
Ministers Reflect Ieuan Wyn Jones | The Institute for Government
Upload
independentCategory
view
2download
0
R
Is
Sa
b
c
d
a
ARRAA
KPTVMBG
1
drprc
ohiiiaa
tf
0d
Brain Research Bulletin 79 (2009) 265–270
Contents lists available at ScienceDirect
Brain Research Bulletin
journa l homepage: www.e lsev ier .com/ locate /bra inresbul l
esearch report
ndividual differences in personality traits reflect structural variance inpecific brain regions
imona Gardinia,b, C. Robert Cloningerc, Annalena Vennerib,d,∗
Department of Neuroscience, University of Parma, ItalyClinical Neuroscience Centre, University of Hull, United KingdomWashington University, Department of Psychiatry, School of Medicine, St Louis, MO, USADepartment of Neuroscience, University of Modena and Reggio Emilia, Italy
r t i c l e i n f o
rticle history:eceived 11 October 2008eceived in revised form 10 January 2009ccepted 14 March 2009vailable online 28 March 2009
eywords:ersonalityPQ
a b s t r a c t
Personality dimensions such as novelty seeking (NS), harm avoidance (HA), reward dependence (RD) andpersistence (PER) are said to be heritable, stable across time and dependent on genetic and neurobio-logical factors. Recently a better understanding of the relationship between personality traits and brainstructures/systems has become possible due to advances in neuroimaging techniques. This MagneticResonance Imaging (MRI) study investigated if individual differences in these personality traits reflectedstructural variance in specific brain regions. A large sample of eighty five young adult participants com-pleted the Three-dimensional Personality Questionnaire (TPQ) and had their brain imaged with MRI. Avoxel-based correlation analysis was carried out between individuals’ personality trait scores and grey
oxel-based morphometryRI
rainrey matter
matter volume values extracted from 3D brain scans. NS correlated positively with grey matter volumein frontal and posterior cingulate regions. HA showed a negative correlation with grey matter volume inorbito-frontal, occipital and parietal structures. RD was negatively correlated with grey matter volumein the caudate nucleus and in the rectal frontal gyrus. PER showed a positive correlation with grey mat-ter volume in the precuneus, paracentral lobule and parahippocampal gyrus. These results indicate thatindividual differences in the main personality dimensions of NS, HA, RD and PER, may reflect structural
areas
variance in specific brain. Introduction
Biosocial theories of personality indicate that some of theimensions of temperament are heritable and linked to neu-obiological markers [8,9]. Familial, clinical, longitudinal andsychobiological research has contributed to better clarify theelationship between these personality traits and their biologicalorrelates.
In the original model by Cloninger [9], three main dimensionsf personality were the focus of research, i.e. novelty seeking (NS),arm avoidance (HA) and reward dependence (RD). Novelty seek-
ng refers to an individual’s tendency to action behaviours and
t is expressed as the genetic tendency to exploratory activityn response to novelty, impulsive decision making, extravagantpproach to cues of reward and quick loss of temper. Harm avoid-nce refers to an individual’s tendency to inhibit behaviours, and∗ Corresponding author at: Clinical Neuroscience Centre, University of Hull, Cot-ingham Road, Hull HU6 7RX, England, UK. Tel.: +44 1482 465558;ax: +44 1482 465599.
E-mail address: [email protected] (A. Venneri).
361-9230/$ – see front matter © 2009 Elsevier Inc. All rights reserved.oi:10.1016/j.brainresbull.2009.03.005
.© 2009 Elsevier Inc. All rights reserved.
is expressed as an innate tendency to caution, apprehensivenessand pessimism. It manifests as pessimistic worry in anticipation offuture problems, passive avoidant behaviour, fear of uncertainty,shyness of strangers and rapid fatigability. Reward dependencerefers to an inherited tendency towards the maintenance of ongo-ing behaviours previously associated with reinforcement and itmanifests as sentimentality, persistence, social attachment anddependence on the approval of others. The Three-dimensional Per-sonality Questionnaire (TPQ) devised by Cloninger [8] measuresthese three personality dimensions, each of which includes foursubscales. Evidence from clinical and experimental studies hasalso indicated that persistence (PER), one of the subscales usedto measure reward dependence, represents a fourth independentpersonality dimension [20]. Persistence is defined as the ability togenerate and maintain arousal and motivation internally, in theabsence of immediate external reward. People who achieve highpersistence scores are generally industrious, diligent, hard working,
ambitious, overachieving and perfectionist.The TPQ was designed based on neurobiology of functional brainsystems, rather than by factor analysis which ignores informationabout the evolution, structure, and function of the brain. It wasdesigned based on studies of within-person differences in learning,

2 arch B
riirthfau
disclcscstat[hal(
tndc[se
ssdensrsriifspft(wpNpNnisgFafn
2.3. Procedure
66 S. Gardini et al. / Brain Rese
ather than between-person differences in behaviour. Factor analyt-cally derived tests assume linear relations among variables, whichs doubtful for brain systems characterized by non-linear dynamicalelationships. As a result the TPQ, but not factor analytically derivedests, provide realistic testable models of specific psychobiologicalypotheses regarding brain systems regulating basic emotions ofear (HA), anger (NS) and affiliation/disgust (RD). Use of the TPQnd later the Temperament and Character Inventory (TCI) has stim-lated extensive work on brain function [7,8].
Early research indicates that individual variations of each of theimensions of personality as defined by the TPQ are largely genet-
cally determined [8,9,11]. Personality dimensions, therefore, areaid to be heritable, stable across time and there also is some indi-ation that they might be modulated by normal variance in theevel of functioning of the different neurotransmitter systems, espe-ially by variations in the expression of the central monoamineystems. Individual differences in novelty seeking were found asso-iated with variability in the level of activity of the dopaminergicystem; individual differences in harm avoidance appeared linkedo variability in the level of activity of the serotoninergic system;nd individual differences in reward dependence seemed linkedo variability in the level of activity of the noradrenergic system8–10,18,27]. Moreover, harm avoidance and reward dependenceave been found to be associated with polymorphisms of serotoninnd dopamine related genes such as the serotonin transporter-inked promoter region (5-HTTLPR) and the dopamine transporterDAT1) gene [27].
Indirect support for a link between variance in neurotransmit-er systems and personality traits can be found in evidence fromeuropsychiatric disorders. A high proportion of individuals whoevelop neuropsychiatric symptoms such as depression [34], sui-idal behaviour [35], bipolar disorder [31], violent schizophrenia17], drug addiction [12], pathological and social gambling [32] andocial anxiety disorder [26] have personality scores which fall at thextreme ends of the normal distribution for each personality trait.
More recently, Cloninger’s theoretical propositions have foundome empirical support in the evidence of a few neuroimagingtudies which have correlated scores of some of the personalityimensions measured with the TPQ or with other instruments withither regional brain blood flow (rCBF) or brain metabolism. Sig-ificant correlation was found with rCBF values, acquired usingingle photon emission computed tomography (SPECT), in thoseegions which are the neural substrates of the monoaminergicystems [40]. In detail, novelty seeking correlated positively withCBF values in the anterior cingulate and anterior and posteriornsula; harm avoidance correlated negatively with rCBF valuesn the parahippocampal, precentral, postcentral, superior frontal,usiform and inferior temporal gyri; and reward dependencehowed a negative correlation with rCBF values in the parahip-ocampal, anterior cingulate, anterior insula, precentral, superiorrontal, middle frontal, superior temporal gyri and precuneus. A fur-her [18F] fluorodeoxyglucose (FDG) positron emission tomographyPET) study found that NS, HA, RD scores correlated significantlyith cerebral glucose metabolism values in paralimbic and tem-oral regions [46]. The finding of a negative correlation betweenS scores and metabolic activity in the substantia nigra gave sup-ort to the hypothesis that individuals who score highly on theS dimension might have low levels of dopaminergic activity. Theegative correlation between HA scores and metabolism values
n temporo-frontal regions was explained as indicating that highcores on this dimension might reflect low levels of serotoniner-
ic activity, a neurotransmitter system linked to anxiety disorders.inally, the positive correlation between RD scores and metabolicctivity in the orbito-frontal cortex was seen as providing supportor the hypothesis that high RD scores might correspondingly sig-ify high responsiveness of those structures involved with rewardulletin 79 (2009) 265–270
mechanisms. These results have been subsequently replicated byanother [18F] FDG PET study of brain metabolism [22]. In a func-tional magnetic resonance imaging (fMRI) study, Gusnard et al. [20]found that individual differences in persistence were associatedwith differential brain activation levels in specific regions of thelateral orbital and medial prefrontal cortex and of the ventral stria-tum in response to a task in which people were required to makeself referential judgments. With a correlational analysis the authorsdemonstrated that the people with the highest persistence scoreshad increases in activation in orbitomedial prefrontal cortex andventral striatum, while those with the lowest persistence scoreshad decreases in activation in these regions.
There is also some indication that there is a relationship betweenvariability in morphology of specific brain structures and variancein personality trait scores. For example, scores on an anxiety relatedpersonality trait (i.e. harm avoidance) were found to negatively cor-relate with variance in regional grey matter volume (obtained bythree-dimensional MRI acquisition) in a large sample of healthyadults [45]. This study also found that only in females there wasa negative correlation between HA scores and grey matter volumein the anterior prefrontal cortex, supporting the hypothesis thatindividual differences in personality traits might be biologicallydetermined and associated with volumetric variability in specificregions of the brain. This study, however, did not extend its inves-tigation to other personality traits (NS, RD, PER).
Taken together the above evidence suggests that even if theneurobiological substrates of each specific personality trait are notclearly defined, it is possible to find meaningful and replicable asso-ciations between regional brain functional/structural variance andvariance in personality. The available findings seem to suggest thatthe observed variance in the normal range of expression of person-ality traits might be linked to neurodevelopmental variability inspecific brain structures. The present prospective study, therefore,was designed to investigate in a large sample of young adults andusing the technique of voxel-based correlation, whether individ-ual differences in the four main personality dimensions measuredby the TPQ (e.g. novelty seeking, harm avoidance, reward depen-dence and persistence) reflect variance in regional grey mattervolume.
2. Methods and materials
2.1. Participants
Eighty-five young adult participants (58 males, 27 females, mean age 32.69, SD6.48, mean education 14.15, SD 4.92) took part in this experiment. All participantswere recruited from the general public and from the staff and student populationof the University of Modena and Reggio Emilia in Italy. Consent was obtained fromeach participant prior to the experiment and ethical approval was granted by thelocal Ethics Committee.
2.2. Materials
The Three-dimensional Personality Questionnaire [8] was used. The TPQ is aself-report questionnaire which includes 100 items. Participants respond with trueor false to each item. No time constraints are given to complete the test. The cur-rent version of the TPQ measures four main dimensions of temperament: noveltyseeking, harm avoidance, reward dependence and persistence. NS and HA includefour subscales, RD includes three subscales and PER, once considered a subscale ofRD, is now scored independently as a fourth personality dimension in its own right.Individual scores for each of these four dimensions of temperament were used inthe correlational analyses carried out in this study.
Participants were asked to fill in the TPQ and to provide written consent toparticipate in a Magnetic Resonance Imaging (MRI) scanning session. Four differentscores were computed for each participant, reflective of the degree of expressionof the four main dimensions of personality measureable with the TPQ, i.e. noveltyseeking, harm avoidance, reward dependence and persistence.
arch Bulletin 79 (2009) 265–270 267
2
Pw2fl
fuUwmcuvssaocSwp
tscap
3
3
aT
3
agg
3
a
Fr
visual attention to the periphery of the visual field [13,28,33] andin implicit attention [19]. The cortical structures in BA 6 containpremotor and supplementary motor areas whose primary role is
S. Gardini et al. / Brain Rese
.4. Structural MRI scanning: acquisition and statistical analysis
Three-dimensional T1-weighted MRI images were acquired on the same 3Thilips Intera MRI system with a Turbo Field Echo sequence. Voxel dimensionsere 1 mm × 1 mm × 1 mm and field of view was 256 mm with a matrix size of56 × 256 × 124. Total acquisition time was 4 min and 41 s (TR 9.9 ms, TE 4.6 ms andip angle 8◦).
A number of preprocessing steps were carried out to isolate the grey matterrom the 3D T1-weighted structural scans before performing the statistical analysissing SPM5 (Wellcome Trust Centre for Neuroimaging, University College London,K). In detail, to correct for global differences in brain shape, structural images werearped to standard stereotactic space and segmented to extract grey matter, whiteatter and cerebrospinal fluid. The grey matter segments were then modulated to
orrect for changes in volume induced by non-linear normalization and smoothedsing a Gaussian filter set at 12 mm to reduce possible error from between-subjectariability in local anatomy and render the data more normally distributed. Finally,moothed grey matter segments were entered into a voxel-based multiple regres-ion analysis to investigate linear correlations between grey matter concentrationnd the personality dimensions indices obtained on the TPQ. Age, number of yearsf education and gender were also included in the model as covariates. Statisti-al significance level was set at false discovery rate (FDR) corrected p < 0.05 usingPM5. This method of correction for multiple comparisons was preferred to family-ise error (FWE) correction to avoid false rejection of true positives as well as falseositives.
Since scores on the RD and PER dimensions were correlated, each personalityrait was entered in a separate regression model for analysis. Four multiple regres-ion analysis models were carried out. Age, education and gender were entered asonfounds in each model and personality scores for NS, HA, RD and PER in turns the variable of interest. Positive and negative correlations between individualersonality trait scores and grey matter volume values were computed.
. Results
.1. Novelty seeking (NS)
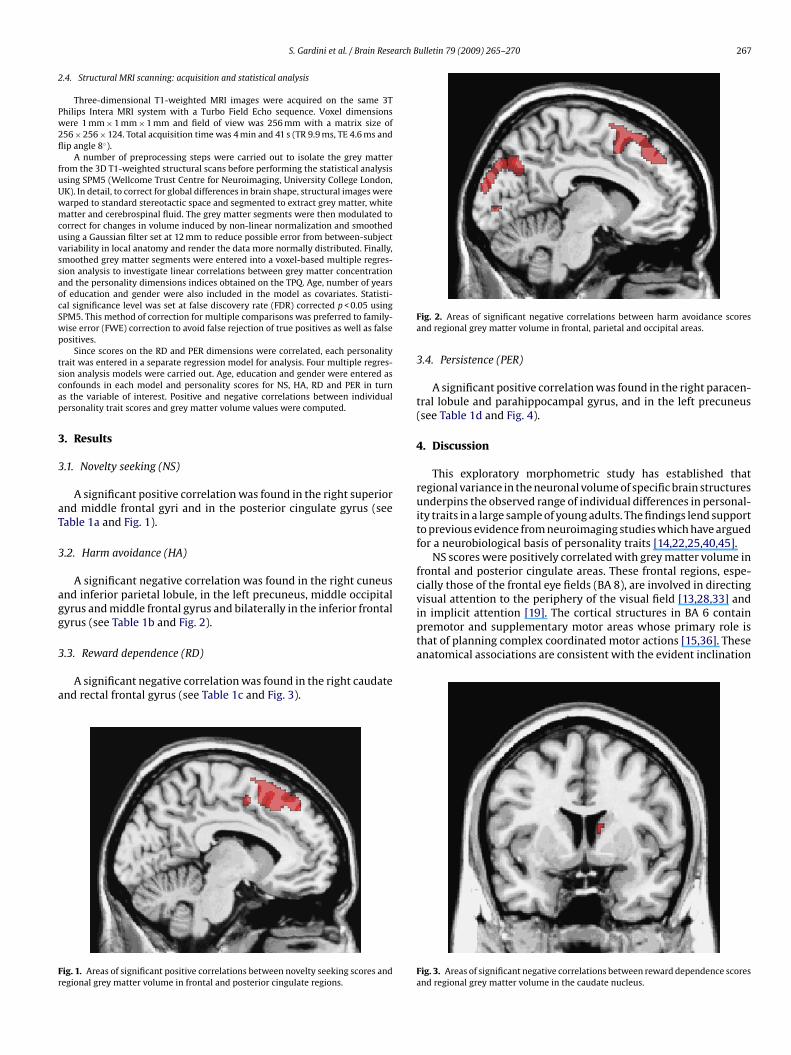
A significant positive correlation was found in the right superiornd middle frontal gyri and in the posterior cingulate gyrus (seeable 1a and Fig. 1).
.2. Harm avoidance (HA)
A significant negative correlation was found in the right cuneusnd inferior parietal lobule, in the left precuneus, middle occipitalyrus and middle frontal gyrus and bilaterally in the inferior frontalyrus (see Table 1b and Fig. 2).
.3. Reward dependence (RD)
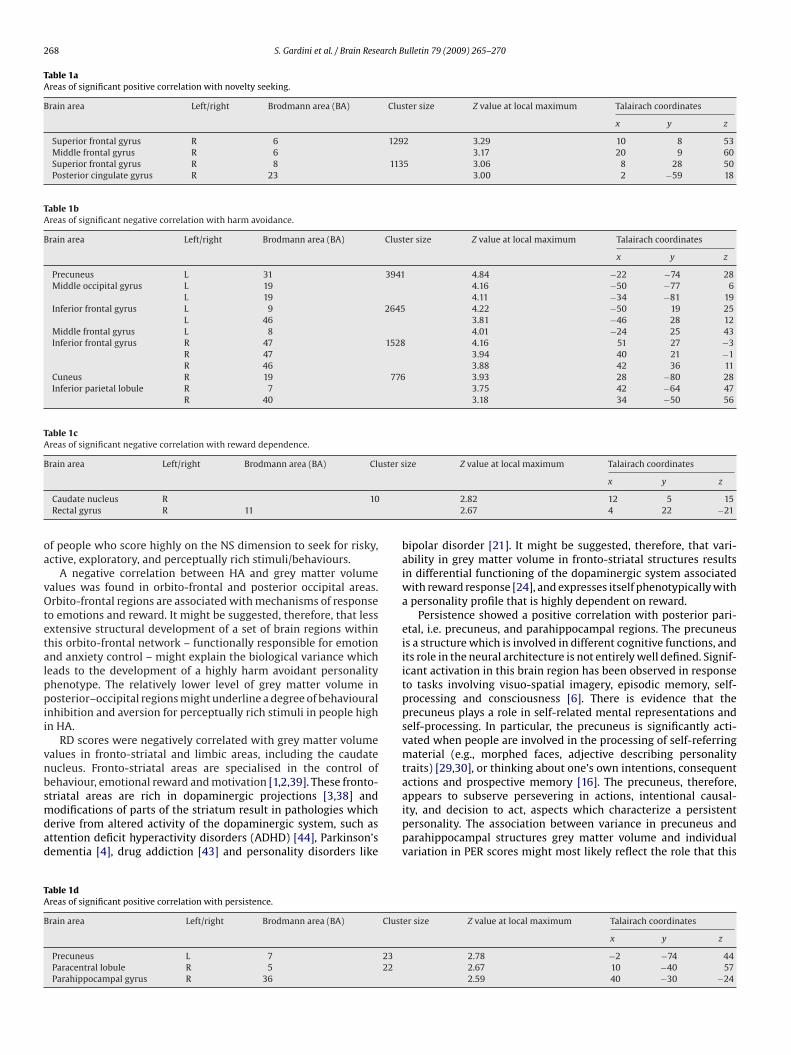
A significant negative correlation was found in the right caudatend rectal frontal gyrus (see Table 1c and Fig. 3).
ig. 1. Areas of significant positive correlations between novelty seeking scores andegional grey matter volume in frontal and posterior cingulate regions.
Fig. 2. Areas of significant negative correlations between harm avoidance scoresand regional grey matter volume in frontal, parietal and occipital areas.
3.4. Persistence (PER)
A significant positive correlation was found in the right paracen-tral lobule and parahippocampal gyrus, and in the left precuneus(see Table 1d and Fig. 4).
4. Discussion
This exploratory morphometric study has established thatregional variance in the neuronal volume of specific brain structuresunderpins the observed range of individual differences in personal-ity traits in a large sample of young adults. The findings lend supportto previous evidence from neuroimaging studies which have arguedfor a neurobiological basis of personality traits [14,22,25,40,45].
NS scores were positively correlated with grey matter volume infrontal and posterior cingulate areas. These frontal regions, espe-cially those of the frontal eye fields (BA 8), are involved in directing
that of planning complex coordinated motor actions [15,36]. Theseanatomical associations are consistent with the evident inclination
Fig. 3. Areas of significant negative correlations between reward dependence scoresand regional grey matter volume in the caudate nucleus.
268 S. Gardini et al. / Brain Research Bulletin 79 (2009) 265–270
Table 1aAreas of significant positive correlation with novelty seeking.
Brain area Left/right Brodmann area (BA) Cluster size Z value at local maximum Talairach coordinates
x y z
Superior frontal gyrus R 6 1292 3.29 10 8 53Middle frontal gyrus R 6 3.17 20 9 60Superior frontal gyrus R 8 1135 3.06 8 28 50Posterior cingulate gyrus R 23 3.00 2 −59 18
Table 1bAreas of significant negative correlation with harm avoidance.
Brain area Left/right Brodmann area (BA) Cluster size Z value at local maximum Talairach coordinates
x y z
Precuneus L 31 3941 4.84 −22 −74 28Middle occipital gyrus L 19 4.16 −50 −77 6
L 19 4.11 −34 −81 19Inferior frontal gyrus L 9 2645 4.22 −50 19 25
L 46 3.81 −46 28 12Middle frontal gyrus L 8 4.01 −24 25 43Inferior frontal gyrus R 47 1528 4.16 51 27 −3
R 47 3.94 40 21 −1R 46 3.88 42 36 11
Cuneus R 19 776 3.93 28 −80 28Inferior parietal lobule R 7 3.75 42 −64 47
R 40 3.18 34 −50 56
Table 1cAreas of significant negative correlation with reward dependence.
Brain area Left/right Brodmann area (BA) Cluster size Z value at local maximum Talairach coordinates
oa
vOtetalppii
vnbsmdad
TA
B
Caudate nucleus R 10Rectal gyrus R 11
f people who score highly on the NS dimension to seek for risky,ctive, exploratory, and perceptually rich stimuli/behaviours.
A negative correlation between HA and grey matter volumealues was found in orbito-frontal and posterior occipital areas.rbito-frontal regions are associated with mechanisms of response
o emotions and reward. It might be suggested, therefore, that lessxtensive structural development of a set of brain regions withinhis orbito-frontal network – functionally responsible for emotionnd anxiety control – might explain the biological variance whicheads to the development of a highly harm avoidant personalityhenotype. The relatively lower level of grey matter volume inosterior–occipital regions might underline a degree of behavioural
nhibition and aversion for perceptually rich stimuli in people highn HA.
RD scores were negatively correlated with grey matter volumealues in fronto-striatal and limbic areas, including the caudateucleus. Fronto-striatal areas are specialised in the control ofehaviour, emotional reward and motivation [1,2,39]. These fronto-triatal areas are rich in dopaminergic projections [3,38] and
odifications of parts of the striatum result in pathologies whicherive from altered activity of the dopaminergic system, such asttention deficit hyperactivity disorders (ADHD) [44], Parkinson’sementia [4], drug addiction [43] and personality disorders like
able 1dreas of significant positive correlation with persistence.
rain area Left/right Brodmann area (BA) Clust
Precuneus L 7 23Paracentral lobule R 5 22Parahippocampal gyrus R 36
x y z
2.82 12 5 152.67 4 22 −21
bipolar disorder [21]. It might be suggested, therefore, that vari-ability in grey matter volume in fronto-striatal structures resultsin differential functioning of the dopaminergic system associatedwith reward response [24], and expresses itself phenotypically witha personality profile that is highly dependent on reward.
Persistence showed a positive correlation with posterior pari-etal, i.e. precuneus, and parahippocampal regions. The precuneusis a structure which is involved in different cognitive functions, andits role in the neural architecture is not entirely well defined. Signif-icant activation in this brain region has been observed in responseto tasks involving visuo-spatial imagery, episodic memory, self-processing and consciousness [6]. There is evidence that theprecuneus plays a role in self-related mental representations andself-processing. In particular, the precuneus is significantly acti-vated when people are involved in the processing of self-referringmaterial (e.g., morphed faces, adjective describing personalitytraits) [29,30], or thinking about one’s own intentions, consequentactions and prospective memory [16]. The precuneus, therefore,appears to subserve persevering in actions, intentional causal-
ity, and decision to act, aspects which characterize a persistentpersonality. The association between variance in precuneus andparahippocampal structures grey matter volume and individualvariation in PER scores might most likely reflect the role that thiser size Z value at local maximum Talairach coordinates
x y z
2.78 −2 −74 442.67 10 −40 572.59 40 −30 −24

S. Gardini et al. / Brain Research B
Fr
stl
msimwmcicirMabspdpt
mtsmdbbuuio[
ocCmfarmi
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ig. 4. Areas of significant positive correlation between persistence scores andegional grey matter volume in the precuneus.
tructure plays in the process of self-representative memory whichakes place during the normal behaviour of waiting for reward andearning.
The positive and negative associations between regional greyatter volume variance and individual NS, HA, RD and PER
cores in this study were found in young adults. An alternativenterpretation of the present findings might be that any observed
orphological brain variability, rather than a predisposing factorhich determines individual differences in personality traits,ight be a byproduct of differences in personality traits. In this
ase a specific neuro-morphological characteristic would be result-ng from repeated behaviours and environmental exposures in theourse of life. This possible alternative interpretation, however,s not fully supported by developmental behavioural geneticsesearch of personality traits (see for example [23,37]. LongitudinalRI studies from childhood to adulthood would be needed to
ddress the nature versus nurture influence on the relationshipetween regional brain morphology and personality trait expres-ion. If taken together with other findings in this field, it is morelausible that the present evidence indicates that genotypicallyefined variability in regional brain morphology expresses itselfhenotypically as individual differences in personality traits, buthis evidence is however, far from conclusive.
The strength of these results is potentially influenced by theethod that was used. Criticisms of the voxel-based morphome-
ry technique include concerns that segmentation inaccuracy mayurvive the automated processes of extraction of grey matter. Theethod, however, can be used to obtain a correlation between
ifferences in grey matter density/volume values and continuousehavioural data, by treating each individual voxel value in therain as a continuous variable. The method appears reliable whensed to identify the neural substrate of specific cognitive functionssing a correlational approach and has been validated by establish-
ng sound associations between brain regions and specific aspectsf cognitive performance in healthy [41] and pathological samples5,42,47].
In summary, the present results support and extend the findingsf previous research [40,45,46]. The earlier findings that an asso-iation exists between personality dimensions as defined by theloninger model, neurotransmitters’ function and regional brainetabolism are extended to suggest that the range of individual dif-
erences in novelty seeking, harm avoidance, reward dependencend persistence observed in the normal population reflects theange of structural variance observed in the normal brain regionalorphology which then manifests itself behaviourally as inter-
ndividual differences in personality traits.
[
ulletin 79 (2009) 265–270 269
References
[1] A.K. Afifi, The basal ganglia: a neural network with more than motor function,Semin. Pediatr. Neurol. 10 (2003) 3–10.
[2] A. Aron, H. Fisher, D.J. Mashek, G. Strong, H. Li, L.L. Brown, Reward, motiva-tion, and emotion systems associated with early-stage intense romantic love, J.Neurophysiol. 94 (2005) 327–337.
[3] V. Bragulat, M.L. Paillere-Martinot, E. Artiges, V. Frouin, J.B. Poline, J.L. Mar-tinot, Dopaminergic function in depressed patients with affective flattening orwith impulsivity: [18F]fluoro-L-dopa positron emission tomography study withvoxel-based analysis, Psychiatry Res. 154 (2007) 115–124.
[4] E. Broussolle, C. Dentresangle, P. Landais, L. Garcia-Larrea, P. Pollak, B. Croisile,O. Hibert, F. Bonnefoi, G. Galy, J.C. Froment, D. Comar, The relation of putamenand caudate nucleus 18F-Dopa uptake to motor and cognitive performances inParkinson’s disease, J. Neurol. Sci. 166 (1999) 141–151.
[5] P.D. Bruen, W.J. McGeown, M.F. Shanks, A. Venneri, Neuroanatomical corre-lates of neuropsychiatric symptoms in Alzheimer’s disease, Brain 131 (2008)2455–2463.
[6] A.E. Cavanna, M.R. Trimble, The precuneus: a review of its functional anatomyand behavioural correlates, Brain 129 (2006) 564–583.
[7] C.R. Cloninger, Feeling Good: The Science of Well Being, Oxford University Press,New York, 2004.
[8] C.R. Cloninger, A systematic method for clinical description and classificationof personality variants. A proposal, Arch. Gen. Psychiatry 44 (1987) 573–588.
[9] C.R. Cloninger, A unified biosocial theory of personality and its role in the devel-opment of anxiety states, Psychiatr. Dev. 4 (1986) 167–226.
10] C.R. Cloninger, D.M. Svrakic, T.R. Przybeck, A psychobiological model of temper-ament and character, Arch. Gen. Psychiatry 50 (1993) 975–990.
11] D.E. Comings, R. Gade-Andavolu, N. Gonzalez, S. Wu, D. Muhleman, H. Blake,M.B. Mann, G. Dietz, G. Saucier, J.P. MacMurray, A multivariate analysis of 59candidate genes in personality traits: the temperament and character inventory,Clin. Genet. 58 (2000) 375–385.
12] K.P. Conway, R.J. Kane, S.A. Ball, J.C. Poling, B.J. Rounsaville, Personality, sub-stance of choice, and polysubstance involvement among substance dependentpatients, Drug Alcohol Depend. 71 (2003) 65–75.
13] J. de Fockert, G. Rees, C. Frith, N. Lavie, Neural correlates of attentional capturein visual search, J. Cogn. Neurosci. 16 (2004) 751–759.
14] T. Deckersbach, K.K. Miller, A. Klibanski, A. Fischman, D.D. Dougherty, M.A.Blais, D.B. Herzog, S.L. Rauch, Regional cerebral brain metabolism correlatesof neuroticism and extraversion, Depress. Anxiety 23 (2006) 133–138.
15] M.P. Deiber, R.E. Passingham, J.G. Colebatch, K.J. Friston, P.D. Nixon, R.S. Frack-owiak, Cortical areas and the selection of movement: a study with positronemission tomography, Exp. Brain Res. 84 (1991) 393–402.
16] H.E. den Ouden, U. Frith, C. Frith, S.J. Blakemore, Thinking about intentions,Neuroimage 28 (2005) 787–796.
17] A. Fresan, R. Apiquian, H. Nicolini, J.J. Cervantes, Temperament and character inviolent schizophrenic patients, Schizophr. Res. 94 (2007) 74–80.
18] G. Gerra, A. Zaimovic, M. Timpano, U. Zambelli, M. Begarani, G.F. Marzocchi, M.Ferri, R. Delsignore, F. Brambilla, Neuroendocrine correlates of temperamenttraits in abstinent opiate addicts, J. Subst. Abuse 11 (2000) 337–354.
19] M.H. Grosbras, T. Paus, Transcranial magnetic stimulation of the human frontaleye field: effects on visual perception and attention, J. Cogn. Neurosci. 14 (2002)1109–1120.
20] D.A. Gusnard, J.M. Ollinger, G.L. Shulman, C.R. Cloninger, J.L. Price, D.C. VanEssen, M.E. Raichle, Persistence and brain circuitry, Proc. Natl. Acad. Sci. U.S.A.100 (2003) 3479–3484.
21] T. Hajek, N. Carrey, M. Alda, Neuroanatomical abnormalities as risk factors forbipolar disorder, Bipolar Disord. 7 (2005) 393–403.
22] Y. Hakamata, M. Iwase, H. Iwata, T. Kobayashi, T. Tamaki, M. Nishio, K. Kawa-hara, H. Matsuda, N. Ozaki, S. Honjo, T. Inada, Regional brain cerebral glucosemetabolism and temperament: a positron emission tomography study, Neu-rosci. Lett. 396 (2006) 33–37.
23] N. Heiman, M.C. Stallings, S.E. Young, J.K. Hewitt, Investigating the genetic andenvironmental structure of Cloninger’s personality dimensions in adolescence,Twin Res. 7 (2004) 462–470.
24] S.D. Iversen, L.L. Iversen, Dopamine: 50 years in perspective, Trends Neurosci.30 (2007) 188–193.
25] D.L. Johnson, J.S. Wiebe, S.M. Gold, N.C. Andreasen, R.D. Hichwa, G.L. Watkins,L.L. Boles Ponto, Cerebral blood flow and personality: a positron emissiontomography study, Am. J. Psychiatry 156 (1999) 252–257.
26] T.B. Kashdan, S.G. Hofmann, The high-novelty-seeking, impulsive subtype ofgeneralized social anxiety disorder, Depress. Anxiety 25 (2008) 535–541.
27] S.J. Kim, Y.S. Kim, H.S. Lee, S.Y. Kim, C.H. Kim, An interaction between the sero-tonin transporter promoter region and dopamine transporter polymorphismscontributes to harm avoidance and reward dependence traits in normal healthysubjects, J. Neural Transm. 113 (2006) 877–886.
28] J.M. Kincade, R.A. Abrams, S.V. Astafiev, G.L. Shulman, M. Corbetta, Anevent-related functional magnetic resonance imaging study of voluntary andstimulus-driven orienting of attention, J. Neurosci. 25 (2005) 4593–4604.
29] T.T. Kircher, M. Brammer, E. Bullmore, A. Simmons, M. Bartels, A.S. David, The
neural correlates of intentional and incidental self processing, Neuropsycholo-gia 40 (2002) 683–692.30] T.T. Kircher, C. Senior, M.L. Phillips, P.J. Benson, E.T. Bullmore, M. Brammer,A. Simmons, S.C. Williams, M. Bartels, A.S. David, Towards a functional neu-roanatomy of self processing: effects of faces and words, Brain Res. Cogn. BrainRes. 10 (2000) 133–144.

2 arch B
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
70 S. Gardini et al. / Brain Rese
31] S.T. Loftus, J.L. Garno, J. Jaeger, A.K. Malhotra, Temperament and characterdimensions in bipolar I disorder: a comparison to healthy controls, J. Psychiatr.Res. 42 (2008) 1131–1136.
32] G. Martinotti, S. Andreoli, E. Giametta, V. Poli, P. Bria, L. Janiri, The dimen-sional assessment of personality in pathologic and social gamblers: therole of novelty seeking and self-transcendence, Compr. Psychiatry 47 (2006)350–356.
33] A. Miki, T. Nakajima, S. Miyauchi, M. Takagi, H. Abe, Functional magnetic reso-nance imaging of the frontal eye fields during saccadic eye movements, NipponGanka Gakkai Zasshi 100 (1996) 541–545.
34] Y. Ono, J. Ando, N. Onoda, K. Yoshimura, T. Momose, M. Hirano, S. Kanba, Dimen-sions of temperament as vulnerability factors in depression, Mol. Psychiatry 7(2002) 948–953.
35] M. Pompili, Z. Rihmer, H.S. Akiskal, M. Innamorati, P. Iliceto, K.K. Akiskal, D.Lester, V. Narciso, S. Ferracuti, R. Tatarelli, E. De Pisa, P. Girardi, Temperamentand personality dimensions in suicidal and nonsuicidal psychiatric inpatients,Psychopathology 41 (2008) 313–321.
36] P.E. Roland, B. Larsen, N.A. Lassen, E. Skinhoj, Supplementary motor area andother cortical areas in organization of voluntary movements in man, J. Neuro-physiol. 43 (1980) 118–136.
37] K.J. Saudino, Behavioral genetics and child temperament, J. Dev. Behav. Pediatr.
26 (2005) 214–223.38] D.J. Scott, M.M. Heitzeg, R.A. Koeppe, C.S. Stohler, J.K. Zubieta, Variations in thehuman pain stress experience mediated by ventral and dorsal basal gangliadopamine activity, J. Neurosci. 26 (2006) 10789–10795.
39] R. Sinha, C. Lacadie, P. Skudlarski, B.E. Wexler, Neural circuits underlying emo-tional distress in humans, Ann. N Y Acad. Sci. 1032 (2004) 254–257.
[
ulletin 79 (2009) 265–270
40] M. Sugiura, R. Kawashima, M. Nakagawa, K. Okada, T. Sato, R. Goto, K. Sato, S.Ono, T. Schormann, K. Zilles, H. Fukuda, Correlation between human personalityand neural activity in cerebral cortex, Neuroimage 11 (2000) 541–546.
41] L.K. Tyler, W. Marslen-Wilson, E.A. Stamatakis, Dissociating neuro-cognitivecomponent processes: voxel-based correlational methodology, Neuropsycholo-gia 43 (2005) 771–778.
42] A. Venneri, W.J. McGeown, H.M. Hietanen, C. Guerrini, A.W. Ellis, M.F. Shanks,The anatomical bases of semantic retrieval deficits in early Alzheimer’s disease,Neuropsychologia 46 (2008) 497–510.
43] N.D. Volkow, G.J. Wang, J.S. Fowler, J. Logan, M. Jayne, D. Franceschi, C. Wong,S.J. Gatley, A.N. Gifford, Y.S. Ding, N. Pappas, “Nonhedonic” food motivation inhumans involves dopamine in the dorsal striatum and methylphenidate ampli-fies this effect, Synapse 44 (2002) 175–180.
44] T.M. Wellington, M. Semrud-Clikeman, A.L. Gregory, J.M. Murphy, J.L. Lan-caster, Magnetic resonance imaging volumetric analysis of the putamen inchildren with ADHD: combined type versus control, J. Atten. Disord. 10 (2006)171–180.
45] H. Yamasue, O. Abe, M. Suga, H. Yamada, H. Inoue, M. Tochigi, M. Rogers, S.Aoki, N. Kato, K. Kasai, Gender-common and -specific neuroanatomical basis ofhuman anxiety-related personality traits, Cereb. Cortex 18 (2008) 46–52.
46] T. Youn, I.K. Lyoo, J.K. Kim, H.J. Park, K.S. Ha, D.S. Lee, K.Y. Abrams, M.C. Lee,
J.S. Kwon, Relationship between personality trait and regional cerebral glu-cose metabolism assessed with positron emission tomography, Biol. Psychol.60 (2002) 109–120.47] G. Zamboni, E.D. Huey, F. Krueger, P.F. Nichelli, J. Grafman, Apathy and dis-inhibition in frontotemporal dementia: insights into their neural correlates,Neurology 71 (2008) 736–742.
Copyright © 2022 FDOKUMEN




















