
Identification of two powdery mildew fungi, Oidium neolycopersici sp. nov. and O. lycopersici,...
14
684 Levente KISS 1 *, Roger T. A. COOK 2 , Gregory S. SAENZ 3 , James H. CUNNINGTON 4 , Susumu TAKAMATSU 5 , Ian PASCOE 6 , Marc BARDIN 7 , Philippe C. NICOT 7 , Y. SATO 8 and Amy Y. ROSSMAN 9 " Plant Protection Institute, Hungarian Academy of Sciences, P.O. Box 102, H-1525 Budapest, Hungary. # Central Science Laboratory, Ministry of Agriculture Fisheries and Food, York YO41 1LZ, UK. $ Department of Biology, University of New Mexico, Albuquerque, NM 87131, USA. % Department of Applied Biology}Biotechnology, RMIT University, Melbourne 3001, Australia. & Faculty of Bioresources, Mie University, Tsu 514-8507, Japan. ’ Institute for Horticultural Development, Victoria 3176, Australia. ( INRA, Unite U de Pathologie Ve U ge U tale, B.P. 94, 84143 Montfavet cedex, France. ) College of Technology, Toyama Prefectural University, Toyama 939-0398, Japan. * USDA-ARS, Systematic Botany & Mycology Laboratory, Beltsville, MD 20705, USA. E-mail : lkiss!nki.hu Received 15 August 2000 ; accepted 9 January 2001. A world-wide study of the Oidium species causing economic damage on tomato has identified two taxa using classical morphological, scanning electron microscope (SEM) and molecular phylogenetic analyses. The material consisted of a total of 25 tomato powdery mildew isolates and 29 herbarium specimens coming from all continents where tomatoes are grown. A taxon with non-catenate conidia widespread in Europe, Africa, North and South America and Asia was identified as an O. subgen. Pseudoidium species (teleomorph : Erysiphe sect. Erysiphe). Formerly mistaken for O. lycopersicum (or O. lycopersici), it is now recognised as a distinct species, O. neolycopersici sp. nov. A phylogenetic analysis of the internal transcribed spacer (ITS) sequences of the ribosomal DNA (rDNA) indicated that O. neolycopersici is closely related to Erysiphe macleayae, E. aquilegiae and other Pseudoidium species. Only a taxon with catenate conidia was found on Australian specimens. This was identified as a species of O. subgen. Reticuloidium (teleomorph : Golovinomyces sp.). Phylogenetic analysis of the rDNA ITS sequences showed that this species is closely related to O. longipes infecting eggplant. Because it is most likely to be the same species as the original O. lycopersicum, which was actually first described in Australia, this is here neotypified as O. lycopersici. INTRODUCTION An apparently new powdery mildew disease caused by an Oidium anamorph has appeared recently on tomato (Lyco- persicon esculentum), on both greenhouse and field crops, and has spread rapidly around the world (Whipps, Budge & Fenlon 1998, Mieslerova & Lebeda 1999). It is distinct from Leveillula taurica (Le ! v.) Arnaud 1921, the powdery mildew fungus having an Oidiopsis anamorph and long known to affect tomato in warmer regions. However, the published data on the nature of the conidiogenesis of the Oidium species associated with these new emerging epidemics are often contradictory making the identification questionable in the absence of any known teleomorph. Sometimes the conidia have been described as maturing singly and at other times as maturing in a chain. The nature of the conidiogenesis is a stable character within species, yet within Oidium there are two groups with fundamentally different types of conidio- * Corresponding author. genesis. One is a homogeneous group referred to in the literature as ‘ pseudoidium-type ’ where conidia mature one at a time and the other is a heterogeneous group, ‘ euoidium- type ’, belonging to several genera having in common conidia which mature in a chain (Blumer 1967, Hammett 1977, Boesewinkel 1980, Braun 1987, 1995). Thus, in the recent tomato powdery mildew outbreaks, at least two species of Oidium were reported sometimes even in the same geo- graphical region. A recent comprehensive scanning electron microscope (SEM) study (Cook, Inman & Billings 1997) has provided a new tool for the identification of powdery mildew anamorphs based on the surface patterns of their conidia and septa. It was subsequently supported by phylogenetic analyses of nuclear ribosomal DNA (rDNA) internal transcribed spacer (ITS) sequences in the Erysiphaceae which have provided a large data set for identification of powdery mildew species (Takamatsu, Hirata & Sato 1998, Saenz & Taylor 1999, Takamatsu et al. 1999, Mori, Sato & Takamatsu 2000). Therefore, the classical morphological criteria and host range data used in identi- Mycol. Res. 105 (6) : 684–697 (June 2001). Printed in the United Kingdom. Identification of two powdery mildew fungi, Oidium neolycopersici sp. nov. and O. lycopersici, infecting tomato in different parts of the world
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Identification of two powdery mildew fungi, Oidium neolycopersici sp. nov. and O. lycopersici,...

684
Levente KISS1*, Roger T. A. COOK2, Gregory S. SAENZ3, James H. CUNNINGTON4, Susumu TAKAMATSU5,
Ian PASCOE6, Marc BARDIN7, Philippe C. NICOT7, Y. SATO8 and Amy Y. ROSSMAN9
" Plant Protection Institute, Hungarian Academy of Sciences, P.O. Box 102, H-1525 Budapest, Hungary.#Central Science Laboratory, Ministry of Agriculture Fisheries and Food, York YO41 1LZ, UK.$Department of Biology, University of New Mexico, Albuquerque, NM 87131, USA.%Department of Applied Biology}Biotechnology, RMIT University, Melbourne 3001, Australia.& Faculty of Bioresources, Mie University, Tsu 514-8507, Japan.' Institute for Horticultural Development, Victoria 3176, Australia.( INRA, UniteU de Pathologie VeUgeU tale, B.P. 94, 84143 Montfavet cedex, France.)College of Technology, Toyama Prefectural University, Toyama 939-0398, Japan.*USDA-ARS, Systematic Botany & Mycology Laboratory, Beltsville, MD 20705, USA.
E-mail : lkiss!nki.hu
Received 15 August 2000 ; accepted 9 January 2001.
A world-wide study of the Oidium species causing economic damage on tomato has identified two taxa using classical
morphological, scanning electron microscope (SEM) and molecular phylogenetic analyses. The material consisted of a total of 25
tomato powdery mildew isolates and 29 herbarium specimens coming from all continents where tomatoes are grown. A taxon with
non-catenate conidia widespread in Europe, Africa, North and South America and Asia was identified as an O. subgen. Pseudoidium
species (teleomorph : Erysiphe sect. Erysiphe). Formerly mistaken for O. lycopersicum (or O. lycopersici), it is now recognised as a distinct
species, O. neolycopersici sp. nov. A phylogenetic analysis of the internal transcribed spacer (ITS) sequences of the ribosomal DNA
(rDNA) indicated that O. neolycopersici is closely related to Erysiphe macleayae, E. aquilegiae and other Pseudoidium species. Only a
taxon with catenate conidia was found on Australian specimens. This was identified as a species of O. subgen. Reticuloidium
(teleomorph : Golovinomyces sp.). Phylogenetic analysis of the rDNA ITS sequences showed that this species is closely related to
O. longipes infecting eggplant. Because it is most likely to be the same species as the original O. lycopersicum, which was actually first
described in Australia, this is here neotypified as O. lycopersici.
INTRODUCTION
An apparently new powdery mildew disease caused by an
Oidium anamorph has appeared recently on tomato (Lyco-
persicon esculentum), on both greenhouse and field crops, and
has spread rapidly around the world (Whipps, Budge & Fenlon
1998, Mieslerova & Lebeda 1999). It is distinct from Leveillula
taurica (Le! v.) Arnaud 1921, the powdery mildew fungus
having an Oidiopsis anamorph and long known to affect
tomato in warmer regions. However, the published data on
the nature of the conidiogenesis of the Oidium species
associated with these new emerging epidemics are often
contradictory making the identification questionable in the
absence of any known teleomorph. Sometimes the conidia
have been described as maturing singly and at other times as
maturing in a chain. The nature of the conidiogenesis is a
stable character within species, yet within Oidium there are
two groups with fundamentally different types of conidio-
* Corresponding author.
genesis. One is a homogeneous group referred to in the
literature as ‘pseudoidium-type ’ where conidia mature one at
a time and the other is a heterogeneous group, ‘euoidium-
type ’, belonging to several genera having in common conidia
which mature in a chain (Blumer 1967, Hammett 1977,
Boesewinkel 1980, Braun 1987, 1995). Thus, in the recent
tomato powdery mildew outbreaks, at least two species of
Oidium were reported sometimes even in the same geo-
graphical region.
A recent comprehensive scanning electron microscope
(SEM) study (Cook, Inman & Billings 1997) has provided a
new tool for the identification of powdery mildew anamorphs
based on the surface patterns of their conidia and septa. It was
subsequently supported by phylogenetic analyses of nuclear
ribosomal DNA (rDNA) internal transcribed spacer (ITS)
sequences in the Erysiphaceae which have provided a large data
set for identification of powdery mildew species (Takamatsu,
Hirata & Sato 1998, Saenz & Taylor 1999, Takamatsu et al.
1999, Mori, Sato & Takamatsu 2000). Therefore, the classical
morphological criteria and host range data used in identi-
Mycol. Res. 105 (6) : 684–697 (June 2001). Printed in the United Kingdom.
Identification of two powdery mildew fungi, Oidiumneolycopersici sp. nov. and O. lycopersici, infecting tomatoin different parts of the world

L. Kiss and others 685
fication (Braun 1987, 1995) can now be supplemented with
both SEM and molecular methods.
This confluence of methods giving mutually supporting
results has triggered a complex revolution in powdery mildew
taxonomy. All ‘pseudoidium ’ anamorphs were shown to
belong to a homogeneous group and are now referred to a
single subgenus of Oidium viz O. subgen. Pseudoidium.
However, ‘euoidium ’ anamorphs are heterogeneous and were
shown to belong to six separate subgenera of Oidium viz O.
subgen. : Oidium, Reticuloidium, Striatoidium, Graciloidium,
Fibroidium, and Setoidium (Cook et al. 1997, Braun et al. 2001).
(Since ‘euoidium ’ is not a homogeneous group, we suggest
replacing this term with ‘catenate Oidium species ’.) These
findings have necessitated a re-structuring of many of the
teleomorphic genera. For instance, Erysiphe is now reserved
for all the teleomorphs of O. subgen. Pseudoidium thus
absorbing the old genera of Microsphaera and Uncinula which
are relegated to sections of Erysiphe. The remaining (chain
forming) members of the old genus Erysiphe have now been
subdivided into Golovinomyces having O. subgen. Reticuloidium
anamorphs, and Neoerysiphe having O. subgen. Striatoidium
anamorphs (Braun 1999, Braun & Takamatsu 2000).
The uncertainty regarding the identity of the causal agents
of tomato powdery mildew poses a major problem for plant
breeders. There is an urgent need to breed resistant tomato
cultivars against the new disease because all cultivars grown
world-wide are susceptible (Burgerjon et al. 1990, Neshev
1993, Lindhout, Pet & van der Beek 1994, Ciccarese et al.
1998, Huang et al. 2000). However, if different powdery
mildew species are involved, different mechanisms of resistance
could complicate the breeding strategy as in the case of certain
melon lines which could be infected by two distinct powdery
mildew species (Epinat, Pitrat & Bertrand 1993). Consequently,
there is a practical need for an exact identification of novel
pathogens for breeding purposes.
Reports suggesting that the pathogen was a catenate
Oidium species came from four continents : Australia (Price
1981, Wicks & Clare 1981) ; Asia : Japan (Abiko 1978) and
India (Kumar et al. 1995) ; Europe : Switzerland (Corbaz 1990,
1993), Bulgaria (Neshev 1993), and Hungary (Kiss 1996) ; and
North America : Canada (Be! langer & Jarvis 1994), and the
USA (Arredondo et al. 1996, Karasevicz & Zitter 1996, White
et al. 1997, Pernezny & Sonoda 1998). The names Erysiphe
cichoracearum DC. 1805 (Abiko 1978, Wicks & Clare 1981,
Corbaz 1990, 1993), Oidium sp. (Price 1981, Neshev 1993)
and Erysiphe sp. (Be! langer & Jarvis 1994, Kiss 1996, Pernezny
& Sonoda 1998) were used for these anamorphs.
The situation is even more complicated with Pseudoidium.
Reports describing this taxon on tomato came from two
continents : Europe : UK (Whipps et al. 1998), Netherlands
(Noordeloos & Loerakker 1989), Germany (Gabler, Gerlach &
Braun 1990), the Czech Republic (Lebeda & Rod 1990),
Greece (Vakalounakis & Papadakis 1992), Hungary (Milotay
& Dormanns-Simon 1997), Switzerland (Bolay 1998) and
Spain (Olalla & Tore! s 1998) ; and North America : USA (Smith,
Douglas & LaMondia 1997) and Canada (Bains, Bennypaul &
Mirza 1999).
Noordeloos & Loerakker (1989) redescribed a Pseudoidium,
infecting tomato in the Dutch glasshouses since the 1980s, as
‘O. lycopersicum Cooke & Massee emend. ’. They based this on
the oldest description of a powdery mildew on tomato
collected in southern Australia (Cooke & Massee 1888),
although this name was previously excluded from the
Erysiphales accepted by Braun (1987) who rightly regarded it
as a nomen ambiguum. Noordeloos & Loerakker (1989) were
able to find and illustrate an interpretable anamorphic mildew
from the holotype, and demonstrate that it was an Oidium
species. Unfortunately, the holotype specimen was in poor
condition and Noordeloos & Loerakker (1989) were unable to
determine the nature of the conidiogenesis of this taxon.
Although they adopted the name O. lycopersicum for the
European tomato powdery mildew fungus, they did not
provide convincing evidence that it was the same species as
the Dutch Pseudoidium anamorph. The holotype of Cooke &
Massee’s O. lycopersicum was lost after this taxonomic study
(I. Russ, pers. comm.) and there is no more original material.
Clearly, use of the name O. lycopersicum is in need of
clarification and neotypification.
Based on specimens from the UK and Czech Republic,
Whipps et al. (1998) and Lebeda & Mieslerova (1999)
suggested that recent outbreaks of powdery mildew on
tomato were caused by a single species of Pseudoidium and
that this anamorph was often misidentified as a catenate
Oidium species because its conidia clung together in ‘pseudo-
chains ’ in high relative humidity. This could explain why the
first reports of the pathogen in the UK (Fletcher et al. 1998)
were not positively identified as either a Pseudoidium or a
catenate Oidium species. The name O. lycopersicum (Whipps et
al. 1998), or its orthographically corrected form, O. lycopersici
(Lebeda & Mieslerova 1999), was recommended for this
species. However, these authors did not compare the
morphology of their local specimens with specimens from
other geographical regions. Thus, a further comprehensive
study is needed to identify the powdery mildews infecting
tomato throughout the world.
The present work combines the use of morphological and
molecular tools to identify the Oidium species infecting
tomato. Its objectives are to : (1) compare the morphological
characteristics of Oidium tomato pathogens from all over the
world, using light microscopy and SEM of both fresh
materials and herbarium specimens ; (2) determine and analyze
the rDNA ITS sequences of these specimens ; (3) study the
mating type of different isolates in vitro ; and (4) to solve the
nomenclatural problems associated with tomato powdery
mildew anamorphs.
MATERIALS AND METHODS
Powdery mildew materials
Twenty-five Oidium tomato powdery mildew isolates were
examined as fresh material (Table 1). The European, African
and Caribbean isolates have been maintained for years in
different laboratories at INRA, PPI and WAU, respectively
(Table 1). Isolates maintained at INRA were kept in Petri
dishes on surface disinfected cotyledons of Lagenaria ciceracia
cv. Minibottle on mannitol sucrose (MS) agar (mannitol,
20 g l−", sucrose, 10 g l−", agar, 7 g l−", tetracycline hydro-
Powdery mildew fungi on tomato 686
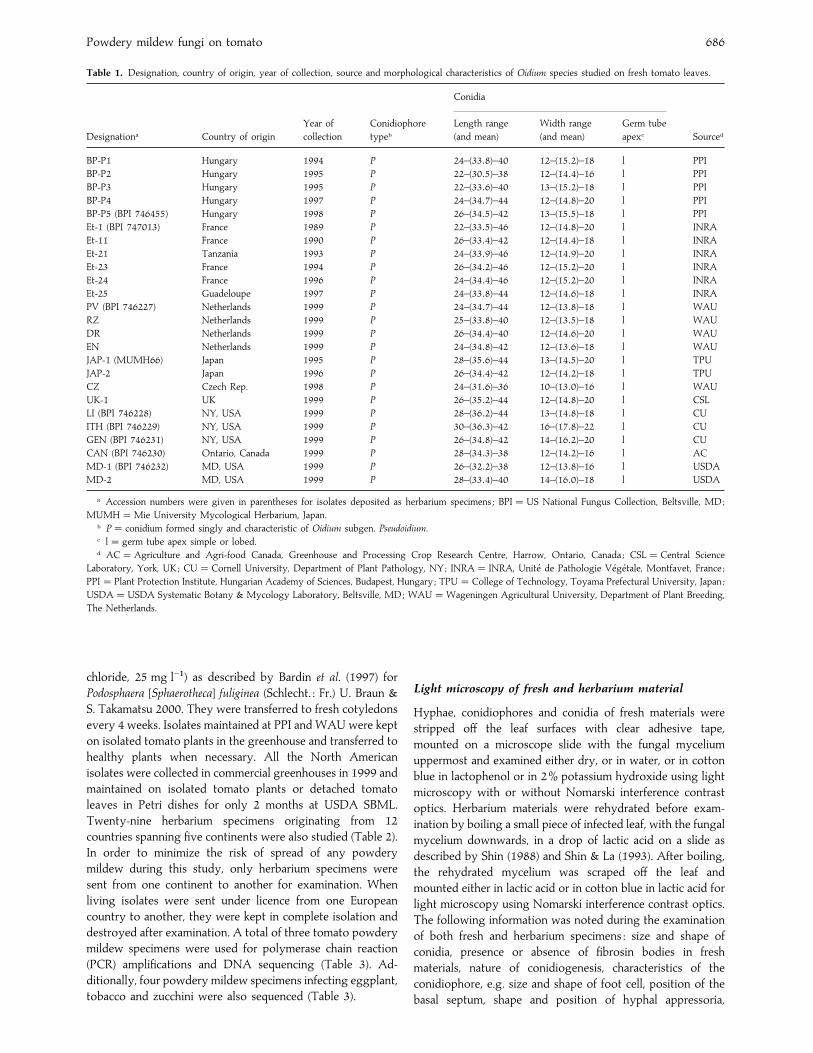
Table 1. Designation, country of origin, year of collection, source and morphological characteristics of Oidium species studied on fresh tomato leaves.
Designationa Country of origin
Year of
collection
Conidiophore
typeb
Conidia
Sourced
Length range
(and mean)
Width range
(and mean)
Germ tube
apexc
BP-P1 Hungary 1994 P 24–(33±8)–40 12–(15±2)–18 l PPI
BP-P2 Hungary 1995 P 22–(30±5)–38 12–(14±4)–16 l PPI
BP-P3 Hungary 1995 P 22–(33±6)–40 13–(15±2)–18 l PPI
BP-P4 Hungary 1997 P 24–(34±7)–44 12–(14±8)–20 l PPI
BP-P5 (BPI 746455) Hungary 1998 P 26–(34±5)–42 13–(15±5)–18 l PPI
Et-1 (BPI 747013) France 1989 P 22–(33±5)–46 12–(14±8)–20 l INRA
Et-11 France 1990 P 26–(33±4)–42 12–(14±4)–18 l INRA
Et-21 Tanzania 1993 P 24–(33±9)–46 12–(14±9)–20 l INRA
Et-23 France 1994 P 26–(34±2)–46 12–(15±2)–20 l INRA
Et-24 France 1996 P 24–(34±4)–46 12–(15±2)–20 l INRA
Et-25 Guadeloupe 1997 P 24–(33±8)–44 12–(14±6)–18 l INRA
PV (BPI 746227) Netherlands 1999 P 24–(34±7)–44 12–(13±8)–18 l WAU
RZ Netherlands 1999 P 25–(33±8)–40 12–(13±5)–18 l WAU
DR Netherlands 1999 P 26–(34±4)–40 12–(14±6)–20 l WAU
EN Netherlands 1999 P 24–(34±8)–42 12–(13±6)–18 l WAU
JAP-1 (MUMH66) Japan 1995 P 28–(35±6)–44 13–(14±5)–20 l TPU
JAP-2 Japan 1996 P 26–(34±4)–42 12–(14±2)–18 l TPU
CZ Czech Rep. 1998 P 24–(31±6)–36 10–(13±0)–16 l WAU
UK-1 UK 1999 P 26–(35±2)–44 12–(14±8)–20 l CSL
LI (BPI 746228) NY, USA 1999 P 28–(36±2)–44 13–(14±8)–18 l CU
ITH (BPI 746229) NY, USA 1999 P 30–(36±3)–42 16–(17±8)–22 l CU
GEN (BPI 746231) NY, USA 1999 P 26–(34±8)–42 14–(16±2)–20 l CU
CAN (BPI 746230) Ontario, Canada 1999 P 28–(34±3)–38 12–(14±2)–16 l AC
MD-1 (BPI 746232) MD, USA 1999 P 26–(32±2)–38 12–(13±8)–16 l USDA
MD-2 MD, USA 1999 P 28–(33±4)–40 14–(16±0)–18 l USDA
a Accession numbers were given in parentheses for isolates deposited as herbarium specimens ; BPI¯US National Fungus Collection, Beltsville, MD;
MUMH¯Mie University Mycological Herbarium, Japan.b P¯ conidium formed singly and characteristic of Oidium subgen. Pseudoidium.c l¯ germ tube apex simple or lobed.d AC¯Agriculture and Agri-food Canada, Greenhouse and Processing Crop Research Centre, Harrow, Ontario, Canada ; CSL¯Central Science
Laboratory, York, UK; CU¯Cornell University, Department of Plant Pathology, NY; INRA¯ INRA, Unite! de Pathologie Ve! ge! tale, Montfavet, France ;
PPI¯ Plant Protection Institute, Hungarian Academy of Sciences, Budapest, Hungary ; TPU¯College of Technology, Toyama Prefectural University, Japan ;
USDA¯USDA Systematic Botany & Mycology Laboratory, Beltsville, MD; WAU¯Wageningen Agricultural University, Department of Plant Breeding,
The Netherlands.
chloride, 25 mg l−") as described by Bardin et al. (1997) for
Podosphaera [Sphaerotheca] fuliginea (Schlecht. : Fr.) U. Braun &
S. Takamatsu 2000. They were transferred to fresh cotyledons
every 4 weeks. Isolates maintained at PPI and WAU were kept
on isolated tomato plants in the greenhouse and transferred to
healthy plants when necessary. All the North American
isolates were collected in commercial greenhouses in 1999 and
maintained on isolated tomato plants or detached tomato
leaves in Petri dishes for only 2 months at USDA SBML.
Twenty-nine herbarium specimens originating from 12
countries spanning five continents were also studied (Table 2).
In order to minimize the risk of spread of any powdery
mildew during this study, only herbarium specimens were
sent from one continent to another for examination. When
living isolates were sent under licence from one European
country to another, they were kept in complete isolation and
destroyed after examination. A total of three tomato powdery
mildew specimens were used for polymerase chain reaction
(PCR) amplifications and DNA sequencing (Table 3). Ad-
ditionally, four powdery mildew specimens infecting eggplant,
tobacco and zucchini were also sequenced (Table 3).
Light microscopy of fresh and herbarium material
Hyphae, conidiophores and conidia of fresh materials were
stripped off the leaf surfaces with clear adhesive tape,
mounted on a microscope slide with the fungal mycelium
uppermost and examined either dry, or in water, or in cotton
blue in lactophenol or in 2% potassium hydroxide using light
microscopy with or without Nomarski interference contrast
optics. Herbarium materials were rehydrated before exam-
ination by boiling a small piece of infected leaf, with the fungal
mycelium downwards, in a drop of lactic acid on a slide as
described by Shin (1988) and Shin & La (1993). After boiling,
the rehydrated mycelium was scraped off the leaf and
mounted either in lactic acid or in cotton blue in lactic acid for
light microscopy using Nomarski interference contrast optics.
The following information was noted during the examination
of both fresh and herbarium specimens : size and shape of
conidia, presence or absence of fibrosin bodies in fresh
materials, nature of conidiogenesis, characteristics of the
conidiophore, e.g. size and shape of foot cell, position of the
basal septum, shape and position of hyphal appressoria,
L. Kiss and others 687
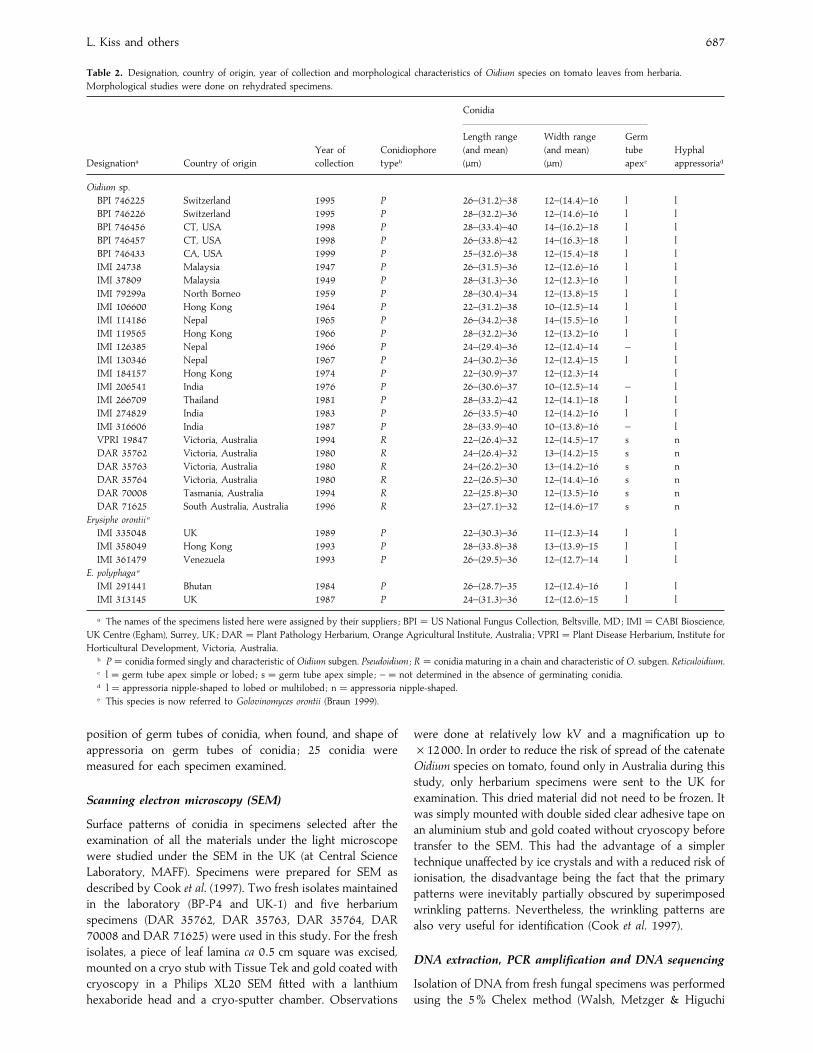
Table 2. Designation, country of origin, year of collection and morphological characteristics of Oidium species on tomato leaves from herbaria.
Morphological studies were done on rehydrated specimens.
Designationa Country of origin
Year of
collection
Conidiophore
typeb
Conidia
Hyphal
appressoriad
Length range
(and mean)
(µm)
Width range
(and mean)
(µm)
Germ
tube
apexc
Oidium sp.
BPI 746225 Switzerland 1995 P 26–(31±2)–38 12–(14±4)–16 l l
BPI 746226 Switzerland 1995 P 28–(32±2)–36 12–(14±6)–16 l l
BPI 746456 CT, USA 1998 P 28–(33±4)–40 14–(16±2)–18 l l
BPI 746457 CT, USA 1998 P 26–(33±8)–42 14–(16±3)–18 l l
BPI 746433 CA, USA 1999 P 25–(32±6)–38 12–(15±4)–18 l l
IMI 24738 Malaysia 1947 P 26–(31±5)–36 12–(12±6)–16 l l
IMI 37809 Malaysia 1949 P 28–(31±3)–36 12–(12±3)–16 l l
IMI 79299a North Borneo 1959 P 28–(30±4)–34 12–(13±8)–15 l l
IMI 106600 Hong Kong 1964 P 22–(31±2)–38 10–(12±5)–14 l l
IMI 114186 Nepal 1965 P 26–(34±2)–38 14–(15±5)–16 l l
IMI 119565 Hong Kong 1966 P 28–(32±2)–36 12–(13±2)–16 l l
IMI 126385 Nepal 1966 P 24–(29±4)–36 12–(12±4)–14 – l
IMI 130346 Nepal 1967 P 24–(30±2)–36 12–(12±4)–15 l l
IMI 184157 Hong Kong 1974 P 22–(30±9)–37 12–(12±3)–14 l
IMI 206541 India 1976 P 26–(30±6)–37 10–(12±5)–14 – l
IMI 266709 Thailand 1981 P 28–(33±2)–42 12–(14±1)–18 l l
IMI 274829 India 1983 P 26–(33±5)–40 12–(14±2)–16 l l
IMI 316606 India 1987 P 28–(33±9)–40 10–(13±8)–16 – l
VPRI 19847 Victoria, Australia 1994 R 22–(26±4)–32 12–(14±5)–17 s n
DAR 35762 Victoria, Australia 1980 R 24–(26±4)–32 13–(14±2)–15 s n
DAR 35763 Victoria, Australia 1980 R 24–(26±2)–30 13–(14±2)–16 s n
DAR 35764 Victoria, Australia 1980 R 22–(26±5)–30 12–(14±4)–16 s n
DAR 70008 Tasmania, Australia 1994 R 22–(25±8)–30 12–(13±5)–16 s n
DAR 71625 South Australia, Australia 1996 R 23–(27±1)–32 12–(14±6)–17 s n
Erysiphe orontii e
IMI 335048 UK 1989 P 22–(30±3)–36 11–(12±3)–14 l l
IMI 358049 Hong Kong 1993 P 28–(33±8)–38 13–(13±9)–15 l l
IMI 361479 Venezuela 1993 P 26–(29±5)–36 12–(12±7)–14 l l
E. polyphaga e
IMI 291441 Bhutan 1984 P 26–(28±7)–35 12–(12±4)–16 l l
IMI 313145 UK 1987 P 24–(31±3)–36 12–(12±6)–15 l l
a The names of the specimens listed here were assigned by their suppliers ; BPI¯US National Fungus Collection, Beltsville, MD; IMI¯CABI Bioscience,
UK Centre (Egham), Surrey, UK; DAR¯ Plant Pathology Herbarium, Orange Agricultural Institute, Australia ; VPRI¯ Plant Disease Herbarium, Institute for
Horticultural Development, Victoria, Australia.b P¯ conidia formed singly and characteristic of Oidium subgen. Pseudoidium ; R¯ conidia maturing in a chain and characteristic of O. subgen. Reticuloidium.c l¯ germ tube apex simple or lobed ; s¯ germ tube apex simple ; –¯ not determined in the absence of germinating conidia.d l¯ appressoria nipple-shaped to lobed or multilobed ; n¯ appressoria nipple-shaped.e This species is now referred to Golovinomyces orontii (Braun 1999).
position of germ tubes of conidia, when found, and shape of
appressoria on germ tubes of conidia ; 25 conidia were
measured for each specimen examined.
Scanning electron microscopy (SEM)
Surface patterns of conidia in specimens selected after the
examination of all the materials under the light microscope
were studied under the SEM in the UK (at Central Science
Laboratory, MAFF). Specimens were prepared for SEM as
described by Cook et al. (1997). Two fresh isolates maintained
in the laboratory (BP-P4 and UK-1) and five herbarium
specimens (DAR 35762, DAR 35763, DAR 35764, DAR
70008 and DAR 71625) were used in this study. For the fresh
isolates, a piece of leaf lamina ca 0±5 cm square was excised,
mounted on a cryo stub with Tissue Tek and gold coated with
cryoscopy in a Philips XL20 SEM fitted with a lanthium
hexaboride head and a cryo-sputter chamber. Observations
were done at relatively low kV and a magnification up to
¬12000. In order to reduce the risk of spread of the catenate
Oidium species on tomato, found only in Australia during this
study, only herbarium specimens were sent to the UK for
examination. This dried material did not need to be frozen. It
was simply mounted with double sided clear adhesive tape on
an aluminium stub and gold coated without cryoscopy before
transfer to the SEM. This had the advantage of a simpler
technique unaffected by ice crystals and with a reduced risk of
ionisation, the disadvantage being the fact that the primary
patterns were inevitably partially obscured by superimposed
wrinkling patterns. Nevertheless, the wrinkling patterns are
also very useful for identification (Cook et al. 1997).
DNA extraction, PCR amplification and DNA sequencing
Isolation of DNA from fresh fungal specimens was performed
using the 5% Chelex method (Walsh, Metzger & Higuchi

Powdery mildew fungi on tomato 688
Table 3. Designations, host plants, countries of origin and database accession numbers of the powdery mildew rDNA ITS sequences used in this study.
Fungus Host plant Designation and source Country of origin Database accession number
Arthrocladiella mougeotii Lycium chinense MUMH135a Japan AB022380
Erysiphe aquilegiae var. ranunculi Cimifuga simplex TPU495a Japan AB015929
E. convolvuli Convolvulus arvensis UC1512307a UK AF011298
E. cruciferarum Arabidopsis thaliana UEA1b UK AF031283
E. macleayae Macleaya cordata TPU1873a Japan AB016048
E. polygoni Polygonum arenastrum UC1512295a USA AF011307
E. weigelae Weigela hortensis TPU1669a Japan AB015931
E. blasti e Lindera umbellata MUMH2a Japan AB015918
E. juglandis e Pterocarpa rhoifolia TPU1745a Japan AB015928
E. platani e Platanus racemosa UC1512309a USA AF011311
E. pulchra var. japonica e Cornus controversa MUMH90a Japan AB015924
Golovinomyces biocellatus f Monarda fistulosa DNAc USA AF011291
G. cichoracearum f,g Arabidopsis thaliana UCSC1a USA AF031282
G. cichoracearum d,f,g Cucurbita pepo Ecr-3b France AF229017
G. cichoracearum d,f,g C. pepo Ecr-1b France F229016
G. cichoracearum f Eupatorium chinense MUMH37a Japan AB000934
G. magnicellulatus f Phlox paniculata DNAc USA AF011303
G. orontii f Agastache pallida var. pallida UC1512314a USA AF011304
G. orontii f Arabidopsis thaliana MGHb USA AF009176
G. orontii f Delphinium belladonna UC1512293a USA AF011305
G. orontii d,f Nicotiana tabacum BP-1TOBb Hungary AF229013
G. orontii f N. tabacum L.I.b Japan AB022413
G. sordidus f Plantago major DNAc USA AF011309
Oidium longipes d Solanum melongena VPRI 22141a Switzerland AF250777
O. lycopersici d Lycopersicon esculentum VPRI 19847a Australia AF229021
O. neolycopersici d L. esculentum Et-1b France AF229019
O. neolycopersici d L. esculentum VPRI 20724a The Netherlands AF229015
a Herbarium specimens at the following Herbaria : MUMH¯Mie University Mycological Herbarium, Japan ; TPU¯Toyama Prefectural University
Herbarium, Japan ; UC¯University of California Herbarium, CA; VPRI¯ Plant Disease Herbarium, Institute for Horticultural Development, Victoria,
Australia.b Isolate maintained on living plants or detached leaves.c DNA extracted from fresh material, no voucher specimen deposited.d New ITS sequence obtained for this study.e Genus formerly known as Microsphaera.f Genus formerly known as Erysiphe.g These isolates cannot belong to G. cichoracearum, because this species is resticted to the host family Compositae (Braun 1987). Therefore, they probably
belong to G. orontii.
1991) with modifications as described in Saenz & Taylor
(1999). This extract was autoclaved for 20 min, and centrifuged
for 10 min at 15000 g. Serial dilutions of 1}10 and 1}100 of
the supernatant were then used as template DNA for
polymerase chain reaction (PCR) amplifications. DNA from
herbarium specimens was isolated by placing scrapings of
mycelium and conidiophores into a 5% Chelex solution
containing 0±001% Triton X100, and incubating in a water
bath for 30 min at 94 °C. A 1 µl aliquot from this mixture was
used as the template.
For DNA from fresh specimens, PCR amplifications utilised
the ITS5 and ITS4 primers pairs (White et al. 1990). The
amplified fragment spanned the entire ITS region. Each
reaction was subjected to 40 thermal cycles with the following
parameters : denaturation at 94° for 1 min, annealing at 53°for 1 min, extension at 72° for 1 min. The ITS region from the
herbarium specimens was amplified using the following two
primers, PMITS1 (5-TCGGACTGGCCTCAGGGAGA-3) and
PMITS2 (5-TCACTCGCCGTTACTGAGGT-3). These pri-
mers have been shown to have some specificity for the ITS
region of powdery mildews (Cunnington & Takamatsu,
unpubl.). In these reactions, 6% Tween 20 was used as a PCR
enhancer (Demeke & Adams 1992) and the thermal cycling
times were 1 min at 94°, 1 min 65° and 2 min 72°, for 35
cycles.
Amplified products were electrophoresed on a 2% agarose
gel and then cleaned using QIAquick spin columns (Qiagen,
Chatsworth, CA). DNA was resuspended into about 30 µl of
sterile water. Ethidium bromide-stained DNA PCR products
were visually compared to standards in order to optimise the
concentration for cycle sequencing. Some specimens were
cycle sequenced using an ABI Prism BigDye Terminator Cycle
Sequencing Kit (Perkin–Elmer, Foster City, CA) and were read
by the DNA Sequencing Facility, Monash University, Victoria,
Australia. Other PCR purified amplicons were sent to the
DNA Sequencing Facility, Center for Agricultural Biotech-
nology, at the University of Maryland, for cycle sequencing,
polyacrylamide gel electrophoresis, and data collection.
Primers ITS1, ITS2, ITS3, ITS4, and ITS5, were used to
sequence the ITS region (White et al. 1990).
Data analysis
Two data sets were created for phylogenetic analysis, one
consisting of the Pseudoidium species and the other consisting
of the catenate Oidium species. The powdery mildews used in

L. Kiss and others 689
this study are listed in Table 3. Newly obtained sequences
were combined with selected sequences from Saenz & Taylor
(1999), Takamatsu et al. (1999), and Mori et al. (2000). The
alignments are available upon request (gsaenz!unm.edu). All
gaps were included in the analysis. The data sets were
analysed using the parsimony and neighbour-joining methods
found in PAUP* (version 4±0b3a ; Swofford 1999). Parsimony
analysis was conducted using a heuristic search using the Tree
Bisection Reconnection (TBR) branch swapping option and
employing the random stepwise addition of sequences option
beginning with a random seed number and 100 replications.
MaxTrees was set to auto-increase, saving all optimal trees
and swapping on all trees. Distance analysis was performed
using the neighbour-joining method. Pairwise distances
between taxa was determined by using both the Kimura two-
parameter (K2P ; Kimura 1980) model for nucleotide substi-
tutions, and the Hasegawa-Kishino-Yano (HKY85; Hasegawa
et al. 1985) model for nucleotide substitutions and unequal
base frequencies. Branch support was determined by bootstrap
analysis (Felsenstein 1985) using 1000 replications. Decay
indices or Bremer support (Bremer 1988) was determined by
Autodecay version 4.0 (Eriksson 1998). Maximum likelihood
analyses were conducted using Phylip 3.572 (Felsenstein
1993). The transition}transversion (ts}tv) ratios for each data
set were estimated using PAUP*. These ts}tv ratio numbers
were then used in the Phylip maximum likelihood (DNAML)
analyses. The DNAML settings included searching for the
best tree, using global rearrangements, randomising the input
order, and jumbling ten times.
Crossing different isolates in vitro
Six different monoconidial tomato powdery mildew isolates
(Et1, Et11, Et22, Et24, Et25, BP-P1), maintained in vitro at
INRA (Table 1), were paired with each other in all possible
combinations on surface disinfested Lagenaria ciceracia
cotyledons on MS agar as described in Bardin et al. (1997) for
cucumber powdery mildew isolates. Cotyledons were ex-
amined 3–4 weeks after inoculation for cleistothecia pro-
duction.
RESULTS
Microscopic observations
Microscopic examination of tomato powdery mildew patho-
gens clearly showed that all the European, North American,
African and Caribbean isolates, maintained alive in different
laboratories, and also all the herbarium specimens obtained
from Europe, North and South America and Asia were
anamorphs of Pseudoidium because conidia always matured
singly on the conidiophores (Fig. 1). Surprisingly, all the
herbarium specimens obtained from Australia represented
another anamorph where conidia clearly matured in chains
and thus belonged to catenate Oidium species (Fig. 2). Striking
differences were also observed in the shape and size of both
conidia and germ tubes of conidia, structure of the
conidiophores and shape of hyphal appressoria of these two
tomato powdery mildew anamorphs (Tables 1–2).
1 2
3 4
Figs 1–4. Light microscope micrographs of Oidium spp. removed
from herbarium specimens of tomato after rehydration in lactic acid.
Fig. 1. Conidiophore of an Oidium species (O. subgen. Pseudoidium)
here re-described as O. neolycopersici, collected in Malaysia in 1947
(IMI 24738). Bar¯ 30 µm. Fig. 2. Conidiophores of a catenate
Oidium species (O. subgen. Reticuloidium), here neotypified as
O. lycopersici, collected in South Australia in 1980 (DAR 35763).
Bar¯ 40 µm. Fig. 3. Germinating conidium of O. neolycopersici from
1964 (IMI 106600). Bar¯ 10 µm. Fig. 4. Germinating conidia of
O. lycopersici from 1994 (DAR 70008). Bar¯ 30 µm.
All the anamorphs of Pseudoidium studied were characterised
by ellipsoid-ovoid or doliform conidia with mean conidial
lengths generally greater than 30 µm and germ tubes
terminating in lobed, or rarely simple, apices (Fig. 3). In
contrast, conidia of all the Australian catenate Oidium species
were smaller, with mean conidial lengths less than 30 µm.
These conidia were cylindrical, with one, or sometimes two,
germ tubes terminating in simple apices (Fig. 4). Conidial
lengths and widths for all the specimens are shown in Tables
1–2.
Differences were also observed in the structure and
dimensions of the conidiophores. First, in the catenate species
the basal septum of the foot cells was always displaced at a
distance of 7–27 µm from the point of branching. In
Pseudoidium species, such displacement of the basal septa was
not observed. Second, the total length of the conidiophores in
Pseudoidium species measured 58–115 µm (mean : 91±5 µm).
They consisted of 2–3 cells attached to a 29–74 µm (mean :
42±4 µm) long foot-cell that was either cylindrical or sometimes
inflated in the middle and constricted at the base. Conidia
were always produced singly, but sometimes remained
attached to each other in high relative humidity forming
‘pseudo-chains ’ of 2–6 conidia. In catenate anamorphs, the

Powdery mildew fungi on tomato 690
5 6
87
Figs 5–8. Scanning electron micrographs of herbarium specimens of O. (subgen. Reticuloidium) lycopersici, from Australia. Bars¯ 5 µm.
Fig. 5. Conidia in a chain. Fig. 6. Fibrillar pattern on end wall of conidium. Fig. 7. Conidium with a polygonal, reticulate pattern of
wrinkling of outer wall. Fig. 8. Simple, unlobed appressoria (indicated by arrows) on hypha.
total length of the conidiophores was greater, 150–210 µm
(mean : 185±7 µm), measured from the point of branching, with
a long, 100–145 µm (mean : 124±4 µm), foot-cell followed by
2–4 shorter cells. Conidia were produced in real chains
consisting of 3–7 conidium initials and mature conidia.
Hyphal appressoria were also different. In the Pseudoidium
anamorphs, they were as expected, simple to lobed or even
multi-lobed, opposite or spread along the hyphae. They
contrasted with the inconspicuous or nipple-shaped appres-
soria situated singly or in sequence in the Australian catenate
anamorphs.
Scanning electron microscopy
Cook et al. (1997) have shown that herbarium specimens,
though not as informative as fresh material, can reveal useful
information under the SEM. For the Australian herbarium
material, the SEM study revealed long conidial chains (Fig. 5)
whose septa showed a fibrillar pattern (Fig. 6) and whose
outer walls bore polygonal}reticulate wrinkling patterns (Fig.
7). Simple, inconspicuous unlobed appressoria were found
with difficulty on the hyphae (Fig. 8). These features are
diagnostic for one of the catenate Oidium species described as
O. subgen. Reticuloidium (Cook et al. 1997).
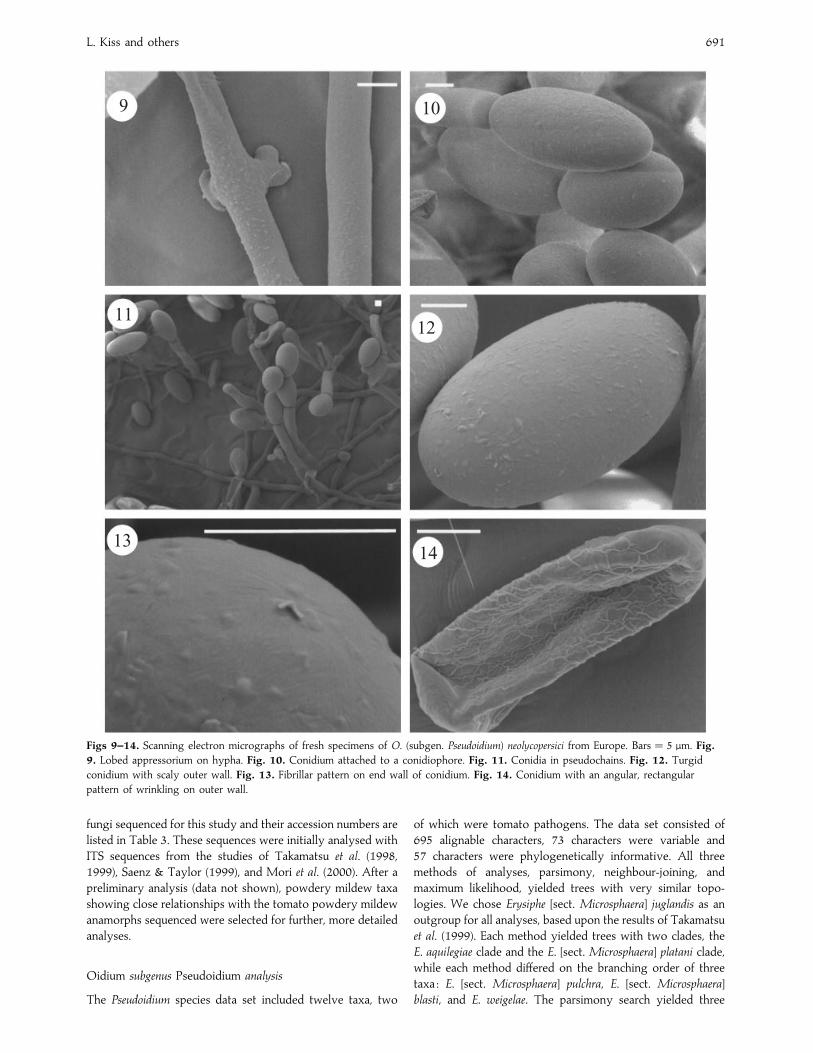
For the Pseudoidium species, two fresh isolates, one from
Hungary, BP-P4, and the other from England, UK-1, were
examined under the SEM. In contrast to the Australian
pathogens, the appressoria were conspicuous and moderately
lobed (Fig. 9). A close examination of the nature of the
conidiogenesis confirmed that conidia matured singly in these
fungi (Fig. 10), as also revealed by light microscopy (Fig. 1).
However, conidia produced singly sometimes remained
attached to each other forming pseudo-chains consisting of
2–3 (Fig. 11), rarely 6 or more conidia. This was also observed
under the light microscope in all the fresh materials studied.
The outer walls of turgid conidia were smooth to somewhat
scaly (Fig. 12), and the septa had a fibrillar pattern (Fig. 13).
When wrinkled, outer walls developed an angular}rectangular
pattern (Fig. 14). Apart from a somewhat smoother outer wall
than usual, this set of patterns is sufficiently diagnostic for O.
subgen. Pseudoidium (Cook et al. 1997, Braun 1999), now the
sole anamorph for the redescribed holomorph genus Erysiphe
sensu stricto (Braun et al. 2001, Braun & Takamatsu 2000).
Thus, the SEM study confirmed that the non-catenate and
catenate anamorphs seen under the light microscope clearly
represent two different Oidium species which can infect
tomato. All the European, Asian, American and African
specimens studied fell into the anamorph genus O. subgen.
Pseudoidium, whereas all the Australian specimens studied fell
into the genus O. subgen. Reticuloidium (a more precise taxon
than the heterogeneous ‘euoidium ’).
Molecular phylogenetic studies
A total of three rDNA ITS sequences of powdery mildew
anamorphs infecting tomato were determined. The data of the
L. Kiss and others 691
9 10
1112
1413
Figs 9–14. Scanning electron micrographs of fresh specimens of O. (subgen. Pseudoidium) neolycopersici from Europe. Bars¯ 5 µm. Fig.
9. Lobed appressorium on hypha. Fig. 10. Conidium attached to a conidiophore. Fig. 11. Conidia in pseudochains. Fig. 12. Turgid
conidium with scaly outer wall. Fig. 13. Fibrillar pattern on end wall of conidium. Fig. 14. Conidium with an angular, rectangular
pattern of wrinkling on outer wall.
fungi sequenced for this study and their accession numbers are
listed in Table 3. These sequences were initially analysed with
ITS sequences from the studies of Takamatsu et al. (1998,
1999), Saenz & Taylor (1999), and Mori et al. (2000). After a
preliminary analysis (data not shown), powdery mildew taxa
showing close relationships with the tomato powdery mildew
anamorphs sequenced were selected for further, more detailed
analyses.
Oidium subgenus Pseudoidium analysis
The Pseudoidium species data set included twelve taxa, two
of which were tomato pathogens. The data set consisted of
695 alignable characters, 73 characters were variable and
57 characters were phylogenetically informative. All three
methods of analyses, parsimony, neighbour-joining, and
maximum likelihood, yielded trees with very similar topo-
logies. We chose Erysiphe [sect. Microsphaera] juglandis as an
outgroup for all analyses, based upon the results of Takamatsu
et al. (1999). Each method yielded trees with two clades, the
E. aquilegiae clade and the E. [sect. Microsphaera] platani clade,
while each method differed on the branching order of three
taxa : E. [sect. Microsphaera] pulchra, E. [sect. Microsphaera]
blasti, and E. weigelae. The parsimony search yielded three

Powdery mildew fungi on tomato 692
Erysiphe polygoni ex Polygonum, United States
1
E. convolvuli ex Convolvulus, United States
E. cruciferarum ex Arabidopsis, United Kingdom
Erysiphe* platani ex Platanus, United States
E. weigelae ex Weigela, Japan
E. aquilegiae ex Cimifuga, Japan
E. macleayae ex Macleaya, Japan
Oidium neolycopersici ex Lycopersicon, France
O. neolycopersici ex Lycopersicon, The Netherlands
E.* blasti ex Lindera, Japan
E.* pulchra ex Cornus, Japan
E.* juglandis ex Pterocarpa, Japan
5 NucleotideSubstitutions
2
479
2
69
2
8
100
286
1
Fig. 15. Phylogenetic analysis of the internal transcribed spacer (ITS) region of the ribosomal RNA gene for 12 Pseudoidium anamorphs.
The tree is one of three most parsimonious trees, which was found using a heuristic search employing the random stepwise addition
option of PAUP* phylogenetic analysis (Swofford 1999). The tree also has the highest likelihood of the three most parsimonious trees.
Percentages of parsimony analysis of 1000 bootstrapped data sets supporting specific branches are given below the branches. Branches
& 95% are strongly supported and are shown in boldface (Felsenstein 1985). Bootstrap values below 50% are not shown. Decay
indices or Bremer support (Bremer 1988) is shown above the branches. Erysiphe *juglandis was used to root the tree. Powdery mildews
on tomato are shown in boldface. The numbers adjacent to the right of the tree correspond to clades discussed in the text : 1¯ the E.
platani clade, 2¯ the Erysiphe aquilegiae clade. Taxa marked with *, indicate genera formerly known as Microsphaera.
equally most parsimonious trees (MPT). The trees were 200
steps in length with a consistency index (CI) of 0±770, a
retention index (RI) of 0±620, and a rescaled consistency index
(RC) of 0±477 (Farris 1989). The three trees were the result of
the inclusive branching order of E. polygoni and E. blasti. Two
supported clades resulted from the bootstrap analysis, the
E. aquilegiae clade was strongly supported (100%), and the
E. platani clade received minor support (69%) (Fig. 15). The E.
aquilegiae clade (Fig. 15) consisted of E. aquilegiae, E. macleayae,
and the two O. subgen. Pseudoidium tomato powdery mildews,
one specimen from France and the other from Netherlands.
The two Pseudoidium isolates from tomato were well supported
(86%) as sister taxa, and were nested within the E. aquilegiae
clade. The E. platani clade consisted of E. platani, E. cruciferarum,
E. convolvuli, and E. polygoni. Decay analysis, or Bremer
support, gave strong support to the E. aquilegiae clade (8),
while the E. platani clade received minor support (2). The clade
containing the two Pseudoidium isolates from tomato received
minor support (2). Both neighbour-joining analyses, utilising
either the K2P or the HKY85 models of nucleotide
substitutions, yielded identical trees (trees not shown).
Bootstrap support indices for each neighbour-joining analysis
were also nearly identical, and the overall support for the
E. platani clade and the E. aquilegiae clade, including internal
branches, increased as compared to the parsimony bootstrap
indices. The E. aquilegiae clade was again strongly supported
(100%), while the support for the E. platani clade increased
from 69% (parsimony) to 87%. The two Pseudoidium isolates
from tomato were again well supported (87%) as sister taxa,
while E. macleayae was strongly supported (99%) as a close
relative. The maximum likelihood analysis yielded a tree very
similar to the parsimony and neighbour joining analyses, in
that both the E. aquilegiae and E. platani clades appeared.
Catenate Oidium (O. subgenus Reticuloidium) analysis
The catenate Oidium data set contained fifteen taxa, including
a tomato powdery mildew specimen from Australia. The data

L. Kiss and others 693
Oidium longipes ex Solanum “eggplant”, Switzerland 1
Golovinomyces* magnicellulatus ex Phlox, United States
Arthrocladiella mougeotii ex Lycium, Japan
G.* sordidus ex Plantago, United States
5 Nucleotide changes per1000 positions
2
3
O. lycopersici ex Lycopersicon tomato, Australia
G.* orontii ex Delphinium, United States
G.* orontii ex Agastache, United States
G.* orontii ex Nicotiana tobacco, Hungary
G.* cichoracearum ex Arabidopsis, United States
G.* orontii ex Nicotiana tobacco, Japan
G.* cichoracearum ex Cucurbita “zucchini”, France
G.* orontii ex Arabidopsis, United States
G.* cichoracearum ex Cucurbita “zucchini”, France
G.* cichoracearum ex Eupatorium, Japan
84
54
100
100
100
86
100
53
96
65
94
63
G.* biocellatus ex Monarda, United States
Fig. 16. Neighbour joining analysis of the ITS region of the ribosomal RNA gene for 15 catenate Oidium species belonging to
subgenus Reticuloidium. The neighbour joining tree was found using the Kimura 2-parameter model for nucleotide substitutions. Branch
lengths correspond to pairwise distances between taxa. Branch support was determined by 1000 bootstrapped data sets, shown above
the branches. Branches¯ 95% are shown in bold. Arthrocladiella mougeotii was used to root the tree. Powdery mildews on tomato are
shown in boldface. The numbers adjacent to the right of the tree correspond to clades discussed in the text : 1¯ the Golovinomyces*
magnicellulatus clade, 2¯ the G.* biocellatus clade, 3¯ the G.* orontii clade. Taxa marked with *, indicate genera formerly known as
Erysiphe. Note that the three sequences labelled G. cichoracearum in clade 3 are more likely to have come from isolates of G. orontii (see
Table 3).
set consisted of 629 alignable characters, 42 characters were
variable and 59 characters were phylogenetically informative.
All three methods of analyses, parsimony, neighbor-joining,
and maximum likelihood, yielded trees with nearly identical
topologies. We chose Arthrocladiella mougeotii (anamorph :
O. subgen. Graciloidium) as an outgroup for all analyses, based
upon the results of Mori et al. (2000). Each method yielded
trees with three clades (containing anamorphs of O. subgen.
Reticuloidium species) : the Golovinomyces magnicellulatus clade
(note that Golovinomyces was formally referred to as Erysiphe),
the G. biocellatus clade, and the G. orontii clade. The G.
magnicellulatus clade consisted of G. magnicellulatus, and two
solanaceous Oidium species : O. longipes from eggplant in
Switzerland, and O. lycopersici from tomato in Australia. The
G. biocellatus clade consisted of G. biocellatus and two G. orontii
isolates. The G. orontii clade consisted of three G. orontii
isolates, three G. cichoracearum isolates, believed to be G.
orontii also, and a G. sordidus isolate. The parsimony search
yielded 18 equally most parsimonious trees (MPT). The trees
were 147 steps in length with a consistency index (CI) of
0±816, a retention index (RI) of 0±827, and a rescaled
consistency index (RC) of 0±675. The 18 trees were the result
of the inclusive branching order of the seven taxa in the G.
orontii clade (trees not shown). Three supported clades
resulted from the bootstrap analysis, the G. magnicellulatus
clade received moderate support (78%). The two solanaceous
Oidium species within the G. magnicellulatus clade were
strongly supported as sister taxa (100%). The G. biocellatus
clade was strongly supported (100%). The G. orontii clade
was well supported (94%). However, the branching order of
these closely related powdery mildews was inclusive. Both
neighbour-joining analyses, utilising either the K2P or the
HKY85 models of nucleotide substitutions, yielded identical
trees (Fig. 16). Bootstrap support indices for each neighbour-
joining analysis were also nearly identical, and the overall
support for the three clades, including internal branches,

Powdery mildew fungi on tomato 694
increased as compared to the parsimony bootstrap indices.
Using the K2P distance model, the bootstrap support for the
G. magnicellulatus clade increased from 77% (parsimony) to
84%, and again the two solanaceous Oidium species were
strongly supported (100%) as sister taxa. The G. biocellatus
clade was again strongly supported (100%). The support for
the G. orontii clade increased from 93% (parsimony) to 100%,
with the branching order in the K2P tree nearly duplicating
the branching order in the MPT with the highest likelihood.
The maximum likelihood analysis yielded a tree with topology
nearly identical to the parsimony and neighbour joining
analyses (tree not shown).
Crossing different isolates in vitro
No cleistothecia or cleistothecium initials were produced in
any pairs of the Pseudoidium isolates crossed in vitro.
Cleistothecia have never been reported for either the
O. subgen. Pseudoidium anamorphs infecting tomato (Whipps
et al. 1998) or O. subgen. Reticuloidium (Price 1981, Wicks &
Clare 1981).
DISCUSSION
To the best of our knowledge, this study includes all the
known tomato powdery mildew material, available either as
herbarium specimens deposited in international collections or
as living isolates maintained on tomato in different labora-
tories. Their identity was investigated using morphological
characteristics, SEM conidial surface patterns and molecular
analysis. The results show that, in spite of the many reports
of catenate Oidium species being involved, all the recent
outbreaks of tomato powdery mildew reported from outside
Australia were caused by a single anamorph belonging to
Oidium subgen. Pseudoidium. In contrast, the causal agent of
the Australian tomato powdery mildew was always an
O. subgen. Reticuloidium species which curiously was not found
on any tomato outside Australia.
Despite an editorial policy requiring the deposition of
herbarium specimens (Rossman 1985), none of the authors
reporting catenate Oidium species as causal agents of tomato
powdery mildew outside Australia deposited any voucher
specimens in international collections. Reports of such
anamorphs from outside Australia could not be investigated
such as those from Japan (Abiko 1978), Switzerland (Corbaz
1990, 1993), Bulgaria (Neshev 1993), Canada (Be! langer &
Jarvis 1994), India (Kumar et al. 1995), the USA (Arredondo et
al. 1996, Karasevitz & Zitter 1996, White et al. 1997, Pernezny
& Sonoda 1998), etc. In order to overcome the lack of
specimens, we obtained fresh or herbarium specimens from
many geographical regions (see Tables 1–2) and found only
Pseudoidium anamorphs in them. Moreover, we have re-
examined the tomato powdery mildew pathogens reported as
catenate Oidium species from Hungary (Kiss 1996) and the UK
(Cook et al. 1997) and found only Pseudoidium anamorphs.
Similarly, Bolay (1998) reported that only a Pseudoidium and
never a catenate Oidium species has been found on tomato in
Switzerland, contrary to Corbaz (1990, 1993) who reported
E. cichoracearum. As suggested by Whipps et al. (1998) and
Lebeda & Mieslerova (1999), authors reporting catenate
Oidium species outside Australia may have been misled by the
pseudo-chains of a Pseudoidium produced in high relative
humidity.
Molecular phylogenetic studies completely corroborated
the morphological data. Recent phylogenetic analysis of
powdery mildew ITS sequence data (Saenz & Taylor 1999,
Mori et al. 2000) fully supported the revised anamorphic
taxonomy of Cook et al. (1997), and further drew attention to
the importance of the anamorphic state in determining the
evolutionary relationships within the Erysiphaceae. Preliminary
analysis of the three tomato powdery mildew specimens
clearly showed that the Australian catenate Oidium species
was not phylogenetically related to the two isolates of a
Pseudoidium. Based on these results, two data sets were created
to analyze each phylogenetic lineage separately.
Phylogenetic analyses of the Pseudoidium isolates, including
the two from tomato, resulted in two supported clades : the
E. aquilegiae clade and the E. [sect. M.] platani clade (Fig. 15).
The two specimens of Pseudoidium from tomato, collected in
France and The Netherlands, grouped as sister taxa within the
E. aquilegiae clade as their rDNA ITS sequences were nearly
identical ; this was also reported by Jones et al. (2000). Another
molecular study analysing the amplified fragment length
polymorphism (AFLP) in five Pseudoidium isolates collected
from tomato in three different European countries revealed
only minor genetic differences among them (Huang, Lindhout
& Niks 1998). These results suggest that there is little genetic
variability among the Pseudoidium anamorphs causing the
recent outbreaks of tomato powdery mildew world-wide.
Phylogenetic analyses of the catenate Oidium species
resulted in three supported clades : the G. magnicellulatus clade,
the G. biocellatus clade, and the G. orontii clade (Fig. 16). The
Australian O. lycopersici grouped strongly with another
solanaceous powdery mildew, O. longipes, within the G.
magnicellulatus clade. These results confirm that the anamorph
of the Australian tomato powdery mildew belongs to O.
subgen. Reticuloidium. The unexpectedly close phylogenetic
relationship between the Australian O. lycopersici on tomato
and the Swiss O. longipes on eggplant might be reflected by
similarities in their morphology. Both have very long
conidiophores with displaced basal septa. However, it is
the foot cells of the conidiophore that are very long in
O. lycopersici and the second and}or the third cells that are
exceptionally long in O. longipes. According to Noordeloos &
Loerakker (1989), O. longipes cannot infect tomato.
In order to examine the phylogenetic relationship between
G. orontii infecting tobacco world-wide and the Australian
catenate Oidium species on tomato, Japanese and Hungarian
tobacco powdery mildew ITS sequences were included in our
phylogenetic analysis (Table 3). As expected from the
differences between their morphology, the tobacco and the
Australian tomato powdery mildew fungi grouped in two
different clades (Fig. 16). Tobacco was repeatedly reported to
be infected with tomato powdery mildew in host range
studies (e.g. Fletcher et al. 1988, Corbaz 1990, 1993, Smith et
al. 1997, Huang et al. 2000), but Whipps et al. (1998)
suggested that these findings might have resulted from a
contamination of tobacco with G. orontii. In our trials in a

L. Kiss and others 695
greenhouse and a climate chamber no cross-infection was
observed between G. orontii on tobacco and the Pseudoidium
species on tomato (Kiss 1996).
That the Australian tomato powdery mildew anamorph
belongs to sect. Reticuloidium, and not to sect. Pseudoidium,
raises nomenclatural problems. The Pseudoidium species was
redescribed as O. lycopersicum by Noordeloos & Loerakker
(1989) based on the original species description (Cooke &
Massee 1888), and the now lost holotype, the only type
specimen available for this name. Without being able to
determine the nature of the conidiogenesis, Noordeloos &
Loerakker (1989) considered that the Australian O. lycopersicum
was a Pseudoidium species compatible with the tomato
pathogen found in Dutch greenhouses in the eighties. In
contrast, our studies indicated that only Reticuloidium
anamorphs occur on tomato in southern Australia. Comparison
of the morphology of the Australian tomato powdery mildew
fungus with the illustration of Noordeloos & Loerakker (1989)
shows several points of comparison. Firstly, the conidiophores
are rather long and have strongly displaced basal septa in both
cases. Secondly, the mycelial appresoria are clustered, and
essentially unlobed. Finally, the type locality of O. lycopersicum
falls within south-eastern Australia, from which only a
Reticuloidium is known on tomato. This is considered sufficient
grounds to redescribe and neotypify Cooke & Massee’s
species below, at the same time correcting its orthography to
O. lycopersici. The neotype is chosen partly because it was
collected within 200 km of the lost holotype in the state of
Victoria, and also because it is the specimen sequenced in the
present study.
Oidium lycopersici Cooke & Massee, Grevillea 16 : 114(1888).
Colonies occurring on the abaxial and adaxial surfaces of
leaves, on petioles, stems and calyces of tomato. Mycelium
either conspicuous and white in some specimens, or extremely
sparse and associated with a hypersensitive necrotic spot in
other specimens. Hyphae straight to undulate, 4–6 µm wide,
hyphal cells 51–66 µm long. Mycelial appressoria well
developed, simple nipple-shaped or weakly lobed, single or
aggregated in serial clusters of up to 8 appressoria.
Conidiophores inserted medianly on the conidiophore mother
cell, moderately to extremely long, basal septum displaced by
15–45 µm, conidiophore stipe consisting of a single long cell
95–180 µm long, occasionally two cells, the basal cell usually
longer than 80 µm, stipe attenuated downwards, 9–10 µm
wide at the basal septum, 10–12 µm wide at the apical septum.
Conidia borne in chains of 3–5 conidia, edge line sinuate,
conidia lacking fibrosin bodies, elliptical to doliform to sub-
cylindrical, falling into two distinct classes, the majority of
conidia small, 25–32¬14–16 µm (in lactic acid mounts), mean
less than 30 µm long, but with a small proportion of much
larger conidia ranging from 35–45 µm long (believed to be
‘double conidia ’ in which a delimiting septum has failed to
form – such conidia germinate with germ-tubes from the
middle as well as the ends). Small conidia in water mounts
from fresh specimens are of similar length but up to 22 µm
wide and broadly ellipsoid. Conidia in lactic acid mounts from
dried herbarium specimens (Shin & La 1993) narrower
(14–16 µm wide) and sub-cylindrical. Primary germ tubes on
host tissue and glass arising from the shoulder, germ tubes
simple, clavate, 23–30¬6–8 µm, basal septum displaced by
2–8 µm. Secondary germination arising either from the body
of the conidium or from the base of the primary germ tube,
below the basal septum.
Typus : Australia : ‘Upper Yarra ’, (K-holotypus lost fide curator,
pers comm.) ; Victoria : Timmering, on L. esculentum cv. ‘Paycetter ’
leaves, 1994, I. Pascoe (VPRI 19847-neotypus hic designatus).
Habitat : On living leaves of Lycopersicon esculentum in
greenhouses, also in the field.
Distribution : Australia.
Molecular data : The nuclear rDNA ITS sequence of the
neotype is deposited in GenBank as AF229021.
Additional specimens examined : see Table 2. Morphological studies
were also carried out on over 50 additional specimens (deposited at
VPRI) collected from south eastern Australia (Pascoe, unpubl.).
Oidium neolycopersici L. Kiss, sp. nov.
Etym. : neo-, new; and from the host Lycopersicon esculentum.
Mycelium album, supra, interdum infra folia in statu vivo, rare in
caulibus crescens. Appressoria lobata vel multilobata, opposita vel
effusa dissita. Conidiophora 58–(91±5)–115 µm longa, cellulae basis
cylindricae, 29–(42±4)–74 µm longae. Conidia 22–(33±5)–46¬10–
(13±7)–20 µm, ellipsoidea-ovoidea vel doliiformia, singula aut si
humida 2–6 in pseudocatenas, sine corpore fibrosina.
Mycelium white, thin, covering the upper and occasionally the
lower surfaces of the leaves, and also the stems. Hyphae
hyaline, septate, 4–8 µm wide, appressoria distinct, lobed to
multi-lobed, rarely nipple-shaped, opposite or dispersed.
Conidiophores erect, 58–115 µm (mean : 91±5 µm) long, foot-
cells cylindrical or sometimes inflated in the middle and
constricted at the base, 29–74 µm (mean : 42±4 µm) long,
followed by 1–2 shorter cells or a single cell of about the same
length. Conidia produced singly, or, in high relative humidity,
in pseudo-chains of 2–6 conidia, 22–46 (mean : 33±5)¬10–20
(mean : 13±7) µm, ellipsoid-ovoid or doliform, without fibrosin
bodies, germ tubes arising from an end or side of the
conidium, substraight or curved, apex enlarged, lobed, rarely
simple, germ tubes increasing in width from base to top.
Typus : France : Aigue Mortes, on Lycopersicon esculentum, 1989,
P. C.Nicot Et1 (BPI 747013-holotypus, HAL-isotypus). Living material
maintained at INRA, Unite! de Pathologie Ve! ge! tale, Avignon, France.
Habitat : On living leaves and stems of Lycopersicon
esculentum in greenhouses, also in the field.
Distribution : Nearly circumglobal wherever tomatoes are
grown.
Molecular data : The nuclear rDNA ITS sequence of isolate
Et1 was deposited in GenBank under the accession number
AF229019.
Additional specimens examined : See Tables 1–2.
This widespread new species has long been confused with the
geographically restricted O. ‘ lycopersicum ’ Cooke & Massee

Powdery mildew fungi on tomato 696
1888 (corrected to O. lycopersici by Lebeda & Mieslerova
1999). At present, no species name exists for the Pseudoidium
anamorph infecting tomato and so we have provided a formal
description and new name here. Although most data in the
recent phytopathological literature suggested that O. neolyco-
persici had arisen fairly recently on tomato (e.g. Whipps et al.
1998), our data show that this is not the case. We have studied
herbarium specimens collected since 1947 (Table 1) which can
now be clearly identified as O. neolycopersici (Fig. 1). So, this
species has been infecting tomato for a long time, at least in
Asia, the origin of all the herbarium specimens older than
50 yr. Unfortunately, we could not obtain any European
herbarium specimens collected before the first European
outbreaks of tomato powdery mildew (Blancard 1988, Fletcher
et al. 1988), although the disease was repeatedly reported in
the mycological literature since 1900. So, we could not
determine if the pathogens reported as anamorphs of Erysiphe
spp. infecting tomato in different European countries (Salmon
1900, Jaczewski 1927, Blumer 1933, 1967, Hammarlund 1945)
were identical with O. neolycopersici. Nor could we obtain any
specimens of Oidium on tomato reported from Sri Lanka
(Petch 1922), Peru (Abbott 1931) and Argentina (Godoy
1939).
Oidium anamorphs on tomato, although poorly identified in
the older literature, sometimes caused severe problems in
different parts of the world long before the recent outbreaks
(e.g. Godoy 1939). However, there is only one doubtful
record on tomato seedlings (Farr et al. 1989) in North America
before the 1990s (Be! langer & Jarvis 1994) suggesting that O.
neolycopersici, now widespread in both the USA and Canada,
was introduced to North America only very recently.
This study has successfully brought together supporting
evidence from entirely separate areas of expertise, i.e. those
involving classical morphological, molecular and SEM tech-
niques. Thus, the joint use of these three methods is now
highly recommended for the precise identification of any
newly discovered powdery mildew anamorphs.
ACKNOWLEDGEMENTS
The authors are grateful to the following scientists for providing fungal
materials : Adrien Bolay, Raymond Cerkauskas, Mike Davis, David M.
Gadoury, Cai-Cheng Huang, David Kalb, Margaret T. McGrath, Nina
Shishkoff, and Victoria L. Smith. The curators of IMI and DAR, John C. David
and Michael Priest, respectively, are also acknowledged for the loan of
materials. We are especially indebted to Uwe Braun for helpful suggestions
on the taxonomic aspects. We also thank Jim Plaskowitz for assistance in
preparing Figs 1–4. The studies of L. Kiss were partly supported by a
Fulbright fellowship and a Ja! nos Bolyai fellowship and a grant (OTKA
F032931) of the Hungarian Scientific Research Fund and those of R. T. A.
Cook by the Plant Health Divison of the Ministry of Agriculture, Fisheries
and Food, UK. G. S. Saenz wishes to thank Laura Salter for advice with
Maximum Likelihood analysis, and Donald O. Natvig and John W. Taylor, for
access to computers and phylogenetic programs. Much of the morphological
study of over 50 specimens of O. lycopersici was carried out by Siva Sivapalan
and Vyrna Beilharz (Institute for Horticultural Development, Knoxfield,
Australia).
REFERENCES
Abbott, E. V. (1931) Further notes on plant diseases in Peru. Phytopathology
21 : 1061–1071.
Abiko, K. (1978) Influence of temperature and humidity on development of
tomato powdery mildew. Proceedings of the Kansai Plant Protection Society
20 : 49–52.
Arredondo, C. R., Davis, R. M., Rizzo, D. M. & Stahmer, R. (1996) First report
of powdery mildew of tomato in California caused by an Oidium sp. Plant
Disease 80 : 1303.
Bains, P. S., Bennypaul, H. & Mirza, M. (1999) First report of powdery mildew
of greenhouse-grown tomatoes in Alberta, Canada. Plant Disease 83 : 488.
Bardin, M., Nicot, P. C., Normand, P. & Lemaire, J. M. (1997) Virulence vari-
ation and DNA polymorphism in Sphaerotheca fuliginea, causal agent of
powdery mildew of cucurbits. European Journal of Plant Pathology 103 :
545–554.
Be! langer, R. R. & Jarvis, W. R. (1994) Occurrence of powdery mildew
(Erysiphe sp.) on greenhouse tomatoes in Canada. Plant Disease 78 : 640.
Blancard, D. (1988) Maladies de la Tomate, observer, identifier, lutter. INRA
Editions, Paris.
Blumer, S. (1933) Die Erysiphaceen Mitteleuropas mit besonderer Beru$ ck-sichtigung der Schweiz. Beitrage zur Kryptogamenflora der Schweiz 7(1) :
1–483.
Blumer, S. (1967) Echte Mehltaupilze (Erysiphaceae). Fisher, Jena.
Boesewinkel, H. J. (1980) The morphology of the imperfect states of powdery
mildews (Erysiphaceae). Botanical Review 46 : 167–224.
Bolay, A. (1998) Les oidiums de la tomate et de l’aubergine en Suisse. Revue
suisse de Viticulture, Arboriculture et Horticulture 30 : 373–378.
Braun, U. (1987) A monograph of the Erysiphales (powdery mildews). Beihefte
zur Nova Hedwigia 89 : 1–700.
Braun, U. (1995) The Powdery Mildews (Erysiphales) of Europe. Gustav Fischer,
Jena.
Braun, U. (1999) Some critical notes on the classification and generic concept
of the Erysiphaceae. Schlechtendalia 3 : 49–55.
Braun, U., Cook, R. T. A., Inman, A. J. & Shin, H.-D. (2001) The taxonomy of
the powdery mildew fungi. In The Powdery Mildews: a comprehensive treatise
(R. R. Be! langer, A. Dik & W. Bushnell, eds) : in press. American
Phytopathological Society Press, St Paul, MN.
Braun,U. &Takamatsu, S. (2000) Phylogeny of Erysiphe,Microsphaera,Uncinula
(Erysipheae) and Cystotheca, Podosphaera, Sphaerotheca (Cystotheceae) inferred
from rDNA ITS sequences – some taxonomic consequences. Schlechtendalia
4 : 1–33.
Bremer, K. (1988) The limits of amino acid sequence data in angiosperm
phylogenetic reconstruction. Evolution 42 : 795–803.
Burgerjon, A., Nicot, P. C., Bertrand, F. & Blancard, D. (1990) Early powdery
mildew of greenhouse-grown tomatoes in France. Phytopathology 80 : 1063.
Ciccarese, F., Amenduni, M., Schiavone, D. & Cirulli, M. (1998) Occurrence
and inheritance of resistance to powdery mildew (Oidium lycopersici) in
Lycopersicon species. Plant Pathology 47 : 417–419.
Cook, R. T. A., Inman, A. J. & Billings, C. (1997) Identification and classifica-
tion of powdery mildew anamorphs using light and scanning electron
microscopy and host range data. Mycological Research 101 : 975–1002.
Cooke, M. C. & Massee, G. (1888) Oidium lycopersicum. Grevillea 16 : 114.
Corbaz, R. (1990) L’oidium de la tomate, une maladie nouvelle en Suisse. Revue
suisse de Viticulture, Arboriculture et Horticulture 22 : 159–161.
Corbaz, R. (1993) Extension d’un oidium des Cucurbitace! es (Erysiphe
cichoracearum) a la tomate. Revue suisse de Viticulture, Arboriculture et
Horticulture 25 : 389–391.
Demeke, T. & Adams, R. P. (1992) The effects of plant polysaccharides and
buffer additives on PCR. Biotechniques 12 : 332.
Epinat, C., Pitrat, M. & Bertrand, F. (1993) Genetic analysis of resistance of 5
melon lines to powdery mildews. Euphytica 65 : 135–144.
Eriksson, T. (1998) Autodecay. Version 4.0. Distributed by the author,
Department of Botany, Stockholm University, Stockholm.
Farr, D. F., Bills, G. F., Chamuris, G. P. & Rossman, A. Y. (1989) Fungi on Plants
and Plant Products in the United States. American Phytopathological Society
Press, St Paul, MN.
Farris, J. S. (1989) The retention index and the rescaled consistency index.
Cladistics 5 : 417–419.
Felsenstein, J. (1985) Confidence limits on phylogenies : an approach using
the bootstrap. Evolution 39 : 783–791.
Felsenstein, J. (1993) PHYLIP (Phylogeny Inference Package). Version 3.5.
Distributed by the author, Department of Genetics, University of
Washington, Seattle, WA.

L. Kiss and others 697
Fletcher, J. T., Smewin, B. J. & Cook, R. T. A. (1988) Tomato powdery mildew.
Plant Pathology 37 : 594–598.
Gabler, J., Gerlach, W. & Braun, U. (1990) Epidemisches Auftreten eines
Mehltaus an Tomaten in der DDR. Nachrichtenblatt des Deutschen
Pflanzenschutzdiens (Braunschweig) 42 : 94–95.
Godoy, E. F. (1939) El oidium del tomate. Revista Argentina de Agronomia 6 :
49–52.
Hammarlund, C. (1945) Beitrage zur Revision einiger imperfekter Mehltau-
Arten. Erysiphe polyphaga nov. sp. Botaniska Notiser 1945 : 101–108.
Hammett, K. R. W. (1977) Taxonomy of Erysiphaceae in New Zealand. New
Zealand Journal of Botany 15 : 687–711.
Hasegawa,M., Kishino, H. &Yano, T. (1985) Dating of the human-ape splitting
by a molecular clock of mitochondrial DNA. Journal of Molecular Evolution
22 : 160–174.
Huang, C.-C., Lindhout, P. & Niks, R. E. (1998) Genetic differences in
powderymildews prevailing recently on tomato.Abstracts of the International
Congress of Plant Pathology ’98, Edinburgh, 2.2.18. [Abstract.]
Huang, C.-C., Biesheuvel, J., Lindhout, P. & Niks, R. E. (2000) Host range of
Oidium lycopersici occurring in the Netherlands. European Journal of Plant
Pathology 106 : 465–473.
Jaczewski, A. A. (1927) [Guide Book to the Identification of Fungi.] Leningrad.
[In Russian.]
Jones, H. E., Whipps, J. M., Thomas, B. J., Carver, T. L. W. & Gurr, S. J. (2000)
Initial events in the colonisation of tomatoes by Oidium lycopersici, a
distinct powdery mildew fungus of Lycopersicon species. Canadian Journal of
Botany 78 : 1361–1366.
Karasevicz, D. M. & Zitter, T. A. (1996) Powdery mildew occurrence on
greenhouse tomato plants in New York. Plant Disease 80 : 709.
Kimura, M. (1980) A simple method for estimating evolutionary rates of base
substitutions through comparative studies of nucleotide sequences. Journal
of Molecular Evolution 16 : 111–120.
Kiss, L. (1996) Occurrence of a new powdery mildew fungus (Erysiphe sp.) on
tomatoes in Hungary. Plant Disease 80 : 224.
Kumar, V., Singh, B., Sugha, S. K.&Basandrai, A. K. (1995) Sources of resistance
to tomato powdery mildew. Indian Journal of Mycology and Plant Pathology
25 : 172–174.
Lebeda, A. & Mieslerova, B. (1999) Identification, occurrence and host range of
tomato powdery mildew (Oidium lycopersici) in the Czech Republic. Acta
Phytopathologica et Entomologica Hungarica 34 : 13–25.
Lebeda, A. & Rod, J. (1990) [Padli rajcat – nova nebezpecna choroba.]
Zahradnictvi 15, 313–315. [In Czech.]
Lindhout, P., Pet, G. & van der Beek, H. (1994) Screening wild Lycopersicon
species for resistance to powdery mildew (Oidium lycopersicum). Euphytica
72 : 43–49.
Mieslerova, B. & Lebeda, A. (1999) Taxonomy, distribution and biology of the
tomato powdery mildew (Oidium lycopersici). Journal of Plant Diseases and
Protection 106 : 140–157.
Milotay, P. & Dormanns-Simon, E. (1997) Powdery mildew on tomato in
Hungary and some possible sources of resistance. In Abstracts of the XIII
Meeting of the EUCARPIA Tomato Working Group, Jerusalem : 60. [Abstract.]
Mori, Y., Sato, Y. & Takamatsu, S. (2000) Evolutionary analysis of the powdery
mildew fungi using nucleotide sequences of the nuclear ribosomal DNA.
Mycologia 92 : 74–93.
Neshev, G. (1993) Powdery mildew (Oidium sp.) on tomatoes in Bulgaria.
Phytoparasitica 21 : 339–343.
Noordeloos, M. E. & Loerakker, W. M. (1989) Studies in plant pathogenic
fungi – II. On some powdery mildews (Erysiphales) recently recorded from
the Netherlands. Persoonia 14 : 51–60.
Olalla, L. & Tore! s, J. A. (1998) First report of powdery mildew of tomato
caused by an Erysiphe sp. in Spain. Plant Disease 82 : 592.
Pernezny, K. & Sonoda, R. M. (1998) Powdery mildew of field-grown tomato
in Florida. Plant Disease 82 : 262.
Petch, T. (1922) Additions to Ceylon fungi. II. Annals of the Royal Botanic
Gardens, Peradeniya 7 : 279–322.
Price, T. V. (1981) Powdery mildew of tomato in Australia. Australasian Plant
Pathology 10 : 38–40.
Rossman, A. Y. (1985) Editorial policy should require voucher specimens !
Plant Disease 69 : 187.
Saenz, G. S. & Taylor, J. W. (1999) Phylogeny of the Erysiphales (powdery
mildews) inferred from internal transcribed spacer ribosomal DNA
sequences. Canadian Journal of Botany 77 : 150–168.
Salmon, E. S. (1900) A monograph of the Erysiphaceae. Memoirs of the Torrey
Botanical Club 9 : 1–292.
Shin, H. D. (1988). Erysiphaceae of Korea. PhD thesis, Seoul National University.
Shin, H. D. & La, Y.-J. (1993) Morphology of edge lines of chained immature
conidia on conidiophores in powdery mildew fungi and their taxonomic
significance. Mycotaxon 46 : 445–451.
Smith, V. L., Douglas, S. M. & LaMondia, J. A. (1997) First report of powdery
mildew of tomato caused by an Erysiphe sp. in Connecticut. Plant Disease
81 : 229.
Swofford, D. L. (1999) PAUP * Phylogenetic analysis using parsimony ( * and other
methods). Version 4.0. Sinauer Associates, Sunderland, MA.
Takamatsu, S., Hirata, T. & Sato, Y. (1998) Phylogenetic analysis and predicted
secondary structures of the rDNA internal transcribed spacers of the
powdery mildew fungi (Erysiphaceae). Mycoscience 39 : 441–453.
Takamatsu, S., Hirata, T., Sato, Y. & Nomura, Y. (1999) Phylogenetic relation-
ships of Microsphaera and Erysiphe section Erysiphe (powdery mildews)
inferred from the rDNA ITS sequences. Mycoscience 40 : 259–268.
Vakalounakis, D. J. & Papadakis, A. (1992) Occurrence of a new powdery
mildew of greenhouse tomato in Greece, caused by Erysiphe sp. Plant
Pathology 41 : 372–373.
Walsh, P. S., Metzger, D. A. & Higuchi, R. (1991) ChelexR 100 as a medium
for simple extraction of DNA for PCR-based typing from forensic material.
BioTechniques 10 : 506–513.
Whipps, J. M., Budge, S. P. & Fenlon, J. S. (1998) Characteristics and host range
of tomato powdery mildew. Plant Pathology 47 : 36–48.
White, T. J., Bruns, T., Lee, S. & Taylor, J. (1990) Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR
Protocols : a guide to methods and applications (D. H. Gelfand, J. J. Sninsky &
T. J. White, eds) : 315–322. Academic Press, New York.
White, J. F., Johnston, S. A., Wang, C.-L. & Chin, C.-K. (1997) First report of
powdery mildew in greenhouse-grown tomatoes in New Jersey. Plant
Disease 81 : 227.
Wicks, T. J. & Clare, B. G. (1981) Powdery mildew on tomatoes. Australasian
Plant Pathology 10 : 36–37.
Corresponding Editor : D. T. Mitchell




















