
On how high performers keep cool brains in situations of cognitive overload

Journal of Chemical Neuroanatomy 21 (2001) 75–93
Immunodetection of Parkin protein in vertebrate and invertebratebrains: a comparative study using specific antibodies
Judith M. Horowitz a,b, Vita A. Vernace b, Jason Myers b,c, Michal K. Stachowiak b,c,David W. Hanlon d, Gregory S. Fraley e, German Torres b,*
a Medaille College, Social Sciences, Buffalo, NY 14214, USAb Molecular and Structural Neurobiology and Gene Therapy Program, State Uni6ersity of New York at Buffalo, Buffalo, NY 14214, USA
c Department of Anatomy and Cell Biology, State Uni6ersity of New York at Buffalo, Buffalo, NY 14214, USAd Oncogene Research Products, Cambridge, MA 02142, USA
e Program in Neuroscience and Department of Veterinary and Comparati6e Anatomy, Pharmacology and Physiology,Washington State Uni6ersity, Pullman, WA 99164, USA
Received 8 August 2000; received in revised form 2 October 2000
Abstract
Parkin is an intracellular protein that plays a significant role in the etiopathogenesis of autosomal recessive juvenileparkinsonism. Using immunoblot methods, we found Parkin isoforms varying from 54 to 58 kDa in rat, mouse, bird, frog andfruit-fly brains. Immunocytochemical studies carried out in rats, mice and birds demonstrated multiple cell types bearing thephenotype for Parkin throughout telencephalic, diencephalic, mesencephalic and metencephalic brain structures. While in someinstances Parkin-containing neurons tended to be grouped into clusters, the majority of these labeled nerve cells were widelyscattered throughout the neuraxis. The topographical distribution and organizational pattern of Parkin within major functionalbrain circuits was comparable in both rats and mice. However, the subcellular localization of Parkin was found to varysignificantly as a function of antibody reactivity. A consistent cytoplasmic labeling for Parkin was observed in rodent tissueincubated with a polyclonal antibody raised against the human Parkin protein and having an identical amino-acid sequence withthat of the rat. In contrast, rodent tissue alternately incubated with a polyclonal antibody raised against a different region of thesame human Parkin protein but having 10 mismatched amino-acid sequence changes with those of the rat and mouse, resultedin nuclear labeling for Parkin in rat but not mouse neurons. This difference in epitope recognition, however, was reversed whenmouse brain tissue was heated at 80°C, apparently unmasking target epitopes against which the antisera were directed.Collectively, these results show a high degree of conservation in the cellular identity of Parkin in animals as different asdrosophilids and mammals and points to the possibility that the biochemical specificities of Parkin, including analogous functionalroles, may have been conserved during the course of evolution. © 2001 Elsevier Science B.V. All rights reserved.
Keywords: Aldehyde fixatives; Evolution; Antigen retrieval; Drosophila ; Dopamine; Parkinson’s disease
www.elsevier.com/locate/jchemneu
1. Introduction
Parkinson’s disease (PD) is a neurodegenerative dis-order characterized by akinesia, tremor and distur-bances of gait and posture. PD brain pathology istypified by the degeneration of the pars compacta of the
substantia nigra (SNc), and to a lesser extent, the locuscoeruleus. SNc neurons synthesize the neurotransmitterdopamine (DA) and a consequence of the selective lossof these mesencephalic neurons is a decrease of DAcontent at innervating forebrain sites, such as the cau-date nucleus and putamen. These brain areas, alongwith neighboring neural circuits are an integral part ofthe basal ganglia, a group of forebrain nuclei facilitat-ing or inhibiting cortical-based motor functions. Thecause of DA cell death in PD is unknown, but anumber of factors such as mutations and exposure toenvironmental toxins have been implicated in the etiol-
* Corresponding author. Present address: Behavioral NeuroscienceProgram, Department of Psychology, State University of New Yorkat Buffalo (SUNY), Park Hall Box 604110, Buffalo, NY 14260, USA.Tel.: +1-716-6453650, ext.: 678; fax: +1-716-6453801.
E-mail address: [email protected] (G. Torres).
0891-0618/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved.PII: S 0 8 9 1 -0618 (00 )00111 -3

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9376
ogy of this brain disease. For example, the identifica-tion of parkin as a gene for autosomal recessive juvenileparkinsonism (AR-JP) and the demonstration that mu-tations within its protein-product leads to (i) degenera-tion of DA neurons, and (ii) deficits in motor functionsupport the hypothesis that some forms of PD can beattributed to deletional or point mutations within spe-cific gene-coding regions. Studies of AR-JP patientsshow that deletions of either exon 4 or exons 3–7 of theparkin gene lead to a defective cellular phenotype thatmay constitute a loss of protein function (Hattori et al.,1998a; Kitada et al., 1998). The gene locus of parkin islocalized to chromosome 6q25.2-q27 as demonstratedby linkage analysis and encodes a novel protein of 465amino-acids (Parkin; predicted molecular weight(MW)=51 652 Da) containing a RING (zinc) finger-like motif at the C-terminal portion of the protein; amotif often seen in proteins involved in gene expression(Hattori et al., 1998b; Kitada et al., 1998). Althoughthe functional significance of Parkin is unknown, it isthought that loss or partial loss of Parkin could lead toaberrations in proteolytic pathways involved in thedegradation of short-lived regulatory proteins, such asCyclins and Synucleins (Leroy et al., 1998; Horowitz etal., 1999; Wang et al., 1999). It is conceivable then, thataccumulation or aggregation of such proteins may pro-duce a metabolic instability within SNc neurons therebyaffecting the fidelity of basal ganglia DA neuro-transmission.
Expression of the native state of Parkin in humanbrain is considerable. Northern blot analysis, using poly(A)+ RNA with exon 7 as a molecular probe, shows a4.5-kb transcript expressed in the cerebral cortex, cau-date nucleus, putamen, thalamus, SN and cerebellum(Kitada et al., 1998). Further, immunocytochemicalanalysis of the human brain shows discrete Parkin-im-munoreactivity (IR) in cytoplasm and neuronal pro-cesses (Kitada et al., 1998). Subcellular fractionationstudies of cortical homogenates are congruent with thelatter findings as Parkin is found in both the cytoplasmand golgi complex (Shimura et al., 1999). Parkinprotein (predicted MW=�50 kDa) is also present inextracts of rat cortex, hypothalamus, SN and cerebel-lum (Horowitz et al., 1999). Immunocytochemical stud-ies using affinity-purified polyclonal antibodies againsthuman Parkin are compatible with this suggestion asstrong Parkin-IR is detected throughout the rat brainparenchyma, particularly in DA-based circuits. Assuch, it is conceivable that Parkin plays a critical role inshaping the metabolic stability of proteins in rodentnerve cells as well. The fact that human Parkin peptide-directed antisera cross-react with rat Parkin proteinindicates striking sequence homology and apparent fea-tures of similar subcellular distribution. However, al-though Horowitz et al. (1999) show Parkin localized tothe cytoplasm using a specific polyclonal antibody from
Chemicon (AB5112), subsequent preliminary studiesusing a different polyclonal antibody from Oncogene(Ab-1) show Parkin localization to be associated withthe nuclear envelope. This discrepancy must be resolvedif we are going to use rodents for gaining insightsrelevant to the function and regulation of humanParkin under normal and pathophysiological condi-tions. Thus, we have used the aforementioned antibod-ies to determine the precise subcellular localization ofParkin to rat and mouse brains. One of the polyclonalantibodies used was a 19-amino-acid peptide obtainedfrom Chemicon with the following 1 letter code se-quence: RILGEEQYNRYQQYGAEEC; it correspondsto amino-acid sequence 305–323 of the human Parkinmolecule. The other antibody used was an 18-amino-acid peptide obtained from Oncogene with the follow-ing 1 letter code sequence: NATGGDDPRNAA-GGCERE; it corresponds to amino-acid sequence 81–98 of the human Parkin molecule. In addition, we haveexamined the regional distribution of Parkin-IRthroughout rat (Rattus nor6egicus) and mouse (Musmusculus) brains thus extending our earlierimmunocytochemical observations of Parkinlocalization to rat cortex, SN and cerebellum (Horowitzet al., 1999). Finally, in order to appreciate theextent of conservation of Parkin across different spe-cies, we have examined the presence of human proteinisoforms in avian (Taeniopygia guttata), frog (Xenopuslae6is) and fruit-fly (Drosophila melanogaster)brains.
2. Methods
2.1. Animals
Wild-type male and female Drosophila (of the Can-ton-S strain; Carolina Biological, Burlington, NC; n=80/sex) were raised on standard medium with dry yeastadded. All experiments were conducted on 4–8 day-oldflies during the light phase of the light:dark cycle. Adultmale and female frogs (n=3) were kept in groups offive at room temperature (20°C) in dechlorinated waterwith free access to calf liver and amphibian sticks. Maleand female zebra finches (n=4/sex) approximately 5months of age were caged in groups of five and main-tained ad libitum with bird seed and water. Adult maleand female C57BL/6 mice (Harlan Sprague-Dawley,Indianapolis, IN; n=7/sex) approximately four–sixmonths of age were housed in groups of four and raisedon standard laboratory murine chow. Adult (3 monthsold) male and female Long-Evans rats (Harlan SpragueDawley, Indianapolis, IN; n=7/sex) were housed ingroups of three, with free access to water and standardlaboratory rat chow. These animals were maintained ona 12:12 light:dark cycle with ambient temperature of

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 77
23–24°C and 60% humidity. Male and female verte-brate and invertebrate animals were used in all experi-ments as we failed to detect gross sex-dependentdifferences in Parkin-IR throughout respective brainmaterial. All testing procedures were carried out inaccordance with the NIH Guide for the Care and Useof Laboratory Animals, and with approval from theUniversity at Buffalo and Washington State UniversityIACUCs.
2.2. Western blotting
Decapitation of Drosophila was accomplished as de-scribed previously (Torres and Horowitz, 1998). Briefly,fruit-flies were anesthetized at −20°C for approxi-mately 2 min and with a sharp steel razor blade, aprecise cut was made between the subesophageal sec-tion of the head and the prothoracic neuromere. Flyheads were immediately placed in conical microtubesthat had been frozen on dry ice. Tissue samples werethen stored at −80°C until homogenization. Frogswere also anesthetized by deep body hypothermia anddecapitated with a standard guillotene. Brains werequickly removed from the calvaria and immediatelystored in conical microtubes at −80°C. Birds androdents were decapitated under CO2 anesthesia andbrains rapidly removed from skulls. Neural tissue wasgrossly dissected (on ice) into cortex (or its equivalent,the archistriatum, in the avian brain), hypothalamus,SN (or its equivalant, nucleus tractus pedunculo-ponti-nus, par compacta [TPc], in the avian brain) and cere-bellum. Immediately after dissection, brain sampleswere placed into conical microtubes and stored at −80°C until homogenization. Prior to Western blotting,brain protein content was analyzed using the experi-mental protocol provided by Bio-Rad protein assays(Bio-Rad Laboratories, Hercules, CA). Bovine serumalbumin was used as a standard (1 mg/ml) and tissuesamples were analyzed by spectrophotometry at 595nm.
To detect Parkin protein by immunoblotting, braintissue samples were separately homogenized in low saltlysing buffer (20 mM Tris-HCl, 150 nM NaCl, 1 mMMgCl, 5 mM EDTA, 2 mM EGTA, 1% NP40 and0.5% deoxycholate) with protease inhibitors (10 mMleupeptin, 1 mM PMSF, 1 mM aprotinin and 5 mg/mlpepstatin A) included in the buffer solution. The ex-tracts were centrifuged at 10 000×g for 10 min and thesupernatant aliquoted and measured for protein con-tent. Aliquots of appropriate brain tissue (100 mgprotein) were mixed with equal amounts of loadingbuffer containing 200 mM Tris, 8% SDS, 0.4% bro-mophenol blue, 40% glycerol and 5% 2-mercap-toethanol. Samples were heated at 95°C for 10 min andthen immediately loaded on 8% SDS-polyacrylamidegels, electrophoresed and transferred to nitrocellulose
membranes. Nitrocellulose blots were probed using rab-bit polyclonal antisera raised against opposite ends ofthe human Parkin protein (Chemicon, Temecula, CA;Oncogene, Cambridge, MA) diluted to a concentrationof 1 mg/ml. Parkin was detected using goat anti-rabbitsecondary antibodies (Pierce, IL) diluted to a concen-tration of 0.02 mg/ml, and immunopositive bands visual-ized by chemiluminescence. Prestained standards forblotting analysis consisted of known proteins withMWs ranging from approximately 200 to 35 kDa (Bio-Rad Laboratories, Hercules, CA).
2.3. Reprobing nitrocellulose membranes for Westernblotting
After visualization of Parkin-IR with chemilumines-cence methods, each nitrocellulose membrane waswashed 4×5 min in tris-buffer solution (TBS) andincubated for 30 min at 50°C in a stripping bufferreagent consisting of 62.5 mM tris-HCl (pH=6.8), 2%SDS and 100 mM 2-mercaptoethanol (as referenced byNEN Life Science, Boston, MA). After incubation withthe stripping buffer solution, nitrocellulose membraneswere washed again 6×5 min in TBS and reprobedalternately with either AB5112 or Ab-1 antibodies de-pending on the original probing conditions.
2.4. Brain tissue fractionation for Western blotting
Fresh rat and mouse SN was homogenized separatelyby mortar and pestle in a fractionation solution con-taining 10 mM Hepes buffer (pH 7.9), 10 mM KCl, 0.1mM EDTA, 0.1 mM EGTA, 1 mM DTT and 0.5 mMPMSF. Immediately after homogenization, rodentbrain samples were allowed to sit on ice for 10 min.Tissue was then aliquoted into 1 ml conical microtubescontaining 0.6% NP40, centrifuged at 4°C for 1 min(600×g) and the cytoplasmic material aliquoted andexamined under a microscope (10× ) for the absence ofnuclear material. Samples containing pure cytoplasmwere then stored at −80°C until blotting. The remain-ing nuclear pellet was washed 3×2 min with 0.35%lysing buffer to remove any remaining cytoplasmicmaterial (Stachowiak et al., 1996). High salt buffer (0.4M) was added to the nuclear pellet which was thensonicated for 1 min. Aliquots (15 ml) were then storedat −80°C for subsequent Western blot analysis.
2.5. Immunocytochemistry
Rats and mice used for the immunocytochemicalexperiments were deeply anesthetized with a ketaminemixture (150 ml/100 g body weight) consisting of ke-tamine (100 mg/ml solution), acepromazine (20 mg/mlsolution) and xylazine (20 mg/ml solution). Animalswere then perfused intracardially first with heparinized

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9378
saline followed by 4% paraformaldehyde in Na2PO4
buffer solution (pH 7.2). Zebra finches were deeplyanesthetized with equithesin (0.05 ml/bird; intramus-cular). Birds were then perfused intracardially with 5ml of 0.1 M phosphate buffer, followed by no morethan 40 ml of 4% paraformaldehyde. Avian brainswere excised from the calvaria and postfixedovernight at 4°C in the above fixative. Brains werethen hardened in 10% sucrose overnight and embed-ded in 8% gelatin. Gel-blocks were allowed to set,first at room temperature for 1 h and then at 4°C for3 h. After this time, gel-blocks were immersed for 48h in 4% paraformaldehyde containing 10% sucrose.Finally, gel-blocks were hardened overnight at 4°C in20% sucrose and then in 25% sucrose the followingday.
The immunocytochemical methods have been de-scribed in detail previously and are only briefly sum-marized below (Torres et al., 1992). Respective rodentbrains were fixed in the same 4% formaldehyde solu-tion for 5 days at 4°C and then stored in 20% su-crose (dissolved in 0.01 M sodium phosphate buffer)overnight. Frozen vertebrate brains were mounted ona sliding microtome and cut into 40 mm coronal sec-tions from rostral striata to cerebella. Cut brain sec-tions were collected in a cold cryoprotectant solution(0.05 M sodium phosphate buffer pH 7.3, 30% ethyl-ene glycol, and 20% glycerol) and stored at −20°Cuntil prepared for standard immunocytochemistryprocedures. Free-floating brain sections were incu-bated for 48 h at 4°C in rabbit polyclonal IgGs(AB5112 and Ab-1) raised against the human Parkinprotein. The antibodies were diluted 1:1000 in potas-sium-phosphate buffer solution (KPBS) with 1% nor-mal goat serum and 0.3% Triton X-100. Next, brainsections were rinsed in KPBS and incubated with sec-ondary biotinylated goat anti-rabbit IgGs (1:1500 di-lution; Vector Laboratories, Burlingame, CA) for 60min. Subsequently, brain sections were incubated atroom temperature with an Avidin–Biotin–PeroxidaseComplex (Vectastain ABC Elite Kit; Vector Labora-tories) for 60 min. After several rinses in KPBS andacetate-imidazole buffer, brain sections were devel-oped in a mixture containing tris-imidazole buffer,Nickel(II) Sulfate·6H2O and the chromagen 3,3’-di-aminobenzidine tetrahydrochloride (DAB) along with1% H2O2. Following the developing phase, brain sec-tions were again washed in KPBS, mounted ontogelatin–chrome–alum-coated slides, allowed to dryovernight, dehydrated through graded concentrationsof ethanol, cleared in xylenes, and coverslipped withDPX mountant (Electron Microscopy Sciences, Ft.Washington, PA). Specificity for both antibodies wasinstituted in the form of positive controls (i.e. pre-in-cubation of the primary antibody with the immuniz-ing Parkin peptide) and negative controls (i.e.
replacing the primary antibody with normal rabbitserum).
For the simultaneous detection of Parkin proteinusing AB5112 and Ab-1 antibodies, double labelingimmunocytochemistry was carried out as previouslydescribed (Torres et al., 1993). In brief, immediatelyafter the labeling of neuronal populations with theAB5112 antibody by peroxidase activity, rat andmouse brain sections were rinsed in KPBS and thenincubated with the Ab-1 antibody for 48 h at 4°C asdescribed above. After incubation with primary andsecondary antibodies, rodent brain tissues were incu-bated with the Avidin–Biotin–Peroxidase Complex,also as described above. Thereafter, brain sectionswere washed with KPBS and developed with DAB.Next, coronal sections were rinsed in KPBS, mountedonto subbed slides, dehydrated through gradedethanols, cleared with xylenes and coverslipped withDPX mountant. Under these experimental conditions,the presence of cytoplasmic Parkin-IR was evident asa brown reaction product, whereas nuclear Parkin-IRwas visualized as a crisp, blue/black reaction productthroughout the rodent brain. Finally, to help identifyrelevant anatomical structures of interest, as well asto define topographical regions of rat and mousebrain, some tissue sections were counterstained withNeutral Red. Visualization of Parkin immunolabelingthroughout functional brain circuits was accomplishedwith bright-fiel microscopy using an Olympus micro-scope equipped with a (10 mm2) grid reticule at mag-nifications of 10× , 20× , and 60× . Relativeintensities of Parkin-IR were arbitrarily defined ashaving either a strong, medium, weak, or very weakcytoplasmic- or nuclear-IR. For these types of experi-ments, Parkin immunolabeling was recognized underbright-field microscopy and its topographical distribu-tion, organizational pattern and cytoarchitectoniccomposition was characterized using rodent brain at-lases.
2.6. Antigen retrie6al by tissue heating in sodiumcitrate buffer
To unmask native Parkin antigens in brain tissue, asubset of rat, mouse, and avian brains were washedfirst with KPBS and then incubated in a sodium cit-rate buffer solution (10 mM; pH 9.0; Sigma Chemi-cals, St. Louis, MO) for 30 min in a water-bath setto 80°C (Jiao et al., 1999). The aforementionedsodium citrate buffer solution containing brain sec-tions was then allowed to cool for 30 min to roomtemperature. Thereafter, brain sections were washedin a KPBS solution containing 1% Carnation drymilk. Brain sections were then processed for detectionof Parkin using standard avidin–biotin immunoperox-idase protocols.

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 79
2.7. Procedures for postmortem, unfixed rodent brains
A subset of rats and mice were killed by CO2 expo-sure and then placed at room temperature for 6 h aspreviously described (Torres et al., 1992). After this 6 hpostmortem delay, rat and mouse brains were removedfrom the calvaria and immediately placed in 4% formal-dehyde phosphate buffer (pH 7.2) for 5 days. There-after, these brains were placed in 20% sucrose in 0.1 Msodium phosphate buffer for 2 days or until the brainssank. Frozen coronal brain sections were cut on asliding microtome and cryoprotected until processed forlocalization of native Parkin antigens to brain cells.Prior to incubation with either AB5112 or Ab-1 anti-bodies, selected free-floating tissue sections from post-mortem brains were briefly treated (10 min) withsodium-m-periodate (1.07 g in 50 ml tris-buffer saline)to inactivate endogenous peroxidases associated withred blood cells in unperfused tissue. Brain sections werethen washed in KPBS and processed for immunocyto-chemistry as described above.
2.8. Statistical analysis
Data were expressed as mean9SEM of MWs fromimmunopositive Western blots. Adjusted volume opti-cal densities were performed by scanning Western blotfilms with the aid of a Bio-Rad densitometer scannerand a statistical computer program, which measuredthe area (mm2) and volume (optical densities×mm2) ofParkin-specific immunopositive bands. When appropri-ate, ANOVAs were performed followed by Scheffe’spost hoc tests for mean comparisons between groups.Statistically significant differences were defined as P00.05.
3. Results
3.1. Immunoblot analyses of Parkin protein in6ertebrate and in6ertebrate brains
Tissue extracts from rat, mouse, avian, frog andfruit-fly brains were subjected to Western blot analysisusing affinity purified antibodies generated against syn-thetic peptides corresponding to amino-acids 305–323(AB5112) and 81–98 (Ab-1) of the human Parkinprotein. Several immunopositive bands were identifiedwhen incubated with the AB5112 antibody, with majorbands thought to be specific for Parkin in the MWrange of 54–58 kDa in all animals tested (Fig. 1).Similar bands were also detected with the Ab-1 anti-body, suggesting therefore that both antibodies recog-nize a similar protein. Further, the sizes of these bandsare consistent with the predicted MW for human andrat Parkin (Horowitz et al., 1999; Shimura et al., 1999).
In rat, mouse and avian brains Parkin protein wasfound consistently in the cortex, hypothalamus, SN andcerebellum (Figs. 1 and 2). Due to the intensity of thebands and the use of the chemiluminescence methodswith the AB5112 antibody, the majority of labeledbands appeared as a single broad band. The MWs ofthe various bands were roughly equivalent in all brainregions tested (PE0.05), with the 54–58 kDa bandsexhibiting the darkest labeling and thus the greatestrelative abundance. Bands of 75 kDa or higher found invertebrate brains may likely correspond to post-transla-tional modifications of Parkin. Confirmation of thesehypotheses will require additional studies.
Parkin protein was also detected in frog and fruit-flybrain homogenates. However, due to experimental con-straints, immunoblot analysis of Parkin was confinedexclusively to whole brain as opposed to representativebrain regions. Nevertheless, tissue extracts preparedfrom amphibians and insects showed that AB5112 (andAb-1) antibodies recognized several bands with MWsof :55 kDa in both frog and Drosophila. Although theamino-acid sequence of Parkin in frogs and fruit-flies isunknown, identified bands in these animals could repre-sent Parkin proteins. Additional high and low MWbands detected in frog and fruit-fly brains are thoughtto be unprocessed and/or glycosylated Parkin isoforms,respectively. While there was some variability in thegels from animal to animal, the 55 kDa bands found infrog and fruit-fly brain were consistently detected in
Fig. 1. Western blotting of Parkin in SN and whole brains fromdifferent vertebrate and invertebrate organisms. A rabbit polyclonalantibody (AB5112; 1 mg/ml) raised against a synthetic peptide corre-sponding to amino-acids 305–323 from the human protein sequencerecognized several immunopositive bands in all brain extracts. Themost salient bands migrated at :54–58 kDa and are roughlyequivalent with the predicted MW of human Parkin (�52 kDa). Thisexperiment was replicated three times with similar results. MWmarkers in kDa are shown on the left.

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9380
Fig. 2. Western blotting of Parkin in brain structures of three vertebrate species. A rabbit polyclonal antibody (AB5112; 1 mg/ml) raised againsta synthetic peptide corresponding to amino-acids 305–323 from the human protein sequence recognized several immunopositive bands in brainextracts. Two prominent bands between 54 and 58 kDa are noted in all brain structures tested. Additional prominent bands are also identifiedin avian brain extracts with apparent MWs of 40 and 46 kDa. This experiment was replicated three times with similar results. MWs in kDa areshown on the left.
three different blots with similar results. It should benoted that in all immunoblotting procedures describedhere, Western blot membranes were stripped and thenreprobed with the alternate polyclonal antibody, thusassuring equal loading and equal transfer of the Parkinprotein. In addition, pre-incubation of the primaryantibody with the immunizing Parkin peptide (i.e., pos-itive control), or replacing the primary antibody withnormal rabbit serum (i.e., negative control), completelyabolished cellular-IR (data not shown). These experi-mental steps suggest apparent specificity for bothParkin antibodies.
3.2. Immunoblot analyses of Parkin protein in wholebrain homogenates and subcellular fractions
To examine whether Parkin is present in the cyto-plasm, nucleus or both cellular compartments, fraction-ation of rat and mouse SN neurons into theirsubcellular components was undertaken. As shown inFig. 3, isolation of rat cytoplasm yielded MW bands ofParkin in the range of 56–63 kDa when incubated withthe AB5112 antibody. Similar bands were also iden-tified in mouse cytoplasmic material, thus indicating
that the AB5112 antibody recognizes two bands in thecytoplasm with MWs of approximately 54 and 63 kDa.The sizes of these bands are consistent with predictedMWs for whole brain extracts (Fig. 3). Nuclei from ratSN showed a single band of �56 kDa when incubatedwith the AB5112 antibody, whereas isolated nucleifrom mouse SN exhibited a strong doublet band of�60 kDa. The ratio of relative Parkin abundance (asadjusted volume optical densities; OD/mm2) associatedwith the cytoplasm was approximately the same as therelative Parkin abundance associated with the nucleususing the AB5112 antibody. For example, the ratio ofnuclear:cytoplasmic Parkin in rat was 0.5, and in mousethe ratio was 0.44. This difference in ODs is too small(PE0.05) to indicate differences in protein amountsbetween subcellular compartments.
It has been reported that the Ab-1 antibody recog-nizes Parkin in nuclei of human neurons after immuno-cytochemical analysis (Charleston et al., 1999). Here,we examined the location of Parkin protein to bothcytoplasmic and nuclear compartments within rat andmouse SN cells. Bands of approximately 54 and 57 kDawere identified in the cytoplasm of rat SN; additionalbands of higher (e.g. 79 kDa) and lower (e.g. 35 kDa)

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 81
MWs were also identified. Similarly in mouse SN, theAb-1 antibody used in the Westerns recognized cyto-plasmic bands of approximately 55 and 56 kDa withadditional lower MW bands in the range of 33 and 35kDa. Isolated nuclei from rat SN exhibited a faint bandof �56 kDa, whereas immunoblot analysis of isolatednuclear preparations from mouse SN yielded bands of55 and 56 kDa (Fig. 3). The sizes of these bands areconsistent with predicted MWs for rodent Parkinprotein. The ratio of relative Parkin abundance (asadjusted volume optical densities; OD/mm2) associatedwith the cytoplasm was approximately the same as therelative Parkin abundance associated with the nucleususing the Ab-1 antibody. For example, the ratio ofnuclear:cytoplasmic Parkin in rat was 1.0, and in mousethe ratio was 0.7. This difference in ODs is too small(PE0.05) to indicate differences in protein amountsbetween subcellular compartments. Taken together,these data indicate that both AB5112 and Ab-1 anti-bodies identify the presence of Parkin protein in bothcytoplasmic and nuclear material of mammalian SNneurons.
3.3. Immunocytochemical analysis of Parkin protein in6ertebrate brains
AB5112 and Ab-1 antibodies were used to identifyclusters of Parkin-containing neurons. Under normalexperimental conditions (i.e., no surgery or pharmaco-logical manipulations; only daily handling), rats andmice exhibited strong Parkin-IR throughout the brainparenchyma. When brain material was incubated withthe AB5112 antibody, the near absence of IR in thenuclei indicated that Parkin’s distribution was primarilycytoplasmic. This is consistent with the reported iden-tity of Parkin as a cytoplasmic protein in both humanand rat brains (Horowitz et al., 1999; Shimura et al.,1999). Examination of cells positive for Parkin showeda punctate pattern of cytoplasmic IR that was particu-larly noticeable at high magnification. Microscopicanalysis also revealed Parkin-IR to be localized withinfine caliber fibers and dendritic processes. Although theintensity of IR varied from cell to cell, a noticablepattern of IR for Parkin was found in several telen-cephalic, diencephalic, mesencephalic and meten-
Fig. 3. Comparison of Parkin immunopositive subcellular isoforms as recognized by AB5112 and Ab-1 antibodies. Both of the aforementionedantibodies identified Parkin fragments within cytoplasmic and nuclear material isolated from rat and mouse SN. These immunopositive bands arecongruent with bands expressed in whole tissue extracts. Both AB5112 and Ab-1 antibodies recognized prominent Parkin subcellular isoforms withapparent MWs of 54 and 56 kDa. The AB5112 antibody invariably yielded relatively more prominent bands in cytoplasmic and nuclearcompartments than comparable bands obtained from the Ab-1 antibody. Additional immunopositive bands of high and low MWs are alsoidentified in both subcellular compartments. MWs in kDa are shown on the left.

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9382
Table 1Anatomical distribution and relative density of Parkin-IR in rodentbraina
Ab-1dBrain region examinedb AB5112c
Medial forebrain bundle ++++Cingulate cortex, area 1 +++++++
++++++++Frontal cortex 1Parietal cortex, area 1 ++++ ++++
+++++++Piriform cortex++++Accumbens nucleus, core +++++++Caudate putamen
++++Olfactory tubercle ++++++++++++Lateral septal nucleus, intermediate
+++Lateral septal nucleus, dorsal ++++Field CA1, 3 of Ammon’s horn ++ ++++
++++Zona incerta+++Central amygdaloid nucleus +++++++Paraventricular hypothalamic nucleus
++Dorsomedial hypothalamic nucleus ++++++++++Ventromedial hypothalamus
++Arcuate hypothalamic nucleus ++++++++Mediodorsal thalamic nucleus ++++++++++Lateral habenular nucleus, medial++++Reticular thalamic nucleus ++++++++++Paraventricular thalamic nucleus
++Tuberomammillary nucleus +++++++++Premammillary nucleus, dorsal
++Submammillothalamic nucleus ++++Precommissural nucleus +++++
+++++++Substantia nigra (reticular/compact)++Hilus dentate gyrus +++
+++Entorhinal cortex ++++++++Dorsal raphe nucleus
++Central gray, medial ++++++Medial parabrachial nucleus +++++Primary fissure
a Relative abundance of Parkin-IR in rat and mouse brain. Arbi-trary scale: ++++ = strong density, +++ = medium density,++ = weak density, + = very weak density.
b Nomenclature derived from rodent brain atlases (Sidman et al.,1971; Paxinos and Watson, 1986).
c Affinity purified polyclonal antibody corresponding to amino-acids 305–323 of the human Parkin protein. Parkin-IR is entirelycytoplasmic.
d Affinity purified polyclonal antibody corresponding to amino-acids 81–98 of the human Parkin protein. Parkin-IR is entirelynuclear. The distribution of Parkin-IR is not an exhaustive survey ofall cells bearing the signal for the protein.
laterally displaced medium-sized neurons of the caudateputamen. A dense aggregation of small, oval cells hav-ing a similar cytoplasmic-IR profile for Parkin was alsoobserved in the paraventricular nucleus of the hypotha-lamus, paraventricular thalamic nucleus and lateralhabenular nucleus. In these latter regions, the cellularaggregations were significantly more abundant in com-parable areas of the mouse brain than in those of therat. The arcuate nucleus, median eminence and interpe-duncular nucleus were also recipients of strong Parkin-IR as the number and density of Parkin-neurons wasconsiderable. In laminar structures such as the cortexand hippocampus, numerous Parkin-containing cellswere observed in both rats and mice. At this level,small- and medium-sized neurons were located mainlylateral to limbic areas of the frontal cortex, and ventralto the polymorph layer of the dentate gyrus. Over thenext 200–300 mm in the caudal direction of rat andmouse brains, the SN, ventral tegmental area, centralgray area and dorsal raphe nucleus contained numerousParkin-positive cells (Fig. 4B). Although the transmitterphenotype of Parkin-positive cells in the SN andraphe nucleus has not been completely determined,based on our previous findings (Horowitz et al.,1999) at least some of these neurons appear to corre-spond to DA- and serotonin-containing cells. Morecaudally, Parkin-IR was observed in the pontine nucleiand cerebellum of both rat and mouse with findings oflarge, presumably Purkinje, cells bearing Parkin in cy-toplasm and dendrites. Collectively, these results sup-port a distribution of Parkin-IR in rodent brain that isconsiderable, and that the observed location of theprotein to neurons appears to be cytoplasmic ratherthan nuclear, in experiments using the AB5112 anti-body.
Comparison of adjacent rodent brain sections, pro-cessed for Parkin using the Ab-1 antibody, revealedthat the vast majority of rat cells exhibited a nuclearlabeling, whereas those of the mouse retained a strongcytoplasmic-IR feature (Fig. 5A–C, Fig. 6A–B). In ratbrain, numerous cells were seen to be nuclear-positivefor Parkin and were inhomogeneously distributed eitherin densely packed clusters or isolated throughout thebrain parenchyma. Further, using the Ab-1 antibody,Parkin-IR appeared to be more widespread and encap-sulated a greater number of brain regions than thoseobtained from tissue samples incubated with theAB5112 antibody. The distribution of nuclear Parkin inrat brain not previously described included the medialforebrain bundle, nucleus accumbens, cingulate cortex,tuberomammillary nucleus, ventral medial and dorso-medial hypothalamus, submammillothalamic nucleus,precommissural nucleus, primary fissures of the cerebel-lum and lateroventral periolivary nucleus. The topo-graphical distribution of Parkin-IR in mouse brain as
cephalic subdivisions of the rodent brain. A compari-sion of Parkin-IR was made by arbitrarily definingbrain regions with strong (+ + + + ), medium (+ ++ ), weak (+ + ) or very weak (+ ) cellular labeling.These results are presented in Table 1.
In coronal sections of rat and mouse brain incubatedwith the AB5112 antibody, a distinct aggregation ofdarkly-stained, Parkin-positive cells were observed inthe prefrontal cortex, piriform cortex and lateral septalnucleus (Fig. 4A). These cells tended to be groupedtogether and were easily distinguished from the more

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 83
deduced from the Ab-1 antibody was similar to thatdescribed for brain sections processed with the AB5112antibody. However, unlike the rat brain, mouse brainsamples incubated with the Ab-1 antibody revealed alarge and striking collection of Parkin-containing cellsin the medial habenular nucleus, fasciculus retroflexusand interpeduncular nucleus (Fig. 7A–C). These neu-rons were both medium and small in size and formed adense aggregation of cytoplasmic Parkin-IR whichbranched into fine caliber fibers and dendrites from thesoma. This pattern of Parkin distribution in mousebrain was highly reproducible and similar to the re-ported distribution of Parkin-IR generated from theAB5112 antibody.
3.4. Parkin-IR in relation to AB5112 and Ab-1antibodies: double-labeling immunocytochemistry andthe unmasking of antigens
The spatial distribution of Parkin in rat brain wasalso examined in double-labeling experiments. A num-ber of restricted subpopulations of labeled neuronsappeared to contain both phenotypic markers (i.e. cyto-plasmic- and nuclear-IR). Although not completelyidentical, the topographical distribution of Parkin-con-taining cells in rat brain corresponded well to IR de-duced from both AB5112 and Ab-1 antibodies. Briefly,the cortex, hippocampus, ventral medial hypothalamus,several thalamic nuclei and parabrachial nucleus
Fig. 4. Representative bright-field photomicrographs depicting Parkin-IR in rat brain. Coronal brain sections (40 mm) were incubated with thepolyclonal AB5112 antibody (1 mg/ml) and processed for peroxidase activity as described in the methods section. Note that Parkin-IR is entirelycytoplasmic in (A) frontal cortex, area 2 and (B) SN (reticular). Magnifications 20× .

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9384
Fig. 5.

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 85
showed distinctive complements of colocalized IR forParkin in the cytoplasm and nucleus with no apparentpreferential association to one cell type (Fig. 8A–B). Ingeneral, neurons positively labeled for cytoplasmicParkin (AB5112 antibody) displayed a punctate brownIR, while those labeled for nuclear Parkin (Ab-1 anti-body) exhibited a dark IR reaction around the nuclearenvelope. It should be noted that little or no overlap ofthe two phenotypic markers was observed in sometelencephalic regions of the rat brain, including thenucleus accumbens, caudate nucleus and putamen.Concomitant with the colocalization of cytoplasmicand nuclear Parkin-IR to some rat neurons, an addi-tional feature of this experiment was the observationthat tissue processed with the Ab-1 antibody exhibiteda more extensive and widespread pattern of labelingthan comparable tissue incubated with the AB5112antibody. It should be emphasized that the quality ofParkin-IR in brain material was not apparently alteredby double labeling immunocytochemistry as cellular-IRwas similar to that observed after incubation withindividual AB5112 or Ab-1 antibodies.
It is well known that treatment of brain tissue withaldehyde-containing fixatives can modify the reactivityof proteins to antibodies. To minimize the masking ofantigens by aldehyde fixation, several methods havebeen used to enhance antigen retrieval and thus facili-tate exposure of target epitopes to antisera (Shi et al.,1996; Newman and Gentleman, 1997). We have usedthe method of Jiao et al. (1999), which employs water-bath heating for antigen retrieval, to test the possibilitythat differences in Parkin distribution between rats andmice may be due to masking of native antigens byaldehyde fixation. By this heating method, we foundthat the cellular distribution of Parkin within mousebrain yielded a nuclear pattern of IR after incubationwith the Ab-1 antibody (Fig. 9A–B). It should be notedthat this nuclear distribution was observed within thesame brain regions previously displaying cytoplasmic-IR. In contrast, a cytoplasmic pattern of labeling wasretained when mouse tissue was processed with theAB5112 antibody (data not shown). One consistentdifference, however, was the absence of changes in thecellular distribution of Parkin-IR in rat brain afterantigen retrieval by water-bath heating. Incubation ofrat tissue with both AB5112 and Ab-1 antibodies al-ways produced a cytoplasmic and nuclear labeling,respectively (see Table 2 for summary). Interestingly, asimilar pattern of cellular distribution for Parkin wasalso noted in avian brain previously heated. That is,after antigen retrieval by water-bath heating, Parkin-IR
was observed in the cytoplasm as recognized by theAB5112 antibody, and nuclear-IR after incubation withthe Ab-1 antibody (Fig. 10A–C). The functional conse-quences of such striking differences between species inthe cellular distribution of Parkin remain unclear.
To test for the possibility that pretreatment of rodentbrains with chemical fixatives and detergents mightchange the reactivity of native proteins to antibodies,we obtained brain material from rats and mice that hadbeen left intact (i.e. without aldehyde fixation) at roomtemperature for 6 h after death. This experimentalprocedure, which owing to its capacity to impede ax-onal transport, has been assumed to allow for completeassessment of peptide antigenicity in situ (Torres et al.,1992). In coronal brain sections heated at 80°C andincubated separately with AB5112 and Ab-1 antibodies,Parkin-IR was strongly visible in many cell typesthroughout the neuraxis. Comparison of adjacent sec-tion immunolabeled alternately with the aforemen-tioned antibodies revealed that the integrity ofParkin-IR was well preserved, with no apparent loss ofcytoplasmic or nuclear staining intensity. Pretreatmentof coronal brain sections with sodium-m-periodate sat-isfactorily eliminated most (endogenous) peroxidasesand minimized discernible blood vessels within thebrain parenchyma (Fig. 11A–C). Overall, the distribu-tion of Parkin-containing neurons in rat and mousebrains corresponded well to that reported in rodentstranscardially perfused with aldehyde fixatives. We in-terpret this finding to mean that aldehyde fixationtreatment does not significantly alter the secondary ortertiary structure of Parkin protein(s). Thus, we con-clude that differences in the subcellular localization ofParkin between rats and mice do not depend on themasking of antigens by aldehyde fixation, but rather aredue to differences related to antibody reactivity.
4. Discussion
Aberrations in the native state of human Parkinresult in loss-of-function and thus, a defective behav-ioral phenotype. Indeed, mutations of the aboveprotein have been associated with AR-JP, a brain dis-ease with selective degeneration of DA neurons withoutLewy body formation (Kitada et al., 1998). Given themoderate similarity of Parkin with Ubiquitin, a proteininvolved in intracellular proteolysis, it is hypothesizedthat Parkin may play a major role in the degradation ofshort-lived proteins as well (Shimura et al., 1999, 2000).The fact that we have detected Parkin in both verte-
Fig. 5. Representative bright-field photomicrographs depicting Parkin-IR in rat brain. Coronal brain sections (40 mm) were incubated with thepolyclonal Ab-1 antibody (1 mg/ml) and processed for peroxidase activity as described in the methods section. Note that Parkin-IR is entirelynuclear in (A) piriform cortex (B) hippocampus (CA1, 3 and dentate gyrus), and (C) arcuate nucleus of the hypothalamus. Magnifications 10× .

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9386
Fig. 6. Representative bright-field photomicrographs of mouse coronal brain sections (40 mm) through the (A) frontal cortex, area 2, and (B) SN(reticular). Brain tissue was collected from mice and processed for standard immunocytochemistry with AB5112 (A) and Ab-1 (B) polyclonalParkin antibodies (1 mg/ml). Note that in both instances, the immunolabeling for Parkin generated from AB5112 and Ab-1 antibodies was entirelycytoplasmic. The spatial distribution and organizational pattern of Parkin-IR in mouse brain, however, was similar to that reported for rat brain.Magnifications 20× .
brate and invertebrate brains adds credence to thehypothesis that Parkin may be playing a universal rolein controlling, for example, the metabolic stability ofproteins in many cell types. This finding also points tothe possibility that the biochemical specificities ofParkin, including analogous functional role(s), may
have been preserved during the course of evolution.Clearly, the detection of Parkin fragments by Westernblotting in brains from organisms as diverse as fruit-flies, frogs and birds is congruent with this evolutionarypossibility. Indeed, it is already known that both insectsand vertebrates have many homologous genes that code
Fig. 7. Bright-field photomicrographs of representative mouse coronal brain sections (40 mm) incubated with the Ab-1 antibody for Parkin (1mg/ml). Ab-1-based immunolabeling yielded an extensive filling of neuronal fibers in the (A) lateral habenula nucleus, (B) fasciculus retroflexus and(C) a complete neuronal filling in the interpeduncular nucleus. One consistent difference in these experiments, however, was the absence of thistype of Parkin-IR in rat brain. The presence of extensive filling of neuronal processes in discrete regions of the mouse brain appears to reflect agenuine immunolabeling for Parkin since the same (Ab-1) antibody yielded a similar pattern of cellular-IR in unfixed mice brains. Magnifications10× .

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 87
Fig. 7. (Continued)

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9388
for proteins that are evolutionary related (Clark, 1999;Hirth and Reichert, 1999). With the entire fly genomesequence in hand, it would be possible for instance, toelucidate cellular pathways within which wild-typeparkin gene(s) act. In this regard, Drosophilamelanogaster is an attractive model organism for study-ing pathways and identifying genes that regulate univer-sally important processes; processes which mightotherwise be difficult to elucidate in vertebrates. Forinstance, it has been recently shown that fruit-fliesexpress ariadne-1, a novel and vital Drosophila generequired for neural development (Aguilera et al., 2000).Of interest, the ariadne-1 gene may be the vertebrateParkin homolog or at least, a member of a conservedfamily of genes whose multiple domains include aRING-finger protein product. Comparative genomicsmay therefore enable the transfer of informationrelated to Parkin function and regulation to humanswith important consequences for understanding whymutations within the parkin gene lead to DA cellpathology.
4.1. Western blotting analysis for Parkin-IR
Western blotting experiments showed Parkin residingin many regions of rodent and avian brain. For exam-ple, our experiments identified Parkin isoforms of :54–58 kDa in cortex, hypothalamus, SN andcerebellum. There were additional bands labeled forParkin in the aforementioned brain regions suggestingpre- or post-transcriptional modifications of the Parkinprotein. It should be noted that the MW of frog, bird,mouse and rat Parkin appears to be similar to thepredicted MW (�51 kDa) for human Parkin (Kitadaet al., 1998). Thus, we hypothesize that the identifiedisoforms found in vertebrate brains indeed correspondto Parkin protein(s). We cannot absolutely rule out thepossibility that there may be species differences, northat proteolytic Parkin fragments might have been pro-duced as a result of tissue degradation. However, braintissue from every vertebrate (and invertebrate)organism used here was frozen immediately upon de-capitation and treated with protease inhibitorsduring homogenization. Fractionation of rat andmouse SN cells into their subcellular componentsshowed Parkin to be localized to both cytoplasmicand nuclear microenvironments as demonstrated byWestern methods. This parallel evidence suggeststhat Parkin may be acting at multiple cellular compart-ments to carry out putative regulated proteolyticfunctions. Previous work has suggested that Parkinmay be localized to either cytoplasmic or nuclearmaterial, but not to both cellular compartments(Charleston et al., 1999; Horowitz et al., 1999; Shimuraet al., 1999). This could be attributable to technicalfactors such as changes (e.g. crosslinking) in the sec-
ondary or tertiary structure of proteins following alde-hyde-based fixation (Dapson, 1993). Such crosslinkingappears to mask antigenic epitopes to antisera therebyaltering the specificity of IR to individual subcellularcomponents. This reasoning may not be applicable toWestern blotting as fixation is absent from this methodand sample heating is implemented to render antigenicepitopes accessible to antibodies directed against them.In light of this evidence, it seems reasonable to con-clude that Parkin protein is localized to both cytoplas-mic and nuclear compartments within rat and mouseneurons.
4.2. Neuroanatomical distribution of Parkin-IR
Our results confirm and extend what was previouslyknown about the distribution of Parkin in rat brain(Horowitz et al., 1999). Further, we have providedevidence that mouse brains have a similar regionaldistribution of Parkin, and that avian brains also con-tain identifiable groups of cells labeled positive for theabove protein. These comparative experiments providesubstantial evidence for a conservative structural orga-nization of Parkin in many cell types. Indeed, theexistence of such a similarity suggests that a compara-ble structural organization of Parkin was alreadypresent in the ancestors of amniotes. Thus, indepen-dently of the origin and nature of Parkin evolution,Parkin protein might play a similar, yet fundamental,role in vertebrate brains. In rats and mice, strongParkin-IR was noted throughout the brain parenchymaincluding the cortex, hippocampus, hypothalamus, tha-lamus, SN and cerebellum. The widespread intracellulardistribution of Parkin in rodent brains implies that theregulatory influences of the protein are not exerteddifferentially, nor are they associated with a particularsubset of neurons sharing a given biochemical pheno-type. Instead, it suggests a complex and uniform influ-ence of Parkin in multiple neural circuits, which on thebasis of connectivity and/or functional associationswith additional cell groups, are involved in a myriad ofphysiological and behavioral processes. We also notedthat Parkin-IR was localized to either the cytoplasm ornucleus within rat and mouse neurons, and that such adivergence in spatial localization was dependent on twoindependent criteria: (i) the reactivity of the antibodyand (ii) the antigen retrieval method used. The cellulardistribution pattern for Parkin was mainly cytoplasmicwhen rat tissue was incubated with the AB5112 anti-body. These results validate our previous findings(Horowitz et al., 1999) and those of Shimura et al.(1999) indicating Parkin antibodies mainly recognizinga cytoplasmic protein within rat and human brains.However, our current findings are incongruent withthose generated from Charleston et al. (1999) as theydetect Parkin protein in nuclear material. It might be

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 89
argued that differences between the studies are simplydue to antibody cross-reactivity variables. This, however,seems less likely because the sequence of the immunizingpeptide used by Chemicon to generate the antibody isidentical in rat (1 letter code sequence: RILGEEQYN-RYQQYGAEEC) and human (1 letter code sequence:RILGEEQYNRYQQYGAEEC). Thus, minimal differ-ences in antibody reactivity between the two mammalianspecies should be expected. Further, the specificity for the
Parkin AB5112 antibody was confirmed by immunoblotmethods (see above) thus precluding the possibility thatthe Parkin antibody was binding to epitopes not foundin the original peptide sequence. Parallel evidence alsoshows considerable amino-acid sequence identity be-tween rat and human Parkin (D’Agata et al., 2000; Guet al., 2000). Such a comparison at the amino-acid levelsuggests that the AB5112 antibody recognizes a Parkinprotein in closely related species.
Fig. 8. Colocalization of Parkin-IR in rat brain. Selected group of neurons labeled positive for both cytoplasmic (AB5112) and nuclear (Ab-1)phenotypic markers. Cytoplasmic material shows a brown reaction product for the protein, whereas nuclear material exhibits a crisp, blue-blackreaction product for Parkin. Note examples of doubly labeled neurons in the (A) lateral mammillary nucleus and (B) parabrachial nucleus(medial). Magnifications 60× .
J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9390
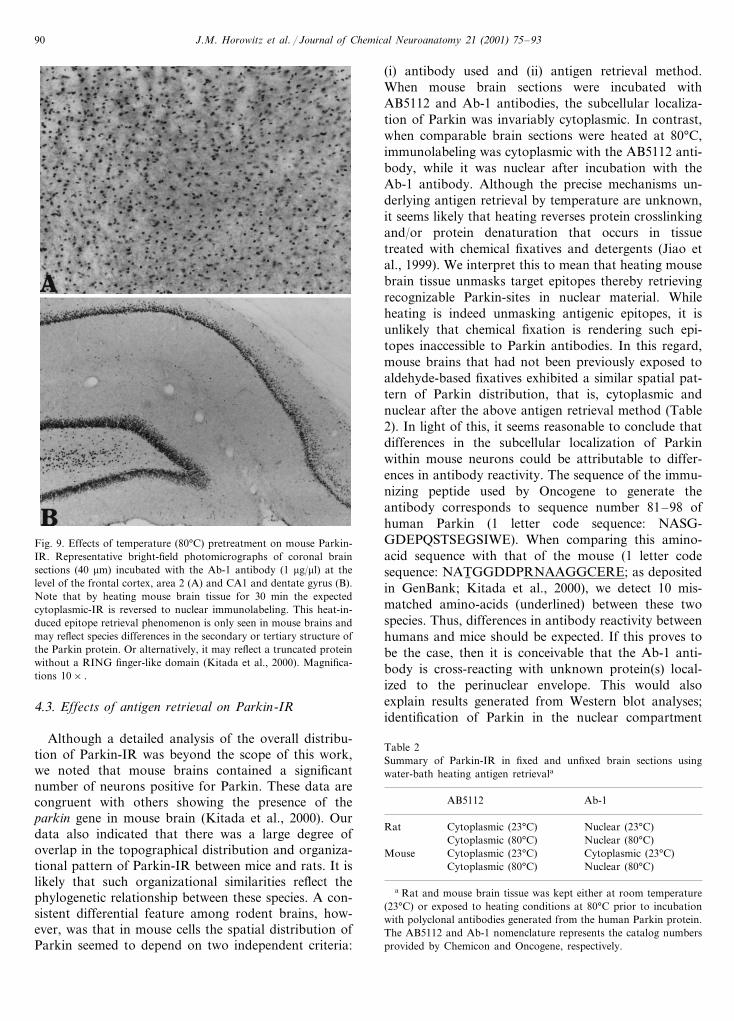
Fig. 9. Effects of temperature (80°C) pretreatment on mouse Parkin-IR. Representative bright-field photomicrographs of coronal brainsections (40 mm) incubated with the Ab-1 antibody (1 mg/ml) at thelevel of the frontal cortex, area 2 (A) and CA1 and dentate gyrus (B).Note that by heating mouse brain tissue for 30 min the expectedcytoplasmic-IR is reversed to nuclear immunolabeling. This heat-in-duced epitope retrieval phenomenon is only seen in mouse brains andmay reflect species differences in the secondary or tertiary structure ofthe Parkin protein. Or alternatively, it may reflect a truncated proteinwithout a RING finger-like domain (Kitada et al., 2000). Magnifica-tions 10× .
(i) antibody used and (ii) antigen retrieval method.When mouse brain sections were incubated withAB5112 and Ab-1 antibodies, the subcellular localiza-tion of Parkin was invariably cytoplasmic. In contrast,when comparable brain sections were heated at 80°C,immunolabeling was cytoplasmic with the AB5112 anti-body, while it was nuclear after incubation with theAb-1 antibody. Although the precise mechanisms un-derlying antigen retrieval by temperature are unknown,it seems likely that heating reverses protein crosslinkingand/or protein denaturation that occurs in tissuetreated with chemical fixatives and detergents (Jiao etal., 1999). We interpret this to mean that heating mousebrain tissue unmasks target epitopes thereby retrievingrecognizable Parkin-sites in nuclear material. Whileheating is indeed unmasking antigenic epitopes, it isunlikely that chemical fixation is rendering such epi-topes inaccessible to Parkin antibodies. In this regard,mouse brains that had not been previously exposed toaldehyde-based fixatives exhibited a similar spatial pat-tern of Parkin distribution, that is, cytoplasmic andnuclear after the above antigen retrieval method (Table2). In light of this, it seems reasonable to conclude thatdifferences in the subcellular localization of Parkinwithin mouse neurons could be attributable to differ-ences in antibody reactivity. The sequence of the immu-nizing peptide used by Oncogene to generate theantibody corresponds to sequence number 81–98 ofhuman Parkin (1 letter code sequence: NASG-GDEPQSTSEGSIWE). When comparing this amino-acid sequence with that of the mouse (1 letter codesequence: NAT6 GGDDPRNAAGGCERE; as depositedin GenBank; Kitada et al., 2000), we detect 10 mis-matched amino-acids (underlined) between these twospecies. Thus, differences in antibody reactivity betweenhumans and mice should be expected. If this proves tobe the case, then it is conceivable that the Ab-1 anti-body is cross-reacting with unknown protein(s) local-ized to the perinuclear envelope. This would alsoexplain results generated from Western blot analyses;identification of Parkin in the nuclear compartment
4.3. Effects of antigen retrie6al on Parkin-IR
Although a detailed analysis of the overall distribu-tion of Parkin-IR was beyond the scope of this work,we noted that mouse brains contained a significantnumber of neurons positive for Parkin. These data arecongruent with others showing the presence of theparkin gene in mouse brain (Kitada et al., 2000). Ourdata also indicated that there was a large degree ofoverlap in the topographical distribution and organiza-tional pattern of Parkin-IR between mice and rats. It islikely that such organizational similarities reflect thephylogenetic relationship between these species. A con-sistent differential feature among rodent brains, how-ever, was that in mouse cells the spatial distribution ofParkin seemed to depend on two independent criteria:
Table 2Summary of Parkin-IR in fixed and unfixed brain sections usingwater-bath heating antigen retrievala
AB5112 Ab-1
Cytoplasmic (23°C)Rat Nuclear (23°C)Cytoplasmic (80°C) Nuclear (80°C)
Mouse Cytoplasmic (23°C) Cytoplasmic (23°C)Cytoplasmic (80°C) Nuclear (80°C)
a Rat and mouse brain tissue was kept either at room temperature(23°C) or exposed to heating conditions at 80°C prior to incubationwith polyclonal antibodies generated from the human Parkin protein.The AB5112 and Ab-1 nomenclature represents the catalog numbersprovided by Chemicon and Oncogene, respectively.

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 91
Fig. 11. Representative bright-field photomicrographs of coronalbrain sections through the paraventricular nucleus of the hypothala-mus (A, B), and CA3 of the hippocampus (C). Brain tissue wascollected from rats left intact (i.e., without aldehyde fixation) at roomtemperature for 6 h after death. Prior to incubation with the Ab-1antibody (1 mg/ml), brain sections (40 mm) were treated with eitherKPBS (A) or sodium-m-periodate (B, C) and then exposed to anoptimal temperature of 80°C. Note that Parkin-IR in hypothalamicand hippocampal neurons is well preserved in unfixed rodent brains(A–C), and its predicted nuclear expression is not affected by agonalor postmortem variables. Magnifications 20× .
Fig. 10. Effects of temperature (80°C) pretreatment on rat (A), mouse(B) and bird (C) Parkin-IR. Representative bright-field photomicro-graphs of coronal brain sections (40 mm) incubated with the Ab-1antibody (1 mg/ml) at the level of the cerebellum. Note that nuclearParkin-IR is found in cerebellar lobules of all three vertebrate species.The effectiveness of the heating method for antigen retrieval indicatesthat heating modifies target epitopes in both cytoplasmic and nuclearmaterial of mouse (and avian) brain. Magnifications 20× .

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–9392
may be due to differences in antibody reactivity. Al-though the generality of this conclusion is uncertain,the results support the possibility that at least some ofthe IR within mouse nuclear material was the result ofbinding to a similar (Parkin) epitope. This issue needsto be further investigated before a coherent descriptionof Parkin distribution within mammalian cells can beformulated.
By comparing amino-acid sequences from GenBank,we find that mouse Parkin contains 464 amino-acids,whereas rat Parkin consists of 458 amino-acids. Thisrepresents considerable sequence homology (94% be-tween mice and rats) with their human counterpart(84% homology between mice and humans, and 84%homology between rats and humans). This high degreeof homology suggests that rodents may be used tostudy whether Parkin protein is required for mainte-nance of mature DA neuron function. In this regard,the identification of an altered protein involved in thepathogenesis of PD raises some outstanding questions.For example, we can ask whether decreased expressionof Parkin may increase the sensitivity of nigral DAneurons to neurotoxin-induced injury, or whetherchanges in Parkin concentrations may impair the self-repairing process of DA cells to chemical insult. Also,we can ask whether transgenic mice generated with amutant Parkin develop overt signs of Parkinson. Fi-nally, it would be of interest to determine whether localor regional Parkin protein concentrations are dimin-ished as a function of age, thus rendering the cellularmicroenvironment more susceptible to neurotoxin-in-duced injury. Of course, the development of a radioim-munoassay would facilitate the quantitativemeasurements of Parkin in these latter studies, all ofwhich are currently on-going projects in our laborato-ries. In conclusion, the main findings of this study arethat (i) Parkin is found in vertebrate (e.g. rat, mouse,bird, frog) and invertebrate (e.g. fruit-flies) organisms,(ii) Parkin-IR is extensive and widespread in rat andmouse brains, (iii) the cellular distribution of Parkin-IRappears to be both cytoplasmic and nuclear, (iv) thereis a modest colocalization of cytoplasmic and nuclearParkin within specific neuronal networks, and (v) anti-gen retrieval by water-bath heating unmasks Parkinepitopes in mouse and avian, but not rat brains.
Acknowledgements
This study was supported by a National ParkinsonFoundation Grant to J.M.H., A Parkinson’s DiseaseFoundation Grant and a National Science FoundationGrant to M.K.S., and a Multidisciplinary ResearchPilot Project Program from SUNY at Buffalo to G.T.The authors would like to thank Christopher Goulahfor his excellent technical assistance.
References
Aguilera, M., Oliveros, M., Martinez-Padron, M., Barbas, J.A.,Ferrus, A., 2000. Ariadne-1: a vital Drosophila gene is required indevelopment and defines a new conserved family of RING-fingerproteins. Genetics 155, 1231–1244.
Charleston, K.P., Meister, B., Hanlon, D.W., 1999. Immunohisto-chemical localization of parkin. Soc. Neurosci. Abs. 25, 19–27.
Clark, M.S., 1999. Comparative genomics: the key to understandingthe human genome project. BioEssays. 21, 121–130.
D’Agata, V., Zhao, W., Cavallaro, S., 2000. Cloning and distributionof the rat Parkin mRNA. Mol. Brain Res. 75, 345–349.
Dapson, R.W., 1993. Fixation for the 1990s: a review of needs andaccomplishments. Biotechnol. Histochem. 68, 75–82.
Gu, W.-J., Abbas, N., Zarate Lagunes, M., Parent, A., Pradier, L.,Bohme, G.A., Agid, Y., Hirsch, E.C., Raisman-Vozari, R., Brice,A., 2000. Cloning of rat Parkin cDNA and distribution of Parkinin rat brain. J. Neurochem. 74, 1773–1776.
Hattori, N., Kitada, T., Matsumine, H., Asakawa, S., Yamamura,Y., Yoshino, H., Kobayashi, T., Yokochi, M., Wang, M., Yori-taka, A., Kondo, T., Kuzuhara, S., Nakamura, S., Simizu, N.,Mizuno, Y., 1998a. Molecular genetic analysis of a novel Parkingene in Japanese families with autosomal recessive juvenileparkinsonism: evidence for variable homozygous deletions in theParkin gene in affected individuals. Ann. Neurol. 44, 935–941.
Hattori, N., Matsumine, H., Asakawa, S., Kitada, T., Yoshino, H.,Elibol, B., Brookes, A.J., Yamamura, Y., Kobayashi, T., Wang,M., Yoritaka, A., Minoshima, S., Shimizu, N., Mizuno, Y.,1998b. Point mutations (Thr240Arg and Ala311Stop) in theParkin gene. Biochem. Biophys. Res. Comm. 249, 754–758.
Hirth, F., Reichert, G., 1999. Conserved genetic programs in insectand mammalian brain development. BioEssays. 21, 677–684.
Horowitz, J.M., Myers, J., Stachowiak, M.K., Torres, G., 1999.Identification and distribution of Parkin in rat brain. NeuroRe-port 10, 3393–3397.
Jiao, Y., Sun, Z., Fusco, F.R., Kimble, T.D., Meade, C.A., Cuthber-ston, S., Reiner, A., 1999. A simple and sensitive antigen retrievalmethod for free-floating and slide-mounted tissue. J. Neurosci.Meth. 93, 149–162.
Kitada, T., Asakawa, S., Hattori, N., Matsumine, H., Yamamura,Y., Minoshima, S., Yokochi, M., Mizuno, Y., Shimizu, N., 1998.Mutations in the Parkin gene cause autosomal recessive juvenileparkinsonism. Nature 392, 605–608.
Kitada, T., Asakawa, S., Minoshima, S., Mizuno, Y., Shimizu, N.,2000. Molecular cloning, gene expression and identification of asplicing variant of the mouse Parkin gene. Mamm. Gen. 11,417–421.
Leroy, E., Anastasopoulos, D., Konitsiotis, S., Lavedan, C., Poly-meropoulos, M., 1998. Deletions in the Parkin gene and geneticheterogeneity in a Greek family with early onset Parkinson’sdisease. Hum. Genet. 103, 424–427.
Newman, S.J., Gentleman, S.M., 1997. Microwave antigen retrievalin formaldehyde-fixed human brain tissue. Meth. Mol. Biol. 72,145–152.
Paxinos, G., Watson, C., 1986. The Rat Brain Atlas in StereotaxicCoordinates. Academic Press, New York.
Shi, S.R., Cote, J.J., Young, L., Cote, R.J., Taylor, C.R., 1996. Useof pH 9.5 tris–HCl buffer containing 5% urea for antigen re-trieval immunohistochemistry. Biotechnol. Histochem. 71, 190–196.
Shimura, H., Hattori, N., Kubo, S., Yoshikawa, M., Kitada, T.,Matsumine, H., Asakawa, S., Minoshima, S., Yamamura, Y.,Simizu, N., Mizuno, Y., 1999. Immunohistochemical and subcel-lular localization of Parkin protein: absence of protein in autoso-mal recessive juvenile Parkinsonism patients. Ann. Neurol. 45,668–672.

J.M. Horowitz et al. / Journal of Chemical Neuroanatomy 21 (2001) 75–93 93
Shimura, H., Hattori, N., Kubo, S., Mizuno, Y., Asakawa, S.,Minoshima, S., Shimizu, N., Iwai, K., Chiba, T., Tanaka, K.,Suzuki, T., 2000. Familial Parkinson disease gene product, Parkinis a ubiquitin-protein ligase. Nature Gene. 25, 302–305.
Sidman, R.L., Angevine, J.B., Taber Pierce, E., 1971. Atlasof the Mouse Brain and Spinal Cord. Harvard University Press,MA.
Stachowiak, M.K., Maher, P., Mordechai, E., Joy, A., Stachowiak,E.W., 1996. Nuclear localization of fibroblast growth factor re-ceptors is regulated by multiple signals in adrenal medullary cells.Mol. Biol. Cell 7, 1299–1317.
Torres, G., Horowitz, J.M., 1998. Activating properties of cocaine
and cocaethylene in a behavioral preparation of Drosophilamelanogaster. Synapse 29, 148–161.
Torres, G., Lee, S., Rivier, C., 1992. Immunocytochemical and in situhybridization detection of hypothalamic neuropeptides from post-mortem unfixed rat brains. Neurosci. Lett. 146, 96–100.
Torres, G., Lee, S., Rivier, C., 1993. Ontogeny of the rat hypothala-mic nitric oxide synthase and colocalization with neuropeptides.Mol. Cell. Neurosci. 4, 155–163.
Wang, M., Hattori, N., Matsumine, H., Kobayashi, T., Yoshino, H.,Morioka, A., Kitada, T., Asakawa, S., Minoshima, S., Shimizu,N., Mizuno, Y., 1999. Polymorphism in the Parkin gene insporadic Parkinson’s disease. Ann. Neurol. 45, 655–658.
Copyright © 2022 FDOKUMEN




















