
In vitro interactions between several species of harmful algae and haemocytes of bivalve molluscs
23
After online publication, subscribers (personal/institutional) to this journal will have access to the complete article via the DOI using the URL: If you would like to know when your article has been published online, take advantage of our free alert service. For registration and further information, go to: http://www.springerlink.com. Due to the electronic nature of the procedure, the manuscript and the original figures will only be returned to you on special request. When you return your corrections, please inform us, if you would like to have these documents returned. Dear Author Here are the proofs of your article. • You can submit your corrections online, via e-mail or by fax. • For online submission please insert your corrections in the online correction form. Always indicate the line number to which the correction refers. • You can also insert your corrections in the proof PDF and email the annotated PDF. • For fax submission, please ensure that your corrections are clearly legible. Use a fine black pen and write the correction in the margin, not too close to the edge of the page. • Remember to note the journal title, article number, and your name when sending your response via e-mail or fax. • Check the metadata sheet to make sure that the header information, especially author names and the corresponding affiliations are correctly shown. • Check the questions that may have arisen during copy editing and insert your answers/corrections. • Check that the text is complete and that all figures, tables and their legends are included. Also check the accuracy of special characters, equations, and electronic supplementary material if applicable. If necessary refer to the Edited manuscript. • The publication of inaccurate data such as dosages and units can have serious consequences. Please take particular care that all such details are correct. • Please do not make changes that involve only matters of style. We have generally introduced forms that follow the journal’s style. • Substantial changes in content, e.g., new results, corrected values, title and authorship are not allowed without the approval of the responsible editor. In such a case, please contact the Editorial Office and return his/her consent together with the proof. • If we do not receive your corrections within 48 hours, we will send you a reminder. • Your article will be published Online First approximately one week after receipt of your corrected proofs. This is the official first publication citable with the DOI. Further changes are, therefore, not possible. • The printed version will follow in a forthcoming issue. Please note http://dx.doi.org/10.1007/s10565-011-9186-6
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of In vitro interactions between several species of harmful algae and haemocytes of bivalve molluscs

After online publication, subscribers (personal/institutional) to this journal will haveaccess to the complete article via the DOI using the URL:
If you would like to know when your article has been published online, take advantageof our free alert service. For registration and further information, go to:http://www.springerlink.com.
Due to the electronic nature of the procedure, the manuscript and the original figureswill only be returned to you on special request. When you return your corrections,please inform us, if you would like to have these documents returned.
Dear Author
Here are the proofs of your article.
• You can submit your corrections online, via e-mail or by fax.
• For online submission please insert your corrections in the online correction form.
Always indicate the line number to which the correction refers.
• You can also insert your corrections in the proof PDF and email the annotated PDF.
• For fax submission, please ensure that your corrections are clearly legible. Use a fine
black pen and write the correction in the margin, not too close to the edge of the page.
• Remember to note the journal title, article number, and your name when sending your
response via e-mail or fax.
• Check the metadata sheet to make sure that the header information, especially author
names and the corresponding affiliations are correctly shown.
• Check the questions that may have arisen during copy editing and insert your
answers/corrections.
• Check that the text is complete and that all figures, tables and their legends are included.
Also check the accuracy of special characters, equations, and electronic supplementary
material if applicable. If necessary refer to the Edited manuscript.
• The publication of inaccurate data such as dosages and units can have serious
consequences. Please take particular care that all such details are correct.
• Please do not make changes that involve only matters of style. We have generally
introduced forms that follow the journal’s style.
• Substantial changes in content, e.g., new results, corrected values, title and authorship are
not allowed without the approval of the responsible editor. In such a case, please contact
the Editorial Office and return his/her consent together with the proof.
• If we do not receive your corrections within 48 hours, we will send you a reminder.
• Your article will be published Online First approximately one week after receipt of your
corrected proofs. This is the official first publication citable with the DOI. Further
changes are, therefore, not possible.
• The printed version will follow in a forthcoming issue.
Please note
http://dx.doi.org/10.1007/s10565-011-9186-6

AUTHOR'S PROOF
_____________________________________________________________________________________
Please note: Image will appear in color online but will be printed in black and white._____________________________________________________________________________________
Metadata of the article that will be visualized in OnlineFirst
1 Article Title In vitro interactions between several species of ha rmful algae and haemocytes of bivalve molluscs
2 Article Sub- Title
3 Article Copyright - Year
Springer Science+Business Media B.V. 2011 (This will be the copyright line in the final PDF)
4 Journal Name Cell Biology and Toxicology
5
Corresponding Author
Family Name Hégaret
6 Particle
7 Given Name Hélène
8 Suffix
9 Organization Université de Bretagne Occidentale—IUEM, LEMAR CNRS UMR 6539
10 Division
11 Address Place Nicolas Copernic, Technopôle Brest Iroise, Plouzané 29280, France
12 Organization University of Connecticut
13 Division Department of Marine Sciences
14 Address 1080 Shennecossett Road, Groton 06340, CT, USA
15 Organization Université de Bretagne Occidentale—IUEM, LEMAR CNRS UMR 6539
16 Division
17 Address Place Nicolas Copernic, Technopôle Brest Iroise, Plouzané 29280, France
18 e-mail [email protected]
19
Author
Family Name Silva
20 Particle da
21 Given Name Patricia Mirella
22 Suffix
23 Organization Université de Bretagne Occidentale—IUEM, LEMAR CNRS UMR 6539
24 Division
25 Address Place Nicolas Copernic, Technopôle Brest Iroise, Plouzané 29280, France
26 Organization Universidade Federal de Sergipe
27 Division Centro de Ciências Biológicas e da Saúde, Núcleo de Engenharia de Pesca
28 Address Cidade Universitária Prof. José Aloísio de Campos, Rua Mal. Rondon s/n, São Cristóvão CEP 49100-000, Sergipe, Brazil
29 Organization Universidade Federal de Sergipe
Page 1 of 3Springer Metadata to PDF File
2/9/2011file://C:\WMS\Springer\Metadata2PDF\temp\CBT19186.htm

AUTHOR'S PROOF
30 Division Centro de Ciências Biológicas e da Saúde, Núcleo de Engenharia de Pesca
31 Address Cidade Universitária Prof. José Aloísio de Campos, Rua Mal. Rondon s/n, São Cristóvão CEP 49100-000, Sergipe, Brazil
32 e-mail
33
Author
Family Name Wikfors
34 Particle
35 Given Name Gary H.
36 Suffix
37 Organization NOAA
38 Division Northeast Fisheries Science Center, National Marine Fisheries Service
39 Address 212 Rogers Avenue, Milford 06460, CT, USA
40 e-mail
41
Author
Family Name Haberkorn
42 Particle
43 Given Name Hansy
44 Suffix
45 Organization Université de Bretagne Occidentale—IUEM, LEMAR CNRS UMR 6539
46 Division
47 Address Place Nicolas Copernic, Technopôle Brest Iroise, Plouzané 29280, France
48 e-mail
49
Author
Family Name Shumway
50 Particle
51 Given Name Sandra E.
52 Suffix
53 Organization University of Connecticut
54 Division Department of Marine Sciences
55 Address 1080 Shennecossett Road, Groton 06340, CT, USA
56 e-mail
57
Author
Family Name Soudant
58 Particle
59 Given Name Philippe
60 Suffix
61 Organization Université de Bretagne Occidentale—IUEM, LEMAR CNRS UMR 6539
62 Division
63 Address Place Nicolas Copernic, Technopôle Brest Iroise, Plouzané 29280, France
64 e-mail
65 Received
Page 2 of 3Springer Metadata to PDF File
2/9/2011file://C:\WMS\Springer\Metadata2PDF\temp\CBT19186.htm

AUTHOR'S PROOF
Schedule
21 June 2010
66 Revised
67 Accepted 2 February 2011
68 Abstract Harmful algal blooms (HABs) can have both lethal and sublethal impacts on shellfish. To understand the possible roles of haemocytes in bivalve immune responses to HABs and how the algae are affected by these cells (haemocytes), in vitro tests between cultured harmful algal species and haemocytes of the northern quahog (= hard clam) Mercenaria mercenaria, the soft-shell clam Mya arenaria, the eastern and pacific oysters Crassostrea virginica and Crassostrea gigas and the Manila clam Ruditapes philippinarum were carried out. Within their respective ranges of distribution, these shellfish species can experience blooms of several HAB species, including Prorocentrum minimum, Heterosigma akashiwo, Alexandrium fundyense, Alexandrium minutum and Karenia spp.; thus, these algal species were chosen for testing. Possible differences in haemocyte variables attributable to harmful algae and also effects of haemolymph and haemocytes on the algae themselves were measured. Using microscopic and flow cytometric observations, changes were measured in haemocytes, including cell morphology, mortality, phagocytosis, adhesion and reactive oxygen species (ROS) production, as well as changes in the physiology and the characteristics of the algal cells, including mortality, size, internal complexity and chlorophyll fluorescence. These experiments suggest different effects of the several species of harmful algae upon bivalve haemocytes. Some harmful algae act as immunostimulants, whereas others are immunosuppressive. P. minimum appears to activate haemocytes, but the other harmful algal species tested seem to cause a suppression of immune functions, generally consisting of decreases in phagocytosis, production of ROS and cell adhesion and besides cause an increase in the percentage of dead haemocytes, which could be attributable to the action of chemical toxins. Microalgal cells exposed to shellfish haemolymph generally showed evidence of algal degradation, e.g. loss of chlorophyll fluorescence and modification of cell shape. Thus, in vitro tests allow a better understanding of the role of the haemocytes and the haemolymph in the defence mechanisms protecting molluscan shellfish from harmful algal cells and could also be further developed to estimate the effects of HABs on bivalve molluscs in vivo.
69 Keywords separated by ' - '
Bivalve - Haemocyte - Harmful algal - HAB - Immune response - Phagocytosis
70 Foot note information
Page 3 of 3Springer Metadata to PDF File
2/9/2011file://C:\WMS\Springer\Metadata2PDF\temp\CBT19186.htm

AUTHOR'S PROOF
UNCORRECTEDPROOF
1
23
4 In vitro interactions between several species of harmful algae5 and haemocytes of bivalve molluscs
6 Hélène Hégaret & Patricia Mirella da Silva &
7 Gary H. Wikfors & Hansy Haberkorn &
8 Sandra E. Shumway & Philippe Soudant
9 Received: 21 June 2010 /Accepted: 2 February 201110 # Springer Science+Business Media B.V. 2011
11
12 Abstract Harmful algal blooms (HABs) can have13 both lethal and sublethal impacts on shellfish. To14 understand the possible roles of haemocytes in15 bivalve immune responses to HABs and how the
16algae are affected by these cells (haemocytes), in17vitro tests between cultured harmful algal species18and haemocytes of the northern quahog (= hard19clam) Mercenaria mercenaria, the soft-shell clam20Mya arenaria, the eastern and pacific oysters21Crassostrea virginica and Crassostrea gigas and22the Manila clam Ruditapes philippinarum were23carried out. Within their respective ranges of24distribution, these shellfish species can experience25blooms of several HAB species, including Proro-26centrum minimum, Heterosigma akashiwo, Alexan-27drium fundyense, Alexandrium minutum and28Karenia spp.; thus, these algal species were chosen29for testing. Possible differences in haemocyte30variables attributable to harmful algae and also31effects of haemolymph and haemocytes on the algae32themselves were measured. Using microscopic and33flow cytometric observations, changes were mea-34sured in haemocytes, including cell morphology,35mortality, phagocytosis, adhesion and reactive oxy-36gen species (ROS) production, as well as changes in37the physiology and the characteristics of the algal38cells, including mortality, size, internal complexity39and chlorophyll fluorescence. These experiments40suggest different effects of the several species of41harmful algae upon bivalve haemocytes. Some42harmful algae act as immunostimulants, whereas43others are immunosuppressive. P. minimum appears44to activate haemocytes, but the other harmful algal45species tested seem to cause a suppression of46immune functions, generally consisting of decreases47in phagocytosis, production of ROS and cell
Cell Biol ToxicolDOI 10.1007/s10565-011-9186-6
H. Hégaret : S. E. ShumwayDepartment of MarineQ1 Sciences, University of Connecticut,1080 Shennecossett Road,Groton, CT 06340, USA
P. M. da Silva :H. Haberkorn : P. SoudantUniversité de Bretagne Occidentale—IUEM,LEMAR CNRS UMR 6539,Place Nicolas Copernic, Technopôle Brest Iroise,29280 Plouzané, France
G. H. WikforsNortheast Fisheries Science Center,National Marine Fisheries Service, NOAA,212 Rogers Avenue,Milford, CT 06460, USA
Present Address:H. Hégaret (*)Université de Bretagne Occidentale—IUEM,LEMAR CNRS UMR 6539,Place Nicolas Copernic, Technopôle Brest Iroise,29280 Plouzané, Francee-mail: [email protected]
Present Address:P. M. da SilvaCentro de Ciências Biológicas e da Saúde, Núcleo deEngenharia de Pesca, Universidade Federal de Sergipe,Cidade Universitária Prof. José Aloísio de Campos,Rua Mal. Rondon s/n,CEP 49100-000 São Cristóvão, Sergipe, Brazil
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011

AUTHOR'S PROOF
UNCORRECTEDPROOF
48 adhesion and besides cause an increase in the percentage49 of dead haemocytes, which could be attributable to the50 action of chemical toxins. Microalgal cells exposed to51 shellfish haemolymph generally showed evidence of52 algal degradation, e.g. loss of chlorophyll fluorescence53 and modification of cell shape. Thus, in vitro tests allow54 a better understanding of the role of the haemocytes and55 the haemolymph in the defence mechanisms protecting56 molluscan shellfish from harmful algal cells and could57 also be further developed to estimate the effects of58 HABs on bivalve molluscs in vivo.
59 Keywords Bivalve . Haemocyte . Harmful algal .
60 HAB . Immune response . Phagocytosis
61 Introduction
62 Internal defence in invertebrates, such as bivalve63 molluscs, is mainly based upon a non-adaptive, non-64 specific, innate immune system (Janeway 1994).65 More recently, Rowley and Powell (2007) demon-66 strated the possibility of a specific immune response67 of invertebrates, suggesting the potential existence of68 an acquired immune response. The innate immune69 system, however, remains the more recognized and70 understood among invertebrate groups although het-71 erogeneous and complex (Loker et al. 2004). In72 molluscs, innate immune responses are mediated by73 haemocytes, the main defence cells, and humoural74 factors, such as antimicrobial peptides (Cheng 1996;75 Hine 1999; Bachère et al. 2004), lysozymes, lectins76 and the alternative complement pathway (Medzhitov77 and Janeway 2002). Indeed, as pathogens encounter78 the external protective barrier of the mollusc, the host79 recognizes their specific molecular pattern (pathogen-80 associated molecular patterns) by its pattern recogni-81 tion proteins (Medzhitov and Janeway 2002) and82 initiates haemocyte-mediated responses such as83 phagocytosis and oxidative burst to accomplish84 complete elimination of invading pathogens.85 Numerous publications have demonstrated the86 effects of pathogens, pesticides and other toxic87 pollutants on the immune system of bivalves (Chu88 and Lapeyre 1993a, b; Anderson et al. 1995; Lapeyre89 et al. 1995; Anderson 1996, 1999; Fisher et al. 1999,90 2000; Oliver et al. 2001; Fournier et al. 2001; Sauvé91 et al. 2002; Hamoutene et al. 2004; Villalba et al.92 2004; Gagnaire et al. 2006; Bado-Nilles et al. 2008;
93Morga et al. 2009; Hannam et al. 2010a, b), and94among the environmental agents that may activate or95modulate the immune system of bivalves are harmful96or toxic microalgae (Hégaret and Wikfors 2005a, b;97Hégaret et al. 2007a, b; da Silva et al. 2008; Galimany98et al. 2008a, b; Haberkorn et al. 2010a). Harmful algal99blooms occur routinely in locations where bivalve100molluscs are present and can have profound effects,101including mass mortalities, leading to both economic102and ecological impacts (Shumway 1990; Burkholder1031998; Hoagland et al. 2002; Matsuyama and Shumway1042009). There are many different species of harmful105algae which invoke various toxic or noxious mecha-106nisms. These microalgae can be toxic to shellfish and107also to human consumers of biotoxin-contaminated108shellfish (reviewed in Shumway 1990; Landsberg1092002). As bivalve molluscs are filter feeders, these110harmful algae can contact gill, digestive-epithelial and111other tissues during ingestion, which allows for112interaction with haemocytes found throughout the113bivalve open circulatory system.114Harmful algal cells have been observed in tissues of115bay scallops (Leibovitz et al. 1984; Wikfors and116Smolowitz 1993). Therefore, it may be feasible for117bivalves to identify algal cells as foreign invaders. The118most likely effect of harmful algae upon the bivalve119immune system is through release of biochemical toxins120(i.e. saxitoxins, venerupin, gymnodimine, brevetoxins;121reviewed by Shumway 1990; Landsberg 2002), haemo-122lytic toxins (Jenkinson and Arzul 2000), other extracel-123lular, organic compounds (Twiner et al. 2004, 2005;124Gentien et al. 2007) or reactive oxygen species125(Marshall et al. 2005a, b). Toxins or toxic compounds126associated with HABs are species specific. Accordingly,127assessing the direct interactions between haemocytes128and individual species of harmful algae can provide129insights into the responses of bivalves as they are130exposed to harmful algal blooms.131Experiments reported here document in vitro inter-132actions between haemocytes of (1) the northern quahog,133or hard clam, Mercenaria mercenaria, the soft-shell134clam Mya arenaria and the eastern oyster Crassostrea135virginica interacting with three harmful algal species:136Alexandrium fundyense, Heterosigma akashiwo and137Prorocentrum minimum; (2) the Pacific oyster Cras-138sostrea gigas and the harmful dinoflagellate Alexan-139drium minutum and (3) the Manila clam Ruditapes140philippinarum and the harmful algae Karenia sell-141iformis and Karenia mikimotoi. These species-specific
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
142 combinations were selected as they represent commer-143 cial shellfish species and dominant HAB species in the144 Northeastern United States and western France that can145 interact in the natural environment. These interactions146 were also selected based upon observations of feeding147 behaviour in various HAB–bivalve pairs that revealed148 differences (Hégaret et al. 2007b). An understanding of149 these in vitro interactions, i.e. in the absence of150 confounding physiological and environmental effects151 possible during in vivo exposures, can help to identify152 the specific roles of the haemocytes in defence153 mechanisms when living bivalves are exposed to154 HABs in the environment.
155 Materials and methods
156 Bivalve molluscs
157 Northern quahogs (hard clams, M. mercenaria Lin-158 naeus—shell length (s.l.), 50–60 mm) and soft-shell159 clams (M. arenaria Linnaeus, s.l. 60–80 mm) were160 collected in Milford Harbor, CT, USA. Eastern161 oysters, C. virginica Gmelin (s.l. 50–60 mm), were162 received from Fisher’s Island Oyster Farm, Fisher’s163 Island, NY, USA. Bivalves were acclimated for at164 least 1 week and maintained in 18°C and 33 ppt with165 a continual flow of seawater in the Milford Laboratory166 prior to experiments. Manila clams, R. philippinarum167 Adams and Reeve (s.l. 35–45 cm), were collected the168 Morbihan Golfe in Brittany (NW France). Manila clams169 were acclimated for at least 1 week with a continual170 flow of seawater of 35 ppt at 16°C prior to experiments.171 Pacific oysters, C. gigas (Thunberg 1793; 60–70 mm),172 were collected from bay of Brest (Brittany, France) and173 acclimated for 1 week with continual flow of filtered174 (1 μm) sea water of 35 pptQ2 at 16°C.
175 Algal cultures
176 The algal species to which quahogs, soft-shell clams and177 eastern oysters were exposed were obtained from the178 NOAA, Milford Laboratory (CT, USA) collection: A.179 fundyense Balech (strain BF2, isolated from the Gulf of180 Maine, USA), P. minimum (Pavillard) Schiller (strain181 JA-98-01, isolated from the Choptank River, MD,182 USA) and H. akashiwo (Hada) Hada ex Sournia (strain183 OL, isolated from NJ, USA). The RHODO strain of184 Rhodomonas sp. was used as a non-toxic, control alga.
185Cultures of A. fundyense were grown in F/2-enriched186(Guillard and Ryther 1962; Guillard 1975) Milford187seawater; H. akashiwo and Rhodomonas sp. were188cultured in E-medium (Ukeles 1973) and P. minimum189was grown in EDL7 medium, a modified version of the190enriched seawater E-medium that contains L-1 trace191metals, double the EDTA of the standard E formula-192tion, KNO3 rather than NaNO3 and soil extract. The193algae were maintained at 20°C on a 12:12-h light/dark194cycle and used in log-phase.195The dinoflagellates K. (= Gymnodinium) mikimotoi196(Miyake and Kominami ex Oda) Hansen et Moestrup197(Stock GM95TIN, isolated in 1995 at Tinduff, Rade de198Brest, France) and K. selliformis (= Gymnodinium199maguelonnense, Strain GM94GAB, isolated from Gulf200of Gabès, Tunisia) were obtained from the Dyneco201Department of IFREMER (Brest, France) and grown in202the IUEM laboratory in sterile, 6-L carboys. Medium203used for these cultures was F/2-enriched, seawater204from the Argenton hatchery, filtered to 1 μm and205autoclaved. Cultures of Karenia spp. were maintained206at 18–20°C on a 12:12-h light/dark cycle and used in207log-phase.208Cultures of A. minutum Halim (strain AM89BM,209isolated in 1989 in the bay of Morlaix, Brittany,210France) and Heterocapsa triquetra (Ehrenberg) Stein211(strain HT99PZ, isolated in 1999 in the bay of212Morlaix, Brittany, France) were grown in 1-l batch213culture using autoclaved seawater filtered to 1 μm and214supplemented with L1 medium (Guillard and Hargraves2151993) and maintained at 16±1°C, with a dark/light216cycle of 12:12 h. Cells of A. minutum and H. triquetra217were harvested in exponential growth phase after21812 days of culture.219Algal cell densities were determined by haemocy-220tometer counts under light microscope. For all experi-221ments, algal densities in exposure tubes were adjusted to222a concentration corresponding to a cell density ten times223higher than a natural bloom to simulate the concentration224of cells that occurs during filtration: 104 cells ml−1 for A.225fundyense (Shumway et al. 1988; Townsend et al.2262005), 105 cells ml−1 for H. akashiwo (Rensel and227Whyte 2004), 105 cells ml−1 for P. minimum (Hégaret228and Wikfors 2005a), 5.104 cells ml−1 for A. minutum229(REPHY; Haberkorn 2009) and 105 cells ml−1 for230Rhodomonas sp. and 5.104 cells ml−1 for H. triquetra as231controls. Concentrations of algae were adjusted by232diluting the cells in their spent medium (filtered at2330.2 μm). Filtrate was obtained by centrifuging (200×g,
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
234 5 min, 16°C) the culture before filtering supernatant235 using syringe filters (0.2 μm diameter). In the case of236 K. mikimotoi and K. selliformis, the concentration of237 algal cells was 4×103 cells ml−1 for the first238 experiment and 7×103 cells ml−1 for the second239 experiment; these densities could not be higher as they240 corresponded to the highest cell counts achieved in the241 cultures of K. selliformis. For the first experiment, K.242 mikimotoi was diluted 2.2 times in F/2 medium, to243 reach the same concentration as K. selliformis for the244 haemocyte exposures.245 For analyses involving the effect of the media on the246 haemocytes, the algal samples were filtered (0.2 μm247 filter), and the spent media (culture media after248 filtration/elimination of the algal cells) were collected249 for analyses. Algal sizes ranged from approximately250 20–25 μm diameter for A. fundyense, A. minutum, H.251 triquetra and P. minimum to 15 μm for H. akashiwo252 and Rhodomonas sp., which also corresponds to the253 size of the haemocytes. The harmful algal species used254 in this study all have demonstrated toxicity to finfish or255 shellfish (Shumway and Cucci 1987; Erard-Le-Denn et256 al. 1990; Luckenbach et al. 1993; Wikfors and257 Smolowitz 1993, 1995; Arzul et al. 1995; Lush et al.258 1996; Jenkinson and Arzul 2000; Guillou et al. 2002;259 Bricelj et al. 2005).
260 Experimental design
261 Bivalve haemocytes from five or six individual262 animals were exposed to the different species of
263harmful algae (Table 1) for 1, 2, 3 or 4 h (n=5–6).264Results of the first set of experiments indicated that265the effects of the harmful algal cells on haemocytes266and vice versa occurred very rapidly, often before2674 h of incubation (data not shown), but that the268results had reached a stable end point by 4 h. Thus,269results after 4 h of interaction are presented here.270Control analyses were also carried out on haemo-271cytes in 0.2-μm filtered seawater (FSW) only.272As the different algal species were cultured in273different seawater media, possible effects of the274media on the haemocytes were also tested to275ascertain whether the observed effect was attribut-276able to the algae itself or the media in which it was277grown. To accomplish this, haemocytes were278exposed to each culture medium at a quantity279equivalent to microalgal culture exposures.
280Flow cytometric analyses
281Haemocyte analyses
282Haemolymph was withdrawn from the adductor283muscle of bivalves using a 5- or 1-ml syringe, then284screened through 80-μm mesh and stored temporarily285in microcentrifuge tubes on ice before use. Haemo-286cyte analyses were conducted on haemolymph col-287lected from individual bivalves.288Haematoimmunological parameters measured289were haemocyte characterization, in terms of size–290FSC detector and internal complexity–SSC detector
t1.1 Table 1 Harmful Q3algae–bivalve haemocyte interactions tested
t1.2 Mercenariamercenaria
Mya arenaria Crassostreavirginica
Crassostrea gigas Ruditapesphilippinarum
t1.3 Alexandrium fundyense Culture, media Culture Culture
t1.4 Alexandrium minutum Culture, media
t1.5 Heterocapsa triquetra Culture, media
t1.6 Heterosigma akashiwo Culture, media Culture Culture
t1.7 Prorocentrum minimum Culture, media Culture Culture
t1.8 Rhodomonas Culture Culture Culture
t1.9 Karenia selliformis Culture, media
t1.10 Karenia mikimotoi Culture
Empty cells are the interactions that were not assessed
Culture the whole culture was tested, media the spent medium was tested
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
291 according to Hégaret et al. (2003a), as well as some292 of their immune functions:
293 (a) Haemocyte mortality, as percentage of dead294 haemocytes, using propidium iodide (Sigma,295 final concentration 20 μg/ml) according to296 Hégaret et al. (2003b)297 (b) Phagocytosis of fluorescent microbeads (Fluo-298 resbrite YG Microspheres, 2.00 μm, Polyscien-299 ces) by haemocytes, as percentage of highly300 phagocytic (>2 beads) haemocytes according to301 Hégaret et al. (2003b)302 (c) Haemocyte production of reactive oxygen spe-303 cies (ROS) with potential to kill non-self,304 engulfed particles was assessed using 2′,7′-305 dichlorofluorescein diacetate (Sigma) described306 in Buggé et al. (2007)307 (d) Adhesion of the haemocytes was measured by308 assessing the proportion of haemocytes that detach309 from the surface of experimental chambers after310 incubation with potential toxins as previously311 described for clams by Choquet et al. (2003).312 The assay was conducted in 24-well plates
313 We used a FACScalibur or FACScan (BD Biosciences,314 San Jose, CA, USA) flow cytometer for all haemocyte315 analyses.
316 Algal analyses
317 Characterization of microalgal cells (size–FSC detector,318 complexity–SSC detector and chlorophyll fluorescence–319 FL3 detector) was assessed by flow cytometry for each320 exposure. The percentage of dead algal cells was also321 assessed by flow cytometry using Sytox Green nucleic322 acid stain (Molecular Probes, S7020; Veldhuis et al.323 1997), which selectively stains dead algal cells, with324 fluorescence detected by the cytometer FL1 detector.325 To analyse interactions between haemocytes and326 harmful algae, microscopic observations, as well as327 flow cytometric analyses, were conducted for five to six328 replicates. The density of algae compare to haemocytes329 was from one to ten haemocytes per algal cell,330 according to the combination of species tested. The331 volume ratio was 1:3 (haemolymph/algal culture).
332 Statistical analyses
333 Results were analysed statistically with then t tests,334 contrasting FSW controls with each treatment after
3354 h of incubation, using Statgraphics Plus statistical336software (Manugistics, Inc., Rockville, MD, USA).
337Results
338Preliminary experiments confirmed that no algal339species was phagocytic, engulfing neither beads nor340haemocytes—activities that could have confounded341interpretation of cytograms.
342Effect of the media on haemocytes of the several343bivalve species
344The haemocyte variables of quahogs, soft-shell clams345and eastern oysters were assessed with haemocytes346incubated with FSW (control), F/2-enriched Milford347seawater (A. fundyense medium) and E-medium348(Rhodomonas sp., H. akashiwo and P. minimum349media). The haemocyte parameters of Manila clams350also were assessed with haemocytes incubated with351FSW (control) and F/2 medium (Karenia spp.352medium). These analyses confirmed that any effects353of algal culture were attributable to the algae354themselves and not to the media in which the cells355were grown. Only one haemocyte variable was356affected by the E-medium: The percentage of phago-357cytic haemocytes decreased when quahog haemocytes358were incubated in E-medium. None of the other359haemocyte parameters tested was affected by the360presence of the media compared to haemocytes361incubated in FSW (t test, P>0.05). Primary, flow362cytometric values for parameters measured when363haemocytes were exposed to FSW are presented in364Table 2.
365Quahog (= hard clam) haemocytes—whole cultures366and cell-free (= spent) media—of A. fundyense,367H. akashiwo, P. minimum and Rhodomonas sp.368as control
369Haemocytes exposed to FSW or to Rhodomonas sp.370did not show any measurable differences after 4 h of371incubation (Fig. 1); thus, effects of spent media from372these algae were not assessed. Only the whole culture373of the raphidophyte H. akashiwo caused mortality of374haemocytes after 4 h (Fig. 1). Haemocyte morphology375was affected only by P. minimum, which induced a376slight decrease in size and complexity (Fig. 1) of the
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
377 haemocytes. Adhesion of haemocytes was signifi-378 cantly inhibited by cultures of A. fundyense (Fig. 1).379 Similarly, haemocytes incubated in H. akashiwo spent380 medium were less able to adhere, whereas the spent
381medium of P. minimum enhanced the adhesion of382haemocytes (Table 3). The whole culture and spent383media of the dinoflagellates A. fundyense and P.384minimum caused significant decreases in percentages
RhodomonasAlexandrium fundyenseHeterosigma akashiwoProrocentrum minimum
Per
cen
tag
e o
f th
e co
ntr
ol
*
** * * *
Size Complexity Phagocytosis ROSProduction
Mortality Adhesion
Immune function
700
600
500
300
200
100
0
Fig. 1 Effects of in vitroexposure of whole cultureof Rhodomonas sp., A.fundyense, H. akashiwo andP. minimum uponhaemocyte parameters ofNorthern quahogs M.mercenaria (results arepresented as a percentage ofthe FSW control(mean, ±SE), cf. Table 2 forcontrol values; asteriskindicates a significant effect,t test, P<0.05)
t2.1 Table 2 Control values for the several haemocyte parameters of the five bivalve species tested, after 4 h of incubation with FSW,mean and SE, n=5
t2.2 Percentage of dead haemocytes Percentage of phagocytosis Percentage of adhered cells
t2.3 Mean SE Mean SE Mean SE
t2.4 Crassostrea gigas 5.48 2.86 8.19 1.07 35.80 12.47
t2.5 Crassostrea virginica 1.63 0.38 19.56 3.10
t2.6 Mercenaria mercenaria 8.02 2.16 35.36 1.21 94.93 3.14
t2.7 Mya arenaria 7.12 1.60 10.71 1.43 87.87 2.63
t2.8 Ruditapes philippinarum 14.73 2.51 24.19 2.73 81.10 6.04
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF385 of phagocytic haemocytes (Fig. 1; Table 3); however,
386 the effect observed with P. minimum may be387 attributable to the E-medium and not to the algae388 themselves, as E-medium alone also inhibited phago-389 cytosis. No significant effect of any harmful algal390 species tested on a loss of haemocyte counts within391 the tube during the time of exposure or on production392 of ROS (Table 3; Fig. 1) was found.393 Total haemolymph from quahogs M. mercenaria394 had significant effects on the harmful algal species395 (Table 4). Quahog haemolymph affected the mor-396 phology of some algal cells species, e.g. the com-397 plexity of A. fundyense increased. Microscopic
398observations showed the presence of numerous399temporary cysts of A. fundyense when the algal cells400were incubated with quahog haemolymph (Fig. 2);401the algal cells were losing cell wall plates and402transforming into temporary cysts. The size of H.403akashiwo increased, whereas its chlorophyll fluores-404cence and complexity decreased in the presence of405quahog haemolymph, and H. akashiwo cells appeared406to lose shape and chlorophyll fluorescence and to407degrade very quickly. Chlorophyll fluorescence of the408three algal species tested (H. akashiwo, P. minimum409and A. fundyense) also decreased significantly in the410presence of quahog haemolymph. Quahog haemo-
t4.1 Table 4 Effect of the haemocytes of several molluscan bivalve species on harmful algal species
t4.2 Harmful algal species Mollusc bivalve species Algal parameters tested
t4.3 Size Complexity Fluorescence Mortality
t4.4 Mean SE Mean SE Mean SE Mean SE
t4.5 Alexandrium fundyense Crassostrea virginica 108.5 3.3 115.1a 3.9 97.9a 0.7 335.2a 73.6
t4.6 Mercenaria mercenaria 106.9 6.4 115.1a 2.6 100.6 1.4 54.0 15.1
t4.7 Mya arenaria 111.4a 2.6 106.8a 1.8 99.7 1.9 260.7a 58.4
t4.8 Heterosigma akashiwo Crassostrea virginica 100.3 1.3 100.7 2.8 93.9a 2.1 309.8 146.0
t4.9 Mercenaria mercenaria 121.9a 6.6 81.8 11.1 88.2a 2.3 424.1a 76.1
t4.10 Mya arenaria 114.3a 4.4 86.8a 1.8 83.9a 3.9 5,799.8a 2,344.3
t4.11 Prorocentrum minimum Crassostrea virginica 102.8a 1.2 96.5 1.4 98.4a 0.7 64.4a 10.9
t4.12 Mercenaria mercenaria 109.7 6.7 116.5 6.9 89.9a 1.6 198.2a 26.5
t4.13 Mya arenaria 96.6 10.4 105.2 3.7 96.5 2.8 491.6a 104.4
t4.14 Karenia mikimotoi Ruditapes philippinarum 156.8a 4.1 174.4a 4.1 89.6a 2.0
t4.15 Karenia selliformis Ruditapes philippinarum 171.5a 10.0 274.5a 30.3 97.5 8.9
Results are presented as a percentage of the FSW controla Indicates a significant effect, t test, P<0.05, n=5–6
t3.1 Table 3 Effects of the cell-free (=spent) media of algal species on haemocyte variables of quahogs M. mercenaria
t3.2 Haemocyte parameters of Mercenaria mercenaria tested
t3.3 Size Complexity Mortality Phagocytosis Production ROS Adhesion
t3.4 Mean SE Mean SE Mean SE Mean SE Mean SE Mean SE
t3.5 A. fundyense medium 91.7 10.7 97.0 7.8 81.3 42.3 80.8a 10.7 119.6 27.6 86.3 3.9
t3.6 H. akashiwo medium 96.7 7.8 91.6 12.4 40.7 15.2 63.9a 14.0 97.3 13.2 31.6a 19.4
t3.7 P. minimum medium 99.3 2.2 90.3 5.3 77.0 14.8 60.0a 3.8 139.0 31.4 114.1a 12.6
Results are presented as a percentage of the FSW controla Indicates a significant effect, t test, P<0.05, n=5
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
411 lymph also increased the percentage of dead algal412 cells in P. minimum and H. akashiwo; conversely, the413 percentage of dead A. fundyense cells decreased.414 Microscopic observations also indicated the presence415 of red (= chlorophyll) fluorescence in individual416 haemocytes incubated with H. akashiwo (Fig. 3) and417 the presence of aggregates of haemocytes surrounding418 P. minimum cells (Fig. 4).
419Soft-shell clam haemocytes—whole cultures420of A. fundyense, H. akashiwo and P. minimum421and Rhodomonas sp. as control
422Haemocytes from soft-shell clams M. arenaria ex-423posed to FSWor to Rhodomonas sp. did not show any424measurable differences after 4 h of incubation, except425for a significant increase in adhesion of haemocytes426(Fig. 5). Haemocytes from soft-shell clam exposed to427three different species of harmful algae showed some428significant effects as well. The production of ROS by429soft-shell clam haemocytes (Fig. 5) significantly430increased in the presence of H. akashiwo. Conversely,431A. fundyense and P. minimum, which did not affect the432production of ROS by haemocytes, significantly433decreased the percentage of phagocytic haemocytes434(Fig. 5). The haemocyte adhesion (Fig. 5) was not435affected by any of the harmful algae tested. The436percentage of dead haemocytes after incubation with437A. fundyense and P. minimum did not change either438(Fig. 5). Flow cytometric studies did not allow439assessment of the effect of the H. akashiwo upon440haemocyte viability; algae incubated with the haemo-441cytes lost chlorophyll fluorescence and shape; there-442fore, it was not possible to distinguish between the red443fluorescence of chlorophyll and the red fluorescence444of dead haemocytes (stained with propidium iodide).445The observations of the cytograms plotting haemo-446cytes and H. akashiwo indicated that the algal cells447either lost complexity or fluorescence or that the448haemocytes engulfed algal cells, thereby acquiring449chlorophyll fluorescence.
Fig. 2 Temporary cyst of A. fundyense (arrow) exposed toquahog haemocytes (arrowhead; scale bar=20 μm)
Fig. 4 Aggregates of quahog haemocytes surrounding P.minimum cells (scale bar=20 μm)
Fig. 3 Cells of H. akashiwo engulfed by haemocytes ofquahogs M. mercenaria. Arrow indicates chlorophyll fluores-cence inside haemocytes (scale bar=20 μm)
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
450 During this study, the effects of soft-shell clam451 haemolymph on algal cells were also assessed452 (Table 4). For the three algal species tested, the453 percentage of dead algal cells increased significantly454 in the presence of soft-shell clam haemolymph.455 Morphology and chlorophyll content in the dinofla-456 gellate P. minimum were not affected by the presence457 of soft-shell clam haemolymph, but A. fundyense cells458 had significantly higher mean complexity and size in459 the presence of clam haemolymph. Finally, H.460 akashiwo was most affected by soft-shell clam461 haemolymph; counts of H. akashiwo cells in the462 tubes incubated with haemocytes strongly decreased463 whereas mortality increased, simultaneously, the size464 of the algal cells increased and the chlorophyll465 content and the complexity decreased significantly.
466 Oyster haemocytes—whole cultures of A. fundyense,467 H. akashiwo and P. minimum and Rhodomonas sp.468 as control
469 Results of experimental co-incubations of Eastern470 oyster C. virginica haemocytes and three microalgal
471species are presented Fig. 6. Two or three populations472of haemocytes were detected with the flow cytometer:473granulocytes and large and small hyalinocytes, which474were not always distinguishable. Haemocytes exposed475to Rhodomonas sp. did not show any significant476changes from the FSW control. Results showed that477A. fundyense and H. akashiwo caused an increase in478the percentage of dead haemocytes (Fig. 6). Moreover,479A. fundyense inhibited phagocytosis by oyster haemo-480cytes (but not significantly; Fig. 6). P. minimum had no481significant effects on oyster haemocytes.482Cells of A. fundyense in contact with oyster483haemolymph tended to lose chlorophyll fluorescence,484but percentages of dead algal cells and internal485complexity both increased (Table 4). These observa-486tions are consistent with microscopic observations487wherein we observed that A. fundyense, in the488presence of haemolymph, was often in the form of489temporary cysts and not vegetative cells (Fig. 2). In490the presence of oyster haemolymph, the cells of H.491akashiwo showed loss of chlorophyll fluorescence,492but microscopic observations indicated that the algal493cells were still motile. The dinoflagellate P. minimum
Rhodomonas
Alexandrium fundyense
Heterosigma akashiwo
Prorocentrum minimum
Per
cen
tag
e o
f th
e co
ntr
ol
**
*
*
Size Complexity Phagocytosis ROSProduction
Mortality Adhesion
Immune function
750
650
550
200
100
0
Fig. 5 Effects of in vitroexposure of whole cultureof Rhodomonas sp., A.fundyense, H. akashiwoand P. minimum uponhaemocyte parameters ofsoft-shell clams M. arenaria(results are presented as apercentage of the FSWcontrol (mean, ±SE), cf.Table 2 for control values;asterisk indicates a significanteffect, t test, P<0.05)
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol
AUTHOR'S PROOF
UNCORRECTEDPROOF
494 lost chlorophyll fluorescence and increased in size495 when incubated in oyster haemolymph. The presence496 of haemolymph also caused a decrease in the number497 of P. minimum cells, as well as a decrease in the498 percentage of dead algal cells. Microscopic observa-499 tions indicated that oyster haemocytes and P. mini-500 mum cells did not seem to have any major cell-to-cell501 interactions; the P. minimum cells remained motile.
502 Manila clam haemocytes—whole culture503 of K. mikimotoi and whole culture and cell-free504 (= spent) medium of K. selliformis
505 As K. selliformis had a much stronger effect on clam506 haemocytes than K. mikimotoi, the experiment only507 assessed the effect of K. selliformis spent medium on508 Manila clams R. philippinarum haemocytes. Results509 of experimental co-incubations of Manila clam hae-510 mocytes and the two Karenia species show a511 significant effect of K. selliformis on haemocyte
512morphology, with a decrease in complexity (Fig. 7).513Although haemocyte size was not affected, haemocyte514functions of Manila clams exposed to K. selliformis515were affected; percentage of dead haemocytes increased516significantly (Fig. 7) while the percentage of phago-517cytic haemocytes and production of ROS and the518adhesion decreased (Fig. 7). The alga K. mikimotoi had519a much smaller impact on clam haemocytes but still520inhibited adhesion (Fig. 7) and suppressed production521of ROS (Fig. 7). The effect of the spent medium of K.522selliformis on clam haemocytes was not as pronounced523as for the whole culture but still caused a significant524inhibition of the production of ROS and increased the525mortality of haemocytes (Fig. 7).526Cells of Karenia spp. incubated with haemolymph527of Manila clams for 4 h reacted with a highly528significant increase in size and complexity (Table 4).529The chlorophyll fluorescence also tended to decrease530in both Karenia species, but the trend was only531significant for K. mikimotoi (Table 4).
Rhodomonas
Alexandrium fundyense
Heterosigma akashiwo
Prorocentrum minimum
* *
Size Complexity Phagocytosis ROS Production
Mortality Adhesion
Immune function
1400
1200
1000
400
200
0
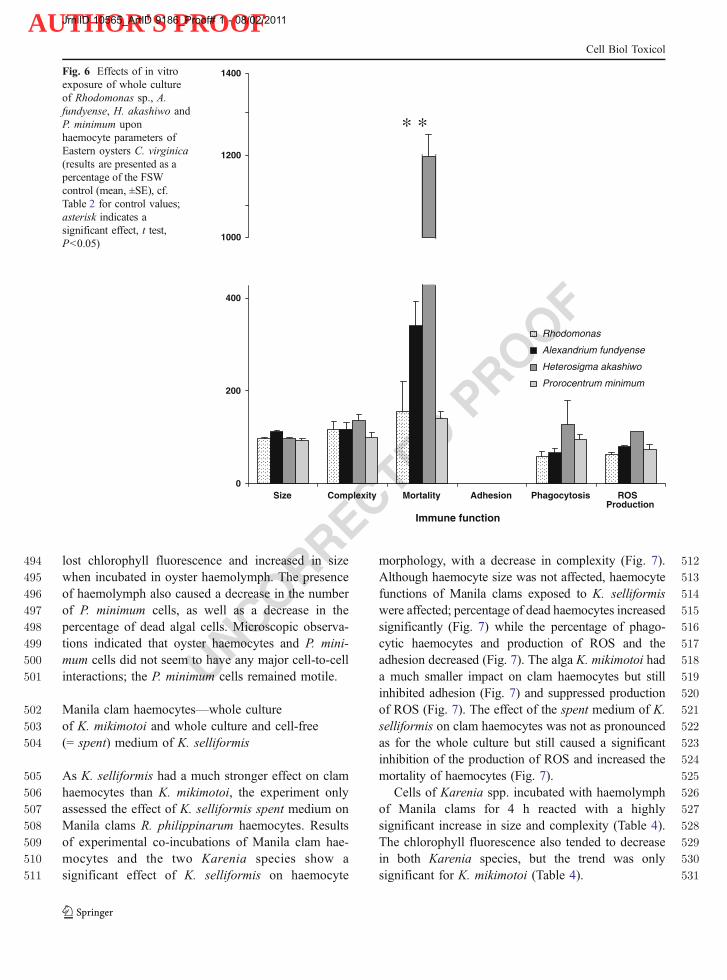
Fig. 6 Effects of in vitroexposure of whole cultureof Rhodomonas sp., A.fundyense, H. akashiwo andP. minimum uponhaemocyte parameters ofEastern oysters C. virginica(results are presented as apercentage of the FSWcontrol (mean, ±SE), cf.Table 2 for control values;asterisk indicates asignificant effect, t test,P<0.05)
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF532 Pacific oyster haemocytes—whole culture and cell-free
533 (= spent) medium of A. minutum and H. triquetra
534 Three populations of haemocytes were detected535 using flow cytometry: granulocytes, hyalinocytes536 and agranulocytes, but results presented here only537 show effects on the whole haemocyte population.538 Exposure for 4 h to H. triquetra or its spent medium539 did not significantly affect haemocyte parameters
540tested in comparison to the control (Fig. 8). Results541show that A. minutum and its spent medium,542however, caused an increase in the percentage of543dead haemocytes and a decrease in their internal544complexity (Fig. 8). Moreover, the whole culture of545A. minutum significantly inhibited production of546ROS by oyster haemocytes, whereas inhibition of547the production of ROS by A. minutum spent medium548was not significant (Fig. 8).
Karenia selliformisKarenia mikimotoiKarenia selliformis medium
*
* *
* * * * * *
Complexity ROS Production
Mortality Adhesion
Immune function
Phagocytosis
350
300
250
200
150
100
50
0
Per
cen
tag
e o
f th
e co
ntr
ol
Fig. 7 Effects of in vitroexposure of whole cultureof K. selliformis and K.mikimotoi and of the spentmedium of K. selliformisupon haemocyte parametersof Manila clams R.philippinarum (results arepresented as a percentage ofthe FSW control (mean, ±SE),cf. Table 2 for control values;asterisk indicates a significanteffect, t test,P<0.05)
H. triquetra
H. triquetra medium
A. minutum
A. minutum medium
* **
*
*
Complexity ROS ProductionMortality Adhesion
Immune function
Phagocytosis
Per
cen
tag
e o
f th
e co
ntr
ol
220
200
180
160
140
120
100
80
60
Fig. 8 Effects of in vitro exposure of whole cultures and spentmedia of H. triquetra and A. minutum upon haemocyteparameters of Pacific clams C. gigas (results are presented as
a percentage of the FSW control (mean, ±SE), cf. Table 2 forcontrol values; asterisk indicates a significant effect, t test, P<0.05)
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
549 Discussion
550 Results of the present study demonstrated that a non-551 harmful, microalgal species does not modulate the552 immune response of bivalve molluscs. In contrast, the553 harmful algal species investigated did cause changes in554 haemocytes, both morphological and functional, that can555 be expected to alter the physiological status and immuno-556 logical responses of the shellfish. Our results also indicate557 that different harmful algal species may be affected558 differently when exposed to shellfish haemolymph.
559 Effect of harmful algal cells on bivalve haemocytes560 and their functions
561 Taken together, our results suggest two main562 response patterns when haemocytes are incubated563 with harmful algae cells. Some harmful algae can564 act as immunostimulants (e.g. P. minimum) whereas565 others can cause a suppression of immune functions.566 Haemocytes of soft-shell clams and quahogs suffered567 a decrease in percentage of highly phagocytic568 haemocytes in the presence of P. minimum, though569 this may be due to the culture medium itself in the570 case of quahogs. Quahog haemocyte morphology571 changed during incubation with P. minimum. Such572 modification might be related to algal cells engulf-573 ment, as suggested by the acquisition of red574 fluorescence. Quahog haemocytes exposed in vitro575 to P. minimum form aggregates surrounding algal576 cells, confirming previous observations of Hégaret et577 al. (2008a), probably to isolate P. minimum cells578 from the host tissues. Similarly, in vivo exposures of579 quahogs, Manila clams, mussels and bay scallops580 exposed to P. minimum showed depression of581 phagocytosis, induced aggregation of haemocytes582 surrounding the harmful algae and a large inflam-583 matory response characterised by a massive infiltra-584 tion of haemocytes into the intestine (Wikfors and585 Smolowitz 1993; Hégaret and Wikfors 2005a;586 Galimany et al. 2008a; Hégaret et al. 2009, 2010).587 Present in vitro results support the hypothesis that588 this massive migration of haemocytes through the589 intestinal epithelium occurred to surround and isolate590 (via encapsulation) the cells of P. minimum. The591 active migration and activation of haemocytes might592 be induced and regulated by soluble molecules,593 released either by algal cells or by intestinal affected594 cells. Grzebyk et al. (1997) reported the presence of
595a water-soluble toxin in P. minimum cultures, and596Wikfors (2005) suggested the existence of some still597uncharacterized toxins. Such molecules might then598act as activating chemical agents.599Contrastingly with clams, P. minimum cells did not600induce any immunological effect over oyster haemo-601cytes. Indeed, neither inhibition of phagocytosis nor602formation of aggregates could be detected, as previ-603ously reported in vivo (Hégaret and Wikfors 2005a, b).604Conversely, Wikfors and Smolowitz (1995) sug-605gested that P. minimum cells interfere with cellular606digestive processes. Indeed, eastern oysters, C. virgin-607ica, showed accumulation bodies within absorptive608cells of the digestive tubules, indicating that P.609minimum cells interfere with cellular digestive processes.610Similarly, Pacific oysters, C. gigas, exposed in vivo to611Prorocentrum rhathymum also had a reduction of the gut612tubule epithelium, and some affected oysters displayed613thinned digestive tubules containing sloughed cells614(Pearce et al. 2005).615The in vitro exposure of Manila clam haemo-616cytes to K. selliformis caused a decrease in haemo-617cyte internal complexity. This loss of internal618complexity could indicate a possible degranulation619of haemocytes, releasing immunoactive enzymes620into the haemolymph (Cheng 1996), or be a621consequence of algal cytotoxicity. Indeed, adhesion,622phagocytosis and production of ROS of Manila clam623haemocytes were depressed, while the percentage of624dead cells increased. Similarly, production of ROS625and adhesion were altered in Manila clams exposed626to both Karenia species. The spent medium of K.627selliformis itself induced immune suppression, al-628though in a lesser extent than algal cells. This629indicates that the effect of K. selliformis is mainly630attributable to the cells themselves, either through631direct, physical contact with the cells (Uchida et al.6321999) or more likely from a fast-degrading toxin633released by the cells (Gentien et al. 2007). Karenia634sp. cells are known to produce allelopathic and635ichtyotoxic compounds (Gentien and Arzul 1990)636but also haemolysins, which in vitro induce red637blood cells lysis (Arzul et al. 1995; Fossat et al.6381999; Sola et al. 1999; Jenkinson and Arzul 2000)639and fast-degrading toxins responsible for autotox-640icity in K. mikimotoi cultures (Gentien et al. 2007).641Cells of K. selliformis also produce a well-642characterised toxin referred to as gymnodimine (Seki643et al. 1995, 1996; Mackenzie et al. 1996), which
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
644 caused death of oyster larvae exposed to whole645 cultures, culture filtrates or sonicated cell extracts of646 Karenia sp. after 7 to 24 h of exposure (Mackenzie647 et al. 1996). Gymnodimine, however, showed no648 haemolytic activity or cytotoxicity to mouse blood649 cells (Seki et al. 1996), and its mechanism of toxicity650 remains unclear. It is then unclear whether the651 observed effects on haemocytes could be attributable652 to gymnodimine, Karenia sp. cells or both. These in653 vitro exposures also revealed an intermediate reac-654 tion of haemocytes to K. mikimotoi, compared to K.655 selliformis, which was previously observed in vivo656 (Hégaret et al. 2007a). Jenkinson and Arzul (2000)657 also showed intermediate haemolytic properties in K.658 mikimotoi, compared to K. selliformis, in exposures659 of horse red blood cells.660 The whole culture of the raphidophyte H. aka-661 shiwo caused a very large increase in the percentage662 of dead haemocytes from both quahogs and oysters,663 which was not significant with the spent medium664 indicating that the presence of the algal cells appeared665 necessary for cytotoxicity. Wang et al. (2006) showed666 an increase in mortality of scallop larvae exposed to667 H. akashiwo and suggested that its glycocalyx668 structures could be responsible for its toxicity. These669 glycocalyx structures were also reported to strongly670 inhibit swimming activity of brine shrimps, Artemia671 salina (Yan et al. 2004). Both entire culture and spent672 medium of H. akashiwo, however, triggered a673 decrease of phagocytosis from quahogs, which could674 be explained by the release of some chemical675 compounds by H. akashiwo, which may gradually676 accumulate and affect the ability of quahog haemo-677 cytes to adhere and phagocyte. Another hypothesis678 for reduced phagocytosis of fluorescent microbeads679 may be that haemocytes have been occupied engulf-680 ing harmful algal cells, as suggested by the chloro-681 phyll fluorescence observed inside haemocytes.682 Moreover, H. akashiwo cells produce ROS (Marshall683 et al. 2005a) and secrete organic compounds (Twiner684 et al. 2004, 2005) that affect the metabolic activity of685 mammalian cells within a few hours and might then686 similarly affect bivalve haemocytes.687 In vitro incubation of the dinoflagellates A.688 fundyense and A. minutum with bivalve haemocytes689 tends to induce immunosuppression. Phagocytosis of690 quahogs and soft-shell clams, as well as adhesion of691 quahog haemocytes, were depressed. Haemocytes of692 the Pacific oyster displayed a decreased internal
693complexity and an inhibited production of ROS.694Finally, both Alexandrium species increased haemo-695cyte mortality in both oyster species, C. virginica and696C. gigas. Alexandrium spp. are known to produce697paralytic shellfish toxins (PST), responsible for698potentially lethal toxicity in mammals. Ford et al.699(2008), however, reported a decrease of adhesion and700phagocytosis of Manila clam and soft-shell clam701haemocytes exposed in vitro to a non-PST producing702Alexandrium tamarense strain, whereas another PST703producing strain did not have any significant effect on704these immune functions. Pacific oysters, C. gigas,705exposed in vivo to Alexandrium catenella had an706increase in percentage of dead haemocytes, which707was not the case neither for C. virginica nor for C.708gigas, respectively, exposed in vivo to A. fundyense709and A. minutum (Hégaret et al. 2007b; Haberkorn et710al. 2010b). Arzul et al. (1999) showed haemolytic,711allelopathic and toxic activities of Alexandrium sp.712over other algae and suggested the presence of PST-713independent, chemical substances responsible for714these effects. Moreover, a recent study reported that715Alexandrium leei can secrete soluble polar ichthyo-716toxin(s), independent from PST which can cause717lesions in and death of fishes (Tang et al. 2007).718The production and release of chemical substances719might then be responsible for the in vitro effects of720Alexandrium sp. on bivalve haemocytes. Contrast-721ingly, in vivo exposure of C. virginica and C. gigas to722A. fundyense and A. minutum, respectively, did not723induce immunosuppression nor toxicity (Hégaret et724al. 2007b; Haberkorn et al. 2010b). Temporary cysts725of A. fundyense and A. minutum have been observed726in the stomach, digestive gland and biodeposits of727bivalves (Shumway et al. 2006; Galimany et al.7282008b; Hégaret et al. 2008b; Haberkorn et al.7292010b), indicating that the algal cells may transform730into cysts as they pass through the digestive system.731The release of toxic substances might then be732reduced, causing less effect on bivalve tissues and733haemocytes during in vivo exposures compared with734in vitro experiments.
735Effects of bivalve haemolymph on algae
736The cellular morphology (size and complexity) of737each algal species measured with flow cytometry738changed when incubated with bivalve haemolymph.739The most noticeable morphological modification
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
740 occurred with haemolymph of quahogs and oysters741 incubated with A. fundyense. Indeed, microscopic742 observations revealed the presence of temporary743 cysts which were not observed during incubation of744 algal cells with FSW. Temporary cysts are non-745 motile cells, surrounded by a pellicle and are746 produced by vegetative cells shedding their theca747 (ecdysis) in response to short-term or sudden adverse748 conditions. Thus, the transformation of A. fundyense749 cells into temporary cysts can be explained by a750 protective response to haemocytes. Similar in vitro751 observations of temporary cyst formation were made752 previously by Hégaret et al. (2008a), and temporary753 cysts of A. fundyense have been observed in vivo in754 the stomachs and biodeposits of bivalve molluscs755 (Persson et al. 2006; Hégaret et al. 2007c, 2008b;756 Galimany et al. 2008b).757 In the majority of the HAB–bivalve pairs investi-758 gated in the present study, harmful algal cells lost759 chlorophyll fluorescence over time as they were760 incubated with haemolymph. The trend is particularly761 extreme with H. akashiwo, for which cells incubated762 with haemolymph of soft-shell clams and, to a certain763 extent, quahogs were degraded very quickly. Disap-764 pearance of degraded cells may be attributable to765 phagocytosis by the haemocytes, which would766 explain the decrease in concentration of H. akashiwo767 cells when incubated with haemolymph. These768 results would also confirm the observations of769 Hégaret et al. (2008a), who observed haemocytes770 with red fluorescence, probably of chlorophyll from771 previously engulfed H. akashiwo cells. Conversely,772 when exposed to oyster haemolymph, the degrada-773 tion of the H. akashiwo cells did not seem as active.774 This is consistent with the partial shell closure and775 reduced filtration of oysters exposed to H. akashiwo776 (Hégaret et al. 2007c), which would limit contact777 with haemocytes.
778 Can an in vitro experiment be a good proxy779 for in vivo exposures?
780 Very few publications presenting results from in vivo781 exposure of bivalve molluscs to harmful algae can be782 found in the literature, which limits the possible783 comparisons between the same bivalve–HAB pairs in784 vitro and in vivo. It is, therefore, not always easy to785 assess whether or not in vitro experiments can be786 good proxies for in vivo exposures.
787In the present study, an incubation time of 4 h was788sufficient to detect the responses of haemocytes and789harmful algal cells to co-incubation, confirming790previous observations of in vitro interactions between791bivalve haemocytes with harmful algal cells (Hégaret792et al. 2008a) and chemical pollutants (Anderson et al.7931997; Gomez-Mendikute et al. 2002, Sauvé et al.7942002; Gagnaire et al. 2003, 2004, 2006).795The effects of P. minimum were previously tested796in various in vivo bivalve–HAB interactions with797eastern oysters C. virginica (Hégaret and Wikfors7982005a, b), bay scallops Argopecten irradians (Hégaret799and Wikfors 2005a), blue mussels Mytilus edulis800(Galimany et al. 2008a), quahogs M. mercenaria801(Hégaret et al. 2010) and Manila clams R. philippi-802narum (Hégaret et al. 2009). Our results indicated that803in vitro interactions between haemocytes and P.804minimum are good proxies for in vivo experiments805and show consistently different haemocyte responses,806according to the bivalve tested, showing no measur-807able effects on fundamental haemocyte functions in808oysters, a decrease in phagocytosis associated to the809formation of large haemocyte aggregates surrounding810the algal cells in quahogs, Manila clams and blue811mussels (Galimany et al. 2008a; Hégaret et al. 2009,8122010). In addition, previous studies (Hégaret and813Wikfors 2005a, b) also demonstrated that laboratory,814in vivo experiments exposing bivalves to cultures of815P. minimum were good proxies for field HAB816exposures. Thus, an in vitro exposure between P.817minimum and haemocytes of a bivalve might be used818as a proxy for in vivo, natural and artificial exposures.819Exposure of Manila clam haemocytes in vitro to820K. mikimotoi and K. selliformis affected haemocyte821functions, reducing adhesion, phagocytosis and822production of ROS, while increasing the percentage823of dead haemocytes. Additionally, exposure to K.824selliformis in vitro caused a decrease in complexity825of Manila clam haemocytes, which confirms the in826vivo exposure to both Karenia species (Hégaret et al.8272007b; da Silva et al. 2008). Flattened haemocytes828were also observed in A. irradians exposed in vivo to829K. mikimotoi, which tends to confirm present results830(Smolowitz and Shumway 1997). Thus, in vitro831exposure to Karenia spp. could also be used as a832proxy for in vivo exposures. However, in vivo833experimental exposures of clams to Karenia spp.834resulted in fewer dead haemocytes (Hégaret et al.8352007b). Harmful algal cell densities used in vivo
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
836 were lower than for in vitro exposures, by a factor of837 4 to 8, which may explain the difference in responses838 between in vitro and in vivo exposures. The toxins839 produced by Karenia spp. might also desintegrate840 before reaching the haemocytes in vivo, consistent841 with the hypothesis of a fast-degrading toxin842 (Gentien et al. 2007).843 The percentage of dead haemocytes in C. gigas844 exposed in vitro to A. minutum increased whereas the845 percentage of dead haemocytes during in vivo846 exposure did not (Haberkorn et al. 2010a, b).847 Moreover, in vivo exposure of C. gigas to A. minutum848 resulted in immunostimulation, characterised by in-849 flammatory response (Haberkorn et al. 2010a),850 whereas in vitro exposure did not show significant851 effects of A. minutum, besides a higher mortality and852 a decrease of the production of ROS. Such an853 inhibition of the production of ROS was similarly854 measured in C. gigas exposed in vivo to A. minutum855 in May, but the contrary was observed a month before856 on the same oyster population (Haberkorn et al.857 2010b). These contradictory results underline the858 difficulties to extrapolate in vitro effects to in vivo859 exposure of Alexandrium sp. on oysters.860 The dinoflagellates A. fundyense (this study; Hégaret861 et al. 2008a) and A. tamarense (Ford et al. 2008) in862 vitro also caused depressed phagocytosis and adhesion863 in clams (quahogs and soft-shell clams). Unfortunately,864 no in vivo exposures have been conducted with any of865 the clam species tested in vitro. Similarly, haemocytes866 of blue mussels exposed in vivo to A. fundyense were867 morphologically different, with lower size and com-868 plexity than the control, but no major changes of the869 haemocyte functions could be observed (Galimany et al.870 2008b), but no data of in vitro exposure of haemocytes871 of mussels to Alexandrium sp. are available. Thus,872 further experiments are needed to ascertain whether or873 not in vitro experiments involving Alexandrium sp. can874 be good proxies for in vivo responses.
875 Conclusion
876 This study analysed the effects in vitro of harmful877 algal cells on haemocytes of several bivalve molluscs.878 Despite differences between the several harmful algae879 tested, results show that haemocytes, the immune880 defence cells in bivalves, respond to HABs in a881 somewhat species-specific manner, but HAB species
882often cause a consistent profile of immunomodulation883in most bivalve species. HAB exposure generally is884associated with increases in the percentages of dead885haemocytes and decreases in haemocyte phagocytosis,886production of ROS and adhesion (immunosuppression).887In most cases, the production and secretion of chemical888substances by algal cells is probably responsible for889such effects. Contrastingly, some harmful algal species890(e.g. P. minimum) can act as immunostimulants,891activating a protective cellular immune response in892bivalves.893This study also indicates that haemolymph and894haemocytes can have measurable effects on algal895cells, including changes in shape, chlorophyll896fluorescence and mortality. Yet, further investiga-897tions will be needed to fully understand the cellular898and molecular mechanisms of toxicity of harmful899algae to bivalves and their effects on haemocyte900responses.901Finally, our results compared to data from the902literature suggested that in vitro analyses “have the903potential to serve” as proxies for in vivo analyses,904although further studies should be developed to905carefully assess the correspondence between in vitro906and in vivo interactions for some bivalve/algae pairs.907908Acknowledgements We thank Nelly Le Goïc, Lise Raimbault,909Madeleine Gonçalvez, Jennifer Alix and Mark Dixon for their910help during the experiments and the preparation of the911manuscript; we also thank Ludovic Donaghy for his help912preparing the manuscript. This work was supported by the913Lerner Gray Fund for Marine Research from the American914Museum of Natural History, the National Shellfisheries Associ-915ation, Sigma Xi, Connecticut Sea Grant, and by USEPA/916ECOHAB grant number 523792.
917References 918
919Anderson RS. Interactions of Perkinsus marinus with humoral920factors and hemocytes of Crassostrea virginica. J Shellfish921Res. 1996;15:127–34.922Anderson RS. Lack of hemocyte chemiluminescence stimula-923tion by Perkinsus marinus in eastern oysters Crassostrea924virginica with dermo disease. J Aquat Anim Health.9251999;11:179–82.926Anderson RS, Burreson EM, Paynter KT. Defense responses of927hemocytes withdrawn from Crassostrea virginica infected928with Perkinsus marinus. J Invert Pathol. 1995;66:82–9.929Anderson RS, Brubacher LL, Calvo LMR, Burreson EM,930Unger MA. Effect of in vitro exposure to tributyltin on931generation of oxygen metabolites by oyster hemocytes.932Environ Res. 1997;74:84–90.
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
933 Arzul G, Gentien P, Bodennec G, Toularastel F, Youenou A,934 Crassous MP. Comparison of toxic effects in Gymnodi-935 nium cf. nagasakiense polyunsaturated fatty acids. In:936 Lassus P, Arzul G, Erard E, Gentien P, Mercaillou-Le-Baut937 C, editors. Harmful marine algal blooms. Paris: Lavoisier;938 1995. p. 395–400.939 Arzul G, Seguel M, Guzman L, Erard-Le Denn E. Comparison940 of allelopathic properties in three toxic Alexandrium941 species. J Exper Mar Biol Ecol. 1999;232:285–95.942 Bachère E, Gueguen Y, Gonzalez M, de Lorgeril J, Garnier943 J, Romestand B. Insights into the anti-microbial defense944 of marine invertebrates: the penaeid shrimps and the945 oyster Crassostrea gigas. Immunol Rev. 2004;198:149–946 68.947 Bado-Nilles A, Gagnaire B, Thomas-Guyon H, Le Floch S,948 Renault T. Effects of 16 pure hydrocarbons and two oils949 on haemocyte and haemolymphatic parameters in the950 Pacific oyster, Crassostrea gigas (Thunberg). Toxicol In951 Vitro. 2008;22(6):1610–7.952 Bricelj V, Connell L, Konoki K, MacQuarrie S, Scheuer T,953 Catterall W, et al. Sodium channel mutation leading to954 saxitoxin resistance in clams increases risk of PSP. Nature.955 2005;434:763–6.956 Buggé D, Hégaret H, Wikfors GH, Allam B. Oxidative burst in957 hard clam (Mercenaria mercenaria) haemocytes. Fish958 Shellfish Immunol. 2007;23:188–96.959 Burkholder JM. Implications of harmful microalgae and960 heterotrophic dinoflagellates in management of sustainable961 marine fisheries. Ecol Appl. 1998;8:S37–62.962 Cheng TC. Hemocytes: Forms and functions. In: Kennedy VS,963 Newell RIE, Eble AF, editors. The Eastern oyster964 Crassostrea virginica. College Park: Maryland Sea Grant;965 1996. p. 299–333.966 Choquet G, Soudant P, Lambert C, Nicolas JL, Paillard C.967 Reduction of adhesion properties on Ruditapes philippinarum968 hemocytes exposed to Vibrio tapetis. Dis Aquat Org.969 2003;57:109–16.970 Chu FLE, Lapeyre JF. Development of disease caused by the971 parasite, Perkinsus marinus and defense-related hemo-972 lymph factors in three populations of oysters from the973 Chesapeake Bay, USA. J Shellfish Res. 1993a;12:21–7.974 Chu FLE, Lapeyre JF. Perkinsus marinus susceptibility and975 defense-related activities in eastern oysters Crassostrea976 virginica—temperature effects. Dis Aquat Org. 1993b;977 16:223–34.978 da Silva P, Hégaret H, Lambert C, Wikfors GH, Le Goïc N,979 Shumway SE, et al. Immunological responses of the980 Manila clam (Ruditapes philippinarum) with varying981 parasite (Perkinsus olseni) burden, during a long term982 exposure to the harmful alga, Karenia selliformis and983 possible interactions. Toxicon. 2008;51:563–73.984 Erard-Le-Denn E, Morlaix M, Dao JC. Effects of Gyrodinium985 cf. aureolum on Pecten maximus (post larvae, juveniles986 and adults). In: Graneli E, Sundstrom B, Edler L,987 Anderson DM, editors. Toxic marine phytoplankton.988 Elsevier: Amsterdam; 1990. p. 132–6.989 Fisher WS, Oliver LM, Walker WW, Manning CS, Lytle TF.990 Decreased resistance of eastern oysters (Crassostrea991 virginica) to a protozoan pathogen (Perkinsus marinus)992 after sublethal exposure to tributyltin oxide. Mar Environ993 Res. 1999;47:185–201.
994Fisher WS, Oliver LM, Winstead JT, Long ER. A survey995of oysters Crassostrea virginica from Tampa Bay,996Florida: associations of internal defense measurements997with contaminant burdens. Aquat Toxicol. 2000;51:115–99838.999Ford SE, Bricelj MV, Lambert C, Paillard C. Deleterious effects of1000a non PST bioactive compound(s) from Alexandrium1001tamarense on bivalve hemocytes. Mar Biol. 2008;154:1432–1002793.1003Fossat B, Porthe-Nibelle J, Sola P, Masoni A, Gentien P,1004Bodennec G. Toxicity of fatty acid 18: 5n3 from1005Gymnodinium cf. mikimotoi: II. Intracellular pH and KC1006uptake in isolated trout hepatocytes. J Appl Toxicol.10071999;19:275–8.1008Fournier M, Pellerin J, Clermont Y, Morin Y, Brousseau P.1009Effects of in vivo exposure of Mya arenaria to organic and1010inorganic mercury on phagocytic activity of hemocytes.1011Toxicol. 2001;161:201–11.1012Gagnaire B, Renault T, Bouilly K, Lapegue S, Thomas-Guyon1013H. Study of atrazine effects on pacific oyster, Crassostrea1014gigas, haemocytes. Curr Pharm Des. 2003;9:193–9.1015Gagnaire B, Thomas-Guyon H, Renault T. In vitro effects of1016cadmium and mercury on Pacific oyster, Crassostrea1017gigas (Thunberg), haemocytes. Fish Shellfish Immunol.10182004;16:501–12.1019Gagnaire B, Thomas-Guyon H, Burgeot T, Renault T. Pollutant1020effects on Pacific oyster, Crassostrea gigas (Thunberg),1021hemocytes: screening of 23 molecules using flow cytom-1022etry. Cell Biol Toxicol. 2006;22:1–14.1023Galimany E, Sunila I, Hégaret H, Ramón M, Wikfors GH.1024Pathology and immune response of the blue mussel1025(Mytilus edulis L.) after an exposure to the harmful1026dinoflagellate Prorocentrum minimum. Harmful Algae.10272008a;7:630–8.1028Galimany E, Sunila I, Hégaret H, Ramón M, Wikfors GH.1029Experimental exposure of the blue mussel (Mytilus edulis,1030L.) to the toxic dinoflagellate Alexandrium fundyense:1031histopathology, immune responses, and recovery. Harmful1032Algae. 2008b;7:702–11.1033Gentien P, Arzul G. Exotoxin production by Gyrodinium cf1034aureolum (Dinophyceae). J Mar Biol Assoc UK. 1990;103570:571–81.1036Gentien P, Lunven M, Lazure P, Youenou A, Crassous MP.1037Motility and autotoxicity in Karenia mikimotoi (Dinophy-1038ceae). Phil Trans R Soc B. 2007;362:1937–46.1039Gomez-Mendikute A, Etxeberria A, Olabarrieta I, Cajaraville1040MP. Oxygen radicals production and actin filament1041disruption in bivalve haemocytes treated with benzo(a)1042pyrene. Mar Environ Res. 2002;54:431–6.1043Grzebyk D, Denardou A, Berland B, Pouchus YF. Evidence of1044a new toxin in the red-tide dinoflagellate Prorocentrum1045minimum. J Plankton Res. 1997;19:1111–24.1046Guillard RRL. Culture of phytoplankton for feeding marine1047invertebrates. In: Smith WL, Chanley MH, editors. Culture1048of marine invertebrate animals. New York: Plenum; 1975.1049p. 26–60.1050Guillard RRL, Hargraves PE. Stichochrysis immobilis is a1051diatom, not a chrysophyte. Phycologia. 1993;32:234–6.1052Guillard RRL, Ryther JH. Studies of marine planktonic diatoms. I.1053Cyclotella nana Hustedt and Detonula confervacea Cleve.1054Can J Microbiol. 1962;8:229–39.
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
1055 Guillou L, Nezan E, Cueff V, Denn EEL, Cambon-Bonavita MA,1056 Gentien P, et al. Genetic diversity and molecular detection of1057 three toxic dinoflagellate genera (Alexandrium, Dinophysis,1058 and Karenia) from French coasts. Protist. 2002;153:1059 223–38.1060 HaberkornH. Impact du dinoflagellé toxique,Alexandrium minutum,1061 sur l’huître creuse, Crassostrea gigas: approche intégrative.1062 Ph.D. thesis. Université de Bretagne Occidentale. 2009.1063 Haberkorn H, Lambert C, Le Goïc N,Moal J, Suquet M, Guéguen1064 M, et al. Effects of Alexandrium minutum exposure on1065 nutrition-related processes and reproductive output in1066 oysters Crassostrea gigas. Harmful Algae. 2010a;9:427–39.1067 Haberkorn H, Lambert C, Le Goïc N, Guéguen M, Moal J,1068 Palacios E, et al. Effects of Alexandrium minutum1069 exposure on physiological and hematological variables of1070 diploid and triploid oysters, Crassostrea gigas. Aquat1071 Toxicol. 2010b;97:96–108.1072 Hamoutene D, Payne JF, Rahimtula A, Lee K. Effect of water1073 soluble fractions of diesel and an oil spill dispersant1074 (Corexit 9527) on immune responses in mussels. Bull1075 Environ Contam Toxicol. 2004;72:1260–7.1076 Hannam ML, Bamber SD, Galloway TS, Moody AJ, Jones1077 MB. Effects of the model PAH phenanthrene on immune1078 function and oxidative stress in the haemolymph of the1079 temperate scallop Pecten maximus. Chemosphere.1080 2010a;78(7):779–84.1081 Hannam ML, Bamber SD, Galloway TS, Moody AJ, Jones1082 MB. Functional immune response in Pecten maximus:1083 combined effects of a pathogen-associated molecular1084 pattern and PAH exposure. Fish Shellfish Immunol.1085 2010b;28(1):249–52.1086 Hégaret H, Wikfors GH. Time-dependent changes in hemocytes1087 of eastern oysters, Crassostrea virginica, and northern bay1088 scallops, Argopecten irradians irradians, exposed to a1089 cultured strain of Prorocentrum minimum. Harmful Algae.1090 2005a;4:187–99.1091 Hégaret H, Wikfors GH. Effects of natural and field-simulated1092 blooms of the dinoflagellate Prorocentrum minimum upon1093 hemocytes of eastern oysters, Crassostrea virginica, from1094 two different populations. Harmful Algae. 2005b;4:201–9.1095 Hégaret H, Wikfors GH, Soudant P. Flow-cytometric analysis1096 of haemocytes from eastern oysters, Crassostrea virginica,1097 subjected to a sudden temperature elevation I. Haemocyte1098 types and morphology. J Exper Mar Biol Ecol. 2003a;1099 293:237–48.1100 Hégaret H, Wikfors GH, Soudant P. Flow cytometric analysis1101 of haemocytes from eastern oysters, Crassostrea virginica,1102 subjected to a sudden temperature elevation II. Haemocyte1103 functions: aggregation, viability, phagocytosis, and respi-1104 ratory burst. J Exper Mar Biol Ecol. 2003b;293:249–65.1105 Hégaret H, Wikfors GH, Soudant P, Lambert C, Shumway SE,1106 Bérard JB, et al. Toxic dinoflagellates (Alexandrium1107 fundyense and A. catenella) have minimal apparent effect1108 on oyster hemocytes. Mar Biol. 2007a;152:441–7.1109 Hégaret H, da Silva P, Wikfors GH, Lambert C, De Bettignies T,1110 Shumway SE, et al. Hemocyte responses of Manila clams,1111 Ruditapes philippinarum, with varying parasite, Perkinsus1112 olseni, severity to toxic-algal exposures. Aquat Toxicol.1113 2007b;84:469–79.1114 Hégaret H, Wikfors GH, Shumway SE. Diverse feeding1115 responses of five species of bivalve mollusc when exposed
1116to three species of harmful algae. J Shellfish Res.11172007c;24:549–59.1118Hégaret H, Wikfors GH, Shumway SE. In vitro interactions1119between several species of harmful algae and hemocytes1120of bivalve molluscs. In: Moestrup O, editor. Proceedings1121of the 12th International Conference on Harmful Algae,1122Copenhagen, Denmark. 2008a. p. 356–359.1123Hégaret H, Shumway SE, Wikfors GH, Pate S, Burkholder JM.1124Potential transport of harmful algae through relocation of1125bivalve molluscs. Mar Ecol Prog Ser. 2008b;361:169–79.1126Hégaret H, da Silva P, Sunila I, Shumway SE, DixonMS, Alix JH,1127et al. Perkinsosis in the Manila clam Ruditapes philippina-1128rum affects responses to the harmful-alga, Prorocentrum1129minimum. J Exper Mar Biol Ecol. 2009;371:112–20.1130Hégaret H, Smolowitz RM, Sunila I, Shumway SE, Alix JH,1131Dixon MS, et al. Combined effects of a parasite, QPX, and1132the harmful-alga, Prorocentrum minimum on northern1133quahogs, Mercenaria mercenaria. Mar Environ Res.11342010;69:337–44.1135Hine PM. The inter-relationships of bivalve haemocytes. Fish1136Shellfish Immunol. 1999;9:367–85.1137Hoagland P, Anderson DM, Kaoru Y, White AW. The economic1138effects of harmful algal blooms in states: estimates, assess-1139ment issues, and information. Estuaries. 2002;25:819–37.1140Janeway CA. The role of microbial pattern recognition in self:1141nonself discrimination in innate and adaptive immunity. In:1142Hoffmann JA, Janeway CA, Natori S, editors. Phylogenetic1143perspectives in immunity: the insect host defence. Austin:1144Landes; 1994. p. 115–22.1145Jenkinson I, Arzul G. Mitigation by cysteine compounds of1146rheotoxicity, cytotoxicity and fish mortality caused by the1147dinoflagellates, Gymnodinium mikimotoi and G. mague-1148lonnense. In: Hallegraeff G, editor. Harmful algal blooms.1149Paris: Intergovernmental Oceanographic Commission of1150UNESCO; 2000. p. 461–4.1151Landsberg JH. The effects of harmful algal blooms on aquatic1152organisms. Rev Fish Sci. 2002;10:113–390.1153Lapeyre JF, Chu FLE, Meyers JM. Hemocytic and humoral1154activities of eastern and Pacific oysters following chal-1155lenge by the protozoan Perkinsus marinus. Fish Shellfish1156Immunol. 1995;5:179–90.1157Leibovitz L, Schott EF, Karney RC. Diseases of wild, captive and1158cultured scallops. J World Maricult Soc. 1984;15:269–83.1159Loker ES, Adema CM, Zhang SM, Kepler TB. Invertebrate1160immune systems—not homogeneous, not simple, not well1161understood. Immunol Rev. 2004;198(1):10–24.1162Luckenbach MW, Sellner KG, Shumway SE, Greene K. Effects1163of two bloom forming dinoflagellates, Prorocentrum1164minimum and Gyrodinium uncatenum, on the growth and1165survival of the eastern oyster, Crassostrea virginica1166(Gmelin 1791). J Shellfish Res. 1993;12:411–5.1167Lush GJ, Hallegraeff GM, Munday BL. High toxicity of the red1168tide dinoflagellate Alexandrium minutum to the brine shrimp1169Artemia salina. In: Yasumoto T, Oshima Y, Fukuyo Y,1170editors. Harmful and toxic algal blooms. Paris: UNESCO;11711996. p. 389–92.1172Mackenzie L, Haywood A, Adamson J, Truman P, Till D.1173Gymnodimine contamination in shellfish in New Zealand.1174In: Yasumoto T, Oshima Y, Fukuyo Y, editors. Harmful1175and toxic algal blooms. Paris: Intergovernmental Oceano-1176graphic Commission of UNESCO; 1996. p. 97–100.
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
1177 Marshall JA, de Salas M, Oda T, Hallegraeff G. Superoxide1178 production by marine microalgae—I. Survey of 37 species1179 from 6 classes. Mar Biol. 2005a;147:533–40.1180 Marshall JA, Ross T, Pyecroft S, Hallegraeff G. Superoxide1181 production by marine microalgae—II. Towards under-1182 standing ecological consequences and possible functions.1183 Mar Biol. 2005b;147:541–9.1184 Matsuyama Y, Shumway SE. Impacts of harmful algal blooms1185 on shellfisheries aquaculture. In: Burnell G, Allan G, eds.1186 New technologies in aquaculture. Improving production1187 efficiency, quality and environmental management. Cam-1188 bridge: Woodhead. 2009; 580–609.1189 Medzhitov R, Janeway CA. Decoding the patterns of self and1190 nonself by the innate immune system. Science. 2002;296:298–1191 300.1192 Morga B, Arzul I, Chollet B, Renault T. Infection with the1193 protozoan parasite Bonamia ostreae modifies in vitro1194 haemocyte activities of flat oyster Ostrea edulis. Fish1195 Shellfish Immunol. 2009;26(6):836–42.1196 Oliver LM, Fisher WS, Winstead JT, Hemmer BL, Long E.1197 Relationships between tissue contaminants and defense-1198 related characteristics of oysters (Crassostrea virginica)1199 from five Florida bays. Aquat Toxicol. 2001;55:203–22.1200 Pearce I, Handlinger JH, Hallegraeff GM. Histopathology in1201 Pacific oyster (Crassostrea gigas) spat caused by the1202 dinoflagellate Prorocentrum rhathymum. Harmful Algae.1203 2005;4:61–74.1204 Persson A, Smith BC, Wikfors GH, Quilliam M. Grazing on toxic1205 Alexandrium fundyense resting cysts and vegetative cells by1206 the eastern oyster (Crassostrea virginica). Harmful Algae.1207 2006;5:678–84.1208 Rensel JE, Whyte JNC. Finfish mariculture and harmful algal1209 blooms. In: Hallegraeff G, Anderson DM, Cembella A,1210 editors. Manual on harmful microalgae. Paris: UNESCO;1211 2004. p. 693–722.1212 Rowley AF, Powell A. Invertebrate immune systems specific, quasi-1213 specific, or nonspecific? J Immunol. 2007;179:7209–14.1214 Sauvé S, Brousseau P, Pellerin J, Morin Y, Senecal L, Goudreau P,1215 et al. Phagocytic activity of marine and freshwater bivalves:1216 in vitro exposure of hemocytes to metals (Ag, Cd, Hg and1217 Zn). Aquat Toxicol. 2002;58:189–200.1218 Seki T, Satake M, Mackenzie L, Kaspar HF. T. Y. Gymnodi-1219 mine, a new marine toxin of unprecedented structure1220 isolated from New Zealand oysters and the dinoflagellate,1221 Gymnodinium sp. Tetrahedron Lett. 1995;36:7093–6.1222 Seki T, Satake M, Mackenzie L, Kaspar HF, Yasumoto T.1223 Gymnodimine, a novel toxic imine isolated from the1224 Foveaux strait oysters and Gymnodinium sp. In: Yasumoto1225 T, Oshima Y, Fukuyo Y, editors. Harmful and toxic algal1226 blooms. Paris: Intergovernmental Oceanographic Commis-1227 sion of UNESCO; 1996. p. 495–8.1228 Shumway SE. A review of the effects of algal blooms on shellfish1229 and aquaculture. J World Aquac Soc. 1990; 21:65–104.1230 Shumway SE, Cucci TL. The effect of the toxic Protogonyaulax1231 tamarensis on the feeding and behavior of bivalve molluscs.1232 Aquat Toxicol. 1987;10:9–27.1233 Shumway SE, Sherman-Caswell S, Hurst JW. Paralytic shell-1234 fish poisoning in Maine: monitoring a monster. J Shellfish1235 Res. 1988;7:643–52.1236 Shumway SE, Burkholder JM, Springer J. Effects of the1237 estuarine dinoflagellate Pfiesteria shumwayae (Dinophy-
1238ceae) on survival and grazing activity of several shellfish1239species. Harmful Algae. 2006;5(4):442–58.1240Smolowitz R, Shumway SE. Possible cytotoxic effects of the1241dinoflagellate, Gyrodinium aureolum, on juvenile bivalve1242mollusks. Aquac Intern. 1997;5(4):291–300.1243Sola F,Masoni A, Fossat B, Porthe-Nibelle J, Gentien P, Bodennec1244G. Toxicity of fatty acid 18: 5n3 from Gymnodinium cf.1245mikimotoi: I. Morphological and biochemical aspects on1246Dicentrarchus labrax gills and intestine. J Appl Toxicol.12471999;19:279–84.1248Tang YZ, Kong LS, Holmes MJ. Dinoflagellate Alexandrium leei1249(Dinophyceae) from Singapore coastal waters produces a1250water-soluble ichthyotoxin. Mar Biol. 2007;150:541–9.1251Townsend DW, Pettigrew NR, Thomas AC. On the nature of1252Alexandrium fundyense blooms in the Gulf of Maine.1253Deep-Sea Res Part Ii-Topical Studies in Oceanography.12542005;52:2603–30.1255Twiner MJ, Dixon SJ, Trick CG. Extracellular organics from1256specific cultures of Heterosigma akashiwo (Raphidophy-1257ceae) irreversibly alter respiratory activity in mammalian1258cells. Harmful Algae. 2004;3:173–82.1259Twiner MJ, Chidiac P, Dixon SJ, Trick CG. Extracellular organic1260compounds from the ichthyotoxic red tide alga Heterosigma1261akashiwo elevate cytosolic calcium and induce apoptosis in1262Sf9 cells. Harmful Algae. 2005;4:789–800.1263Uchida T, Toda S, Matsuyama Y, Yamaguchi M, Kotani Y, Honjo1264T. Interactions between the red tide dinoflagellates Hetero-1265capsa circularisquama and Gymnodinium mikimotoi in1266laboratory culture. J Exper Mar Biol Ecol. 1999;241:285–99.1267Ukeles R. Continuous culture—a method for the production of1268unicellular algal foods. In: Stein JR, editor. Handbook of1269phycological methods: culture methods and growth meas-1270urements. Cambridge: Cambridge University Press; 1973.1271p. 233–55.1272Veldhuis MJW, Cucci TL, Sieracki ME. Cellular DNA content of1273marine phytoplankton using two new fluorochromes: taxo-1274nomic and ecological implications. J Phycol. 1997;33:527–41.1275Villalba A, Reece KS, Ordas MC, Casas SM, Figueras A.1276Perkinsosis in molluscs: a review. Aquat Liv Res.12772004;17:411–32.1278Wang LP, Yan T, Zhou MJ. Impacts of HAB species Heterosigma1279akashiwo on early development of the scallop Argopecten1280irradians Lamarck. Aquac. 2006;255:374–83.1281Wikfors GH. A review and new analysis of trophic interactions1282between Prorocentrum minimum and clams, scallops, and1283oysters. Harmful Algae. 2005;4:585–92.1284Wikfors GH, Smolowitz RM. Detrimental effects of a Prorocen-1285trum isolate upon hard clams and bay scallops in laboratory1286feeding studies. In: Smayda TJ, Shimizu Y, editors. Toxic1287phytoplankton blooms in the sea. New York: Elsevier; 1993.1288p. 447–52.1289Wikfors GH, Smolowitz RM. Experimental and histological studies1290of four life-history stages of the eastern oyster, Crassostrea1291virginica, exposed to a cultured strain of the dinoflagellate1292Prorocentrum minimum. Biol Bull. 1995;188:313–28.1293Yan T, Zhou MJ, Fu M, Wang Y, Yu F, Li RC. The toxicity1294study on Heterosigma akashiwo using an Artemia bioas-1295say. In: Hall S, Etheridge S, Anderson D, Kleindinst J,1296Zhu M, Zou Y, editors. Harmful algae management and1297mitigation, APEC #204-MR-04.2. Singapore: Asia-Pacific1298Economic Cooperation; 2004. p. 220–5.
1299
JrnlID 10565_ArtID 9186_Proof# 1 - 08/02/2011
Cell Biol Toxicol

AUTHOR'S PROOF
UNCORRECTEDPROOF
AUTHOR QUERIES
AUTHOR PLEASE ANSWER ALL QUERIES.
Q1. Please check if authors’ affiliations were correctly presented.Q2. “35 ppt ppt” here was changed to “35 ppt”. Please check if appropriate.Q3. There were modifications made in Tables 1–4. Please check if appropriate.




















