
in e ac ions, pe sis ence and coe is ence mechanisms ... - UFRN
160
IN8E7AC8IONS, PE7SIS8ENCE AND COE<IS8ENCE MECHANISMS OF NEC8A7I:O7O9S BA8S AND 8HE PLAN8S 8HE= FEED ON, IN A SEASONALL= D7= 87OPICAL FO7ES8 IN NO78HEAS8E7N B7A>IL Eugenia Cordero Schmidt Orientador: Dr. Eduardo Martins Venticinque Co-orientador: Dr. Bernal Rodríguez Herrera
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of in e ac ions, pe sis ence and coe is ence mechanisms ... - UFRN

INTERACTIONS, PERSISTENCE ANDCOEXISTENCE MECHANISMS OFNECTARIVOROUS BATS AND THE PLANTS THEYFEED ON, IN A SEASONALLY DRY TROPICALFOREST IN NORTHEASTERN BRAZILEugenia Cordero SchmidtOrientador: Dr. Eduardo Martins VenticinqueCo-orientador: Dr. Bernal Rodríguez Herrera

INTERAÇÕES, MECANISMOS DE PERSISTÊNCIA E COEXISTÊNCIA DE
MORCEGOS NECTARÍVOROS E DAS PLANTAS DAS QUAIS SE
ALIMENTAM EM UMA FLORESTA TROPICAL SAZONAL SECA NO
NORDESTE BRASILEIRO
INTERACTIONS, PERSISTENCE AND COEXISTENCE MECHANISMS OF
NECTARIVOROUS BATS AND THE PLANTS THEY FEED ON, IN A
SEASONALLY DRY TROPICAL FOREST IN NORTHEASTERN BRAZIL
Eugenia Cordero Schmidt
Orientador: Dr. Eduardo Martins Venticinque
Universidade Federal do Rio Grande do Norte
Coorientador: Dr. Bernal Rodríguez Herrera
Universidad de Costa Rica
Tese apresentada ao Programa de Pós
Graduação em Ecologia da Universidade
Federal do Rio Grande do Norte, como parte do
requerimento para obtenção do título de Doutora
em Ecologia.
Natal, Março 2020

3
Schmidt, Eugenia de Jesus Cordero. Interações, mecanismos de persistência e coexistência demorcegos nectarívoros e das plantas das quais se alimentam emuma floresta tropical sazonal seca no Nordeste brasileiro /Eugenia de Jesus Cordero Schmidt. - Natal, 2020. 160 f.: il.
Tese (Doutorado ) - Universidade Federal do Rio Grande doNorte. Centro de Biociências. Programa de Pós-graduação emEcologia. Orientador: Prof. Dr. Eduardo Martins Venticinque. Coorientador: Prof. Dr. Bernal Rodríguez Herrera.
1. Caatinga - Tese. 2. Chiroptera - Tese. 3. Dieta - Tese. 4.Fenologia - Tese. 5. Pólen - Tese. 6. Reprodução - Tese. I.Venticinque, Eduardo Martins. II. Herrera, Bernal Rodríguez.III. Universidade Federal do Rio Grande do Norte. IV. Título.
RN/UF/BSCB CDU 574
Universidade Federal do Rio Grande do Norte - UFRNSistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial Prof. Leopoldo Nelson - -Centro de Biociências - CB
Elaborado por KATIA REJANE DA SILVA - CRB-15/351

1

AGRADECIMENTOS
Que fase doida, incrível, intensa, pesada e leve. Muito aprendizado, autoconhecimento e muito pra agradecer!
Juank, mi persona favorita! Comenzamos esto juntos y lo terminamos juntos. GRACIAS por tu amor, compañía, comida, cuidados (en los varios problemas aleatorios que me han pasado) y por tu increíble paciencia. GRACIAS por tu fuerte trabajo en campo, las manejadas, las carreras, las embarcadas, por tu compañerismo y por tus fotos INCREIBLES (95% das fotos desta tese foram tiradas por ele!). ¡GRACIAS por equilibrarme y por hacerme reír todos los días! ¡GRACIAS POR EXISTIR!
Familia, ma, pa, Gaby y todos mis hermanos y sobrinos perros. ¡GRACIAS por el apoyo IN-CON-DI-CIO-NAL! GRACIAS por todos los viajes, cenas, cajitas sorpresas por correo… Que bárbaros, sin ustedes y sus miles de ayudas emocionales, espirituales y económicas no hubiera logrado esto. ¡Me hace muy feliz saber que están orgullosos de mi! GRACIAS tíos, primos y abuelitos, me han hecho mucha falta, pero nunca he dejado de sentirlos cerca.
GRACIAS a mis queridos amigos ticos biologeos, Sol, Marce, Cristin (¡MIL GRACIAS SIEMPRE por tu disponibilidad y ayuda para revisar mis cosas!), Jose, Moritzio, Jenny, Kathe, Maggie, Elena y Adri (¡¡da Silva!! Te volviste brasileña! Gracias por el cariño!). A mis queridos termitas peruanas Adri, Bala y Bompe, los extraño muchísimo!
OBRIGADA Bernal e Dadão. Formamos uma equipe de trabalho muito forte e completa, baseada em amizade e carinho. Obrigada por todos esses muitooos anos de trabalho juntos, aprendi muito com vocês e agradeço sua paciência! Bernal, muchas gracias por abrirme la puerta hacia Brasil, donde fui tan bien recibida por Dadāo, los quiero mucho! Ragde e Marina, por associações acadêmicas e conjugais, acabei fazendo duas grandes amigas! Ragde, sos um ser de luz! Marina, mulherão cheia de criatividade e força (vou sentir falta das nossas conversas pós-reunião!!).
OBRIGADA amigos da alma e irmãos brasileiros! Marin (aprendi muito de você, obrigada pela amizade pura), Caro (gracias por las comiditas, risaditas e por ser um exemplo de força!), vocês são mis hermanas favoritas! Pocas (MULHERÃO incrível! Exemplo de pessoa!) Dante (mi amigo, chegou pra nos

6
alegrar a vida mais ainda!), Caramujo (figuraaaaaaaaa! Valeu pelas risadas!), Ricard (tua paz e transparência são muito legais!), Dardo (que falta hacés!!!!!!!!!), Isa (queridaaaaaaaaa você é uma pessoa MUITO legal e única, saudade!!), Juanpi, Tamara, Dri, Guiga, JB, Gustav, Helder, Carol, Dukin saudades dos churrascos e risadas com vocês! OBRIGADA a todos os cachorros que me ajudaram com minha carência e saúde mental, Tucumā obrigada por ter me proporcionado tanta felicidade, Pretis (você é incrível!), Lampião, Judith, Paçoca, Malú e Nina.
OBRIGADA Vermiculitas, vocês viraram elementos fundamentais da minha vida! Obrigada pelas risadas SEMPRE, pelos conselhos e momentos de “lua na sinceridade”. Obrigada pelas constantes manifestações de afeto disfarçadas de bullying (saibam que meu nível de português é intermediário por causa de vocês). Helo obrigada por tua leveza!!!! Estar com você sempre me recarrega com energias positivas e isso é muito difícil de achar! Eli, minha querida leonina, campeã do bullying, professora de yoga, artista, a minha amiga mais descolada... vou sentir falta de te-incomodar e rir com você! Julia, sempre vou te agradecer por ter trazido a Tucumā na minha vida! Você é uma das pessoas mais confiáveis que conheço, corazón peludo pero no mucho! Tenho muito carinho e admiração por todas vocês!
“Lab” 2020 OBRIGADA pelas risadas e conselhos! Foi muito bom compartilhar com vocês os sofrimentos e conquistas!!! OBRIGADA Palito pela força sempre, pelos vários campos juntos com conversas super legais, por me transmitir uma paixão fortíssima pela Caatinga. Você é um pesquisador exemplar e dedicado, te admiro muito! Você vai longe e com certeza vai levar a Caatinga junto! OBRIGADA Virginita pela tua energia sempre legal e leve! Obrigada pelas ajudas em campo, você fez com que essa última fase de coleta em campo fosse muito mais legal e engraçada, obrigada pelas conversas, conselhos e parceria, você é uma pessoa muito especial! Fer, obrigada pelos dados de precipitação, pelas palhaçadas e pelas imitações incrivelmente semelhantes à realidade haha. Continua com essa perseverança e dedicação, você vai longe! Ellen, te conheci pouco, mas o que sei é que você é uma mulher muito forte e perseverante, vai que vai!
OBRIGADA ao Programa de Pós-graduação em Ecologia da UFRN. Agradeço a todos os professores que se tornaram grandes amigos: Carlinhos, Gis, Márcio, Liana, Serginho, Ádrian e Marília (que não foi muito minha professora, mas sim uma amiga muito querida! Vou sentir falta das tretas nas jogatinas!).

7
OBRIGADA ao Laboratório de Investigação de Matrizes Vegetais Energéticas (LIMVE), a Elaine, ao pessoal do Herbário da UFRN. Obrigada ao Laboratório de Micromorfologia Vegetal - LAMIV UEFS, MUITO obrigada Francisco, Paulino, Vivi, Luis, Camila (rainha das Acanthaceae todas!) vocês são proffesionais de primeira e muito queridos!
OBRIGADA aos morcególogos queridos, grandes pesquisadores e pessoas: Enrico, muito obrigada pelo apoio e confiança sempre. Você foi fundamental na nossa caminhada com morcegos no nordeste brasileiro! Eder, Marília, Marianita, Julia, Marlon, Messi, Patrício, Valeria foi sempre um prazer compartilhar e trocar ideias com vocês nos congressos, reuniões e suas respetivas festas!
OBRIGADA meus fiéis companheiros ao longo de este processo todo: Deus, música, praia, meditação, exercício, café e demais plantas que alimentaram meu corpo e espírito!
OBRIGADA Caatinga. Ela foi a minha principal educadora neste enorme processo de aprendizado. Ela me ensinou a ser humilde e respeitosa. Me mostrou paisagens deslumbrantes e imagens duras e inesquecíveis. Me permitiu estudar parte da sua riqueza e me apresentou o povo mais forte, resiliente e generoso que já conheci! OBRIGADA Seu João (meu pai no mato, forte, sorridente e conhecedor!), Darquinha (obrigada pela tua amizade e comidas incríveis), Jusara (<3), Pedrinho (ajuda firme e conversas no mato), dona Das Dores (e seus cafés milagrosos), seu Lorival (e seus abraços e sorrisos apaziguadores), obrigada por me deixarem ser parte da sua família, vocês são incríveis! OBRIGADA dona Creusa, seu Cícero, Laysa e Irene, obrigada por sempre ter um lugar pra nos acolher, comida deliciosa e conversas gostosas!! OBRIGADA a todos nossos companheiros, guias e amigos de campo: Juninho (ainda estou impressionada com tua força!), Edinho (teu sorriso, tua energia positiva e tranquilidade são admiráveis), Valtenci (tuas risadas e histórias sempre surpreendentes! E tua família acolhedora e querida!), Iatagan (outro pai! Obrigada pelo cuidado, conselhos, bolachas, goiabada com açúcar, água gelada, risadas, e por compartilhar sempre teu conhecimento interminável!), Veio, Titico, Mario, Geilson e Geison.
OBRIGADA aos funcionários das Unidades de Conservação que nos acolheram e facilitaram nosso trabalho: Lucia (uma mulher forte, centrada, e muito querida!!!), Rielson (e sua agradável família!), e Leonardo do Parque Nacional Furna Feia; Mauro dos Anjos gestor da FLONA de Açu; George e

8
Geraldo da ESEC do Seridó. Diego Bento (grande parceria!) do Centro de Pesquisa e Conservação de Cavidades (CECAV).
Um agradecimento especial a uma pessoa que se tornou fundamental na reta final do doutorado, Fabiana Lopes (a querida Bia!). Obrigada pela grande confiança depositada em mim e pelo apoio, forca e paciência. Vamos fazer grandes coisas juntas!
OBRIGADA morcegos!! Animais fascinantes, incríveis e interessantíssimos! Obrigada por me permitir invadir sua vida, sua casa e sua privacidade! Amo vocês! Obrigada as mais de 600 pessoas que participaram nas oficinas de educação ambiental com morcegos, obrigada pelo interesse em conhece-los um pouquinho melhor!
A ciência definitivamente não é gasto, é investimento!
O presente trabalho foi realizado com apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Código de Financiamento 001; também com o apoio fundamental do CNPq (Pesquisador Visitante Especial-PVE - Projeto de Ecologia e Conservação de Morcegos na Caatinga Potiguar: 401467 / 2014-7), The Rufford Small Grants Foundation, e finalmente obrigada Wildlife Conservation Society Brasil pelo apoio na logística de campo.
“Se nada ficar destas páginas, algo, pelo menos, esperamos que permaneça: nossa confiança no povo. Nossa fé nos homens e na criação de um mundo em
que seja menos difícil amar”- Paulo Freire.

1 2 3 4
GENERAL
10 About this doctorate
11 What we found
12 General introduction
Chapter I. SHEDDING LIGHT ON NECTARIVOROUS BAT – PLANT NETWORKS:
INTERACTIONS FROM CAATINGA REVEALS HIGHLY GENERALIZED
ASSOCIATIONS AND TEMPORAL STABILITY
19 Abstract
21 Introduction
23 Methods
29 Results
30 Discussion
35 References
47 Figures and tables
52 Supporting information
Chapter II. MECHANISMS MEDIATING NECTAR-FEEDING BATS COEXISTENCE AND
PERSISTENCE THROUGH THE SEASONAL RHYTHM OF CAATINGA
64 Abstract
66 Introduction
69 Methods
76 Results
82 Discussion
93 References
105 Figures and tables
115 Supporting information
Chapter III. FEMALE REPRODUCTION PATTERNS IN CAATINGA ARE INFLUENCED
BY PRECIPITATION AND CACTACEAE RESOURCE AVAILABILITY
120 Abstract
122 Introduction
125 Methods
129 Results
134 Discussion
139 References
146 Figures and tables
151 Supporting information
SUMMARY
158 What we learned about nectar-feeding bats in Caatinga

10
5
Coletamos os dados principalmente no município de Lajes, no estado do Rio Grande do Norte, mas para representar melhor a heterogeneidade da Caatinga desse pouco explorado estado, incluímos outros seis municípios (amostrados em menor escala). Coletamos dados em sete tipos diferentes de habitat: 1) Carnaubais, 2) Enclaves de floresta úmida, 3) Caatinga média, 4) Pomares, 5) Caatinga ripária, 6) Afloramentos rochosos e 7) Caatinga arbustiva.
Em diferentes períodos entre maio de 2015 e fevereiro de 2019, cobrindo a sazonalidade tão característica da Caatinga.
Usamos redes de neblina para capturar os morcegos nectarívoros. Anotamos: a hora de captura, espécie, sexo, medidas eco-morfológicas (peso, antebraço, focinho e asas). Coletamos dois tipos de amostras: fezes e pólen de seus pelos (com cubos de gelatina glicerinada). Para caracterizar a dieta das espécies: identificamos os tipos de pólen e itens alimentares complementares ao néctar. Também coletamos dados fenológicos de floração de nove espécies de árvores, arbustos e cactos para saber quais recursos estavam disponíveis para os morcegos e quando.
Nos aprofundamos na vida de quatro espécies de morcegos nectarívoros: Glossophaga soricina (Phyllostomidae: Glossophaginae), Lonchophylla inexpectata, Lonchophylla mordax e Xeronycteris vieirai (Phyllostomidae: Lonchophyllinae). Descrevemos suas redes de interações com plantas (capítulo 1), exploramos seus mecanismos de persistência e coexistência ao longo do tempo e estações do ano (capítulo 2) e, finalmente, descrevemos os padrões de reprodução dos morcegos nectarívoros (capítulo 3).

11
6
Anoura geoffroyi, G. soricina, Linexpectata, L. mordax e X. vieirai interagiram com 31 espécies de plantas. A rede mostrou um padrão altamente generalizado de interações, consistente ao longo das estações e anos. Os morcegos apresentaram alta sobreposição nas interações, o que contribuiu para a generalização ecológica (baixa especialização e modularidade) e padrão não aninhado. Os padrões de interação generalizados podem ser uma condição necessária para a persistência de morcegos nectarívoros e de suas plantas especializadas em ambientes variáveis como a Caatinga.
Glossophaga soricina, L. inexpectata, L. mordax e X. vieirai coexistem e persistem ao longo do tempo (anos e estações) mediados por uma mistura de mecanismos: 1) Particionamento temporal (diferenças nas capturas ao longo da noite). 2) Diferenças eco-morfológicas (peso, antebraço, focinho e proporções das asas). 3) Particionamento de recursos (flexibilidade da dieta consumindo pelo menos 20 espécies de plantas além de pólen, insetos e tecidos vegetais). A persistência temporal dos morcegos foi facilitada pela continuidade e complementaridade dos recursos florais disponíveis ao longo do ano. No entanto, as Cactaceae são um recurso fundamental na Caatinga, espécies de cactos foram usadas em alta frequência por todos os morcegos.
Dois padrões reprodutivos foram observados para as espécies de morcegos nectarívoros na Caatinga, padrão bimodal em G. soricina e aparentemente em L. inexpectata (poucos dados reprodutivos) e multimodal em L. mordax e X. vieirai. A precipitação afetou positivamente a probabilidade de ocorrência de gravidez em L. mordax e X. vieirai. Nenhuma das variáveis testadas influencia a gravidez de G. soricina. A disponibilidade de recursos florais de Cactaceae do mês anterior, afetaram positivamente a probabilidade de ocorrência de lactação para todos os morcegos nectarívoros, com algumas diferenças para determinadas espécies de cactos.

12
7 GENERAL INTRODUCTION 8
Seasonally dry tropical forest (SDTF) are considered a harsh environment 9
due to their high temperature, scarce and variable rainfall that cause seasonal 10
restrictions on the availability of water and food resources (Dirzo et al., 2011). 11
SDTFs are distribuited in disjunct patches throughout the world. They vary greatly 12
physiognomically influenced by local climate, soil and topography (Pennington et 13
al., 2009) which has hampered the overall definition of SDTFs and the 14
delimitation of the biomes and ecosystems within them. In Neotropics the majority 15
of SDTFs occurs in South America with the largest and most continuous 16
fragments in Bolivia and Brazil (Sánchez-Azofeifa and Portillo-Quintero, 2011). 17
Animals and plants that occur in SDTFs developed adaptations to deal 18
with these conditions. For example, most of the plants are facultative or obligatory 19
deciduous species. Thorny species are abundant and their flowering and fruiting 20
phenologies are markedly seasonal with a shorter growing period (Murphy and 21
Lugo, 1986; Pennington et al., 2000). In the case of mammals, several 22
mechanisms about how they cope with and survive in SDTFs were synthesized 23
by Stoner and Timm (2011). Broadly, they divided the mechanisms in 24
physiological (changes in body temperature, seasonal torpor or hibernation, 25
water conservation, and delayed reproduction), and behavioral adaptations 26
(dietary flexibility, long and short distance migrations, activity patterns, 27
seasonality of reproduction). 28
It has been proven that bats can occupy arid and semiarid forests mainly 29
due to long distance migrations strategies and short distance habitat shifts 30
(Fleming et al., 2003), diet flexibility (Heithaus et al., 1975; Soriano et al., 1991; 31
Tschapka et al., 2008) and presenting a close relationships with keystone plants 32
which are regular resources, such as cactus and agaves (Sosa and Soriano 33
1996; Valiente-Banuet et al., 1996, 1997, Stoner et al., 2003). 34
Nectar-feeding bats are found frequently in dry areas and are the main 35
agents of pollination for hundreds of plant species, some of which are completely 36

13
dependent on them for reproduction (Bolzan et al., 2015). They are known to 37
pollinate over 528 species of plants worldwide (Kunz et al., 2011) and are 38
responsible for the pollination of 13.1% of the plants present in a Caatinga region 39
(Machado and Lopes, 2004). This percetage is considered a high when 40
compared to other ecosystems ( 3% in Cerrado and humid forests Silberbauer-41
Gottsberger and Gottsberger,1988; Bawa et al., 1985). 42
Caatinga is the largest continuous SDTF area in northeastern Brazil, with 43
around 800,000 km2, covering 11% of the country (Silva et al., 2018). It is a 44
mosaic of vegetation types, ranging from habitats with non-thorny arboreal 45
habitats to xeric habitats, where thorny shrubs, cactus and bromeliads 46
predominate (Andrade-Lima, 1981). Caatinga is the most populated semiarid 47
region in the world, harboring over 28.6 million people (Silva et al., 2018). 48
Unfortunately, Caatinga experience gradual but persistent degradation 49
processes (Ribeiro et al., 2015). Caatinga harbors a high diversity and endemism 50
in both flora and fauna with outstanding adaptations to the extreme abiotic 51
conditions (Coe and de Sousa, 2014; Ribeiro et al., 2015). It is home to about 52
5000 species of angiosperms, 300 of which are endemic (Giulietti et al., 2004) 53
and 153 species of mammals with ten endemic species (Paglia et al., 2012; 54
Gutierrez and Marinho-Filho, 2017). More than a half of the mammalian diversity 55
in the Caatinga is represented by bats, a total of 90 species (59% of all mammals 56
occurring in the Caatinga) have been recorded of which nine are known to be 57
nectar-feeding bats (belonging to Glossophaginae and Lonchophyllinae 58
subfamilies) (Carvalho-Neto et al., 2016; Gutierrez and Marinho-Filho, 2017). 59
In this thesis I seek to understand the ecology of the ensemble of 60
nectarivorous bats and their persistence, coexistence and reproduction patterns 61
in the northeastern part of Caatinga, at the Rio Grande do Norte (RN) state. In 62
the first chapter I describe how the network of interactions between nectarivorous 63
bats and the plants they feed on is structured and evaluate its temporal dynamics 64
across seasons. In the second chapter, I test for three possible coexistence 65
mechanisms for the nectar-feeding bats: 1) Temporal partitioning 2) 66
Ecomorphological differences and 3) Resource partitioning in a Caatinga 67
fragment in the municipality of Lajes. In the third chapter, I described the female 68
reproductive patterns of nectar-feeding bats and inspect the effects of 69
precipitation and Cactaceae resource availability. 70

14
71 REFERENCES 72
Andrade-Lima D (1981) The caatinga dominium. Rev Bras Bot 4:149–153 73 Bawa, K. S., Perry, D. R., & Beach, J. H. (1985). Reproductive biology of 74
tropical lowland rain forest trees. I. Sexual systems and incompatibility 75 mechanisms. American journal of botany, 72(3), 331-345. 76
Bolzan, D. P., Pessôa, L. M., Peracchi, A. L., & Strauss, R. E. (2015). 77 Allometric patterns and evolution in Neotropical nectar-feeding bats (Chiroptera, 78 Phyllostomidae). Acta Chiropterologica, 17(1), 59-73. 79
Carvalho-Neto, F. G., da Silva, J. R., Santos, N., Rohde, C., Garcia, A. C. 80 L., & Montes, M. A. (2016). The heterogeneity of Caatinga biome: an overview of 81 the bat fauna. Mammalia, 81(3), 257-264. 82
Coe, H. H. G., & Sousa, L. O. F. (2014). The Brazilian" Caatinga": ecology 83 and vegetal biodiversity of a semiarid region. Dry Forests: Ecology, Species 84 Diversity and Sustainable Management, 1, 81-103. 85
Dirzo, R., Young, H. S., Mooney, H. A., & Ceballos, G. (2011). Seasonally 86 dry tropical forests: ecology and conservation. Island Press. 87
Fleming, T. H., Eby, P., Kunz, T. H., & Fenton, M. B. (2003). Ecology of 88 bat migration. In: Bat ecology, 156, 164-65. 89
Giulietti, A. M., Bocage Neta, A. L., Castro, A. A. J. F., Gamarra-Rojas, C. 90 F. L., Sampaio, E. V. S. B., Virgínio, J. F., ... & Harley, R. M. (2004). Diagnóstico 91 da vegetação nativa do bioma Caatinga. Biodiversidade da Caatinga: áreas e 92 ações prioritárias para a conservação. 93
Gutiérrez, E. E., & Marinho-Filho, J. (2017). The mammalian faunas 94 endemic to the Cerrado and the Caatinga. ZooKeys, (644), 105. 95 Heithaus, E. R., Fleming, T. H., & Opler, P. A. (1975). Foraging patterns and 96 resource utilization in seven species of bats in a seasonal tropical forest. Ecology, 97 56(4), 841-854. 98
Kunz, T. H., Braun de Torrez, E., Bauer, D., Lobova, T., & Fleming, T. H. 99 (2011). Ecosystem services provided by bats. Annals of the New York Academy 100 of Sciences, 1223(1), 1-38. 101
Machado, I. C., & Lopes, A. V. (2004). Floral traits and pollination systems 102 in the Caatinga, a Brazilian tropical dry forest. Annals of Botany, 94(3), 365-376. 103 Murphy, P. G., & Lugo, A. E. (1986). Ecology of tropical dry forest. Annual review 104 of ecology and systematics, 17(1), 67-88 105
Paglia, A. P., Da Fonseca, G. A., Rylands, A. B., Herrmann, G., Aguiar, L. 106 M., Chiarello, A. G., ... & Mendes, S. L. (2012). Lista Anotada dos Mamíferos do 107 Brasil 2ª Edição/Annotated Checklist of Brazilian Mammals. Occasional papers 108 in conservation biology, 6, 1-82. 109
Pennington, T. R., Prado, D. E., & Pendry, C. A. (2000). Neotropical 110 seasonally dry forests and Quaternary vegetation changes. Journal of 111 Biogeography, 27(2), 261-273. 112

15
Pennington, R. T., Lavin, M., & Oliveira-Filho, A. (2009). Woody plant 113 diversity, evolution, and ecology in the tropics: perspectives from seasonally dry 114 tropical forests. Annual Review of Ecology, Evolution, and Systematics, 40, 437-115 457. 116
Ribeiro, E. M., Arroyo-Rodríguez, V., Santos, B. A., Tabarelli, M., & Leal, 117 I. R. (2015). Chronic anthropogenic disturbance drives the biological 118 impoverishment of the Brazilian Caatinga vegetation. Journal of Applied 119 Ecology, 52(3), 611-620. 120
Rojas-Martínez, A., Valiente-Banuet, A., Del Coro Arizmendi, M., 121 Alcántara-Eguren, A., & Arita, H. T. (1999). Seasonal distribution of the long-122 nosed bat (Leptonycteris curasoae) in North America: does a generalized 123 migration pattern really exist?. Journal of Biogeography, 26(5), 1065-1077. 124
Silva, J. M. C., Leal, I. R., & Tabarelli, M. (Eds.). (2018). Caatinga: the 125 largest tropical dry forest region in South America. Springer. 126
Soriano, P. J., Sosa, M., & Rossell, O. (1991). Hábitos alimentarios de 127 Glossophaga longirostris Miller (Chiroptera: Phyllostomidae) en una zona árida 128 de los Andes venezolanos. Rev. Biol. Trop, 39(2), 263-268Sosa, M., & Soriano, 129 P. J. (1993). Solapamiento de dieta entre Leptonycteris curasoae. Rev. Biol. 130 Trop, 41(3), 529-532. 131
Sánchez-Azofeifa, G. A., & Portillo-Quintero, C. (2011). Extent and drivers 132 of change of Neotropical seasonally dry tropical forests. In Seasonally Dry 133 Tropical Forests (pp. 45-57). Island Press, Washington, DC. 134
Silberbauer-Gottsberger, I. and Gottsberger, G. (1988). A polinização de 135 plantas do cerrado. Revista Brasileira de Biologia, vol. 48, p. 651-663. 136
Silva, J. M. C., Leal, I. R., & Tabarelli, M. (Eds.). (2018). Caatinga: the 137 largest tropical dry forest region in South America. Springer. 138
Stoner, K. E., Karla, A. S., Roxana, C. F., & Quesada, M. (2003). 139 Population dynamics, reproduction, and diet of the lesser long-nosed bat 140 (Leptonycteris curasoae) in Jalisco, Mexico: implications for conservation. 141 Biodiversity & Conservation, 12(2), 357-373. 142
Stoner, K. E., & Timm, R. M. (2004). Tropical dry-forest mammals of Palo 143 Verde: Ecology and conservation in a changing landscape. University of 144 California Press, Berkeley. 145
Sosa, M., & Soriano, P. J. (1993). Solapamiento de dieta entre 146 Leptonycteris curasoae. Rev. Biol. Trop, 41(3), 529-532. 147
Stoner, K. E., & Timm, R. M. (2011). Seasonally dry tropical forest 148 mammals: Adaptations and seasonal patterns. In Seasonally Dry Tropical 149 Forests (pp. 85-106). Island Press, Washington, DC. 150
Tschapka, M., Sperr, E. B., Caballero-Martínez, L. A., & Medellín, R. A. 151 (2008). Diet and cranial morphology of Musonycteris harrisoni, a highly 152 specialized nectar-feeding bat in western Mexico. Journal of Mammalogy, 89(4), 153 924-932. 154
van Schaik, C. P., Terborgh, J. W., & Wright, S. J. (1993). The phenology 155 of tropical forests: adaptive significance and consequences for primary 156 consumers. Annual Review of ecology and Systematics, 24(1), 353-377 157

16
Valiente-Banuet, A., Arizmendi, M. D. C., Rojas-Martínez, A., & 158 Domínguez-Canseco, L. (1996). Ecological relationships between columnar cacti 159 and nectar-feeding bats in Mexico. Journal of Tropical Ecology, 12(1), 103-119. 160
Valiente-Banuet, A., Rojas-Martínez, A., Arizmendi, M. D. C., & Dávila, P. 161 (1997). Pollination biology of two columnar cacti (Neobuxbaumia mezcalaensis 162 and Neobuxbaumia macrocephala) in the Tehuacan Valley, central 163 Mexico. American Journal of Botany, 84(4), 452-455. 164 165 166

17
167

18
168 169
Shedding light on nectarivorous bat-plant networks: interactions from 170
Caatinga reveals highly generalized associations and temporal stability 171
172
Eugenia Cordero-Schmidt1, Pietro Kiyoshi Maruyama2, Juan Carlos Vargas-173 Mena1, Paulino Pereira Oliveira3, Francisco de Assis R. Santos3, Rodrigo A. 174 Medellín4, Bernal Rodriguez-Herrera5 and Eduardo M. Venticinque1 175 176 177 1Departamento de Ecologia, Universidade Federal do Rio Grande do Norte, 178 59078900 Lagoa Nova Natal, RN, Brazil; E-mail: [email protected] 179 2 Centro de Síntese Ecológica e Conservação, Departamento de Genética, 180 Ecologia e Evolução - ICB, Universidade Federal de Minas Gerais, Belo 181 Horizonte, MG, Brazil. 182 3 Laboratório de Micromorfologia Vegetal, Universidade Estadual de Feira de 183 Santana, 44036-900, Novo Horizonte, BA, Brazil 184 4 Instituto de Ecología, Universidad Nacional Autónoma de México, 70-275, 185 04510 México D. F., México 186 5 Escuela de Biología, Universidad de Costa Rica, 2060 Montes de Oca, San 187 José, Costa Rica 188 189

19
ABSTRACT 190
Seasonally Dry Tropical Forests have low and variable precipitation regimes 191 causing a seasonal pattern in the supply of feeding resources, controlling species 192 composition and plant–animal interactions. Vertebrates, especially nectarivorous 193 bats are important pollinators in STDFs and other tropical ecosystems, including 194 the Brazilian Caatinga. Nevertheless, community-level interaction network 195 studies considering nectarivorous bats are scarce in the literature, which hinders 196 a more comprehensive understanding of plant-pollinator interactions. Here, we 197 describe a nectarivorous bat-plant interaction network from the Caatinga and 198 evaluated its temporal dynamics across seasons and years, considering the 199 highly variable environmental conditions in this ecosystem. To do so, we mist-200 netted nectarivorous bats from 2015 to 2019, sampled pollen loads on their 201 bodies and built a pollen transportation network between bats and plants. Five 202 species of nectar-feeding bats interacted with 31 plant species. This network 203 showed a highly generalized pattern of interactions, which was consistent across 204 repeated seasons and years. Most commonly sampled nectarivorous bat species 205 showed high levels of interaction overlap, which contributed to ecological 206 generalization (low specialization and modularity) and lack of nestedness. 207 Specialized chiropterophilous plants and plants that do not show specialized bat 208 pollination traits were equally important components of the interaction network. 209 The ecologically and phenotypically generalized interaction patterns found may 210 be a necessary condition for the persistence of nectarivorous bats and their 211 specialized plants in the environmentally harsh and variable environments of 212 Caatinga. Studying neglected groups of pollinators and their interaction networks 213 may shed new insights into our general understating of plant-pollinator 214 relationships, and more studies considering bats and other nocturnal pollinators 215 need to be conducted. Specifically, the underappreciated generalization on 216 interaction with plants call for studies testing for the effectiveness of bats in 217 pollinating specialized and non-specialized plants they visit. 218 219 220 Key words: Chiropterophily; Pollen transport; Pollination; Pollination syndrome; 221
Seasonal Dry Tropical Forest; Temporal variation 222
223

20
GRAPHICAL ABSTRACT 224

21
INTRODUCTION 225
Many vertebrates are key pollinators, especially birds and bats (Fleming & 226
Muchhala 2008; Ratto et al. 2018). Recently, the field of pollination ecology has 227
greatly benefited from the use of interaction network approaches when 228
characterizing plant and pollinator interactions at the community level (Knight et 229
al. 2018). Despite many advances that the interaction networks theory and 230
methods brought, the literature in plant-pollination networks has shortcomings, 231
for instance, neglecting some functional group of pollinators from tropical regions 232
(Vizentin-Bugoni et al. 2018). Bats are involved in frugivory and nectarivory 233
interactions with plants throughout the Neotropics (Mello et al. 2019). However, 234
bat-plant pollination interactions have been poorly explored, especially when 235
compared to birds which have been more frequently studied using the network 236
approach (Zanata et al. 2017). Bat-flower interactions at the community scale are 237
largely absent in the network literature (Vizentin-Bugoni et al. 2018). To our 238
knowledge, only two recent studies have evaluated Old-World nectarivorous bats 239
and their interaction networks (Stewart & Dudash 2017; Sritongchuay, Hughes & 240
Bumrungsri, 2019). However, Old-World (Pteropodidae) and New-World 241
nectarivorous bats (Phyllostomidae) have different evolutionary histories, even 242
though they show many convergent traits and ecological similarities (Fleming, 243
Geiselman & Kress, 2009). 244
Conversely, bat pollination has evolved independently many times in 245
different lineages of tropical and subtropical plants (Fleming, Geiselman & Kress, 246
2009), and includes hundreds of ecological and economically important plant 247
species (Kunz, Braun de Torrez, Bauer, Lobova & Fleming., 2011). Nectar-248
feeding bats are typically associated with large robust flowers or inflorescences, 249

22
with light-colored flowers with long and numerous stamens, copious amounts of 250
nectar, large quantities of pollen, and sometimes producing unpleasant odors that 251
attract nectarivorous bats (Tschapka & Dressler, 2002). This set of floral traits 252
characterizes the pollination syndrome of chiropterophily (Fægri & van der Pijl 253
1979), and specialized bat-pollinated plants seem to be especially dependent on 254
their pollinators for sexual reproduction (Ratto et al. 2018). 255
Considering the importance of bats as pollinators in the Tropics, and 256
especially in some ecosystems, studies on nectarivorous bat-plant networks 257
could bring new insights for the understanding of plant-pollinator interactions. 258
Seasonally Dry Tropical Forests (SDTF) are endangered ecosystems (Janzen 259
1988) with a remarkable diversity of lifeforms and functional groups of both flora 260
and fauna (Dirzo, Young, Mooney & Ceballos, 2011). Low and variable rainfall 261
patterns in SDTFs means that in most cases plant-related resources are found in 262
temporal pulses (Chesson et al. 2004). This seasonal pattern of resource 263
availability seems to be a key factor controlling species composition and therefore 264
influencing niche dynamics and coexistence patterns of consumer populations 265
(Kneitel & Chase, 2004). Despite this irregularity, plants from STDFs present high 266
levels of dependence on biotic vectors for their pollination and seed dispersal 267
(Quesada et al. 2009; Leal, Lopes, Machado & Tabarelli, 2017), indicating a tight 268
plant-animal relationship controlling ecosystem dynamics. 269
Caatinga is the largest SDTF ecosystem in South America with over 800 270
000 km2 located in northeastern Brazil, which harbors a rich biota with high level 271
of endemism (Silva, Leal & Tabarelli, 2018). Here, the proportion of bat-pollinated 272
plants in communities are outstandingly high, representing 11-13.1% of the flora; 273
which is much higher than in other tropical plant communities (Machado & Lopes, 274

23
2004; Quirino & Machado, 2014). Caatinga is home to 9 nectar-feeding bat 275
species including two endemic species (Moratelli & Dias, 2015; Carvalho-Neto et 276
al. 2017), but information about their interactions with plants is scarce (Cordero-277
Schmidt et al. 2017; Silva, Neves, Guedes, Almeida & Brasil-Sato, 2019). 278
In this study, we describe community-level nectarivorous bat-plant 279
interaction networks from the Neotropics and evaluate its dynamics across time 280
in two scales: (1) seasons and (2) years. Because of marked seasonality, plant–281
animal interactions in SDTFs are probably much more sensitive to phenological 282
patterns, including potential changes in the structure of interactions networks 283
(Souza et al. 2018). Although studies evaluating temporal dynamics in pollination 284
networks are not very common, previous studies with bat (Sritongchuay et al. 285
2019) and multi-taxa pollination networks (Souza et al. 2018) from seasonal 286
tropical areas have shown that interaction networks are more specialized in the 287
dry season when floral resources are less abundant. Hence, we also expected 288
variation according to seasons in bat-plant networks from Caatinga, with higher 289
specialization in the dry season. In contrast, pollination networks may show 290
surprisingly similar structural properties from year to year (Alarcón et al. 2008), 291
and we expected our networks to also maintain the same general structural 292
patterns across the years. Finally, considering the high number of 293
chiropterophilous plants in Caatinga (Machado & Lopes 2004), we expected that 294
bats would rely on their diet primarily on the species of plants with 295
chiropterophilous syndrome characteristics (Fleming et al. 2009). 296
297
METHODS 298
Study site 299

24
We conducted the study in the state of Rio Grande do Norte, Brazil, with sampling 300
mostly conducted in the municipality of Lajes, and also in six other municipalities: 301
Cerro Corá, Assu, Felipe Guerra, Baraúna, Martins and Serra Negra do Norte 302
(Supporting Information Figure S1). We spread our samples across these 303
different municipalities to encompass the heterogeneity of Caatinga vegetation, 304
composed of seven habitat types (Supporting Information Table S1, Figure S2): 305
Riparian (7 sampling sites), Shrubby Caatinga (7), Medium Caatinga (6), Rocky 306
outcrops (3), Copernicia groves (2), Humid forest enclave (1), and Orchards (1). 307
For surveyed sites we recorded general environmental characteristics and noted 308
the dominant plant species to classify them into habitat types described by Mares, 309
Willig, Streilein & Lacher (1981) and Prado (2003) (Supporting Information Figure 310
S2 and Table S3). 311
The climate type in the region is semi-arid (Köppen’s BSh), with the wet 312
season lasting approximately six months from January to June and the dry 313
season from July to December (following Oliveira, Silva & Lima, 2017, Northern 314
semiarid subregion). We collected data during two periods, between May and 315
October 2015, and between May 2017 and June 2019. 316
Nectar-feeding bat captures 317
We sampled bats for 151 nights, placing 80 m to 120 m of mist nets per night 318
over six hours from sunset to midnight. All mist nets were placed at ground level 319
and along preexisting trails and near flowering plants or known/ suspected roosts. 320
The total netting effort was 310.8925 m2h (Straube & Bianconi, 2002). We 321
identified the species in the field using systematic keys and species diagnosis 322
(Díaz, Solari, Aguirre, Aguiar & Barquez, 2016). All bats were released after data 323
collection except for the individuals that were impossible to identify in the field or 324

25
that represented a new report for the area, which were deposited in the Adalberto 325
Varela Mammalogy Collection (CMAV) of the Universidade do Rio Grande do 326
Norte. All procedures for capture, handling and collection of bats met the 327
guidelines of the American Society of Mammalogists for the use of wild mammals 328
in research (Sikes et al., 2016), and the legal Brazilian requirements of 329
conservation and animal welfare. Fieldwork was authorized by 330
MMA/ICMBio/SISBIO under permits 48325-2. To recognize recaptured 331
individuals during the same sampling period, bats were marked by cutting a small 332
portion of fur (1 cm) from the lower back of each individual with a curved 333
dissecting scissor. 334
Pollen collection and identification 335
We collected pollen samples from each captured bat’s fur, wings, legs and 336
uropatagium using a single glycerin jelly cube (3-4 mm) per individual, which was 337
later mounted on glass microscope slides. Jelly were prepared with glycerin and 338
phenol for preservation and safranin for staining (Voigt, Kelm, Bradley & 339
Ortmann, 2009). To avoid cross-contamination from one sample to another, 340
special care was taken by cleaning the worktable, hands and tweezers with 341
alcohol after finishing the handling of each bat. Besides, the cloth bags in which 342
the bats were held after capture were always clean and never reused until they 343
were washed. 344
We identified pollen under a light microscope (magnification 400-1000x, 345
Leica DM500) to the lowest possible taxonomic level through comparison with 346
reference collections. To identify the pollen types, we used a collection 347
specialized on the pollen of the Caatinga vegetation from the Palynotheca of the 348
LAMIV, UEFS (Plant Micromorphology Laboratory, Universidade Estadual de 349

26
Feira de Santana), and a reference collection that we made from local plants in 350
bloom during our fieldwork. We also used pollen catalogs for additional reference 351
(Palacios-Chávez, Ludlow-Wiechers & Villanueva, 1991; Roubik & Moreno 1991; 352
Santos, Watanabe & Hamburgo-Alves, 1997; Carreira & Barth 2003; Melhem et 353
al. 2003; Silva, Santos & Lima, 2016). 354
We scored the presence/absence of each pollen type on individual bats 355
and used the number of times each pollen type was found in a specific bat 356
species as a measure of interaction frequency (Sazatornil et al. 2016). To 357
minimize the effects of possible contaminations, i.e., pollen grains placed in the 358
flowers by other pollinators before the visit of the bats, we established a minimum 359
number of pollen grains according to size categories (following Heithaus, Fleming 360
& Opler, 1975). In the present study, we established three criteria based on pollen 361
size and count (Supporting Information Table S4). 362
We then gathered information from the literature for the following plant and 363
floral traits to characterize the floral biology and pollination syndromes of plants 364
sampled: Flowering season, flower opening (day, night or crepuscular), blossom 365
class (according Fægri & van der Pijl, 1971), corolla color, flower size, flower size 366
categories (according to Machado & Lopes, 2004), and pollen size (according to 367
Erdtman, 1952) (Supporting Information Table S5). 368
Network analyses 369
We built networks for seasons (dry - July to December, wet - January to June), 370
years of sampling (2015, 2017, 2018, each year from June to October) and only 371
for the municipality of Lajes. We calculated four network metrics that characterize 372
distinct aspects of the interaction patterns. We calculated (1) connectance, the 373
ratio of observed and possible links in the network, which is an estimate of how 374

27
interactions are constrained within the community (Jordano, 1987); (2) 375
complementary specialization (H2') that measures how species restrict their 376
interactions from those randomly expected based on the availability of partners 377
(Blüthgen et al. 2006); (3) nestedness using NODF index (Almeida-Neto, 378
Guimaraes, Guimaraes, Loyola & Ulrich, 2008), which quantifies how interactions 379
of specialized species are subsets of the interactions of the more generalist 380
species in the networks; (4) modularity (Q) using the DIRTLPA+ algorithm 381
(Beckett 2016), which quantifies the tendency of species forming subgroups of 382
interacting species. Due to the stochastic nature of the optimization algorithm, we 383
repeated the analysis 30 times and kept the highest Q value as the optimal 384
solution (Beckett, 2016). To assess the significance of these network-level 385
metrics, we contrasted the observed values to those generated by null models. 386
For NODF, we used the r1 model from the vegan package, which uses the 387
column (bats) marginal frequencies as probabilities to distribute the presence of 388
interactions (Oksanen et al. 2019). For quantitative indices, including H2' and Q, 389
we used the r2dtable model from the bipartite package (Dormann et al. 2008) that 390
constrains the marginal totals and network size. We considered a metric 391
significant by assessing whether the observed value was greater than 95% of the 392
simulated values from the null models (p<0.05). 393
We also calculated three species-level indices from the interaction 394
network: (1) plant degree, which is the number of bat species each plant species 395
interacted with; (2) species strength, quantified as the sum of the proportions of 396
interactions performed by a given species across all its interacting partners, 397
measuring the extent to which bat species depend on a specific plant species 398
(Bascompte, Jordano & Olesen, 2006); (3) species-level specialization d', which 399

28
quantifies how interaction frequencies of a given species deviate concerning the 400
availability of interaction partners in the network, defined by their marginal totals 401
(Blüthgen, Menzel & Blüthgen, 2006). Calculations of all network-related indices 402
were conducted with the bipartite package version 2.13 (Dormann, Gruber & 403
Fruend, 2008). We used these indices to contrast plants showing characteristics 404
in accordance with chiropterophily syndrome, to those showing characteristics 405
associated with other pollination syndromes, e.g., diurnal flower opening. We 406
used the three species level indices as response variables in three distinct 407
Generalized Linear Models (GLMs) for each index, with a categorical predictor 408
assigning plant species as chiropterophilous or not. We assumed a Poisson 409
distribution of data for the GLM with degree, and Gaussian distribution for GLMs 410
with species strength and specialization (Zuur et al. 2009). We only used the 411
pollen types that were identified to species level in this analysis. To attain 412
significance values for each model, we used the function Anova in car package 413
version 3.0-4 (Fox and Weisberg, 2011). 414
Finally, we also estimated the sampling completeness of the networks 415
(Chacoff et al. 2012; Vizentin-Bugoni et al. 2016). We took each combination of 416
a bat and plant species as an equivalent of ‘species’ and the frequency of each 417
pairwise interaction as their ‘abundances’, and then calculated the ratio between 418
observed and estimated interaction diversity (Vizentin-Bugoni et al. 2016). We 419
used the Chao 1 estimator from the iNEXT package to estimate the richness of 420
pairwise interactions from the observed data (Hsieh, Ma & Chao, 2014). All 421
analysis was conducted in R, version 3.6.1 (R Core Team, 2019). 422
423
RESULTS 424

29
Nectar-feeding bats 425
Five species of nectar-feeding bats were captured (Figure 1), with a total of 650 426
captures (Supporting Information Table S6). Glossophaga soricina was the most 427
abundant (n= 252) and found in most localities (L=6) and Habitat Types (HT=6) 428
during both seasons, followed by Lonchophylla mordax (n= 173 L=5, HT=4). The 429
two species that had the lowest occurrence were Xeronycteris vieirai (n= 134, 430
L=1, HT= 2) and Lonchophylla inexpectata (n= 103, L=1, HT=2); while Anoura 431
geoffroyi was the least abundant (n= 6, L=2, HT=2) during both seasons. 432
Bat-plant interactions 433
We collected a total of 608 glycerin jelly samples, from which 121 did not contain 434
any pollen grain. From the remaining 487 samples, 54% belonged to the dry 435
season and 46% to the rainy season (A. geoffroyi n= 5; G. soricina n=178; L. 436
mordax n= 137; L. inexpectata n= 76; X. vieirai n=91). A total of 31 pollen types 437
(Figure 2) were recorded in the bats from 19 genera and 14 plant families (Figure 438
3). In average, we recorded 2.56 ± 1.70 pollen types (min. = 1, max. = 10), per 439
sample. Cactaceae (eight pollen types) was the most frequent plant family, 440
followed by Acanthaceae and Fabaceae (four each). We were unable to identify 441
three pollen types; however, they were only rarely recorded (1.6% of samples). 442
Three additional pollen types were identified only up to the family level (Fabaceae 443
type, Sapindaceae type and Malpighiaceae type), and two pollen types up to the 444
genus level (Pseudobombax type, and Ipomoea type). 445
The interaction network comprised of 1250 interactions between bat and 446
plant species (pollen types). The overall network showed a highly generalized 447
interaction pattern, with a high value of connectance, and low but significant 448
values of specialization and modularity. Thus, although there is a non-random 449

30
pattern on how interactions are organized, there is very little partitioning of 450
interactions between species. Moreover, because most bats are equally 451
generalist, we did not find significant nestedness in the network (Table 1). The 452
two seasonal networks comprised a slightly smaller number of plant species (26 453
in the dry and 27 in the rainy season), indicating some but not strong seasonality 454
in flowering species richness. These networks showed similar structure to the 455
overall network, and no strong variation between the seasons (Table 1). 456
Moreover, there was no strong variation among the distinct years of sampling, 457
and the network considering only the most sampled locality, Lajes, showed a 458
similar pattern as the overall network (Table 1). 459
When comparing chiropterophilous plants to other plants used by bats, 460
there was no difference in any of the three species level indices (Figure 4). 461
Finally, we found that our networks were all reasonably well sampled, with 462
sampling completeness from 65% to 87% (Table 1). 463
464
DISCUSSION 465
The nectarivorous bat-plant network from Caatinga showed a highly generalized 466
pattern of interactions, which was consistent across the seasons and years. Most 467
commonly sampled nectarivorous bat species showed high levels of interaction 468
overlap, which contributed to ecological generalization (low specialization and 469
modularity) and lack of significant nestedness. Some plant species that do not 470
show characteristics in accordance with chiropterophily syndrome were equally 471
important in the interaction network to those that were more specialized. This 472
implies bats may also be generalists regarding floral traits of the plants they 473

31
interact with, characterizing these pollinators as phenotypic and functional 474
generalists (Ollerton, Killick, Lamborn, Watts & Whiston, 2007; Armbruster 2017). 475
Generalization in pollination interactions is regarded as conferring 476
resilience to both plants and pollinators to fluctuation on their respective 477
resources (Waser et al. 1996). Flowering phenology in Caatinga is highly 478
variable, dependent on somewhat unpredictable rainfall (Machado & Barros, 479
1997), hence generalization may be favored. Within bats, generalist species may 480
opportunistically switch their diet temporarily according to availability of mass 481
flowering plants, in contrast to more specialist ones that remain loyal to plants 482
with longer flowering periods (Stewart & Dudash 2018). When considering the 483
five most important plants for each nectar-feeding bat (>10% of samples with 484
pollen type), we found all bats interacting with plants showing distinct flowering 485
phenologies, i.e. both plants with continuous and short flowering pulses. So, 486
nectarivorous bats were all similarly generalized and do not seem to show 487
differences in feeding strategy regarding plant phenologies. 488
Differences in rostrum and tongue size have been linked to differences in 489
nectar extraction efficiency among bat species, but this does not necessarily lead 490
to floral niche partitioning (Gonzalez-Terrazas et al. 2012). Although there may 491
be some exceptions with extreme morphological specialists (Muchhala & 492
Thomson, 2009), long rostrum/tongue in bats may have evolved so that species 493
get efficient access to the broadest range of the local flower resources in the 494
community (Gonzalez-Terrazas et al. 2012). In our bat community, X. vieirai has 495
the longest (11.9 ± 0.7 mm) and G. soricina the shortest snout (7.3 ± 0.6 mm), 496
but both species show high interaction overlap (Fig. 1). Morphological matching 497
can be an important mechanism generating niche partitioning and resource 498

32
specialization in ecological networks (Vázquez et al. 2009, Sonne et al. 2020). 499
While nectar acquisition trade-offs lead to the partitioning of interactions among 500
species in plant-hummingbird (Maruyama et al. 2014, Maglianesi et al. 2015), 501
and -hawkmoth networks (Sazatornil et al. 2016), this might not the case in 502
nectarivorous bats. Nevertheless, detailed studies on eco-morphology between 503
pollinators and plants in the Caatinga are still missing and are of great interest as 504
it may show distinct mechanisms structuring their interactions according to 505
pollination systems. 506
Our study adds many new records of interactions between bats and 507
flowers for Caatinga (Bredt et al. 2012; Cordero-Schmidt et al. 2017; Silva et al. 508
2019). Most of the new plant records and around 30% of the plants in the pollen 509
transport network overall present floral traits associated typically to bird and bee 510
pollination (van der Pijl, 1961). These non-chiropterophilous plants were similarly 511
important in the network as were plants showing characteristics in accordance 512
with chiropterophily syndrome, a trend also described previously for plant-513
hummingbird interactions networks in Caatinga (Las-Casas et al. 2012) and the 514
neighboring Cerrado’s savanna ecosystem (Maruyama et al. 2013). Interestingly, 515
such generalization may ensure the persistence of pollinator populations in 516
climatically less stable environments (Waser et al. 1996). 517
Moreover, such seldom-appreciated opportunistic use of generalist 518
flowers by bats may provide the opportunity for new evolutionary paths in some 519
groups of plants. Many closely related plant species are pollinated by bats and 520
birds, and some even have both as pollinators with distinct contribution to plant 521
reproduction (Sazima et al. 1994; Muchhala, 2007). Additionally, the transition 522
from bird to bat pollination seems the most common evolutionary pathways in 523

33
distinct groups (Tripp & Manos, 2008, Abrahamczyk, Souto-Vilarós, Renner 524
2014). In this sense, that some of the sampled non-chiropterophilous plants are 525
commonly used by bats, such as M. zehntneri (ornithophily) which comprised 526
22.8% of the samples in total, emphasizes the ability of bats to include plants in 527
their diet opportunistically. Species of Melocactus have short diurnal anthesis, 528
with flowers lasting only some hours until the end of the day (Romão, Hughes, 529
Vieira & Fontes, 2007; Locatelli & Machado, 1999). For bats to use the 530
nectar/pollen resources of these plants, visits to flowers must occur right after 531
they start foraging (17:30-18:30). Melocactus flowers are abundant year-round in 532
Caatinga, associated with distinct floral visitors such as birds, lizards and insects 533
(Taylor, 1991; Leal, Lopes & Machado, 2006), and now we also include bats in 534
this list. The opportunistic use of plants we recorded suggest that studies focusing 535
on the floral biology of diurnal flowers may find “surprises” by potentially including 536
crepuscular and nocturnal observation, as these may uncover novel and 537
unexpected interactions. Because the methodology we used does not allow us to 538
characterize effective pollination, experiments and observations on visitation 539
rates are necessary to better understand the role of nectar-feeding bats in the 540
reproduction and evolutionary paths of the non-chiropterophilous plants. 541
Cactaceae, is often regarded as closely linked to New World bats, and 542
two species, Pilosocereus pachycladus and P. gounellei with specialized bat 543
flowers and relatively extended flowering phenology (E. Cordero-Schmidt pers. 544
obs.) were important component of the network. Species in the family are well 545
adapted to arid and semiarid environments, and its predictability in providing 546
resources contribute to the persistence of fauna (Anderson, 2001). Many cacti 547
species from STDFs in the New World depend heavily on pollination by one or 548

34
more species of Phyllostomid bats (Valiente-Banuet, Arizmendi, Rojas-Martínez 549
& Domínguez-Canesco, 1996; Nassar, Beck, Sternberg & Fleming, 2003), and 550
the same pattern seems to occur in the Brazilian Caatinga. Some species of bats 551
are even defined as obligate cactophilic species (Fleming & Valiente-Banuet 552
2002). The overlapping geographic range between some nectar-feeding bats and 553
cactus-rich environments is an important aspect indicating the importance of this 554
interaction (Simmons and Wetterer, 2002). One of our bat species, Xeronycteris 555
vieirai, has recently been classified as endemic to the dry diagonal crossing the 556
central portion of Brazil, including Caatinga and Cerrado (Dias & Oliveira, 2019), 557
and was also the species that most frequently interacted with Cactaceae species 558
in both dry and rainy seasons, suggesting it is potentially a cactophilic species. 559
In conclusion, the observed generalized and temporarily stable interaction 560
patterns indicated that bat-plant interactions may be somewhat robust to current 561
human-driven disturbance impacting Caatinga (González-Varo et al. 2013, Silva, 562
Leal & Tabarelli, 2017). However, our studied interactions also include some 563
endemic species (X. vieirai and L. inexpectata), which may be more threatened. 564
Considering the relative scarcity of bat-flower networks and the importance of 565
bats as pollinators for native and crop plants (Ratto et al. 2018, Sheherezade et 566
al. 2019; Tremlett et al. 2019) we urge that more networks studies be conducted 567
with bat pollination, so that insights on the mechanisms structuring this and other 568
pollination interactions may be uncovered. As reported here, there is still much to 569
be learned and we think more unexpected findings, such as the importance of 570
non-chiropterophilous plants, might happen. Given the deep links between plants 571
and bats in this ecosystem, Caatinga should be considered a top priority for 572
conservation efforts. 573

35
REFERENCES 574
Abrahamczyk, S., Souto-Vilarós, D., & Renner, S. S. (2014). Escape from 575
extreme specialization: passionflowers, bats and the sword-billed 576
hummingbird. Proceedings of the Royal Society B: Biological Sciences, 577
281(1795), 20140888. 578
Alarcón, R., Waser, N. M., & Ollerton, J. (2008). Year-to-year variation in the 579
topology of a plant–pollinator interaction network. Oikos, 117(12), 1796-580
1807. 581
Almeida-Neto, M., Guimaraes, P., Guimaraes Jr, P. R., Loyola, R. D., & Ulrich, 582
W. (2008). A consistent metric for nestedness analysis in ecological 583
systems: reconciling concept and measurement. Oikos, 117(8), 1227-584
1239. 585
Anderson, E. F., & Brown, R. (2001). The cactus family (Vol. 776). Portland: 586
Timber press. 587
Armbruster, W. S. (2017). The specialization continuum in pollination systems: 588
diversity of concepts and implications for ecology, evolution and 589
conservation. Functional ecology, 31(1), 88-100. 590
Bredt, A., Uieda, W., & Pedro, W. A. (2012). Plantas e morcegos na recuperação 591
de áreas degradadas e na paisagem urbana. Brasília: Rede de sementes 592
do Cerrado, 273. 593
Carreira, L. M. M. & Barth, O. M. (2003). Atlas de pólen da vegetação de Canga 594
da Serra de Carajás (Pará, Brasil), Belém, Brazil: Mus. Para. Emílio 595
Goeldi. 596

36
Carvalho-Neto, F. G., da Silva, J. R., Santos, N., Rohde, C., Garcia, A. C. L., & 597
Montes, M. A. (2017). The heterogeneity of Caatinga biome: an overview 598
of the bat fauna. Mammalia, 81(3), 257-264. 599
Chacoff, N. P., Vázquez, D. P., Lomáscolo, S. B., Stevani, E. L., Dorado, J., & 600
Padrón, B. (2012). Evaluating sampling completeness in a desert plant–601
pollinator network. Journal of Animal Ecology, 81(1), 190-200. 602
Chesson, P., Gebauer, R. L., Schwinning, S., Huntly, N., Wiegand, K., Ernest, M. 603
S., ... & Weltzin, J. F. (2004). Resource pulses, species interactions, and 604
diversity maintenance in arid and semi-arid 605
environments. Oecologia, 141(2), 236-253. 606
Cordero-Schmidt, E., Barbier, E., Vargas-Mena, J. C., Oliveira, P. P., Santos, F. 607
A. R., Medellín, R. A., ... & Venticinque, E. M. (2017). Natural history of 608
the Caatinga endemic Vieira's flower bat, Xeronycteris vieirai. Acta 609
chiropterologica, 19(2), 399-408. 610
Díaz, M. M., Solari, S., Aguirre, L. F., Aguiar, L., & Barquez, R. M. (2016). Clave 611
de Identificación de los murciélagos de Sudamérica–Chave de 612
identificação dos morcegos da America do Sul. Tucumán, Argentina: 613
Yerba Buena. 614
Dirzo, R., Young, H. S., Mooney, H. A., & Ceballos, G. (2011). Seasonally dry 615
tropical forests: ecology and conservation. Island Press. 616
Dormann, C.F., Gruber B. & Fruend, J. (2008). Introducing the bipartite Package: 617
Analysing Ecological Networks. R news Vol 8/2, 8 - 11. 618
Erdtman, G. (1952). Pollen morphology and plant taxonomy. New York, 6-24. 619
Fægri, K. & van der Pijl, L. 1971. The Principles of Pollination Ecology. Pergamon 620
Press, Oxford. 621

37
Fægri, K., & van der Pijl, L. (1979). Pollination in angiosperms. The principles of 622
pollination ecology. Pergamon Press, Oxford, 13-33. 623
Fleming, T. H., & Valiente-Banuet, A. (Eds.). (2002). Columnar cacti and their 624
mutualists: evolution, ecology, and conservation. University of Arizona 625
Press. 626
Fleming, T. H., & Muchhala, N. (2008). Nectar-feeding bird and bat niches in two 627
worlds: pantropical comparisons of vertebrate pollination systems. Journal 628
of Biogeography, 35(5), 764-780. 629
Fleming, T. H., Geiselman, C., & Kress, W. J. (2009). The evolution of bat 630
pollination: a phylogenetic perspective. Annals of botany, 104(6), 1017-631
1043. 632
Fox, J., Weisberg, S., 2011. An R companion to applied regression, Second 633
edition. SAGE Publications. 634
Gonzalez-Terrazas, T. P., Medellin, R. A., Knörnschild, M., & Tschapka, M. 635
(2012). Morphological specialization influences nectar extraction 636
efficiency of sympatric nectar-feeding bats. Journal of Experimental 637
Biology, 215(22), 3989-3996. 638
González-Varo J.P., Biesmeijer J.C., Bommarco R., Potts S., Schweiger O., 639
Smith H., …, Vilà M. (2013) Combined effects of global change pressures 640
on animal-mediated pollination. Trends in Ecology and Evolution 28: 524-641
530. 642
Heithaus, E. R., Fleming, T. H., & Opler, P. A. (1975). Foraging patterns and 643
resource utilization in seven species of bats in a seasonal tropical 644
forest. Ecology, 56(4), 841-854. 645

38
Hsieh, T. C., Ma, K. H., & Chao, A. (2014). CRAN - Package iNEXT: iNterpolation 646
and EXTrapolation for species diversity. R package version 2.0. Available 647
at: http://chao.stat.nthu.edu.tw/blog/software-download. 648
Janzen, D. H. (1988). Tropical dry forests. Biodiversity, 130-137. 649
Jordano, P. (1987). Patterns of mutualistic interactions in pollination and seed 650
dispersal: connectance, dependence asymmetries, and coevolution. Am. 651
Nat. 129: 657–677. 652
Kneitel, J. M., & Chase, J. M. (2004). Trade-offs in community ecology: linking 653
spatial scales and species coexistence. Ecology letters, 7(1), 69-80. 654
Knight, T. M., Ashman, T. L., Bennett, J. M., Burns, J. H., Passonneau, S., & 655
Steets, J. A. (2018). Reflections on, and visions for, the changing field of 656
pollination ecology. Ecology letters, 21(8), 1282-1295. 657
Kunz, T. H., Braun de Torrez, E., Bauer, D., Lobova, T., & Fleming, T. H. (2011). 658
Ecosystem services provided by bats. Annals of the New York Academy 659
of Sciences, 1223(1), 1-38. 660
Las-Casas, F. M. G., Azevedo Júnior, S. M., & Dias Filho, M. M. (2012). The 661
community of hummingbirds (Aves: Trochilidae) and the assemblage of 662
flowers in a Caatinga vegetation. Brazilian Journal of Biology, 72(1), 51-663
58. 664
Leal, I. R., Lopes, A. V., Machado, I. C., & Tabarelli, M. (2017). Plant–animal 665
interactions in the caatinga: overview and perspectives. In J. M. C Silva, I. 666
Leal, M. Tabarelli (Eds.), Caatinga (pp. 255-278). Cham, Switzerland: 667
Springer. 668

39
Locatelli, E., & Machado, I. C. S. (1999). Comparative study of the floral biology 669
in two ornithophilous species of Cactaceae: Melocactus zehntneri and 670
Opuntia palmadora. Bradleya, 1999(17), 75-86. 671
Machado, I. C., Barros, L. M., & Sampaio, E. V. (1997). Phenology of caatinga 672
species at Serra Talhada, PE, northeastern Brazil. Biotropica, 29(1), 57-673
68. 674
Machado, I. C., & Lopes, A. V. (2004). Floral traits and pollination systems in the 675
Caatinga, a Brazilian tropical dry forest. Annals of Botany, 94(3), 365-376. 676
Machado, I. C., Lopes, A. V., Leite, A. V., & de Brito Neves, C. (2006). Cleome 677
spinosa (Capparaceae): Polygamodioecy and pollination by bats in urban 678
and Caatinga areas, northeastern Brazil. Botanische Jahrbücher, 127(1), 679
69-82. 680
Maglianesi, M. A., Böhning-Gaese, K., & Schleuning, M. (2015). Different 681
foraging preferences of hummingbirds on artificial and natural flowers 682
reveal mechanisms structuring plant–pollinator interactions. Journal of 683
Animal Ecology, 84(3), 655-664. 684
Mares, M. A., M. R. Willig, K. E. Streilein, and T. E. Lacher. 1981. The mammals 685
of northeastern Brazil: a preliminary assessment. Annals of the Carnegie 686
Museum of Natural History, 50:81–137. 687
Maruyama, P. K., Oliveira, G. M., Ferreira, C., Dalsgaard, B., & Oliveira, P. E. 688
(2013). Pollination syndromes ignored: importance of non-ornithophilous 689
flowers to Neotropical savanna hummingbirds. Naturwissenschaften, 690
100(11), 1061-1068. 691
Maruyama, P. K., Vizentin-Bugoni, J., Oliveira, G. M., Oliveira, P. E., & 692
Dalsgaard, B. (2014). Morphological and spatio-temporal mismatches 693

40
shape a neotropical savanna plant-hummingbird network. Biotropica, 694
46(6), 740-747. 695
Melhem, T. S., Cruz-Barros, M. A. V., Corrêa, A. M. S., Makino-Watanabe, H., 696
Silvestre-Capelato, M. S. F. & Gonçalves-Esteves, V. L. (2003). 697
Variabilidade polínica em plantas de Campos do Jordão (São Paulo, 698
Brasil). Bol. Inst Bot., 16, 1–104. 699
Mello, M. A., Felix, G. M., Pinheiro, R. B., Muylaert, R. L., Geiselman, C., 700
Santana, S. E., ... & Stevens, R. D. (2019). Insights into the assembly rules 701
of a continent-wide multilayer network. Nature ecology & evolution, 1-8. 702
Moratelli, R., & Dias, D. (2015). A new species of nectar-feeding bat, genus 703
Lonchophylla, from the Caatinga of Brazil (Chiroptera, 704
Phyllostomidae). ZooKeys, (514), 73. 705
Muchhala, N. (2007). Adaptive trade-off in floral morphology mediates 706
specialization for flowers pollinated by bats and hummingbirds. The 707
American Naturalist, 169(4), 494-504. 708
Muchhala, N., & Thomson, J. D. (2009). Going to great lengths: selection for long 709
corolla tubes in an extremely specialized bat–flower 710
mutualism. Proceedings of the Royal Society B: Biological 711
Sciences, 276(1665), 2147-2152. 712
Nassar, J. M., Beck, H., Sternberg, L. D. S., & Fleming, T. H. (2003). Dependence 713
on cacti and agaves in nectar-feeding bats from Venezuelan arid 714
zones. Journal of Mammalogy, 84(1), 106-116. 715

41
Oksanen, Jari, F. G. Blanchet, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, 716
P… & H. Wagner (2019). vegan: Community Ecology Package. R package 717
version 2.5-6. https://CRAN.R-project.org/package=vegan 718
Oliveira, P. T., e Silva, C. S., & Lima, K. C. (2017). Climatology and trend analysis 719
of extreme precipitation in subregions of Northeast Brazil. Theoretical and 720
applied climatology, 130(1-2), 77-90. 721
Ollerton, J., Killick, A., Lamborn, E., Watts, S., & Whiston, M. (2007). Multiple 722
meanings and modes: on the many ways to be a generalist 723
flower. Taxon, 56(3), 717-728. 724
Palacios-Chávez, R., Ludlow-Wiechers, B., & Villanueva, G. (1991). Flora 725
palinológica de la reserva de la biosfera de Sian Ka'an, Quintana Roo, 726
México. Chetumal, CIQRO. 727
Prado, D. E. 2003. As caatingas da América do Sul. In I. R. Leal, M. Tabarelli, & 728
J. M. C. da Silva (Eds.), Ecologia e Conservação da Caatinga (pp. 3-74). 729
Recife, Brazil: Ed. Universitária da UFPE. 730
Quesada, M., Sanchez-Azofeifa, G. A., Alvarez-Anorve, M., Stoner, K. E., Avila-731
Cabadilla, L., Calvo-Alvarado, J., ... & Gamon, J. (2009). Succession and 732
management of tropical dry forests in the Americas: Review and new 733
perspectives. Forest Ecology and Management, 258(6), 1014-1024. 734
Quirino, Z. G. M., & Machado, I. C. (2014). Pollination syndromes in a Caatinga 735
plant community in northeastern Brazil: seasonal availability of floral 736
resources in different plant growth habits. Brazilian Journal of 737
Biology, 74(1), 62-71. 738
Ratto, F., Simmons, B. I., Spake, R., Zamora-Gutierrez, V., MacDonald, M. A., 739
Merriman, J. C., ... & Dicks, L. V. (2018). Global importance of vertebrate 740

42
pollinators for plant reproductive success: a meta-analysis. Frontiers in 741
Ecology and the Environment, 16(2), 82-90. 742
Romão, R. L., Hughes, F. M., Vieira, A. M. C., & Fontes, E. C. (2007). 743
Autoecologia de Cabeça-de-frade (Melocactus ernestii Vaupel) em duas 744
áreas de afloramentos na Bahia. Revista Brasileira de Biociências, 5(1), 745
738-740. 746
Roubik, D. W., & Moreno, P. (1991). Pollen and spores of Barro Colorado Island. 747
Panamá. 748
Santos, F. A. R., Watanabe, H. M., & Hamburgo Alves, J. L. (1997). Pollen 749
morphology of some Cactaceae of North-Eastern 750
Brazil. Bradleya, 1997(15), 84-98. 751
Santos, J. C., I. R. Leal, J. S. Almeida-Cortez, G.W. Fernandes, & M. Tabarelli. 752
(2011). Caatinga: the scientific negligence experienced by a dry tropical 753
forest. Tropical Conservation Science, 4(3), 276-286. 754
Sargent, R. D., & Ackerly, D. D. (2008). Plant–pollinator interactions and the 755
assembly of plant communities. Trends in Ecology & Evolution, 23(3), 756
123-130. 757
Sazatornil, F. D., Moré, M., Benitez-Vieyra, S., Cocucci, A. A., Kitching, I. J., 758
Schlumpberger, B. O., ... & Amorim, F. W. (2016). Beyond neutral and 759
forbidden links: morphological matches and the assembly of mutualistic 760
hawkmoth–plant networks. Journal of Animal Ecology, 85(6), 1586-1594. 761
Sazima, M., Sazima, I., & Buzato, S. (1994). Nectar by day and night: 762
Siphocampylus sulfureus (Lobeliaceae) pollinated by hummingbirds and 763
bats. Plant Systematics and Evolution, 191(3-4), 237-246. 764

43
Sikes, R. S., & Animal Care and Use Committee of the American Society of 765
Mammalogists. (2016). 2016 Guidelines of the American Society of 766
Mammalogists for the use of wild mammals in research and 767
education. Journal of Mammalogy, 97(3), 663-688. 768
Silva, F. H. M., Santos, F. A. R. & Lima, L. C. L. (2016) Flora polínica das 769
caatingas: Estação Biológica de Canudos (Canudos, Bahia, Brasil). Feira 770
de Santana, Micron Bahia. 771
Silva, J. M. C., Leal, I. R., & Tabarelli, M. (Eds.). (2017). Caatinga: the largest 772
tropical dry forest region in South America. Cham, Switzerland: Springer. 773
Silva, S. S. P. D., Neves, Q. F., Guedes, P. G., Almeida, J. C. D., & Brasil-Sato, 774
M. D. C. (2019). Diet and parasites of Lonchophylla inexpectata Moratelli 775
& Dias, 2015 (Chiroptera, Lonchophyllinae) in the threatened Caatinga 776
scrublands from Ceará (northeastern Brazil). Studies on Neotropical 777
Fauna and Environment, 1-7. 778
Simmons, N. B., & Wetterer, A. L. (2002). Phylogeny and convergence in 779
cactophilic bats. In: Columnar cacti and their mutualists: evolution, 780
ecology, and conservation, 87-121. 781
Sonne, J., Vizentin-Bugoni, J., Maruyama, P. K., Araujo, A. C., Chávez-782
González, E., Coelho, A. G., Cotton, P. A., Marín-Gómez, O. H., Lara, C., 783
Lasprilla, L. R., Machado, C. G., Maglianesi, M. A., Malucelli, T. S., Martín 784
González, A. M., Oliveira, G. M., Oliveira, P. E., Ortiz-Pulido, R., Rocca, 785
M. A., Rodrigues, L. C., Sazima, I., Simmons, B. I., Tinoco, B., Varassin, 786
I. G., Vasconcelos, M. F., O’Hara, B., Schleuning, M., Rahbek, C., Sazima, 787
M. & Dalsgaard, B. (2020). Ecological mechanisms explaining interactions 788
within plant–hummingbird networks: morphological matching increases 789

44
towards lower latitudes. Proceedings of the Royal Society B, 287(1922), 790
20192873. 791
Souza, C. S., Maruyama, P. K., Aoki, C., Sigrist, M. R., Raizer, J., Gross, C. L., 792
& de Araujo, A. C. (2018). Temporal variation in plant–pollinator networks 793
from seasonal tropical environments: Higher specialization when 794
resources are scarce. Journal of Ecology, 106(6), 2409-2420. 795
Sritongchuay, T., Hughes, A. C., & Bumrungsri, S. (2019). The role of bats in 796
pollination networks is influenced by landscape structure. Global Ecology 797
and Conservation, 20, e00702. 798
Stewart, A. B., & Dudash, M. R. (2017). Flower-visiting bat species contribute 799
unequally toward agricultural pollination ecosystem services in southern 800
Thailand. Biotropica, 49(2), 239-248. 801
Stewart, A. B., & Dudash, M. R. (2018). Foraging strategies of generalist and 802
specialist Old World nectar bats in response to temporally variable floral 803
resources. Biotropica, 50(1), 98-105. 804
Straube, F. C., & Bianconi, G. V. (2002). Sobre a grandeza e a unidade utilizada 805
para estimar esforço de captura com utilização de redes-de-806
neblina. Chiroptera Neotropical, 8(1-2), 150-152. 807
Taylor, N. P. (1991). The genus Melocactus (Cactaceae) in Central and South 808
America. Bradleya, 1991(9), 1-81. 809
Tremlett, C. J., Moore, M., Chapman, M. A., Zamora-Gutierrez, V., & Peh, K. S. 810
H. Pollination by bats enhances both quality and yield of a major cash crop 811
in Mexico. Journal of Applied Ecology, 57(3), 450-459. 812

45
Tripp, E. A., & Manos, P. S. (2008). Is floral specialization an evolutionary dead-813
end? Pollination system transitions in Ruellia (Acanthaceae). Evolution: 814
International Journal of Organic Evolution, 62(7), 1712-1737. 815
Tschapka, M., & Dressler, S. (2002). Chiropterophily: On bat-flowers and flower-816
bats. Curtis's Botanical Magazine, 19(2), 114-125. 817
Valiente-Banuet, A., Arizmendi, M. D. C., Rojas-Martínez, A., & Domínguez-818
Canseco, L. (1996). Ecological relationships between columnar cacti and 819
nectar-feeding bats in Mexico. Journal of Tropical Ecology, 12(1), 103-820
119. 821
Van der Pijl, L. (1961). Ecological aspects of flower evolution. II. Zoophilous 822
flower classes. Evolution, 15(1), 44-59. 823
Vázquez, D. P., Blüthgen, N., Cagnolo, L., & Chacoff, N. P. (2009). Uniting 824
pattern and process in plant–animal mutualistic networks: a review. Annals 825
of botany, 103(9), 1445-1457. 826
Vizentin-Bugoni, J., Maruyama, P. K., Debastiani, V. J., Duarte, L. D. S., 827
Dalsgaard, B., & Sazima, M. (2016). Influences of sampling effort on 828
detected patterns and structuring processes of a Neotropical plant–829
hummingbird network. Journal of Animal Ecology, 85(1), 262-272. 830
Vizentin-Bugoni, J., Maruyama, P. K., de Souza, C. S., Ollerton, J., Rech, A. 831
R., & Sazima, M. (2018). Plant-pollinator networks in the tropics: a review. In W. 832
Dáttilo & V. Rico-Gray (Eds.), Ecological networks in the tropics (pp. 73-833
91). Cham, Switzerland: Springer 834
Waser, N. M., Chittka, L., Price, M. V., Williams, N. M., & Ollerton, J. (1996). 835
Generalization in pollination systems, and why it matters. Ecology, 77(4), 836
1043-1060. 837

46
Zanata, T. B., Dalsgaard, B., Passos, F. C., Cotton, P. A., Roper, J. J., 838
Maruyama, P. K., et al. (2017). Global patterns of interaction specialization 839
in bird–flower networks. Journal of Biogeography, 44(8), 1891-1910. 840
Zuur, A., Ieno, E. N., Walker, N., Saveliev, A. A., & Smith, G. M. (2009). Mixed 841
effects models and extensions in ecology with R. Springer Science & 842
Business. 843

844
Table 1. Network properties of plant – bat pollination networks from Caatinga of Rio Grande do Norte state, northeast Brazil. * indicates 845 significant network metrics in comparison to null models (see Methods for detail). 846 847
Network Bat Plant Interactions Connectance Specialization
(H'2)
Nestedness
(NODF)
Modularity
(Q)
Sampling Completeness
(%)
All 5 31 1250 0.66 0.08* 63.5 0.13* 84.3 Dry 5 26 675 0.6 0.11* 62.1 0.16* 77.6
Rainy 5 27 574 0.56 0.09* 67.0 0.15* 64.9 Lajes 4 24 1117 0.88 0.05* 47.2 0.11* 80.9 2015 4 16 151 0.61 0.16* 62.4 0.19* 76.4 2017 4 17 188 0.72 0.15* 69.3 0.20* 87.2 2018 4 21 244 0.73 0.11* 65.8 0.17* 70.4
848

849
850
Figure 1. Interaction network between plants and nectar-feeding bats from 851 Caatinga of Rio Grande do Norte state, northeast Brazil. The size of the lines and 852 the thickness of the boxes indicate the interaction frequency. Plant species not 853 shown (less frequent ones) are, in order: Jatropha mollissima, Bauhinia 854 cheilantha, Melocactus ernestii, Coursetia rostrata, Pilosocereus piauhyensis, 855 Fabaceae sp1, Harpochilus neesianus, Sapindaceae sp., Undetermined2, 856 Combretum leprosum, Aloe vera, Malpighiaceae sp1., Monocot sp, Pilosocereus 857 tuberculatus and Undetermined4. 858 859 860 861 862

49
863 Figure 2. Pollen types found in bat body and fur samples in Caatinga. 864 Acanthaceae: A Dicliptera ciliaris; B Harpochilus neesianus; C Haporchilus 865 paraibanus; D Ruellia asperula; Bignoneaceae: E Tabebuia aurea; 866 Bromeliaceae: F Encholirium spectabile; Cactaceae: G Cereus jamacaru; H 867 Melocactus ernestii; I Melocactus zehntneri; J Pilosocereus chrysostele; K 868 Pilosocereus gounellei; L Pilosocereus pachycladus; M Pilosocereus 869 piauhyensis; N Pilosocereus tuberculatus; Capparaceae: O Cyanophalla hastata; 870 Cleomaceae: P Tarenaya spinosa; Combretaceae: Q Combretum leprosum; 871 Convolvulacae: R Ipomoea type. Euphorbiacae: S Jatropha mollissima; 872 Fabaceae: T Bauhinia cheilantha; U Bauhinia pentandra; V Fabaceae type; 873 Malvaceae: W Helicteris baruensis; X Pseudobombax type. (Scale bar 10 µm) 874

50
875 Figure 3. Flowers of some of the bat-visited plants. Chiropterophilous species A-876 L: A. Cyanophalla hastata; B. Tareyana spinosa; C. Bauhinia cheilantha (Photo: 877 Rubens Teixeira de Queiroz); D. Bauhinia pentandra (Photo: Carmen Paixão); E. 878 Helicteris baruensis (Photo: Francisco Farriols Sarabia), F. Harpochilus 879 neesianus (Photo: Mario Adelmo Varejão Silva); G. Haporchilus paraibanus 880 (Photo: Emanoel Messias); H. Pilosocereus piauhyensis; I. Pilosocereus 881 chrysostele; J. Pilosocereus gounellei; K. Pilosocereus pachycladus; L. 882 Pilosocereus tuberculatus (Photo: Victor Lima). Melittophilous species M-Q: M. 883 Tabebuia aurea; N. Combretum leprosum (Photo: M. B. I. Lojola); O. Coursettia 884 rostrata (Photo: Domingos Cardoso); P. Aloe vera (Photo: Jamie Morrow); Q. 885 Jatropha mollissima. Ornithophilous species R-V: R. Dicliptera ciliaris (Photo: 886 Olivier Gaubert), S. Ruellia asperula; T. Melocactus zehntneri; U. Melocactus 887 ernestii. Phalaenophilous species: V. Cereus jamacaru. Mixed: W. Encholirium 888 spectabile with Lonchophylla mordax visiting its flowers. 889

51
890
Figure 4. Comparison of species-level network indices between chiropterophilous 891 plants (N= 12 species) and plants showing other pollination syndromes (N=9). 892 The boxplots depict the median and the quartiles for all the metrics. None of the 893 comparisons were significant. 894 895

52
SUPPORTING INFORMATION 896 897
898 Figure S1. A) Map of the sampling sites in Rio Grande do Norte state, 899 northeastern Brazil. B) Area of occurrence of Caatinga ecosystem in Brazil. 900
901

53
Table S1. Sampling sites of bat captures in Caatinga of Rio Grande do Norte from 902 2015-2019. *Conservation Units, other sites were located in unprotected private 903 properties 904 905
Municipality Sampling Site Latitude Longitude Habitat Assu FLONA de Açu - Sitio cables -5.576278 -36.954722 Shrubby Caatinga *Açu National Forest FLONA de Açu - Sitio carnaubal -5.537781 -36.962528 Copernicia groves
FLONA de Açu - Sitio medio -5.562993 -36.956627 Shrubby Caatinga
Baraúna Sitio Coqueiros (Buffer area) -5.022500 -37.447583 Orchards *Furna Feia National Park PARNA Furna Feia - Atolados -5.045963 -37.510515 Medium Caatinga PARNA Furna Feia - Sitio 1 -5.038146 -37.559421 Medium Caatinga PARNA Furna Feia - Sitio letreiro -5.055785 -37.537123 Medium Caatinga Cerro Corá Sitio Araras do Bala, velho -6.080221 36.317938 Riparian Caatinga Serra São João -6.045830 -36.287671 Shrubby Caatinga Felipe Guerra Sitio Passagem Funda -5.572606 -37.675224 Copernicia groves Lajedo do Rosario -5.561032 -37.663018 Rocky outcrops Lajedo do Urubu -5.572894 -37.652178 Rocky outcrops Sitio Barra -5.577348 -37.574361 Riparian Caatinga Lajes Açude Santa Rosa -5.831779 -36.202887 Riparian Caatinga Fazenda Santo Antonio -5.798484 -36.240754 Shrubby Caatinga Sitio Amarante -5.798667 -36.139528 Riparian Caatinga Serra do Feiticeiro-Minas -5.747957 -36.174944 Shrubby Caatinga
Sitio Juazeiro -5.765694 -36.215472 Riparian Caatinga Sitio Bom Fim -5.810528 -36.120556 Riparian Caatinga Sitio Tapuá -5.818944 -36.201389 Shrubby Caatinga Martins Casa de pedra -6.071420 -37.885478 Medium Caatinga Brejo -6.063889 -37.936472 Humid forest enclave Serra ESEC do Seridó - Açude e trilha -6.580697 -37.255378 Riparian Caatinga *Seridó Ecological Station ESEC do Seridó - Junco -6.599227 -37.249024 Medium Caatinga ESEC do Seridó - Lajedo -6.575139 -37.267417 Rocky outcrops ESEC do Seridó - Pe da serra -6.565667 -37.264667 Medium Caatinga ESEC do Seridó - Savana -6.573081 -37.256470 Shrubby Caatinga
906

54
Figure S2. Sampled habitat types. A. Copernicia groves, palm stands dominated 907 by Copernicia prunifera (carnauba) of 10-20m high near river courses, lagoons 908 or lakes. B. Humid forest enclave, evergreen and semideciduous forest enclaves 909 found on mountains (>500m altitude) surrounded by Caatinga vegetation with a 910 canopy of 10-20m with abundant trees, vines and epiphytes. C. Medium 911 Caatinga, xerophytic trees of 7-15m tall with a close canopy during rainy season 912 and with a variable density in the arboreal layers. D. Orchards with non-native 913 species like Mangifera indica, Musa sp., and Artocarpus heterophyllus 914 surrounded by Caatinga vegetation. E. Riparian Caatinga, alongside rivers, 915 lakes, lagoons and artificial dams. F. Rocky outcrops, outcroppings of calcareous 916

55
or granitic rocks with patches of Caatinga vegetation. G. Shrubby Caatinga, with 917 dispersed trees (3-8m tall) in a matrix of bushes and open areas with annual 918 herbs and grasses, cacti and also bromeliads. 919 920 Table S3. Description of the sampled Caatinga habitat types 921 Habitat Description Medium Caatinga forest Xerophytic trees of 7-15m tall with a close canopy during rainy season and
with a variable density in the arboreal layers. Common plant species are Poincianella pyramidalis, Mimosa tenuiflora, Ziziphus joazeiro, Myracrodruon urundeuva, Auxemma oncocalyx, Jatropha mollissima, Bauhinia forficata, Cereus jamacaru, Bromelia laciniosa. These forests are rare in RN and restricted to hillsides or canyons and in protected areas.
Shrubby Caatinga Dispersed trees (3-8m tall) of Aspidosperma pirifolium, Tabebuia aurea, Amburana cearensis in a matrix of bushes of Poincianella pyramidalis, Mimosa tenuiflora, Jatropha mollissima, Ruellia asperula, and Cnidosculus urens, and open areas with annual herbs and grasses like Aristida sp. Cacti are the most common components, Cereus jamacaru, Melocactus zehtneri, Tacinga inamoena, T. palmadoura, Pilosocereus gounellei, P. chrysostele and P. pachycladus, the latter a tree-like cacti found in large densities in some localities (e.g. Lajes); also terrestrial bromeliad Bromelia laciniosa and Encholirium spectabile are found.
Riparian Caatinga Plant composition varies among localities but, in general, riparian caatingas in RN are a mixture of Shrubby Caatinga with components of Medium Caatinga forest. However, it presents typical plant species associated to water bodies that are found alongside rivers, lakes, lagoons and artificial dams such as Licania rigida, Tabebuia aurea, Cleome spinosa. Trees of P. pyramidalis, M. tenuiflora, Prosopis juliflora, Z. joazeiro, shrubs as Jatropha mollissim., Ruellia asperula, and cacti C. jamacaru, P. gounellei and P. pachycladus can be found as well.
Copernicia Forest Palm stands dominated by Copernicia prunifera (carnauba) of 10-20m high near river courses, lagoons or lakes with a varzea characteristic during the rainy season. During the rainy season the understory is covered with herbaceous species.
Rocky outcrops Outcroppings of calcareous or granitic rocks that varies in complexity from simple unbroken rock faces to a complex of many fissured rock faces studded with patches of Caatinga vegetation including cacti (Pilosocereus gounellei), terrestrial bromeliads (Encholirium spectabile, Bromelia lacinosa), Senna sp and C. urens shrubs, and dispersed trees of P. pyramidalis and Tabebuia aurea trees are commonly found.
Brejo de Altitude Evergreen and semideciduous forest enclaves found on mountains (>500m altitude) surrounded by Caatinga vegetation with a canopy of 10-20m with trees of Ficus sp., Brosium sp., Copaifera sp., Senegalia polyphylla, Hymenaea sp., Contortisiliquum sp., Anacardium occidentale, Entherolobium sp., Psidium guajava, Spondias sp., Annona squamosa; Syagrus coronata palms, Ruellia sp. bushes, Cereus jamacaru cacti, with Cayoponia sp., Ipomea sp., and Mucuna sp vines and epiphyte bromeliads and orchids with non-native species like Mangifera indica, Musa sp., and Artocarpus heterophyllus are found.
922
923

56
Table S4. Criteria established to consider pollen contamination in the samples 924 Criteria *Pollen grains Family/Genus Observation Anemophilous or autogamous
1-2 Poaceae, Asteraceae and Amaranthaceae were disregarded
Sigle pollen in single sample
1 Malvaceae sp1., Bromeliaceae sp1., and Justicia rubrobracteata, were disregarded
Attention should be paid to these plants as possible resources for bats in future studies
**Small pollen grains
≤10 Tarenaya spinosa (Jacq.) Raf. (basionym Cleome spinosa Jacq. Criteria used by Sperr et al. 2011
Not applied to samples of individuals captured during the first half hour of sampling (1730 h-1800 h), we considered that the few pollen grains present in their fur was from the previous night and not contamination.
**Medium pollen grains
≤5 Combretum, Coursetia, Cynophalla, Dicliptera and Tabebuia
**Large and very large pollen grains
≤ 5 Bauhinia, Cereus, Encholirium, Harpochilus, Jatropha, Melocactus, Pilosocereus and Pseudobombax
*Number of pollen grains per sample that were considered contamination 925 **Size categories in Erdtman 1952 926 927
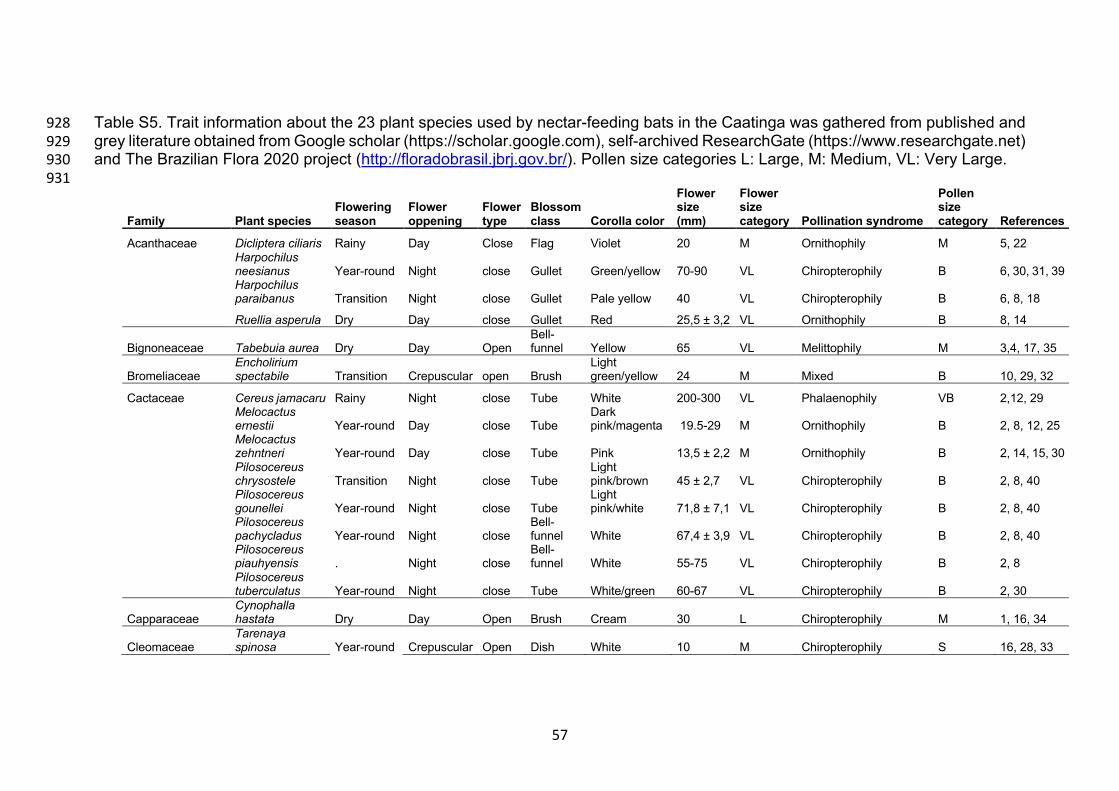
57
Table S5. Trait information about the 23 plant species used by nectar-feeding bats in the Caatinga was gathered from published and 928 grey literature obtained from Google scholar (https://scholar.google.com), self-archived ResearchGate (https://www.researchgate.net) 929 and The Brazilian Flora 2020 project (http://floradobrasil.jbrj.gov.br/). Pollen size categories L: Large, M: Medium, VL: Very Large. 930 931
Family Plant species Flowering season
Flower oppening
Flower type
Blossom class Corolla color
Flower size (mm)
Flower size category Pollination syndrome
Pollen size category References
Acanthaceae Dicliptera ciliaris Rainy Day Close Flag Violet 20 M Ornithophily M 5, 22
Harpochilus neesianus Year-round Night close Gullet Green/yellow 70-90 VL Chiropterophily B 6, 30, 31, 39
Harpochilus paraibanus Transition Night close Gullet Pale yellow 40 VL Chiropterophily B 6, 8, 18
Ruellia asperula Dry Day close Gullet Red 25,5 ± 3,2 VL Ornithophily B 8, 14
Bignoneaceae Tabebuia aurea Dry Day Open Bell-funnel Yellow 65 VL Melittophily M 3,4, 17, 35
Bromeliaceae Encholirium spectabile Transition Crepuscular open Brush
Light green/yellow 24 M Mixed B 10, 29, 32
Cactaceae Cereus jamacaru Rainy Night close Tube White 200-300 VL Phalaenophily VB 2,12, 29
Melocactus ernestii Year-round Day close Tube
Dark pink/magenta 19.5-29 M Ornithophily B 2, 8, 12, 25
Melocactus zehntneri Year-round Day close Tube Pink 13,5 ± 2,2 M Ornithophily B 2, 14, 15, 30
Pilosocereus chrysostele Transition Night close Tube
Light pink/brown 45 ± 2,7 VL Chiropterophily B 2, 8, 40
Pilosocereus gounellei Year-round Night close Tube
Light pink/white 71,8 ± 7,1 VL Chiropterophily B 2, 8, 40
Pilosocereus pachycladus Year-round Night close
Bell-funnel White 67,4 ± 3,9 VL Chiropterophily B 2, 8, 40
Pilosocereus piauhyensis . Night close
Bell-funnel White 55-75 VL Chiropterophily B 2, 8
Pilosocereus tuberculatus Year-round Night close Tube White/green 60-67 VL Chiropterophily B 2, 30
Capparaceae Cynophalla hastata Dry Day Open Brush Cream 30 L Chiropterophily M 1, 16, 34
Cleomaceae Tarenaya spinosa Year-round Crepuscular Open Dish White 10 M Chiropterophily S 16, 28, 33

58
Combretaceae Combretum leprosum Rainy Day Open Dish White 6 S Melittophily M 2, 8, 21
Euphorbiaceae Jatropha mollissima Year-round Day open Dish Yellow 5 S Melittophily/Psychophily VB 7, 27, 30, 37
Fabaceae Bauhinia cheilantha Transition Night close Flag White 50-60 VL Chiropterophily VB 9, 28, 30
Bauhinia pentandra Rainy Night close Flag White/green 40-50 VL Chiropterophily VB 2, 8
Coursetia rostrata Rainy Day close Flag White 30-35 VL Melittophily M 2, 8, 13, 19
Malvaceae Helicteres baruensis Rainy Crepuscular close Flag White-green 30 L Chiropterophily M 2, 8, 11, 38
Xanthorrhoeaceae Aloe vera Dry Day close Tube Yellow 26-34 L Melittophily/Ornithophily B 2, 8, 36 932 1. Aguirre De la Hoz, A. C., & Ruiz Zapata, T. (2017). Capparaceae Juss. from Atlántico deparment, Colombia. Ciencia
en Desarrollo, 8(1), 51-69.
2. Anderson, E. F., & Brown, R. (2001). The cactus family (Vol. 776). Portland: Timber press.
3. Barros, M. G. (2001). Pollination ecology of Tabebuia aurea (Manso) Benth. & Hook. and T. ochracea (Cham.) Standl.(Bignoniaceae) in Central Brazil cerrado vegetation. Brazilian Journal of Botany, 24(3), 255-261.
4. Barbosa-Filho, J. M., Lima, S. A., Camorim, E. L., de Sena, K. X. F., Almeida, J. R. G., da-Cunha, V. L., ... & Braz-Filho, R. (2004). Botanical study, phytochemistry and antimicrobial activity of Tabebuia aurea:(with 1 table & 1 figure). Phyton (Buenos Aires), 73, 221-228.
5. Côrtes, A. L. A., & Rapini, A. (2013). Justicieae (Acanthaceae) do semiárido do estado da Bahia, Brasil. Hoehnea, 40(2), 253-292.
6. Costa-Lima, J. L., & Chagas, E. C. D. O. (2019). A revision of Harpochilus sheds light on new combinations under Justicia (Acanthaceae). Phytotaxa, 393(2), 119-130.
7. Diogo, I. J. S., Martins, F. R., Verola, C. F., & Costa, I. R. D. (2016). Variation in plant-animal interactions along an elevational gradient of moist forest in a semiarid area of Brazil. Acta Botanica Brasilica, 30(1), 27-34.
8. Erdtman, G. (1952). Pollen morphology and plant taxonomy. New York, 6-24.
9. Fonseca Vaz, A. M. S., & Azevedo Tozzi, A. M. G. (2003). Bauhinia ser. Cansenia (Leguminosae: Caesalpinioideae) no Brasil. Rodriguésia, 54(83), 55-143.
10. Forzza, R. C., & Zappi, D. (2011). Side by side: two remarkable new species of Encholirium Mart. ex Schult. & Schult. f.(Bromeliaceae) found in the Cadeia do Espinhaço, Minas Gerais, Brazil. Kew Bulletin, 66(2), 281.
11. Goldberg, L. (2009). Patterns of nectar production and composition, and morphology of floral nectaries in Helicteres guazumifolia and Helicteres baruensis (Sterculiaceae): two sympatric species from the Costa Rican tropical dry forest. Revista de Biología Tropical, 57, 161-177.
12. Gomes, V. G. N., Quirino, Z. G. M., & Machado, I. C. (2014). Pollination and seed dispersal of Melocactus ernestii Vaupel subsp. ernestii (Cactaceae) by lizards: an example of double mutualism. Plant Biology, 16(2), 315-322.
13. Lavin, M. (1988). Systematics of Coursetia (Leguminosae-Papilionoideae). Systematic Botany Monographs, 1-167.

59
14. Leal, F. C., Lopes, A. V., & Machado, I. C. (2006). Polinização por beija-flores em uma área de caatinga no Município de Floresta, Pernambuco, Nordeste do Brasil. Revista Brasileira de Botânica, 29(3), 379-389.
15. Locatelli, E., & Machado, I. C. S. (1999). Comparative study of the floral biology in two ornithophilous species of Cactaceae: Melocactus zehntneri and Opuntia palmadora. Bradleya, 1999(17), 75-86.
16. Machado, I. C., Lopes, A. V., Leite, A. V., & de Brito Neves, C. (2006). Cleome spinosa (Capparaceae): Polygamodioecy and pollination by bats in urban and Caatinga areas, northeastern Brazil. Botanische Jahrbücher, 127(1), 69-82.
17. Mendes, D. O. F., L. F. P. Mendes, E. O. Souza & C. Aoki, 2017. Flores de paratudo (Tabebuia aurea) (Bignoniaceae) como recurso alimentar para aves no Pantanal Sul, Brasil. Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais 12(2): 295-299.
18. Monteiro, F. K. S., Fernando, E. M. P., Lucena, M. D. F. D. A., & Melo, J. I. M. (2018). A new species of northeastern Brazilian endemic genus Harpochilus (Acanthaceae). Phytotaxa, 358(3), 289-294.
19.Queiroz, L. P., & Lavin, M. (2011). Coursetia (Leguminosae) from Eastern Brazil: nuclear ribosomal and chloroplast DNA sequence analysis reveal the monophyly of three Caatinga-inhabiting species. Systematic Botany, 36(1), 69-79.
20. Queiroz, J. A., Quirino, Z. G. M., Lopes, A. V., & Machado, I. C. (2016). Vertebrate mixed pollination system in Encholirium spectabile: a bromeliad pollinated by bats, opossum and hummingbirds in a tropical dry forest. Journal of Arid
21. Quirino, Z. G. M., & Machado, I. C. (2001). Biologia da polinização e da reprodução de três espécies de Combretum Loefl.(Combretaceae). Revista Brasileira de Botânica, 24(2), 181-193.
22. Quirino, Z. G. M., & Machado, I. C. (2014). Pollination syndromes in a Caatinga plant community in northeastern Brazil: seasonal availability of floral resources in different plant growth habits. Brazilian Journal of Biology, 74(1), 62-71.
23. Rathod, A. H., Parmar, S. K., Vaghela, P. O., Sheikh, W. A., Shinde, A. S., & Kalaskar, S. R. (2014). Floral and reproductive phenology of Aloe vera. Bioscan, 9(2), 723-726.
24. Rede de catálogos polínicos online. disponível em: < http://chaves.rcpol.org.br/ >. acesso em: 1/11/2019 25. Romão, R. L., Hughes, F. M., Vieira, A. M. C., & Fontes, E. C. (2007). Autoecologia de Cabeça-de-frade (Melocactus ernestii Vaupel) em duas áreas de
afloramentos na Bahia. Revista Brasileira de Biociências, 5(1), 738-740.
26. Rocha, E. A., Machado, I. C., & Zappi, D. C. (2007). Floral biology of Pilosocereus tuberculatus (Werderm.) Byles & Rowley: a bat pollinated cactus endemic from the “Caatinga” in northeastern Brazil1. Bradleya, 2007(25), 129-145.
27. Santos, M. J., Machado, I. C., & Lopes, A. V. (2005). Biologia reprodutiva de duas espécies de Jatropha L.(Euphorbiaceae) em Caatinga, Nordeste do Brasil. Revista Brasileira de Botânica, 28(2), 361-373.
28. Santos, F. A. R., Novaes, D., & de Queiroz, L. P. (2012). Pollen of Bauhinia L. and Phanera Lour.(Leguminosae-Caesalpinioideae) from the Brazilian Caatinga. American Journal of Plant Sciences, 3(07), 909.
29. Silva, T. D. S., Felix, L. P., & Melo, J. I. M. D. (2015). Bromeliaceae and Orchidaceae on rocky outcrops in the Agreste Mesoregion of the Paraíba State, Brazil. Hoehnea, 42(2), 345-365.
30. Silva, F. H. M., Santos, F. A. R., & Lima, L. C. L. (2016). Flora Polínica das Caatingas: Estação Biológica de Canudos (Canudos, Bahia, Brasil). Feira de Santana: Micron.
31. Silva Monteiro, F. K., Pinto, A. S., da Costa, F. C. P., & de Melo, J. I. M. (2018). A Taxonomic Synopsis of Acanthaceae Juss. Native to Paraíba State, Brazil. Harvard Papers in Botany, 23(2), 189-204.
32. Silva, J., Rocha, L. H. S., Jorge, J. P. S., Sousa, P. H. P., Santos, R. L., & Freire, E. M. X. (2018). Floral visitors and potential pollinators of a rupicolous bromeliad (Pitcairnioideae) in the Brazilian semiarid. Neotropical Biology and Conservation, 13(2), 101-110.
33. Soares Neto, R. L., Magalhães, F. Á. L., Tabosa, F. R. S., Moro, M. F., Costa e Silva, M. B., & Loiola, M. I. B. (2014). Flora of Ceará state, Brazil: Capparaceae. Rodriguésia, 65(3), 671-684.

60
34. Soares Neto, R. L., & Jardim, J. G. (2015). Capparaceae in the state of Rio Grande do Norte, Brazil. Rodriguésia, 66(3), 847-857.
35. Souza, C. N., Rezende, A. A., & Gasparino, E. C. (2019). Pollen morphology of Bignoniaceae from Brazilian forest fragments and its systematic significance. Palynology, 43(2), 333-347.
36. Velásquez-Arenas, R., & Imery-Buiza, J. (2008). Fenología reproductiva y anatomía floral de las plantas Aloe vera y Aloe saponaria (Aloaceae) en Cumaná, Venezuela. Revista de biología tropical, 56(3), 1109-1125.
37. Viana, B. F., Neves, E. L. D., & MachadoI, I. C. (2011). Sistemas de polinização e de reprodução de três espécies de Jatropha (Euphorbiaceae) na Caatinga, semi-árido do Brasil.
38. von Helversen, O., & Voigt, C. C. (2002). Glossophagine bat pollination in Helicteres baruensis (Sterculiaceae). Polinización mediante murciélagos Glossophaginae en Helicteres baruensis (Sterculiaceae). Ecotropica., 8(1), 23-30.
39. Vogel, S., Machado, I.C., Lopes, A.V., (2004). Harpochilus neesianus and other novel cases of chiropterophily in neotropical Acanthaceae. Taxon 53, 55e60.
40. Rocha, E. A., Domingos-Melo, A., Zappi, D. C., & Machado, I. C. Reproductive biology of columnar cacti: are bats the only protagonists in the pollination of Pilosocereus, a typical chiropterophilous genus?. Folia Geobotanica, 1-18.
933

61
Table S6. Nectar-feeding bat captures throughout the study period 934 935
(a) Captures per year Year (Sampling effort) Anoura
geoffroyi Glossophaga
soricina Lonchophylla inexpectata
Lonchophylla mordax
Xeronycteris vieirai
Total
2015 (44163,75 m2h) 1 48 12 27 33 121 2017 (103646,25 m2h) . 72 26 49 29 176 2018 (143623,75 m2h) 4 99 50 81 55 289 2019 (15663,75 m2h) 1 27 9 15 12 64
TOTAL 6 246 97 172 129 650
(b) Captures per season Season (Sampling
effort) Anoura
geoffroyi Glossophaga
soricina Lonchophylla inexpectata
Lonchophylla mordax
Xeronycteris vieirai
Total
Rainy (140717,5 m2h) 1 99 45 97 62 304 Dry (166380 m2h) 5 147 52 75 67 346
(c) Captures per municipality and habitat type
Municipality (sampling effort)
Habitat type Anoura geoffroyi
Glossophaga soricina
Lonchophylla inexpectata
Lonchophylla mordax
Xeronycteris vieirai
*Açu (14198,75 m2h) Copernicia groves
. . . 1 .
Copernicia groves
. 10 . . .
*Baraúna(21220 m2h) Medium . 7 . . . Orchards . . . . . Shrubby . . . . .
Cerro Corá (5086,25 m2h)
Riparian . 2 . . .
Shrubby . . . . . Felipe Guerra (30907,5
m2h) Copernicia
groves 1 . . .
Medium . 5 . . . Riparian . 5 . 3 . Rocky
outcrops 18 . 13 .
Lajes (202600 m2h) Riparian 1 79 43 47 10 Shrubby 109 54 100 119
Martins (14267,5 m2h)
Humid forest enclave
5 10 . . .
Medium . . . 1 . *Serra Negra do Norte
(22612,5 m2h) Medium . . . . .
Riparian . . . . . Rocky
outcrops . . . 7 .
Shrubby . . . . . *Protected areas
936 937

62
938
939

63
940
941 942
Mechanisms mediating nectar-feeding bats coexistence and persistence 943
through the seasonal rhythm of Caatinga 944
945
Eugenia Cordero-Schmidt1, Juan Carlos Vargas-Mena1, Paulino Pereira-946 Oliveira2, Francisco de Assis R. Santos2, Bernal Rodriguez-Herrera3 and 947 Eduardo M. Venticinque1 948 949 950 1 Departamento de Ecologia, Universidade Federal do Rio Grande do Norte, 951 59078900 Lagoa Nova Natal, RN, Brazil; E-mail: [email protected] 952 2 Laboratório de Micromorfologia Vegetal, Universidade Estadual de Feira de 953 Santana, 44036-900, Novo Horizonte, BA, Brazil 954 3 Escuela de Biología, Universidad de Costa Rica, 2060 Montes de Oca, San 955 José, Costa Rica 956 957

64
ABSTRACT 958
Species coexistence is mediated by ecological differences in space, time 959 and/or complementarity of food resources and these attributes are linked to each 960 species physiology, morphology, and behavior. Coexisting nectar-feeding bats 961 have been observed in most neotropical regions and up to eight species have 962 been observed in a single locality, and up to five species co-occur in Caatinga. 963 We tested three coexisting mechanisms between nectar-feeding bat species: a) 964 Temporal partitioning by comparing differences in capture rates between years, 965 seasons and between hours within the same nights. b) Ecomorphological 966 differences by measuring body weight, forearm size, snout length and wing 967 morphology. c) Resource partitioning by identifying pollen in fur samples and 968 determining the composition of their feces. We studied a nectar-feeding bat 969 ensemble in an area of Caatinga in Rio Grande do Norte state for over two years 970 and collected phenological data on nine plant species to document resource 971 availability through time. We capture a total of 443 individuals belonging to four 972 nectar-feeding bat species: Glossophaga soricina, Lonchophylla inexpectata, L. 973 mordax and Xeronycteris vieirai. The mechanisms detected are complementary 974 and seem to be explained by temporal partitioning (differences in captures 975 throughout the night), by ecomorphological differences (size, weight, snout 976 length, wing aspect ratio) and by resource partitioning (diet flexibility feeding on 977 at least 20 plant species, and insects). The temporal persistence of nectar bats 978 was facilitated by the continuity and complementarity of floral resources 979 availability during both seasons. Cactaceae is a keystone resource in Caatinga 980 since it was the most used family (five cacti species used in a high frequency) by 981 all nectar-feeding bats. 982 983
984
Keywords: Eco-morphology; Keystone resources; Phenology; Resource-985
partitioning; Seasonally Dry Tropical Forest; Night-sharing 986
987

65
GRAPHICAL ABSTRACT 988
989
990
Nectar-feeding bats persist and coexist year-round both in thewet season & dry season by:
Ecomorphological:- Weight and size
- Snout length
- Wing Aspect ratio
Temporal partitioning:
- Differences in captures throughout
the hours of the night
Diet flexibility:- 24 plant species- Insects- Pollen- Plant tissues
Keystone resource:- Cactaceae- Pilosocereus pachycladus
Glossophaga soricina
Lonchophylla mordax
Xeronycteris vieirai
Lonchophylla inexpectata
9
8
76
10
Res
ourc
e pa
rtitio
ning
1112

66
INTRODUCTION 991
Tropics are packed with coexisting species, and understanding the mechanisms 992
enabling this have interested ecologists for decades. Coexistence may be even 993
more intriguing between species performing same ecological roles, such as 994
pollinators (Palmer, Stanton & Young, 2003). Pianka (1974) noted that species 995
coexistence is mediated by ecological differences in space, time and/or 996
complementarity of food resources and these attributes are linked to each 997
species physiology, morphology, and behavior (Jorgensen & Fath, 2014). Bats 998
have been an ideal group to deepen the understanding of coexistence and 999
competition mechanisms, as suites of evolutionary sister species may commonly 1000
be encountered in the same locality (Findley, 1993). 1001
The known fauna of nectar-feeding bats (hereafter ‘nectar bats’) in the 1002
Neotropics is composed of 55 species (Solari & Martínez-Arias, 2014, Moratelli 1003
& Dias, 2015), divided in the subfamilies Lonchophyllinae (19 species) and 1004
Glossophaginae (36 species). Coexisting nectar bats have been observed in 1005
most neotropical regions and up to eight species have been observed in a single 1006
locality (Fleming & Muchhala 2008). There are several adaptations linked to 1007
nectar feeding species and small differences among them can mediate 1008
mechanisms of coexistence between the species of nectar bats (Gonzales-1009
Terrazas et al., 2012). 1010
Nectar bats present long narrow snouts with reduced dentition coupled 1011
with long tongue with hair-like papillae in the tip of the tongue (in Glossophaginae) 1012
or deep lateral grooves along the entire length of the tongue (in Lonchophyllinae) 1013
(Tschapka, Gonzalez-Terrazas, & Knörnschild, 2015). Nectar bats are generally 1014
light weighted and have specific proportions in their wings that aid in hovering 1015

67
while feeding (Findley, Studier & Wilson, 1972). The mentioned morphological 1016
characteristics limits access to certain floral resources and nectar extraction 1017
efficiency contributing to coexistence by resource partition (Gonzales-Terrazas 1018
et al., 2012). 1019
Physiologically, nectar bats possess high basic metabolic rates and high 1020
daily energy requirements, likewise they are able to digest and metabolize nectar 1021
and pollen quickly (Datzmann, von Helversen & Mayer, 2010). In nature nectar 1022
bats encounter a wide variety of resources with different energetic cost-benefits. 1023
Bats mostly feed on chiropterophilous flowers (nocturnal, robust with light colors 1024
and strong scent), but have been reported to use non-chiropterophilous as well 1025
(Vogel, Lopes & Machado, 2005; Fadini et al., 2018; Cordero-Schmidt in prep.). 1026
It has been proven that the resource selection is not random and that it is highly 1027
influenced by flower attraction, access and rewards (amounts of nectar and 1028
pollen and nectar sugar concentrations) (Tschapka, & Dressler, 2002) and that 1029
the use of certain resources could be species-specific (Winter & von Helversen, 1030
1998; Tschapka et al., 2008; Sperr et al., 2011; Gonzales-Terrazas et al., 2012), 1031
once again contributing with nectar bats coexistence. 1032
Diet plasticity has proven to be another important mechanism of 1033
coexistence and persistence of populations of nectar bats in areas with resource 1034
seasonality (Heithaus et al., 1975; Soriano et al., 1991; Tschapka et al., 2008). 1035
Certain nectarivorous species can also feed on insects and fruits at times of low 1036
availability of floral resources (Reid, 2009). Finally, species turnover has also 1037
been documented for coexistence in bats (Fleming, Muchala & Ornelas, 2005). 1038
Though, a mixture of all the mentioned mechanisms is what generally explains 1039
the coexistence of the species. 1040

68
Testing coexistence is a challenging task, especially in the highly 1041
diversified tropics. Mechanisms modeling nectar bat co-occurence have been 1042
fairly well documented in arid and semi-arid areas (Valiente-Banuet et al., 1996; 1043
Petit, 1996; Nasar et al., 2003). Nine species of nectar-feeding bats are known to 1044
inhabit Caatinga, the largest Seasonal Tropical Dry Forest in America located in 1045
northeastern Brazil, but information about these species is scarce. 1046
The coexistence of nectar bat species in the highly variable environment 1047
of Caatinga is particularly puzzling considering the low and variable precipitation 1048
regulating plant phenologies (Machado, Barros & Sampaio, 1997; Lima & Rodal, 1049
2010) and thus species composition and behavior. The present study is the first 1050
to focus on Caatinga nectar bat ensemble using an ecological approach of 1051
coexistence and persistence of species over time. We tested three coexisting 1052
mechanisms between nectar bat species: a. Temporal partitioning (between 1053
years, seasons and between hours within the same nights). b. Ecomorphological 1054
differences (snout length and wing morphology). c. Resource partitioning (diet 1055
composition). 1056
In markedly seasonal ecosystems, climatic variables are good predictors 1057
of population behavior (Janzen, 1973). Hence, considering Caatinga’s temporal 1058
variability reflected in the seasonality of the availability of food resources, our 1059
hypothesis was that we would find seasonal differences both in the number of 1060
nectar bat species coexisting as well as in the resources used by them. We 1061
expected that at times of less resources available (driest months) the composition 1062
of the ensemble would be different, and the remaining species would have a more 1063
similar diet among them. On the contrary, in periods with more resources (wettest 1064
months) there would be more co-existing nectar bats which would have a more 1065

69
differentiated diet (considering used plant species and complementing items) and 1066
the use of resources would be explained by ecomorphological differences among 1067
the bat species. 1068
METHODS 1069
Study site 1070
The study was carried out in the Caatinga in the municipality of Lajes, Rio Grande 1071
do Norte (RN) state, northeastern Brazil (5°48'46.95"S 36°10'34.60"W). Our three 1072
sampling sites were located within the largest (53,500 ha; 200–400 mamsl) 1073
continuous fragment of Caatinga vegetation in the RN state. Lajes is located in 1074
the Northern semiarid region (following Oliveira et al., 2017) with an average 1075
rainfall of 399.31 mm (precipitation data from 2009 to 2019). The wet season is 1076
from January to June and the dry season from July to December. 1077
During our study, the sum of the accumulated rainfall was 313.2 mm in 1078
2017, the rainiest month was February (111.9 mm) and the driest months were 1079
August to November with zero precipitation. In 2018 the accumulated rainfall was 1080
351.65 mm in 2018 (rainiest April with 108 mm and the driest August, September 1081
and November with zero precipitation). All precipitation data were extracted from 1082
the EMPARN (acronym in Portuguese Rio Grande do Norte Agricultural 1083
Research Corporation) database. Vegetation in Lajes is Hyperxerophilous 1084
Caatinga type, a dry vegetation, with large Cactaceae presence and smaller and 1085
scattered plants. Presenting a predominance of deciduous herbaceous and 1086
shrubby plant species (Farias, 2016). 1087
Bat sampling 1088
Bat captures was conducted monthly, in a total of 57 mist-netting nights 1089
(approximately 3 nights per month), between February 2017 and February 2019 1090

70
(except November 2017 and May 2018), including 30 nights in the dry season 1091
and 27 in the wet season. Mist nets were placed for over six hours (from sunset 1092
to midnight), using 80 to 120 m of mist nets per night. Total sampling effort was 1093
155991.25 m2h (total area of the mist-nets multiplied by the by the time they were 1094
set; Straube and Bianconi, 2002). We identified the species in the field using 1095
systematic keys and species diagnosis (Díaz et al., 2016). All procedures for 1096
capture, handling and collection of bats met the guidelines of the American 1097
Society of Mammalogists for the use of wild mammals in research (Sikes, 2016), 1098
and the legal Brazilian requirements of conservation and animal welfare. 1099
Fieldwork was authorized by MMA/ICMBio/SISBIO under permits 48325-2. Bats 1100
were marked by cutting a small portion of fur from the back of each individual to 1101
recognize recaptured individuals during the same sampling period. 1102
a. Temporal partition and species turnover 1103
The time of capture was recorded for each bat individual; this information was 1104
used as a proxy for differences in the species activity as a coexistence 1105
mechanism. We tested for significant differences in the capture rates between 1106
years, seasons and hours within the night. For this analysis we considered 20 1107
sampled periods (9 months for the wet season, 11 for the dry). First, we did a 1108
Non-Metric Multidimensional Scaling (NMDS) over a Bray Curtis dissimilarity 1109
matrix of the nectar bat species. Prior to the analysis the bat capture data was 1110
corrected by effort as follows: [Captures N / (mist netting hours * meters2) * 1000]. 1111
The ordering adjustment was measured by stress value. The temporal species 1112
composition of the nectar bat ensemble between years, seasons and hours was 1113
tested by applying a MANOVA to the NMDS first and second axis scores values. 1114

71
After MANOVA, one-way ANOVAs were applied to identify in which axes the 1115
variation was significant. 1116
Differences between years, seasons and hours within the nights were 1117
tested for statistical significance using a one-way ANOVA followed by Tukey’s 1118
Honest Significant difference test. We additionally performed a MANOVA to see 1119
significant differences across all variables using Wilks's Lambda Test. 1120
b. Eco-morphological differences 1121
-Body weight and size 1122
We sexed each captured nectar bat, weighed all adult individuals (except 1123
pregnant females) with a dynamometer (precision: 1 g), and measured their 1124
forearm length using calipers (precision: 0.1 mm). 1125
-Snout length 1126
The length of the snout is an indication of specialization in nectarivorous diet, the 1127
longer, the more specialized (Freeman, 1995). A ruler (precision: 0.5 mm) was 1128
used to measure the distance between the center of the eye and the tip of the 1129
lower lip (Tschapka, 2004). 1130
-Wing morphology 1131
To explore if the coexistence mechanisms of nectar bats are related to 1132
differences in their flight style (maneuverability and hovering) and foraging 1133
behavior (ability to fly longer distances in search of resources), we collected three 1134
wing measurements following Tschapka (2004). 1135
We measured the length of the 3rd and 5th finger by using a ruler 1136
(precision: 0.5 mm). Wing proportions were quantified using Aspect Ratio Index 1137
(ARI) and Wing Tip Index (WTI). Following the morphologic properties of bat 1138
wings (Findley, Studier & Wilson, 1972), migratory and high-speed species have 1139

72
high ARI and WTI. These characteristics favors long distance foraging. On the 1140
other hand, species with good ability to maneuver and hover have a low ARI. 1141
Both indices are independent of body size and of each other. 1142
Differences in morphological variables (weight, forearm, ARI, WTI and 1143
snout) between nectar bat species were tested for statistical significance using a 1144
one-way ANOVA followed by Tukey’s Honest Significant difference test. 1145
c. Resource Partition 1146
-Resource availability 1147
To detect the availability of floral resources for one year (December 2017-1148
November 2018, except for May 2018), we followed monthly the phenology of 1149
152 individuals of 9 species (Supporting Information Table S1). Plant species 1150
selection was based on previous field observations of resource use by bats. We 1151
established 15 transects of 100 x 5 m following pre-existing trails, where the 1152
species of trees, shrubs, cacti and bromeliads were identified, and individuals of 1153
each species were counted for abundance estimation in each transect. We 1154
selected at least two adult individuals of each species and tagged them with 1155
fluorescent flagging tape and numbered aluminum plates. Data collected for each 1156
individual were presence or absence of flowers (both buds and open flowers were 1157
considered). 1158
Botanical samples of the species were collected for proper identification 1159
and deposited in the herbarium of the Universidade Federal do Rio Grande do 1160
Norte. Pollen samples were collected directly from flowers available during the 1161
sampling period with jelly-glycerin cubes in order to make a reference pollen 1162
collection. 1163
A matrix of presence and absence of flowering species by month was 1164

73
developed; we summed the number of species flowering in each month. The 1165
dates of the observations were converted into angles, the day 01 / January = 0°, 1166
and sequentially until December 31=360 °, with intervals of approximately 30°. 1167
The flowering phenophase was analyzed using circular statistics using the R 1168
program with the packages Circular (Lund, Agostinelli & Agostinelli, 2017), and 1169
CircStats (Lund and Agostinelli, 2018). The mean angle (M) or average date is 1170
the time of the year around which the dates of flowering occurred for most 1171
species. The Rayleigh (p) test was performed to verify flowering seasonality for 1172
data with unimodal or uniform distribution, whereas Rao’s Spacing Test was 1173
performed when data showed bi or multimodal patterns. The vector ‘r’ indicates 1174
the temporal concentration of events and vary from 0 (when phenological activity 1175
is distributed uniformly throughout the year) to 1 (when phenological activity is 1176
concentrated around one single date or time of year). 1177
- Flower visits 1178
We constructed species accumulation curves on nectar bats captured and the 1179
pollen types found in the samples of the individuals to establish whether the diet 1180
sampling was completed, or otherwise, to know its representativeness of the total 1181
estimated plant species (Moreno and Halffter, 2001). 1182
We collected pollen samples from each captured bat’s fur, wings, legs and 1183
uropatagium using a single glycerin jelly cube (Voigt et al., 2009) per individual, 1184
which were later mounted on glass microscope slides. To avoid cross-1185
contamination from one sample to another, special care was taken by cleaning 1186
the worktable, hands and tweezers with alcohol after finishing the handling of 1187
each individual bat, additionally, cloth bags used to keep the nectar bats were 1188
always clean. 1189

74
We identified pollen types under a light microscope (magnification 40-1190
100x, Leica DM500) to the lowest possible taxonomic level through comparison 1191
with reference collections. To identify the pollen types, we used a collection 1192
specialized on pollen of the Caatinga vegetation from the Palynotheca of the 1193
LAMIV, UEFS (Plant Micromorphology Laboratory, Universidade Estadual de 1194
Feira de Santana, and a reference collection that we made from local plants in 1195
bloom during our fieldwork. We also used pollen catalogues for additional 1196
reference (Palacios-Chávez, Ludlow-Wiechers & Villanueva, 1991; Roubik & 1197
Moreno 1991; Santos, Watanabe & Hamburgo-Alves, 1997; Carreira & Barth 1198
2003; Melhem et al., 2003; Silva, Santos & Lima, 2016). 1199
We noted the identity of the pollen types and number of times each pollen 1200
type was found in a specific bat species as a measure of the frequency of 1201
resource use. Small amounts of pollen can contaminate the samples; thus, we 1202
established a minimum number of pollen grains according to size categories 1203
(Erdtman, 1952; Heithaus, Fleming & Opler, 1975). Medium, large and very large 1204
pollen types (Combretum, Coursetia, Cynophalla, Dicliptera and Tabebuia, 1205
Bauhinia, Cereus, Encholirium, Harpochilus, Jatropha, Melocactus, Pilosocereus 1206
and Pseudobombax) represented in a sample by less than five pollen grains were 1207
considered contamination. For small pollen types (Tareyana spinosa) less than 1208
10 pollen grains were considered contamination. 1209
To detect possible patterns of associations and differences within the 1210
plants used by nectar bat species in the dry and wet season, we performed a 1211
Correspondence Analysis (CA). The CA is an analysis for displaying the rows 1212
and columns of a matrix as points in dual low dimensional vector space 1213
(Greenacre, 1984). The graphical representation of data generated by CA allows 1214

75
visual examination of the correlations among the plant species (row labels) and 1215
bat species (column labels) in the dry and wet season. On the plot, the closer two 1216
rows are to each other, the more similar their residuals, additionally, the closer 1217
the labels are to the origin (where the x- and y-axes are both at 0), the less 1218
distinct they probably are. The length of the lines connecting row/column label to 1219
the origin indicates the association between nectar bats species and one or more 1220
plant species (longer lines indicates higher associations). Finally, the angle 1221
formed between the two lines indicate association, where angles <90 indicate 1222
higher association, 90 degrees angle indicate no relationship and 180 degrees 1223
indicate negative associations. 1224
We performed an analysis of similarity (ANOSIM) to test if there were 1225
significant differences in the diet between species. To measure the similarity, we 1226
used the Bray Curtis dissimilarity index. Significance levels were adjusted by a 1227
Bonferroni sequential correction. To help with the interpretation of the results 1228
obtained with the ANOSIM, we performed a Similarity Percentages (SIMPER) 1229
analysis using the Bray-Curtis method. This analysis breaks down the 1230
contribution of each species of plant to the observed dissimilarity between the 1231
nectar-feeding bat species diets. It allows to identify the species that are most 1232
important in creating the observed pattern of dissimilarity. Analyses were 1233
performed in PAST 3.0. 1234
-Diet composition 1235
To infer if diet plasticity of the different nectar bat species could be 1236
considered as a mechanism of persistence and coexistence, we collected fecal 1237
samples and characterized their contents using a microscope (magnification 40-1238
100x, Leica DM500). We categorized the contents of each fecal sample by the 1239

76
presence or absence of pollen, insect fragments, and “plant tissue” which are 1240
unidentified elements that appear to be of plant origin such as leaves, pericarp, 1241
hairs of cactus areolas, trichomes, etc. 1242
All the individuals captured were fed with sugar water at different times 1243
throughout the handling process, to maintain their energy requirements and 1244
reduce the risk of death due to stress. Additionally, it promotes the production 1245
and excretion of feces while collecting the data, since bats have a fast transit time 1246
of food ingestion-defecation (Muscarella and Fleming, 2007). Fecal samples 1247
were placed in micro tubes for preservation and subsequent analysis. 1248
As mentioned before, snout length is a proxy of specialization, the longer 1249
the snout, the less force in the bite, thus limiting the ability to consume harder 1250
food items, such as insects and fruits (Heithaus, 1982). We correlated the snout 1251
length (mm) of the four nectar bat species and established a Fecal Composition 1252
Index grouping fecal sample in three categories A. Pollen: samples with ≥ 1 pollen 1253
types; B. Pollen & insects: with both ≥ 1 pollen types and insect fragments and/or 1254
moth scales; C Mix: with ≥ 1 pollen types, insect fragments and plant tissues. The 1255
Fecal Composition Index is the score of the first axe of an NMDS analysis with 1256
the percentages of fecal composition by species. In the NMDS ordination we 1257
used the Bray Curtis dissimilarity index to calculate the distance between fecal 1258
species composition. 1259
RESULTS 1260
Nectar-feeding bat ensemble 1261
We captured a total of 443 nectar bats. The local nectar bat ensemble consisted 1262
of four resident species (present year-round) (Figure 1), one Glossophaginae 1263
species, Glossophaga soricina (dry n= 87, wet n=53), and three Lonchophyllinae 1264

77
species, Lonchophylla mordax (dry n=57, wet n=63), Xeronycteris vieirai (dry 1265
n=46, wet n=50) and Lonchophylla inexpectata (dry n=44, wet n=43). 1266
Temporal partitioning 1267
-Changes in species composition and temporal partitioning between seasons and 1268
years 1269
There were no significant differences between the captures rates for all species 1270
(F = 0.710; DF = 3,72; P = 0.548), between seasons (F = 0.655; DF = 1,72; P = 1271
0.420) and the interaction between species * season (F = 0.470; DF = 3.72; P = 1272
0.703) (Supporting Information Table S2). 1273
The time dynamics of capture rates were strongly correlated between X. 1274
vierai and L inexpectata (r = 0.83), between L. mordax and L. inexpectata (r = 1275
0.71) and between X. vierai and L. mordax (r = 0.69). Glossophaga soricinia 1276
presented a capture dynamic uncoupled with L. inexpectata and L. mordax 1277
(Supporting Information Table S3). There were no significant differences among 1278
the community composition within the years (MANOVA with Wilks's 1279
Lambda=0.96393; F=0.29939; df=2,16; p=0.74533) and the seasons (MANOVA 1280
with Wilks's Lambda=0.91777; F=0.76156; df=2,17; p=0.48222) (Respectively, 1281
Supporting Information Table S4A and S4B and Figure 2A and 2B). 1282
-Temporal partition between hours within the night 1283
There were significant differences (MANOVA with Wilks's Lambda=0.44649; 1284
F=2.15177; df=12,52; p=0.02883) among the community composition between 1285
the hours of the nights (Figure 3A). Some subtle differences are observed 1286
between the number of the different species (Figure 3B). However, after 21:00 1287
the captures of all species remain low but stable. In the univariate ANOVA, the 1288
first axis from the NMDS over a Bray Curtis dissimilarity matrix of the nectar bat 1289

78
species captured within the night showed a significant difference between the bat 1290
community over the hours (F=4.301; df=6,27; p=0.00362), but the second axis 1291
showed no significant difference (F=0.47422; df=6,27; p=0.82138). The first axis 1292
was related to a decrease in activity of all species. Xeronycteris vierai showed a 1293
less pronounced peak in early evening activity (Supporting Information Table S5). 1294
a. Ecomorphological measurements 1295
Significant differences were found in the morphology of the four nectar bat 1296
species Table 1). Body size: Considering weight and forearm, the biggest species 1297
is X. vieirai (respectively x̄=11.87 ± 1.77 g; x̄=38.47 ± 0.96 mm), followed by G. 1298
soricina (x̄=9.98 ± 1.29 g; x̄=35.89 ± 1.26 mm), and finally L. mordax (x̄=8.73 ± 1299
1.21 g; x̄=35.38 ± 1,67 mm), and L. inexpectata (x̄=8.47 ± 1.10 g; x̄=35.06 ± 1.46 1300
mm). 1301
Wing morphology: Wing Tip Index (WTI) was the only morphological 1302
measurement that was not significantly different between species, and Aspect 1303
Ratio Index (ARI) was significantly different for X. vieirai when compared to the 1304
other species. 1305
Snout length: The four nectar bat species could be grouped into three size 1306
categories with significant differences, X. vieirai (x̄=11.99 ± 0.55 mm) has the 1307
longest snout of all, followed by the two species with intermediate size, L. mordax 1308
(x̄=8.84 ± 0.66 mm) and L. inexpectata (x̄=8.99 ± 0.70 mm), finally G. soricina 1309
with the snout significantly shorter (x̄=7.33 ± 0.56 mm). 1310
Resource Partition 1311
-Resource availability 1312
The studied plant community of this Caatinga area presented a constant and 1313
complementary resource offer throughout the year (Figure 4, Table 2). All the 1314

79
nine species included in this phenological analysis were used as a feeding 1315
resource by nectar bats. Considering the number of species and the intensity of 1316
resources availability, the months with the greatest supply are December, 1317
January and February. Cereus jamacaru and Jatropha mollissima flower only in 1318
the wet season. Lowest resource availability occurred in the driest period in 1319
September, October and November. Phenological complementarity is observed 1320
throughout the year (Figure 4, Supplementary Material Table S1). 1321
- Flower visits 1322
We identified pollen types in a total of 443 samples, of which 140 were from G. 1323
soricina, 120 from L. mordax, 96 of X. vieirai and 87 from L. inexpectata. The diet 1324
of the four nectar bats studied was fairly well represented in our samples 1325
(Supporting Information Figure S6). They are feeding on at least 24 plant species 1326
belonging to 11 families. The used plant species show different flower size, 1327
blossom class (flower shape), nectar’s volume and sugar concentration (Table 1328
3). 1329
The nectar bats used more species of the Cactaceae family (5 species), 1330
followed by Acanthaceae and Fabaceae, each with four species. Although most 1331
of the plants were used by all species of nectar bats, differences in the frequency 1332
of use of resources were found (Table 3). The most similar diets were between 1333
L. mordax and L. inexpectata in both seasons and between L. inexpectata and 1334
G. soricina in the wet season (Table 4). Lonchophylla mordax used 23 species 1335
(except J. mollissima), 21 species used by G. soricina (except: Ipomoea sp., H. 1336
neesianus and J. mollissima), likewise X. vieirai used 21 species (except B. 1337
cheilantha, A. vera and an unidentified pollen type), finally the species that fed 1338

80
on fewer plant species was L. inexpectata that used 20 species (except H. 1339
neesianus, A. vera and two unidentified pollen types). 1340
Correspondence analysis separated our nectar-feeding bat species 1341
according to their diet per season. Axis1 accounted 54.2% of the variance (Table 1342
5) and separated bat species by season, additionally, Axis2 (13.4%) separated 1343
X. vieirai from the other bat species (Figure 5). There are several plant species 1344
located in the middle (i.e., near the origin) of the two-dimension space, these 1345
plants are used by all bat species during both seasons. The longest lines in the 1346
different seasons indicate seasonality of resource availability of the plant species, 1347
and/or the importance of the resource for a specific bat species in each season. 1348
Ipomoea type was mostly used by L. inexpectata, while R. asperula was 1349
mostly used by L. mordax in the dry season. On the other hand, H. neesianus 1350
was mostly used by L. mordax and an unidentified leguminosae species 1351
(Fabaceae type) was mostly used by X. vieirai. Glossophaga soricina’s diet 1352
variated greatly seasonally. In the dry season G. sorcina was associated with 1353
most of the commonly used plant species (closer to the origin) T. spinosa and J. 1354
mollissima, in the wet season the G. soricina was the only bat species to use A. 1355
vera, C. rostrata and C. hastata. 1356
The differences between the diet of the nectar-feeding bat species are 1357
better represented in the Axis2 (13.4%) and Axis3 (12.1%) (Figure 6). The four 1358
nectar bat species were divided by genus into three groups and not by sub 1359
families that would be expected by their tongue characteristics. Lonchophylla 1360
sister species have a more similar diet between them, especially during the wet 1361
season and differed more in the dry season. The diet of Xeronycteris differs 1362
greatly from the other nectar-feeding bat species in both seasons (Figure 5 and 1363

81
6). Xeronycteris vieirai’s diet diverges the most with L. mordax in the dry season, 1364
and with G. soricina in the wet season. The ANOSIM (Table 6) revealed 1365
differences in the diet between the four species of nectar-feeding bats (R = 0.036, 1366
p = 0.035). 1367
The contribution of each plant species (%) to the dissimilarity between the 1368
diet of each nectar-feeding bat species in each season are summed in Table 7 1369
(SIMPER). The first eight species of consumed plants explain a little more than 1370
50% of the dissimilarity in the diet of the nectarivores. Season related 1371
dissimilarities in the use was observed in C. jamacaru and H. neesianus (mostly 1372
used in the wet season) and H. baruensis and E. spectabile (in the dry season). 1373
Bat taxonomic related dissimilarities in resource use was observed in P. 1374
pachycladus, P. gounellei and T. spinosa (Supporting Information S7). Where P. 1375
gounellei and P. pachycladus (Cactaceae) were the only two species that were 1376
used year-round with the highest frequency of use by all nectar bat species. 1377
Nonetheless, P. gounellei was mostly used by Lonchophyllinae species and P. 1378
pachycladus mostly by X. vieirai. Tareyana spinosa was also used year-round 1379
but was used differently by nectar bat species in the different seasons 1380
-Diet composition 1381
We collected 125 fecal samples (G. soricina n=23; L. inexpectata n=33; L. 1382
mordax n=29 and X. vieirai n=40) (Figure7). The four species of nectar bat 1383
species are feeding on the same elements (pollen, insects and plant tissue) 1384
during both seasons. However, G. soricina in addition to these elements also 1385
consumed cactus fruits. We found P. pachycladus seeds in the body of two 1386
individuals (fruits were not included in the analysis since the seeds were not found 1387
in the fecal samples but in the bats fur near the anus). Fecal samples of all 1388

82
species presented pollen, however for most species we found pollen mixed with 1389
insect fragments and/or plant tissues (which could be leaves, pericarp, pulp, 1390
cactus areolas hairs and/or trichomes). The large amount of pollen, insects and 1391
plant tissues present in several samples indicates that the species are consuming 1392
these items to complement their diet. A higher proportion of fecal samples with 1393
insect fragments (mostly Lepidoptera scales) were documented in the wet 1394
season for all nectar bat species. 1395
There are significant differences between the snout length of the different 1396
nectar bat species and fecal composition (Figure 8). The fecal composition of 1397
G.soricina (shortest snout) was dominated by pollen and insects. No fecal 1398
samples of G. soricina contained exclusively pollen, contrary to X. vieirai (longest 1399
snout) who presented the highest percentage of samples composed exclusively 1400
of pollen, which could be considered a suggestion for its diet specialization in 1401
nectarivory/pollinivory (Figure 7, 8). The Lonchophylla species, with their 1402
intermediate snout size, presented fecal composition more similar to G. soricina. 1403
DISCUSSION 1404
The nectar-feeding bat ensemble in this Caatinga area consists of four resident 1405
species coexisting and persisting year-round, thus flowing with the seasonal 1406
rhythm of Caatinga’s resources offer. The nectar bat species exhibit similar 1407
ecologies in sympatry yet showing subtle but significant differences in 1408
morphological attributes (weight, forearm, snout and wing aspect ratio), in 1409
capture rates within the night and also differences in resource use. The studied 1410
nectar bat ensemble presents some sort of specialization on a ‘primary’ food type 1411
(nectar and pollen of Cactaceae) while retaining omnivorous capabilities to 1412

83
exploit in different frequency supplementary food items such as insects, plant 1413
tissues and nectar of ‘secondary’ plant species. 1414
Nectar-feeding bat ensemble 1415
The nectar bat ensemble in the Caatinga of Lajes is species rich when compared 1416
to other Caatinga areas. Previous studies on the structure of bat communities in 1417
Caatinga reported ensembles of nectar bats mostly comprised by two species 1418
(Silva, Guedes & Peracchi, 2001; Gregorin et al., 2008; Beltrão et al., 2015; 1419
Soares et al., 2018a; 2018b; Feijó & Rocha 2017), three species (Sá-Neto & 1420
Marinho-Filho, 2013; Rocha et al., 2015; Silva et al., 2015) and less frequently by 1421
four (Rocha, Ruiz-Esparza & Ferrari, 2017) and five species (Cordero-Schmidt et 1422
al., 2017). 1423
The studied ensemble is mainly conformed by Lonchophyllinae species (L. 1424
mordax, L. inexpectata and Xeronycteris vieirai) and a single Glossophaginae 1425
species (Glossophaga soricina). Anoura geoffroyi (Glossophaginae) is another 1426
nectar bat species reported for several localities in Caatinga (Gregorin et al., 1427
2008; Silva et al., 2015; Rocha et al., 2017; Feijó & Rocha, 2017; Soares et al., 1428
2018b) including our study site (Cordero-Schmidt et al., 2017), withal, A. geoffroyi 1429
captures are generally low. We considered this species as "occasional" or rare 1430
species of our studied nectar bat ensemble, but due to the lack of captures during 1431
the study period our discussion of this species is limited. 1432
Glossophaga soricina is one of the most commonly encountered nectar 1433
bat species in the Neotropics (Fleming, Muchhala & Ornelas, 2005) and so it was 1434
in our study. Its flexible diet, generalized habitat requirements and its ability to 1435
defend feeding territories is what makes G. soricina successful and widespread 1436
in both preserved and highly modified habitats (Lemke, 1985; Clare et al., 2014). 1437

84
This species is the second biggest (weight and forearm) in our ensemble, it has 1438
the shortest snout and their wings showed the lowest ARI and WTI. 1439
Lonchophylla inexpectata is a recently described poorly known Caatinga 1440
endemic species (Moratelli & Dias, 2015). Moratelli & Dias (2015) stated this 1441
species was for many years misidentified as L. mordax due to their morphological 1442
resemblance, therefore we could expect that its distribution and abundance have 1443
been underestimated in other areas. Lonchophylla mordax despite being a 1444
relatively common species in Caatinga (Sá-Neto & Marinho-Filho, 2013; Rocha 1445
et al., 2015), there is little information on its biology and diet (Bredt, Uieda & 1446
Pedro, 2012). The morphological similarities between both Lonchophylla species 1447
were confirmed with our data on the species weight, forearm, ARI, WTI and snout 1448
length. However, some differences in the use of resources were observed mainly 1449
during the wettest months (higher resource availability). 1450
Xeronycteris vieirai has recently been classified as endemic to the dry 1451
diagonal crossing central Brazil including Caatinga and Cerrado (Dias & Oliveira, 1452
2019). Information on the natural history of X. vieirai has been compiled by 1453
Cordero-Schmidt et al., (2017) and Gomes et al., (2018). Its recent increase in 1454
distribution may indicate that this species is widely distributed, however it seems 1455
to be a locally rare species in some areas of Caatinga and relatively common in 1456
others (Cordero-Schmidt et al. in prep.). In comparison with the other species of 1457
nectar bats, this species is the largest (weight and forearm), it also has the 1458
longest snout and the highest ARI. This species can be classified as “obligate 1459
cactophile” (sensu Simmons & Wetterer, 2002) due to their predominantly 1460
dependence on cactus products (nectar and pollen) through the year. 1461

85
Palmer, Staton & Young (2003) stated that the mechanisms allowing 1462
species coexistence should be considered as a set of mechanisms and not as 1463
individual ones. We discuss three non-exclusive mechanisms potentially 1464
facilitating coexistence and persistence of G. soricina, L. inexpectata, L. mordax 1465
and X. vieirai in Caatinga. 1466
Temporal persistence mediated by resource availability and keystone 1467
plants 1468
Seasonal changes in the composition of nectar bats have been observed in 1469
different types of habitats in the tropics, with altitudinal or latitudinal migrations 1470
(Sosa & Soriano, 1993; Tschapka, 2004), some species behave as common 1471
persistent residents (core) and others that come and go (satellite) related to 1472
resource availability (Fleming, Muchala & Ornelas, 2005). In Caatinga, Rocha et 1473
al. (2015) observed that two species (G. soricina and L. mordax) were captured 1474
year-round, nonetheless, they were significantly more abundant in the wet 1475
season. Authors linked nectar bats year-round presence to the local abundance 1476
of specific chiropterophilic resources. We did not observe any seasonal changes 1477
in the composition and abundance of nectar bats in our studied Caatinga area. 1478
In the same way we observed available floral resources throughout the year. 1479
The continuity of the floral resources offered by plants in Caatinga has 1480
been suggested to be a plant strategy to maintain pollinator populations year-1481
round (Machado, Barros & Sampaio, 1997; Lima & Rodal, 2010; Quirino & 1482
Machado, 2014). This has been verified for populations of hummingbirds (Las-1483
Casas, Azevedo & Dias, 2012) and for bees (Novais, Absy & Santos, 2014) and 1484
now our study confirms it for nectar bats. In other semiarid areas of the 1485
neotropics, Cactaceae and Agavaceae species have been pointed as keystone 1486

86
species for maintaining nectar-feeding bat populations in lean times (Fleming, 1487
Nuñez, & Sternberg,1993; Sosa & Soriano, 1993; Petit, 1996). In our study site 1488
Pilosocereus pachycladus (Cactaceae), displayed floral resources throughout 1489
the year (though with higher resource availability in the wet season) and was the 1490
most used plant species by the four nectar bat species, besides, this cactus is 1491
one of the most abundant plant species in the area. Species of the genus 1492
Pilosocereus have been highlighted by having close relations with bats in 1493
Caatinga (Rocha, Machado & Zappi, 2007; Rocha et al., 2019). Together with P. 1494
pachycladus other additional four Cactaceae species (P. gounellei, P. 1495
chrysostele, C. jamacaru and M. zehntneri) were central plant species in the diet 1496
of the nectar bat ensemble during both seasons. We suggest that the Cactaceae 1497
family, with particular emphasis on P. pachycladus, are keystone plant resources 1498
in Caatinga. Peres (2010) described keystone species attributes such as local 1499
abundance, temporarily reliable availability of resources and extreme generalism. 1500
Most of the consumed Cactaceae present local abundance (except for C. 1501
jamacaru and P. chrysostele), year-to-year predictable phenology (comparing 1502
this study data to personal observations in the field for the past four years), plus 1503
they are consumed by all nectar bat species, in addition to other vertebrate 1504
species (Leal et al., 2017). 1505
Temporal partitioning within the night and bat mobility 1506
Temporally based resource niche partitioning, such as differences in the hours of 1507
activity in the exploration of resources is expected for bats that feed on quickly 1508
renewable resources (Aguiar & Marinho-Filho, 2004), such as nectar in the 1509
flowers. This behavior has been observed in nectar bats (Solmsen, 1998). In our 1510
study while we captured the four species of nectar bats throughout the night, 1511

87
some differences were observed suggesting that time partitioning could be a 1512
coexisting mechanism in our ensemble. Xeronycteris vieirai was the species that 1513
presented a more differentiated pattern. This species showed a low but constant 1514
capture rate throughout the night, this could be related to its size and therefore 1515
to its energy requirements and the characteristics of the nectar they are feeding 1516
off. 1517
Foraging modes and habitat use in bats are mainly determined by wing 1518
shape and size (Findley, Studier & Wilson, 1972) and by body size (Norberg, 1519
1994). A big body size and faster flight can be considered as an asset when less 1520
resources are available and sparse, (Norberg, 1994), such might be the case for 1521
X. vieirai. This species is the largest species in our study and it is also the most 1522
specialist in nectar/pollen diet. Nectar specialists are forced by flowering plants 1523
to continuously search for new nectar sources since nectar rewards are barely 1524
large enough to satisfy bats energetic requirements (Voigt & Speakman, 2007). 1525
The sugar concentration of the consumed nectar must be taking into 1526
consideration when understanding the energetics involved in increased foraging 1527
time (Ayala-Berdon et al., 2011), such as the presented by X. vieirai. Pilosocereus 1528
pachycladus was the most used plant species by X. vieirai (present in 67.54% of 1529
the samples), this species has a diluted nectar (11-17 % of sugar concentration 1530
Rocha et al., 2019). Our results are in accordance with Ayala-Berdon et al. (2011) 1531
where the volumetric intake of low sugar concentrated nectars (5-15 %) explains 1532
the increase in the total nightly feeding time. 1533
Xeronycteris vieirai showed the highest ARI (longer and slender wings), 1534
this characteristic has been discussed for other nectar-feeding bat species as the 1535
ability to fly faster and respond quickly to season changes in local resource 1536

88
availability (Sperr et al., 2011). During the dry season (less resource available) in 1537
our study, X. vieirai was the species that fed the most on H. neesianus and 1538
Pseudobombax type, these plant species were not observed within our 1539
phenology transects, nor close to our sampling sites (˜5 km), suggesting X. vieirai 1540
is capable of flying longer distance looking for resources. 1541
The lower ARI (shorter and wider wings) presented by G. soricina, L. 1542
inexpectata and L. mordax may promote lift at low speeds and ease maneuvering 1543
(Findley, Studier & Wilson, 1972). Even small differences in wing proportions 1544
between nectar bats have proven to be particularly important in the energy 1545
demanding foraging efficiency (Tschapka, 2004). Melocactus zehntneri, D. ciliaris 1546
and R. asperula are ornithophilic species with smaller and more delicate flowers 1547
when compared to a typical chiropterophilus species with big and robust flowers. 1548
Feeding on these small flowered species probably would require better hovering 1549
aptitudes (Fleming and Muchhala, 2008) such as the observed in G. soricina, and 1550
the Lonchophylla species inferred by their ARI values. Moreover, capture rate of 1551
these species is higher during the first hour after sunset (18:00 hrs.); to be able 1552
to feed on the nectar of the mentioned ornithophilic species, bats must visit them 1553
right after they start foraging. The nectar of these plant species could be the right 1554
intake nectar bats need to get their night started. Welch & Suarez (2008) found 1555
that G. soricina can use recently ingested sugars to provide ∼78% of the fuel 1556
required for oxidative metabolism during their energetically expensive hovering 1557
flight. 1558
Although studies on the energy requirements and foraging of each species 1559
in relation to the spatial and temporal availability of resources are necessary, the 1560
nightly temporal partitioning together with morphological differences (weight size 1561

89
and wing indexes) observed in the four nectar bat species could be supporting 1562
another mechanism of coexistence for nectar bats in Caatinga. 1563
Resource partitioning and use mediated by ecomorphological differences 1564
Ecomorphological patterns between plant-pollinator reflects at least two 1565
processes: interspecific competition and mutualistic coevolution (Palmer, Stanton 1566
& Young, 2003). These processes are common among coexisting hummingbird 1567
species and have been proven to be rare or absent for nectar bat species (von 1568
Helversen & Winter, 2005). Yet, small morphological differences may allow 1569
sympatric nectar bats to exploit different microhabitats, resulting in the 1570
consumption of different food items and resource partitioning (Tschapka, 2004; 1571
Gonzales-Terrazas et al., 2012). In our study bats are feeding on pollen, insects 1572
and other unidentified plant tissues with regularity along the different seasons 1573
and some species-specific differences in use of these items were observed 1574
indicating a possible resource partitioning strategy, 1575
-Snout length and diet composition 1576
Lonchophylla inexpectata, L. mordax and X. vieirai included more insects during 1577
the wet season coinciding with the insect’s maximum abundance in Caatinga 1578
(Vasconcellos et al., 2010). On the other hand, G. soricina consumed similar 1579
proportion of insects during both seasons. Clare et al. (2014) determined that G. 1580
soricina’s diet flexibility is explained by both morphology (shorter snout and high 1581
maneuverability ease by low wing ARI) and sensory characteristics (low-intensity 1582
echolocation). Species with shorter snout vary their diet the most contrary to 1583
longer snouts with a more nectar restricted diet (Solmsen, 1998). 1584
The diet and snout of X. vieirai differentiated the most when compared to 1585
the other nectar bat species. Even though X. vieirai also feeds on insects, it does 1586

90
so in a smaller proportion as the other nectar bat species, also explained by the 1587
extremely reduced dentition (Nogueira et al., 2014) in comparisson to the other 1588
studied nectar bat species. Further, it seems to be the most specialized in nectar 1589
feeding which coincides with the inference made by Gregorin and Ditchfield 1590
(2005) and Cordero-Schmidt et al. (2017). Xeronycteris vieirai was also the 1591
species that included pollen in its diet the most (in 63.63% of the samples) and 1592
through the year. Pollen is an important source of proteins, vitamins, and 1593
minerals, but the efficiency digestion of pollen contents is different within bat 1594
species (Herrera & Martínez, 1998). Authors stated that bat species that regularly 1595
include pollen in their diet, as the case of X. vieriai in our study, should be more 1596
efficient at extracting pollen contents than species that ingest pollen only 1597
seasonally presenting another hint of resource partitioning as a coexistence 1598
facilitator. 1599
Glossphaga soricina complemented its nectar-feeding diet with pollen, 1600
insects and other plant tissues, it was the only species that fed on P. pachycladus 1601
fruits. We expected to find more evidence of fruit use as it is a resource commonly 1602
used by nectarivorous bats when nectar resources are scarce (Fleming, Nuñez 1603
& Sternberg, 1993; Godínez-Alvarez & Valiente-Banuet, 2000), but since there is 1604
nectar and pollen are available through the year, it seems that nectar bat species 1605
can rely on a flower-resource based diet. 1606
-Nectar sugar concentration and resource use 1607
Tschapka, Gonzales-Terrazas & Knörnschild (2015) proposed that differences in 1608
nectar efficiency extraction linked to different tongue morphologies between 1609
species belonging to Glossophaginae and Lonchophyllinae could mediate other 1610
coexistence mechanism; where Glossophaginae species would feed more on 1611

91
high sugar concentration nectar extracted more efficiently with ‘viscous dipping’ 1612
mechanism, contrary to Lonchophyllinae feeding on low sugar concentration 1613
nectar using a ‘pumping-tongue’ mechanism. Our data does not demonstrate a 1614
clear pattern with respect to this taxonomic mediated coexistence mechanism, 1615
since most plant species with different sugar concentration are used by both 1616
subfamilies with a similar frequency. Nonetheless, when looking into species-1617
specific capacity to reach energetic balance it can be elucidated by the resources 1618
the nectar bat species are feeding on considering the nectar’s sugar 1619
concentration and composition (Ayala-Berdon et al., 2011). Authors highlighted 1620
two mechanisms to obtain energy balance ‘Compensatory feeding’ where 1621
species maintain a constant energy intake in spite of food quality. The species 1622
with this type of energetic balance generally feeds on high sugar concentrated 1623
nectar but also present a wider nectar niche. This could be the case of L. mordax 1624
who feeds the most on the two plant species that presented the highest sugar 1625
concentration (C. jamacaru 30%, and R. aperula 29.25%), at the same time L. 1626
mordax also feeds on more plant species (23) when compared to the other nectar 1627
bats (20-21 plant species). 1628
The uncoupled observed activity dynamic of G. soricina with L. inexpectata 1629
and L. mordax, together with the fact that is the nectar bat species that feed 1630
mostly of low sugar concentrated plant species, might be in accordance to 1631
‘Physiological constrains’ mechanism proposed by Ayala-Berdon & Schondube 1632
(2011). Physiological constrains occurs when species limit the amount of energy 1633
they obtain when feeding on dilute nectars (i.e. less than 15% sugar 1634
concentration). Authors explained that species feeding on dilute nectar must visit 1635
more flowers but at the same time they should also reduce their flight time, finding 1636

92
this way an energetic balance. It would be expected that species with this 1637
physiological constrains will forage in smaller areas with high resource 1638
abundance (Ayala-Berdon et al., 2011). To further conclude energy-mediated 1639
coexisting mechanisms, future studies should focus on better characterizing the 1640
plant specie’s nectar concentration and composition, as well as exploring the 1641
physiological constrains such as gut size, sugar absorption and digestion rates 1642
and water absorption. This energetic characterization of resource use, as well as 1643
observations on foraging behavior would give us valuable information about the 1644
role of nectar bat species in pollen movement and pollination in Caatinga. 1645
Pollination focused studies are urgent since Caatinga suffers from intense 1646
and chronic degradation and vegetation destruction since it is inhabited by 1647
approximately 28.6 million people (Silva et al., 2018). The destruction of the 1648
native vegetation results in the decrease in food resources for different animal 1649
populations, and consequently have a dramatical effect in the environmental 1650
services provided by them (Kearns & Inouye, 1997). 1651
CONCLUSIONS 1652
With this study we were able to have the first insight on the mechanisms of 1653
persistence and coexistence of four species of nectar bats in Caatinga. The 1654
mechanisms detected are complementary and seem to be explained by temporal 1655
partitioning (differences in captures throughout of the night), by ecomorphological 1656
differences (size, weight, snout length, wing aspect ratio) and by resource 1657
partitioning (diet flexibility). The four species of nectar bats present a flexible diet, 1658
consuming more than 20 species of plants with different concentrations of sugar 1659
and with both, chiropterophilic and non-chiropterophilic characteristics. They also 1660
include pollen, insects and other plant tissues. On the other hand, contrary to our 1661

93
expectations, there were no changes in species composition mechanism in the 1662
period of least available resources. Additionally, no evidence of an evolutionary 1663
mediated mechanism (differences in nectar concentration linked to nectar bat 1664
subfamily) was found. 1665
The temporal persistence of nectar bats was facilitated by the continuity 1666
and complementarity of floral resources available during both seasons, however 1667
we highlight the importance of the Cactaceae family in Caatinga. Cacti should be 1668
considered keystone resources and therefore conservation initiatives should 1669
consider sites with high abundance and diversity of Cactaceae as a conservation 1670
priority. 1671
1672
REFERENCES 1673
Aguiar, L. M. D. S., & Marinho-Filho, J. (2004). Activity patterns of nine 1674
phyllostomid bat species in a fragment of the Atlantic Forest in southeastern 1675
Brazil. Revista Brasileira de Zoologia, 21(2), 385-390. 1676
Ayala-Berdon, J., Rodríguez-Peña, N., Orduña-Villaseñor, M., Stoner, K. E., 1677
Kelm, D. H., & Schondube, J. E. (2011). Foraging behavior adjustments 1678
related to changes in nectar sugar concentration in phyllostomid bats. 1679
Comparative Biochemistry and Physiology Part A: Molecular & Integrative 1680
Physiology, 160(2), 143-148. 1681
Ayala-Berdon, J., & Schondube, J. E. (2011). A physiological perspective on 1682
nectar-feeding adaptation in phyllostomid bats. Physiological and 1683
Biochemical Zoology, 84(5), 458-466. 1684

94
Beltrão, M. G., Zeppelini, C. G., Fracasso, M. P. A., & Lopez, L. C. S. (2015). Bat 1685
inventory in a Caatinga area in Northeastern Brazil, with a new occurrence 1686
in the state of Paraíba. Neotropical Biology and Conservation, 10(1), 15-20. 1687
Bredt, A., Uieda, W., & Pedro, W. A. (2012). Plantas e morcegos na recuperação 1688
de áreas degradadas e na paisagem urbana. Brasília: Rede de sementes 1689
do Cerrado, 273. 1690
Carreira, L. M. M. and Barth, O. M. (2003). Atlas de pólen da vegetação de 1691
Canga da Serra de Carajás (Pará, Brasil), Belém, Brazil: Mus. Para. 1692
Emílio Goeldi. 1693
Clare, E. L., Goerlitz, H. R., Drapeau, V. A., Holderied, M. W., Adams, A. M., 1694
Nagel, J., ... & Brock Fenton, M. (2014). Trophic niche flexibility in 1695
Glossophaga soricina: how a nectar seeker sneaks an insect 1696
snack. Functional Ecology, 28(3), 632-641. 1697
Cordero-Schmidt, E., Barbier, E., Vargas-Mena, J. C., Oliveira, P. P., Santos, F. 1698
D. A. R., Medellín, R. A., ... & Venticinque, E. M. (2017). Natural history of 1699
the Caatinga endemic Vieira's flower bat, Xeronycteris vieirai. Acta 1700
chiropterologica, 19(2), 399-408. 1701
Datzmann, T., von Helversen, O., & Mayer, F. (2010). Evolution of nectarivory in 1702
phyllostomid bats (Phyllostomidae Gray, 1825, Chiroptera: Mammalia). 1703
BMC Evolutionary Biology, 10(1), 165. 1704
Díaz, M. M., Solari, S., Aguirre, L. F., Aguiar, L., & Barquez, R. M. (2016). Clave 1705
de Identificación de los murciélagos de Sudamérica–Chave de 1706
identificação dos morcegos da America do Sul. Tucumán, Argentina: 1707
Yerba Buena. 1708

95
Dias, D., & de Oliveira, M. B. (2019). First Record of Xeronycteris vieirai Gregorin 1709
& Ditchfield, 2005 (Chiroptera, Phyllostomidae) for the Cerrado Biome. 1710
Oecologia Australis. 1711
EMPARN- Empresa de Pesquisa Agropecuária do Rio Grande do Norte. Rainfall 1712
monitoring: accumulated monthly rainfall per year by municipality. Retrieved 1713
from:http://meteorologia.emparn.rn.gov.br:8181/monitoramento/monitoram1714
ento.php 1715
Erdtman, G. (1952). Pollen morphology and plant taxonomy. New York, 6-24. 1716
Fadini, R. F., Fischer, E., Castro, S. J., Araujo, A. C., Ornelas, J. F., & de Souza, 1717
P. R. (2018). Bat and bee pollination in Psittacanthus mistletoes, a genus 1718
regarded as exclusively hummingbird-pollinated. Ecology, 99(5), 1239-1719
1241. 1720
Farias Bezerra, G. R. (2016). Geomorfologia da Serra do Feiticeiro em 1721
Lajes/RN. Revista de Geociências do Nordeste, 2, 22-32. 1722
Feijó, A., & da Rocha, P. A. (2017). Morcegos da Estação Ecológica Aiuaba, 1723
Ceará, nordeste do Brasil: uma unidade de proteção integral na 1724
caatinga. Mastozoología neotropical, 24(2), 333-351. 1725
Findley, J. S., Studier, E. H., & Wilson, D. E. (1972). Morphologic properties of 1726
bat wings. Journal of Mammalogy, 53(3), 429-444. 1727
Findley, J. S. (1993). Bats: a community perspective. CUP Archive. 1728
Fleming, T. H., Nuñez, R. A., & Sternberg, L. D. S. L. (1993). Seasonal changes 1729
in the diets of migrant and non-migrant nectarivorous bats as revealed by 1730
carbon stable isotope analysis. Oecologia, 94(1), 72-75. 1731

96
Fleming, T. H., Muchhala, N., & Ornelas, J. F. (2005). New world nectar-feeding 1732
vertebrates: community patterns and processes. Contribuciones 1733
mastozoológicas en homenaje a Bernardo Villa, 163. 1734
Fleming, T. H., & Muchhala, N. (2008). Nectar-feeding bird and bat niches in two 1735
worlds: pantropical comparisons of vertebrate pollination systems. Journal 1736
of Biogeography, 35(5), 764-780. 1737
Freeman, P. W. (1995). Nectarivorous feeding mechanisms in bats. Biol. J. Linn. 1738
Soc. Lond. 56, 439-463. 1739
Greenacre, M. J. (1984). Correspondence analysis. London: Academic Press. 1740
Gregorin, R., Carmignotto, A. P., & Percequillo, A. R. (2008). Quirópteros do 1741
Parque Nacional da Serra das Confusões, Piauí, nordeste do 1742
Brasil. Chiroptera Neotropical, 14(1), 366-383. 1743
Gregorin, R., & Ditchfield, A. D. (2005). New genus and species of nectar-feeding 1744
bat in the tribe Lonchophyllini (Phyllostomidae: Glossophaginae) from 1745
northeastern Brazil. Journal of Mammalogy, 86(2), 403-414. 1746
Godínez-Alvarez, H., & Valiente-Banuet, A. (2000). Fruit-Feeding Behavior of the 1747
Bats Leptonycteris curasoae and Choeronycteris mexicana in Flight Cage 1748
Experiments: Consequences for Dispersal of Columnar Cactus Seeds1. 1749
Biotropica, 32(3), 552-557. 1750
Gomes, L. A., Maas, A. C., Godoy, M. S., Martins, M. A., Pedrozo, A. R., & 1751
Peracchi, A. L. (2018). Ecological considerations on Xeronycteris vieirai: an 1752
endemic bat species from the Brazilian semiarid 1753
macroregion. Mastozoología Neotropical, 25(1), 81-88. 1754
Gonzalez-Terrazas, T. P., Medellin, R. A., Knörnschild, M., & Tschapka, M. 1755
(2012). Morphological specialization influences nectar extraction efficiency 1756

97
of sympatric nectar-feeding bats. Journal of Experimental Biology, 215(22), 1757
3989-3996. 1758
Heithaus, E. R., Fleming, T. H., & Opler, P. A. (1975). Foraging patterns and 1759
resource utilization in seven species of bats in a seasonal tropical 1760
forest. Ecology, 56(4), 841-854. 1761
Heithaus, E. R. (1982). Coevolution between bats and plants. In Ecology of bats 1762
(pp. 327-367). Springer, Boston, MA. 1763
Herrera M, L. G., & Martínez Del Río, C. (1998). Pollen digestion by New World 1764
bats: effects of processing time and feeding habits. Ecology, 79(8), 2828-1765
2838. 1766
Janzen, D. H. 1973. Sweep samples of tropical foliage insects: effects of 1767
seasons, vegetation types, elevation, time of day, and insularity. Ecology 1768
54:687-708. 1769
Jorgensen, S. E., & Fath, B. D. (2014). Encyclopedia of ecology. Newnes. 1770
Kearns, C. A., & Inouye, D. W. (1997). Pollinators, flowering plants, and 1771
conservation biology. Bioscience, 47(5), 297-307. 1772
Kunz, T. H. (1973). Resource utilization: temporal and spatial components of bat 1773
activity in central Iowa. Journal of Mammalogy, 54(1), 14-32. 1774
Las-Casas, F. M. G., Azevedo Júnior, S. M., & Dias Filho, M. M. (2012). The 1775
community of hummingbirds (Aves: Trochilidae) and the assemblage of 1776
flowers in a Caatinga vegetation. Brazilian Journal of Biology, 72(1), 51-58. 1777
Leal, I. R., Lopes, A. V., Machado, I. C., & Tabarelli, M. (2017). Plant–animal 1778
interactions in the caatinga: overview and perspectives. In Caatinga (pp. 1779
255-278). Springer, Cham. 1780

98
Lemke, T. O. (1985). Pollen carrying by the nectar-feeding bat Glossophaga 1781
soricina in a suburban environment. Biotropica, 107-111. 1782
Lima, A. L. A., & Rodal, M. J. N. (2010). Phenology and wood density of plants 1783
growing in the semi-arid region of northeastern Brazil. Journal of Arid 1784
Environments, 74(11), 1363-1373. 1785
Lima, A. L. A., & Rodal, M. J. N. (2010). Phenology and wood density of plants 1786
growing in the semi-arid region of northeastern Brazil. Journal of Arid 1787
Environments, 74(11), 1363-1373. 1788
Lund, U., Agostinelli, C., & Agostinelli, M. C. (2017). Package 1789
‘circular’. Repository CRAN. 1790
Lund, U., & Agostinelli, C. CircStats: Circular Statistics, from “Topics in Circular 1791
Statistics”(2001). R package version 0.2-6, 2018. 1792
Machado, I. C., Barros, L. M., & Sampaio, E. V. (1997). Phenology of caatinga 1793
species at Serra Talhada, PE, northeastern Brazil. Biotropica, 29(1), 57-68. 1794
Melhem, T. S., Cruz-Barros, M. A. V., Corrêa, A. M. S., Makino-Watanabe, H., 1795
Silvestre-Capelato, M. S. F. & Gonçalves-Esteves, V. L. (2003). 1796
Variabilidade polínica em plantas de Campos do Jordão (São Paulo, 1797
Brasil). Bol. Inst Bot., 16, 1–104. 1798
Moratelli, R., & Dias, D. (2015). A new species of nectar-feeding bat, genus 1799
Lonchophylla, from the Caatinga of Brazil (Chiroptera, 1800
Phyllostomidae). ZooKeys, (514), 73. 1801
Moreno, C. E., & Halffter, G. (2001). On the measure of sampling effort used in 1802
species accumulation curves. Journal of Applied Ecology, 38(2), 487-490. 1803
Muscarella, R., & Fleming, T. H. (2007). The role of frugivorous bats in tropical 1804
forest succession. Biological reviews, 82(4), 573-590. 1805

99
Nassar, J. M., Beck, H., Sternberg, L. D. S., & Fleming, T. H. (2003). Dependence 1806
on cacti and agaves in nectar-feeding bats from Venezuelan arid 1807
zones. Journal of Mammalogy, 84(1), 106-116. 1808
Nogueira, M. R., Gregorin, R., & Peracchi, A. L. (2014). Emended diagnosis of 1809
Xeronycteris vieirai (Mammalia: Chiroptera), with the first record of 1810
polyodontia for the genus. Zoologia (Curitiba), 31(2), 175-180. 1811
Norberg U.M. 1994. Wing design, Flight Performance, and Habitat Use in Bats. 1812
In: Ecological Morphology (Wainwright P.C. & Reilly S. M., eds.), pp. 205-1813
239. The University of Chicago Press, Chicago and London. 1814
Novais, J. S. D., Absy, M. L., & Santos, F. D. A. R. D. (2014). Pollen types 1815
collected by Tetragonisca angustula (Hymenoptera: Apidae) in dry 1816
vegetation in Northeastern Brazil. European Journal of Entomology, 111(1). 1817
Oliveira, P. T., e Silva, C. S., & Lima, K. C. (2017). Climatology and trend analysis 1818
of extreme precipitation in subregions of Northeast Brazil. Theoretical and 1819
applied climatology, 130(1-2), 77-90. 1820
Palacios-Chávez, R., Ludlow-Wiechers, B., & Villanueva, G. (1991). Flora 1821
palinológica de la reserva de la biosfera de Sian Ka'an, Quintana Roo, 1822
México. Chetumal, CIQRO. 1823
Palmer, T. M., Stanton, M. L., & Young, T. P. (2003). Competition and 1824
coexistence: exploring mechanisms that restrict and maintain diversity 1825
within mutualist guilds. The American Naturalist, 162(S4), S63-S79. 1826
Peres, C. A. (2000). Identifying keystone plant resources in tropical forests: the 1827
case of gums from Parkia pods. Journal of Tropical Ecology, 16(2), 287-1828
317. 1829

100
Petit, S., & Pors, L. (1996). Survey of columnar cacti and carrying capacity for 1830
nectar-feeding bats on Curaçao. Conservation Biology, 10(3), 769-775. 1831
Pianka, E. R. (1974). Niche overlap and diffuse competition. Proceedings of the 1832
National Academy of Sciences, 71(5), 2141-2145. 1833
Quirino, Z. G. M., & Machado, I. C. (2014). Pollination syndromes in a Caatinga 1834
plant community in northeastern Brazil: seasonal availability of floral 1835
resources in different plant growth habits. Brazilian Journal of 1836
Biology, 74(1), 62-71. 1837
Reid, F. A. 2009. A Field Guide to The Mammals of Central America & 1838
Southeast Mexico. 2nd ed. Oxford University Press, New York, 346 pp. 1839
Rocha, E. A., Machado, I. C., & Zappi, D. C. (2007). Floral biology of Pilosocereus 1840
tuberculatus (Werderm.) Byles & Rowley: a bat pollinated cactus endemic 1841
from the “Caatinga” in northeastern Brazil1. Bradleya, 2007(25), 129-145. 1842
Rocha, P. A., Ruiz-Esparza, J., de Souza Ribeiro, A., & Ferrari, S. F. (2015). 1843
Species diversity and seasonal variation in the composition of a bat 1844
community in the semi-arid brazilian caatinga. Acta Scientiarum. Biological 1845
Sciences, 37(2), 197-203. 1846
Rocha, P. A., Ruiz-Esparza, J., & Ferrari, S. F. (2017). Differences in the structure 1847
of the bat community between a cloud forest refuge and a surrounding semi-1848
arid Caatinga scrubland in the northeastern Brazil. Journal of arid 1849
environments, 151, 41-48. 1850
Rocha, E. A., Domingos-Melo, A., Zappi, D. C., & Machado, I. C. (2019). 1851
Reproductive biology of columnar cacti: are bats the only protagonists in the 1852
pollination of Pilosocereus, a typical chiropterophilous genus?. Folia 1853
Geobotanica, 1-18. 1854

101
Roubik, D. W., & Moreno, P. (1991). Pollen and spores of Barro Colorado Island. 1855
Panamá. 1856
Sá-Neto, R. J., & Marinho-Filho, J. (2013). Bats in fragments of xeric woodland 1857
caatinga in Brazilian semiarid. Journal of Arid Environments, 90, 88-94. 1858
Santos, F. D. A. R., Watanabe, H. M., & de Hamburgo Alves, J. L. (1997). Pollen 1859
morphology of some Cactaceae of North-Eastern 1860
Brazil. Bradleya, 1997(15), 84-98. 1861
Sikes, R. S., & Animal Care and Use Committee of the American Society of 1862
Mammalogists. (2016). 2016 Guidelines of the American Society of 1863
Mammalogists for the use of wild mammals in research and 1864
education. Journal of Mammalogy, 97(3), 663-688. 1865
Silva, D. S., Guedes, P. G., & Peracchi, A. L. (2001). Levantamento preliminar 1866
dos morcegos do Parque Nacional de Ubajara (Mammalia, Chiroptera), 1867
Ceará, Brasil. Revista Brasileira de Zoologia, 18(1), 139-144. 1868
Silva, S. S., Dias, D., Martins, M. A., Guedes, P. G., de Almeida, J. C., da Cruz, 1869
A. P., ... & Peracchi, A. L. (2015). Bats (Mammalia: Chiroptera) from the 1870
caatinga scrublands of the Crateus region, northeastern Brazil, with new 1871
records for the state of Ceará. Mastozoología neotropical, 22(2), 335-348. 1872
Silva, F. H. M., Santos, F. A. R. & Lima, L. C. L. (2016) Flora polínica das 1873
caatingas: Estação Biológica de Canudos (Canudos, Bahia, Brasil). Feira 1874
de Santana, Micron Bahia. 1875
Simmons, N. B., & Wetterer, A. L. (2002). Phylogeny and convergence in 1876
cactophilic bats. Columnar cacti and their mutualists: evolution, ecology, 1877
and conservation, 87-121. 1878

102
Soares, F. A., Da Rocha, P. A., Bocchiglieri, A., & Ferrari, S. F. (2018a). Structure 1879
of a bat community in the xerophytic Caatinga of the state of Sergipe, 1880
Northeastern Brazil. Mammalia, 83(2), 125-133. 1881
Soares, F., da Rocha, P. A., Ferrari, S. F., Acioli, T., & Sobrinho, J. P. P. (2018b). 1882
Bats (Mammalia, Chiroptera) from an area of Caatinga in southwestern 1883
Bahia, Brazil. Pesquisa e Ensino em Ciências Exatas e da Natureza, 2(1), 1884
7-16. 1885
Solari, S., & Martínez-Arias, V. (2014). Cambios recientes en la sistemática y 1886
taxonomía de murciélagos Neotropicales (Mammalia: 1887
Chiroptera). Therya, 5(1), 167-196. 1888
Solmsen, E. H. (1998). New world nectar-feeding bats: biology, morphology and 1889
craniometric approach to systematics. Zoologisches Forschungsinstitut und 1890
Museum Alexander Koenig Bonn. Ed 44. Hamburg, Germany. 1891
Soriano, P. J., Sosa, M., & Rossell, O. (1991). Hábitos alimentarios de 1892
Glossophaga longirostris Miller (Chiroptera: Phyllostomidae) en una zona 1893
árida de los Andes venezolanos. Revista de Biología Tropical, 263-268. 1894
Sosa, M., & Soriano, P. J. (1993). Solapamiento de dieta entre Leptonycteris 1895
curasoae y Glossophaga longirostris (Mammalia: Chiroptera). Revista de 1896
Biología Tropical, 529-532. 1897
Sperr, E. B., Caballero-Martinez, L. A., Medellin, R. A. and Tschapka, M. (2011). 1898
Seasonal changes in species composition, resource use and reproductive 1899
patterns within a guild of nectar-feeding bats in a west Mexican dry forest. 1900
J. Trop. Ecol. 27, 133-145. 1901

103
Straube, F. C., & Bianconi, G. V. (2002). Sobre a grandeza e a unidade utilizada 1902
para estimar esforço de captura com utilização de redes-de-1903
neblina. Chiroptera Neotropical, 8(1-2), 150-152. 1904
Tschapka, M., & Dressler, S. (2002). Chiropterophily: On bat-flowers and flower-1905
bats. Curtis's Botanical Magazine, 19(2), 114-125. 1906
Tschapka, M. (2004). Energy density patterns of nectar resources permit 1907
coexistence within a guild of Neotropical flower-visiting bats. Journal of 1908
Zoology, 263(1), 7-21. 1909
Tschapka, M., Sperr, E., Caballero-Martinez, L. A. and Medellin, R. A. (2008). 1910
Diet and cranial morphology of Musonycteris harrisoni, a highly specialized 1911
nectar-feeding bat in western Mexico. J. Mammal. 89, 924-931. 1912
Tschapka, M., Gonzalez-Terrazas, T. P., & Knörnschild, M. (2015). Nectar uptake 1913
in bats using a pumping-tongue mechanism. Science advances, 1(8), 1914
e1500525. 1915
Valiente-Banuet, A., Arizmendi, M. D. C., Rojas-Martínez, A., & Domínguez-1916
Canseco, L. (1996). Ecological relationships between columnar cacti and 1917
nectar-feeding bats in Mexico. Journal of Tropical Ecology, 12(1), 103-119. 1918
Vasconcellos, A., Andreazze, R., Almeida, A. M., Araujo, H. F., Oliveira, E. S., & 1919
Oliveira, U. (2010). Seasonality of insects in the semi-arid Caatinga of 1920
northeastern Brazil. Revista Brasileira de Entomologia, 54(3), 471-476. 1921
Vogel, S., Lopes, A.V., and Machado, I.C. (2005). Bat pollination in the NE 1922
Brazilian endemic Mimosa lewisii: an unusual case and first report for the 1923
genus. Taxon 54, 693e700. 1924

104
Voigt, C. C., & Speakman, J. R. (2007). Nectar-feeding bats fuel their high 1925
metabolism directly with exogenous carbohydrates. Functional 1926
Ecology, 21(5), 913-921. 1927
Voigt C., D. Kelm, B. Bradley & S. Ortmann. 2009. Dietary Analysis of plat-visiting 1928
bats. In: T. H. Kunz & S. Parsons (Eds.), Ecological and Behavioral 1929
Methods for the Study of Bats (2nd ed., pp. 593-609). Baltimore: Johns 1930
Hopkins University Press. 1931
von Helversen, O. and Winter, Y. (2005). Glossophagine bats and their flowers: 1932
costs and benefits for plants and pollinators. In Bat Ecology (ed. T. H. Kunz 1933
and M. B. Fenton), pp. 346-398. Chicago, IL: The University of Chicago 1934
Press. 1935
Welch, K. C., & Suarez, R. K. (2008). Dietary sugar as a direct fuel for flight in 1936
the nectarivorous bat Glossophaga soricina. Journal of Experimental 1937
Biology, 211(3), 310-316. 1938
Winter, Y., & Von Helversen, O. (1998). The energy cost of flight: do small bats 1939
fly more cheaply than birds? Journal of Comparative Physiology B, 168(2), 1940
105-111. 1941
1942

105
1943 1944 Figure 1. Local nectar-feeding bat guild: A. Glossophaga soricina; B. 1945 Lonchophylla inexpectata; C. Lonchophylla mordax; D. Xeronycteris vieirai. 1946 1947 1948
1949 1950 Figure 2. Graphic representation of the first two axes of a NMDS performed for 1951 all species of nectar-feeding bats captures between years (A) and seasons (B) 1952 first axis retained 68.6% of the total variation and second axis 45.3% 1953 Stress=0.096. 1954 1955

106
1956 1957 Figure 3. Capture rates from 18:00 to 24:00. A. Nectar-feeding bat capture 1958 composition per hour, and B. Captured individuals for each species. 1959

107
Table 1A. Morphological measurements (W= Weight, FA= Forearm, ARI=Aspect 1960 Ratio Index, WTI=Wing Tip Index and snout) of the four nectar-feeding bat 1961 species. N= individuals with data, B. ANOVA of morphological measurements of 1962 the four nectar-feeding bat species. 1963 1964
A. Glossophaga soricina Lonchophylla inexpectata W (g) FA (mm) ARI WTI Snout W (g) FA (mm) ARI WTI Snout N 117 126 126 126 126 65 69 69 69 69 Minimum 5.00 30.00 1.26 1.34 6.00 5.00 31.00 1.26 1.37 8.00 Maximum 17.00 39.50 2.27 2.23 10.00 13.00 38.00 2.31 2.16 10.00 Mean 9.98 35.89 2.11 1.83 7.33 8.47 35.06 2.11 1.84 8.99 SD 1.29 1.26 0.12 0.09 0.56 1.10 1.46 0.12 0.10 0.70
Lonchophylla mordax Xeronycteris vieiriai W (g) FA (mm) ARI WTI Snout W (g) FA (mm) ARI WTI Snout N 90 107 107 107 107 68 74 74 74 74 Minimum 5.00 23.30 1.76 1.32 7.00 6.50 35.50 2.08 1.69 11.00 Maximum 11.50 38.00 2.33 2.88 11.00 16.00 40.50 2.45 2.06 14.00 Mean 8.73 35.38 2.12 1.85 8.84 11.87 38.47 2.17 1.90 11.99 SD 1.21 1.67 0.07 0.13 0.66 1.77 0.96 0.05 0.05 0.55
1965
B. Type III SS df Mean
Squares F-ratio p-value R2 Parwise difference
Weight 579.99 3 193.33 78.42 <0.001 0.39 Xv > Gs > Li =Lm Error 917.13 372 2.47
Forearm 551.08 3 183.69 96.50 <0.001 0.44 Xv > Gs > Li =Lm Error 708.15 372 1.90
ARI 0.20 3 0.07 7.64 <0.001 0.06 Xv > Lm = Li = Gs Error 3.29 372 0.01
WTI 0.07 3 0.02 1.22 0.31 0.01 Equal Error 7.09 372 0.02
Snout 1018.92 3 339.64 896.38 <0.001 0.88 Xv > Li = Lm > Gs Error 140.96 372 0.3789
1966 1967

108
1968 D 1969
1970 1971 Figure 4. Circular histogram of monthly (except May 2018) flowering phenology. 1972 The height of the bars represents the number of species in bloom per month. A. 1973 All studied plant species. B. Four chiropterophilous plant species: Pilosocereus 1974 chrysostele, P. gounellei, P. pachycladus and Tareyana spinosa. C. Five non-1975 chiropterophilous species: R. asperula, T. aurea, C. jamacaru, M. zehntneri and 1976 J. mollissima. D. Linear representation of flowering phenology of nine plant 1977 species in Caatinga. The solid line indicates the moments where > 50% of the 1978 monitored individuals had flowers; Dotted line indicates moment where < 50% of 1979 individuals had flowers; Blank sections indicate individuals with no observed 1980 available flowers. 1981 1982 Table 2. Circular statistical analyses testing for the occurrence of seasonality on 1983 flowering phenological behavior of nine plant species in Caatinga. The average 1984 date (angle) is the time of the year around which the dates of flowering occurred 1985 for most species. Rayleigh test was performed for significance of the mean angle. 1986 1987
Plants N Average date (Angle) r Z Rayleigh
(p) Rao (p)
Chiropterophilous 33 86.21 0.120 0.475 . <0.001
Non-chiropterophilous 30 6.97 0.124 0.467 0.630 .
All 63 47.93 0.094 0.559 0.571 . 1988 1989
Chiropterophilous Dec17 Jan18 Feb18 Mar18 Apr18 May18 Jun18 Jul18 Aug18 Sep18 Oct18 Nov18
Tareyana spinosaPilosocereus chrysostele
Pilosocereus gounellei Pilosocereus pachycladus
Non-Chiropterophilous
Ruellia asperulaTabebuia aurea
Cereus jamacaruMelocactus zehntneri
Jatropha mollissima
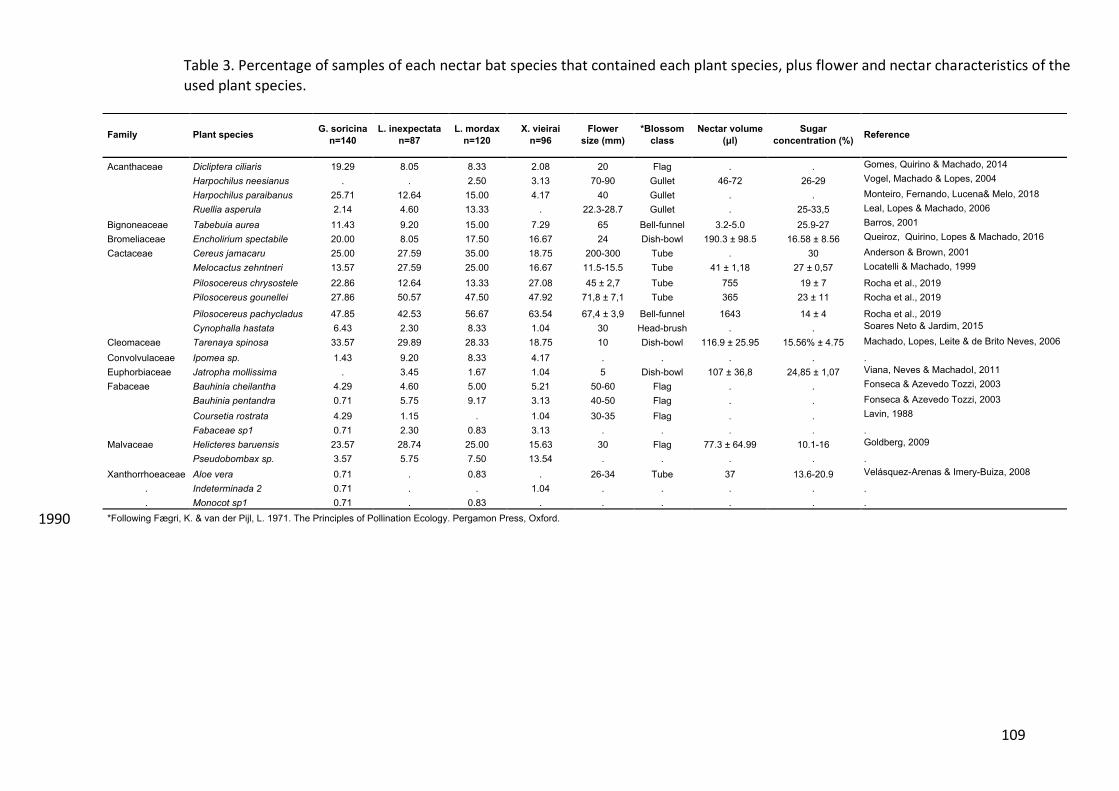
109
1990
Family Plant species G. soricina
n=140 L. inexpectata
n=87 L. mordax
n=120 X. vieirai
n=96 Flower
size (mm) *Blossom
class Nectar volume
(μl) Sugar
concentration (%) Reference
Acanthaceae Dicliptera ciliaris 19.29 8.05 8.33 2.08 20 Flag . . Gomes, Quirino & Machado, 2014
Harpochilus neesianus . . 2.50 3.13 70-90 Gullet 46-72 26-29 Vogel, Machado & Lopes, 2004
Harpochilus paraibanus 25.71 12.64 15.00 4.17 40 Gullet . . Monteiro, Fernando, Lucena& Melo, 2018 Ruellia asperula 2.14 4.60 13.33 . 22.3-28.7 Gullet . 25-33,5 Leal, Lopes & Machado, 2006
Bignoneaceae Tabebuia aurea 11.43 9.20 15.00 7.29 65 Bell-funnel 3.2-5.0 25.9-27 Barros, 2001
Bromeliaceae Encholirium spectabile 20.00 8.05 17.50 16.67 24 Dish-bowl 190.3 ± 98.5 16.58 ± 8.56 Queiroz, Quirino, Lopes & Machado, 2016
Cactaceae Cereus jamacaru 25.00 27.59 35.00 18.75 200-300 Tube . 30 Anderson & Brown, 2001 Melocactus zehntneri 13.57 27.59 25.00 16.67 11.5-15.5 Tube 41 ± 1,18 27 ± 0,57 Locatelli & Machado, 1999
Pilosocereus chrysostele 22.86 12.64 13.33 27.08 45 ± 2,7 Tube 755 19 ± 7 Rocha et al., 2019 Pilosocereus gounellei 27.86 50.57 47.50 47.92 71,8 ± 7,1 Tube 365 23 ± 11 Rocha et al., 2019
Pilosocereus pachycladus 47.85 42.53 56.67 63.54 67,4 ± 3,9 Bell-funnel 1643 14 ± 4 Rocha et al., 2019 Cynophalla hastata 6.43 2.30 8.33 1.04 30 Head-brush . . Soares Neto & Jardim, 2015
Cleomaceae Tarenaya spinosa 33.57 29.89 28.33 18.75 10 Dish-bowl 116.9 ± 25.95 15.56% ± 4.75 Machado, Lopes, Leite & de Brito Neves, 2006
Convolvulaceae Ipomea sp. 1.43 9.20 8.33 4.17 . . . . . Euphorbiaceae Jatropha mollissima . 3.45 1.67 1.04 5 Dish-bowl 107 ± 36,8 24,85 ± 1,07 Viana, Neves & MachadoI, 2011
Fabaceae Bauhinia cheilantha 4.29 4.60 5.00 5.21 50-60 Flag . . Fonseca & Azevedo Tozzi, 2003
Bauhinia pentandra 0.71 5.75 9.17 3.13 40-50 Flag . . Fonseca & Azevedo Tozzi, 2003
Coursetia rostrata 4.29 1.15 . 1.04 30-35 Flag . . Lavin, 1988
Fabaceae sp1 0.71 2.30 0.83 3.13 . . . . . Malvaceae Helicteres baruensis 23.57 28.74 25.00 15.63 30 Flag 77.3 ± 64.99 10.1-16 Goldberg, 2009
Pseudobombax sp. 3.57 5.75 7.50 13.54 . . . . . Xanthorrhoeaceae Aloe vera 0.71 . 0.83 . 26-34 Tube 37 13.6-20.9 Velásquez-Arenas & Imery-Buiza, 2008
. Indeterminada 2 0.71 . . 1.04 . . . . .
. Monocot sp1 0.71 . 0.83 . . . . . . *Following Fægri, K. & van der Pijl, L. 1971. The Principles of Pollination Ecology. Pergamon Press, Oxford.
Table 3. Percentage of samples of each nectar bat species that contained each plant species, plus flower and nectar characteristics of the used plant species.

110
Table 4. Bray-Curtis dissimilarity matrix comparing diet between nectar-feeding 1991 bat species based on pollen samples. Darker blue and highest value indicate the 1992 most dissimilar diets, whilst darker red and lowest value indicate the most similar 1993 diets. GS: Glossophaga soricina, LI: Lonchophylla inexpectata, LM: Lonchophylla 1994 mordax and XV: Xeronycteris vieirai. 1995 1996
1997 1998 1999 Table 5. Results of Correspondence Analysis of the relationships between four 2000 species of nectar-feeding bat and the 24 plants species they feed on in the dry 2001 and wet season. 2002 2003
2004 2005
GS_DRY LI_DRY LM_DRY XV_DRY GS_WET LI_WET LM_WETGS_DRYLI_DRY 0.41LM_DRY 0.33 0.22XV_DRY 0.37 0.32 0.32GS_WET 0.47 0.58 0.46 0.53LI_WET 0.47 0.41 0.38 0.45 0.29LM_WET 0.52 0.54 0.54 0.59 0.33 0.29XV_WET 0.53 0.49 0.43 0.41 0.37 0.31 0.36
Axis 1 2 3 4 5 6 7Eigenvalue 0.21 0.05 0.05 0.03 0.03 0.01 0.01% of total 54.25 13.46 12.15 8.04 6.91 3.48 1.71
Cumulative 54.25 67.71 79.86 87.91 94.81 98.30 100.00
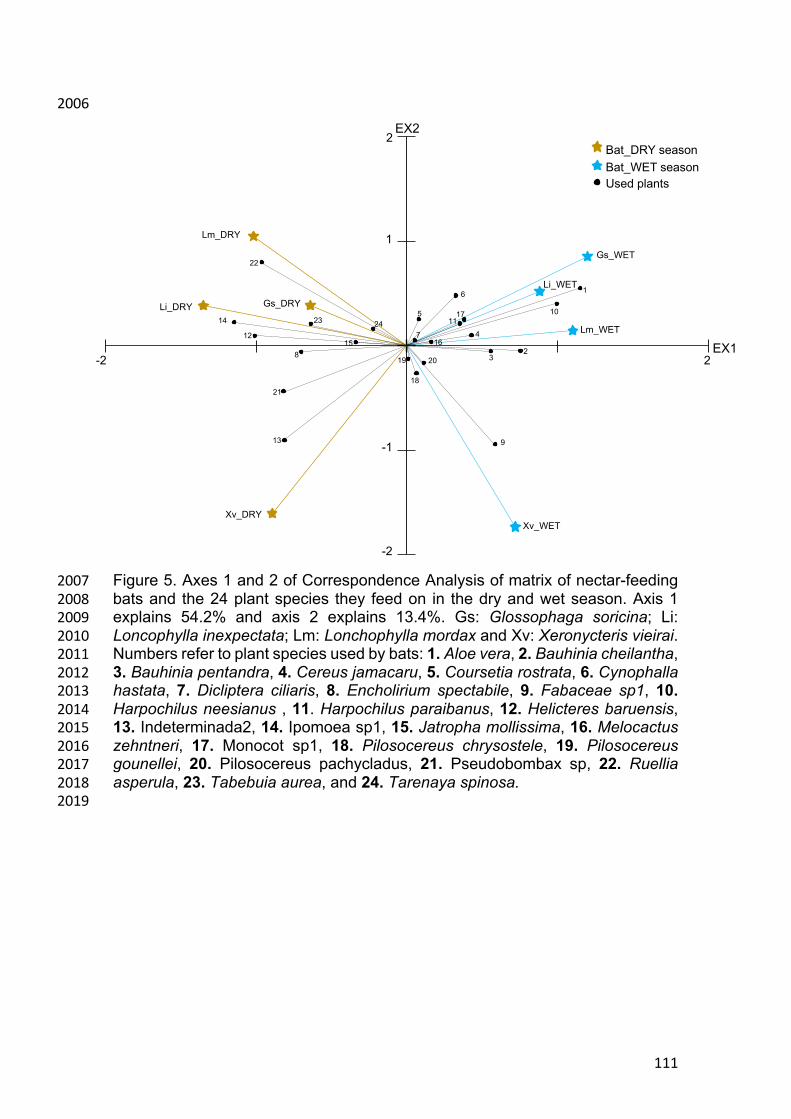
111
2006
Figure 5. Axes 1 and 2 of Correspondence Analysis of matrix of nectar-feeding 2007 bats and the 24 plant species they feed on in the dry and wet season. Axis 1 2008 explains 54.2% and axis 2 explains 13.4%. Gs: Glossophaga soricina; Li: 2009 Loncophylla inexpectata; Lm: Lonchophylla mordax and Xv: Xeronycteris vieirai. 2010 Numbers refer to plant species used by bats: 1. Aloe vera, 2. Bauhinia cheilantha, 2011 3. Bauhinia pentandra, 4. Cereus jamacaru, 5. Coursetia rostrata, 6. Cynophalla 2012 hastata, 7. Dicliptera ciliaris, 8. Encholirium spectabile, 9. Fabaceae sp1, 10. 2013 Harpochilus neesianus , 11. Harpochilus paraibanus, 12. Helicteres baruensis, 2014 13. Indeterminada2, 14. Ipomoea sp1, 15. Jatropha mollissima, 16. Melocactus 2015 zehntneri, 17. Monocot sp1, 18. Pilosocereus chrysostele, 19. Pilosocereus 2016 gounellei, 20. Pilosocereus pachycladus, 21. Pseudobombax sp, 22. Ruellia 2017 asperula, 23. Tabebuia aurea, and 24. Tarenaya spinosa. 2018 2019
-2 2EX1
-2
-1
1
2EX2
Gs_DRYLi_DRY
Lm_DRY
Xv_DRY
Gs_WET
Lm_WET
Xv_WET
1
23
4
5
6
7
8
9
1011
12
13
14
15 16
17
18
19 20
21
22
23 24
Li_WET
Used plantsBat_WET seasonBat_DRY season

112
Figure 6. Axes 2 and 3 of Correspondence Analysis of matrix of nectar-feeding 2020 bat and the 24 plants species they feed on in the dry and wet season. Axis 2 2021 explains 13.4% and axis 3 explains 12.1%. Gs: Glossophaga soricina; Li: 2022 Loncophylla inexpectata; Lm: Lonchophylla mordax and Xv: Xeronycteris vieirai. 2023 Numbers refer to plant species used by bats: 1. Aloe vera, 2. Bauhinia cheilantha, 2024 3. Bauhinia pentandra, 4. Cereus jamacaru, 5. Coursetia rostrata, 6. Cynophalla 2025 hastata, 7. Dicliptera ciliares, 8. Encholirium spectabile, 9. Fabaceae sp1, 10. 2026 Harpochilus neesianus , 11. Harpochilus paraibanus, 12. Helicteres baruensis , 2027 13. Indeterminada2, 14. Ipomoea sp1, 15. Jatropha molíssima, 16. Melocactus 2028 zehntneri, 17. Monocot sp1, 18. Pilosocereus chrysostele, 19. Pilosocereus 2029 gounellei, 20. Pilosocereus pachycladus, 21. Pseudobombax sp, 22. Ruellia 2030 asperula, 23. Tabebuia aurea, and 24. Tarenaya spinosa. 2031 2032 Table 6. ANOSIM for differences in diet composition among the different nectar-2033 feeding bat species. 2034
2035 G. soricina L. inexpectata L. mordax
L. inexpectata 0.3293 L. mordax 0.1622 0.3658 X. vieirai 0.1687 0.0553 0.0124
2036
-2 1 2EX2
-2
2
3EX3
Gs_DRY
Li_DRY
Lm_DRY
Xv_DRY
Gs_WET
Li_WET
Lm_WET
Xv_WET
1
2
3
4
5
6
78
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Used plantsBat_WET seasonBat_DRY season

113
Table 7. Plant species consumed by the nectar-feeding bats and its relative contribution to the Bray-Curtis dissimilarity index; values 2037 calculated by the SIMPER Method. Values for each species are mean abundances. First eight species (grey background) explains 2038 54.3% of the dissimilarity in the bat species. 2039 2040
2041
Pollen type Av. dissim Contrib. %
Cumulative %
Mean Gs_DRY
Mean Li_DRY
Mean Lm_DRY
Mean Xv_DRY
Mean Gs_WET
Mean Li_WET
Mean Lm_WET
Mean Xv_WET
Cereus jamacaru 2.61 8.23 8.23 5.58 4.76 6.61 3.47 13.70 12.40 14.40 10.80Pilosocereus pachycladus 2.61 8.23 16.46 14.50 11.40 16.30 19.40 19.20 16.30 16.60 27.50Helicteres baruensis 2.29 7.23 23.69 7.43 9.52 5.73 6.25 0.00 0.00 0.00 0.00Pilosocereus gounellei 2.11 6.65 30.33 9.29 15.20 13.20 16.70 9.59 18.30 14.40 18.30Encholirium spectabile 1.98 6.24 36.57 8.92 5.71 5.73 7.64 0.69 1.31 0.54 1.67Tarenaya spinosa 1.95 6.16 42.74 13.80 9.52 11.00 9.03 6.85 10.50 4.81 4.17Harpochilus paraibanus 1.88 5.93 48.67 8.92 2.86 3.52 2.08 8.90 9.80 9.09 5.00Harpochilus neesianus 1.79 5.64 54.30 0.00 0.00 0.00 0.69 8.22 5.23 5.35 0.83Pilosocereus chrysostele 1.62 5.11 59.41 7.06 3.81 3.96 9.72 8.90 4.58 3.74 10.00Pseudobombax sp. 1.54 4.87 64.28 1.86 4.76 3.52 7.64 0.00 0.00 0.54 1.67Tabebuia aurea 1.52 4.81 69.08 4.83 6.67 6.17 4.86 2.05 0.65 2.14 0.00Ipomea sp. 1.51 4.76 73.84 0.74 7.62 4.41 2.78 0.00 0.00 0.00 0.00Melocactus zehntneri 1.35 4.24 78.08 3.72 7.62 5.73 4.86 6.16 10.50 9.09 7.50Ruellia asperula 1.21 3.81 81.89 1.12 2.86 7.05 0.00 0.00 0.65 0.00 0.00Dicliptera ciliaris 1.20 3.78 85.67 6.69 1.90 2.64 0.69 2.74 0.65 4.28 4.17Cynophalla hastata 1.02 3.23 88.90 2.60 0.95 3.08 0.69 6.16 3.27 2.14 0.83Bauhinia pentandra 0.99 3.14 92.03 0.00 1.90 0.44 0.00 0.69 1.96 5.35 2.50Bauhinia cheilantha 0.94 2.95 94.99 0.00 0.00 0.88 0.69 4.11 2.61 2.14 3.33Coursetia rostrata 0.47 1.47 96.45 2.23 0.95 0.00 0.69 1.37 0.65 1.60 0.00Fabaceae sp1 0.39 1.23 97.68 0.00 0.00 0.00 0.69 0.00 0.00 1.60 1.67Jatropha mollissima 0.38 1.21 98.89 0.00 1.90 0.00 0.69 0.00 0.65 1.07 0.00Aloe vera 0.13 0.42 99.31 0.00 0.00 0.00 0.00 0.69 0.00 0.54 0.00Indeterminada 2 0.12 0.38 99.68 0.37 0.00 0.00 0.69 0.00 0.00 0.00 0.00Monocot sp1 0.10 0.32 100.00 0.37 0.00 0.00 0.00 0.00 0.00 0.54 0.00

114
2042 Figure 7. Percentage of fecal samples of each nectar-feeding bat species 2043 with different contents. Gs: Glossophaga soricina Ndry=9, Nwet=14; Li: Lonchophylla 2044 inexpectata Ndry=11, Nwet=22; Lm: Lonchophylla mordax Ndry=8, Nwet=21; 2045 Xeronycteris vieirai Ndry=22, Nwet=18. Examples of each category type A. Sample 2046 with at least three pollen types. B. At least two pollen types, insect fragments and moth 2047 scales. C. One pollen type and unidentified green plant tissue. D. Pollen types, moth 2048 scales and unidentified plant tissue. 2049 2050
2051 Figure 8. Relationship between the snout length of the four nectar-feeding bat species 2052 and their diet considering fecal contents. Fecal composition index included three 2053 categories (A. Pollen; B. Pollen & Insects; C. Mix). 2054

115
SUPPORTING INFORMATION 2055 2056 Table S1. Plant species with flowering data (N), growth habit and the relative 2057 abundance of each species in the 15 transects in Lajes, RN. 2058 2059
2060 2061 Table S2. Comparison between capture rates between seasons (dry and wet) and the 2062 four species of nectarivorous bat in the municipality of Lajes RN 2063 2064
Source Type III SS df Mean Squares F-ratio p-value
Season 0.32837 1 0.32837 0.65581 0.42071 Bat species 1.06774 3 0.35591 0.71083 0.54869 Season*Bat
species 0.70723 3 0.23574 0.47083 0.70356
Error 36.05082 72 0.50071 2065 Table S3. Pearson correlation of the capture rates of each nectarivorous bat species 2066 in the municipality of Lajes, RN. Bold values represent significant correlations after the 2067 Bonferroni criterion is applied 2068 2069
G. soricina L. inexpectata L. mordax L. inexpectata 0.50
L. mordax 0.53 0.71
X. vieirai 0.57 0.83 0.69 2070 2071
Family Species Pollination syndrome N AbundanceAcanthaceae Ruellia asperula (Mart. ex Nees) Lindau Ornithophily 2 20Bignoniaceae Tabebuia aurea (Silva Manso) Benth. & Hook.f. ex S.Moore Melittophily 2 5Cactaceae Cereus jamacaru DC. Phalaenophily 9 17
Pilosocereus gounellei (F.A.C.Weber) Byles & Rowley Chiropterophily 40 341Pilosocereus pachycladus F.Ritter Chiropterophily 64 369Pilosocereus chrysostele (Vaupel) Byles & G.D.Rowley Chiropterophily 6 20Melocactus zehntneri (Britton & Rose) Luetzelb. Ornithophily 12 53
Cleomaceae Tarenaya spinosa (Jacq.) Raf. Chiropterophily 5 236Euphorbiaceae Jatropha mollissima (Pohl) Baill. Melittophily/Psychophily 12 302
116
Table S4. MANOVA of the bat composition between years (A) and seasons (B) at 2072 Caatinga in Lajes, Northeast Brazil. 2073 2074
A. Years 2075
2076 B. Seasons 2077
2078 2079 Table S5. Spearman Correlation Matrix between nectar bats captured within the night 2080 2081 Species NMDS I NMDS II G. soricina -0.784 0.122 L. inexpectata -0.669 0.137 L. mordax -0.704 -0.080 X. vieirai -0.448 0.409 Total -0.774 0.163
2082 2083 2084 2085
Source Type III SS df Mean Squares F-ratio p-value
NMDS I 0.01335 1 0.01335 0.43946 0.51627 Error 0.51639 17 0.03038
NMDS II 0.00294 1 0.00294 0.16821 0.68683 Error 0.29733 17 0.01749
Wilks's Lambda=0.96393; F=0.29939; df=2,16; p=0.74533
Source Type III SS df MS F-ratio p-value
NMDS I 0.00952 1 0.00952 0.31455 0.58182 Error 0.54486 18 0.03027
NMDS II 0.01999 1 0.01999 1.25251 0.27779 Error 0.28729 18 0.01596
Wilks's Lambda=0.91777; F=0.76156; df=2,17; p=0.48222

117
2086 Figure S6. Accumulation curve of pollen types present in the diet of each nectarivorous 2087 bat species. 2088 2089
2090 Figure S7. Percentage of resource used by Glossophaginae (G. soricina) and 2091 Lonchophyllinae (L. inexpectata, L. mordax and X. vieirai) species organized by the 2092 plant’s nectar sugar concentration ranging from the highest 30% to the lowest 13% .2093 2094 2095

118
2096

119
2097
C H A P T E R 3 2098
2099
Female reproduction patterns in Caatinga are influenced by precipitation and 2100
Cactaceae resource availability 2101
2102
Eugenia Cordero-Schmidt1, Juan Carlos Vargas-Mena1, Bernal Rodriguez-Herrera3 2103
and Eduardo M. Venticinque1 2104 2105 1Departamento de Ecologia, Universidade Federal do Rio Grande do Norte, 59078900 2106
Lagoa Nova Natal, RN, Brazil; E-mail: [email protected] 2107 2Escuela de Biología, Universidad de Costa Rica, 2060 Montes de Oca, San José, 2108
Costa Rica 2109
2110

120
ABSTRACT 2111 2112 The life history strategies of animals are significantly determined by the frequency and 2113 timing of their reproductive events (Crichton & Krutzsch, 2000), but reproduction is not 2114 an easy task or cheap one and it is influenced by abiotic and biotic factors. Considering 2115 that sympatric species with similar adaptations and food habits provide ideal models 2116 for comparative studies in reproduction, in this study we aim to describe the 2117 reproductive patterns of G. soricina, L. inexpectata, L. mordax and X. vieirai, and 2118 answer three main questions: 1. What is the effect of: A. precipitation and B. 2119 Cactaceae flowers availability on the reproductive patterns of the species? 2. Do they 2120 affect differently the four nectar bat species? 3. Will the diet composition (pollen types 2121 load and other items in fecal samples) be different among nectar-feeding female bats 2122 according to the reproductive status in which they are found? We detected two types 2123 of reproductive patterns for the nectar bat species in Caatinga, bimodal pattern in G. 2124 soricina and apparently L. inexpectata and polymodal in L. mordax and X. vieirai. 2125 Precipitation positively affected the probability of occurrence of pregnancy for L. 2126 mordax and X. vieirai, but not for G. soricina (none of the tested variables influenced 2127 the pregnancy of G. soricina. The availability of floral resources of Cactaceae from the 2128 previous month, positively affected the probability of occurrence of lactation for all 2129 nectar bat species, with some differences of significance for certain cacti species. The 2130 fate of both nectar bats and Cactaceae in Caatinga are closely linked and so are the 2131 other animals that also depended on these keystone resources, including people. 2132 Conservation initiatives should consider areas with high abundance and diversity of 2133 Cactaceae as a priority. 2134 2135
Keywords: Chiroptera; Diet; Phenology; Seasonal Dry Tropical Forest; Resource 2136
partitioning 2137
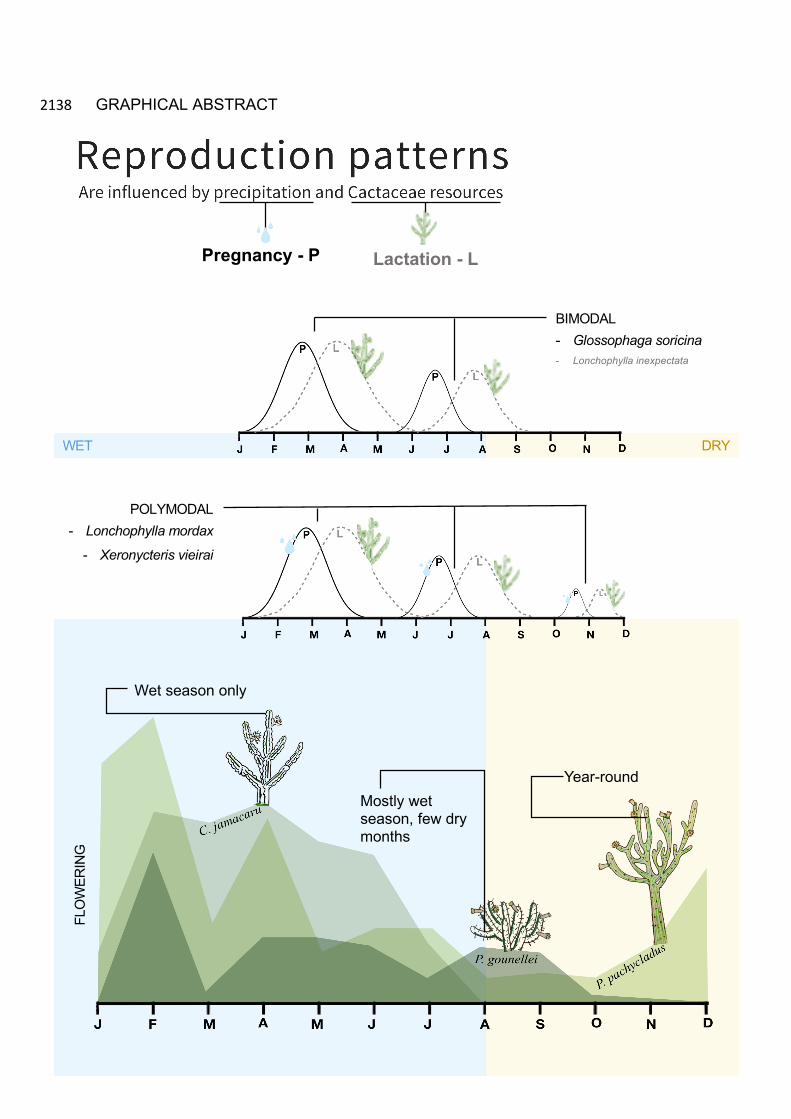
121
GRAPHICAL ABSTRACT 2138
BIMODAL- Glossophaga soricina- Lonchophylla inexpectata
POLYMODAL- Lonchophylla mordax
- Xeronycteris vieirai
Wet season only
Mostly wet season, few dry months
Year-round
FLO
WER
ING
Pregnancy - P Lactation - L
DRYWET

122
INTRODUCTION 2139
Essentially all species have the same purpose in life, to reproduce and pass their genes 2140
from generation to generation, but this is not an easy task, nor a cheap one. Bats life-2141
history is characterized by longevity, multiple reproductive events, delayed sexual 2142
maturity and small litters with one to two well developed (up to 43% of maternal body 2143
mass) offspring (Kurta & Kunz, 1987). 2144
After decades of study on animal reproduction, scientists identified the factors 2145
triggering reproductive periods, and also the ones limiting them. For bats, calcium (for 2146
skeletal growth), nitrogen (protein) and energy (carbohydrates and fat) are three 2147
fundamental factors during pregnancy but mainly during lactation considering it is the 2148
most energetically costly phase of the reproductive cycle (Barclay, 1994; Racey et al., 2149
2000; Voigt, 2003). From a nutritional and physiological perspective, nectar-feeding 2150
bat’s reproduction is noteworthy as their main diet is composed by carbohydrates and 2151
for them to meet the nutritional constrains, they are required to ingest more food and 2152
complement it with protein rich pollen and insects (Herrera et al., 2001; Tschapka, 2153
2005). 2154
Bat reproduction triggering factors vary according to latitude and habitat types 2155
(Racey & Entwistle, 2000). In temperate areas, photoperiod and temperature are 2156
determinant aspects, in the tropics precipitation is fundamental, but in all regions, 2157
parturitions are synchronized with periods of greater resource availability (Fleming, 2158
Hooper & Wilson, 1972; Sperr et al., 2011). This synchrony is expected to be even 2159
more strong in habitats where food resources vary seasonally (Fleming, Hooper & 2160
Wilson, 1972), such as in Caatinga (Machado, Barros & Sampaio, 1997). 2161
Willig (1985) first described the reproduction patterns of bats in Caatinga and 2162
noted that the long-term pattern of rainfall seems to be the dominant factor molding 2163

123
bats reproductive patterns. In other Seasonally Dry Tropical Forests (SDTF) 2164
reproduction of nectar-feeding bats (hereafter ‘nectar bats’) have been intimately 2165
linked to cactus resources (nectar, pollen and fruits) availability (Sosa & Soriano, 1996; 2166
Petit, 1997; Martino, Arends & Aranguren, 1998). Authors highlighted the dependence 2167
on nectar bats reproduction on cactus and vis versa. 2168
Cactaceae is a family endemic to America (Anderson, 2001) and Brazil stands 2169
out as one of the countries with greatest cactus diversity. It has 270 species (70.4% 2170
are endemic) of which 97 species occur in Caatinga (43.2% are endemic) (Taylor and 2171
Zappi, 2002; Ortega-Baes and Godínez-Alvarez, 2006). Cactaceae reproductive 2172
phenology in SDTFs is known to be affected by precipitation both negative or positive, 2173
or even neutrally for different cacti species (Petit, 2001; Cruz & Pavón, 2013), but this 2174
information about Caatingas’ cacti is scarce. 2175
Four basic reproductive patterns have been stablished characterizing the 2176
number of peaks in the proportion of pregnant (hereafter ‘P’) females in a population 2177
of a species or ensemble and the associated lactating (hereafter ‘L’) peaks, if more 2178
than one peak is observed they can be classified as primary and secondary as 2179
suggested by Durant et al. (2013). The patterns are: Amodal (absence of peaks in 2180
pregnancy or lactation); Unimodal (one annual P peak followed by one L peak); 2181
Bimodal (two P peaks associated with two L peaks); and Polymodal (≥ 3 P peaks 2182
accompanied by L peaks). To date, most information on the reproductive patterns of 2183
Neotropical nectar bats have been gathered for SDTFs and provided information for 2184
four genus Glossophaga, Leptonycteris and on less stand for Anoura and 2185
Musonycteris (Fleming et al., 1972; Ramirez-Pulido et al., 1993; Sosa & Soriano, 2186
1996; Petit 1997; Martino et al. 1998; Sperr et al., 2011; but see Zortea, 2003; 2187
Tschapka, 2005 for other habitat type). Most nectar bats present seasonal bimodal 2188

124
polyestrous pattern. Willig (1985) presented pioneering data on G. soricina 2189
reproduction in Caatinga, and only little information has been recently added for the 2190
Vieira’s flower bat (Cordero-Schmidt et al., 2017). 2191
Glossophaga soricina, Lonchophylla inexpectata, Lonchophylla mordax and 2192
Xeronycteris vieiria has been documented to persist and coexist in an area of Caatinga 2193
in the state of Rio Grande do Norte (RN) in northeast Brazil (Cordero-Schmidt et al., 2194
in prep. here Chapter 2). Their diet is composed by at least 31 plant species belonging 2195
to 14 families. Withal, the most used plant family for the four nectar-feeding bats 2196
species was Cactaceae. In this same Caatinga area, at least seven species of cacti 2197
coexist, Cereus jamacaru DC, Pilosocereus chrysostele (Vaupel) Byles & 2198
G.D.Rowley, Pilosocereus gounellei (F.A.C.Weber) Byles & Rowley, Pilosocereus 2199
pachycladus F.Ritter, Melocactus zehntneri (Britton & Rose) Luetzelb and other two 2200
Tacinga Britton & Rose species not used by nectar bats. Cordero-Schmidt et al., (in 2201
prep. here Chapter 1 and 2) have stressed the importance of Cactaceae floral 2202
resources (both pollen and nectar) for the persistence over time of local nectar-feeding 2203
bat ensemble. But is this tight relationship strong enough to influence reproduction 2204
patterns of nectar-feeding female bats in Caatinga? 2205
Considering that sympatric species with similar adaptations and food habits 2206
provide ideal subjects for comparative studies in reproduction, in this study we aim to 2207
describe the reproductive patterns of G. soricina, L. inexpectata, L. mordax and X. 2208
vieirai, and answer three main questions: 1. What is the effect of: A. precipitation and 2209
B. Cactaceae flowering on the female reproductive patterns of the species? 2. Do 2210
they affect differently the four nectar bat species? 3. Will the diet composition (pollen 2211
types load and other items in fecal samples) be different among nectar-feeding female 2212
bats according to the reproductive status in which they are found? 2213

125
In bats energetic investment in offspring is optimized in relation to food 2214
abundance (Racey & Entwistle, 2000), we expect that the reproductive patterns, as in 2215
other SDTFs (Petit, 1997; Martino et al., 1998) will be influenced by both precipitation 2216
and higher resource abundance. Pregnancies will be better explained by precipitation 2217
as a predictor to synchronize their lactation parturition/lactation with the periods of 2218
maximum cacti resource availability to increase their reproductive success. 2219
Nonetheless we expect to find subtle differences between the nectar-feeding bat 2220
species as a way to avoid competition for resources. Additionally, we expect pregnant 2221
and lactating females to have a diet more similar to each other and that will be different 2222
from the diet of non-reproductive females since foraging behavior efficiency can be 2223
impaired by the extra weight of either the final phase of pregnancy or females that 2224
usually fly with their young (Norberg & Rayner, 1987). 2225
METHODS 2226
Study site and climate 2227
This study was conducted in Caatinga, in the municipality of Lajes in the Rio Grande 2228
do Norte (RN) state, located in northeastern Brazil (5°48'46.95"S 36°10'34.60"W). 2229
Vegetation in Caatinga is very diverse for climatic, edaphic, topographic, and anthropic 2230
reasons (Silva, Leal & Tabarelli, 2018). It is characterized by deciduous leaves during 2231
the dry season and frequent or abundant presence of thorny shrubs and Cactaceae. 2232
In a 10-year time series (2009-2019) of the precipitation of the municipality of 2233
Lajes there was a great monthly and year-to-year variation of precipitation (Supporting 2234
Information Figure S1). In Lajes the mean accumulated rainfall was 399.31 mm; 2009 2235
was the rainiest year with a cumulative total of 984.1 mm and 2012 the driest year with 2236
a cumulative total of 99 mm. All precipitation data were extracted from the EMPARN 2237

126
(acronym in Portuguese Rio Grande do Norte Agricultural Research Corporation) 2238
database. 2239
Bat data 2240
We captured bats on 3-day monthly fieldtrips from May to October 2015 for a total of 2241
31 nights (netting effort 43920 m2h) and from February 2017 to February 2019 (except 2242
for November 2017 and May 2018) for a total of 58 mist-netting nights (netting effort 2243
150175 m2h). We mist-netted from 17:30 to midnight and to increase the proportion of 2244
nectar bats caught we set the mist-nets near available flowering plants when possible. 2245
We identified bats to species level using systematic key and species diagnosis 2246
(Gregorin & Ditchfield, 2005; Moratelli & Dias, 2015; Díaz et al. 2016). We determined 2247
age following Brumet-Rossinni & Wilkinson (2009), looking into the epiphyseal-2248
diaphyseal fusion, pelage coloration and tooth wear. 2249
Reproduction data 2250
We externally inspected females to identify the reproductive status. They were 2251
categorized as: non-reproductive – NR: with flat abdomen, nonapparent nipples (post-2252
lactating females with no milk produced and new fur growth surrounding the nipples 2253
were included in the NR category); pregnant – P: pregnancy was detected by lower 2254
abdominal palpation and general body shape); lactating- L: enlarged nipples with no 2255
hair surrounding them, with milk secretion verified by light pressure on the nipples. 2256
All procedures for capture, handling and collection of bats met the guidelines of 2257
the American Society of Mammalogists for the use of wild mammals in research 2258
(Sikes, 2016), and the legal Brazilian requirements of conservation and animal 2259
welfare. Fieldwork was authorized by MMA/ICMBio/SISBIO under permits 48325-2. 2260
Prior to the analysis the bat capture data was corrected by effort as follows: 2261
! !"#%&'()*"+,-./-*0,**/012%"+-∗#!" ∗ 1000 2262

127
Diet 2263
Dietary information on the nectar bat species were obtained from pollen samples 2264
collected from the fur, wings, legs and uropatagium using a single glycerin jelly cube 2265
(3-4 mm) per individual (Voigt, Kelm, Bradley & Ortmann, 2009), which were later 2266
mounted on glass microscope slides. Pollen types were identify to the lowest possible 2267
taxonomic level using a light microscope (magnification 40-100x, Leica DM500) by 2268
comparing our samples with a reference pollen collection from the study sites and with 2269
other pollen catalogues (Palacios-Chávez, Ludlow-Wiechers & Villanueva, 1991; 2270
Roubik & Moreno 1991; Santos, Watanabe & Hamburgo-Alves, 1997; Carreira & Barth 2271
2003; Melhem et al., 2003; Silva, Santos & Lima, 2016). Additionally, we collected 2272
fecal samples and characterized their contents using a microscope (magnification 40-2273
100x, Leica DM500). We categorized the contents of each fecal sample by the 2274
presence or absence of pollen, insect fragments, and “plant tissue” which are 2275
unidentified elements that appear to be of plant origin such as leaves, pericarp, hairs 2276
of the areolas of the cactus, trichomes, etc. 2277
Cactaceae flower resources 2278
We established 15 transects of 100 * 5 m following pre-existing trails, where we tagged 2279
110 randomly chosen reproductive individuals of three species of Cactaceae (nine 2280
Cereus jamacaru - Mandacaru, 40 Pilosocereus gounellei - Xique-xique and 61 2281
Pilosocereus pachycladus - Facheiro). We recorded the individuals with flowers (both 2282
buds and open flowers) available at the time we were mist-netting for bats and the 2283
abundance of each species in the transects. Availability of resource per hectare was 2284
calculated as follows: 2285
Resourceavailabilitypermonth = 5Floweringindv.Observedindv. < ∗ 5Sppabundance
Aream! ∗ 10.000ha. < 2286
2287

128
Studied Cacti species 2288
-Cereus jamacaru DC, commonly known as Mandacaru (Supporting Information 2289
Figure S2). Is a widely distributed Caatinga endemic species, present both in natural 2290
and anthropic areas. It is a columnar cactus with many erect branches forming dense 2291
crowns, individuals can have a height of up to 10 m with distinct trunks to 60 cm in 2292
diameter (Anderson, 2001). Flowers are tubular and very large with 20-30 cm long and 2293
18-20 cm in diameter, solitary, nocturnal and white colored. Individuals can produce 1 2294
to 87 flowers, the opening of all the buds is given over 3 days, once a month during 2295
the rainy period (Zanina, 2013), the blooming of this species is considered massive. 2296
Its pollination syndromes involve sphingophily and chiropterophily (Quirino, 2006). 2297
- Pilosocereus gounellei (F.A.C.Weber) Byles & Rowley, commonly known as Xique-2298
xique (Supporting Information Figure S3). It is a widely distributed Caatinga endemic 2299
species, mostly common on sandy soil and rocky outcrops (Rocha and Agra, 2002). It 2300
is a shrubby cactus, rarely treelike, 0.5-4 m high with numerous erect, inclined and 2301
even horizontal stems (Anderson, 2001). Flowers are tubular and have a narrower and 2302
longer flower-tube the other Pilosocereus species with 7.18 cm long and 2.31cm in 2303
diameter. Individuals have seven to 17 open flowers per night and their flowers last 2304
one day (Rocha et al., 2019). Its pollination syndromes involve sphingophily (Rocha 2305
et al., 2019) and probably chiropterophily (Zappi, 1994; Cordero-Schmidt et al., 2017). 2306
- Pilosocereus pachycladus F.Ritter, commonly known as Facheiro (Supporting 2307
Information Figure S4). It is widely distributed Caatinga endemic species, occurs more 2308
frequently in the top of the mountains (Lucena et al., 2015a). It is a columnar cactus 2309
with many erect branches, individuals can have a height of 2 to 10 m or more 2310
(Anderson, 2001). The flower’s shape is bell/funnel and have a size of approximately 2311
6.74 cm and a diameter of 2.7 cm. Individuals have one to nine open flowers per night 2312

129
and their flowers last one day (Rocha et al., 2019). Its pollination syndromes involve 2313
chiropterophily (Quirino, 2006). 2314
Analysis 2315
We applied simple and multiple logistic regression models to estimate the probability 2316
of occurrence of pregnant and lactating females of G. soricina, L. mordax and X. vieirai 2317
(L. inexpectata was excluded from the analysis due to lack of data on captured 2318
reproductive females). Candidate models included the following variables: 2319
precipitation (Precip.), one month anticipated precipitation (T-1), and the availability of 2320
floral resources of three Cactaceae species P. pacycladus (Pp), Pilosocereus 2321
gounellei (Pg), and Cereus jamacaru (Cj), and one month anticipated the availability 2322
of floral resources of this species (t-1). Model selection was performed using Akaike 2323
Information Criterion (AIC) (Williams et al., 2002). The normalized likelihood that a 2324
model fits the data better (Akaike weights) was used to choose which model was most 2325
likely to be the best to explain the observed data. The lower the value of AIC scores 2326
(∆AIC < 2), the more plausible the model adjustment is to the data. Analysis were 2327
performed in the software SYSTAT version 12. 2328
After selecting the best models, we used the estimative of parameter for 2329
selected logistic models to explain the effect in both pregnancy and lactation of the 2330
studied nectar bat species. 2331
RESULTS 2332
Precipitation 2333
Among the three sampled years, 2015 was the driest with a cumulative total of 277.6 2334
mm, followed by 2017 with 313.2 mm and 2018 with 351.6 mm (Figure 1). The rainiest 2335
months were February (x̄=75.03 mm), March (x̄= 62.06 mm) and April (x̄= 84.90 mm). 2336
The driest months were August, September and November with zero mm. 2337

130
Cactaceae resource availability 2338
The months of greatest flower availability of the three species of cactus coincides with 2339
the months with the highest rainfall (Figure 1). The primary flowering peak of the three 2340
species is overlapped and occurs in February. Pilosocereus pachycladus and C. 2341
jamacaru presented a secondary peak in April, contrary to P. gounellei that kept the 2342
number of individuals blooming similarly from April to September. Cereus jamacaru is 2343
the most seasonal species blooming only in the rainiest period (January-July). 2344
Pilosocereus gounellei bloom during the rainy season and extends its flowering during 2345
few dry months (August-September) and P. pachycladus is the only species that 2346
remains blooming throughout the year. 2347
Reproductive patterns and diet 2348
Glossophaga soricina 2349
We captured a total of 14 pregnant females, nine lactating and 76 non-2350
reproductive females (Table 1). 2351
Reproductive pattern 2352
Glossophaga soricina presented a seasonal bimodal pattern (Figure 2A). The primary 2353
pregnancy peak is observed in May (late rainy period) and the second peak in 2354
December (end of the dry period). Lactation peaks occurred approximately two 2355
months after each pregnancy peak, the primary peak occurred in July. 2356
None of the tested variables (precipitation, nor Cactaceae resource availability with 2357
their respective month anticipation) had an effect on pregnancies of G. soricina 2358
(Supplementary information Table S5). Cactaceae flower resource availability of the 2359
previous month (t-1) had a positive effect on lactating females of G. soricina (Pp(t-1) 2360
Estimate=0.004; Pg(t-1) Estimate=0.002; Cj(t-1) Estimate=0.12; ΔAIC= 0.66; 2361

131
wAIC=13.06; Table 2). Cereus jamacaru had a marginally more significant effect than 2362
the other cacti species (Table S5). 2363
Diet 2364
The females of G. soricina fed on 18 plant species including five species of cactus 2365
(Supporting Information Table S8). This species used P. pachycladus in a similar 2366
proportion in its different reproductive status. During lactation (L), 80% of the samples 2367
included the five species of cacti, with M. zehntneri and P. pachycladus being the most 2368
used resource and they supplemented their diet with two species, 30% of the samples 2369
contained Harpochilus paraibanus (Acanthaceae) and 10% Encholirium spectabile 2370
(Bromeliaceae). 2371
We can observe that either pregnant, lactating or non-reproductive females 2372
used the same complementary resources (pollen, insects and plant tissues) and the 2373
use of these food items does not seem to be determined by precipitation (Figure 4). 2374
However, due to the low number of fecal samples of each reproductive status it is 2375
difficult to draw any conclusions regarding the use of complementary food items. 2376
Lonchophylla inexpectata 2377
We captured a total of five pregnant females, one lactating and 51 non-2378
reproductive (Table 1). 2379
Reproductive pattern 2380
The first peak of pregnancy was observed in February (Figure 2B). The few pregnant 2381
captured females might suggest a seasonal bimodal pattern, nonetheless, all 2382
observed peaks are based on small samples and thus not robust enough to conclude 2383
a pattern. 2384
Diet 2385

132
More than 60% of their samples were composed by Cactaceae pollen and 2386
complemented their diet with 12 other plant species (Figure 3; Supporting Information 2387
Table S8). Lonchophylla inexpectata also complemented their diet with pollen, insects 2388
and plant tissues and it seems that the species included both pollen and insects in the 2389
drier periods in June and August (Figure 4). Due to the low capture of reproductively 2390
active females we cannot have proper conclusions about the diet of this species in the 2391
different reproductive status. 2392
Lonchophylla mordax 2393
We captured a total of 17 pregnant females, 19 lactating and 51 non-2394
reproductive females (Table 1). 2395
Reproductive pattern 2396
Lonchophylla mordax presented a polymodal pattern (Figure 2C) with one primary 2397
pregnancy peak in April (second precipitation peak), a secondary peak in December-2398
January (transition between dry and rainy period) and occasional pregnancies in June 2399
and August (beginning of the dry period). 2400
Precipitation had a strong significant effect in pregnancy of L. mordax (Precip. 2401
Estimate=0.02; ΔAIC= 0; wAIC=36.12) (Table 2). Cactaceae flower resource 2402
availability of the previous month (of the three species) had a positive significant effect 2403
on lactation in L. mordax but was most significantly influenced by P. pachycladus 2404
previous month flowering availability (Pp(t-1) Estimate=0.004; ΔAIC= 0; wAIC=42.64) 2405
(Table 2). 2406
Diet 2407
Females of L. mordax fed on 20 plant species including five species of cactus 2408
(Supporting Information Table S8). Lonchophylla mordax is the nectar bat that used 2409
higher proportion of non-cacti species, still, Cactaceae predominates in the diet mainly 2410

133
of pregnant females (80%) and lactation (60%). During lactation L. mordax used a 2411
similar proportion of C. jamacaru, P. gounellei and P. pachycladus. 2412
Fecal samples of pregnant females presented feces with exclusively pollen in 2413
August and April (Figure 4). Females diet included a higher proportion of insects in the 2414
rainiest periods and used higher proportion of pollen and other plant tissues in the 2415
driest period. 2416
Xeronycteris vieirai 2417
We captured a total of seven pregnant females, eleven lactating and 48 non-2418
reproductive females (Table 1). 2419
Reproductive pattern 2420
Xeronycteris vieirai seems to have a polymodal pattern based on the three lactation 2421
peaks in March, May (primary peak) and July. Pregnancy peaks are not as discernible 2422
as lactation peaks (Figure 2D). 2423
Precipitation had a significant positive effect on pregnancy in X. vieirai (Precip. 2424
Estimate=0.01; ΔAIC= 0; wAIC=20.19) (Table 2). Previous month flower availability of 2425
C. jamacaru had a significant positive effect on lactating females (Cj(t-1) Estimate=0.12; 2426
ΔAIC= 0; wAIC=24.87) (Table 2) the second most significant model was flower 2427
availability of P. gounellei of the previous month (Pg(t-1) Estimate=0.01; ΔAIC= 0; 2428
wAIC=12.97;) (Table 2). 2429
Diet 2430
Females of X. vieirai fed on 14 plant species including five species of cactus 2431
(Supporting Information Table S8). Xeronycteris vieirai was the nectar bat the used a 2432
higher proportion of cactus (≥80%) in their diet in all reproductive status, the higher 2433
proportion was used by pregnant females (>90%) where they only complemented their 2434
diet with Bauhinia pentandra present in 14% of the samples. During lactation the most 2435

134
used cactus species was C. jamacaru (30%), followed by approximately 20% of P. 2436
pachycladus and P. gounellei, and used only two species of non-cacti plants, 2437
Encholirium spectabile (18%) and B. pentandra (9%). Pregnant and non-reproductive 2438
females used a similar proportion of P. pachycladus and P. gounellei (around 30%), 2439
M. zehntneri (10%) and P. chrysostele (10%). 2440
Fecal samples from the driest months (January, June and August to December) 2441
included almost exclusively pollen, independently of their reproductive status (Figure 2442
4). In February (rainiest month) pregnant, lactating and non-reproductive females 2443
complemented their diet with insects and pollen. 2444
The diet of the females of the four studied nectar bats, we can see that regardless of 2445
their reproductive status, more than 50% of female’s diets are composed of 2446
Cactaceaea (Figure 3). 2447
DISSCUSSION 2448
The life history strategies of animals are significantly determined by the 2449
frequency and timing of their reproductive events (Crichton & Krutzsch, 2000). We 2450
detected two types of reproductive patterns for the nectar bat species in Caatinga, 2451
bimodal pattern in G. soricina and polymodal in L. mordax and X. vieirai. Precipitation 2452
positively affected the probability of occurrence of pregnancy for L. mordax and X. 2453
vieirai. None of the tested variables (precipitation nor Cactaceae resources 2454
availability) influenced the pregnancy of G. soricina. On the other hand, the availability 2455
of floral resources of Cactaceae from the previous month, positively affected the 2456
probability of occurrence of lactation for all nectar bat species, with some differences 2457
of significance for certain cacti species. 2458
The phenological flowering patterns of sympatric cactus species have been 2459
studied in various SDTFs where both synchronous and asynchronous, inter and intra-2460

135
specific blooms have been observed (Soriano et al., 1991; Silvius, 1995; Locatelli & 2461
Machado, 1996b; Sosa & Soriano 1996; Petit, 1997; Petit, 2001; Ruiz et al., 2000; 2462
McIntosh, 2002). Authors have mentioned that this phenological variation throughout 2463
the year favors the maintenance of pollinators and dispersers, mainly in environments 2464
with high seasonality. Also, the periods of flowering overlap can favor pollination, since 2465
an increase in resources could also be reflected pollinators abundance (Kudo, 2006). 2466
The studied cacti species seem to present both strategies since their flotation peaks 2467
overlap in the rainy season furthermore each species showed a different flowering 2468
frequency throughout the year (C. jamacaru-rainy season only; P. gounellei - mainly 2469
rainy season and extends during the first months of the dry season; P. pachycladus 2470
year-round availability). This phenological variability is influencing not only the 2471
persistence of the species in the area over time (Cordero-Schmidt in prep. Chapter 2), 2472
but also influencing the occurrence of lactation events. 2473
Pregnancy and rainfall 2474
Caatinga is an environment dominated by seasonal changes, where resources 2475
vary according to precipitation fluctuations (Lima & Rodal, 2010) thus creating more 2476
suitable periods to reproduce. The long period of gestation of bats limits their ability 2477
to react quickly to short-term fluctuations but respond to predictable seasonal changes 2478
by which to optimize their time of reproduction (Heideman, 2000). Bimodal and 2479
polymodal polyestry patterns have shown a clear relationship in enviroments 2480
presenting unimodal rainfall (Heideman, 2000) as observed in our studied Caatinga 2481
area. Lonchophylla mordax and X. vieirai presented most of their pregnancy events 2482
within the propitious rainy season and lower pregnancy events and non-breeding 2483
during suboptimal dry seasons. Differently, none of the variables used in our models 2484
were able to explain the patterns in pregnancy of G. soricina, maybe this is due to its 2485

136
generalist’s behavior (Estrada & Coates-Estrada, 2001). Glossophaga soricina is the 2486
nectar bat species with more information gathered about its reproductive patterns in 2487
different habitats. This species has a bimodal pattern independent of habitat type 2488
(Fleming 1972; Willig, 1985; Ramirez-Pulido et al., 1993; Stoner 2001; Zortea, 2003; 2489
Tschapka, 2005), but the time when the pregnancy and lactation peaks occurs can 2490
vary geographically (Durant et al., 2013), our data corroborates this. In the Caatinga 2491
of RN state G. soricina shows a bimodal pattern, with a peak pregnancy peaks in May 2492
(rainy season) and December (dry season) followed by two lactation peaks in February 2493
(rainy season) and July (transition between rainy to dry season). In other SDTFs G. 2494
soricina’s reproductive patterns have been explained according to the availability of 2495
resources and their great flexibility in the diet (Sperr et al., 2011). 2496
Lactation and Cactaceae resource availability 2497
As mentioned before, lactation is the most energetically costly reproductive status, 2498
hence seems logic that the most important factor in determining the timing of this 2499
event’s occurrence is resource availability (Racey & Entwistle, 2000). Lactation 2500
overlap occurred year-round; but the primary peaks of L. mordax and X. vieirai do not 2501
overlap and are separated by approximately one month (Figure S9). Such a difference 2502
in the timing of reproduction emphasizes the importance of diet in determining 2503
reproductive timing, and reinforce that food availability, rather do climatic factors, 2504
ultimately underlies the timing of reproductive events in tropical bat species (Racey & 2505
Entwistle, 2000). July is the only month where the lactation of the three studied nectar 2506
bat species seems to overlap the most. July is also the last month where there is 2507
flowering overlap of the three cacti species and the month where a last peak in rain is 2508
observed (Figure 1). The fact that there is no overlap of the main lactation peaks of 2509
the three species seems to indicate a mechanism to avoid competition (Ramirez-2510

137
Pulido et al., 1993). Considering that all cacti species were equally available for all bat 2511
species, another possible evidence of competition avoidance is the distinct influence 2512
of the different cacti species on the different nectar bat species. Glossophaga soricina 2513
lactation events were better explained by C. jamacaru, L. mordax by P. pachycladus 2514
and X. vieirai by C. jamacaru and P. gounellei. It seems that the nectar bat species 2515
are partitioning resources during the delicate phase of lactation. 2516
Bats generally give birth to well-developed large infants, about 26.6% of the 2517
mother’s body mass in phyllostomids (Kurta and Kunz, 1987). Females of G. soricina 2518
and X. vieirai were caught several times carrying their young, this behavior was never 2519
observed in the Lonchophylla species; this behavior probably affects the species 2520
foraging behavior and dislocation capacity and thus the diet composition (Kurta and 2521
Kunz, 1987). Lonchophylla mordax included more species of plants during lactation 2522
(14 plant species) than G. soricina and X. vieirai, this might be explained by greater 2523
access to different flowers because they are not carrying their young while foraging. 2524
But at the same time L. mordax lactating females may be obligated to forage where 2525
most resources are concentrated or forage near the maternity roosts (Sperr et al., 2526
2011). However, it is necessary to study this in more detail and in other areas to see 2527
if it is a repeating pattern for these species. 2528
Fecal contents of X. vieirai females seem to show some sort of seasonality, 2529
where regardless of their reproductive status the females include more insects in their 2530
diet during the rainy season and include more pollen in the dry season. In Caatinga, 2531
the rainiest months of the year are the months with the greatest abundance of insects 2532
(Vasconcellos et al., 2010) and this could explain the increase in the frequency of this 2533
item in the diet of X. vieirai. Cordero-Schmidt (in prep. Chapter 2) found that X. vieirai 2534
is the most specialized nectar/pollen-feeding bat in this Caatinga area, especially 2535

138
when compared to G. soricina which is the most generalist species, capable to actively 2536
look for insect preys (Clare et al., 2014). In general we can affirm that the females of 2537
the species of nectar bats complement their diet mainly with pollen, which is an 2538
important source of tyrosine, an amino acid found in abundance in the milk of small 2539
mammals and acts as growth stimulant (Howell, 1974), followed by insects and plant 2540
tissues, thus obtaining the inputs of protein, nitrogen among others, necessary for the 2541
different reproductive states. But the low number of fecal samples does not allow us 2542
to have any robust conclusions about the way the species are supplementing their diet 2543
according to their reproductive status. 2544
Conservation concerns 2545
Both abiotic (precipitation) and biotic resources availability (Cactaceae) influence the 2546
reproduction events in nectar-feeding bats in Caatinga. Sadly both factors are 2547
threatened by climate change. Predictive models about the potential impact of climate 2548
change in the Caatinga indicate that precipitation will decrease 22% by 2100 and that 2549
drier sites show impoverished plant assemblages that are more sensitive to chronic 2550
disturbances such as wood extraction, cattle ranching and subsistence agriculture 2551
(Rito et al., 2017). In other SDTFs the range reductions of particular species, lower 2552
abundance, extirpation, extinction, and increased competition in environments altered 2553
by humans are having severe negative consequences on the structure and 2554
composition of mammal communities (Dirzo et al., 2011). Moreover, changes in 2555
flowering phenology caused by habitat fragmentation have shown to change bat 2556
pollination patterns and to negatively affect the reproductive outcome of the plants 2557
they pollinate (Quesada et al. 2004). 2558
Cactaceae have intrinsic characteristics that make them vulnerable to habitat 2559
fragmentation, loss of habitat quality, mining, trade and illegal collection (Zappi et al., 2560

139
2011). Their life cycle is slow and only small proportion of the annual seed production 2561
will germinate and survive (Ortega-Baes et al., 2010). Cacti resources are very 2562
important not only for wildlife but for humans as well. Cactaceae species are widely 2563
used by human populations in Caatinga, its uses vary and range from construction (for 2564
making doors, windows, boards,laths, etc), food for cattle, goats and sheep, religious, 2565
medicinal, ornamental and veterinary use, among others (Lucena et al., 2015b). 2566
The fate of both nectar bats and Cactaceae in Caatinga are closely linked and 2567
so are the other animals that also depended on this keystone resources including 2568
people. The loss of cacti species could certainly affect the supply of important 2569
environmental services such as pollination and seed dispersal by animals (Zappi, et 2570
al., 2011). Conservation initiatives should consider areas with high abundance and 2571
diversity of Cactaceae as a priority. 2572
2573
REFERENCES 2574
Alves, J. J. A., de Araújo, M. A., & do Nascimento, S. S. (2009). Degradação da 2575
Caatinga: uma investigação ecogeográfica. Revista Caatinga, 22(3), 126-135. 2576
Barclay, R. M. (1994). Constraints on reproduction by flying vertebrates: energy and 2577
calcium. The American Naturalist, 144(6), 1021-1031. 2578
Brunet-Rossinni A. K. & G. S. Wilkinson. (2009). Methods for age estimation and the 2579
study of senescence in bats. In: Ecological and behavioral methods for the study 2580
of bats, 315-325. 2581
Cordero-Schmidt, E., Barbier, E., Vargas-Mena, J. C., Oliveira, P. P., Santos, F. D. A. 2582
R., Medellín, R. A., ... & Venticinque, E. M. (2017). Natural history of the Caatinga 2583
endemic Vieira's flower bat, Xeronycteris vieirai. Acta chiropterologica, 19(2), 2584
399-408. 2585

140
Clare, E. L., Goerlitz, H. R., Drapeau, V. A., Holderied, M. W., Adams, A. M., Nagel, 2586
J., ... & Brock Fenton, M. (2014). Trophic niche flexibility in G lossophaga 2587
soricina: how a nectar seeker sneaks an insect snack. Functional Ecology, 28(3), 2588
632-641. 2589
Crichton, E. G., & Krutzsch, P. H. (Eds.). (2000). Reproductive biology of bats. 2590
Academic Press. 2591
Dirzo, R., Young, H. S., Mooney, H. A., & Ceballos, G. (2011). Seasonally dry tropical 2592
forests: ecology and conservation. Island Press. 2593
Durant, K. A., Hall, R. W., Cisneros, L. M., Hyland, R. M., & Willig, M. R. (2013). 2594
Reproductive phenologies of phyllostomid bats in Costa Rica. Journal of 2595
Mammalogy, 94(6), 1438-1448. 2596
EMPARN- Empresa de Pesquisa Agropecuária do Rio Grande do Norte. Rainfall 2597
monitoring: accumulated monthly rainfall per year by municipality. Retrieved 2598
from:http://meteorologia.emparn.rn.gov.br:8181/monitoramento/monitoramento.2599
php 2600
Estrada, A., & Coates-Estrada, R. (2001). Species composition and reproductive 2601
phenology of bats in a tropical landscape at Los Tuxtlas, Mexico. Journal of 2602
Tropical Ecology, 17(5), 627-646. 2603
Feming, T. H .• E. T. Hooper. and D. E. Wilson. 1972. Three central American bat 2604
communities: structure. reproductive cycles and movement pattems. 2605
Heideman, P. D. (2000). Environmental regulation of reproduction. In Reproductive 2606
biology of bats (pp. 469-499). Academic Press. 2607
Herrera M, L. G., Hobson, K. A., Leticia, M. M., Nicte, R. P., Germán, M. C., & VÍctor, 2608
S. C. (2001). Sources of protein in two species of phytophagous bats in a 2609

141
seasonal dry forest: evidence from stable-isotope analysis. Journal of 2610
Mammalogy, 82(2), 352-361. 2611
Howell, D. J. (1974). Bats and pollen: physiological aspects of the syndrome of 2612
chiropterophily. Comparative Biochemistry and Physiology Part A: 2613
Physiology, 48(2), 263-276. 2614
Kudo, G. (2006). Flowering phenologies of animal-pollinated plants: reproductive 2615
strategies and agents of selection. Ecology and evolution of flowers, 139-158. 2616
Lima, A. L. A., & Rodal, M. J. N. (2010). Phenology and wood density of plants growing 2617
in the semi-arid region of northeastern Brazil. Journal of Arid 2618
Environments, 74(11), 1363-1373. 2619
Locatelli, E., & Machado, I. C. S. (1999a). Floral biology of Cereus fernambucensis: a 2620
sphingophilous cactus of restinga. Bradleya, 1999(17), 86-95. 2621
Locatelli, E., & Machado, I. C. S. (1999b). Comparative study of the floral biology in 2622
two ornithophilous species of Cactaceae: Melocactus zehntneri and Opuntia 2623
palmadora. Bradleya, 1999(17), 75-85. 2624
Lucena, E. A. R. M. D. (2007). Fenologia, biologia da polinização e da reprodução de 2625
Pilosocereus Byles & Rowley (Cactaceae) no nordeste do Brasil. 2626
Lucena, C. M., Ribeiro, J. E., Nunes, E. N., Meiado, M. V., Quirino, Z. G., Casas, A. 2627
L. E. J. A. N. D. R. O., & Lucena, R. F. (2015a). Distribuição local de Cereus 2628
jamacaru Dc. subsp. jamacaru e Pilosocereus pachycladus F. Ritter subsp. 2629
pernambucoensis, Gaia scientia (2015). Edição Especial Cactaceae. 9(2): 97-2630
103. 2631
Lucena, C. M., Carvalho, T. K., Ribeiro, J. E., Quirino, Z. G., Casas, A., & Lucena, R. 2632
F. P. (2015b). Conhecimento botânico tradicional sobre cactáceas no semiárido 2633
do Brasil. Gaia scientia, 9(2), 77-99 2634

142
Machado, I. C., Barros, L. M., & Sampaio, E. V. (1997). Phenology of caatinga species 2635
at Serra Talhada, PE, northeastern Brazil. Biotropica, 29(1), 57-68. 2636
Martino, A., Arends, A., & Aranguren, J. (1998). Reproductive pattern of Leptonycteris 2637
curasoae Miller (Chiroptera: Phyllostomidae) in northern 2638
Venezuala. Mammalia, 62(1), 69-76. 2639
McIntosh, M. E. (2002). Flowering phenology and reproductive output in two sister 2640
species of Ferocactus (Cactaceae). Plant Ecology, 159(1), 1-13. 2641
Norberg, U. M., & Rayner, J. M. (1987). Ecological morphology and flight in bats 2642
(Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy 2643
and echolocation. Philosophical Transactions of the Royal Society of London. B, 2644
Biological Sciences, 316(1179), 335-427. 2645
Ortega-Baes, P., & Godínez-Alvarez, H. (2006). Global diversity and conservation 2646
priorities in the Cactaceae. Biodiversity & Conservation, 15(3), 817-827. 2647
Ortega-Baes, P., Sühring, S., Sajama, J., Sotola, E., Alonso-Pedano, M., Bravo, S., & 2648
Godínez-Alvarez, H. (2010). Diversity and conservation in the cactus family. 2649
In Desert plants (pp. 157-173). Springer, Berlin, Heidelberg. 2650
Quesada, M., Stoner, K. E., Lobo, J. A., Herrerias-Diego, Y., Palacios-Guevara, C., 2651
Munguía-Rosas, M. A., ... & Rosas-Guerrero, V. (2004). Effects of Forest 2652
Fragmentation on Pollinator Activity and Consequences for Plant Reproductive 2653
Success and Mating Patterns in Bat-pollinated Bombacaceous Trees 1. 2654
Biotropica, 36(2), 131-138. 2655
Petit, S. (1997). The diet and reproductive schedules of Leptonycteris curasoae 2656
curasoae and Glossophaga longirostris elongata (Chiroptera: Glossophaginae) 2657
on Curaçao. Biotropica, 29(2), 214-223. 2658

143
Petit, S. (2001). The reproductive phenology of three sympatric species of columnar 2659
cacti on Curaçao. Journal of Arid Environments, 49(3), 521-531. 2660
Quirino, Z. (2006). Fenologia, síndromes de polinização e dispersão e recursos florais 2661
de uma comunidade de Caatinga no cariri paraibano. 2662
Racey, P. A., & Entwistle, A. C. (2000). Life-history and reproductive strategies of bats. 2663
In Reproductive biology of bats (pp. 363-414). Academic Press. 2664
Ramírez-Pulido, J., Armella, M. A., & Castro-Campillo, A. (1993). Reproductive 2665
patterns of three neotropical bats (Chiroptera: Phyllostomidae) in Guerrero, 2666
Mexico. The Southwestern Naturalist, 24-29. 2667
Rito, K. F., Arroyo-Rodríguez, V., Queiroz, R. T., Leal, I. R., & Tabarelli, M. (2017). 2668
Precipitation mediates the effect of human disturbance on the Brazilian Caatinga 2669
vegetation. Journal of Ecology, 105(3), 828-838. 2670
Ruiz, A., Santos, M., Cavelier, J., & Soriano, P. J. (2000). Phenological study of cacti 2671
in the arid region of Tatacoa, Colombia. Biotropica, 32(3), 397-407. 2672
Silva, J. M. C., Leal, I. R., & Tabarelli, M. (Eds.). (2018). Caatinga: the largest tropical 2673
dry forest region in South America. Springer. 2674
Silvius, K. M. (1995). Avian consumers of cardon fruits (Stenocereus griseus: 2675
Cactaceae) on Margarita Island, Venezuela. Biotropica, 96-105. 2676
Soriano, P. J., Sosa, M., & Rossell, O. (1991). Hábitos alimentarios de Glossophaga 2677
longirostrisMiller (Chiroptera: Phyllostomidae) en una zona árida de los Andes 2678
venezolanos. Revista de Biología Tropical, 263-268. 2679
Sosa, M., & Soriano, P. J. (1996). Resource availability, diet and reproduction in 2680
Glossophaga longirostris (Mammalia: Chiroptera) in an arid zone of the 2681
Venezuelan Andes. Journal of Tropical Ecology, 12(6), 805-818. 2682

144
Sperr, E. B., Caballero-Martinez, L. A., Medellin, R. A. and Tschapka, M. (2011). 2683
Seasonal changes in species composition, resource use and reproductive 2684
patterns within a guild of nectar-feeding bats in a west Mexican dry forest. J. 2685
Trop. Ecol. 27, 133-145. 2686
Stoner, K. E. (2001). Differential habitat use and reproductive patterns of frugivorous 2687
bats in tropical dry forest of northwestern Costa Rica. Canadian Journal of 2688
Zoology, 79(9), 1626-1633. 2689
Taylor, N. P., & Zappi, D. (2002). Distribuição das espécies de Cactaceae na 2690
caatinga. Vegetação & Flora da Caatinga. Recife: Associação de Plantas do 2691
Nordeste–APNE, 123-125. 2692
Tschapka, M. (2005). Reproduction of the Bat Glossophaga commissarisi 2693
(Phyllostomidae: Glossophaginae) in the Costa Rican Rain Forest During 2694
Frugivorous and Nectarivorous Periods 1. Biotropica: The Journal of Biology and 2695
Conservation, 37(3), 409-415. 2696
Vasconcellos, A., Andreazze, R., Almeida, A. M., Araujo, H. F., Oliveira, E. S., & 2697
Oliveira, U. (2010). Seasonality of insects in the semi-arid Caatinga of 2698
northeastern Brazil. Revista Brasileira de Entomologia, 54(3), 471-476. 2699
Voigt, C. C. (2003). Reproductive energetics of the nectar-feeding bat Glossophaga 2700
soricina (Phyllostomidae). Journal of Comparative Physiology B, 173(1), 79-85. 2701
Willig, M. R. (1985). Reproductive patterns of bats from caatingas and cerrado biomes 2702
in northeast Brazil. Journal of Mammalogy, 66(4), 668-681. 2703
Zanina, D. N. (2013). Quem poliniza Cereus jamacaru? (Master's thesis, Universidade 2704
Federal do Ceará, Fortaleza, Brazil). Retrieved from 2705
http://repositorio.ufc.br/handle/riufc/17133 2706

145
Zappi, D., Taylor, N., Ribeiro-Silva, S., Machado, M., Moraes, E. D., Calvente, A., ... 2707
& Aona, L. (2011). Plano de ação nacional para conservação das cactáceas. 2708
Instituto Chico Mendes de Conservação da Biodiversidade, Brazil. 2709
Zortéa, M. (2003). Reproductive patterns and feeding habits of three nectarivorous 2710
bats (Phyllostomidae: Glossophaginae) from the Brazilian Cerrado. Brazilian 2711
Journal of Biology, 63(1), 159-168. 2712
2713
2714 2715

146
2716
2717
2718 Figure 1. Monthly flowering individuals per hectare of the sampled three Cactaceae 2719 species and monthly precipitation. 2720 2721

147
Table 1. Monthly sampling effort, monthly and average precipitation during the sampling periods and the captured individuals 2722 categorized in five reproductive status: NRF- Non-Reproductive Females; P- Pregnant; L-Lactating. Cells are filled with the number 2723 of captured individuals, · accounts for zero. 2724 2725
2726

148
2727 Figure 2. Reproductive pattern of the four nectar bat species synthesized from 2 2728 sampling periods: May-October 2015 and February 2017 to February 2019 (except for 2729 November 2017 and May 2018). The patterns are based on monthly proportions of: 2730 Pregnant, Lactating and Non-Reproductive females; all corrected by monthly sampling 2731 effort. I. Female reproduction patterns in relation to mean monthly precipitation. II. 2732 Females reproductive pattern in relation to three Cactaceae species flowering 2733 phenologies (P.p: Pilosocereus pachycladus; C.j: Cereus jamacaru; P.g: Pilosocereus 2734 gounellei). 2735
2736
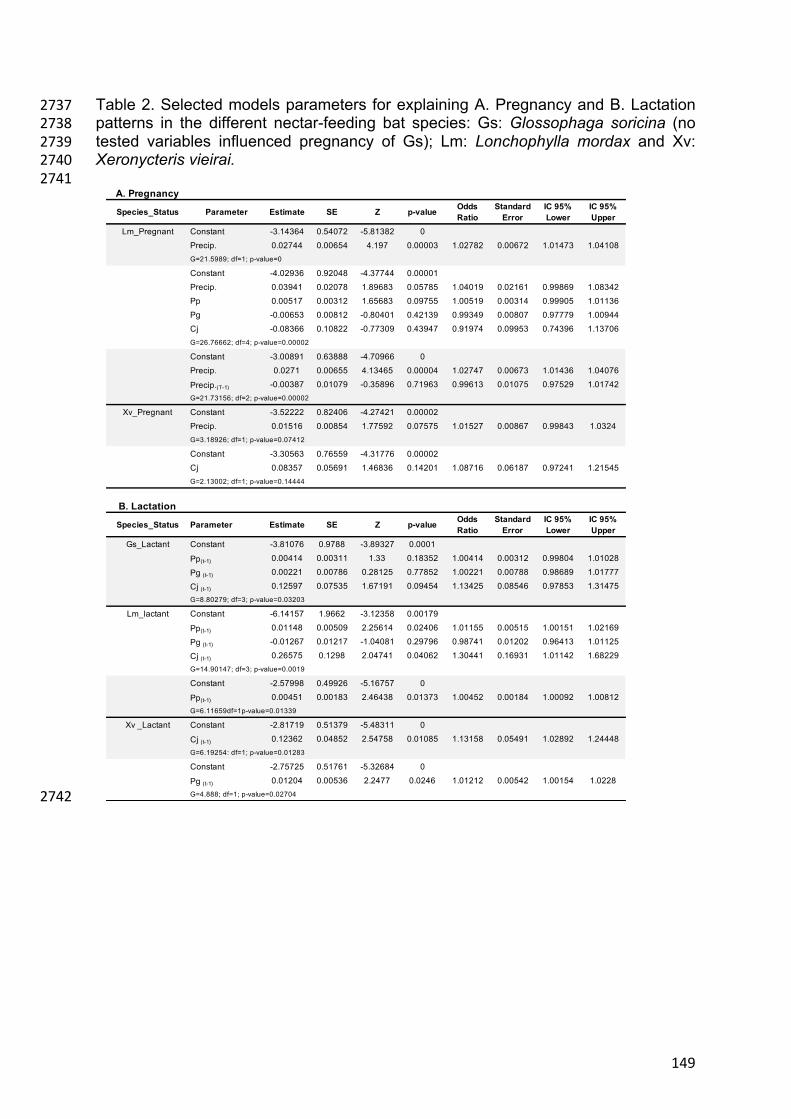
149
Table 2. Selected models parameters for explaining A. Pregnancy and B. Lactation 2737 patterns in the different nectar-feeding bat species: Gs: Glossophaga soricina (no 2738 tested variables influenced pregnancy of Gs); Lm: Lonchophylla mordax and Xv: 2739 Xeronycteris vieirai. 2740 2741
2742
A. Pregnancy
Species_Status Parameter Estimate SE Z p-value Odds Ratio
Standard Error
IC 95% Lower
IC 95% Upper
Lm_Pregnant Constant -3.14364 0.54072 -5.81382 0Precip. 0.02744 0.00654 4.197 0.00003 1.02782 0.00672 1.01473 1.04108G=21.5989; df=1; p-value=0
Constant -4.02936 0.92048 -4.37744 0.00001Precip. 0.03941 0.02078 1.89683 0.05785 1.04019 0.02161 0.99869 1.08342Pp 0.00517 0.00312 1.65683 0.09755 1.00519 0.00314 0.99905 1.01136Pg -0.00653 0.00812 -0.80401 0.42139 0.99349 0.00807 0.97779 1.00944Cj -0.08366 0.10822 -0.77309 0.43947 0.91974 0.09953 0.74396 1.13706G=26.76662; df=4; p-value=0.00002
Constant -3.00891 0.63888 -4.70966 0Precip. 0.0271 0.00655 4.13465 0.00004 1.02747 0.00673 1.01436 1.04076
Precip.(T-1) -0.00387 0.01079 -0.35896 0.71963 0.99613 0.01075 0.97529 1.01742G=21.73156; df=2; p-value=0.00002
Xv_Pregnant Constant -3.52222 0.82406 -4.27421 0.00002Precip. 0.01516 0.00854 1.77592 0.07575 1.01527 0.00867 0.99843 1.0324G=3.18926; df=1; p-value=0.07412
Constant -3.30563 0.76559 -4.31776 0.00002Cj 0.08357 0.05691 1.46836 0.14201 1.08716 0.06187 0.97241 1.21545G=2.13002; df=1; p-value=0.14444
B. Lactation
Species_Status Parameter Estimate SE Z p-value Odds Ratio
Standard Error
IC 95% Lower
IC 95% Upper
Gs_Lactant Constant -3.81076 0.9788 -3.89327 0.0001
Pp(t-1) 0.00414 0.00311 1.33 0.18352 1.00414 0.00312 0.99804 1.01028
Pg (t-1) 0.00221 0.00786 0.28125 0.77852 1.00221 0.00788 0.98689 1.01777
Cj (t-1) 0.12597 0.07535 1.67191 0.09454 1.13425 0.08546 0.97853 1.31475G=8.80279; df=3; p-value=0.03203
Lm_lactant Constant -6.14157 1.9662 -3.12358 0.00179
Pp(t-1) 0.01148 0.00509 2.25614 0.02406 1.01155 0.00515 1.00151 1.02169
Pg (t-1) -0.01267 0.01217 -1.04081 0.29796 0.98741 0.01202 0.96413 1.01125
Cj (t-1) 0.26575 0.1298 2.04741 0.04062 1.30441 0.16931 1.01142 1.68229G=14.90147; df=3; p-value=0.0019
Constant -2.57998 0.49926 -5.16757 0
Pp(t-1) 0.00451 0.00183 2.46438 0.01373 1.00452 0.00184 1.00092 1.00812G=6.11659df=1p-value=0.01339
Xv _Lactant Constant -2.81719 0.51379 -5.48311 0
Cj (t-1) 0.12362 0.04852 2.54758 0.01085 1.13158 0.05491 1.02892 1.24448G=6.19254: df=1; p-value=0.01283
Constant -2.75725 0.51761 -5.32684 0
Pg (t-1) 0.01204 0.00536 2.2477 0.0246 1.01212 0.00542 1.00154 1.0228G=4.888; df=1; p-value=0.02704

150
2743 Figure 3. Females’ diet composition observed in the pollen samples of the four nectar 2744 bat species different reproductive status. NR- Non-Reproductive; P- Pregnant; L-2745 Lactating. The proportion of each non-cacti species used is represented in each grey 2746 compartment. Number inside ( ) is the number of samples from each reproductive 2747 status. 2748
Figure 4. Monthly females’ diet composition observed in the fecal samples of the four 2749 nectar bat species different reproductive status. Glossophaga soricina N= 14 (Non-2750 reproductive-NR= 9; Pregnant-P=2; Lactating-L=3). Lonchophylla inexpectata N=15 2751 (NR= 14; P= 1). Lonchophylla mordax N=17 (NR= 7; P=6; L= 4). Xeronycteris vieirai 2752 N=22 (NR= 11; P=2 L= 9). 2753
2754

151
SUPPORTING INFORMATION 2755 2756 2757 2758
2759 2760 Figure S1. Mean monthly precipitation from 2009 to 2019 (shaded light-blue area) in 2761 the municipality of Lajes in northeastern Brazil. Rainiest year was 2009 (blue dashed 2762 line), driest year 2012 (black dashed line), sampled years precipitation are 2763 represented by continuous colored lines. 2764

152
Figure S2. Cereus jamacaru DC, common name Mandacarú, Cactaceae 2765

153
Figure S3. Pilosocereus gounellei (F.A.C.Weber) Byles & Rowley, common name 2766 Xique- xique Cactaceae 2767

154
Figure S4. Pilosocereus pachycladus F.Ritter, common name Facheiro, Cactaceae 2768

155
Table S5. Selection models of logist regressions for Glossophaga soricina pregnancy 2769 and lactation. AIC - Akaike Information Criterion value; ΔAIC - difference in the Akaike 2770 Information Criterion between each model and the best model; W AIC - Akaike weight 2771 and w AIC um - cumulative weight of AIC. 2772 2773
2774
Glossophaga soricinaA. Pregnancy models AIC Δ AIC w AIC w AIC cumNull model 61.33866 0 18.18 18.18Precip.(T-1) 62.00041 0.66175 13.06 31.23Cj (t-1) 62.00947 0.67081 13.00 44.23Pp 62.8168 1.47814 8.68 52.91Pg 63.27697 1.93831 6.90 59.81Cj 63.29118 1.95252 6.85 66.66Precip. 63.32209 1.98343 6.74 73.40Pp(t-1) 63.3276 1.98894 6.72 80.12Pg (t-1) 63.33841 1.99975 6.69 86.81Precip. + Precip.(T-1) 63.93893 2.60027 4.95 91.77Pp(t-1) + Pg(t-1) + Cj(t-1) 64.34065 3.00199 4.05 95.82Precip.(T-1) + Pp(t-1) + Pg(t-1) + Cj(t-1) 65.27891 3.94025 2.53 98.35Pp + Pg + Cj 66.80748 5.46882 1.18 99.53Precip. + Pp + Pg + Cj 68.65995 7.32129 0.47 100.00
B. Lactation models AIC Δ AIC w AIC w AIC cumPp(t-1) + Pg(t-1) + Cj(t-1) 49.34502 0 74.77 74.77Cj (t-1) 54.22654 4.88152 6.51 81.28Pp(t-1) 54.85174 5.50672 4.76 86.05Cj 55.44522 6.1002 3.54 89.59Pg 55.57414 6.22912 3.32 92.91Precip. 56.66901 7.32399 1.92 94.83Precip. + Precip.(T-1) 57.21488 7.86986 1.46 96.29Null model 58.2465 8.90148 0.87 97.16Pg (t-1) 58.47649 9.13147 0.78 97.94Precip.(T-1) 58.7374 9.39238 0.68 98.62Pp + Pg + Cj 58.86454 9.51952 0.64 99.26Pp 59.42353 10.07851 0.48 99.75Precip. + Pp + Pg + Cj 60.7285 11.38348 0.25 100.00Precip.(T-1) + Pp(t-1) + Pg(t-1) + Cj(t-1) **** unsatisfactory solution

156
Table S6. Selection models of logist regressions for Lonchophylla mordax pregnancy 2775 and lactation. AIC - Akaike Information Criterion value; ΔAIC - difference in the Akaike 2776 Information Criterion between each model and the best model; W AIC - Akaike weight 2777 and w AIC um - cumulative weight of AIC. 2778 2779
2780 2781
Lonchophylla mordaxA. Pregnancy models AIC Δ AIC w AIC w AIC cumPrecip. 71.02498 0 36.12 36.12Precip. + Pp + Pg + Cj 71.85726 0.83228 23.82 59.94Precip. + Precip.(T-1) 72.89232 1.86734 14.20 74.14Pp(t-1) + Pg(t-1) + Cj(t-1) 73.08742 2.06244 12.88 87.02Precip.(T-1) + Pp(t-1) + Pg(t-1) + Cj(t-1) 74.92053 3.89555 5.15 92.17Pp + Pg + Cj 75.48289 4.45791 3.89 96.06Pp 75.76236 4.73738 3.38 99.44Pg 79.68772 8.66274 0.47 99.91Pg (t-1) 84.29037 13.26539 0.05 99.96Cj 85.8122 14.78722 0.02 99.98Pp(t-1) 86.72477 15.69979 0.01 100.00Null model 90.62388 19.5989 0.00 100.00Precip.(T-1) 92.1174 21.09242 0.00 100.00Cj (t-1) 92.59177 21.56679 0.00 100.00
B. Lactation models AIC Δ AIC w AIC w AIC cumPp(t-1) 87.1826 0 42.64 42.64Precip. 90.29003 3.10743 9.02 51.65Pp(t-1) + Pg(t-1) + Cj(t-1) 90.60521 3.42261 7.70 59.36Precip. + Precip.(T-1) 90.79375 3.61115 7.01 66.37Cj 91.01386 3.83126 6.28 72.64Null model 91.29919 4.11659 5.44 78.09Pg (t-1) 91.53587 4.35327 4.84 82.92Precip.(T-1) 91.95728 4.77468 3.92 86.84Precip.(T-1) + Pp(t-1) + Pg(t-1) + Cj(t-1) 92.33949 5.15689 3.24 90.08Cj (t-1) 92.41795 5.23535 3.11 93.19Pg 92.81246 5.62986 2.55 95.74Pp 92.88179 5.69919 2.47 98.21Pp + Pg + Cj 94.69184 7.50924 1.00 99.21Precip. + Pp + Pg + Cj 95.15429 7.97169 0.79 100.00

157
Table S7. Selection models of logist regressions for Xeronycteris vieirai pregnancy 2782 and lactation. AIC - Akaike Information Criterion value; ΔAIC - difference in the Akaike 2783 Information Criterion between each model and the best model; W AIC - Akaike weight 2784 and w AIC um - cumulative weight of AIC. 2785 2786 2787
2788
Xeronycteris vieiraiA. Pregnancy models AIC Δ AIC w AIC w AIC cumPrecipitation 37.27039 0 20.19 20.19Cj 38.32964 1.05925 11.89 32.07Null model 38.45965 1.18926 11.14 43.21Pg (t-1) 38.76249 1.4921 9.57 52.79Precip. + Precip.(T-1) 38.94167 1.67128 8.75 61.54Pp 39.01898 1.74859 8.42 69.96Pg 39.34525 2.07486 7.15 77.12Pp(t-1) 39.84975 2.57936 5.56 82.68Precip.(T-1) 40.23503 2.96464 4.58 87.26Cj (t-1) 40.23887 2.96848 4.58 91.84Pp + Pg + Cj 41.20642 3.93603 2.82 94.66Pp(t-1) + Pg(t-1) + Cj(t-1) 41.41201 4.14162 2.55 97.20Precip. + Pp + Pg + Cj 42.04871 4.77832 1.85 99.05Precip.(T-1) + Pp(t-1) + Pg(t-1) + Cj(t-1) 43.39088 6.12049 0.95 100.00
B. Lactation models AIC Δ AIC w AIC w AIC cumCj (t-1) 55.45911 0 24.87 24.87Pg (t-1) 56.7636 1.30449 12.96 37.83Pp(t-1) + Pg(t-1) + Cj(t-1) 56.84886 1.38975 12.42 50.25Pp + Pg + Cj 57.55236 2.09325 8.73 58.98Precip. + Pp + Pg + Cj 57.69148 2.23237 8.15 67.13Precip.(T-1) 57.89418 2.43507 7.36 74.49Cj 58.52586 3.06675 5.37 79.86Precip.(T-1) + Pp(t-1) + Pg(t-1) + Cj(t-1) 58.78662 3.32751 4.71 84.57Precip. + Precip.(T-1) 59.06233 3.60322 4.11 88.68Null model 59.65166 4.19255 3.06 91.73Pp(t-1) 59.84916 4.39005 2.77 94.50Pp 59.97883 4.51972 2.60 97.10Precip. 60.92345 5.46434 1.62 98.72Pg 61.39118 5.93207 1.28 100.00

158
Table S8. Plants species used by females of each nectar bat species in the different 2789 reproductive status. NR- Non-Reproductive Females; P- Pregnant; L-Lactating. The 2790 single lactating pollen sample of L. inexpectata did not contained any pollen types. 2791 2792
2793 2794
2795
2796
Figure S9. Proportion of lactating females of the three nectar-feeding bats. The 2797
underlined months indicate the primary lactation peak for the different bat species. 2798
2799
Family Plant species G. soricina L. inexpectata L. mordax X. vieiraiAcanthaceae Harpochilus paraibanus NR | P | L NR NR | P | L NR
Dicliptera ciliaris NR | P NR NR | P | L NRRuellia asperula NR NR NR | L NRHarpochilus neesianus ・ ・ NR ・
Bignoneaceae Tabebuia aurea NR NR NR | L NRBromeliaceae Encholirium spectabile NR | P | L NR NR | P | L LCactaceae Pilosocereus pachycladus NR | P | L NR | P NR | P | L NR | P | L
Pilosocereus gounellei NR | P | L NR | P NR | P | L NR | P | LMelocactus zehntneri NR | P | L NR | P NR | P | L NR | P | LCereus jamacaru NR | P | L NR | P NR | P | L NR | P | LPilosocereus chrysostele NR | P | L P NR | P | L NR | P
Capparaceae Cynophalla hastata NR ・ NR | P ・
Cleomaceae Tareyana spinosa NR | P NR NR ・
Convolvulaceae Ipomoea sp. ・ NR NR | L ・
Euphorbiaceae Jatropha mollissima ・ NR ・ NRFabaceae Bauhinia cheilantha NR | P NR NR | P ・
Bauhinia pentandra NR ・ NR | L P | LCoursetia rostrata NR ・ ・ ・
Fabaceae sp. ・ NR NR ・
Malvaceae Helicteres baruensis NR | P NR | P NR | P | L NRPseudobombax ・ NR NR | L NR
Xanthorrhoeaceae Aloe vera NR ・ ・ ・
. Monocot sp. NR ・ NR ・
Total used plant species 18 17 20 14

159
- At 2800 2801 At least five species can co-occur:
- Anoura geoffroyi, the rarest. Mostly captured in the humid forest enclave and one individual was captured in a riparian Caatinga. This species feeds on at least eight plant species.
- Glossophaga soricina, the most common and abundant. Captured in all sampled localities and types of habitat. It feeds on 26 plant species and complements its diet mostly with insects, but also with pollen and plant tissues.
- Lonchophylla inexpectata, a Caatinga endemic, this species had the second lowest occurrence. Only captured in the locality of Lajes in riparian and shrubby Caatinga. It feeds on 20 plant species and complements its diet with insects, pollen and plant tissues.
- Lonchophylla mordax, the second most common and abundant species. Captured in Copernicia groves, Medium, Shrubby and Riparian Caatinga and in Rocky outcrops. It feeds on 28 plant species and complements its diet with insects, pollen and plant tissues.
- Xeronycteris vieirai, endemic to the dry diagonal crossing central Brazil. Captured only in Lajes, in riparian and shrubby Caatinga. It feeds on 21 plant species and complements its diet mainly with pollen but also with insects and plant tissues.
- The interaction networks between nectar bats and plants is highly generalized and showed high overlap. This associations are stable through the seasons and years.
- Nectar bats showed ecological and phenotypical generalization in the plants they include in their diets.
- The species coexisted and persisted year-round in Caatinga by presenting temporal partitioning within the hours of the night, resource partitioning facilitated by ecomorphological differences and diet flexibility.
- Temporal persistence of nectar bats was facilitated by the continuity and complementarity of floral resources available through the year and the high use of keystone resources.
- Female reproduction patterns of the studied nectar-feeding bats are bimodal (G. soricina and apparently L. inexpectata) and polymodal (L. mordax and X. vieirai).
- Pregnancy was influenced by precipitation in L. mordax and X. vieirai but not for G. soricina.
- Lactation was influenced by Cactaceae resource availability of the previous month for the three nectar bat species.
- Cactaceae are keystone resources for nectar bats in Caatinga. At least five species were widely used, but Pilosocereus pachycladus stood out. Conservation initiatives should consider sites with high abundance and diversity of Cactaceae as a conservation priority.

160
2802




















