
Relative efficacy of strobilurin formulations in the control of downy mildew of sunflower
10
Relative efficacy of strobilurin formulations in the control of downy mildew of sunflower J. Sudisha • S. R. Niranjana • S. L. Sukanya • R. Girijamba • N. Lakshmi Devi • H. Shekar Shetty Received: 16 July 2009 / Accepted: 12 May 2010 / Published online: 29 May 2010 Ó Springer-Verlag 2010 Abstract The efficacy of three commercial formulations of strobilurins, viz., trifloxystrobin, kresoxim-methyl, and azoxystrobin was evaluated against sunflower downy mil- dew disease caused by Plasmopara halstedii under labora- tory, greenhouse, and field conditions. Complete inhibition of sporangial sporulation, zoospore release, and motility was observed with 2 lg ml -1 in trifloxystrobin, and 5 lg ml -1 for azoxystrobin and kresoxim-methyl. Seed treatment with different concentrations of strobilurins enhanced seed ger- mination and seedling vigor of sunflower to varying degrees compared to control. Highest seed germination was recorded at 10 lg ml -1 in kresoxim-methyl, and maximum seedling vigor was noticed with trifloxystrobin at 30 lg ml -1 . The effect of strobilurins was tested as seed treatment, foliar application, and seed treatment followed by foliar applica- tion. Under greenhouse conditions none of the concentra- tions used, either as seed treatment and foliar application, were phytotoxic. For the three strobilurins, the seed treat- ment along with foliar application enhanced the protection of the plants as compared to only the treatment of seeds. Foliar spray treatments alone provided an intermediate control of the disease. Trifloxystrobin showed a better effect than kresoxim-methyl and azoxystrobin. Disease curative activity of trifloxystrobin was higher compared to kresoxim-methyl and azoxystrobin. Tested fungicides when applied on adaxial leaf surface showed partial translaminar activity, and disease inhibition was marginal. Loss of trifloxystrobin and azox- ystrobin activity over time was low, indicating stable rain- fastness residual activity. A trend in protection against downy mildew disease similar to greenhouse results was evident in the field trials. This is the first report of strobilurins exhibiting high activity against P. halstedii and is a prom- ising fungicide for controlling sunflower downy mildew disease by seed treatment and foliar spray. Keywords Helianthus annuus L. Plasmopara halstedii b methoxyacrylic acid group formulations Seed and foliar treatments Disease management Introduction Sunflower (Helianthus annuus L.), one of the important oil seed crops, faces the problem of crop loss due to diseases caused by microorganisms. Downy mildew (Plasmopara halstedii (Farlow) Berl. & de Toni occurs destructively wher- ever sunflower is grown. Sunflower downy mildew is reported in India, and caused severe epidemics in cvs. Morden and EC 68414 in Maharashtra during 1984–1989, and subsequently, in Karnataka and Andhra Pradesh (Agrawal et al. 1991). It is a soil and seed-borne pathogen. Mayee and Patil (1986) reported a field occurrence of the sunflower downy mildew disease in Marathwada region in Maharashtra state of India, with yield losses up to 80%. Disease management strategies include use of resistant hybrids/cultivars, chemical control, and cultural practices. Sunflower hybrids resistant to downy mildew are available, but new pathogenic races of the fungus are being formed in nature, making questionable the use Communicated by M. Traugott. J. Sudisha S. R. Niranjana (&) S. L. Sukanya R. Girijamba H. Shekar Shetty Downy Mildew Research laboratory, Department of Studies in Biotechnology, University of Mysore, Manasagangotri, Mysore, Karnataka 570006, India e-mail: [email protected] N. Lakshmi Devi Department of Studies in Microbiology, University of Mysore, Manasagangotri, Mysore, Karnataka 570006, India 123 J Pest Sci (2010) 83:461–470 DOI 10.1007/s10340-010-0316-3
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Relative efficacy of strobilurin formulations in the control of downy mildew of sunflower

Relative efficacy of strobilurin formulations in the controlof downy mildew of sunflower
J. Sudisha • S. R. Niranjana • S. L. Sukanya •
R. Girijamba • N. Lakshmi Devi • H. Shekar Shetty
Received: 16 July 2009 / Accepted: 12 May 2010 / Published online: 29 May 2010
� Springer-Verlag 2010
Abstract The efficacy of three commercial formulations
of strobilurins, viz., trifloxystrobin, kresoxim-methyl, and
azoxystrobin was evaluated against sunflower downy mil-
dew disease caused by Plasmopara halstedii under labora-
tory, greenhouse, and field conditions. Complete inhibition
of sporangial sporulation, zoospore release, and motility was
observed with 2 lg ml-1 in trifloxystrobin, and 5 lg ml-1
for azoxystrobin and kresoxim-methyl. Seed treatment with
different concentrations of strobilurins enhanced seed ger-
mination and seedling vigor of sunflower to varying degrees
compared to control. Highest seed germination was recorded
at 10 lg ml-1 in kresoxim-methyl, and maximum seedling
vigor was noticed with trifloxystrobin at 30 lg ml-1. The
effect of strobilurins was tested as seed treatment, foliar
application, and seed treatment followed by foliar applica-
tion. Under greenhouse conditions none of the concentra-
tions used, either as seed treatment and foliar application,
were phytotoxic. For the three strobilurins, the seed treat-
ment along with foliar application enhanced the protection of
the plants as compared to only the treatment of seeds. Foliar
spray treatments alone provided an intermediate control of
the disease. Trifloxystrobin showed a better effect than
kresoxim-methyl and azoxystrobin. Disease curative activity
of trifloxystrobin was higher compared to kresoxim-methyl
and azoxystrobin. Tested fungicides when applied on adaxial
leaf surface showed partial translaminar activity, and disease
inhibition was marginal. Loss of trifloxystrobin and azox-
ystrobin activity over time was low, indicating stable rain-
fastness residual activity. A trend in protection against
downy mildew disease similar to greenhouse results was
evident in the field trials. This is the first report of strobilurins
exhibiting high activity against P. halstedii and is a prom-
ising fungicide for controlling sunflower downy mildew
disease by seed treatment and foliar spray.
Keywords Helianthus annuus L. � Plasmopara halstedii �b methoxyacrylic acid group formulations �Seed and foliar treatments � Disease management
Introduction
Sunflower (Helianthus annuus L.), one of the important oil
seed crops, faces the problem of crop loss due to diseases
caused by microorganisms. Downy mildew (Plasmopara
halstedii (Farlow) Berl. & de Toni occurs destructively wher-
ever sunflower is grown. Sunflower downy mildew is reported
in India, and caused severe epidemics in cvs. Morden and EC
68414 in Maharashtra during 1984–1989, and subsequently, in
Karnataka and Andhra Pradesh (Agrawal et al. 1991). It is a soil
and seed-borne pathogen. Mayee and Patil (1986) reported a
field occurrence of the sunflower downy mildew disease in
Marathwada region in Maharashtra state of India, with yield
losses up to 80%. Disease management strategies include use
of resistant hybrids/cultivars, chemical control, and cultural
practices. Sunflower hybrids resistant to downy mildew
are available, but new pathogenic races of the fungus are
being formed in nature, making questionable the use
Communicated by M. Traugott.
J. Sudisha � S. R. Niranjana (&) � S. L. Sukanya �R. Girijamba � H. Shekar Shetty
Downy Mildew Research laboratory, Department of Studies
in Biotechnology, University of Mysore, Manasagangotri,
Mysore, Karnataka 570006, India
e-mail: [email protected]
N. Lakshmi Devi
Department of Studies in Microbiology, University of Mysore,
Manasagangotri, Mysore, Karnataka 570006, India
123
J Pest Sci (2010) 83:461–470
DOI 10.1007/s10340-010-0316-3

of formerly resistant cultivars in a particular area
(Molinero-Ruiz et al. 2002). Fungicidal seed treatment is a
common practice to control downy mildews and is deemed
economical. The efficacy of metalaxyl against different
oomycetes is a widely reported phenomenon. Metalaxyl seed
treatment (2 g a.i./kg) has been reported to control downy
mildew in sunflower (Patil et al. 1991). Apron 35 SD (6 g a.i./
kg) seed treatment provides maximum control of the sunflower
downy mildew disease (Melero-vara et al. 1982). Fungicides
with definite systemic and long-lasting properties (e.g., met-
alaxyl or related compounds) are of significance in controlling
the disease. Some authors have reported the rapidity with
which resistance to phenylamide fungicides has arisen in nat-
ural populations of oomycetes (Parra and Ristaino 1998).
Resistance to phenylamide fungicides prior to their commer-
cial use has been detected in Pseudoperonospora cubensis,
Phytophthora infestans, and Plasmopara viticola fungicides
(Gisi et al. 2000). The development of metalaxyl-resistant
strains in oomycetes is attributed mainly to genetic uniformity
of the host cultivar and genetic variability in the pathogen
(Parra and Ristaino 2001). However, over-use of the fungicide
has led to the selection of pathogen strains resistant to pesti-
cides, the occurrence of resistant isolates of the pathogen was
first reported under greenhouse (Oros and Viranyi 1984). The
resistance of the oomycete has been found at doses of metal-
axyl lower than 2 g a. i./kg seed in Hungary, USA, and Turkey
(Gulya et al. 1999; Viranyi et al. 1992) and at commercial doses
of 2 g a.i./kg seed in France and Spain (Albourie et al. 1998;
Molinero-Ruiz et al. 2003). Mefenoxam was marketed 10 years
later than metalaxyl, and it contains the biologically active
enantiomer of the racemic fungicide metalaxyl (Shetty 1998).
Both fungicides are used worldwide against sunflower downy
mildew. Resistance to mefenoxam has also been already
reported in natural populations of the oomycetes Peronospora
parasitica and Phytophthora capsici (Parra and Ristaino 1998;
2001; Lamour and Hausbeck 2000). The existence of resis-
tance to mefenoxam in P. halstedii from Spain as well as from
the USA is reported (Molinero-Ruiz et al. 2005; 2008). It is also
suggested that isolates of P. halstedii with resistance to met-
alaxyl may also be resistant to mefenoxam (Molinero-Ruiz
et al. 2005). Different levels of resistance to metalaxyl and
mefenoxam have been observed in P. halstedii with increasing
doses of both fungicides (Molinero-Ruiz et al. 2008).
Pesticides that are adapted to prevent or cure oomycete
plant pathogens either do not exist or are not economically
feasible for marginal farmers (Attard et al. 2007). These
limitations associated with the routinely used strategies
emphasize a search for alternative chemical for sunflower
downy mildew disease management.
Strobilurins, a new class of fungicidal compounds, were
first discovered or isolated from wood-decaying Basidio-
mycete species (Anke et al. 1977). Strobilurins represent bmethoxyacrylic acid group of natural products, which have
become an integral part of disease management programmes
(Bartlett et al. 2002). Strobilurins belong to a new class of
fungicides with a mode of action that is different from that of
the metalaxyl and mefenoxam (Phenyl amide fungicide
(PAF)), iprovalicarb (Carboxylic acid amides) and cyazof-
amid (phenylimidazole chemical class). Mode of action
studies have shown that strobilurins act against mitochon-
drial cytochrome bc1 at the Qo site (Wiggins and Jager
1993) and iprovalicarb does not affect respiration (Stenzel
et al. 1998). Whereas cyazofamid blocks electron transfer in
the mitochondrial cytochrome bc1 complex by binding the
Qi center of the enzyme (Mitani et al. 2001). The most
exciting characteristics about the strobilurins are their
spectrum of activity (Sauter et al. 1999; Herms et al. 2003).
The PAFs are postulated to be site specific, specifically
inhibiting uridine incorporation into RNA polymerase-1
(Sukul and Spiteller 2000), and it was not surprising during
the late 1970s the application of PAFs build up resistance in
susceptible populations (Davidse et al. 1991).
Strobilurins have broad-spectrum activity against Asco-
mycetes, Basidiomycetes, Fungi Imperfecti, and Oomyce-
tes. These fungicides have also been used for the control of
fungal isolates resistant to other fungicides with novel mode
of action and used in low rates (Bartlett et al. 2002). These
strobilurins proved excellent in yield increase and quality of
agricultural produce (Margot et al. 1998; Bartlett et al.
2002; Beck et al. 2002). The recommended applications
include foliar application, seed treatment, and in-furrow
applications. In a short span of time, they have represented
10% of the global fungicidal market (Bartlett et al. 2002)
and led to major changes in disease control programmes by
registering on cereals, turfgrass, grapevines, potatoes, fruit,
nut, and vegetable crops. So far studies with commercially
available strobilurins formulation viz., trifloxystrobin,
kresoxim-methyl, and azoxystrobin to control sunflower
downy mildew have not been attempted.
In this study different experiments were conducted to
evaluate the efficacy of the three strobilurin fungicides on
the pathogen P. halstedii in laboratory. Further, optimiza-
tion of fungicides concentration was carried out based on
their effect on sunflower seed development. The optimized
concentrations were then evaluated both under greenhouse
and field conditions for their effect on downy mildew
disease protection. In addition, the nature and stability of
the test fungicides was also evaluated.
Materials and methods
Test fungicides
Strobilurin fungicides, viz., Trifloxystrobin; benzene acetic
acid; a-(methoxyimino)-2-[[[[1-[3-(trifluoromethyl)phenyl]
462 J Pest Sci (2010) 83:461–470
123

ethylidene] amino]oxy]- methyl]-, methylester, (E,E) (Flint
50 WG) from Bayer, Kresoxim-methyl; methyl (E)-methoxy-
imino-[-(o-tolyloxy)-o-tolyl]acetate) (Stroby 50 WG)
developed by BASF and Azoxystrobin; (E)-2-{2-[6-(2-cy-
anophenoxy) pyrimidin-4-yloxy]phenyl}-3-methoxyacry-
late) (Amistar 250 SC) a product of Syngenta, were used for
this study.
Host and pathogen
Sunflower seeds of the highly susceptible cultivar Morden
were used. The disease response of this cultivar was con-
firmed by screening experiments in the experimental plot
and also under greenhouse conditions. The pathogen
P. halstedii isolated from Morden cultivar and maintained
on the same cultivar under field conditions was used in this
study for all inoculation experiments.
Collection of sporangia and preparation of inoculum
Downy mildew-infected leaves from the Morden cultivar
were collected in the evening, the existing sporangia were
washed in running tap water, and the leaves were cut into
small pieces and placed in humidity chamber prepared by
lining the interiors of petri dishes 50 cm 9 30 cm 9 12 cm
sizes with a wet double layer of blotting paper. These
chambers were kept at 20�C and[95% RH in the dark in an
incubator for 6–7 h. After 7 h, to encourage pathogen
sporulation, the incubator was programmed to 15–18�C the
temperature at which sporangia remain intact and viable for
at least 12 h. Sporangia produced on the leaves were har-
vested into distilled water, the spore load was adjusted to
4 9 104 zoospores ml-1 using a hemocytometer, and used
as a source of inoculum in greenhouse studies (Cohen and
Sackston 1973).
In vitro studies—biological activity
Inhibition of Plasmopara halstedii sporangia
(zoosporangiogenesis)
Downy mildew-infected leaves from Morden cultivar were
collected, the existing sporangia were washed, surface
wetness was removed and 1 cm2 leaf area was smeared
with different dilutions (0.1, 0.5, 1, 2, and 5 lg ml-1) of
azoxystrobin, kresoxim-methyl, and trifloxystrobin for
30 min. Sterile distilled water treatment to infected leaves
(1 cm2) served as control. Leaves were incubated in moist
chambers for 12 h and observed for sporulation. Sporangia
from each treatment were harvested in 1 ml sterile distilled
water and spore load was assessed using a hemocytometer
with four replicates.
Inhibition of Plasmopara halstedii zoospore release and
motility
Sporangial suspension of P. halstedii prepared as per the
procedures of Cohen and Sackston (1973) and adjusted
to 1.5 9 104 sporangia ml-1 using a hemocytometer was
treated with 0.1, 0.5, 1, 2, and 5 lg ml-1 of trifloxystrobin,
kresoxim-methyl, and azoxystrobin in 1:1 ratio (v/v) and
kept for 15 min in dark conditions. Later, the treated spo-
rangial suspension was loaded to a cavity block glass slides
(76 mm 9 25 mm 9 1.25 mm) thick with ground edges
and polished round depression (15 mm diameter and 1 mm
depth) at one side. The experiment consisted of four rep-
licates each. The cavity block slides from each treatment
were observed for zoospore release by counting the empty
and intact sporangia microscopically.
The zoospore suspension of P. halstedii was adjusted
to 4 9 104 zoospores ml-1 using a hemocytometer, then
treated with the above concentrations of the test fungicides
in 1:1 ratio (v/v) and kept for 15 min in dark conditions. To
assess zoospore motility in each treatment they were har-
vested and loaded to cavity block glass slides as explained
earlier. The experiment consisted of four replicates each
(cavity block slides) observed under microscope. Relative
percentage of zoospore motility was calculated based on
numbers of motile zoospores released from the sporangia
in each fungicide treatment and was compared to the
control.
Effect of strobilurins seed treatment on sunflower seed
development
Seeds of ‘Morden’ were treated with different concentra-
tions of trifloxystrobin, kresoxim-methyl, and azoxystrobin
viz., 0.5, 2, 5, 10, and 30 lg ml-1 (400 seeds in 4 ml
solution and shaken until seeds were fully coated). Four
replicates of 100 seeds per treatment were placed equi-
distantly on three layers of moistened blotter disks in the
Perspex plates to evaluate percentage germination (Anon-
ymous 1993) and another set of seeds treated with similar
concentrations were subjected to between paper towel
method. The brown germination paper was soaked in dis-
tilled water, and fifty seeds were placed equidistant apart
on the paper. Another presoaked brown germination paper
was placed onto the first brown germination paper so that
the seeds were held in position. The paper towels were
rolled with polythene wrapping to prevent drying of the
towels, then incubated in an incubation chamber at
25 ± 2�C for 7 days. The experiment consisted of four
replications of 100 seeds (50 seeds in eight towels). Dis-
tilled water treated seeds served as control. After 7 days,
the towels were unrolled and the number of seeds germi-
nated was counted and represented as a percentage.
J Pest Sci (2010) 83:461–470 463
123

Seedling vigor was analyzed after 7 days of incubation, the
length of the root and shoot of each individual seedling was
measured. The vigor index (VI) was calculated using the
formula (Abdul Baki and Anderson 1973).
Vigor index ¼ seed germinationð%Þ � ½Mean root length
þmean shoot length�
The concentrations of the fungicides, which did not have
inhibitory effects on seed germination and vigor index,
were used for further studies.
Greenhouse studies
Effect of seed treatment and foliar spray with strobilurin
fungicides on downy mildew disease incidence upon
inoculation with Plasmopara halstedii
The fungicide treatments consisted of seed dressing with
trifloxystrobin (5 lg ml-1), kresoxim-methyl (5 lg ml-1)
and azoxystrobin (10 lg ml-1), seed dressing at the same
doses followed by foliar spray of trifloxystrobin
(10 lg ml-1), kresoxim-methyl (30 lg ml-1) and azox-
ystrobin (30 lg ml-1) and foliar spray alone. Treated seeds
were sown in earthen pots containing 2:1:1 red soil, sand
(red sandy soil) and manure. Four-day-old seedlings were
whorl inoculated with P. halstedii zoospore suspension
(4 9 104 zoospores ml-1) for three consecutive days once
every morning at around 6 AM. In the whorl inoculation
method, droplets of P. halstedii zoospores were dropped
onto the leaf whorl formed by the emerging seedlings and
allowed to flow down to the base. The challenge-inoculated
plants were maintained under greenhouse conditions
(25 ± 2�C, 95% relative humidity).
Strobilurins foliar spray was applied to inoculated plants
(before disease expression) to near run-off on the 15th day
after seedling emergence. Distilled water seed treatment or
foliar sprayed batches served as controls. Each treatment
consisted of 25 plants in four replications. These were
arranged in a randomized complete block design. Plants
showing downy mildew symptoms such as chlorosis of
leaves, growth of the pathogen and dwarfing were con-
sidered as infected. Disease incidence was recorded when
the plants were 30 days old. Percent downy mildew pro-
tection was calculated using the formula,
Fungicide nature
Curative action
Twenty-one-day old downy mildew-infected (sporulating)
sunflower plants were sprayed with trifloxystrobin (10 lg/
ml-1), kresoxim-methyl (30 lg/ml-1), and azoxystrobin
(30 lg/ml-1) till near run-off and plants were maintained
under greenhouse conditions (25 ± 2�C, 95% relative
humidity). The infected plants sprayed with distilled water
served as control. Each treatment consisted of 25 plants in
four replications. These were arranged in a randomized
complete block design. Observations were made for inhi-
bition of the pathogen in the form of recovery of plants
from disease.
Translaminar activity
The fungicides viz., trifloxystrobin (10 lg ml-1), kresox-
im-methyl (30 lg ml-1) and azoxystrobin (30 lg ml-1)
were applied to 21-day-old downy mildew-infected
(sporulating) plants on adaxial leaf surfaces by surface
smearing, in order to study the translaminar activity. In
addition fungicides were also smeared on the abaxial leaf
surface to study its direct effect on disease and on both
leaf surfaces to study the combined effect. Infected plants
smeared with distilled water served as controls. Each
treatment consisted of 25 plants in four replications and
the experiment was conducted in greenhouse conditions
(25 ± 2�C, 95% relative humidity). The plants were
arranged in a randomized complete block design. The
treated plants were examined for disease symptoms after
5 days of inoculation.
Rainfastness: residual activity
Ten-day old greenhouse plants were sprayed with suspen-
sions of trifloxystrobin (10 lg ml-1), kresoxim-methyl
(30 lg ml-1), and azoxystrobin (30 lg ml-1). Treated
plants were subjected to artificial rain in sleeves by sprin-
kling with tap water for 2 h, dried overnight, and inoculated
after 14 h with P. halstedii (4 9 104 zoospores ml-1).
Distilled water sprinkled plants served as controls. The
% protection ¼ % downy mildew in untreated plant�% downy mildew in strobilurin treated plants
% downy mildew inuntreated plants� 100
464 J Pest Sci (2010) 83:461–470
123

plants were arranged in a randomized complete block
design. The inoculated plants were kept in greenhouse
conditions (25 ± 2�C, 95% relative humidity) and exam-
ined for disease symptoms after 5 days of inoculation.
Field studies
Effect of test fungicides on sunflower downy mildew
disease
Three different field experiments viz., seed dressing, seed
dressing followed by foliar spray, and foliar spray with
the test fungicides, trifloxystrobin, kresoxim-methyl, and
azoxystrobin were conducted in the disease plot in a ran-
domized complete block design with four replicates per
plot, each replicate consisting of four rows of 25 plants.
Each rows with 5 m long and 75 cm apart (between rows)
with 25 cm plant spacing. The disease area had been
infested with oosporic inoculum for over one decade. Field
trials were conducted in the experimental station of
Department of Biotechnology, University of Mysore
(N26�180, E73�300, 817 m altitude, red loam soil) during
monsoon. The concentrations used were the same as in the
greenhouse studies. Foliar application of fungicides was
made on the 10th day after seedling emergence using hand
sprayers. Distilled water treated seed or sprayed plants
served as controls. Downy mildew disease incidence was
recorded 30th day onwards.
Statistical analysis
In vitro studies, biological activity experiments consisted
of four replicates each (hemocytometer and cavity block
slides). In the case of seed development, the experiment
consisted of four replications of 100 seeds, and values were
expressed as mean ± standard error subjected to arcsine
transformation and analysis of variance (ANOVA) using
SPSS Inc. 16.0. Significant effects of treatments were
determined by the magnitude of the F-value (P \ 0.005).
Treatment means were separated by Tukey’s HSD (Hon-
estly Significant Differences) test. Greenhouse experiments
were conducted in 4 replicates of 25 plants each (100
plants), and field experiments were conducted in 4 repli-
cates with each replicate consisting of four rows of 25
plants (400 plants) with randomized complete block design
and were analyzed separately for each experiment and
subjected to arcsine transformation and analysis of vari-
ance (ANOVA) using SPSS Inc. 16.0. Significant effects of
treatments were determined by the magnitude of the F-
value (P \ 0.005). Treatment means were separated by
Tukey’s HSD test.
Results
Biological activity
Inhibition of sporangia
A significant (P \ 0.005) dose-dependent sporulation inhi-
bition was noticed with all the three strobilurin formulations,
at 0.5, 1, 2, and 5 lg ml-1 concentrations (Table 1). Com-
plete inhibition of sporulation was obtained with 2 lg ml-1
of trifloxystrobin, while for azoxystrobin and kresoxim-
methyl it was noticed at the highest concentration
(5 lg ml-1). Further, a significant inhibition of 13000 spo-
rangia cm-2 was noticed in trifloxystrobin at 0.1 lg ml-1.
However, the production of sporangia in treatments with
kresoxim-methyl and azoxystrobin at the lowest dose
(0.1 lg ml-1) was not significantly different from that on the
control.
Inhibition of zoospore release and motility
All the tested concentrations proved to be very effective in
inhibiting zoospore release and arresting zoospore motility
(P \ 0.005) (Table 1). At the lowest concentration of
0.1 lg ml-1, percent of zoospore release of 7, 10, and 26%
and zoospore motility of 61, 52, and 45% were recorded in
treatments of trifloxystrobin, kresoxim-methyl, and azox-
ystrobin, respectively, as compared to that of the control
which recorded 85% zoospore release and 100% zoospore
motility. A dose-dependant inhibition pattern of zoospore
release and motility was observed among treatments.
Trifloxystrobin and kresoxim-methyl showed complete
inhibition of zoospore release and zoospore motility at
2 lg ml-1 concentration and were not significantly dif-
ferent, wherein azoxystrobin treatment showed complete
inhibition of zoospore release and motility at 5 lg ml-1.
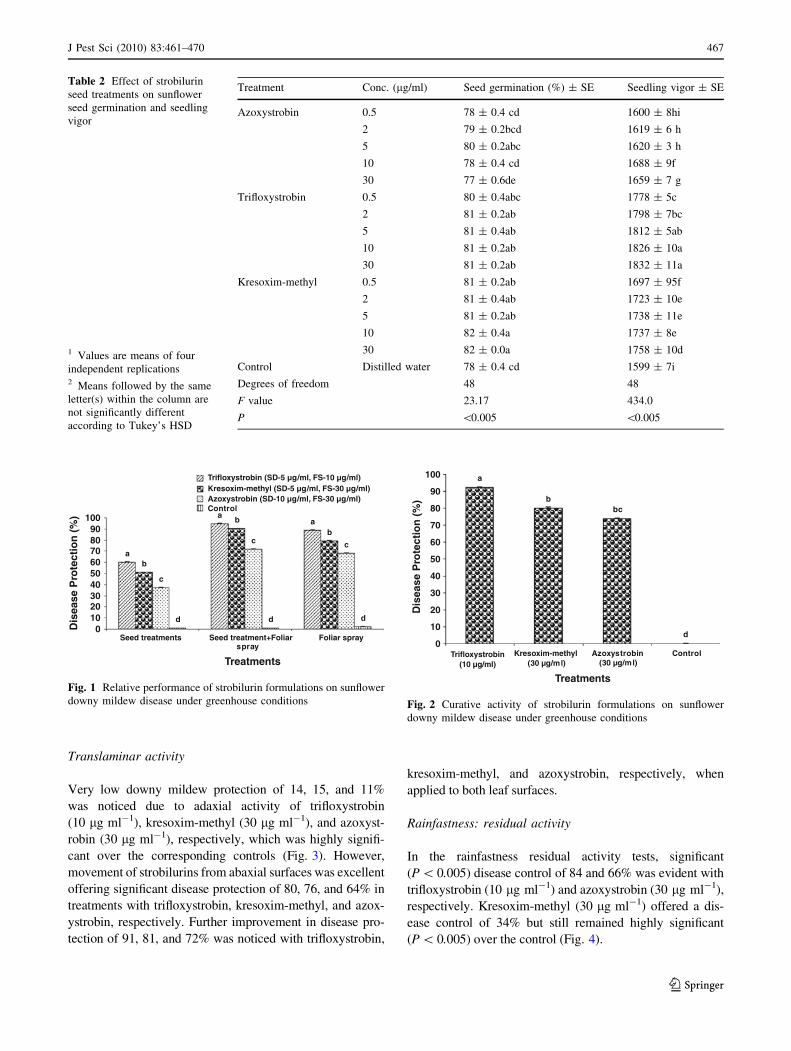
Sunflower seed development assessment
Seed treatment with different concentrations of strobilurins
revealed significant (P \ 0.005) enhanced seed germina-
tion and seedling vigor to varying degrees compared to the
control (Table 2). Maximum germination of 82% was
recorded at 10 and 30 lg ml-1 in kresoxim-methyl. In the
case of trifloxystrobin maximum germination of 81% was
noticed at concentrations 2–30 lg ml-1 and was not sig-
nificantly different with kresoxim-methyl (0.5, 2, and
5 lg ml-1) concentrations. While for azoxystrobin seed
treatment, maximum germination of 80% was noticed at
5 lg ml-1, followed by 2 lg ml-1 which recorded 79%
germination. The concentrations 0.5 and 10 lg ml-1 tested
J Pest Sci (2010) 83:461–470 465
123
in this treatment were not significantly different and were
par with the control (78% germination). Further, inhibition
of seed germination was noticed in azoxystrobin treatments
when tested at 30 lg ml-1. A similar trend was noticed for
vigor index in all the tested fungicides. For each of the
fungicide the vigor index of 1599 in control was enhanced
to maximum of 1832 at 30 lg ml-1 of trifloxystrobin, 1758
at 30 lg ml-1 of kresoxim-methyl, and 1688 at 10 lg ml-1
of azoxystrobin.
Greenhouse studies
Effect of seed treatment and foliar spray with strobilurin
fungicides on downy mildew disease incidence
None of the strobilurin treatments produced phytotoxic
symptoms of necrosis, epinasty, hyponasty, wilting, leaf-tip
injury, or vein clearing. Among seed treatments, trifloxyst-
robin was significantly (P \ 0.005) superior when compared
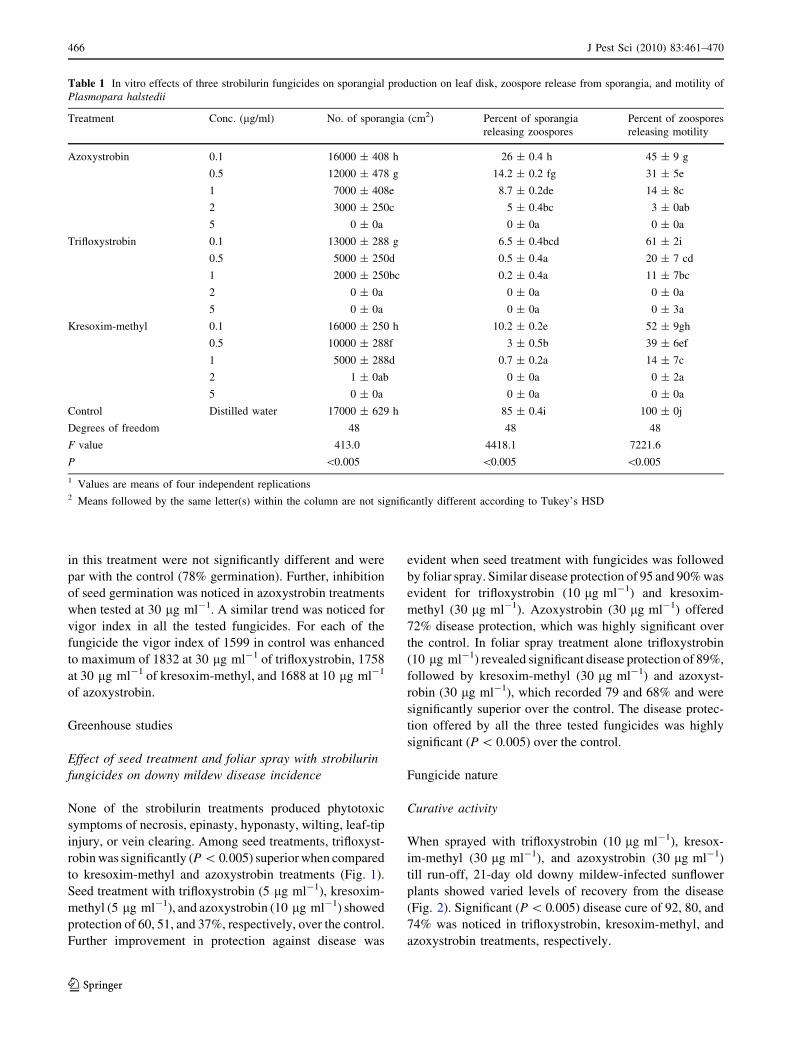
to kresoxim-methyl and azoxystrobin treatments (Fig. 1).
Seed treatment with trifloxystrobin (5 lg ml-1), kresoxim-
methyl (5 lg ml-1), and azoxystrobin (10 lg ml-1) showed
protection of 60, 51, and 37%, respectively, over the control.
Further improvement in protection against disease was
evident when seed treatment with fungicides was followed
by foliar spray. Similar disease protection of 95 and 90% was
evident for trifloxystrobin (10 lg ml-1) and kresoxim-
methyl (30 lg ml-1). Azoxystrobin (30 lg ml-1) offered
72% disease protection, which was highly significant over
the control. In foliar spray treatment alone trifloxystrobin
(10 lg ml-1) revealed significant disease protection of 89%,
followed by kresoxim-methyl (30 lg ml-1) and azoxyst-
robin (30 lg ml-1), which recorded 79 and 68% and were
significantly superior over the control. The disease protec-
tion offered by all the three tested fungicides was highly
significant (P \ 0.005) over the control.
Fungicide nature
Curative activity
When sprayed with trifloxystrobin (10 lg ml-1), kresox-
im-methyl (30 lg ml-1), and azoxystrobin (30 lg ml-1)
till run-off, 21-day old downy mildew-infected sunflower
plants showed varied levels of recovery from the disease
(Fig. 2). Significant (P \ 0.005) disease cure of 92, 80, and
74% was noticed in trifloxystrobin, kresoxim-methyl, and
azoxystrobin treatments, respectively.
Table 1 In vitro effects of three strobilurin fungicides on sporangial production on leaf disk, zoospore release from sporangia, and motility of
Plasmopara halstedii
Treatment Conc. (lg/ml) No. of sporangia (cm2) Percent of sporangia
releasing zoospores
Percent of zoospores
releasing motility
Azoxystrobin 0.1 16000 ± 408 h 26 ± 0.4 h 45 ± 9 g
0.5 12000 ± 478 g 14.2 ± 0.2 fg 31 ± 5e
1 7000 ± 408e 8.7 ± 0.2de 14 ± 8c
2 3000 ± 250c 5 ± 0.4bc 3 ± 0ab
5 0 ± 0a 0 ± 0a 0 ± 0a
Trifloxystrobin 0.1 13000 ± 288 g 6.5 ± 0.4bcd 61 ± 2i
0.5 5000 ± 250d 0.5 ± 0.4a 20 ± 7 cd
1 2000 ± 250bc 0.2 ± 0.4a 11 ± 7bc
2 0 ± 0a 0 ± 0a 0 ± 0a
5 0 ± 0a 0 ± 0a 0 ± 3a
Kresoxim-methyl 0.1 16000 ± 250 h 10.2 ± 0.2e 52 ± 9gh
0.5 10000 ± 288f 3 ± 0.5b 39 ± 6ef
1 5000 ± 288d 0.7 ± 0.2a 14 ± 7c
2 1 ± 0ab 0 ± 0a 0 ± 2a
5 0 ± 0a 0 ± 0a 0 ± 0a
Control Distilled water 17000 ± 629 h 85 ± 0.4i 100 ± 0j
Degrees of freedom 48 48 48
F value 413.0 4418.1 7221.6
P \0.005 \0.005 \0.005
1 Values are means of four independent replications2 Means followed by the same letter(s) within the column are not significantly different according to Tukey’s HSD
466 J Pest Sci (2010) 83:461–470
123
Translaminar activity
Very low downy mildew protection of 14, 15, and 11%
was noticed due to adaxial activity of trifloxystrobin
(10 lg ml-1), kresoxim-methyl (30 lg ml-1), and azoxyst-
robin (30 lg ml-1), respectively, which was highly signifi-
cant over the corresponding controls (Fig. 3). However,
movement of strobilurins from abaxial surfaces was excellent
offering significant disease protection of 80, 76, and 64% in
treatments with trifloxystrobin, kresoxim-methyl, and azox-
ystrobin, respectively. Further improvement in disease pro-
tection of 91, 81, and 72% was noticed with trifloxystrobin,
kresoxim-methyl, and azoxystrobin, respectively, when
applied to both leaf surfaces.
Rainfastness: residual activity
In the rainfastness residual activity tests, significant
(P \ 0.005) disease control of 84 and 66% was evident with
trifloxystrobin (10 lg ml-1) and azoxystrobin (30 lg ml-1),
respectively. Kresoxim-methyl (30 lg ml-1) offered a dis-
ease control of 34% but still remained highly significant
(P \ 0.005) over the control (Fig. 4).
Table 2 Effect of strobilurin
seed treatments on sunflower
seed germination and seedling
vigor
1 Values are means of four
independent replications2 Means followed by the same
letter(s) within the column are
not significantly different
according to Tukey’s HSD
Treatment Conc. (lg/ml) Seed germination (%) ± SE Seedling vigor ± SE
Azoxystrobin 0.5 78 ± 0.4 cd 1600 ± 8hi
2 79 ± 0.2bcd 1619 ± 6 h
5 80 ± 0.2abc 1620 ± 3 h
10 78 ± 0.4 cd 1688 ± 9f
30 77 ± 0.6de 1659 ± 7 g
Trifloxystrobin 0.5 80 ± 0.4abc 1778 ± 5c
2 81 ± 0.2ab 1798 ± 7bc
5 81 ± 0.4ab 1812 ± 5ab
10 81 ± 0.2ab 1826 ± 10a
30 81 ± 0.2ab 1832 ± 11a
Kresoxim-methyl 0.5 81 ± 0.2ab 1697 ± 95f
2 81 ± 0.4ab 1723 ± 10e
5 81 ± 0.2ab 1738 ± 11e
10 82 ± 0.4a 1737 ± 8e
30 82 ± 0.0a 1758 ± 10d
Control Distilled water 78 ± 0.4 cd 1599 ± 7i
Degrees of freedom 48 48
F value 23.17 434.0
P \0.005 \0.005
a
aa
b
b
b
c
c c
dd d0
102030405060708090
100
Seed treatments Seed treatment+Foliar spray
Foliar spray
Treatments
Dis
ease
Pro
tect
ion
(%
)
Trifloxystrobin (SD-5 µg/ml, FS-10 µg/ml)Kresoxim-methyl (SD-5 µg/ml, FS-30 µg/ml)Azoxystrobin (SD-10 µg/ml, FS-30 µg/ml)Control
Fig. 1 Relative performance of strobilurin formulations on sunflower
downy mildew disease under greenhouse conditions
a
bbc
d0
10
20
30
40
50
60
70
80
90
100
Kresoxim-methylTrifloxystrobin(30 µg/ml)
Azoxystrobin (30 µg/ml)
Control
Treatments
Dis
ease
Pro
tect
ion
(%
)
(10 µg/ml)
Fig. 2 Curative activity of strobilurin formulations on sunflower
downy mildew disease under greenhouse conditions
J Pest Sci (2010) 83:461–470 467
123
Field studies
Effect of test fungicides on sunflower downy mildew
disease
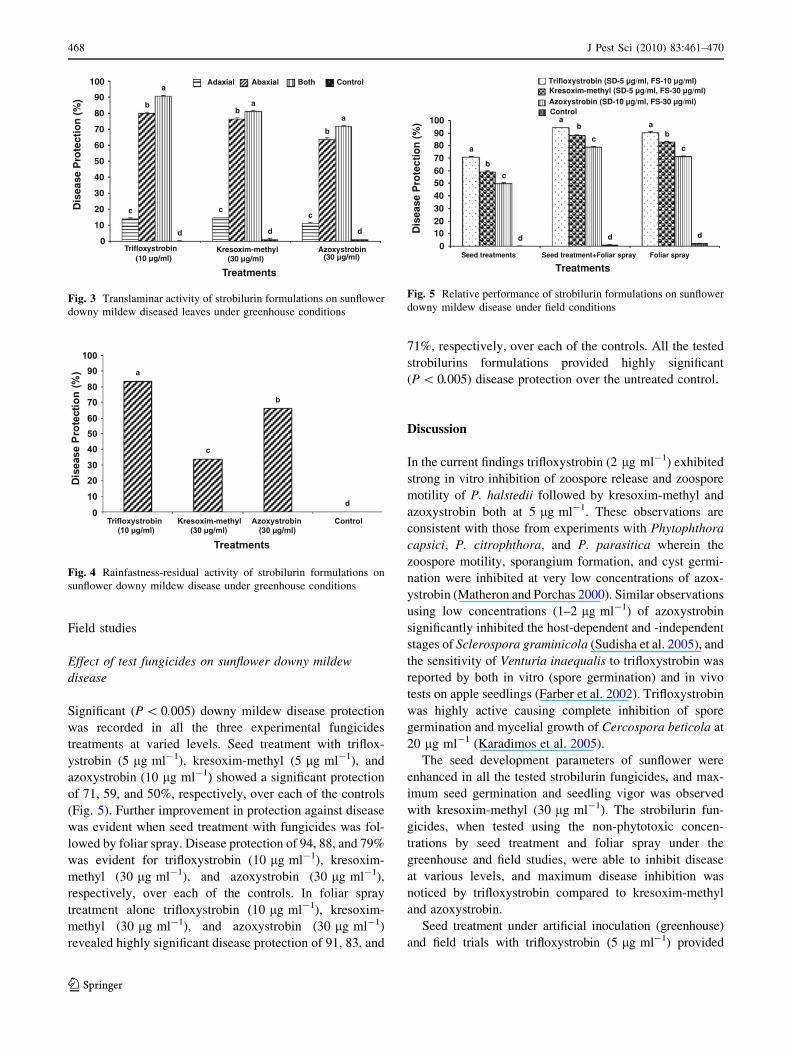
Significant (P \ 0.005) downy mildew disease protection
was recorded in all the three experimental fungicides
treatments at varied levels. Seed treatment with triflox-
ystrobin (5 lg ml-1), kresoxim-methyl (5 lg ml-1), and
azoxystrobin (10 lg ml-1) showed a significant protection
of 71, 59, and 50%, respectively, over each of the controls
(Fig. 5). Further improvement in protection against disease
was evident when seed treatment with fungicides was fol-
lowed by foliar spray. Disease protection of 94, 88, and 79%
was evident for trifloxystrobin (10 lg ml-1), kresoxim-
methyl (30 lg ml-1), and azoxystrobin (30 lg ml-1),
respectively, over each of the controls. In foliar spray
treatment alone trifloxystrobin (10 lg ml-1), kresoxim-
methyl (30 lg ml-1), and azoxystrobin (30 lg ml-1)
revealed highly significant disease protection of 91, 83, and
71%, respectively, over each of the controls. All the tested
strobilurins formulations provided highly significant
(P \ 0.005) disease protection over the untreated control.
Discussion
In the current findings trifloxystrobin (2 lg ml-1) exhibited
strong in vitro inhibition of zoospore release and zoospore
motility of P. halstedii followed by kresoxim-methyl and
azoxystrobin both at 5 lg ml-1. These observations are
consistent with those from experiments with Phytophthora
capsici, P. citrophthora, and P. parasitica wherein the
zoospore motility, sporangium formation, and cyst germi-
nation were inhibited at very low concentrations of azox-
ystrobin (Matheron and Porchas 2000). Similar observations
using low concentrations (1–2 lg ml-1) of azoxystrobin
significantly inhibited the host-dependent and -independent
stages of Sclerospora graminicola (Sudisha et al. 2005), and
the sensitivity of Venturia inaequalis to trifloxystrobin was
reported by both in vitro (spore germination) and in vivo
tests on apple seedlings (Farber et al. 2002). Trifloxystrobin
was highly active causing complete inhibition of spore
germination and mycelial growth of Cercospora beticola at
20 lg ml-1 (Karadimos et al. 2005).
The seed development parameters of sunflower were
enhanced in all the tested strobilurin fungicides, and max-
imum seed germination and seedling vigor was observed
with kresoxim-methyl (30 lg ml-1). The strobilurin fun-
gicides, when tested using the non-phytotoxic concen-
trations by seed treatment and foliar spray under the
greenhouse and field studies, were able to inhibit disease
at various levels, and maximum disease inhibition was
noticed by trifloxystrobin compared to kresoxim-methyl
and azoxystrobin.
Seed treatment under artificial inoculation (greenhouse)
and field trials with trifloxystrobin (5 lg ml-1) provided
ccc
b
bb
a
a
a
ddd0
10
20
30
40
50
60
70
80
90
100
Trifloxystrobin Kresoxim-methyl Azoxystrobin
Treatments
Dis
ease
Pro
tect
ion
(%
)Adaxial Abaxial Both Control
(10 µg/ml) (30 µg/ml) (30 µg/ml)
Fig. 3 Translaminar activity of strobilurin formulations on sunflower
downy mildew diseased leaves under greenhouse conditions
a
c
b
d0
10
2030
405060
7080
90100
Trifloxystrobin (10 µg/ml)
Kresoxim-methyl(30 µg/ml)
Azoxystrobin (30 µg/ml)
Control
Treatments
Dis
ease
Pro
tect
ion
(%)
Fig. 4 Rainfastness-residual activity of strobilurin formulations on
sunflower downy mildew disease under greenhouse conditions
a
aa
b
bb
c
cc
d d d0
102030405060708090
100
Seed treatments Seed treatment+Foliar spray Foliar spray
Treatments
Dis
ease
Pro
tect
ion
(%
)
Trifloxystrobin (SD-5 µg/ml, FS-10 µg/ml)Kresoxim-methyl (SD-5 µg/ml, FS-30 µg/ml)Azoxystrobin (SD-10 µg/ml, FS-30 µg/ml)Control
Fig. 5 Relative performance of strobilurin formulations on sunflower
downy mildew disease under field conditions
468 J Pest Sci (2010) 83:461–470
123

superior average disease protection of 65%, followed by
kresoxim-methyl (5 lg ml-1) and azoxystrobin
(10 lg ml-1) which offered 55 and 43% disease protection,
respectively. Trifloxystrobin (10 lg ml-1) as foliar spray
alone recorded higher disease protection (average 90%) than
that of seed treatment, further the combination of both seed
treatment and foliar spray of trifloxystrobin provided highly
significant average disease protection of 94% compared to
individual treatments alone (65% and 90% for seed treatment
and foliar spray, respectively). A similar trend in disease
protection was also observed in kresoxim-methyl and azox-
ystrobin with foliar spray alone and combination treatments.
Trifloxystrobin had stronger curative action (92%) than
kresoxim-methyl (80%) and azoxystrobin (74%). Transla-
minar activity of the tested fungicides was superior mainly
by abaxial modes compared to adaxial means, and this
property of strobilurins has been reported previously
(Hermann et al. 1998; Reuveni 2001). Movement of stro-
bilurins from abaxial surfaces was excellent and offered
significant disease protection in treatments with trifloxyst-
robin, kresoxim-methyl, and azoxystrobin, respectively,
over the control. The foliar uptake of strobilurin fungicides
from both abaxial and adaxial surfaces is rapid due to their
distribution around the application site either by local or
systemic action (Margot et al. 1998). The variation in the
activity of strobilurin fungicides in the above experiments
was due to different biological properties that consequently
confer a wide range of biokinetic behavior both inside the
plants and around its external surfaces (Bartlett et al. 2002).
Strong translaminar activity was recorded by trifloxyst-
robin with high disease protection when applied on abax-
ial-infected leaf surfaces. Similar strong translaminar
activity of trifloxystrobin was observed in sugar beet
infected with Cercospora beticola (Karadimos et al. 2005).
Further, rainfastness residual activity by trifloxystrobin
and azoxystrobin was significant in disease control which
was evident by relatively small loss of both formulations.
Though kresoxim-methyl had good curative activity, it
recorded higher loss of formulation on leaf surfaces resulting
in lesser disease protection. This result clearly indicates the
loss of kresoxim-methyl formulation which was relatively
high compared to trifloxystrobin and azoxystrobin.
In this study all three strobilurin formulations tested
exhibit strong in vitro inhibition of the pathogen, an
increase of sunflower seeds germination and vigor, curative
effect, rainfastness residual activity (except for kresoxim-
methyl), and moderate translaminar activity.
This is the first report of strobilurin fungicides viz.,
trifloxystrobin, kresoxim-methyl, and azoxystrobin exhib-
iting high activity against P. halstedii under laboratory,
greenhouse, and field conditions. These fungicides are very
promising for sunflower downy mildew disease manage-
ment by seed treatment which is feasible and economical,
and they also can be used in foliar treatments. Further
investigations are needed to establish the most efficient
sequential treatments to include strobilurins for sunflower
downy mildew disease management. In conclusion, the
application of strobilurin fungicides as seed treatment and
also by foliar treatments could be a beneficial component
of integrated disease management of sunflower. These
strobilurin fungicides, apart from their action against sun-
flower downy mildew pathogen, are good plant growth
promoters, which is an added advantage for any practical
agricultural system.
Acknowledgments The authors thank Dr. T. S. Thind, Professor,
Department of Plant Pathology, Punjab Agricultural University,
Ludhiana, Punjab, India for supplying the experimental fungicides
and Indian Council of Agricultural Research (ICAR) through All
India Co-ordinated Pearl Millet Improvement Project (AICPMIP),
Government of India for laboratory and field facilities provided at
University of Mysore.
References
Abdul Baki AA, Anderson JD (1973) Vigor determination in soybean
seed by multiple criteria. Crop Sci 13:630–633
Agrawal SC, Gupta RK, Prasad KVV (1991) A case of downy mildew
of sunflower in Madhya Pradesh. J Oilseeds Res 8:126
Albourie JM, Tourvieille J, de Tourvieille Labrouhe D (1998)
Resistance to metalaxyl in isolates of the sunflower pathogen
Plasmopara halstedii. Eur J Plant Pathol 104:235–242
Anke T, Oberwinkler F, Steglich W, Schramm G (1977) The
strobilurins new antifungal antibiotics from the basidiomycete
Strobiluris tenacellus. J Antibiotics 30:806–810
Anonymous (1993) International rules for seed testing. Seed Sci
Technol 13:309–333
Attard A, Gourgues M, Galiana E, Panabieres F, Ponchet M, Keller H
(2007) Strategies of attack and defense in plant oomycete
interactions, accentuated for Phytophthora parasitica dastur
(syn. P. nicotianae Breda de Haan). J Plant Physiol. doi:
10.1016/j.jplph.2007.06.011
Bartlett DW, Clough JM, Godwin JR, Hall AA, Hamer M, Parr-
Dobrzanski B (2002) The strobilurin fungicides. Pest Manag Sci
58:649–662
Beck C, Oerke EC, Dehne HW (2002) Impact of strobilurins on
physiology and yield formation of wheat. Meded Rijksuniv Gent
Fak Landbouwkd Toegep Biol Wet 67(2):181–187
Cohen Y, Sackston WE (1973) Factors affecting infection of
sunflowers by Plasmopara halstedii. Can J Bot 51:15–22
Davidse LC, van den Berg-Velthuis GCM, Mantel BC, Jespers ABK
(1991) Phenylamides and Phytophthora. In: Lucas JA, Shattock
RC, Shaw DS, Cooke LR (eds) Phytophthora. British Mycolog-
ical Society, Cambridge, pp 349–360
Farber RB, Chin KM, Leadbitter N (2002) Sensitivity of Venturia
inaequalis to trifloxystrobin. Pest Manag Sci 58(3):261–267
Gisi U, Chin KM, Knapova G, Kung Farber R, Mohr U, Parisi S,
Sierotzki H, Steinfeld U (2000) Recent developments in
elucidating modes of resistance to phenylamide, DMI and
strobirulin fungicides. Crop Protection 19:863–872
Gulya TJ, Draper M, Harbour J, Holen C, Knodel J, Lamey A, Mason
P (1999) Metalaxyl resistance in sunflower downy mildew in
North America. Proc. 21st Sunf. Res. Workshop, Jan 14 & 15,
1999, Fargo ND, USA, pp 2–7
J Pest Sci (2010) 83:461–470 469
123

Hermann D, Fischer W, Knauf-Baider G, Steinemann A, Margot P,
Gisi U, Laird D (1998) Behavior of the new strobilurin fungicide
trifloxystrobin on and in plants. Phytopathology 88:S37
Herms S, Seehaus K, Koehle H, Conrath U (2003) A strobilurin
fungicide enhances the resistance of tobacco against tobacco
mosaic virus and Pseudomonas syringae pv tabaci. Plant Physiol
130:120–127
Karadimos DA, Karaoglanidis GS, Klonari TK (2005) Biological
activity and physical modes of action of the Qo inhibitor
fungicides trifloxystrobin and pyraclostrobin against Cercosporabeticola. Crop Protection 24:23–29
Lamour KH, Hausbeck MK (2000) Mefenoxam insensitivity and the
sexual stage of Phytophthora capsici in Michigan cucurbit fields.
Phytopathology 90:396–400
Margot P, Huggenberger F, Amrein J, Weiss B (1998) A new broad-
spectrum strobilurin fungicide. Brighton Crop Prot Conf Pest
Disease 2:375–382
Matheron ME, Porchas M (2000) Impact of azoxystrobin, dim-
ethomorph, fluazinam and fosetyl-Al and metalaxyl on growth,
sporulation and zoospore cyst germination of three Phytophthorasp. Plant Dis 84:454–458
Mayee CD, Patil MA (1986) Downy mildew of sunflower. Indian
Pytopathol 39:314
Melero-vara JM, Garcia-Baudin C, Lopez-Herrera CJ, Jimenez-Diaz
RM (1982) Control of sunflower downy mildew with metalaxyl.
Plant Dis 66:132–135
Mitani S, Araki S, Takii Y, Ohisma T, Matsuo N, Miyoshi H (2001)
The biochemical mode of action of the novel selective fungicide
cyazofamid: specific inhibition of mitochondrial complex III in
Pythium spinosum. Pestic Biochem Physiol 71:107–115
Molinero-Ruiz ML, Domı́nguez J, Melero-Vara JM (2002) Races of
isolates of Plasmopara halstedii from Spain and studies on their
virulence. Plant Dis 86:736–740
Molinero-Ruiz ML, Melero-Vara JM, Gulya TJ, Dominguez J (2003)
First report of resistance to metalaxyl in downy mildew of
sunflower caused by Plasmopara halstedii in Spain. Plant Dis
87:749
Molinero-Ruiz ML, Dominguez J, Gulya TJ, Melero-Vara JM (2005)
Reaction of field populations of sunflower downy mildew
(Plasmopara halstedii) to metalaxyl and mefenoxam. Helia
28:65–74
Molinero-Ruiz ML, Cordon-Torres MM, Melero-Vara JM, Domin-
guez J (2008) Resistance to metalaxyl and to metalaxyl-M in
populations of Plasmopara halstedii causing sunflower downy
mildew. Can J Plant Pathol 30:97–105
Oros G, Viranyi F (1984) Resistance of Plasmopara halstedii to
metalaxyl in the greenhouse. Temperate Downy Mildew News-
letter 3:22–23
Parra G, Ristaino JB (1998) Insensitivity to Ridomil Gold (mefen-
oxam) found among field isolates of Phytophthora capsicicausing Phytophthora blight on bell pepper in North Carolina
and New Jersey. Plant Dis 82:711
Parra G, Ristaino JB (2001) Resistance to mefenoxam and metalaxyl
among field isolates of Phytophthora capsici causing Phytoph-thora blight of bell pepper. Plant Dis 85:1069–1075
Patil MA, Phad H, Ramtirthkar MS, Phad HB (1991) Chemical
control of sunflower downy mildew caused by Plasmoparahalstedii. Indian Phytopathol 44:325–327
Reuveni M (2001) Activity of trifloxystrobin against powdery and
downy mildew diseases of grapevines. Can J Plant Pathol 23:52–
59
Sauter H, Steglich W, Anke T (1999) Strobilurins: evolution of new
class of active substances. Angew Chem Int Ed 38:1328–1349
Shetty K (1998) Managing metalaxyl-resistance in Plasmoparahalstedii from a chemical company perspective. Symposium
III, Sunflower Downy Mildew, Jan 13 & 14, 1998, Fargo ND,
USA, pp 74–78
Stenzel K, Pontzen R, Seitz T, Tiemann R, Witzenberger A (1998)
SZX 722: a novel systemic oomycete fungicide. Proc Brighton
Crop Prot Conf—Pests and Diseases, BCPC, Farnham, Surrey,
UK, pp 361–374
Sudisha J, Amruthesh KN, Deepak SA, Shetty NP, Sarosh BR, Shetty
HS (2005) Comparative efficacy of strobilurin fungicides against
downy mildew disease of pearl millet. Pesticide Biochem
Physiol 81:188–197
Sukul P, Spiteller M (2000) Metalaxyl: persistence, degradation,
metabolism and analytical methods. Rev envir Contam toxicol
164:1–26
Viranyi F, Gulya TJ, Masirevic S (1992) Races of Plasmoparahalstedii in Central Europe and their metalaxyl sensitivity. Proc
13th Int Sunf Conf, Vol I, Sept 7–11, 1992, Pisa, Italy, pp 865–
868
Wiggins TE, Jager BJ (1993) Mode of action of the new methoxy-
acrylate antifungal agent ICIA5504. Biochem Soc Trans 22:68S
470 J Pest Sci (2010) 83:461–470
123




















