
Improvement of the nutritional value of rotifers by varying the type and concentration of oil and...
13
Aquaculture 147 (1996) 93-105 Improvement of the nutritional value of rotifers by varying the type and concentration of oil and the enrichment period C. Rodriguez a, J.A. P6rez a, M.S. Izquierdo b, J.R. Cejas ‘, A. Boltios a, A. Lorenzo a7* a Departumento de Biolo~iu Animul. Fucultud de Bichgiu, UkwGkul de Lu Luguna, 38270 La Lo~unu. Tenerfle, Conury I.dunds, Spin b Depurtumento de Biologh, Fucultud de Ciencius de1 Mar. Uniuersidud de Lus Pulmus de Grun Cunuriu. Tujiru Buju, 35019 Las Pulmus de Grun Cunuriu, Cunury Islond~. Spin ’ Centro Oceunogrcifico de Cunurius, Institute Espuiiol de Oceunopw& Curreteru de Sun AndrGs .s/ n. 38120 S/C de Teneri,k, Tenerijk. Cunury I.tlund~s, Spuin Accepted 24 June I996 Abstract Several enrichment conditions (type and concentration of oil in culture medium and time of enrichment) were tested in order to study their influence on rotifer (Bra&onus plicatilis) lipid and fatty acid composition as well as on its population profile. Rotifers pre-fed on baker’s yeast (Suc~haromyces cereuisiae) were enriched with lipid emulsions containing triacylglycerols (TAG) or methyl ester (ME) mixtures at three different concentrations. Rotifers were sampled after 0 (yeast-fed rotifer), 3, 6, I2 and 24h enrichment. At every concentration and time assayed, the highest lipid levels were obtained when using TAG. In spite of the differences existing in saturated and monounsaturated fatty acid composition between the enrichment emulsions, rotifers tended to maintain the level of these fatty acids independently of the enrichment period and oil concentration. Increasing the enrichment period rather than the amount of oil present in the medium was found to be more efficient in increasing the n - 3 highly unsaturated fatty acids (n - 3 HUFA) level in rotifers. Rotifers reflected a better incorporation of eicosapentaenoic acid (EPA, 20:5n - 3) than of docosahexaenoic acid (DHA, 22:6n - 3), regardless of the ratio between * Correspondence to: Dr.A. Lorenzo HemBndez, Fisiologia Animal, Departamento de Biologia Animal, Facultad de Biologia, Universidad de La Laguna, 38271 Tenerife, Canary Islands, Spain. 0044-8486/96/$lS.O0 Copyright 0 1996 Elsevier Science B.V. All rights reserved. PI1 SOO44-8486(96)0 1397-X
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Improvement of the nutritional value of rotifers by varying the type and concentration of oil and...

Aquaculture 147 (1996) 93-105
Improvement of the nutritional value of rotifers by varying the type and concentration of oil and the
enrichment period
C. Rodriguez a, J.A. P6rez a, M.S. Izquierdo b, J.R. Cejas ‘, A. Boltios a, A. Lorenzo a7*
a Departumento de Biolo~iu Animul. Fucultud de Bichgiu, UkwGkul de Lu Luguna, 38270 La Lo~unu.
Tenerfle, Conury I.dunds, Spin
b Depurtumento de Biologh, Fucultud de Ciencius de1 Mar. Uniuersidud de Lus Pulmus de Grun Cunuriu.
Tujiru Buju, 35019 Las Pulmus de Grun Cunuriu, Cunury Islond~. Spin
’ Centro Oceunogrcifico de Cunurius, Institute Espuiiol de Oceunopw& Curreteru de Sun AndrGs .s/ n. 38120
S/C de Teneri,k, Tenerijk. Cunury I.tlund~s, Spuin
Accepted 24 June I996
Abstract
Several enrichment conditions (type and concentration of oil in culture medium and time of enrichment) were tested in order to study their influence on rotifer (Bra&onus plicatilis) lipid and fatty acid composition as well as on its population profile. Rotifers pre-fed on baker’s yeast (Suc~haromyces cereuisiae) were enriched with lipid emulsions containing triacylglycerols (TAG) or methyl ester (ME) mixtures at three different concentrations. Rotifers were sampled after 0 (yeast-fed rotifer), 3, 6, I2 and 24h enrichment. At every concentration and time assayed, the highest lipid levels were obtained when using TAG. In spite of the differences existing in saturated and monounsaturated fatty acid composition between the enrichment emulsions, rotifers tended to maintain the level of these fatty acids independently of the enrichment period and oil concentration. Increasing the enrichment period rather than the amount of oil present in the medium was found to be more efficient in increasing the n - 3 highly unsaturated fatty acids (n - 3 HUFA) level in rotifers. Rotifers reflected a better incorporation of eicosapentaenoic acid (EPA, 20:5n - 3) than of docosahexaenoic acid (DHA, 22:6n - 3), regardless of the ratio between
* Correspondence to: Dr.A. Lorenzo HemBndez, Fisiologia Animal, Departamento de Biologia Animal,
Facultad de Biologia, Universidad de La Laguna, 38271 Tenerife, Canary Islands, Spain.
0044-8486/96/$lS.O0 Copyright 0 1996 Elsevier Science B.V. All rights reserved.
PI1 SOO44-8486(96)0 1397-X

94 C. Rodriguez et al./Aquaculture 147 (1996) 93-105
both fatty acids in the emulsions. The use of ME as an enrichment source produced greater mortality in egg-cawing females than when using TAG.
Keywords: Rotifers; Enrichment conditions; Lipid emulsions; n - 3 HUFA; EPA/DHA
1. Introduction
Marine fish larvae have been shown to require n - 3 highly unsaturated fatty acids (n - 3 HUFA) such as eicosapentaenoic acid (EPA, 22:5n - 3) and docosahexaenoic acid (DHA, 22:6n - 3) for their development and survival (Fujita, 1979; Izquierdo, 1988; Koven et al., 1989, 1992; Rodriguez et al., 1993, 1994). The rotifer Brachionus
plicatilis has been extensively used as live food for rearing fish larvae due to its appropriate size, rapid production rate and suitability for mass culture under controlled conditions (Watanabe et al., 1983a). Although rotifers have been found to synthesize some n - 3 HUFA (Lubzens et al., 1985) the amount accumulated is small and insufficient to fulfil the possible demand of the fish larvae. In this sense, various enrichment techniques are used in hatcheries to increase rotifer n - 3 HUFA content, such as feeding with algae prior to introduction into fish tanks, through the addition of oil emulsions to a baker’s yeast (Sacccharumyces cereuisiae) diet (Watanabe et al., 1983b) or directly into rotifer tanks (Watanabe et al., 1982). Previous studies have shown that fatty acid composition of rotifers is readily affected by the fatty acid content of the diet (Rainuzzo et al., 1989) and that the content of 20:5n - 3 and 22:6n - 3 in the rotifers chiefly determines their nutritional value as living food (Watanabe et al., 1989; Rodriguez et al., 1993, 1994). The EPA and DHA levels in rotifers depend not only on the level of these fatty acids present in the lipid emulsion but also on the enrichment conditions (Izquierdo, 1988; Rainuzzo et al., 1994).
The aim of the present study was to investigate the incorporation of different groups of fatty acids and particularly n - 3 HUFA by rotifers, when varying the type of oil, its concentration in the culture medium and the enrichment period. The intention was to determine which combination of these variables would be most suitable for application in larval production.
2. Material and methods
2.1. Experimental design
Rotifers which had been cultured on baker’s yeast in the Centro Oceanografico de Canarias of the Spanish Oceanographic Institute were divided into six 40 1 tanks filled with filtered 30 g 1-l salinity sea water at a density of approximately 150 individuals ml-’ and enriched by the direct method described by Watanabe et al. (1982). The culture was strongly aerated and the water temperature ranged between 17-19°C.
Lipid emulsions were prepared by mixing 100 ml of sea water, 1 g of raw egg yolk, 0.5 g of soybean lecithin and sufficient oil to obtain concentrations of 4, 8 and 12% oil

C. Rodriguez et nl./Aquncuhure 147 (1996) 93-105 95
in rotifers medium. Rotifers from three tanks were enriched with 4, 8 or 12% of triacylglycerols (TAG) containing 40% n - 3 HUFA. The other three tanks received methyl ester (ME) containing 81% n - 3 HUFA prepared at the same three concentra- tions mentioned above. Both oils were supplied by Nippai Chemical Company, Tokyo.
For each concentration of oil, one sample of rotifers (2 g> was taken after 3, 6, 12 and 24h.’ A sample of rotifers exclusively fed on yeast was also taken. Rotifers were concentrated with a 60 pm scoop mesh, thoroughly washed with freshwater, carefully dried with filter paper and stored at - 40°C until analysis. Each experiment was carried out in triplicate.
Before sample collection, the population profile of one replicate was studied by estimating the number of rotifers ml-’ and the percentage of egg-carrying females.
2.2. Analytical procedures
Total lipid and fatty acid composition of each sample were determined. Total lipid was extracted after homogenization in chloroform-methanol (2: 1, v/v> as described by Folch et al. (1957). Fatty acid mixtures were prepared from the crude lipids by saponification with KOH. Unsaponifiable matter was determined gravimetrically. Fatty- acid MEs were prepared by transesterification with borontrifluoride in methanol accord- ing to Metcalfe and Schmitz (1961). The fatty acid MEs were analysed in a Shimadzu GC-14A gas chromatograph (Shimadzu Corp., Kyoto, Japan) equipped with a flame ionization detector (250°C) and a Supelcowax-IO fused-silica capillary column (30 m x 0.32 mm I.D., Supelco, Bellefonte, PA, USA) using helium as carrier gas. The initial temperature was 180°C for 10 min followed by a thermal gradient to 215°C at 2.5 C min-’ and then maintained for 12 min. Individual fatty acid MEs were identified by reference to authentic standards and to a well characterized fish oil. The MEs were quantified using a Shimadzu C-R4A recording integrator.
2.3. Statistical analysis
All biochemical results are given as mean f s.e.m. and were subjected to a one-way analysis of variance (ANOVA) (Zar, 1984). Differences between means were studied using Tukey’s multiple range test (P < 0.05) (Sokal and Rohlf, 1981). Biological data are expressed as percentage of variation after 24 h.
3. Results
The fatty acid content of rotifers fed baker’s yeast as well as of the oils used for the rotifer enrichment regimes (TAG and ME) are shown in Table 1. Total lipid accounted for 13.15% of the dry weight (% DW) of the initial rotifers, the predominant fatty acids being 16: 1 n - 7, 18: 1 n - 9 and 16:0. n - 3 And n - 6 fatty acids were almost absent.
Comparing the fatty acid composition of the two enrichment sources, the content of saturated and monounsaturated fatty acids in TAG were 18.89% and 25.74%, respec- tively, while the ME did not contain saturated fatty acids and had only a very low
% C. Rodriguez et aL/Aquaculture 147 (1996) 93-105
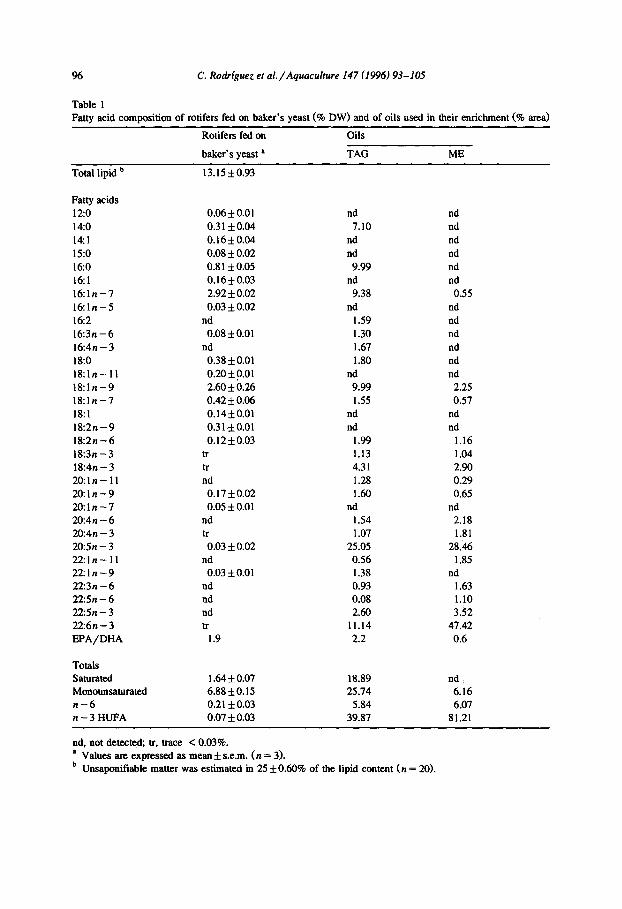
Table 1 Fatty acid composition of rotifers fed on baker’s yeast (% DW) and of oils used in their enrichment (I area)
Rotifers fed on Oils
baker’s yeast ’ TAG ME
Total lipid b 13.15*0.93
Fatty acids 12:o 14:o 14:l 15:o 16:O 161 16:ln-7 16:ln-5 162 16:3n -6 16:4n-3 18:o 18:ln-11 18:ln-9 18:ln-7 18:l 18:2n-9 18:2n-6 18:3n-3 18:4n - 3 20:ln- 11 20:ln-9 20:ln-7 20~411-6 20:4n - 3 20:5n - 3 22:ln-11 22:ln-9 22:3n-6 22:5n - 6 22:5n-3 22:6n-3 EPA/DHA
0.06 f 0.01 0.3 1 f 0.04 0.16*0.04 0.08 f 0.02 0.8 1 f 0.05 0.16kO.03 2.92 f 0.02 0.03 f 0.02
nd 0.08!~0.01
nd 0.38 f 0.01 0.2o*p.o1 2.60 f 0.26 0.42 f 0.06 0.14*0.01 0.31~*0.01 0.12f0.03
tr tr nd 0.17kO.02 0.05 f 0.0 1
nd tr 0.03 f 0.02
nd 0.03 f 0.01
nd nd nd tr
1.9
nd nd 7.10 nd
nd nd nd nd
9.99 nd nd nd
9.38 0.55 nd nd
1.59 nd 1.30 nd 1.67 nd 1.80 nd
nd nd 9.99 2.25 1.55 0.57
nd nd nd nd
1.99 1.16 1.13 1.04 4.31 2.90 1.28 0.29 1.60 0.65
nd nd 1.54 2.18 1.07 1.81
25.05 28.46 0.56 1.85 1.38 nd 0.93 1.63 0.08 1.10 2.60 3.52
11.14 47.42 2.2 0.6
Totals Saturated Monounsaturated n-6 n-3 HUFA
1.64 f 0.07 6.88zkO.15 0.2 1 f 0.03 0.07 l 0.03
18.89 nd 25.74 6.16
5.84 6.07 39.87 81.21
nd, not detected; tr, trace < 0.03%. a Values are expressed as mean It s.e.m. (n = 3). b Unsaponifiable matter was estimated in 25 f 0.60% of the lipid content (n = 20).

C. Rodrguez et al./Aquaculture 147 (1996) 93-105 97
Table 2 Total lipid content of rotifers enriched with TAG (% DW ‘)
Enrichment Oil concentration in rotifers medium (%) period (h) 4 8 12
0 13.15*0.93 a 13.1.5~0.93 B 13.15rto.93 B 3 20.91 k 1.94 b 23.14* 1.16 b 26.50 f 2.66 b 6 20.19 f 0.50 b 21.97* 1.14 b 27.19 f 3.23 b
12 24.405 1.72 b 24.88 f 2.56 b 32.21 f4.14 b 24 25.93 f 1.26 b 30.46 f 3.87 b 36.94k2.13 b
Values are expressed as mean f s.e.m. from a sample with n = 3. Means in the same column with different letters are significantly different (P < 0.05). ’ Unsaponifiable matter was estimated in 25 f 0.60% of the lipid content as a mean of 20 samples.
amount of monoenes (6.16%). The TAG had 40% n - 3 HUFA in which the amount of EPA was twice that of DHA (EPA/DHA ratio of 2.2) while the ME contained 81% n - 3 HUFA with an EPA/DHA ratio of 0.6. EPA and total n - 6 fatty acids series were almost equally present in both oils.
Total lipid content of rotifers increased, in general terms, over time and with the elevation of the amount of oil present in the culture medium (Tables 2 and 3). Considering total lipid present in rotifers as a function of the enrichment period, the increases were significant at 3 h for TAG and at 6 to 12 h for MEL However, these significant increases were not observed when raising the oil concentration. The amount of lipid present in rotifers for each time and concentration was generally lower when using ME than TAG. For each oil, the highest total lipid content was obtained with 12% oil and 24 h enrichment.
Fig. l(a, b and c> illustrate variation with time of the main fatty acid groups in rotifers enriched with TAG at concentrations of 4, 8 and 12%, respectively. In spite of the high amount of saturated fatty acids present in the oil (18.89%), this group varied little over time in the enriched rotifers. Monounsaturated fatty acids (25.74% of the oil) rose significantly after 3 h of enrichment at all the concentrations assayed. n - 6 Fatty acids, present in the oil at only 5.84% were significantly incorporated into the rotifers after 3 h enrichment at every concentration but, as occurred with monoenes, longer
Table 3 Total lipid content of rotifem enriched with ME (% DW ‘)
Enrichment Oil concentration in rotifem medium (0) period (h) 4 8
0 13.15Ito.93 = 13.15 f0.93 .?
3 19.73rt 1.78 a 19.64rt2.80 ’ 6 21.98rt 1.05 b 19.80 2.70 f a
12 23.43 f 2.39 b 24.59 f 2.60 b 24 23.23 + 1.41 b 30.08 f 0.20 b
12
13.15rto.93 a 18.66 f 0.67 ’ 21.4Srt 1.65 b 25.41 f 1.51 b 31.67& 1.33 b
Values am expressed as mean f s.e.m. from a sample with n = 3. Means in the same column with different ptters are significantly different (P < 0.05).
Unsaponifiable matter was estimated in 25 rtO.60% of the lipid content as a mean of 20 samples.

98 C. Rodriguez et al./Aquaculture 147 (1996) 93-105
w
(4
6-
0 3 6 d 12 15 18 21 24
0 3 6 9 12 15 18 21 24
8-
:/ il
6
-i-_:
b
6 9 12 is lk 2’1 2h
Enrichment period (h)
0 Saturates r-i t Monoenes - n-6 + n-3 HUFA
Fig. 1. Time course variation of fatty acid composition in rotifers enriched with 4% (a), 8% (b) or 12% (c) of a triacylglycerols mixture (TAG).Values in the same group of fatty acids with different letters are significantly different (P < 0.05). Error bars give the standard error of the mean of three replicates.

C. Rodriguez et al./Aquaculture 147 (1996) 93-105 99
0 3 6 9 12 15 IX 21 24
12 -__ r
0
0 Saturates + Monoenes + n-6 * n-3 HUFA
0 3 6 9 12 15 18 21 24
Enrichment period (h)
Fig. 2. Time course variation of fatty acid composition in rotifers enriched with 4% (a), 8% (b) or 12% (c) of a methyl esters mixture (ME).Values in the same group of fatty acids with different letters are significantly different (P < 0.05). Error bars give the standard error of the mean of three replicates.
100 C. Rodriguez et al./Aquacubure 147 (1996) 93-105
Table 4 EPA/DHA ratio of rotifers enriched with TAG
Enrichment Oil concentration in rotifers medium (%) period (h) 4 8 12
3 2.8*0.7 3.9f0.2 2.8fO.l 6 3.8 f 0.3 3.8 f 0.5 3.3 f 0.3
12 2.9kO.2 3.3 f 0.0 2.9f0.2 24 2.9kO.4 2.9f0.4 2.8 f 0.3
Values are expressed as mean f s.e.m. from a sample with R = 3. Rows and columns lacking letters mean no significant differences found (P < 0.05).
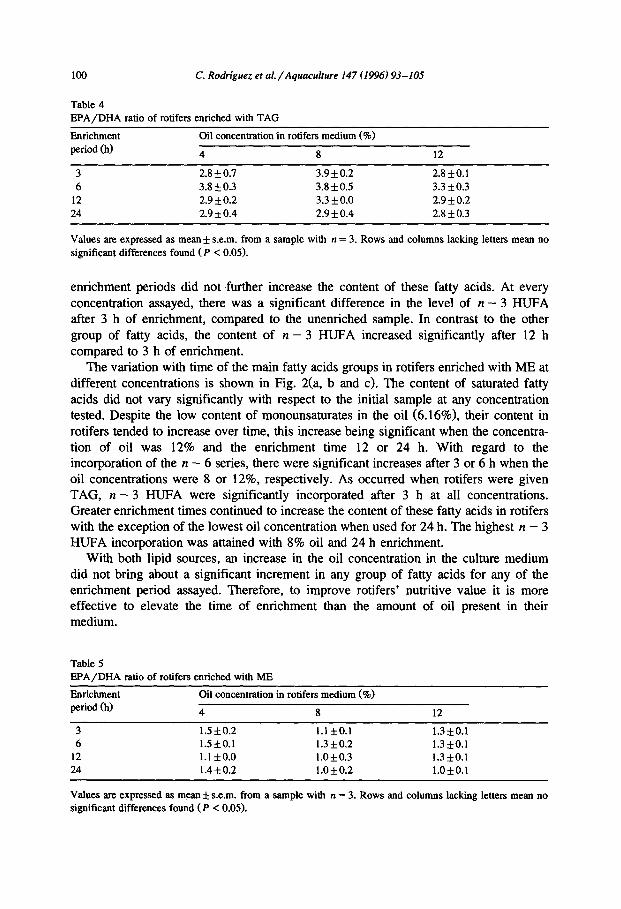
enrichment periods did not further increase the content of these fatty acids. At every concentration assayed, there was a significant difference in the level of n - 3 HUFA after 3 h of enrichment, compared to the unenriched sample. In contrast to the other group of fatty acids, the content of n - 3 HUFA increased significantly after 12 h compared to 3 h of enrichment.
The variation with time of the main fatty acids groups in rotifers enriched with ME at different concentrations is shown in Fig. 2(a, b and c). The content of saturated fatty acids did not vary significantly with respect to the initial sample at any concentration tested. Despite the low content of monounsaturates in the oil (6.16%), their content in rotifers tended to increase over time, this increase being significant when the concentra- tion of oil was 12% and the enrichment time 12 or 24 h. With regard to the incorporation of the n - 6 series, there were significant increases after 3 or 6 h when the oil concentrations were 8 or 12%, respectively. As occurred when rotifers were given TAG, n - 3 HUFA were significantly incorporated after 3 h at all concentrations. Greater enrichment times continued to increase the content of these fatty acids in rotifers with the exception of the lowest oil concentration when used for 24 h. The highest n - 3 HUFA incorporation was attained with 8% oil and 24 h enrichment.
With both lipid sources, an increase in the oil concentration in the culture medium did not bring about a significant increment in any group of fatty acids for any of the enrichment period assayed. Therefore, to improve rotifers’ nutritive value it is more effective to elevate the time of enrichment than the amount of oil present in their medium.
Table 5 EPA/DHA ratio of rotifers enriched with ME
Enrichment Oil concentration in rotifers medium (%I period (h) 4 8 12
3 l.Sf0.2 1.1*0.1 1.3fO.l 6 1..5*0.1 1.3kO.2 1.3 *0.1
12 1.1 fO.O l.OjzO.3 1.3*0.1 24 1.4f0.2 1.0*0.2 1.0*0.1
Values are expressed as mean f s.e.m. from a sample with n = 3. Rows and columns lacking letters mean no significant differences found (P < 0.05).
C. Rodriguez et al/Aquacuhue I47 (1996) 93-105 101
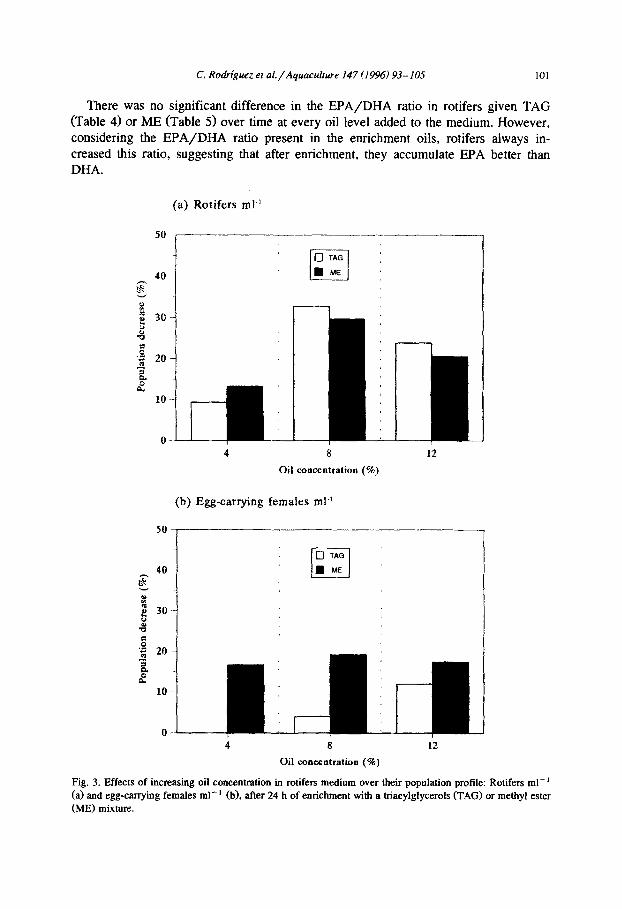
There was no significant difference in the EPA/DHA ratio in rotifers given TAG (Table 4) or ME (Table 5) over time at every oil level added to the medium. However, considering the EPA/DHA ratio present in the enrichment oils, rotifers always in- creased this ratio, suggesting that after enrichment, they accumulate EPA better than DHA.
(a) Rotifers ml-’
1 8
-I 12
Oil concentration (%)
(b) Egg-carrying females m1”
4 I3 12
Oil concentralion (%)
Fig. 3. Effects of increasing oil concentration in rotifers medium over their population profile: Rotifers ml- ’ (a) and egg-carrying females ml-’ (b), after 24 h of enrichment with a triacylglycerols (TAG) or methyl ester
(ME) mixture.

102 C. Rodriguez et al./Aquaculture 147 (1996) 93-105
Fig. 3(a and b) show the results of a preliminary study on the effect of oil concentration on rotifer population profiles after 24 h enrichment. When enriching with 4% of both lipid sources the population density underwent slight modifications over time while oil concentrations of 8 and 12%, produced sharper decreases. After 24 h, the reductions in the number of rotifers ml-’ were 9.3, 32.7 and 23.8% for TAG at concentrations of 4, 8 and 12%, respectively, and of 13.2, 29.7 and 20.5% for ME. Therefore, independently of the type of enrichment oil, rotifer mortality was similar at each concentration and was less marked when the percentage of oil in the medium was lowest (Fig. 3a). The amount of egg-carrying females decreased slightly upon emich- ment with 8 and 12% of TAG; this decrease was more notable at all concentrations when the enrichment source was ME (Fig. 3b).
4. Discussion
The rapid increase in total lipid content of rotifers observed during the enrichment process was similar to that reported by other authors in experiments carried out with lipid emulsions (Watanabe et al., 1983b; Izquierdo et al., 1989; Rainuzzo et al., 1994) or commercial diets (Femhdez et al., 1987).
In general terms, the fatty acid content of rotifers reflects that of the enrichment emulsions (Izquierdo, 1988; Rainuzzo et al., 1989, 1994; Rodriguez et al., 1993, 1994). However, certain patterns can be observed in the incorporation of the different groups bf fatty acids, probably due to their metabolic activities or enzymatic affinities. In this regard, the levels of monoenes present in rotifers enriched with TAG or ME were similar, despite the fact that the TAG monoenes component was considerable while that of ME was small. Likewise, the content of saturated fatty acids tended to remain constant in spite of not being present among the components of the ME emulsion. The behaviour of rotifers with respect to n - 6 and n - 3 fatty acids seemed to differ from the saturated and monoenes. In this case, after 3 h of enrichment, n - 6 fatty acids attained values up to 10 times greater than that of the initial rotifers and, as other authors have pointed out, n - 3 HUFA fatty acids were also assimilated to a great extent in rotifers after 3 h of enrichment (Rainuzzo et al., 1989).
To achieve increments of n - 3 HUFA in rotifers, it is more effective to increase the enrichment time than the amount of oil present in the culture medium. However, after 24 h of enrichment the content of n - 3 HUFA decreased when the oil concentration was at a minimum (4%). This seems to indicate that 4% oil was insufficient when the enrichment period was prolonged. Walford and Lam (19871, using microcapsules, recommended enrichment times over 5 h and observed maximum assimilation of n - 3 HUFA after 12 h. Watanabe (1993) suggested enrichment for about 6 h, considering the optimum enrichment period to be 12 h for rotifers and nauplii of Artemia fed on lipid emulsions. McEvoy et al. (1995) showed that enrichment periods of 24 h increased the risk of perioxidation of polyunsaturated fatty acids in Artemiu enrichment medium. These authors suggested that potentially toxic oxidation products could negatively affect the live prey population and possibly the performance of larval fish preying on them. In addition, minimizing the time required for enrichment reduces the cost associated with live food production (Sorgeloos et al., 1986; Southgate and Lou, 1995).

C. Rodriguez et al./Aquacuiture 147 11996) 93-105 103
In the present work, TAG and ME emulsions containing 4% oil both generated approximately 4% II - 3 HUFA on a dry weight basis of rotifers after 12 h of enrichment. The literature indicates that this amount should be sufficient to meet the requirements of marine fish larvae (Le Milinaire et al., 1983; Izquierdo, 1988; Koven, 1991). Oil concentrations above 8% are not recommended for the wellbeing of the rotifer population or from an economic standpoint. Although further replication of the population study is required, the results presented in Fig. 3 indicate that when the oil concentration in the medium was 4%, the mortality was lower for both lipid sources than when using 8 or 12%. This could be due to the fact that an excess of oil in the medium could lead to water quality degradation and to enhanced bacterial growth or, as occurs in fish (Dabrowski and Poczycznski, 1988) an excess of lipid in the diet could be harmful to the rotifers.
On the other hand, the results also showed that the incorporation of n - 3 HUFA was lower when the rotifers were enriched with ME than with TAG. The reduced incorpora- tion observed may not only be due to the type of oil used but also to the excess n - 3 HUFA present therein. Leger (1985) has shown that an excess of n - 3 HUFA affects lipid metabolism in fish, hindering the incorporation of these fatty acids. This could also account for the reduced incorporation of total lipid when rotifers were enriched with ME. At the same time, rotifers given ME might yield lower n - 3 HUFA levels due to their previously mentioned difficulty in accumulating DHA with respect to EPA, since the emulsion was very rich in the first fatty acid. Furthermore, production of toxic methanol during digestion of ME also could produce a negative effect on the population and their enrichment.
Previous studies have shown that the EPA/DHA ratio in the diet is important for larval development (Mourente et al., 1993; Reitan et al., 1994; Rodriguez, 1994). Therefore, the assimilation capacities of live prey for both fatty acids should be taken into account when using them for larval feeding. Considering the EPA and DHA contributions in oils, it seems that in the present study, rotifers accumulate EPA better than DHA. This was manifested particularly when rotifers were enriched with ME since the EPA/DHA ratio was maintained between 1 and 1.5 (Table 5) despite the fact that the oil presented twice as much DHA than EPA. This greater accumulation of EPA with respect to DHA was previously observed in rotifers by Rainuzzo et al. (19941, but its cause is still unclear. Rotifers are found mainly in freshwater, their original habitat (Dormer, 1966), filtering phytoplankton and organisms that feed on these microalgae. Since in the majority of freshwater microalgae, EPA is the dominant fatty acid among the n - 3 HUFA (Stewart, 19741, rotifers enzymatic system could be adapted to incorporate EPA more readily than DHA. Similar results have been described for Arfemia, which accumulates DHA with difficulty even when its diet is rich in this fatty acid (Hinchcliffe and Riley, 1972; Izquierdo, 1988). Recently, McEvoy et al. (1995) have shown that the EPA/DHA ratios in Anemia fed on three different emulsions were always considerably higher than the EPA/DHA ratios in the original enrichment medium. It was thought that a possible explanation for the lower accumulation of the DHA with respect to EPA might be a faster autoxidation rate of DHA in the enrichment media. However, examination of the EPA/DHA ratio of the Artemia medium over time negated this hypothesis. Instead, it appears that Artemiu might metabolize DHA to a

104 C. Rodriguez et al./Aquaculture 147 (1996) 93-105
larger extent than EPA. Due to the importance of DHA for marine fish larvae development (Izquierdo et al., 1989; Watanabe, 1993; Rainuzzo et al., 1994; Rodriguez, 1994) it is important to consider that, in rotifers, this fatty acid is accumulated to a lesser extent than EPA.
In summary, the results of the present work indicate that in order to obtain increased n - 3 HUFA content in rotifers, it is more effective to prolong the enrichment period than to increase the amount of oil present in the culture medium. The best results in terms of nutritive value and population state of rotifers, were obtained with oil concentrations between 4 and 8%, and an enrichment period of no longer than 12 h. Since n - 3 HUFA incorporation was lower when using ME and greater mortality of egg-carrying females occurred with this lipid source, it is considered more advisable to utilize TAG. Moreover, some authors have reported marine fish larvae mortalities related to the use of ME as a live prey enrichment source (Izquierdo, 1988; Rodriguez, 1994).
Acknowledgements
This project was funded by the Autonomous Canary Islands Government (Project No. 28/01.06.88) and a research grant from the Spanish Education and Science Ministry (F.P.I. 1988).
We are most grateful to the Nippai Chemical Company, Tokyo, Japan for providing the experimental oils used in this study.
References
Dabrowski, K. and Poczycznski, P., 1988. Laboratory experiment and mass rearing of coregonid fish fed exclusively on dry diets. Aquaculture, 69: 307-316.
Dormer, J., 1966. Rotifers. Frederick Warne, London and New York, 80 pp. Femhdez, M.J., Ferreiro, M.T., Labarta, U. and Munilla, R., 1987. Influencia de dietas emiquecedoras en la
calidad nutritiva de Brachionus plicarilis y Anemia sp. Cuad. Marisq. Publ. TIC., 12: 579-584. Folch, J., Lees, M. and Sloane Stanley, G.H., 1957. A simple method for the isolation and purification of total
lipids from animal tissues. J. Biol. Chem., 226: 497-509. Fujita, S., 1979. Culture of red sea bream, Pagrw major, and its food. In: E. Styczynska-Jurewicz, T. Backiel,
E. Jaspers and G. Persoone (Editors), Cultivation of Fish Fry and Its Live Food. Euro. Maricult. Sot. Special Publ. No. 4, Bredene, Belgium, pp. 183-197.
Hinchcliffe, P.R. and Riley, J.P., 1972. The effect of diet on the component fatty acid composition of Artemia salina. J. Mar. Biol. Assoc. UK, 52: 203-211.
Izquierdo, M.S., 1988. Estudio de 10s requerimientos de Lidos grasos esenciales en latvas de peces marinos. ModificaciQ de la composici6n lipidica de las presas. PhD Thesis, University of La Laguna, Spain, 205
PP. Izquierdo, M.S., Watanabe, T., Takeuchi, T., Arakawa, T. and Kitajima, C., 1989. Optimal EFA levels in
Artemia to meet the EFA requirements of red seabream (Pagrus major). Proc. Third Int. Symp. Feeding and Nutr. in Fish. Toba (Japan). Aug. 28-Sept. 1, pp. 221-232.
Koven, W.M., 1991. The combined effect of dietary n - 3 highly unsaturated fatty acids and age, on growth, survival and lipid composition in larval gilthead seabream (Sparus aurora; Perciformes, Teleostei). PhD Thesis, Hebrew University, Israel, 130 pp.
Koven. W.M., Kissil, G.Wm. and Tandler, A., 1989. Lipid and n-3 HUFA requirement of Sparus aurata larvae during starvation and feeding. Aquaculture, 79: 185- 191.

C. Rodriguez et al./Aquaculture 147 (1996) 93-105 105
Koven, W.M., Tandler, A., Kissil, GWm. and Sklan, D., 1992. The importance of n - 3 highly unsaturated
fatty acids for growth in larval Sparus uurata and their effect on survival, lipid composition and size
distribution. Aquaculture, 104: 91-104.
Leger, C., 1985. Digestion, absorption and transport of lipids. In: C.B. Cowey, A.M. Mackie and J.G. Bell
(Editors), Nutrition and Feeding in Fish. Academic Press, London, pp. 299-33 1.
JA Milinaire, C., Gatesoupe, F.J. and Stephan, G., 1983. Approche du besoin quantitative en acides gras longs
polyinsatmebs de la serie n - 3 chez la larve de turbot (Scophrulmus maximus). C. R. Acad. SC. Paris, 29&B): 917-920.
Lubzens, E., Marko, A. and Tietz, A., 1985. De novo synthesis of fatty acids in the rotifer Brclchionus plicafilis. Aquaculture, 47: 27-37.
McEvoy, L.A., Navarro, J.C., Bell, J.G. and Sargent, J.R., 1995. Autoxidation of oil emulsions during the
Artemia enrichment process. Aquaculture, 134: IOI- 112.
Metcalfe, L.D. and Schmitz. A.A., 1961. The rapid preparation of fatty acid ester for gas chromatographic
anlysis. Anal. Chem., 33: 363-364.
Mourente, G., Rodriguez, A., Tocher. D.R. and Sargent, J.R., 1993. Effects of dietary docosahexaenoic acid
(DHA; 22:6n - 3) on lipid and fatty acid compositions and growth in gilthead seabream (Spurus aurufu L.)
larvae during first feeding. Aquaculture, 112: 79-98.
Ramuzzo, J.R., Olsen, Y. and Rosenlund, G., 1989. The effect of enrichment diets on the fatty acid
composition of the rotifer Brachionus plicutilis. Aquaculture, 79: 157- 161.
Rainuzzo, J.R., Reitan, K.I. and Olsen, Y., 1994. Effect of short- and long-term lipid enrichment on total
lipids, lipid class and fatty acid composition in rotifers. Aquacult. hit., 2: 19-32.
Reitan, K.I., Rainuzzo, J.R. and Olsen, Y., 1994. Influence of lipid composition of live feed on growth,
survival and pigmentation of turbot larvae. Aquacult. Int., 2: 33-48.
Rodriguez, C., 1994. Estudio de 10s requerimientos de dcidos grasos esenciales de la dorada europea (Sparus uurufa, L.) durante las dos primeras semanas de alimentacibn. PhD Thesis, University of La Laguna,
Spain, 284 pp.
Rodriguez, C., Perez, J.A., Izquierdo, M.S., Mora, J., Lorenzo, A. and Femattdez-Palacios, H., 1993. Essential
fatty acid requirements of larval gilthead sea bream, Sparus aurafu CL.). Aquacult. Fish. Manage., 24:
295-304.
Rodriguez, C., Perez, J.A., Lorenzo, A., Izquierdo, M.S. and Cejas, J.R., 1994. n - 3 HUFA requirement of
larval gilthead seabream Sparus uwafu when using high levels of eicosapentaenoic acid. Comp. B&hem.
Physiol., 107A: 693-698.
Sokal, R.R. and Rohlf, F.J., 1981. Biometry. W.H. Freeman, New York, 859 pp.
Sorgeloos, P., Lavens, P., Leger, P., Tackaert, W. and Vetsichele, D., 1986. Manual for the culture and use of
brine shrimp Artemicr in aquaculture. State University of Ghent, Belgium, 319 pp.
Southgate, P.C. and Lou, D.C., 1995. Improving the n-3 HUFA composition of Artemin using microcap-
sules containing marine oils. Aquaculture, 134: 91-99.
Stewart, W.P., 1974. Algal physiology and biochemistry. In: W.P. Stewart (Editor), Botanical Monographs,
Vol. 10. Blackwell Scientific, Oxford, 989 pp.
Walford, J. and Lam, T.J., 1987. Effect of feeding with microcapsules on the content of essential fatty acids in
the live foods for the larvae of marine fishes. Aquaculture, 61: 219-230.
Watanabe, T., 1993. Importance of docosahexaenoic acid in marine larval fish. J. World Aquacult. Sot.. 24:
152- 161.
Watanabe, T., Ohta, M., Kitajima, C. and Fuji& S., 1982. Improvement of dietary value of brine shrimp
Anemia salina for fish larvae by feeding them on w3 highly unsaturated fatty acids. Bull. Jpn. Sot. Sci. Fish., 48: 1775-1782.
Watanabe, T., Kitajima, C. and Fujita, S., l983a. Nutritional values of live organisms used in Japan for mass
propagation of fish: A review. Aquaculture, 34: 1 l.5- 143.
Watanabe, T., Tamiya, T.. Oka, A., Hirata, M., Kitajima, C. and Fuji& S., 1983b. Improvement of dietary
value of live foods for fish larvae by feeding them on w3 highly unsaturated fatty acids and fat-soluble vitamins. Bull. Jpn. Sot. Sci. Fish., 49: 471-479.
Watanabe, T., Izquierdo. M., Takeuchi, T., Satoh, S. and Kitajima, C., 1989. Comparison between eicosa-
pentaenoic and docosahexaenoic acids in terms of essential fatty acid efficacy in larval red seabream.
Nippon Suisan Gakkaishi, 55: l635- 1640.
Zar, J.H., 1984. Biostatistical analysis. Prentice-Hall, Englewood Cliffs, NJ, 718 pp.




















