
Analysis of Apple Fruit (Malus domestica Borkh.) Quality ...
Upload
independentCategory
view
0download
0
Physiologia Plantarum 128: 604–617. 2006 Copyright ª Physiologia Plantarum 2006, ISSN 0031-9317
Induction of polyphenol gene expression in apple(Malus x domestica) after the application of adioxygenase inhibitorThilo C. Fischer,a,† Heidrun Halbwirth,b,† Susanne Roemmelt,c,† Emidio Sabatini,d,†
Karin Schlangen,b,† Carlo Andreotti,d Francesco Spinelli,d Guglielmo Costa,d Gert Forkmann,a
Dieter Treutterc and Karl Stichb,*
aLehrstuhl fur Zierpflanzenbau, Department fur Pflanzenwissenschaften, Technische Universitat Munchen Weihenstephan, Am Hochanger 4,
D-85350 Freising, GermanybInstitut fur Verfahrenstechnik, Umwelttechnik und Technische Biowissenschaften, Technische Universitat Wien, Getreidemarkt 9/1665,
A-1060 Vienna, AustriacFachgebiet Obstbau, Department fur Pflanzenwissenschaften, Technische Universitat Munchen Weihenstephan, Alte Akademie 16,
D-85350 Freising, GermanydDipartimento Colture Arboree, Universita di Bologna, via Fanin 46, I-40127 Bologna, Italy
Correspondence
*Corresponding author,
e-mail: [email protected]
Received 24 March 2006; revised 12
June 2006
doi: 10.1111/j.1399-3054.2006.00787.x
A comprehensive study of the complex polyphenol biosynthesis in developing
leaves of apple (Malus domestica) was performed comprising gene expression,
enzyme activities and polyphenol composition. During leaf development, an
early increase in gene expression was observed for phenylalanine ammonialyase (PAL, EC 4.3.1.5), chalcone synthase (CHS, EC 2.3.1.74), flavanone 3-
hydroxylase (FHT, EC 1.14.11.9) and dihydroflavonol 4-reductase/flavanone
4-reductase (DFR/FNR, EC 1.1.1.219). Their enzyme activities showed
a corresponding trend during the time course. A parallel set of experiments
was carried out with leaves treated with prohexadione-Ca (ProCa), which is an
enzyme inhibitor of 2-oxoglutarate dependent dioxygenases (2-ODDs). ProCa is
known to induce changes in polyphenol biosynthesis, which are accompanied
bya reduced incidenceof fire blight and scab, the twomajor pome fruit diseases.The application of ProCa led to an increase in activities of PAL, CHS, FHT and
DFR/FNR, which was based on an enhanced gene expression. In contrast, an
inhibition of gene expression was detected for anthocyanidin synthase (EC
1.14.11.19). These effects are interpreted as a feedback regulation by changed
polyphenol levels. Because of the inhibition of the 2-ODDs FHT and flavonol
synthase (EC 1.14.11.23), some pronounced changes in polyphenol composi-
tion were observed. Eriodictyol, the substrate of FHT, accumulated as
eriodictyol-7-O-glucoside and 6$-O-trans-p-coumaroyleriodictyol 3#-O-glu-coside. In addition, the 3-deoxycatechin luteoliflavan was formed which is not
present in untreated apple leaves. Hence, beyond the redirection of polyphenol
biosynthesis by the enzyme inhibitor, changed polyphenol levels obviously
cause a distinct induction of gene expression by feedback regulation.
Abbreviations – 2-ODD, 2-oxoglutarate dependent dioxygenase; ANS, anthocyanidin synthase; cDNA, complementary DNA;
CHS, chalcone synthase; DFR, dihydroflavonol 4-reductase; DW, dryweight; EDTA, ethylenediaminetetraacetic acid; FHT, flavanone
3-hydroxylase (¼F3H); FLS, flavonol synthase; FNR, flavanone 4-reductase; F3#H, flavonoid 3#-hydroxylase; GA, gibberellic acid;HPLC, high-performance liquid chromatography; mRNA, messenger RNA; NAR, naringenin; PAL, phenylalanine ammonium lyase;
ProCa, prohexadione-Ca; rDNA, ribosomal DNA; SDS, sodium dodecyl sulphate.
†Contributed equally to the results.
604 Physiol. Plant. 128, 2006

Introduction
The constitutive phenolic compounds of apple leaves
(Malus domestica) have been investigated in detail
(Gutmann et al. 1990, Mayr et al. 1995, Schieber et al.
2001, Treutter 2001). Although some of them, in
particular proanthocyanidins, were shown to play animportant part in resistance against pathogens (Feucht and
Treutter 1999, Feucht et al. 1996, 1998, Mayr et al. 1997,
Michalek et al. 1998, Picinelli et al. 1995), knowledge
about their biosynthesis in apple leaves is still limited
(Fischer et al. 2003,Halbwirth et al. 2003). Little is known
about resistance induction under field conditions, and the
accumulation of resistance-related polyphenols during
leaf development is poorly investigated.Recently, interest has focused on possibilities tomodify
the polyphenol composition to induce resistance to
pathogens (Halbwirth et al. 2006, Norelli and Miller
2004, Rademacher et al. 2000a, 2000b). One effective
tool for themodification of the polyphenol composition is
the application of enzyme inhibitors such as prohex-
adione-Ca (ProCa), which was originally developed as
a growth regulator. As a 2-oxoglutarate analogue, itinhibits 3b-hydroxylation of gibberellic acidGA20,which
is catalysed by a 2-oxoglutarate dependent dioxygenase
(2-ODD), and thus impedes the formation of the active
gibberellic acid GA1 (Rademacher et al. 1992, 2000). In
addition, 2-ODDs involved in the polyphenol biosynthe-
sis such as flavanone 3-hydroxylase (FHT), flavonol
synthase (FLS) and anthocyanidin synthase (ANS) are
affected (Halbwirth et al. 2006). Treatments with ProCacause an effective inhibition of FHT in apple leaves,
which leads to changes in the polyphenol composition
and to the occurrence of 3-deoxyflavonoids, which are
not formed in untreated apple leaves (Fischer et al. 2003,
Halbwirth et al. 2003, Roemmelt et al. 2003). 3-
Deoxyflavonoids are known to contribute to resistance in
maize and sorghum species (Lo et al. 1999, Mueller-
Harvey and Reed 1992, Lopes 1993, Snyder andNicholson1990, Styles and Ceska 1972, Tenkouano et al. 1993), and
a strong anti-microbiological activity of the 3-deoxyflavo-
noid luteoforol (3-deoxyleucocyanidin) could be demon-
strated (Spinelli et al. 2005). ProCa-treated apple trees
showed reduced susceptibility to fire blight and scab (Bazzi
et al. 2003). Thus, the induction of 3-deoxyflavonoid
formation, in particular of luteoforol, may provide a very
strong defence mechanism against plant pathogens.Apart from FHT inhibition by ProCa (Halbwirth et al.
2006) and the fact that apple dihydroflavonol 4-reductase
(DFR) shows also a flavanone 4-reductase (FNR) activity
(Fischer et al. 2003), little information was available on
the underlying molecular mechanisms for the observed
resistance induction. We here provide the first study,
which investigates the effect of ProCa on the polyphenol
biosynthesis at the levels of gene expression, enzyme
activity and polyphenol accumulation. This knowledge
will form the basis of strategies for an effective defence
management against fire blight and scab.
Materials and methods
Plant material
The experiments were carried out on M. domestica cv.‘Golden Delicious’ in May 2000 during periods of
uniform and sunny weather conditions. One hundred
2-year-old scions grafted on ‘M9’ rootstocks kept in the
orchard of the Department of Arboriculture, University of
Bologna, were used. Half of the plants were sprayed once
in the evening with 250 ppm ProCa until the leaf surfaces
were completely wetted, whilst the control was treated
with water. The first three leaves were labelled. Youngest(leaf 1), second (leaf 2) and third leaves (leaf 3) were
collected separately. Representative samples of 15 leaves
were collected in total for each developmental stage and
time from five different trees and distributed for analyses
of polyphenol composition, enzyme activities and gene
expression, respectively. Samples were drawn before (0 h)
and five times after treatment (4 h, 1 day, 2 days, 5 days,
10 days). The plant material was frozen in liquid nitrogenand kept at 280�C. For high-performance liquid chroma-
tography (HPLC) analysis, the material was freeze dried.
Chemicals
For spray application of ProCa, the formulation BAS 125
10 W (Regalis�, BASF AG, Ludwigshafen, Germany),
a wettable granular containing 10% by weight of ProCa
was used. [2-14C]-Malonyl-coenzyme A (55 mCimmol21) was purchased from Amersham International
(Freiburg, Germany). [14C]-Naringenin (NAR) was pre-
pared as described (Britsch and Grisebach 1985), using
recombinant chalcone synthase (CHS) (Schroeder, Uni-
versity of Freiburg, Germany). [14C]-Eriodictyol was
synthesized from [14C]-NAR (Forkmann and Stotz
1981), using F3#H-activity of microsomal preparations
from Tagetes erecta. [14C]-Dihydrokaempferol was pre-pared from [14C]-NARwith recombinant FHT (Halbwirth
et al. 2006) and used for preparation of [14C]-dihydro-
quercetin (Forkmann and Stotz 1981), with microsomal
preparations from T. erecta.
Analyses of phenolic compounds
Determination and identification of phenolic compounds
were performed according to Roemmelt et al. (2003). For
Physiol. Plant. 128, 2006 605

extraction of phenolic compounds, lyophilised leaves
were ground in a ball mill. One milligram of the fine
powder was extracted with 500 ml of 100% methanol
containing 6-methoxyflavone (0.1 mg ml21) as an inter-
nal standard for 30 min in a cooled water bath during
sonication. After centrifugation the supernatant wasevaporated, the residue redissolved in small quantities
ofmethanol and injected forHPLC analysis. The phenolic
compounds were separated on a column (250 � 4 mm
inner diameter, I.D.) prepackedwithHypersil ODS, 3 mm
particle size, following a stepwise gradient, using
mixtures of solvent A (formic acid, 5% in water) and
solvent B (methanol), from 95:5 (v/v) to 10:90 (v/v), with
a flow rate of 0.5 ml min21 (Treutter et al. 1994). Thegradient profile usedwas 0–5 min, isocratically, 5%B; 5–
15 min, 5–10% B; 15–30 min, isocratically, 10% B; 30–
50 min, 10–15% B; 50–70 min, isocratically, 15% B;
70–85 min, 15–20% B; 85–95 min, isocratically, 20% B;
95–110 min, 20–25% B; 110–140 min, 25–30% B; 140–
160 min, 30–40% B; 160–175 min, 40–50% B; 175–
190 min, 50–90% B. For the selective estimation of
flavans, a postcolumn derivatisation method was em-ployed (Treutter et al. 1994). Flavanones, catechins and
procyanidins aswell as the novel compound luteoliflavan
were identified according to their chromatographic
behaviour on HPLC and thin layer chromatography and
in comparison with previously isolated standards (Mayr
et al. 1995, Roemmelt et al. 2003). Flavonols were
selectively detected at 360 nm and identified by their
ultraviolet absorbance spectra with a diode array de-tector. Quantificationwas performed as follows: catechin
and epicatechin were available as standards; procyani-
dins were calculated as procyanidin B2, luteoliflavan as
catechin, flavonols as rutin and flavanones as eriodictyol-
7-O-glucoside. For the HPLC determination of the
dihydrochalcone glycoside phloridzin the extract was
diluted 1:200 with methanol and the phloridzin was
analysed using a short column (12.5 � 4 mm I.D.)
prepacked with LiChrospher 100 RP18, 5 mm particle
size and a gradient ranging from 40 to 90% aqueous
methanol (Leser and Treutter 2005).
Determination of enzyme activities
Enzyme preparations and assays were performed as
previously described (Halbwirth et al. 2002). Crude
extracts were passed through a gel chromatography
column (Sephadex G25) to remove low molecular
compounds. This step was particularly important for the
determination of FHT activity from ProCa-treated leaves
because remnants of the inhibitor were shown to disturb
the analyses. Specific activities were calculated based onthe protein content, whichwas determined by amodified
Lowry procedure (Sandermann and Strominger 1972)
using bovine serum albumin as a standard. For compar-
ison of the datameasured on treated anduntreated leaves,
the activities at time 0 were taken as 100% and were
calculated relative to the activity at time 0.
Molecular cloning of complementary DNAs
The partial complementary DNA (cDNA) of phenylala-
nine ammonia lyase (PAL) and the full size clones of FHT,
DFR, FLS, ANS and an 18S rDNA fragment were isolated
from M. domestica cv. ‘M9’, a widely used rootstock
cultivar. The partial cDNA of CHS was obtained from
M.domestica cv. ‘Weirouge’. Isolation ofmessenger RNA
(mRNA) was performed using the mMACS mRNA Iso-lation Kit� (Miltenyi Biotec, 752-01; Bergisch Gladbach,
Germany). Reverse transcription was done with the
SuperScript II� Reverse Transcriptase (Gibco BRL 1864)
and an oligo-dT-anchor-primer (Table 1). The following
reverse transcription-PCR (RT-PCR) conditions were
used: 1.5 min 94�C, 30� (30 s, 94�C; 1 min, 45–60�C;2 min, 72�C), 7 min 72�C. RT-PCR products obtained
with the optimal annealing conditions (45–60�C) were
Table 1. PCR primers used for the cloning of complementary DNAs. ANS, anthocyanidin synthase; PAL, phenylalanine ammonium lyase; CHS, chalcone
synthase; DFR, dihydroflavonol 4-reductase.
Primer Sequence Reference for sequence
oligo-dT-anchor-primer GAC CAC GCG TAT CGA TGT CGA C(T)16V –
5# PAL GTC GAC GAG CAG CAC AAT CAG G Davies and Bradley, Genbank X68126
3# PAL ATG CAG CAT GTA AAC CGT GAC G Davies and Bradley, Genbank X68126
5# CHS ATG ATG TAC CAG CAG GGG TGC Podivinsky et al. 1993, X68977
3# CHS CTT CAA GCA GCC ACG CTG TG Podivinsky et al. 1993, X68977
5# ANS CGA GTA ATA TAC TAG CTGAG Lee et al. 1998, Genbank AF117269
3# ANS ATT AGG ACG ATA GTT CAC AAC Lee et al. 1998, Genbank AF117269
5# 18S rDNA GAC TGT GAA ACT GCG AAT GG Kim et al., Genbank AF179400
3# 18S rDNA GTA AGT TTC AGC CTT GCG ACC Kim et al., Genbank AF179400
5# degen DFR GGX TTYATH GGB TCW TGG CTY RTC ATG A Amino acid motif GF(I/V)GSWL(V/I)M
3# degen DFR TCD AYX GCH CCH XYR WAC ATR TCC TC Amino acid motif EDM XXG AXX
606 Physiol. Plant. 128, 2006

chosen for cloning,whichwas performedwith the ‘TOPO
TA Cloning� Kit’ (Invitrogen, 45-0071; Karlsruhe, Ger-
many). By this PCR-approach the fragments of PAL and
CHS cDNAs as well as the full size cDNA of ANS were
cloned. Sequences were determined by a commercial
supplier to verify the identity of the cloned cDNAs.Primers for RT-PCR amplification of PAL, CHS and ANS
and the sequence references are given in Table 1. TheM.
domestica M9 ANS is available under the accession
number DQ156905. Full size cDNAs of M. domestica
M9 DFR, FHT and FLS were available from previous
studies (Fischer et al. 2003, Halbwirth et al. 2006,
accession no AY227728, AY965340, AY965343). An
18S rDNA-fragment was amplified fromM. domestica cv.‘M9’ genomic DNA using primers that were derived from
the 18S rDNA sequence known from Pyrus pyrifolia
(Table 1). This fragmentwas used as a hybridization probe
to quantify and normalize the amount of gel-loaded total
RNA. DFR fragments were PCR amplified from cDNA
derived from ProCa-treated leaves and untreated control
leaves. The degenerated DFR primers were derived from
conserved amino acid sequences (Table 1).
Northern analysis
Gene expression studies were performed by Northern
analysis. Total RNAwas prepared from first leaves of apple
plants using the RNeasy Kit� (Qiagen; Hilden, Germany)
and stored for later use at –80�C. Denaturing 1% agarose
gels were run with 20 mM 3-morpholinopropane sulfonicacid,MOPS (pH7.0), 8 mM sodiumacetate (NaAc), 1 mM
ethylenediaminetetraacetic acid (EDTA), 6.6% formalde-
hydeand0.5 mg ml21 ethidiumbromide. Fivemicrograms
of RNAwas loaded after addition of 0.33 volumes of 37%
formaldehyde and 0.33 volumes of 100 mM MOPS (pH
7.0), 40 mM NaAc, 5 mM EDTA, heating to 65�C,subsequent cooling on ice, addition of 0.33 volumes of
TE/bromophenol blue and parallel prerun of gel electro-phoresis. All samples of one time course were loaded on
the same gel. Gel electrophoresis was performed at 40 V.
The gel was washed several times with H2O and photo-
graphed to check equal RNA loading. Further washings
were carried out for 20 min in 50 mM NaOH, 20 min in
H2O and 45 min in 20 � SSC. Membrane transfer was
performed with 20 � SSC on Immobilon Nylon N1�
membrane (Millipore, Schwalbach, Germany) over night.After blotting the membrane was briefly washed with
6 � SSC, air-dried and baked for 2 h at 80�C. DNA-
labelling was performed using the RediprimeTM II kit (RPN
1633, Amersham). Hybridizations were performed with32P-labelledDNA in 2 � SSC at 63�C for at least 15 h. The
blot was washed three times with 0.2 � SSC/1% sodium
dodecyl sulphate (SDS) at 63�C. Exposition was performed
with a phosphor-imager (Fuji BAS 1000 Bio-Imaging
Analyser, screens: BAS-MS 2040, Fuji). For the analysis,
the program TINA 2.09 (Raytest, Straubenhardt, Germany)
was used. For quantification equal areas were defined for
the bands, and resulting values were corrected for the
background. After hybridization with the gene-specificprobe, the blots were additionally hybridized with 32P-
labelled 18S rDNA. The resulting values were used for
normalization of the gene-specific values. The values for
the time0were takenas100% for the treated anduntreated
time course, respectively.
Southern analysis
Genomic DNA was prepared from young apple leaves
using the DNeasy� Plant DNA Kit (Qiagen). Five micro-
grams of DNA each was digested with 50 U restriction
enzyme (BamHI, EcoRI, HindIII, SalI, SacI and XbaI) in
the respective buffer for 5 h at 37�C. The restricted DNA
was ethanol precipitated, redissolved in 20 ml TE at 65�Cand used for agarose gel electrophoresis (1% agarose,
1 mg ethidium bromide l21, TAE buffer, 30 V). The gelwas soaked in 0.25 M HCl for 15 min afterward, rinsed
withwater,washed twice in 0.5 MNaOH/1.5 MNaCl for
20 min, rinsed with water and washed two times in 1 M
Tris pH 7.4/1.5 M NaCl for 20 min. Blotting transfer was
done with 0.4 M NaOH on Immobilon Nylon N1�
membrane (Millipore) overnight. After blotting the
membrane was briefly washed with 5 � SSPE, it was
air-dried and baked for 30 min at 80�C. Hybridizationswere performed with 32P-labelled DNA (see Northern
analysis) in hybridization buffer (3 � SSPE/0.02% Ficoll/
0.02% polyvinyl pyrrolidone (40 000 D)/0.1% SDS/
50 mg l21 preboiled calf thymus DNA) at 63�C for at
least 15 h. The blot waswashed twice with 2 � SSPE/1%
SDS and oncewith 2 � SSPE/0.1%SDS at 63�C for 5 min
each. Exposition and analysis were performed as
described for Northern analysis.
Results
The experiments were performed under orchard con-
ditions on the economically important apple cultivar
‘Golden Delicious’ to provide data representing natural
conditions. Polyphenol gene expression, activity of the
corresponding enzymes and polyphenol accumulationwere studied from first leaves of apple shoots over a time
course of 10 days. Because of the accumulation of
disturbing substances in RNA preparations from the
second and third leaves, quantification of gene expres-
sion was only possible in the first leaves. Enzyme
activities and corresponding polyphenol composition
were determined also from second and third leaves.
Physiol. Plant. 128, 2006 607

Cloning of cDNAs and gene number determination
Full sized cDNAs fromM. x domesticaM9were availablefor DFR, FHT and FLS from previous work (Fischer et al.
2003, Halbwirth et al. 2006). The function of these genes
had been confirmed by expression of the recombinant
enzymes. PAL and CHS cDNA fragments and a full sized
ANS cDNAwere RT-PCR-amplified from M9 leaf cDNA
using available sequence information, cloned and
confirmed by sequencing. To test whether further DFR/
FNR genes are induced after ProCa treatment, whichcould be responsible for the 3-deoxyflavonoid formation,
DFR/FNR fragments of approximately 900-bp length
were PCR amplified with degenerated DFR primers from
ProCa-treated leaves and untreated control leaves of the
cultivar ‘Golden Delicious’, cloned and sequenced.
There was only little sequence variation in comparison
with the known DFR/FNR sequence of M9, and variation
was even less for the sequences recovered from thetreated leaves (data not shown). No hint was found by this
approach for induction of further DFR/FNR genes after
ProCa treatment. The cloned cDNAs of the various
polyphenol genes were used as hybridization probes for
Southern analyses and for the gene expression studies
performed by Northern analyses.
Southern analyses of ‘M9’ apple genomicDNAcleaved
with six different restriction enzymes revealed theexistence of small multigene families for all the genes
studied except ANS (Table 2). For some genes that had
been included in previous studies on apple fruits,
comparable results of small multigene families were also
obtained for ‘Jonathan’ apple (Honda et al. 2002) and
‘Fuji’ apple (Kim et al. 2003). Most apple cultivars are
highly heterozygous. Therefore, some bands in Southern
analysis could be because of restriction fragment lengthpolymorphisms between parent lines, which would
reduce the definite gene number of the multigene family.
Gene expressions in first leaves
The expression studies included cDNAsof six genes of the
polyphenol pathway. 18S rDNA fragment hybridizationwas performed for quantification and normalization of
the total amount of RNA loaded on the gels (Fig. 1). For
comparison of the data for treated and untreated leaves,
the values for time ‘0 h’ were taken as 100%, all other
data refer to this, respectively (Fig. 2).
During the first 2 days, untreated unrolling first leaves
showed an increase in gene expression for PAL, CHS, FHT
and DFR/FNR, whereas FLS und ANS genes showed onlyminor changes (Fig. 2, dashed lines). Surprisingly, the
expression of all six genes was markedly decreased on
day 5, but thereafter expression increased again. In
ProCa-treated leaves, inductive effects on gene expres-
sions were observed (Fig. 2, full lines). CHS, FHT and
DFR/FNR showed strong induction of gene expression
during the first days,whereas for PAL only a late induction
effect was found. FLS showed only a weak and lateinduction, and ANS gene expression even decreased
slightly. As observed for untreated leaves, a—partly
considerable—decrease in expression of all six genes
was measured on day 5, followed by a late increase.
Despite filing of meteorological data during the orchard
experiment, no environmental influence could be iden-
tified that could explain the decrease at day 5.
Enzyme activities in leaves
The enzymatic studies focused on the activities of PAL,
CHS, FHT and DFR/FNR. FLS and ANS activities could
not be detected with the enzyme preparations from apple
leaves. For all enzymes, specific enzyme activities were
calculated per total protein concentration. For compar-
ison of the datameasured on treated anduntreated leaves,the activities at time 0 were taken as 100% and were
calculated relative to the activity at time 0.
During the development of untreated unrolled first
leaves, an early increase in PAL, CHS and FHT enzyme
activities was observed (Fig. 3 left, dashed lines), which
corresponded to the respective gene expressions (Fig. 2,
dashed lines). Treatment with ProCa (Fig. 3 left, full lines)
led to an increase in the activities of PAL, FHT, DFR/FNRand CHS in comparison with the untreated first leaves.
Most pronounced was a late increase in PAL activity by
ProCa treatment. The time courses of enzyme activities in
first leaves closely followed the time courses of gene
expression (Fig. 2). However, for FHT the pronounced
increase in gene expression was reflected only by a
moderate increase in enzyme activity. In treated and
untreated leaves, a decrease in the activities of all fourenzymeswasmeasured at day 5 (Fig. 3), as observed also
for gene expression.
All enzyme activities were still present in the enzyme
preparations of second and third leaves. During leaf
development, a distinct decrease in protein concentration
was observed from 1.4 mg ml21 enzyme preparation in
first leaves to 0.7 mg ml21 enzyme preparation in third
Table 2. Estimation of gene copy numbers in Malus x domestica cv.
Golden Delicious, each relying on Southern analysis of six different
genomicdigests. PAL, phenylalanineammonium lyase;CHS, chalcone syn-
thase; FHT, flavanone 3-hydroxylase; DFR, dihydroflavonol 4-reductase;
FLS, flavonol synthase; ANS, anthocyanidin synthase.
Gene PAL CHS FHT DFR FLS ANS
Estimated number of homologues 3-4 4-5 2 3-5 2-3 1
608 Physiol. Plant. 128, 2006
leaves. Because protein concentration and reaction rates
simultaneously decreased over the investigation period of
10 days, specific activities (kat kg21 protein) of all enzymes
remained rather stable. Time courses for first, second and
third leaf enzyme activities are quite similar. Generally, the
inductive effect of ProCa could be seen in all three leaves.
For the enzymatic investigations, ProCa was removed
by gel chromatography from the enzyme preparations todetermine the FHTenzyme activity in the absence of the
enzyme inhibitor. However, the in vivo inhibition could
be demonstrated by omission of the gel chromatography
step in the enzyme preparation. When ProCa residues
were not removed from the enzyme preparations of
treated leaves, a distinct FHTenzyme inhibition lasting for
at least 5 days after treatment of the leaves with ProCa
was observed (Fig. 4). In these assays, the FHT inhibitionmasks the enhanced FHTactivity,which is the result of the
induced FHT gene expression.
Composition and concentration of phenoliccompounds in developing apple leaves
Seventeen main, constitutively formed phenolic com-
pounds of apple leaves were analysed. This included
hydroxycinnamic acids (p-coumaric acid, chlorogenic
acid, p-coumaroylglucose), dihydrochalcones (phloretin,
phloridzin, 4-O-cis-p-coumaroyl-phloridzin, 4-O-trans-
p-coumaroylphloridzin), the flavanone naringenin 7-O-
glucoside, derivatives of the flavonol quercetin (hyperin,
isoquercitrin, rutin, quercitrin), flavan 3-ols (catechin,
epicatechin) and derived polymeric procyanidins (B2, B5
and E-B5). In addition, three compounds occurring onlyafter ProCa treatment (Roemmelt et al. 2003) were
determined: the flavanones eriodictyol-7-O-glucoside
and 6$-O-trans-p-coumaroyleriodictyol 3#-O-glucoside
and the 3-deoxycatechin luteoliflavan (Fig. 5). The poly-
phenol contents given in Figs 6 and 7 are based on dry
weight (DW).
Dihydrochalcones are the main phenolic compounds
in apple leaves. Their concentrations remained quitestable during the whole investigation period, showing
values between 200 and 250 mg g21 DW in first leaves,
150–250 mg g21 DW in second leaves and 150–200 mg
g21 in third leaves. The flavonol content was lowest in
the youngest leaves and increased slowly during leaf
development (Fig. 6 first row, dashed lines). The catechin
content was rather low (below 0.15 mg g21 DW) and
remained quite constant during the developmental
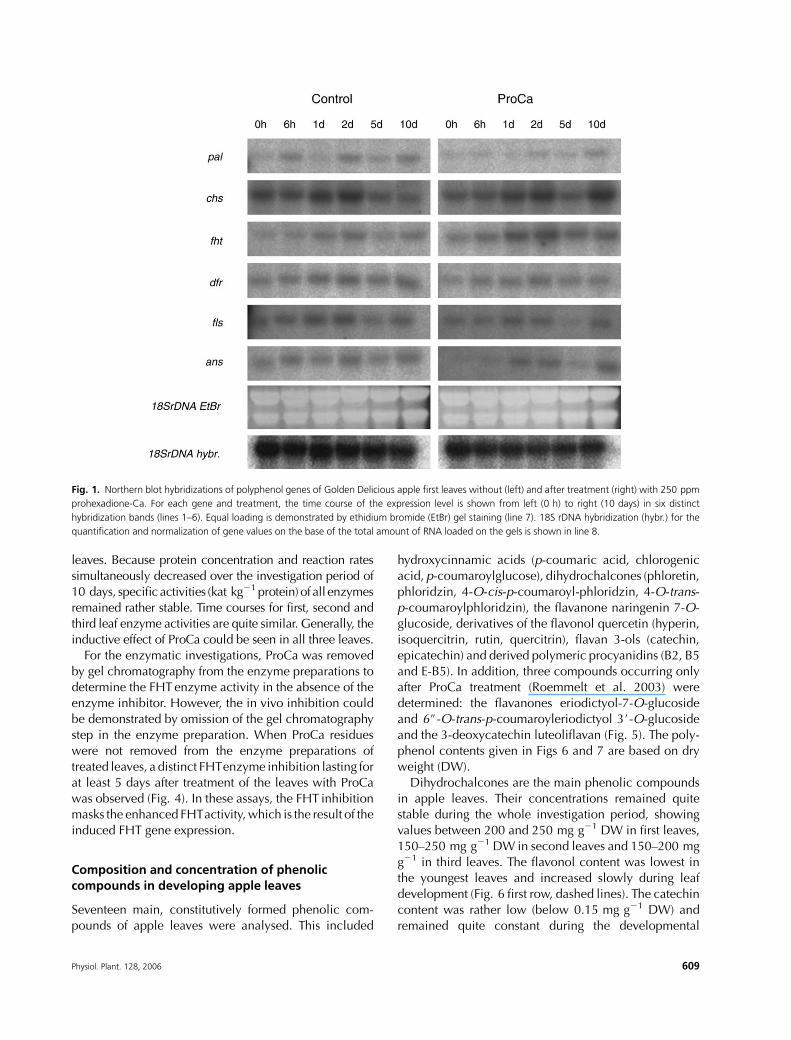
Fig. 1. Northern blot hybridizations of polyphenol genes of Golden Delicious apple first leaves without (left) and after treatment (right) with 250 ppm
prohexadione-Ca. For each gene and treatment, the time course of the expression level is shown from left (0 h) to right (10 days) in six distinct
hybridization bands (lines 1–6). Equal loading is demonstrated by ethidium bromide (EtBr) gel staining (line 7). 18S rDNA hybridization (hybr.) for the
quantification and normalization of gene values on the base of the total amount of RNA loaded on the gels is shown in line 8.
Physiol. Plant. 128, 2006 609

process, while epicatechin and the derived procyanidinsincreased during the investigation period (Fig. 6, dashed
lines). Hydroxycinnamic acids also increased during the
investigation period from about 1 to 5 mg g21 DW.
Pronounced changes in the concentrations of the
constitutive phenolic compounds were observed after
ProCa treatment. Naringenin 7-O-glucoside concen-
tration was markedly increased (Fig. 7 first row). The
flavonols showed much lower contents in treated leavesduring the whole period of the experiment (Fig. 6 first
row). Catechin and epicatechin concentrations markedly
increased, particularly in first and second leaves (Fig. 6
second and third row, full lines). The formation of the
oligomeric flavan 3-ols (procyanidins) slightly increased
in first leaveswhile therewas no effect in second and third
leaves (Fig. 6 last row, full lines). In contrast, procyanidin
E-B5 decreased in all leaves treated with ProCa (data notshown). The content of hydroxycinnamic acids was only
weakly affected by ProCa, and the content of dihydro-
chalcones was not influenced at all (data not shown).
In addition, treatmentwithProCa led todistinct changesin the polyphenol composition and concentration. In
agreement with earlier studies (Roemmelt et al. 1999,
2003), three novel compounds were detected. In first and
second leaves, considerable amounts of luteoliflavanwere
formed, reaching a maximum of approximately 1 mg g21
DW5 days after treatment (Fig. 7 second row, full lines). In
third leaves, much lower amounts of luteoliflavan were
found (up to 0.2 mg g21 DW). Furthermore, the formationof eriodictyol glycosides was induced by ProCa. Highest
concentrations were observed in the youngest leaves
where they reached up to 1.4 mg g21 DW 10 days after
treatment (Fig. 7 third row, full lines). In third leaves,
eriodictyol glycosides accumulated only to a small extent.
Discussion
The effect of ProCa on the polyphenol composition
in apple leaves and the related beneficial effects
for plant health have been intensively investigated
PALfirst leaf
00 2 4 6 8 10
100
200
300
400
500
Time course (days)
0 2 4 6 8 10
Time course (days)
% r
elat
ive
% r
elat
ive
% r
elat
ive
0 2 4 6 8 10
Time course (days)
0 2 4 6 8 10
Time course (days)
0 2 4 6 8 10
Time course (days)0 2 4 6 8 10
Time course (days)
0
100
200
300
400
500
% r
elat
ive
0
100
200
300
400
500
0
100
200
300
400
500
% r
elat
ive
0
100
200
300
400
500
% r
elat
ive
0
100
200
300
400
500
CHSfirst leaf
FHTfirst leaf
DFRfirst leaf
FLSfirst leaf
ANSfirst leaf
Fig. 2. Time course of polyphenol gene expression in apple first leaveswithout (dashed lines) and after treatment (full lines)with 250 ppmprohexadione-
Ca. Relative gene expressions (%) were calculated with reference to the value measured for 0 h.
610 Physiol. Plant. 128, 2006

before (Halbwirth et al. 2003, Roemmelt et al. 2003).
However, there have been few studies on how the
enzymes and genes from the polyphenol pathway are
affected by ProCa (Fischer et al. 2003, Halbwirth et al.
2006). The parallel sets of data are valuable to under-
stand the changes in the levels of constitutive compound
and the formation of novel compounds after ProCa
treatment.
Polyphenol biosynthesis in young apple leaves
Generally, gene expressions and enzyme activities
showed parallel time courses, whereas changes in the
phenolic contentwere only observedwith a certain delay.
As the enzyme activities in leaves of different ages
showed quite similar time courses, it must be concluded
that the observed time courses are because of environ-
mental factors rather than leaf development. This is
PALfirst leaf
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
0
0 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
00 5 10
50
100
150
200
250
Time course (days)
% r
elat
ive
PALsecond leaf
PALthird leaf
CHSfirst leaf
CHSsecond leaf
CHSthird leaf
FHTfirst leaf
DFRfirst leaf
FHTsecond leaf
DFRsecond leaf
FHTthird leaf
DFRthird leaf
Fig. 3. Time course of polyphenol enzyme activities in apple first (left), second (middle) and third (right) leaves without (dashed lines) and after treatment
(full lines) with 250 ppm prohexadione-Ca. Relative enzyme activities (%) were calculated with reference to the value measured for OH.
Physiol. Plant. 128, 2006 611

supported by the high enzyme activities observed even in
youngest leaves on day 0,which indicate an early onset of
polyphenol biosynthesis in the still closed leaf. Further-
more, no distinct correlation could be observed between
the accumulation of phenolic compounds and their
positioning in the sequence of the polyphenol pathway.
For example, the formation of phloridzin and catechin,
which represent an early and a late biosynthetic step
(Fig. 5), respectively, could both already be determined in
the youngest leaves, and no significant further increases
gel chromatography
00 2 4 6 8 10
20406080
100120140160
Time course (days)0 2 4 6 8 10
Time course (days)
% r
elat
ive
no gel chromatography
020406080
100120140160
% r
elat
ive
Fig. 4. Time course of flavanone 3-hydroxylase (FHT) activity in apple first leaveswithout (dashed lines) and after treatmentwith 250 ppmprohexadione-
Ca (ProCa) (full lines). Left: Assays performed with enzyme preparations purified via gel chromatography (residual ProCa removed) showed strongly
induced FHT activities (corresponding to induced FHT gene expression) for ProCa applications. Right: Assays with enzyme preparations not purified by
a gel chromatography step clearly showed the FHTenzyme inhibition by remaining ProCa from the leaf treatments, reflecting the in vivo conditions.
Luteoliflavan 5-O-glucoside(3-Deoxycatechin)
Luteoliflavan(3-Deoxycatechin)
O
OH
OH
OH
HO
O
OR
FLS
ANS
PAL
HO
OH
O
p-Coumaric acid
Phenylalanine
O
O
OH
OH
OH
HO
COOHHO
Chlorogenic acid
HO
O
OGlu
p-Coumaroyl glucose
OH
OH
HO OH
O
Phloretin (Dihydrochalcone)
Phloridzin(Dihydrochalcone)
HO OH
OOGlu
O O
OH4-O-p-Coumaroyl-phloridzin
(Dihydrochalcone)
Epicatechin
Procyanidins
DFR
OGluO
O
OH
OH Naringenin(Flavanone)Naringenin 7-O-glucoside
(Flavanone)
Eriodictyol(Flavanone)
OGluO
O
OH
OH
OH
Eriodictyol 7-O-glucoside(Flavanone)
Dihydroquercetin(Dihydroflavonol)
Flavan 3, 4-diol
Catechin
CHS/CHI
FNR=DFR
Quercetin 3-O-Glycosides(Flavonol)
6’’-O-trans-p-Coumaroyl-eriodictyol 3’glucoside
(Flavanone)
Luteoforol (Flavan 4-ol)
O
OH
OH
OH
HO
FHT
O
OH
OH
OH
HO
OH
O
OH
OH
OH
HO
OH
Fig. 5. Overviewon the polyphenol biosynthesis in apple leaves. Compounds occurring exclusively after prohexadione-Ca application aremarked in grey.
Dashed arrows indicate unknown biosynthetical steps.
612 Physiol. Plant. 128, 2006

Flavonolsleaf 1
00 2 4 6 8 10
2
4
6
8
10
12
14
Time course (days)
0 2 4 6 8 10Time course (days)
0 2 4 6 8 10Time course (days)
0 2 4 6 8 10
Time course (days)
Mg
g–1D
W
00 2 4 6 8 10
2
4
6
8
10
12
14
Time course (days)M
gg–1
DW
00 2 4 6 8 10
2
4
6
8
10
12
14
Time course (days)
Mg
g–1D
W
Catechinleaf 1
0
0.2
0.4
0.6
0.8
0.2
0.4
0.6
0.8
Mg
g–1D
W
0 2 4 6 8 10
Time course (days)
0
0.2
0.4
0.6
0.8
Mg
g–1D
W
0 2 4 6 8 10Time course (days)
0
0.2
0.4
0.6
0.8
Mg
g–1D
WEpicatechin
leaf 1
Flavonolsleaf 2
Catechinleaf 2
Epicatechinleaf 2
Flavonolsleaf 3
Catechinleaf 3
Epicatechinleaf 3
0
1
Mg
g–1D
W
0 2 4 6 8 10Time course (days)
0.2
0.4
0.6
0.8
0
1
Mg
g–1D
W
0 2 4 6 8 10Time course (days)
0.2
0.4
0.6
0.8
0
1
Mg
g–1D
W
Procyanidinsleaf 1
Procyanidinsleaf 2
Procyanidinsleaf 3
0
1
2
3
4
5
Mg
g–1D
W
0 2 4 6 8 10Time course (days)
0
1
2
3
4
5
Mg
g–1D
W
0 2 4 6 8 10
Time course (days)
0
1
2
3
4
5
Mg
g–1D
W
Fig. 6. Time course of flavonol and flavan 3-ol contents in apple first (left), second (middle) and third (right) leaves without (dashed lines) and after
treatment (full lines) with 250 ppm prohexadione-Ca.
Physiol. Plant. 128, 2006 613

were observed. In contrast, concentrations of flavonols,
epicatechin and derived procyanidins slightly increased
during leaf development. This coincides with previous
findings on very young shoot tips (Roemmelt et al. 2003).
Polyphenol biosynthesis in ProCa-treated leaves
After treatment with ProCa, two different effects on the
polyphenol pathway were observed. On the one hand,
partial inhibition of the 2-ODDs FHT, FLS and ANS led to
distinct changes of the polyphenol spectrum in the leaves.
On the other hand, therewas a general increase in enzymeactivities of thepolyphenol pathwaybasedonanenhanced
expression of their genes. This is an interesting effect of
ProCa treatment, which has not been described before.
Changes of the polyphenol spectrum
The inhibition of FHT primarily results in an accumula-
tion of flavanones, which are further modified viaglycosylation and acylation. This leads to the increased
formation of the constitutively present naringenin 7-O-
glucoside and to the occurrence of the two novel
compounds eriodictyol-7-O-glucoside and 6$-O-trans-
p-coumaroyleriodictyol 3#-O-glucoside (Roemmelt et al.
2003). Derivatives of hydroxycinnamic acid start to
accumulate 3 days later than flavanone derivatives as
they are involved in upstream steps and thus are affectedwith a certain delay by substrate tailback. The late
increase in hydroxycinnamic acids up to 10 days after
treatment reflects the late induction of PAL.
Naringenin 7-O-glucoside leaf 1
0.00 2 4 6 8 10
0 2 4 6 8 10
0 2 4 6 8 10
0.2
0.4
0.6
0.8
Time course (days)
mg
g–1D
W
0.00 2 4 6 8 10
0.2
0.4
0.6
0.8
Time course (days)
mg
g–1D
W
0.00 2 4 6 8 10
0.2
0.4
0.6
0.8
Time course (days)
mg
g–1D
W
Naringenin 7-O-glucosideleaf 2
Naringenin 7-O-glucosideleaf 3
Eriodictyol glycosidesleaf 1
0.0
0.4
0.8
1.2
1.6
Time course (days)
mg
g–1D
W
0 2 4 6 8 100.0
0.4
0.8
1.2
1.6
Time course (days)
mg
g–1D
W
0 2 4 6 8 100.0
0.4
0.8
1.2
1.6
Time course (days)
mg
g–1D
W
Eriodictyol glycosidesleaf 2
Eriodictyol glycosidesleaf 3
Luteoliflavanleaf 1
0.0
0.4
0.8
1.2
Time course (days)
mg
g–1D
W
0 2 4 6 8 100.0
0.4
0.8
1.2
Time course (days)
mg
g–1D
W
0 2 4 6 8 100.0
0.4
0.8
1.2
Time course (days)
mg
g–1D
W
Luteoliflavanleaf 2
Luteoliflavanleaf 3
Fig. 7. Time course of induced compounds after treatment with 250 ppm prohexadione-Ca in apple leaves (full lines) in comparison with untreated
controls (dashed lines).
614 Physiol. Plant. 128, 2006

Another consequence of the accumulation of flava-
nones is the formation of luteoliflavan (3-deoxycatechin)
from eriodictyol via the anti-microbial intermediate
luteoforol (3-deoxyleucocyanidin) occurring as early as
1 day after treatment and decreasing after 5 days.
Luteoliflavan and luteoforol belong to the rare class of3-deoxyflavonoids, which are commonly not formed in
Rosaceous species (Roemmelt et al. 2003). Previous
studies on Zea mays, Sinningia cardinalis (Halbwirth
et al. 2003,Winefieldet al. 2005) andSorghum sp. (Lo et al.
1999) suggested that natural occurrence of 3-deoxyfla-
vonoids is correlated with a low or even absent FHT
activity. Apple leaves generally show strong FHTactivities.
Accordingly, the spectrum of polyphenols is shifted fromthe common series toward the rare 3-deoxy-series only
when FHT enzyme activity is artificially reduced by
treatment with ProCa.
The accumulation of luteoliflavan can be regarded as
a marker for the sensitivity of the leaves with respect to
their inducible polyphenol biosynthesis. In previous
studies (Roemmelt et al. 2003), where very young shoot
tips were treated with ProCa, luteoliflavan concentrationreached up to 8 mg g21 DW. In our study, the maximum
values for first and second leaveswere 1 mg g21DW, and
only 0.2 mg g21 DW for third leaves on day 5 after
treatment. The restricted accumulation in third leaves
occurs despite an enhanced activity of polyphenol
enzymes (Fig. 3). This may be because of the reduced
substrate supply for PAL in older leaves compared with
younger ones where a higher sink potential can beassumed. The reduced synthesis of luteoliflavan from
eriodictyol via luteoforol may therefore be explained
by a regulation at the level of substrate supply as proposed
by Jones and Hartley (1999) and Margna (1977). This is
an important feature with respect to the defensive role
of these compounds and to resistance induction by
ProCa.
Interestingly, the FHT inhibition did not result ina simultaneous decrease for all 3-hydroxyflavonoids.
Most pronounced was the decrease in flavonol contents
after treatment. This confirms the simultaneous inhibition
of FHT and FLS (two bottlenecks in the pathway), which
results in a rapid turnover of the soluble flavonols. After
overcoming theminimum, the flavonol curves of first and
second leaves parallel those of the untreated controls.
Only in the third leaf, where the effect of ProCa wasgenerallyweakest, the flavonol levels of the treated leaves
reached those of the untreated ones.
The flavan 3-ols catechin and epicatechin accumulate
on a very low level. Surprisingly, their concentrations rise
after ProCa treatment in the first and second leaves but
their amounts remain very low. In case of catechin, this
can easily be explained by the inhibition of FLS and ANS
(concurring reactions) leading to a rise in catechin,
despite the diminished precursor synthesis because of
reduced FHTactivity (one bottleneck in the pathway). The
flavan 3-ol epicatechin, however, is producedvia the FHT
and ANS reactions (two bottlenecks in the pathway).
Thus, the observed rise in epicatechin concentration isnot yet understood.
Influence of ProCa treatment on polyphenol gene
expression and corresponding enzyme activities
Inhibition of theMalus FHTand FLS by ProCawas already
shown in vitro with leaf enzyme extracts as well as with
the recombinant enzymes (Halbwirth et al. 2002, 2006).In our study, the inhibition of FHT by residual ProCa
(Fig. 4 right) in the enzyme assays was avoided (Fig. 3,
Fig. 4 left) by generally purifying the enzyme preparations
from ProCa by gel chromatography. However, the in vivo
inhibition could be visualized in vitro by using enzyme
preparations obtained without removing of residual
ProCa by gel chromatography (Fig. 4, right). The enzyme
inhibition observed in these assays indicates that thereis no complete blockage but a strong reduction of
FHT activity during the first 5 days. Later on, the ProCa
inhibition decreases, which seems to reflect degradation
of ProCa. This interpretation is supported by the coinci-
dence with the peak of the ProCa-induced luteoliflavan
also occurring at day 5.
Apart from the quantitative and qualitative changes in
the polyphenol spectrum, the second striking observationafter ProCa treatment was the unexpected, pronounced
increase in gene expression and enzyme activities. On
the one hand, the severe changes in the phytohormone
balance caused by ProCa treatment (Rademacher 2000)
may result in changes in thewhole secondarymetabolism
and thus may be responsible for the observed influence
on polyphenol gene expression and enzyme activity.
On the other hand, the changes in polyphenol levelsafter ProCa application might be responsible for the
changes in the gene expression by feedbackmechanisms.
This seems especially evident for FHT. Such influences
of intermediates on the expression of genes and corre-
sponding enzymes in the polyphenol pathway have been
discussed before (Dedaldechamp andUhel 1999, Kubasek
et al. 1992, Pelletier et al. 1999).
With respect to the induction of gene expression, it is aninteresting question, if genes usually not expressed in
apple leaves are induced after ProCa treatment. Especially
theDFR/FNR is the keyenzyme for the alternative pathway
leading to 3-deoxyflavonoids. However, using degener-
ated primers against conserved motifs of DFR/FNR
sequences for RT-PCR no hint was found for induction of
additional DFR/FNR genes by ProCa treatment.
Physiol. Plant. 128, 2006 615

Conclusions
Our study provided an in-depth insight into the formation
of resistance-related compounds in apple leaves. On the
one hand, the constitutive polyphenols catechin, epica-techin and derived procyanidins are built very early in leaf
development, prior to leaf unrolling. All polyphenol
enzymes investigated were already present in young
unrolled leaves.On the other hand, it could be shown that
inhibition of the 2-ODDs FHT, FLS and ANS by ProCa
induces the formation of 3-deoxyflavonoids. The 3-
deoxycatechin luteoliflavan serves as an indicator for the
rather unstable precursor 3-deoxyflavonoid luteoforol,which is anti-microbially active. Apart from the 2-ODD
inhibition, a strong induction of polyphenol gene expres-
sion occurs. Most likely, this is because of a feedback
mechanism relying on polyphenol concentrations. The
stimulating effect on polyphenol gene expression may be
of practical relevance for future applications.
Acknowledgements – These investigations were supported
by the European Union Commission (QLK5-CT-1999-01583).
The authors are grateful to Laszlo Janvari for design and supply
of degenerated DFR primers. The authors thank Christine
Statnik and Jurgen Greiner for their excellent technical
assistance. Special thanks go to Thorsten Strissel for his
support during the performance of the enzymatic investiga-
tions. Finally, the authors would like to thank Eva Meggeneder
for critically reading the manuscript.
References
Bazzi C, Messina CH, Tortoreto L, Stefani E, Bini F,
Brunelli A, Andreotti C, Sabatini E, Spinelli F, Costa G,
Hauptmann S, Stammler G, Doerr S, Marr J, Rademacher
W (2003) Control of pathogen incidence in pome fruits
and other horticultural crop plants with prohexadione-Ca.
Eur J Hortic Sci 68: 108–114
Britsch L, Grisebach H (1985) Improved preparation and
assay of chalcone synthase. Phytochemistry 24:
1975–1976
Dedaldechamp F, Uhel C (1999) Induction of anthocyanin
synthesis in non-pigmented grape cell suspension by
acting on DFR substrate availability or precursor level.
Enzyme Microb Technol 25: 316–321
Feucht W, Treutter D (1999) The role of flavan-3-ols
and proanthocyanidins in plant defense. In: Inderjit, D,
Foy CL (eds), Principles and Practices in Plant
Ecology. CRC Press, Boca Raton, Florida, pp
307–338
Feucht W, Treutter D, Christ E (1996) Shot hole disease in
sour cherry: defensive flavanol barriers. Z Pflanzenkr
Pflanzenschutz 103: 279–287
Feucht W, Treutter D, Schwalb P (1998) Principles of barrier
formation of scab-infected apple fruits. Z Pflanzenkr
Pflanzenschutz 105: 394–403
Fischer TC, Halbwirth H, Meisel B, Stich K, Forkmann G
(2003) Molecular cloning, substrate specificity of the
functionally expressed dihydroflavonol 4-reductases from
Malus domestica and Pyrus communis cultivars and the
consequences for polyphenol metabolism. Arch Biochem
Biophys 412: 223–230
Forkmann G, Stotz G (1981) Genetic control of flavanone
3-hydroxylase activity and flavonoid 3#-hydroxylaseactivity in Antirrhinum majus (snapdragon). Z Naturforsch
C J Biosci 36C: 411–416
Gutmann M, Treutter D, Feucht W (1990) Analysis of phenols
of cultivars and wild species of apple by HPLC-CRD
techniques. Mitt Klosterneuburg 40: 80–83
Halbwirth H, Fischer TC, Meisel B, Kampan W, Rademacher
W, Forkmann G, Treutter D, Costa G, Bazzi C, Stich K
(2002) Biochemical and molecular biological
investigations with respect to induction of fire blight
resistance in apple and pear by transiently altering the
polyphenol metabolism with specific enzyme inhibitors.
Acta Hortic 590: 485–492
Halbwirth H, Fischer TC, Roemmelt S, Spinelli F, Schlangen
K, Peterek S, Sabatini E, Messina C, Speakman JB,
Andreotti C, Rademacher W, Bazzi C, Costa G, Treutter D,
Forkmann G, Stich K (2003) Induction of antimicrobial
3-deoxyflavonoids in pome fruit trees controls fire blight.
Z Naturforsch C J Biosci, 58C: 765–770
Halbwirth H, Fischer TC, Schlangen K, Rademacher W,
Schleifer KJ, Forkmann G, Stich K (2006) Screening for
inhibitors of the 2-oxoglutarate dependent dioxygenases:
flavanone 3b-hydroxylase and flavonol synthase.
Plant Sci 171: 194–205
Honda C, Kotoda N, Wada M, Kondo S, Kobayashi S,
Soejima J, Zhang Z, Tsuda T, Moriguchi T (2002)
Anthocyanin biosynthetic genes are coordinately
expressed during red coloration in apple skin. Plant
Physiol Biochem 40: 955–962
Jones CG, Hartley SE (1999) A protein competition model of
phenolic allocation. Oikos 86: 27–44
Kim SH, Lee JR, Hong ST, Yoo YK, An G, Kim SR (2003)
Molecular cloning and analysis of anthocyanin
biosynthesis genes preferentially expressed in apple skin.
Plant Sci 165: 403–413
Kubasek WL, Shirley BW, McKillop A, Goodman HM, Briggs
W, Ausubel FM (1992) Regulation of flavonoid
biosynthetic genes in germinating Arabidopsis seedlings.
Plant Cell 4: 1229–1236
Lee JR, Hong ST, Yoo YG, Kim SR (1998) Molecular cloning
and expression of anthocyanin biosynthesis genes from
‘Fuji apple’. Direct sequence submission to GenBank
Leser C, Treutter D (2005) Effects of nitrogen supply on
growth, contents of phenolic compounds and pathogen
(scab) resistance of apple trees. Physiol Plant 123: 49–56
616 Physiol. Plant. 128, 2006

Lo SC, De Verdier K, Nicholson RL (1999) Accumulation of
3-deoxyanthocyanidin phytoalexins and resistance to
Colletotrichum sublineolum in sorghum. Physiol Mol Plant
Pathol 55: 263–273
Lopes AMQ (1993) Phytoalexins in sorghum and their role in
interactions with phytopathogenic fungi. Summa
Phytopathol 19: 59–61
Margna U (1977) Control at the level of substrate supply—an
alternative in the regulation of phenylpropanoid
accumulation in plant cells. Phytochemistry 16: 419–426
Mayr U, Treutter D, Santos-Buelga C, Bauer H, Feucht W
(1995) Developmental changes in the phenol
concentrations of ‘Golden Delicious’ apple fruits and
leaves. Phytochemistry 38: 1151–1155
Mayr U, Michalek S, Treutter D, Feucht W (1997) Phenolic
compounds of apple and their relationship to scab
resistance. J Phytopathol 145: 69–75
Michalek S, Mayr U, Treutter D, Lux-Endrich A, Gutmann M,
Feucht W, Geibel M (1998) Role of flavan-3-ols in
resistance of apple trees to Venturia inaequalis. Acta
Hortic 484: 535–539
Mueller-Harvey I, Reed JDJ (1992) Identification of phenolic
compounds and their relationships to in-vitro digestibility
of sorghum leaves from bird-resistant and
non-bird-resistant varieties. Sci Food Agric 60: 179–196
Norelli JL, Miller SS (2004) Effect of prohexadione-calcium
dose level on shoot growth and fire blight in young apple
trees. Plant Dis 88: 1099–1106
Pelletier MK, Burbulis IE, Winkel-Shirley B (1999) Disruption
of specific flavonoid genes enhances the accumulation of
flavonoid enzymes and end-products in Arabidopsis
seedlings. Plant Mol Biol 40: 45–54
Picinelli A, Dapena E, Mangas JJ (1995) Polyphenolic pattern
in apple tree leaves in relation to scab resistance. A
preliminary study. J Agric Food Chem 43: 2273–2278
Podivinsky E, Bradley JM, Davis KM (1993) Sequence
announcements. Plant Mol Biol 21: 737–738
Rademacher W (2000) Growth retardants: effects on
gibberellin biosynthesis and other metabolic pathways.
Annu Rev Plant Physiol Plant Mol Biol 51: 501–531
Rademacher W, Temple-Smith KE, Griggs DI, Hedden P
(1992) The mode of action of acylcyclohexanediones—a
new type of growth retardant. In: Karssen CM, Van Loon
LC, Vreulgdenhil (eds), Progress in Plant Growth
Regulation. Kluwer Academic, Dordrecht, The
Netherlands, pp 571–577
Rademacher W, Speakman JB, Ammermann E, Stammler G,
Creemers P (2000a) Increasing crop resistance to
phytopathogenic fungi. PCT Int Appl. CODEN: PIXXD2
WO 2000078144 A1 20001228, 22 pp
Rademacher W, Speakman JB, Ammermann E, Jabs T,
Herbers K (2000b) Method for preparing transgenic plants
with reduced flavanone-3-hydroxylase activity and
enhanced resistance to phytopathogenic fungi and
bacteria. Ger Offen. GWXXBX DE19927575 A1
20001221, 8 pp
Roemmelt S, Treutter D, Speakman JB, Rademacher W
(1999) Effects of prohexadione-Ca on the flavonoid
metabolism of apple with respect to plant resistance
against fire blight. Acta Hortic 489: 359–363
Roemmelt S, Zimmermann N, Rademacher W, Treutter D
(2003) Formation of novel flavonoids in apple (Malus x
domestica) treated with the 2-oxoglutarate-dependent
dioxygenase inhibitor prohexadione-Ca. Phytochemistry
64: 709–716
Sandermann H, Jr, Strominger JL (1972) Purification and
properties of C 55—isoprenoid alcohol phosphokinase
from Staphylococcus aureus. J Biol Chem 247:
5123–5131
Schieber A, Keller P, Carle R (2001) Determination of
phenolic acids and flavonoids of apple and pear by
high-performance liquid chromatography. J Chromatogr
910: 265–273
Snyder BA, Nicholson RL (1990) Synthesis of phytoalexins in
Sorghum as a site-specific response to fungal ingress.
Science 248: 1637–1639
Spinelli F, Speakman JB, Rademacher W, Halbwirth H,
Stich K, Costa G (2005) Luteoforol, a flavan 4-ol, is
induced in pome fruits by prohexadione-calcium and
shows phytoalexin-like properties against Erwinia
amylovora and other plant pathogens. Eur J Plant Pathol
112: 133–142
Styles ED, Ceska O (1972) Flavonoid pigments in genetic
strains of maize. Phytochemistry 11: 3019–3021
Tenkouano A, Miller FR, Hart GE, Frederiksen RA,
Nicholson RL (1993) Phytoalexin assay in juvenile
sorghum: an aid to breeding for anthracnose resistance.
Crop Sci 33: 243–248
Treutter D (2001) Biosynthesis of phenolic compounds
and its regulation in apple. Plant Growth Regul 34:
71–89
Treutter D, Santos-Buelga C, Gutmann M, Kolodziej H (1994)
Identification of flavan-3-ols and procyanidins by high
performance liquid chromatography and chemical
reaction detection. J Chromatogr 667: 290–297
Winefield CS, Lewis DH, Swinny EE, Zhanga H, Arathoon
HS, Fischer TC, Halbwirth H, Stich K, Gosch C, Forkmann
G, Davies KM (2005) Mechanism of 3-deoxyanthocyanin
biosynthesis in Sinningia cardinalis. Physiol Plant 124:
419–430.
Edited by A. Crozier
Physiol. Plant. 128, 2006 617
Copyright © 2022 FDOKUMEN


















![Synthesis and inhibition study of monoamine oxidase, indoleamine 2,3-dioxygenase and tryptophan 2,3-dioxygenase by 3,8-substituted 5H-indeno[1,2-c]pyridazin-5-one derivatives](https://static.fdokumen.com/doc/165x107/6343bf46fc30a9d0e204e609/synthesis-and-inhibition-study-of-monoamine-oxidase-indoleamine-23-dioxygenase.jpg)
