
Influence of R and S enantiomers of 1-octen-3-ol on gene ...
15
Vol.:(0123456789) 1 3 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991 https://doi.org/10.1007/s10295-019-02168-4 GENETICS AND MOLECULAR BIOLOGY OF INDUSTRIAL ORGANISMS - ORIGINAL PAPER Influence of R and S enantiomers of 1‑octen‑3‑ol on gene expression of Penicillium chrysogenum Guohua Yin 1,2 · Yuliang Zhang 1 · Maojie Fu 1 · Sui Sheng T. Hua 3 · Qixing Huang 1 · Kayla K. Pennerman 2 · Guangxi Wu 4 · Wayne M. Jurick II 5 · Samantha Lee 2 · Lijing Bu 6 · Hui Zhao 1 · Joan W. Bennett 2 Received: 30 April 2018 / Accepted: 23 March 2019 / Published online: 28 March 2019 © Society for Industrial Microbiology and Biotechnology 2019 Abstract Inhibition of spore germination offers an attractive and effective target for controlling fungal species involved in food spoilage. Mushroom alcohol (1-octen-3-ol) functions as a natural self-inhibitor of spore germination for many fungi and, therefore, provides a useful tool for probing the molecular events controlling the early stages of fungal growth. In Penicillium spp., the R and S enantiomers of 1-octen-3-ol delayed spore germination and sporulation in four species of Penicillium involved in soils of fruit and grains, but to different degrees. Because of its well-annotated genome, we used Penicillium chrysogenum to perform a comprehensive comparative transcriptomic analysis of cultures treated with the two enantiomers. Altogether, about 80% of the high-quality reads could be mapped to 11,396 genes in the reference genome. The top three active pathways were metabolic (978 transcripts), biosynthesis of secondary metabolites (420 transcripts), and microbial metabolism in diverse environments (318 transcripts). When compared to the control, treatment with (R)-(-)-1-octen-3-ol affected the transcription levels of 91 genes, while (S)-(+)-1-octen-3-ol affected only 41 genes. Most of the affected transcripts were annotated and predicted to be involved in transport, establishment of localization, and transmembrane transport. Alternative splicing and SNPs’ analyses indicated that, compared to the control, the R enantiomer had greater effects on the gene expression pattern of Penicillium chrysogenum than the S enantiomer. A qRT-PCR analysis of 28 randomly selected differentially expressed genes confirmed the transcriptome data. The transcriptomic data have been deposited in NCBI SRA under the accession number SRX1065226. Keywords Volatile organic compounds (VOCs) · (R)-(-)-1-octen-3-ol · (S)-(+)-1-octen-3-ol · Transcriptome analysis · Fungal storage contaminants · Penicillium chrysogenum Introduction Postharvest diseases caused by filamentous fungi are a globally significant problem [37, 48]. Blue mold rots caused by Penicillium expansum and other Penicillium spp. are the most common and economically important postharvest fruit rot diseases worldwide [ 39]. Annual losses due to blue mold can result in millions of dollars in the USA alone [ 40]. Penicillium spp. also produce mycotoxins such as citrinin and patulin that pose a food safety issue [2], especially for children when they are pro- cessed into products such as apple juice and applesauce [18, 48]. The interaction of fungal postharvest pathogens with stored fruits begins with spore germination and sub- sequent penetration of host tissues via wounds caused dur- ing harvest and shipping. From a food quality perspective, Guohua Yin and Yuliang Zhang have contributed equally to this work. Electronic supplementary material The online version of this article (https://doi.org/10.1007/s10295-019-02168-4) contains supplementary material, which is available to authorized users. * Guohua Yin [email protected] * Hui Zhao [email protected] Extended author information available on the last page of the article Downloaded from https://academic.oup.com/jimb/article/46/7/977/5996811 by guest on 31 May 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Influence of R and S enantiomers of 1-octen-3-ol on gene ...

Vol.:(0123456789)1 3
Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991 https://doi.org/10.1007/s10295-019-02168-4
GENETICS AND MOLECULAR BIOLOGY OF INDUSTRIAL ORGANISMS - ORIGINAL PAPER
Influence of R and S enantiomers of 1‑octen‑3‑ol on gene expression of Penicillium chrysogenum
Guohua Yin1,2 · Yuliang Zhang1 · Maojie Fu1 · Sui Sheng T. Hua3 · Qixing Huang1 · Kayla K. Pennerman2 · Guangxi Wu4 · Wayne M. Jurick II5 · Samantha Lee2 · Lijing Bu6 · Hui Zhao1 · Joan W. Bennett2
Received: 30 April 2018 / Accepted: 23 March 2019 / Published online: 28 March 2019 © Society for Industrial Microbiology and Biotechnology 2019
AbstractInhibition of spore germination offers an attractive and effective target for controlling fungal species involved in food spoilage. Mushroom alcohol (1-octen-3-ol) functions as a natural self-inhibitor of spore germination for many fungi and, therefore, provides a useful tool for probing the molecular events controlling the early stages of fungal growth. In Penicillium spp., the R and S enantiomers of 1-octen-3-ol delayed spore germination and sporulation in four species of Penicillium involved in soils of fruit and grains, but to different degrees. Because of its well-annotated genome, we used Penicillium chrysogenum to perform a comprehensive comparative transcriptomic analysis of cultures treated with the two enantiomers. Altogether, about 80% of the high-quality reads could be mapped to 11,396 genes in the reference genome. The top three active pathways were metabolic (978 transcripts), biosynthesis of secondary metabolites (420 transcripts), and microbial metabolism in diverse environments (318 transcripts). When compared to the control, treatment with (R)-(-)-1-octen-3-ol affected the transcription levels of 91 genes, while (S)-(+)-1-octen-3-ol affected only 41 genes. Most of the affected transcripts were annotated and predicted to be involved in transport, establishment of localization, and transmembrane transport. Alternative splicing and SNPs’ analyses indicated that, compared to the control, the R enantiomer had greater effects on the gene expression pattern of Penicillium chrysogenum than the S enantiomer. A qRT-PCR analysis of 28 randomly selected differentially expressed genes confirmed the transcriptome data. The transcriptomic data have been deposited in NCBI SRA under the accession number SRX1065226.
Keywords Volatile organic compounds (VOCs) · (R)-(-)-1-octen-3-ol · (S)-(+)-1-octen-3-ol · Transcriptome analysis · Fungal storage contaminants · Penicillium chrysogenum
Introduction
Postharvest diseases caused by filamentous fungi are a globally significant problem [37, 48]. Blue mold rots caused by Penicillium expansum and other Penicillium spp. are the most common and economically important postharvest fruit rot diseases worldwide [39]. Annual losses due to blue mold can result in millions of dollars in the USA alone [40]. Penicillium spp. also produce mycotoxins such as citrinin and patulin that pose a food safety issue [2], especially for children when they are pro-cessed into products such as apple juice and applesauce [18, 48]. The interaction of fungal postharvest pathogens with stored fruits begins with spore germination and sub-sequent penetration of host tissues via wounds caused dur-ing harvest and shipping. From a food quality perspective,
Guohua Yin and Yuliang Zhang have contributed equally to this work.
Electronic supplementary material The online version of this article (https ://doi.org/10.1007/s1029 5-019-02168 -4) contains supplementary material, which is available to authorized users.
* Guohua Yin [email protected]
* Hui Zhao [email protected]
Extended author information available on the last page of the article
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022

978 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
germination is the crucial step, because, as soon as vis-ible hyphae are observed, a product is spoiled [16]. The morphological aspects of germination have been well described for several plant pathogens [28]; however, the molecular events controlling this process have not been elucidated. Inhibition of spore germination via fumigation with safe volatile compounds provides an attractive target to manage postharvest rots during storage. It is envisioned that new spore germination inhibitors can be combined with existing fumigants such as 1-methylcyclopropene [4] to provide control of products rotted by Penicillium molds.
Volatile organic compounds (VOCs) are a chemically diverse group of low-molecular-weight compounds that evaporate easily at room temperature (20–25 °C) and serve biologically targeted and diverse functions as semiochemi-cals, antimicrobials, and defense response chemicals [21, 24]. Several biogenic VOCs have been used as environmen-tally sound and effective agents for fumigation control of postharvest pathogens [31]. For example, (E)-2-hexenal vapors inhibit Aspergillus flavus growth in corn [13] and also provide control of P. expansum, the major causal agent of blue mold disease in pears [32]. Both 1-hexanol and (E)-2-hexanol inhibit Fusarium avenaceum and Fusarium graminearum growth on wheat-chick pea rotations [12].
The most abundant volatile emitted by fungi and the agent responsible for their stereotypical “mushroom/moldy” odor is the aliphatic alcohol commonly called “mushroom alco-hol” (1-octen-3-ol) (CAS Registry Number: 3391-86-4) [18, 48]. It is an eight-carbon compound characteristic of fungi [15, 35] and many bacteria [47], produced as a degradation product of linoleic acid [14, 45]. Chitarra et al. [8] identi-fied 1-octen-3-ol as a volatile self-inhibitor in Penicillium paneum after finding that dense suspensions of conidia showed poor germination. The addition of 4 mM 1-octen-3-ol to the media resulted in a germination frequency of 2% after 7 h, while 83% of the conidia germinated in the control [7]. Mushroom alcohol also functions as a self-inhibitor of spore germination in several other species of filamentous fungi such as Aspergillus niger and Penicillium chrysoge-num [51]. Moreover, this compound is also fungistatic in Aspergillus nidulans [19] and Lecanicillium fungicola [3].
Mushroom alcohol is generally regarded as safe (GRAS) by the Food and Drug Administration (https ://iaspu b.epa.gov/sor_inter net/regis try/subst reg/searc handr etrie ve/subst ances earch /searc h.do?detai ls=displ ayDet ails&selec tedSu bstan ceId=42737 ), which is used commercially as a flavor compound [53] and environmentally as a lure for biting insects [9, 25]. Since 1-octen-3-ol has a chiral carbon, there are two optical isomers or enantiomers: (R)-(-)-1-octen-3-ol (R) and (S)-(+)-1-octen-3-ol (S). The R enantiomer exhibits more of the typical aroma of fresh mushrooms, while the S form has a musty, grassy, or moldy aroma [10]. While both are formed by the oxidative breakdown of linoleic acid, the
R form predominates in many mushrooms where it is enzy-matically produced [53].
The long-term goal of our study is to develop an under-standing of the molecular mechanisms underlying the ability of mushroom alcohol to inhibit spore germination and fungal growth. Therefore, we employed RNA-seq, a well-developed approach for interpreting the functional elements of the genome and which has been used to describe the molec-ular events surrounding storage decay of fruits caused by Penicillium expansum [1] and P. digitatum [26]. As more information can be obtained from experimentally accessible organisms, we chose to focus on P. chrysogenum which has the best assembled and annotated genome in the genus, and which is closely related to P. expansum and other blue rot species [20, 36].
In this study, we first exposed seven Penicillium strains to volatile phase R-1-octen-3-ol and S-1-octen-3-ol, and investigated the ability of these compounds to inhibit spore germination and colony radial growth. Having determined that both enantiomers were effective on P. chrysogenum, we employed RNA-seq to perform a comprehensive com-parative analysis of the transcriptomes after cultures had been treated separately with either of the two enantiomers of 1-octen-3-ol. We then used bioinformatics analyses to annotate the differently expressed genes (DEGs) and puta-tively to identify the cellular pathways in which they are involved. These data will help to guide future studies on the molecular mechanisms that lead to inhibition of fungal growth. In addition, our research provides information that can be used to develop rational strategies for finding other safe and effective spore germination and growth inhibitors for the control of blue molds that cause postharvest decay of apples, pears, and other economically important fruit crops.
Materials and methods
Chemicals
Chemical standards of (R)-(-)-1-octen-3-ol (99.6%) and (S)-(+)-1-octen-3-ol (99.9%) were obtained from Bedoukian Research Inc (USA). Potato dextrose agar (PDA), glycerol, and Tween 80 were purchased from Sigma (USA). Cover-slips, “I” plates, Petri dishes, and Parafilm were purchased from VWR International (USA).
Effects of two enantiomers of 1‑octen‑3‑ol on Penicillium isolates
Six Penicillium isolates (P. expansum G10, P. expansum R19, P. expansum R21, P. solitum SA, P. solitum NJ1, and P. paneum G9) were obtained from the Food Quality Laboratory, Beltsville Agricultural Research Center (USA).
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022

979Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
Isolate P. chrysogenum P001 was maintained by our lab at Rutgers University [51]. Exposure of germinating conidia to 1-octen-3-ol enantiomers at 0.4 mM/L was performed according to the method reported by Yin et al. [51] with slight modifications. Three separate cultures of each Penicil-lium isolate were cultivated on PDA in Petri dishes at 26 °C. The conidia were harvested 3 days later in 10 mL of 10% glycerol and 1% Tween 80, and pooled and stored in ster-ile 50 mL tubes at 4 °C (maintained for less than 1 week). The concentration of viable spores was determined by serial dilution (10−4–10−7) with sterile water, plated on PDA, and colonies were counted 3 days after inoculation. One half of an “I” plate with PDA was inoculated with enough viable spores to obtain 10–30 colonies. On the other side of the division, an aliquot of R or S 1-octen-3-ol was placed on a sterile glass coverslip and to yield 0.4 mM/L. Sterilized water was used as the control. The plates were wrapped with two layers of Parafilm and placed in 4 L glass containers. Number of colonies, colony diameter, and conidiation were noted 3 days after incubation at 26 °C. The experiment was done twice, with three replicates each time (n = 6). Student’s t test was used to determine statistical significance of the differences among the treatment groups.
Penicillium chrysogenum preparation and transcriptome sequencing
Penicillium chrysogenum was cultured on PDA at 25 °C for 3 days and then transferred to 250 mL Erlenmeyer flasks containing 100 mL liquid potato dextrose medium with 2 mM/L (R)-(-)-1-octen-3-ol, (S)-(+)-1-octen-3-ol or an equal volume of sterilized water as the control group (Fig. S1). The cultures were also supplemented with 100 ng/L ampicillin, 50 ng/L kanamycin, and 50 ng/L streptomycin to prevent bacterial contamination. Cultures were grown for 120 h at 25 °C and 200 rpm. After cultivation, total RNA was extracted using TRIzol Reagent (Life Technologies) according to the manufacturer’s instruction. To remove any traces of genomic DNA from RNA extractions, total RNA was treated with RNase-free DNase (Promega) as instructed by the manufacturer. Total RNA integrity and quantity were evaluated using the 2100 Bioanalyzer RNA Nano 6000 Kit (Agilent Technologies) and the Qubit RNA Assay Kit (Life Technologies). RNA samples had integrity values between 7.6 and 9.0. Libraries were prepared for sequencing accord-ing to the Illumina TruSeq protocols using the Illumina TruSeq RNA Sample Prep Kit. Isolated mRNA was sheared with ions to about 200 nt fragments and was reverse-tran-scribed into cDNA. We then used the TruSeq PE Cluster Kit v3 to perform end repair, added an ‘A’-base to the blunt ends, and ligated the cDNA to paired-end adaptors. The cDNA samples were amplified through 15 cycles of PCR. The amplified products were subjected to electrophoresis
on a 2% Certified Low-Range Ultra-Agarose gel (Bio-Rad) and sizes suitable for Illumina sequencing were purified. These purified products were quantified using PicoGreen (Life Technologies) and a TBS-380 Mini-Fluorometer (Pro-mega) before loading onto an Illumina cBot system for clus-ter generation by bridge PCR amplification. Sequencing was performed on an Illumina HiSeq 2500 sequencing platform at Major Biosystem (China). Length of sequencing reads was 2 × 101 bp.
Transcriptome assembly and bioinformatics analysis
We trimmed all the raw reads according to published meth-ods [50]. After quality control, we mapped all the clean reads to the P. chrysogenum reference genome (http://genom e.jgi.doe.gov/Pench 1/Pench 1.home.html) [46] using TopHat (http://topha t.cbcb.umd.edu/) [43, 44]. Duplicate reads and gene coverage degree, VennDiagram, Gene Ontology, KEGG database, Cluster of Orthologous Groups of proteins (COG), SNPs, etc. were all analyzed as described in our previous paper [50].
The qRT‑PCR evaluation of DEGs
The qRT-PCR primers were designed using Primer 5 to evaluate the DEGs discovered by RNA-seq as described in the previous work [50]. The tubulin and actin genes were used as endogenous controls. All the gene-specific primers are listed in Supplementary Table S2.
Results
Effects of 1‑octen‑3‑ol enantiomers on Penicillium species
Seven strains of Penicillium (Penicillium chrysogenum P001, Penicillium expansum G10, Penicillium expansum R19, Pen-icillium expansum R21, Penicillium solitum SA, Penicillium solitum NJ1, and Penicillium paneum G9) known to cause spoilage of fruits or grains were used in our study. All seven strains were exposed to the enantiomers of 1-octen-3-ol to determine the comparative effects on spore germination and growth. The R form of 1-octen-3-ol significantly reduced spore germination in five out of seven isolates, while the S enantiomer resulted in significant reduction only for P. chrysogenum 001 and P. solitum SA (Fig. 1a). All strains, except for P. expansum G10 and P. solitum NJ1, yielded a significant reduction in the number of colony forming units (CFUs) in response to the volatiles. Colony diameters were significantly smaller in the presence of either form of 1-octen-3-ol, for all seven isolates (Fig. 1b). Furthermore, there were statistically significant differences (p < 0.05)
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022

980 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
between the R and S treatments for the number of CFUs for P. expansum R19, P. expansum R21, and P. solitum SA; and for the colony diameter for P. solitum SA (not indicated in figure). Both enantiomers delayed sporulation of all isolates for at least 2 days compared to the control (data not shown).
Sequencing read quality and mapping to reference genome
We selected P. chrysogenum for RNA-seq analysis, because it is the best studied and annotated genome in the genus [36, 46]. The RNA-seq data counts and qualities of P. chrysoge-num P001 cultures exposed to R or S 1-octen-3-ol are sum-marized in Table 1. After trimming the raw reads by SeqPrep and Sickle software using default parameters, over 99% of the clean reads had a quality ≥ Q20 (error rate < 1%); over
Fig. 1 Effects of R and S enantiomers of 1-octen-3-ol on the number of CFUs (a) and colony diameter (b) of seven Penicillium isolates. Comparisons of colony forming units (a) and colony diameters (b) among the control (black), R-1-octen-3-ol (yellow), and S-1-octen-3-ol (red) treatments are shown. Error bars represent one standard deviation above the mean. Asterisks (*) indicate a significant differ-
ence between the R or S treatment compared to the control (p < 0.05). There were statistically significant differences (p < 0.05) between the R and S treatments for the number of colony forming units for strains P. expansum R19, P. expansum R21, and P. solitum SA; and for the colony diameter of P. solitum SA (not indicated in figure)
Table 1 Sequencing read counts and quality
Control: added equal volume of sterile water; R: (R)-(-)-1-octen-3-ol; S: (S)-(+)-1-octen-3-ol; the length of sequencing is 2 × 101 bp, e.g. 101 bp/read and paired-end sequencing
Treatments Raw reads (No.) Raw bases (bp) ≥ Q20 (%) ≥ Q30 (%) Trimmed reads (No.) Trimmed bases (bp) ≥ Q20 (%) ≥ Q30 (%)
Control 36,480,974 3,684,578,374 93.57 93.57 33,772,290 3,304,053,748 99.15 94.72R 40,653,692 4,106,022,892 94.27 94.27 38,118,340 3,726,250,033 99.16 94.74S 30,375,266 3,067,901,866 93.81 93.81 28,251,370 2,761,257,013 99.21 95.03
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022
981Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
94% were ≥ Q30 (error rate < 0.1%). About 80% of clean reads could be mapped to the reference genome (Table 2).
Gene and pathway functional annotation of mapped transcripts
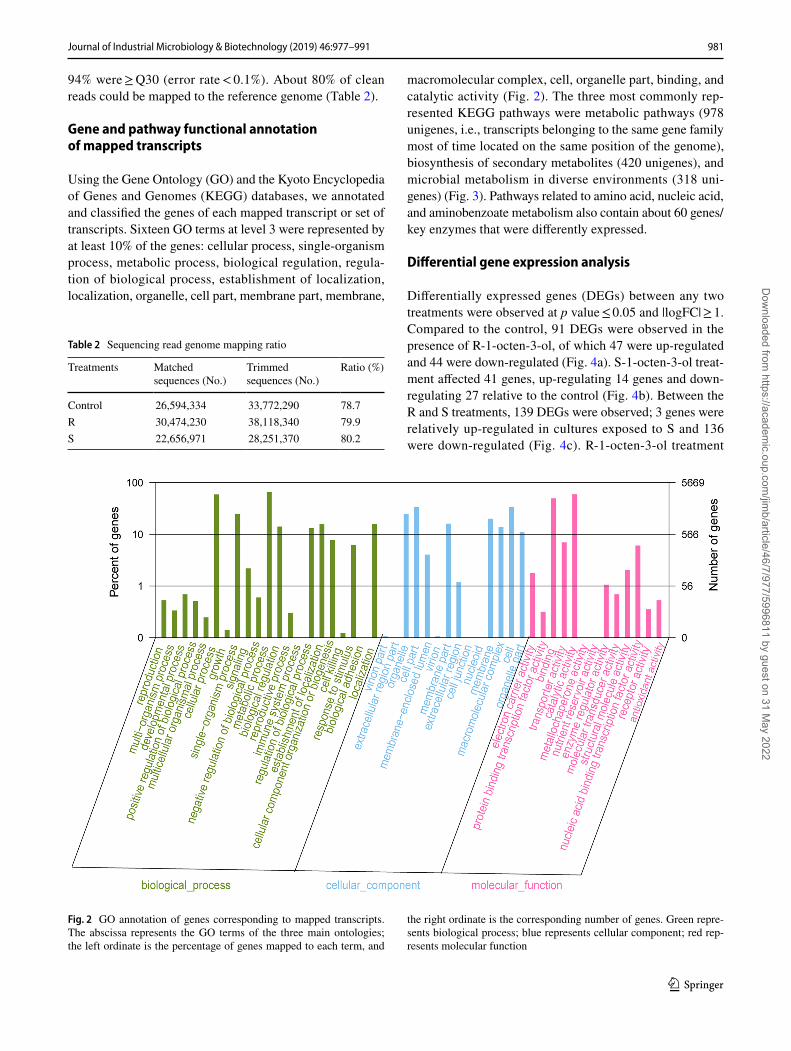
Using the Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) databases, we annotated and classified the genes of each mapped transcript or set of transcripts. Sixteen GO terms at level 3 were represented by at least 10% of the genes: cellular process, single-organism process, metabolic process, biological regulation, regula-tion of biological process, establishment of localization, localization, organelle, cell part, membrane part, membrane,
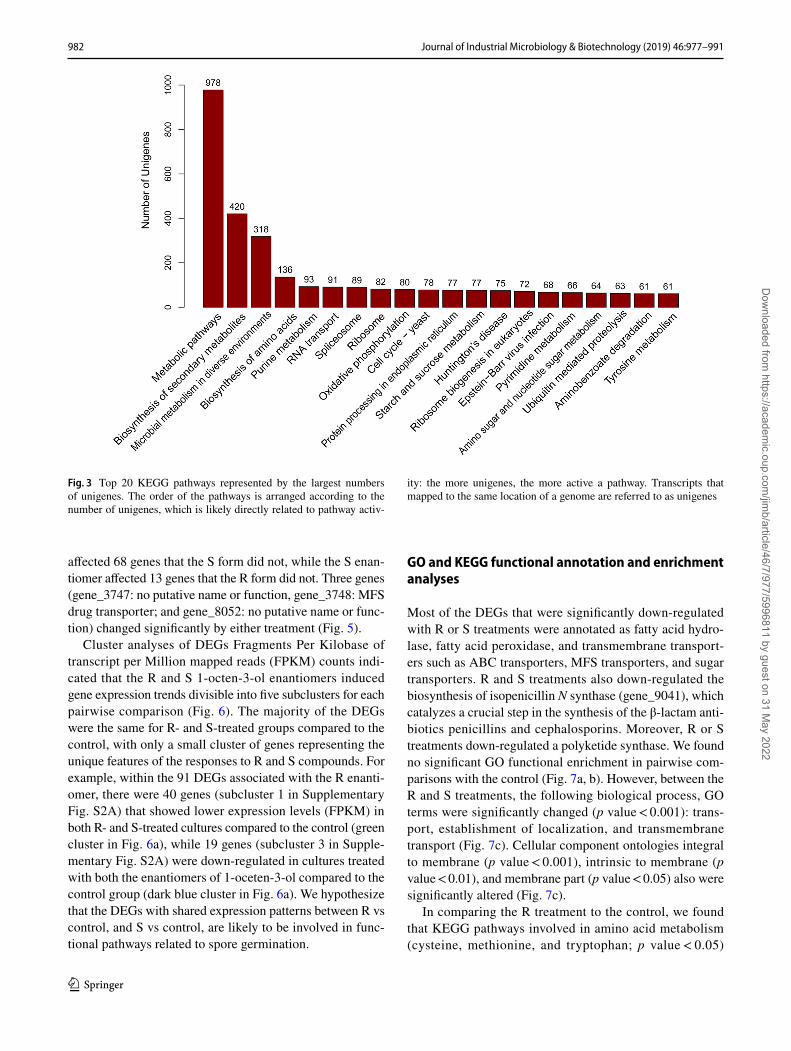
macromolecular complex, cell, organelle part, binding, and catalytic activity (Fig. 2). The three most commonly rep-resented KEGG pathways were metabolic pathways (978 unigenes, i.e., transcripts belonging to the same gene family most of time located on the same position of the genome), biosynthesis of secondary metabolites (420 unigenes), and microbial metabolism in diverse environments (318 uni-genes) (Fig. 3). Pathways related to amino acid, nucleic acid, and aminobenzoate metabolism also contain about 60 genes/key enzymes that were differently expressed.
Differential gene expression analysis
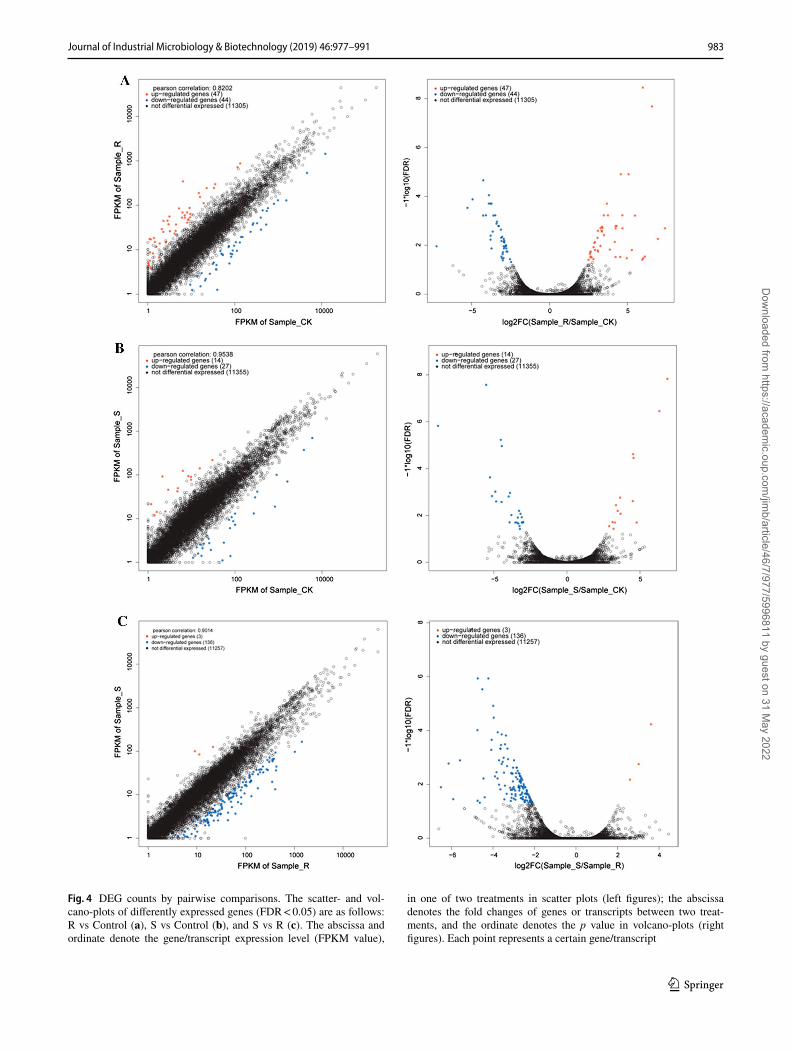
Differentially expressed genes (DEGs) between any two treatments were observed at p value ≤ 0.05 and |logFC| ≥ 1. Compared to the control, 91 DEGs were observed in the presence of R-1-octen-3-ol, of which 47 were up-regulated and 44 were down-regulated (Fig. 4a). S-1-octen-3-ol treat-ment affected 41 genes, up-regulating 14 genes and down-regulating 27 relative to the control (Fig. 4b). Between the R and S treatments, 139 DEGs were observed; 3 genes were relatively up-regulated in cultures exposed to S and 136 were down-regulated (Fig. 4c). R-1-octen-3-ol treatment
Table 2 Sequencing read genome mapping ratio
Treatments Matched sequences (No.)
Trimmed sequences (No.)
Ratio (%)
Control 26,594,334 33,772,290 78.7R 30,474,230 38,118,340 79.9S 22,656,971 28,251,370 80.2
Fig. 2 GO annotation of genes corresponding to mapped transcripts. The abscissa represents the GO terms of the three main ontologies; the left ordinate is the percentage of genes mapped to each term, and
the right ordinate is the corresponding number of genes. Green repre-sents biological process; blue represents cellular component; red rep-resents molecular function
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022
982 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
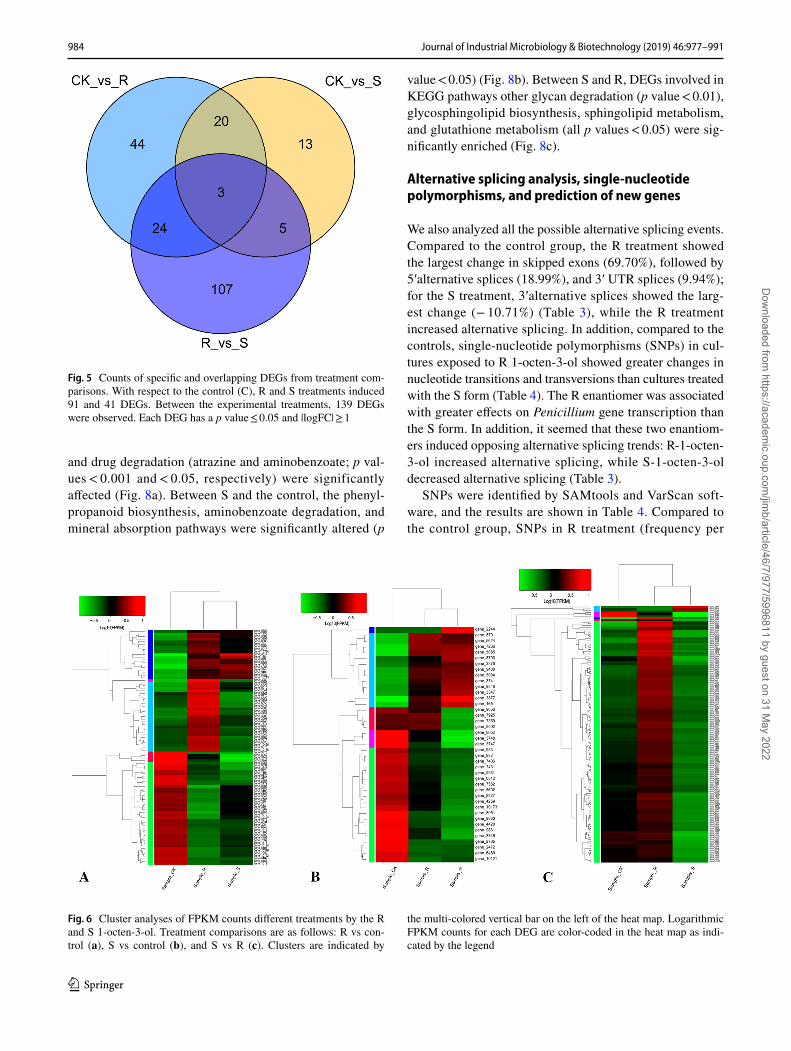
affected 68 genes that the S form did not, while the S enan-tiomer affected 13 genes that the R form did not. Three genes (gene_3747: no putative name or function, gene_3748: MFS drug transporter; and gene_8052: no putative name or func-tion) changed significantly by either treatment (Fig. 5).
Cluster analyses of DEGs Fragments Per Kilobase of transcript per Million mapped reads (FPKM) counts indi-cated that the R and S 1-octen-3-ol enantiomers induced gene expression trends divisible into five subclusters for each pairwise comparison (Fig. 6). The majority of the DEGs were the same for R- and S-treated groups compared to the control, with only a small cluster of genes representing the unique features of the responses to R and S compounds. For example, within the 91 DEGs associated with the R enanti-omer, there were 40 genes (subcluster 1 in Supplementary Fig. S2A) that showed lower expression levels (FPKM) in both R- and S-treated cultures compared to the control (green cluster in Fig. 6a), while 19 genes (subcluster 3 in Supple-mentary Fig. S2A) were down-regulated in cultures treated with both the enantiomers of 1-oceten-3-ol compared to the control group (dark blue cluster in Fig. 6a). We hypothesize that the DEGs with shared expression patterns between R vs control, and S vs control, are likely to be involved in func-tional pathways related to spore germination.
GO and KEGG functional annotation and enrichment analyses
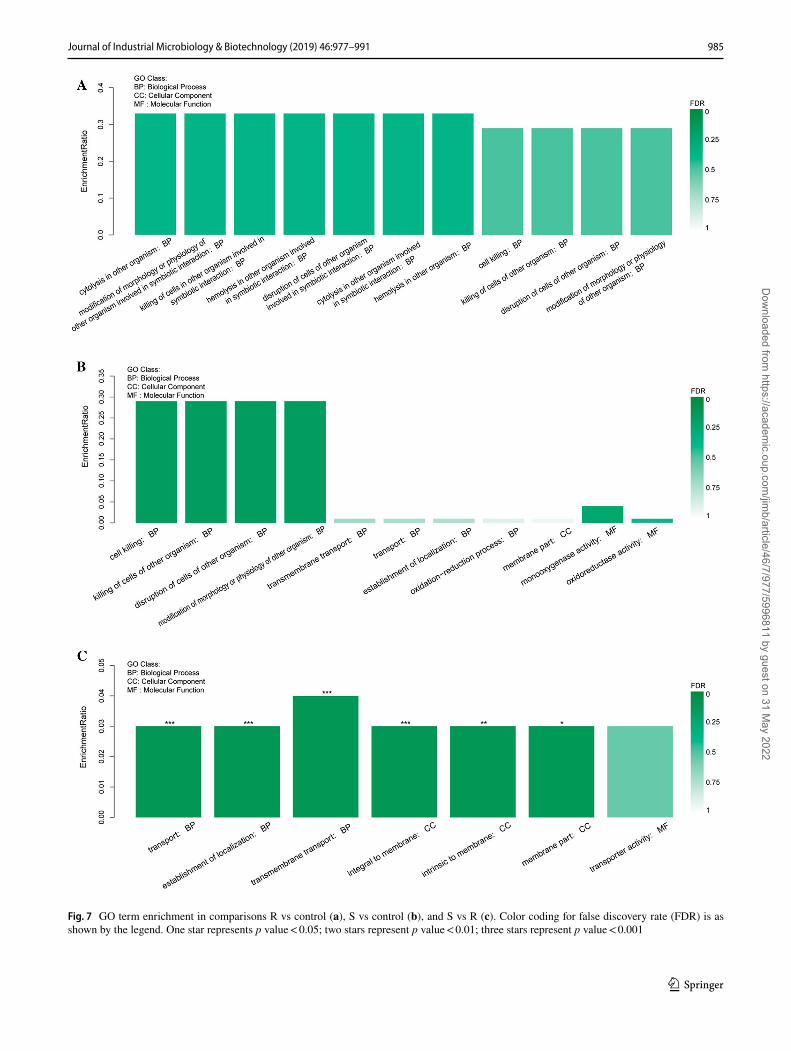
Most of the DEGs that were significantly down-regulated with R or S treatments were annotated as fatty acid hydro-lase, fatty acid peroxidase, and transmembrane transport-ers such as ABC transporters, MFS transporters, and sugar transporters. R and S treatments also down-regulated the biosynthesis of isopenicillin N synthase (gene_9041), which catalyzes a crucial step in the synthesis of the β-lactam anti-biotics penicillins and cephalosporins. Moreover, R or S treatments down-regulated a polyketide synthase. We found no significant GO functional enrichment in pairwise com-parisons with the control (Fig. 7a, b). However, between the R and S treatments, the following biological process, GO terms were significantly changed (p value < 0.001): trans-port, establishment of localization, and transmembrane transport (Fig. 7c). Cellular component ontologies integral to membrane (p value < 0.001), intrinsic to membrane (p value < 0.01), and membrane part (p value < 0.05) also were significantly altered (Fig. 7c).
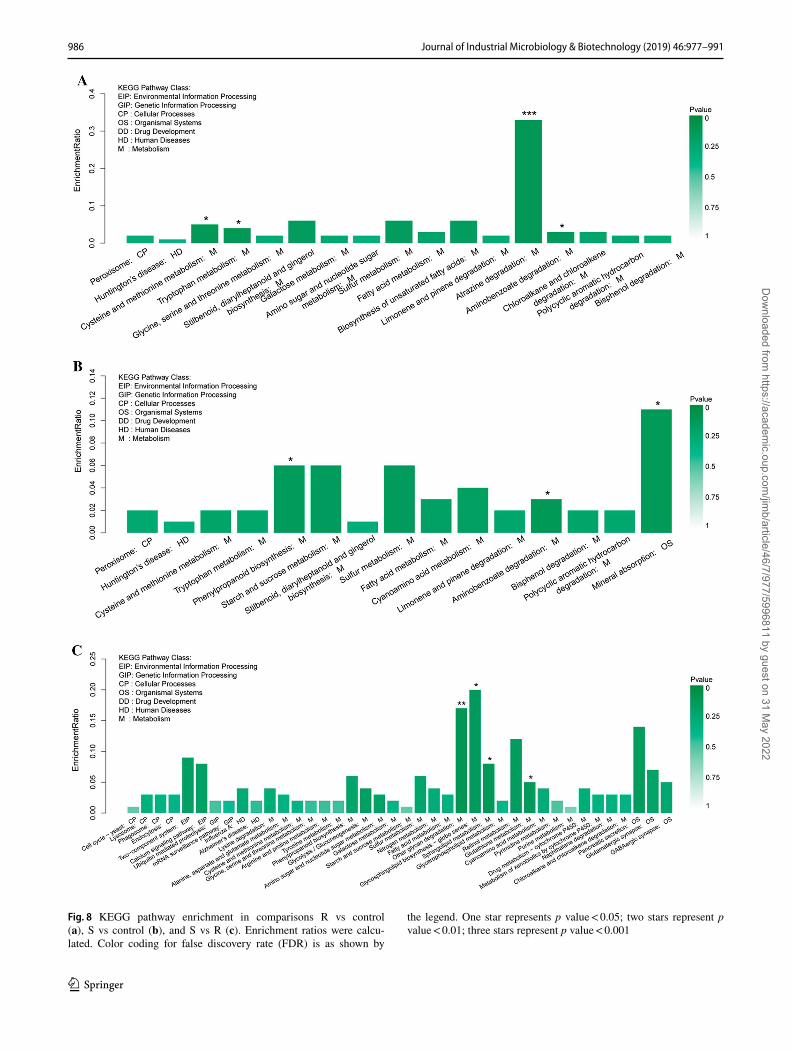
In comparing the R treatment to the control, we found that KEGG pathways involved in amino acid metabolism (cysteine, methionine, and tryptophan; p value < 0.05)
Fig. 3 Top 20 KEGG pathways represented by the largest numbers of unigenes. The order of the pathways is arranged according to the number of unigenes, which is likely directly related to pathway activ-
ity: the more unigenes, the more active a pathway. Transcripts that mapped to the same location of a genome are referred to as unigenes
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022
983Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
Fig. 4 DEG counts by pairwise comparisons. The scatter- and vol-cano-plots of differently expressed genes (FDR < 0.05) are as follows: R vs Control (a), S vs Control (b), and S vs R (c). The abscissa and ordinate denote the gene/transcript expression level (FPKM value),
in one of two treatments in scatter plots (left figures); the abscissa denotes the fold changes of genes or transcripts between two treat-ments, and the ordinate denotes the p value in volcano-plots (right figures). Each point represents a certain gene/transcript
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022
984 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
and drug degradation (atrazine and aminobenzoate; p val-ues < 0.001 and < 0.05, respectively) were significantly affected (Fig. 8a). Between S and the control, the phenyl-propanoid biosynthesis, aminobenzoate degradation, and mineral absorption pathways were significantly altered (p
value < 0.05) (Fig. 8b). Between S and R, DEGs involved in KEGG pathways other glycan degradation (p value < 0.01), glycosphingolipid biosynthesis, sphingolipid metabolism, and glutathione metabolism (all p values < 0.05) were sig-nificantly enriched (Fig. 8c).
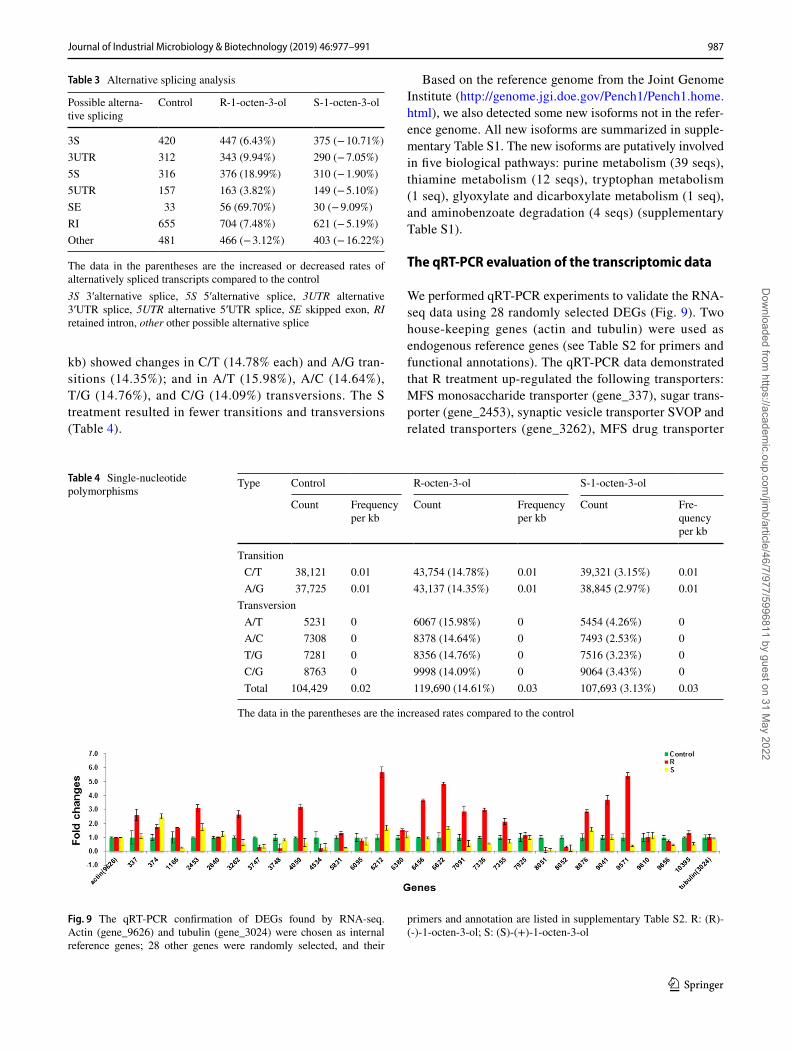
Alternative splicing analysis, single‑nucleotide polymorphisms, and prediction of new genes
We also analyzed all the possible alternative splicing events. Compared to the control group, the R treatment showed the largest change in skipped exons (69.70%), followed by 5′alternative splices (18.99%), and 3′ UTR splices (9.94%); for the S treatment, 3′alternative splices showed the larg-est change (− 10.71%) (Table 3), while the R treatment increased alternative splicing. In addition, compared to the controls, single-nucleotide polymorphisms (SNPs) in cul-tures exposed to R 1-octen-3-ol showed greater changes in nucleotide transitions and transversions than cultures treated with the S form (Table 4). The R enantiomer was associated with greater effects on Penicillium gene transcription than the S form. In addition, it seemed that these two enantiom-ers induced opposing alternative splicing trends: R-1-octen-3-ol increased alternative splicing, while S-1-octen-3-ol decreased alternative splicing (Table 3).
SNPs were identified by SAMtools and VarScan soft-ware, and the results are shown in Table 4. Compared to the control group, SNPs in R treatment (frequency per
Fig. 5 Counts of specific and overlapping DEGs from treatment com-parisons. With respect to the control (C), R and S treatments induced 91 and 41 DEGs. Between the experimental treatments, 139 DEGs were observed. Each DEG has a p value ≤ 0.05 and |logFC| ≥ 1
Fig. 6 Cluster analyses of FPKM counts different treatments by the R and S 1-octen-3-ol. Treatment comparisons are as follows: R vs con-trol (a), S vs control (b), and S vs R (c). Clusters are indicated by
the multi-colored vertical bar on the left of the heat map. Logarithmic FPKM counts for each DEG are color-coded in the heat map as indi-cated by the legend
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022
985Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
Fig. 7 GO term enrichment in comparisons R vs control (a), S vs control (b), and S vs R (c). Color coding for false discovery rate (FDR) is as shown by the legend. One star represents p value < 0.05; two stars represent p value < 0.01; three stars represent p value < 0.001
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022
986 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
Fig. 8 KEGG pathway enrichment in comparisons R vs control (a), S vs control (b), and S vs R (c). Enrichment ratios were calcu-lated. Color coding for false discovery rate (FDR) is as shown by
the legend. One star represents p value < 0.05; two stars represent p value < 0.01; three stars represent p value < 0.001
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022
987Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
kb) showed changes in C/T (14.78% each) and A/G tran-sitions (14.35%); and in A/T (15.98%), A/C (14.64%), T/G (14.76%), and C/G (14.09%) transversions. The S treatment resulted in fewer transitions and transversions (Table 4).
Based on the reference genome from the Joint Genome Institute (http://genom e.jgi.doe.gov/Pench 1/Pench 1.home.html), we also detected some new isoforms not in the refer-ence genome. All new isoforms are summarized in supple-mentary Table S1. The new isoforms are putatively involved in five biological pathways: purine metabolism (39 seqs), thiamine metabolism (12 seqs), tryptophan metabolism (1 seq), glyoxylate and dicarboxylate metabolism (1 seq), and aminobenzoate degradation (4 seqs) (supplementary Table S1).
The qRT‑PCR evaluation of the transcriptomic data
We performed qRT-PCR experiments to validate the RNA-seq data using 28 randomly selected DEGs (Fig. 9). Two house-keeping genes (actin and tubulin) were used as endogenous reference genes (see Table S2 for primers and functional annotations). The qRT-PCR data demonstrated that R treatment up-regulated the following transporters: MFS monosaccharide transporter (gene_337), sugar trans-porter (gene_2453), synaptic vesicle transporter SVOP and related transporters (gene_3262), MFS drug transporter
Table 3 Alternative splicing analysis
The data in the parentheses are the increased or decreased rates of alternatively spliced transcripts compared to the control3S 3′alternative splice, 5S 5′alternative splice, 3UTR alternative 3′UTR splice, 5UTR alternative 5′UTR splice, SE skipped exon, RI retained intron, other other possible alternative splice
Possible alterna-tive splicing
Control R-1-octen-3-ol S-1-octen-3-ol
3S 420 447 (6.43%) 375 (− 10.71%)3UTR 312 343 (9.94%) 290 (− 7.05%)5S 316 376 (18.99%) 310 (− 1.90%)5UTR 157 163 (3.82%) 149 (− 5.10%)SE 33 56 (69.70%) 30 (− 9.09%)RI 655 704 (7.48%) 621 (− 5.19%)Other 481 466 (− 3.12%) 403 (− 16.22%)
Table 4 Single-nucleotide polymorphisms
The data in the parentheses are the increased rates compared to the control
Type Control R-octen-3-ol S-1-octen-3-ol
Count Frequency per kb
Count Frequency per kb
Count Fre-quency per kb
Transition C/T 38,121 0.01 43,754 (14.78%) 0.01 39,321 (3.15%) 0.01 A/G 37,725 0.01 43,137 (14.35%) 0.01 38,845 (2.97%) 0.01
Transversion A/T 5231 0 6067 (15.98%) 0 5454 (4.26%) 0 A/C 7308 0 8378 (14.64%) 0 7493 (2.53%) 0 T/G 7281 0 8356 (14.76%) 0 7516 (3.23%) 0 C/G 8763 0 9998 (14.09%) 0 9064 (3.43%) 0 Total 104,429 0.02 119,690 (14.61%) 0.03 107,693 (3.13%) 0.03
Fig. 9 The qRT-PCR confirmation of DEGs found by RNA-seq. Actin (gene_9626) and tubulin (gene_3024) were chosen as internal reference genes; 28 other genes were randomly selected, and their
primers and annotation are listed in supplementary Table S2. R: (R)-(-)-1-octen-3-ol; S: (S)-(+)-1-octen-3-ol
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022

988 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
(gene_4059), MFS multidrug transporter (gene_6212 and gene_7355), MFS multidrug resistance transporter (gene_9041), and ABC multidrug transporter (gene_10395). Phthiocerol synthesis polyketide synthase type I PpsC (gene_1166), linoleate 10R-lipoxygenase (gene_6380), transcription factor IIIB 60 kDa subunit (gene_6456), short chain dehydrogenase (gene_6622), transcriptional regulator CRZ1 (gene_7091), azole resistance protein 1 (gene_7336), isopenicillin N synthase, and related dioxy-genases (gene_8876) and gene_9571 (no putative name or function) were also up-regulated by R treatment. S treat-ment down-regulated expression of vacuolar protein sorting-associated protein 27 (gene_5831) and gene_9656 (no puta-tive name or function). The gene for fatty acid hydroxylase (gene_374) was up-regulated by both R and S enantiom-ers. Gene_3747 (no putative name or function), MFS drug transporter (gene_3748), allantoate permease (gene_4534), superoxide dismutase (gene_8051), and gene_8052 (no puta-tive name or function) were all significantly down-regulated by treatment with either enantiomer. Polyphenol oxidase 4 (gene_2640), polyamine transporter 2 (gene_7925), and high-affinity glucose transporter ght2 (gene_9610) did not show significant changes with R and S treatments. Overall, the qRT-PCR data agreed with 20 out of 28 of the tested RNA-Seq results (supplementary Table S3) [1].
Discussion and conclusion
In our search to find effective methods to control postharvest fungal plant pathogens, we focused on deciphering the earli-est stage of decay, i.e., germination of a pathogen’s spores. If an environmentally safe, volatile compound can be for-mulated to inhibit this step, it would provide an effective strategy to control postharvest decay of fruit and vegetable crops, and simultaneously block the potential for mycotox-ins biosynthesis. We selected 1-octen-3-ol (“mushroom alcohol”), for detailed study, because it is known to be a natural self-inhibitor of fungal spore germination. Exposure to exogenous 1-octen-3-ol affects growth, conidiation, and germination in many fungi [3, 7, 30, 33, 34]. For Aspergil-lus niger and Penicillium chrysogenum, (R)-(-)-1-octen-3-ol imposed the greatest impact on fungal growth compared to (S)-(+)-1-octen-3-ol and the racemic form [51]. In other published studies, 1-octen-3-ol and 10-oxo-trans-8-decenoic acid inhibited the mycelial growth of Penicillium expansum at concentrations 1.25 mM/L or greater. It was further noted that 1-octen-3-ol had mild effects on the plasma membrane and interfered with essential metabolic processes, such as swelling and germination of the conidia, in a reversible man-ner. The authors noted that the compounds may serve as sources of natural food preservatives [34].
In this study, we observed a differential effect of R and S 1-octen-3-ol on colony morphology and conidial germina-tion on Penicillium spp. To investigate the molecular causes for these differences, we screened genes for their involve-ment in response to either enantiomer of this natural spore inhibitor. We observed that many transporters and other membrane constituents were enriched, indicating the impor-tance of membrane trafficking for the 1-octen-3-ol response and suggesting that changes in the levels of membrane trans-porters may affect the balance of nutrients and metabolites involved in spore germination and growth (Fig. 10). By con-ducting transcriptomic analysis of P. chrysogenum, a species with a well-annotated genome, we have begun the molecular dissection of the early events of fungal growth that eventu-ally lead to the spoilage of stored products. Similar results have been obtained from other transcriptomic studies using VOCs demonstrating significant changes in expression of genes involved in localization and transport [6, 52]. Further investigation will be required to determine if transmembrane proteins are involved in 1-octen-3-ol signaling and biologi-cal response via functional analysis of single gene deletion mutants.
Entomologists have shown that 1-octen-3-ol attracts bit-ing insects such as mosquitoes, although the chemical itself does not kill insects [9]. Furthermore, they have shown that the enantiomers of 1-octen-3-ol function differently in insects. For example, odorant receptors of Aedes aegypti are selectively more sensitive to the R form [5, 17, 41]. Ambly-omma variegatum, A. aegypti, and Culex quinquefasciatus show more behavioral changes with the R form [11, 27], whereas, in Cis bolete, the female beetles are more attracted to the S form [42]. Recently, an odorant receptor (AaegOR8) known to occur on the maxillary palps of mosquitoes was investigated in a heterologous system and demonstrated to be selectively sensitive to (R)-(-)-1-octen-3-ol. Lesser responses were stimulated by (S)-(+)-1-octen-3-ol and various structural analogs. Features such the chain length, position of unsaturation, functional groups, and the enantio-meric composition are crucial in eliciting responses from the peripheral system [17]. In other studies, it has been shown that 1-octen-3-ol serves as the natural ligand of bovine odor-ant-binding protein [38]. In addition, 1-octen-3-ol disrupts dopamine packaging and uptake, and causes neurodegenera-tion in both Drosophila melanogaster and human cell lines [22, 23]. Furthermore, in human embryonic stem cells, the S enantiomer is more cytotoxic than the R form [23].
1-Octen-3-ol is a degradation product of linoleic acid [10, 49]. Many fungi and plants produce linoleic acid, which is an essential fatty acid for animals, including many insects. In fungi, lipoxygenases and lyases are hypothesized to degrade linoleic acid to produce 1-octen-3-ol, but definitive experimental evidence is lacking [29]. A recent study indicates that intact Aspergillus flavus
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022

989Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
conidia produced low amounts of C8 volatiles and that repeated freeze–thaw treatments, resulting in increased permeability of the cell membrane, resulted in more pro-duction of (R)-(-)-1-octen-3-ol. Curiously, knockout or RNAi suppression of any of the five fatty acid oxygenases hypothesized to promote 1-octen-3-ol production (PpoA, PpoB, PpoC, PpoD, or lipoxygenase) did not affect the formation of the C8 volatile [29].
Data from our current investigation provide foundational information to target specific genes for functional analyses to better understand fundamental processes of volatile sign-aling in fungi. P. chrysogenum RNA transcripts accumu-lated differently due to exogenous exposure to either the R enantiomer or S enantiomer of 1-octen-3-ol. To our knowl-edge, this is the first report to characterize the molecular mechanisms underlying differential 1-octen-3-ol enantiomer responses in fungi. We hope that these data will serve as a platform for research that seeks to translate these basic find-ing into rational methods of fungal pathogen control.
In conclusion, our growth data reinforce earlier studies, demonstrating that mushroom alcohol has inhibitory effects on germination and colony growth of blue molds, and our transcriptome analyses supports the hypothesis that genes involved in membrane trafficking are important to the inhibi-tory effects. We plan to use this information to guide the development of safe, environmentally friendly biofumigants for suppression of postharvest pathogens. Because several blue mold species produce mycotoxins, one implication of our research is that fumigation by 1-octen-3-ol might
provide a safe method for lowering toxin contamination of these important food crops.
Acknowledgements This work was primarily funded by the Special Fund for Agro-scientific Research in the Public Interest of the Peo-ple’s Republic of China (Grant No. 201403075) and partially funded by the USDA-ARS Cooperative Agreement (Grant No. 2-47012). The funders played no role in experimental design, execution, data analysis, or decision to publish. Use of a company or product name by the U.S. Department of Agriculture does not imply approval or recommenda-tion of the product to the exclusion of others that may also be suitable.
References
1. Ballester AR, Marcet Houben M, Levin E, Sela N, Selma Lázaro C, Carmona L, Wisniewski M, Droby S, González Candelas L, Gabaldón T (2015) Genome, transcriptome, and functional analy-ses of Penicillium expansum provide new insights into second-ary metabolism and pathogenicity. Mol Plant Microbe Interact 28(3):232–248
2. Bennett JW, Klich M (2003) Mycotoxins. Clin Microbiol Rev 16:497–516
3. Berendsen RL, Kalkhove SI, Lugones LG, Baars JJ, Wösten HA, Bakker PA (2013) Effects of the mushroom-volatile 1-octen-3-ol on dry bubble disease. Appl Microbiol Biotechnol 97(12):5535–5543
4. Blankenship SM, Dole JM (2003) 1-Methylcyclopropene: a review. Postharvest Biol Technol 28(1):1–25
5. Bohbot JD, Dickens JC (2009) Characterization of an enanti-oselective odorant receptor in the yellow fever mosquito Aedes aegypti. PLoS One 4(9):e7032
6. Chang PK, Hua SST, Sarreal SBL, Li RW (2015) Suppression of aflatoxin biosynthesis in Aspergillus flavus by 2-phenylethanol
Fig. 10 Putative pathway of 1-octen-3-ol release and uptake. 1-octen-3-ol is biosynthesized by the oxidative breakdown of linoleic acid in fungi. Many different transporters in the membrane possibly are involved in its emission or intake
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022

990 Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
is associated with stimulated growth and decreased degradation of branched-chain amino acids. Toxins 7(10):3887–3902
7. Chitarra GS, Abee T, Rombouts FM, Dijksterhuis J (2005) 1-Octen-3-ol inhibits conidia germination of Penicillium paneum despite of mild effects on membrane permeability, res-piration, intracellular pH, and changes the protein composition. FEMS Microbiol Ecol 54(1):67–75
8. Chitarra GS, Abee T, Rombouts FM, Posthumus MA, Dijkster-huis J (2004) Germination of Penicillium paneum conidia is regulated by 1-octen-3-ol, a volatile self-inhibitor. Appl Environ Microbiol 70(5):2823–2829
9. Cilek JE, Hallmon CF, Johnson R (2011) Semi-field comparison of the BG Lure, nonanal, and 1-octen-3-ol to attract adult mos-quitoes in northwestern Florida. J Am Mosquito Control Assoc 27(4):393–397
10. Combet E, Eastwood DC, Burton KS, Henderson J (2006) Eight-carbon volatiles in mushrooms and fungi: Properties, analysis, and biosynthesis. Mycoscience 47:317–326
11. Cook JI, Majeed S, Ignell R, Pickett JA, Birkett MA, Logan JG (2011) Enantiomeric selectivity in behavioural and electro-physiological responses of Aedes aegypti and Culex quinque-fasciatus mosquitoes. Bull Entomol Res 101(5):541–550
12. Cruz AF, Hamel C, Yang C, Matsubara T, Gan Y, Singh AK, Kuwada K, Ishii T (2012) Phytochemicals to suppress Fusar-ium head blight in wheat-chickpea rotation. Phytochemistry 78:72–80
13. De Lucca AJ, Carter Wientjes CH, Boué S, Bhatnagar D (2011) Volatile trans-2-hexenal, a soybean aldehyde, inhibits Aspergil-lus flavus growth and aflatoxin production in corn. J Food Sci 76(6):M381–M386
14. Dijkstra FY, Wiken TO (1976) Studies on mushroom flavours. 1. Organoleptic significance of constituents of the cultivated mush-room, Agaricus bisporus. Zeitschrift fur Lebensmittel-Untersu-chung und-Forschung 160(3):255-62
15. Fischer G, Dott W (2003) Relevance of airborne fungi and their secondary metabolites for environmental, occupational and indoor hygiene. Arch Microbiol 179:75–82
16. Garcia D, Ramos AJ, Sanchis V, Marín S (2009) Predicting myco-toxins in foods: a review. Food Microbiol 26(8):757–769
17. Grant AJ, Dickens JC (2011) Functional characterization of the octenol receptor neuron on the maxillary palps of the yellow fever mosquito, Aedes aegypti. PLoS One 6(6):e21785
18. Harrison MA (1988) Presence and stability of patulin in apple products: a review. J Food Saf 9(3):147–153
19. Herrero Garcia E, Garzia A, Cordobés S, Espeso EA, Ugalde U (2011) 8-Carbon oxylipins inhibit germination and growth, and stimulate aerial conidiation in Aspergillus nidulans. Fungal Biol 115(4):393–400
20. Houbraken J, Samson RA (2011) Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud Mycol 70:1–51
21. Hung R, Lee S, Bennett JW (2015) Fungal volatile organic com-pounds and their role in ecosystems. Appl Microbiol Biotechnol 99(8):3395–3405
22. Inamdar AA, Hossain MM, Bernstein AI, Miller GW, Richardson JR, Bennett JW (2013) Fungal-derived semiochemical 1-octen-3-ol disrupts dopamine packaging and causes neurodegeneration. Proc Natl Acad Sci 110(48):19561–19566
23. Inamdar AA, Moore JC, Cohen RI, Bennett JW (2012) A model to evaluate the cytotoxicity of the fungal volatile organic compound 1-octen-3-ol in human embryonic stem cells. Mycopathologia 173(1):13–20
24. Korpi A, Järnberg J, Pasanen A (2009) Microbial volatile organic compounds. Crit Rev Toxicol 39:139–193
25. Luntz AM (2003) Arthropod semiochemicals: mosquitoes, midges and sealice. Biochem Soc Trans 31(1):128–133
26. Marcet Houben M, Ballester AR, de la Fuente B, Harries E, Marcos JF, González Candelas L, Gabaldón T (2012) Genome sequence of the necrotrophic fungus Penicillium digitatum, the main postharvest pathogen of citrus. BMC Genom 13(1):1
27. McMahon C, Guerin PM, Syed Z (2001) 1-octen-3-ol isolated from bont ticks attracts Amblyomma variegatum. J Chem Ecol 27(3):471–486
28. Mendgen K, Deising H (1993) Infection structures of fungal plant pathogens—a cytological and physiological evaluation. New Phy-tol 124(2):193–213
29. Miyamoto K, Murakami T, Kakumyan P, Keller NP, Matsui K (2014) Formation of 1-octen-3-ol from Aspergillus flavus conidia is accelerated after disruption of cells independently of Ppo oxy-genases, and is not a main cause of inhibition of germination. Peer J 2:e395
30. Nemcovic M, Jakubikova L, Viden I, Farkas V (2008) Induction of conidiation by endogenous volatile compounds in Trichoderma spp. FEMS Microbiol Lett 284(2):231–236
31. Neri F, Mari M, Brigati S (2006) Control of Penicillium expansum by plant volatile compounds. Plant Pathol 55(1):100–105
32. Neri F, Mari M, Menniti A, Brigati S (2006) Activity of trans-2-hexenal against Penicillium expansum in ‘Conference’ pears. J Appl Microbiol 100(6):1186–1193
33. Noble R, Dobrovin Pennington A, Hobbs PJ, Pederby J, Rodger A (2009) Volatile C8 compounds and pseudomonads influ-ence primordium formation of Agaricus bisporus. Mycologia 101(5):583–591
34. Okull DO, Beelman RB, Gourama H (2003) Antifungal activity of 10-oxo-trans-8-decenoic acid and 1-octen-3-ol against Peni-cillium expansum in potato dextrose agar medium. J Food Prot 66(8):1503–1505
35. Pasanen P, Korpi A, Kalliokoski P, Pasanen A (1997) Growth and volatile metabolite production of Aspergillus versicolor in house dust. Environ Int 162:263–272
36. Peng Q, Yuan Y, Gao M, Chen X, Liu B, Liu P, Wu Y, Wu D (2014) Genomic characteristics and comparative genomics analy-sis of Penicillium chrysogenum KF-25. BMC Genom 15(1):1
37. Pitt JI, Hocking AD, Diane A (2009) Fungi and food spoilage. Springer, Berlin
38. Ramoni R, Vincent F, Grolli S, Conti V, Malosse C, Boyer FD, Nagnan Le Meillour P, Spinelli S, Cambillau C, Tegoni M (2001) The insect attractant 1-octen-3-ol is the natural ligand of bovine odorant-binding protein. J Biol Chem 276(10):7150–7155
39. Rosenberger D (1990) Blue mold. Compend Apple Pear Dis 1:54–55
40. Rosenberger DA (1997) Recent research and changing options for controlling postharvest decays of apples. In: Proc. Harvest-ing, Handling, and Storage Workshop. Northeast Reg. Agric. Eng. Serv. Publ. NRAES-112. Cornell University, Ithaca, NY
41. Syed Z, Leal WS (2007) Maxillary palps are broad spectrum odor-ant detectors in Culex quinquefasciatus. Chem Senses 32:727–738
42. Thakeow P, Angeli S, Weissbecker B, Schutz S (2008) Antennal and behavioral responses of Cis boleti to fungal odor of Trametes gibbosa. Chem Senses 33(4):379–387
43. Trapnell C, Pachter L, Salzberg SL (2009) TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25(9):1105–1111
44. Trapnell C, Roberts A, Goff L, Pertea G, Kim D, Kelley DR, Pimentel H, Salzberg SL, Rinn JL, Pachter L (2012) Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat Protoc 7(3):562–578
45. Tressl R, Bahri D, Engel K (1982) Formation of eight-carbon and ten-carbon components in mushrooms (Agaricus campestris). J Agric Food Chem 30:89–93
46. Van den Berg MA, Albang R, Albermann K, Badger JH, Daran JM, Driessen AJ, Garcia-Estrada C, Fedorova ND, Harris DM, Heijne WH et al (2008) Genome sequencing and analysis of the
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022

991Journal of Industrial Microbiology & Biotechnology (2019) 46:977–991
1 3
filamentous fungus Penicillium chrysogenum. Nat Biotechnol 26(10):1161–1168
47. Viehweg S, Schmitt R, Schmidtlorenz W (1989) Microbial spoil-age of refrigerated fresh broilers.VII. Production of off odors from poultry skin by bacterial isolates. Lebensmittel-Wissenschaft und-Technologie 22:356–367
48. WHO (1990) Patulin. WHO Food Addit Ser 26:143–165 49. Wurzenberger M, Grosch W (1984) The formation of 1-octen-3-ol
from the 10-hydroperoxide isomer of linoleic acid by a hydrop-eroxide lyase in mushrooms (Psalliota bispora). Biochimica et Biophysica Acta 794(1):25–30
50. Yang F, Zhang Y, Huang Q, Yin G, Pennerman KK, Yu J, Liu Z, Li D, Guo A (2015) Analysis of key genes of jasmonic acid mediated signal pathway for defense against insect damages by comparative transcriptome sequencing. Sci Rep 5:16500
51. Yin G, Padhi S, Lee S, Hung R, Zhao G, Bennett JW (2015) Effects of three volatile oxylipins on colony development in two
species of fungi and on Drosophila larval metamorphosis. Curr Microbiol 71(3):347–356
52. Yung PY, Grasso LL, Mohidin AF, Acerbi E, Hinks J, Seviour T, Marsili E, Lauro FM (2016) Global transcriptomic responses of Escherichia coli K-12 to volatile organic compounds. Sci Rep 6:19899
53. Zawirska Wojtasiak R (2004) Optical purity of (R)-(-)-1-octen-3-ol in the aroma of various species of edible mushrooms. Food Chem 86(1):113–118
Publisher’s Note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Affiliations
Guohua Yin1,2 · Yuliang Zhang1 · Maojie Fu1 · Sui Sheng T. Hua3 · Qixing Huang1 · Kayla K. Pennerman2 · Guangxi Wu4 · Wayne M. Jurick II5 · Samantha Lee2 · Lijing Bu6 · Hui Zhao1 · Joan W. Bennett2
1 Key Laboratory of Biology and Genetic Resources of Tropical Crops, Ministry of Agriculture, Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, Hainan, China
2 Department of Plant Biology and Pathology, Rutgers, The State University of New Jersey, New Brunswick, NJ 08901, USA
3 Department of Agriculture, ARS, Western Regional Research Center, Albany, CA 94710, USA
4 Department of Energy, Oak Ridge Institute for Science and Education, Oak Ridge, TN 37831, USA
5 Department of Agriculture, Food Quality Laboratory, ARS, Beltsville Agricultural Research Center, Beltsville, MD 20705, USA
6 Department of Biology, Center for Evolutionary and Theoretical Immunology CETI, University of New Mexico, Albuquerque, NM 87131, USA
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/46/7/977/5996811 by guest on 31 May 2022




















