
Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride...
7
Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride by HS-SPME-GC-MS Norbert Stoppacher a , Bernhard Kluger a , Susanne Zeilinger b , Rudolf Krska a , Rainer Schuhmacher a, ⁎ a Center for Analytical Chemistry, Department for Agrobiotechnology (IFA-Tulln), University of Natural Resources and Applied Life Sciences, Vienna, Konrad Lorenz Strasse 20, A-3430 Tulln, Austria b Institute of Chemical Engineering, Research Area of Gene Technology and Applied Biochemistry, Vienna University of Technology, Getreidemarkt 9/166-5, A-1060 Wien, Austria abstract article info Article history: Received 30 December 2009 Received in revised form 10 March 2010 Accepted 10 March 2010 Available online 16 March 2010 Keywords: Gas chromatography–mass spectrometry Headspace Trichoderma atroviride Microbial volatile organic compounds Fusaric acid Solid phase microextraction In the present study we describe a method, which is based on solid phase microextraction (SPME) coupled to gas chromatography–mass spectrometry (GC-MS) and which can be used for the profiling of microbial volatile organic compounds (MVOCs) in the headspace (HS) of cultures of filamentous fungi. The method comprises the following successive steps: 1. growth of the fungus on a solid culture medium directly in headspace vials, 2. measurement of volatiles by HS-SPME-GC-MS, 3. deconvolution of mass spectra, 4. identification of volatiles by comparison of measured, deconvoluted mass spectra and linear temperature programmed retention indices (LTPRI) on two stationary GC phases with database entries and LTPRI published in the literature, and 5. profiling of the identified MVOCs. The developed method was successfully applied to cultures of the biocontrol fungus Trichoderma atroviride. An in-house library consisting of mass spectra and LTPRI values of fungal VOCs was established and used to study the profiles of MVOCs of this fungus. In total, 25 different MVOCs were identified by applying strict criteria (spectral match factor at least 90% and a maximum relative deviation of LTPRI of ± 2% from literature values). The MVOCs were assigned to the compound classes of alcohols, ketones, alkanes, furanes, pyrones (mainly the bioactive 6-pentyl-alpha-pyrone), mono- and sesquiterpenes, 13 of which have never been reported to be produced by Trichoderma spp. before. Eleven of these volatiles have been additionally confirmed using authentic standards. Finally, time course experiments and cultivation of T. atroviride in the presence of the mycotoxin fusaric acid demonstrated the potential of the method to study the dynamics of MVOC profiles as well as the effect of different environmental/biological conditions on the expression of MVOCs of filamentous fungi. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Microbial volatile organic compounds (MVOCs) appear as inter- mediate and end products of various metabolic pathways and belong to numerous structure classes such as mono- and sesquiterpenes, alcohols, ketones, lactones, esters or C 8 compounds (e.g. Schnürer et al., 1999; Korpi et al., 2009). These metabolites have been shown to be involved in different biological processes such as biocontrol or communication between microorganisms and their living environ- ment. They can mediate defence against predators, parasites and diseases, and may be produced for competition between species and to facilitate reproductive processes (Sivasithamparam and Ghisal- berti, 1998). MVOCs of the filamentous biocontrol fungus Tricho- derma, for example, act antibiotically against plant pathogenic moulds and can confer plant growth promoting effects (Vinale et al., 2008) as well as systemic resistance to plants, thus rendering plants less vulnerable to fungal pathogens (Harman et al., 2004). Determination of volatile fungal metabolites usually is done by gas chromatographic (GC) methods and has been described for different fungi such as Aspergillus, Fusarium, Mucor, Penicillium and Trichoderma. After cultivation of the fungi in liquid (Pinches and Apps, 2007) or on solid growth medium (Nemčovič et al., 2008) volatiles can be extracted in different ways, e.g. with organic solvents (Zeppa et al., 1990; Reithner et al., 2005; Claydon et al., 1987), solid phase extraction using C 18 or silica gel columns (Keszler et al., 2000), online gas enrichment on adsorption tubes (Wheatley et al., 1997) or various headspace techniques: e.g. closed loop stripping analysis (Meruva et al., 2004), dynamic headspace (purge and trap) (Deetae et al., 2007) and solid phase microextraction (Fiedler et al., 2001; Demyttenaere et al., 2004; Van Lancker et al., 2008). The latter technique can be automated and used for the direct and non-invasive extraction of volatiles above living fungal cultures. It therefore offers the potential of monitoring fungal VOC profiles over time and for different growth conditions. After extraction and GC separation on non-polar to medium-polar stationary phases, the constituents of complex mixtures of MVOCs can Journal of Microbiological Methods 81 (2010) 187–193 ⁎ Corresponding author. Tel.: + 43 2272 66280 407; fax: + 43 2272 66280 403. E-mail address: [email protected] (R. Schuhmacher). 0167-7012/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.mimet.2010.03.011 Contents lists available at ScienceDirect Journal of Microbiological Methods journal homepage: www.elsevier.com/locate/jmicmeth
Transcript of Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride...

Journal of Microbiological Methods 81 (2010) 187–193
Contents lists available at ScienceDirect
Journal of Microbiological Methods
j ourna l homepage: www.e lsev ie r.com/ locate / jmicmeth
Identification and profiling of volatile metabolites of the biocontrol fungusTrichoderma atroviride by HS-SPME-GC-MS
Norbert Stoppacher a, Bernhard Kluger a, Susanne Zeilinger b, Rudolf Krska a, Rainer Schuhmacher a,⁎a Center for Analytical Chemistry, Department for Agrobiotechnology (IFA-Tulln), University of Natural Resources and Applied Life Sciences, Vienna, Konrad Lorenz Strasse 20,A-3430 Tulln, Austriab Institute of Chemical Engineering, Research Area of Gene Technology and Applied Biochemistry, Vienna University of Technology, Getreidemarkt 9/166-5, A-1060 Wien, Austria
⁎ Corresponding author. Tel.: +43 2272 66280 407;E-mail address: [email protected] (R.
0167-7012/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.mimet.2010.03.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 30 December 2009Received in revised form 10 March 2010Accepted 10 March 2010Available online 16 March 2010
Keywords:Gas chromatography–mass spectrometryHeadspaceTrichoderma atrovirideMicrobial volatile organic compoundsFusaric acidSolid phase microextraction
In the present study we describe a method, which is based on solid phase microextraction (SPME) coupled togas chromatography–mass spectrometry (GC-MS) and which can be used for the profiling of microbialvolatile organic compounds (MVOCs) in the headspace (HS) of cultures of filamentous fungi. The methodcomprises the following successive steps: 1. growth of the fungus on a solid culture medium directly inheadspace vials, 2. measurement of volatiles by HS-SPME-GC-MS, 3. deconvolution of mass spectra, 4.identification of volatiles by comparison of measured, deconvoluted mass spectra and linear temperatureprogrammed retention indices (LTPRI) on two stationary GC phases with database entries and LTPRIpublished in the literature, and 5. profiling of the identified MVOCs.The developed method was successfully applied to cultures of the biocontrol fungus Trichoderma atroviride.An in-house library consisting of mass spectra and LTPRI values of fungal VOCs was established and used tostudy the profiles of MVOCs of this fungus. In total, 25 different MVOCs were identified by applying strictcriteria (spectral match factor at least 90% and a maximum relative deviation of LTPRI of ±2% from literaturevalues). The MVOCs were assigned to the compound classes of alcohols, ketones, alkanes, furanes, pyrones(mainly the bioactive 6-pentyl-alpha-pyrone), mono- and sesquiterpenes, 13 of which have never beenreported to be produced by Trichoderma spp. before. Eleven of these volatiles have been additionallyconfirmed using authentic standards. Finally, time course experiments and cultivation of T. atroviride in thepresence of the mycotoxin fusaric acid demonstrated the potential of the method to study the dynamics ofMVOC profiles as well as the effect of different environmental/biological conditions on the expression ofMVOCs of filamentous fungi.
fax: +43 2272 66280 403.Schuhmacher).
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Microbial volatile organic compounds (MVOCs) appear as inter-mediate and end products of various metabolic pathways and belongto numerous structure classes such as mono- and sesquiterpenes,alcohols, ketones, lactones, esters or C8 compounds (e.g. Schnürer etal., 1999; Korpi et al., 2009). Thesemetabolites have been shown to beinvolved in different biological processes such as biocontrol orcommunication between microorganisms and their living environ-ment. They can mediate defence against predators, parasites anddiseases, and may be produced for competition between species andto facilitate reproductive processes (Sivasithamparam and Ghisal-berti, 1998). MVOCs of the filamentous biocontrol fungus Tricho-derma, for example, act antibiotically against plant pathogenic mouldsand can confer plant growth promoting effects (Vinale et al., 2008) as
well as systemic resistance to plants, thus rendering plants lessvulnerable to fungal pathogens (Harman et al., 2004).
Determination of volatile fungal metabolites usually is done by gaschromatographic (GC) methods and has been described for differentfungi such as Aspergillus, Fusarium, Mucor, Penicillium and Trichoderma.After cultivation of the fungi in liquid (Pinches and Apps, 2007) or onsolid growthmedium (Nemčovič et al., 2008) volatiles can be extractedin differentways, e.g. with organic solvents (Zeppa et al., 1990; Reithneret al., 2005; Claydon et al., 1987), solid phase extraction using C18 orsilica gel columns (Keszler et al., 2000), online gas enrichment onadsorption tubes (Wheatley et al., 1997) or various headspacetechniques: e.g. closed loop stripping analysis (Meruva et al., 2004),dynamic headspace (purge and trap) (Deetae et al., 2007) and solidphase microextraction (Fiedler et al., 2001; Demyttenaere et al., 2004;Van Lancker et al., 2008). The latter technique can be automated andused for the direct and non-invasive extraction of volatiles above livingfungal cultures. It therefore offers the potential of monitoring fungalVOC profiles over time and for different growth conditions.
After extraction and GC separation on non-polar to medium-polarstationary phases, the constituents of complexmixtures of MVOCs can

188 N. Stoppacher et al. / Journal of Microbiological Methods 81 (2010) 187–193
be detected by flame ionisation detection (FID) (Elke et al., 1999) or –nowadays most widely used – mass spectrometry (MS) (Hynes et al.,2007). Mass spectrometric detection offers the possibility to identifyindividual volatiles from complex mixtures. Structure characterisa-tion and confirmation of identity is usually achieved by comparison ofmass spectra with library spectra, the determination of chromato-graphic retention indices and/or the determination of authenticstandards in parallel (Jeleń, 2003; Ruther, 2000; Oprean et al., 1998;Oprean et al., 2001).
In the present study, Trichoderma atroviride ATCC 74058 (alsoknown as P1) served as model organism to investigate the profiles ofvolatile fungal substances. Filamentous soil fungi of this genusfrequently live in association with plant roots. Some species ofTrichoderma are able to parasitize plant pathogenic moulds such asBotrytis cinerea or Rhizoctonia solani (mycoparasitism) (Howell, 2003)and to confer increased growth and systemic resistance to plants(Harman et al., 2004; Brunner et al., 2005). Owing to these beneficialeffects, some strains of e.g. T. harzianum, T. atroviride and T. asperellumare used as plant protection agents for the biocontrol of moulds and asplant growth promoters in agriculture, fruit growing and vegetablegardening (Harman et al., 2004; Harman, 2006; Verma et al., 2007).The capability of Trichoderma spp. to produce a great number ofvolatile (e.g. pyrones, sesquiterpenes) and non-volatile secondarymetabolites (e.g. peptaibols) has been reviewed recently (Reino et al.,2008). Volatile secondary metabolites have been demonstrated toplay a key role in mycoparasitism of Trichoderma as well as in itsinteraction with plants (e.g. Vinale et al., 2008). Surprisingly,however, only few studies on MVOC profiles of Trichoderma havebeen published which addressed the effect of different culture mediaon the type of volatiles produced by Trichoderma (e.g. Wheatley et al.,1997) or the biological function of selected volatiles of this fungus(Nemčovič et al., 2008).
Here, we present the development of an automated HS-SPME GC-MS based analytical method for the direct profiling of living fungalcultures and its application to the biocontrol fungus T. atroviride Thedeveloped method has been demonstrated to be a powerful tool tostudy the dynamics as well as the influence of the Fusariummycotoxinfusaric acid on the MVOC profile of this fungus.
2. Material and methods
2.1. Chemicals and standards
n-Hexane (95% purity) was obtained from J.T. Baker (Deventer,The Netherlands) and potato-dextrose agar (PDA) was purchasedfrom Merck (Darmstadt, Germany). 2-Heptanone (99%), 3-octanone(N98%), 3-octanol (95%), 1-octen-3-ol (98%), 2-nonanone (99%), 2-undecanone (99%), trans-nerolidol (85%), alpha-terpinene (95%),phenylethyl alcohol (99%), 6-pentyl-alpha-pyrone (96%), alpha-phellandrene, fusaric acid (99%) and the alkane standard solution(mixture of C8–C20 in hexane) were bought from Sigma-Aldrich(Vienna, Austria).
2.2. Cultivation of T. atroviride
T. atroviride strain ATCC 74058 (also known as P1) was used forthis study and grown on potato-dextrose agar PDA at 28 °C for 7 days.From the fungal mycelium on culture plates, discs, 5 mm in diameter,were placed to the centre of slants consisting of 5 mL of PDA in 20-mLheadspace (HS) vials. The vials were closed with sterile cotton plugsand rubber straps for aerobic cultivation without oxygen limitation.The vials were incubated in the absence of light at 22 °C for definedtime periods up to 5 days.
For investigating the influence of fusaric acid on the profile ofMVOCs produced by T. atroviride, a stock solution of 1 g/L fusaric acidwas prepared in doubly distilled water and filtered through a 0.2 µm
PES membrane filter. An aliquot of the sterile stock solution wasadded to liquefied PDA medium to give a final concentration of100 mg/L before pipetting 5 ml PDA into HS vials.
2.3. Analysis of standards and blanks
For confirmation of the identified volatiles, standard solutions of 2-heptanone, 1-octen-3-ol, 3-octanone, 3-octanol, alpha-terpinene, 2-nonanone, 2-undecanone, nerolidol, alpha-phellandrene, phenylethylalcohol and 6-pentyl-alpha-pyrone (6-PP) were mixed and dilutedwith hexane to concentrations between 0.05 and 5000 mg/L persubstance. 20 µl of the standard solutions were pipetted onto thesurface of 5 mL of solid PDA in 20-mL HS vials and allowed to diffuseinto the agar overnight, which resulted in final concentrationsbetween 0.0002 and 20 mg per litre agar (assuming homogenicdistribution in the agar). Then the vials were sealed with gas-tightsilicone/teflon septa and analysed by HS-SPME-GC-MS as describedbelow.
For experimental determination of LTPRI values, 10 µL of an alkanestandard mixture (C8–C20 containing 40 mg/L of each alkane in n-hexane) were pipetted into empty HS vials and analysed on the HP-5MS and the HP-Innowax columns accordingly. PDA-filled and emptyHS vials were used to record background (blank) chromatograms toevaluate substances originating from the culture medium, bleeding ofthe GC stationary phase or the SPME fibre coating.
2.4. Sample preparation and time course experiments
Three individual time course experiments were performed withthe same time schedule. For each of two experiments with maximumcultivation periods of 5 days, 9 groups of samples were analysed, at 4replicates (independently grown fungal cultures of T. atroviride) each.In the time course experiment with fusaric acid, 8 groups of sampleswere analysed at 3 replicates, againwithmaximum cultivation time of5 days. For each fungal culture, a single GC-MS measurement wascarried out. All cultures of a time course experiment were inoculatedat the same time and incubated at 22 °C in the dark for 8 h, 23 h, 32 h,47 h, 56 h, 71 h, 80 h, 95 h and 104 h respectively. After cultivation forthe specified time periods, fungal volatiles were removed from the HSvials by purging with synthetic air (filtered through 0.2 µm PTFEMidisart BV membrane filter (Sartorius, Göttingen, Germany)) for30 s. Then, the vials were sealed with screw-caps containing gas-tightsilicone/teflon septa and incubated at 22 °C for further 6 h prior toinstrumental GC-MS analysis.
2.5. HS-SPME-GC-MS analysis
After 15 min of equilibration at 30 °C, extraction of volatiles fromthe headspace of the fungal cultures was carried out by the aid of aGerstel MPS 2XL autosampler (Gerstel, Mülheim, Germany).
For selection of the appropriate fibre coating, the followingmaterialswith the respective thicknesses were tested: polydimethylsiloxane(PDMS), 100 µm; PDMS/divinylbenzene (DVB), 65 µm; carboxene(CAR)/PDMS, 85 µm; polyacrylate (PA), 85 µm and DVB/CAR/PDMS,50/30 µm (Supelco, Vienna, Austria). For all subsequent experiments anSPME fibrewith 65 µmPDMS/DVB coatingwas used. For extraction, thefibre was penetrated by 21 mm into HS vials for 30 min withoutagitation. The volatiles bound to the fibre were desorbed for 2 min in asplit-/splitless injector (splitless mode 250 °C, fibre penetration depth57mm; headspace inlet glass liner, 1.5 mm i.d. (Supelco, Vienna,Austria)), of anAgilent 6890 Ngas chromatograph coupled to anAgilent5975 mass selective detector (MSD) (Agilent, Waldbronn, Germany).Column 1: HP5-MS 30 m×0.25 mm×0.25 µm (Agilent, Waldbronn,Germany): this column was also used for the experiments with fusaricacid. Column 2: HP-Innowax 30 m×0.32 mm×0.5 µm (Agilent, Wald-bronn,Germany).Helium(5.0)wasused as carrier gas at a constantflow
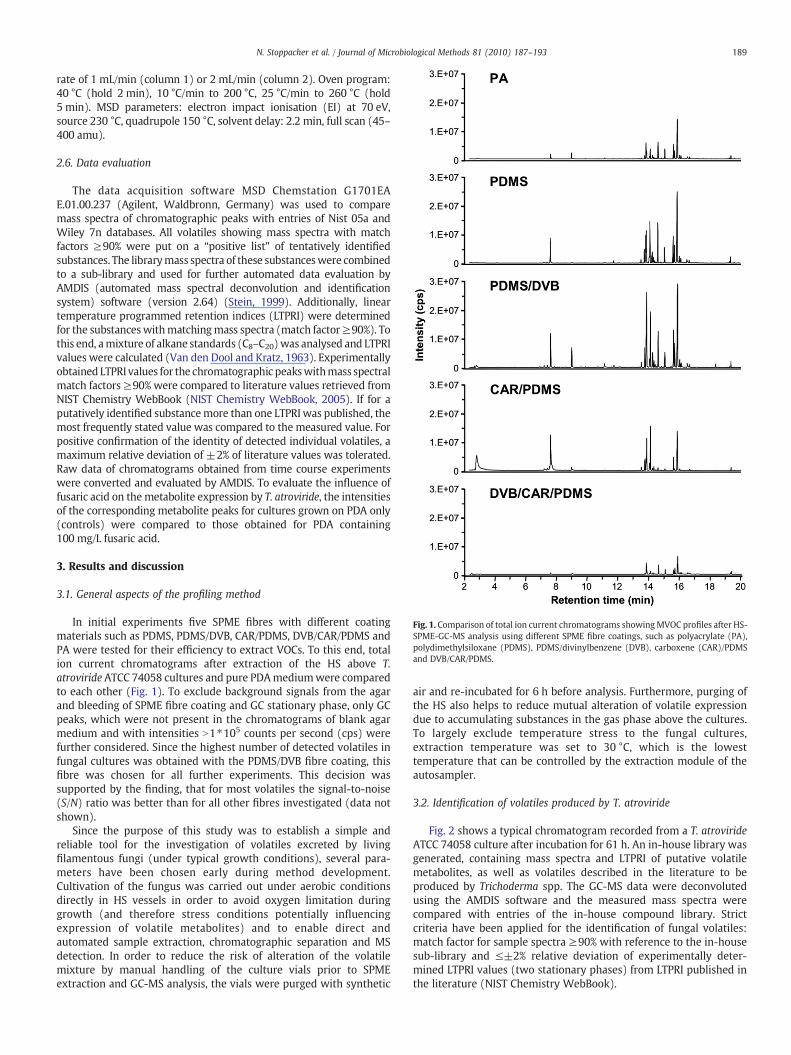
Fig. 1. Comparison of total ion current chromatograms showingMVOC profiles after HS-SPME-GC-MS analysis using different SPME fibre coatings, such as polyacrylate (PA),polydimethylsiloxane (PDMS), PDMS/divinylbenzene (DVB), carboxene (CAR)/PDMSand DVB/CAR/PDMS.
189N. Stoppacher et al. / Journal of Microbiological Methods 81 (2010) 187–193
rate of 1 mL/min (column 1) or 2 mL/min (column 2). Oven program:40 °C (hold 2 min), 10 °C/min to 200 °C, 25 °C/min to 260 °C (hold5 min). MSD parameters: electron impact ionisation (EI) at 70 eV,source 230 °C, quadrupole 150 °C, solvent delay: 2.2 min, full scan (45–400 amu).
2.6. Data evaluation
The data acquisition software MSD Chemstation G1701EAE.01.00.237 (Agilent, Waldbronn, Germany) was used to comparemass spectra of chromatographic peaks with entries of Nist 05a andWiley 7n databases. All volatiles showing mass spectra with matchfactors ≥90% were put on a “positive list” of tentatively identifiedsubstances. The librarymass spectra of these substanceswere combinedto a sub-library and used for further automated data evaluation byAMDIS (automated mass spectral deconvolution and identificationsystem) software (version 2.64) (Stein, 1999). Additionally, lineartemperature programmed retention indices (LTPRI) were determinedfor the substanceswithmatchingmass spectra (match factor≥90%). Tothis end, amixture of alkane standards (C8–C20)was analysed and LTPRIvalues were calculated (Van den Dool and Kratz, 1963). Experimentallyobtained LTPRI values for the chromatographic peakswithmass spectralmatch factors≥90% were compared to literature values retrieved fromNIST Chemistry WebBook (NIST Chemistry WebBook, 2005). If for aputatively identified substancemore than one LTPRI was published, themost frequently stated value was compared to the measured value. Forpositive confirmation of the identity of detected individual volatiles, amaximum relative deviation of ±2% of literature values was tolerated.Raw data of chromatograms obtained from time course experimentswere converted and evaluated by AMDIS. To evaluate the influence offusaric acid on themetabolite expression by T. atroviride, the intensitiesof the corresponding metabolite peaks for cultures grown on PDA only(controls) were compared to those obtained for PDA containing100 mg/L fusaric acid.
3. Results and discussion
3.1. General aspects of the profiling method
In initial experiments five SPME fibres with different coatingmaterials such as PDMS, PDMS/DVB, CAR/PDMS, DVB/CAR/PDMS andPA were tested for their efficiency to extract VOCs. To this end, totalion current chromatograms after extraction of the HS above T.atroviride ATCC 74058 cultures and pure PDAmediumwere comparedto each other (Fig. 1). To exclude background signals from the agarand bleeding of SPME fibre coating and GC stationary phase, only GCpeaks, which were not present in the chromatograms of blank agarmedium and with intensities N1⁎105 counts per second (cps) werefurther considered. Since the highest number of detected volatiles infungal cultures was obtained with the PDMS/DVB fibre coating, thisfibre was chosen for all further experiments. This decision wassupported by the finding, that for most volatiles the signal-to-noise(S/N) ratio was better than for all other fibres investigated (data notshown).
Since the purpose of this study was to establish a simple andreliable tool for the investigation of volatiles excreted by livingfilamentous fungi (under typical growth conditions), several para-meters have been chosen early during method development.Cultivation of the fungus was carried out under aerobic conditionsdirectly in HS vessels in order to avoid oxygen limitation duringgrowth (and therefore stress conditions potentially influencingexpression of volatile metabolites) and to enable direct andautomated sample extraction, chromatographic separation and MSdetection. In order to reduce the risk of alteration of the volatilemixture by manual handling of the culture vials prior to SPMEextraction and GC-MS analysis, the vials were purged with synthetic
air and re-incubated for 6 h before analysis. Furthermore, purging ofthe HS also helps to reduce mutual alteration of volatile expressiondue to accumulating substances in the gas phase above the cultures.To largely exclude temperature stress to the fungal cultures,extraction temperature was set to 30 °C, which is the lowesttemperature that can be controlled by the extraction module of theautosampler.
3.2. Identification of volatiles produced by T. atroviride
Fig. 2 shows a typical chromatogram recorded from a T. atrovirideATCC 74058 culture after incubation for 61 h. An in-house library wasgenerated, containing mass spectra and LTPRI of putative volatilemetabolites, as well as volatiles described in the literature to beproduced by Trichoderma spp. The GC-MS data were deconvolutedusing the AMDIS software and the measured mass spectra werecompared with entries of the in-house compound library. Strictcriteria have been applied for the identification of fungal volatiles:match factor for sample spectra ≥90% with reference to the in-housesub-library and ≤±2% relative deviation of experimentally deter-mined LTPRI values (two stationary phases) from LTPRI published inthe literature (NIST Chemistry WebBook).
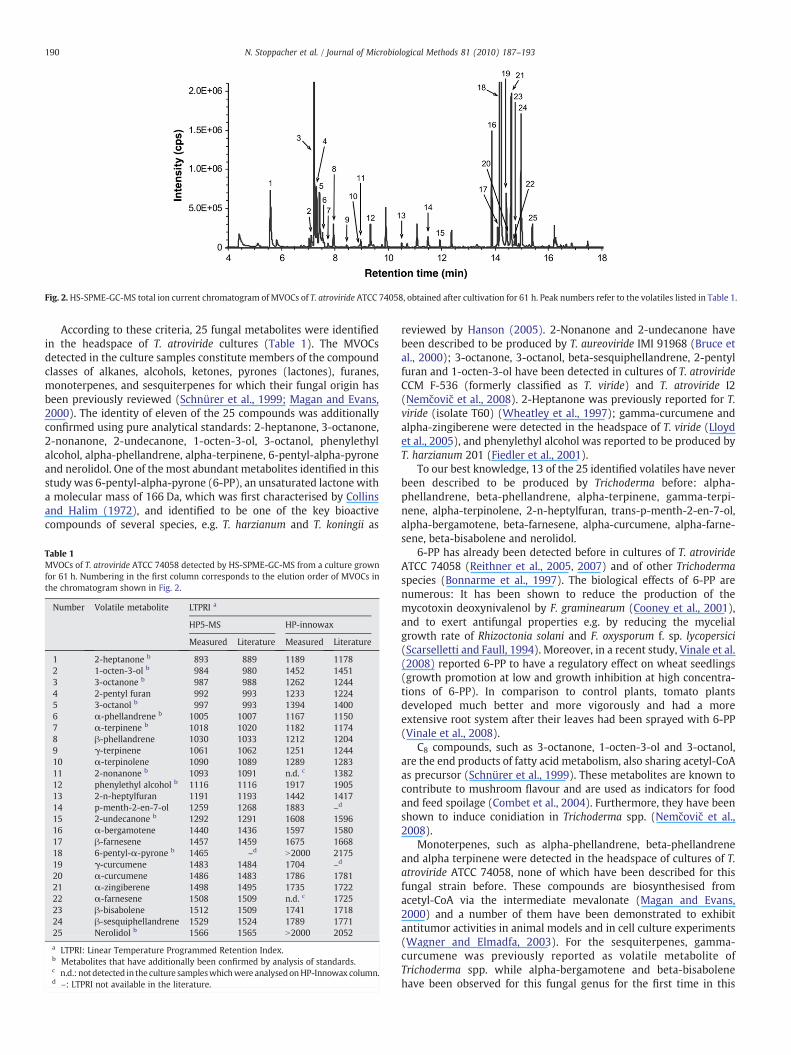
Fig. 2.HS-SPME-GC-MS total ion current chromatogram of MVOCs of T. atroviride ATCC 74058, obtained after cultivation for 61 h. Peak numbers refer to the volatiles listed in Table 1.
190 N. Stoppacher et al. / Journal of Microbiological Methods 81 (2010) 187–193
According to these criteria, 25 fungal metabolites were identifiedin the headspace of T. atroviride cultures (Table 1). The MVOCsdetected in the culture samples constitute members of the compoundclasses of alkanes, alcohols, ketones, pyrones (lactones), furanes,monoterpenes, and sesquiterpenes for which their fungal origin hasbeen previously reviewed (Schnürer et al., 1999; Magan and Evans,2000). The identity of eleven of the 25 compounds was additionallyconfirmed using pure analytical standards: 2-heptanone, 3-octanone,2-nonanone, 2-undecanone, 1-octen-3-ol, 3-octanol, phenylethylalcohol, alpha-phellandrene, alpha-terpinene, 6-pentyl-alpha-pyroneand nerolidol. One of the most abundant metabolites identified in thisstudy was 6-pentyl-alpha-pyrone (6-PP), an unsaturated lactone witha molecular mass of 166 Da, which was first characterised by Collinsand Halim (1972), and identified to be one of the key bioactivecompounds of several species, e.g. T. harzianum and T. koningii as
Table 1MVOCs of T. atroviride ATCC 74058 detected by HS-SPME-GC-MS from a culture grownfor 61 h. Numbering in the first column corresponds to the elution order of MVOCs inthe chromatogram shown in Fig. 2.
Number Volatile metabolite LTPRI a
HP5-MS HP-innowax
Measured Literature Measured Literature
1 2-heptanone b 893 889 1189 11782 1-octen-3-ol b 984 980 1452 14513 3-octanone b 987 988 1262 12444 2-pentyl furan 992 993 1233 12245 3-octanol b 997 993 1394 14006 α-phellandrene b 1005 1007 1167 11507 α-terpinene b 1018 1020 1182 11748 β-phellandrene 1030 1033 1212 12049 γ-terpinene 1061 1062 1251 124410 α-terpinolene 1090 1089 1289 128311 2-nonanone b 1093 1091 n.d. c 138212 phenylethyl alcohol b 1116 1116 1917 190513 2-n-heptylfuran 1191 1193 1442 141714 p-menth-2-en-7-ol 1259 1268 1883 –d
15 2-undecanone b 1292 1291 1608 159616 α-bergamotene 1440 1436 1597 158017 β-farnesene 1457 1459 1675 166818 6-pentyl-α-pyrone b 1465 –d N2000 217519 γ-curcumene 1483 1484 1704 –d
20 α-curcumene 1486 1483 1786 178121 α-zingiberene 1498 1495 1735 172222 α-farnesene 1508 1509 n.d. c 172523 β-bisabolene 1512 1509 1741 171824 β-sesquiphellandrene 1529 1524 1789 177125 Nerolidol b 1566 1565 N2000 2052
a LTPRI: Linear Temperature Programmed Retention Index.b Metabolites that have additionally been confirmed by analysis of standards.c n.d.: not detected in the culture sampleswhichwere analysedonHP-Innowax column.d –: LTPRI not available in the literature.
reviewed by Hanson (2005). 2-Nonanone and 2-undecanone havebeen described to be produced by T. aureoviride IMI 91968 (Bruce etal., 2000); 3-octanone, 3-octanol, beta-sesquiphellandrene, 2-pentylfuran and 1-octen-3-ol have been detected in cultures of T. atrovirideCCM F-536 (formerly classified as T. viride) and T. atroviride I2(Nemčovič et al., 2008). 2-Heptanone was previously reported for T.viride (isolate T60) (Wheatley et al., 1997); gamma-curcumene andalpha-zingiberene were detected in the headspace of T. viride (Lloydet al., 2005), and phenylethyl alcohol was reported to be produced byT. harzianum 201 (Fiedler et al., 2001).
To our best knowledge, 13 of the 25 identified volatiles have neverbeen described to be produced by Trichoderma before: alpha-phellandrene, beta-phellandrene, alpha-terpinene, gamma-terpi-nene, alpha-terpinolene, 2-n-heptylfuran, trans-p-menth-2-en-7-ol,alpha-bergamotene, beta-farnesene, alpha-curcumene, alpha-farne-sene, beta-bisabolene and nerolidol.
6-PP has already been detected before in cultures of T. atrovirideATCC 74058 (Reithner et al., 2005, 2007) and of other Trichodermaspecies (Bonnarme et al., 1997). The biological effects of 6-PP arenumerous: It has been shown to reduce the production of themycotoxin deoxynivalenol by F. graminearum (Cooney et al., 2001),and to exert antifungal properties e.g. by reducing the mycelialgrowth rate of Rhizoctonia solani and F. oxysporum f. sp. lycopersici(Scarselletti and Faull, 1994). Moreover, in a recent study, Vinale et al.(2008) reported 6-PP to have a regulatory effect on wheat seedlings(growth promotion at low and growth inhibition at high concentra-tions of 6-PP). In comparison to control plants, tomato plantsdeveloped much better and more vigorously and had a moreextensive root system after their leaves had been sprayed with 6-PP(Vinale et al., 2008).
C8 compounds, such as 3-octanone, 1-octen-3-ol and 3-octanol,are the end products of fatty acid metabolism, also sharing acetyl-CoAas precursor (Schnürer et al., 1999). These metabolites are known tocontribute to mushroom flavour and are used as indicators for foodand feed spoilage (Combet et al., 2004). Furthermore, they have beenshown to induce conidiation in Trichoderma spp. (Nemčovič et al.,2008).
Monoterpenes, such as alpha-phellandrene, beta-phellandreneand alpha terpinene were detected in the headspace of cultures of T.atroviride ATCC 74058, none of which have been described for thisfungal strain before. These compounds are biosynthesised fromacetyl-CoA via the intermediate mevalonate (Magan and Evans,2000) and a number of them have been demonstrated to exhibitantitumor activities in animal models and in cell culture experiments(Wagner and Elmadfa, 2003). For the sesquiterpenes, gamma-curcumene was previously reported as volatile metabolite ofTrichoderma spp. while alpha-bergamotene and beta-bisabolenehave been observed for this fungal genus for the first time in this
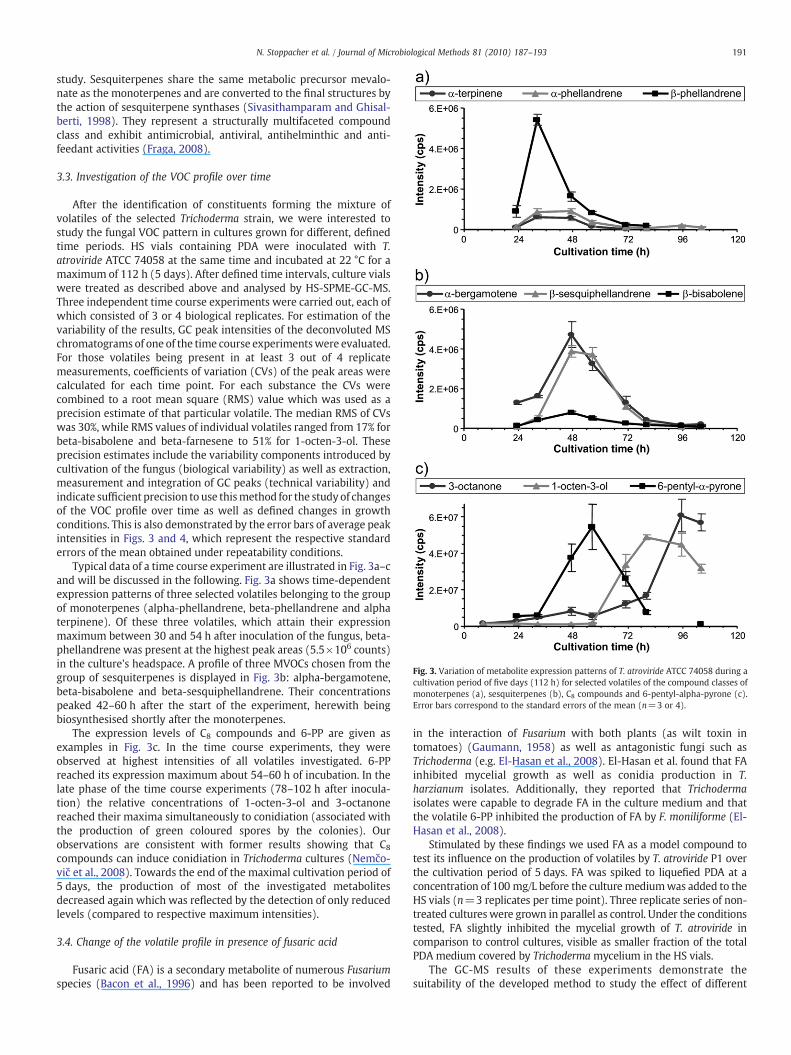
Fig. 3. Variation of metabolite expression patterns of T. atroviride ATCC 74058 during acultivation period of five days (112 h) for selected volatiles of the compound classes ofmonoterpenes (a), sesquiterpenes (b), C8 compounds and 6-pentyl-alpha-pyrone (c).Error bars correspond to the standard errors of the mean (n=3 or 4).
191N. Stoppacher et al. / Journal of Microbiological Methods 81 (2010) 187–193
study. Sesquiterpenes share the same metabolic precursor mevalo-nate as the monoterpenes and are converted to the final structures bythe action of sesquiterpene synthases (Sivasithamparam and Ghisal-berti, 1998). They represent a structurally multifaceted compoundclass and exhibit antimicrobial, antiviral, antihelminthic and anti-feedant activities (Fraga, 2008).
3.3. Investigation of the VOC profile over time
After the identification of constituents forming the mixture ofvolatiles of the selected Trichoderma strain, we were interested tostudy the fungal VOC pattern in cultures grown for different, definedtime periods. HS vials containing PDA were inoculated with T.atroviride ATCC 74058 at the same time and incubated at 22 °C for amaximum of 112 h (5 days). After defined time intervals, culture vialswere treated as described above and analysed by HS-SPME-GC-MS.Three independent time course experiments were carried out, each ofwhich consisted of 3 or 4 biological replicates. For estimation of thevariability of the results, GC peak intensities of the deconvoluted MSchromatograms of one of the time course experimentswere evaluated.For those volatiles being present in at least 3 out of 4 replicatemeasurements, coefficients of variation (CVs) of the peak areas werecalculated for each time point. For each substance the CVs werecombined to a root mean square (RMS) value which was used as aprecision estimate of that particular volatile. The median RMS of CVswas 30%, while RMS values of individual volatiles ranged from 17% forbeta-bisabolene and beta-farnesene to 51% for 1-octen-3-ol. Theseprecision estimates include the variability components introduced bycultivation of the fungus (biological variability) as well as extraction,measurement and integration of GC peaks (technical variability) andindicate sufficient precision to use thismethod for the study of changesof the VOC profile over time as well as defined changes in growthconditions. This is also demonstrated by the error bars of average peakintensities in Figs. 3 and 4, which represent the respective standarderrors of the mean obtained under repeatability conditions.
Typical data of a time course experiment are illustrated in Fig. 3a–cand will be discussed in the following. Fig. 3a shows time-dependentexpression patterns of three selected volatiles belonging to the groupof monoterpenes (alpha-phellandrene, beta-phellandrene and alphaterpinene). Of these three volatiles, which attain their expressionmaximum between 30 and 54 h after inoculation of the fungus, beta-phellandrene was present at the highest peak areas (5.5×106 counts)in the culture's headspace. A profile of three MVOCs chosen from thegroup of sesquiterpenes is displayed in Fig. 3b: alpha-bergamotene,beta-bisabolene and beta-sesquiphellandrene. Their concentrationspeaked 42–60 h after the start of the experiment, herewith beingbiosynthesised shortly after the monoterpenes.
The expression levels of C8 compounds and 6-PP are given asexamples in Fig. 3c. In the time course experiments, they wereobserved at highest intensities of all volatiles investigated. 6-PPreached its expression maximum about 54–60 h of incubation. In thelate phase of the time course experiments (78–102 h after inocula-tion) the relative concentrations of 1-octen-3-ol and 3-octanonereached their maxima simultaneously to conidiation (associated withthe production of green coloured spores by the colonies). Ourobservations are consistent with former results showing that C8compounds can induce conidiation in Trichoderma cultures (Nemčo-vič et al., 2008). Towards the end of the maximal cultivation period of5 days, the production of most of the investigated metabolitesdecreased again which was reflected by the detection of only reducedlevels (compared to respective maximum intensities).
3.4. Change of the volatile profile in presence of fusaric acid
Fusaric acid (FA) is a secondary metabolite of numerous Fusariumspecies (Bacon et al., 1996) and has been reported to be involved
in the interaction of Fusarium with both plants (as wilt toxin intomatoes) (Gaumann, 1958) as well as antagonistic fungi such asTrichoderma (e.g. El-Hasan et al., 2008). El-Hasan et al. found that FAinhibited mycelial growth as well as conidia production in T.harzianum isolates. Additionally, they reported that Trichodermaisolates were capable to degrade FA in the culture medium and thatthe volatile 6-PP inhibited the production of FA by F. moniliforme (El-Hasan et al., 2008).
Stimulated by these findings we used FA as a model compound totest its influence on the production of volatiles by T. atroviride P1 overthe cultivation period of 5 days. FA was spiked to liquefied PDA at aconcentration of 100 mg/L before the culture mediumwas added to theHS vials (n=3 replicates per time point). Three replicate series of non-treated cultureswere grown in parallel as control. Under the conditionstested, FA slightly inhibited the mycelial growth of T. atroviride incomparison to control cultures, visible as smaller fraction of the totalPDA medium covered by Trichoderma mycelium in the HS vials.
The GC-MS results of these experiments demonstrate thesuitability of the developed method to study the effect of different
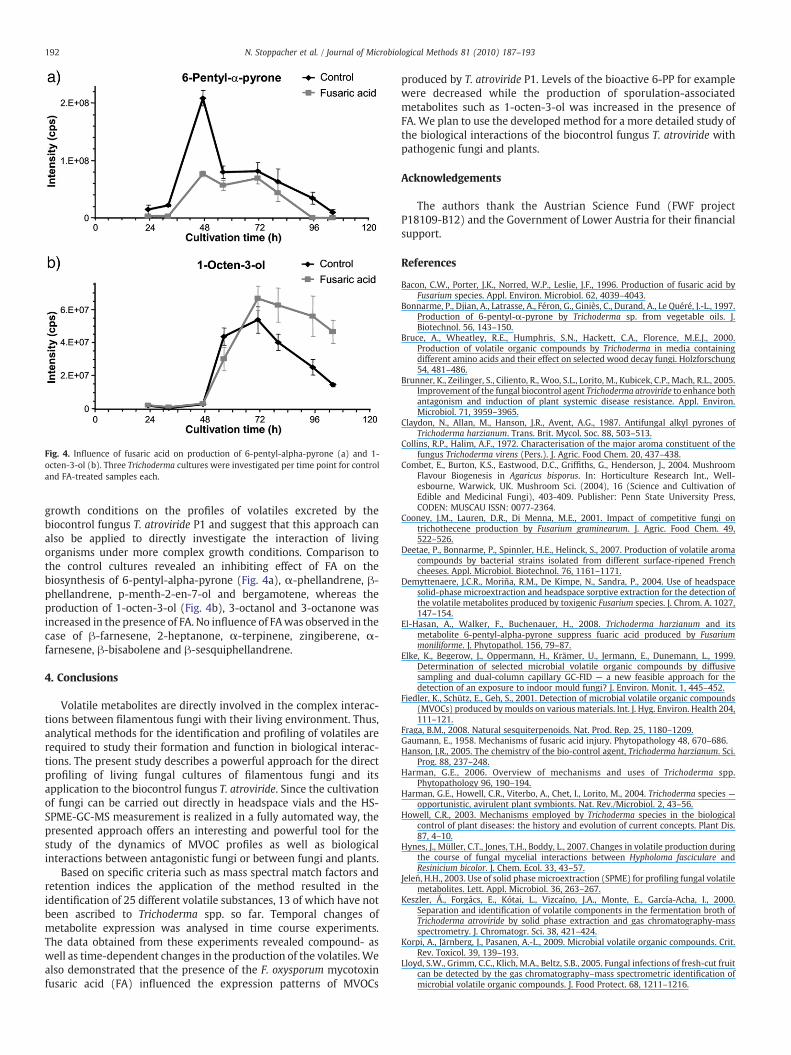
Fig. 4. Influence of fusaric acid on production of 6-pentyl-alpha-pyrone (a) and 1-octen-3-ol (b). Three Trichoderma cultures were investigated per time point for controland FA-treated samples each.
192 N. Stoppacher et al. / Journal of Microbiological Methods 81 (2010) 187–193
growth conditions on the profiles of volatiles excreted by thebiocontrol fungus T. atroviride P1 and suggest that this approach canalso be applied to directly investigate the interaction of livingorganisms under more complex growth conditions. Comparison tothe control cultures revealed an inhibiting effect of FA on thebiosynthesis of 6-pentyl-alpha-pyrone (Fig. 4a), α-phellandrene, β-phellandrene, p-menth-2-en-7-ol and bergamotene, whereas theproduction of 1-octen-3-ol (Fig. 4b), 3-octanol and 3-octanone wasincreased in the presence of FA. No influence of FAwas observed in thecase of β-farnesene, 2-heptanone, α-terpinene, zingiberene, α-farnesene, β-bisabolene and β-sesquiphellandrene.
4. Conclusions
Volatile metabolites are directly involved in the complex interac-tions between filamentous fungi with their living environment. Thus,analytical methods for the identification and profiling of volatiles arerequired to study their formation and function in biological interac-tions. The present study describes a powerful approach for the directprofiling of living fungal cultures of filamentous fungi and itsapplication to the biocontrol fungus T. atroviride. Since the cultivationof fungi can be carried out directly in headspace vials and the HS-SPME-GC-MS measurement is realized in a fully automated way, thepresented approach offers an interesting and powerful tool for thestudy of the dynamics of MVOC profiles as well as biologicalinteractions between antagonistic fungi or between fungi and plants.
Based on specific criteria such as mass spectral match factors andretention indices the application of the method resulted in theidentification of 25 different volatile substances, 13 of which have notbeen ascribed to Trichoderma spp. so far. Temporal changes ofmetabolite expression was analysed in time course experiments.The data obtained from these experiments revealed compound- aswell as time-dependent changes in the production of the volatiles. Wealso demonstrated that the presence of the F. oxysporum mycotoxinfusaric acid (FA) influenced the expression patterns of MVOCs
produced by T. atroviride P1. Levels of the bioactive 6-PP for examplewere decreased while the production of sporulation-associatedmetabolites such as 1-octen-3-ol was increased in the presence ofFA. We plan to use the developed method for a more detailed study ofthe biological interactions of the biocontrol fungus T. atroviride withpathogenic fungi and plants.
Acknowledgements
The authors thank the Austrian Science Fund (FWF projectP18109-B12) and the Government of Lower Austria for their financialsupport.
References
Bacon, C.W., Porter, J.K., Norred, W.P., Leslie, J.F., 1996. Production of fusaric acid byFusarium species. Appl. Environ. Microbiol. 62, 4039–4043.
Bonnarme, P., Djian, A., Latrasse, A., Féron, G., Giniès, C., Durand, A., Le Quéré, J.-L., 1997.Production of 6-pentyl-α-pyrone by Trichoderma sp. from vegetable oils. J.Biotechnol. 56, 143–150.
Bruce, A., Wheatley, R.E., Humphris, S.N., Hackett, C.A., Florence, M.E.J., 2000.Production of volatile organic compounds by Trichoderma in media containingdifferent amino acids and their effect on selected wood decay fungi. Holzforschung54, 481–486.
Brunner, K., Zeilinger, S., Ciliento, R., Woo, S.L., Lorito, M., Kubicek, C.P., Mach, R.L., 2005.Improvement of the fungal biocontrol agent Trichoderma atroviride to enhance bothantagonism and induction of plant systemic disease resistance. Appl. Environ.Microbiol. 71, 3959–3965.
Claydon, N., Allan, M., Hanson, J.R., Avent, A.G., 1987. Antifungal alkyl pyrones ofTrichoderma harzianum. Trans. Brit. Mycol. Soc. 88, 503–513.
Collins, R.P., Halim, A.F., 1972. Characterisation of the major aroma constituent of thefungus Trichoderma virens (Pers.). J. Agric. Food Chem. 20, 437–438.
Combet, E., Burton, K.S., Eastwood, D.C., Griffiths, G., Henderson, J., 2004. MushroomFlavour Biogenesis in Agaricus bisporus. In: Horticulture Research Int., Well-esbourne, Warwick, UK. Mushroom Sci. (2004), 16 (Science and Cultivation ofEdible and Medicinal Fungi), 403-409. Publisher: Penn State University Press,CODEN: MUSCAU ISSN: 0077-2364.
Cooney, J.M., Lauren, D.R., Di Menna, M.E., 2001. Impact of competitive fungi ontrichothecene production by Fusarium graminearum. J. Agric. Food Chem. 49,522–526.
Deetae, P., Bonnarme, P., Spinnler, H.E., Helinck, S., 2007. Production of volatile aromacompounds by bacterial strains isolated from different surface-ripened Frenchcheeses. Appl. Microbiol. Biotechnol. 76, 1161–1171.
Demyttenaere, J.C.R., Moriña, R.M., De Kimpe, N., Sandra, P., 2004. Use of headspacesolid-phase microextraction and headspace sorptive extraction for the detection ofthe volatile metabolites produced by toxigenic Fusarium species. J. Chrom. A. 1027,147–154.
El-Hasan, A., Walker, F., Buchenauer, H., 2008. Trichoderma harzianum and itsmetabolite 6-pentyl-alpha-pyrone suppress fuaric acid produced by Fusariummoniliforme. J. Phytopathol. 156, 79–87.
Elke, K., Begerow, J., Oppermann, H., Krämer, U., Jermann, E., Dunemann, L., 1999.Determination of selected microbial volatile organic compounds by diffusivesampling and dual-column capillary GC-FID — a new feasible approach for thedetection of an exposure to indoor mould fungi? J. Environ. Monit. 1, 445–452.
Fiedler, K., Schütz, E., Geh, S., 2001. Detection of microbial volatile organic compounds(MVOCs) produced by moulds on various materials. Int. J. Hyg. Environ. Health 204,111–121.
Fraga, B.M., 2008. Natural sesquiterpenoids. Nat. Prod. Rep. 25, 1180–1209.Gaumann, E., 1958. Mechanisms of fusaric acid injury. Phytopathology 48, 670–686.Hanson, J.R., 2005. The chemistry of the bio-control agent, Trichoderma harzianum. Sci.
Prog. 88, 237–248.Harman, G.E., 2006. Overview of mechanisms and uses of Trichoderma spp.
Phytopathology 96, 190–194.Harman, G.E., Howell, C.R., Viterbo, A., Chet, I., Lorito, M., 2004. Trichoderma species —
opportunistic, avirulent plant symbionts. Nat. Rev./Microbiol. 2, 43–56.Howell, C.R., 2003. Mechanisms employed by Trichoderma species in the biological
control of plant diseases: the history and evolution of current concepts. Plant Dis.87, 4–10.
Hynes, J., Müller, C.T., Jones, T.H., Boddy, L., 2007. Changes in volatile production duringthe course of fungal mycelial interactions between Hypholoma fasciculare andResinicium bicolor. J. Chem. Ecol. 33, 43–57.
Jeleń, H.H., 2003. Use of solid phase microextraction (SPME) for profiling fungal volatilemetabolites. Lett. Appl. Microbiol. 36, 263–267.
Keszler, Á., Forgács, E., Kótai, L., Vizcaíno, J.A., Monte, E., García-Acha, I., 2000.Separation and identification of volatile components in the fermentation broth ofTrichoderma atroviride by solid phase extraction and gas chromatography-massspectrometry. J. Chromatogr. Sci. 38, 421–424.
Korpi, A., Järnberg, J., Pasanen, A.-L., 2009. Microbial volatile organic compounds. Crit.Rev. Toxicol. 39, 139–193.
Lloyd, S.W., Grimm, C.C., Klich, M.A., Beltz, S.B., 2005. Fungal infections of fresh-cut fruitcan be detected by the gas chromatography–mass spectrometric identification ofmicrobial volatile organic compounds. J. Food Protect. 68, 1211–1216.

193N. Stoppacher et al. / Journal of Microbiological Methods 81 (2010) 187–193
Magan, N., Evans, P., 2000. Volatiles as an indicator of fungal activity and differentiationbetween species, and the potential use of electronic nose technology for earlydetection of grain spoilage. J. Stored Prod. Res. 36, 319–340.
Meruva, N.K., Penn, J.M., Farthing, D.E., 2004. Rapid identification of microbial VOCsfrom tobacco molds using closed-loop stripping and gas chromatography/time-of-flight mass spectrometry. J. Ind. Microbiol. Biotechnol. 31, 482–488.
Nemčovič, M., Jakubíková, L., Víden, I., Vladimír, F., 2008. Induction of conidiation byendogenousvolatile compounds in Trichoderma spp. FEMS Microbiol. Lett. 284,231–236.
NIST Chemistry WebBook, 2005. Nist Standard Reference Database http://webbook.nist.gov/chemistry (retrieved 12/28/2009).
Oprean, R., Tamas, M., Sandulescu, R., Roman, L., 1998. Essential oils analysis. I.Evaluation of essential oils composition using both GC and MS fingerprints. J.Pharm. Biomed. Anal. 18 (4–5), 651–657.
Oprean, R., Oprean, L., Tamas, M., Sandulescu, R., Roman, L., 2001. Essential oils analysis.II. Mass spectra identification of terpene and phenylpropane derivatives. J. Pharm.Biomed. Anal. 24 (5–6), 1163–1168.
Pinches, S.E., Apps, P., 2007. Production in food of 1, 3-pentadiene and styrene byTrichoderma species. Int. J. Food Microbiol. 116, 182.
Reino, J.L., Guerrero, R.F., Hernández-Galán, R., Collado, I.G., 2008. Secondary metabolitesfrom species of the biocontrol agent Trichoderma. Phytochem. Rev. 7, 89–123.
Reithner, B., Brunner, K., Schuhmacher, R., Peissl, P., Seidl, V., Krska, R., Zeilinger, S.,2005. The G protein α subunit Tga1 of Trichoderma atroviride is involved inchitinase formation and differential production of antifungal metabolites. FungalGenet. Biol. 42, 749–760.
Reithner, B., Schuhmacher, R., Stoppacher, N., Pucher, M., Brunner, K., Zeilinger, S., 2007.Signaling via the Trichoderma atroviride mitogen-activated protein kinase Tmk1differentially affects mycoparasitism and plant protection. Fungal Genet. Biol. 44,1123–1133.
Ruther, J., 2000. Retention index database for identification of general green leafvolatiles in plants by coupled capillary gas chromatography-mass spectrometry. J.Chromatogr. A 890, 313–319.
Scarselletti, R., Faull, J.L., 1994. In vitro activity of 6-pentyl-α-pyrone, a metabolite ofTrichoderma harzianum, in the inhibition of Rhizoctonia solani and Fusariumoxysporum f. sp. Lycopersici. Mycol. Res. 98, 1207–1209.
Schnürer, J., Olsson, J., Börjesson, T., 1999. Fungal volatiles as indicators of food andfeeds spoilage. Fungal Genet. Biol. 27, 209–217.
Sivasithamparam, K., Ghisalberti, E.L., 1998. Trichoderma and Gliocladium. In: G.E.,Kubicek, C.P. (Eds.), Harman Vol. 1. Taylor & Francis Ltd., London, pp. 139–188.
Stein, S.E., 1999. An integrated method for spectrum extraction and compoundidentification from gas chromatography/mass spectrometry data. J. Am. Soc. MassSpectrom. 10, 770–781.
Van den Dool, H., Kratz, P.D., 1963. A generalization of the retention index systemincluding linear temperature programmed gas–liquid partition chromatography. J.Chromatogr. A 11, 463–471.
Van Lancker, F., Adams, A., Delmulle, B., De Saeger, S., Moretti, A., Van Peteghem, C., DeKimpe, N., 2008. Use of headspace SPME-GC-MS for the analysis of the volatilesproduced by indoor molds grown on different substrates. J. Environ. Monit. 10,1127–1133.
Verma, M., Brar, S.K., Tyagi, R.D., Surampalli, R.Y., Valéro, J.R., 2007. Antagonistic fungi,Trichoderma spp.: panoply of biological control. Biochem. Engineer. J. 37, 1–20.
Vinale, F., Sivasithamparam, K., Ghisalberti, E.L., Marra, R., Barbetti, M.J., Li, H., Woo, S.L.,Lorito, M., 2008. A novel role for Trichoderma secondary metabolites in theinteractions with plants. Physiol. Mol. Plant Pathol. 72, 80–86.
Wagner, K.-H., Elmadfa, I., 2003. Biological relevance of terpenoids: overview focusingon mono-, di- and tetraterpenes. Ann. Nutr. Metab. 47, 95–106.
Wheatley, R., Hackett, C., Bruce, A., Kundzewicz, A., 1997. Effect of substratecomposition on production of volatile organic compounds from Trichoderma spp.Inhibitory to wood decay fungi. Int. Biodeter. Biodegr. 39, 199–205.
Zeppa, G., Allegrone, G., Barbeni, M., Guarda, P.A., 1990. Variability in the production ofvolatile metabolites by Trichoderma viride. Ann. Microbiol. 40, 171–176.




















