
Influence of transient shade periods on the effects of drought on photosynthesis, carbohydrate...
10
Environmental and Experimental Botany 58 (2006) 75–84 Influence of transient shade periods on the effects of drought on photosynthesis, carbohydrate accumulation and lipid peroxidation in sunflower leaves Maria Jo˜ ao Correia ∗ , Maria Leonor Os ´ orio, J ´ ulio Os ´ orio, Isabel Barrote, M´ onica Martins, Maria Manuela David Faculdade de Engenharia de Recursos Naturais (CDCTPV), Universidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal Received 23 June 2005; accepted 23 June 2005 Abstract The effects of a slow-imposing two-week soil drying period, and subsequent re-watering, on leaf water potential (Ψ ), gas exchange rates, chlorophyll fluorescence and on the concentrations of malondialdehyde (MDA) and non-structural carbohydrates (starch, hexose and sucrose) were determined in mature leaves of sunflower plants growing under controlled-environmental conditions. To assess how transient shade periods, associated with increased cloud cover, may influence drought-induced effects on carbon assimilation, measurements were carried out both in plants kept under the photosynthetic photon flux density (PPFD) prevailing during the growth period and stress imposition (750 mol m −2 s −1 ), and in plants subjected to a 5-h long period under a lower PPFD (200 mol m −2 s −1 ). In plants kept under high PPFD, Ψ , stomatal conductance (g), net CO 2 uptake rate (A), the quantum yield of photosystem II electron transport (Φ e ), the photochemical efficiency of open PSII reaction centres (F v /F m ) and the diurnal accumulation of total non-structural carbohydrates (TNC) were significantly depressed at the end of the soil drying period, whereas non-photochemical quenching (NPQ), the concentrations of MDA and the predawn pools of soluble sugars were found to increase. Under high-light level, drought-induced effects on lipid peroxidation, chlorophyll fluorescence parameters and gas exchange rates were fully reversed upon re-watering. However, the inhibition of diurnal accumulation of TNC still persisted two days following re-watering, suggesting that carbohydrate export rates were enhanced following stress relief. An overall positive effect was found upon transferring water-stressed plants to low light level, as indicated by the increases in Ψ , intrinsic water use efficiency (A/g), Φ e and F v /F m , as well as the reversal of drought-induced enhancement of both NPQ and MDA concentration. Despite g being similar in shaded well-watered and re-watered plants, the latter exhibited net CO 2 uptake rates below those found in well-watered leaves under the same light conditions, together with a diurnal decrease in the concentration of TNC that was mainly attributable to the depletion of starch and sucrose. These results indicate that, contrasting with the positive effects of shading on water-stressed plants, low PPFD may negatively affect the recovery of net photosynthesis following stress relief. © 2005 Elsevier B.V. All rights reserved. Keywords: Drought; Helianthus annuus; Non-structural carbohydrates; Photosynthesis; Lipid peroxidation; Shade Abbreviations: A, net carbon dioxide uptake rate; C i , intercellular carbon dioxide concentration; ETR, electron transport rate; F v /F m , maximal photo- chemical efficiency of photosystem II; F v /F m , photochemical efficiency of open PSII reaction centres; g, stomatal conductance for water vapour diffu- sion; MDA, malondialdehyde; NPQ, non-photochemical quenching; PPFD, photosynthetic photon flux density; PSII, photosystem II; q p , photochemical quenching; TNC, total non-structural carbohydrates; Φ e , quantum yield of photosystem II electron transport; Ψ , leaf water potential ∗ Corresponding author. Tel.: +351 289800932; fax: +351 289818419. E-mail address: [email protected] (M.J. Correia). 1. Introduction Water deficit is a major constraint to plant growth and pro- ductivity, one of the earliest effects of soil drying being the decline in photosynthetic carbon assimilation, resulting from stomatal and mesophyll limitations (Lawlor, 2002; Flexas et al., 2004). However, the extent of drought-induced depres- sion in photosynthesis, and its underlying causes, may be dependent on the superimposition of other environmental stress conditions (Chaves, 1991). As water-stress episodes 0098-8472/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.envexpbot.2005.06.015
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Influence of transient shade periods on the effects of drought on photosynthesis, carbohydrate...

Environmental and Experimental Botany 58 (2006) 75–84
Influence of transient shade periods on the effects of drought onphotosynthesis, carbohydrate accumulation and lipid
peroxidation in sunflower leaves
Maria Joao Correia ∗, Maria Leonor Osorio, Julio Osorio, Isabel Barrote,Monica Martins, Maria Manuela David
Faculdade de Engenharia de Recursos Naturais (CDCTPV), Universidade do Algarve,Campus de Gambelas, 8005-139 Faro, Portugal
Received 23 June 2005; accepted 23 June 2005
Abstract
The effects of a slow-imposing two-week soil drying period, and subsequent re-watering, on leaf water potential (Ψ ), gas exchange rates,chlorophyll fluorescence and on the concentrations of malondialdehyde (MDA) and non-structural carbohydrates (starch, hexose and sucrose)were determined in mature leaves of sunflower plants growing under controlled-environmental conditions. To assess how transient shadeperiods, associated with increased cloud cover, may influence drought-induced effects on carbon assimilation, measurements were carriedout both in plants kept under the photosynthetic photon flux density (PPFD) prevailing during the growth period and stress imposition(750 �mol m−2 s−1), and in plants subjected to a 5-h long period under a lower PPFD (200 �mol m−2 s−1). In plants kept under high PPFD, Ψ ,stomatal conductance (g), net CO2 uptake rate (A), the quantum yield of photosystem II electron transport (Φe), the photochemical efficiency ofopen PSII reaction centres (F ′
v/F′m) and the diurnal accumulation of total non-structural carbohydrates (TNC) were significantly depressed at
the end of the soil drying period, whereas non-photochemical quenching (NPQ), the concentrations of MDA and the predawn pools of solublesugars were found to increase. Under high-light level, drought-induced effects on lipid peroxidation, chlorophyll fluorescence parameters andgas exchange rates were fully reversed upon re-watering. However, the inhibition of diurnal accumulation of TNC still persisted two daysfollowing re-watering, suggesting that carbohydrate export rates were enhanced following stress relief. An overall positive effect was foundupon transferring water-stressed plants to low light level, as indicated by the increases in Ψ , intrinsic water use efficiency (A/g), Φe and F ′
v/F′m,
as well as the reversal of drought-induced enhancement of both NPQ and MDA concentration. Despite g being similar in shaded well-wateredand re-watered plants, the latter exhibited net CO2 uptake rates below those found in well-watered leaves under the same light conditions,together with a diurnal decrease in the concentration of TNC that was mainly attributable to the depletion of starch and sucrose. These resultsindicate that, contrasting with the positive effects of shading on water-stressed plants, low PPFD may negatively affect the recovery of net
photosynthesis following stress relief.© 2005 Elsevier B.V. All rights reserved.Keywords: Drought; Helianthus annuus; Non-structural carbohydrates; Photosynth
Abbreviations: A, net carbon dioxide uptake rate; Ci, intercellular carbondioxide concentration; ETR, electron transport rate; Fv/Fm, maximal photo-chemical efficiency of photosystem II; F ′
v/F′m, photochemical efficiency of
open PSII reaction centres; g, stomatal conductance for water vapour diffu-sion; MDA, malondialdehyde; NPQ, non-photochemical quenching; PPFD,photosynthetic photon flux density; PSII, photosystem II; qp, photochemicalquenching; TNC, total non-structural carbohydrates; Φe, quantum yield ofphotosystem II electron transport; Ψ , leaf water potential
∗ Corresponding author. Tel.: +351 289800932; fax: +351 289818419.E-mail address: [email protected] (M.J. Correia).
1
ddsesds
0098-8472/$ – see front matter © 2005 Elsevier B.V. All rights reserved.doi:10.1016/j.envexpbot.2005.06.015
esis; Lipid peroxidation; Shade
. Introduction
Water deficit is a major constraint to plant growth and pro-uctivity, one of the earliest effects of soil drying being theecline in photosynthetic carbon assimilation, resulting fromtomatal and mesophyll limitations (Lawlor, 2002; Flexas
t al., 2004). However, the extent of drought-induced depres-ion in photosynthesis, and its underlying causes, may beependent on the superimposition of other environmentaltress conditions (Chaves, 1991). As water-stress episodes
7 and Exp
itifvopiwPItme
rcpfivdcriwdsIdetidt2osaoiCspsthepdsmr
pBe
pdtedscaKatuthet
aatwlbp5trsa
2
2
oiaTetita((s
tt1
6 M.J. Correia et al. / Environmental
n field conditions are often accompanied by strong light,he interaction between high irradiance and drought has beennvestigated in a number of studies. Ogren and Oquist (1985)ound no interaction between drought and irradiance, pro-ided the leaves were properly light-acclimated. However,ther studies have shown that water deficits predispose thehotosynthetic apparatus to photoinhibition, and drought-nduced inhibition of photosynthetic activity is exacerbatedhen leaves are dehydrated under strong light (Bjorkman andowles, 1984; Masojıdek et al., 1991; Behera et al., 2002).n contrast, Valladares and Pearcy (2002) recently reportedhat the capacity of plants to withstand a severe drought
ight be decreased under shade conditions, rather then beingnhanced.
On studies undertaken to asses the influence of lightegime on the effects of drought, light level is often keptonstant throughout the experimental period. In contrast, thehotosynthetic photon flux density varies temporally undereld conditions. Cloud cover is one of the major causes ofarying sunlight intensity under field conditions, and the inci-ence of transient shade periods, associated with increasedloud cover, increases during precipitation events. Therefore,e-watering of rain-fed plants is expected to occur underrradiances lower than those prevailing during growth andater deficit development. The impact of those transientecreases in irradiance on carbon assimilation under droughttress and after stress relief has been scarcely addressed.n a study in which sunflower plants were subjected to aays-long shade period during water deficits imposition, Bent al. (1987) found that the extent of drought-induced reduc-ion in the CO2-saturated rate of photosynthesis was greatern leaves kept at high-light than in shaded leaves, and norought-induced changes in photochemistry was detected inhe latter. Results obtained in a recent work (Sofo et al.,004), in which the shading treatment was extended through-ut the entire re-watering period, also showed that partialhade alleviates the oxidative load on leaves, and leads tofaster recovery of photosynthesis. Another likely effect
f decreased irradiance would be the limitation of drought-nduced accumulation of sugars (Munns and Weir, 1981).arbohydrate build-up in leaves can lead to decreased expres-
ion of photosynthetic genes, and hence to a depression inhotosynthetic activity (Paul and Foyer, 2001). Therefore,hade-induced limitation in sugar accumulation is expectedo alleviate end-product inhibition of photosynthesis, whichas been reported to occur under drought conditions (Souzat al., 2004). The above mentioned studies point out to aositive effect of shading on photosynthetic activity underrought conditions, but it is conceivable that the depres-ion in the capacity of the leaves to produce carbohydratesay limit the recovery of growth rate after drought stress
elief.
When leaves are subjected to low light, a decreased res-iratory rate is essential to allow a positive carbon balance.ecause acclimation of leaf respiratory rates to the new lightnvironment may take a few days (Noguchi et al., 2001), tem-
oags
erimental Botany 58 (2006) 75–84
orary carbon imbalance is likely to occur following suddenecreases in irradiance, as respiration continues at high ratesypical for high-light grown leaves. This may contribute toxplain why switching high-light grown leaves to low irra-iance conditions has been found to be associated with aubstantial decrease in foliar levels of total non-structuralarbohydrates (Veneklaas and den Ouden, 2005), and to neg-tively affect leaf instantaneous water use efficiency (Fay andnapp, 1995). However, the relative magnitude of the neg-
tive effects of short-term (minutes-long) shade periods onranspiration and photosynthesis, and hence the way waterse efficiency is affected, has been found to be dependent onhe watering regime (Fay and Knapp, 1998). These resultsighlight the importance to proceed studies on the influ-nce of sudden decreases in light level on plant responseso drought stress.
The present work was undertaken in order to contribute tobetter understanding of the short-term effects of low irradi-nce on the response of plants to drought. With that purpose,he effects of soil drying, and subsequent re-watering, onater status, gas exchange rates, chlorophyll fluorescence,
ipid peroxidation and accumulation of non-structural car-ohydrates were determined in mature leaves of sunflowerlants, either kept under growth irradiance or subjected to a-h long period of partial shade. To our knowledge, the simul-aneous effects of temporary shading on leaf gas exchangeates, chloroplast photochemistry, leaf oxidative damage andugar accumulation in water-stressed leaves has not beenssessed so far.
. Materials and methods
.1. Plant material and growth conditions
Helianthus annuus L. seeds (cv. Giant) were soakedvernight and allowed to germinate on wet tissue paper,n darkness. They were then placed in 3 l pots filled with
mixture of unfertilised peat and vermiculite (1:1, v/v).he plants (one per pot) were grown in a controlled-nvironment cabinet (Fitoclima 16.000 EHVP, Aralab, Por-ugal) under a photoperiod of 12 h, with light provided byncandescent and fluorescent lamps, supplying a photosyn-hetic photon flux density (PPFD) of about 750 �mol m−2 s−1
t the top of the plants. The temperature was 18 ◦C/25 ◦Cnight/day) and the relative humidity was 70%/60%night/day). Watering was done using a complete nutrientolution.
From sowing until the beginning of water deficit imposi-ion, all the pots were regularly brought to field capacity (eachwo days). The onset of water deficit imposition took place9 days after sowing, and afterwards the pots were watered
n a daily basis. Water-stress was imposed by replacing onlybout half the water lost by evapotranspiration (determinedravimetrically). After 17 days of soil drying, the water-tressed pots were re-watered to field capacity. In the case
and Exp
oeepapwp
rwstp(o
posw
2r
pC
ctmm
ia(cbcrtwlssipimPcet(w
(ic(wwa
2n
daEtid(u(ratbiwoTma5uttbAic(
pe8meAtwtsf
M.J. Correia et al. / Environmental
f the plants referred to as well-watered, all the water lost byvapotranspiration was replaced by watering throughout thentire experimental period. Measurements and sampling oflants subjected to water deficit took place 10 and 17 daysfter the onset of soil drying. Re-watered plants were sam-led 24 and 48 h following re-watering. Well-watered plantsere sampled the days preceding sampling of water-stressedlants.
Two light treatments were imposed in each wateringegime: half of the plants were kept in the growth chamberhere growth took place (high-light treatment); the plants
ubjected to the low-light treatment were transferred, athe end of the dark period that preceded the day of sam-ling, to another growth chamber in which PPFD was lower200 �mol m−2 s−1). The temperature regime was the samen the two growth chambers.
A different set of plants was sampled on each day (fivelants per treatment), and all measurements were undertakenn fully expanded, non-senescent leaves. Unless otherwisetated, all measurements were carried out 5 h after the lightsere turned-on (midday).
.2. Measurements of leaf water status, gas exchangeates and chlorophyll fluorescence
Leaf water status was assessed by measuring leaf waterotential (Ψ ) using a pressure chamber (PMS Instruments,orvallis, OR, USA).
Stomatal conductance for water vapour diffusion (g), netarbon dioxide uptake rate (A), and intercellular CO2 concen-ration (Ci) were determined using a portable gas exchangeeasuring system (HCM-1000, H. Walz, Effeltrich, Ger-any).Chlorophyll fluorescence measurements were undertaken
mmediately before the lights were turned-on (predawn), andt midday, using a portable pulse-modulated fluorometerPAM-2000, H. Walz). The maximal photochemical effi-iency of photosystem II (PSII) was estimated, at predawn,y the fluorescence ratio Fv/Fm of dark-adapted leaves, cal-ulated from F0 (basal fluorescence) and Fm (maximal fluo-escence), with Fv being the variable fluorescence. The pho-ochemical efficiency of open PSII reaction centres (F ′
v/F′m)
as calculated by measuring the same parameters as above onight-adapted leaves (F ′
0 and F ′m). After recording the steady-
tate value of fluorescence of light adapted leaves (Fs), aaturating flash was imposed to determine the maximal leveln the light-adapted state (F ′
m). Afterwards, a black cloth waslaced over the leaf, and a far-red pulse (3 s) was providedn order to fully oxidise PSII during the measurement of the
inimal level in the light-adapted state (F ′0). The effective
SII quantum efficiency in light-adapted leaves (φe) wasalculated by the (F ′
m − Fs)/F ′m = (�F/F ′
m) ratio (Genty
t al., 1989). As described by Krall and Edwards (1992),he electron transport rate (ETR) was estimated as ETR =�F/F ′m) × PPFD × 0.5 × 0.84. The leaf absorbance usedas 0.84 because that is the most common value for C3 plants
aHpg
erimental Botany 58 (2006) 75–84 77
Bjorkman and Demmig, 1987). The photochemical quench-ng (qp), which was used as an estimate of the fraction of openentres, was calculated according to Bilger and Schreiber1986), whereas the Stern–Volmer equation (Cornic, 1994)as used to calculate non-photochemical quenching (NPQ),hich is an estimate of the thermal energy dissipation
t PSII.
.3. Quantification of malondialdehyde andon-structural carbohydrates
The level of oxidative damage to lipids was assessed byetermining the concentration of malondialdehyde (MDA),product of the oxidation of polyunsaturated fatty acids.xtraction of leaf material, measurements and procedures
o determine the amount of MDA equivalents, corrected fornterfering compounds in leaf extracts, were performed asescribed by Hodges et al. (1999). In short, leaf materialapproximately 100 mg fresh mass) was powdered in liq-id nitrogen and suspended in 1 ml 20% trichloroacetic acidTCA). Each sample had two replicates, two blanks and aeference. Replicates were obtained after homogenizationnd centrifugation (3000 × g at 4 ◦C, during 10 min), byhe addition of 500 �l of supernatant to 1.5 ml of a thio-arbituric acid (TBA) solution made in TCA (0.5% TBAn 20% TCA, v/v). In the case of the blanks, the sampleas replaced by 500 �l of 20% TCA. The reference wasbtained by adding 500 �l of supernatant to 1.5 ml 20%CA. All the samples, references and blanks were sub-itted to a heat/cool cycle and again centrifuged (10 min
t 10,000 × g and room temperature), the absorbances at32, 600 and 440 nm being determined in the supernatantsing a spectrophotometer Shimadzu UV-160. MDA con-ent was calculated by correcting for compounds otherhan MDA, which absorb at 532 nm, such as anthocyanins,y subtracting the absorbance at 532 nm of the reference.bsorbances measured at 600 and 440 nm allowed to take
nto account interference due to non-specific turbidity andarbohydrates, respectively, as described by Hodges et al.1999).
Soluble sugars and starch were quantified in leaf discsunched at predawn and at midday. Soluble sugars werextracted from intact leaf discs, with 80% (v/v) ethanol, at0 ◦C, for 20 min. Sucrose, fructose and glucose were theneasured in the ethanol extract using a spectrophotometric
nzyme-coupled assay, as described by David et al. (1998).fter the extraction of soluble sugars, the leaf discs were
hen used for starch analysis. After being subjected to aashing step, the leaf discs were homogenised with water,
he resulting suspension was autoclaved, and subsequentlyubjected to hydrolysis (Stitt et al., 1989). Glucose releasedrom starch hydrolysis was quantified using a colorimetric
ssay modified to be used in 96 well plates, as described inatterscheid and Willenbrink (1991). Starch in each sam-le, expressed as glucose equivalents, was determined usinglucose solutions as standard. The sum of soluble sugars
7 and Experimental Botany 58 (2006) 75–84
a(
2
wSr
cjhittItdweckc
jetaacAu
c
3
3
icwfidtr
abwwwt
Fig. 1. Leaf water potential (Ψ ) determined in sunflower plants well-watered(WW), after 10 days (S10) or 17 days (S17) of soil drying, and following24 h (R1) or 48 h (R2) of re-watering. Means for plants transferred to low-lt‘w
pt
3
aweAewdb
a5aIs3tpctpup
8 M.J. Correia et al. / Environmental
nd starch is referred to as total non-structural carbohydratesTNC).
.4. Data analysis
Statistical analysis and graphic display were performedith SPSS (Release 12.0, SPSS Inc., Chicago, IL) andigmaPlot (Version 8.02, SPSS Inc.) software packages,espectively.
Data from leaf water status, leaf gas exchange rates,hlorophyll fluorescence and lipid peroxidation were sub-ected to one-way ANOVA after testing for normality andomogeneity of variance. If the ANOVA yielded a signif-cant F value (P < 0.05), then the four individual means ofhe water-stress and re-watering treatments were comparedo the well-watered control using a Dunnett post hoc test.n case of heterogeneity of variances the Kruskal–Wallisest was used for non-parametric analysis of variance, andifferences between watering regimes means were testedith Kruskal–Wallis multiple-comparison Z-value tests. At
ach watering regime, independent-samples t-tests wereomputed to compare the calculated means between plantsept under high-light and those transferred to low-lightonditions.
Data from non-structural carbohydrates were also sub-ected to one-way ANOVA. At each watering regime, differ-nces between the two light regimes and predawn means wereested using Dunnett post hoc tests. In addition, differencest predawn between water-stress and re-watering treatmentsnd the well-watered control were tested in the same way. Inase of heteroscedastic data, Kruskal–Wallis non-parametricNOVA and Kruskal–Wallis multiple-comparison tests weresed instead.
All differences were considered to be statistically signifi-ant if P < 0.05.
. Results
.1. Leaf water status
Restricting the water supplied through watering resultedn the development of a substantial leaf water deficit in thease of plants kept under high-light: when compared to well-atered plants, leaf Ψ decreased only by 0.2 MPa during therst 10 days of limited water supply, but at the end of soilrying period Ψ of water-stressed leaves was 0.9 MPa lowerhan in well-watered ones. Following re-watering Ψ quicklyecovered to control levels.
As shown in Fig. 1, decreasing irradiance positivelyffected leaf water potential, shade-induced increase in Ψ
eing higher in the case of droughted plants than in well-
atered ones. Consequently, the difference in Ψ betweenell-watered and droughted plants, although not eliminated,as attenuated upon switching to low irradiance conditions:he difference in Ψ between well-watered and water-stressed
pArt
ight conditions (dashed bars) that are significantly different (P < 0.05) fromhose for plants kept under high-light (white bars) are marked with symbol*’. Values shown are mean ± standard error of five replicates, except for theell-watered controls in which n = 10.
lants did not exceed 0.6 MPa in the case of plants transferredo low light conditions.
.2. Leaf gas exchange rates
In plants kept under high-light conditions, neither stom-tal conductance (g) nor net carbon dioxide uptake rate (A)ere significantly affected after 10 days of soil drying. How-
ver, as shown in Fig. 2, by the end of the soil drying period,and g were strongly depressed, the values of both param-
ters decreasing to about 10% of values determined in well-atered plants. No significant drought-induced change wasetected at the level of the intercellular concentration of car-on dioxide (Ci) measured under high-light conditions.
In the case of the more severely stressed plants, stom-tal conductance and net photosynthesis increased by about0 and 75%, respectively, in response to shading (Fig. 2),lthough those changes were not statistically significant.n contrast, upon transferring well-watered and moderatelytressed plants to low-light conditions, g decreased by about5%, the concomitant depression in A being even more accen-uated: net photosynthesis of shaded leaves of well-wateredlants was below 30% of values determined under high-lightonditions. As a consequence of the opposite response of Ao shading exhibited by well-watered and severely stressedlants, the negative effects of soil drying on A were alleviatednder low-light conditions: when compared to well-wateredlants under the same light conditions, severely stressed
lants kept under high-light exhibited a 10-fold decline in, whereas under shade conditions A was only halved inesponse to soil drying. The light conditions also influencedhe way A/g ratio and Ci were affected by soil drying: in

M.J. Correia et al. / Environmental and Exp
Fig. 2. Stomatal conductance for water vapour diffusion (g), net carbondioxide uptake rate (A) and intercellular carbon dioxide concentration (Ci)determined in sunflower plants well-watered (WW), after 10 days (S10) or 17days (S17) of soil drying, and following 24 h (R1) or 48 h (R2) of re-watering.Means for plants transferred to low-light conditions (dashed bars) that aresignificantly different (P < 0.05) from those for plants kept under high-light(white bars) are marked with symbol ‘*’. Values shown are mean ± standarderror of five replicates, except for the well-watered controls in which n = 10.
si(vid
sswtedd(utssttlvo
3
mfltos
rpgredriDmcwadprl
lwio
erimental Botany 58 (2006) 75–84 79
everely stressed plants under shade conditions, A/g rationcreased by 10-fold (data not shown) and Ci was halvedFig. 2) compared to well-watered plants, whereas those twoariables did not vary significantly in response to water deficitn the case of the plants kept under the light level prevailinguring the growth period and stress imposition.
Despite the strong drought-induced depression in photo-ynthesis and stomatal conductance, the negative effects ofoil drying on A and g, measured under growth irradiance,ere no longer detected 24 h following re-watering. In con-
rast, re-watered plants transferred to low-light conditionsxhibited net CO2 uptake rates significantly lower than thoseetermined in well-watered plants under the same light con-itions, despite full stomatal reopening following re-wateringFig. 2). Actually, on the day following re-watering, net CO2ptake rates determined in shaded leaves were 25% lowerhan those determined in severely stressed plants under theame light conditions (Fig. 2). The different extent of photo-ynthetic recovery following stress relief was associated withhe enhancement of the negative effects of shading on net pho-osynthesis following re-watering: upon transferring plants toow-light conditions, A decreased to about 25 and 10% of thealues determined in plants kept under high-light in the casef well-watered and re-watered plants, respectively.
.3. Chlorophyll fluorescence
Despite the severity of drought-induced reduction in A, theaximal photochemical efficiency of PSII, estimated by theuorescence ratio Fv/Fm of dark-adapted leaves, was close
o maximum (varying between 0.84 and 0.87), irrespectivef the watering and light regimes to which the plants wereubjected (Fig. 3).
The photochemical activity of light adapted leaves alsoemained unaffected after 10 days of limited water sup-ly. However, in more severely stressed plants kept underrowth irradiance, the photochemical efficiency of open PSIIeaction centres (F ′
v/F′m ratio), the quantum yield of PSII
lectron transport (φe), and the electron transport rate (ETR)ecreased significantly, although moderately: the F ′
v/F′m
atio was decreased by 10%, whereas the extent of drought-nduced depression in φe and ETR reached about 15%.rought-induced restriction in φe was accompanied by aoderate increase in the reduction state of QA, as indi-
ated by a 5% decrease in photochemical quenching (qp),hereas non-photochemical quenching (NPQ) increased by
bout 40%, indicating the enhancement of thermal energyissipation at the PSII. Drought-induced changes in chloro-hyll fluorescence were fully reversible upon water deficitelief, all the fluorescence parameters returning to pre-stressevels on the day following re-watering.
As shown in Fig. 3, upon transferring the plants to low-
ight conditions, φe, the F ′v/F′m ratio and qp increased,
hereas NPQ decreased, the extent of shade-induced changesn fluorescence parameters increasing with the severityf water-stress. Consequently, when measured under low
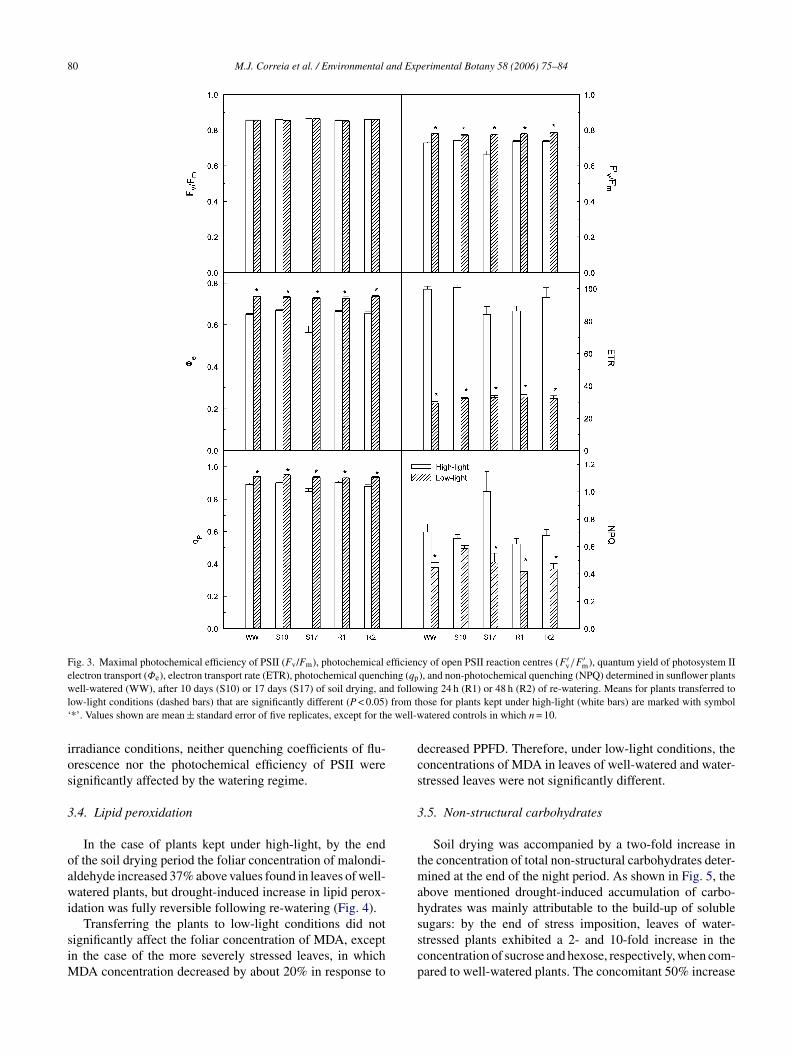
80 M.J. Correia et al. / Environmental and Experimental Botany 58 (2006) 75–84
Fig. 3. Maximal photochemical efficiency of PSII (Fv/Fm), photochemical efficiency of open PSII reaction centres (F ′v/F
′m), quantum yield of photosystem II
electron transport (Φe), electron transport rate (ETR), photochemical quenching (qp), and non-photochemical quenching (NPQ) determined in sunflower plantsw d followl from th‘ e well-w
ios
3
oawi
siM
dcs
3
tmah
ell-watered (WW), after 10 days (S10) or 17 days (S17) of soil drying, anow-light conditions (dashed bars) that are significantly different (P < 0.05)*’. Values shown are mean ± standard error of five replicates, except for th
rradiance conditions, neither quenching coefficients of flu-rescence nor the photochemical efficiency of PSII wereignificantly affected by the watering regime.
.4. Lipid peroxidation
In the case of plants kept under high-light, by the endf the soil drying period the foliar concentration of malondi-ldehyde increased 37% above values found in leaves of well-atered plants, but drought-induced increase in lipid perox-
dation was fully reversible following re-watering (Fig. 4).
Transferring the plants to low-light conditions did notignificantly affect the foliar concentration of MDA, exceptn the case of the more severely stressed leaves, in which
DA concentration decreased by about 20% in response to
sscp
ing 24 h (R1) or 48 h (R2) of re-watering. Means for plants transferred toose for plants kept under high-light (white bars) are marked with symbolatered controls in which n = 10.
ecreased PPFD. Therefore, under low-light conditions, theoncentrations of MDA in leaves of well-watered and water-tressed leaves were not significantly different.
.5. Non-structural carbohydrates
Soil drying was accompanied by a two-fold increase inhe concentration of total non-structural carbohydrates deter-
ined at the end of the night period. As shown in Fig. 5, thebove mentioned drought-induced accumulation of carbo-ydrates was mainly attributable to the build-up of soluble
ugars: by the end of stress imposition, leaves of water-tressed plants exhibited a 2- and 10-fold increase in theoncentration of sucrose and hexose, respectively, when com-ared to well-watered plants. The concomitant 50% increase
M.J. Correia et al. / Environmental and Exp
Fig. 4. Concentrations of malondialdehyde (MDA) determined in leaves ofsunflower plants well-watered (WW), after 10 days (S10) or 17 days (S17) ofsoil drying, and following 24 h (R1) or 48 h (R2) of re-watering. Means forplants transferred to low-light conditions (dashed bars) that are significantlydifferent (P < 0.05) from those for plants kept under high-light (white bars)ar
iiiftfsbt
cttaitpaatnr
Fapdi
re marked with symbol ‘*’. Values shown are mean ± standard error of fiveeplicates, except for the well-watered controls in which n = 10. t
fo
ig. 5. Concentrations of starch, hexose, sucrose and total non-structural carbohydfter 10 days (S10) or 17 days (S17) of soil drying, and following 24 h (R1) or 48lants kept under high-light (white bars), and on those transferred to low-light condetermined at predawn (black bars) are marked with symbol ‘*’. Values shown are mn which n = 10.
erimental Botany 58 (2006) 75–84 81
n the concentration of starch was not statistically signif-cant, but the concentration of this insoluble carbohydratencreased following re-watering, reaching values about two-old higher than those determined in well-watered plants byhe end of the night that followed stress relief. Two daysollowing re-watering the predawn pools of both hexose andtarch returned to pre-stress levels, but significant differencesetween well-watered and re-watered plants still persisted aso their predawn concentrations of sucrose.
In the case of plants kept under growth irradiance, theoncentration of TNC in well-watered leaves increased bywo-fold from predawn to midday (Fig. 5). The above men-ioned carbohydrate build-up was mainly attributed to theccumulation of starch and hexose, whose concentrationsncreased by two- and six-fold, respectively, from predawno midday. As shown in Fig. 5, by the end of the soil dryingeriod no significant difference was found between predawnnd midday concentrations of TNC, due to drought-inducedbolishment of diurnal build-up of starch and decrease inhe extent of hexose accumulation. The inhibition of diur-al accumulation of TNC still persisted two days followinge-watering (Fig. 5).
In the case of well-watered and water-stressed plantsransferred to low irradiance conditions, no significant dif-erence was detected between predawn and midday valuesf starch and soluble sugars concentrations. However, on the
rates (TNC) determined in leaves of sunflower plants well-watered (WW),h (R2) of re-watering. Means for concentrations determined at midday, initions (dashed bars), that are significantly different (P < 0.05) from meansean ± standard error of five replicates, except for the well-watered controls

8 and Exp
dadwp
4
4i
m(teesttpttΦ
twiwgpapctr(
drdnoenttdpruoSado
ba
iipepdft1elbmAebw(ttfr
toidTtisQ(sbcpta
4s
paiip
2 M.J. Correia et al. / Environmental
ay following re-watering, the concentrations of TNC werepproximately halved, from predawn to midday. The referredecrease in the concentration of carbohydrates was associatedith the diurnal depletion of starch and sucrose, the latter stillersisting two days following re-watering (Fig. 5).
. Discussion
.1. Effects of water deficits in plants kept under growthrradiance conditions
Although the restriction in water supply was extended forore than two weeks, leaf Ψ never dropped below −1.5 MPa
Fig. 1), indicating a level of leaf water deficit less severehan that reported for field-grown sunflower plants (Freedent al., 1991; Pankovic et al., 1999). Despite that, by thend of the soil drying period, net CO2 uptake rate wastrongly depressed (Fig. 2). As shown in Fig. 3, PSII pho-ochemistry was also negatively affected, as indicated byhe decrease in the quantum yield of PSII electron trans-ort (Φe), but the ratio Fv/Fm did not change, indicatinghat non-reversible photoinhibitory damage to PSII reac-ion centres did not occur. Drought-induced depression in
e was largely a result of decreased efficiency of excita-ion energy capture by open PSII reaction centres (F ′
v/F′m),
hat may reflect light-induced non-photochemical quench-ng (Baker, 1991). In accordance, a significant rise in NPQas observed in the more severely stressed plants kept underrowth irradiance (Fig. 3). These results provide evidence thatart of the excitation energy in PSII antennae was dissipateds heat, through the activation of non-radiative dissipativerocesses, such as those associated with the xanthophyllsycle (Demmig-Adams and Adams, 1992). That may con-ribute to avoid photoinhibitory damage, and to the fastecovery of photosynthetic activity following stress reliefFig. 2).
Since incident PPFD was kept constant, drought-inducedecrease in Φe was accompanied by a similar decrease in theate of electron transport (Fig. 3). However, the extent of ETRepression was much smaller than the concomitant decline inet CO2 uptake rate (Fig. 2). Similar results were obtained inther studies (Flexas et al., 1998; Wingler et al., 1999; Osoriot al., 2006), and were interpreted as indicating an increase inon-assimilatory electron transport processes, such as pho-orespiration and the Mehler-peroxidase reaction. Althoughhese processes may have a protective role by decreasing theegree of reduction of the PSII acceptor side and the risk ofhotoinhibition, both processes result in the production ofeactive oxygen species (Foyer et al., 1994). In plants keptnder high-light, a significant increase in foliar concentrationf MDA was detected by the end of soil drying period (Fig. 4).
ince MDA is a product of lipid peroxidation (Hodges etl., 1999; Sofo et al., 2004), these results indicate that watereficit imposed to sunflower plants resulted in a moderatexidative stress at the whole leaf level, similarly to what hassIct
erimental Botany 58 (2006) 75–84
een reported for other species (Behera et al., 2002; Jiangnd Zhang, 2002).
In accordance with the strong drought-induced depressionn A (Fig. 3), the extent of carbohydrates accumulation dur-ng the day was much higher in the leaves of well-wateredlants than in leaves of severely stressed plants, but the latterxhibited higher predawn levels of TNC than well-wateredlants (Fig. 5). The increase in the predawn pool of carbohy-rates observed in severely stressed plants is likely to resultrom decreased rates of export, which have been reportedo be inhibited under water deficits conditions (Quick et al.,992; Pelleschi et al., 1997). Drought-induced inhibition ofxport rates apparently persisted during the first night fol-owing re-watering, since the pool of foliar TNC determinedy the end of that night did not decrease below values deter-ined at midday, before the plants were re-watered (Fig. 5).fterwards however, carbohydrate export rates were appar-
ntly promoted by re-watering. This contention is supportedy the observation that, in contrast to well-watered plants, re-atered plants exhibited no daytime carbohydrate build-up
Fig. 5), although net CO2 uptake rates were similar on thewo groups of plants (Fig. 2). That apparent increase in day-ime export rates may be related with increased sink demandor photo-assimilate due to growth recovery after stresselief.
Water deficit imposed in the present work also affectedhe contribution of soluble sugars and starch to the total poolf carbohydrates: as shown in Fig. 5, in response to soil dry-ng, the contribution of starch to the predawn pool of TNCecreased, and the diurnal starch build-up was abolished.hese results are in agreement with previous studies showing
hat water deficits inhibit starch synthesis and induce a shiftn the partitioning of newly formed carbohydrates towardsoluble sugars (Freeden et al., 1991; Zrenner and Stitt, 1991;uick et al., 1992). In accordance with Zrenner and Stitt
1991), who found that the inhibitory effect of drought ontarch synthesis was rapidly reversed following rehydration,y the end of the first night after re-watering starch con-entration increased above values found in severely stresslants, this increase in starch content being proportional tohe concomitant decrease in the concentration of soluble sug-rs (Fig. 5).
.2. Influence of shading on the effects of soil drying andubsequent re-watering
The results obtained in the present work show an overallositive effect of short-term shading on droughted plants. Inccordance with Sofo et al. (2004), who evidenced the abil-ty of shading in limiting drought-induced oxidative damagen field-grown olive trees, drought-induced increase in lipideroxidation was fully reversed upon the transfer of water-
tressed sunflower plants to low-light conditions (Fig. 4).n addition, no drought induced-inactivation of the photo-hemical activity of PSII was detected in sunflower plantsransferred to low-light conditions (Fig. 3), similarly to what
and Exp
hs
oiasri(mbotanas
srsolwraCtw
ffercThmoA(witCwTitsiisip
cit
pmttrbawkwobdpdw
A
3aUa
R
B
B
B
B
B
B
C
C
C
M.J. Correia et al. / Environmental
as been previously found in others studies with this plantpecies (Ben et al., 1987; Pankovic et al., 1999).
Shade-induced reversal of drought-induced inactivationf photochemical efficiency was accompanied by increasedntrinsic water use efficiency, as evidenced by Ci beingpproximately halved following the transfer of severelytressed plants to low irradiance conditions (Fig. 2). Theseesults are apparently contradictory with previous reportsndicating that shading negatively affects water use efficiencyFay and Knapp, 1995). These apparent contradictory resultsay be conciliated taking into account that, similarly to what
een previously found by Fay and Knapp (1998), the effectsf shading on A/g in sunflower leaves were dependent onhe watering regime: when well-watered, moderately stressednd re-watered plants were transferred to shade conditions,et photosynthesis was inhibited to a higher extent than stom-tal conductance, in contrast with what occurred in moreeverely stressed plants (Fig. 2).
Contrasting with the positive effects of shading on water-tressed plants, low irradiance may negatively affect theecovery of net CO2 uptake rates following re-watering. Ashown in Fig. 2, stomatal reopening following re-wateringccurred irrespective of the light conditions to which theeaves were subjected, but the recovery of A after stress reliefas strongly dependent on light conditions: whereas full
ecovery of net photosynthetic rate occurred on the first dayfter re-watering on plants kept under growth irradiance, netO2 uptake rates determined in re-watered plants transferred
o low-light did not surpassed half the values determined inell-watered plants under a similar light level.Since the persistence of drought-induced inhibition of A
ollowing rehydration were restricted to measurements per-ormed in plants transferred to low-light, we might hypoth-sise that respiratory processes were enhanced followinge-watering. Haupt-Herting et al. (2001) reported that mito-hondrial respiration in the light is inhibited by water-stress.o our knowledge, the effect of rehydration on respirationas not been assessed so far, although increased demand foraintenance respiration may be expected after stress relief, in
rder to support the repair of dehydration-induced damages.s to photorespiration, according to Cornic and Fresneau
2002), an absolute increase in photorespiration is expectedhen leaves are dehydrated under limiting light. Drought-
nduced increases in photorespiration have been attributedo depleted Ci due to stomatal closure (Wingler et al., 1999;ornic and Fresneau, 2002; Noctor et al., 2002). In the presentork, stomata fully reopened following re-watering (Fig. 2).herefore, even if photorespiration had been increased dur-
ng the period of low g, it should be expected to returno control values following re-watering and concomitanttomatal reopening. However, the occurrence of drought-nduced increase in mesophyll diffusive resistance may result
n depressed chloroplastic CO2 concentration irrespective oftomatal resistance (Flexas et al., 2004). Whether drought-nduced limitation in mesophyll conductance occurred in theresent study, and persisted following stress relief, then it isD
erimental Botany 58 (2006) 75–84 83
onceivable the maintenance of a high photorespiration raten re-watered leaves, despite the relief of stomatal limitationo CO2 diffusion.
It is note worthy that, under low-light conditions, the totalool of foliar carbohydrates was halved from predawn toidday on the day following re-watering (Fig. 5). This deple-
ion of foliar carbohydrate can not be entirely explained byhe negative effect of shading on carbon assimilation, sincee-watered plants under low-light exhibited A values lowut positive (Fig. 2). Therefore, these results may indicaten enhancement of carbohydrate export rates following re-atering, as already suggested by the data obtained on plantsept under high-light. It is important to note that these resultsere obtained by measurements performed in a different setf plants each day, whereas field-grown rain-fed plants maye subjected to several consecutive cloudy days during rehy-ration periods. It should deserve future investigation theossibility of the cumulative effects of prolonged shadinguring re-watering leading to foliar carbohydrate starvation,hich in turn may limit growth recovery.
cknowledgments
This work was supported by the project POCTI/AGR/6145, which was funded by the Foundation for Sciencend Technology (FCT, Portugal) and by FEDER (Europeannion). The authors thank Joao Azedo-Silva for his technical
ssistance in this research.
eferences
aker, N.R., 1991. Possible role of photosystem II in environmental pertur-bations of photosynthesis. Physiol. Plant 81, 563–570.
ehera, R.K., Misha, P.C., Choudhury, N.K., 2002. High irradiance andwater stress induce alterations in pigment composition and chloroplastactivities of primary wheat leaves. J. Plant Physiol. 159, 967–973.
en, G.Y., Osmond, C.B., Sharkey, T.D., 1987. Comparisons of photo-synthetic responses of Xanthium strumarium and Helianthus annuusto chronic and acute water stress in sun and shade. Plant Physiol. 84,476–482.
ilger, W., Schreiber, U., 1986. Energy-dependent quenching of dark-levelchlorophyll fluorescence in intact leaves. Photosyn. Res. 10, 303–308.
jorkman, O., Powles, S.B., 1984. Inhibition of photosynthetic reactionsunder water stress: interaction with light stress. Planta 161, 490–504.
jorkman, O., Demmig, B., 1987. Photon yield of O2 evolution and chloro-phyll fluorescence characteristics at 77 K among vascular plants ofdiverse origin. Planta 170, 489–504.
haves, M.M., 1991. Effects of water deficits on carbon assimilation. J. Exp.Bot. 42, 1–16.
ornic, G., 1994. Drought stress and high light effects on leaf photosynthesis.In: Baker, N.R., Bowyer, J.R. (Eds.), Photoinhibition of Photosynthesis:from Molecular Mechanisms to the Field. BIOS Scientific Publishers,Oxford, pp. 297–313.
ornic, G., Fresneau, C., 2002. Photosynthetic carbon reduction and carbon
oxidation cycles are the main electron sinks for photosysthem II activityduring a mild drought. Ann. Bot. 89, 887–894.avid, M.M., Coelho, D., Barrote, I., Correia, M.J., 1998. Leaf age effects onphotosynthetic activity and sugar accumulation in droughted and rewa-tered Lupinus albus plants. Aust. J. Plant Physiol. 25, 299–306.

8 and Exp
D
F
F
F
F
F
F
G
H
H
H
J
K
L
M
M
N
N
O
O
P
P
P
Q
S
S
S
V
V
W
4 M.J. Correia et al. / Environmental
emmig-Adams, B., Adams, W.W., 1992. Photoprotection and otherresponses of plants to high light stress. Annu. Rev. Plant Physiol. PlantMol. Biol. 43, 599–626.
ay, P.A., Knapp, A.K., 1995. Stomatal and photosynthetic responses toshade in sorghum, soybean and eastern gamagrass. Physiol. Plant 94,613–620.
ay, P.A., Knapp, A.K., 1998. Responses to short-term reductions in light insoybean leaves: effects of leaf position and drought stress. Int. J. PlantSci. 159, 805–811.
lexas, J., Escalona, J.M., Medrano, H., 1998. Down-regulation of photo-synthesis by drought under field conditions in grapevine leaves. Aust. J.Plant Physiol. 25, 893–900.
lexas, J., Bota, J., Loreto, F., Cornic, G., Sharkey, T.D., 2004. Diffusive andmetabolic limitations to photosynthesis under drought and salinity in C3
plants. Plant Biol. 6, 269–279.oyer, C.H., Lelandais, M., Kunert, K.J., 1994. Photooxidative stress in
plants. Physiol. Plant 92, 696–717.reeden, A.L., Gamon, J.A., Field, C.B., 1991. Responses of photosynthe-
sis and carbohydrate-partitioning to limitations in nitrogen and wateravailability in field-grown sunflower. Plant Cell Environ. 14, 963–970.
enty, B., Briantais, J.M., Baker, N.R., 1989. The relationship between thequantum yield of photosynthetic electron transport and quenching ofchlorophyll fluorescence. Biochim. Biophys. Acta 990, 87–92.
atterscheid, G., Willenbrink, J., 1991. Mikroplattenleser zur enzymatis-chen zuckerbestimung. BioTec. Analytik 4, 46–48.
aupt-Herting, S., Klug, K., Fock, H.P., 2001. A new approach to measuregross CO2 fluxes in leaves. Gross CO2 assimilation, photorespiration,and mitochondrial respiration in the light in tomato under drought stress.Plant Physiol. 126, 388–396.
odges, D., DeLong, J.M., Forney, C.F., Prange, R.K., 1999. Improving thethiobarbituric acid-reactive-substances assay for estimating lipid per-oxidation in plant tissues containing anthocyanin and other interferingcompounds. Planta 207, 604–611.
iang, M., Zhang, J., 2002. Water stress-induced abscisic acid accumula-tion triggers the increased generation of reactive-oxygen species andup-regulates the activities of antioxidant enzymes in maize leaves. J.Exp. Bot. 53, 2401–2410.
rall, J.P., Edwards, G.E., 1992. Relationship between photosystem II activ-ity and CO2 fixation in leaves. Physiol. Plant 86, 180–187.
awlor, D.M., 2002. Limitation to photosynthesis in water-stressed leaves:stomata vs. metabolism and the role of ATP. Ann. Bot. 89, 871–885.
asojıdek, J., Trivedi, S., Halshaw, L., Alexiou, A., Hall, D.O., 1991. Thesynergistic effect of drought and light stresses in sorghum and pearlmillet. Plant Physiol. 96, 198–207.
unns, R., Weir, R., 1981. Contribution of sugars to osmotic adjustment in
elongating and expanded zones of wheat leaves during moderate waterdeficits at two light levels. Aust. J. Plant Physiol. 8, 93–105.octor, G., Veljovic-Jovanovic, S., Driscoll, S., Novitskaya, L., Foyer, C.H.,2002. Drought and oxidative load in the leaves of C3 plants: a predomi-nant role for photorespiration? Ann. Bot. 89, 841–850.
Z
erimental Botany 58 (2006) 75–84
oguchi, K., Nakajima, N., Terashima, I., 2001. Acclimation of leaf res-piratory properties in Alocasia odora following reciprocal transfers ofplants between high- and low-light environments. Plant Cell Environ.24, 831–839.
¨ gren, E., Oquist, G., 1985. Effects of drought on photosynthesis, chloro-phyll fluorescence and photoinhibition susceptibility in intact willowleaves. Planta 166, 380–388.
sorio, M.L., Breia, E., Rodrigues, A., Osorio, J., Le Roux, X., Daudet, F.,Ferreira, I., Chaves, M.M., 2006. Limitations to carbon assimilation bymild drought in nectarine trees growing under field conditions. Environ.Exp. Bot. 55, 235–247.
ankovic, D., Sakac, Z., Kevresan, S., Plesnicar, M., 1999. Acclimation tolong-term water deficit in the leaves of two sunflower hybrids: photo-synthesis, electron transport and carbon metabolism. J. Exp. Bot. 50,127–138.
aul, M.J., Foyer, C.H., 2001. Sink regulation of photosynthesis. J. Exp. Bot.52, 1383–1400.
elleschi, S., Rocher, J.P., Prioul, J.L., 1997. Effect of water restriction oncarbohydrate metabolism and photosynthesis in mature maize leaves.Plant Cell Environ. 20, 493–503.
uick, W.P., Chaves, M.M., Wendler, R., David, M., Rodrigues, M.L., Pas-sarinho, J.A., Pereira, J.S., Adcock, M.D., Leegood, R.C., Stitt, M.,1992. The effect of water stress on photosynthetic carbon metabolism infour species grown under field conditions. Plant Cell Environ. 15, 25–35.
ofo, A., Dichio, B., Xiloyannis, C., Masia, A., 2004. Effects of differ-ent irradiance levels on some antioxidant enzymes and on malondi-aldehyde content during rewatering in olive tree. Plant Sci. 166, 293–302.
ouza, R.P., Machado, E.C., Silva, J.A.B., Lagoa, A.M.M.A., Silveira,J.A.G., 2004. Photosynthetic gas exchange, chlorophyll fluorescence andsome associated metabolic changes in cowpea (Vigna unguiculata) dur-ing water stress and recovery. Environ. Exp. Bot. 51, 45–56.
titt, M., MaC Lilley, R., Gerhardt, R., Heldt, H.W., 1989. Determination ofmetabolite levels in specific cells and subcellular compartments of plantleaves. Meth. Enzymol. 174, 518–636.
alladares, F., Pearcy, R.W., 2002. Drought can be more critical in the shadethan in the sun: a field study of carbon gain and photo-inhibition in aCalifornian shrub during a dry El Nino year. Plant Cell Environ. 25,749–759.
eneklaas, E., den Ouden, F., 2005. Dynamics of non-structural carbohy-drates in two Ficus species after transfer to deep shade. Environ. Exp.Bot. 54, 148–154.
ingler, A., Quick, W.P., Bungard, R.A., Bailey, K.J., Lea, P.J., Leegood,R.C., 1999. The role of photorespiration during drought stress: an anal-
ysis utilizing barley mutants with reduced activities of photorespiratoryenzymes. Plant Cell Environ. 22, 361–373.renner, R., Stitt, M., 1991. Comparison of the effect of rapidly and graduallydeveloping water-stress on carbohydrate metabolism in spinach leaves.Plant Cell Environ. 14, 939–946.




















