
Functional annotation of putative hypothetical proteins from Candida dubliniensis
Upload
independentCategory
view
1download
0
JOURNAL OF CLINICAL MICROBIOLOGY, Feb. 2002, p. 556–574 Vol. 40, No. 20095-1137/02/$04.00�0 DOI: 10.1128/JCM.40.2.556–574.2002Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Identification of Four Distinct Genotypes of Candida dubliniensis andDetection of Microevolution In Vitro and In Vivo
Sarah F. Gee,1 Sophie Joly,2 David R. Soll,2 Jacques F. G. M. Meis,3 Paul E. Verweij,3Itzhack Polacheck,4 Derek J. Sullivan,1 and David C. Coleman1*
Microbiology Research Unit, Department of Oral Medicine and Oral Pathology, School of Dental Science and Dublin DentalHospital, Trinity College, University of Dublin, Dublin 2, Republic of Ireland1; Department of Biological Sciences, University ofIowa, Iowa City, Iowa 522422; Department of Medical Microbiology and Infectious Diseases, Canisius Wilhelmina Hospital and
University Medical Center, 6532 SZ Nijmegen, The Netherlands3; and Department of Clinical Microbiology and Infectious Diseases,The Hebrew University-Hadassah Medical Center, Jerusalem, Israel4
Received 31 August 2001/Returned for modification 18 November 2001/Accepted 25 November 2001
The present study investigates further the population structure of Candida dubliniensis and its ability toexhibit microevolution. Using 98 isolates (including 80 oral isolates) from 94 patients in 15 countries, weconfirmed the existence of two distinct populations within the species C. dubliniensis, designated Cd25 groupI and Cd25 group II, respectively, on the basis of DNA fingerprints generated with the C. dubliniensis-specificprobe Cd25. The majority of Cd25 group I isolates (48 of 71, 67.6%) were from human immunodeficiency virus(HIV)-infected individuals, whereas the majority of Cd25 group II isolates (19 of 27, 70.4%) were fromHIV-negative individuals (P < 0.001). Nucleotide sequence analysis of the internal transcribed spacer (ITS)regions of the rRNA genes from 19 representative isolates revealed the presence of four separate genotypes. Allof the Cd25 group I isolates tested belonged to genotype 1, while the Cd25 group II population was comprisedof three distinct genotypes (genotypes 2 to 4), which corresponded to distinct clades within the Cd25 group IIpopulation. These findings were confirmed using genotype-specific PCR primers with 70 isolates. We alsoshowed that C. dubliniensis can exhibit microevolution in vivo and in vitro as occurs in other yeast species. DNAfingerprinting using the C. dubliniensis probes Cd25, Cd24, and Cd1 and karyotype analysis of multiple oralisolates recovered from the same specimen from each of eight separate patients revealed microevolution in sixof eight of the clonal populations. Similarly, sequential clonal isolates from various anatomical sites in twoseparate patients exhibited microevolution. Microevolution was also shown to occur when two clinical isolatessusceptible to fluconazole were exposed to the drug in vitro. The epidemiological significance of the four C.dubliniensis genotypes and the ability of C. dubliniensis to undergo microevolution has yet to be established.
Candida dubliniensis was first identified as a novel Candidaspecies in 1995. It is closely related to Candida albicans, withwhich it shares many phenotypic characteristics, including theability to produce germ tubes and chlamydospores and resis-tance to cycloheximide (57). As a result of these similarities, inthe past many isolates of C. dubliniensis have been misidenti-fied as C. albicans, hampering comprehensive epidemiologicalanalysis of this species (56). In recent years a number of testshave been developed which have permitted the rapid and re-liable identification of C. dubliniensis in clinical samples.
Phenotype-based tests such as the analysis of carbohydrateassimilation profiles using commercial yeast identification sys-tems (38), growth temperature (39), differentially colored pri-mary colonies on CHROMagar Candida medium (8, 34),chlamydospore production and colony morphology on bird-seed agar (1, 53), and molecular tests based on PCR technol-ogy (10, 11, 20, 35, 58) have all been shown to be useful in theidentification of C. dubliniensis. Using these techniques, C.dubliniensis isolates have been identified throughout the worldin a wide variety of patient cohorts (1, 16, 17, 19, 28, 29, 40, 42,
54). Originally associated with oral carriage and oral candidi-asis in human immunodeficiency virus (HIV)-infected andAIDS patients (3, 29, 30, 57), C. dubliniensis has more recentlybeen identified in cases of systemic disease in Europe, theUnited States, and Australia (4, 28, 31, 46).
In an attempt to improve our understanding of the epide-miology of C. dubliniensis, in 1999 Joly et al. (17) developedand characterized three complex DNA fingerprinting probesfor C. dubliniensis, one of which (Cd25) was species specific.The remaining two probes (Cd1 and Cd24) reacted weaklywith C. albicans DNA. As well as providing a useful means ofinvestigating the epidemiology of C. dubliniensis, the existenceof the species-specific sequences contained within Cd25 pro-vided further evidence that C. dubliniensis is a distinct species.
The Cd1 and Cd24 probes were shown to be useful fordetecting in vitro and in vivo microevolutionary events,whereas Cd25 yielded stable complex fingerprint patterns suit-able for the comparison of strains in epidemiological studies.This probe was used to fingerprint 57 independent C. dublini-ensis isolates from 11 countries (17). Computer-assisted anal-ysis of the fingerprints obtained using the software packageDENDRON (17, 52) demonstrated that the C. dubliniensisisolates could be clearly divided into two separate groups,groups I and II. Group I isolates comprised 86% of thosetested and were all closely related (average similarity coeffi-cient [SAB] � 0.80), whereas the group II isolates, which com-
* Corresponding author. Mailing address: Microbiology ResearchUnit, Department of Oral Medicine and Oral Pathology, School ofDental Science, Trinity College, University of Dublin, Dublin 2, Re-public of Ireland. Phone: 353 1 6127276. Fax: 353 1 6127295. E-mail:[email protected].
556
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

prised the remaining 14%, constituted a less closely relatedclade (average SAB � 0.47).
The purpose of the present study was to investigate furtherthe genetic diversity of C. dubliniensis by analysis of Cd25-generated fingerprint profiles using a larger number of isolatesfrom around the world taken from a broader range of anatomicsites. Secondly, this study sought to determine if the two groupsidentified on the basis of Cd25-generated fingerprint profileanalysis represent specific genotypes which can be differenti-ated based on nucleotide sequence analysis of the internaltranscribed spacer regions (ITS) of the rRNA gene cluster.The third objective of this study was to further investigate thephenomenon of microevolution in C. dubliniensis, both in vivoand in vitro, using the three fingerprinting probes described byJoly et al. (17).
MATERIALS AND METHODS
C. dubliniensis clinical isolates and culture conditions. Ninety-eight isolates ofC. dubliniensis from 94 separate individuals in 15 different countries were in-cluded in this study (Table 1). In addition, multiple single-colony isolates of C.dubliniensis recovered from the same clinical specimen obtained from eightseparate individuals (patients A through H, Table 1) were also studied.
For each population, cells were sampled directly from the site of colonizationby plating swab specimens from the mid-dorsum of the tongue onto CHROM-agar Candida medium (8, 34). After 48 h of incubation at 37°C, single-colonyisolates were presumptively identified as C. dubliniensis on the basis of their darkgreen coloration (8). Definitive identification was confirmed by the inability ofthe isolates to grow at 45°C (39) and by their substrate assimilation profiles usingthe API ID 32C yeast identification system (bioMérieux, Marcy l’Etoile, France)as described previously (38). Two of these individuals, patients D and E, eachharbored two distinct strains of C. dubliniensis, as determined by DNA finger-printing analysis (see Results section).
In a separate analysis, serial isolates were recovered from two separate pa-tients in the Netherlands over a defined time period (Table 2) (31). In the firstcase, a patient receiving chemotherapy for relapsed nasopharyngeal rhabdomyo-sarcoma yielded C. dubliniensis from a variety of anatomic sites (including blood)over a period of 5 months. In the second case, a patient suffering from graft-versus-host disease following an allogeneic hematopoietic stem cell transplantalso yielded a series of C. dubliniensis isolates in samples taken over a period of10 days. Following each sampling, isolates were identified as C. dubliniensis onthe basis of lack of growth at 45°C, dark green colony color on CHROMagarCandida medium, poor hybridization with the C. albicans fingerprinting probeCa3, and randomly amplified polymorphic DNA (RAPD) fingerprint patternanalysis (31). In each case a single colony was chosen for further analysis.
Isolates were routinely cultured on potato dextrose agar (PDA; Oxoid) me-dium, pH 5.6, at 37°C. For liquid culture, isolates were grown in yeast extract-peptone-dextrose (YPD) broth at 37°C in an orbital incubator (Gallenkamp,Leicester, United Kingdom) at 200 rpm.
Chemicals, enzymes, radioisotopes, and oligonucleotides. Analytical-grade ormolecular biology-grade chemicals were purchased from Sigma-Aldrich IrelandLtd. (Tallaght, Dublin, Ireland), BDH (Poole, Dorset, United Kingdom), orRoche Diagnostics Ltd. (Lewes, East Sussex, United Kingdom). Enzymes werepurchased from the Promega Corporation (Madison, Wis.) and from New En-gland Biolabs (Beverly, Mass.) and used according to the manufacturers’ instruc-tions. [�-32P]dATP (6,000 Ci mmol�1; 222 TBq mmol�1) was purchased fromAmersham International Plc. (Little Chalfont, Buckinghamshire, United King-dom). Custom-synthesized oligonucleotides were purchased from Sigma-Geno-sys Biotechnologies (Europe) Ltd. (Pampisford, Cambridgeshire, United King-dom).
Hybridization analysis and computer-assisted analysis of DNA fingerprintprofiles. Southern blot hybridization of restriction endonuclease-digestedgenomic DNA from C. dubliniensis isolates was performed as described previ-ously (57). EcoRI-digested DNA was electrophoresed through 0.65% (wt/vol)agarose gels for 16 h at 65 V and transferred by capillary blotting to nylonmembrane filters (Osmonics, Westborough, Mass.) as described by Sullivan et al.(57). Hybridization reactions were carried out under high-stringency conditionswith 32P-labeled probes labeled by random primer labeling (57).
The C. dubliniensis probes Cd25, Cd24, and Cd1 were used for fingerprintingisolates of C. dubliniensis. The complex probe Cd25 was used for fingerprinting
independent isolates of C. dubliniensis, whereas Cd24 and Cd1 were used toassess microevolution within individual strains (17). Computer-assisted analysesof hybridization patterns were performed using the DENDRON software pack-age version 2.3 (Solltech, Iowa City, Iowa) as described previously (17, 51, 52).DNA from the C. dubliniensis isolate CM6 (57) was used as a reference on eachgel used for computer-assisted analysis. Similarity coefficients (SABs) based onband position alone were calculated for each pairwise combination of isolatepatterns according to the formula SAB � 2E/(2E � a � b), where E is the numberof bands shared by strains A and B, a is the number of bands unique to A, andb is the number of bands unique to B (52). An SAB of 0.0 represents totallydifferent patterns with no correlated bands, an SAB of 1.00 represents identicalpatterns, and SABs ranging from 0.01 to 0.99 represent patterns with increasingproportions of bands at the same positions.
PCR amplification. The ITS1/ITS4 primer pair (Table 3) was used to amplifythe internal transcribed spacer 1 and 2 (ITS1 and ITS2, respectively) regions andthe intervening 5.8S rRNA gene of C. dubliniensis isolates. These primers arecomplementary to conserved regions of the fungal 18S and 28S rRNA genes,respectively, which flank the ITS1 and ITS2 regions (62). Specific restrictionendonuclease cleavage sites were incorporated into the primers to facilitatecloning of amplified products (Table 3). PCR amplifications were performedusing the High Fidelity PCR System (Roche, Lewes, East Sussex, United King-dom) following the recommendations of the manufacturer. In brief, each 100-�lreaction contained 1� Expand High Fidelity buffer; dATP, dTTP, dCTP, anddGTP at 200 �M each; 2.6 U of Expand High Fidelity PCR System enzyme mix;300 nM ITS1 and 300 nM ITS4; 1.5 mM MgCl2; and 100 ng of template DNA.Amplification reactions were carried out in a Perkin Elmer Cetus DNA thermalcycler with initial denaturation for 3 min at 95°C, followed by 35 cycles of 1 minat 95°C, 1 min at 58°C, and 2 min at 72°C, and a final extension for 10 min at72°C. Following amplification, the amplimers were purified using a PromegaPCR extraction kit and cloned into pBlusescript II KS(�) by conventionalmethods (47).
PCR identification of isolates belonging to C. dubliniensis genotypes 1, 2, 3,and 4 was performed using the primer pairs G1F/G1R, G2F/G2R, G3F/G3R,and G4F/G4R, respectively (Table 3). These primers were designed based onobserved differences in the nucleotide sequence of the ITS1 and ITS2 regions ofisolates belonging to each of the four C. dubliniensis genotypes (see Resultssection). The genotype 4-specific forward and reverse primers were designedwith a base mismatch at their 3� ends to increase their specificity (Table 3) (13).Each PCR was carried out with one pair of these genotype-specific primers andthe universal fungal primers RNAF/RNAR (13), which amplify approximately610 bp from all fungal large-subunit rRNA genes and were used as an internalpositive control.
PCR was carried out using Taq DNA polymerase (Promega). Each 100-�lPCR for genotypes 1, 2, and 3 contained 1� reaction buffer; dATP, dTTP, dCTP,and dGTP at 200 �M each; 2.5 U of Taq polymerase; a 300 nM concentration ofeach primer; 1.5 mM MgCl2; and either 100 ng of purified template DNA orrapidly prepared template DNA obtained by boiling yeast cells. Template DNAobtained by the latter method was prepared by boiling a single 48-h-old PDA-grown colony in 50 �l of sterile ultrapure H2O for 10 min. After boiling, thedebris was pelleted by centrifugation, and the DNA contained in 25 �l of thesupernatant was used as the template for subsequent PCRs. The PCR cyclingconditions were initial denaturation for 3 min at 95°C, followed by 30 cycles of 1min at 95°C, 1 min at 50°C, and 20 s at 72°C, and a final extension for 10 min at72°C. Optimum conditions for genotype 4-specific PCR were determined empir-ically. These were as above except that 20 �M deoxynucleoside triphosphates(dNTPs) were used and the annealing temperature was 55°C.
Following PCR, 5-�l aliquots of the amplification mixture from genotype 1-,2-, and 3- specific PCRs and 10 �l of the amplification mixture from genotype4-specific PCR were separated by electrophoresis through 2.4% (wt/vol) agarosegels containing 0.5 �g of ethidium bromide per ml and were visualized on a UVtransilluminator. Due to the lower dNTP concentration, less PCR product isproduced in the genotype 4-specific PCR compared to the other three genotype-specific PCRs.
DNA sequencing. Nucleotide sequence analysis was performed by the dideoxychain-terminating method of Sanger et al. (48) using an automated AppliedBiosystems 377 DNA sequencer and dye-labeled terminators (Applied Biosys-tems, Foster City, Calif.). The sequencing primers used were the M13 forwardand reverse primers. Sequence alignments were carried out using the Clustal Wsequence analysis computer software package (59).
Karyotype analysis. Yeast chromosomes were prepared in agarose plugs bythe method of Vazquez et al. (60) and resolved in 1.4% (wt/vol) agarose gels bypulsed-field gel electrophoresis using the CHEF-Mapper system (Bio-Rad, Rich-mond, Calif.). Saccharomyces cerevisiae chromosome preparations (Bio-Rad)
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 557
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

TABLE 1. C. dubliniensis isolates used in this studya
C. dubliniensis isolate AIDS or HIV status Sample Yr of isolation Country of origin Reference
CD71 � Oral 1994 Argentina 54CM1 � Oral 1991 Australia 57CM2 � Oral 1991 Australia 57CM4 � Oral 1991 Australia 57CM5 � Oral 1991 Australia 57
CM6 � Oral 1992 Australia 57Can 6 � Oral 1995 Canada 39Can 1 � Oral 1996 Canada 39Can 3 � Oral 1996 Canada 39Can 4 � Oral 1996 Canada 39
Can 9 � Oral 1996 Canada 39Can 13 � Oral 1996 Canada 39CD516 � Oral 1996 Finland 39CD96.29 � Oral 1996 Germany 39CD96.34 � Oral 1996 Germany 39
CD96.54 � Oral 1996 Germany 39CD96.63 � Oral 1996 Germany 39CD159 � Oral 1995 Greece 39CBS2747 � Sputum 1952 Netherlands 31CBS8500 � Blood 1998 Netherlands 31
CBS8501 � Blood 1998 Netherlands 31CD98923 � Oral 1998 India 1CD36 � Oral 1988 Ireland 57CD500 AIDS Oral 1988 Ireland 39CD33 � Oral 1989 Ireland 57
CD38 � Oral 1989 Ireland 57CD503 � Oral 1989 Ireland 39CD505 � Oral 1989 Ireland 39CD506 AIDS Oral 1989 Ireland This studyCD57 � Vagina 1992 Ireland 32
CD507 (A, 18) AIDS Oral 1992 Ireland This studyCD509 � Oral 1992 Ireland 39CD510 AIDS Oral 1994 Ireland 39CD511 � Oral 1995 Ireland 39CD512 � Oral 1995 Ireland 39
CD513 � Oral 1995 Ireland This studyCD514 (B, 11) � Oral 1995 Ireland This studyCD517 � Oral 1996 Ireland 39CD518 � Oral 1996 Ireland 39CD519 (C, 8) AIDS Oral 1997 Ireland This study
CD520 � Oral 1997 Ireland This studyCD521 � Oral 1997 Ireland 39CD522 � Oral 1997 Ireland 39CD523A (D, 17) � Oral 1997 Ireland This studyCD523B (D, 3)
CD524 � Oral 1997 Ireland This studyCD525 � Oral 1997 Ireland This studyCD526 � Oral 1998 Ireland This studyCD527A (E, 15) AIDS Oral 1998 Ireland This studyCD527B (E, 2)
CD528 (F, 20) AIDS Oral 1998 Ireland This studyCD529 (G, 18) AIDS Oral 1998 Ireland This studyCD530 � Oral 1998 Ireland This studyCD531 � Oral 1998 Ireland This studyCD532 � Oral 1998 Ireland This study
CD533 � Blood 1998 Ireland This studyCD536 (H, 10) � Oral 1998 Ireland This studyCD542 � Oral 1999 Ireland This studyCD543 AIDS Oral 1999 Ireland This studyCD2000 � Oral 2000 Ireland 1
Continued on following page
558 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

were included on each gel as reference standards. The electrophoresis bufferused was 1� TBE (Tris-borate-EDTA) (47) and was maintained at 14°C usingbuffer recycling through a Bio-Rad minichiller (model 1000). Gels were electro-phoresed at 4 V/cm for 72 h with an initial switch time of 40 s and a final switchtime of 600 s, with a ramping factor of �2.379 and an included angle of 110°.Following electrophoresis, gels were stained with 0.5 �g of ethidium bromide perml for 15 min, destained in distilled water, and viewed on a UV transilluminator.
The C. dubliniensis isolate CD33 (57) was used as a reference isolate for com-paring karyotype profiles on different gels.
Exposure of isolates of C. dubliniensis to fluconazole in vitro. One hundredfluconazole-susceptible colonies of each of the C. dubliniensis strains CD514 andCD36 (Table 1) were inoculated onto yeast peptone dextrose (YPD) agar me-dium (per liter: 10 g of yeast extract [Oxoid], 20 g of peptone [Difco], 20 g ofglucose, and 15 g of Bacto agar [Difco], pH 5.5) containing fluconazole at a
TABLE 1—Continued
C. dubliniensis isolate AIDS or HIV status Sample Yr of isolation Country of origin Reference
CD2003 � Oral 2000 Ireland 1CD2004 � Oral 2000 Ireland 1p6265 � Sputum 1999 Israel 40p6785 � Urine 1999 Israel 40p7276 � Respiratory tract 1999 Israel 40
p7858 � Vagina 1999 Israel This studyp7890 � Respiratory tract 1999 Israel This studyp7852 � Vagina 1999 Israel This studyp7507 � Sputum 1999 Israel This studyp7718 � Wound 1999 Israel This study
CD534 � N/A N/Ab Japan 1CD19398 � Oral 1998 Norway 1CD94191 � Oral 1994 Spain 39CD2491 � Oral 1994 Spain 39CD94234 � Oral 1994 Spain 39
CD941026 � Oral 1994 Spain 39CD94895 � Oral 1994 Spain 39CD94208 � Oral 1994 Spain 39CD94924 � Oral 1994 Spain 39CD96104 � Oral 1995 Spain 42
Co4 � Oral 1993 Switzerland 3Co5 � Oral 1993 Switzerland 3Co7 � Oral 1993 Switzerland 3P2 � Oral 1993 Switzerland 3P7 � Oral 1993 Switzerland 3
P21 � Oral 1993 Switzerland 3P27 � Oral 1993 Switzerland 3NCPF3108 � Stool 1957 United Kingdom 57CD75089 N/A Oral 1973 United Kingdom 39CD75043 N/A Oral 1975 United Kingdom 39
CD75004 N/A Oral 1975 United Kingdom 39CD538 � Stool 1986 United Kingdom 39CD539 AIDS Oral 1994 United Kingdom 39CD540 � Sputum 1997 United Kingdom 39CD541 � Blood 1997 United Kingdom 39
m26b � Oral 1995 United Kingdom 39m262b � Oral 1995 United Kingdom 39m196cd � Oral 1995 United Kingdom 39
a There were multiple isolates of CD507, CD514, CD519, CD523, CD527, CD528, CD529, and CD536. These isolates were recovered from the same oral specimen,in each case, from patients A to H. The patient designation and the number of isolates of each strain examined are indicated in parentheses after the strain name, e.g.,CD507 (A, 18) indicates there were 18 isolates of the strain CD507 examined and that this strain came from patient A. Two separate C. dubliniensis strains detectedon the basis of Cd25 fingerprint profiles were recovered from patients D and E, and these were termed CD523A and CD523B (patient D) and CD527A and CD527B(patient E). Multiple clonal isolates of strains CBS8500 (six isolates) and CBS8501 (15 isolates) were recovered from various body sites of two separate Dutch patients.Isolates CM1 and CM2 (57) and isolates Can4 and Can6 were both recovered from the same patients at separate clinical evaluations (39). The C. dubliniensis type strainCD36 is lodged with the American Type Culture Collection (accession number MYA-646) and with the British National Collection of Pathogenic Fungi (accessionnumber NCPF 3949). Isolates CBS2747, CBS8500, and CBS8501 are lodged with the Centraalbureau voor Schimmelcultures. Ten of the isolates investigated here wereincluded in the Joly et al. study (17), including the seven Cd25 group I isolates CD36, Co4, Co5, CM1, CM2, CM4, and CM6 and the three Cd25 group II isolatesCD75089, CD75043, and CD75004. The provenance of specific clusters of isolates highlighted in color in Fig. 1 was as follows: Australian isolates CM1, CM2, CM4,CM5, and CM6, highlighted in dark blue, were from patients at the Fairfield Hospital, Melbourne, Victoria; Canadian isolates Can3, Can4, Can6, and Can9, highlightedin blue, were from patients at the Centre hospitalier de l’Université Laval, Quebec City; Irish isolates CD36, CD38, CD503, CD507, CD522, CD2000, and CD2003,highlighted in green, were from patients at the Dublin Dental Hospital, whereas isolate CD57 was isolated from a patient at St. James’s Hospital, Dublin; Spanishisolates CD94924, CD941026, and CD94208, highlighted in red were from patients at the Hospital de Cruces, Bizkaia, Spain, whereas isolate CD94895 was isolatedfrom a patient at the Clínicas Odontológicas, University of the Basque Country, Bilbao, Spain.
b N/A, not available.
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 559
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

concentration of 0.5 �g/ml and incubated for 48 h at 37°C. Each colony was thenaseptically transferred using sterile toothpicks onto fresh YPD agar containing0.5 �g of fluconazole per ml and incubated for a further 48 h at 37°C. Eachcolony was then further subcultured twice as above on YPD agar containing
fluconazole at 1, 5, 10, 50, 60, and 70 �g/ml. At 50 �g/ml, derivatives of only 24of 100 of the CD514 colonies initially selected still grew; 20 of these were selectedfor further study and termed CD514Flu50 derivatives. At 70 �g of fluconazole perml, only 13 of 100 of the CD514 colonies still grew; all of these were selected forfurther study and termed CD514Flu70 derivatives. Derivatives of all 100 CD36colonies initially selected were obtained which were capable of growing on 70 �gof fluconazole per ml; 20 of these were selected for further study and termedCD36Flu70 derivatives. Similar experiments were also performed with C. dublini-ensis CD514 and CD36, which were repeatedly subcultured, as above, but in theabsence of fluconazole.
Antifungal susceptibility test methods. Microdilution susceptibility testingwith fluconazole was carried out by the method of Rodriguez-Tudela and Mar-tinez-Suárez on RPMI medium supplemented with 2% (wt/vol) glucose (44),which is a modification of the broth macrodilution method outlined in theNational Committee for Clinical Laboratory Standards document M-27P (33).The NCCLS-recommended quality control strains Candida krusei ATCC 6258and Candida parapsilosis ATCC 22019 were used (33). Fluconazole powder wasa gift from Pfizer Pharmaceuticals (Sandwich, Kent, United Kingdom).
RESULTS
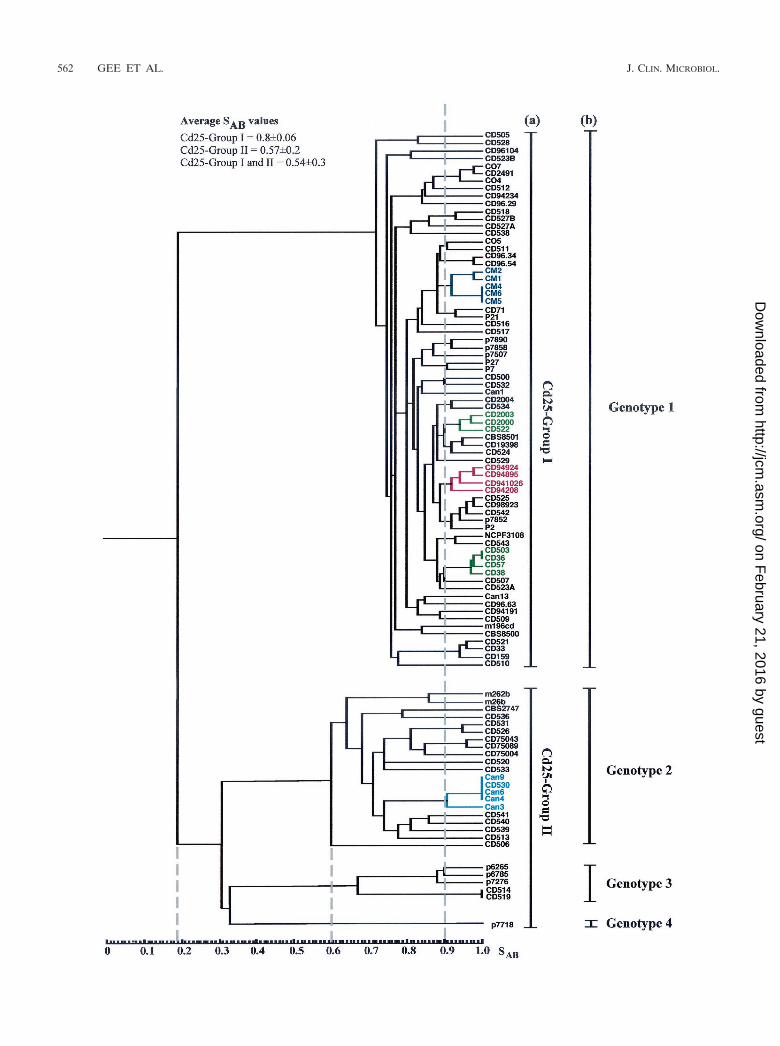
DNA fingerprint analysis of a diverse collection of C. dub-liniensis isolates. In order to investigate the range of geneticdiversity among C. dubliniensis isolates, a collection of 98 iso-lates recovered from 94 separate individuals in 15 differentcountries (Table 1) were fingerprinted with the C. dubliniensis-specific complex probe Cd25. The resulting complex hybridiza-tion profiles were subjected to computer-assisted analysis withthe fingerprint profile analysis software package DENDRON.Similarity coefficient (SAB) values were computed for everypossible pairwise combination of isolates, and these data wereused to construct a dendrogram showing the relationships be-tween all of the isolates in the collection (Fig. 1).
Examination of the dendrogram revealed that the isolatescould be clearly divided by a node at an SAB value of 0.19 intotwo main populations of isolates. The first population of iso-
TABLE 2. C. dubliniensis serial isolates recovered from varioussamples from two Dutch patientsa
Patientno. Isolate Date of isolation
(day-mo-yr) Sample
1 CBS 8501.1 26-06-1995 OralCBS 8501.2 17-07-1995 BronchialCBS 8501.3 25-07-1995 FecalCBS 8501.4 4-08-1995 OralCBS 8501.5 7-08-1995 SputumCBS 8501.6 11-08-1995 OralCBS 8501.7 13-08-1995 FecalCBS 8501.8 13-08-1995 BloodCBS 8501.9 13-08-1995 BloodCBS 8501.10 13-08-1995 BloodCBS 8501.11 14-08-1995 OralCBS 8501.12 14-08-1995 OralCBS 8501.13 18-08-1995 OralCBS 8501.14 19-08-1995 WoundCBS 8501.15 21-11-1995 Oral
2 CBS 8500.1 8-01-1996 FecalCBS 8500.2 12-01-1996 AscitesCBS 8500.3 12-01-1996 BloodCBS 8500.4 15-01-1996 BloodCBS 8500.5 15-01-1996 BloodCBS 8500.6 18-01-1996 Blood
a Serial isolates were recovered from two separate patients in the Netherlands(31). Patient 1 yielded C. dubliniensis from a variety of anatomic sites over aperiod of 5 months while receiving chemotherapy for relapsed nasopharyngealrhabdomyosarcoma. Patient 2 also yielded C. dubliniensis over a period of 10days while suffering from graft-versus-host disease following an allogeneic he-matopoietic stem cell transplant.
TABLE 3. Nucleotide sequence of PCR primers used to amplify specific regions of C. dubliniensis DNA
Primer Sequencea Purpose Reference
ITS1 5�-CGGAATTCTCCGTAGGTGAACCTGCGG-3� Forward primer for amplifying the ITSb region 62ITS4 5�-CGGAATTCTCCTCCGCTTATTGATATGC-3� Reverse primer for amplifying the ITS region 62G1F 5�-TTGGCGGTGGGCCCCTG-3� Forward primer for amplifying approx. 330 bp of the
ITS region from genotype 1 isolatesThis study
G1R 5�-AGCATCTCCGCCTTATA-3� Reverse primer for amplifying approx. 330 bp of theITS region from genotype 1 isolates
This study
G2F 5�-CGGTGGGCCTCTACC-3� Forward primer for amplifying approx. 330 bp of theITS region from genotype 2 isolates
This study
G2R 5�-CATCTCCGCCTTACC-3� Reverse primer for amplifying approx. 330 bp of theITS region from genotype 2 isolates
This study
G3F 5�-TTGGTGGTGGGCTTCTG-3� Forward primer for amplifying approx. 330 bp of theITS region from genotype 3 isolates
This study
G3R 5�-GCAATCTCCGCCTTACC-3� Reverse primer for amplifying approx. 330 bp of theITS region from genotype 3 isolates
This study
G4Fc 5�-GGCCTCTGCCTGCCGCCAGAGGATG-3� Forward primer for amplifying approx. 330 bp of theITS region from genotype 4 isolates
This study
G4Rc 5�-AGCAATCTCCGCCTTACT-3� Reverse primer for amplifying approx. 330 bp of theITS region from genotype 4 isolates
This study
RNAF 5�-GCATATCAATAAGCGGAGGAAAAG-3� Forward primer for amplifying approx. 610 bp of thelarge ribosomal subunit gene, used here as aninternal positive control in genotype-specific PCR
13
RNAR 5�-GGTCCGTGTTTCAAGACG-3� Reverse primer for amplifying approx. 610 bp of thelarge ribosomal subunit gene, used here as aninternal positive control in genotype-specific PCR
13
a Underlining indicates EcoRI restriction enzyme sites incorporated into the primer to facilitate cloning.b The ITS includes ITS1, 5.8S rRNA gene, and ITS2. The locations of the various GF and GR primer sequences within the ITS region are shown in Fig. 2a.c The G4F and G4R primers each differed from the ITS genotype 4 sequence (shown in Fig. 2a) by a single base-pair change at the 3� end to improve specificity.
560 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

lates was designated Cd25 group I (71 isolates), and the secondpopulation of isolates was designated Cd25 group II (27 iso-lates) (Fig. 1). These findings confirm and extend the findingsof a previous study with Cd25-generated fingerprint profilesobtained with a collection of 57 independent C. dubliniensisisolates from 11 different countries, in which the isolates wereclearly separated into two distinct groups at an SAB node valueof 0.24 (17). In the present study, the total collection of isolateshad an average SAB value of 0.54 � 0.30, Cd25 group I isolateshad an average SAB value of 0.80 � 0.06, and the Cd25 groupII isolates had an average SAB value of 0.57 � 0.20. At an SAB
threshold value of 0.60, four distinct clades are evident (Fig. 1).Cd25 group I isolates form a distinct clade, whereas Cd25group II isolates consist of three separate clades.
Multiple isolates of the same C. dubliniensis strain (withidentical or very closely related Cd25 fingerprint profiles) re-covered from the same clinical specimen for each of eightseparate individuals (patients A to H, Table 1) yielded SAB
values of �0.93 in each case (see below). Closely related clus-ters of isolates (e.g., SAB values of 0.9) are evident in thedendrogram. While many of these clusters have no obvioussignificance, in some cases clusters correspond to the hospitalof isolation and/or the country of origin of the isolates (Fig. 1and Table 1). For example, there are two clusters that com-prise solely isolates from Dublin, Ireland (highlighted in greenin Fig. 1), all of the Australian isolates which were obtained inthe same Melbourne hospital clustered closely together (high-lighted in dark blue in Fig. 1), four of the Spanish isolateswhich were obtained in two separate clinics or hospitals clus-tered together (highlighted in red in Fig. 1), and there is acluster of Canadian isolates from the same hospital with oneIrish isolate (highlighted in light blue in Fig. 1). There are alsoseveral examples of pairs of isolates recovered in the samehospital from different patients with an SAB value above 0.9(Fig. 1), including oral isolates CD75089 and CD75043, (SAB
� 0.98), recovered from patients in a hospital in Leeds, UnitedKingdom, and a pair of Irish oral isolates, CD518 and CD527B(SAB � 0.93), recovered in the Dublin Dental Hospital (Fig. 1).
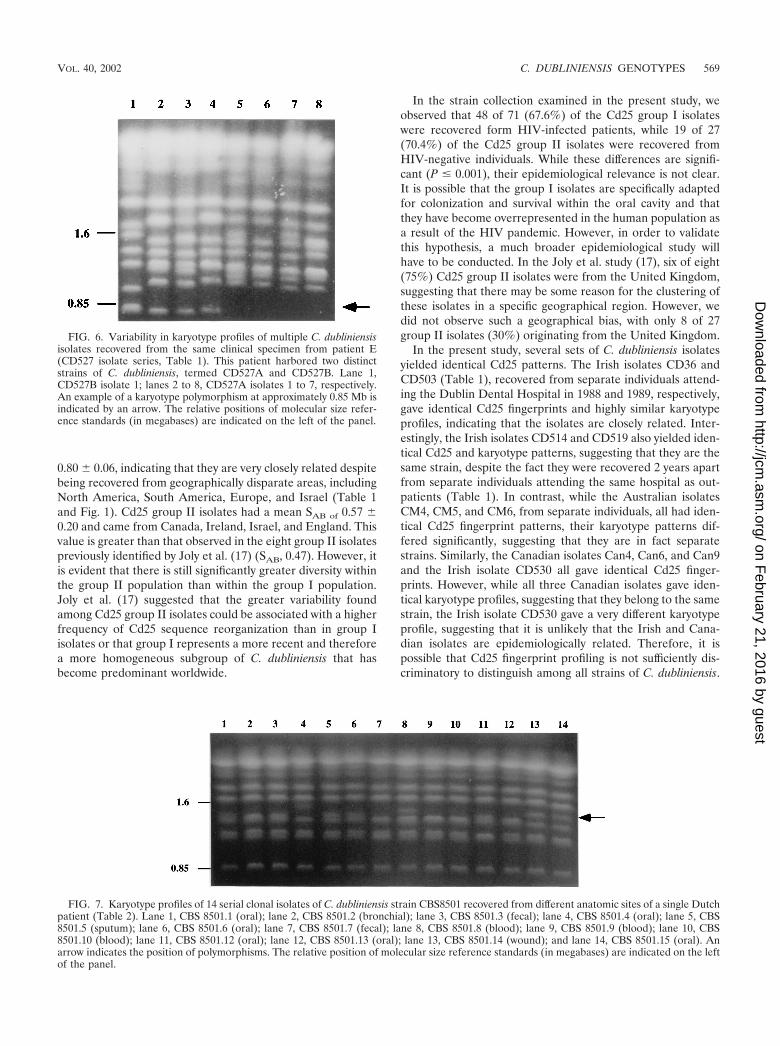
There were several examples where isolates recovered fromseparate individuals yielded identical Cd25 fingerprint profiles,suggesting that these individuals harbored identical or veryclosely related strains. In order to confirm this, these isolateswere also fingerprinted by karyotype analysis. Three AustralianCd25 group I isolates (CM4, CM5, and CM6, Fig. 1 and Table1) yielded identical Cd25 patterns; however, Sullivan et al.previously showed that these isolates yielded distinctly differ-ent karyotype patterns with �5 band differences (57), suggest-ing that these isolates are not necessarily the same strain. TheIrish Cd25 group I isolates CD36 and CD503 (Fig. 1 and Table1) yielded identical Cd25 fingerprint patterns and similarkaryotype patterns (five karyotype bands in common but dif-fered by three bands), the Irish Cd25 group II isolates CD514and CD519 also yielded identical Cd25 fingerprint patternsand karyotype patterns. This suggests that the isolates in eachpair are the same strain in each case.
In addition, the Canadian isolates Can4, Can6 (both recov-ered from the same patient at different times), and Can9 andthe Irish isolate CD530 (Table 1) yielded identical Cd25 fin-gerprint patterns (Fig. 1) despite the fact that the isolates wererecovered in different countries. The isolates Can4, Can6, and
Can9 also yielded identical karyotype patterns, strongly sug-gesting that they do in fact belong to the same strain. However,the Irish isolate CD530 yielded a very different karyotype pat-tern (only one karyotype band in common) from the Canadianisolates, suggesting that this isolate, which is epidemiologicallyunrelated to the Canadian isolates, is highly unlikely to be thesame strain.
The majority (48 of 71, 67.6%) of the Cd25 group I isolateswere recovered from HIV-infected patients, whereas the ma-jority (19 of 27, 70.4%) of the Cd25 group II isolates wererecovered from HIV-negative individuals (these include iso-lates CD75089, CD75043, and CD75004, which were recoveredbetween 1973 and 1975 from a cardiac unit from patientsassumed for the purpose of this study to be HIV negative)(Table 1). Overall, 23 of 27 (85%) of the Cd25 group II isolateswere recovered from individuals in Ireland, the Netherlands,the United Kingdom, or Israel. Eight of the 11 English isolatesincluded in this study belonged to Cd25 group II, and of these,7 were from HIV-negative individuals. A minority (17 of 98,17.3%) of the C. dubliniensis isolates in the total collectionstudied were nonoral isolates, and of these, 8 of 17 (47%)belonged to Cd25 group II.
Nucleotide sequence analysis of C. dubliniensis ITS regions.In order to determine whether the separation of isolates intoCd25 groups I and II on the basis of Cd25 fingerprint patternswas reflected by differences at the nucleotide sequence level,the nucleotide sequences of the ITS1 and ITS2 regions and theintervening 5.8S rRNA encoding DNA of 19 of the C. dublini-ensis isolates analyzed by DNA fingerprinting were deter-mined. The 19 isolates examined (Table 1) included 7 Cd25group I isolates (CD507, CD33, CD505, CD516, CD2491, P2,and CD96.54) and 12 Cd25 group II isolates (Can6, CD539,CD540, CD75043, CD520, CD506, p7718, CD514, CD519,p6265, p6785, and p7276). These 19 isolates were originallyselected because they represented a diverse range of isolatesfrom the four clades observed in Fig. 1, and each yielded adifferent fingerprint profile with the Cd25 probe, apart fromCD514 and CD519, which yielded identical fingerprints.
PCR amplification reactions were performed with templateDNA from each of the isolates using the ITS1/ITS4 primer pair(Table 3), which were designed to specifically amplify the ITSregion and flanking sequences. Specific restriction endonucle-ase cleavage sites were incorporated into the primers to facil-itate cloning of amplified products (Table 3). A single am-plimer of approximately 520 bp was obtained with templateDNA from each of the 19 isolates. Amplimer DNA from eachisolate was digested with restriction endonucleases and clonedinto vector plasmid pBluescript, and the nucleotide sequenceof both strands of the cloned insert was determined.
The ITS region (from the first nucleotide of ITS1 to the lastnucleotide of ITS2) of the seven Cd25 group I isolates wasfound to be 453 bp in length, whereas the corresponding ITSregion of the 12 Cd25 group II isolates varied between 451 and452 in length. Four sequence types were observed among the19 isolates sequenced, and the corresponding isolates weredesignated genotypes 1 to 4 (Fig. 2a). The seven Cd25 group Iisolates sequenced belonged to genotype 1 (ITS region 453 bplong; EMBL database accession no. AJ311895), six of the Cd25group II isolates sequenced belonged to genotype 2 (ITS re-gion 451 bp long; EMBL database accession no. AJ311896),
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 561
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from
562 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

five of the Cd25 group II isolates sequenced belonged to ge-notype 3 (ITS region 452 bp long; EMBL database accessionno. AJ311897), and the final Cd25 group II isolate sequencedbelonged to genotype 4 (ITS region 452 bp long; EMBL data-base accession no. AJ311898) (Fig. 2a).
Genotype 1 isolates differed from genotype 2 isolates at fivenucleotide positions, from the five genotype 3 isolates at sixnucleotide positions, and from the single genotype 4 isolate atsix nucleotide positions (Fig. 2a). Six of the seven genotype 1isolates examined yielded identical sequences; the remainingisolate exhibited a single base difference (C changed to a T) atnucleotide position 413. Similarly, five of the six genotype 2isolates examined yielded identical sequences, whereas oneisolate showed a single base difference (C changed to a T) atnucleotide position 92. The five genotype 3 isolates yieldedidentical sequences.
A phylogenetic neighbor-joining tree (45) based on the ITSsequences of the four C. dubliniensis genotypes and C. albicanswas generated using Clustal W (Fig. 2b). Each individual C.dubliniensis genotype in this tree corresponds to a distinctclade in the dendrogram shown in Fig. 1. The Cd25 group IIisolates separated into three distinct clades in the dendrogram,and these subgroups correspond to ITS genotypes 2, 3, and 4(Fig. 1). The genotype 3 and 4 isolates separate from thegenotype 2 isolates at an SAB node of 0.3. The genotype 4isolate separated from the genotype 3 isolates at an SAB nodeof 0.32. Since the order of data input by the unweighted pairgroup method with arithmetic mean, as used by the Dendronsoftware program, can affect branching and thus affect thestability of clusters in a dendrogram (2), data input was ran-domized 100 times and 100 separate dendrograms were gen-erated. In these dendrograms, the four genotypes always clus-tered as four distinct clades (Fig. 1), demonstrating theirstability.
Analysis of the nucleotide sequences of the ITS region ofgenotypes 1 to 4 indicated differences in the presence or ab-sence of specific restriction endonuclease cleavage sites (Fig.2a). Thus, there is a unique ApaI restriction enzyme sitepresent in the genotype 1 sequence only. A unique HaeIIIrestriction site is present in the sequences from genotypes 1, 2,and 4 which is absent in the corresponding genotype 3 se-quence (Fig. 2a). The presence of these restriction enzymecleavage sites can be used to distinguish genotypes 1 and 3 bydigestion of the 520-bp ITS amplimers generated with theITS1/ITS4 primers (data not shown). However, genotypes 2and 4 cannot be distinguished from each other based on theserestriction sites (Fig. 2a).
In order to be able to rapidly identify C. dubliniensis isolatesbelonging to each of the four genotypes determined by ITS
region sequencing, genotype-specific primer pairs were de-signed based on nucleotide sequence differences observed inthe ITS1 and ITS2 regions (Fig. 2, Table 3). Each genotype-specific primer pair was used separately to amplify a DNAfragment of approximately 330 bp from C. dubliniensis tem-plate DNA from each of 43 genotype 1 isolates (Cd25 group Iisolates), all 21 genotype 2 isolates, all five genotype 3 isolates,and the single genotype 4 isolate. PCRs also contained thefungal universal primers RNAF/RNAR (13), which amplify aproduct of approximately 610 bp from the fungal large-subunitrRNA gene and serve as an internal positive control. While allC. dubliniensis isolates should produce a product of approxi-mately 610 bp with the RNAF/RNAR primers, only C. dub-liniensis isolates belonging to the correct genotype should yielda product of approximately 330 bp with the appropriate gen-otype-specific primer pair.
All 70 C. dubliniensis isolates tested yielded the 610-bp prod-uct resulting from amplification with the fungal universal prim-ers. However, only the 43 genotype 1 isolates tested yielded aproduct of approximately 330 bp with the genotype 1 primerpair (G1F/G1R) (Fig. 3). Similarly, only the 21 genotype 2isolates yielded a product of approximately 330 bp with thegenotype 2 primer set (G2F/G2R), only the five genotype 3isolates yielded a product of approximately 330 bp with thegenotype 3 primer set (G3F/G3R), and only the single geno-type 4 isolate yielded a product of approximately 330 bp withthe genotype 4 primer set (G4F/G4R) (Fig. 3).
Karyotype analysis of C. dubliniensis isolates. Eighty-eight ofthe C. dubliniensis isolates fingerprinted with the Cd25 probewere also investigated by karyotype analysis, including 65 Cd25group I isolates and 23 Cd25 group II isolates (including 17 ITSgenotype 2 isolates, the 5 ITS genotype 3 isolates, and thesingle ITS genotype 4 isolate). All 23 Cd25 group II isolatestested had a chromosome-sized DNA band of approximately1.7 Mb present in their karyotype profiles that was absent inthe corresponding profiles of all but 5 (5 of 65, 7.6%) of theCd25 group I isolates tested. Furthermore, the majority (60 of65, 92.3%) of the Cd25 group I isolates tested had a chromo-some-sized DNA band of approximately 1.75 Mb present intheir karyotype profiles that was absent in the correspondingprofiles of all 23 Cd25 group II isolates tested (Fig. 4). A1.7-Mb band was also present in the five Cd25 group I isolateslacking the band of approximately 1.75 Mb.
In vivo microevolution in populations of C. dubliniensis. (i)Genetic variation in multiple isolates of C. dubliniensis fromthe same clinical specimen. To assess the genetic relatednessof populations of C. dubliniensis isolates from particular indi-viduals, multiple single-colony isolates recovered from thesame oral specimen from each of eight separate patients at-
FIG. 1. Dendrogram generated from the similarity coefficients (SABs) computed for every possible pairwise combination of 98 isolates from 94patients fingerprinted with Cd25. The origins of the isolates are shown in Table 1. At an SAB of 0.19 (short dashed vertical line), the isolates aredivided into two main populations, termed Cd25 groups I and II. At an SAB of 0.6 (short dashed vertical line), four distinct clades are evident; oneclade corresponds to Cd25 group I, whereas the other three clades are found within Cd25 group II. Multiple isolates of the same C. dubliniensisstrain recovered from the same clinical specimen for each of eight separate individuals yielded SAB values of � 0.93; the SAB value of 0.9 (dashedvertical line) was therefore chosen as an arbitrary threshold value for describing clusters. The cluster of Australian isolates is highlighted in darkblue, two clusters consisting of Irish isolates only are highlighted in green, a cluster of four Spanish isolates is highlighted in red; and a cluster ofCanadian isolates and one Irish isolate is highlighted in light blue. The Cd25 groups (a) and the ITS genotypes (b) of the isolates are shown tothe right of the dendrogram. All 27 of the Cd25 group II isolates and 43 of 71 of the Cd25 group I isolates had their genotypes determined bygenotype-specific PCR.
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 563
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from
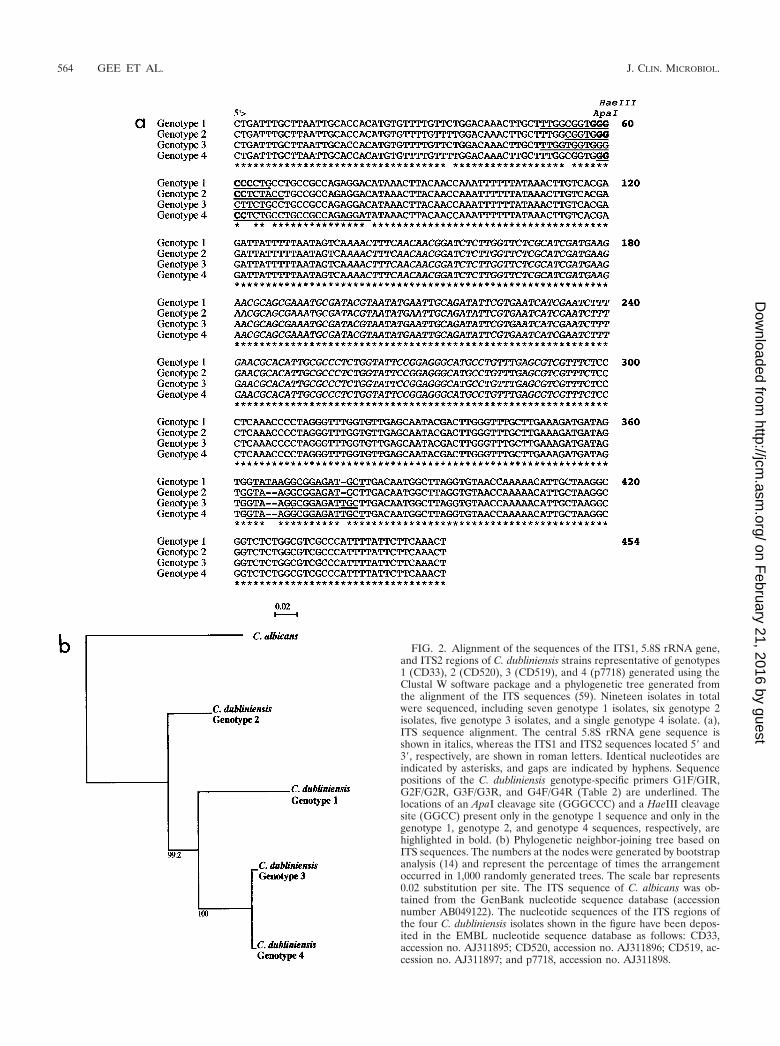
FIG. 2. Alignment of the sequences of the ITS1, 5.8S rRNA gene,and ITS2 regions of C. dubliniensis strains representative of genotypes1 (CD33), 2 (CD520), 3 (CD519), and 4 (p7718) generated using theClustal W software package and a phylogenetic tree generated fromthe alignment of the ITS sequences (59). Nineteen isolates in totalwere sequenced, including seven genotype 1 isolates, six genotype 2isolates, five genotype 3 isolates, and a single genotype 4 isolate. (a),ITS sequence alignment. The central 5.8S rRNA gene sequence isshown in italics, whereas the ITS1 and ITS2 sequences located 5� and3�, respectively, are shown in roman letters. Identical nucleotides areindicated by asterisks, and gaps are indicated by hyphens. Sequencepositions of the C. dubliniensis genotype-specific primers G1F/GIR,G2F/G2R, G3F/G3R, and G4F/G4R (Table 2) are underlined. Thelocations of an ApaI cleavage site (GGGCCC) and a HaeIII cleavagesite (GGCC) present only in the genotype 1 sequence and only in thegenotype 1, genotype 2, and genotype 4 sequences, respectively, arehighlighted in bold. (b) Phylogenetic neighbor-joining tree based onITS sequences. The numbers at the nodes were generated by bootstrapanalysis (14) and represent the percentage of times the arrangementoccurred in 1,000 randomly generated trees. The scale bar represents0.02 substitution per site. The ITS sequence of C. albicans was ob-tained from the GenBank nucleotide sequence database (accessionnumber AB049122). The nucleotide sequences of the ITS regions ofthe four C. dubliniensis isolates shown in the figure have been depos-ited in the EMBL nucleotide sequence database as follows: CD33,accession no. AJ311895; CD520, accession no. AJ311896; CD519, ac-cession no. AJ311897; and p7718, accession no. AJ311898.
564 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

tending the Dublin Dental Hospital, either with (patients A, C,D, and E, Table 1) or without (patients B, F, G, and H, Table1) signs of oral candidiasis, were fingerprinted with the com-plex probe Cd25. All of the isolates tested from six of eightpatients studied (patients A, B, C, F, G, and H, Table 1) werefound to be clonal in each case (mean SAB for isolates in eachpopulation, 0.93) (Table 4). Patients D and E were eachfound to harbor two separate strains of C. dubliniensis, basedon the Cd25 fingerprint profiles obtained (examples from pa-tient D are shown Fig. 5A). The SAB values calculated for thetwo different strains present in patients D (strains CD523Aand CD523B, Table 1) and E (strains CD527A and CD527B,Table 1) were 0.76 and 0.80, respectively.
None of the isolates tested from patients B, C, and H yieldedany variants with the Cd25 probe (for each population, themean SAB was 1; Table 4). However, minor hybridization banddifferences were observed in the profiles of the clonal isolatesfrom patients A, F, and G (mean SAB of 0.987 � 0.021, 0.992� 0.011, and 0.994 � 0.016, respectively) and in the profiles ofthe isolates of the most abundant strain in each case frompatients D and E (mean SAB of 0.937 � 0.072 and 0.985 �0.028, respectively) (Fig. 5A, Table 4). No variation in theCd25 fingerprint profiles of the minority strains from patient D(strain CD523B, three isolates) or E (strain CD527B, twoisolates) was observed.
Joly et al. (17) previously reported that the Cd1 and Cd24 C.dubliniensis fingerprinting probes were superior to Cd25 forassessing variability within a strain over time. In the presentstudy, Cd1 and Cd24 were used separately to fingerprint theclonal populations from patients A to H. Using these probes,minor band variation, or microevolution, was observed in thepopulations from patients A, D, E, F, and G, as also seen withCd25, but in addition, microevolution was also observed in thepopulation from patient H but not in the populations frompatients B and C (Table 4). Furthermore, the hybridizationpatterns obtained with probes Cd24 and Cd1 readily distin-guished the two strains detected in the population of isolates
from patients D and E using the Cd25 probe (data not shown).Figure 5B shows examples of the variability detected amongclonal isolates from patient A with the Cd24 probe. As withCd25, no variation in the Cd24 and Cd1 fingerprint profiles ofthe minority strains of patient D (strain CD523B, three iso-lates) and E (strain CD527B, two isolates) was observed.
Isolates from each of the populations recovered from pa-tients A to H were also examined by karyotype analysis. Vari-ation in the karyotype profile was observed in the same isolatepopulations that exhibited microevolution using Cd25 (pa-tients A, D, E, F, and G) but not in the populations frompatients B, C, and H (Table 4). Furthermore, the karyotypepatterns obtained also distinguished the two strains detected inthe population of isolates from patients D (data not shown)and E using the Cd25 probe (Fig. 6, lanes 1 and 2). The clonalisolates belonging to the majority strain from patient E (strainCD527A, Table 1) showed the greatest variability in karyotypepattern, with nine different patterns detected in 15 isolatesexamined (Table 4), examples of which are shown in Fig. 6,lanes 2 to 8. Variability was observed in the karyotype patternof one of the three isolates of the minority strain of patient D(data not shown), although no variability was observed in theminority strain of patient E (data not shown).
The isolates from patients A to D and F to H were allsusceptible to fluconazole (�8 �g/ml), while the isolates frompatient E (CD527) varied in their susceptibility to fluconazole.Four of the 15 isolates of the predominant strain from patientE (CD527A) had reduced susceptibility to fluconazole (MIC,16 �g/ml), whereas the remaining 11 isolates were susceptible
FIG. 3. Agarose gel showing ethidium bromide-stained amplimers(approximately 330-bp products) from PCRs with C. dubliniensis gen-otype-specific primer sets G1F/G1R (lanes 2 to 5), G2F/G2R (lanes 6to 9), G3F/G3R (lanes 10 to 13), and G4F/G4R (14 to 17) and thefungal universal primers RNAF and RNAR (610-bp product). Theamplimers shown in the lanes were obtained from template DNA fromC. dubliniensis isolates as follows: lanes 2, 6, 10, and 14, CD505 (ge-notype 1); lanes 3, 7, 11, and 15, CD536 (genotype 2); lanes 4, 8, 12,and 16, CD514 (genotype 3); and lanes 5, 9, 13, and 17, p7718 (geno-type 4). Lanes 1 and 19, molecular size reference markers (100-bpladder); lane 18, negative control reaction lacking template DNA butincluding RNAR and RNAF primers.
FIG. 4. Karyotype analysis of C. dubliniensis isolates. The karyo-types shown correspond to C. dubliniensis genotype 1 isolates CD33(lane 1) and CD94924 (lane 2) and genotype 2 isolates Can6 (lane 3),Can3 (lane 4), and CD541 (lane 5). The position of a chromosome-sized band of approximately 1.75 Mb present in the genotype 1 isolateprofiles but absent in genotype 2 isolate profiles is indicated by anarrow. The position of a chromosome-sized band of approximately 1.7Mb present in the genotype 2 isolate profiles but absent in the majorityof genotype 1 isolate profiles is also indicated by an arrow. Genotypes3 and 4 lacked a band at 1.75 Mb and, like the genotype 2 isolates, hada band at approximately 1.7 Mb (data not shown). The relative posi-tions of molecular size reference markers (in megabases) are shown onthe left.
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 565
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

to fluconazole (MIC, �1 �g/ml). The four isolates with re-duced susceptibility to fluconazole and one isolate that wassusceptible to fluconazole (MIC, 1.0 �g/ml) had Cd24 and Cd1patterns which differed from the patterns of the remaining 10susceptible isolates. The karyotype profiles of the four isolateswith reduced susceptibility to fluconazole and one isolate thatwas susceptible to fluconazole (MIC, 1.0 �g/ml) all lacked aband at approximately 0.85 Mb (Fig. 6, lanes 5 to 8) that waspresent in the remaining 10 susceptible isolates (Fig. 6, lanes 2to 4). The two isolates from the minority strain from patient E(CD527B) were susceptible to fluconazole (MIC, 1 �g/ml).
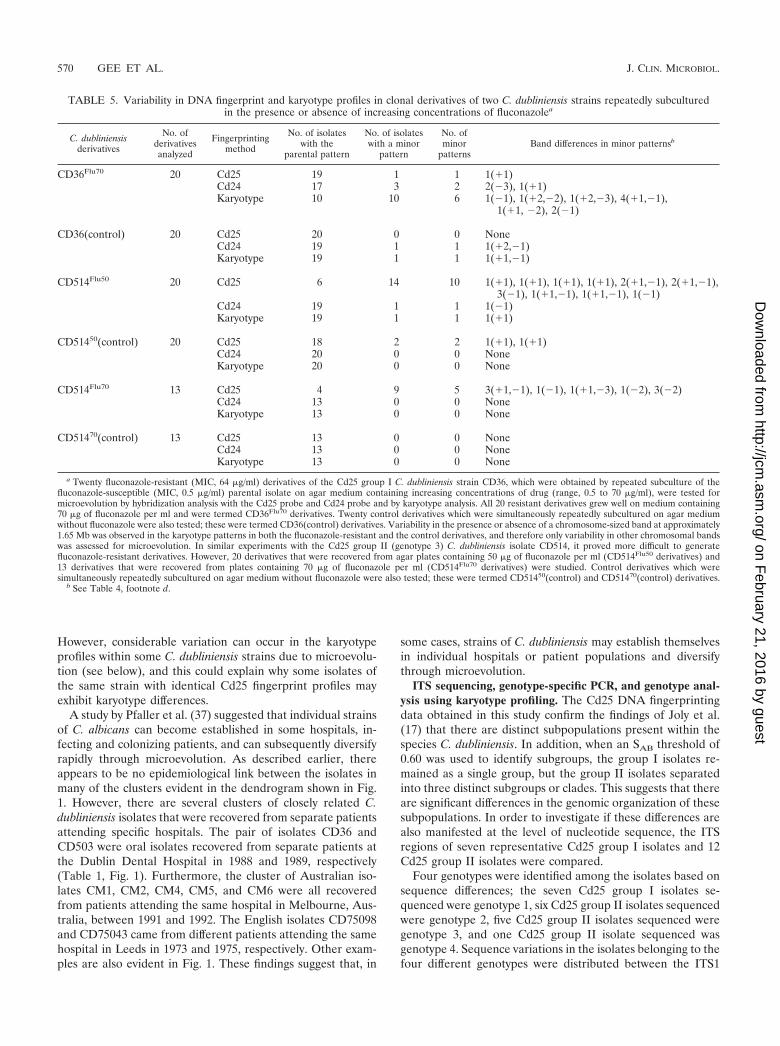
(ii) Genetic variation in serial isolates of C. dubliniensis.Serial isolates of C. dubliniensis from two separate patientswere also fingerprinted. These two cases of systemic infectioncaused by C. dubliniensis were recently reported by Meis et al.(31). For one of the patients concerned, samples were takenfrom a variety of anatomic locations, including oral, bronchial,fecal, wound, and blood samples, during a 5-month period(Table 2). The first sample was taken from the oral cavity,which had signs of oral candidosis. Other samples (includingsputum, fecal, oral, and wound samples) were subsequentlytaken during the following 5 months for surveillance purposes.Blood samples were also taken when the patient had a fever,which was refractory to antibiotic therapy, suggesting a possi-ble fungal infection. C. dubliniensis was recovered in combina-tion with C. albicans from all of the samples, with the exceptionof the blood cultures, which yielded C. dubliniensis only.
In each case, a single C. dubliniensis colony was taken fromthe primary isolation plate and fingerprinted. Using the Cd25probe, all 15 isolates recovered from different clinical samplesyielded very similar fingerprint patterns (average SAB � 0.98 �0.03) belonging to Cd25 group I, suggesting that the candi-demia was endogenously acquired. Fingerprinting using theprobes Cd25, Cd24, and Cd1 (data not shown) and karyotypeanalysis (Fig. 7) revealed the presence of minor polymor-phisms indicative of microevolution.
In a second patient, C. dubliniensis isolates were recoveredfrom four blood, one fecal, and one ascites sample taken overa period of 10 days. All six isolates were found to belong to asingle Cd25 group I strain using the Cd25 probe. Fingerprint-ing with the Cd25 (average SAB � 0.99 � 0.02) and Cd1 probesand karyotype analysis showed that there were minor variantspresent; however, no variation was observed with the Cd24probe (data not shown).
In vitro microevolution in clonal populations of C. dublini-ensis exposed to fluconazole. It has previously been shown thatfluconazole resistance can be induced in C. dubliniensis by invitro exposure to the drug (32). In a similar experiment, 20fluconazole-resistant (MIC, 64 �g/ml) derivatives of the Cd25group I (ITS genotype 1) C. dubliniensis strain CD36, termedCD36Flu70, were obtained by repeated subculture of the flu-conazole-susceptible (MIC, 0.5 �g/ml) parental isolate on agarmedium containing increasing concentrations of drug (range,0.5 to 70 �g/ml). All 20 resistant derivatives grew well onmedium containing 70 �g of fluconazole per ml.
Twenty control derivatives, termed CD36(control), were si-multaneously repeatedly subcultured on agar medium withoutfluconazole. These CD36Flu70 and CD36(control) derivativeswere tested for microevolution by hybridization analysis withthe Cd25, Cd24, and Cd1 probes and by karyotype analysis. A
single variant (Table 5) was detected in the hybridization pat-terns of the 20 CD36Flu70 derivatives with the Cd25 probewhile no variants were observed in the control derivatives (Ta-ble 5). Three variants (Table 5) were detected in the hybrid-ization patterns of the CD36Flu70 derivatives with the Cd24probe while only one variant (Table 5) was observed with thecontrol derivatives. Variability was observed in the Cd1 hybrid-ization patterns in both the fluconazole-resistant and controlderivatives (data not shown).
Variability in the presence or absence of a chromosome-sized band of approximately 1.65 Mb was observed in thekaryotype patterns in both the fluconazole-resistant and thecontrol derivatives (Fig. 8A and 8B). This band was present in5 of 20 of the CD36Flu70 derivatives and 11 of 20 of the controlderivatives. In addition, 18 separate colonies of the parentalCD36 strain from the same PDA plate culture were also fin-gerprinted by karyotype analysis, and 12 of 18 colonies had aband at approximately 1.65 Mb (data not shown). However,apart from variability in this band at 1.65 Mb, only 1 of 20 ofthe control derivatives showed variability in other karyotypebands, compared with 10 of 20 of the CD36Flu70 derivatives(Fig. 8A and 8B). These results suggested that exposure of C.dubliniensis CD36 to fluconazole in vitro might be associatedwith increased levels of genetic variation, manifested aschanges in Cd24 fingerprint and/or karyotype profiles, partic-ularly the latter.
In similar experiments using the Cd25 group II (ITS geno-type 3) C. dubliniensis isolate CD514, it proved more difficult togenerate fluconazole-resistant derivatives. The majority of the100 colonies plated on increasing concentrations of flucon-azole grew very poorly; at 50 �g of fluconazole per ml, only 24out of the original 100 colonies grew, and at 70 �g of flucon-azole per ml, only 13 colonies grew.
Twenty derivatives recovered from agar plates containing 50�g of fluconazole per ml (CD514Flu50 derivatives) and 13 de-rivatives recovered from plates containing 70 �g of fluconazoleper ml (CD514Flu70 derivatives) were investigated for geneticvariation. Most of the CD514Flu50 derivatives and CD514Flu70
derivatives grew poorly on medium containing 50 �g and 70 �gof fluconazole per ml, respectively. Four each of theCD514Flu50 and the CD514Flu70 derivatives exhibited dose-dependent fluconazole susceptibility (MIC, 16 �g/ml), one ofthe CD514Flu70 derivatives expressed fluconazole resistance(MIC, 64 �g/ml), while all the remaining derivatives werefluconazole susceptible (MIC, 8 �g/ml). No variability wasobserved in any of the fluconazole-exposed or control deriva-tives with the Cd1 probe. Variability was observed in onefluconazole-exposed derivative (Table 5) with the Cd24 probe,but no variability was observed in any of the control derivatives(Table 5). However, minor variations were detected in somefluconazole-exposed derivatives with the Cd25 probe, includ-ing susceptible derivatives and derivatives with reduced sus-ceptibility (Fig. 8C, Table 5). None of the control derivatives ofCD514 which were subcultured in the absence of drug exhib-ited Cd24 or Cd1 fingerprint or karyotype variations, but twoexhibited a minor band variation in the Cd25 fingerprint pat-tern (Table 5).
One of the fluconazole-exposed derivatives of CD514 exhib-ited a minor band change in the karyotype profile. It is inter-esting that when multiple single-colony isolates of CD514 orig-
566 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

TA
BL
E4.
Variability
inD
NA
fingerprintand
karyotypeprofiles
ineight
clonalpopulationsa
ofC
.dubliniensisisolates
detectedw
iththe
Cd25,C
d24,and
Cd1
probesand
bykaryotype
analysis
PatientStrain
bProbe
orkaryotypeanalysis
No.of
isolatestested
Avg
SA
Bc
No.of
isolatesw
iththe
predominant
pattern
No.of
isolatesw
itha
minor
pattern
No.of
minor
patternsB
anddifferences
inm
inorpatterns d
AC
D507
(I,1)C
d2518
0.987�
0.02115
33
1(�1),1(�
1),1(�1)
Cd24
810
53(�
1,�1),4(�
1,�1),1(�
1),1(�2,�
2),1(�2,�
1)C
d10
1818
All18
isolateshad
adifferent
patternK
aryotype5
136
3(�1,�
1),1(�1),2(�
1,�2),5(�
1),1(�1,�
2),1(�2,�
1)
BC
D514
(II,3)C
d2511
1.0011
00
None
Cd24
110
0N
oneC
d111
00
None
Karyotype
110
0N
one
CC
D519
(II,3)C
d258
1.008
00
None
Cd24
80
0N
oneC
d18
00
None
Karyotype
80
0N
one
De
CD
523A(I,1)
Cd25
170.937
�0.072
125
42(�
3),1(�3,�
1),1(�1),1(�
1,�1)
Cd24
143
31(�
1),1(�3,�
2),1(�1)
Cd1
413
113(�
1),1(�2,�
2),1(�1),1(�
1),1(�1),1(�
1),1(�1,�
3),1(�3,�
2),1(�
2,�1),1(�
3,�2),1(�
1,�1)
Karyotype
161
11(�
3,�
2)
Ee
CD
527A(I,1)
Cd25
150.985
�0.028
132
21(�
1,�2),1(�
1)C
d2410
52
4(�1),1(�
2)C
d110
51
5(�1)
Karyotype
312
91(�
1,�1),2(�
1,�1),3(�
2,�1),1(�
3,�2),1(�
3,�2),1(�
3,�2),1(�
3,�3),
1(�2,�
1),1(�2,�
2)
FC
D528
(I,1)C
d2520
0.992�
0.01117
31
3(�1)
Cd24
812
27(�
1),5(�2,�
1)C
d19
116
1(�1,�
2),3(�1,�
1),3(�1,�
1),2(�1,�
2),1(�2,�
1),1(�1,�
1)K
aryotype9
113
5(�1),4(�
1),2(�3,�
1)
GC
D529
(I,1)C
d2518
0.994�
0.01617
11
1(�1)
Cd24
171
11(�
1,�1)
Cd1
513
62(�
1),3(�1),3(�
1,�1),2(�
1,�1),1(�
1,�2),2(�
2,�1)
Karyotype
135
41(�
1),1(�1),2(�
1,�1),1(�
1,�1)
HC
D536
(II,2)C
d2510
1.0010
00
None
Cd24
91
11(�
1)C
d15
54
1(�1,�
3),2(�1),1(�
1,�1),1(�
1)K
aryotype10
00
None
aM
ultiplesingle-colony
isolatesrecovered
fromthe
same
oralspecimen
foreach
ofeight
individualsw
ereinvestigated.
bT
heC
d25group
(Ior
II)and
thegenotype
(1,2,or3)
tow
hicheach
strainbelongs
areindicated
inparentheses.
cSimilarity
coefficient(SA
B )values
were
calculatedfrom
theC
d25-generatedhybridization
patternsfor
allpossiblecom
binationsofisolate
pairsin
eachpopulation
usingthe
DE
ND
RO
Nsoftw
arepackage.T
hem
eanaverage
SA
Bvalue
forthe
isolatesin
eachpopulation
isshow
n.d
Each
variantpatternw
asassessed
accordingto
thenum
berofbands
differingfrom
thepredom
inantpattern,e.g.,(�1),a
bandaddition;(�
1),aband
loss;(�1,�
1),aband
additionand
aband
loss.Num
bersin
frontoftheparentheses
referto
thenum
berofisolates
with
theparticular
variantpattern,e.g.,2(�2,�
1)indicates
thattwo
isolateshad
thesam
epattern
thatdifferedfrom
thepredom
inantpatternby
theaddition
oftwo
bandsand
theloss
ofone
band.ePatients
Dand
Ew
ereeach
foundto
harbortw
odistinctstrains
ofC.dubliniensison
thebasis
ofCd25
fingerprintprofiles,oneofw
hichpredom
inatedin
eachcase.T
heC
.dubliniensisisolatepopulation
frompatientD
studiedconsisted
of17isolates
ofstrainC
D523A
and3
isolatesofstrain
CD
523B(T
able1),w
hereasthe
C.dubliniensis
isolatepopulation
studiedfrom
patientEconsisted
of15isolates
ofstrainC
D527A
and2
isolatesofstrain
CD
527B(T
able1).N
oneof
theC
D523B
orC
D527B
isolatesshow
edvariation
infingerprint
patternsw
ithany
ofthe
threeprobes
used,andonly
oneisolate
ofC
D523B
showed
am
inorvariation
inkaryotype
profile(data
notshow
n).
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 567
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

inally recovered from the same clinical specimen (and theidentical strain CD519) were examined by Cd25, Cd24, andCd1 fingerprinting and by karyotype analysis, no variation wasdetected.
DISCUSSION
Cd25 fingerprint analysis of C. dubliniensis isolates fromdisparate geographic locations. Analysis of hybridization pat-terns generated by complex DNA probes has proven to be oneof the most effective methods for investigating the epidemiol-ogy of several species of Candida, since it fulfills many of therequirements for performing computer-assisted broad epide-miological studies (12, 18, 24, 50, 51, 52). In 1999 Joly et al.(17) used the moderately repetitive C. dubliniensis-specificDNA fingerprinting probe Cd25 to investigate 57 independentC. dubliniensis isolates and found that the isolates appeared tofall into two distinct populations. One of the purposes of thepresent study was to investigate the findings of Joly et al. andto examine further the population structure of C. dubliniensisusing a larger number of isolates from a broader geographicalrange than used in the original study.
Ninety-eight isolates of C. dubliniensis from 94 separate in-
dividuals obtained from 15 countries around the world werestudied, including 10 of the isolates from the study of Joly et al.(17) (Table 1). These were subjected to DNA fingerprintinganalysis using the Cd25 fingerprinting probe. Computer-as-sisted analysis of the fingerprint patterns obtained showed thatthe isolates separated clearly into two populations correspond-ing to the two groups described by Joly et al. (17). The firstpopulation of isolates was designated Cd25 group I, and thesecond population was designated Cd25 group II (Fig. 1). Asimilar form of dichotomy was observed previously in C. albi-cans by Pujol et al. (41), who used the complex probe Ca3 todistinguish major groups in C. albicans. These groups weresubsequently confirmed by Lott et al. (25) using microsatellitesand IS1 analysis. In addition, a recent study using the complexC. parapsilosis fingerprinting probe Cp3–13 confirmed the ex-istence of three major subgroups of C. parapsilosis (12), orig-inally suggested on the basis of restriction fragment lengthpolymorphism analysis (49), random amplified polymorphicDNA analysis (21), karyotype analysis (26), isoenzyme profiles,and ITS sequencing (22).
In the present study, the majority (72%) of the isolatesexamined belonged to Cd25 group I, with a mean SAB value of
FIG. 5. DNA fingerprinting patterns of multiple C. dubliniensis isolates recovered from the same clinical specimen from patients A and D.(A) Probe Cd25-generated hybridization patterns of EcoRI-digested genomic DNA from 10 single-colony isolates of C. dubliniensis from patientD (Table 1). This patient harbored two distinct strains of C. dubliniensis, termed CD523A and CD523B. Lane 1, C. dubliniensis reference strainCM6 (57); lane 2, CD523B isolate 1; lanes 3 to 10, CD523A isolates 1 to 8, respectively; lane 11, CD523B isolate 2. Examples of polymorphismsin the profiles of CD523A isolates are indicated by arrows. (B) Probe Cd24-generated hybridization patterns of EcoRI-digested genomic DNAfrom 11 single-colony isolates of C. dubliniensis from patient A. This patient harbored a single strain of C. dubliniensis (CD507 isolate series, Table1). Lane 1, C. dubliniensis reference strain CM6; lanes 2 to 12, CD507 isolates 1 to 11, respectively. Polymorphisms in the profiles are indicatedby arrows. The relative positions of molecular size reference markers (in kilobases) are shown on the left of the panels.
568 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from
0.80 � 0.06, indicating that they are very closely related despitebeing recovered from geographically disparate areas, includingNorth America, South America, Europe, and Israel (Table 1and Fig. 1). Cd25 group II isolates had a mean SAB of 0.57 �0.20 and came from Canada, Ireland, Israel, and England. Thisvalue is greater than that observed in the eight group II isolatespreviously identified by Joly et al. (17) (SAB, 0.47). However, itis evident that there is still significantly greater diversity withinthe group II population than within the group I population.Joly et al. (17) suggested that the greater variability foundamong Cd25 group II isolates could be associated with a higherfrequency of Cd25 sequence reorganization than in group Iisolates or that group I represents a more recent and thereforea more homogeneous subgroup of C. dubliniensis that hasbecome predominant worldwide.
In the strain collection examined in the present study, weobserved that 48 of 71 (67.6%) of the Cd25 group I isolateswere recovered form HIV-infected patients, while 19 of 27(70.4%) of the Cd25 group II isolates were recovered fromHIV-negative individuals. While these differences are signifi-cant (P � 0.001), their epidemiological relevance is not clear.It is possible that the group I isolates are specifically adaptedfor colonization and survival within the oral cavity and thatthey have become overrepresented in the human population asa result of the HIV pandemic. However, in order to validatethis hypothesis, a much broader epidemiological study willhave to be conducted. In the Joly et al. study (17), six of eight(75%) Cd25 group II isolates were from the United Kingdom,suggesting that there may be some reason for the clustering ofthese isolates in a specific geographical region. However, wedid not observe such a geographical bias, with only 8 of 27group II isolates (30%) originating from the United Kingdom.
In the present study, several sets of C. dubliniensis isolatesyielded identical Cd25 patterns. The Irish isolates CD36 andCD503 (Table 1), recovered from separate individuals attend-ing the Dublin Dental Hospital in 1988 and 1989, respectively,gave identical Cd25 fingerprints and highly similar karyotypeprofiles, indicating that the isolates are closely related. Inter-estingly, the Irish isolates CD514 and CD519 also yielded iden-tical Cd25 and karyotype patterns, suggesting that they are thesame strain, despite the fact they were recovered 2 years apartfrom separate individuals attending the same hospital as out-patients (Table 1). In contrast, while the Australian isolatesCM4, CM5, and CM6, from separate individuals, all had iden-tical Cd25 fingerprint patterns, their karyotype patterns dif-fered significantly, suggesting that they are in fact separatestrains. Similarly, the Canadian isolates Can4, Can6, and Can9and the Irish isolate CD530 all gave identical Cd25 finger-prints. However, while all three Canadian isolates gave iden-tical karyotype profiles, suggesting that they belong to the samestrain, the Irish isolate CD530 gave a very different karyotypeprofile, suggesting that it is unlikely that the Irish and Cana-dian isolates are epidemiologically related. Therefore, it ispossible that Cd25 fingerprint profiling is not sufficiently dis-criminatory to distinguish among all strains of C. dubliniensis.
FIG. 6. Variability in karyotype profiles of multiple C. dubliniensisisolates recovered from the same clinical specimen from patient E(CD527 isolate series, Table 1). This patient harbored two distinctstrains of C. dubliniensis, termed CD527A and CD527B. Lane 1,CD527B isolate 1; lanes 2 to 8, CD527A isolates 1 to 7, respectively.An example of a karyotype polymorphism at approximately 0.85 Mb isindicated by an arrow. The relative positions of molecular size refer-ence standards (in megabases) are indicated on the left of the panel.
FIG. 7. Karyotype profiles of 14 serial clonal isolates of C. dubliniensis strain CBS8501 recovered from different anatomic sites of a single Dutchpatient (Table 2). Lane 1, CBS 8501.1 (oral); lane 2, CBS 8501.2 (bronchial); lane 3, CBS 8501.3 (fecal); lane 4, CBS 8501.4 (oral); lane 5, CBS8501.5 (sputum); lane 6, CBS 8501.6 (oral); lane 7, CBS 8501.7 (fecal); lane 8, CBS 8501.8 (blood); lane 9, CBS 8501.9 (blood); lane 10, CBS8501.10 (blood); lane 11, CBS 8501.12 (oral); lane 12, CBS 8501.13 (oral); lane 13, CBS 8501.14 (wound); and lane 14, CBS 8501.15 (oral). Anarrow indicates the position of polymorphisms. The relative position of molecular size reference standards (in megabases) are indicated on the leftof the panel.
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 569
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from
However, considerable variation can occur in the karyotypeprofiles within some C. dubliniensis strains due to microevolu-tion (see below), and this could explain why some isolates ofthe same strain with identical Cd25 fingerprint profiles mayexhibit karyotype differences.
A study by Pfaller et al. (37) suggested that individual strainsof C. albicans can become established in some hospitals, in-fecting and colonizing patients, and can subsequently diversifyrapidly through microevolution. As described earlier, thereappears to be no epidemiological link between the isolates inmany of the clusters evident in the dendrogram shown in Fig.1. However, there are several clusters of closely related C.dubliniensis isolates that were recovered from separate patientsattending specific hospitals. The pair of isolates CD36 andCD503 were oral isolates recovered from separate patients atthe Dublin Dental Hospital in 1988 and 1989, respectively(Table 1, Fig. 1). Furthermore, the cluster of Australian iso-lates CM1, CM2, CM4, CM5, and CM6 were all recoveredfrom patients attending the same hospital in Melbourne, Aus-tralia, between 1991 and 1992. The English isolates CD75098and CD75043 came from different patients attending the samehospital in Leeds in 1973 and 1975, respectively. Other exam-ples are also evident in Fig. 1. These findings suggest that, in
some cases, strains of C. dubliniensis may establish themselvesin individual hospitals or patient populations and diversifythrough microevolution.
ITS sequencing, genotype-specific PCR, and genotype anal-ysis using karyotype profiling. The Cd25 DNA fingerprintingdata obtained in this study confirm the findings of Joly et al.(17) that there are distinct subpopulations present within thespecies C. dubliniensis. In addition, when an SAB threshold of0.60 was used to identify subgroups, the group I isolates re-mained as a single group, but the group II isolates separatedinto three distinct subgroups or clades. This suggests that thereare significant differences in the genomic organization of thesesubpopulations. In order to investigate if these differences arealso manifested at the level of nucleotide sequence, the ITSregions of seven representative Cd25 group I isolates and 12Cd25 group II isolates were compared.
Four genotypes were identified among the isolates based onsequence differences; the seven Cd25 group I isolates se-quenced were genotype 1, six Cd25 group II isolates sequencedwere genotype 2, five Cd25 group II isolates sequenced weregenotype 3, and one Cd25 group II isolate sequenced wasgenotype 4. Sequence variations in the isolates belonging to thefour different genotypes were distributed between the ITS1
TABLE 5. Variability in DNA fingerprint and karyotype profiles in clonal derivatives of two C. dubliniensis strains repeatedly subculturedin the presence or absence of increasing concentrations of fluconazolea
C. dubliniensisderivatives
No. ofderivativesanalyzed
Fingerprintingmethod
No. of isolateswith the
parental pattern
No. of isolateswith a minor
pattern
No. ofminor
patternsBand differences in minor patternsb
CD36Flu70 20 Cd25 19 1 1 1(�1)Cd24 17 3 2 2(�3), 1(�1)Karyotype 10 10 6 1(�1), 1(�2,�2), 1(�2,�3), 4(�1,�1),
1(�1, �2), 2(�1)
CD36(control) 20 Cd25 20 0 0 NoneCd24 19 1 1 1(�2,�1)Karyotype 19 1 1 1(�1,�1)
CD514Flu50 20 Cd25 6 14 10 1(�1), 1(�1), 1(�1), 1(�1), 2(�1,�1), 2(�1,�1),3(�1), 1(�1,�1), 1(�1,�1), 1(�1)
Cd24 19 1 1 1(�1)Karyotype 19 1 1 1(�1)
CD51450(control) 20 Cd25 18 2 2 1(�1), 1(�1)Cd24 20 0 0 NoneKaryotype 20 0 0 None
CD514Flu70 13 Cd25 4 9 5 3(�1,�1), 1(�1), 1(�1,�3), 1(�2), 3(�2)Cd24 13 0 0 NoneKaryotype 13 0 0 None
CD51470(control) 13 Cd25 13 0 0 NoneCd24 13 0 0 NoneKaryotype 13 0 0 None
a Twenty fluconazole-resistant (MIC, 64 �g/ml) derivatives of the Cd25 group I C. dubliniensis strain CD36, which were obtained by repeated subculture of thefluconazole-susceptible (MIC, 0.5 �g/ml) parental isolate on agar medium containing increasing concentrations of drug (range, 0.5 to 70 �g/ml), were tested formicroevolution by hybridization analysis with the Cd25 probe and Cd24 probe and by karyotype analysis. All 20 resistant derivatives grew well on medium containing70 �g of fluconazole per ml and were termed CD36Flu70 derivatives. Twenty control derivatives which were simultaneously repeatedly subcultured on agar mediumwithout fluconazole were also tested; these were termed CD36(control) derivatives. Variability in the presence or absence of a chromosome-sized band at approximately1.65 Mb was observed in the karyotype patterns in both the fluconazole-resistant and the control derivatives, and therefore only variability in other chromosomal bandswas assessed for microevolution. In similar experiments with the Cd25 group II (genotype 3) C. dubliniensis isolate CD514, it proved more difficult to generatefluconazole-resistant derivatives. However, 20 derivatives that were recovered from agar plates containing 50 �g of fluconazole per ml (CD514Flu50 derivatives) and13 derivatives that were recovered from plates containing 70 �g of fluconazole per ml (CD514Flu70 derivatives) were studied. Control derivatives which weresimultaneously repeatedly subcultured on agar medium without fluconazole were also tested; these were termed CD51450(control) and CD51470(control) derivatives.
b See Table 4, footnote d.
570 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

and ITS2 regions, while the 5.8S rDNA regions were identical(Fig. 2a). These differences correlated exactly with the clustersidentified in the dendrogram generated from SAB values ob-tained from Cd25 fingerprinting. Genotype 1 and genotype 2isolates separate from each other at an SAB node value of 0.19,and the five genotype 3 isolates and the single genotype 4isolate separate from the genotype 2 isolates at an SAB node of0.3. The genotype 4 isolate separates from, the genotype 3isolates at an SAB node of 0.32.
The separation of C. dubliniensis into four genotypes basedon the ITS sequence data, together with the SAB node valuesfor the four genotypes, strongly suggests that C. dubliniensisconsists of at least four distinct populations. Indeed, it is alsoconceivable that analysis of a larger number of isolates could
reveal additional genotypes. Three distinct genotypes have alsobeen identified in C. parapsilosis using several methods, includ-ing ITS region sequence analysis and hybridization with therepeat sequence probe Cp3–13 (12, 22). However, there werefar more substantial sequence differences observed betweenthe C. parapsilosis genotypes (ranging from 82 to 88%) thanfound in the present study with C. dubliniensis (Fig. 2a). In C.parapsilosis, most differences were observed in ITS1, while the5.8S rDNA regions were identical and ITS2 showed minimaldifferences (22).
Karyotype analysis could not consistently distinguish thefour genotypes. Two major karyotype patterns were foundbased on the presence or absence of chromosome-sized bandsat approximately 1.7 and 1.75 Mb (Fig. 4). While all genotype2, 3, and 4 isolates analyzed had a band at 1.7 Mb, the majorityof genotype 1 isolates lacked this band but had a band at 1.75Mb. However, there were exceptions to this.
In order to determine if the genotype subgroupings of C.dubliniensis have any epidemiological significance, it will benecessary to be able to rapidly and accurately identify isolatesbelonging to each genotype in large collections of isolates.Differentiation of C. dubliniensis isolates into ITS genotypesfollowing nucleotide sequencing or into Cd25 groups usingfingerprint analysis is time-consuming and expensive. To sim-plify matters, a genotype-specific PCR assay was designedbased on sequence differences in the ITS region between the
FIG. 8. Karyotype and Cd25 hybridization patterns of isolates of C.dubliniensis exposed to fluconazole in vitro. (A) Karyotype profiles ofC. dubliniensis isolate CD36 and derivatives (CD36Flu70) exposed tofluconazole in vitro. Lane 1, C. dubliniensis parental isolate CD36;lanes 2 to 11, CD36Flu70 derivatives. The relative positions of molec-ular size reference markers (in megabases) are shown on the left of thepanel. (B) Karyotype profiles of C. dubliniensis isolate CD36 andCD36(control) derivatives repeatedly subcultured in the absence offluconazole. Lane 1, C. dubliniensis parental isolate CD36; lanes 2 to11, CD36(control) derivatives. The arrow indicates the position of achromosome-sized band of approximately 1.65 Mb which was presentin 11 of 20 of the CD36(control) derivatives tested, examples of whichare shown here in lanes 2, 4, 9, and 10. The relative positions ofmolecular size reference markers (in megabases) are shown on the leftof the panel. (C) Cd25 hybridization patterns of C. dubliniensis isolateCD514 and derivatives (CD514Flu50)exposed to fluconazole in vitro.Lane 1, C. dubliniensis parental isolate CD514; lanes 2 to 11,CD514Flu50 derivatives. An arrow indicates the position of polymor-phisms. The relative positions of molecular size reference markers (inkilobases) are shown on the left of the panel.
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 571
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

four genotypes (Fig. 2a and 3). Using this procedure, the ge-notype and hence the Cd25 fingerprint group of an isolatecould be determined in a matter of 5 h. At present we are alsoin the process of investigating whether there are any pheno-typic differences among the four genotypes which could facil-itate their identification in clinical samples.
Microevolution in vivo. Several previous studies with DNAfingerprinting probes demonstrated microevolution in clonalpopulations of Candida tropicalis (18), C. parapsilosis (12),Candida glabrata (24), C. albicans (23), and Cryptococcus neo-formans. (55). In the present study, the genetic homogeneity ofeight oral populations of C. dubliniensis isolates recoveredfrom the same clinical specimen from each of eight separatepatients was examined by fingerprint analysis using probesCd25, Cd24, and Cd1 and by karyotype analysis. Two of theisolate populations examined (from patients B and C, Table 1)exhibited identical fingerprint patterns, in each case, with allthree probes and by karyotype analysis. This suggests thatthese two patients harbored an identical genotype 3 strainwithout any detectable fingerprint differences. Of the other sixisolate populations, five exhibited fingerprint profile variationwith the Cd25 fingerprinting probe, six exhibited variation withthe probes Cd24 and Cd1, and five of them exhibited variationby karyotype analysis (Fig. 5 and 6, Table 4). These resultsdemonstrated that significant in vivo genetic variation, or mi-croevolution, occurred in these six isolate populations, as thethree fingerprinting probes hybridized to different regions ofthe genome (17) and karyotype analysis examines gross chro-mosomal organization. It has been suggested previously thatthe ability of C. dubliniensis to exhibit microevolution can re-sult in the generation of altered genotypes, some of which mayhave a selective advantage under rapidly changing environ-mental conditions (32).
The population of 17 clonal isolates from patient E (Table 1)was found to consist of both fluconazole-susceptible (13 iso-lates; MIC, 1 �g/ml) isolates and isolates with reduced sus-ceptibility to fluconazole (4 isolates; MIC, 16 �g/ml). Theother six isolate populations from patients A to D and F to H(Table 1) were all found to be susceptible to fluconazole.Interestingly, the four isolates from patient E which exhibitedreduced susceptibility to fluconazole all yielded variable fin-gerprint profiles with the Cd24 and Cd1 probes and lacked achromosome-sized band of approximately 0.85 Mb comparedto the fluconazole-susceptible isolates. Only 1 of the 13 flucon-azole-susceptible isolates from patient E showed variable Cd24and Cd1 fingerprints and a similar karyotype profile to theisolates with reduced susceptibility to fluconazole. Similar find-ings were reported by White et al. (61), who used the Ca3fingerprinting probe to analyze a collection of serial clonalisolates of C. albicans from an HIV-infected patient and ob-served microevolution that was associated with the acquisitionof fluconazole resistance.
In the present study, clonal isolates of C. dubliniensis recov-ered from serial samples from different anatomical locations,including blood, in two additional patients were also examinedfor microevolution by fingerprint analysis with the Cd25, Cd24,and Cd1 probes and by karyotype analysis. In the case of thefirst patient, minor polymorphisms indicative of microevolu-tion were detected with all three probes and by karyotypeanalysis. In the case of the second patient, minor polymor-
phisms were detected with the Cd25 and Cd1 probes and bykaryotype analysis.
All of these results confirmed that microevolution occursfrequently in clonal populations of C. dubliniensis in vivo andthat in some circumstances genetic variation may be associatedwith phenotypic changes, such as susceptibility to fluconazole.
Microevolution in vitro. It has been shown previously thatexposure of C. dubliniensis isolates to fluconazole in vitro rap-idly leads to the development of decreased susceptibility to thisdrug (32). Furthermore, it has also been noted that in somecases the development of fluconazole resistance is associatedwith minor changes in karyotype pattern (32). In the presentstudy, we investigated the effect of exposure to fluconazole onmicroevolution in the genotype 1 C. dubliniensis isolate CD36and the genotype 3 isolate CD514. One hundred colonies ofeach strain were subcultured (i) on agar plates containingincreasing concentrations of fluconazole in vitro and (ii) oncontrol plates without drug. The colonies of CD36 readilydeveloped resistance to fluconazole upon exposure, whereasthe colonies of CD514 grew very poorly on drug-containingmedium and did not develop resistance as easily.
Derivatives of both strains grown on drug-free medium andon medium containing either 50 or 70 �g of fluconazole per mlwere fingerprinted with the probes Cd25, Cd24, and Cd1,which were used as markers for independent genetic variationbecause they have no known relationship to drug resistance,and also by karyotype analysis. Derivatives of CD514 grown onmedium containing either 50 or 70 �g of fluconazole per ml(CD514Flu50 and CD514Flu70, respectively) did not developresistance to fluconazole very readily; however, they showed ahigh level of variation with the probe Cd25 (14 of 20 of theCD514Flu50 and 9 of 13 of the CD514Flu70 derivatives showedvariants), but very little with Cd24 (one variant) or Cd1 (novariants). The CD36Flu70 derivatives had decreased suscepti-bility to fluconazole but did not exhibit a significant degree ofvariation with the probe Cd25 (one variant).
It is interesting to speculate that the variation observed in theCd25 fingerprints of the CD514Flu50 and CD514Flu70 derivativesoccurred as a direct response of the cells to the environmentalstress of exposure to fluconazole. However, despite this variation,it would appear that, unlike the CD36Flu70 derivatives, derivativesof CD514 could not develop drug resistance as readily. With theexception of one of the chromosomes, possibly chromosome R,there was only one chromosomal change detected in the karyo-types of CD36(control) derivatives not exposed to drug. Incontrast, in 10 of 20 of the CD36Flu70 derivatives tested, karyo-type changes were observed. None of the CD514 control de-rivatives or the CD514Flu70 derivatives and only one of theCD514Flu50 derivatives showed karyotype variation (Table 5).
The results described above show that the response of C.dubliniensis to fluconazole exposure in vitro is accompanied byan increased frequency of genomic variation but that the extentand manifestation of this variation may be strain or genotypedependent. This variation is associated with changes in repet-itive sequence elements (detected with repetitive sequence-containing probes) and in chromosomal rearrangements (de-tected by karyotype variation). It has been suggested that C.albicans cells undergoing changes at repetitive sequence loci atunusually high frequency may also be undergoing parallel high-
572 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

frequency changes at other genetic loci directly involved indrug susceptibility (52).
Whether there is a direct link between the ability of C.dubliniensis to generate fluconazole-resistant derivatives andits ability to undergo genomic microevolution is not clear.However, the acquisition of phenotypic traits in tandem withchromosomal variation has been reported previously in C. al-bicans and C. glabrata. Chromosomal rearrangements and de-letions have been associated with phenotypic switching in C.albicans (43). Moreover, the fluconazole resistance phenotypeof a C. glabrata isolate has been linked to the duplication of thechromosome carrying the ERG11 gene, encoding the flucon-azole target enzyme (27). This duplication resulted in the al-tered expression of approximately 100 proteins, including theoverexpression of ERG11 (27). Furthermore, Perepnikhatakaet al. (36) demonstrated that exposure of C. albicans to flu-conazole resulted in the nondisjuntion of two specific chromo-somes in drug-resistant mutants, each obtained by an indepen-dent mutational event. This finding establishes a role forchromosomal changes as a possible general response of C.albicans cells in the development of drug resistance.
In contrast, in the present study, the C. dubliniensis CD36derivatives obtained following exposure to fluconazole exhib-ited increased karyotype variability, but this showed no consis-tent association with drug resistance. However, of the 15 clonalisolates of C. dubliniensis CD527A recovered from the sameclinical specimen from patient E (Table 1), 5 lacked a chro-mosome-sized band of approximately 0.85 Mb, and 4 of thesevariants exhibited reduced susceptibility to fluconazole. Cowenet al. also showed no consistent association between flucon-azole resistance and alterations in DNA fingerprints andkaryotype profiles in experimental populations of drug-resis-tant C. albicans (9). Repetitive sequence (RPS)-containing el-ements have been implicated in the variability of the size andnumber of chromosomes in C. albicans (5, 6, 7, 15). C. dub-liniensis contains sequences homologous to RPS dispersedthroughout its genome (17), and it is possible that these ele-ments may be responsible, at least in part, for the observed vari-ability in the karyotypes of isolates, both in vivo and in vitro.
The diverse range of geographical and clinical sources of theC. dubliniensis isolates described in this paper is further evi-dence of the prevalence of this species throughout the world.However, a definitive wide-ranging study is required to deter-mine the true epidemiological significance of C. dubliniensis.Now, with the availability of effective DNA fingerprintingprobes and a rapid genotype identification PCR test, we hopeto be able to initiate a comprehensive analysis of the popula-tion dynamics of this novel species.
ACKNOWLEDGMENTS
This work was supported by Irish Health Research Board grants41/96 and 04/99 to D.C.C. and NIH grant A12392 to D.R.S.
REFERENCES
1. Al Mosaid, A., D. Sullivan, I. F. Salkin, D. Shanley, and D. C. Coleman.2001. Differentiation of Candida dubliniensis from Candida albicans on Staibagar and caffeic acid-ferric citrate agar. J. Clin. Microbiol. 39:323–327.
2. Backeljau, T., L. De Bruyn, H. DeWolf, K. Jordaens, S. Van Dongen, and B.Winnepenninckx. 1996. Multiple UPGMA and neighbor-joining trees andthe performance of some computer packages. Mol. Biol. Evol. 13:309–313.
3. Boerlin, P., F. Boerlin-Petzold, C. Durussel, M. Addo, J.-L. Pagani, J.-P.Chave, and J. Bille. 1995. Cluster of oral atypical Candida albicans isolates
in a group of human immunodeficiency virus-positive drug users. J. Clin.Microbiol. 33:1129–1135.
4. Brandt, M. E., L. H. Harrison, M. Pass, A. N. Sofair, S. Huie, R.-K. Li, C. J.Morrison, D. W. Warnock, and R. H. Hajjeh. 2000. Candida dubliniensisfungemia: the first four cases in North America. Emerg. Infect. Dis. 6:46–49.
5. Chibana, H., S. Iwaguchi, M. Homma, A. Chindamporn, Y. Nakagawa, andK. Tanaka. 1994. Diversity of tandemly repetitive sequences due to shortperiodic repetitions in the chromosomes of Candida albicans. J. Bacteriol.176:3851–3858.
6. Chindamporn, A., Y. Nakagawa, M. Homma, H. Chibana, M. Doi, and K.Tanaka. 1995. Analysis of the chromosomal localization of the repetitivesequences (RPSs) in Candida albicans. Microbiology 141:469–476.
7. Chindamporn, A., Y. Nakagawa, I. Mizuguchi, H. Chibana, M. Doi, and K.Tanaka. 1998. Repetitive sequences (RPSs) in the chromosomes of Candidaalbicans are sandwiched between two novel stretches, HOK and RB2, com-mon to each chromosome. Microbiology 144:849–857.
8. Coleman, D., D. Sullivan, B. Harrington, K. Haynes, M. Henman, D. Shan-ley, D. Bennett, G. Moran, C. McCreary, and L. O’Neill. 1997. Molecular andphenotypic analysis of Candida dubliniensis: a recently identified specieslinked with oral candidosis in HIV-infected and AIDS patients. Oral Dis.3(Suppl. 1):S96–S101.
9. Cowen, L. E., D. Sanglard, D. Calabrese, C. Sirjusingh, J. B. Anderson, andL. M. Kohn. 2000. Evolution of drug resistance in experimental populationsof Candida albicans. J. Bacteriol. 182:1515–1522.
10. Donnelly, S. M., D. J. Sullivan, D. B. Shanley, and D. C. Coleman. 1999.Phylogenetic analysis and rapid identification of Candida dubliniensis basedon analysis of ACT1 intron and exon sequences. Microbiology 145:1871–1882.
11. Elie, C. M., T. J. Lott, E. Reiss, and C. J. Morrison. 1998. Rapid identifi-cation of Candida species with species-specific DNA probes. J. Clin. Micro-biol. 36:3260–3265.
12. Enger, L., S. Joly, C. Pujol, P. Simonson, M. Pfaller, and D. R. Soll. 2001.Cloning and characterization of a complex DNA fingerprinting probe forCandida parapsilosis. J. Clin. Microbiol. 39:658–669.
13. Fell, J. 1993. Rapid identification of yeast species using three primers in apolymerase chain reaction. Mol. Mar. Biol. Biotechnol. 3:174–180.
14. Felsenstein, J. 1985. Confidence limits on phylogenies: an approach usingthe bootstrap. Evolution 39:783–791.
15. Iwaguchi, S.-I., M. Homma, H. Chibana, and K. Tanaka. 1992. Isolation andcharacterization of a repeated sequence (RPS1) of Candida albicans. J. Gen.Microbiol. 138:1893–1900.
16. Jabra-Rizk, M. A., A. A. Baqui, J. I. Kelley, W. A. Falkler, W. G. Merz, andT. F. Meiller. 1999. Identification of Candida dubliniensis in a prospectivestudy of patients in the United States. J. Clin. Microbiol. 37:321–326.
17. Joly, S., C. Pujol, M. Rysz, K. Vargas, and D. R. Soll. 1999. Development andcharacterization of complex DNA fingerprinting probes for the infectiousyeast Candida dubliniensis. J. Clin. Microbiol. 37:1035–1044.
18. Joly, S., C. Pujol, K. Schröppel, and D. R. Soll. 1996. Development of twospecies-specific fingerprinting probes for broad computer-assisted epidemi-ological studies of Candida tropicalis. J. Clin. Microbiol. 34:3063–3071.
19. Kamei, K., M. J. McCullough, and D. A. Stevens. 2000. Initial case ofCandida dubliniensis infection from Asia: nonmucosal infection. Med. My-col. 38:81–83.
20. Kurzai, O., W. J. Heinz, D. J. Sullivan, D. C. Coleman, M. Frosch, and F. A.Mühlschlegel. 1999. Rapid PCR test for discriminating between Candidaalbicans and Candida dubliniensis isolates using primers derived from thepH-regulated PHR1 and PHR2 genes of C. albicans. J. Clin. Microbiol.37:1587–1590.
21. Lehmann, P. F., D. Lin, and B. A. Lasker. 1992. Genotypic identification andcharacterization of species and strains within the genus Candida by usingrandom amplified polymorphic DNA. J. Clin. Microbiol. 30:3249–3254.
22. Lin, D., L.-C. Wu, M. G. Rinaldi, and P. F. Lehmann. 1995. Three distinctgenotypes within Candida parapsilosis from clinical sources. J. Clin. Micro-biol. 33:1815–1821.
23. Lockhart, S. R., J. J. Fritch, A. S. Meier, K. Schröppel, S. Srikantha, R.Galask, and D. R. Soll. 1995. Colonizing populations of Candida albicans areclonal in origin but undergo microevolution through C1 fragment reorgani-zation as demonstrated by DNA fingerprinting and C1 sequencing. J. Clin.Microbiol. 33:1501–1509.
24. Lockhart, S. R., S. Joly, C. Pujol, J. D. Sobel, M. A. Pfaller, and D. R. Soll.1997. Development and verification of fingerprinting probes for Candidaglabrata. Microbiology 143:3733–3746.
25. Lott, T. J., D. A. Cogan, B. P. Holloway, R. Fundyga, and J. Arnold. 1999.Towards understanding the evolution of the human commensal yeast Can-dida albicans. Microbiology 145:1137–1143.
26. Lott, T. J., R. J. Kuykendall, S. F. Welbel, A. Pramanik, and B. A. Lasker.1993. Genetic heterogeneity in the yeast Candida parapsilosis. Curr. Genet.23:463–467.
27. Marichal, P., H. Vanden Bossche, F. C. Odds, G. Nobels, D. W. Warnock, V.Timmerman, C. Van Broeckhoven, S. Fay, and P. Mose-Larsen. 1997. Mo-lecular biological characterization of an azole-resistant Candida glabrataisolate. Antimicrob. Agents Chemother. 41:2229–2237.
VOL. 40, 2002 C. DUBLINIENSIS GENOTYPES 573
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from

28. Marriott, D., M. Laxton, and J. Harkness. 2001. Candida dubliniensis can-didemia in Australia. Emerg. Infect. Dis. 7:479.
29. McCullough, M., B. Ross, and P. Reade. 1995. Characterization of geneti-cally distinct subgroup of Candida albicans strains isolated from oral cavitiesof patients infected with human immunodeficiency virus. J. Clin. Microbiol.33:696–700.
30. Meiller, T. F., M. A. Jabra-Rizk, A. A. Baqui, J. I. Kelley, V. I. Meeks, W. G.Merz, and W. A. Falkler. 1999. Oral Candida dubliniensis as a clinicallyimportant species in HIV-seropositive patients in the United States. OralSurg. Oral Med. Oral Pathol. Oral Radiol. Endod. 88:573–580.
31. Meis, J. F., M. Ruhnke, B. E. De Pauw, F. C. Odds, W. Siegert, and P. E.Verweij. 1999. Candida dubliniensis candidemia in patients with chemother-apy-induced neutropenia and bone marrow transplantation. Emerg. Infect.Dis. 5:150–153.
32. Moran, G. P., D. J. Sullivan, M. C. Henman, C. E. McCreary, B. J. Har-rington, D. B. Shanley, and D. C. Coleman. 1997. Antifungal drug suscep-tibilities of oral Candida dubliniensis isolates from human immunodeficiencyvirus (HIV)-infected and non-HIV-infected subjects and generation of stablefluconazole-resistant derivatives in vitro. Antimicrob. Agents Chemother.41:617–623.
33. National Committee for Clinical Laboratory Standards. 1992. Referencemethod for broth dilution antifungal susceptibility testing for yeasts. Pro-posed Standard M27-P. National Committee for Clinical Laboratory Stan-dards, Villanova, Pa.
34. Odds, F. C., and R. Bernaerts. 1994. CHROMagar Candida, a new differ-ential isolation medium for presumptive identification of clinically importantCandida species. J. Clin. Microbiol. 32:1923–1929.
35. Park, S., M. Wong, S. A. E. Marras, E. W. Cross, T. E. Kiehn, V. Chaturvedi,S. Tyagi, and D. S. Perlin. 2000. Rapid identification of Candida dubliniensisusing a species-specific molecular beacon. J. Clin. Microbiol. 38:2829–2836.
36. Perepnikhatka, F., F. J. Fischer, M. Niimi, R. A. Baker, R. C. Cannon, Y.-K.Wang, F. Sherman, and E. Rustchenko. 1999. Specific chromosome alter-ations in fluconazole-resistant mutants of Candida albicans. J. Bacteriol.181:4041–4049.
37. Pfaller, M. A., S. R. Lockhart, C. Pujol, J. A. Swails-Wenger, S. A. Messer,M. B. Edmond, R. N. Jones, R. P. Wenzel, and D. R. Soll. 1998. Hospitalspecificity, region specificity, and fluconazole resistance of Candida albicansbloodstream isolates. J. Clin. Microbiol. 36:1518–1529.
38. Pincus, D. H., D. C. Coleman, W. R. Pruitt, A. A. Padhye, I. F. Salkin, M.Geimer, A. Bassel, D. J. Sullivan, M. Clarke, and V. Hearn. 1999. Rapididentification of Candida dubliniensis with commercial yeast identificationsystems. J. Clin. Microbiol. 37:3533–3539.
39. Pinjon, E., D. Sullivan, I. Salkin, D. Shanley, and D. Coleman. 1998. Simple,inexpensive, reliable method for differentiation of Candida dubliniensis fromCandida albicans. J. Clin. Microbiol. 36:2093–2095.
40. Polacheck, I., J. Strahilevitz, D. Sullivan, S. Donnelly, I. F. Salkin, and D. C.Coleman. 2000. Recovery of Candida dubliniensis from nonhuman immuno-deficiency virus-infected patients in Israel. J. Clin. Microbiol. 38:170–174.
41. Pujol, C., S. Joly, S. R. Lockhart, S. Noel, M. Tibayrenc, and D. R. Soll. 1997.Parity among the randomly amplified polymorphic DNA method, multilocusenzyme electrophoresis, and Southern blot hybridization with the moder-ately repetitive probe Ca3 for fingerprinting Candida albicans. J. Clin. Mi-crobiol. 35:3248–3258.
42. Quindos, G., A. J. Carrillo-Munoz, M. P. Arevalo, J. Salgado, R. Alonso-Vargas, J. M. Rodrigo, M. T. Ruesga, A. Valverde, J. Peman, E. Canton, E.Martin-Mazuelos, and J. Ponton. 2000. In vitro susceptibility of Candidadubliniensis to current and new antifungal agents. Chemotherapy 46:395–401.
43. Ramsey, R., B. Morrow, and D. R. Soll. 1994. An increase in switchingfrequency correlates with an increase in recombination of the ribosomalchromosomes of Candida albicans strain 3153A. Microbiology 140:1525–1531.
44. Rodriguez-Tudela, J. L., and J. V. Martinez-Suárez. 1995. Defining condi-tions for microbroth antifungal susceptibility tests: influence of RPMI andRPMI-2% glucose on the selection of endpoint criteria. J. Antimicrob. Che-mother. 35:739–749.
45. Saitou, N., and M. Nei. 1987. The neighbour joining method for reconstruct-ing phylogenetic trees. Mol. Biol. Evol. 4:406–425.
46. Salesa, R., M. D. Moragues, R. Sota, J. Peman, G. Quindos, and J. Ponton.2001. Specific antibody response in a patient with Candida dubliniensis fun-gemia. Rev. Iberoam. Micol. 18:42–44.
47. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratorymanual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,N.Y.
48. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencing withchain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467.
49. Scherer, S., and D. A. Stevens. 1987. Application of DNA typing methods toepidemiology and taxonomy of Candida species. J. Clin. Microbiol. 25:675–679.
50. Scherer, S., and D. A. Stevens. 1988. A Candida albicans dispersed, repeatedgene family and its epidemiologic applications. Proc. Natl. Acad. Sci. USA85:1452–1456.
51. Schmid, J., E. Voss, and D. R. Soll. 1990. Computer-assisted methods forassessing strain relatedness in Candida albicans by fingerprinting with themoderately repetitive sequence Ca3. J. Clin. Microbiol. 28:1236–1243.
52. Soll, D. R. 2000. The ins and outs of DNA fingerprinting the infectious fungi.Clin. Microbiol. Rev. 13:332–370.
53. Staib, P., and J. Morschhäuser. 1999. Chlamydospore formation on Staibagar as a species-specific characteristic of Candida dubliniensis. Mycoses42:521–524.
54. Sullivan, D., K. Haynes, J. Bille, P. Boerlin, L. Rodero, S. Lloyd, M. Hen-man, and D. Coleman. 1997. Widespread geographic distribution of oralCandida dubliniensis strains in human immunodeficiency virus-infected indi-viduals. J. Clin. Microbiol. 35:960–964.
55. Sullivan, D., K. Haynes, G. Moran, D. Shanley, and D. Coleman. 1996.Persistence, replacement, and microevolution of Cryptococcus neoformansstrains in recurrent meningitis in AIDS patients. J. Clin. Microbiol. 34:1739–1744.
56. Sullivan, D. J., G. Moran, S. Donnelly, S. Gee, E. Pinjon, B. McCartan, D. B.Shanley, and D. C. Coleman. 1999. Candida dubliniensis: an update. Rev.Iberoam. Micol. 16:72–76.
57. Sullivan, D. J., T. J. Westerneng, K. A. Haynes, D. E. Bennett, and D. C.Coleman. 1995. Candida dubliniensis sp. nov.: phenotypic and molecularcharacterisation of a novel species associated with oral candidosis in HIV-infected individuals. Microbiology 141:1507–1521.
58. Tamura, M., K. Watanabe, T. Imai, Y. Mikami, and K. Nishimura. 2000.New PCR primer pairs specific for Candida dubliniensis and detection of thefungi from the Candida albicans clinical isolates in Japan. Clin. Lab. 46:33–40.
59. Thompson, J. D., D. G. Higgins, and T. J. Gibson. 1994. CLUSTAL W:improving the sensitivity of progressive multiple sequence alignment throughsequence weighting, position-specific gap penalties and weight matrix choice.Nucleic Acids Res. 22:4673–4680.
60. Vazquez, J. A., A. Beckley, J. D. Sobel, and M. J. Zervos. 1991. Comparisonof restriction enzyme analysis and pulsed-field gradient gel electrophoresis astyping systems for Candida albicans. J. Clin. Microbiol. 29:962–967.
61. White, T. C., M. A. Pfaller, M. G. Rinaldi, J. Smith, and S. W. Redding. 1997.Stable azole drug resistance associated with a substrain of Candida albicansfrom an HIV-infected patient. Oral Dis. 3(Suppl. 1):S102–S109.
62. White, T. J., T. D. Bruns, S. B. Lee, and J. W. Taylor. 1990. Amplificationand direct sequencing of fungal ribosomal RNA genes for phylogenetics, p.315–322. In M. A. Innis, D. H. Gelfand, J. J. Sininsky, and T. J. White (ed.),PCR protocols: a guide to methods and applications. Academic Press, Inc.,San Diego, Calif.
574 GEE ET AL. J. CLIN. MICROBIOL.
on February 21, 2016 by guest
http://jcm.asm
.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















