
IDENTIFICATION OF TWO NOVEL CVC DOMAIN - TSpace
87
IDENTIFICATION OF TWO NOVEL CVC DOMAIN- CONTAINING HOMEOBOX GENES Jens Looser A thesis submitted in conformity with the requirements for the degree of Master of Science Graduate Department of Molecular and Medical Genetics University of Toronto O Copyright by Jens Looser 1995
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of IDENTIFICATION OF TWO NOVEL CVC DOMAIN - TSpace

IDENTIFICATION OF TWO NOVEL CVC DOMAIN-
CONTAINING HOMEOBOX GENES
Jens Looser
A thesis submitted in conformity with the requirements
for the degree of Master of Science
Graduate Department of Molecular and Medical Genetics
University of Toronto
O Copyright by Jens Looser 1995

National Library 1*1 of Canada Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques 395 Wellington Street 395, rue Weilington ûîtawa ON K1A ON4 Onawa ON K1A ONO Canada canada
The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distribute or seU reproduire, prêter, distribuer ou copies of this thesis in microform, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/film, de
reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts fiom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

IDENTIFICATION OF TWO NOVEL CVC DOMAIN-
CONTAINING HOMEOBOX GENES.
Master of Science, 1995,
Jens Looser,
Graduate Department of Molecular and Medical Genetics,
University of Toronto.
ABSTRACT
The retinal C M 0 gene encodes a protein with both a homeodomain of the paired-
like type and a novel evolutionarily conserved protein domain of unknown function, the
CVC domain. In this thesis, 1 report the isolation of CHXIO- 2, a human gene related to
CHXIO. The CHXIO-1 cDNA was isolated from a screen for CHXIO homologues
expressed in adult human retina. CHXIO-I is expressed at low levels in the inner nuclear
and ganglion ce11 layers of the adult retina Chxl71, a murine CHXIO-1 homologue, differs
from CHXIO-1 in sequence and abundance. Genomic Southem analysis showed that
although Chxl7i is the closest mouse homologue of CHXIO-1, a more closely related
human gene may exist. This work demonstrates that multiple CVC domain-containing
homeobox genes are expressed in the mammalian retina. Like C M O , they are likely to
contribute to retinal development and maintenance.

ACKNO WLEDGMENTS
Sitting at my cornputer (a Mac, not some lousy IBM!) now after everything is over 1
try to remember everyone that helped me in completing this monumental piece. Suddenly
my brain feels as einpty as it has been for the last three months while 1 worked on the
following pages. Therefore, in case 1 forget your name in the following lines please forgive
me.
1 guess 1 wiii take the obvious ones fïrst. Thanks to d l past and present inernbers of
the McInnes Lab (in dphabetical order: Anna Soltyk, Bryan Snow, Carol Freund, Dingfang
Bu, Elizabeth Garami, Geoff Clarke, Ivy Liu, Jakub Noviik, Joe Zhou, Lin Liu, Lynda
Ploder, Midhat Osman, Roger Bascom) for their various contributions to iny work, the
cnimby remainders of my social life or rny booze-independent headaches. Special thanks to
Geoff Clarke for Our crash-and-bum science talks and the fact that he actually succeeded (at
times) in making me think.
Thanks to the University of Toronto, the Research lnstitute at the Hospital for Sick
Children and the Medical Research Council of Canada for the scholarships 1 received.
Thanks to everyone in the Hospital and the Department, especially Janet Rossant,
Johanna Rommens, Lee Montgomery, Lynn Mar and Vivien Measday, for uncountable
favors and advice on various aspects of life.
A particular thanks to Rod Mclnnes (old buddy, old pal) for taking the
(retrospectively seen not so big, eh!) risk of accepting some foreigner who burst into his
off~ce two summers ago. Thanks for teaching me aspects of d t i n g and speakinç that 1 did
not cal1 my own before being chained to one of your benches. 1 also appreciated your
support of my future plans even though they did not quite match yours.
Thanks to everyone who had to deal with my whining over the last two years,
especially during the last crack down on the thesis. Special thanks to Viçky Aneliunas (for
teaçhing me proper table manners), Pat Derkis (for taking my mind of work), Lian de
Lotbinière (for actuaily admiring what 1 am doing) and Christina Zeidler (for k ing herself).
Last and most importantly 1 would like to thank my family and fnends over in good
old Germûny. Your love, understanding and support enabled me to come and stay in
Canada. As a token of my love 1 want to dedicate this thesis to my parents for everything
they have ever done for me.

TABLE OF CONTENTS
Abstract
Acknowledgmen ts
Table of Contents
List of Figures
CHAPTER 1: INTRODUCTION
1. RETINA
1 . 1 Physiology of the adult mamrnalian retina
1.2 Development of the marnmalian retina
2. HOMEOBOX GENES
2.1 Discovery and Charactenzation of the homeobox
2.2 Phylogeny of the homeobox
2.3 The Paxfprd class of homeobox genes
2.4 Pax6/eyeiess; Master contd genes of eye development
3. ChxlO
3.1 Ch10 biology in the mammalian retina
3.2 Ch10 mutations and ocular retardution
3.3 Ch10 homologues in rnice and goldfish
4. RATIONALE FOR THIS PROJECT

CHAPTER 2: ORIGINAL RESEARCH
INTRODUCTION
EXPERIMENTAL PROCEDURES
RESULTS
1. Attribution of data
2. Human retinal screen
3. OTX3, a novel orthdenticle homologue
4. Human LIM homeobox gene LH2
5 CHXIO-1, a novel CHXlO homologue
5.1 Cloning
5.2 Putative protein domains
5.3 Mapping to the chromosomes
5.4 Northem analysis on total RNA
5.5 Northern analysis on poly-(A)+ RNA
5.6 In situ hybridizations
6. Isolation of the murine CHXIO-1 orthologue. C h 1 71
6.1 Cloning
6.2 Putative protein domains
6.3 Northem analysis
6.4 Phylogenetic relationship to CHXIO-1
DISCUSSION
CHAPTER 3: FUTURE DIRECTIONS
LIST OF CITATIONS

LIST OF FIGURES
Figure 1 - 1
Figure 1-2
Figure 1-3
Figure 1-4
Figure 1-5
Figure 1-6
Figure 2- 1
Figure 2-2
Figure 2-3
Figure 2-4
Figure 2-5
Figure 2-6
Figure 2-7
Figure 2-8
Figure 2-9
Figure 2- 10
Figure 2- 1 1
Figure 2- 12
Figure 2- 13
Figure 2-14
Figure 2- 15
Retinal layer structure
Murine eye development
ChxlO compared to Px/prd homeobox genes
C M 0 in situ hybridizations
ocular retardution phenotypes
previously known Chri0 homologues
Flow chart of the human retinal screen
Class L,2 and 3 plaque pure phages
OTX3 alignments
CHXlO-1 clone rnap and cDNA
chx 10- l/chx 10: aiignment and peptide structures
Mapping of CHXIO-1 io the chromosomes
CHXlO-1 Northern analysis
CHXIO-1 in situ hybridizations
Ch171 clone map and cDNA
c h 17 llchx 10- 1 : dignment and peptide structures
Chxl71 Nort hem analy sis
Evolutionary conservation of chx 10
Aiignment of al1 CVC homeobox genes
Mouse genomic Southem analysis
Human genomic Southem analysis

CHAPTER 1: INTRODUCTION
In this thesis, 1 present the results of a screen for homologues of the 'retinal'
homeobox gene ChxlO. In this chapter, 1 present a brief overview of: 1. selected aspects
of neuroretinal biology, 2. homeobox-containing genes in general and, in more detail, the
Pax/prd class, and 3. the biology of Ch10 and its importance for the retina.
1. RETINA
1.1 Physiology of the adult mammalian retina
Light absorption in vertebrates occurs in a thin sheet of neural tissue called the
neuroretina, which lines the back of the eye cup. Within this thin sheet of neural tissue,
visual image information is captured, transduced into a neuronal signal and sent to the
visual centers of the brain. Six major classes of neuronal cells (photoreceptors, horizontal,
amacrine, interplexiform, bipolar and ganglion) and two classes of glial cells (Müller and
astrocyte) are found in the retina (Figure 1-1 .). The neuronal ce11 nuclei are organized into
three well defined parallel nuclear layers, which are separated by two plexiform layers that
host the synaptic processes (Adler and Farber, 1986; Rowe, 199 1).
Retinal organization is the inverse of what one might intuitively suspect, in that the
photoreceptors are facing away from the light source and are covered by multiple retinal ce11
layers (Figure 1- 1). Light that enters the eye therefore has to travel through al1 three
nuclear layers before being captured by the photoreceptors. Distal to the photoreceptors
lies the retinal pigment epithelium (RPE), a neuroepithelial layer of single ce11 thickness.
The pigment granules in this layer help absorb scattered light and thereby enhance the
image quality. Additionally, the RPE serves to transport metabolites and nutrients to the
photoreceptors as well as to promote phagocytosis of shed discs from the outer segments
of the photoreceptor cells. The inner segments of the photoreceptor cells, which contain
the major metabolic machinery of the ce11 and the nucleus, make up the outer nuclear layer.

Vitreal Border
Figure 1-1
Schematic representation of the mammaiian retina with aii its ce11 types. Abbreviation of
ceU narnes: astrocyte (A), bipolar (B), cone (C), horizontal (H), interplexiform a), Müller
(M), rod (R). Taken from Adler and Farber (1986).

Photoreceptors extend their synaptic termini into the outer plexiform layer (OPL), which
separates the outer nuclear layer (ONL) from the inner nuclear layer (NL). Here, synapses
between photoreceptor cells and bipolar cells form via the horizontal cells. The bipolar
cells lie in the inner nuclear layer together with four other major ce11 classes (horizontal,
interplexiform, arnacrine and displaced ganglion). Distal to the inner nuclear layer is the
inner plexiform layer (IPL) which contains the direct or indirect synapses between the
interneurons and the ganglion cells in the most proximal layer, the ganglion ce11 layer. On
the level of both the inner and outer plexifon layer visual information is being integrated
and modulated by horizontal and amacrine cells. respectively. Al1 ganglion cells project
their axons towards the optic disk where they converge to form the optic nerve, which
serves as the sole pathway for transmission of visual information to the brain (Adler and
Farber, 1986; Robinson, 199 1 ; Rowe, 199 1 )
1.2 Development of the mammalian retina
RetinaI structures arise from the germinal neuroepithelium, and their neuronal
development follows the same principles that have been defined for the rest of the CNS.
The precise time at which the eye primordia become induced is still in question, but studies
in amphibians indicate that this event may occur as early as gastrulation (Spemann, 1938).
Additionally Spemann's work shows that a well defined area of the anterior neural plate
seems to be largely committed to generating the eye at the neural plate stage. This
cornmitment, however, is not complete until the optic vesicle stage, since transplantation of
this neural plate region with more posterior mesoderm leads to the formation of more
posterior structures (Aldennann, 1935).
The first visible signs of eye development are observed at the neural plate stage,
which corresponds to embryonic day eight (E8) in rnice. Two small indentations, called
the optic pits, or sulci, form bilaterally on the dorsal surface of the neural plate (Figure 1-
2A). As the neural plate thickens and curls up on itself to fuse into the neural tube, the
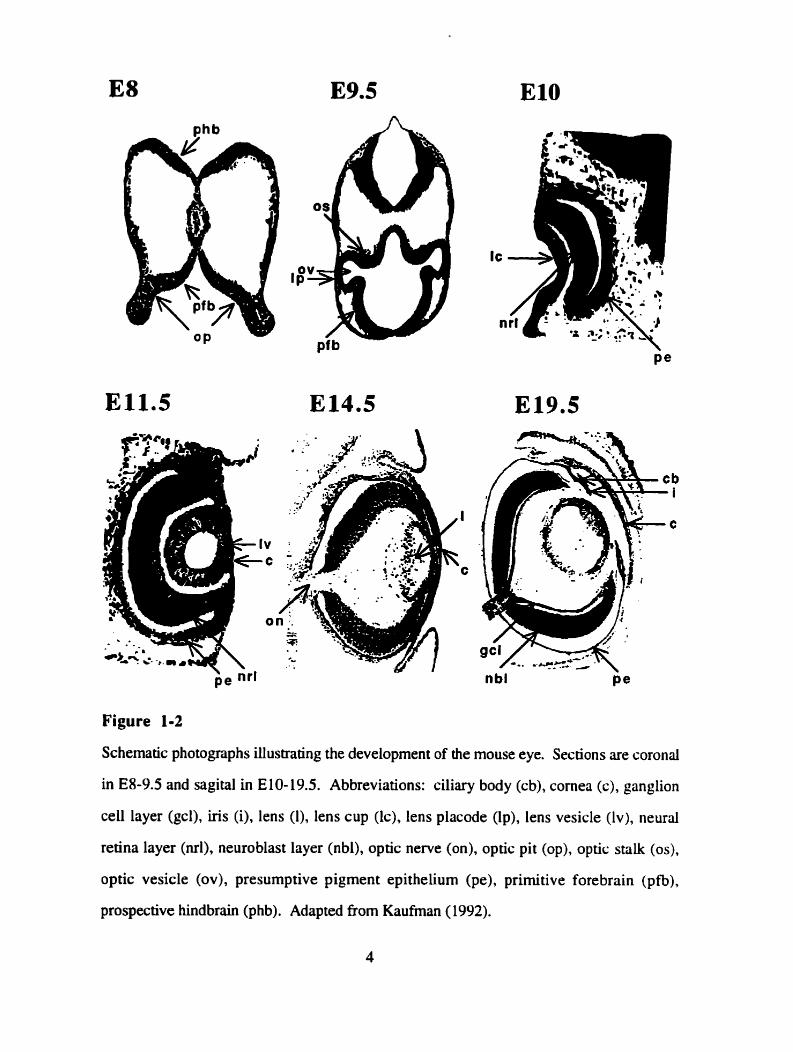
Figure 1-2
Schematic photographs illustrating the development of the mouse eye. Sections are coronal
in E8-9.5 and sagitd in E10-19.5. Abbreviations: ciliary body (cb), comea (c), ganglion
ce11 layer (gcl), iris (i), lens (l), lens cup (lc), lens placode Op), lens vesiçle (IV), neural
retina layer (ml), neuroblast layer (nbl), optic nerve (on), optic pit (op), optic stalk (os),
optic vesicle (ov), presumptive pigment epithelium (pe), primitive forebrain (pfb),
prospective hindbrain (phb). Adapted fiom Kaufman ( 19%).

optic vesicles form as lateral outgrowths of the indentations of the diencephalon, the optic
pits. At day E9 (in mice) the optic vesicle will have extended enough to establish physical
contact with the overlying ectoderm (Figure 1-2B). Mesenchymai cells, which initially fil1
the space between the optic vesicle and the overlying surface ectoderm, partially degenerate
as the two ce11 layers establish physicai contact. Those mesenchymal ceus that remain will
give rise to the muscles of the pupil, the pigment cells of the iris, the ciliary body and the
choroid (Robinson, 1991). The development of the eye placode and the lens disk, as local
thickenings of the optic vesicle and the surface ectoderm, respectively, depends on the
establishment of physical contact between both tissues. The optic vesicle will subsequentiy
invaginate to form the the optic cup (Figure 1-2C). This invagination occurs slightly off-
center, resulting in the formation of a groove dong the ventral wall of the optic stalk. This
groove, the optic fissure, deepens as the ventral optic cup thickens to complete the cup
structure and, in the aduli eye, will serve as the exit point of ganglion ce11 axons and retinal
blood vessels (Robinson, 199 1).
The inner and the outer cup layer assume different fates &er the invagination of the
optic vesicle. Proliferation suddenly decreases in the outer cup layer leading to the
formation of a single-ce11 layer, the future retinal pigment epithelium (Figure 1-2D).
Continuing proliferation in the inner layer of the optic cup, however, leads to the formation
of an apparently homogeneous mass of cells called the neuroblast layer (Figure 1-2E). All
neuronal cells found in the adult retina, as well as the Müller glial cells, onginate from the
neuroblast layer. Only the second type of glial cells, the astrocytes, migrate into the eye,
via the optic nerve (Reh, 1992).
Mitotically active undifferentiated cells of the developing retina go through M-phase
at the outer limiting membrane (OLM), and through S-phase in the inner part of the
neuroblast layer. From there, they return to the OLM for their next division. After their
final mitosis, neurons migrate inwards to assume their proper position within the retina.
Ce11 differentiation is thought to occur after the last rnitosis but prior to migration (Hinds

and Hinds, 1978; Tumer and Cepko, 1988). The birth of neuronal cells has been found to
occur in two waves in al1 mammalian species. With rninor variations between species,
ganglion cells, horizontal cells, cone cells and amacrine cells constitute the first wave,
while bipolar cells, rod photoreceptors and Müller glial cells make up the second (Sidman,
1961; Young, 1985). In dl species studied to date lineage analysis suggests that these cells
arise from multipotent precursor cells (Cepko, 1992; Wetts and Fraser, 1988).
Coincident with strong proliferation is neuronal ce11 death as an integral part of
normal retinal development (Silver and Hughes, 1973). More than half of al1 generated
neurons have died by the time adulthood is reached, most likely because they failed to
establish synapses (Robinson, 199 1).
The fnst morphologicai signs of ce11 differentiation in the neuroblast layer are found
at E l 1 in mice. This early differentiation occurs in ganglion ce11 precunors, and thus the
ganglion ce11 layer is the first layer to separate from the neuroblast layer. The remaining
layers form simultaneously and lead to the formation of the multilayered adult retina
(Figure 1-2F). Retinal differentiation in the mouse is not complete until the second
postnatal week, concurrent with the opening of the eyes (Sidman, 1961; Hinds and Hinds,
1974, 1978, 1979; Young, 1985).
Complex developmental processes, such as the morphological changes in the
developing retina outlined above, require complex genetic control. Work of the last ten
years has led to the identification of multiple classes of developmental regulators. One such
class consists of transcriptional regulators that, based on functional and structural
characteris tics, have been named homeobox genes.

2. HOMEOBOX GENES
2.1 Discover y and Characterization of the homeobox
The discovery and study of the homeotic genes has allowed more molecular insight
into developmental processes than might have been guessed 15 years ago. Most of the
pioneering work was done in Drosophila rnelanogaster due to the relative ease of genetic
studies. The narne Homeosis was originally coined by Bateson (1894) for ~ansforrnations
of body segments 'into the likeness of something else'. In 1923 Bridges identified the first
homeotic mutation in flies. Due to the transformation of the rudimentary pair of wings, the
halteres, into an actual second pair of wings, he named the mutation bifhorux (Bridges and
Morgan, 1923). The second homeotic mutant, Antennapedia, was identified by Gehring
in 1967, as a spontaneous mutation that converted the antenna into an additional pair of legs
(Gehring, 1967). The fact that mutations in single genes led to transformation of body
segments into the likeness of others led Lewis to postulate that these homeotic genes might
represent a tandem array of conserved genes (Lewis, 1978).
The ability to clone the genes responsible for these mutations came with the advent
of chromosome walking, and was used to clone both the bithorax (Bender et al., 1983),
and the Antennapedia locus (Garber et al., 1983). Complementary DNA's that were
located to the genomic region surrounding the Antennapedia locus exhibited cross-
hybndization when used on genomic blots, the first evidence suggesting the existence of
conserved sequences predicted by Lewis (1978). Sequence analysis of the isolated cDNAs
revealed that al1 of them share a common motif of 180 nucleotides, which was termed the
homeobox due to its occurrence in homeotic genes. By analogy, the encoded protein
domain was named the homeodomain. The homeotic genes in Drosophila were found to be
organized into two complexes, the Antennapedia and bithora complexes (Gehring, 1994).
The fîst clues as to the function of the homeodomain came from the identification
of homology to DNA binding motifs of the al and a 2 yeast mating type (MAT) proteins
(Shepherd et al., 1984) and the helix-turn-helix motifs (Scott et al., 1989) of prokaryotic

transcriptional regulators (Pabo and Sauer, 1984). The presence of these putative DNA
binding domains indicated a possible involvement in transcriptional regulation. in recent
years homeodomain proteins have been proven to be transcriptional regulators with
sequence specific DNA binding properties. Homeodomains bind DNA as monomers with
high affinity (Desplan et al. 1985). The ability of the homeodomain to regulate
transcription has been demonstrated in ce11 culture for frz, Ubx and Abd-B (Jaynes and
O'Farrell, 1988; Thali et al., 1988) and in vivo for the regulation of hunchback by bicoid
(Driever and Nuesslein-Vollhard. 1989). In vitro target sites were found to have a
conserved 5'-TAAT-3' core sequence, even for relatively divergent homeodomains. Initial
homeodomain swap experiments showed that the developmental specificity of
homeodomain protein function resides in the homeodomain (Kuziora and McGinnis,
1989). In some cases, however, the homeodomain requires additional regions of the
protein to achieve its developmental specificity (Mann and Hogness, 1990). Mutagenesis
studies demonstrated that much of the DNA binding specificity resides in amino acid 50 of
the homeodomain (Treisman, 1989).
Structural analysis of homeodornains has shown that the homology with regulatory
motifs is not restricted to the sequence level. Nuclear magnetic resonance (NMR)
spectroscopy for Antennapedia (Antp) has shown that the homeodomain forms three a-
helices, with helices two and three exhibiting a helix-tum-helix motif (Qian et al., 1989).
NMR studies of Antp bound to its target sequence showed that helix three, the so-called
recognition helix, contacts DNA in the major groove. The N-terminal arm of the
homeodomain contacts bases in the minor groove and the loop between helix 1 and 2
establishes contact with the phosphate backbone. While residues that are absolutely
conserved among al1 homeodomains contact the 5'-TAAT-3' core sequence, residue 50, the
ninth residue of the recognition helix, contacts the two nucleotides immediately 5' of the
core, providing additional binding specificity (Otting et al., 1990). These findings have

been confumed by the X-ray crystallography data with the engraiied homeodomain and the
MAT a2 homeodomain (Kissinger et al., 1990; Wollberger et al., 199 1).
2.2 Phylogeny of the horneobox
One of the biggest surprises in homeobox research was the finding that homeobox
genes are not confined to huitnies. An initial evolutionary survey showed that these genes
also exist in vertebrates, including chicken, rnice and man (McGinnis et al., 1984). To
date, homeobox genes have also been identified in orgmisms as divergent as sponges,
yeast and plants (Murtha et al., 199 1 ; Seimiya et al., 1994).
Subsequent work demonstrated that homeobox genes were not only present in
many organisms, but that the structural organization of the homeotic clusters had also been
conserved. Clustering comparable to Drosophila was found in nematode, mice and man
(Gehring, 1994). Whereas invertebrates contain only one or two clusters, mamtnals such
as mice and man have four homeotic clusters in their genome. These clusters seem to have
aisen from an Anfp type prototype gene through tandem duplications (Graham et al., 1989;
Boncinelli et al., 1988 and 199 1). In mice and man each of the four clusters, designated
Hox a through d, consists of 9 or 10 homeobox genes which c m be grouped into 13
paralogous groups. Astoundingly, the physical order of the paraiogous groups in the
cluster has been conserved between fruitfly and mammals (Duboule and Dolle, 1989;
Graham et al., 1989). Al1 the genes in the mouse homeobox cluster are transcribed in the
same orientation (Boncinelli et al., 1991) and their physical order in the cluster appears to
have functional significance, in that expression of the genes at the 3' end of the cluster is
developrnentally earlier, more anterior and more responsive to retinoic acid (Robertis,
1994). Two forms of selective forces have been suggested to be necessary for the
conservation of the cluster organization. One is cis-regulatory elements that are shared
between adjacent genes in the cluster (Peifer et al., 1987). The second may be that higher
order chromatin structures within the cluster are essential targets of trans-activators

(Jürgens, 1985; Gyurkovicz et al, 1990; Campbell et al., 1995). The extreme degree of
conservation at the nucleotide and genornic level suggests that very different body plans
might be achieved, both in insects and vertebrates, by evolutionarily conserved gene
networks that possibly have similar regulatory interactions.
Expression of the Hox genes in mice was found to demarcate specific regions of the
embryo, rather than tissues or ce11 types which are established later in development
(Gehring, 1994). Genes of the Hox clusters are not expressed anterior to the hindbrain,
and therefore they cannot be involved in the development of the more anterior CNS
structures of forebrain, rnidbrain and anterior part of the hindbrain.
To date, a large number of homeobox genes have been identified that, based on
their expression pattern, may contribute to the development of the antenor structures of the
CNS. Almost one third of the 30 distinct homeodomain classes known to date have been
implicated in some aspect of head development (Kappen et al., 1993; Robertis, 1994). Of
these the Pax/Prd, POU and LIM homeodomain proteins are charactenzed by additional
protein domains, while the Dlx, Otx, Emx, Gsc, Msx and the Nkx proteins are classified
solely on the basis of their homeodomain sequences (Robertis, 1994). Arnong these, the
Pax/prd class has received particular attention due to its involvement in genetic disease in
flies, rnice and man.
2.3 Padprd class of homeobox genes
The Paxlprd class of homeobox genes is characterized by the paired domain, which
was initially identified as a conserved region between the Drosophila paired and goosebeny
proteins. The paired dornain spans 128 amino acids (Bopp et al., 1986) and has been
shown to possess sequence-specific DNA binding activity (Chalepakis et al., 199 1 ;
Treisman et al., 1991). While the paired domain is characteristic of al1 nine marnmalian
Pax proteins, only four of them also contain a homeodomain. AU four of them encode a
serine in the crucial position nine of the recognition helix (Kappen. 1993). The vertebrate

Pax genes are thought to have arisen from the paired gene through tandem duplications
(Gruss and Walter, 1992). The Pax genes are dispersed throughout the genome,
suggesting that the chromosomal environment is unimportant in dictating their expression
pattern (Strachan and Read, 1994).
Studies of the in vitro target sequences of the Padprd genes have reveaied a
surprising difference between Pax and Hox homeodomain function. Many DNA binding
proteins bind DNA as dimers or higher order complexes. However, most Hox
homeodomains were found to bind DNA as monomers. The prd homeodornain on the
other hand has been shown to bind cooperatively to palindromic binding sites.
Cooperativity was also found upon exchange of the ninth residue of the paired
homeodomain from serine (S9), to glutamine (Q9) or lysine (Kg), residues commonly
found in other homeodomain classes. However, this amino acid exchange results in a
change in the magnitude of cooperativity and the binding specificities (Wilson et al., 1993).
Although the cooperativi ty found for the paired homeodomain was shown to occur on
DNA and not in solution, its basis is not known. Possible mechanisms for the
cooperativity include direct protein-protein interactions between the homeodomains, or
allosteric changes in DNA shape after binding of the first homeodomain (Wilson et al.,
1993). New data suggests that the changes in DNA shape occurring when paired
homeodomains bind are superimposable on those found for non-paired homeodomains,
implicating that direct protein-protein interactions are a more likely primary mechanisrn of
cooperativity (Desplan, un publis hed resul ts).
Pax genes are expressed primarily during embryonic development and al1 except
Pax1 and Pax9 exhibit a distinct spatial and temporal expression pattern in the developing
nervous system. Al1 the homeobox-containing Pax genes have been suggested to be
involved in the specification of positional information, possibly the A-P and D-V axis in the
developing spinal cord and brain (Strachan and Read, 1994; Chalepakis et al., 1993).

Five of the nine mammalian Pax genes, narnely Paxl. P d , Pax3. P d and P d ,
were found to be mutated in previously described disease loci in several species, and four
of these phenotypes involve the brain (Gruss and Walther, 1992; Read, 1995). The fact
that none of the marnrnalian Hox genes could be assigned to any disease locus has been
suggested to be due to the redundancy of the four conserved clusters (Strachan and Read,
1994). Most of the identified natural Pax mutations are loss-of-function mutations,
resulting in developmental defects, while the only gain-of-function mutation identified to
date confirms the oncogenic properties that have been shown in cell-culture studies (Galili
et al., 1993, Strachan and Read 1994). Mutation analysis and ectopic expression
experiments have recently established that one of the Pax genes is essential and suffîcient
for eye developmen t .
2.4 Pax 6/eyeless; Master control genes of eye development
Par6 encodes a paired domain and a S9 homeodomain. Expression is detected in
the optic pit as early as E8.5, and later extends into the eye vesicle, optic stalk, retina and
also the lens, comea, and ectoderm from which they develop. P d is therefore expressed
in al1 the structures of the developing eye (Gruss and Walther, 1992). Mutations in the
Pax6 gene and its orthologues were found to be responsible for eyeless in Drosophila
(Quiring et al., 1994), Small eye in mice (Hogan et al., 1988) and Aniridia in humans
(Chalepakis et al., 1993). Small eye animals, in the heterozygous state, have reduced body
size, smali eyes, vacuolated lenses, cataracts, abnomally folded retinae and a reduced RPE
layer. Homozygotes completely lack eye and nasal tissues and die postnatally due to the
failure to breathe. Aniridia is an autosomal dominant disorder with variable expressivity,
which is characterized by partial or complete absence of the iris and abnormal lens, retina
and optic nerve.
The fact that eye development is controlled by orthologous genes in both vertebrates
and insects suggests that eye morphogenesis may be under similar genetic control in al1

higher eukaryotes. Pnx6/eyeless is not only essential but also sufficient for eye
development in Drosophila. Ectopic expression of eyeless or Pax6 in Drosophila imagina1
discs results in the formation of ectopic eyes, with normal morphology and a full
complement of differentiated cells (Halder et al., 1995). This finding suggests that eyeless
is the master control gene for eye development in flies. Although Pax6 has been shown to
be crucial for proper mouse eye development, some rudimentary eye structures do develop.
Thus, Pax6 may not be a master regulator of eye development in mamrnals, even though it
can functionally substitute for eyeless in Drosophila. In contrast to the fruit fly, various
rnembers of the Pax/paired family as well as other transcription factors (Liu et al., 1994)
have been shown to be expressed in the mammalian eye. Consequently vertebrate eye
morphogenesis may require the orchestrated activities of other members of the Paxlprd
family, such as the homeobox gene CkiO, in addition to regulatory genes of other types.
3. C h x l O
3.1 ChxlO biology in the mammalian retina
Ch10 identifies a new subclass of prd-like homeodomain proteins. It was
originally isolated from an adult human retinal cDNA library in a differential hybridization
scnen for evolutionarily conse~ed genes that are expressed abundantly and specifically in
the retina (Bascom et al., 1992). The human and mouse cDNA's both contain an open
reading frarne encoding a protein of 361 amino acids which contains a homeodomain,
putative transactivation domains (proline rich, serinelthreonine rich, acidic), and a putative
nuclear targeting sequence (Liu et al. 1994). Comparison of the Chxi0 sequence with
those in Genbank revealed that the gene most similar to this novel clone is a Caenorhbditis
elegans homeobox gene, cailed cehl0 (Hawkins and McGhee, 1990; Svendsen and
McGhee, 1995), leading to the designation of CHXIO (Ç.elegans-like Homeoboz m. The similarity between the two polypeptides extends into a 57 arnino acid region
immediately C-terminal to the homeodomain (Figure 1-3A) which has also k e n identified

Name Homeodomain Sequence Identity
Figure 1-3
(A) Protein dot-matrix alignment of chxlO and its closest homologue cehl0. Alignment
was done on Geneworks 2.1 with a window size of 10 amino acids and cut-off at 70%
amino acid identity. The chxlO and cehl0 proteins are 361 and 344 amino acids in length,
respectively . (B) Alignment of the c h 1 0 homeodomain with similar homeodomains of the Paxlpaired
class of homeobox genes. Mismatches are shown and respective amino acid identities to
chx 10 are indicated. Adapted from Liu et al. (1994).

in the goldfish cDNA Vsxl as a 54 amino acid region (Levine et al., 1994). Due to the
high degree of evolutionary conservation from nematode to man, Svendsen and McGhee
(1995) suggested that this 54-57 amino acid residue sequence represents a functional
protein domain and named it the CVC-domain, for the first three clones containing this
domain (çhxlO, xsx 1, ~ehlO). Based on the overall homology in the homeodomain
sequence, Ch10 belongs to the PaxlPrd farnily of horneobox genes (Figure 1-3B). The
predicted chx 10 protein also contains an octapeptide sequence as do most of the Pax genes,
but it does not contain a paired domain. Among the PaxfPrd family mernbers, chxIO is
very similar to the Drosophila aristaless protein, with 75% amino acid identity in the
homeodomain and additional conservation in outside regions, which suggests that these
genes might have developed from a cornrnon ancestor (Liu et al., 1994).
The pattern of Ch10 expression in mouse development suggested that it may play
significant roles in the formation of the retina. Northem analysis in adult mouse tissues
revealed that ChxlO expression in the retina is abundant but not specific, since weak
hybridization was also observed in pons, medulla and spinal cord (Liu et al., 1994).
Immunofluorescence, using a polyclonal antibody raised to the N-terminus of the human
protein, showed that expression in the retina is further restricted. The chx10 protein is
observed only in the inner nuclear layer, in the nuclei of bipolar cells and amacrine cells,
and possibly in al1 ce11 types of this layer. Occasional weakly staining cells were observed
in the ganglion ce11 layer and are believed to be displaced amacrine cells. Within the inner
nuclear layer expression seems to occur in a gradient with higher levels at the outer border
(Liu et al., 1994).
The developmental expression of the murine Ch10 gene was studied by 3% in
situ hybridizations (Liu et al., 1994). Transcripts were first detected at E9.5 ai the tip of
the evaginating optic vesicle (these cells constitute the presumptive neuroretina), while no
staùiing was observed in the overlying head ectoderm which will give rise to the lem, or in
the presumptive optic stalk (Figure 1-4B). As the optic vesicle invaginates to fonn the

Figure 1-4
In situ hybridization with the ChxlO probe in the mouse retina. A, C, E, G are
micrographs taken under bright field illumination, B, D, F, H are rnicrographs taken under
dark field illumination. (A, B) A transverse section of the forebrain of a 9.5 dpc mouse
embryo at the optic vesicle stage. (C, D) A section of a 1 1.5 dpc embryonic mouse eye at
the optic stage. (E, F) A section of a 16.5 dpc embryonic mouse eye. (G, H) An adult
mouse retina section. Abbreviations: forebrain (0, ganglion ce11 layer (gcl), inner nuclear
layer (hl), lens (l), neuroblast layer (nbl), neuroretinal layer (ml), outer nuclear layer (onl),
optic stak (os), optic vesicle (ov), pigment epithelium (pe). Taken from Liu et al. (1994).

optic cup, Ch10 expression remains confined to the inner layer, the actively proliferating
neuroblast layer (Figure 1-4D/F). Upon differentiation and layer formation, expression is
extinguished in the ganglion and the photoreceptor ce11 layers, but remains in the inner
nuclear layer (Figure 1 -4H).
Outside the eye, the C h i 0 transcnpt is found in the hindbrain and spinal cord. At
E 1 1 Chxi0 is expressed throughout the posterior neural tube, from the rhombencephalic
isthmus into the spinal cord. Expression in the media1 pons and medulla oblongata
correlates with regions where cranial motor nuclei and reticular nuclei develop. in addition,
expression was observed at the border of the caudal thalamus and the pretectal area of the
rostral midbrain as well as lateral to the median groove of the upper lip. However the
chx10 protein has not yet been detected in these structures in adult mouse tissues using
immunofluorescence (D. van der Kooy and R. McInnes, unpublished observation).
The developmental regulation of Ch10 expression was studied by quantitative
Northern analysis on rabbit retinae. Transcripts were first detected at a time point
correlating with optic cup formation. Levels rapidly increase 3-4 fold as horizontal cells,
bipolar cells, amacrine cells and cone photoreceptors form. While rod outer segment
differentiate, expression drops to the constant level observed in the adult (Liu et al., 1994).
The highly restricted temporal and spatial pattern of expression of Chxi0 suggests
possible functions for the protein in the developing and mature mammalian retina. Liu et
al. (1994) proposed that ChxlO might specify ce11 identity within the inner nuclear layer
and give positional cues for lamination. They also speculated that absence of ChxlO
expression in the outer nuclear layer and the ganglion ce11 layer rnight be a result of, or a
requirement for, the formation of those layers. Functional concepts like these are usually
addressed by means of creating a mouse that lacks a hinctional gene product. In this case
however, the gene did not have to be targeted, since a natural mutation in ChxlO was
identified.

3.2 ChxlO mutations and ocular retardation
Ocrlur retardation (or) is a recessive mouse mutation that was initially described by
Truslove (1962) as a spontaneous mutation amongst a CBAICSïBL colony of mice.
Homozygous mice are viable. but born blind with small eyes that have thin and poorly
differentiated retinae (Figure 1 - S m ) . Additionally, the mutant developing retinae show
decreased morphogenetic ce11 death, cataracts, optic nerve aplasia and microphthalmia. The
disease is truly recessive in that no phenotype has ever been described for the heterozygous
animals (Robb et al., 1978).
Molecular analysis showed that a mutation in Chxl O is responsible for the ocular
retardation phenotype. Initially mapped to mouse chromosome 2, Chri0 was found to lie
physically close to the ocular retardation locus (B. Taylor, M. Burmeister and R. McInnes,
unpublished). In an attempt to refine the mapping with a CA-repeat of genomic ChxlO,
none of the 170 outbred mouse crosses displayed recombination between the CA-repeat
and ocular retordution, suggesting that Ch10 might be causative for the ocular retardation
phenotype (M. Burmeister and R. McInnes, unpublished). Andysis of the Chri0 locus in
genomic DNA from ocular retardation Nce, did not reveal any gross rearrangernents.
Sequence analysis of the Chxi0 locus in one of the disease alleles, 04 showed a C to A
conversion in codon 176. This change converts a tyrosine to a premature stop codon,
resulting in a truncation of the protein within the homeodomain, indicating that mutations in
Chxl O mutation are causative for the ocular retardution phenotype. Immunofluorescence
showed that no c h 1 0 protein is ever detected in the orJ retina at any stage of development,
even at times such as El 1 when the mutant eye is morphologically normal although
possibly slightly reduced in size.
The morphological abnormalities in oJ rnice suggest a critical requirement for the
ch10 protein in eye development. The o d mutation initialiy manifests at E10.5, at which
time there is a virtual absence of morphogenic ce11 death with only 2% of the normal
number of necrotic cells being detected in the mutant neuroblast layer. Mutant retinae
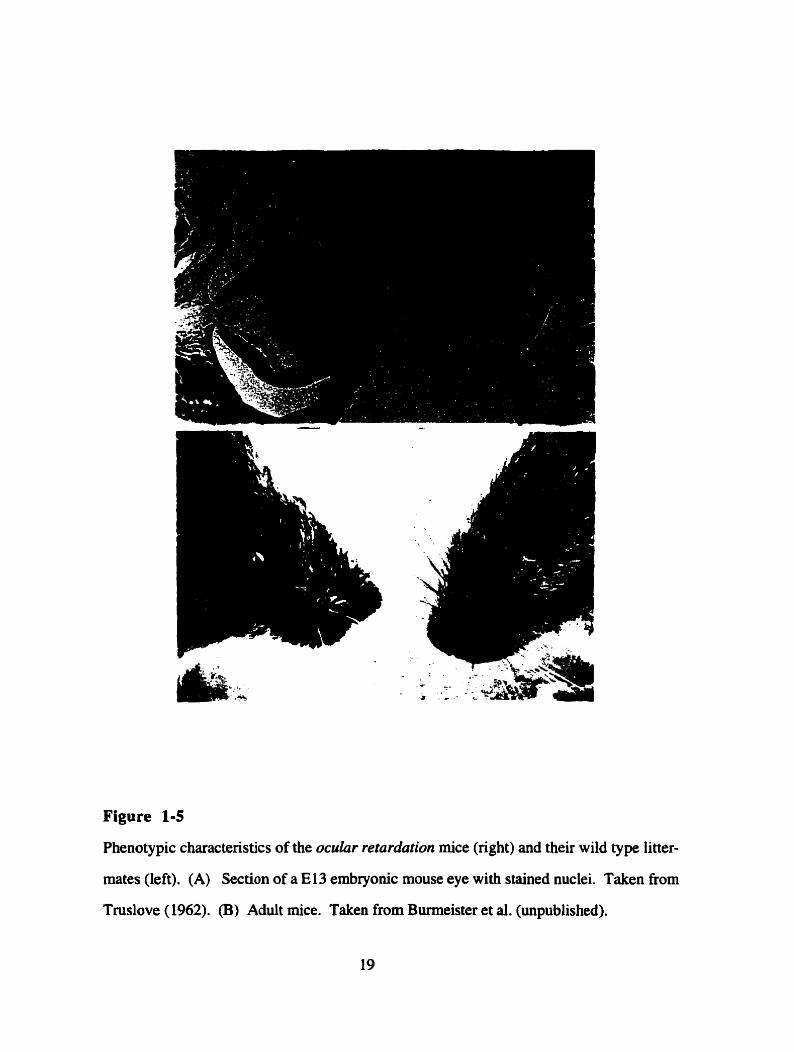
Figure 1-5
Phenotypic characteristics of the ocular retardation mice (right) and their wild type litter-
mates (left). (A) Section of a El3 embryonic mouse eye with stained nuclei. Taken fiom
Tmslove (1962). (B) Aduit mice. Taken from Burmeister et al. (unpublished).

cease to develop, stay single layered and only develop a rudimentary optic fïber tayer
(Figure M A ) . The use of ce11 type specific antibodies, indicated that many major ce11
types were present, except bipolar cells. Thus, rod photoreceptors, ganglion cells and
amacrine cells are present and situated in their correct location, but neither of two bipolar
ce11 markers detected staining, suggesting that these cells are either absent or poorly
differentiated. This suggests that in addition to being necessary for general proliferation
and celi death in the developing retina, ChxlO appears to be required for the ce11 fate
determination of at least bipolar cell progenitors (M. Hankin and R. McInnes, unpublished
observations).
The identification of the mutation in ocular retardation adds to the group of mouse
mutants with eye disease phenotype for which mutations have been assigned (Pax6, Gii3,
Rrra, rds and Mi). The mutant phenotype in ocular retardation, however, is restricted to
the eye and is only detected in homozygous animals. Both the severity and the specificity
of the disease phenotype suggest that Ch10 is crucial for the proper development of the
mamrnalian retina. Evidence from our laboratory and others now suggests that chx 10
specifies a novel subclass of paired-like homeodomain proteins.
Throughout the following text 1 will be referring to homology between cDNAs or
proteins in the sense of sequence similarîty, without infemng close common ancestry. The
term orthologues will be used to describe genes from distinct species that represent the
most recent bifurcation of one common ancestral gene.
3.3 ChxlO homologues in mice and goldfish
Genomic ChxlO homologues were isolated in a screen of a genornic DNA library
for the Ch10 gene. Using a full-length mouse ChxlO cDNA as the probe, genornic
clones covering the entire ChxlO locus were obtained. In addition, a second class of
clones was observed which hybridized specifically, but less strongly, to the Ch10 cDNA.
These clones fell into three different classes, arbitrarily named 15 1, 17 1 and 48 1, and one
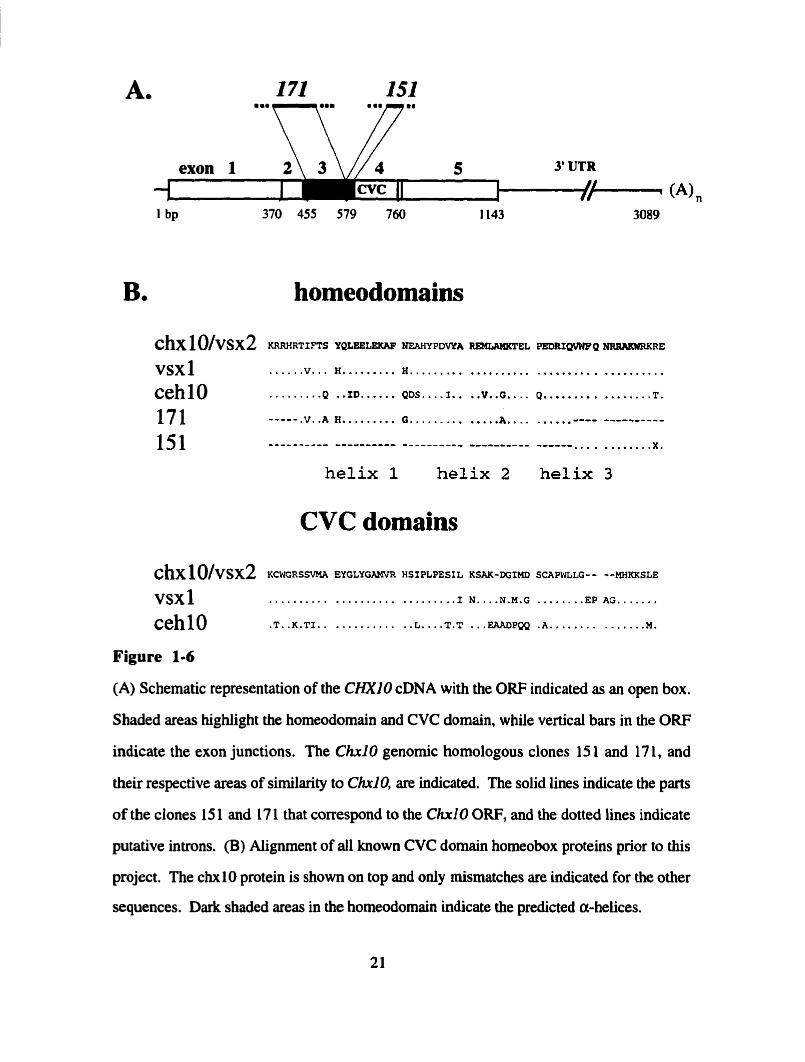
151
3' UTR
370 455 579 760 1143 /+ (A),,
3089
B. homeodomains
C ~ X 1 O / V S X ~ KRRHRTIFTS YQLeBLElW? NEAliYPDWA -EL PWRXQVWFQ
...... .............................. vsx 1 V... Il......... H.........
. * .... .. . . ........ ceh 10 .........Q XD...... QDs 1.. v..o.. Q......... T.
171 ----- .v..A H. ........ G . . . . . ......... A.. . . ..,.,,-..-- -i-----ir-
151 ---..*_-a-- ---------- --------*- -_-------- ------ . . . a . . . . . . . . X .
helix 1 helix 2 helix 3
CVC domains
C ~ X ~ ( ) / v s x ~ KCWGRSnmR EYGLVGAMVR HSIPLPESIL KSAK-DGIHD SCAPWLLG- - --MHKKSLE
............................. V S X ~ 1 N....N.M.G ........ EP AG....... ............ ... ....... cehl0 .T..K.T.ï.. L....T.T EAADPQQ . A . . . . . . . . M.
Figure 1-6
(A) Schematic representation of the CHXlO cDNA with the ORF indicated as an open box.
Shaded areas highlight the homeodomain and CVC domain, while vertical bars in the ORF
indicate the exon junctions. The Chrl O genomic homologous clones 15 1 and 17 1, and
their respective areas of similarîty to CMO, are indicated. The solid lines indicate the parts
of the clones 15 1 and 17 1 that correspond to the CM0 ORF, and the dotted lines indicate
putative introns. (B) Aiignment of al1 known CVC domain homeobox proteins prior to this
project. The chxlO protein is show on top and only mismatches are indicated for the other
sequences. Da& shaded areas in the homeodomain indicate the predicted a-helices.

representative of each class was malyzed. Sequence analysis within the shonest
hybridizing fragments showed the existence of potential open reading frames. In al1 three
clones these partial ORFs displayed significant homology to the ChxlO homeodomain and
displayed some conserved splice junctions with Ch10 (Figures 1-6A/B; J. Novak and R.
McInnes, unpublished).
Ch10 homologues have also been isolated from the goldfish retina. Levine and
Schechter (1993) canied out a PCR screen for homeobox genes in the goldfish retina and
identified Vsxl and Vsx2, which both encode a homeodomain and a CVC domain. The
vsx2 protein has 100% amino acid identity to ch10 in homeodornain and CVC domain and
is therefore likely to be the ChxlO orthologue of goldfish. Vsxl, on the other hand, shows
significant changes in both domains, and thus represents a Vsx2/ChrlO homologue (Figure
1 -6B; Levine et ai., 1994). Remarkably, Vsxl is expressed in the inner nuclear layer of the
goldfish retina, a location like that of ChrlO. This result indicates that these genes are
likely to be true homologues and that the homology extends beyond the nucleotide and
amino acid level to functional characteristics.
4. RATIONALE FOR THIS PROJECT
The isolation of partial mouse genornic ChxlO homologues by Jakub Novak and
the cloning of one vertebrate retinal homologue by Levine et al (1994), suggested that
ChxlO homologues are likely to exist in mammals. Homologous genes arise through
duplications of ancestral genes, which leads to functionally redundant units that
subsequentiy diverge in sequence, expression pattern, and function. Due to this process,
highly homologous clones ofien overlap considerably in their expression patterns (Strachan
and Read, 1994). For this reason, ChxlO homologues are likely to be expressed in the
retina, as is already indicated for Vsxl. Mammalian Ch10 homologues, or alternatively a
mammalian Vsxl orthologue, may therefore be represented in a retinal cDNA library,
which is why the screen for these homologues was undertaken.

CHAPTER 2: ORIGINAL RESEARCH
INTRODUCTION
This chapter describes my work on the identification of human C H X I O
homologues. To identiQ such genes, a human retinal cDNA library was screened with a
human CHXlO probe at high and low stringency. Human CHXlO clones were eliminated
from the screen using PCR analysis for CHXIO 3' UTR sequences. PCR-negative clones
were plaque purified to homogeneity and phage inserts were subcloned into a suitable
sequencing vector. Of the clones that have been analyzed to date, three were found to
encode homeodomain proteins, two of them novel (CHXIO-I and OTX3) and one
representing the previously identified human LIM homeobox gene LH2 (Xu et al., 1993).
To date 30 clones remain to be subcloned and anaiyzed. Sequence analysis of the CHXIO-
1 cDNA established its close hornology to the CHXlO cDNA in homeodomain and CVC
domain. The expression of CHXIO-1 in adult human tissues was analyzed by Northem
blotting and preliminary in situ hybridization, which showed that CHXIO-I expression
levels are far below those of CHXlO in adult retina. In addition CHXIO-I exhibits a
distinct layer specificity from CHXlO in that i t is expressed at equal levels in the inner
nuclear and the ganglion ceIl layer (Liu et al., 1994).
Cloning of a homologous cDNA from a mouse retinal library led to the
identification of the cDNA encoded by the 17 1 genomic locus, Ch.171. Sequence analysis
for Ch171 established that it is quite distinct from CHNO-1 in nucleotide and predicted
protein sequence. In contrast to CHXIO-1 in humans, C h i 7 1 is expressed at high levels
in the adult mouse retina. Genomic Southem blots were perfomed in order to establish the
phylogenetic relationship between CHXIO-1 and Chxl7l. This analysis showed that no
mouse sequence is more similar to CHXIO-I than Chxl7I is. However it seems that
CHXI O- 1 is not the closest homologue to ChxI7l in human genomic DN A, suggesting
that a human CHX17I cDNA might exist.

EXPERIMENTAL PROCEDURES
Human retinal cDNA library
tissue source: adult neuroretina
primer: oligo-dT
arnplified: no
vector: hgt 10
cloning site: EcoM
host: BNN 102 or C600 hfIA
independent recombinants: 2.106
selection: tetracycline
source: Jeremy Nathans
reference: Bascom et al., 1992
Human fetal brain (cortex) cDNA library (Stratagene #936206)
tissue source: 17- 18 week late abortion, normal femde
primer: oligo dT and random
size selected: SSkb, average 1 .O kb
amplified: once
primary plaques: 2- 106
estimated background: 4% non-recombinants
vector: WIi
cloning site: EcoRi
helper phage: R408 or VCSM 13
host: XL1-Blue
selection: tetracyciine
source: Stratagene, La Jolla, CA.

Mouse brain cDNA library
tissue source: adult mouse cortex
primer: oligo dT and random
size selected: average s ix 1 kb
amplified: yes
vector: AZAPII
cloning site: EcoM
host: XLl Blue
selection: tetracycline
source: Hayden M., UBC
Mouse newborn eye cDNA library
tissue source: Mouse eye PO-3
primer: oligo dT
size selected: >1 kb, average size 1.5-2.0 kb
amplified: once
background: >O. 1 %
number of independent clones: 1 1.6- 106
vector: pcDNA, derivative of pCDM8
cloning site: BstXI (destroyed)
host: MC10611P3
selection: ampicilllli
source: Yang Xianjie, Harvard Meàical School
Mouse retinal cDNA library (h BALBZ)
tissue source: 1 month-old Balbk mice

primer: oligo dT
amplifieci: once
vector: Charon BS(-)
cloning site: HindIII and EcoRI
host: Kg02 or LE392
selection: none
source: Anand Swaroop
reference: Qais Farjo et al., 1993.
Phage library screening
Phage were diluted in SM buffer (Sarnbrook et al., 1989). Host bacteria were
cultured overnight at 370C in 0.2% maltose LB with lOmM MgCI2, then harvested and
resuspended to an OD600 of 0.6- 1.0 in lOmM MgS04 (Sambrook et al., 1989).
Adsorptions were carried out at 370C for 20 minutes. Bacteria and adsorbed phage were
mixed with top agarose (0.7% agarose in LB, 1OmM MgCl2) which had been melted and
cooled to 50%, plated onto 1.5% agar LB plates, and incubated at 370C for 6 to 12 hours.
Hybond-N+ membranes (Amersham) were used for plaque blotting as outlined in Ausubel
et al. (1989): replica filters were incubated twice for 5 minutes in denaturing solution (0.5
M NaOH, 1.5 M NaCI), twice for 5 min in neutralizing solution (1.5 M NaCI, 0.5 M Tris
pH 7 3 , 5 min in 2x SSC (Sarnbrook et al., 1989), fixed for 5 min in 0.4 M NaOH, and
rinsed for 30 sec in 5x SSC. Filters were prehybridized for a minimum of 3 hours in 30%
or 50% formamide, lx Denhardt's solution, 5x SSPE, 0.1 % SDS and 100 mg/ml
denatured salmon-sperm DIVA at 420C (Sarnbrook et al., 1989). Hybridizations were
carried out for a minimum of 8 hours at 420C in buffer of the same composition as the
prehybridization buffer with the addition of 10% w/v dextran sulphate with approxirnately
1-2x106 cpm/rnl of hybridization buffer. Probes were labeled to a specific activity of
- 1x109 cpm/pg by the random hexanucleotide prirning method (Feinberg and Vogelstein,

1983) using [ ~ - ~ ~ P I ~ c T P (NEN). Labeled probes were spun on Sephadex G-50
(Pharmacia) columns that had been prewashed with an equal volume of 0 . 0 1 ~ TE pH 8.0,
0.1% SDS, denatured with 0.2 volumes of 1 M NaOH for 5- 10 min, and neutralized by
adding Tris pH 7.5 to a final concentration of 0.5 M before being added to hybridization
buffer. Membranes were washed twice for 20 min in 2x SSC, 0.2% SDS at room
temperature for both stringencies and twice for 20-30 min in either 2x SSC, 0.2% SDS at
500C for low stringency or 0 . 2 ~ SSC, 0.2% SDS at 65OC for high stringency. Washed
membranes were exposed to XAR5 X-ray films (Kodak) with intensifying screens at
-7OOC.
Phage DNA analysis
Phage DNA was prepared according to Grossberger ( 1987). For analytical
purposes 1 pg of phage DNA was digested with 10-20 U of restriction enzymes using
conditions specified by the enzyme manufacturers. Digests were loaded on 0.7-18
agarose gels in lx TAE (Sambrook et al., 1989) and electrophoresed in 1 x TAE.
Photographs were taken with a Siratagene Eagle Eye photodocumentation system.
Plasmid library screening
The transformed plasmid library was diluted in LB and plated onto numbered 132-
mm Hybond-N+ membranes on top of LB agar plates containing ampicillin (100 pg/ml) at
a density of 80,000 colonies per plate, and incubated upside down for 12 hours at 370C.
Replica filters were prepared according to Ausubel et al. (1989) and incubated for a few
hours until the colonies had just appeared. DNA on the replica lifts was denatured,
neutralized and fixed as recornmended by Ausubel et al. (1989). Filters were then soaked
in 2X SSC for 5 min and washed in 5x SSC, 0.5% SDS and 1mM EDTA for 30 min at
650C to remove ce11 debris. Prehybridizations, hybridizations and washes were done as
outlined above for phage library screening.

Subcloning
Cloning vector (pBluescript KS(-), Stratagene)was linearized and dephosphorylated
as outlined in Sambrook et al. (1989). Digests of phage DNA vector were electrophoresed
on 0.7% agarose gel in l x TAE. Selected fragments were cut out and DNA purified from
the gels using either GENECLEAN (BI0 101, CA) or QIAEX (Qiagen, CA) purification
kits as specified by the supplier. The DNA was resuspended in 5pl of lx TE pH 7.5. The
yield of DNA was estimated by the band intensity of a 1 pl aliquot in a lx TAE gel using a
k - H W Z size ladder as quantitation standard. Roughly 50 ng of vector and enough phage
insett for a molar ratio of 1 to 2-3 were rnixed with 0.5 pl of T4 Ligase (BRL) and 1 pl of
5x T4 ligase buffer (BRL) in a total volume of 5 pl, and the reactions incubated 3- 12 hours
at room temperature. 1-2 pl of the reaction mixture were used to transform 25 pl of DHSa
competent cells (BRL) as specified by the manufacturer. The transformed cells were plated
ont0 LB plates with 0.08 mg/ml X-gal, 0.2 rnM IPTG, and 100 pg/ml ampicillin, and
incubated ovemight at 370C.
CHXlO 3' UTR PCR
The 250 bp 3' UTR fragment of CHXlO was amplified by PCR (Saiki et al., 1988)
as single fragment from 5 CI of the pnmary library picks in at total volume of 25 pl using
the primer set 5'-AACAATTGGGAGCACATAGCC-3' and 5 ' -
AAG'ITGCAACGCACïTGCTC-3'. The components used in the amplification were: 50
mM KCI, 10 mM Tris pH 8.3, 1.5 rnM MgCl2, 0.011 gelatin, 200 pM of each dNTP,
and 200 ng of each primer. In a Perkin Elmer themocycler, an initial denaturation was
carried out for 5 min at 980C, followed by 35 cycles. Taq polymerase (10 units) and light
minera1 oil were added after the initial denaturation step. The amplification profile for the
PCR was 950C for 30 seconds, 5S°C for 30 seconds and 720C for 30 seconds.

CHX10-1 genomic PCR
The 164 bp fragment of the CHXIO-1 open reading frarne was amplified by PCR
(Saiki et al., 1988) as single fragment from 50 ng of human genomic DNA in a total
volume of 50 pl using the primer set 5'-CCATGGGGATGATAAGGAAGC-3' and 5'-
GAGGTCAATAGCCACATCTTCC-3'. The components used in the initial amplification
were: 50 m M KCl, 10 mM Tris pH 8.3, 1 .S mM MgC12,0.01% gelatin, 200 FM of each
dNTP, and 200 ng of each primer. In a Perkin Elmer themocycler. an initial denaniration
was carried out for 5 minutes at 94OC, followed by 35 cycles. Taq polymerase (IOU) and
light mineral oil were added after the initial denaturation step. The amplification profile for
the PCR was 94OC for 40 seconds, 600C for 30 seconds and 720C for 30 seconds.
PCR for the genetic mapping was done using a MouseMamster Hybrid Panel (#2
from Coriell Ce11 Repositories, Panel#2, Carnden, NJ, USA). The components used in the
initial amplification were: 67 mM Tris pH 8.8, 6.7 m M MgCI2, 10 m M P- mercaptoethanol, 16.6 rnM (NH&S04,6.8 pM EDTA, 170 pg/rnl BSA, l .S mM dNTPs
each, 10% DMSO, and 200 ng of each primer. Initial denaturation, amounts of Taq
polymerase, and the amplification profile were as described above.
Plasmid DNA analysis
Plasmid DNA was prepared by the boiling prep method from Sambrook et al.
(1989) with slight modifications (J. Rornrnens, pers. comm.). In brief, bacteria were
collected from 3 ml of ovemight culture. The pellet of bacteria was resuspended in 250 pl
of STET and 30 pl of lysozyme (10 mg/rnl) were added. Bacteria were placed in a rolling
boil for 55-60 seconds. After centrihigaiion the supernatant was precipitated in two
volumes of ethanol, incubated at room temperature for 5 minutes and centrifuged in a
micro-centrifuge for 5 minutes. The pellet was resuspended in 300 pl of 0.3 M Sodium
acetate pH 7-7.5 and 750 pl of ethanol were added (room temperature for 5-15 minutes).

After centrifugation the pellet was washed in 70% ethanol, re-centrifuged and air-dried at
room temperature for 5- 10 minutes.
Sequence analysis
DNA was sequenced using the T7 polymerase DNA sequencing kit from
Phannacia. Sequencing reactions were carried out with [ ~ ~ S I ~ A T P (NEN) according to the
Pharmacia protocol, except that reactions were performed in a Nunc micro-titer plate.
Reactinns were run on 6% polyacrylamide gels (acrylamide/N'Nt-bis-methylene-
acrylamide ratio 36.5/1,0.053% APS, 0.052% TEMED) at 1500-2000 V, gels were fixed
with a 10% methanol, 10% glacial acetic acid solution, dried at 800C and exposed to an X-
ray film (Kodak). Sequences were compareci to released sequences at Genbank using the
BLAST server ([email protected]; Altschul et al., 1990). Sequence alignrnents
were performed on Geneworks 2.1 from Intelligenetics using standard parameters.
Map location of known human or mouse phenotypes
The chromosomal location of human genetic diseases or mouse phenotypes were
determined using the on-line Mendelian inheritance in man (OMIM) server at the Genome
Data Base (http://gdbwww.gdb.org/gdbdoc/topq.html) or the Jackson Laboratory Mouse
Locus catalog (http://www.infonnatics.jax.org/mgd.html), with a Netscape 0.96 software
package.
Eye donations
Human eyes for retinal RNA preparations were obtained from the Ontario Eye Bank
(Toronto).

Total RNA purification
Total RNA was prepared from tissue samples using the T W O L method (GIBCO
BRL, Cat.#15596, 1992).
Poly-(A)+ RNA purification
Poly-(A)+ RNA was prepared from total RNA using a Oligotex-dT purification kit
(Qiagen, Catalog #70022). Purification is based on hybridization of the poly-(A) tail to
oligo-dT primers that are coupled to a solid phase matrix. Since hybridization depends on
high salt concentration, the poly-(A)+ RNA is subsequently eluted in low ionic strenght
buffer.
Northern blot analysis
Total RNA and poly-(A)+ RNA gels were prepared and blotied according to
Sambrook et al. (1989). Hybridizations were carried out in the high stringency
hybridization buffer under the conditions described above, and washed using the same high
stringency washing conditions.
Tissue preparation for in situ hybridization
The neuroretina layer was removed from human eyes (Ontario Eye Bank) and
rinsed once in PBS, followed by a two hour fixation in 4% paraformaldehyde in PBS at
room temperature. Sarnples were dehydrated by passage through an ethanol series (70%,
85%, 95%, two times 100%) and embedded in paraKin. Retinai sections (-7pm thickness)
were cut and placed on pretreated, RNAse fkee slides. The slides were subsequently baked
ovemight in an oven at 55-600C (Y.Zhou, pers. c o r n ) .

Digoxygenin in situ hybridization
Digoxigenin labeled sense and antisense nboprobes were generated by in vitro
translation of a non-conserved part of the CHXIO-1 cDNA as probe (nucleotide 758-
1696). In situ hybridization were carried out following a protocol from Boehringer
Mannheim.
Genomic Southern Blot snalysis
Human and mouse genornic DNA were digested, electrophoresed and blotted as
outlined in Sambrook et al. (1989). Hybridizations were done in the buffer described
above at 50% and 40% formamide concentrations, while membranes were washed at 650C
in 0 . 2 ~ SSC, 0.2% SDS and 550C in 2x SSC, 0.2% SDS, respectively.

RESULTS
1. Attribution of data
Primary screen of 1.5- 106 p h (Lin Liu)
PCR analysis and plaque purification for class 1 (David diCiommo and Melanie Schumach)
Physical mapping of the OTX3 gene to chromosome 19q13.3 (Aiessandra Duncan)
Physical mapping of CHXIO-1 to chromosome 20 (Catherine Duff)
Physical mapping of CHXIO- I to chromosome 20p 1 1.2 1 q 1 1.2 1 (Henry Heng)
Northem analysis for Ch171 (Bryan Snow)
2. Human retinal screen for CHXZO homologues
Three million plaque forrning units (phi) of an unamplified oligo-dT primed adult
human retinal cDNA library were screened by hybridization to a 680 nucleotide Apal
fragment of the human CM10 cDNA, which spans the homeobox and the CVC-box.
Duplicate lifts were screened at high and low stringency (see Figure 2- 1) and clones were
grouped according to the presence or absence of a signal at either stringency (Figure 2- 1).
Three classes of clones were identified: Class 1 clones, which gave signals in low
stringency only, Class 2 clones, with signai at low and high stringency, but with markedly
reduced signals at high stringency, and Class 3 clones, with almost equally strong signals
under both conditions. Based on the high degree of sequence similarity between CHXlO
and both mouse clone 17 1 and goldfish V s x l , a true CHXI0 homologue should be
expected to appear in Classes two and three.
In order to exclude CHXI0 clones from the screen, ail primary picks were screened
for CHXIO-specific sequences by PCR, using primers to the CHXlO 3' UTR,
imrnediately upstream of the poly-(A) tail. Most clones from an oligo-dT primed library
should contain this part of the cDNA. Al1 primary picks of Classes 2 and 3, totaling 450,
and Class 1, totaiing 85, were screened using this PCR approach. The expected band size
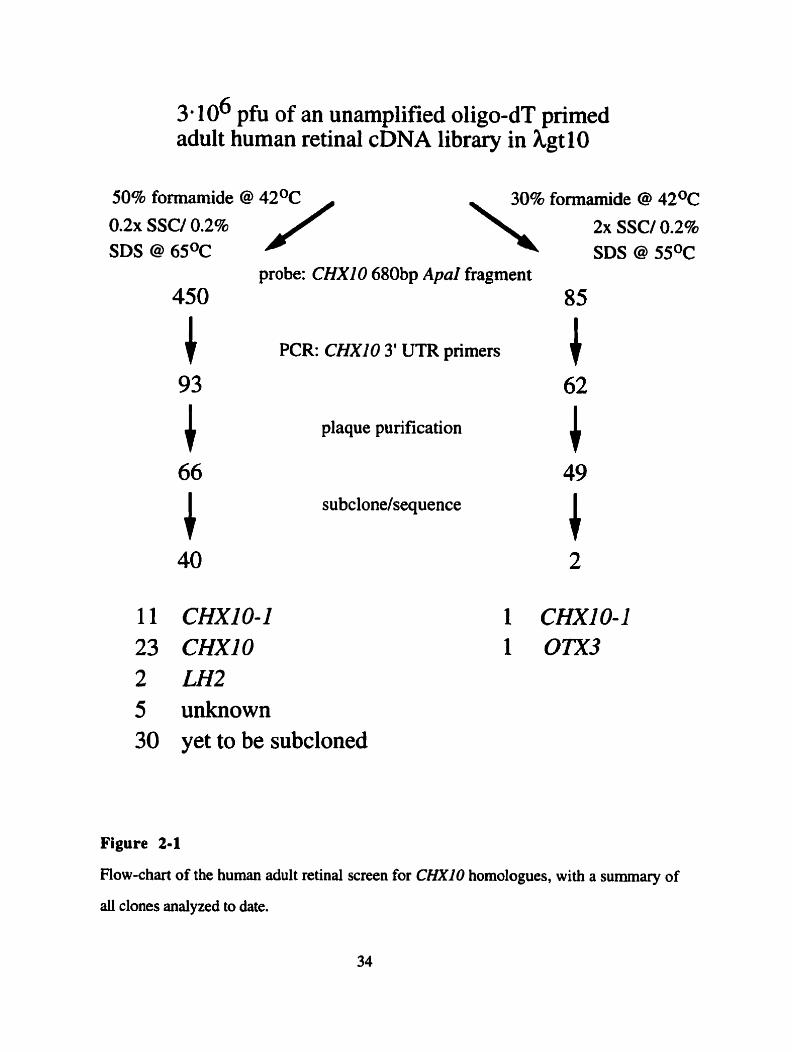
3 a 106 pfu of an unamplified oligo-dT primed adult human retinal cDNA library in hgt 10
0 . 2 ~ SSC/ 0.2% SDS @ 6SoC
probe: CHXlO 680bp ApaI fragment 85
PCR: CHXlO 3' UTR primers l plaque purification
subclonelsequence
11 CHXIO-1 23 CHXlO 2 LH2 5 unknown 30 yet to be subcloned
Figure 2-1
Flow-chart of the human adult retinal screen for CHXlO homologues, with a summary of
di clones analyzed to date.

of 144 bp was reproducibly found for 357 of the 450 Class 2/3 clones and 23 of the 85
Class 1 clones, suggesting them to be CHXlO cDNAs. Ninety-three of the high stringency
clones were shown not to contain this most 3' part of the CHXlO cDNA, and 66 were
plaque purified to homogeneity (Figure 2- 1). Similarly, 49 of the 62 PCR negative Class
1 clones were plaque purified to homogeneity (Figure 2-1).
Plaque pure phage of Classes 1,2 and 3 were dot-blotted on a lawn of host bacteria
and incubated uniil phage plaques becarne visible. Multiple lifts were hybridized to the 680
nucleotide Apal fragment of the human C M 1 0 cDNA at high and low stringency. This
analysis confinned the initial classification (Figure 2-2). Subsequently phage DNA was
prepared, inserts were excised and gel purified. and subcloned into pBluescript for
sequence analysis. My studies of 25 of these clones have led to the identification of three
homeobox containing cDNAs, OTXJ, LH2 and CHXIO-1 , which are described in the
follow ing text.
3. OTX3, a novel orthodenticle homologue
The clone described in this section was actually isolated from a previous low
stringency screen for CHXIO homologues that was initiated by Midhat Osman and
Elisabeth Garami. In general, this screen was not technically satisfactory, but it did yield
one homeobox-containing clone. Due to the conditions used to isolate this clone, it is
categorized here as a Class 1 clone. Sequence analysis revealed that this clone encoded a
homeodomain-containing ORF that was tmncated at the 5' end. Due to this truncation only
a partial homeobox sequence is available, which is highly homologous to the mouse Otx
homeoboxes, with 81% nucleotide identity to Otxl and 77% nucleotide identity to 01x2
(Figure 2-3A). Cornparisons were only made to the mouse genes since the sequence of the
human genes has not been made public. The changes in the nucleotide sequence result in
the substitution of four of the 36 amino acids of the homeodomain that are encoded in this
cDNA clone (Figure 2-3B). This finding suggests clearly that this cDNA represents a

Figure 2-2
A Dot-blot of 59 plaque pure phage of Classes 1, 2 and 3 were hybridized to a 680
nucleotide Apal fragment of the human CHXlO cDNA probed with 680ApaI at low
stnngency (see Figure 2-1) in panel (A) and high stringency in panel (B). Typical
representatives of the isolated clones, are clone #l for CHXIO, clone #97 for C H X l O- I
and clone #22 for LH2.

A. Consensus CGA . GGGAG . G . AC. AC. TT . AC. . G .. CA CAGCT . GACG T .... G . . . . . . . . T ... c . 60
O a l otx2
. . . C. .... C .C..C..C.. C..GC.CT.. ..... G.... .GCTC.AGGC Gt7i'G.TCG.A
... A. .... A .G..G..T.. T..TA.GG.. ..... C.... .TCTG.AAGC TCTG.TTG.C
. Consensus . .G. c . C. . T ACCCAGAC . T CT. .... .G . GA. GAGGTGG C.CT. m . AT CAA. TGCC. 120
OTX3 GT.V.C.AG. ........ G. ..ATGCCC.T . . G....... .T..G..G.. . . . TC. ... T ot~f AA.A.T.GC. ........ A. ..TCATGC.C . . G....... .A..C..G.. . . . CC....A 01x2 AA.A.C.GG. . . . . . . . . A. ..TCATGA.G .. A....... .A..G..A.. ... CT....A
Consensus GA. TCCAG . G T . CAGGT . TG GTT . MGM. CG. . G . GC . A A. TGC . G . CA . CAGC . . CAG I 80
om3 .. G... ..G. .T.....T.. ... C.....C ..GA.G..T. .A...A.G.. G....GA... Ou1 . . G.. . . . A. .C.....T.. ... C.....C ..CC,C..C. .G...C.C.. G....AG... Oa2 .. A.....G. .G.....A.. ... T.....T ..AA.A..T. .G...c.c.. A....AG...
otx 1 otx2
otx3 otx 1 otx2
ou3 otx 1 otx2
otx3 otx 1 otx2
oix3 otx 1 otx2
YAREEVALKI NLPESRVQVW FKNRRAKCRQ QRQQQKQQQQ PPGGQAKARP AKRRA----- 7 1 FMREEVALXI NLPESRVQVW FKNRRARCRQ QQQ------- -SGSGTKSRP AKKKSSPVRE 117 FMREWALKr NLPESRVQVW FKNRRAKCRQ QQQQQ----- QNGGQNKVRP AKXKSSPARE 90
WSPASESPLX EAQRAG---- LVASG--PSL TSAPYAMTYA PASAF---CS SPSAYG---- 159 WSPASISPGS APASVSVPEP LAAPSNTSCM QRSAAGAAT AAASYPMSYG QGGSYGQGYP 237 WSPASISPLS DP-------- LSTSS--SCM QRS-YPMTYL QA----------- SGYSQGYA 160
-SPSSYFSGL D--PYLSPMV PQLGGPALSP LSGPSVGFSL AQSP---TSL ---SG----- 205 TPSSSYFGGV DCSSYLAPMH SHHHPHQLSP MAPSSMAGHH HHHPWHPL SQSSGHHHHH 296 -GSTSYFGGM DCGSYLTPMH HQLPGPG-AT LSPMGTNAVT SHLNQSPASL ---ST----- 210
QSYG- ---AYSP-VD SLEFKDPTG- ----m3KFTY NPMDPLDYKD Q-SAWKPQIL. 248 HHHHHQGYGG SGLAFNS -AD CLDYKEPGAA AASSAWKfrNF NSPDCLDYKD Q- ASWRPQVt . 354 ---a- QGYGA SSLGFNSTTD CLDYKDQTA- ----SWKLNF NA-DCLDYKD QTSSWKFQVL. 3 9
Figure 2-3
Multiple alignments of 0TX3 with its two closest homologues Otxl and Oa2. (A)
Nucleotide alignment of the respective homeobox sequences. (B) Amino acid alignrnent of
the respective 5' truncated peptides, with identical areas shaded.

novel homeodomain protein, which we named OTX3. In situ hybridization (ISH) mapped
this gene to human 19q 13.3 (A. Duncan, Queens University, pers. cornm.). The different
map locations of OTXl (12~13, Kastury et al., 1994) and OTX2 (14q21-22, Kastury et
al., 1994) confirm the conclusion that 0TX3 represents a novel gene. No diseases
involving eye or neural tissues have been linked to this location in humans. Similarly, no
abnormal mouse phenotypes map to the homologous syntenic region of the mouse genome,
chromosome 7, between 1 and 22 centi Morgans (CM).
4. Human LIM homeobox gene t H 2
Sequence analysis of clones #22 and #29 showed that they both represent part of
the previously identified cDNA for the human LIM homeodomain gene LH2 (Xu, et al.
1993). The homeobox of this gene has 601 nucleotide identity to the CHXIO homeobox
and no significant similarity to any other region of the C M 0 cDNA.
5. CHXIO-2, a novel CHXIO-like homeobox gene
5.1 Cloning
A novel CHNO-like cDNA was initially identified from the overlapping clones #4
and #28. This cDNA contains both a homeobox and a CVC box, and was named CHXIO-
1, due to its overall similarity to CHXIO. Reprobing of the plaque pure phage dot blots at
high stringency showed that another 14 clones hybridized strongly to both #4 and #28 and
thus most likely represented the sarne cDNA (data not shown). Subcloning and sequencing
of seven additional clones enabled the identification of a 1947 bp long composite cDNA
(Figure 2-4AB). The CHXIO-1 transcript contains an intemal EcoRI site which resulted
in the loss of the most 3' 250 bp in many of the clones (Figure 2-4A), most likely when
excising the insert from the EcoRI cloning site. This fiagrnent most likely constitutes part
of the cDNA since it has been isolated from three unique clones. The CHXIO-1 cDNA
encodes a full length OW. In the 5' UTR the CHXIO-1 cDNA contains an in-fiame ochre

CAAGCGCGAC GTCTCTGGAT CCCGTTTGCT TGCTiGGAA CCATGACCGG CCGGGACTCG CmCCGACG GGCGCACTAG CAGCAGGGCG 90 - M T G R D 5 L S D G R T S S R A CTGGTGCCTG GCGGTTCCCC TAGGGGCTCG CGCCCCGGGG ClTCGCCATC ACGGACCTGC TGGGCTTGGA GGCCGAGCTG CCCGGCGCCC 180 L V P G G S P R G S R P G A S P S R T C W A W R P S C P A P GCTGGCCCAC GACAGGGATC TGGCTGCGAG GGTCCGGCAG TCGCGCCGTG CCCGGGCCCG GGCTTGACGG CTCCAGCTGG CGCGTGGGGG 270 A G P G Q G S G C E G P A V A P C P G P G L T A P A G A W G CCCTACCGCT GGGACTCGGC CTCCTCTGTG GCTTCGGCAC GCAGCCGCCG GCGCCGCTCG AGCACCCTGC CTGCTCCTAG CGGACGTGCC 360 P Y R W D S A S S V A S A R S R R R R S S T L P A P 5 G R A GTTCCTGCCG CCCAGGGGCC CCGAGCCCGC TGCCCCGCTG GCTCCCAGCC GTCCGCCGCC TCCGCTCGGC CGCCAGAAGC GCAACGCAGC 450 V P A A Q G P R A R C P A G S Q P S A A C A R P P E A - , Q - R - S GTCTCCACGT CCGATGAGGA CAGCCAGTCT GAAGACAGGA ATGACCTAAA GGCATCCCCC ACCTTGGGCA AGAGGAAGM%@GCG~'C 540 V S T S D E D S Q S E D R N D L K A S P T L G K R K ~ ~ R ~ ~ R ~ ~ ~ , AGGACACjiTt $ACTGCTCAa CCAGCTGGM GAGTTCGAGA A6GCATTCAC;~CC;CACGCCÇAC TACCCyCAtE.TbTATCC5~~~~CCT~ 630
.-i.'y2.~ . j= :A .H'' Q-L.:E*, :E- ~,E.:Y - ' : ~ : . ~ : , ~ : : , ~ ~ L ; ' ~ ~ ' . - ~$.p;~$jY~;&'~ . +. &.:gr. 6 - w CKAGff CCC CWGACCGG ATACAGGTCT :Gm..AA ; c c G ~ G $ G c ~ * Q $ ~ ~ ~ A c c ~ ~ G M GCGCTGCGGC 720 ; k : 1 v w - : Q ~ & R W GGCAGCAGCG TGATGGCCCA GTACGGGCTG TACGGCGCCA- TGGTGCGCCA CTGCATCCCG CTGCCCAGAC TCCGTGCTCA ACTGCCCGAG 810 G S S V M A E Y G L Y G A M V R H C I P L P R L R A Q L P E GCCGGCCTGC TGGGCTCCTG CGCGCCCTGG CTCCTGGGGA TGCATAAAAA ATCCATGGGG ATGATAAGGA AGCCAGGMG TGAAGATAAG 900 G G L L G S C A P W L L G W H K K s u c M I R K P G S E D K TTGGCAGGAC TCTGGGGCTC TGACCACTTC AAAGAAGGTT CTAGCCAGAG TGAGTCACGA TCACAGAGAG GCTCAGATAA AGTGAGCCCT 990 L A G L W G S D H F K E G S S Q S E S G S Q R G S D K V S P GAGAATGGCT TGGAACATGT GGCTATTCAC CTCTCCAGCT CTGCCCGGCA GGAGACCAAG AAAGTGCACC CTCGGGCTGG TGCtCAAGGA 1080 E N G L E D V A I D L S S S A R Q E T K K V H P G A G A Q G GGCTCCAACT CCACGGCACT CGAGGGGCCC CAGCCAGGGA ACCTGGGAGC CACATGAGAC CCACAGGTCC CACTGTCAAA GGCTGAAAAT 1170 G S N S T A L E G P Q P G K V C A T . GTGCATATTC CTCACTGGCA T m C C A A A A GACAAAAATT GTCCAAGACA TATACTCTCA G m G A T r T CITCTGACAA TGTCAAGATA 1260 AAATGCAATG CCACTTGCTT TAAGAGGACA GATGAGTGAC OAGGGAAAA TTCTAf fTCA TTCTAGAATG GAAACGGITC CTTAATGGCA 1350 CTTTAACCAG TTAACTTGGT GCAAAACTTT TWCTCTT TTGTAGCTTG AGCAGGGAGG ATCCGAAGïT CAAAAATGAT TTCACTGCTG 1440 GGCATCATTT GAGATGTGTG GTTCAATGTA CTGTATAGAT GAATAGGTTA GGTCTGATGT CCTCATTCTC AATTAGGAAA TATAAAATTG 1530 TTTAATATCC AGAAGAATCT CATATCATCA TAGTGAAGAC TCCATACAGA CACATGAATG AATTATATK T C m A C C T T GAACTTGGCC 1620 T T G G A T A ï l l C A A f m C r r GTTAGGGAGT AATTTTGTGT TTTCTTGACT CTCACrrGAT AGGCTATCCA TATTGGAATT CAAAGAAACC 1710 CATACTCTAG T C G m C T G C A T T T C C m A AAAATAGCAT GGGATCATAT TTAAACAATT GTTGGATAGC TGGAAACAGT C r r m A C G T 1 8 0 CAGACCAGGC T i T K G T C l T TCAAAATTTG TGATTGAAAG C A C m A G T G A C r r G G A l l l TGTAAATCTT T C C m C C T G C G f l l C C T T T 1090 GCCATTTATT TATTCTTTAA A A U C A T ITTGTGGT GT GTTAAAAAAA AAAAAAA 1947
Figure 2-4
(A) Schematic representation of the CHXIO-1 cDNA and al l clones that were isolated from
the human retinal screen. (B) Nucleotide and predicted amino acid sequence of the human
CHXlO-1 cDNA. Shaded areas represent the homeobox (dark) and the CVC box (light).
The upstream in-frame stop codon is double underlined. The putative poly-adenylation
signal in the 3' UTR is underlined.

stop codon (TAA) nine nucleotides upstream of the start codon at nucleotide 1 (ATG). The
sequences surrounding the ATG (GAACCATGA) correspond well to the Kozak sequence
(CC(80%A, 2O%G)CCATGG(40%)) for translational starts (Kozak, 1984). The open
reading frame between nucleotide 1 and the opal stop codon (TGA) at nucleotide 1092
encodes a predicted protein of 364 amino acids, with predicted molecular weight of 39
kDa. At the nucleotide level the CHXIO-1 cDNA is 54% identical to CHXlO over the
entire ORF, with the highest conservation in the homeobox (78% nucleotide identity) and
the CVC box (77% nucleotide identity). The CHXIO-1 3' UTR contains a poly-
adenylation site at nucleotide 1869, 22 nucleotides upstream of a poly-(A) tail,
corresponding well with the average distance of 1 1-30 nucleotides (Lewin, 1990).
CHXIO-1 encodes a homeodomain of the prd-like type (Bürglin, 1994; residues
163 and 222), with a glutamine at position 50; the residue at this position has been shown
to be crucial in determining the DNA binding specificity (Treisman et al., 199 1). The
homeodomain sequence is most sirnilar to the chxlO homeodomain with 92% amino acid
identity and less similar to the cehl0 homeodomain with 78% arnino acid identity (Figure
2-5A). The chx 10-1 protein also contains a CVC-domain (residue 223-276), immediately
C-terminal to the homeodomain, with 70% amino acid identity to the chxlO CVC domain
and 6 1 % amino acid identity to the cehl0 CVC domain. Outside of the homeodomain and
CVC domain chx 10 and chx 10- l are highly dissirnilar with 17% amino acid identity in the
N-termini and 19% amino acid identity in the C-termini. In these outside regions, the
chxl0-1 protein is slightly more sirnilar to the ceh10 proiein in the N-terminus (22% amino
acid identity) but slightly less similar in the C-terminus (12% amino acid identity).
5.2 Putative protein domains
Comparison of the chxl0-1 peptide to sequences in Genbank led to the
identification of several other putative protein domains. The predicted chxlO-1 protein
contains sequences that resemble putative activation domains (Figure 2-5B). N-tenninal to
chxl0-1 - - nuciear targeting
N--~CVC= -C octapeptide
O 100 200 300 aa
Figure 2-5
O
100 -
chxlO 200 -
300- aa
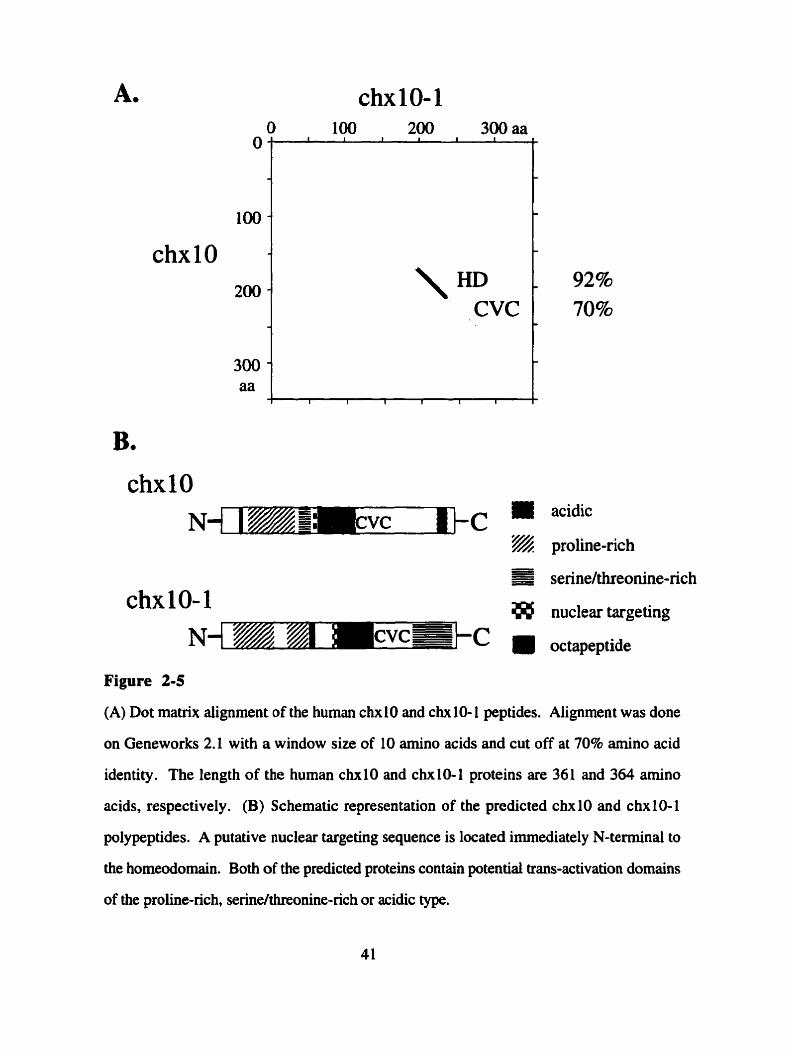
(A) Dot matrix alignrnent of the human chx 10 and chx 10- 1 peptides. Alignment was done
on Geneworks 2.1 with a window size of 10 amino acids and cut off at 70% amino acid
I 1
\ HD CVC
I I I 7 I I
identity. The length of the human chxlO and chxl0-1 proteins are 361 and 364 amino
-
-
- 92% 70% -
C
acids, respectively. (B) Schematic representation of the predicted chxlO and chxlO-1
polypeptides. A putative nuclear targeting sequence is located immediately N-terminal to
the homeodomain. Both of the predicted proteins contain potential trans-activation domains
of the proline-rich serinelthreonine-rich or acidic type.

the horneodomain is a region with 24% proline content, reminiscent of the proline-rich
activation domains found in CTFlR/3 (Mitchell and Tijan, 1989; Mermod et al., 1989).
Similar to CTFI, this region is split up into two blocks (residue 18-77 and 100-131).
Interestingly, two irnperfect repeats (CEAPAGPG and AECPGPG), comparable to those
described in the AP-2 proline-rich region, can be found in block 1 (Williams and Tijan,
199 1). C-terminal to the proline-rich region is a 12 residue sequence (residues 140 to 15 1)
that contains 50% acidic amino acids (two glutamic acids and four aspartic acids),
comparable to the longer acidic activation domain found in the AP-I/Jun/TFIID proteins
(Mitchell and Tijan, 1989). In the C-terminus of chxl0-I there is a 31 amino acid region
(residue 300-330) with 32% serinehhreonine content resembling the trans-activation
domain described for the POU homeodomain protein GHF-1 (Theill et ai., 1989). In
addition, this region is also fairly acidic (248 between 284-326, with 5 glutamic acids and
4 aspartic acids). The chx 10- 1 protein aiso encodes a putative nuclear targeting sequence,
as defined by Dingwall and Laskey (1991). This sequence, between residues 160-167
(KRKKRRHR), extends five amino acids into the homeodomain.
5.3 Mapping to the chromosomes
In order to determine whether CHXIO-1 might be implicated in human disease, the
gene was mapped by both PCR and in situ hybridization. PCR primers were devised for
use on a chromosome-hybrid panel. Since this technique requires amplification from
genomic DNA. it is best to devise primers within one exon. 1 assumed that the genomic
structure of CHXIO-1 might be quite similar to CHXIO, and therefore primers were
constructed to a region of CHXIO-1 (nucleotides 820-840 and 959-980) corresponding to
exon 4 of the CHXlO gene (Liu et al., 1994). 1 demonstrated that these primers ampli@ a
fragment of the predicted size (160 bp) from human genomic DNA. These pnmers were
found to ampli@ a single band of 160 bp from chromosome 20 of a Coriell PCR panel of
human chromosome hybrids (C. Duff, Hospital for Sick Children, pers. comrn.). This

result was confirmed and refined by Henry Heng (Hospital for Sick Children) using clone
#4 (Figure 2-4A) for fluorescent-in situ-hybridization (FISH). A strong signal was
observed on human chromosome 20, in a band corresponding to the region of 20p 1 1.2 1 -
q 1 1.2 1 (Figure 2-6A/B).
5.4 Northern analysis on total RNA
In order to determine the tissue specificity of CHXI0-I expression in adult human
tissues, a multi-tissue Northern blot, containing 5 pg of total RNA from retina, brain, liver,
kidney, spleen, fibroblasts and lymphoblasts, was hybridized with a CHXIO-1 probe
(nucleotide 758- 1698) at high stringency. No signal was detected in any of the tissues after
a 2 day exposure. Even with a 10 day exposure, only a faint band corresponding to
roughly 2 kb, could be recognized in the retinal RNA lane, which is absent from the other
lanes (Figure 2-7A). The low abundance of the CHXIO-I transcript is in sharp contrast to
the high level of expression of CHXlO (Liu et al., 1994), but was confirmed upon
reprobing of the Northem membrane with a CHXlO probe, which reproduced the strong
signal seen by Liu et al. (1 994) after only ovemight exposure (data not shown).
5.5 Northern analysis on poly-(A)+ RNA
To confirm the expression of CHXIO-1 in the retina, and to measure the size of the
transcript, blots of poly-(A)+ RNA blots were performed. Poly-(A)+ RNA was prepared
from 120 pg of total RNA from human lymphoblasts and retina, leading to an estimated
yield of 2-3 pg of poly-(A)+ RNA. Hybridization with the CHXIO-f probe at high
stringency revealed a distinct band of 2.05-2.25 kb specific to the retina lane only (Figure
2-7B). This corresponds well with the predicted CHXIO-1 cDNA length of 1950 bp,
when considering an average poly-(A) tail of roughly 200 (A)'s, as found for other
transcripts (Lewin, 1990).
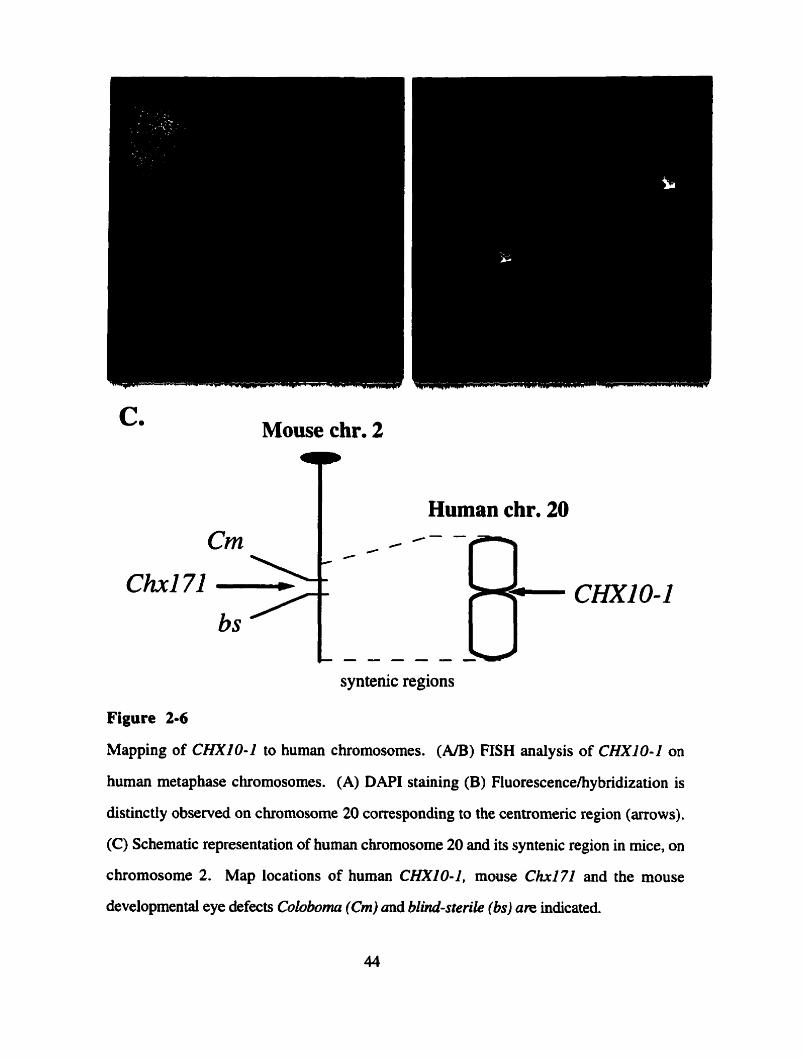
C. Mouse chr. 2
t
syntenic regions
Figure 2-6
Mapping of CHX10-2 to human chromosomes. (NB) FISH analysis of CHXIO-1 on
human metaphase chromosomes. (A) DAPI staining (B) Fluorescencelhybridization is
distinctiy observed on chromosome 20 corresponding to the centromeric region (arrows).
(C) Schematic representation of human chromosome 20 and its syntenic region in mice, on
chromosome 2. Map locations of human CHXIO-1. mouse Ch172 and the mouse
developmental eye defects Coloboma (Cm) und blind-sterile (bs) are indicated.

Figure 2-7
Northem analysis of CHXIO-1 in adult human tissues. (A) Total RNA blot after 10 day
exposure shows a faint signal in only the retina lane. (B) Poly-(A)+ RNA blot after
ovemight exposure exhibits a distinct band in only the retina lane.

5.6 In s i tu hybridizations
In order to detexmine whether CHXlO-1 is expressed in a layer specific manner
within the adult retina, in situ hybridizations were performed. Using paraformaldehyde
fixed sections of adult human retinae, staining was observed specifically with the antisense
but not the sense probe. The staining was observed in the inner nuclear layer and the
ganglion ce11 layer (Figure 2-8).
6. isolation of mutine CHXlO-1: Chx171
6.1 Cloning
Due to the difficulty of examining gene expression in man, and the relative ease of
studying development in the mouse, 1 decided to isolate the mouse orthologue of CHXIO-
1. However neither two million plaque forming units of an adult mouse brain library (M.
Hayden, pers.comrn.) nor one million colony fomiing units of a newborn eye library (Y.
Xinajie, pers.cornrn.) produced any positives when screened with clone #28 (see Figure 2-
4A) at high stringency. However 220 hybridizing clones were observed when screening
2.6 million plaque forming units of an adult mouse retina library (A. Swaroop,
pers.cornm.). Subcloning and sequencing of 1 1 arbitra~ily picked clones showed that they
al1 represented the same cDNA (Figure 2-9A). Sequence analysis of the two longest clones
MR3 1D and MR42, lead to the identification of a 2.6 kb long cDNA (Figure 2-LOB). This
cDNA may not be full-length since clone MR27 exhibits a 3' UTR that is 0.8 kb longer.
However, this 3' UTR sequence was not included in the analysis since it has only been
found in one clone. Cornparisons with Our database revealed that the isoiated cDNA
exhibited 100% nucleotide identity to the open reading frame fragment of 123 base-pairs
within the genomic clone 171, leading to the narne Chxl71. The 5' UTR of the cDNA
contains an in-frame arnber stop codon (TAG) 18 nucleotides upstream of the Start
(GAAGCATGA), which corresponds well to the Kozak sequence (CC(80%A,
20%G)CCATGG(40%)) for translational starts (Kozak, 1984). The open reading frame

Figure 2-8
In situ hybridizations of sections of adult human retina with CHXIO-1 probes. (A) Some
non-specific hybridization to the sense probe is observed in the imer nuclear layer (INL).
(B) Specific hybridization is observed in the i ~ e r nuclear (N) and the ganglion ceil layer
(GCL) after hybridization with the antisense probe. The bar in (A) represents 100 Pm.

B. GAAilCAAGC GAGGACTCAA CTCCGGCTCC T A G C O A A G GACTGAGAAG CATGACTGGA CGGGATGGGC llTCGGATGC GCGCTCCAGG - M T G R D G L S D A R S R
AGTAGAGCCC TGGCGCCAGG CTGTCCTCCC ACCGGCTCTC GCCnCCAAG C m G C C A T C AATGACCTGC TGGGATfGGA GGCACACCTG S R A L A P C C P P T G S R L R S F A 1 N O L 1 G L E A D L CCAACTCCGG CGGAGCCTGG GCTAAGATCC AACAGCGGAG ATCCAGCCGA GGCAATAGGG TCGGGACCGG GACCGGCGCC AGGACTCTGC P T P A E P G L R S N S G D P A € A I G S G P G P G P G L C GGCTCCTGCC CTGCTCGCGG CGCTCTCCCG CTGGGCCTGG CGCTCCTCTG CGGCTTCGCG GCGCAGCCCC CTTCCGTGCC GCCGCCGCTC G S C P A R G A L P L G L G L L C G F G A Q P P S V P P P L GTGCGCGCTG CCTGCTCCTG CCCGACTGCG GCTTCTGCCA TCTGCAGGAC CCGAGCCCTG GTCTCCCCCA GGGCCCGGTC CACCCGCGCC V R A A C S C P T A A S A I C R T R A L V S P R A R S T R A TGCGCTCGGC AGCCAGCAGC GCAGCGGAGC GTCTCCACGT CGGATGGGCA CAGTCCATCT CMGAAAAGA ATGACCCGAA GATGTCCCTT
AGAAAACCAG AAAGTGAAGA CAAGTTGGCA GGACTCTGGG AGTTTGACCA TCTCAAAAAG GGTGCTAATA AGGATGAGGT GGACCCTGAG R K P E S E D K L A G L W E F D H L K K G A N K D E V D P E AGGGGGCCAG ACGAAACAAC CCAGAACCCT GAGAATAGCT TGCAGGATGT GGCCATTGAC CTGTCCAGCT CTfCCAGGCA GGAGACTAAG R G P O E T T Q N P E N S L E D V A I 0 L S S S S R Q E T K AAAATGCCCC CAGGGTCCAG TACTCAGCTG CCCCAGCCCC AACAGGTGGG AGCCTCATGA GACTGACAGA TTGCGTCCCC CAAAGCTGGA
K H P P G 5 S T Q L P Q P Q Q V G A S . CAAACACCTT CAGTTYGATT l lCTCTCTGA AATCTCTGGA TAAAAAGCAA CACTACAACA CTGAATAAAC AATGACGTAG GGAAAGTTCC C m A G f f C T AGAGTGGGGA ACATKATAC CACCTCTCAA GCCAGTCAGT TTGGTCTAAA ATCTTGTTï l TCCTTAGTGC AATCTGAACA TGGAAGGTTT GAAGTTCAAA CAAAATTTTA GGGATGGGGT ATGACGTCGC TTAGTTAATA GTACACAAGA AGGCCTGGAT TCAATACCCG GCACCACTCA AACCAGATAC AGTAGCACGC ACCTGCATCC CAGCACTCAG GGGCCGGAGA CAAGAGATCA GATGTTCAAG AGCATCCCAG AGAGAGAGAG AGACAGAGAG AGACAGAGAG AGATTATTAC TCCCTAGCAT TATTfAAGAT GTGGTTCACT CATTAAAGAG GGCTGGTCTG ACCTCCTCAC TCTAAGTTAG AATATTTGGT TTGGTTl?GG TGAAATGGG TTCTfGCTGT GCAGCCCAGG CICGCTTCAT TTTCATGATC TCCTGCCTCA GTCTCTCTAG TGCTGGGTTA CAAGTClGAA TATAATTGTT ATACCATCTC TAGGCTTCAG AGGAGTGAAT GAATCAATGA ATGAATGACT GCTATAiTAC CATTCCTTGA GCTGGGTCfA GGACATCTCT GTCTTTGTGA TAGGTGATM TTTTGTCrrC TCAGTCACn G A T G G G C n GAATATTCGA GGTATAAACA ACAAAGAAAG CCACAAGCCC CTCCTCTCTG AAGTCACCTG GAGCCATCTT TAAGCAAl lG TTAAATATCT GGAGACTCTC TGAAGGTCAG ACCCAACTTC TTGGTCTTTT TGAATTTATG TTTGAAAGCA C m A G T A A C TTGCATTCAT GAATGilGTC TTCCCTGTAT T T C r m G C C ATTTATTTGT TAITTAAAAA TAAATGTGTC GTGTGTCTGG CTCAAAGCTA CTfCCTGGCC CCTCAGTAGG CTCTCCCCrr ATGCCCCGIl TTAGTCTGGA AACATCCCGA GCAAGCTGTC TTCAGCCCG CAGAACTTAG CACAGGGTCT GTGTCATGTA CTGTGTGGCA GTAGCCCTGG ClTi7GGAGT TCAGAGGCTG AAGAAGGGTT G T C T G t m G GGCAGCTTCC TCTGCAGATT ATCTCAGGAA CCCATCAAGT TAGGCTGTCC CCACCITCAC CCTACTCTCT 'I'TAGATAAGA AAACGGGCAG T C T C n C C l T TAGTCTAGGT TTGTCCTATG TTAGTTCCAT A G T W A C A C TCTATATCAA TGAACfATTA ACCCTGTGAG ATGATGGTGA AATGTTGCTA GGTGAGAAGC CCrrAGAGGG GGAAAGAGAT AAGAGGAAAT A A A G A T G m AAAGTACACT GAAAATAAAA TAAATAATAT TCGAAAAAAA AAAAA
Figure 2-9
(A) Schematic representation of the Chxl71 cDNA and al1 the isolated clones. (B)
Nucleotide and predicted amino acid sequence of the Mouse CM71 cDNA. Shaded areas
represent the homeobox (dark) and the CVC box (Iight). The in-frame stop codon
upstream of the translation start site is doubly underlined and the predicted octapeptide
singly underlined. Putative poly-adenylation signals in the 3' UTR are underlined.

between nucleotide 1 (ATG) and the opal stop codon at nucleotide 1086 (Tee) exhibits
74% nucleotide identity with the nucleotide sequence of the CHXlO-I ORF. The ORF
encodes a predicted protein of 362 arnino acids, with predicted molecular weight of 39
kDa. The 3' UTR contains a poly-adenylation site at residues 2548 and 2574 ( M A A A ) ,
46 and 20 nucleotides upstream of a poly-(A) tail, respectively. The second poly-
adenylation site at nucleotide 2574 corresponds better with the average distance of 1 1-30
nucleotides (Lewin, 1990).
The chxl7 1 polypeptide contains a homeodomain of the prd-like type (Bürglin,
1994), situated between residues 170 and 229. The homeodomain of chxl71 is 97%
identical to that of chx10-1 and less similar to the ceh10 homeodomain with 78% amino
acid identity. Like dl known CVC domain-containing homeodomain proteins, the CVC
domain of chxl7 1 is situated immediately C-terminal to the homeodomain, and is 80%
identical to the CVC domain of chx 10- 1 and 6 1% identical to the CVC domain of ceh 10.
In addition, chxl7 1 encodes an octapeptide of the HSIDGILG type (residue 3 1-38:
FAINDLLG) as it has k e n found in some members of the Pax/prd class of homeodomain
proteins including chxlO (Noll, 1993) and is similar in seven out of eight residues to the
octapeptides of chxl0, cehl0 and vsxl (Liu et al., 1994; Svendson and McGhee, 1995,
Levine et al., 1994). The chxlïl protein and the chxl0-1 protein are 42% identical in their
N-termini and 62% identical in their C-termini. Over the eotire length of the predicted
protein chx 17 l is 60% identical to chx 10- 1. The complex of homeodomain and CVC-
domain is found 7 amino acids more C-terminal in chxl71 than in the chxl0-1 peptide
(Figure 2- 10A).
6.2 Putative protein domains of chu171
The predicted chx17 1 protein contains sequences that resemble putative activation
domains (Figure 2-10B). N-terminal to the homeodomain is a 81 amino acid region
(residues 22-102) with 21% proline-content, reminiscent of the proline-rich activation
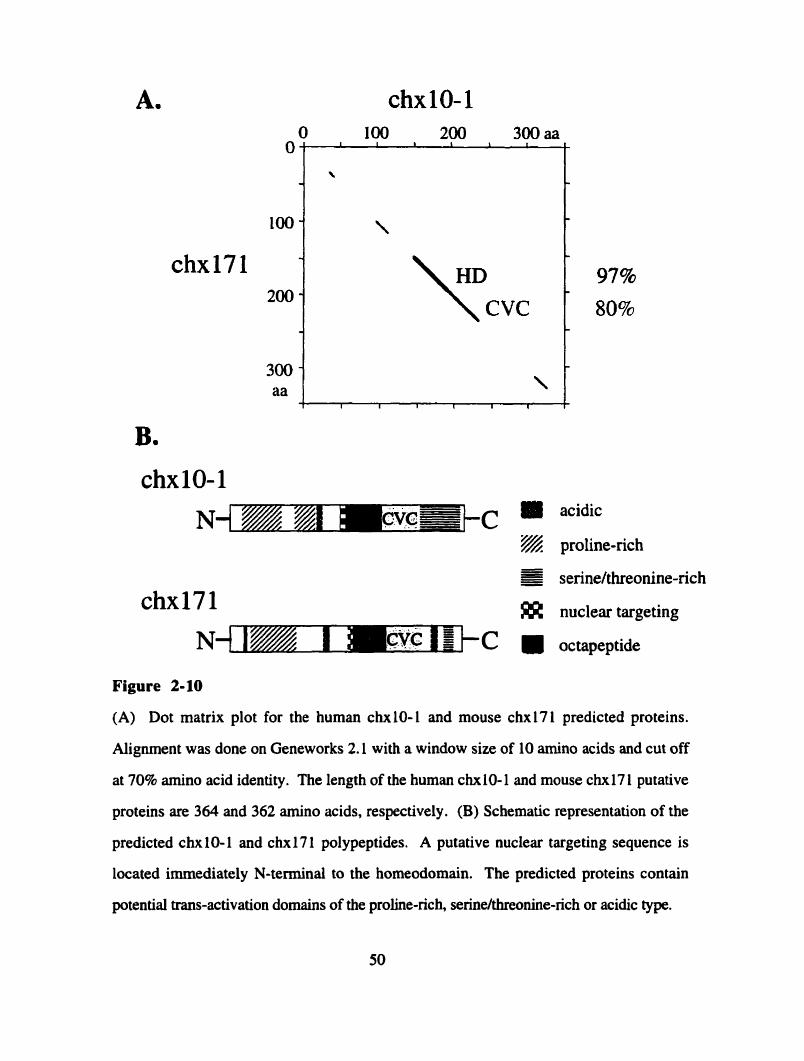
acidic
proline-rich - - s - - serinehhreonine-rich -
nuclear targeting
octapeptide
Figure 2-10
(A) Dot matrix plot for the human chxl0- 1 and mouse chxl7 1 predicted proteins.
Alignment was done on Geneworks 2.1 with a window size of 10 amino acids and cut off
at 70% amino acid identity. The length of the human chxl0-1 and mouse c h 1 7 1 putative
proteins are 364 and 362 amino acids, respectively. (B) Schematic representation of the
predicted chx 10- 1 and chx 17 1 polypeptides. A putative nuclear targeting sequence is
located immediately N-temiinal to the homeodomain. The predicted proteins contain
potential trans-activation domains of the probe-rich, seriw/threonine-rich or acidic type.

domains found in CTFIR/3 (Mitchell and Tijan, 1989; Memod et al., 1989; Williams and
Tijan, 199 1). C-terminal to the proline-rich domain, is an 1 1 residue region (residues 148-
158) that contains 45% acidic amino acids (2 glutamic acids and 3 aspartic acids),
comparable to the longer acidic activation domain found in the AP-I/Jun/TFIID (Mitchell
and Tijan, 1989). An acidic dornain is also found C-terminal to the homeodomain, in a 22
amino acid region (residues 308-329) with 41% acidic amino acids (five glutamic acids and
four aspartic acids). Further towards the C-terminus is a 17 iunino acid region with 4145
serine/threonine content, resembling the Iarger trans-activation domain found in the POU
homeodomain protein GHF-I (Theill et al., 1989). A putative nuclear targeting sequence
(Dingwall and Laskey, 199 1) identicai to that of chx 10- 1, is situated between residues 167-
174 (KRKKRRHR), this sequence extends five amino acids into the homeodomain.
6.3 Northern analysis
Chxl71 (nucleotides 1269 to 2337) was hybridized to a Northem blot of mouse
tissues, with 5 pg of total RNA from retina, brain, heart, liver, kidney, lung, testis, spleen
at high stringency. Transcripts were detected in the retina lane after only overnight
exposure at levels comparable to those of ChrlO, at a position corresponding to a length of
-3.1 kb (Figure 2-1 1; B. Snow, pers. comm.).
6.4 Phylogenetic relationship to C H X I 0-1
Sequence alignments between CHXIO-1 and CM71 left doubt whether they really
do represent orthologues or actually are homologues. Compared to the sequence
conservation of CHXI0 across species (Figure 2-12) it seems striking that CHNO-1 and
Ch171 have such a relatively weak conservation of sequence outside the homeodomain
and the CVC domain. In addition, the presence of 2 amino acid differences in the
homeodomain (60 amino acids) and 10 amino acid differences in the CVC domain (54
amino acids) is unexpected, particularly since several of these changes are nonconservative
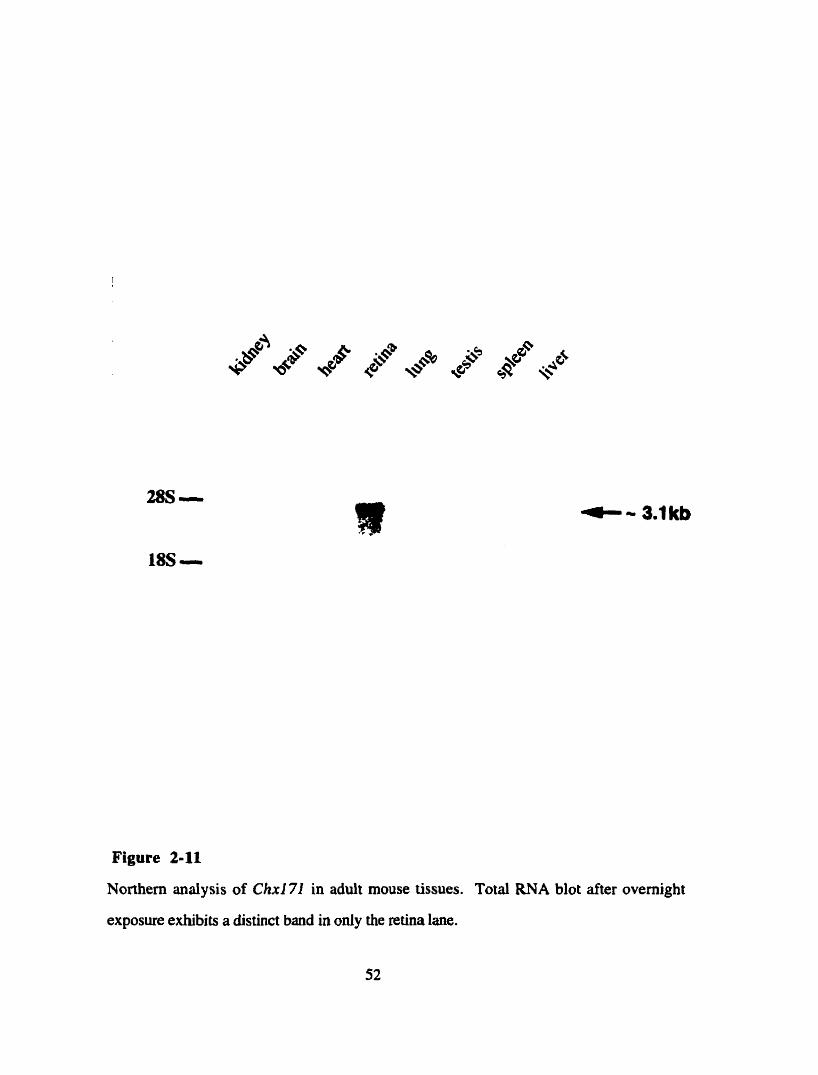
Figure 2-11
Northem analysis of Ch171 in adult mouse tissues. Total RNA Mot after overnight
exposure exhibits a distinct band in only the retina lane.

chxlO homeodomains
Human -RTI
Mouse . . . . . . . . .
Chicken .........
Goldfish . . . . . . . . .
chxlO CVC domains Human KCWCRS- EYGLYG- HSIPLPESII KS-ImS C A P W L L G m KSLE
...................................................... Mouse Chic ken ..................................... .E. ............ v.
...................................................... Goldfis h
Figure 2-12
Multiple protein alignments demonstrating the sequence conservation of the chxlO
homeodomain and CVC domain between goldfish, chicken, mice and man. Dots indicate
identical amino acid sequences, while mismatches in the amino acid sequence are indicated.
Dark shaded areas in the homeodomain indicate the predicted a-helices.

(Figure 2- 13). In order to determine the relationship between CHXIO-2 and Chxl71
duplicate mouse genomic blots were hybridized with full-length probes of either cDNA.
When using high stnngency conditions only Ch.171 hybridized to distinct bands in the
mouse genome, CHXIO-1 did not hybridize (Figure 2-14). Under slightly lowered
stringency (40% formamide) CHXIO-1 hybridized to bands which matched exactly the
band sizes detected by Chri71 (Figure 2- 14), suggesting that Chx271 is the locus most
closely related to CHXIO-1 in the mouse genome.
This finding does not necessarily imply that CHXIO-1 is the locus most closely
related to C h i 71 in humans. In order to determine this, duplicate human genomic blots
were hybridized with full-length probes of either cDNAs. At slightly lowered stringency
(40% formamide) CHXlO-1 detected only one single band (Figure 2- 1 S), whereas,
surprisingly Chxl71 detected repetitive sequences in human genomic DNA at these
conditions. Disregarding the elevated background oii the Chxl71 blot, it is clear that
several bands are detected, including those of CHXIO-1. Even when high stringency
washing conditions were used, bands that do not correspond to the CHXIO or the CHXIO-
2 locus remain visible (Figure 2-158). This finding indicates that human genomic DNA
contains at least one additional locus that is at least as sirnilar to Chxl71 as is CHXiO-1.
DISCUSSION
A multitude of screens for homeobox containing genes have been described in the
past several years. Whereas many attempts were based on the isolation of homologous
clones of Drosophila homeobox genes (e.g. Simeone et al., 1992), other screens for novel
sequences were performed using degenerate PCR approaches (e.g. Levine and Schechter,
1993). However most of the screens were performed on whole-embryo cDNA, which
potentially leads to two problems. First, highiy tissue specific homeobox genes might be
underrepresented and therefore rnissed. Second, homeobox genes that are involved oniy in
late differentiation events might not be represented in early embryonic cDNA libraries.
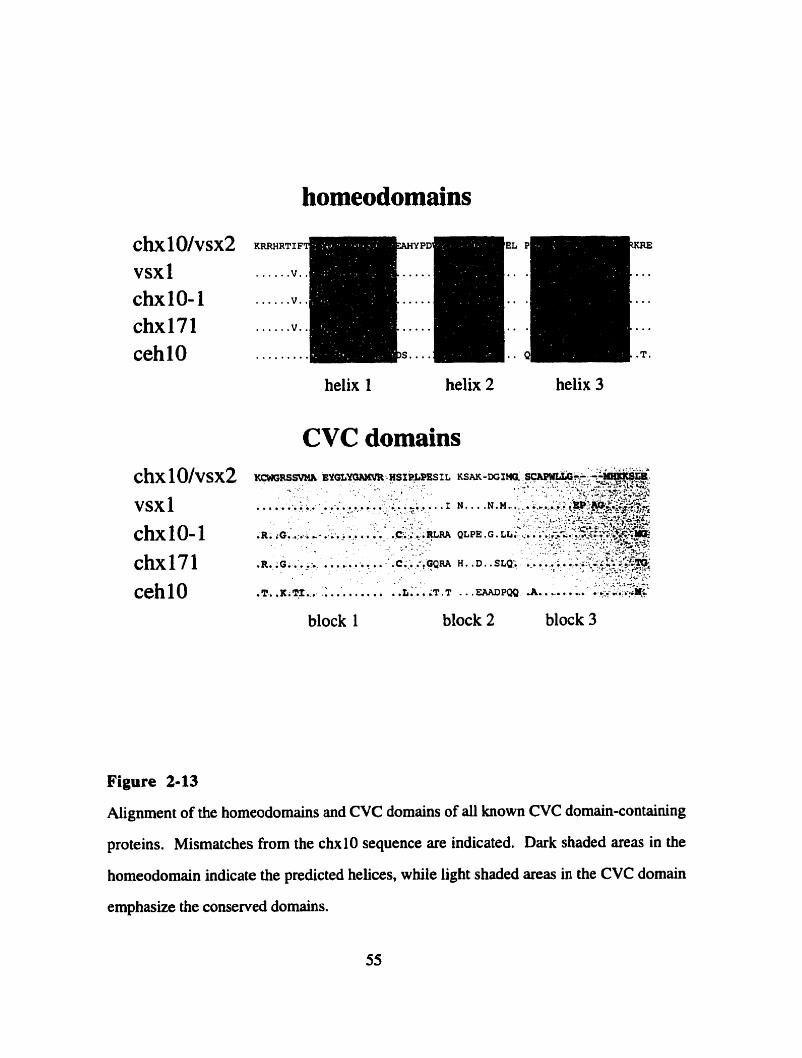
homeodomains
chx101vsx2 vsx 1 chxl0-l chxl71 cehl0
helix 1 helix 2 helix 3
CVC domains chx lOIvsx2 vsxl chxl0-1 chxl71 cehl0
KSAK
N...
QLPE
H. .D
block 1 block 2 block 3
Figure 2-13
Alignment of the homeodomains and CVC domains of ail known CVC domain-containhg
proteins. Mismatches from the chxlO sequence are indicated. Dark shaded areas in the
homeodomain indicate the predicted helices, while light shaded areas in the CVC domain
emphasize the consewed domains.
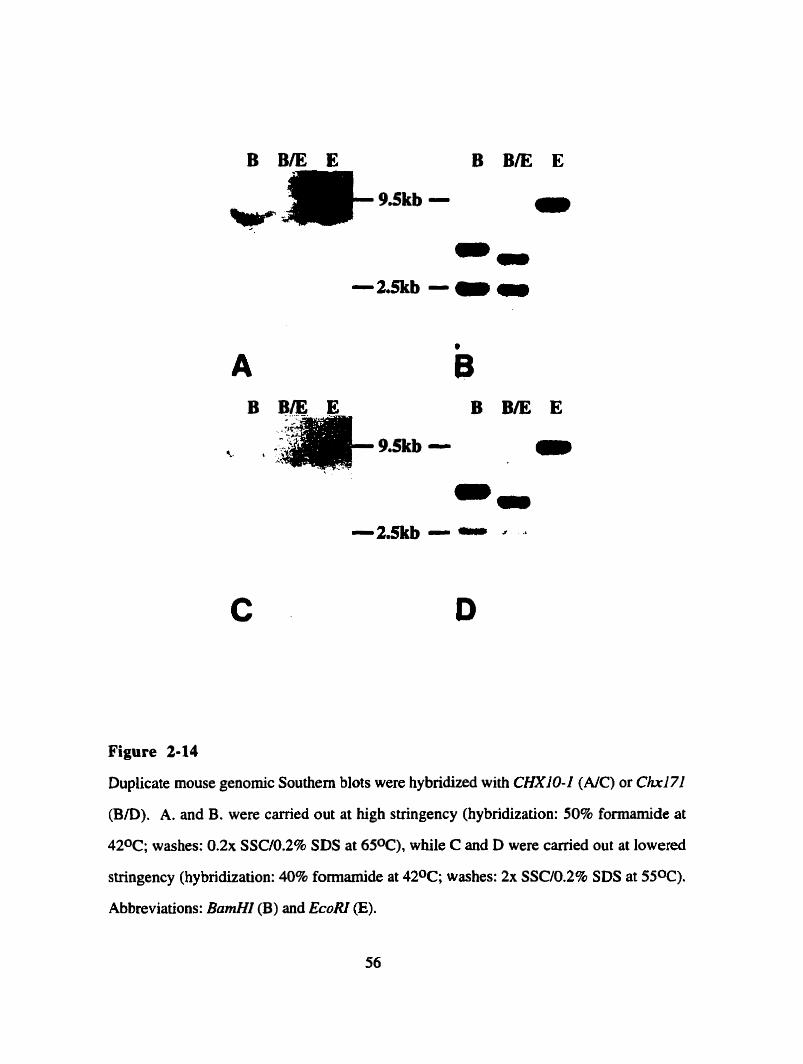
Figure 2-14
Duplicate mouse genomic Southem blots were hybridized with CHXîO-I (AIC) or Chi71
(B/D). A. and B. were carried out at high stringency (hybridization: 50% formamide at
420C; washes: 0 . 2 ~ SSC/O.P% SDS at 650C), while C and D were carried out at lowered
stringency (hybridization: 40% formamide at 420C; washes: 2x SSCIO.Z% SDS at 550C).
Abbreviations: BamHI (B) and EcoRI (E).
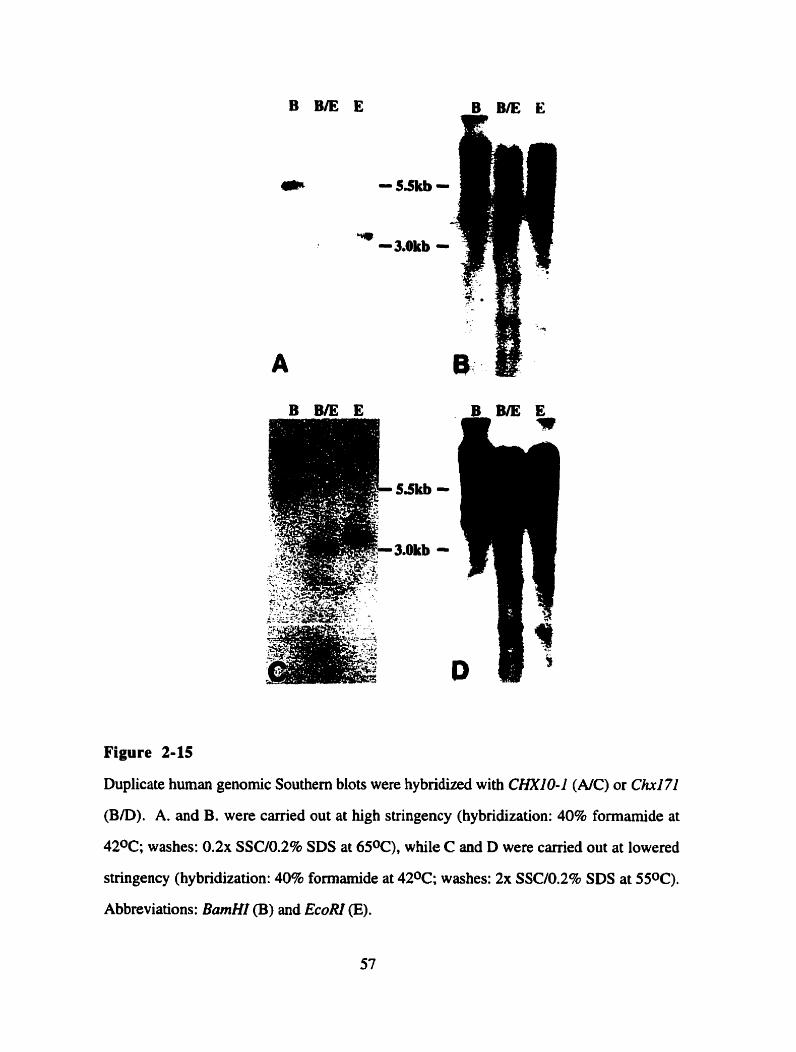
Figure 2-15
Duplicate human genomic Southern blots were hybridized with CHXIO-1 (AIC) or Ch171
(B/D). A. and B. were carried out at high stringency (hybridization: 40% formamide at
420C; washes: 0 . 2 ~ SSC/0.2% SDS at 650C), while C and D were carried out at lowered
stringency (hybridization: 40% formamide at 420C; washes: 2x SSC/O.2% SDS at 550C).
Abbreviations: BamHI (B) and EcoRI (E).

Exarnples of this type are the retinal homeobox genes, since the retina constitutes only a
minute part of the whole organism and some differentiation events do not occur until after
birth particularly in rodents (Sidman, 1961). Deitcher et al. ( 1994) tried to address the
problem of retina-specific homeobox genes by conducting a PCR-screen for homeobox
genes in the chick retina. However, their screen seemed to be technically problematic, in
that when using the PCR clone to screen a chick retinal cDNA library they were unable to
isolate the corresponding cDNA and only isolated a homologous cDNA. In addition, they
failed to identify chick CHXI O, which should have been amplified with the employed
primers and had not been published at the time (R.McInnes, pers. comm).
The data outlined in the introduction suggested that more retinal homeobox genes
are Iikely to exist. We were particularly interested in the isolation of close CHXlO
homologues, such as a human Vsxl orthologue, that would display almost complete amino
acid identity in the homeodomain. Using the library screening technique described, 1 have
been able to isolate two novel homeoboxîontaining cDNAs, namely OTX3 and CHXIO-1,
from an adult human retinal library. While the OïX3 is only distantly related, CHXIO-I
displays al1 the features of a close CHXlO homologue. However, due to the non-
embryonic stage of the libraiy, this screen also might not saturate the retina for homeobox
genes, a drawback accepted because of the demonstrated quality of the library, and because
CHXIO, although expressed in the adult, is also critical to mouse development (Burmeister
et al., unpublished).
The PCR approach that was used to distinguish clones of CHXlO from its
homologues, and thereby reduce the number of clones that had to be analyzed, does harbor
some potential problems. First, it will select not only CHXlO homologues, but also
CHXlO clones that lack the 3' UTR, or that have a rearrangement of the 3' UTR; such
clones have indeed k e n found in those selected. Second, if CHXI0 homologues share a
highly conserved regulatory element in the part of the 3' UTR that was amplified, the PCR
method rnight produce a sirnilar size band for actual homologues, which might thereby be

ignored in further analysis. However, based on sequence analysis we do not have any
indication of conservation in the 3' UTR of known C H X l O orthologues (man, chicken,
mouse). Even if such conserved sequences did exist, clones that have a 3' UTR
rearrangment should still be identifiable. In retrospect it might have been a good idea to
perform an additional PCR, with a primer set located in close proximity to the ORE Since
the screen was done with a probe covenng two thirds of the ORF, such a site would have
been less likely to be rearranged and therefore might have led to a more definitive
identification of CHXlO clones,
The identified human LIM homeobox gene LH2 is only 60% identical to the
CHXlO homeobox at the nucleotide level, showing clearly that the screening strategy was
sensitive enough to allow divergent sequences to be identified. The published expression
pattern of this gene includes high levels during lymphoid ce11 and CNS development, with
expression in the retina frorn optic cup io adult stages (Xu et al., 1993). Since this gene
has already been substantially characterized and does not represent a CHXIO homologue, it
was not chosen for further study.
The isolation of an OTX-like clone was unexpected, since substantial research
effort has been devoted to identifying the vertebrate homologues of orthodenticle, and
mammals were thought to oniy have two homologues (Simeone et al., 1992). Recently
another murine orthodenticle homologue, named Orthopedia. has been described by
Simeone et al. (1994). However, this clone differs substantially from the otx
homeodomains in many residues, including the ninth residue of the recognition helix, and
therefore can not be considered a true orthodenticle homologue. The partial cDNA that was
isolated in the screen described here contains a truncated homeobox with a high degree of
similarity to OtxlL2 at the nucleotide level, mostly differing at wobble-positions. The
nucleotides encoding the first 9 amino acids of the OTX3 cDNA differ strongly from the
other Otx homeoboxes, and are likely to be an artifact created by the technique used to
constnict the library. This method allowed snap-back structures to occur after first strand

cDNA synthesis, and therefore the 5' ends of some cDNAs might contain sequences that
do not correspond to the actual transcript (J. Nathans et al., pers. comm.). If these 9
amino acids are ignored, the remaining protein exhibits 88% amino acid identity to both o a
homeodomains. Although the rest of the protein is highly divergent, it does exhibit three
conserved blocks, when compared to otx 1 and 01x2 (Figure 2-3B); the significance of these
regions is unknown. The alignments clearly show that the OTX3 cDNA is more closely
related to mouse Otxl than to mouse 01x2. Al1 the alignments are based on the mouse
cDNAs of Otxl and 2, because, except for the 100% identical homeodomains, sequences
for the human OTX genes are unavailable.
CHXZO-1
The isolation of CHXIO-1, with both a homeobox and a CVC-box, confirmed Our
expectation that true CHXlO homologues might exist. The homeodomain sequence is
highly conserved with CHXIO, having 92% amino acid identity with only five differing
residues. Only one of these changes occurs within the helix-tum-helix motif, between
helix 2 and the mm, while the other four changes occur at the beginning and end of helix 1,
in regions that have been shown to be highly variable amongst homeobox gene farnily
members (Figure 2-13; Kappeii et al., 1993). Although no function has yet been assigned
to the CVC domain, its conservation in another homeodornain protein reiterates its probable
func tional significance . Interestingly, the predicted chxlO-1 peptide contains regions of high proline,
serinefthreonine and acidic amino acid content reminiscent of the putative activation
domains found in chx 10. Their arrangement within the peptide, however, is quite distinct
from chx 10 (Figure 2-5B). This different arrangement of protein domains suggests that
chxl0- l has acquired distinct functional properties from the chx 10 peptide.
Retinal expression of CHXIO-I in the adult is rnuch lower than that of CHXIO
(Figure 2-7). This finding is not necessarily surprising, since for most homeobox genes,

the highest expression is found during embryonic development, and this may be the case
for CHXIO-I as well. Al1 the murine Pax genes manifest down-regulation with
progressing developrnent, and two of the nine Pax genes are not expressed in the adult at
d l (Chalepakis et al., 1993).
Although the results of the in situ hybridization can only be regarded to be
preliminary, they allow for an interesting observation (Figure 2-8). CHXID-I was found
to be expressed in the inner nuclear and ganglion ce11 layers of adult human retina. Since
C m 0 was found to be specific to the inner nuclear layer in adult retina (Liu et al., 1994),
the expression pattern of CHWO- I and ChxlO are very similar. This is rather interesting,
in that many homologous homeobox genes such as OmlR have similar expression patterns
(Simeone et al., 1992), presumably reflecting a common origin. If confirmed in
subsequent expression studies, the result with CHXIO-I suggests that, like CHXIO,
CHXlO-I may also be involved in speciQing ce11 fate within the developing retina.
Human chromosome 20 does not contain any obvious disease loci in the region to
which CHXIO-I was mapped (Figure 2-6B). However, the homologous region of mouse
chromosome 2, contains two developmental eye diseases, Coloboma (Cm) and blind-sterile
(bs), as well as the locus of the ChrI O-homologous genomic clone 17 1 (B.Taylor, pers.
comm.).
Coloboma is a semidominant disease originally described in the offspring of an
irradiated ((C3HMeH x 1OlM) F1) mde. Although homozygotes have never been
described, the eyes of heterozygotes are small at birth and a ventral segment of their
choroid is absent. The eyeballs are rotated ventrally and affected animals have abnormal
posture, head-shaking, and circling, indicating additionai underlying neuronal defects
(Searle, 1966).
Blind-sterile mice arose spontaneously in an AKR/J strain. Homozygotes at day
El6 have bilateral nuclear cataracts that are present in the center of the lem, eyes that are
slightly smaller than normal, and glossy coats (Vamum, 1983). Adult males copulate

normally but have smaller than normal testis, with a reduced number of sperm, which, if
formed, completely lack acrosomes (Sotomayer and Handel, 1986).
C h x l 7 1
When a mouse retina library was screened at high stringency for the murine
CHXIO-I orthologue, only one cDNA was identified. This cDNA appears to represent the
transcnpt encoded by the 17 1 genornic locus. Although the CHNO-1 and Chri71 cDNAs
exhibit only 74% nucleotide identity over the whole ORF, both the homeodomain and the
CVC domain are well conserved, with 974 and 80% amino acid identity, respectively.
Over the entire length of the predicted proteins, however, they only exhibit 60% amino acid
identity (Figure 2- 1 OA).
This degree of divergence seemed higher than expected for orthologous
homeodomain proteins. In a study on the evolutionary conservation of Hox genes Kappen
et al. (1989) found an arithmetic mean of 10.7 nucleotide changes between human and
mouse orthologous homeoboxes. Even considering the standard deviation of +/- 4.5
nucleotides, this is much lower than the 24 nucleotide changes found between the CHXIO-
I and C h 1 71 homeoboxes. Since CHXIO-I and Ch271 do not belong to the Hox-class.
this finding might not necessarily represent the evolutionary pressure acting on these genes.
However, additional evidence indicates that these two proteins might be homologues rather
than orthologues. First, although chx 17 1 and chx 10- 1 contain similar activation domains,
their arrangements within the predicted proteins are quite distinct. This might indicate that
these proteins have acquired different funciional properties (Figure 2-IOB). Second,
aithough chxl7 1 is two amino acid residues shorter than chxl0- 1, its homeodomain and
CVC-domain complex are located hirther towards the C-terminus. Third, the variable
block 2 of the chx171 CVC domain displays only 25% identity to block 2 of chxl0-1, even
though blocks 1 and 3 are 95% conserved (Figure 2-13). In conclusion, Chxl7I and
C H X I O - 1 map to homologous chromosomal regions, but they differ in sequence,

transcript length, abundance and organization of predicted sequence motifs, to a degree
unexpected for orthologous homeobox genes of man and mouse.
In order io detennine whether C h 1 71 and CHXlO-1 represent homologues or
orthologues, their respective loci were examined in mouse and human genornic DNA.
Probing mouse genomic Southern blots with both the human CHXIO-1 and the mouse
Ch171 cDNAs showed that both appear to recognize the same genomic locus (Figure 2-
14). This result suggests that the screen of the mouse retinal cDNA library was successful,
in that the retinally expressed (at least in adult retina) sequence most sirnilar to CHXIO-1 in
the mouse genome, other than ChxlO, has indeed been identified. Probing human genomic
Southern blots with both the human CHXIO-1 and the rnouse Chxl7l cDNAs showed that
Chxl71 hybridizes to the human CHXIO-1 genomic locus and at least one other locus.
The hybridizing bands remain even after the hybndization stringency is increased (Figure
2-15). This result presents us with the possibility that the Ch171 and CHXIO-1 genes are
independent products of a duplication event of an ancestral gene, and that Chri 71 has a
human orthologue (CHXI 71).
The degree of sequence divergence between Ch171 and CHXIO-1 is consistent
with the occurrence of a duplication event. Dayhoff and Barker (1976) reponed a
cornparison of the extent of divergence between proteins that were derived from gene
duplications. They found that due to reduced selective pressure, substitution rates for one
of the genes are strongly elevated, presumably allowing the rapid acquisition of a different
function. This might explain why, considering the fairly recent divergence point between
man and mice of around 50 million years ago (Dickerson, 197 l), chxl0- 1 and chxl7 1 have
diverged 40% throughout the entire protein and 14% within the conserved homeodomain
and the CVC-domains.
Mer sequencing two thirds of the cDNAs from the human retinal cDNA screen that
1 conducted, no human CHXI71 cDNA has yet been cloned. Due to the demonstrated
retioa-specificity and abundance of Chxi71 in the rnouse it seems unlikely, if CHXI 71

exists, that it is not expressed in human retina. One possible explanation is that instead of a
change of the spatial expression pattern versus CHXI O- I , C M 71 might have acquired a
distinct developmental timing after the duplication event. A precedent for this type of event
is shown by Pax2 and P a 8 . Whereas both of these genes are expressed in the developing
neural tube and hindbrain in identical patterns, only Pax2 expression persists past E13.5
into the adult (Chalepakis, 1993).
With the isolation of the two clones (CHX10-1 and Chxl71) described in this
thesis, the group of CVC domain homeodomain genes has enlarged to one invertebrate and
four vertebrate members (Figure 2- 13). Although no function has yet been assigned to the
CVC domain, it could be analogous to class-specific domains found in paired- and POU-
homeobox genes which are involved in DNA binâing (Robertis, 1994), or the LM-domain
which is thought to be involved in protein-protein interactions (Sanchez-Garcia and
Rabbitts, 1994).
Alignments of the CVC domains of al1 known CVC domain-containing
homeodomain proteins show that the two lateral blocks (residues 1-27 and 40-59) are
highly conserved, while the middle region (residues 28-39) is highly variable (Figure 2-
13). Although block 2 does not display any conservation of particular amino acids, its size
is conserved, indicating that the spacing between blocks 1 and 3 seems to be crucial for
CVC domain îunction. Among al1 the vertebrate genes, block 2 has 11 residues length in
vertebrates, while that of the Celegans gene cehl0 is 10 residues in length (Figure 2-13).
Al1 the CVC-domain homeobox genes encode a homeodomain of the paired-like
type with a glutamine as residue nine of the recognition heüx (Bürglin, 1994). In a study
by Wilson et ai. (1993) these homeodomains were shown to be capable of cooperative
dimerization on their target sequences. In fact, paired homeodomains of the Q9 type
exhibited the highest degree of cooperativity, ranging up to 300-fold and were able to
heterodimerize with Paxlprd homeodomains of the S9-, and the K9-type. The observation
of dimerization for Pdprd-type homeodomain proteins, as well as the larger palindromic

DNA binding sequence may reflect the means by which the CVC domain-containing
proteins attain their functional specificity.
Al1 the CVC homeobox genes studied to date have been shown to be expressed in
neurons. More precisely, as far as data exist, al1 the vertebrate members are expressed in
the retina in a layer specific manner. Vsxl, CHXlO and CHXI 0-1 have been shown to be
expressed in the inner nuclear layer, while CHXIO-I is also expressed in the ganglion ce11
layer. This conservation of expression suggests that the vertebrate members of this
homeobox gene subclass are involved in the establishment or maintenance of specific ce11
types of the inner nuclear layer, or in the definition of the inner nuclear layer as a specific
domain within the retina. The Celegans member of this homeobox gene group, cehl0,
was found to be expressed in the AIY intemeuron, which receives synaptic input from the
thennosensitive AFD cell. The AFD ce11 has, due to its morphology and location, been
suggested to be potentiail y photosensitive (Svendson and McGhee, 1995). Although we
await further Chxl71 and CHXIO-I expression studies, members of the CVC homeobox
gene subclass may be involved in the determination and maintenance of sensory
interneurons.

CHAPTER 3: FUTURE DIRECTIONS
This thesis describes the cloning and partial characterization of two novel CVC
domain-containing homeobox genes. Further elucidation of the exact expression pattern of
both genes and comparison to the known patterns of expression of ChlO, Vsxl and
cehl0, should provide some insight into the function of this group of homeobox genes in
retinal development. In situ hybridization and immunohistochemistry cm be used to
determine the ce11 types in which these proteins are expressed.
C H X l O- I
The CHXlO-l gene has been shown to be highly divergent in sequence from
ChrlPI. In particular its abundance in the adult retina is very low compared to that of
Chxl71. More detailed expression studies of CHXIO-1 and Chxl71 will be necessary to
determine whether the genes have also diverged in hinction.
In order to determine the funciional equivalency of CHXIO-1 and Chx171, the
retinal layer and ce11 type in which both genes are expressed must be refined. The
localization of the CHXIO-1 transcripts reported as a preliminary result in this thesis will be
confirmed by additional digoxygenin in situ hybridization studies. Determination of the
expression pattem of mouse Ch171 might give an indication on whether CHXIO-I has
adopted a different function. Immunohistochemistry using antibodies that distinguish
chx 17 1 and c hx 10- 1, will provide additional data critical to assessing the relationship
between these two proteins.
In order to detexmine the role of CHXIO-I in transcriptional regulation its binding
sites (both to DNA and other proteins) must be identified and its activation properties
determined. Although the homeodomains of CVC proteins may have the same general
target DNA sequence as do the Pax/prd homeodomains, no activation of transcription was
observed with ch10 using the Pax reporter construct of Wilson et al. (1993) (G. Clarke,

unpublished results). Due to the presence of the CVC domain, binding properties of the
homeodomain might be altered, which is why target sequences must be selected from a
random oligonucleotide sequence library as outlined in Wilson et al. (1993). Using a
reporter construct with this target sequence trans-activating properties of CHXIO-1 could
be studied in Chloramphenicol acetyltransferase (CAT) assays as it has been done for other
homeodomain proteins (Wilson et al., 1993). Comparison of these data with those of
Chxl71 should indicate whether CHXIO-1 has retained the in vitro activation properties of
Chxl7l.
Narrowing down the tirne point of the duplication event might allow a correlation of
the presence of CHXIO-1 and morphological changes in the retina. To do this genomic
zoo-blots should be performed and include species which are phylogenetically closer to the
human than the mouse, such as new and old world monkeys.
Chxl71
In contrast to CHXlO-1, mouse Chxl71 is expressed at high levels, specifically in
the adult retina. To determine the importance of C M 7 1 for mouse retinal development, a
more detailed analysis of the Ch171 expression pattem will be necessary. If these
expression studies suggest a possible involvement of Chxl71 in the development of the
eye, proof of the requirement of Ch1 71 could be obtained by targeting the C h 1 7 1 locus.
Altematively the possibility that Chx171 is mutated in the known eye diseases blind-sterile
and Coloboma could be investigated.
In situ hybridization studies will be needed to define the spatial and temporal
expression pattem of C M 7 1 in mouse development. The focal points of this study should
be the time and place of the onset of expression, the expression pattem in neuronal tissues
and in the developing eye, and the cells in which expression is maintained in the adult
retina. This study will be complemented by irnmunohistochemistry for chxl71, which
should identiQ the specific cells that express chxl7l. Overlaps in the spatial expression

patterns of Chxi71 and ChxlO, rnight suggest both an interplay of the two proteins in
development, and their formation of heterodimen in cells in which they are both expressed.
To permit mutational analysis, or altematively to create a knock-out mouse, the
genomic locus of Ch171 must be isolated. This work has already k e n started and plaque
pure phage are currently king analyzed (C. Freund, unpublished results).
To determine whether mutations in Ch171 cause Coloboma or blind sterile, the
Ch171 location in the genetic map must be further refined. For this purpose the full-
length C h 1 71 cDNA will be used for linkage analysis. If this linkage analysis does not
produce any recombinants with either blind-sterile or Coloboma, it will be necessary to
investigate directly the possible involvement of Chiri 71 in these two diseases. Initially this
problem could be addressed by examining the retinal expression of C M 71 in mutant rnice
by Northern blotting. To characterize the inherent molecular mechanism, one should
initially examine the CM71 locus in genomic DNA from mutant animals, by Southem blot
analysis. If no gross rearrangements are found, small aiterations in the sequence (such as
substitutions) cm be sought by SSCP ancilor direct sequencing, in order to determine the
specific molecular defec t.
Altematively, if C h 1 71 does not seem to be implicated in known genetic diseases,
its requirement in retinal development will have to be detedned by gene targeting. This
project could be done using existing strategies such as those currently employed in our lab
for the targeting of the Rom1 locus (G. Clarke, unpublished results).
CVC domain function and significance
The significance of the CVC domain, a highly conserved sequence element found in
homeobox genes from nematode to man, is unknown but its conservation suggests that it is
likely to be important. The major question will be whether the CVC-domain, like the
homeodomain, binds DNA or altematively functions in the interaction with other proteins.

Research will have to focus on the determination of interacting partners. structural motifs
and its role in transcriptional regulation.
To identify the interacting partners of the CVC domain it will be necessary to
establish whether it interacts with DNA or protein. Interaction with DNA could be
demonstrated by gel-shift assays (using the target site selected for the homeodomain and
CVC domain complex by the Wilson method (1993)), while an interaction with proteins
could be shown by affinity-chromatography. The interacting proteins, possibly other CVC
homedomain proteins, could then be identified using the yeast two-hybrid system.
Since the CVC domain represents a novel domain it is important to determine its
tertiary structure. The relatively small size of the CVC domain should allow both NMR
spectroscopy and X-ray crystallography to be done. If the CVC domain does interact with
DNA, structural analysis should include the complex of the CVC domain bound to DNA.
Structural data might indicate residues critical for hinction and may also explain if the low
degree of conservation of block two arises from this block acting mainly as a spacer
sequence.
Alternatively the CVC domain might represent an activation domain. This
possibility could be determined by observing whether deletion of the CVC domain has a
deleterious or enhancing effect on tram-activation in a CAT-assay.
Human CHXlO homologues
The screening of the human retina cDNA library for CHXlO homologues must be
completed. Although additional novel clones may be identified, one has to consider the
possibility that no human CHXl71 or human VSXl is present in the human genome.
However, since circumstantial evidence suggests that these clones do exist, other strategies
will have to be used to clone these genes. While problems with the tissue specificity might
be circumvented most easily by screening a genomic Library, distinctive feanires such as the
Vsxl alternative exon might allow PCR strategies to be used successfully.

To date 35 clones of the high stringency class (Classes 2 and 3) remain to be
characterized. Additionally there are 48 clones of the low stringency class (Class I) , that
have not been analyzed. Since many rearranged clones have been identified, the initial
anaiysis should include additional PCR with primers specific for the 3' untranslated regions
of both CHXlO and CHXIO- I in order to exclude these known cDNAs. Analysis of these
clones might still lead to the identification of additionai CHXlO homologues.
This screen has not yet produced a human CHX171, either due to problems with
the screening procedure or tissue specificity. It is therefore probably best to screen for
such a clone in a human genornic library, which could be done in conjunction with the
isolation of the respective CHXlO-I locus.
Alignments of Vsxl with C H X i O - 1 and Ch171 clearly suggest that the
mammalian onhologue of Vsxl also remains to be identified. The most distinct feature of
Vsxl is a 12 nucleotide insertion in the CVC domain, which is why 1 have decided to
attempt a PCR based approach. Primers to the 5' end of the horneobox and overlapping
with the alternative exon of the Vsxl CVC box have been constructed, based on the unique
Vsxl sequence and the preferred human codon usage. Used on the uncharacterized clones
of the human retinal screen or alternatively, reverse-transcribed retinal RNA, these primers
should lead to the isolation of V W , if such a gene indeed exists.

LIST OF CITATIONS
Adler, R., and Farber D. (1986). The retina: A mode1 for ce11 biology studies. in Cellular Neurobiology Part 1 (Academic Press Inc.), 5.
Alderman, A.L. (1935). The determination of the eye in the anuran, Hyla regilla. J. Exp. 2001. 70, 205-232.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990). Basic local alignment tool. J. Mol. Biol. 215, 403-410.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith. J.A.,
and Struhl, K. (1989). Current Protocols in Molecular Biology (John Wiley & Sons).
Bascom, R.A., Manara, S., Collins, L., Molday, R.S., Kalnins, V.I. and McInnes, R.R. (1992). Cloning of the cDNA for a novel photoreceptor membrane protein (rom-1)
identifies a disk rim protein farnily implicated in human retinopathies. Neuron 8, 1172-84.
Bateson, W. (1894). Materials for the study of variation treated with especial regard to
discontinuity in the origin of species (McMillan and Co., London).
Bender, W., Spierer, P., and Hogness, D.S. (1983). Chromosomal walking and jumping to isolate DNA from the Ace and rosy loci and the Bithorax complex in Drosophila melanogaster. J. Mol.Bio1. 168, 17-33.
Boncinelli, E., Simeone, A., and Acampora, D. (199 1). HOX gene activation by retinoic
acid. Trends Genet. 7,329-334.
Boncinelli, E., Somma, R., Acampora, M., D'Esposito. M., Faiella, A., and Simeone, A.
(1988). Organization of human homeobox genes. Human Reproduction 3,880-886 .
Bopp, D., Bum, M., Baumgartner, S., Frigeno, G., and Noll, M. (1986). Conservation of a large protein domain in the segmentation gene paired and in functionally related genes of Drosophila. Cell.47, 1033- 1040.

Bridges, C.B., and Morgan, T.H. (1923). The thirdthromosome group mutant character
of Drosophila melanogaster. (Washington: Carnegie Institution of Washington), 137- 139.
Bürglin, T.R. (1994). A comprehensive classification of homeobox genes. In Guidebook to the Homeobox Genes. Denis Duboule ed. (Oxford University Press), 25-72.
Burmeister, M., Novak, J., Basu, S., Ploder, L., Liang. M.-Y., Hawes, N.L., Vidgen, D.. Hoover, F., Goldman, D., Kalnins. V.I., Roderick, T.H., Taylor, B.A., Hankin, M.H., and McInnes R.R. (1995). A nul1 allele in the ChxlO homeobox gene of the ocular retardution mouse. Unpublished.
Campbell, R.B., Sinclair, D.A.R., Couling, M., and Brock, H.W. (1995). Genetic interactions and dosage effects of Pdycomb group genes of Drosophika. Mol. Gen. Genet. 246, 29 1-300.
Cepko, C.L. (1992). Retinal ce11 fate determination. Progress in Retinal Research 12, 1 -
Chalepakis, G., Stoykova, A., Wijnholds, Je, Tremblay, P., and Gruss, P. (1993). Pax;
Gene regulators in the developing nervous system. J. Neurobiol. 24, 136% 1384.
Chalepakis, G., Fritsch, R., Fickenscher, H.9 Deutsch, U., Goulding, M., and Gruss, P.
(1991). The molecular basis of the undulated/Pax-l mutation. Ce11 66,873-884.
Dayhoff, M.O., and Barker, W.C. ( 1976). Mechanisms in molecular evolution: Examples. In Atlas of Protein Sequence and Structure 5. M.O. Dayhoff, ed. (Silver Springs, Md.:
National Biomedical Research foundation), 30 1-3 10.
Deitcher, D.L., Fekete, D.M.9 and Cepko, C.L. (1994). Asymmetnc expression of a novel homeobox gene in vertebrate sensory organs. J. Neurosci. 14,486-498.
Desplan, C., Theis, J., and O'Farrell, P.H. (1985). The Drosophila developmental gene, engrailed, encodes a sequence-specific DNA binding activity. Nature 318,630-635.
Dickerson, R.E. (1971). The structure of cytochrome c and the rates of molecular evolution, J. Mol. Evo. 1, 26-45,

Dingwail, C., and Laskey, R.E. (1991). Nuclear targeting sequences - a consensus? Trends Bioch. Sci. 16,478-48 1.
Driever, W ., and Nüsslein-Vollhard, C. ( 1989). The bicoid protein is a positive regulator of hunchback transcription in the early Drosophila embryo. Nature 33 7, 1 38- 143.
Duboule, D., and DollC, P. (1989). The structural and functional organization of the murine Hox gene family resembles that of Drosophila homeotic genes. EMBO J. 9, 1497-
1505.
Feinberg, A.P., and Vogelstein, B.A. (1983). A technique for radiolabelling DNA
restriction endonuclease fragments to high specific activity . Anal. Biochem. 132,6- 13.
Finkelstein, R., and Boncinelli, E. (1994). Frorn fly to mammalian forebrain: the story of otd and Otx, Trends Genet. 10, 3 10-3 15.
Galili, N., Davis, R.J., Fredericks, W.J., Mukhodpadhay, S., Rauscher III, F.J., Emanuel, B.S., Rovera, G., and Barr, F.G. (1993). Fusion of a fork head domain gene to PAX3 in the solid tumour alveolar rhabdomyosarcoma. Nature 5,230-235.
Garber, R.L., Kuriowa, A., and Gehring, W.J. (1 983). Genomic and cDNA clones of the homeotic locus Antennapedia in Drosophila. EMBO J. 2,2027-2036.
Gehring, W.J. (1994). A history of the homeobox. In Guidebook to the homeobox genes.
Denis Duboule ed.(Oxford University Press), 1- 10.
Gehring, W.J. (1992). The homeobox in perspective. Trends Bioch. Sci. 17,277-280.
Gehring, W.J. (1967). Formation of a complete mid leg with stemopleura in the antennae
region by the mutant Nasobemia (Ns) of Drosophila melanogaster. Arch Julius Klaus Stift Vererbungsforsch. Sozialanthropol. Rassenhyg. 41,44-54.
Graham, A., Papalopulu, N., and h m l a u f , R. (1989). The murine and Drosophila Homeobox gene complexes have common features of organization and expression. Ce11 57, 367-378.

Grossberger, D. (1987). Minipreps of DNA from bactenophage Lambda, Nucl. Acids
Res. 15, 6737.
Gruss, P., and Walther, C. (1992). Pax in development. Cell69, 7 19-722.
Gyurkovics. H., Gausz, J., Kummer, J., and Karch, F. (1990). A new homeotic mutation in the Drosophila bithorax complex removes a boundary separating two domains of
regulation. EMBO J. 9,2579-2585.
Halder, G., Callaerts, P., and Gehring, W .J. (1995). Induction of ectopic eyes by targeted
expression of the eyeless gene in Drosophila. Science 267, 1788-92.
Haw kin. N .C., and McGhee, J. D. ( 1990). Homeobox-containing gnes in the nematode
caenorhabditis elegans. Nucl. Acids Res. 18,6 10 1-6 106.
Hinds, J.W., and Hinds, P.L. (1974). Early ganglion ce11 differentiation in the mouse
retina: An electron microscopic analysis utilizing send sections. Devel. Biol. 37,38 1-4 16.
Hinds, J.W., and Hinds, P.L. (1978). Early development of arnacrine cells in the mouse
retina: An electron rnicroscopic, serial section analysis. J. Comp. Neurol. 179,277-300.
Hinds, J.W., and Hinds, P.L. (1979). Differentiation of photorcepton and horizontal cells
in the embryonic mouse retina: An electron microscopic analysis utilizing senal section. I. Comp. Neurol. 187, 495-5 12.
Hogan, B.L.M., Hirst, E.M.A., Horsburgh, G., and Hetherington, C.M. (1988). Small eye (Sey): a mouse mode1 for the genetic analysis of craniofacial abnormalities.
Development 103, 1 15- 1 19.
Jaynes, J.B., and O'Farrell, P.H. (1988). Activation and repression of transcription by
homoeodomain-containing proteins that bind a c o m o n site. Nature 336,744-9.
Jürgens, G. (1985). A group of genes controlling the spatial expression of the bithorax
complex in Drosophila. Nature 316, 153- 155.

Kappen, C., Schughart, K., and Ruddle, F.H. (1993). Early evolutionary origin of major homeodomain sequence classes. Genomics 18,54-70.
Kappen, C., Schughart, K., and Ruddle, F.H. (1989). Two steps in the evolution of Antennapedia-class vertebrate homeobox genes., Proc. Natl. Acad. Sci. 56,5459-63.
Kastury, K., Druck, T., Huebner, K., Barletta, C., Acampora, D., Simeone, A., Faiella, A., and Boncinelli, E. (1994). Chromosome locations of EMX and OTX genes. Genomics 22, 41-45.
Kaufman, M.H. ( 1992). The Atlas of Mouse Development. (Academic Press, Toronto).
Kissinger, C.R., Liu, B.. Martin-Bianco, E., Komberg, T.B., and Pabo, C.O. (1990). Crystal Structure of an engrailed homeodomain-DNA complex at 2.8A resolution: A framework for understanding homeodornain-DNA interactions. Ce11 63,579-590.
Kuziora, M.A., and McGinnis, W. ( 1989). A homeodomain substitution changes the regulatory specificity of the deformed protein in Drosophila embryos. Ce11 59,56363- 1.
Levine, E.M., Hitchcock, P.F., Glasgow, E., and Schechter, N. ( 19%). Restricted expression of a new paired-class homeobox gene in normal and regenerating adult goldfish
retina. J. Comp. Neurol. 348, 596-606.
Levine, E.M., and Schechter, N. (1993). Homeobox genes are expressed in the retina and brain of adult goldfish, Proc. Natl. Acad. Sci. 90, 2729-2733.
Lewin, B. ( 1990). Genes N. (Ce11 Press, Cambridge, Massachusetts), 178- 179.
Lewis, E.B. (1978). A gene complex controllhg segmentation in Drosophila. Nature 276, 565-570.
Liu, I.S.C., Chen, J., Ploder, L., Vidgen, D., van der Kooy, D., Kalnins V.I., and McInnes R.R. (1994). Developmental expression of a novel rnurine homeobox gene ChxlO: evidence for roles in determination of the neuroretioa and inner nuclear layer. Neuron 13, 377-393.

Mann, R., and Hogness, D.S. (1990). Functional Dissection of Ultrabithorax proteins in
Drosophila melanogaster. Ce11 60,597-6 10.
McGinnis, W., Garber, R.L. Wirz, J., Kuriowa, A., and Gehring, W.J. (1984). A homologous proteintoding sequence in Drosophila homeotic genes and its conservation in other metazoans. Ce11 37,403-408.
Mermod, N., O'Neill, E.A., Kelly, T.J., and Tijan, R. (1989). The proline-rich transcriptional activator of CTF/NF-1 is distinct from the replication and DNA binding domain. Ce11 58, 74 1 -753.
Mitchell, P.J., and Tijan, R. (1989). Transcriptional regulation in rnammalian cells by
sequence-specific DNA binding proteins. Science 245,37 1-378.
Murtha, M.T., Leckman, J.F., and Ruddle, F.H. (1991). Detection of homeobox genes in development and evolution. Proc. Natl. Acad. Sci. 88, 1071 1-107 15.
Nei, M. (1987). Genomic Evolution. In Molecular Evolutionary Genetics. (Columbia
University Press, New York), 1 1 1- 148.
Noll, M. (1993). Evolution and Role of Pax genes, Cun: Opin. Genet. Dev. 3, 595-605.
Otting, G., Qian, Y.Q., Billeter, M., Mueller, M., Affolter, M., Gehring, W.J., and Wuethrich, K. ( 1990). Protein-DNA contacts in the structure of a homeodomain-DNA complex detemiined by nuclear magnetic resonance spectroscopy in solution. EMBO J. 9,
3085-3092.
Pabo, CO. , and Sauer, R.T. ( 1984). Protein-DNA recognition. Ann. Rev. Biochem. 53,
293-32 1.
Peifer, M., Karch, F., and Bender, W. (1987). The bithorax complex: control of
segmental identity . Genes Dev. 1,89 1-898.
Qais, F., Jackson, A.U., Xu, J., Gryzenia, M., Skolnick, C., Agarwal, N., and Swaroop, A. (1993). Molecular characterization of the murine neural retina leucine zipper gene Nrl. Genomics 18,2 16-222.

Qian, Y.G., Billeter, M., Otting, G., Müller, M., Gehring, W.J., and Wüthrich, K. (1989). The structure of the Aniennapedin homedomain determined by NMR spectroscopy in solution: cornparison with prokaryotic repressors. Cell59,573-580.
Quiring, R., Walldorf, U., Kloter, U., and Gehring, W.J. (1994). Homology of the eyeless gene of Drosophila to the Small eye gene in mice and Aniridia in Humans. Science
265, 785-789.
Read, A.P. (1995). Pax genes-paired feet in t h e camps. Nature Genet. 9, 333-334.
Reh, T. (1992). Generation of neuronal diversity in the vertebrate retina, 1992, Determinants of Neuronal Identity. Shankland, S.M., and Macagno, E.R., eds. (New
York, Academic Press), 433-467.
Robertis de, E.M. (1994). The homeobox in ce11 differentiation and evolution. In Guidebook to the homeobox genes. Duboule, D., ed., (Oxford University Press), 1 1-24.
Robinson, S.R. (1991). Development of the rnammalian retina. In Neuroanatomy of the Visual Pathways and their Development. Dreher, B., and Robinson, S.R., eds. In Vision and Visual Dysfunction 3.3. Cronly-Dillon ed. (Macmillan Press Ltd.), 69- 128.
Robb, R.M. , Silver, J., and Sullivan, R.T. (1978). Ocular retardartion (or) in the mouse. Invest. Opthal. Vis. Sci. 17 468-473.
Rowe, M.H. (199 1). Functional organization of the retina. In Neuroanatomy of the Visual Pathways and their Development. Dreher, B., and Robinson, S.R., eds. In Vision and
Visual Dysfunction 3. J. Cronly-Dillon ed. (Macmillan Press Ltd.), 1-69.
Saiki, R.K., Gelfand, D.H., Stoffel, S., Sharf, S.J., Higuchi, R., Hom, G.T., and Erlich, H.A. (1988). Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239,487-49 1.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual. (Cold Spring Harbor).

Sanchez-Garcia, I., and Rabbitts, T.H. (1994). The LIM domain: a new structural motif found in zinc-finger-like proteins. Trends Genet. 10,3 15-320.
Scott, M.P ., Tamkun, J.W ., and Hartzell, G.W., In (1989). The structure and function of
the homeodomain. Biochim. Biophys. Acta 989,2548.
Searle, A.G. (1966). Mouse News Lett. 35,27.
Seimiya, M., Ishiguro, H., Miura, K., Watanabe, Y., and Kuosoawa, Y. (1994).
Homeobox-containing genes in the most primitive metazoa, the sponges, Euro. J. Bioch.
221, 2 19-225.
Shepherd, J.C.W., McGinnis, W., Carrasco, A.E., Robertis de, E.M., and Gehring,
W.J. (1984). Fly and frog homoeo domains show homologies with yeast mating type
regulatory proteins. Nature JI O, 70-7 1.
Sidman, R.L. (1961). Histogenesis of mouse retina studied with HI thymidine. In The
Structure of the Eye. Smelser, G., ed. (New York Academic Press), 487-506.
Silver, J., and Hughes, A.F.W. (1973). The role of ce11 death during morphogenesis of
the mammalian eye. J.Morpho1. 140, 1 59- 170.
Simeone, A., D'Apice, M.R., Nigro, V., Casanova, J., Graziani, F., Acampora, D., and
Avantaggiato, V. (1994). Orthopedia, a novel homeoboxtontaining gene expressed in the
developing CNS of both mouse and Drosophila. Neuron 13,83-101.
Simeone, A., Acampora, D., Gulisano, M., Stornaiuolo, A., and Bonnicelli, E. (1992).
Nested expression domains of four homeobox genes in developing rostral brain. Nature
358, 687-690.
Sotomayer, R.E., and Handel, M.A. (1986). Failure of acrosome assembly in a male
sterile mouse mutant. Biol. Reprod. 34, 17 1- 182.
Spemann, H. (1938). Embryonic development and induction. (New Haven, Conneticut:
Yale University Press).

Strachan, T., and Read, A.P. (1994). Pax genes. Curr. Opin. Genet. Dev. 4,427-438.
Svendsen, P.C., and McGhee, J.D. (1995). The C. elegans neuronally-expressed homeobox gene ceh-10 is closely related to genes expressed in the vertebrate eye.
Development 121, 1253- 1262.
Thaii, M., Müller, M.M., DeLorenzi, M., Matthias, P., and Bienz, M. (1988). Drosophila
homoeotic genes encode transcriptional activators sirnilar to marnmalian OTF-2. Nature
336, 598-60 1.
Theill, L.E., Castrillo, LL., Wu, D., and Karin, M. (1989). Dissection of functional
domains of the pituitary-specific transcription factor GHF-2. Nature 342,945-948.
Treisman, J., Harris, E., and Desplan, C. (1991). The paired box encodes a second DNA- binding domain in the paired homeodomain protein. Genes. Dev. 5,594-604.
Treisman, J., Gonczy, P., Vashishtha, M., Harris, E., and Desplan, C. (1989). A single amino acid cm determine the DNA binding specificity of homeodomain protein. Ce11 59,
553-562.
Truslove, G.M. (1962). A gene causing ocular retardation in the mouse, J. Embryol. Exp. Morphol. 10, 652.
Turner, D.L., and Cepko, C.L. (1988). Histogenesis of mouse retina studied with [%]-
thymidine. In The structure of the Eye. Smelser, G., ed., (New York Acadernic Press),
487-506.
Varnum, D.S. (1983). Blind-sterile: a new mutation on chromosome 2 of the house
mouse. J. Hered. 74, 206-207.
Wetts, R., and Fraser, S.E. (1988). Multipotent precursors can give rise to al1 major ce11 types of the frog retina. Science 239, 1 1 142- 1 145.
Williams, T., and Tijan, R. (1991). Analysis of the DNA binding and activation properties
of the human transcription factor AP-2. Genes Dev. 5,670-682.

Wilson, D., Sheng, G., Lecuit, T., Dostatni, N., and Desplan, C. (1993). Cooperative dimerization of paired class homeodomains on DNA, Genes Dev. 7,2 120-2 134.
Wolberger, C., Vershon, A.K., Liu, B., Johnson, A.D., and Pabo, C.O. (1991). Crystal structure of a Matda2 homeodomain-operator cornplex suggests a general mode1 for
homeodomain-DNA interactions. Ce11 67,s 17-28.
Xu, Y., Baldassare, M., Fisher, P., Rathbun, G., Oltz, E.M., Yancopoulos, G.D., Jessell, T.M., and Alt, F.W. (1993). LH2: A LIM/homoedomain gene expressed in developing lymphocytes and neural cells, Proc. Natl. Acad. Sci. 90,227-23 1.
Young, R.W. (1985). Ce11 differentiation in the Retina of the Mouse. The Anatomical
Record 21 2, 199-205.




















