
The Context and Associations of Monument 3 from La Blanca, Guatemala
Upload
independentCategory
view
1download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Ichthyoplankton spatial pattern in the inner shelf off Bahıa Blanca Estuary, SWAtlantic Ocean
Hoffmeyer Monica Susanaa,b,*, Menendez Marıa Clara a, Biancalana Florencia a, Nizovoy Alicia Mabel c,Torres Eduardo Ramon a,d
a Instituto Argentino de Oceanografıa-IADO (CONICET-UNS), Camino La Carrindanga km 7.5, B8000FWB Bahıa Blanca, Buenos Aires, Argentinab Facultad Regional Bahıa Blanca, Universidad Tecnologica Nacional, 11 de Abril 461, B8000LMI, Bahıa Blanca, Buenos Aires, Argentinac Centro Austral de Investigaciones Cientıficas y Tecnicas, B. Houssay 200, V9410CAB Ushuaia, Tierra del Fuego, Antartida e Islas del Atlantico Sur, Argentinad Universidad Nacional de la Patagonia Austral, Av. Lisandro de la Torre 860, Z9400JZR Rıo Gallegos, Santa Cruz, Argentina
a r t i c l e i n f o
Article history:Received 17 December 2008Accepted 10 July 2009Available online 25 July 2009
Keywords:fish eggsfish larvaedistributionzooplanktonfrontsArgentina
a b s t r a c t
This study focuses on the composition, abundance and distribution of ichthyoplankton in the inner shelfarea off Bahıa Blanca Estuary on the SW Atlantic Ocean during late spring. Eggs and larvae of Brevoortiaaurea, Engraulis anchoita, Parona signata, Sciaenidae spp. – such as Cynoscion guatucupa and Micro-pogonias furnieri –, and Odontesthes argentinensis were found. Species richness was low probably asa result of season and shallow depths. Ichthyoplankton abundance reached values close to 10 000 per10 m�3 (eggs) and 4000 per 10 m�3 (larvae) and displayed a spatial distribution pattern with maximumabundance values restricted to a band parallel to the coast. Differences between egg and larval patterns,probably derived from a different displacement and hydrodynamic behavior, were observed. Egg andlarvae distribution patterns were found related with spawning areas and to directly depend on salinityand mesozooplankton. The larvae distribution pattern, in particular, was found to inversely depend onparticulate organic carbon. In addition, the geographic location of egg and larvae maxima stronglycoincided with a saline front reported for this area in springtime, thus suggesting a direct relationshipwith it.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Variability in biotic and abiotic factors is crucial to ichthyo-plankton spatial and temporal distribution and abundance incoastal zones (Grioche and Koubbi, 1997; Reiss and McConaugha,1999; Espinosa-Fuentes and Flores-Coto, 2004). The most impor-tant biotic factors in fish populations are: competition amongindividuals, seasonal abundance of adults and reproductive strat-egies, all of which make fish spawn in areas and at times thatensure food availability (Espinosa-Fuentes and Flores-Coto, 2004).The reproductive behavior of fish populations in relation to envi-ronmental conditions regulates their spatial and seasonal distri-bution patterns and, as a result, those of their eggs and larvae.
Physical features and meteorological events such as circulationpatterns (Nishimoto and Washburn, 2002; Marrari et al., 2004),
river-estuarine plumes and fronts (Grimes and Finucane, 1991;Kingsford and Suthers, 1994), and mixing processes and winds(Sabates et al., 1989; Govoni and Grimes, 1992; Gray, 1996; Sabatesand Olivar, 1996; Simionato et al., 2008) are the main mechanismsof concentration and dispersion of plankton and ichthyoplankton.Likewise, hydrodynamic conditions behave as regulators of thedynamics of coastal aquatic ecosystems and sometimes contributeto determining a cause–effect relationship between physical factorsand the distribution of organisms (Legendre and Demers, 1984;Legendre et al., 1985).
A high number of surveys over the last 40 years along thecontinental shelf of Argentina, at depths ranging from 40 m to theslope, have greatly contributed to learning about ichthyoplanktonand fish population assemblages (Weiss, 1972; Ciechomski andWeiss, 1974; Ciechomski et al., 1979; Ciechomski and Booman,1983; Cassia and Booman, 1985; Cordo, 1986; Sanchez and Cie-chomski, 1995, among others). However, the characteristics ofichthyoplankton in the innermost shelf regions are comparativelyless known.
In the region known as El Rincon off Bahıa Blanca Estuary, whichis, in fact, our study area, an intense reproductive activity of coastal
* Corresponding author at: Instituto Argentino de Oceanografıa-IADO (CONICET-UNS), Camino La Carrindanga km 7.5, B8000FWB Bahıa Blanca, Buenos Aires,Argentina.
E-mail address: [email protected] (M.S. Hoffmeyer).
Contents lists available at ScienceDirect
Estuarine, Coastal and Shelf Science
journal homepage: www.elsevier .com/locate/ecss
0272-7714/$ – see front matter � 2009 Elsevier Ltd. All rights reserved.doi:10.1016/j.ecss.2009.07.017
Estuarine, Coastal and Shelf Science 84 (2009) 383–392

Author's personal copy
fish such as Micropogonias furnieri (whitemouth croaker), Cynoscionguatucupa (stripped weakfish), Parona signata (parona leath-erjacket), Mustelus schmitti (narrownose smoothhound), andScomber japonicus (chub mackerel) has been recorded in spring(Lasta, 1998; Perrota et al., 1999; 2003). It has also been reportedthat the Argentine anchovy (Engraulis anchoita) moves southwardalong the shelf to spawn between late spring and early summer. Asa result, adults, eggs and larvae of this species have been recordedmainly in the eastern, deeper zone of El Rincon (Ciechomski et al.,1979; Pajaro et al., 1998; Marrari et al., 2004). Although thereproductive biology and the distribution of fish populations in thisshelf region have been thoroughly studied (Ciechomski et al., 1979;Lasta, 1998; Pajaro et al., 1998; Perrota et al., 2003), little is knownto date on fish eggs–larvae assemblage distribution.
In view of the above, this study focuses on the composition,abundance and spatial distribution of ichthyoplankton in the innershelf area off Bahıa Blanca Estuary on the SW Atlantic Ocean, duringlate spring. The purpose of this study was to analyze ichthyo-plankton spatial pattern in relation to some chemical and physicalconditions of the water mass. Oceanographic features in the studyarea were also taken into account.
2. Study area
The study area (39�–40
�030 S and 61
�090–62
�030 W) is a shallow
zone with depths lesser than 23 m within the region known as ElRincon (39
�–41
�S,<40 m depth), which is located in the inner shelf
off Bahıa Blanca Estuary on the SW Atlantic Ocean. El Rincon isconnected with Bahıa Blanca Estuary and Anegada Bay on the west(Perillo, 1994) and it receives fresh water from the western andsouthwestern coasts of the Colorado and Negro rivers, respectively,whereas its water column is vertically homogeneous mainly asa result of tidal action and winds (Cuadrado et al., 2002; Acha et al.,2004).
A coastal horizontal saline front forced by mean circulation overthe continental shelf and bathymetry (Martos and Piccolo, 1988;Piola and Rivas, 1997) has been reported for the spring of November1994 (Lasta, 1998; Acha et al., 2004). This front marks the separa-tion between coastal waters diluted by the discharge from theNegro and Colorado rivers (as well as the plumes of Anegada Bayand Bahıa Blanca Estuary) moving to the north and the more salineshelf waters running from San Matıas Gulf towards the northeast(Martos and Piccolo, 1988; Piola and Rivas, 1997; Lasta, 1998).
3. Materials and Methods
3.1. Sampling survey
Sampling was carried out during the AU-02/93 oceanographicsurvey aboard the Argentine vessel ‘‘El Austral’’ on November 1–4,1993. Ichthyoplankton samples were collected using oblique towsfrom around the bottom to the surface with a 0.53 m-open nethaving a 200 mm mesh. These samples were collected continuously(day and night) from a grid of 32 stations separated by 20 km fromeach other on 11 transects perpendicular to the western coast(Fig. 1). A mechanical flow-meter was used to calculate the watervolume filtered by the net. Samples were preserved in 4% bufferedformalin. Surface temperature and salinity (according to PracticalSalinity Scale) were measured in situ using an Inter Ocean CTD.Concentrations of chlorophyll-a, phaeopigments, and particulateorganic carbon (POC) in surface water samples collected from allthe stations, were determined on board with a Beckman spectro-photometer according to Strickland and Parsons (1968) techniques.
3.2. Laboratory and statistical procedures
In the laboratory fish eggs and larvae were sorted with the aid ofa stereomicroscope. They were identified to the lowest possibletaxon following Matsuura and Olivar (1999). Enumeration waseither by total counting or by 20% aliquots depending on thedensity level in the samples. Due to their remarkable similarities,both eggs and larvae of Cynoscion guatucupa and Micropogoniasfurnieri could not be distinguished from each other. Both species arethe main sciaenids in El Rincon region (Lopez Cazorla, 1987; Lasta,1998; Jaureguizar et al., 2004a).
In the absence of significant differences between the samplescollected at night and those collected during the day, all ichthyo-planktonic data were considered as a single dataset. Egg and larvalabundance data were particularly analyzed using multivariateanalysis with PRIMER-E� software (Clarke and Warwick, 1994).They were transformed using the log10 (Xþ 1) function to enhancethe contribution of the less abundant species. HierarchicalAgglomerative Clustering Analysis was carried out using the Bray-Curtis quantitative similarity index and group average linking.Before defining the final cluster of sample data, outliers corre-sponding to three samples (with a 20% or lower degree of similaritywith others, which also represented the lowest abundances) wereremoved to avoid constraints associated with them (Somarakiset al., 2002).
A stepwise Multiple Linear Regression Analysis (MLRA) usingSPSS� software was also carried out on the two abundance datasets(eggs and larvae) to explore the relationships between
Fig. 1. Map of the study area showing the location and labels of sampling stations. Thestations with a black rectangle below correspond to Group I from cluster. The stationswithout a rectangle below correspond to Group II (see Results section).
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392384

Author's personal copy
ichthyoplankton and some biotic and abiotic measured variables.Biotic variables (eggs, larvae and mesozooplankton abundances)were Log10 (xþ 1) transformed before carrying out all the analyses.Egg (EA) and larval abundance (LA) were the dependent variables(Y) whereas temperature (Temp), salinity (Sal), chlorophyll-a (Ch-a),phaeopigments (Phae), particulate organic carbon (POC) and mes-ozooplankton abundance (MA), were the explanatory or predictorvariables (X1.Xj). The latter showed null or low correlationsignificance values (Pearson’s coefficient) among themselves. Thesignificance of the squared multiple correlation coefficients R2 weretested through F statistics as follows:
F ¼R2
Y
1� R2Y
� N � J � 1J
where N is the number of observations¼ 32 and J is the number ofexplanatory variables¼ 6.
Surface data of environmental variables were related with ich-thyoplankton and mesozooplankton abundance data estimatedfrom oblique integrated samples, assuming that the water columnis vertically homogeneous as a result of tidal mixing (Cuadradoet al., 2002). The mesozooplankton collected during the sameoceanographic survey (AU-02/93) has been recently studied byHoffmeyer (unpublished results) and the abundance data from thesame survey were used in the present research.
4. Results
The highest temperatures were recorded at the coastal stationsin the north of the study area (Fig. 2A). Salinity values were lowestin the north of the mouth of Colorado River, giving rise to a saline-diluting zone towards the north as a result of Colorado River’splume (Figs. 1 and 2B). Chlorophyll-a values were highest in thenorth of the mouth of Colorado River and off Anegada Bay,decreasing towards the outer stations with the exception of station22 (Fig. 2C). Phaeopigments were highest in the northern area ofthe mouth of Colorado River and in the zone adjacent to BahıaBlanca Estuary (Fig. 2D). POC showed the highest values at stationsoff the mouth of this estuary (Fig. 2E) while the highest meso-zooplankton abundances were found at the outer stations of thestudy area (Fig. 2F).
Five fish families were represented in the samples, namelyClupeidae, Engraulidae, Carangidae, Sciaenidae, and Atherinidae.Eggs and larvae of Brevoortia aurea (Brazilian menhaden), Engraulisanchoita, Parona signata, and Sciaenidae spp. and larvae of Odon-testhes argentinensis (marine silverside) were found in the studyarea. P. signata and Sciaenidae eggs showed the highest frequencyof occurrence (84.37 and 59.37%, respectively).
The most frequently occurring larvae were Engraulis anchoita(59.37%), Brevoortia aurea, Parona signata, and Sciaenidae (eachwith 50%) (Table 1). E. anchoita larvae were most abundant (453.59per 10 m�3) whereas Odontesthes argentinensis larvae were theleast abundant (1.88 per 10 m�3) (Table 1). B. aurea showed thehighest average abundance of eggs (432.28 per 10 m�3) whileE. anchoita showed the lowest (1.28 per 10 m�3) average abundanceof eggs.
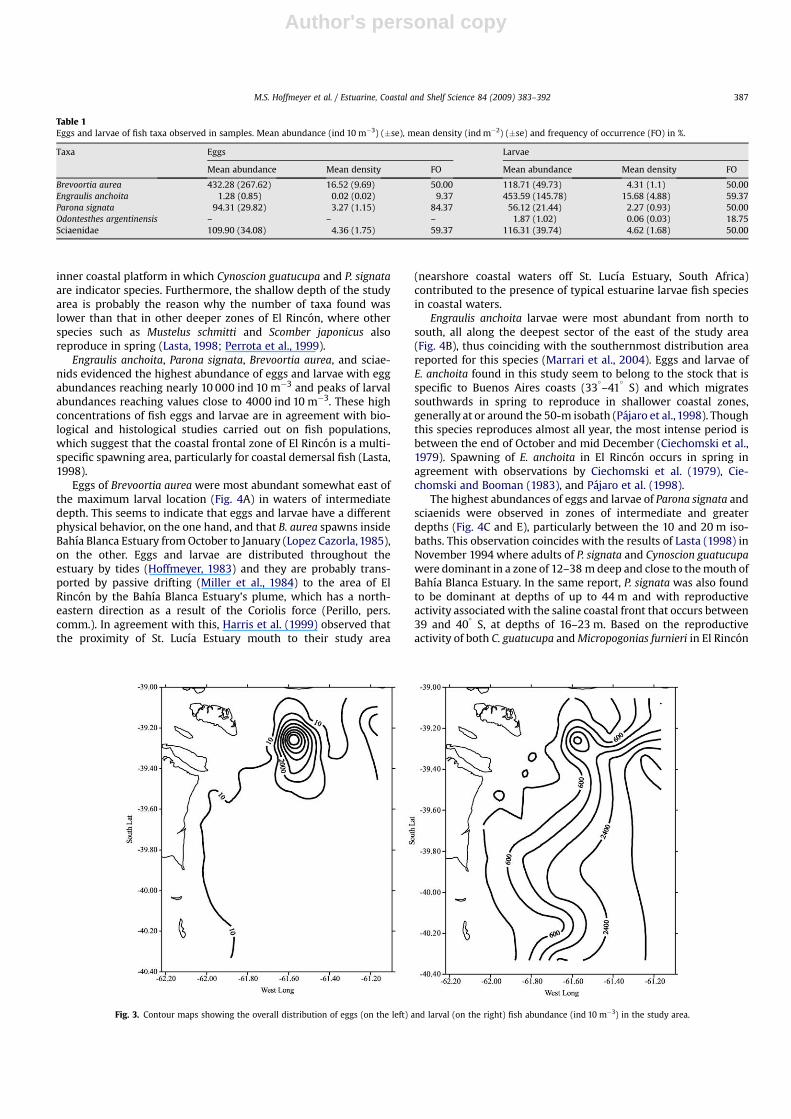
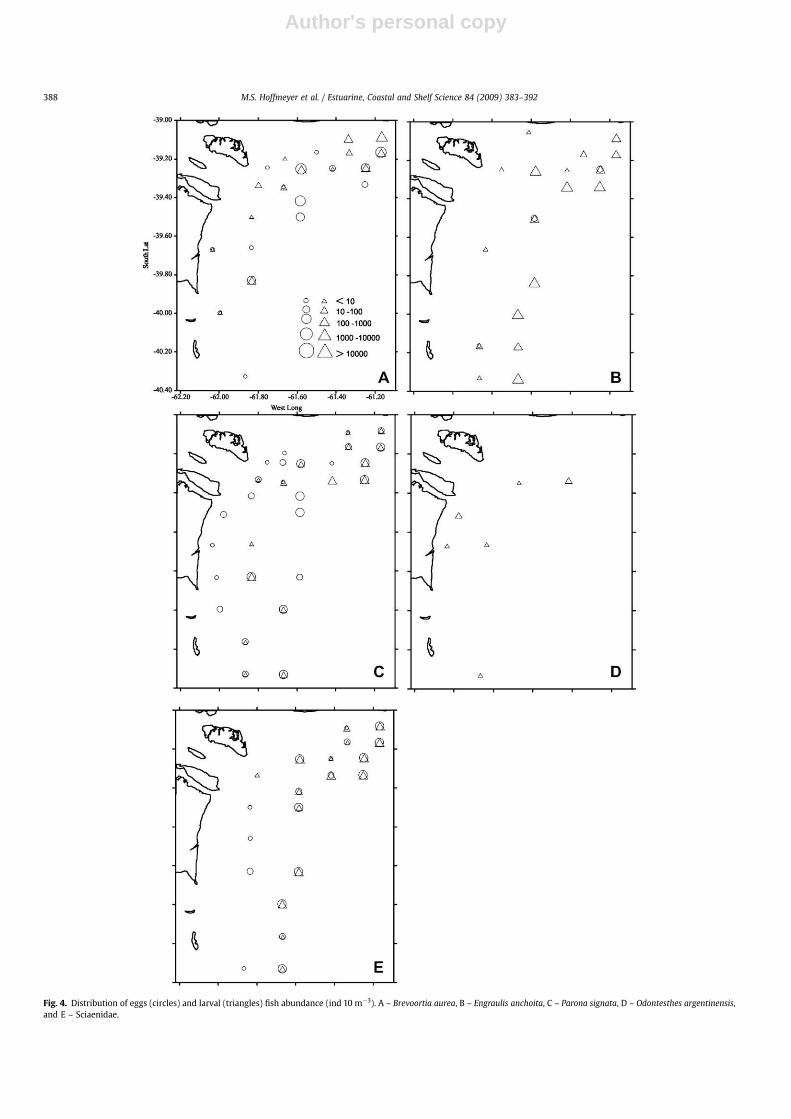
The highest egg abundance values were restricted to a smallzone close to the mouth of Bahıa Blanca Estuary while larvae weremost abundant offshore, decreasing towards the coast (Fig. 3). Eggsof Brevoortia aurea were most abundant in the outer zone towardsthe north of the study area, with maximum values at station 11(8.078 ind 10 m�3), followed by stations 19 and 5 (2718 and1944 ind 10 m�3, respectively) (Fig. 4A, see Fig. 1). The lowestabundances (up to 2 ind 10 m�3) were at the deepest coastalstations. The larval abundance pattern was similar to that of eggs
(ranging from 3 to 1.145 per 10 m�3) although the highest abun-dances were detected more to the north compared to the eggabundance pattern. Engraulis anchoita eggs were detected only atstations 13, 22, and 32 at low values ranging between 2 and 20 per10 m�3 (Fig. 4B). The highest abundance of E. anchoita larvae wasobserved in the north and south of the outer zone of the study areaat stations 14, 35, and 31, with values of 2955, 2438, and 1857 per10 m�3, respectively. In general, the lowest abundances (1–9 ind10 m�3) were in the zone close to the coast.
The distribution of Parona signata eggs was fairly homogeneousalthough abundance at deep stations was somewhat higher than atcoastal stations (Fig. 4C). The highest abundance of P. signata larvaewas observed in the outer zones (stations 13–15 and 27) withvalues ranging between 261 and 922 per 10 m�3. The lowestabundances were most frequently found in the areas of lowerdepths. Sciaenidae eggs and larvae were detected mainly in theouter zone of the study area. Egg abundances ranged between 2 and762 per 10 m�3 whereas larval abundances ranged between 2 and842 per 10 m�3 (Fig. 4D). Odontesthes argentinensis larvae evi-denced very low abundances and were found only at four coastalstations (16, 20, 23, 32) and at two outer stations (5 and 25), withvalues varying between 3 and 30 per 10 m�3 (Fig. 4E).
Cluster analysis of overall egg plus larval abundance datadetermined the formation of two sample groups at a 40% of simi-larity. The cluster plot is not shown. However, the two definedgroups of stations can be seen in Fig. 1. Group I was composed offifteen samples located in the outer zone whereas Group II wascomposed of fourteen samples located in the shallower inner zone,adjacent to Bahıa Blanca Estuary and Anegada Bay. Group I revealedthe highest average abundances for all taxa, and was dominated byEngraulis anchoita larvae followed by Brevoortia aurea eggs whereasGroup II showed very low average abundances (Table 2).
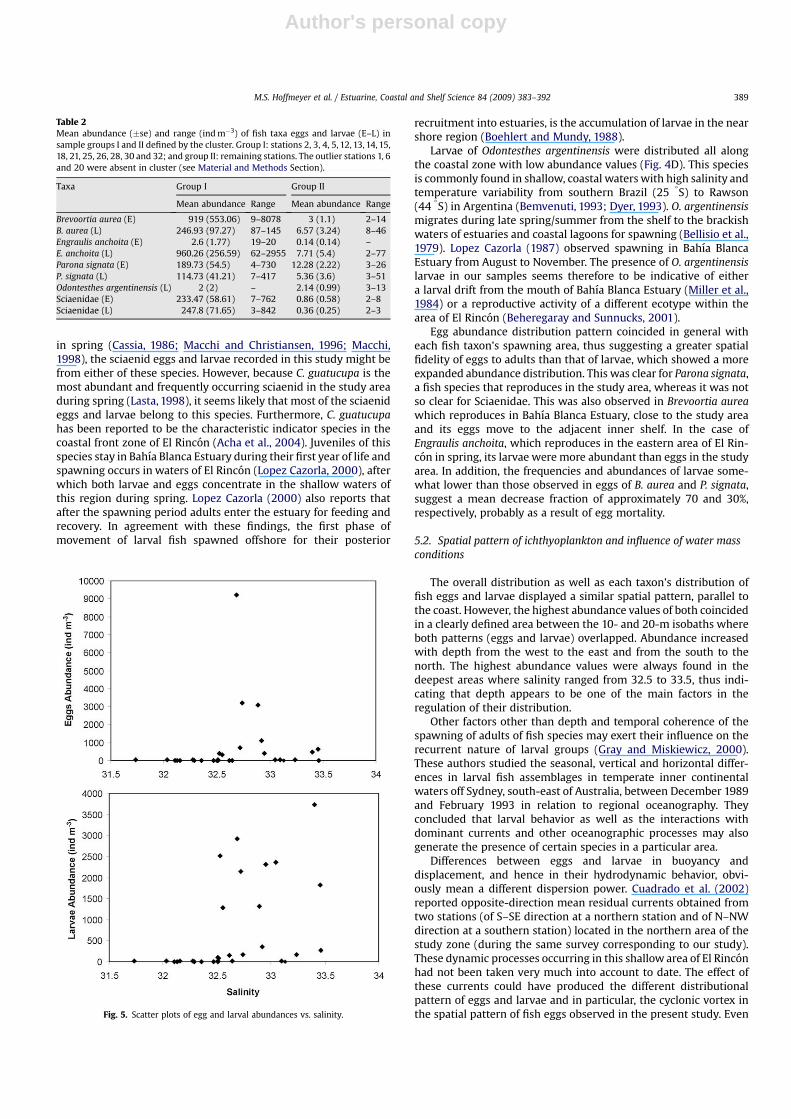
Egg and larval distribution patterns showed strong relationshipswith salinities between 32.5 and 33.5 (Fig. 5) and mesozooplanktonabundance, but not with POC concentration. The stepwise MultipleLinear Regression Analysis corroborated these trends revealingdirect relationships of eggs and larvae with these variables but aninverse relationship of larvae with particulate organic carbon(POC). The selected models for assessing the environmental inter-dependence of eggs and larvae were: Eggs (EA)¼�45.78þ 1.017MAþ 1.28 Sal, R2¼ 0.48 (F¼ 3.87, p 0.05*), and Larvae(LA)¼�44.7� 0.001 POCþ 1.35 Salþ 0.71 MA, R2¼ 0.60 (F¼ 6.28,p 0.01**). The b values of the explanatory variables in both modelsindicate the different degree of importance of each relationship.The estimated R2 values although are lower than the minimalpredictive value (0.65) suggested by Prairie (1996), are reasonableexplanatory percentages regarding the observational nature ofthese data.
5. Discussion
5.1. Ichthyoplankton composition and its abundance distribution inrelation with specific spawning areas
Results from the present study indicate the presence of eggs ofone estuarine fish (Brevoortia aurea), two demersal fish (Paronasignata and sciaenids), and only one pelagic fish (Engraulis anchoita)in the study area. They also reveal the presence of larvae of all thesefish species as well as of Odontesthes argentinensis, a coastal estu-arine species (Lopez Cazorla, 1987). The low ichthyoplanktonrichness observed could be explained as a result of the reproduc-tion of these few species during spring (Lasta et al., 1998; Jaur-eguizar et al., 2004a). In this respect, and based on their relativeabundance, Angelescu and Prenski (1987) and Jaureguizar et al.(2004b) identified for El Rincon, a fish assemblage typical of the
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392 385

Author's personal copy
Fig. 2. Contour maps. A – temperature (T�C), B – salinity, C – chlorophyll-a content (mg m�3), D – phaeopigments concentration (mg m�3), E – particulate organic carbon
concentration (mg m�3), and F – mesozooplankton abundance (ind m�3).
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392386
Author's personal copy
inner coastal platform in which Cynoscion guatucupa and P. signataare indicator species. Furthermore, the shallow depth of the studyarea is probably the reason why the number of taxa found waslower than that in other deeper zones of El Rincon, where otherspecies such as Mustelus schmitti and Scomber japonicus alsoreproduce in spring (Lasta, 1998; Perrota et al., 1999).
Engraulis anchoita, Parona signata, Brevoortia aurea, and sciae-nids evidenced the highest abundance of eggs and larvae with eggabundances reaching nearly 10 000 ind 10 m�3 and peaks of larvalabundances reaching values close to 4000 ind 10 m�3. These highconcentrations of fish eggs and larvae are in agreement with bio-logical and histological studies carried out on fish populations,which suggest that the coastal frontal zone of El Rincon is a multi-specific spawning area, particularly for coastal demersal fish (Lasta,1998).
Eggs of Brevoortia aurea were most abundant somewhat east ofthe maximum larval location (Fig. 4A) in waters of intermediatedepth. This seems to indicate that eggs and larvae have a differentphysical behavior, on the one hand, and that B. aurea spawns insideBahıa Blanca Estuary from October to January (Lopez Cazorla, 1985),on the other. Eggs and larvae are distributed throughout theestuary by tides (Hoffmeyer, 1983) and they are probably trans-ported by passive drifting (Miller et al., 1984) to the area of ElRincon by the Bahıa Blanca Estuary’s plume, which has a north-eastern direction as a result of the Coriolis force (Perillo, pers.comm.). In agreement with this, Harris et al. (1999) observed thatthe proximity of St. Lucıa Estuary mouth to their study area
(nearshore coastal waters off St. Lucıa Estuary, South Africa)contributed to the presence of typical estuarine larvae fish speciesin coastal waters.
Engraulis anchoita larvae were most abundant from north tosouth, all along the deepest sector of the east of the study area(Fig. 4B), thus coinciding with the southernmost distribution areareported for this species (Marrari et al., 2004). Eggs and larvae ofE. anchoita found in this study seem to belong to the stock that isspecific to Buenos Aires coasts (33
�–41
�S) and which migrates
southwards in spring to reproduce in shallower coastal zones,generally at or around the 50-m isobath (Pajaro et al., 1998). Thoughthis species reproduces almost all year, the most intense period isbetween the end of October and mid December (Ciechomski et al.,1979). Spawning of E. anchoita in El Rincon occurs in spring inagreement with observations by Ciechomski et al. (1979), Cie-chomski and Booman (1983), and Pajaro et al. (1998).
The highest abundances of eggs and larvae of Parona signata andsciaenids were observed in zones of intermediate and greaterdepths (Fig. 4C and E), particularly between the 10 and 20 m iso-baths. This observation coincides with the results of Lasta (1998) inNovember 1994 where adults of P. signata and Cynoscion guatucupawere dominant in a zone of 12–38 m deep and close to the mouth ofBahıa Blanca Estuary. In the same report, P. signata was also foundto be dominant at depths of up to 44 m and with reproductiveactivity associated with the saline coastal front that occurs between39 and 40
�S, at depths of 16–23 m. Based on the reproductive
activity of both C. guatucupa and Micropogonias furnieri in El Rincon
Table 1Eggs and larvae of fish taxa observed in samples. Mean abundance (ind 10 m�3) (�se), mean density (ind m�2) (�se) and frequency of occurrence (FO) in %.
Taxa Eggs Larvae
Mean abundance Mean density FO Mean abundance Mean density FO
Brevoortia aurea 432.28 (267.62) 16.52 (9.69) 50.00 118.71 (49.73) 4.31 (1.1) 50.00Engraulis anchoita 1.28 (0.85) 0.02 (0.02) 9.37 453.59 (145.78) 15.68 (4.88) 59.37Parona signata 94.31 (29.82) 3.27 (1.15) 84.37 56.12 (21.44) 2.27 (0.93) 50.00Odontesthes argentinensis – – – 1.87 (1.02) 0.06 (0.03) 18.75Sciaenidae 109.90 (34.08) 4.36 (1.75) 59.37 116.31 (39.74) 4.62 (1.68) 50.00
Fig. 3. Contour maps showing the overall distribution of eggs (on the left) and larval (on the right) fish abundance (ind 10 m�3) in the study area.
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392 387
Author's personal copy
Fig. 4. Distribution of eggs (circles) and larval (triangles) fish abundance (ind 10 m�3). A – Brevoortia aurea, B – Engraulis anchoita, C – Parona signata, D – Odontesthes argentinensis,and E – Sciaenidae.
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392388
Author's personal copy
in spring (Cassia, 1986; Macchi and Christiansen, 1996; Macchi,1998), the sciaenid eggs and larvae recorded in this study might befrom either of these species. However, because C. guatucupa is themost abundant and frequently occurring sciaenid in the study areaduring spring (Lasta, 1998), it seems likely that most of the sciaenideggs and larvae belong to this species. Furthermore, C. guatucupahas been reported to be the characteristic indicator species in thecoastal front zone of El Rincon (Acha et al., 2004). Juveniles of thisspecies stay in Bahıa Blanca Estuary during their first year of life andspawning occurs in waters of El Rincon (Lopez Cazorla, 2000), afterwhich both larvae and eggs concentrate in the shallow waters ofthis region during spring. Lopez Cazorla (2000) also reports thatafter the spawning period adults enter the estuary for feeding andrecovery. In agreement with these findings, the first phase ofmovement of larval fish spawned offshore for their posterior
recruitment into estuaries, is the accumulation of larvae in the nearshore region (Boehlert and Mundy, 1988).
Larvae of Odontesthes argentinensis were distributed all alongthe coastal zone with low abundance values (Fig. 4D). This speciesis commonly found in shallow, coastal waters with high salinity andtemperature variability from southern Brazil (25
�S) to Rawson
(44�S) in Argentina (Bemvenuti, 1993; Dyer, 1993). O. argentinensis
migrates during late spring/summer from the shelf to the brackishwaters of estuaries and coastal lagoons for spawning (Bellisio et al.,1979). Lopez Cazorla (1987) observed spawning in Bahıa BlancaEstuary from August to November. The presence of O. argentinensislarvae in our samples seems therefore to be indicative of eithera larval drift from the mouth of Bahıa Blanca Estuary (Miller et al.,1984) or a reproductive activity of a different ecotype within thearea of El Rincon (Beheregaray and Sunnucks, 2001).
Egg abundance distribution pattern coincided in general witheach fish taxon’s spawning area, thus suggesting a greater spatialfidelity of eggs to adults than that of larvae, which showed a moreexpanded abundance distribution. This was clear for Parona signata,a fish species that reproduces in the study area, whereas it was notso clear for Sciaenidae. This was also observed in Brevoortia aureawhich reproduces in Bahıa Blanca Estuary, close to the study areaand its eggs move to the adjacent inner shelf. In the case ofEngraulis anchoita, which reproduces in the eastern area of El Rin-con in spring, its larvae were more abundant than eggs in the studyarea. In addition, the frequencies and abundances of larvae some-what lower than those observed in eggs of B. aurea and P. signata,suggest a mean decrease fraction of approximately 70 and 30%,respectively, probably as a result of egg mortality.
5.2. Spatial pattern of ichthyoplankton and influence of water massconditions
The overall distribution as well as each taxon’s distribution offish eggs and larvae displayed a similar spatial pattern, parallel tothe coast. However, the highest abundance values of both coincidedin a clearly defined area between the 10- and 20-m isobaths whereboth patterns (eggs and larvae) overlapped. Abundance increasedwith depth from the west to the east and from the south to thenorth. The highest abundance values were always found in thedeepest areas where salinity ranged from 32.5 to 33.5, thus indi-cating that depth appears to be one of the main factors in theregulation of their distribution.
Other factors other than depth and temporal coherence of thespawning of adults of fish species may exert their influence on therecurrent nature of larval groups (Gray and Miskiewicz, 2000).These authors studied the seasonal, vertical and horizontal differ-ences in larval fish assemblages in temperate inner continentalwaters off Sydney, south-east of Australia, between December 1989and February 1993 in relation to regional oceanography. Theyconcluded that larval behavior as well as the interactions withdominant currents and other oceanographic processes may alsogenerate the presence of certain species in a particular area.
Differences between eggs and larvae in buoyancy anddisplacement, and hence in their hydrodynamic behavior, obvi-ously mean a different dispersion power. Cuadrado et al. (2002)reported opposite-direction mean residual currents obtained fromtwo stations (of S–SE direction at a northern station and of N–NWdirection at a southern station) located in the northern area of thestudy zone (during the same survey corresponding to our study).These dynamic processes occurring in this shallow area of El Rinconhad not been taken very much into account to date. The effect ofthese currents could have produced the different distributionalpattern of eggs and larvae and in particular, the cyclonic vortex inthe spatial pattern of fish eggs observed in the present study. Even
Table 2Mean abundance (�se) and range (ind m�3) of fish taxa eggs and larvae (E–L) insample groups I and II defined by the cluster. Group I: stations 2, 3, 4, 5, 12, 13, 14, 15,18, 21, 25, 26, 28, 30 and 32; and group II: remaining stations. The outlier stations 1, 6and 20 were absent in cluster (see Material and Methods Section).
Taxa Group I Group II
Mean abundance Range Mean abundance Range
Brevoortia aurea (E) 919 (553.06) 9–8078 3 (1.1) 2–14B. aurea (L) 246.93 (97.27) 87–145 6.57 (3.24) 8–46Engraulis anchoita (E) 2.6 (1.77) 19–20 0.14 (0.14) –E. anchoita (L) 960.26 (256.59) 62–2955 7.71 (5.4) 2–77Parona signata (E) 189.73 (54.5) 4–730 12.28 (2.22) 3–26P. signata (L) 114.73 (41.21) 7–417 5.36 (3.6) 3–51Odontesthes argentinensis (L) 2 (2) – 2.14 (0.99) 3–13Sciaenidae (E) 233.47 (58.61) 7–762 0.86 (0.58) 2–8Sciaenidae (L) 247.8 (71.65) 3–842 0.36 (0.25) 2–3
Fig. 5. Scatter plots of egg and larval abundances vs. salinity.
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392 389

Author's personal copy
though other explanations could be provided in this respect, itcould also be hypothesized that a concentrated fish spawningoccurred in the vortex zone from where eggs and larvae were laterdispersed to shallow zones by passive drift. It is known that gyresand coastal eddies with variable persistence on a seasonal scalebehave as excellent fish egg and larval retention mechanisms(Nishimoto and Washburn, 2002). A clockwise gyre-like motion inthe deepest waters adjacent to our study area in El Rincon has beensuggested (Piola and Rivas, 1997). This circulation pattern, inagreement with Marrari et al. (2004), could be another retentionmechanism for fish eggs and larvae in this coastal region.
Surface temperature and salinity showed opposite gradientsfrom the mouth of Bahıa Blanca Estuary and Anegada Bay towardsthe deepest zones of the inner shelf. This demonstrates the influ-ence of a cross-shelf effect from the mouth and plumes of theseenvironments and the inner shelf. The surface gradients of thesevariables agree with those reported for the entire water columndata obtained through profiles in the same survey indicating somevertical mixing (Cuadrado et al., 2002). However, a slight stratifi-cation of both salinity and temperature in El Rincon, in front ofAnegada Bay and Bahıa Blanca Estuary was observed in November1994 (Lasta, 1998). A homogeneous plume of Colorado River char-acterized by high concentrations of suspended matter, low salinity,and re-suspended matter, running in a NE direction towards the ElRincon coastal region was also reported for the same period of timeof our survey: November 1993 (Cuadrado et al., 2002). The influ-ence of this plume was also observed in the surface data of salinityand inorganic nutrient-POC concentration analyzed in this study.Also, the highest surface POC values coincided with the highestconcentration values of suspended matter in the water columnreported by Cuadrado et al. (2002). The gradients of these variablesfrom Bahıa Blanca Estuary, Colorado River, and Anegada Bay to theEl Rincon seem to have contributed to the formation of a turbidityfront. However, maximal egg and larval abundances were detectedin the eastern sector of the study area in agreement with themaximal values of mesozooplankton abundance and salinity andthe minimal POC concentrations. Accordingly, results derivedmainly from the b values of explanatory variables in models fromMultiple Linear Regression Analyses, corroborate the above-mentioned relationships.
Interestingly, part of the saline inner shelf front described byGuerrero and Piola (1997), Lasta (1998) and Acha et al. (2004), islocated in the area where maximal egg and larval abundance valueswere found. This front divides the inner shelf in El Rincon regioninto two sub-areas with different salinities (i.e. the inner area haslower salinity than the outer area) and constitutes the site of themost important demersal fish spawning reported to date (Lasta,1998). The presence of ichthyoplankton in this area could be due tothe influence that this front exerts concentrating plankton andseston, both of which are trophic resources considerably importantfor fish larvae (Mann and Lazier, 1996; Grioche and Koubbi, 1997;Guerrero and Piola, 1997; Piola and Rivas, 1997; Lee et al., 2005).Thehigh densities of eggs and larvae found in the present study agreewell with observations on the concentration of larvae and repro-ductive activity of Micropogonias furnieri, Cynoscion guatucupa andParona signata along the bottom saline front of the Rıo de la PlataEstuary, a northern area on Argentina’s coasts (Berasategui et al.,2004; Simionato et al., 2008). However, in the absence of additionalstations seaward the front, we cannot demonstrate the presence ofa complete relationship between this frontal system and theobserved ichthyoplankton spatial pattern. Unlike observationsfrom the present research, Grimes and Finucane (1991) describedthe spatial distribution and abundance of ichthyoplankton insurface waters around the Mississippi River discharge plume. Theseauthors sampled the three water masses in the plume area – plume
water, Gulf of Mexico shelf water and frontal water – and theyobserved that the hydrodynamic convergence at the zone of thefront most likely accounts for the highest ichthyoplankton abun-dance found. High microzooplankton concentrations (mostlycopepod nauplii) have also been reported from this frontal region ofthe Mississippi plume (Dagg and Whitledge, 1991). Thus, highzooplankton concentrations in frontal waters produce the co-occurrence of high ichthyoplankton concentrations (Grimes andFinucane, 1991). Copepod nauplii, copepodites and other micro-zooplankton are known rich food resources that can sustainaggregations of larval and small juvenile fish (see Hunter andKimbrell, 1980; Peterson and Ausubel, 1984; Govoni and Chester,1990). For instance, nauplii and copepodites of the copepod Para-calanus parvus are suitable food for anchovy (Engraulis anchoita)larvae in the Argentinean shelf (Vinas and Ramırez, 1996).
Similarly to what occurs with ichthyoplankton spatial distribu-tion in the majority of coasts all over the world, the spatial patternobserved in the study area during late spring 1993, was primarilyassociated with the reported specific fish taxa spawning areas(Bellisio et al., 1979; Ciechomski et al., 1979; Ciechomski and Boo-man, 1983; Lopez Cazorla, 1985, 1987; Cassia, 1986; Macchi andChristiansen, 1996; Lasta, 1998; Lasta et al., 1998; Macchi, 1998;Pajaro et al., 1998; Jaureguizar et al., 2004a; Marrari et al., 2004).
As a common phenomenon in several coastal areas (Dagg andWhitledge, 1991; Grimes and Finucane, 1991; Grioche and Koubbi,1997; Gray and Miskiewicz, 2000; Berasategui et al., 2004; Leeet al., 2005; Simionato et al., 2008), the geographic location ofmaximal egg–larval densities were coincident with the position ofa coastal estuarine saline front described for this region in spring-time, which evidences this bio-physical and trophic linkage.
Finally, the direct relationships detected between egg–larvaldistribution, and salinity–mesozooplankton gradients and thesmall inverse relationship only between larvae and POC as well asthe influence of depth, currents and plumes of Bahıa Blanca Estuary,Colorado River and Anegada Bay on egg and larval distribution, areother factors which also could induce the observed ichthyo-plankton spatial pattern.
6. Conclusions
Based on our results, it can be concluded that the study area in ElRincon region off Bahıa Blanca Estuary is an important nurseryground for Brevoortia aurea, Engraulis anchoita, Parona signata,Odontesthes argentinensis and Sciaenidae spp. during spring. Takentogether, our findings demonstrate the occurrence of high densitiesof zooplankton and ichthyoplankton in synchronism with spawn-ing of various fish populations such as Cynoscion guatucupa andMicropogonias furnieri, and P. signata, which are commerciallyimportant fishery species (Lasta, 1998). They therefore confirmobservations on other coastal areas in which a coherence patternbetween eggs–larvae and the spawning area of adults has also beenrecorded. It was also possible to observe the influence of oceano-graphic physical factors, particularly of a frontal zone and currents,on the ichthyoplankton spatial pattern. Still, further detailedstudies are necessary to demonstrate which of the environmentalfactors detected in the present study, are crucial to control theparticular spatial pattern of ichthyoplankton observed in thestudied area during spring.
Acknowledgements
This study was supported by grant PIP N�
3003/00 to M.S.H.from the National Research Council (CONICET). Authors areindebted to the onboard personnel of the oceanographic ship ‘‘ElAustral’’ during the November 1993 cruise (AU-02/93). Thanks are
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392390

Author's personal copy
due to W.D. Melo for designing the maps and to the staff working atthe Laboratorio de Quımica Marina, Instituto Argentino de Ocean-ografıa (IADO), for providing the physical and chemical variabledata. Thanks are also due to some anonymous reviewers that hel-ped to improve the first versions of this Ms.
References
Acha, E.M., Mianzan, H.W., Guerrero, R.A., Favero, M., Bava, J., 2004. Marine fronts atthe continental shelves of Austral South America. Physical and ecologicalprocesses. Journal of Marine Systems 44, 83–105.
Angelescu, V., Prenski, L.B., 1987. Ecologıa trofica de la merluza comun del MarArgentino (Merlucciidae) Merluccius hubbsi. Parte 2 Dinamica de la alimenta-cion analizada sobre la base de las condiciones ambientales, la estructura y lasevaluaciones de los efectivos en su area de distribucion. Contribucion Cientıfica561. Instituto Nacional de Investigacion y Desarrollo Pesquero (INIDEP), Mar delPlata, Argentina, p. 205.
Beheregaray, L.B., Sunnucks, P., 2001. Fine-scale genetic structure, estuarine colo-nization and incipient speciation in the marine silverside fish Odontesthesargentinensis. Molecular Ecology 10, 2849–2866.
Bellisio, N., Lopez, R., Torno, A., 1979. Peces marinos patagonicos. Subsecretarıa dePesca, Secretarıa de Estado de Intereses Marıtimos. Ministerio de Economıa dela Republica Argentina, Buenos Aires, p. 279.
Bemvenuti, M.A., 1993. Redescriçao do peixe-rei Odontesthes argentinensis (Valen-ciennes) Pisces: Atherinidae, na costa do Rio Grande do Sul. Atlantica 15, 17–35.
Berasategui, A.D., Acha, E.M., Fernandez Araoz, N.C., 2004. Spatial patterns of ich-thyoplankton assemblages in the Rıo de la Plata Estuary (Argentina–Uruguay).Estuarine, Coastal and Shelf Science 60, 599–610.
Boehlert, G.W., Mundy, B.C., 1988. Roles of behavioral and physical factors in larvaland juvenile recruitment to estuarine nursery areas. American Fisheries SocietySymposium 3, 51–67.
Cassia, M.C., 1986. Reproduccion y fecundidad de la pescadilla de red (Cynoscionstriatus. Publicacion de la Comision Tecnica Mixta del Frente Marıtimo 1, 191–203.
Cassia, M.C., Booman, C.I., 1985. Distribucion del ictioplancton en el Mar Argentinoen los anos 1981–1982. Physis, A 43, 91–111.
Ciechomski, J.D., Booman, C.I., 1983. Distribucion cuantitativa de huevos y larvas deanchoıta (Engraulis anchoita) en la plataforma continental, frente a las costas dela Argentina y Uruguay en el ciclo anual 1981/1982. Contribucion Cientıfica 431.Instituto Nacional de Investigacion y Desarrollo Pesquero (INIDEP), Mar delPlata, Argentina, p. 14.
Ciechomski, J.D., Sanchez, R.P., Ehrlich, M.D., Lasta, C.A., 1979. Distribucion dehuevos y larvas de anchoıta (Engraulis anchoita) en el Mar Argentino en difer-entes epocas del ano y evaluacion de sus efectivos de adultos desovantes.Contribucion Cientıfica 379. Instituto Nacional de Investigacion y DesarrolloPesquero (INIDEP), Mar del Plata, Argentina, p. 14.
Ciechomski, J.D., Weiss, G., 1974. Distribucion de huevos y larvas de merluza,Merluccius merluccius hubbsi, en las aguas de la plataforma de la Argentina yUruguay en relacion con la anchoıta, Engraulis anchoita, y las condicionesambientales. Physis, A 33, 185–198.
Clarke, K.R., Warwick, R.M., 1994. Change in Marine Communities: an Approach toStatistical Analysis and Interpretation. Plymouth Marine Laboratory, UnitedKingdom, p. 144.
Cordo, H.D., 1986. Estudios biologicos sobre peces costeros con datos de dos cam-panas de investigacion realizadas en 1981. III La pescadilla de red (Cynoscionstriatus). Publicacion de la Comision Tecnica Mixta del Frente Marıtimo, Com.Tec. Mix. Fr. Mar., Montevideo, Uruguay 1, 15–27.
Cuadrado, D.G., Piccolo, M.C., Perillo, G., 2002. Hydrography of the inner shelfoffshore Bahıa Blanca Estuary, Argentina. Thalassas 18, 45–56.
Dagg, M.J., Whitledge, T.E., 1991. Concentrations of copepod nauplii associated withthe nutrient-rich plume of the Mississippi River. Continental Shelf Research 11,409–423.
Dyer, B.S., 1993. A Phylogenetic Study of Atheriniform Fishes with a SystematicRevision of the South American Silversides (Atherinomorpha, Atherinopsinae,Sorgentinini). Ph.D. thesis, University of Michigan, Ann Arbor, Michigan, USA,p. 596.
Espinosa-Fuentes, L., Flores-Coto, C., 2004. Cross-shelf and vertical structure ofichthyoplankton assemblages in continental shelf waters of the Southern Gulfof Mexico. Estuarine, Coastal and Shelf Science 59, 333–352.
Govoni, J.J., Chester, A.J., 1990. Diet composition of larval Leiostomus xanthurus in theMississippi river plume. Journal of Plankton Research 12, 819–830.
Govoni, J.J., Grimes, C.B., 1992. The surface accumulation of larval fishes by hydro-dynamic convergence within the Mississippi River plume front. ContinentalShelf Research 12, 1265–1276.
Gray, C.A., 1996. Do thermoclines explain the vertical distributions of larval fishes inthe dynamic coastal waters of south-eastern Australia? Marine FreshwaterResearch 47, 183–190.
Gray, C.A., Miskiewicz, A.G., 2000. Larval fish assemblages in south-east Australiancoastal waters: seasonal; and spatial structure. Estuarine, Coastal and ShelfScience 50, 549–570.
Grimes, C.B., Finucane, J.H., 1991. Spatial distribution and abundance of larval andjuvenile fish, chlorophyll and macrozooplankton around the Mississippi River
discharge plume, and the role of the plume in fish recruitment. Marine EcologyProgress Series 75, 109–119.
Grioche, A., Koubbi, P., 1997. A preliminary study of the influence of a coastal frontalstructure on ichthyoplankton assemblages in the English Channel. ICES Journalof Marine Science 54, 93–104.
Guerrero, R.A., Piola, A.R., 1997. Masas de agua de la Plataforma Continental. In:Boschi, E.E. (Ed.), El Mar Argentino y sus recursos pesqueros. Tomo 1- Ante-cedentes historicos de las exploraciones en el mar y las caracterısticas ambi-entales. Instituto Nacional de Investigacion y Desarrollo Pesquero (INIDEP), Mardel Plata, Argentina, pp. 107–118.
Harris, S.A., Cyrus, D.P., Beckley, L.E., 1999. The Larval fish assemblage in nearshorecoastal waters off the St Lucia Estuary, South Africa. Estuarine, Coastal and ShelfScience 49, 789–811.
Hoffmeyer, M.S., 1983. Zooplancton del area interna de la Bahıa Blanca (BuenosAires, Argentina) I. Composicion faunıstica. Historia Natural 3, 73–94.
Hunter, J.F., Kimbrell, C.A., 1980. Early life history of Pacific mackerel, Scomberjaponicus. Fishery Bulletin 78, 89–101.
Jaureguizar, A., Guerrero, R., Lasta, C., 2004a. Estructura y dinamica de las asocia-ciones ıcticas demersales costeras (34
�–41
�S), e influencia ambiental sobre su
estructuracion espacial. Informe Tecnico Interno 64. Instituto Nacional deInvestigacion y Desarrollo Pesquero (INIDEP), Mar del Plata, Argentina, p. 32.
Jaureguizar, A.J., Menni, R., Guerrero, R., Lasta, C., 2004b. Environmental factorsstructuring fish communities of the Rio de la Plata estuary. Fisheries Research66, 195–211.
Kingsford, M.J., Suthers, I.M., 1994. Dynamic estuarine plumes and fronts: impor-tance to small fish and plankton in coastal waters of NSW, Australia. Conti-nental Shelf Research 14, 665–672.
Lasta, C.A. (Ed.), 1998. Resultados de una campana de evaluacion de recursosdemersales costeros de la provincia de Buenos Aires y del litoral uruguayo.Noviembre, 1994. Instituto Nacional de Investigacion y Desarrollo Pesquero(INIDEP), Mar del Plata, Argentina, p. 141. Informe Tecnico 21.
Lee, O., Nash, R.D.M., Danilowicz, B.S., 2005. Small-scale spatio-temporal variabilityin ichthyoplankton and zooplankton distribution in relation to a tidal-mixingfront in the Irish Sea. ICES Journal of Marine Science 62, 1021–1036.
Legendre, L., Demers, S., 1984. Towards dynamic biological oceanography andlimnology. Canadian Journal of Fisheries and Aquatic Sciences 41, 2–19.
Legendre, P.S., Dallot, S., Legendre, L., 1985. Succession of species within a commu-nity: chronological clustering, with applications to marine and freshwaterzooplankton. American Naturalist 125, 257–288.
Lopez Cazorla, A., 1985. Edad, crecimiento y comportamiento migratorio de Bre-voortia aurea (Agassiz, 1829) (Osteichthyes, Clupeidae) de Bahıa Blanca(Argentina). Investigaciones Pesqueras 49, 297–313.
Lopez Cazorla, A., 1987. Contribucion al conocimiento de la ictiofauna marina delarea de Bahıa Blanca. Thesis doctoral, Universidad Nacional de La Plata, La Plata,Argentina, p. 247.
Lopez Cazorla, A., 2000. Age structure of the population of weakfish Cynoscionguatucupa (Cuvier) in the Bahıa Blanca waters, Argentina. Fisheries Research 46,279–286.
Macchi, G.J., 1998. Preliminary estimate of spawning frequency and batch fecundityof stripped weakfish, Cynoscion striatus, in coastal waters off Buenos Airesprovince. Fishery Bulletin 96, 375–381.
Macchi, G.J., Christiansen, H.E., 1996. Analisis temporal del proceso de maduracion ydeterminacion de la incidencia de atresias en la corvina rubia (Micropogoniasfurnieri). Frente Marıtimo 16 (A), 93–101.
Mann, K.H., Lazier, J.R.N., 1996. Dynamics of Marine Ecosystems. Biological–PhysicalInteractions in the Oceans. Blackwell Scientific Publications, Boston, p. 466.
Marrari, M., Vinas, M.D., Martos, P., Hernandez, D., 2004. Spatial patterns of mes-ozooplankton distribution in the Southwestern Atlantic Ocean (34
�–41
�S)
during austral spring: relationship with the hydrographic conditions. ICESJournal of Marine Science 61, 667–679.
Martos, P., Piccolo, M.C., 1988. Hydrography of the Argentine continental shelfbetween 38
�and 42
�S. Continental Shelf Research 8, 1043–1056.
Matsuura, Y., Olivar, M.P., 1999. Fish larvae. In: Boltovskoy, D. (Ed.), South AtlanticZooplankton. Backhuys Publishers, Leiden, pp. 1445–1496.
Miller, J.M., Reed, J.P., Pietrafesa, L.J., 1984. Patterns, mechanisms and approaches tothe study of migrations of estuarine-dependent fish larvae and juveniles. In:McCleave, J.D., Arnold, G.P., Dodson, J.J., Neill, W.H. (Eds.), Mechanisms ofMigration in Fishes. Plenum Press, New York, pp. 209–225.
Nishimoto, M.M., Washburn, L., 2002. Patterns of coastal eddy circulation andabundance of pelagic juvenile fish in the Santa Barbara Channel, California, USA.Marine Ecology Progress Series 241, 183–199.
Pajaro, M., Sanchez, R.P., Aubone, A., 1998. Mortalidad embrionaria por canibalismoen la anchoıta argentina Engraulis anchoita Hubbs y Marini, 1935. Boletın delInstituto Espanol de Oceanografıa 14, 81–97.
Perillo, G., 1994. Estudio oceanografico de la plataforma interior frente al estuariode Bahıa Blanca. Contribucion Cientıfica 311. Instituto Argentino de Ocean-ografıa (IADO), Bahıa Blanca, Argentina, p. 54.
Perrota, R.G., Madirolas, A., Vinas, M.D., Akselman, R., Guerrero, R., Sanchez, F.,Lopez, F., Castro Machado, F., Machi, G., 1999. La caballa (Scomber japonicus) y lascondiciones ambientales en el area bonaerense de ‘‘El Rincon’’ (39
�–40
�300 S).
Agosto, 1996. Informe Tecnico 26. Instituto Nacional de Investigacion y DesarrolloPesquero (INIDEP), Mar del Plata, Argentina, p. 29.
Perrota, R.G., Vinas, M.D., Madirolas, A., Reta, R., Akselman, R., Castro Machado, F.J.,David Garciarena, A., Macchi, G.J., Moriondo Danovaro, P., Llanos, V.,Urteaga, J.R., 2003. La caballa (Scomber japonicus) y las condiciones
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392 391

Author's personal copy
ambientales en el area ‘‘El Rincon’’ (39�–40
�300 S) del Mar Argentino. Sep-
tiembre, 2000. Informe Tecnico 54. Instituto Nacional de Investigacion yDesarrollo Pesquero (INIDEP), Mar del Plata, Argentina, p. 25.
Peterson, W.T., Ausubel, S.J., 1984. Diets and selective feeding by larvae of Atlanticmackerel S comber scombrus on zooplankton. Marine Ecology Progress Series 17,65–75.
Piola, A.R., Rivas, A.L., 1997. Corrientes en la plataforma continental. In: Boschi, E.E.(Ed.), El Mar Argentino y sus recursos pesqueros. Tomo 1. Antecedentes his-toricos de las exploraciones en el mar y las caracterısticas ambientales. InstitutoNacional de Investigacion y Desarrollo Pesquero (INIDEP), Mar del Plata,Argentina, p. 132.
Prairie, Y.T., 1996. Evaluating the predictive power of regression models. CanadianJournal of Fisheries and Aquatic Science 53, 490–492.
Reiss, C.S., McConaugha, J.R., 1999. Cross-frontal transport and distribution of ich-thyoplankton associated with Chesapeake Bay plume dynamics. ContinentalShelf Research 19, 151–170.
Sabates, A., Olivar, M.P., 1996. Variation of larval fish distributions associated withvariability in the location of a shelf slope front. Marine Ecology Progress Series135, 11–20.
Sabates, A., Zabala, M., Garcıa-Rubıes, A., 1989. Larval fish communities in theMedes Islands Marine Reserve (north-west Mediterranean). Journal of PlanktonResearch 25, 1034–1046.
Sanchez, R.P., Ciechomski, J.,1995. Spawning and nurserygrounds of pelagic fish speciesin the sea-shelf off Argentina and adjacent areas. Scientia Marina 59, 455–478.
Simionato, C.G., Berasategui, A., Meccia, V.L., Acha, M., Mianzan, H., 2008. Shorttime-scale wind forced variability in the Rıo de la Plata Estuary and its role onichthyoplankton retention. Estuarine, Coastal and Shelf Science 76, 211–226.
Somarakis, S., Drakopolous, P., Filippou, V., 2002. Distribution and abundance of larvalfish in the northern Aegean Sea – eastern Mediterranean – in relation to earlysummer oceanographic conditions. Journal of Plankton Research 24, 339–358.
Strickland, J.D.H., Parsons, T.R., 1968. A Practical Handbook of Sea-water Analysis.Journal Bulletin 167. Fisheries Research Board of Canada, p. 311.
Vinas, M.D., Ramırez, F.C., 1996. Gut analysis of first-feeding anchovy larvae fromPatagonian spawning area in relation to food availability. Archives of FisheryMarine Research 43, 231–256.
Weiss, G., 1972. Consideraciones sobre la distribucion de huevos de peces en elplancton de las aguas de la plataforma del sector bonaerense y de Uruguay.Physis 83, 613–630.
M.S. Hoffmeyer et al. / Estuarine, Coastal and Shelf Science 84 (2009) 383–392392
Copyright © 2022 FDOKUMEN




















