
Trichoderma atroviride PHR1, a Fungal Photolyase Responsible for DNA Repair, Autoregulates Its Own...
12
Published Ahead of Print 1 June 2007. 2007, 6(9):1682. DOI: 10.1128/EC.00208-06. Eukaryotic Cell Benjamin A. Horwitz and Alfredo Herrera-Estrella Gloria M. Berrocal-Tito, Edgardo U. Esquivel-Naranjo, Autoregulates Its Own Photoinduction Photolyase Responsible for DNA Repair, PHR1, a Fungal Trichoderma atroviride http://ec.asm.org/content/6/9/1682 Updated information and services can be found at: These include: REFERENCES http://ec.asm.org/content/6/9/1682#ref-list-1 at: This article cites 49 articles, 19 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on April 13, 2014 by guest http://ec.asm.org/ Downloaded from on April 13, 2014 by guest http://ec.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Trichoderma atroviride PHR1, a Fungal Photolyase Responsible for DNA Repair, Autoregulates Its Own...

Published Ahead of Print 1 June 2007. 2007, 6(9):1682. DOI: 10.1128/EC.00208-06. Eukaryotic Cell
Benjamin A. Horwitz and Alfredo Herrera-EstrellaGloria M. Berrocal-Tito, Edgardo U. Esquivel-Naranjo, Autoregulates Its Own Photoinduction Photolyase Responsible for DNA Repair,
PHR1, a FungalTrichoderma atroviride
http://ec.asm.org/content/6/9/1682Updated information and services can be found at:
These include:
REFERENCEShttp://ec.asm.org/content/6/9/1682#ref-list-1at:
This article cites 49 articles, 19 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

EUKARYOTIC CELL, Sept. 2007, p. 1682–1692 Vol. 6, No. 91535-9778/07/$08.00�0 doi:10.1128/EC.00208-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Trichoderma atroviride PHR1, a Fungal Photolyase Responsible forDNA Repair, Autoregulates Its Own Photoinduction�
Gloria M. Berrocal-Tito,1† Edgardo U. Esquivel-Naranjo,1Benjamin A. Horwitz,2 and Alfredo Herrera-Estrella1*
Laboratorio Nacional de Genomica para la Biodiversidad, Cinvestav Campus Guanajuato, Km 9.6 Libramiento Norte CarreteraIrapuato-Leon, A.P. 629, Irapuato 36500, Guanajuato, Mexico,1 and Department of Biology,
Technion—Israel Institute of Technology, Haifa 32000, Israel2
Received 30 June 2006/Accepted 18 May 2007
The photolyases, DNA repair enzymes that use visible and long-wavelength UV light to repair cyclobutanepyrimidine dimers (CPDs) created by short-wavelength UV, belong to the larger photolyase-cryptochrome genefamily. Cryptochromes (UVA–blue light photoreceptors) lack repair activity, and sensory and regulatory roleshave been defined for them in plants and animals. Evolutionary considerations indicate that cryptochromesdiverged from CPD photolyases before the emergence of eukaryotes. In prokaryotes and lower eukaryotes, somephotolyases might have photosensory functions. phr1 codes for a class I CPD photolyase in Trichodermaatroviride. phr1 is rapidly induced by blue and UVA light, and its photoinduction requires functional blue lightregulator (BLR) proteins, which are White Collar homologs in Trichoderma. Here we show that deletion of phr1abolished photoreactivation of UVC (200 to 280 nm)-inhibited spores and thus that PHR1 is the maincomponent of the photorepair system. The 2-kb 5� upstream region of phr1, with putative light-regulatedelements, confers blue light regulation on a reporter gene. To assess phr1 photosensory function, fluenceresponse curves of this light-regulated promoter were tested in null mutant (�phr1) strains. Photoinduction ofthe phr1 promoter in �phr1 strains was >5-fold more sensitive to light than that in the wild type, whereas inPHR1-overexpressing lines the sensitivity to light increased about 2-fold. Our data suggest that PHR1 mayregulate its expression in a light-dependent manner, perhaps through negative modulation of the BLRproteins. This is the first evidence for a regulatory role of photolyase, a role usually attributed tocryptochromes.
DNA photolyases are able to use sunlight for DNA repairdue to the fact that they have two chromophores, namely, acatalytic one which is a flavin (reduced flavin adenine dinucle-otide) and a light-harvesting one which is either methyl tetra-hydrofolate (MTHF) or another flavin. Photolyases bind todamaged DNA, and upon excitation of their flavin, they gen-erate a dimer radical which splits, restoring the damaged pairof neighboring pyrimidines. Photolyases, which exist in almostall forms of life, carry out this repair reaction on their own,with no need for partners.
Cryptochromes sense sunlight using the same chromophoresas photolyases, but they lack repair activity. They have anadditional C-terminal domain (8) and act as photoreceptorsand regulators of specific responses, usually in coordinationwith other proteins. Plant cryptochromes act as blue-UVAlight receptors (1, 21, 30, 32, 50). Along with phytochromes andphototropins (10), they control growth, gene expression, andcircadian rhythms (46) in response to light. In Drosophila mela-
nogaster, too, cryptochrome acts as a blue-UVA light receptorand, together with visual pigments, entrains the circadian os-cillator by light (9, 14). The mammalian cryptochromes areintegral components of the negative feedback loop of the cir-cadian oscillator, and this function may be independent of light(20, 38, 48). There is genetic evidence for a photoreceptive rolefor cryptochromes in the mammalian retina, particularly forphotocontrol of gene expression in the superchiasmatic nuclei(44). A new cryptochrome class, DASH, which includes cryp-tochromes with no C-terminal extension, has been reportedrecently. Members of this class include cry from Synechocystissp., cry1 from Vibrio cholerae, and Atcry3 from Arabidopsisthaliana (6, 29, 49). The existence of DASH cryptochromessupports the hypothesis that cryptochromes evolved before thedivergence of eukaryotes and prokaryotes (28) and also sup-ports our proposition (27) that in bacteria and lower eu-karyotes, we might find photosensory functions in cyclobutanepyrimidine dimer (CPD) photolyases. Indeed, the reverse hasbeen reported recently, as vertebrate orthologues of DASHcryptochromes show residual CPD photolyase activity (13).Fungal photolyases might therefore provide some of the sen-sory functions attributed to cryptochromes in bacteria, plants,and animals.
In fungi, blue light is central to physiology, modulating thedirection and rate of growth, inducing mycelial pigmentation,inducing or repressing asexual or sexual reproduction, andentraining the circadian rhythm. The term “cryptochrome” wasinitially applied to plant, fungal, and Drosophila blue-UVA
* Corresponding author. Mailing address: Laboratorio Nacional deGenomica para la Biodiversidad, Cinvestav Campus Guanajuato, Km9.6 Libramiento Norte Carretera Irapuato-Leon, A.P. 629, Irapuato36500, Guanajuato, Mexico. Phone: 52 462 6239600. Fax: 52 4626245849. E-mail: [email protected].
† Present address: UMR Interactions Plantes-Microorganismes etSante Vegetale, Institute National de la Recherche Agronomique,Centre de Recherche de Sophia-Antipolis, 06903 Sophia AntipolisCedex, France.
� Published ahead of print on 1 June 2007.
1682
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

action spectra (11, 17, 19, 34). With the identification of Neu-rospora crassa WC1 and WC2 as a likely photoreceptor com-plex (3) which has the properties expected of a light-regulatedtranscription factor, at least in vitro (15, 22, 25), fungal bluelight receptors are no longer entirely “cryptic.” There is no lackof multiple photoreceptor candidates in the Neurospora ge-nome (16), including the PAS domain protein VIVID, whichgates photoresponses (23) and is required for adaptation (43).A second photolyase/cryptochrome gene has been annotatedin the N. crassa genome. This gene can be classified in theDASH cryptochrome group due to its close identity toSynechocystis cry (13, 16; G. M. Berrocal-Tito, unpublishedresults). The photobiological functions of fungal opsin andcryptochrome genes have yet to be defined.
The photolyase gene phr1 of the filamentous fungus Tri-choderma atroviride is strongly and rapidly up-regulated by bluelight (4, 5). Putative light regulatory elements have been de-scribed for the phr1 promoter. Blue light regulator genes (blr1and blr2) coding for White Collar (WC) homologs were char-acterized in Trichoderma, and they are necessary for phr1 pho-toinduction (7).
In this study, we show that the wavelength dependence ofphr1 photoinduction indicates a blue-UVA light photoreceptorand that phr1 codes for a functional photolyase. Furthermore,a lack of photoreactivation in �phr1 strains confirmed the roleof phr1 as the major component of the DNA photorepairsystem. To assess its possible photosensory role, we measuredthe light sensitivity for induction of phr1 in transgenic lineslacking or overexpressing the corresponding gene. In order tomonitor phr1 light induction in null mutants, a reporter genedriven by the phr1 promoter was introduced. phr1 photoinduc-tion in null mutant and multicopy (MC) strains showed a shiftto lower fluences than that of the wild-type (WT) strain, sug-gesting that phr1 autoregulates its transcription. PHR1 couldhave the putative BLR1/BLR2 complex as a target, modulatingits activity. We further demonstrate that deletion of phr1clearly alters the expression of the recently described blugenes, a set of genes up-regulated by light.
MATERIALS AND METHODS
Transgenic fungal strains and nucleic acid and protein methods. To obtainnull strains, a disruption construct was made as follows. The 5�-flank (2 kb) and3�-flank (1.73 kb) fragments of phr1 were generated by PCR, using pT3T7Thphr15.8 as a template. The 5� flank, with PstI/NdeI sites, was obtained using primersPL5 and PL6. The 3� flank, with BamHI/HindII sites, was obtained with primersPL9 and PL10. The green fluorescent protein (GFP) coding region (0.8 kb), withNdeI/SalI sites, was amplified using synthetic GFP as a template, with primersPL7 and PL8. A 1.4-kb fragment containing the hph gene was obtained fromeither pCSN44 or pUCATPH by digestion with the SalI/BamHI enzymes. All ofthese fragments were introduced by one-step ligation into a pBlueScript SKvector at PstI/HindIII sites, and the construct was named pC16. The primersequences were as follows: PL5, 5�GCA ACT GCA GCG CTT CTT GTC ACTTTC T3�; PL6, 5�GTA GAT CCA TAT GTT ACA CAC GGT GGA TTC TTAAAT GAT AAT3�; PL7, 5�GTA GAT CCA TCA TGG TGA GCA AGG GCGAGG AG3�; PL8, 5�ACG CGT CGA CGC AAG ACC GGC AAC AGG ATTCAA3�; PL9, 5�AGC TGG ATC CAT TGG ATA TCG AAA ACG GTT CATC3�; and PL10, 5�GAT CAA GCT TGT CGG AAG AGA TCA CCA GTG TTTT3�. The construct was transformed as either circular plasmid DNA or linearDNA amplified from the plasmid by PCR with primers PL5 and PL10, by particlebombardment, or by the polyethylene glycol-Ca2� method. MC strains wereobtained by cotransformation of spores of T. atroviride IMI 206040 with theplasmids pHat� (8.9 kb) and pT3T7phr1 5.8 (8.5 kb) by particle bombardment asdescribed previously (36). The bombarded plates were overlaid with 400 �g/mlhygromycin B in agar. Transformed strains selected on hygromycin at 200 �g/ml
were subjected to monosporic subculture. Null strain candidates were analyzedby PCR for the presence of phr1, using primers PL11 (5� TGC AAG CGG GACAAT CAG TTC3�) and PL12 (5� AAT CCC CAG CCA CCG TTG TTG3�). Todetect the presence of gfp, we used primers PL7 and PL8 (see above), and forhph, we used primers HygF (5� GAG GGC GAA GAA TCT CGT GC3�) andHygR (5� CAC TGA CGG TGT CGT CCA TC3�). For Southern analysis, 5 �gof total DNA was digested with SalI, resolved in a 0.8% agarose gel, and blottedonto a nylon membrane. For Northern analysis, total RNA was isolated usingTriReagent (MRC) following the manufacturer’s protocol. Thirty micrograms ofRNA was separated by electrophoresis on denaturing formaldehyde gels, blottedonto Hybond N� membranes (Amersham), hybridized in 7% sodium dodecylsulfate (SDS), 250 mM phosphate, pH 7, and washed twice for 20 min each timein 5% SDS, 20 mM phosphate, followed by a 5- to 10-min wash with 1% SDS, 20mM phosphate. Signals were measured using a phosphorimager (Fuji) and Tinav.2.1 software and normalized to total RNA detected by hybridization with anrRNA genomic fragment or by quantitation of ethidium bromide fluorescence onstained gels prior to transfer. In some earlier experiments, signals were quanti-fied by scanning autoradiograms and integration using NIH Image 1.63 software.
To obtain total cell protein extracts, mycelium was lyophilized, ground to apowder, and extracted in Laemmli sample buffer with 1 mM phenylmethylsul-fonyl fluoride by vortex mixing with acid-washed glass beads. The extract washeated for 5 min to 100°C and centrifuged for 5 min at 18,000 � g, and 160�g/lane was separated by SDS-polyacrylamide gel electrophoresis, blotted ontoa nitrocellulose membrane, and probed with antiserum to Neurospora photolyaseat a 1:500 dilution; the second antibody was goat anti-rabbit (GIBCO) diluted1:5,000, and detection was done by ECL (Amersham).
Real-time PCR. Approximately 1 �g total RNA was treated with 1 Kunitz unitof RNase-free DNase (QIAGEN) for 20 min at 37°C in New England Biolabsrestriction enzyme buffer 4 (total volume, 10 �l), followed by 15 min at 65°C toinactivate the DNase. Four microliters of this reaction mix was reverse tran-scribed (ImProm-II; Promega) according to the recommended protocol, usingrandom primers; the reaction was done for 1 h at 42°C with 3.75 mM MgCl2,followed by 15 min at 70°C to inactivate the reverse transcriptase. Four micro-liters of a 100-fold dilution of each cDNA (in the linear range for all primer pairs,as tested in preliminary experiments on a pooled cDNA sample) was amplifiedin a 20-�l reaction mix with SYBR green ROX mix (ABgene, Surrey, UnitedKingdom) and 1 pmol of each primer. Primer pairs (forward and reverse [5�-3�])were as follows: CGC CTG GCG AGA TTT TTA CA and ACA GAC ATATGG CCA GTT AAC CAA A for photolyase, GAG CTG AAG GGC ATCGAC TT and CTT GTG CCC CAG GAT GTT G for GFP, and GAT GGG TGTCAA CCA CAA GGA and GCA AGA GGC GTT GGA GAG AAC forglyceraldehyde-3-phosphate dehydrogenase. Reactions were run in duplicate ortriplicate in an Applied Biosystems 7000 cycler. Data were analyzed using the2���CT method (35), with the glyceraldehyde-3-phosphate dehydrogenase primersas the standard primer pair and the maximum fluence as the standard condition.The cycle threshold (CT) values for the two or three replicate PCRs with eachRNA sample were averaged, and then 2���CT values were calculated, followed byaveraging of the independent experiments.
Growth and photoinduction conditions. Standard culture conditions werePDYC medium (24 g/liter potato dextrose broth, 2 g/liter yeast extract, 1.2 g/litercasein hydrolysate; all from Difco) at 25°C. For photoinduction experiments,cultures were grown in liquid PDYC medium as described previously (5) for 36 hin total darkness, after which they were exposed to a pulse of blue light of 540�mol m2 or to the light and irradiance required. Samples were kept in the darkand collected 15 min after exposure under a red safelight. Light of differentwavelength bands was obtained by filtering the output of cool-white fluorescenttubes or a slide projector (equipped with heat filter 115 [Schott]) through thefollowing filters: B, blue Lee filter 183 or a blue acrylic filter (4); V, violet Leefilter; BG, blue-green Lee filter 124; R, red Lee filter 106 or red dichroic filter(OCLI, Santa Rosa, CA); and G, cyan dichroic filter (OCLI) combined withcutoff filter OG 515 (Schott). For UVA, light from a long-wavelength UV lamp(UVP) passed through a wide-band UG11 filter (Corning). Fluence rates wereadjusted with neutral-density filters and measured with a LiCor (Lincoln, NB)quantum photometer (visible region) or UV meter (UVP) for UVA. Fluencesused were 78, 420, and 1,050 �mol m�2.
Complementation of Escherichia coli deficient in photolyase. The coding se-quence of phr1 was obtained in two steps, using the genomic clone pT3T7phr15.8(5) as a template for PCR amplification using Pfu DNA polymerase (Promega).First, two fragments were obtained using the following oligonucleotide pairs,where F indicates sense and R indicates antisense primers: F, 5�CAG GAA TTCGAT GCT CGC GAG GAG CG3�; 2R, 5�GTT CAT ACA GAC ATA TGGCCA G3�; 3F, 5�GGC CAT ATG TCT GTA TGA ACA AGC C3�; and R,5�GTT CGG GCC CTA GAT CCC TCT CTC CAG CC3�. These two products
VOL. 6, 2007 AUTOREGULATION OF FUNGAL PHOTOLYASE 1683
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

were combined and used as a template for amplification using primers F and R(including the start and stop codons, whose locations are indicated in bold),resulting in removal of the single intron in phr1. The 1.9-kb final amplifiedproduct was digested with EcoRI and ApaI and ligated into pBluescript SK(Stratagene), and the construct was verified by sequencing both strands. Thisplasmid was used to transform electrocompetent cells of E. coli SY2 (JM107�phr::Cmr �uvrA::Kmr �recA::Tetr) to ampicillin resistance, and the resultingstrain was tested for photorepair of UV damage. For photorepair assays, bacte-rial colonies were isolated from LB plates with 50 �g/ml ampicillin, 50 �g/mlchloramphenicol, 25 �g/ml kanamycin, and 20 �g/ml tetracycline; overnightcultures were plated at different dilutions on LB with 50 �g/ml ampicillin, driedfor 30 min in a laminar flow hood, and exposed to the indicated UVC fluencesfrom a cross-linker (Hoefer). The UVC fluence rate was reduced with a neutralfilter consisting of several layers of nylon mesh and calibrated with a UVP UVmeter.
RESULTS
UVA and blue light induce phr1 expression. We reportedpreviously that the blue region of the spectrum was effective inthe induction of phr1 expression (4, 5). In order to comparephr1 induction with the other blue light responses, photorepairand photoconidiation, we mapped the wavelength dependenceof phr1 mRNA induction. We used wide-band illumination indefined spectral regions from 300 to 760 nm. Light between300 and 550 nm is effective for phr1 photoinduction. UVA isthe most effective region of the spectrum for phr1 photoinduc-tion, being almost twice as effective as the blue (B) region,followed by the violet (V) and the blue-green (BG) regions.phr1 photoinduction is blind to red light (R) (Fig. 1).
Deletion of phr1 results in a loss of photoreactivation, whilephr1 overexpression increases photoreactivation capacity. The62% identity of the deduced amino acid sequence of PHR1 toNeurospora crassa photolyase suggested that phr1 encodes afunctional photolyase belonging to class I, type I (5). We testedwhether phr1 could complement a CPD photolyase mutant ofE. coli, i.e., strain SY2. Cells harboring phr1 had a UVC sur-vival rate of 3 to 4 orders of magnitude higher than that of cells
carrying a control plasmid after exposure to photoreactivatinglight (Fig. 2), indicating that phr1 indeed confers photorepairactivity.
To answer the question of whether photolyase plays anotherrole in Trichoderma in addition to its DNA repair function, wegenerated strains in which phr1 was not present or was over-expressed. Strains lacking the photolyase gene were obtainedby gene replacement, using the following approach. Fragmentsflanking the coding region of phr1 were fused to the hygromy-cin phosphotransferase gene (hph) as a selectable marker (Fig.3A). This construct was introduced into Trichoderma, and can-didates were screened by PCR. Two transformants, 3.20 and3.3, showed double homologous recombination of the con-struct (phr1 is absent and hph is present), while 3.21 showedectopic integration (Fig. 3B). phr1 MC strains were generatedby the introduction of plasmid pT3T7phr1 5.8 (5), containingthe full-length genomic clone (Fig. 3C). Multiple integration ofthe plasmid was confirmed by Southern analysis. Strains car-rying one (MC8) and two (MC6 and MC7) extra copies of phr1were obtained (Fig. 3D).
In order to determine whether more phr1 copies indeedcorrelated with more PHR1 protein, we analyzed the mRNAand protein levels of phr1 in MC strains. We reported previ-ously that, in Trichoderma, low fluences of blue light rapidlyinduce the expression of phr1 in dark-grown mycelium andconidiophores (5). Consequently, we analyzed whether thephr1 mRNA and protein levels were increased in MC strains inthe dark and under photoinduced conditions. Dark levels ofphr1 mRNA were not detectable in any of the strains, whileunder photoinduced conditions phr1 expression was three- tofivefold higher in MC strains than in the WT (data not shown).Western blot analysis using antiserum against the Neurosporaphotolyase and using dark-grown and photoinduced culturesshowed that PHR1 was almost undetectable in the dark butthat a polypeptide of the expected size was clearly detectable inphotoinduced cultures. This signal was seven- to ninefold moreabundant in the lines with two extra copies, whereas no proteincould be detected in the �phr1 strains 3.20 and 3.3 (Fig. 3E).
�phr1 strains 3.20 and 3.3 were assayed for photoreactiva-
FIG. 1. Wavelength dependence of phr1 induction. Northern anal-ysis was performed from induced cultures by fluences of differentspectral regions. Data from different blue filters were consistent withinthe variability and were combined. The peak transmission wavelengths,in nm, of the filters were as follows: UVA, 360 (300 to 390); violet, 412(330 to 530); blue, 440 (395 to 500) and 458 (380 to 560); blue-green,505 (450 to 560); green, 505 to 600; and red, 585 to 760 (the numbersin parentheses indicate the ranges for �10% transmission). Relativeeffectiveness was calculated from densitometric measurements of phr1and normalized against the 28S rRNA signal, in the linear range (0 to0.6 of maximum) (see Fig. 7). Bars are means with standard errors(SE) for two to six replicates. The photoreactivation action spectrum isshown schematically as a line for comparison.
FIG. 2. Trichoderma phr1 encodes a functional photolyase. E. colilacking CPD photolyase was complemented by phr1. Circles, E. coliSY2 carrying the phr1 coding sequence (Phr1); diamonds, E. coli SY2carrying phr1 cloned in reverse orientation (control). Photoreactiva-tion (L; empty symbols) was induced with white light for 30 min at 30�mol m�2 s�1; dark (D; filled symbols) samples were incubated in totaldarkness following UVC exposure. Each point is the mean of tworeplicates.
1684 BERROCAL-TITO ET AL. EUKARYOT. CELL
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

tion, by reversion of germination inhibition of UVC-inacti-vated spores (41), at different UVC doses. UVC-treated �phr1spores were unable to restore germination after white lighttreatment, while WT spores did (Fig. 4A). As expected, �phr1strains completely lost photoreactivation ability (Fig. 4B). Incontrast, we did not observe an increase in UVC survival ratesfor PHR1-overexpressing strains compared to that for the WT(data not shown), but a significant increase in their photore-activation capacity was detected when either dark- or light-grown spores were assayed under nonsaturating, photoreacti-vating light conditions (Student t test; P 0.05) (Fig. 4C).Other than the loss of photoreactivation, the loss of phr1 inTrichoderma, as in Neurospora (45), conferred no obvious phe-notypes: growth, sporulation, and pigmentation were normal(data not shown).
Light responsiveness in �phr1 mutants and phr1 MC strains.If PHR1, in addition to its CPD photolyase function, has anyphotosensory role, phr1 deletion or overexpression could mod-ify Trichoderma light responsiveness. Therefore, we tested phr1photoinduction in phr1 null and MC strains. For this purpose,in �phr1 strains, the phr1 coding region was replaced with theGFP coding region (Fig. 3A). GFP was inserted immediatelyafter the 2-kb left flank of the construct, which is the regionending at the predicted translation start codon (5). GFP pho-toinduction in the �phr1 strains 3.20 and 3.3 shows that uponreplacement, the upstream region is still able to confer lightregulation on the reporter gene (Fig. 5A). PHR1, therefore, isnot required for its own induction. In an ectopic integrant, thephotoinduction of the resident and transgenic genes was tightlycorrelated (Fig. 5B), indicating that all the regulatory elements
FIG. 3. Construction of transgenic strains. (A) Strategy for replacement of phr1. The construct contains the upstream region of the phr1 codingsequence (5� flank) ligated to the gfp coding region, followed by the hph gene under the control of the trpC promoter and, finally, by the downstreamregion of phr1 (3� flank). The arrow represents the coding region. (B) Identification of phr1-deleted strains (�phr1) by PCR analysis oftransformants. Amplification of phr1 and markers was assayed with genomic DNAs from the indicated strains. 3.20 and 3.3, �phr1 strains; 3.21,ectopic integration strain. (C) Plasmid pT3T7phr1 5.8 map used for obtaining MC strains. Relevant restriction enzyme sites are shown. Black,plasmid region; gray, phr1 clone. (D) Identification of overexpresser strains (MC) by Southern analysis of the phr1 transformants. A blot of SalIdigests of genomic DNAs from the indicated strains was probed with the 5.8-kb phr1 PstI fragment. The 4.8-kb bands correspond to the WT gene,and 3.6-kb bands correspond to the exogenous copy. The 1.2-kb fragment is present in both digests. The densitometric ratio of the 3.6-kb to 4.8-kbbands shows that strains MC6 and MC7 have two extra copies and strain MC8 has one extra copy. V contains the cotransformation vector.(E) PHR1 levels in transgenic strains were determined by Western blotting (top). Ponceau staining is shown as a loading control (bottom). Dark,samples grown in total darkness; light, samples were frozen 30 min after a saturating exposure to blue light (540 �mol m�2).
VOL. 6, 2007 AUTOREGULATION OF FUNGAL PHOTOLYASE 1685
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

are contained within the 2-kb upstream region represented bythe left flank of the construct. Time courses for GFP lightinduction in �phr1 strains and for phr1 photoinduction in MCstrains were similar in general to that for the WT. In the decayphase, however, the time course was altered (Fig. 5C). While in
FIG. 4. Photoreactivation of spores from transgenic lines. (A) Lossof photoreactivation in �phr1 mutant line. One-week-old spores fromthe WT and a �phr1 mutant grown in constant light at 30 �mol m�2
s�1 were either light treated or not, kept in darkness for 16 h, andexamined for germination. The percentage of spore germination rep-resents the percentage of photoreactivation. Control, no light treat-ment; UV, after inactivation by 100 J m�2 UVC; UV � L, exposed for30 min to 40 to 60 �mol m�2 white light irradiation immediately afterUVC treatment. (B) Photoreactivation curves from �phr1 3.3 and 3.20mutant lines. Spores were exposed to the indicated amounts of UVC(x axis) and then either incubated in the dark (D) or exposed tophotoreactivating light treatment (L). (C) Photoreactivation of MClines. Seven-day-old spores from either cultures grown with a 16-hphotoperiod at 30 �mol m�2 s�l irradiance or dark-grown spores fromMC strains (MC6, MC7, and MC8) and controls (WT and V) wereassayed for photoreactivation as described previously (41), except thatthe photoreactivating light exposure was 15 min from a cool-white
fluorescent tube at 40 to 60 �mol m�2 s�1. Bars represent means SE(n � 6). Values are as follows: WT, 23.41 6.17; V, 29.73 7.97;MC6, 47.82 8.32; MC7, 49.72 7.51; and MC8, 59.46 10.65.
FIG. 5. gfp under control of the phr1 promoter is photoinduced inphr1 null strains. (A) Northern analysis was performed on total RNA fromdark-grown (D) or light-induced cultures (L) exposed to a fluence of 540�mol m�2 blue light. 3.3 and 3.20, �phr1 mutants, 3.21, ectopic line.(B) Correlation of gfp and phr1 expression in a cell line carrying both thetransgene and the resident phr1 copy. Each point represents the gfp signal(x axis) and phr1 signal (y axis) of a single RNA sample. (C) Time courseof photoinduction in transgenic lines. Northern analysis was performed ontotal RNA from dark-grown (0) or light-induced cultures exposed to afluence of 540 �mol m�2 blue light. Samples were collected 0, 5, 15, 30,60, and 120 min after the pulse. The phr1 signal value was normalizedagainst the 28S signal. The curves of phr1 induction for the WT and thetransgenic lines are shown. MC6 and MC7 are MC strains, and 3.3 and3.20 are null mutants.
1686 BERROCAL-TITO ET AL. EUKARYOT. CELL
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

the WT and overexpression strains the expression levels at 120min decreased to 25% of the peak, in �phr1 strains theexpression levels were still 65 to 66% of the maximum. Thedeletion strains may have slow decay or may have lost adap-tation, or the stability of the gfp mRNA may be greater thanthat of the endogenous transcript.
The light-regulated GFP was used to test whether PHR1makes any contribution to its own induction. A comparison ofphotoinduction fluence response curves for the gfp transcript in�phr1 strains and for phr1 in WT strains shows that the lightrequirement for half-saturation (k1/2) of �phr1 strains wasshifted to lower values than those for either WT or ectopicstrains (Fig. 6B; Table 1). For individual strains, k1/2 valueswere 13.6 �mol m�2 for strain 3.20 and 19.3 �mol m�2 forstrain 3.3, compared to 32.1 �mol m�2 for GFP transcripts inthe ectopic strain 3.21 and 82.4 �mol m�2 for phr1 transcriptsin the WT. The phr1 promoter light sensitivity was increasedfivefold in the �phr1 strains compared to that of the WT.
Quantitative reverse transcription-PCR was also performed(Fig. 6C), and the exponential fit for deleted strains clearlyshows a shift to lower fluences than those for the WT; however,the shift in light sensitivity appears to be somewhat smallerthan the one deduced by quantification of the hybridizationsignals on Northern blots. k1/2 values for this experiment wereas follows: for �phr1 strains 3.20 and 3.3, 20.45 �mol m�2; andfor the WT, 43.38 �mol m�2.
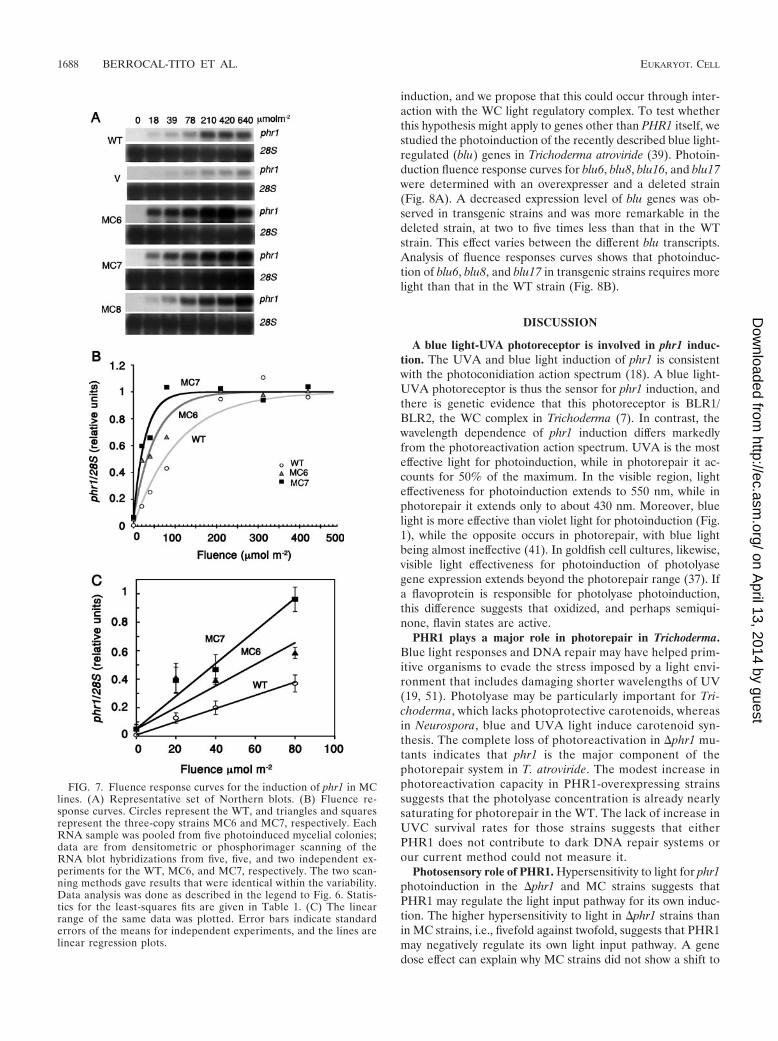
We then compared the fluence response curves for phr1induction in WT and MC strains (Fig. 7A). The k1/2 values ofMC strains were shifted to lower fluences than those for eitherthe WT or the control strain (Fig. 7B; Table 1) carrying onlythe cotransformation vector. The k1/2 value for a strain carryinga copy of the cotransformation vector, 79.6 �mol m�2, wassimilar to that for the WT (82.4 �mol m�2). The strains withthe highest copy number, MC6 and MC7, require less light forphr1 induction than the control, with k1/2 values of 48.2 and32.0 �mol m�2, respectively (Table 1). Analysis of the linearrange of the fluence response curves by linear regression alsoconfirmed that this difference is significant (Fig. 7C). Thesestrains have increased sensitivity to light for phr1 induction, buttheir change in light requirement was less than that for �phr1strains.
These results suggested that PHR1 autoregulates its photo-
FIG. 6. Light sensitivity for induction of gfp under control of thephr1 promoter in �phr1 strains. (A) Northern blot representative ofthe data used to construct the graph in panel B. The fluence of thelight pulse is indicated for each lane. (B) Fluence response curves.Each RNA sample was from one or two mycelial colonies; data arefrom phosphorimager scanning of the RNA blot hybridizations. Thephr1 and gfp signal values were normalized against the rRNA signal,and the data were then normalized for overall changes in hybrid-ization intensity between experiments by dividing them by the meanof the three highest fluence points. Black squares indicate meanswith SE for three independent experiments with two �phr1 lineseach (separate data for each are also indicated). The lines arenonlinear least-squares fits to an exponential model, calculated asdescribed previously (5). Statistics for the least-squares fits aregiven in Table 1. The black line is the fit to the combined �phr1data, and the gray line is a least-squares fit to phr1 fluence responsecurves from three independent experiments on the WT. (C) Real-time PCR data for RNA samples from the same experiments. They axis indicates the transcript abundance of phr1 (WT) or gfp(�phr1) relative to the abundance of gpdh transcript in the samesamples and to the signal at the highest fluence, calculated as 2���CT.
TABLE 1. k1/2 values for WT and different transgenic linesa
StrainNo. ofphr1
copies
k1/2(�mol m�2)
Error(�)
Error(�) df
WT 1 82.4 6.65 6.18 23V 1 79.6 8.00 7.31 12MC6 3 48.2 4.82 4.40 26MC7 3 32.0 2.56 2.37 13MC8 2 77.56 9.26 8.29 5�phr1/phr-gfp 0 15.1 1.52 1.42 39
a k1/2 values were calculated from least-square fits to an exponential saturationmodel (as in Fig. 6 and 7). The errors for k1/2 values were calculated (by inversionof the exponential function used to fit the data) from the estimated errors of theexponential function, determined as standard errors from the means of theresiduals, with df degrees of freedom. V, transformant with cotransformationvector only; MC, multicopy transformants for phr1, as indicated in the secondcolumn. �phr1/phr-gfp indicates gene replacement of phr1 by gfp, and the dataare for gfp transcript levels.
VOL. 6, 2007 AUTOREGULATION OF FUNGAL PHOTOLYASE 1687
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from
induction, and we propose that this could occur through inter-action with the WC light regulatory complex. To test whetherthis hypothesis might apply to genes other than PHR1 itself, westudied the photoinduction of the recently described blue light-regulated (blu) genes in Trichoderma atroviride (39). Photoin-duction fluence response curves for blu6, blu8, blu16, and blu17were determined with an overexpresser and a deleted strain(Fig. 8A). A decreased expression level of blu genes was ob-served in transgenic strains and was more remarkable in thedeleted strain, at two to five times less than that in the WTstrain. This effect varies between the different blu transcripts.Analysis of fluence responses curves shows that photoinduc-tion of blu6, blu8, and blu17 in transgenic strains requires morelight than that in the WT strain (Fig. 8B).
DISCUSSION
A blue light-UVA photoreceptor is involved in phr1 induc-tion. The UVA and blue light induction of phr1 is consistentwith the photoconidiation action spectrum (18). A blue light-UVA photoreceptor is thus the sensor for phr1 induction, andthere is genetic evidence that this photoreceptor is BLR1/BLR2, the WC complex in Trichoderma (7). In contrast, thewavelength dependence of phr1 induction differs markedlyfrom the photoreactivation action spectrum. UVA is the mosteffective light for photoinduction, while in photorepair it ac-counts for 50% of the maximum. In the visible region, lighteffectiveness for photoinduction extends to 550 nm, while inphotorepair it extends only to about 430 nm. Moreover, bluelight is more effective than violet light for photoinduction (Fig.1), while the opposite occurs in photorepair, with blue lightbeing almost ineffective (41). In goldfish cell cultures, likewise,visible light effectiveness for photoinduction of photolyasegene expression extends beyond the photorepair range (37). Ifa flavoprotein is responsible for photolyase photoinduction,this difference suggests that oxidized, and perhaps semiqui-none, flavin states are active.
PHR1 plays a major role in photorepair in Trichoderma.Blue light responses and DNA repair may have helped prim-itive organisms to evade the stress imposed by a light envi-ronment that includes damaging shorter wavelengths of UV(19, 51). Photolyase may be particularly important for Tri-choderma, which lacks photoprotective carotenoids, whereasin Neurospora, blue and UVA light induce carotenoid syn-thesis. The complete loss of photoreactivation in �phr1 mu-tants indicates that phr1 is the major component of thephotorepair system in T. atroviride. The modest increase inphotoreactivation capacity in PHR1-overexpressing strainssuggests that the photolyase concentration is already nearlysaturating for photorepair in the WT. The lack of increase inUVC survival rates for those strains suggests that eitherPHR1 does not contribute to dark DNA repair systems orour current method could not measure it.
Photosensory role of PHR1. Hypersensitivity to light for phr1photoinduction in the �phr1 and MC strains suggests thatPHR1 may regulate the light input pathway for its own induc-tion. The higher hypersensitivity to light in �phr1 strains thanin MC strains, i.e., fivefold against twofold, suggests that PHR1may negatively regulate its own light input pathway. A genedose effect can explain why MC strains did not show a shift to
FIG. 7. Fluence response curves for the induction of phr1 in MClines. (A) Representative set of Northern blots. (B) Fluence re-sponse curves. Circles represent the WT, and triangles and squaresrepresent the three-copy strains MC6 and MC7, respectively. EachRNA sample was pooled from five photoinduced mycelial colonies;data are from densitometric or phosphorimager scanning of theRNA blot hybridizations from five, five, and two independent ex-periments for the WT, MC6, and MC7, respectively. The two scan-ning methods gave results that were identical within the variability.Data analysis was done as described in the legend to Fig. 6. Statis-tics for the least-squares fits are given in Table 1. (C) The linearrange of the same data was plotted. Error bars indicate standarderrors of the means for independent experiments, and the lines arelinear regression plots.
1688 BERROCAL-TITO ET AL. EUKARYOT. CELL
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

higher fluences in photoinduction. Another explanation couldbe that PHR1 has a more complex role than a simple negativeregulator of its light input pathway.
The molecular mechanism by which PHR1 could modulate
its own expression is unknown. Photolyase binds undamagedDNA with a low affinity, but the possibility of direct interactionwith regulatory sequences, including those in its own promoter,must be discarded due to the lack of specificity needed for theinduction of particular genes. The region upstream of phr1 (5)contains elements, such as APE and GATX boxes, similar tothose found in the Neurospora gene promoters that are regu-lated by light through the light-responsive transcriptional acti-vators WC1 and WC2 (12, 15, 22, 47). The Trichoderma BLR1and BLR2 proteins (WC1 and WC2 homologs) are requiredfor photoinduction of phr1. A BLR1 and BLR2 complex is thuslikely to be the photoreceptor providing the major componentof blue light and UVA regulation of phr1 (7). PHR1 mightinteract with the BLR complex to negatively regulate its ownexpression. The decreased expression levels of blu genes andtheir decreased sensitivity to blue light in phr1-overexpressingand phr1-deleted strains could also be explained by an inter-action of PHR1 with the WC complex. Such a PHR1-WCcomplex interaction might be reflected in repression of thephr1 promoter in the dark and by a positive action on photo-induction of blu transcript accumulation.
Is the same PHR1 functional in photorepair and photoregu-lation? The difference in action spectra for photoreactivationand photolyase induction and the fact that photoreactivationrequires 3 orders of magnitude more light than photoresponseled us to postulate that a specific biochemical state of PHR1provides a photosensory function. Light effectiveness for pho-torepair extends to 430 nm, while in photoresponse it extendsto 550 nm. If a flavin from PHR1 is involved somehow inphotolyase photoinduction, this difference suggests that oxi-dized, and perhaps semiquinone, flavin states are more impor-tant for photoinduction than the reduced state privileged forphotorepair. The MTHF chromophore seems dispensable forE. coli photolyase activity, while it appears to be more impor-tant in V. cholerae cryptochrome function. MTHF is stablyattached in VcCry1, and moreover, the energy transfer fromMTHF to FAD in VcCry1 is four times faster than that for E.coli photolyase (42). The MTHF chromophore of PHR1 mighttherefore have a more important role in photosensing than inphotorepair.
Evolutionary and structural considerations for a photolyasephotosensory role. The ancestral gene of the photolyase-pho-toreceptor family is CPD photolyase. Considering that 6-4photolyases from animals and plants are very close to animalcryptochromes, it was proposed that the ancestors of plant andanimal cryptochromes diverged before plants diverged fromanimals (26, 31). In this evolutionary model, plant and animalcryptochromes arose independently by repeated evolution (8).Evolution of the circadian clock would have led to a time-keeping mechanism in which light input is controlled by cryp-tochrome, even in large metazoans that were at less direct riskof UV damage. Finally, in mammals, cryptochrome becamepart of the clock oscillator, having a central but light-indepen-dent role in the negative feedback loop of the clock (20). Therecent identification of prokaryotic cryptochromes belongingto the DASH group has brought up to date the initial propo-sition that cryptochromes diverged from photolyases beforethe origin of eukaryotes (6, 28, 49). Thus, of the five mainsubfamilies, the cluster including 6-4 photolyases and animalcryptochromes and the new DASH cryptochrome group have
FIG. 8. blu gene photoinduction is altered in phr1-overexpressingand -deleted strains. (A) Photoinduction fluence response assays ofphr1, blu6, blu8, blu16, and blu17 were carried out with the overex-presser strain MC7, the �phr1 knockout strain 3.20, and the WT strainby Northern blot hybridization. The fluence of the light pulse is indi-cated for each lane. (B) Fluence response curves. Data are fromdensitometric scanning of the RNA blot hybridizations, normalizedusing gpdh transcript levels in the same blots.
VOL. 6, 2007 AUTOREGULATION OF FUNGAL PHOTOLYASE 1689
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

both photorepair and sensory functions. The plant crypto-chrome cluster, in contrast, has only sensory functions, whilethe class I and class II CPD photolyases are known only to haveDNA repair functions. The structure of cryptochromes resem-bles that of photolyases, and indeed, a recently determined cryp-tochrome structure (6) is similar to that of E. coli photolyase.Thus, it would be surprising, from the evolutionary point of view,if photolyase has no protein interaction partners. DrosophiladCRY exhibits a strong, light-dependent interaction with its part-ner in circadian transcriptional regulation, TIMELESS (9); theability to interact with another member of the transcriptionalregulatory complex persists (and becomes independent of light)when the C-terminal extension is deleted (40), leaving a proteinwhich should have virtually the same structure as photolyase.
The N-terminal “tail” of fungal photolyases as a putativephotosensory domain. A C-terminal domain extending 30 to200 amino acids beyond the region of high homology withphotolyases characterizes most cryptochromes and seems to beimportant for nucleus-cytosol trafficking and protein-proteininteraction. The C-terminal “tail” is sufficient to mimic theaction of light in plants (50). In Drosophila cryptochrome, it isresponsible for light sensitivity, as loss of the C-terminal “tail”results in a constitutive “on” phenotype with respect to itsability to bind to its partner(s) in the circadian transcriptionalregulator complex (40). Fungal photolyases and DASH cryp-tochromes lack this specific additional sequence, but they doextend 70 to 140 amino acids towards the N terminus. In fungalphotolyases, this N-terminal extension contains the putativemitochondrial and nuclear localization signals, while in Cry3, aDASH cryptochrome, it contains a transit peptide for localiza-tion in chloroplasts and mitochondria. This N-terminal exten-sion might confer some photosensory properties (Fig. 9A).Alignment of the N-terminal 140 amino acids of visible light-regulated fungal photolyases, namely, those of T. atroviride (5),N. crassa (Berrocal-Tito, unpublished results), and Fusarium
oxyosporum (2), shows that this region is very well conserved(Fig. 9B). A standard BLAST analysis of this N-terminal re-gion of T. atroviride photolyase does not show any specifichomology, while an RPSI-BLAST analysis against theBLOCKS database (25) shows significant alignments with pre-senilin, amphiphysin, and Sac3/GANP proteins, which have incommon a protein binding ability, suggesting an interactingrole for this region. In addition, putative phosphorylation sitesfor either cyclic AMP- or cyclic GMP-dependent protein ki-nase, protein kinase C, and casein kinase II are found bycomparison to the PROSITE database, at amino acid positions44 to 48 of PHR1. In class I photolyases, light excites MTHF,
FIG. 10. Photolyase light-dependent autoregulation model. Cir-cles, blue light regulators (BRL) 1 and 2; gray box and solid line, phr1promoter and coding regions; black and gray polygons, DNA photol-yase (PHR1) with different photochemical properties; rectangle, pu-tative repressor; lightning, exogenous light. Photolyase would act as amodulator of its own transcription through interaction with the BRLcomplex, recruiting a repressor to the complex in the dark. Lightrelieves repression by an unknown mechanism, resulting in activationof phr1 transcription, which is dependent on BLR1 and BLR2. Inaddition, light activates PHR1 for DNA repair.
FIG. 9. Comparison of N-terminal regions of light-regulated fungal photolyases. (A) Schematic comparison of fungal photolyases andcryptochromes. (B) Alignment of the 140 N-terminal amino acids of light-regulated fungal photolyases. The underlined region of PHR1 showshomology with presenilin and amphiphysin, and the heavy line indicates a phosphorylation site. E. c., E. coli; At, A. thaliana; Nc, N. crassa; Fo, F.oxyosporum; Ta, T. atroviride.
1690 BERROCAL-TITO ET AL. EUKARYOT. CELL
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

and this energy excitation is transferred to FAD, perhaps pro-moting a conformational change in PHR1 that could result inN-terminal modification, for example, phosphorylation.
To explain the findings reported here, we propose the fol-lowing model (Fig. 10), which can be tested by further molec-ular genetic and protein interaction experiments. In thismodel, PHR1 interacts with or is part of a transcriptionalregulatory complex. To be part of this complex, PHR1 musthave specific photochemical properties, resulting, for example,from a posttranslational modification of the N terminus. PHR1present in the dark would exert an inhibitory effect on photo-induction by recruiting an inhibitory protein to the complex;light relieves this repression. This would be completely re-moved in null mutants, leading to the observed increase in lightsensitivity. A weak gfp hybridization signal was detected evenin the dark in some, but not all, Northern blots (just visible inFig. 6A) and was more significant in real-time PCR experi-ments (Fig. 6C). This suggests that a basal level of PHR1might enhance binding of the repressor already present inthe dark. Overexpression might lead to accumulation ofexcess free PHR1 in the nucleus, partially titrating away theinhibitory protein. Thus, the light-dependent interaction ofcryptochrome/photolyase family members with transcrip-tional regulators may predate the divergence of crypto-chromes from photolyases.
ACKNOWLEDGMENTS
We are grateful to Akira Yasui for the E. coli photolyase mutantSY2, to A. P. Eker for antiserum to Neurospora photolyase, to T.Rosales-Saavedra and R. Hadar for preparation of some RNA andprotein samples, and to K. Florian Storch and Charles J. Weitz forhelpful suggestions during early stages of this work.
This study was supported in part by grants from FDI-AT11,CORFO—Chile, and ICGEB (CRP/MEX99-02) to A.H.E. and fromthe Israel Academy of Sciences to B.A.H.
REFERENCES
1. Ahmad, M., and A. R. Cashmore. 1993. HY4 gene of A. thaliana encodes aprotein with characteristics of a blue-light photoreceptor. Nature 366:162–166.
2. Alejandre-Duran, E., T. Roldan-Arjona, R. R. Ariza, and M. RuizRubio.2003. The photolyase gene from the plant pathogen Fusarium oxyosporum f.sp. lycopersici is induced by visible light and �-tomatine from tomato plant.Fungal Genet. Biol. 40:159–165.
3. Ballario, P., P. Vittorioso, A. Magrelli, C. Talora, A. Cabibbo, and G.Macino. 1996. White collar-1, a central regulator of blue light responses inNeurospora, is a zinc finger protein. EMBO J. 15:1650–1657.
4. Berrocal-Tito, G., T. Rosales-Saavedra, A. Herrera-Estrella, and B. Horwitz.2000. Characterization of blue-light and developmental regulation of thephotolyase gene phr1 in Trichoderma harzianum. Photochem. Photobiol.71:662–668.
5. Berrocal-Tito, G., L. Sametz-Baron, K. Eichenberg, B. A. Horwitz, and A.Herrera-Estrella. 1999. Rapid light regulation of a Trichoderma harzianumDNA photolyase gene. J. Biol. Chem. 274:14288–14294.
6. Brudler, R., K. Hitomi, H. Daiyasu, H. Toh, K. Kucho, M. Ishiura, M.Kanehisa, V. A. Roberts, T. Todo, J. A. Tainer, and E. D. Getzoff. 2003.Identification of a new cryptochrome class. Structure, function, and evolu-tion. Mol. Cell 11:59–67.
7. Casas-Flores, S., M. Rios-Momberg, M. Bibbins, P. Ponce-Noyola, and A.Herrera-Estrella. 2004. BLR-1 and BLR-2, key regulatory elements of pho-toconidiation and mycelial growth in Trichoderma atroviride. Microbiology150:3561–3569.
8. Cashmore, A. R., J. A. Jarillo, Y. J. Wu, and D. Liu. 1999. Cryptochromes:blue light receptors for plants and animals. Science 284:760–765.
9. Ceriani, M. F., T. K. Darlington, D. Staknis, P. Mas, A. A. Petti, C. J. Weitz,and S. A. Kay. 1999. Light-dependent sequestration of TIMELESS byCRYPTOCHROME. Science 285:553–556.
10. Christie, J. M., M. Salomon, K. Nozue, M. Wada, and W. R. Briggs. 1999.LOV (light, oxygen, or voltage) domains of the blue-light photoreceptorphototropin (nph1): binding sites for the chromophore flavin mononucle-otide. Proc. Natl. Acad. Sci. USA 96:8779–8783.
11. Corrochano, L. M., and E. Cerda-Olmedo. 1990. Photomorphogenesis inbehavioral and color mutants of Phycomyces. J. Photochem. Photobiol. B6:325–336.
12. Crosthwaite, S., J. Dunlap, and J. Loros. 1997. Neurospora wc-1 and wc-2:transcription, photoresponses, and the origins of circadian rhythmicity. Sci-ence 276:763–769.
13. Daiyasu, H., T. Ishikawa, K.-I. Kuma, S. Iwai, T. Todo, and H. Toh. 2004.Identification of cryptochrome DASH from vertebrates. Genes Cells 9:479–495.
14. Emery, P., W. V. So, M. Kaneko, J. C. Hall, and M. Rosbash. 1998. CRY, aDrosophila clock and light-regulated cryptochrome, is a major contributor tocircadian rhythm resetting and photosensitivity. Cell 95:669–679.
15. Froehlich, A. C., Y. Liu, J. J. Loros, and J. C. Dunlap. 2002. White Collar-1,a circadian blue light photoreceptor, binding to the frequency promoter.Science 297:815–819.
16. Galagan, J. E., S. E. Calvo, K. A. Borkovich, E. U. Selker, N. D. Read, D.Jaffe, W. FitzHugh, L. J. Ma, S. Smirnov, S. Purcell, B. Rehman, T. Elkins,R. Engels, S. Wang, C. B. Nielsen, J. Butler, M. Endrizzi, D. Qui, P. Ianak-iev, D. Bell-Pedersen, M. A. Nelson, M. Werner-Washburne, C. P. Selitren-nikoff, J. A. Kinsey, E. L. Braun, A. Zelter, U. Schulte, G. O. Kothe, G. Jedd,W. Mewes, C. Staben, E. Marcotte, D. Greenberg, A. Roy, K. Foley, J. Naylor,N. Stange-Thomann, R. Barrett, S. Gnerre, M. Kamal, M. Kamvysselis, E.Mauceli, C. Bielke, S. Rudd, D. Frishman, S. Krystofova, C. Rasmussen,R. L. Metzenberg, D. D. Perkins, S. Kroken, C. Cogoni, G. Macino, D.Catcheside, W. Li, R. J. Pratt, S. A. Osmani, C. P. DeSouza, L. Glass, M. J.Orbach, J. A. Berglund, R. Voelker, O. Yarden, M. Plamann, S. Seiler, J.Dunlap, A. Radford, R. Aramayo, D. O. Natvig, L. A. Alex, G. Mannhaupt,D. J. Ebbole, M. Freitag, I. Paulsen, M. S. Sachs, E. S. Lander, C. Nusbaum,and B. Birren. 2003. The genome sequence of the filamentous fungus Neu-rospora crassa. Nature 422:859–868.
17. Galland, P., and H. Senger. 1988. The role of pterins in the photoreceptionand metabolism of plants. Photochem. Photobiol. 48:811–820.
18. Gressel, J., and K. M. Hartmann. 1968. Morphogenesis in Trichoderma:action spectrum of photoinduced sporulation. Planta 79:271–274.
19. Gressel, J., and W. Rau. 1983. Photocontrol of fungal development, p.603–639. In W. Shropshire, Jr., and H. Mohr (ed.), Encyclopedia of plantphysiology. Springer-Verlag, Berlin-Heidelberg, Germany.
20. Griffin, E. A., Jr., D. Staknis, and C. J. Weitz. 1999. Light-independent roleof CRY1 and CRY2 in the mammalian circadian clock. Science 286:768–771.
21. Guo, H., H. Yang, T. C. Mockler, and C. Lin. 1998. Regulation of floweringtime by Arabidopsis photoreceptors. Science 279:1360–1363.
22. He, Q., P. Cheng, Y. Yang, L. Wang, K. H. Gardner, and Y. Liu. 2002. Whitecollar-1, a DNA binding transcription factor and a light sensor. Science297:840–843.
23. Heintzen, C., J. J. Loros, and J. C. Dunlap. 2001. The PAS protein VIVIDdefines a clock-associated feedback loop that represses light input, modu-lates gating, and regulates clock resetting. Cell 104:453–464.
24. Reference deleted.25. Henikof, S., and J. Henikof. 1994. Protein family classification based on
searching a database of blocks. Genomics 19:97–107.26. Hitomi, K., K. Okamoto, H. Daiyasu, H. Miyashita, S. Iwai, H. Toh, M.
Ishiura, and T. Todo. 2000. Bacterial cryptochrome and photolyase: charac-terization of two photolyase-like genes of Synechocystis sp. PCC6803. NucleicAcids Res. 28:2353–2362.
27. Horwitz, B. A., and G. M. Berrocal Tito. 1997. A spectroscopic view of somerecent advances in the study of blue light photoreception. Bot. Acta 110:360–368.
28. Kanai, S., R. Kikuno, H. Toh, H. Ryo, and T. Todo. 1997. Molecular evolu-tion of the photolyase-blue-light photoreceptor family. J. Mol. Evol. 45:535–548.
29. Kleine, T., P. Lockhart, and A. Batschauer. 2003. An Arabidopsis proteinclosely related to Synechocystis cryptochrome is targeted to organelles. PlantJ. 35:93–103.
30. Kleiner, O., S. Kircher, K. Harter, and A. Batschauer. 1999. Nuclear local-ization of the Arabidopsis blue light receptor cryptochrome 2. Plant J. 19:289–296.
31. Kobayashi, Y., T. Ishikawa, J. Hirayama, H. Daiyasu, S. Kanai, H. Toh, I.Fukuda, T. Tsujimura, N. Terada, Y. Kamei, S. Yuba, S. Iwai, and T. Todo.2000. Molecular analysis of zebrafish photolyase/cryptochrome family: twotypes of cryptochromes present in zebrafish. Genes Cells 5:725–738.
32. Lin, C., M. Ahmad, D. Gordon, and A. Cashmore. 1995. Expression of anArabidopsis cryptochrome gene in transgenic tobacco results in hypersensi-tivity to blue, UV-A and green light. Proc. Natl. Acad. Sci. USA 92:8423–8427.
33. Linden, H., and G. Macino. 1997. White collar 2, a partner in blue-lightsignal transduction, controlling expression of light-regulated genes in Neu-rospora crassa. EMBO J. 16:98–109.
34. Lipson, E. D., and B. A. Horwitz. 1991. Photosensory receptors and theirtransduction, p. 1–64. In B. Satir (ed.), Sensory receptors and signal trans-duction, modern cell biology, vol. 7. Alan R. Liss, New York, NY.
35. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expression
VOL. 6, 2007 AUTOREGULATION OF FUNGAL PHOTOLYASE 1691
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from

data using real-time quantitative PCR and the 2���CT method. Methods25:402–408.
36. Lorito, M., C. K. Hayes, A. D. Pietro, and G. E. Harman. 1993. Biolistictransformation of Trichoderma harzianum and Gliocladium virens usingplasmid and genomic DNA. Curr. Genet. 24:349–356.
37. Mitani, H., N. Uchida, and A. Shima. 1996. Induction of cyclobutane pyrim-idine dimer photolyase in cultured fish cells by UVA and blue light. Photo-chem. Photobiol. 64:943–948.
38. Ozgur, S., and A. Sancar. 2003. Purification and properties of humanblue-light photoreceptor cryptochrome 2. Biochemistry 42:2926–2932.
39. Rosales-Saavedra, T., E. U. Esquivel-Naranjo, S. Casas-Flores, P. Martınez-Hernandez, E. Ibarra-Laclette, C. Cortes-Penagos, and A. Herrera-Estrella.2006. Novel light-regulated genes in Trichoderma atroviride: a dissection bycDNA microarrays. Microbiology 152:3305–3317.
40. Rosato, E., V. Codd, G. Mazzotta, A. Piccin, M. Zordan, R. Costa, and C. P.Kyriacou. 2001. Light-dependent interaction between Drosophila CRY andthe clock protein PER mediated by the carboxy terminus of CRY. Curr. Biol.11:909–917.
41. Sametz-Baron, L., G. Berrocal, T. R. Amit, A. Herrera-Estrella, and B. A.Horwitz. 1997. Photoreactivation of UV-inactivated spores of Trichodermaharzianum. Photochem. Photobiol. 65:849–854.
42. Saxena, C., H. Wang, I. H. Kavakli, A. Sancar, and D. Zhong. 2005. Ultrafastdynamics of resonance energy transfer in cryptochrome. J. Am. Chem. Soc.127:7984–7985.
43. Schwerdtfeger, C., and H. Linden. 2003. VIVID is a flavoprotein and servesas a fungal blue light photoreceptor for photoadaptation. EMBO J. 22:4846–4855.
44. Selby, C. P., C. Thompson, T. M. Schmitz, R. N. Van Gelder, and A. Sancar.2000. Functional redundancy of cryptochromes and classical photoreceptorsfor nonvisual ocular photoreception in mice. Proc. Natl. Acad. Sci. USA97:14697–14702.
45. Shimura, M., Y. Ito, C. Ishii, H. Yajima, H. Linden, T. Harashima, A. Yasui,and H. Inoue. 1999. Characterization of a Neurospora crassa photolyase-deficient mutant generated by repeat induced point mutation of the phrgene. Fungal Genet. Biol. 28:12–20.
46. Somers, D. E., P. F. Devlin, and S. A. Kay. 1998. Phytochromes and cryp-tochromes in the entrainment of the Arabidopsis circadian clock. Science282:1488–1490.
47. Talora, C., L. Franchi, H. Linden, P. Ballario, and G. Macino. 1999. Role ofa white collar-1-white collar-2 complex in blue-light signal transduction.EMBO J. 18:4961–4968.
48. van der Horst, G. T., M. Muijtjens, K. Kobayashi, R. Takano, S. Kanno, M.Takao, J. de Wit, A. Verkerk, A. P. Eker, D. van Leenen, R. Buijs, D.Bootsma, J. H. Hoeijmakers, and A. Yasui. 1999. Mammalian Cry1 and Cry2are essential for maintenance of circadian rhythms. Nature 398:627–630.
49. Worthington, E. N., I. H. Kavakli, G. Berrocal-Tito, B. E. Bondo, and A.Sancar. 2003. Purification and characterization of three members of thephotolyase/cryptochrome family blue-light photoreceptors from Vibrio chol-erae. J. Biol. Chem. 278:39143–39154.
50. Yang, H.-Q., Y.-J. Wu, R.-H. Tang, D. Liu, Y. Liu, and A. Cashmore. 2000.The C termini of Arabidopsis cryptochromes mediate a constitutive lightresponse. Cell 103:815–827.
51. Yasuhira, S., and A. Yasui. 1992. Visible light-inducible photolyase genefrom the goldfish Carassius auratus. J. Biol. Chem. 267:25644–25647.
1692 BERROCAL-TITO ET AL. EUKARYOT. CELL
on April 13, 2014 by guest
http://ec.asm.org/
Dow
nloaded from




















