim ately equal
contributions to the binding (Figure 6.12C). In fact, only the f irs t three zinc
fingers of TFULA show the closest similarity w ith the consensus sequences of
other zinc finger p ro teins (Gibson e t aL, 1988; K lug & Rhodes,1987; Jacobs,
1992). The overall TFULA - D N A interaction ap p ea rs m uch m ore complex
and asym m etrical th an originaUy proposed in the early models. A num ber of
recent studies of TFUIA-DNA interaction cum ulatively confirm the view that
there is a high deg ree of non-uniform ity and functional in te rdependency
am ong the zinc fingers (Clemens e t al., 1994; Rawlings e t aL, 1996; Rowland &
Segall, 1996; M cBryant et aL, 1996). Perhaps, p ro te in s w ith rela tive ly large
num bers of zinc fingers should be analyzed differently from the smaU and

201
3 ’ N C
Cri/
IE A
rr •wCOOH
3 C
5 ’ N C
CO•jO
COOH
L&'
6 \
F igure 6.12 M odels for TFIUA - 5S DNA complex. (A) After H ayer & Tullius,
1992. (B) After C lem ens e t al., 1992. (C) After Fairall & Rhodes, 1992.

202
com pact ones, like EGR-1. Therefore, sim ple ex trapolation o f an EGR-l-like
m ode of b inding is no t adequate w hen trying to understand proteins such as
TFHIA.
A series o f "broken-finger" TFHIA m u tan ts w ere created , in w hich
h istid ine residues w ere substitu ted by asparagines to in troduce structural
d isrup tions in each zinc finger (Del Rio e t al., 1993). T hese experim ents
confirm that zinc fingers 1 and 9 are least im portan t for D N A binding, while
the m ost de le terious effect on DNA binding is p roduced w hen finger 3 is
m u ta ted (Del Rio e t al., 1993). Curiously, w hile d isruption o f finger 4 did no t
re su lt in any a ltera tions of the footprint, it d isp layed the second largest
reduction of the b ind ing affinity (Del Rio et al., 1993). T hese results suggest
tha t finger 4 p lays an im portant role of correctly linking the first zinc finger
trip le t w ith the m iddle triplet. The finger sw ap experim ents conducted in our
lab (Zang et al., 1995) also identified fingers 2 ,3 , and 4 as im portan t for IFlilA -
DN A interaction, and agree very well w ith the findings from the "broken
finger" experim ents (Del Rio et al., 1993).
The short sequences connecting zinc fingers, called linkers, are highly
conserved in evo lu tion (Choo & Klug, 1993). Therefore, it w as proposed that
they play an im p o rtan t functional role w hich until recen tly w as not well
understood . A linker of the sequence TGEKP is conserved betw een m any
zinc finger p ro te in s, including EGR-1, but on ly in the linkers of the three
am ino-term inal fingers of TFHIA (Gibson et al., 1988; Jacobs e t al., 1992; Choo
& K lug, 1993). The role of the interfinger linker sequences in the am ino-
te rm in a l three z inc fingers has been assessed in a n u m b er of stud ies
invo lv ing single an d m ultiple am ino acid substitu tions (Sm ith e t al., 1991;
C hoo & Klug, 1993; Clemens e t al., 1994). These su b s titu tio n s result in
red u c tio n of D N A -binding betw een 7- and 24-fold (C hoo & Klug, 1993).

203
In teresting ly , rep lacem ent of the first linker o f TFHIA w ith the linker
connecting zinc fingers 3 and 4 abolished DNA b ind ing (Choo & Klug, 1993).
T his suggests th a t different linkers provide nonequivalent contributions to
th e finger-finger re la tionsh ips. Thus, linker sequences a re im p o rtan t
determ inants of the overall structural integrity of TFHIA and a re required for
high-affinity DNA binding. It rem ains to be established w hether the function
of the linker sequences is to correctly position the zinc fingers relative to the
o ther domains of a protein, provide flexibility/rigidity, or m ake DNA contacts
(the latter is less likely), or any combinations of the above.
Recently, a solution structure of the first th ree zinc fingers of TFHIA
com plexed to its cognate DNA (base pairs 79 to 93 in the ICR num bering
schem e) has been reported (W uttke et al., 1997). N uclear m agnetic resonance
spectroscopy has been em ployed to solve the h ig h reso lu tion structure.
F igure 6.13 shows the amino acid sequence of zf 1-3 and the sequence of the C
block of the 55 DNA element. The structure reveals that the th ree zinc finger
p ep tid e wraps a round the DNA m ajor groove, m uch like the zinc fingers of
EGR-1. However, there are several new features: each zinc finger contacts 4 to
5 base pairs of the DNA, novel am ino acids (such as tryptophan a t position +2
of the finger 1 a-helix and arginine a t position +10 of finger 3) a re involved in
m ak ing direct base contacts, w hile the entire a -he lix of finger 3 is used for
m aking sequence-specific contacts.
In sum m ary, the protein-DNA recognition by the zf 1-3 fragm ent of TFHIA is
m ed ia ted by electrostatic and hydrogen bond ing in terac tions, as well as
struc tu ra l com plem entarity at the protein-DNA interface. All contacts occur
in the major groove of the DNA o n both coding an d noncoding DNA stands
(w ith the m ajority of the contacts m ade to the non-coding strand ). The
contacts to the sugar-phosphate backbone, while no t providing specificity.

204
(a)Strand 1 Strand 2 Helix
c z ^ - i Q â â r 6 10 J! 11 15 20 26 n Î7 42
M K R ( Y ) l £ S F A 0 C G A A ® n ( 3 8 @ k1 L Q A £ L T G E ® P
43 45 » 56 S3 67 22
F P «■: K E E G C E ® G F @ S L H ^ L T ( R ® S L T H T G E ® N
n rS M M »3 M 101F T C O S O G C D L R F @ T ® A @ ] M ( g @ 0 F N @ ]F H N I K
(b) 79 93I S 10 I S
5 ’-T T G G A T G G G A G A C C G - 3 * (noncoding)3 - A A C C T A C C C T C T G G C -5 ' (coding)
30 26 21 16
Figure 6.13 (a) Am ino acid sequence of zf 1-3. The construct consists of
residues 11 to 101 of TFIUA. C onserved zinc lig an d s (Cys and H is) are
u n d e rlin ed ; the P-strands are in d ica ted by a rro w s, an d the a-helix by a
cylinder. Boxes indicate residues involved in m ak ing hydrophilic contacts to
DNA b ases , an d circles denote those involved in con tacting the D N A
phosphate backbone, (b) DNA sequence of the C -block elem ent of th e 5S
RNA gene. Both structural (nucleotides 1-30) an d biochem ical (nucleotides
79-93) num bering schemes are shown.

205
help anchor the zinc fingers to the D N A as well as p roperly orient them in
the m ajor groove.
In d iv id u a l fingers appear to p rov ide nonequ ivalen t contributions to
the overall binding. Thus, finger 3 contributes m ore th an any other finger to
the TFIIIA-DNA in teraction . Indeed , unlike any o th e r zinc finger-D N A
structu res characterized todate, the en tire length o f finger 3 of TFIUA (i.e.
positions -1 to +10) are involved in sequence-specific base contacts. Extensive
m utagenesis data of finger 3 and corresponding D N A site also p o in t to the
critical role of finger 3 in the high affinity binding by TFHIA (Zang et al., 1995;
V eldhoen e t al., 1994; P ieler et al., 1985). An u n u su a l feature of finger 1
in teraction is the extensive netw ork o f hydrophobic interactions fo rm ed by
Trp28 (position+2 in the a-helix). It contacts four bases o n both strands of the
DNA, w h ich has not been previously docum ented fo r any other zinc finger
proteins.
The role of the interfinger linker structures are o f particular in terest as
NMR struc tu re determ ination provides a dynamic v iew of the interaction. It
w as show n previously th a t point m utations in the linker sequences can be
deleterious to DNA b ind ing (Choo & Klug, 1993; C lem ens e t al., 1994). The
present N M R structure show s that linker sequences a d o p t a highly o rdered
structure u p o n the DNA complex form ation thus actively contributing to the
overall s tab ility of the com plex. S im ilarly , p ro te in -p ro te in in te rac tions
betw een zinc fingers indirectly contribute to the h ig h affinity DNA binding.
A nother strik ing feature of the presen t NMR structu re is th a t a surprising ly
large n u m b e r of D N A -con tacting side-chains e x h ib it con fo rm ational
flexibility w ith in the protein-D N A interface w ithout loss of specificity.
In conclusion , the s tu d y gives a m olecu lar leve l p rospective fo r the
accum ulated m utagenesis and footprinting data available for TFHIA.

206
CHAPTER 7.0 THE STUDY OF THE NUCLEIC ACID INTERACTIONS OF
ZINC HNGER 4-7 REGION OF THIIA
7.1 Introduction
In a recently pub lished study, Z an g e t al. (1995) observed th a t the
substitu tion o f the TFIUA zinc fingers 4 to 7 by the equivalent zinc fingers of
an R N A -binding protein p43 produced a 100-fold reduction in the DNA
binding affinity of the resultan t protein construct. This effect can be explained
by either assum ing that a num ber of energetically im portant contacts are lost,
o r tha t th e im proper a lignm en t of th e m u tan t p ro te in over the DN A
precludes high-affinity interaction. Replacem ent of the finger 7 alone, on the
other hand , d id not result in a significant change in the b ind ing affinity (Zang
et al., 1995). Therefore, the rem aining fingers 4, 5, an d 6 m ust possess
im portant determ inants of the DNA-binding specificity.
W e have approached the problem relating to the functional role of the
zinc finger 4 to 6 region in high-affinity nucleic a d d binding by quantitatively
analyzing the properties of w ild-type TFIUA and a set of the m utant form s of
the pro tein containing substitutions of an entire zinc finger, or either halves
of a g iven finger. The do n o r proteins u sed w ere p43 an d W ilm s’ tum or
proteins. Since these donor proteins belong to the same zinc finger fam ily as
does TFIUA, the stereochem ical fold of the resu ltan t m u ta n t p ro te ins is
expected to be m aintained. Thus, the differences in the m easured p ro te in -
nude ic a d d b inding affinities are presum ed to be attributable to the lo st or
gained contacts. p43 protein w as chosen as a donor since it does not have a
D N A -binding activity, and, therefore, w ou ld not interfere w ith the analysis of
the D N A -binding properties of zinc fingers under investigation. Sim ilarly,
the W ilms' tum our protein does not spedfica lly interact w ith either the 55
rRNA nor its gene, and is thus also well su ited for the purposes of the study .

207
The pro teins were purified to hom ogeneity, and assayed for their D N A and
RNA b ind ing abilities using a nitrocellulose filter b ind ing assay. This is a
co llabora tive s tu d y w ith Dr. P au l J. R om aniuk, w ho sy n th esized the
o ligonucleo tides fo r the PC R -m ediated s ite -d irec ted m u tagenesis , and
Kathleen Barilla, w ho constructed a n d purified the Tp series of p ro te ins , as
well as m ade constructs for the expression of the WTF and TFW series of
m u tan ts (see nom encla tu re in th e results sec tio n of this ch ap ter). I
constructed and purified the TF(4-7)WT m utant, carried out the expression
and purification of the WTF and TFW sets of the m u tan t proteins, as well as
carried o u t all the DNA and RNA binding experim ents.
7.2 M aterials and M ethods
7.2.1 Construction of TFIIIA finger sw ap m utants
The TFIUA m utan ts were produced by site directed m utagenesis, which
w as e ither PCR-m ediated, or carried ou t w ith the use of a transform er kit
(Clontech Laboratories, Inc.). P lasm id pT7-TF contain ing the TFIIIA cDNA
(Ginsberg e t al., 1984) was a k ind g ift from Dr. J Tso. The N d e l/B a m H l
fragm en t of pT7-TF w as cloned in to the sam e restric tion sites of the
expression plasm id p E T -llb (Studier e t al., 1990) to yield pTF4 (Veldhoen,
Ph.D. thesis). To facilitate mutagenesis, zinc finger 4 to 7 region of TFIUA was
am plified from pTF4 using prim ers w hich incorporate restriction sites Bgl n
and Xhol for insertion back into TFIIIA cDNA (Table 7.1). The am plified zinc
finger 4-7 fragm ent o f TFHIA was cloned by b lun t-end ligation into Sm al site
of pUC19, and the sequence of the resu lting p lasm id was verified by
sequencing. The resu ltan t plasmid w as designated pUC19-zf4-7 (Figure 7.1A).
To create a TFIUA m utan t in which zinc fingers 4-7 are replaced w ith the zinc
fingers 1-4 of W Tl, specific primers w ere designed to am plify zinc finger

208
T able 7.1 Sequences of oligonucleotides u sed in construction of TFHIA
substitu tion m u tan ts (see details in the text).
Construct name Oligonucleotide sequence Specific Comments
pUCI9-zf4-7 CATATGAAGATCTGCGTCTATGTGT Upstream; Bgl II site ACTAGTATCTCGAGGGCAGAGATA Downstream; Xho I CA site
PÜCI9-WT1-BX AGATCTGCGTCTATGTGTGTGCTTACCCAGGCTGCAAT
Upstream; Bgl II site Downstream; Xho I
Tp4A CATATGAAGATCTGCCCCTTAAAGTGCnTGTTCCGGGCTGTAAGAGAAGCnCAAGAAACACAAT
Replaces first 15 a.a. of TFHIA finger 4 with first 15 a.a. of p43 finger 4
Tp4B AACTGTGGCAAAGCTTTCAGGAAAAAGCGTGCATTAAGGCGTCATCTGTCCGTTCACAGTAATCAGCTGCCATACGAA
Replaces last 15 a.a. of TFHIA finger 4 with last 15 a.a. of p43 finger 4
Tp5A AGTCACACACAGGAGCCGCTATCCGTATGTGATGTTCCAGGCTGTrCCTGGAAGTdTCTTTGCCT
Replaces first 15 a.a. of TFHIA finger 5 with first 15 a.a. of p43 finger 5
Tp5B AAGCGGTTTTCTTCGGTTGCCAAGCTTGTAGCTCATCAAAAACGCCATCGAGGCTATCCCTGC
Replaces last 15 a.a of TFIIIA finger 5 with last 15 a.a. of p43 finger 5
Tp5C TGGAAGTdTCTTCGGTTGCCAAGCTTGTAGCTCATCAAAAACGCCATCGAGGCTATCCCTGC
Swaps TFHIA finger 5 with p43 finger 5

209
Construct name Oligonucleotide sequence Specific Comments
Tp6 A
Tp6 B
WTF4
WTF5
WTF6
TFW4
TFW5
TFW6
GTCCATGCAGGCTATCGCTGCAGTTATGAAGGTTGCCAAACTGTGAGTCCGAdTGGACATTATAC
GGAAAGACITGGACAGCACTGCAGACACACGTGAAAAAACATCCGCTCCTAGCAGTATGTGAT
AAGAGATATAAAAAGCATAACCAGCTGAAGGTGCACCAGTTCAGCCACACTCAAGAGAAACCATAC
AGGTTTTCTCTTCCATCCCGGCTTAAGAGACACGAAAAGGTACATGCAGGTGTGAAACCA
CGAAAGTTCAAGACGTGGACCCTCTATCTTAAGCACGTCGCCGAGTGTCATCAAGATGAAAAGCCCITC
AAAGCATICnTAAACrCAGTCATTTACAGATGCATTCGCGCAAGCACACAGGGCAGCTGCCA
CGGTnTCTAGATCTGACCAGTTAAAACGTCATCAAAGACGCCATACAGGCTATCCC
GTGGGAAAGAGATCTGATCATCTCAAGACACACACAAGAACCCATACGGGCCTAGCAGTATGT
Replaces first 15 a.a. of TFHIA finger 6 with first 15 a.a. of p43 f.6
Replaces last 15 a.a. of TFHIA finger 6 with last 15 a.a. of p43
Replaces last 15 a.a. of W Tl finger 1 with last 15 a.a. of finger 4 of TFHIA
Replaces last 15 a.a. of W Tl finger 2 with last 15 a.a. of finger 5 ofTFHHA
Replaces last 15 a.a. of W Tl finger 3 with last 15 a.a. of finger 6
of TFHIA
Replaces last 15 a.a. of TFHIA finger 4 with last 15 a.a. of finger 1 of WTl
Replaces last 15 a.a. of TFHIA finger 5 with last 15 a.a. of finger 2 of WTl
Replaces last 15 a.a. of TFHIA finger 6
with last 15 a.a. of finger 3 of WTl

210
Bgl II X VO I
1 1 1 1 2 i 3 9 1 1 3'UTR 1'W ^ -W
TFIU A zinc fingers 1
C-term inus
1 . PCR amplify zf4-7
2. Blunt-end ligate into pUC19,
T sequencepUC19-zf4-7
B
W n zinc fingers in a plasmid
1- PCR - amplify the zinc finger region, incorporating Eg\ II and Xho I sites for cloning into full-length TFHIA
2. Clone into pUC19, sequence
pUC19-WTl-BX
Figure 7.1 Schematic diagramm illustrating construction of (A) pUC19-zf4-7;
(B) pUC19-WTl-BX

211
reg ion of W Tl, and incorporate Bgl II and Xho I restriction sites for
subsequen t clon ing into full-length TFIUA cDNA (Table 7.1). The general
PCR protocol w as used as described in C hap ter 3 of this thesis. The PCR
p ro d u ct w as cloned into pUC19, sequenced, an d designated pUC19-WTl-BX
(Figure 7.1B).
M utants Tp5A, Tp5B, TFW4, TFW5, WTF4, WTF5, a n d W TF6 were
generated using PCR-m ediated site directed m utagenesis p ro ced u re (Nelson
& Long, 1989). The nom enclature T p ’ refers to TFIUA m u ta n ts in w hich the
first o r last 15 am ino adds w ith in a given zinc finger w ere rep laced w ith the
f irs t or last 15 am ino acids o f the co rrespond ing f in g e r o f p43. The
nom enclature 'WTF' refers to constructs in w hich TFIUA zinc fingers 4-7
w ere replaced by fingers 1-4 o f the W Tl protein, and, subsequently , a-helical
reg ions of TFIUA fingers 4-6 w ere placed back. TFW' refers to TFIUA
substitu tion m utan ts in which a-helical regions of zinc fingers 4, 5, and 6 are
replaced w ith those of the W Tl zinc fingers 1, 2, and 3.
pUC19-zf4-7 w as used as a tem plate to create m u tan ts Tp5A, Tp5B,
TFW4 and TFW5. pUC19-WTl-BX was used as a tem plate to create m utants
WTF4, WTF5, an d WTF6 . As the first step, the m utagenic sequence (Table
7.1) and a unique flanking prim er site (RUMP) w ere in troduced into the wild
type tem plate D N A (Figure 7.2). The 50 pi PCR reaction contained: PCR buffer
(10 m M Tris-HCl pH 8.3 at 20°C, 1.5 mM MgClz, 25 mM KCl), 50 p g /m l gelatin,
2.5 nM each o f dTTP, dGTP, dCTP, and dATP, 10 pmol FUM P and m utagenic
p rim er, 1 ng tem plate DNA, and 1.25 units T aq DNA po lym erase . The
reaction was overla id with 50 p i mineral oil p rio r to therm ocycling. Twenty
fo u r ro u n d s o f therm al cycling were carried o u t u s in g the follow ing
conditions: d én a tu ra tio n tem pera tu re of 94®C (1.5 m inu tes), an annealing
tem perature of 55°C (1.5 m inutes), and an extension tem perature of 72°C (two

212
Step 1 : Incorporate m utations into the pro tein sequence
FUMP
EcoR I
Bgl II
E & 3m utagenic p rim er
Hind III 'BamH I
X ho l
Extend the PCR-generated m utant tem plate
EcoR I
.XX i- — -----------— ^
Bgl II
Step 2: Amplify the m utant template
UMP 1
I H ind III 3amH I
X hol
■JQL I______
RUP
XX
Figure 7JZ Schematic chart o f the PCR-mediated site-directed
mutagenesis protocol used to create TFIUA substitution m u tan ts

213
m inutes). The second step reaction (80 pi) contained: 1 ng tem plate DN A,
PCR buffer (10 mM Tris-HCl pH 8.3 a t 20=C, 1.5 m M MgClz, 25 mM KCl), 50
p g /m l gelatin, 5 nmol each of dTTP, dGTP, dCTP, dA TP, 1-3 pi step 1 PCR, and
2.5 u n its Taq DNA polym erase. Initially, five ro u n d s of therm ocycling w ere
carried ou t under the following conditions: dénatu ration tem perature of 94°C
(3 m in u te s ) , an an n ea lin g tem p era tu re of 55°C (1.5 m inutes), an d an
extension tem perature o f 72°C (two m inutes). 20 p i reaction buffer containing
20 p m o l universal m utagenic (UMP) an d M13 rev e rse universal p rim ers
(RUP) w ere added, and 25 additional cycles, identical to step 1, were carried
out.
The PCR products were pu rified by phenol: CHCI3 extraction, an d
ethanol-precipitated . The Tp series m u tan t constructs w ere cloned into EcoR
I an d BamH I sites of pUC19, and TFW & WTF constructs were cloned in to
EcoR I and Hind m sites o f pUC19 for sequence verification. The correct
sequences w ere subsequently subcloned into Bgl H and Xho I sites of pTF4 for
p ro te in expression.
M utants Tp4A, Tp4B, Tp5C, Tp6 A, Tp6 B, a n d TFW 6 were created w ith
the u se of the transform er site-directed m utagenesis k it (Clontech laboratories
Inc., 1994) (Figure 7.3). Sequences of m utagenic oligonucleotides are given in
Table 7.1. Sequences o f selection p rim ers are sh o w n in Table 7.2. All the
procedures w ere perform ed as per protocol (Clontech laboratories Inc., 1994).
The re su lta n t constructs w ere subcloned into Bgl II and Xho I sites of pTF4
p lasm id for protein expression.
7.2.2 Expression and purification of TFIIIA proteins
Expression and purification of recom binant TFIIIA proteins was carried
out as described by Del Rio & Setzer (1991). 100 m l LB m edium supplem ented

214
Unique restriction site: Sca I pUC19-
zf4-7
Cloned gene
I. Denatureds DNA
S tu l
Z Anneal primers
primer: 4^----w T I
Selection
Seal
Mutagenic primer
3. Synthesize second strand with T4 DNA polymerase and seal gaps with T4 DNA ligase; primary digestion with selection restriction enzym e
Figure 7.3

215
4 T r a n s f o r m / . ' /i
f i r s t I r a n s t o r t i M i i . i n .
i s o l a t e * D N A
itu
Parentalplasmid
Sca I
Mutatedplasmid
5- Secondary digestion with selection enzyme: Sca I
6 . Transform E.coli Second (final) transformation
Mutatedplasmid
7. Isolate DNA from individualtransformants; sequence to confirm presence of desired mutation
M utatedplasmid
Figure 73 Schematic diagram show ing the strategy for introducing m utations using
the Transformer site-directed mutagenesis kit (Clontech Inc., 1994)

216
T able 7.2 Sequences o f selection prim ers used in transform er m utagenesis
protocol to construct substitu tion m utan ts o f TFIIIA
Nam e of p rim er ■Sgquence Specific com m ents
Trans Oligo Sca I/S tu I GTGACTGGTGAGG T urns w ild type pUC19
CCTCAACCAAGTC Sca I site at 2177 to Stu I
Switch Stu I/S ca I GTGACTGGTGAGT
ACTCAACCAAGTC
T urns Stu I site in pUC19
back to wild type Sca I

217
with 100 ^ g /m l am pid llin w as inoculated w ith 1 ml of overnight culture of E.
coli s tra in BL21(DE3) h arbo ring expression plasm id o f in terest (containing
either w ild type or m u tan t TFHIA gene). The culture w as grow n at 37°C with
shaking a t 300 rpm to an O.D.goo of 0.5 to 0.7. Protein synthesis was initiated
by ad d itio n of ZnS0 4 an d IPTG to final concentrations o f 50 pM an d Im M ,
respectively. After induction for three to four hours, cells were harvested by
centrifugation at 3800 x g for 10 minutes a t 4°C, in a Beckm an JA-20 rotor, and
w ashed w ith 10 ml of ice-cold TAB buffer (20 mM HEPES pH 7.4, 5 mM
M gC lz/ 50 |iM ZnS0 4 , 5 m M DTT, 10% glycerol, 250 m M NaCl). All the
following procedures w ere carried out on ice. The cells were pelleted once
more, and resuspended in 4 m l of TAB? buffer (TAB buffer + 1 mM PMSF).
Cells w ere sonicated using a microtip sonicator (Heat System s-Ultrasonics W-
385) a t se tting four, 50% du ty cycle for 6 x 20 second intervals w ith 1 m inute
cooling be tw een the pu lses. The sonicated cells w ere centrifuged for 20
minutes a t 12000 x g in a Beckman JA-20 rotor. The pellet was resuspended in
1 ml of TABUP buffer (TAB + 5M urea + PMSF) a n d m ixed by inversion
overnight in a coldroom (4°C).
The solubilized extract w as spun in a m icrocentrifuge for 20 m inutes at
14000 X g, and the su p e rn a ta n t rem oved to a new tube. A series of
am m onium sulphate cuts w as started by addition of 0.666 m l of TABAS (TAB
saturated w ith (NH4 )2S0 4 ) to bring the sam ple to 40% saturation, and mixed
by inversion for one hour. The proteins precipitated by the first ro u n d of
salting-out w ere pelleted by centrifugation a t 1 2 0 0 0 x g for 2 0 m inutes, and
discarded. M ore (N H 4 )2S0 4 was added to the rem ain ing su perna tan t to
achieve 80% saturation. The sam ple w as mixed by inversion for one hour,
pelleted b y cen trifuga tion a t 1 2 0 0 0 x g for 2 0 m in u te s , and the pellet
resuspended in 10 ml of TABU-250 (TAB + 5 M urea + 250 mM NaCl). This

218
protein so lu tion was loaded onto BioRex 70 colum n (800 |il) pre-equilibrated
w ith TABU-250. The colum n w as then w ashed w ith 1 m l o f loading buffer,
follow ed by TABU-400 (TAB + 5M urea + 400 mM NaCl). The protein w as
e lu ted from the column in 800 |il of TABU-600 (TAB + 5M urea + 600 m M
NaCl). P urified proteins w ere aliquoted and stored at -70°C. The p u rity of
each protein preparation w as checked on a 10% SDS-PAGE (Figure 7.4 & 7.5).
P ro te in concentrations w ere determ ined by the m ethod of Bradford (1976)
using BSA as a standard.
7J1.3 R adiolabeling of 5S D NA
The 5S rRNA gene w as released from plasm id pXlo (Rom aniuk e t al.,
1987) using EcoRI and HindUl endonucleases of restriction a n d end-labeled
w ith [a-P32]-dATP and the Klenow fragment of DNA polym erase I (Sambrook
e t al., 1989). The details of the labeling protocol are the sam e as in C hapter 3
of this thesis.
7.2.4 Synthesis and R adiolabeling of 55 rRNA
A m ethod of in vitro run-off transcription was used as described by
R om aniuk e t al. (1987). The pXlo plasm id containing 5S rR N A gene w as
d igested w ith D ral which cleaves the DNA a t position +121 relative to the
first nucleotide incorporated in to RNA. The digested D N A was ex tracted
w ith 100 |il of phenol: CHCI3 , and DNA precipitated w ith ethanol. The DNA
pellet was d ried in Speed Vac concentrator, resuspended in deionized w ater,
and stored a t -20°C.
Internally labeled 5S rRNA w as made as described by R om aniuk e t al.
(1987). The labeling reaction had a final volume of 100 pi, an d contained: 40
m M Tris-HCl pH 8.0,15 mM MgCl2, 5 mM DTT, Im M sperm idine, 100 p g /m l

219
8 MW
Figure 7.4 C oom assie blue-stained 15% SDS-poIyacrylam ide gel show ing
TFIUA w ild type a n d m u tan t proteins. Lane designated MW represen ts
protein m olecular w e igh t m arker; lane 1: w ild-type TFUIA; lanes 2-8; Tp4A,
Tp4B, Tp5A, TpSB, Tp5C, Tp6A, Tp6B.

220
LJ7 MW
F igu re 7.5 C oom assie b lue-stained 15% SDS-polyacrylam ide gel show ing
TFUIA w ild type a n d m utan t pro teins. Lane designated M W rep resen ts
p ro te in m olecular w e igh t marker; lane 1: w ild-type TFUIA; lanes 2-4: WTF4,
WTF5, WTF6 ; lanes 5-7: TFW4, TFW5, TFW6 .

221
BSA, 1000 U /m l RNasin, 0.5 mM each ATP, CTP, UTF, 0.0125 mM GTP, 50
liCi [a-P32] g T P (600Ci/m m ol), 1 p.g of tem plate DNA and 0.6 |ig of T7 RNA
polym erase. The reaction w as incubated for 3 hours at 37°C, and purified on
an 8 M urea, 12% polyacrylam ide gel. The gel band containing the labeled
R N A w as excised, eluted overnight in elution buffer an d ethanol precip ita ted
(Sam brook e t al., 1989). The level of radiolabel incorporation was determ ined
u sin g an LKB 1214 Rackbeta Scintillation counter and toluene to w h ich 0.4%
PPO w as added as a scintillant.
7.2.5 N itrocellulose filter b ind ing assays
The b ind ing affinities were quantified using a n itrocellu lose filter
b in d in g assay as described on page 8 8 of this thesis, except the b ind ing
reactions w ere supplem ented w ith 28 |ig /m l of nonspecific com petitor po ly A.
7.3 R esults
7.3.1 Effects o f zinc finger substitu tion m utagenesis o n the DNA b in d in g
activ ity of TFIIIA
The first p a rt of th is study was to determ ine the con tribu tions of
ind iv idua l zinc fingers 4, 5, and 6 to the high affinity DNA binding o f TFUIA.
The em ployed strategy was to replace one half of a zinc finger at a tim e w ith
the corresponding sequences of the p43 protein (Figure 7.6) to assess w hether
the second half (a-helical portion) of the zinc finger sequences, p resu m ed to
be the "recognition" helix by analogy w ith the EGR-1 pro tein , w ould p rovide
a g rea ter contribution to the binding than the first half. A nitrocellulose filter
b in d in g assay w as used to m easure the affinity for each m utan t p ro tein . The
resu lts of the experiments are given in Table 7.3 in a form of relative affinities
to th e w ild-type TFUIA.

222
P43-ZF4TF0IA-ZF4
P43-ZF5TFmA-ZF5
P43-ZF6TFmA-ZF6
a-helix > -
- 1 1 2 3 4 5 6GEAVPLKCFUPGCKASFRKKAALRRHLSVHSN
NIKICVYVCHFENCGKAFKKHNQLKVHQFSHTQ
E PLSVCDVPGCSWKS S SVAKLVAHQKRHRG QLPYECPHEGCDKRFSLPSRLKRHEKVHAG
YRCSYEG CQTVSPTWTALQTHVKK HAG Y PCKKDD s e s FVGKTWTLYLKHVAECHEK
Figure 7.6 Com parison of the amino a d d sequences of zinc fingers 4-6 of
the donor p43 w ith the zinc fingers 4-6 of TFHIA. The structural
cysteines and histidines are outlined; the conserved 'TWT' triplet in
firmer 6 is shown in bold.

223
T able 7 3 T he effects o f TFIIIA zinc finger substitu tion m utations on the DNA
an d RNA b in d in g o f the factor
M u ta n t Relative affinity for 55 Relative affinity for 55
rRNA gene rR N A
Tp4A 0.72 ± 0.14 0.84 ± 0.09
Tp4B 0.59 ± 0.04 0.14 ± 0.02
Tp5A 0.29 ±0.05 0.50 ± 0.06
Tp5B 0.23 ± 0.02 0.82 ± 0 .1 1
Tp5C 0.09 ± 0.002 0.83 ± 0.10
T p 6 A 1.02 + 0.09 0 .8 8 ± 0 .0 2
T p 6 B 1.56 ± 0.09 0.89 ± 0.02
The nom enclature Tp' refers to TFHIA m utants in w hich the first or last 15
am ino acids w ith in a given zinc finger were replaced w ith the first or last 15
eunino a d d s of the corresponding finger of p43. Letters A, B, and C refer to the
first half, second half, o r the whole zinc finger region replaced.
Relative affinities w ere calculated using the m ean values o f the assodation
constants determ ined for the m utant proteins (each value representing the
m ean of th ree independen t determ inations), and com pared against the value
o f the w ild-type TFHIA assodation constant (Ka=1.9 x 1 0^ M 'l for DNA; Ka=l-4
X 109 M -l for RNA).

224
The substitu tions o f either halves of finger 4 resulted in a less th an a
tw o-fold red u c tio n in DN A binding , thus im ply ing that likely no specific
hydrogen-bond contacts w ere d isrupted . F inger 5 m utants p roduced m ore
substantial decreases in the b ind ing ability: the entire finger 5 sw ap m u tan t
(Tp5C) show ed a 10-fold d ro p in binding, while either half resulted in about
four-fold low er affinity. This suggests that som e specific contacts betw een
finger 5 a n d 5S DNA w ere d isrup ted by the m utations. Finally, finger 6
m utan ts sh o w ed an unchanged (in the case of the Tp6 A m utan t), an d an
actually im proved binding (if only by a factor of 1.5) in the case of T p6 B
m utant. It is interesting to note that no single m utan t proved accountable for
the d ram atic reduction in b ind ing seen w h en fingers 4-7 o f TFIIIA are
substitu ted w ith those of p43. Rather, it appears that the in tegrity of each
TFIIIA zinc finger in the 4-6 finger region is required for the m axim um
binding . O f all the m u tan ts tested , finger 5 seem s to p rov ide the m ost
significant contribution to the overall binding affinity to DNA, while finger 4
provides the m ost significant contribution to 5S rRNA binding. The sam ple
b ind ing curves are shown in Figure 7.7.
The nex t tw o sets of m utants were created using Wilms' tum our zinc
fingers as donors (Figure 7.8). Initially, we substitu ted the 4 to 7 region of
TFIIIA w ith zinc fingers 1 to 4 of W Tl. The resu ltan t construct (TF(4-7WT))
d id not have detectable D N A -binding activity, an effect similar to the one
observed w ith the p43 as a donor o f fingers 4-7 (Zang et al., 1995). Again, the
explanation m ay be that this effect is due to the loss of specific protein-D N A
contacts, or to the altered spatial arrangem ent of the zinc fingers in the
context of the full-length protein. In the first set of m utants (WTF4-6), the a -
helical portions of TFIIIA zinc fingers were reintroduced into the TF(4-7)WT
one at a tim e to determ ine precisely which portions of zinc fingers 4-6 were

225
0.8■ Oc3O
<ZQCooco
0.2
0.010*1 0 "
[T FIIIA ], M
1.0
0.8"DC3Oja
0.6<Z
<)/'
QCo 0 .4
O(Su.
0.010*
Figure 7.7 Equilibrium b ind ing c u r y ^ i ^ TFIUA and Tp5B m u tan t proteins
in teracting w ith 5S DNA. Each data po in t is the m ean o f three or m ore
in d ep e n d en t d e te rm in a tio n s, w ith the associated s ta n d a rd d ev ia tio n s
indicated w ith the error bars.

226
a-helix < >- 1 1 2 3 4 5 6
T F I I I A - Z F 4 NIKICVYVCHFENCGECAFKKHNQLKYHQFSHTQW T l - Z F l KRPFMCRYPGCHKRYFKLSHLQMHSRKHTG
T F I I I A - Z F 5 QLPYECPHEGCDKRFSLPSRLKRHEKVHAGWT1-ZF2 EKPYQCDFKDCERRFSRSDQLKRHQRRHTG
T F I I I A - Z F YPCKKDDSCSFVGKTWTLYLKHVAECHQDWT1-ZF3 VKPFQCKT CQRKFSRSDHLKTHTRT HTGKTS
TF 1 1 1 A - Z F 7 LAVCDV CNRKFRHKDYLRDHQKTHEKWT1-ZF4 EKPFSCRWPS CQKKFARSDEVR HHNMHQR
Figure 7.8 Com parison of the amino a d d sequences of zinc fingers 1-4 of
the donor W Tl-ZF with the zinc fingers 4-7 of TFHIA. The structural
cysteines and histidines are shaded.

227
Table 7.4 The effects o f TFIIIA zinc finger substitu tion m utations on the D N A
and RN A b in d in g of the factor
M u ta n t Relative affinity for 5S
rRNA gene
Relative affinity for 5S
rR N A
W TF4 0.06 ± 0.01 <0 .0 1
W TF5 0.04 ± 0.06 <0 .0 1
W TF6 <0 .0 1 <0 .0 1
TFW 4 0.34 ± 0.04 0.16 ± 0 .0 2
TFW 5 0.08 ±0.008 0.48 ± 0.07
TFW 6 0.55 ± 0.037 <0 .0 1
The nom enclature 'WTF' refers to constructs in w hich TFIIIA zinc fingers 4-7
were replaced by fingers 1-4 of the W Tl protein, and, subsequently , a -he lical
reg ions of TFIIIA fingers 4-6 were p laced back. 'T F W refers to TFIIIA
substitu tion m utants in w hich a-helical regions of zinc fingers 4, 5, and 6 are
replaced w ith those of the W Tl zinc fingers 1, 2, and 3.
Relative affinities were calculated using the m ean values o f the association
constants determ ined for the m utant p ro teins (each value represen ting the
m ean o f three independen t determ inations), and com pared against the value
of the w ild-type TFIIIA association constant (Ka=1.9 x 10^ M'^ for DNA; Ka=l-4
X 109 M-l for RNA).

228
necessary to restore the w ild-type DNA-binding activity of TFIIIA. The results
o f filter-b inding assays show n in Table 7.4 reveal th a t no single finger could
rescue the b ind ing of TF(4-7)WT to a significant degree. M utan t WTF6 d id
no t p roduce any im provem ent in binding, suggesting tha t it e ither is the least
im portan t finger for DNA binding, or that it couldn’t establish any contacts
d u e to the im posed struc tu ra l constraints. M utants WTF4 a n d WTF5 d id
show som ew hat im proved binding: their affinities w ere about 17-fold and 22-
fo ld lo w er re la tiv e to the w ild -type TFIIIA, respectively . T his sm all
im provem ent is no t surprising since m ore than one TFIIIA finger is replaced
in a g iven construct.
In the second set of m utants, the a-helical portions of the W Tl zinc
fingers w ere in troduced into the fingers 4, 5, and 6 o f the fu ll-length TFIIIA.
The re su lta n t m u ta n t p ro teins w ere designated TFW4, TFW5, an d TFW6 .
A gain, the largest reduction in b inding affinity to 5S DNA w as seen for the
finger 5 m u tan t (about 13-fold), the next largest - for finger 4 (abou t 3-fold),
follow ed by finger 6 (less that 2-fold). These results are consistent w ith the
effects we observed using other m utants in this study.
7.3.2 Effects o f zinc finger su b s titu tio n m utagenesis on the R N A b ind ing
activity of TFIIIA.
In para lle l to determ in ing the effects of the zinc finger m utants on
DNA binding , w e also m easured the affinities of these m utan t p ro teins for 5S
rRNA. The TF(4-7)WT d id not specifically bind to 5S rRNA, a lthough it
exhibited a fairly h igh nonspecific b inding for the m olecule (Figure 7.9). The
resu lts of the RNA b ind ing stud ies are sum m arized in Tables 7.3 and 7.4.
M ost o f the m utations presented in Table 7.3 (m utants w hich incorporate p43
sequences) d id no t d isru p t RNA binding appreciably, displaying alm ost wild-

229
T3C3O 0.8 -
<zal
0.6
COLO 0 .4 -
co4-»uraL_u_
0 . 2 -
tRNA, M
F ig u re 7.9 C om petition assay for 5S rRNA binding using tRNA^^G as a
com petitor. O pen circles represen t TFIIIA; closed circles represent T F(4-7)WT
proteins. Each data point is the m ean of three independent determ inations,
w ith the associated standard deviations indicated w ith the error bars.

230
type affinities for RNA. The exception is m utan t Tp>4B, w hich sh o w ed an
alm ost 10-fold drop in binding. The substitu tion of the first 15 am ino acids in
finger 5 p roduced a 2-fold reduction in binding, while sw apping its a -he lica l
15 am ino acids resulted in an almost w ild-type affinity. The two categories of
m utants in Table 7.4 (created using W Tl sequences as a donor) reveal the
follow ing: m u tan ts W TF4, WTF5, an d WTF6 d id n o t recover a n y RNA-
b inding activ ity of the TF(4-7)WT protein . This is no t surprising , since an
extensive reg ion of zf 4-7 has been replaced w ith exogenous (W Tl) sequences
in those constructs. The second category of m utants (TFW4-6) show ed tha t
w hile substitu tions o f the a-helical reg ions in fingers 4 and 5 p ro d u ced
approxim ately 6 - and 2 -fold reduction in binding, respectively, replacem ent of
a-helical reg ion in finger 6 abolished RNA binding. The reason w h y th is
resu lt is so d ram atica lly d ifferen t from the one u s in g a s im ila r p43
substitu tion m utan t o f finger 6 is that unlike p43, W T l p ro tein doesn 't have
am ino acid homology to TFEELA. in finger 6 sequence (except for the structural
am ino acids). Figure 7.6. p43, on the other hand, is equ ipped w ith am ino acid
triplet TWT, located a t the tip of finger 6 . This triplet am ino a d d sequence is
conserved betw een IFLLlA and p43 (Joho e t al., 1990; C lem ens et al., 1993). In
fact, these three amino a d d s in finger 6 are the only ones, apart from the zinc-
coord inating cysteines an d histidines an d the structural arom atic residues
that are conserved betw een the two proteins. The role of these am ino acids in
sequence-specific RNA recognition by TFIIIA and p43 needs to be investigated
further.
7.4 D iscussion
Since the dem onstra tion that D N A - and R N A -binding activ ities of
TFIIIA cou ld be separated betw een the different zinc finger subsets of the

231
protein , TFIIIA began to be view ed as a fusion betw een tw o p ro teins th a t
specifically evo lved to interact w ith the tw o different nucleic acids.
H ow ever, the results of m any recent studies conclusively dem onstrate
th a t all the zinc fingers are im portan t for contributing to the overall DNA-
and RN A -binding energy. Just as im portantly, the zinc fingers d o no t provide
equal con tribu tions to the b ind ing to e ither DNA or RNA. A lthough zinc
fingers 1 to 3 w ere show n to dom inate in DNA binding , substitu tion of zinc
fingers 4-7 w ith the ir coun terparts from p43 p ro te in resu lt in a 100-fold
reduction in D N A -binding affinity (Zang e t al., 1995), w hile m ain ta in ing
w ild-type R N A -binding affinity. We undertook this study to determ ine the
contribution o f each zinc finger in the central zinc finger trip le t o f TFIIIA to
specific D N A a n d RNA bind ing . Several sets o f TFIIIA m u ta n ts w ere
generated, in w hich specific zinc finger sequences w ere replaced w ith those of
p43 or W ilm s' tum our proteins. N itrocellulose filter b inding assay was used
to quantitatively determ ine affinities of the m utan t proteins for b o th nucleic
adds.
A nalysis of the D N A -binding d a ta for the Tp series o f m u tan ts
indicated th a t the D N A -binding affinity correlated to that o f the w ild-type
TFIIIA in the follow ing order: Tp6 B > w t TFIIIA = Tp6 A > Tp4A > Tp4B >
Tp5A > Tp5B > Tp5C. The largest reduction in DNA affinity w as observed for
m u tan t Tp5C (10-fold). Sim ilarly, substitu tion of the finger 5 a -h e lix by the
corresponding sequence of the W Tl p ro tein (m utant TFW5) resu lted in a 13-
fold reduction o f D N A -binding affinity. These findings are consistent w ith
the current m odel o f TFIIIA - DNA binding, in w hich finger 5 in teracts w ith
the major groove of the interm ediate elem ent (Clemens et al., 1992). In fact, it
w as show n th a t a zinc finger peptide containing fingers 1-5 b o u n d DNA w ith
higher affinity th an full-length TFIIIA (Clemens e t al., 1994). A dditionally , a

232
strong foo tp rin t w as obtained for finger 5 b ind ing site (C lem ens et al-, 1994).
'Broken finger’ experiments also identified a significant contribution of finger
5 to DNA binding (Del Rio et al., 1993). The structural d isru p tio n of the finger
resulted in alm ost 7-fold decrease in D N A -binding affin ity (Del Rio e t al.,
1993). The ten- to thirteen-fold reduction in DNA b in d in g th a t we observed
reflects a d isrup tion of up to four hydrogen bonds that finger 5 makes in the
in term ediate elem ent of the ICR (assuming tha t an energetic cost of a single
hydrogen bond is reflected in a three-fold d rop in b ind ing affinity, as well as
that energ ies of ind iv idual bonds are additive). M éthy la tion interference
experim ents previously dem onstrated that finger 5 m akes tw o major groove
contacts w ith guanines on the non-coding DNA strand (C lem ens et al., 1992),
w hich also agrees well w ith o u r findings. C om plem entary da ta from the
DNA m utagenesis experiments conducted in ou r laboratory (You et al., 1991;
Veldhoen e t al., 1994) identified that the IE, and , in particu la r, tw o base pair
positions w ith in the IE are im portan t for TFIIIA interaction (GC base pairs 70
and 71). These data agree well w ith the findings that finger 5 is im portant in
prom oter recognition by TFIIIA.
Finger 4 m utants p roduced a relatively m odest decrease in the DNA
binding: betw een 2 - and 3-fold. Clemens et al. (1994) show ed that addition of
finger 4 to the th ree am ino-term inal zinc fingers d o es n o t con tribu te
significantly to DNA binding. M oreover, hydroxy-radical stud ies have show n
that finger 4 m akes closest contacts with the m inor groove of the DNA (Hayes
6 C lem ens, 1992). In terestingly , structural d is ru p tio n of finger 4 in the
broken finger' experim ents low ered the D N A -binding affin ity by as m uch as
9.3-fold (Del Rio e t al., 1993). However, no a lterations in the footprinting
pattern w ere observed for the finger 4 TFIIIA m utan t in the sam e study. It
w as thus assum ed th a t finger 4 does not m ake any specific contacts w ith

233
D N A , bu t ra ther plays a role as a linker w hich positions adjacent fingers over
the 5S DNA. In a recent study, McBryant e t al. (1996) found tha t m utations in
finger 4 DNA binding sequence resulted in a two-fold decrease in b ind ing
affinity , w hich is in concordance w ith ou r data. Interestingly, a b ind ing site
selection and amplification assay perform ed by the sam e g roup (McBryant et
al., 1996) failed to recover w ild-type finger 4 binding site sequence, bu t instead
resu lted in a selection of purine-rich sequence. This finding suggests tha t the
overa ll DNA structure, ra ther than its particu lar sequence is the p rim ary
de term inan t of finger 4 b inding affinity. M oreover, chem ical m odifications
in the major groove of the finger 4 b ind ing site did not affect high-affinity
D N A binding of zf 1-5. These results further support the previously proposed
m odel in which finger 4 binds across the DNA m inor groove (Clemens e t al.,
1992; Hayes & Tillius, 1992).
A lthough both fingers 4 and 6 w ere proposed to in teract w ith the
m in o r groove o f the DNA, the m u ta tions in the tw o fingers p ro d u ced
nonequivalent results: while finger 4 m utan ts had an approxim ately tw o-fold
reduc tion in binding, substitu tions in finger 6 show ed an im proved DNA
b in d in g (Tp series of m utants). This m ay indicate that the tw o zinc fingers
p lay distinct roles in DNA recognition by TFIIIA. There doesn 't seem to be an
unan im ous agreem ent in the literature as to the energetic con tribu tion of
finger 6 to the overall D N A -binding affinity. Thus, C lem ens et al. (1994)
repo rted that addition of finger 6 to the 1-5 zf peptide w eakened its b ind ing to
D N A by about three-fold. However, a loss of the binding affinity was detected
in the broken-finger' experim ents (Del Rio et al., 1993), w hich dem onstrated
a 3.4-fold drop for finger 6 m utant (versus a 9.3-fold drop for finger 4). It is of
in te res t to note that the am ino acid sequence of finger 6 o f TFIIIA deviates
m ost significantly firom the consensus sequence established for fingers 1-3, as

234
w ell as from certain sequence elem ents o f fingers 4 to 9 (Jacobs, 1992).
Perhaps, d u e to this uniqueness finger 6 o f TFIIIA adopts a d istinct m ode of
DNA interaction in the context of the full-length protein.
W ith respect to the high-affinity RNA binding, m ost o f the TFIIIA - p43
m utants (Tp4-6) retained the majority of the RNA-binding affiiüty. It is easily
explained g iven that bo th proteins specialize in b ind ing to the sam e RNA
target, 5S rRNA. In addition, amino a d d s TWT located in the recognition a -
helix of TFIIIA are conserved a t the sam e position in finger 6 of p43, thus
retaining the specific RNA-binding activity . M utants th a t show ed reduced
R N A -binding activity include Tp4B m u ta n t, w hich h ad ab o u t eigh t-fo ld
low er affinity than the w ild-type TFIIIA, and Tp5A m u tan t w ith a two-fold
d rop in affinity. The im portance of finger 4 for RNA b ind ing was previously
dem onstrated by many researchers. Thus, Clem ens et al. (1993) show ed that
deletion of zinc finger 4 from the 4-7 zinc finger peptide resu lted in a 20-fold
d ro p in RNA binding. The fact tha t o u r substitu tion m u tan ts p ro d u ced
sm aller effects m ay be attributable to the use of the full-length TFIIIA protein,
in which the loss of certain contacts m ay be com pensated fo r by the rest o f the
protein. D ifferent m ethods of assaying for affinities can also have an effect on
the m easured affinity values. A recent s tu d y by Friesen & D arby (1997) using
a zf 4-7 zinc finger peptide of TFIIIA dem onstra ted that alan ine substitutions
in zinc fingers 4 or 6 of TFIIIA had a 77- and a 38-fold d ro p in affinity for
RN A , respectively . In con trast, f inger 5 a lan ine su b s titu tio n m u ta n t
p roduced only a m odest change in the RNA binding.
Finally, the most dram atic effect o n RNA binding w as revealed upon
su b stitu tio n o f the 15 a -h e lica l am ino acids in zinc fin g er 6 w ith the
co rrespond ing region of W Tl pro tein (m u tan t TFW6 ). T his substitu tion
abolished specific RNA binding, while reta in ing alm ost w ild -type affinity for

235
DNA. This suggests that th e protein w as active in our assays. A loss of RNA
binding due to m utations in a single zinc finger was not previously observed.
This m ay be d u e to the fact tha t other researchers used dono r proteins th a t
have extensive am ino acid sim ilarity w ith TFIIIA, th u s creating m u tan ts
w hich retain significant levels of binding energy. As m entioned earlier, the
three conserved am ino a d d s a t the tip of finger 6 m ay d irectly contribute to
sequence-specific recogn ition of 5S rR N A . Thus, a lth o u g h the th ree -
d im ensional s tru c tu re as opposed to a sp e d fic sequence of 5S rR N A is
p ro p o sed to be the p rim a ry d e te rm in an t for TFIIIA recognition, som e
sequence-spedfic contacts m ay also con tribu te to the interaction. A good
candidate for such an in teraction may be the loop A reg ion of 5S rR N A ,
w hich was show n to provide direct contacts to the finger 6 (Romaniuk, 1989;
Baudin et al., 1991; You e t al., 1991; Setzer e t al., 1996). The contacts of specific
am ino a d d s in zinc finger 6 of TFIIIA n eed to be assigned to in d iv id u a l
residues in 5S rRNA.
In conclusion , th is s tu d y p ro v id es q u a n tita tiv e analysis o f the
contributions o f the central zinc fingers o f TFIIIA to the high-affinity D N A
and RNA binding . No single zinc finger in the central trip le t proved to be
responsible for the d ram atic , 100-fold red u c tio n in D N A binding affin ity
observed w hen all three zinc fingers w ere replaced. R ather, the central zinc
fingers provide an in tegrated contribution to the DNA binding . In add ition ,
w e found th a t m utations in finger 6 a -helix resu lt in a loss of spedfic RNA
binding. Thus, finger 6 contains major determ inants to su p p o rt high-affinity
R N A b in d in g . The d e ta ils of th is p h e n o m e n o n d eserv e fu r th e r
consideration.

236
8.0 Conclusions
N ucleic a d d b ind ing properties of three zinc finger proteins - W Tl-ZF,
EGR-l-ZF a n d TFIIIA w ere exam ined in the course of m y Ph.D. w ork
described in this thesis. Q uantitative m easurem ents of protein-nucleic a d d
equilibria w ere made using a nitrocellulose filter binding assay.
W ilm s' tum our p ro te in is a putative transcription factor of the C 2H 2
zinc finger fam ily, and has four zinc finger motifs. EGR-1 - a prototypical
m em ber of the same fam ily - has three zinc fingers for w hich consensus DNA
binding site has been defined. Despite the fact that both W Tl and EGR-1 can
bind to the sam e sequence, it was not dear w hat the preferred target sequence
for W Tl w as. In order to identify high affinity binding sites for W Tl an d to
determ ine the role of the additional, first zinc finger of W T l, w e conducted a
binding site selection assay (SAAB). The DNA target site h ad the sequence:
G C G -T G G -G C G -N N N N N . The results of this experim ent reveal th a t the
h ig h affinity b ind ing site for W Tl-ZF had the sequence: GCG-TGG-GCG-
(T /G )(A /G )(T/G )N N . A W TIAFI-ZF protein w hich lacks the first zinc finger
w as used as a control, an d show ed no sequence preferences at any o f the
random ized positions. W Tl-ZF protein had a four-fold higher affinity for the
selected sequence versus the non-selected sequence GCG-TGG-GCG-CCC. The
responsiveness of p ro m o te rs containing e ith e r selected o r non-se lec ted
sequences w as m easured in transien t transfection assays using MTV-CAT
reporter system . These experim ents revealed that the low er affinity CCC site
w h e n p re s e n t in th re e tan d e m ly rep e a te d copies con fers g re a te r
responsiveness to W T l th a n does the h igh affinity TGT site. F u rth e r
experim ents are necessary to clarify these phenomena.
Five p o in t m utations in zinc fingers 2 and 3 of W Tl w ere isolated in
DDS patients. It was therefore suggested that these m utations m ay in terfere

237
w ith the norm al function of W Tl by e ither lowering its affinity for DNA
ta rg e t sites, o r conferring new DNA b ind ing specificities. To test these
possibilities, SAAB experim ents were conducted for the DDS m utant p ro te ins
u sing tw o D N A templates: one w ith finger 2 random ized subsite, the o ther-
w ith finger 3 random ized subsite. None o f the five DDS m utan t p ro te ins
selected new , high-affinity DNA binding sequences. This suggests th a t the
observed DDS phenotype is likely not due to a change in gene ne tw orks
regu la ted by W Tl. M utant R394W had no detectable D N A binding. The
rem aining m utan ts show ed altered DNA b inding selectivities. These m u tan t
p ro teins h ad reduced DN A binding affinities, ranging from 1.4- to 14-fold.
T hus, even sm all changes in DNA-binding affinity can resu lt in a phenotype
seen in DDS patients.
A num ber of equilibrium binding param eters for W Tl-ZF and EG Rl-
ZF proteins w ere m easured using a quantitative nitrocellulose binding assay.
The equilibrium dissociation constants were determ ined to be 1.14 ± 0.2 x 10
-0^ M for W Tl-ZF, and 3.55 ± 0.4 x IQ'09 M for EGRl-ZF p ro te in u n d e r the
fo llow ing conditions: p H 7.5, O.IM KCl a n d incubation a t 22°C. P o in t
m utational anaysis was perform ed to m easure the contribution of each base
p a ir to h ig h affinity b in d in g for the tw o proteins. W hile ex h ib iting
sim ilarities, im portan t différencies were revealed in the b ind ing m echanism s
o f the tw o proteins, w ith respect to both therm odynam ics a n d DNA sequence
preferences. Thus, W Tl-ZF-D N A in terac tion appears to be m ostly an
e n tro p y d r iv e n process w hile EG Rl-ZF-D N A in te rac tio n is d r iv en by
enthalpy. Placing adenine a t position 8 of the consensus b ind ing site resu lted
in 4.5-fold increase in W Tl-ZF D N A -binding affinity w hile the EG R-l-ZF
affinity d ropped by a factor of 12 suggesting that the two pro teins may act on
distinct prom oter sequences.

238
Transcription factor n iA is necessary for the expression o f the 5S rRNA
genes an d the storage of the 5S rRNA molecules. TFIQA has th u s bo th DNA
an d RN A binding activities. The roles of zinc fingers 4 th ro u g h 7 in DNA
an d R N A binding w ere determ ined. The substitu tion of zinc fingers 4-7 of
TFIIIA w ith those of W T l resulted in the loss of D N A binding. The majority
of m u ta tio n s in ind iv idual zinc fingers p roduced m odest effects o n either
D N A or RNA binding. The largest reduction o f D N A binding affinity was
observed w hen finger 5 of TFIIIA w as m utated (10-fold), w hereas substitution
of a n a -h e lica l region o f finger 6 abolished RN A b ind ing activ ity . These
find ings suggest tha t the energetic contributions o f TFIIIA zinc fingers for
b ind ing to either DNA or RNA are no t equal. Thus, finger 6 is indispensable
for h ig h affinity RNA binding, while no single finger in the zf reg ion 4-7 has
the sam e effect on DNA binding of the factor.

239
9.0 Literature cited
Aggarwal, A.K., Rodgers, D.W., Drottar, M., Ptashne, M ., H arrison, S.C. (1988). "Recognition of a DNA operator by the repressor of phage 434: a view at high resolution." Science 242: 899-907.
A nant, S., A xenovich, S.A., M adden, S.L., Rauscher HE, F.J. and Subram anian, K.N. (1994). "N ovel replication inhibitory functions o f th e developm ental reg u la to r / tran scrip tion repressor p ro te in W Tl en co d ed by the W ilms' tum or gene." Oncogene 9: 3113-3126.
A ndersen , J. a n d Delihas, N . (1986). "C harac te rization of R N A -protein in te rac tions in 75 RNP from Xenopus laevis oocytes." T he Journal of Biological C hem istry 261: 2912-2917.
A ndrew s, M. T. an d Brown, D. D. (1987). "Transient activation of oocyte 55 RNA genes in Xenopus em bryos by raising the level of the trans-acting factor TFIIIA." C sii 51: 445-453.
Baird, P.N., G roves, N., Haber, D.A., Housm an, D.E. an d Cow ell J.K. (1992a). "Identification o f m utations in the W Tl gene in tum ours from patien ts w ith the WAGR syndrom e." Oncogene 7: 2141-2149.
Baird, P.N., Santos, A., G roves, N., Jadresic, L. and C ow ell, J.K. (1992b). "C onstitu tional m utations in the W Tl gene in pa tien ts w ith D enys-D rash syndrom e." H u m an M olecular Genetetics 1:301-305.
Baird, P.N. a n d Cowell, J.K. (1993). "A novel zinc finger m u ta tion in a pa tien t w ith D enys-D rash syndrom e." H um an M olecular G enetics 2: 2193- 2194.
B audin, F., Rom by, P., Rom aniuk, P. J., Ehresm ann, B. an d Ehresm ann, C. (1989). "C rosslinking of transcription factor TFIIIA to ribosom al 55 RNA from X. laevis by trans-d iam m ined ich lo rop latinum (II)." N ucleic Acids Research 17: 10035-10046.
Bazett-Jones, D. P. and Brown, M. L. (1989). "Electron M icroscopy reveals that transcription factor TFIIIA bends 55 DNA." M olecular an d Cell Biology 9 (1): 336-341.
B eaudette , N .V ., & Langerm an, N. (1980). CRC C ritic a l R eview s in Biochemistry. CRC Press, Cleveland, OH, pp 145-169.

240
Berg, J. M. (1988). 'T roposed structure for the zinc-binding dom ains from transcrip tion factor HIA and rela ted proteins." Proceeding of the N ational A cadem y of Sciences of the United States of America 85: 99-102.
Berg, J. M. (1990). "Zinc F inger Dom ains: H ypo theses an d C urren t K now ledge." A nnual Review of Biophysics and Biophysical Chem istry 19: 405-421.
Bhat, R., Worley, P., Cole, A., and Baraban, J. (1992). "A ctivation of the zinc finger encoding gene Krox-20 in ad u lt rat brain: com parison w ith zif268." M olecular Brain Research 13: 263-266.
Bickmore, W.A., O ghene, K., Little, M.H., Seawright, A., v an H eyningen, V. an d H astie , N.D. (1992). "M odulation of DNA b ind ing specificity by a lternative splicing of the Wilms tum or w tl gene transcrip t." Science 257: 235-237.
Bieker, J. J. and Roeder, R. G. (1984). 'Thysical properties a n d DNA-binding sto ich iom etry of a 55 gene-specific transcrip tion factor." The Journal of Biological Chem istry 259: 6158-6164.
Bieker, J.J., M artin, P.L., and R oeder, R.G. (1985). "Form ation of a rate- lim iting interm ediate in 55 RNA gene transcription." Cell 40: 119-127.
Bieker, J. J. and Roeder, R. G. (1986). "Characterization of the nucleotide req u ire m e n t for elim ination of the rate-lim iting step in 55 RNA gene transcription." The Tournai of Biological Chemistry 261: 9732-9738.
Blackwell, T. K. and W eintraub, H . (1990). "Differences an d sim ilarities in D N A -binding preferences of M yoD and E2A pro tein com plexes revealed by b ind ing site selection." Science 250: 1104-1110.
Blake, P R. and Sum m ers, M.F. (1994). "Probing the unusually sim ilar metal co o rd in a tio n sites of retroviral zinc fingers an d iron -su lfu r p ro teins by nuclear magnetic resonance." Advances in Biophysical C hem istry 10: 201-228.
Bogenhagen, D. F., Sakonju, 5. and Brown, D. D. (1980). "A control region in the center of the 55 RNA gene directs specific initiation of transcription: H. The 3’ border of the region." Cell 19: 27-35.
Bogenhagen, D. F. and Brown, D. D. (1981). "N ucleotide sequences in Xenopus 55 DNA required for transcription term ination." Cell 24: 261-270.

241
Bogenhagen, D. F., W orm ington, W. M. and Brow n, D. D. (1982). "Stable transcrip tion com plexes of Xenopus 55 RNA genes: a means to m aintain the differentiated state." Cell 28:413-421.
Bonetta, L., K uehn, S.E., Huang, A., Law, D.J., Kalikin, L.M., Koi, M., Reeve,A.E., Brownstein, B.H., Yeger, H., Williams, B.R.G., and Feinberg, A.P. (1990). "W ilm s tum our locus on l lp l3 defined by m ultip le CpG island-associated transcripts." Science 250: 994-997.
Bonventre, J.V., Sukhatm e, V.P., Bamberger, M., O uellette, A.J., and Brown,D. (1991). "Localization of the p ro tein product of the im m ediate early growth response gene, Egr-1 in the kidney after iscem ia and reperfusion ." C ell Regulation 2: 251-260.
Bordereaux, D., Fichelson, S., Tam bourin, P., a n d S. G isselbrecht. (1990). "A lternative splicing of the Evi-1 zinc fingers gene generates m RNAs which differ by the num ber of zinc finger motife." O ncogene 5: 925-927.
Borel, F., Barilla K., Hamilton T., Iskandar, M., Rom aniuk, P. (1996). "Effects o f D enys-D rash syndrom e point m utations on the DNA binding activity of the W ilm s’ tum our suppressor pro tein W Tl." B iochem istry 35: 12070-12076.
Bradeesy, N ., Zabel, B., Schmitt, K., and Pelletier, J. (1994). "W Tl m utations associated w ith incom plete Denys-Drash syndrom e define a dom ain predicted to behave in a dom inant-negative fashoin." G enom ics 2 1 : 663-665.
Bradford, M. M. (1976). "A Rapid and Sensitive M ethod for the Q uantitation of M icrogram Q uan tities of Protein Utilizing the Principle of Protein-D ye Binding." Analytical Biochemistry 72: 248-254.
B rodeur, G.M. (1990). "N euroblastom a: clinical significance of genetic abnorm alities." Cancer Surveys 9: 673-688.
B row n, D. D., C arro ll, D. and Brown, R. D. (1977). "The iso lation and characterization of a second oocyte 5S DNA from Xenopus laevis." Cell 1 2 : 1045-1056.
Brown, R. S., Sander, C. and Argos, P. (1985). "The prim ary structure of transcrip tion factor TFIIIA has 12 consecutive repeats." FEBS (Federation of E uropean Biochemical Societies) Letters 186: 271-274.

242
B ruening, W ., Bardeesy, N ., S ilverm an, B.L., Cohn, R.A., A ronson , A.J., H o u sm an , D., a n d Pelletier, J. (1992). "G erm line in tron ic an d exonic m u ta tio n s in th e W ilm s’ tu m o r gene (W Tl) a ffec tin g u ro g e n ita l developm ent." N atu re Genetics 1 : 144-148.
B ruening, W., and Pelletier, J. (1996). "A non-AUG transla tional in itiation even t generates novel W Tl isoform s." The Tournai of Biological C hem istry 271: 8646-8654.
Call, K.M., G laser, T., Ito, C.Y., Buckler, A.J., Pelletier, J., H aber, D.A., Rose, E.A., Krai, A., Yeger, H., Lewis, W .H., Jones, C. and H ousm an, D.E. (1990). 'Iso la tio n and characterization of a zinc finger polypeptide gene a t the hum an chrom osom e 11 W ilm s’ tum or locus." Cell 60: 509-520.
Cao, X., Koski, R., a n d Gashler, A. M cKiernan, M., M orris, C F., Gaffney, R., H ay, R.V., and Sukhatm e, V.P. (1990). 'Identification and characterization of the EGR-1 gene p roduct, a D N A -binding zinc finger p ro te in induced by differentiation and grow th signals." M olecular and Cellular Biology 10: 1931- 1939.
Caricasole, A., D uarte, A., Larsson, S.H., Hastie, N.D., Little, M., H olm es, G., T odorov , I. and W ard , A. (1996). "RNA binding by the W ilm s tum or supp resso r zinc finger p ro teins." Proceedings of the N ational A cadem y of Sciences of the U nited States of America 93: 7562-7566.
C havrier, P., Zerial, M., Lemaire, P., A lm endral, J., Bravo, R., and Charney, P.(1988). "A gene encoding a p ro tein w ith zinc fingers is activa ted d u ring G O /G l transition in cultured cells." The EMBO (European M olecular Biology O rganization) Journal 7: 29-35.
Cho, Y., G orina, S. Jeffrey, P.D., N.P. Pavletich. (1994). "Crystal structure of a p53 tu m o r su p p re sso r-D N A com plex : u n d e rs ta n d in g tu m o rig e n ic m utations." Science 265; 346.
C hoo, Y. and K lug, A. (1993). "A role in DNA b ind ing for the linker sequences of the first three zinc fingers of TFIIIA." Nucleic Acids Research 21 (15): 3341-3346.
Choo, Y. and Klug, A. (1994a). "Tow ard a code for the interactions of zinc fingers w ith DNA: selection o f random ized fingers d isp layed on phage." P roceedings of the N ational A cadem y of Sciences of the U n ited States of Am erica 91: 11163-11167.

243
Choo, Y. a n d Klug, A. (1994b). "Selection of DNA b ind ing sites fo r zinc fingers u s in g ra tionally random ized DN A reveals coded in te rac tio n s ." P roceedings of the N ational Academ y of Sciences o f the U nited S ta tes of America 91: 11168-11172.
Choo, Y., Sanchez-Garcia, I., and Klug, A. (1994c). "In vivo repression by a site-specific D N A -binding protein designed against an oncogenic sequence." N ature (London) 372: 642-645.
Choo, Y., a n d Klug, A. (1995). "D esigning D N A -binding proteins o n the surface of filamentous phage." C urrent O pinion in Biotechnilogy 6 : 431-436.
Choo, Y., a n d Klug, A. (1997). "Physical basis of a protein-D N A recognition code." C urren t Opinion in Structural Biology 7: 117-125.
C hristensen, J. H., H ansen, P. K., Lillelund, O. and Thogersen, H. C. (1991). "Sequence-Specific B inding of the N -T erm inal Three-F inger F ragm en t of X enopus T ranscrip tion Factor-UIA to the In ternal C on tro l Region o f a 5S RNA G ene." FEBS (Federation of E uropean Biochemical Societies) Letters 281 (1-2): 181-184.
C hristian sen , J., Brown, R. S., Sproat, B. S. and G arre tt, R. A. (1987). "X enopus transcrip tion factor UIA b in d s p rim arily a t junctions be tw een double helical stems and internal loops in oocyte 5S RNA." EMBO (E uropean M olecular Biology Organization) Tournai 6 : 453-460.
Christy, B., Lau, L., and N athans, D. (1988). "A gene activated in m ouse 3T3 cells by serum growth factors encodes a pro tein w ith "zinc finger" sequences." Proceedings of the N ational Academ y of Sciences of the U nited S tates .of America 85: 7857-7861.
Christy, B. an d Nathans, D. (1989). "D N A binding site of the grow th factor- indudb le p ro te in Zif268." Proceedings of the National A cadem y of Sciences of the U nited States of America 8 6 : 8737-8741.
Churchill, M. E. A., Tullius, T. D. and K lug, A. (1990). "M ode of in teraction o f the z inc finger p ro te in TFIIIA w ith a 5S RNA gene of X e n o p u s ." P roceeding o f the N ational Academ y of Sciences of the U nited States__of America 87 (7): 5528-5532.
Ciliberto, G ., Castagnoli, and Cortese, R. (1983a). "T ranscrip tion by RNA polym erase HI." Current Topics in D evelopm ental Biology 18: 59-87.

244
C iliberto , G., Raugei, G., Costanzo, F., Dente, L. and C ortese, R. (1983b). "C om m on a n d in te rchangeab le elem ents in the p ro m o te rs of genes transcribed by RNA polymerase HI." Cell 32: 725-733.
C lem ens, K. R., Liao, X. B., Wolf, V., Wright, P. E. and Gottesfeld, J. M. (1992). "D efinition of the binding sites o f individual zinc fingers in the transcrip tion factor niA-5S RNA gene complex." Proceedings of the N ational Academ y of Sciences of the U nited States of America 89 (22): 10822-10826.
Clem ens, K. R., W olf, V., McBryant, S. J., Zhang, P. H., Liao, X. B., W right, P.E. an d Gottesfeld, J. M. (1993). "Molecular basis for specific recognition of both RNA and D N A by a zinc finger protein." Science 260 (5107): 530-533.
C lem ens, K. R., Z hang , P., Liao, X., M cBryant, S. J., W righ t, P. E. a n d G ottesfeld , J. M. (1994). "Relative contributions of the zinc fingers of transcrip tion factor HIA to the energetics o f DNA b ind ing ." Tournai o f M olecular Biology 244: 23-35.
Colem an, J.E. (1992). "Zinc Proteins: enzymes, storage proteins, transcription factors, and replication proteins." Annual Review of Biochem istry. 61: 897- 964.
C oppes, M.J., Bonetta, L., H uang , A., H oban , P., C hilton-M acN eill, S., Cam pbell, C.E., W eksberg, R., Yeger, H., Reeve, A.E., and W illiam s, B.R.G. (1992a). "Loss o f heterozygosity m apping in W ilm s' tu m o r indicates the involvem ent of th ree distinct regions and a lim ited role for non-disjunction or m itotic recom bination." Genes Chrom osom es Cancer 5: 326-334.
C oppes, M.J., Liefers, G.J., Higuchi, M., Zirm, A.B., Balfe, J.W., and W illiams,B.R.G. (1992b). "Inherited W T l m utations in D enys-D rash syndrom e." Cancer Research 52: 6125-6128.
C oppes, M.J., Cam pbell, C.E. and Williams, B.R. (1993). "The role of W Tl in W ilm s' tum origenesis." FASEB (Federation of A m erican Societies fo r Experim ental Biology) Journal 7: 886-895.
Crosby, S.D., Puetz, J.J., Simburger, K.S., Fahrner, T.J., and M ilbrandt, J. (1991). "The early response gene NGFI-C encodes a zinc finger tran scrip tiona l ac tiva to r and is a m am ber of the GCGGGGGCG (GSG) e lem en t-b ind ing pro tein family." M olecular and Cellular Biology 11: 3835-3841.

245
D’Angio, G.J., Breslow, N ., Bechwith, J.B., Evans, A., Baum, H ., DeLorimier, A., Fernbach, D., H rabovski, E., Jones, B., Kelalis, P., O thersen, B., Tefft, M., an d Thomas, P.R.M. (1989). "T reatm ent of W ilm s' tum or. R esults of the th ird national W ilm s’ tum or study." C ancer 64: 349-360.
D arby, M. K. and Joho, K. E. (1992). "Differential Binding o f Z inc Fingers from Xenopus TFIIIA and p43 to 5S RNA and the 55 RNA Gene." M olecular and Cellular Biology 12 (7): 3155-3164.
Day, M., Fahrner, T., Aykent, S., and M ilbrandt, J. (1990). "The zinc finger protein NGFI-A exists in both nuclear and cytoplasmic forms in nerve grow th factor-stim ulated PC12 cells." The Journal of Biological Chem istry 25: 15253.
Del Rio, S. and Setzer, D. R. (1991). "H igh Yield Purification of Active Transcription Factor UIA Expressed in £ Cali." Nucleic Acids Research 19 (22): 6197-6203.
Del Rio, S. an d Setzer, D. R. (1993a). "The role of z inc fingers in transcriptional activation by transcription factor UIA." Proceedings of the National Academy of Sciences of the U nited States of America 90 (1): 168-172.
Del Rio, S., M enezes, S. R and Setzer, D. R. (1993b). "The function of in d iv id u a l z inc fingers in Sequence-Specific D N A re c o g n itio n by transcription Factor UIA." Journal of M olecular Biology 233 (4): 567-579.
Delihas, N. and A ndersen, J. (1982). "G eneralized struc tu res of the 5S ribosomal RNAs." Nucleic Acids Research 10: 7323-7344.
Delihas, N., A ndersen, J. and Singhal, R. P. (1984). "Structure, function and evolution of 5S ribosom al RNAs." Progress in Nucleic Acid Research and Molecular Biology 31: 161-190.
Denys, P., M alvaux, P., Van Den Berghe, H., Tanghe, W., and Proesm ans, W. (1967). "A sso c ia tio n d u n sy n d ro m e a n a to m o -p a th o lo g iq u e de p se u d o h e rm a p h ro d itism e m ascu lin , d 'u n e tu m e u r de W ilm s, d 'u n e nephropath ie parenchym ateuse et d 'u n m osaïcism e XX/XY." A rchives of French Pediatrics 24: 729-739.
Desjarlais, J. R. and Berg, J. M. (1992). "Tow ard rules relating zinc finger p ro tein sequences and DNA binding site preferences." Proceedings of the N ational Academ y of Sciences of the U nited States of America 89 (16): 7345- 7349.

246
Desjarlais^ J. R. an d Berg, J. M. (1993). "Use of a zinc finger consensus sequence fram ew ork an d specificity ru les to design specific D N A binding p ro te ins." Proceedings of the N ational Academy o f Sciences of the U n ited States o f America 90: 2256-2260.
Dey, B.R., Sukhatm e, V.P., Roberts, A.B., Spom, M B., Rauscher HI, F.J. andKim, S.J. (1994). "R epression of the transform ing g ro w th fac to r-p i gene by the W ilm s’ tum or suppressor W Tl gene product." M olecular E ndocrinology 8:595-602.
D iakun, G. P., Fair all, L. and Klug, A. (1986). "EXAFS study o f the zinc- b ind ing sites in the p ro te in transcription factor HIA." N ature (L ondon) 324: 698-699.
D onehow er, L.A., H arvey, M., Slagle, B.L., M cArthur, M.J., M ontgom ery, Jr.,C .A., Butel. J.S. a n d Bradley, A. (1992). "M ice deficient fo r p53 are developm entally no rm al b u t susceptible to spontaneous tum ors." N a tu re (London) 356: 215-221.
D rash, A ., Sherm an, F., H artm an, W .H . and B lizzard, R.M. (1970). "A sy n d ro m e of p seudoherm aphrod itism , W ilms' tu m o r, h y p e rten sio n , and degenerative renal desease." Tournai o f Pediatrics 76: 585-593.
D rum m ond, I.A., M adden, S.L., Row her-N utter, P., Bell, G.I., Sukhatm e, V.P., and R auscher HI, F.J. (1992). "Repression of the insulin-like g ro w th factor-U gene by the Wilms tum or suppressor W T l." Science 257: 674-678.
D rum m ond , I.A., R upprech t, H.D., R ohw er-N utter. P., L opez-G uiza, J.M., M ad d en , S.L., R auscher III, F.J., a n d Sukhatm e, V.P. (1994). "D N A recogn ition by sp lic ing variants of th e W ilms' tu m o r su p p resso r, W T l." M olecular and Cellular Biology 14: 3800-3809.
Elbert, S.N ., Balt, S.L., H unter, J.P., G ashler, A., Sukhatm e, V.P., a n d W ong,D.L. (1994). "Egr-1 activation in ra t adrenal p h e n y le th an o lam in e N- m ethy ltransferase gene." The Tournai of Biological C hem istry 269: 20885- 20898.
Elrod-Erickson, M., R ould, M.A., N ekludova, L., Pabo, C.O. (1996). "Zif268 p ro tein-D N A complex refined at 1.6 A°: a model system for un d ers tan d in g zinc finger-D N A interactions." S tructure 4: 1171-1180.

247
Engelke, D. R., N g , S.-Y., Shastry, B. S. and Roeder, R. G. (1980). "Sf^cific interaction of a pu rified transcription factor w ith an internal control region of 5S RNA genes." Ceil 19: 717-728.
Englert, C., H ou, X., M aheswaran, S., Bennett, P., Ngw u, C., Re, G.G., G arvin, A.J., Rosner, M.R., and Haber, D A. (1995a). "W Tl suppresses synthesis of the epiderm al g row th factor receptor and induces apoptosis." EMBO (E uropean M olecular Biology Organization) Tournai 14: 4662-4675.
Englert, C., Vidal, M., M aheswaran, S., Ge, Y., Ezzel, R.M., Isselbacher, K.J. and H ab er, D.A. (1995b). "T runcated W Tl m u tan ts a lte r th e subnuc lea r localization of the w ild-type protein." Proceedings of the N ational Academ y of Sciences of the U nited States of America 92: 11960-11964.
E rdm ann, V.A., W olters, J., Pieler, T., D igweed, M., Specht, T., Ulbrich, N. (1987). "Evolution o f organism s and organelles as stud ied by com parative com puter and biochem ical analysis of ribosom al 5S RNA structure ." A nnals N ew York Academ y of Sciences 503: 103-123.
E vans, R.M. (1988). "The s te ro id a n d th y ro id h o rm o n e recep to r superfam ily." Science 240: 889-895.
Fairall, L., Rhodes, D. and Klug, A. (1986). "M apping of the sites of protection on a 58 RNA gene by the Xenopus transcription factor HIA. A m odel for the interaction." Tournai of Molecular Biology 192: 577-591.
Fairall, L., M artin , S. an d Rhodes, D. (1989). "The DNA b ind ing site of the X enopus tran sc rip tio n factor HIA has a non-B-form s tru c tu re ." E M B O (European M olecular Biology Organization) Tournai 8 (6): 1809-1817.
Fairall, L. and R hodes, D. (1992). "A new approach to the analysis of DNase I foo tp rin ting d a ta an d its app lica tion to the TFIIIA /5S D N A com plex." Nucleic Acids Research 20 (18): 4727-4731.
Fairall, L., Schwabe, J. W. R., C hapm an, L., Finch, J. T. an d R hodes, D. (1993). "The crystal structure of a two zinc finger-peptide reveals an extension to the rules for zinc-finger DNA recognition." N ature (London) 366 (6454): 483-487.
Fearon, E.R., a n d Vogelstein, B. (1990). "A genetic m odel for colorectal tum origenesis." Cell 61: 759-767.
Fedoroff, N. V. an d Brown, D. D. (1978). "The nucleotide sequence of oocyte 55 DN A in Xenopus laevis. I. The AT-rich spacer." Cell 13: 701-716.

248
Fraizer, G.E., Bowen-Pope, D.F. an d Vogel, A.M . (1987). "T ranscriptional regu la tion of the h u m an W ilm s’ tum or gene (w tl) ." Tournai o f C ellular Physiology 133:169-174.
Francke, U., Holmes, L.B., Atkins, L., and Riccardi, V.M. (1979). "Aniridia- W ilm s' tum our association: ev id ence for specific de le tion o f l lp lS ." Cytogenetics Cell Genetics 24:185-192.
Frankel, A. D., Berg, J. M. and Pabo, C. O. (1987). "M etal-dependant folding of a sing le zinc finger from transcrip tion factor HIA." P roceed ing of the N ational Academy of Sciences of the United States of America 84: 4841-4845.
Freem ont, P. S. (1993). "The RING finger. A novel p ro tein sequence motif related to the zinc finger." Annals of N ew York Academ y of Sciences 684: 174- 192.
Friesen, W.J. and Darby, M. K. (1997). "Phage d isp lay of RNA b in d in g zinc fingers from transcription factor HIA." The Tournai of Biological Chem istry 272:10994-10997.
Gashler, A.L., Bonthron, D.T., M adden, S.L., Rauscher m , F.J., Collins, T., and Sukhatm e, V.P. (1992). "H um an platelet-derived g row th factor A chain is tran sc rip tio n a lly re p re sse d by th e W ilm s tu m o r su p p re s s o r W T l." Proceedings of the N ational A cadem y of Sciences of the U nited States of America 89: 10984-10988.
G ashler, A.L., Sw am inathan, S., a n d Sukhatm e, V.P. (1993). "A novel repression m odule, an extensive activation dom ain, and a b iparta te nuclear localization signal defined in the im m ediate-early transcrip tion factor EGR- 1." M olecular and Cellular Biology 13: 4556-4571.
Gashler, A.L., and Sukhatm e, V.P. (1995). "Early grow th response protein 1 (EGR-1): Prototype of a zinc finger family of transcrip tion factors." N ucleic Acid Research and M olecular Biology 50: 191-221.
Gessler, M., Poustka, A., Cavenee, W ., Neve, R.L., O rkin, S.H., a n d Bruns, G.A. (1990). "H om ozygous deletion in Wilms' tum ours of a zinc-finger gene identified by chromosome jum ping." N ature fLondon) 343: 774-778.
Gibson, T. J., Postma, J. P. M., Brown, R S. and Argos, P. (1988). "A m odel for the tertia ry structure o f the 28 residue D N A -binding m otif ( 'z inc finger')

249
com m on to m any eukaryotic transcriptional regulatory pro teins." P ro te in Engineering 2 (3): 209-218.
G insberg, A. M., King, B. O. and Roeder, R. G. (1984). "Xenopus 5S Gene T ran sc rip tio n Factor, TFIIIA: C haracterization of a cDNA C lone andM easurem ent of RNA Levels throughout Developm ent." Cell 39: 479-489.
Gogos, J.A., H su, T., Bolton, J., and F.C. Kafatos. (1992). "Sequence discrim ination by alternatively spliced isoform s of a DNA binding zinc finger dom ain." Science 257: 1951-1955.
G oodyer, P., Dehbi, M., Torban, E., Bruening, W. and Pelletier, J. (1995). "R epression of the retinoic acid receptor-alpha gene by the W ilm s’ tum or suppressor gene product, w tl." Oncogene 10:1125-1129.
G uddat, U ., Bakken, A. H. and Pieler, T. (1990). "Protein-M ediated N uclear Export of RNA: 5S rRNA Containing Small RNPs in Xenopus Oocytes." Cell 60 (4): 619-628.
G upta, M.P., Gupta, G., Zak, R., and Sukhatme, V.P. (1991). "EGR-1, a serum - inducible zinc finger protein, regulates transcription of the ra t cardiac alpha- m yosin heavy chain gene." The Tournai of Biological C hem istry 266: 12813- 12816.
Haber, D.A., Sohn, R.L., Buckler, A.J., Pelletier, J., Call, K.M., and H ousm an, D.E. (1991). "A lternative splicing and genom ic s tru c tu re of the W ilm s tum our gene W Tl. " Proceedings of the National Academ y of Sciences of the United States of America 88: 9618-9622.
H aber, D. A., and Buckler, A. J. (1992a). "W Tl: a novel tum or supp resso r gene inactivated in Wilms’ tum our." N ew Biology 4; 97-106.
H aber, D.A., and Housman, D.E. (1992b). "The genetics of W ilm s’ tum or." Advances in Cancer Research 59: 41-68.
H all, R. K. and Taylor, W. L. (1989). "T ranscrip tion Factor HIA G ene Expression in Xenopus Oocytes Utilizes a Transcription Factor Sim ilar to the Major Late Transcription Factor." Molecular and Cellular Biology 9 (11): 5003- 5011.
H allahan, D., Sukhatme, V.P., Sherman, M., V irudachalam , S., Kufe, D., and W eichselbaum , R.R. (1991). 'T ro te in kinase C m ediates X-ray in d u d b ility of

250
nuclear signal transducers EGR-1 a n d Jun ." P roceed ings of the N ational Academy o f Sciences of the United States of America 88: 2156-2160.
H am el, P .A ., P h illip s , R.A., M u n cas te r, M ., and G allie , B. (1993). "Specu la tions on ro les of R bl in tissue-specific d iffe ren tia tio n , tum or initiation, and tum or progression." FASEB ^Federation of American Societies for Experimental Biology) Tournai 7: 846-854.
H am ilton, T.B., Barilla, K.C., and R om aniuk , P.J. (1995). "H igh affinity b inding sites for the W ilm s’ tum or suppresso r protein W T l." Nucleic Acids Research 23: 277-284.
H am ilton, T.B., Borel, F., and Rom aniuk, P.J. (1998). "C om parison of DNA binding characteristics of the related zinc finger proteins W T l and EGR-1." Biochemistry 37: 2051-2058.
Han, K., an d M anley, J. (1993). "Transcriptional repression by the Drosophila even-skipped protein: definition of a m inim al repression dom ain." G enes and Developm ent 7: 491-503.
Hanas, J. S., H azuda, D. J., Bogenhagen, D. F., W u, F. Y.-H. and W u, C.-W. (1983). "Xenopus transcrip tion factor A requires Zinc for b inding to the 5S RNA gene." The Tournai of Biological Chem istry 258: 14120-14125.
H anas, J. S., Bogenhagen, D. F. and W u, C.-W. (1984). "B inding of Xenopus transcription factor A to 5S RNA and to single stranded D N A ." Nucleic Acids Research 12: 2745-2758.
Hansen, P. K., Christensen, J. H., N yborg, J., Lillelund, O. and Thogersen, H. C. (1993). "D issection of the DNA-Binding dom ain of Xenopus laevis TFIIIA - quantitative DNase-I footprinting analysis of specific com plexes betw een a 5S RNA gene fragm ent and N-Term inal fragm ents of TFIIIA containing 3-Zinc Finger, 4-Zinc Finger or 5-Zinc F inger dom ains." Jo u rn a l of M olecu lar Biology 233 (2): 191-202.
H arring ton , M.A., K onicek, B., Song, A., Xia, X.-L., F redericks, W.J. and Rauscher m, F.J. (1993). "Inhibition of colony-stim ulating factor-1 prom oter activity by the product of the Wilms' tum or locus." The Journal of Biological Chem istry 268: 21271-21275.
H arris, H ., M iller, O.J., Klien, G., W orst, P ., and T achibana, T. (1969). "Suppression of m alignancy by cell fusion." N ature fLondon) 223: 363-368.

251
Hastie, N.D . (1994). "The genetics of W ilms' tum or - a case o f d isrup ted developm ent." A nnual Review of Genetics 28: 523-558.
H ayes, J. J. and C lem ens, K. R. (1992a). "Locations of contacts betw een ind iv idual zinc fingers of Xenopus laevis transcrip tion Factor HIA a n d the internal control region o f a 5S RNA gene." Biochemistry 31 (46): 11600-11605.
H ayes, J. J. and T ullius, T. D. (1992b). "Structure of the TF1HA-5S DNA complex." Tournai of M olecular Biology 227 (2): 407-417.
H enry, I., G rand jouan , S., Coullin, P., Barichard, P., H uerre-Jeanpierre , C., G laser, T., Philip, T., Lenoir, G., C haussain , J.L., an d Junien, C. (1989). "Tum or-specific loss o f l lp l5 .5 alleles in d e ll lp lS W ilm s’ tu m o u r and in familial adrenocortical carcinom a." Proceedings of the N ational A cadem y of Sciences USA 86: 3247-3251.
H ew itt, S.M., Fraizer, G.C., Wu, Y.-J., Rauscher in , F.J., and S aunders, G.F. (1996). "D ifferential function of W ilm s’ tum or gene W T l splice isoform s in trnascrip tional regu la tion ." The Journal of Biological C hem istry 271: 8588 - 8592.
Honda, B. M. and Roeder, R. G. (1980). "Association of a 5S gene transcription fac to r w ith 5S R N A a n d a lte re d levels of the factor d u r in g cell differentiation." Cell 22: 119-126.
H ori, H . a n d O saw a, S. (1987). "O rig in and evo lu tion of o rgan ism s as deduced from 5S ribosom al RNA sequences." M olecu lar B iology and Evolution 4 (5): 445-472.
H oubaviy , H.B., U sheva, A., Shenk, T., Burley, S.K. (1996). "C ocrystal s tru c tu re o f YYl b o u n d to the adeno-associa ted v iru s P5 in itia to r." Proceedings of the N a tiona l Academ v of Sciences o f the U nited States of America 93: 13577-13582.
H su, T., Gogos, J.A., K irsh, S.A., an d F.C. Kafatos. (1992). "M ultip le zinc finger form s resulting from developm entally regulated alternative splicing of a transcription factor gene." Science 257: 1946-1950.
Huber, P. W . and W ool, I. G. (1986). 'Identification of the binding site on 5S rRNA for the tran scrip tio n factor HIA: Proposed s tru c tu re of a com m on b ind ing site on 5S rR N A and on the gene." Proceeding of th e N a tio n a l Academ y of Sciences o f the United States_of America 83: 1593-1597.

252
H uber, P. W ., Morii, T., Mei, H. Y. and Barton, J. K. (1991). "S tructural Polym orphism in the M ajor Groove o f a 55 RNA Gene C om plem ents the Zinc F inger Dom ains o f T ranscription Factor HIA." P roceed ings o f the National Academ y of Sciences of the U nited States of America 88 (23); 10801- 10805.
Huff, V., Villalba, F., Strong, L.C., and Saunders, G.F. (1991). "A lteration of the W Tl gene in patients w ith Wilms’ tum or and genitourinary anom alies." Am erican Tournai of H um an Genetics 49: 44.
Isalan, M., Choo, Y., and Klug, A. (1997). "Synergy betw een adjacent zinc fingers in sequence-specific DNA recognition." Proceedings of the N ational Academy o f Sciences of the United States of America 94: 5617-5621.
Jacks, T., R em ington, L., W illiam s, B.O., Schm itt, E.M., H a lach m i, S., Bronson, R.T., and W einberg, R.A. (1994). "Tumor spectrum analysis of p53- m utant m ice." Current Biology 4: 1-7.
Jacobs, G. H . (1992). "Determ ination of the base recognition positions of zinc fingers from sequence analysis." EMBO (E uropean M olecu lar Biology O rganization) Tournai 11 (12): 4507-4517.
Jamieson, A. C., Kim, S. H . and Wells, J. A. (1994). "In vntro selection of zinc fingers w ith altered D N A -binding specificity." B iochem istry 33 (19): 5689- 5695.
Jamieson, A., Hongm ing, W., and Kim, S.-H. (1996). "A zinc finger directory for high-affinity DNA recognition." Biochemistry 93: 12834-12839.
Joho, K.E., Darby, M.K., C raw ford, E.T., an d Brown, D.D. (1990). "A finger pro tein structurally sim ilar to I'FUIA th a t binds exclusively to 5S RNA in Xenopus." Cfiil 61: 293-300.
Joseph, L., Le Beau, M ., and Jam ieson, G. (1988). "M olecular cloning, sequencing, an d m apping of EGR-2, a hum an early g row th response gene encoding a protein w ith "zinc-binding finger ' structure." Proceedings of the National Academ y of Sciences of the U nited States of America 85: 7164-7168.
Kadonaga, J.T., Garner, K.R., Masiarz, F.R., and Tjian, R. (1987). 'Iso la tio n of cDNA encoding transcription factor S p l and functional analysis o f the DNA binding dom ain." CgU 51: 1079-1090.

253
Kassavetis, G. A., Braun, B. R., Nguyen, L. H. and Geiduschek, E. P. (1990). "S. Cerevisiae TFIIIB Is the T ranscrip tion In itiation Factor P ro p e r of RNA Polym erase m. W hile TFIILA and IF lliC Are Assem bly Factors." C ell 60 (2): 235-245.
Kim, C.A., Berg, J.M. (1996). "A 2.2 Â resolution crystal s tru c tu re of a designed zinc finger p ro tein bound to DNA." N atu re S tructural Biology 3: 940-945.
Kim, S. H., D arby, M. K., Joho, K. E. and Brown, D. D. (1990). "The C haracterization o f the TFIILA Synthesized in Som atic Cells o f X enopus laevis" Gene & D evelopm ent 4 (9): 1602-1610.
Kjems, J., Olesen, S. O. and Garrett, R. A. (1985). "Com parison of Eubacterial an d E ukaryotic 5S RNA Structures: A chem ical m odifica tion s tu d y ."Biochemistry 24: 241-250.
Klevit, R. E. (1991). "Recognition of DNA by Cys2, His2 zinc fingers." Science 253 (5026): 1367.
K lug, A. and R hodes, D. (1987). "Z inc fingers: a novel p ro te in m otif for nucleic acid recognitiorL" Trends in Biochemical Sciences 12: 464-469.
K n u d so n , A.G. (1971). "M uta tion an d cancer: s ta tis tica l s tu d y of retinoblastom a." Proceedings of the N ational A cadem y of Sciences of the U nited States of Am erica 68: 820-823.
K nudson, A.G., and Strong, L.C. (1972). "M utation and cancer: a m odel for W ilms' tum or of k idney." Tournai of N ational Cancer Institute 48: 313-324.
Kochoyan, M., H avel, T. F., Nguyen, D. T., Dahl, C. E., K eutm ann, H. T. and W eiss, M. A. (1991a). "Alternating zinc fingers in the hum an m ale associated p ro te in ZFY - 2D N M R structure of a n even finger an d im plications for "jum ping linker" DN A recognition." Biochem istry 30 (14): 3371-3386.
Kochoyan, M., K eutm ann, H. T. and W eiss, M. A. (1991b). "A lternating zinc fingers in the hum an m ale associated p ro te in ZFY - refinem ent o f the NM R structu re of an even finger by selective deuterium labeling and im plications for DNA recognition." Biochemistry 30 (29): 7063-7072.
Kochoyan, M., K eutm aim , H. T. and W eiss, M. A. (1991c). "A lternating zinc fingers in the h um an male-associated p ro tein ZFY - HX3H and HX 4H m otifs encode a local structural switch." Biochemistry 30 (39): 9396-9402.

254
Kochoyan, M ., K eutm ann, H. T. and Weiss, M. A. (1991d). "A rchitectural ru les of the zinc finger motif: Com parative tw o-dim ensional N M R stud ies of na tiv e a n d "arom atic-sw ap" dom ains define a "w eakly p o la r sw itch ”." P roceed ings o f the N ational Academ y of Sciences of the U n ited S tates of America 88: 8455-8459.
Koufos, A., G rundy , P., M organ, K., Aleck, K.A., H adro, T ., L am pkin, B.C., Kalbakji, A ., an d Cavenee, W.K. (1989). "Fam ilial W iedem ann-B eckw ith syndrom e a n d a second Wilms' tum or locus bo th m ap to l lp l5 .5 ." A m erican Tournai of H u m an Genetics 44: 711-719.
K reidberg, J.A., Sariola, H., Loring, J.M., M aeda, M., Pelletier, J., H ousm an, D., a n d Jaenisch, R. (1993). "W Tl is required for early k id n ey developm en t." Cell 74: 679-691.
K riw acki, R. W ., Schultz, S. C., Steitz, T. A. and C aradonna , J. P. (1992). "Sequence-specific recognition of DNA by zinc finger p ep tid es derived from th e tran scrip tion factor S p l." P roceedings o f the N a tio n a l A cadem y of Sciences of the U nited States of America 89 (20): 9759-9763.
Lam aire, P., Revelent, O., Bravo, R , and C ham ay , P. (1988). "Two m ouse genes encod ing po ten tial transcrip tion factors w ith iden tica l D N A -binding dom ains are activated by grow th factors in cu ltu red cells." Proceedings o f the N ational A cadem y of Sciences o f the United States of America 85: 4691-4695.
Larsson, S.H., C haliur, J.-P., M ijagawa, K., Engelkam p, D., R assoulzadegan, M., Ross, A., C uzin , F., van H eyningen, V. and H as tie, N.D. (1995). "Subnuclear localization o f W Tl in splicing o r transcription factor dom ains is regu lated by alternative splicing." Cell 81: 391-401.
Lassar, A. B., M artin, P. L. and Roeder, R. G. (1983). "T ranscription of class m genes: form ation of preinitiation complexes." Science 222: 740-748.
L au, L.F., a n d N athans, D. (1987). "Expression of a se t o f g row th -re la ted im m ediate-early genes in BALB/c 3T3 cells: coordinate reg u la tio n w ith c-fos o r c-myc." Proceedings of the N ational A cadem y of Sciences o f the U n ited States of A m erica 84: 1182-1186.
Lee, M. S., G ippert, G. P., Soman, K. V., Case, D. A. and W right, P. E. (1989). "T hree-d im ensional so lu tion struc tu re of a single zinc fin g er D N A -bind ing dom ain ." Science 245 (4918): 635-637.

255
Lee, M. S., Gottesfeld, J. M. and W right, P. E. (1991). "Zinc is req u ired for folding a n d binding o f a single zinc finger to D N A ." FEES ^Federation of European Biochemical Societies) Letters 279 (2): 289-294.
Lees-M iller ,S.P., and A nderson, C.W. (1991). "The D N A -activated p ro tein kinase, DNA-PK: a potential coordinator of nuclear events." C ancer Cell 3: 341-346.
Lemaire, P., Vesque, C., Schmitt, J., Stunnenberg, H ., Frank, R., a n d C ham ay, P. (1990). "The serum -inducible m ouse gene Krox-24 encodes a sequence- specific transcrip tional activator." M olecular and Cellular Biology 10: 3456- 3467.
Lem erle, J., Voute, P .A ., Tournade, M.F., R odary, C., D elem arre, J.F.M., S irrazin, D., and B urgers, J.M.V. (1983) "Effectiveness of p reo p e ra tiv e ch em o th e rap y in W ilm s' tum or; resu lts of an in te rn a tio n a l society of paediatric oncology (SIOP) clinical trial." Tournai of Clinical O ncology 10: 604- 609.
Levine, M. and Manley, J.L. (1989). "Transcriptional repression o f eukaryotic prom oters." Cell 59: 405-408.
Lewis, W .H., Yeger, H ., Bonetta, L., Chan, H.L.S., Kang, J., Junien, C., Cowell, J., Jones, C., and Dafoe, L.A. (1988). "Hom ozygous deletion of a D N A m arker from chrom osom e l l p l 3 in sporadic W ilm s’ tum or." G enom ics 3: 25-31.
Liao, X. B., Clemens, K. R., Tennant, L., W right, P. E. and G ottesfeld , J. M. (1992). "Specific interaction of the first-three zinc fingers of I'FUIA w ith the internal control region of the Xenopus 5S RNA gene." Tournai o f M olecular Biology 223 (4): 857-871.
Licht, J., G rossel, M., Figge, J., and H ansen, U. (1990). "Drosophila K ruppel protein is a transcriptional repressor." N atu re (Londonl 346: 76-79.
Lim, R. W ., V arnum , B. C., and H erschm an, H. R. (1987). "C lon ing of tetradecanoyl phorbol-ester-induced 'p rim ary response ' sequences an d their expression in density-arrested Swiss 3T3 cells and a TP A non-pro liferative variant." O ncogene 1: 263-270.
Little, M .H ., Prosser, J., Condie, A., Smith, P.J., Van H eyningen, V. a n d Hastie, N.D. (1992). "Zinc finger point m utations w ithin the W Tl gene in W ilm s' tum or p a tien ts ." Proceedings of the N ational A cadem y of Sciences o f the United S tates of America 89: 4791-4795.

256
Little, M .H ., W illiamson, K.A., M annens, M., Kelsey, A., G osden, C., Hastie, N .D . an d V an H eyningen, V. (1993). "Evidence th a t W Tl m uta tions in D enys-D rash syndrom e patients m ay act in a dom inan t-negative fashion." H um an M olecular Genetics 2: 259-264.
Little, M .H., W illiamson, K.A., M armens, M., Kelsey, A., G osden, C., Hastie, N .D ., H einingen, V. van. (1993). "Evidence th a t W Tl m utations in Denys- D rash syndrom e patients may act in a dom inant-negative fashion." H um an M olecular Genetics 2: 259-264.
L ohm an , T., deH aseth , P. L., and Record, M.T. (1980). "Pentalysine- deoxyribonucleic acid interactions: A m odel for the general effects of ion c o n ce n tra tio n s on th e in te rac tions of p ro te in s w ith n u c le ic acids." Biochem istry 19: 3522-3530.
L ohm an, T.M., and M ascotti, D P. (1992). "T herm odynam ics of ligand- nucleic acid interactions." Methods in Enzymology 212: 400-424.
L undin , M., Nehlin, J.O., and Ronne, H. (1994). "Im portance o f a flanking AT-rich region in target site recognition by the GC box-binding zinc finger pro tein M IG l." M olecular and Cellular Biology 14: 1979-1985.
M adden , S.L., Cook, D M., Morris, J.F., G ashler, A., Sukhatm e, V.P. and Rauscher m , J.F. (1991). "Transcriptional repression m ediated by the W Tl WUms tum or gene product. " Science 253: 1550-1553.
M adden, S.L., Cook, D M. and Rauscher HI, F.J. (1993). "A structure-function analysis of transcrip tional repression m ediated by the W Tl, W ilm s tum or suppressor protein." O ncogene 8: 1713-1720.
M ahesw aran, S., Park, S., Bernard, A., Morris, J.F., Rauscher m , F.J., Hill, D.E., and H aber, D A. (1993). "Physical and functional interaction bew een W Tl and p53 proteins." Proceedings of the National A cadem y of Sciences of the U nited States of America 90: 5100-5104.
M ahesw aran, S., Englert, C., Benett, P., Henrich, G., and Haber, D A. (1995). "The W T l gene product stabilizes p53 and inhibits p53-m ediated apoptosis." Genes and Developm ent 9: 2143-2156.
M ao, X. Z. and Darby, M. K. (1993). "A Position-D ependent Transcription- A ctivating dom ain in TFHLA." Molecular Cellular Biology 13 (12): 7496-7506.

257
M artinez, E., Lagna, G. and Roeder, R. G. (1994). "O verlapping transcrip tion by RNA polym erase II an d m of the Xenopus TFIILA gene in som atic cells." The Tournai of Biological C hem istry 269; 25692-25698.
M artinez, J., Georgoff, I., and Levine, A.J. (1991). "C ellular localization and cell cycle regulation by a tem perature-sensitive p53 p ro te in ." G enes and D evelopm ent 5:151-159.
M atsum oto, Y. and K orn, L. J. (1988). "U pstream sequences req u ired for transcrip tion of the TFIILA gene in Xenopus oocytes." Nucleic Acids Research 16:3801-3814.
Maw, M .A., G rundy, P.E., Millow, L.J., Eccles, M.R., D unn, R.S., Sm ith, P.J., Feinberg, A.P., Law , D.J., Paterson , M.C., T elzerow , P.E., C alien , D.F., Thom pson, A.D., R ichards, R.I., an d Reeve, A.E. (1992). "A th ird W ilms' tum or locus on chrom osome 16q." Cancer Research 52: 3094-3098.
M cBryant, S.J., Veldhoen, N., G edulin , B., Leresche, A., Foster, M., W right, P.E., Rom aniuk, P.J. a n d Gottesfeld, J.M. (1995). "In teraction of th e RNA binding fingers of Xenopus transcription factor HIA w ith specific regions of 55 ribosom al RNA." Tournai of M olecular Biology 248: 44-57.
M cBryant, S.J., G edulin , B., Clem ens, K.R., W right, P., and G ottesfeld, J.M. (1996). "A ssessm ent o f major and m inor groove DNA in teractions by the zinc fingers of Xenopus transcription factor HIA." N ucleic A cids Research 24 (13): 2567-2574.
M cConkey, G. A. and Bogenhagen, D. F. (1987). "T ransition m utations w ithin the Xenopus borealis som atic 55 RNA gene can have independen t effects on transcription and TFULA binding." M olecular and C ellular Biology 7: 486-494.
Michael, 5. F., Kilfoil, V. J., Schm idt, M. H., A m ann, B. T. and Berg, J. M.(1992). '^ e t a l b inding and folding properties of a m in im alist Cys2HiS2 zinc finger pep tide ." Proceedings of the N ational A cadem y of Sciences of the United States of America 89 (11): 4796-4800.
M ichalovitz, D., H alevy, O. and O ren, M. (1990). "C onditional inh ib ition of transform ation and of cell proliferation." Cell 62: 671-680.
Miki, Y. (1994). "A strong candidate for the b rea s t an d o v a rian cancer susceptibility gene. " Science 266: 66-71.

258
M ilbrandt, J. (1987). "A nerve grow th factor-induced gene encodes a possible transcrip tional regulatory factor." Science 238: 797-799.
M iller, J. R., C artw right, E. M., Brownlee, G. G., Fedoroff, N. V. an d Brown, D. D. (1978). "The nucleotide sequence of oocyte 55 DNA in Xenopus laevis. H. The GC-rich region." £ d l 13: 717-725.
M iller, J., M cLachlan, A.D. and Klug, A. (1985). "Repetitive zinc-b ind ing dom ains in the p ro te in transcrip tion factor HIA from X enopus oocy tes." EMBO fE uropean M olecular Biology O rganization) Tournai 4: 1609-1614.
M itchell, P.J., a n d T ijian, R. (1989). "T ran sc rip tio n a l reg u la tio n in m am m alian cells by sequence-specific DNA b ind ing proteins." Science 245: 371-378.
M offett, P ., B ruen ing , W., N akagam a, H ., B ardeesy , N ., H o u sm an , D., H ousm an , D.E., and Pelletier, J. (1995). "A ntagonism of W T l activ ity by p ro te in self-association." Proceedings of the N ational Academy of Sciences of the U nited States of America 92: 11105-11109.
M olnar, G., C rozat, A., and Pardee, A.B. (1995). "The im m ediate-early gene Egr-1 regulates the activity of the thym idine kinase prom oter a t the GO - to G1 transition of the cell cycle." M olecular and Cellular Biology 14: 5242-5248.
M orris, J.F., M adden , S.L., Tournay, O.E., Cook, D M ., Sukhatm e, V.P., and Rauscher, F.J. m . (1991). "Characterization of the zinc finger p ro tein encoded by the WTL W ilm s tum or locus." O ncogene 6: 2339-2348.
M uller, H.-J., Skerka, C., Bialonski, A., and Zipfel, P. (1991). "Clone pAT 133 identifies a gene th a t encodes another h u m an m em ber of a class of g row th factor-induced genes w ith alm ost identical zinc-finger dom ains." P roceedings of the N ationa l A cadem y of Sciences of the U n ited States o f A m erica 88: 10079-10083.
N akagam a, H ., H einrich , G., Pelletier, J. and H ousm an , D.E. (1995). "Sequence a n d structu ra l requirem ents for high-affinity DNA b ind ing by the W Tl gene p roduct." M olecular and Cellular Biologv 15: 1489-1498.
N akaseko, Y., N euhaus, D., Klug, A. and Rhodes, D. (1992). "A djacent zinc- finger m otifs in m ultip le zinc-finger pep tides from SWI5 form struc tu ra lly independen t, flexibly linked dom ains." The Tournai o f M olecular Biology 228 (2): 619-636.

259
N ardelli, Gibson, T. J., Vesque, C. and C ham ay, P. (1991). "Base sequence discrim ination by zinc-finger DNA-binding dom ains." N atu re (London) 349 (6305): 175-178.
N ard e lli, J., G ibson, T., and C har nay, P. (1992). "Z inc fin g er - DNA recognition: analysis of base specificity by site-directed m utagenesis." Nucleic Acid Research 20: 4137-4144.
N ehlin , J.O., and Ronne, H. (1990). "Yeast M IG l repressor is re la ted to the m am m alian early g row th response and W ilm s' tum or finger p ro teins." The EMBO (European M olecular Biology Organization) Tournai 9: 2891-2898.
N ekludova, L. & Pabo, C O. (1994). "D istinctive DN A conform ation with en larged major g roove is found in 2In-finger-DNA an d o ther protein-D N A complexes." Proceedings of the National A cadem y o f Sciences o f the United States of America 91: 6948-6952.
N elson , R.M. an d Long, G.L. (1989). "A general m ethod of site-specific m utagenesis u sing a m odification of the Therm us aquaticus po lym erase chain reaction." Analitical Biochemistry 180: 147-151.
N euhaus, D., N akaseko, Y., Schwabe, J. W. R. an d Klug, A. (1992). "Solution s tru c tu re s of tw o Z inc-Finger dom ains from SWI5 o b ta in ed using 2- D im ensional H-1 nuclear m agnetic resonance spectroscopy - a Z inc Finger struc tu re with a 3 rd strand of b-Sheet." The Tournai o f M olecular Biology 228 (2): 637-651.
N guen , H.Q., H offinan-Lieberm ann, B., L ieeberm ann, D A. (1993). "The zinc finger transcription factor EGR-1 is essential for and restricts differentiation along the m acrophage lineage." Cell 72: 197-209.
N ordenskjold, A., Friedm an, E. and Anvret, M. (1994). "W Tl m utations in p a tien ts w ith D enys-D rash syndrom e: a novel m u ta tio n in exon 8 and paternal allele origin." H um an Genetics 93: 115-120.
O 'H allo ran , T. (1993). "T ransition m etals in contro l of gene expression." Science 261: 715-725.
O gaw a, O., Eccles, M R., Szeto, J., McNoe, L.A., Yun, K., Maw, M .A., Smith, P.J., and Reeve, A.E. (1993a). "Itelaxation of insulin-like grow th factor U gene im prin ting im plicated in W ilms’ tum our." N a tu re (London) 362: 749-751.

260
O gaw a, G., Eccles, M R., Yun, K., Mueller, R.F., H oldaw ay, M.D. a n d Reeve A.E. (1993b). "A novel insertional m utation at the th ird zinc finger coding reg ion of the W Tl gene in D enys-D rash syndrom e." H u m an M olecu lar Genetics 2: 203-204.
Om ichinski, J. G., Clore, G. M., Appella, E., Sakaguchi, K. and G ronenbom , A. M. (1990). "H igh-resolution 3-dimensional structure of a single zinc finger from a h um an enhancer b inding protein in solution." B iochem istry 29 (40): 9324-9334.
O m ichinski, J. G., Clore, G. M., Robien, M., Sakaguchi, K., A ppella, E. and G ronenborn , A. M. (1992). "H igh-Resolution Solution S tructu re o f the D ouble Cys2H is2 Zinc Finger from the H um an Enhancer B inding P rotein MBP-1." B ioehem isfrySl (16): 3907-3917.
Pabo, C O., and Sauer, R.T. (1992). "Transcription factors: structural families an d princip les of DNA recognition." Annual Review of B iochem istry 61: 1053-1095.
Palm er, A. G., Ranee, M. and W right, P. E. (1991). "Intram olecular m otions of a zinc finger D N A -binding dom ain from Xfin characterized by p ro ton- detected natural abundance C-12 heteronuclear NMR spectroscopy." Tournai of the A m erican Chemical Society 113 (12): 4371-4380.
Park , S., Tom linson, G., N isen, P. and Haber, D A. (1993). "A ltered trans- activational properties of a m utated W Tl gene product in a W AGR-associated W ilm s' tum or." Cancer Research 53: 4757-4760.
Pârraga, G., H orvath, S. J., Eisen, A., Taylor, W. E., Hood, L., Young, E. T. and Klevit, R. E. (1988). "Z inc-dependent structure of a single finger do m ain of yeast A D R l." Science 241: 1489-1492.
P a tw ardhan , S., Gashler, S., Siegel, M., Chang, L., Joseph, L., Show s, T., Le Beau, M ., and Sukhatm e, V. (1991). "EGR-3, a novel m em ber o f the Egr fam ily o f genes encoding im m ediate-early transcription factors." O ncogene 6: 917-927.
Pavletich, N . P. and Pabo, C. O. (1991). "Zinc Finger-DNA R ecognition - C rystal S tructure of a Zif268-DNA Complex at 2.1Â." Science 252 (5007): 809- 817.

261
Pavletich, N . P. and Pabo, C. O. (1993). "Crystal struc tu re of a 5-Finger GLI- DNA complex - new perspectives on zinc fingers." Science 261 (5129): 1701- 1707.
Pelham, H. R. B. and Brown, D. D. (1980). "A specific transcription factor that can bind e ither the 55 RNA gene or 55 RNA." P roceeding of the N ational Academy of Sciences o f the United States of America 77: 4170-4174.
Pelletier, J., Bruening, W ., Kashtan, C., M auer, M., M anivel, J.C., Striegel, J.E., H oughton, D C., Junien, C., Habib, R., Fouser, L., Fine, R.N., Silverm an, B.L., Haber, D.A., and H ousm an, D. (1991a). "Germline m utations in the W ilms' tum or suppressor gene are associated w ith abnorm al urogenital developm ent in Denis-Drash syndrom e." Cell 67: 437-447.
Pelletier, J., Schalling. M., Buckler, A.J., Rogers, A., H aber, D A. and H ousm an, D. (1991b). "E xpression of the W ilm s tum or gene W T l in the m urine urogenital system ." Genes and D evelopm ent 5: 1345-1356.
Pelletier, J., Bruening, W ., Li, P.P., H aber, D.A., Glaser, T., and H ousm an, D.E. (1991c). "W T l m utations contribute to abnorm al genital system developm ent and hereditary Wilms' tum our." N ature (London) 353: 431-434.
Peterson, R. C., Doering, J. L. and Brown, D. D. (1980). "Characterization of tw o Xenopus somatic 5S DNAs and one m inor oocyte-specific 5S DNA." Cell 20:131-141.
Pfaff, S. L., Hall, R. K., H art, G. C. and Taylor, W. L. (1991). "Regulation of the Xenopus laevis T ranscrip tion Factor HLA G ene D uring Oogenesis an d Early E m bryogenesis - N egative Elem ents R epress the O-TFIIIA P ro m o ter in Embryonic Cells." Developm ental Biology 145 (2): 241-254.
Pfaff, S. L. an d Taylor, W. L. (1992). "C haracterization of a Xenopus Oocyte Factor That Binds to a Developm entally Regulated ds-E lem ent in the TFIILA Gene." Developm ental Biology 151 (1): 306-316.
Picard, B. and W egnez, M. (1979). "Iso lation of a 75 particle from Xenopus laevis oocytes: a 55 R N A -protein com plex ." P roceed ing of the N ational Academ y of Sciences of the United States of America 76: 241-245.
Pieler, T., E rdm ann, V. A. and Appel, B. (1984). "S tructural requirem ents for the in teraction of 5S rR N A w ith the eukaryotic tran scrip tion factor HIA." Nucleic A d d s Research 12: 8393-8406.

262
Pieler, T., Appel, B., O ei, S. L., M entzel, H. and E rdm ann, V. A. (1985a). 'T o in t m u ta tio n a l analysis o f the Xenopus laevis 55 gene p ro m o te r." E M B O (European M olecular Biology O rganization) Tournai 4 (7): 1847-1853.
Pieler, T., Oei, S.-L., H am m , J., Engelke, U. and E rdm ann, V. A. (1985b). "F unctional d om ains o f the Xenopus laevis 55 g ene p rom o te r." E M B O (European M olecular Biology O rganization) Tournai 4: 3751-3756.
Pieler, T., H am m , J. a n d Roeder, R. G. (1987). "The 55 gene in ternal control region is com posed o f three d istinc t sequence e lem en ts, o rgan ized as two functional dom ains w ith variable spacing." Cell 48: 91-100.
Pieler, T. and T heunissen , O. (1993). "TFIIIA: n ine fingers - th ree hands?" TIB5 (Trends in Biochemical 5ciences) 18: 226-230.
Quek, H .H ., Chow, V.T., and Tock, E.P. (1993). "The th ird zinc finger of the W Tl gene is m u ta ted in W ilms' tu m o r but not in a b road range of other urogenital tum ors." A nticancer research 13: 1575-1580.
Q uian, X., Gozan, 5.N ., Yoon, H., Jeon, C., Agarw al, K., Weiss M.A. (1993a) "N ovel z inc finger m o tif in the basal tran scrip tio n m achinery : Three- d im entional NMR stu d ies of the nucleic acid b ind ing dom ain of transcrip tion elongation factor TF115." Biochemistry 32: 9944-9959.
Quian, X., Jeon, C., Yoon, H., Agarwal, K., Weiss, M.A. (1993b) "5 tructure of a new nucleic-acid-binding m otif in eukaryotic transcrip tional elongation factor TFII5." N atu re (London) 365: 277-279.
Rackley, R.R., Flermiken, A.M., K uriyan, N.P., Kessler, P.M., 5toler, M .H ., and W illiams, B.R. (1993). "Expression of the W ilms' tum or supp resso r gene W Tl d u rin g m ouse em bryogenesis." Cell Growth a n d D ifferentiation 4: 1023- 1031.
Rainier, 5., Johnson, L.A., Dobry, C.J., Ping, A.J., G ru n d y , P.E., and Feinberg,A.P. (1993). "R elaxation of im prin ted genes in h u m a n cancer." N a tu re (London) 362: 747-749.
Rauscher, F.J., HI, M orris, J.F., T ournay, O.E., C ook, D M., and C u rran , T. (1990). "Binding of the W ilms tum or locus zinc finger pro tein to the EGR-1 consensus sequence." 5cience 250: 1259-1262.
R auscher, F.J., III. (1993). "The W T l W ilm s tu m o r gene p ro d u c t: a developm entally regu la ted transcrip tion factor in the k idney that fim ctions as

263
a tum or supp resso r." The FASEB (Federation of A m erican Societies for Experim ental Biology) Tournai 7: 896-903.
Raw lings, S.L., M att, G.D., and H uber, P.W. (1996). "Analysis of the binding o f Xenopus transcrip tion factor HIA to oocyte 55 rRNA and to the 55 rRNA gene." The Tournai of Biological Chem istry 271: 869-877.
R ead, D., and J.L. M anley. (1992). "A lternatively spliced transcrip ts of the D roosophila tram track gene encode zinc finger proteins w ith distinct DNA b in d in g specificities." EMBO fE uropean M olecular Biology O rganization) Tournai 11: 1035-1044.
R ebar, E. J. and Pabo, C. O. (1994). "Zinc finger phage - affinity selection of fingers w ith new DNA-Binding specificities." 5cience 263 (5147): 671-673.
Record, M.T., Lohm an, M.L., and H aseth, P. De (1976). "Ion effects on ligand- nucleic acid interactions." Journal of Molecular Biology 107: 145-158.
R eddy, J.C.,Morris, J.C., W ang, J., English, M.A., Haber, D.A., 5hi, Y., and Licht, J. (1995). 'W T l-m ed ia ted transcriptional activation is inh ibited by dom inant negative m u tan t proteins." The Journal of Biological C hem istry 270: 10878- 10884.
R eddy, J.C., and Licht, J.D. (1996). "The W Tl W ilms' tum or suppressor gene: H ow m uch do w e really know?" Biochimica e t Biophysica Acta 1287:1-28.
R eeve, A.E., Eccles, M R., W ilkins, R.J., Bell, G.L, and M illow , L.J. (1985). "E xpression o f insulin-like g row th factor-n transcrip ts in W ilm s tum our." N a tu re (London) 317: 258-260.
R eynolds, W. F. and Gottesfeld, J. M. (1983). "55 rRNA gene transcrip tion fac to r niA alters the helical coitfiguration of DNA." P ro ceed in g of the N ational Academ y of 5ciences of the United 5tates of America 80: 1862-1866.
R hodes, D. a n d K lug, A. (1986). "A n un d erly in g re p e a t in som e transcrip tional control sequences corresponding to half a doub le helical tu rn of DNA." Cell 46: 123-132.
R hodes, D. a n d K lug, A. (1993). "Zinc Fingers." 5 c ien tific A m erican February: 56-65.

264
R iccard i, V.M., Su jansky , E., S m ith , A .C ., a n d F rancke , U. (1978). "C hrom osom al im balance in the an irid ia-W ilm s' tum or association: l i p interstitial deletion." Pediatrics 61: 604-610.
Rollins, M. B., Del Rio, S., Galey, A. L., Setzer, D. R. and A ndrew s, M. T.(1993). "R ole of TFIIIA zinc fingers in v ivo - analysis o f Single-Finger function in developing Xenopus em bryos." M olecular and C ellular Biology 13 (8): 4776-4783.
R om aniuk, P. J. (1985). "Characterization of the RNA b ind ing properties of transcrip tion factor IHA of Xenopus laevis oocytes." Nucleic A cids Research 13 (14): 5369-5387.
R om an iuk , P. J. (1989). "The ro le o f h igh ly conserved sing le-s tranded nucleotides of Xenopus 5S RNA in the binding of transcrip tion factor HIA." Biochem istry 28: 1388-1395.
R om aniuk, P.J. (1990). "Characterization of equilibrium b ind ing of Xenopus tran scrip tio n factor HIA to the 5S RNA gene." The Tournai of Biological C hem istry 265: 17593-17600.
R ow land, O., and Segall, J. (1996). "Interactionof w ild-type and truncated form s o f transcrip tion factor EUA from Saccharomyces cerevisiae w ith the 5S RNA gene." The Tournai of Biological Chem istry 271: 12103-12110.
R u p p rech t, H.D., D rum m ond, I.A., M adden, S.L., R auscher, F.J.ni, and S ukhatm e, V.P. (1994). "The W ilm s' tu m o r su p p resso r gene W T l is negatively au to regu la ted ." The Tournai of Biological C hem istry 269: 6198- 6206.
Russo, M.W., M athny, C., and M ilbrandt, J. (1993). "Transcriptional activity of the zinc finger pro tein NGFI-A is influenced by its interaction w ith cellular factor." Molecular and Cellular Biology 13: 6858-6865.
Ryan, G., Steele-Perkins, V., Morris, J.F., Rauscher HI, F.J. an d Dressier, G.R.(1995). "Repression of Pax-2 by W Tl during norm al kidney developm ent." D evelopm ent 121: 867-875.
Sakai, A., Tadokoro, K., Yanagisawa, H., N agafuchi, S., H osh ikaw a, N., Suzuki, T., Kohsaka, T., Hasegawa, T., N akahori, Y. and Yam ada, M. (1993). "A novel m utation o f the W Tl gene (a tum or suppresso r gene for W ilm s’ tum or) in a pa tien t w ith D enys-D rash synd rom e." H u m an M o le c u la r Genetics 2:1969-1970.

265
Sakonju, S., Bogenhagen, D. F. and Brown, D. D. (1980). "A control reg ion in the center o f the 5S RNA gene directs specific in itiation of transcription: I. The 5' border of the region." Cell 19: 13-25.
Sakonju, S., Brown, D. D., Engelke, D., N g, S.-Y., Shastry, B. S. and Roeder, R. G. (1981). "The binding of a transcrip tion factor to de le tion m utants o f a 58 rRN A gene." CgH 23: 665-669.
Sakonju, S. an d Brown, D. D. (1982). "C ontact po in ts betw een a positive transcription factor and the Xenopus 5S RNA gene." Cell 31: 395-405.
Sam brook, J., Fritsch, E.F., and M aniatis, T. (1989). M olecu lar C lon ing : A Laboratory M anual (Second edition). Cold Spring H arbor, Cold Spring H arbor Laboratory Press.
Sands, M. S. and Bogenhagen, D. F. (1987). "TFIIIA b inds to different dom ains o f 5S RNA and the Xenopus borealis 5S RNA gene." M olecu lar and C ellu lar Biology 7: 3985-3993.
Saw adogo, M. an d R oeder, R. G. (1985). " In terac tion of a gene-specific transcrip tion factor w ith the adenovirus m ajor late p rom oter upstream of the TATA box region." Cell 43: 439-448.
Schm iedeskam p, M., and Klevit, R. (1994). "Zinc finger diversity ." C u rre n t O pinion in Structural Biology 4: 28-35.
Schroth, G. P., Cook, G. R., Bradbury, E. M. and G ottesfeld , J. M. (1989). "T ranscription Factor IHA Induced B ending of the Xenopus Somatic 5S Gene Prom oter." N atu re (London) 340 (6233): 487-488.
Scott, J., Cowell, J., Robertson, M.E., Priestley, L.M., W adey, R., H opkins, B., P ritchard , J., Bell, G.I., Rail, L.B., G raham , C F., a n d K nott, T.J. (1985) "Insu lin -like g ro w th factor-II gene exp ress ion in W ilm s tu m o u rs and em ryonic tissues." N ature (London) 317: 260-262.
Scotto, K. W., Kaulen, H. a n d Roeder, R. G. (1989). "Positive and negative regulation of the gene for transcription factor HIA in Xenopus laevis oocytes." Genes & D evelopm ent 3: 651-662.
Seem an, N .C., Rosenberg, J.M. and Rich, A. (1976). "Sequence-specific recognition of double helical nucleic acids by pro teins." Proceedings o f the N ational Academ y of Sciences of the U nited States of A m erica 73: 804-808.

266
Segall, J., M atsui, T. an d Roeder, R. G. (1980). "M ultiple factors are requ ired for the accurate transcrip tion of purified genes by RNA polym erase HI." T he Tournai o f Biological Chem istry 255 (24): 11986-11991.
Setzer, D. R. and Brown, D. D. (1985). "Formation and stability of the 5S RNA transcrip tion com plex." The Journal of Biological C h em istry 260 (4): 2483- 2492.
Setzer, D R., M enezes, S R., Del Rio, S., H ung, V.S., an d Subram anya, G. (1996). "F unctiona l in teractions b e tw een the zinc fingers of X en o p u s transcrip tion factor IDA during 5S rRNA binding." RNA 2: 1254-1269.
Sharm a, P.M ., Bow m an, M., Yu, B.F., and Sukum ar, S. (1994) "A roden t m odel fo r W ilm s’ tu m o r: em bryonal k idney neop lasm s induced by N- n itroso-N '-m ethyurea. " Proceedings o f the National A cadem y of Sciences of the U nited States of A m erica 91: 9931-9935.
Shastry, B. S., Ng, S.-Y. and Roeder, R. G. (1982). "M ultiple factors involved in the tran sc rip tio n o f class XU genes in Xenopus laevis." The Jou rna l of Biological Chem istry 257: 12979-12986.
Shastry, B. S., H onda, B. M. and Roeder, R. G. (1984). "A ltered levels of a 5S gene-specific transcrip tion factor (TFIIIA) during oogenesis and em bryonic deve lopm en t of Xenopus laevis." The Journal of Biological C h e m is try 259: 11373-11382.
Shi, Y., an d Berg, J. (1995). "Specific DNA-RNA hybrid b inding by zinc finger proteins." Science 268: 282-284.
Silver, P. A., Keegan, L. P., and Ptashne, M. (1984). "A m ino-term inus o f the yeast GAL4 gene p roduct is sufficient for nuclear localization." Proceedings of the N ational A cadem y of Sciences o f the U nited States o f America 81: 5951- 5955.
Sim m ons, D. L., L evy, D. B., Y annoni, Y., and E rikson, R. L. (1989). " Id en tif ic a tio n of a pho rbo l e s te r-rep ress ib le u -s re -in d u c ib le g e n e ." Proceedings of the N ationa l A cadem y of Sciences o f the U nited S tates of Am erica 86: 1178-1182.
Smith, D. R., Jackson, I. J. and Brown, D. D. (1984). "D om ains of the positive transcrip tion factor specific for the Xenopus 5S RNA gene." Cell 37: 645-652.

267
Som ers, W ., U ltsch, M., De-Vos, A., Kossiakoff, A.A. (1994). "The X-ray s tru c tu re o f a g ro w th horm one-pro lac tin receptor com plex." N a tu r e fLondon) 372: 478-481
Stanbridge, E.J. (1990). "H um an tum or suppressor genes." A nnual Review of Genetics 24: 615-657.
Steitz, T.A. (1990). "S tructural studies o f protein-nucleic a d d interaction: the sources of sequence-specific binding." O uaternal Reviews of Biophysics 23 (3): 205-280.
Steitz, T.A. (1993). "S tructural studies of protein-nucleic a d d in teraction." Cam bridge U niversity Press, Cambridge.
S tudier, W.F. an d M offat, B.A. (1986). "Use of bacteriophage T7 RNA polym erase to direct selective high-level expression of d o n e d genes." Tournai of M olecular Biology 189: 113-130.
Studier, F. W ., Rosenberg, A. H., Dunn, J. J. and D ubendorff, J. W. (1990). "G ene E xpression Technology. Use o f T7 RNA polym erase to d irec t expression of d o n ed genes." Methods of Enzymology 185: 60-89.
Suggs, S., K atzow itz, J., Tsai-M orris, C., an d Sukhatme, V. (1990). "cD NA sequence of the hum an early growth response gene Egr-1." Nucleic A cids Research 16: 8835-8846.
Sukhatm e, V.P., K artha, S., Toback, F.G., Taub, R., H oover, R.G., and Tsai- M orris, C.-H. (1987). "A novel earlly g row th response gene rapidly induced by fibroblast, epithelial cell and lymphocyte mitogens." Oncogene Research 1: 343-355.
Sukhatm e, V.P., Cao, X., Chang, L.C., Tsai-M orris, C H ., Stam enkovich, D., Ferreira, P.C.P., Cohen, D R., Edwards, S.A., Shows, T.B., C urran, T., Le Beau, M.M., and A dam son, E D. (1988). "A zinc finger-encoding gene coregulated w ith c-fos d u r in g g ro w th and d iffe ren tia tio n , a n d after c e llu la r depolarization." Cell 53: 37-43.
Suzuki, M. (1993). "C om m on features in DNA recogn ition helices of eukaryotic tran scrip tio n factors." EMBO (E uropean M olecular B iology O rganization) Tournai 12: 3221-3226.
Suzuki, M ., B renner, S.E., G erstein, M ., an d Yagi, N . (1995). "D N A recognition code of transcription factors." Protein engineering 8:319-328.

268
Swirnoff, A.H., and M ilbrandt, J. (1995). "DNA-binding sp e d fid ty of NGFI-A a n d related zinc finger transcription factors." M olecular and Cellular Biology 15: 2275-2287.
Taylor, W., Jackson, I. J., Siegel, N., Kumar, A. and Brown, D. D. (1986). "The developm ental expression o f the gene for TFIILA in Xenopus laevis." N ucleic Acids Research 14: 6185-6195.
T enH arm sel, A ., A uston, R.J., Savenelli, N an d Biggin, M.D. (1993). "C ooperative binding at a distance by even-skipped p ro te in correlates w ith repression and suggests as m echanism of silencing." M olecular and C ellular Biology 13: 2742-2752.
Theunissen, O ., Rudt, P., G uddat, U., Mentzel, H. and Pieler, T. (1992). "RNA a n d DNA b ind ing zinc fingers in Xenopus TFIIIA." Cell 71 (4): 679-690.
T hom pson, C.C. and Evans, R.M. (1989). Proceedings o f the N a t io n a l A cadem y of Sciences of the U nited States of America 86: 3494-3498.
T hukral, S.K., M orrison, M.L., and Young, E.T. (1991). "A lanine scanning site-directed m utagenesis of the zinc fingers of transcrip tion factor A D R l: residues th a t contact DNA and tha t transactivate." P roceed ings o f the N ational Academ y of Sciences of the United States of America 88: 9188-9192.
Thukral, S.K., M orrison, M.L., and Young, E.T. (1992). "M utations in the zinc fin g e rs of A D R l tha t change the specificity of D N A b in d in g a n d transactivation. " Molecular and Cellular Biology 12: 2784-2792.
T on, C.C., H uff, V., Call, K.M., Cohn, S., Strong, L.C., H ousm an, D.E., and Saunders, G.F. (1991). "Smallest region of overlap in W ilms tum or deletions uniquely im plicates an l l p l 3 zinc finger gene as the disease locus." Genom ics 10: 293-297.
Tso, J. Y., Van Den Berg, D. J. and Kom, L. J. (1986). "Structure of the gene for Xenopus transcription factor TFULA." Nucleic Acids Research 14: 2187-2200.
V aranasi, R., Bardeesy, N ., G hahrem ani, M., Petruzzi, M.J., N ow ak, N., A dam , M.A., G rundy, P., Shows, T.B., and Pelletier, J. (1994). "Fine structure analysis of the W Tl gene in sporadic Wilms' tum ors." Proceedings of the N ational A cadem y of Sciences of the United States of America 91: 3554-3558.

269
V eldhoen , N ., You, Q. M ., Setzer, D. R. a n d R om aniuk, P. J. (1994). "C on tribu tion o f individual base pairs to the interaction of TFIUA w ith the Xenopus 55 RN A gene." B iochem istry 33 (24): 7568-7575.
v o n H ippel, F .H ., and Schleich, T. (1969). in S tructure an d S tability of Biological M acromolecules, p p . 417-574, M arcel Dekker Inc., N ew York.
V rana, K. E., Churchill, M. E. A., Tullius, T. D. and Brown, D. D. (1988). "M apping functional regions of transcrip tion factor TFIIIA." M olecular and Cellular Biology 8: 1684-1696.
W ang, Z.-Y., an d Deuel, T. (1992a). "An SI nuclease-sensitive hom opurine- hom opyrim id ine dom ain in the PDGF A -chain prom oter contains a novel b ind ing site for the grow th factor-inducible p ro tein EGR-1." Biochemical and Biophysical Research Com m unications 188: 433-439.
W ang, Z.-Y., M adden, S.L., D euel, T.F., and Rauscher IQ, F.J. (1992b). "The h u m an platelet-derived g row th factor A-chain (PDGF-A) gene is a ta rg e t for rep ression by the W Tl W ilm s tum or p ro te in ." The Journal of Biological Chem istry 267: 21999-22002.
W ang, Z.-Y., Q iu, Q.-Q., Enger, K.T., and D euel, T.F. (1993a). "A second transcrip tionally active D N A -binding site for the Wilms tum or gene p roduct, W T l." Proceedings of the N ational Academy of Sciences of the United States o f America 90: 8896-8900.
W ang, Z.-Y., Q iu, Q.Q., and Deuel, T.F. (1993b). "The W ilm s' tum or gene p ro d u c t W T l activates o r su p p resses tran sc rip tio n th ro u g h se p a ra te functional dom ains." The Tournai of Biological Chem istry 268: 9172-9175.
W ang, Z.-Y., Q iu, Q.Q., H u an g , J., G urrieri, M., and D euel, T.F. (1995). "Products of alternatively spliced transcripts of the Wilms' tum or suppressor gene, w tl , have altered DNA binding specificity and regulate transcrip tion in different w ays." Oncogene 10: 415-422.
W atson, M .A. a n d M ilbrandt, J. (1990). "Expression of the nerve g ro w th fac to r - reg u la te d NGFI-A an d NGFI-B genes in the dev e lo p in g ra t." D evelopm ent 110: 173-183.
W eissm an, B.E., Saxon, P.J., Pasquale, S R., Jones, G.R., G eiser, A.G., an d S tanbridge, E.J. (1987). "In troduction of a norm al hum an chrom osom e 11 in to a W ilm s tum or cell line controls its tum origenic expression." S c ience 236:175-180.

270
W erner, H ., Re, G.G., D rum m ond, I.A., Sukhatm e, V.P., Rauscher HI, F.J., Sens, D.A., G arvin , A.J., LeRoith, D., D., an d Roberts, C.T., Jr. (1993). "Increased expression o f the insulin-like g row th factor I receptor (IGF-I-R) gene in W ilm s tum or is correlated w ith m odula tion of IGF-I-R p rom oter activity b y the W Tl W ilm s' tum or gene p ro d u ct." P roceed ings o f the National A cadem y of Sciences of the United States of America 90: 5828-5832.
W erner, H ., Rauscher m , F.J., Sukhatme, V.P., D rum m ond, I.A., Roberts Jr., C.T., and LeRoith, D. (1994). "Transcriptional repression o f the insulin-like frow th factor I receptor (IGF-I-R) gene by the tum or suppressor W Tl involves b ind ing to sequences b o th upstream and dow nstream o f the IGF-I-R gene transcription start site." The Journal of Biological Chem istry 269: 12577-12582.
W esthof, E., Rom by, P ., Rom aniuk, P. J., Ebel, J.-P., E hresm ann, C. and Ehresm aim , B. (1989). "C om puter m odeling from solu tion data of sp inach chloroplast and of Xenopus laevis somatic and oocyte 5S rR N A s." Tournai of M olecular Biology 207: 417-431.
W ilson, T.E., Day, M.L., Pexton, T., Padgett, K.A., Johnson, M ., and M ilbrandt, J. (1992). "In vivo m uta tiona l analysis of the NGFI-A zinc fingers." T h e Tournai of Biological C hem istry 267: 3718-3724.
W im berly, B., Varani, G. and Tinoco, I. (1993). "The conform ation of Loop-E of eukaryotic 5S Ribosomal RNA." Biochemistry 32 (4): 1078-1087.
W itm an, BCG. (1993). "The retinoblastoma gene: role in cell cycle control and cell d iffe ren tia tio n ." FASEB (F edera tion o f A m erican Societies for Experim ental Biology! Tournai 7: 834-845.
W oiters, J. an d E rdm ann, V. A. (1988). "C om pilation o f 5S rRNA a n d 5S rRNA gene sequences." Nucleic Acids Research 16 (supplem ent): rl-r9.
W orm ington, W. M. (1989). "D evelopm ental Expression and 5S rRN A - Binding A ctiv ity of Xenopus laevis Ribosomal Protein L5." M olecular an d Cellular Biology 9 (12): 5281-5288.
W right, J., G unter, K., M itsuya, H., Irving, S., Kelly, K., a n d Siebenlist, U. (1990). "Expression of a zinc finger gene in HTLV-1 and HTLV-II-transform ed cells." Science 248: 588-591.
W uttke, D.S., Foster, M .P., Case, D.A., Gottesfeld, J.M. and W right, P. (1997). "Solution s truc tu re of the first three zinc fingers of TFIIIA bound to the

271
cognate D N A sequence: determ inants o f affinity and sequence specificity." loum al o f M olecular Biology 273 (1): 183-206.
Xing, Y. Y. and Worcel, A. (1989). "The C-term inal dom ain of transcrip tion factor niA interacts differently with different 5S RNA genes." M olecular and Cellular Biology 9: 499-514.
Ye, Y., R aychaudhuri, B., Gurney, A., Campbell, C.E. and W illiam s, B.R.G.(1996). "Regulation o f W T l by phosphorylationrinhibition of D N A binding, a lte ra tion o f transcrip tional activity and cellu lar translocation ." E M B O (European Molecular Biology Organization) Tournai 15: 5606-5615.
You, Q., Veldhoen, N ., Baudin, F. and Romaniuk, P. J. (1991). "M utations in 5S DNA an d 5S RNA H ave Different Effects on the Binding o f X enopus Transcription Factor IDA." Biochemistry 30 (9): 2495-2500.
Zam betti, G.P., and Levine, A.J. (1993). "A com parison of th e biological activities o f w ild-type and m utant p53." FASEB (Federation o f A m erican Societies for Experimental Biology! Tournai 7: 855-865
Zang, W -Q ., V eldhoen, N. and Rom aniuk, P.J. (1995). "T he effect of substitu tion m utations of transcription factor HIA on nucleic ac id b inding activity." Biochemistry 34: 15545-15552.
Zhang, Y., and Tycko, B. (1992). "Monoallelic expression of the h u m an H19 gene." N atu re Genetics 1: 40-44.


















