

การศึกษาปริมาณสารต้านอนุมูลอิสระและฤทธิÎกา...
128
RMUTSB Acad. J. 8(2) : 187-198 (2020) 187 การศึกษาปริมาณสารต้านอนุมูลอิสระและฤทธิ การยับยังอนุมูลอิสระของนํามะหวด Investigation of antioxidant capacity and antioxidant activity of Ma-huad (Lepisanthes rubiginosa (Roxb.) Leenh.) juice วิชนี มัธยม 1* กานดาวดี โนชัย 1 กัญญา กอแก้ว 1 กาญจนา พิศาภาค 1 และ ดารานัย รบเมือง 1 Wishanee Matthayom 1* , Kandawadee Nochai 1 , Kanya KoKaew 1 , Kanjana Pisapak 1 and Daranai Robmuang 1 บทคัดย่อ มะหวด เป็นไม้ยืนต้นขนาดเล็กอยู ่ในวงศ์เงาะ (Sapindaceae) สามารถพบได้ตามป่าโปร่งของประเทศไทยและประเทศ เพื0อนบ้าน ผลสุกของมะหวดที0มีสีม่วงแดงมีสารสําคัญกลุ ่มแอนโทไซยานิน (anthocyanin) และนอกจากนี?ยังอาจพบสารกลุ ่ม โพลีฟีนอลอื0นๆ ร่วมด้วย งานวิจัยนี?เป็นการศึกษาปริมาณสารต้านอนุมูลอิสระและฤทธิ G ต้านอนุมูลอิสระของนํ?ามะหวดที0เตรียม จากผลมะหวดสุกในอัตราส่วนนํ?าหนักผลมะหวดต่อนํ?ากลั0นเท่ากับ 1:1, 1:2 และ 1:3 ตามลําดับ ใช้เวลาในการต้ม 5-15 นาที (เปลี0ยนช่วงเวลาทุก 5 นาที) โดยควบคุมอุณหภูมิที0 95 องศาเซลเซียส จากนั?นนําไปวิเคราะห์คุณสมบัติทางกายภาพของ นํ?ามะหวดและวิเคราะห์ปริมาณแอนโทไซยานินรวมด้วยวิธี pH-differential ปริมาณฟีนอลิกรวมด้วยวิธี Folin-Ciocalteu และ ฤทธิ G ต้านอนุมูลอิสระด้วยวิธี DPPH และ ABTS ผลการศึกษาพบว่า ตัวอย่าง T3 (อัตราส่วนนํ?าหนักผลมะหวดต่อนํ?าเท่ากับ 1:1 ที0 เวลา 15 นาที) แสดงปริมาณแอนโทไซยานิน ปริมาณฟี นอลิกรวม และฤทธิ G ต้านอนุมูลอิสระด้วยวิธี DPPH และ ABTS สูงกว่าตัวอย่าง อื0นๆ คือ 86.00±1.77 mg CGE/100g of FW, 118.64±1.40 mg GAE/100 g crude extract, 57.78 เปอร์เซ็นต์ และ 96.62 เปอร์เซ็นต์ ตามลําดับ นอกจากนี? ยังพบว่าความสัมพันธ์ระหว่างปริมาณแอนโทไซยานินรวมและปริมาณฟีนอลิกรวมต่อฤทธิ G การ ต้านอนุมูลอิสระด้วยวิธี DPPH (r=0.82 และ 0.88) และ ABTS (r=0.64 และ 0.72) มีความสัมพันธ์กันในระดับสูง ที0ระดับ นัยสําคัญ 0.05 (p>0.05) คําสําคัญ: มะหวด แอนโทไซยานิน ฤทธิ G การยับยั?งอนุมูลอิสระ Abstract Ma-huad (Lepisanthes rubiginosa (Roxb.) Leenh.) is a perennial plant of the Sapindaceae family. This plant has been found in the sparse forest of Thailand and neighboring countries. The color of ripe Ma-huad fruits is red - purple and the active substances found in this fruit are anthocyanins. Furthermore, others polyphenols, may be found as constituents in this plant. In the present study, antioxidant contents and antioxidant activity of Ma-huad 1 คณะวิทยาศาสตร์และเทคโนโลยี มหาวิทยาลัยเทคโนโลยีราชมงคลสุวรรณภูมิ 1 Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi * Corresponding author. E-mail: [email protected] Received: April 27, 2020; Revised: June 25, 2020; Accepted: September 17, 2020
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of การศึกษาปริมาณสารต้านอนุมูลอิสระและฤทธิÎกา...

RMUTSB Acad. J. 8(2) : 187-198 (2020) 187
การศกษาปรมาณสารตานอนมลอสระและฤทธ�การยบย �งอนมลอสระของน �ามะหวด Investigation of antioxidant capacity and antioxidant activity of Ma-huad
(Lepisanthes rubiginosa (Roxb.) Leenh.) juice
วชน มธยม1* กานดาวด โนชย1 กญญา กอแกว1 กาญจนา พศาภาค1 และ ดารานย รบเมอง1
Wishanee Matthayom1*, Kandawadee Nochai1, Kanya KoKaew1, Kanjana Pisapak1 and Daranai Robmuang1
บทคดยอ มะหวด เปนไมยนตนขนาดเลกอยในวงศเงาะ (Sapindaceae) สามารถพบไดตามปาโปรงของประเทศไทยและประเทศ
เพ0อนบาน ผลสกของมะหวดท0มสมวงแดงมสารสาคญกลมแอนโทไซยานน (anthocyanin) และนอกจากน ?ยงอาจพบสารกลม โพลฟนอลอ0นๆ รวมดวย งานวจยน ?เปนการศกษาปรมาณสารตานอนมลอสระและฤทธGตานอนมลอสระของน ?ามะหวดท0เตรยมจากผลมะหวดสกในอตราสวนน ?าหนกผลมะหวดตอน ?ากล0นเทากบ 1:1, 1:2 และ 1:3 ตามลาดบ ใชเวลาในการตม 5-15 นาท (เปล0ยนชวงเวลาทก 5 นาท) โดยควบคมอณหภมท0 95 องศาเซลเซยส จากน ?นนาไปวเคราะหคณสมบตทางกายภาพของ น ?ามะหวดและวเคราะหปรมาณแอนโทไซยานนรวมดวยวธ pH-differential ปรมาณฟนอลกรวมดวยวธ Folin-Ciocalteu และฤทธGตานอนมลอสระดวยวธ DPPH และ ABTS ผลการศกษาพบวา ตวอยาง T3 (อตราสวนน ?าหนกผลมะหวดตอน ?าเทากบ 1:1 ท0เวลา 15 นาท) แสดงปรมาณแอนโทไซยานน ปรมาณฟนอลกรวม และฤทธGตานอนมลอสระดวยวธ DPPH และ ABTS สงกวาตวอยางอ0นๆ คอ 86.00±1.77 mg CGE/100g of FW, 118.64±1.40 mg GAE/100 g crude extract, 57.78 เปอรเซนต และ 96.62 เปอรเซนต ตามลาดบ นอกจากน ? ยงพบวาความสมพนธระหวางปรมาณแอนโทไซยานนรวมและปรมาณฟนอลกรวมตอฤทธGการตานอนมลอสระดวยวธ DPPH (r=0.82 และ 0.88) และ ABTS (r=0.64 และ 0.72) มความสมพนธกนในระดบสง ท0ระดบนยสาคญ 0.05 (p>0.05) คาสาคญ: มะหวด แอนโทไซยานน ฤทธGการยบย ?งอนมลอสระ
Abstract Ma-huad (Lepisanthes rubiginosa (Roxb.) Leenh.) is a perennial plant of the Sapindaceae family. This plant
has been found in the sparse forest of Thailand and neighboring countries. The color of ripe Ma-huad fruits is red - purple and the active substances found in this fruit are anthocyanins. Furthermore, others polyphenols, may be found as constituents in this plant. In the present study, antioxidant contents and antioxidant activity of Ma-huad 1 คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม 1 Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi * Corresponding author. E-mail: [email protected] Received: April 27, 2020; Revised: June 25, 2020; Accepted: September 17, 2020

188 ว.มทรส. 8(2) : 187-198 (2563)
juices were determined. Preparation of Ma-huad juices was done by using the ratio of ripe Ma-huad fruit weight and distilled water into 1:1, 1:2 and 1:3, respectively. The boiling temperature was controlled at 95 oC while the boiling times were changed from 5 to 15 minutes (5-minute intervals). Some physical properties of Ma-huad juices were analyzed. These samples were determined for total anthocyanin content by using pH-differential method, total phenolic content by using Folin-Ciocalteu reagent, antioxidant activity by DPPH and ABTS assays. The results showed that the maximum total anthocyanin contents, total phenolic contents and antioxidant activities from DPPH and ABTS methods were 86.00±1.77 mg CGE/100g of FW, 118.64±1.40 mg GAE/100 g crude extract, 57.78%, and 96.62%, respectively which was obtained from T3 sample (ratio of ripe Ma-huad fruit weight and distilled water was 1:1 at 15 minutes). Total anthocyanin contents and total phenolic contents were highly correlated to the percentage of the inhibition of antioxidant activity both DPPH assay (r=0.82 and 0.88) and also ABTS assay (r=0.64 and 0.72) at 0.05 significance (p>0.05). Keywords: Ma-huad (Lepisanthes rubiginosa (Roxb.) Leenh.), anthocyanin, antioxidant activity
บทนา
ในปจจบนยงคงมการศกษาวจยคนหาพชสมนไพรชนดใหมๆ เพ0อนามาใชประโยชนอยางตอเน0อง ไมวาจะเปนการใชเปนยารกษาโรค การใชเปนสวนประกอบในเคร0องสาอาง หรออาหารเสรม เปนตน พชสมนไพรท0พบตามทองถ0นของไทยหลายชนดนอกจากจะสามารถรบประทานเปนผกหรอผลไมแลวยงมฤทธGท0สาคญทางยามากมาย
มะหวด (Lepisanthes rubiginosa (Roxb.) Leenh) เปนไมยนตนขนาดเลกอยในวงศเงาะ (Sapindaceae) พบไดในปาโปรง สวนหรอบรเวณทงนา สามารถพบไดท0วทกภมภาคของประเทศไทยและประเทศเพ0อนบาน โดยท0วไปชาวบานจะนาใบออนและผลมาบรโภค ตารายาโบราณมการกลาวถงสรรพคณทางยาของพชชนดน ?เชน รากสามารถแกไขพษฝภายใน แกพษรอน ขบพยาธ แกไขท0มพษรอน แกงสวด ปวดศรษะ รกษาโรคผวหนงผ0นคน ผลชวยสมานแผล แกบดมกเลอด แกไขรากสาด แกทองรวง แกปวดมวนในทอง เปนตน
(BGO Plant Databases, The Botanical Garden Organization, n.d.) แมจะมการนาสวนตางๆ ของพช ชนดน ?มาใชประโยชนอยบางแตกยงนบวายงมผศ ก ษ า ค น ค ว า ห า ค ว า ม ร ใ น ร า ย ละ เ อ ย ด ด า นอง คปร ะกอบท าง เคม คอน ขา ง นอย จ งควร มการศกษาพชชนดน ?มากย0งข ?นเพ0อนาไปสการใชประโยชนท0มประสทธภาพมากข ?น ตลอดจนสามารถสงเสรมใหเกดการปลกเพ0อการอนรกษและขยายพนธพชใหเปนท0รจกมากย0งข ?น
สารสาคญท0พบในผลของมะหวดท0มสมวงแดง ช0อวา แอนโทไซยานน (anthocyanins) ซ0งเปนเมดสท0ละลายน ?าได เปนสตามธรรมชาตท0มโครงสรางของโมเลกลอยในกลมฟลาโวนอยด (Chaovanalokit, 2011) และนอกจากน ?ยงอาจพบสารกลมโพลฟนอลอ0นๆ เชน สารประกอบฟนอลก เปนตน (Diaconeasa et al., 2015) สของแอนโทไซยานนจะเปล0ยนไปตามสภาวะความเปนกรด-ดาง แอนโทไซยานนเปนสารท0ใหสต ?งแตสน ?าเงนเขมหรออาจไมมสเลยเม0อในอย

RMUTSB Acad. J. 8(2) : 187-198 (2020) 189
สภาวะดาง (pH>7) เม0ออยในสภาวะท0เปนกลางจะมสมวง (pH=7) และจะเปล0ยนเปนสแดงถงแดงเขมไดในสภาวะเปนกรด (pH<7) (Giusti, & Wrolstad, 2001; Hurtado, Morales, Gonzalez-Miret, & Escudero-Gilete, 2009) สารในกลมแอนโทไซยานน ท0พบบอยมอย 6 ชนด (Cabrita, Fossen, & Andersen, 2000) ไดแก เพลาโกนดน (pelargonidin) ไซยานดน (cyanidin) เดลฟนดน (delphinidin) พโอนดน (peonidin) เพทนดน (petunidin) และ มาลวดน (malvidin) จากงานวจยท0มอยในปจจบน ช ?ใหเหนวาแอนโทไซยานน มสมบตเปนสารตานอนมลอสระ (antioxidant) ท0มฤทธGสงกวาวตามนซและอถงสองเทา (Lourith, Kanlayavattanakul, & Chanpirom, 2009; Aninbon, Banterng, Sanitchon, & Songsri, 2011) นอกจากน ?ยงมสมบตลดอาการอกเสบ (anti-inflammatory) ชวยปกปองหลอดเลอด ลดคอเลสเตอรอลในเลอดลดความเส0ยงของโรคมะเรงและตานไวรส (Wetwitayaklung, Charoenteeraboon, Limmatvapirat, & Phaechamud, 2012)
การวเคราะหปรมาณแอนโทไซยานนสามารถทาไดหลายวธ เชน การวดคาการดดกลนแสงท0ความยาวคล0นหน0งๆ และวธพเอช-ดฟเฟอเรนเชยล ซ0งในปจจบนวธหลงไดรบความนยมมากกวาวธแรกเน0องจากเปนวธท0มความถกตองแมนยาสงกวา (Lee, Durst, & Wrolstad, 2005; Shafazila, Pat, & Lee, 2010) โดยจะทาการวดคาการดดกลนแสงของแอนโทไซยานนในสารละลายบฟเฟอรท0 pH 1.0 และ 4.5 ซ0งแอนโทไซยานนจะมการเปล0ยนแปลงโครงสรางจากรปออกโซเนยม (oxonium form) ใน pH 1.0 กลายเปนรปเฮมคทาล (hemiketal form) ท0 pH 4.5 วธการน ?ทาการวดคาการดดกลนแสงของแอนโทไซยานน ณ ความยาว
คล0นสงสด (λmax) ท0 pH 1.0 และ 4.5 แลวนามาหกลบกนเพ0อเปนการกาจดการดดกลนแสงของสารอ0นๆ ท0ไมใชแอนโทไซยานน และวดคาการดดกลนแสงท0 700 นาโนเมตร เพ0อหกลบคาความขนท0อาจเกดข ?น (Chaovanalokit, 2011)
จากขอมลขางตน ผวจยคาดการณวาการนาผลของมะหวดมาพฒนาเปนเคร0องด0มน ?าผลไมเพ0อเพ0มคณคาทางโภชนาการใหกบผ บรโภค โดยคดเลอกอตราสวนท0เหมาะสมของมะหวดตอการผลตเปนน ?าผลไม และทดสอบหาปรมาณโพลฟนอลรวมและแอนโทไซยานนรวม เพ0อใหไดองคความรในการพฒนาผลตภณฑเคร0องด0มน ?ามะหวดและผลตภณฑใหมๆ ท0มประโยชนตอสขภาพและอาจจะสามารถนาไปพฒนาเปนผลตภณฑจาหนายในเชงพาณชยตอไป
วธการศกษา
การเตรยมตวอยางและศกษาคณภาพเบ �องตนบางประการดวยวธทางกายภาพของน �ามะหวด
นาผลมะหวดสกมาลางน ?าใหสะอาด ผ0งใหแหง นามาแยกเน ?อและเมลด จากน ?นนาเฉพาะสวนเน ?อมาค ?นน ?า โดยกาหนดอตราสวนระหวางน ?าหนกเน ?อมะหวดสกและน ?าหนกน ?าเปนอตราสวน 1:1, 1:2 และ 1:3 นาไปใหความรอนท0อณหภม 95 องศาเซลเซยส แตละอตราสวนจะมการปรบเวลาในการตมเปน 5, 10 และ 15 นาท ตามลาดบ ต ?งท ?งไวใหเยนและนามากรองผานผาขาวบาง จากน ?นนาไปกรองดวยกระดาษกรอง 1 คร ?ง นาไปเกบไวท0อณหภม 0 องศาเซลเซยส จนไดตวอยางจานวน 9 ตวอยาง (T1-T9) สาหรบนาไปวเคราะหคณภาพของน ?ามะหวดในเบ ?องตนดวยวธทางกายภาพ ไดแก คาความเปนกรด-ดาง (pH)

190 ว.มทรส. 8(2) : 187-198 (2563)
การตรวจวดสโดยวดความเขมของส ดวยเคร0 อง spectrocolorimeter ของแขงท ?งหมดท0ละลายได (total soluble solid : TSS) ดวยเคร0อง refractometer รวมท ?งการวเคราะหปรมาณสารตานอนมลอสระและฤทธG
การยบย ?งอนมลอสระตอไป
การวเคราะหปรมาณแอนโทไซยานนรวม วเคราะหโดยใชวธ pH differential (Shafazila,
Pat, & Lee, 2010) เจอจางตวอยางท ?ง 9 ตวอยาง ท0จะทดสอบ 1 สวนใน 9 สวนโดยปรมาตรของสารละลายบฟเฟอร 2 ชนด คอโพแทสเซยมคลอไรดบฟเฟอร (0.025 M KCl, pH 1.0) และอะซเตทบฟเฟอร (0.4 M CH3COONa, pH 4.5) นาสารละลายท0เตรยมไวไปเขยา และวดคาการดดกลนแสงท0 510 นาโนเมตร (A510) และ 700 นาโนเมตร (A700) ดวยเคร0องวด การดดกลนแสง (UV-Vis spectrophotometer) โดยทาการทดลอง 3 ซ ?า นาคาการดดกลนแสงท0ได ไปคานวณหาปรมาณแอนโทไซยานนรวม (total anthocyanin contents : TA) โดยแสดงขอมลปรมาณแอนโทไซยานนรวม ในหนวยของมลลกรมสมมลของ cyanidin-3-glucoside (cyanidin-3-glucoside equivalent: CGE) ตอตวอยางสด (FW) 100 กรม (mg CGE/ 100g of FW) ตามสมการตอไปน ?
( ) ( )1.0 4.5
510 700 510 700pH pHA = A - A - A - A (1)
( )A × Mw × dilution factor × 100
TA content = ε
(2)
โดยท0 A, A510 และ A700 หมายถง ผลตาง
ของคาการดดกลนแสงท0ความยาวคล0นสงสดของ
ตวอยางในสารละลายบฟเฟอร pH 1.0 และ 4.5 คาการดดกลนแสงท0ความยาวคล0นสงสดของ ตวอยางท0 510 นาโนเมตร และ คาการดดกลนแสง ท0ความยาวคล0นสงสดของตวอยางท0 700 นาโนเมตร ตามลาดบ
Mw หมายถง มวลโมเลกลของ cyanidin-3-glucoside ซ0งเทากบ 449.2 g/mol
ε หมายถง โมลารแอบซอบตวต (molar absorptivity) ซ0งข ?นกบชนดของแอนโทไซยานนและตวทาละลาย โดยท0วไป นยมใชคาของ cyanidin-3-glucoside ในสารละลายบฟเฟอร pH 1.0 ซ0งเทากบ 26,900 M.cm
dilution factor หมายถง แฟคเตอรท0เกดจากการเจอจางสารละลาย ซ0งในท0น ?ใชตวอยาง 1 มลลลตร เ จอจางดวยสารละลายบฟเฟอรจนมปรมาตร 10 มลลลตร ใชคา dilution factor เทากบ 10 การวเคราะหปรมาณฟนอลกรวม
วเคราะหปรมาณฟนอลกของสารสกดท ?งหมดโดยใชวธ Folin-Ciocalteu colorimetric method (Lourith, Kanlayavattanakul, & Chanpirom, 2009) โดยนาตวอยางท ?ง 9 ตวอยาง ปรมาตร 0.5 มลลลตร เตมสารละลาย Folin Ciocalteu reagent ปรมาตร 1.8 มลลลตร จากน ?นเตมสารละลาย 7.5 เปอรเซนต Na2CO3 ปรมาตร 1.2 มลลลตร แลวปรบปรมาตรดวยน ?ากล0นใหมปรมาตร 10 มลลลตร ผสมใหเขากน ต ?ง ท ?งไว 60 นาท ท0อณหภมหอง และวดคาการดดกลนแสงท0ความยาวคล0น 765 นาโนเมตร (A765) ดวยเคร0องวดการดดกลนแสง โดยทาการทดลอง 3 ซ ?า เปรยบเทยบคาการดดกลนแสงท0วดไดกบกราฟมาตรฐานของสารละลาย gallic acid (เตรยมสาร

RMUTSB Acad. J. 8(2) : 187-198 (2020) 191
มาตรฐาน gallic acid ท0ความเขมขน 100, 200, 300, 400 และ 500 ไมโครกรม/มลลลตร ในน ?ากล0น) แสดงในรปมลลกรม gallic acid equivalent/100 กรมของสารสกด (mg GAE/100 g crude extract)
การวเคราะหฤทธ�การตานอนมลอสระ วเคราะหโดยวธ DPPH (Lourith, Kanlayavattanakul,
& Chanpirom, 2009) นาตวอยางท ?ง 9 ตวอยางปรมาตร 0.5 มลลลตร เตมสารละลายมาตรฐาน DPPH ปรมาตร 2 มลลลตร ปรบปรมาตรใหเปน 10
มลลลตร ดวยเมทานอล 95 เปอรเซนต เขยาใหสารละลายผสมกน ต ?งท ?งไวใหเกดปฏกรยาในท0มด ณ อณหภมหองเปนเวลา 30 นาท แลวจงนาไปวดคาการดดกลนแสงท0ความยาวคล0น 525 นาโนเมตร (A525) ดวยเคร0 องวดการดดกลนแสง นาคาการดด ก ลน แ สง ข อ ง ตว อ ย า ง เ ป ร ย บ เ ท ย บ กบ ตว ควบคม (control) โดยทาการทดลอง 3 ซ ?า นาคาท0ไดไปคานวณหาเปอรเซนต scavenging activity ตามสมการ
control sample
control
A - ADPPH scavenging activity (%) = × 100
A (3)
Acontrol = คาการดดกลนแสงของตวควบคม Asample = คาการดดกลนแสงของตวอยาง
การวเคราะหฤทธ�การตานอนมลอสระ
วเคราะหโดยวธ ABTS (Kriengsak, Boonprakob, Crosby, Cisneros-Zevallos, & Byrne, 2006) เตรยม
สารละลาย ABTS•+ radical โดยนา 7 mM ABTS (2,2’ -azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) ในน ?ากล0น กบ 2.45 mM K2S2O8 (dipotassium peroxodisulphate) ในอตราสวน 1:1 ผสมใหเขากน ต ?งท ?งไวในท0มดท0อณหภมหอง เปนเวลา 12 ช0วโมง
จากน ?นนาสารละลาย ABTS•+ มาเจอจางในเมทานอล
95 เปอรเซนต ใหสารละลายมคาการดดกลนแสงอยในชวง 7.00±0.05 ท0ความยาวคล0น 734 นาโนเมตร
นาตวอยางท ?ง 9 ตวอยาง ป รมาตร 0.5
มลลลตร เตมสารละลาย ABTS•+ ท0เจอจางแลว 4.5 มลลลตร ต ?งท ?งไวในท0มดท0อณหภมหอง 30 นาท นาไปวดคาการดดกลนแสงเทยบกบตวควบคม ท0ความยาวคล0น 734 นาโนเมตร จากน ?นนาคาการดดกลนแสงท0ไดไปคานวณคา ABTS scavenging activity (%) ตามสมการ
control sample
control
A - AABTS scavenging activity (%) = × 100
A (4)
Acontrol = คาการดดกลนแสงของตวควบคม Asample = คาการดดกลนแสงของตวอยาง
192 ว.มทรส. 8(2) : 187-198 (2563)
เน0องจากกลไกการตานอนมลอสระสามารถเกดไดหลายรปแบบ เชน แบบฟรแรดคล แบบแรดคล-แคตไอออน จงควรทาการศกษาการตานอนมลอสระ 2 วธข ?นไป การวเคราะหทางสถต
นาผลการศกษามาวเคราะหหาความแปรปรวนทางสถต (analysis of variance: ANOVA) และวเคราะหหาคาความแตกตางของคาเฉล0ยโดยวธ Duncan’s new multiple’s range test (DMRT) และหาความสมพนธของขอมลทางสถตดวยวธ Pearson correlation test เพ0อเปนขอมลสาหรบการคดเลอกชดทดลองท0เหมาะสม
ไปศกษาปรมาณน ?ามะหวดตอปรมาณน ?าตาลท0เหมาะสมในการพฒนาเปนน ?ามะหวดพรอมด0มตอไป
ผลการศกษา
ผลการศกษาคณภาพเบ �องตนบางประการดวยวธทางกายภาพของน �ามะหวด
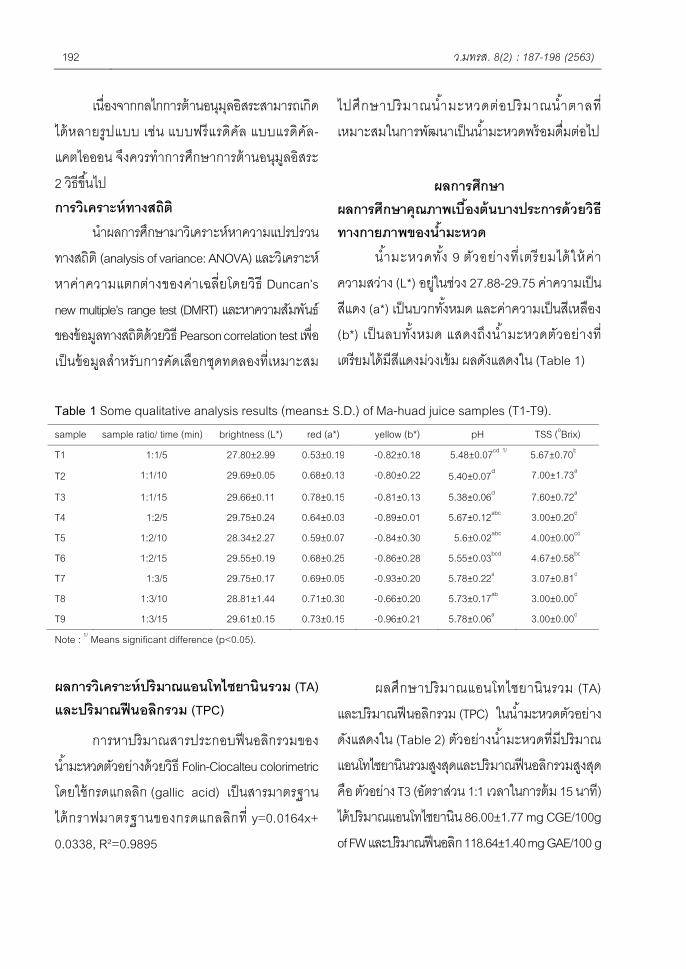
น ?ามะหวดท ?ง 9 ตวอยางท0เตรยมไดใหคาความสวาง (L*) อยในชวง 27.88-29.75 คาความเปนสแดง (a*) เปนบวกท ?งหมด และคาความเปนสเหลอง (b*) เปนลบท ?งหมด แสดงถงน ?ามะหวดตวอยางท0เตรยมไดมสแดงมวงเขม ผลดงแสดงใน (Table 1)
Table 1 Some qualitative analysis results (means± S.D.) of Ma-huad juice samples (T1-T9). sample sample ratio/ time (min) brightness (L*) red (a*) yellow (b*) pH TSS (oBrix) T1 1:1/5 27.80±2.99 0.53±0.19 -0.82±0.18 5.48±0.07cd 1/ 5.67±0.70b
T2 1:1/10 29.69±0.05 0.68±0.13 -0.80±0.22 5.40±0.07d 7.00±1.73a
T3 1:1/15 29.66±0.11 0.78±0.15 -0.81±0.13 5.38±0.06d 7.60±0.72a T4 1:2/5 29.75±0.24 0.64±0.03 -0.89±0.01 5.67±0.12abc 3.00±0.20d T5 1:2/10 28.34±2.27 0.59±0.07 -0.84±0.30 5.6±0.02abc 4.00±0.00cd T6 1:2/15 29.55±0.19 0.68±0.25 -0.86±0.28 5.55±0.03bcd 4.67±0.58bc T7 1:3/5 29.75±0.17 0.69±0.05 -0.93±0.20 5.78±0.22a 3.07±0.81d T8 1:3/10 28.81±1.44 0.71±0.30 -0.66±0.20 5.73±0.17ab 3.00±0.00d T9 1:3/15 29.61±0.15 0.73±0.15 -0.96±0.21 5.78±0.06a 3.00±0.00d Note : 1/ Means significant difference (p<0.05).
ผลการวเคราะหปรมาณแอนโทไซยานนรวม (TA) และปรมาณฟนอลกรวม (TPC)
การหาปรมาณสารประกอบฟนอลกรวมของน ?ามะหวดตวอยางดวยวธ Folin-Ciocalteu colorimetric โดยใชกรดแกลลก (gallic acid) เปนสารมาตรฐาน ไดกราฟมาตรฐานของกรดแกลลกท0 y=0.0164x+ 0.0338, R²=0.9895
ผลศกษาปรมาณแอนโทไซยานนรวม (TA) และปรมาณฟนอลกรวม (TPC) ในน ?ามะหวดตวอยาง ดงแสดงใน (Table 2) ตวอยางน ?ามะหวดท0มปรมาณแอนโทไซยานนรวมสงสดและปรมาณฟนอลกรวมสงสด คอ ตวอยาง T3 (อตราสวน 1:1 เวลาในการตม 15 นาท)ไดปรมาณแอนโทไซยานน 86.00±1.77 mg CGE/100g of FW และปรมาณฟนอลก 118.64±1.40 mg GAE/100 g

RMUTSB Acad. J. 8(2) : 187-198 (2020) 193
crude extract ในขณะท0ตวอยาง T8 เปนตวอยางท0มปรมาณแอนโทไซยานนและปรมาณฟนอลกต0าท0สดได 24.63±0.59 mg CGE/100g of FW และ 20.98±0.04 mg GAE/100 g crude extract ตามลาดบ
Table 2 Total anthocyanins (TA) and total phenolic contents (TPC) (means ± S.D.) of Ma-huad juice samples (T1-T9).
sample total anthocyanins (TA) (mg CGE/100g of FW)
total phenolic content (TPC) (mg GAE/100 g
crude extract) T1 47.38±2.66 62.32±0.38 T2 81.66±6.26 83.88±0.86 T3 86.00±1.77 118.64±1.40 T4 42.75±0.35 43.57±0.04 T5 44.46±3.36 89.21±2.85 T6 42.21±0.30 104.89±1.53 T7 33.36±1.48 35.85±0.14 T8 24.63±0.59 45.96±0.53 T9 33.56±0.94 65.37±0.40
ผลการทดสอบฤทธ�ตานอนมลอสระ ผลการทดสอบฤทธGการตานอนมลอสระดวยวธ
DPPH radical scavenging activity (Table 3) พบวาน ?ามะหวดตวอยาง T3 มฤทธGในการตานอนมลอสระสงท0สด รองลงมาคอตวอยาง T2 และ T1 ตามลาดบ คดเปนรอยละ 57.78, 55.22 และ 54.77 ตามลาดบ สวนตวอยาง T7 และ T8 เปนตวอยางท0มฤทธGในการตานอนมลอสระต0าท0สด คดเปนรอยละ 26.88 และ 28.27 ตามลาดบ ผลการทดสอบฤทธ�ตานอนมลอสระ
ผลการทดสอบฤทธGการตานอนมลอสระดวยวธ ABTS (Table 3) พบวาน ?ามะหวดตวอยาง T3 มฤทธGใน
การตานอนมลอสระสงท0สด รองลงมาคอตวอยาง T2 และ T6 ตามลาดบ คดเปนรอยละ 96.62, 95.38 และ 94.81 ตามลาดบ สวนตวอยาง T7 และ T8 เปนตวอยางท0มฤทธGในการตานอนมลอสระต0าท0สด คดเปนรอยละ 64.68 และ 76.16 ตามลาดบ
Table 3 DPPH and ABTS antioxidant activities (means ± S.D.) of Ma-huad juice samples (T1-T9).
sample % DPPH radical
scavenging activity % ABTS scavenging
activity T1 55.22±0.55 81.05±1.16 T2 54.77±0.73 95.38±1.17 T3 57.78±0.77 96.62±1.40 T4 39.91±0.88 81.77±0.81 T5 47.01±0.82 91.38±1.85 T6 54.49±0.12 94.81±1.53 T7 26.88±0.23 64.68±0.69 T8 28.27±0.59 76.16±0.85 T9 35.36±0.13 79.37±0.40
อภปรายผล
การศกษาคณภาพเบ �องตนบางประการดวยวธทางกายภาพของน �ามะหวด
การศกษาคณภาพเบ ?องตนบางประการดวยวธทางกายภาพของน ?ามะหวด ไดผลดงแสดงใน (Table 1) การวดคาส (L*, a* และ b*) ท0เปนคาท0นยมในการประเมนลกษณะปรากฏของตวอยาง ท0ทาการศกษา คา L* ท0เขาใกล 100 หมายถง ตวอยางมความสวางมากจนเปนสขาวหรอสจาง แตถาคา L* เขาใกล 0 หมายถง ตวอยางมความสวางนอยลง จนเปนสคล ?า คา a* ท0เปนบวก แสดงวาตวอยางเปนสแดง แตคา a* ท0เปนลบแสดงวาตวอยางเปนสเขยว

194 ว.มทรส. 8(2) : 187-198 (2563)
และคา b* ท0เปนบวกแสดงวาตวอยางเปนสเหลอง แตถาคา b* เปนลบแสดงวาตวอยางเปนสน ?าเงน (Choudhury, 2014) น ?ามะหวดท ?ง 9 ตวอยางท0เตรยมไดใหคาความสวาง (L*) อยในชวง 27.88-29.75 คาความเปนสแดง (a*) เปนบวกท ?งหมด และคาความเปนสเหลอง (b*) เปนลบท ?งหมด แสดงถงน ?าม ะ ห ว ด ตว อ ย า ง ท0 เ ต ร ย ม ไ ด ม ส แ ด ง ม ว ง เ ข ม เน0องมาจากแอนโทไซยานนท0เปนรงควตถท0พบในมะหวดท0ใหสแดง น ?าเงน มวง (Chaovanalokit, 2011)
การศกษาปจจยอตราสวนของมะหวดและน ?า ในอตราสวน 1:1, 1:2 และ 1:3 พบวาไมมผลตอคาสของน ?ามะหวด โดยไมมความแตกตางทางสถต (p>0.05) แตมผลตอคาความเปนกรดดาง และปรมาณของแขงท0ละลายไดท ?งหมด (p<0.05) ท ?งน ?เน0องจากปรมาณน ?าท0เพ0มทาใหปรมาณของแขงท0ละลายไดท ?งหมดถกเจอจางลงดวยน ?า ในขณะท0ปจจยระยะเวลาการตมสกดไมมผลตอคาสของน ?ามะหวด และคาความเปนกรดดาง (p>0.05) แตมผลตอปรมาณของแขงท0ละลายไดท ?งหมด (p<0.05) ท ?งน ?เน0องจากระยะเวลาการตมสกดท0เพ0มข ?นอาจทาใหเกดการระเหยของน ?า จงอาจทาใหปรมาณของแขงท ?งหมดเพ0มมากข ?น
ในการพฒนาน ?ามะหวดเพ0อเปนเคร0องด0มน ?าผลไมตอไป จาเปนท0จะตองวดคา pH คาความเปนกรด และสวนประกอบน ?าตาล (องศาบรกซ) เน0องจากตวแปรเหลาน ?มผลตอท ?งรสชาตและอายการเกบรกษาน ?าผลไมจาก (Table 1) จะเหนวา pH ของน ?ามะหวดตวอยางท0มอตราสวนเดยวกนจะใหผลท0ไมแตกตางกนอยางมนยสาคญทางสถต โดยมคาอยในชวง 5.38-5.78 ซ0งเม0อเปรยบเทยบกบปรมาณความเปนกรดดางของน ?า
มะหวดเร0มตน มคาเทากบ 5.34 อยในชวงใกลเคยงกน คาความเขมขนของน ?าผลไม (TSS หรอ oBrix) บอกถงปรมาณของแขงท0ละลายไดในน ?าท ?งหมดใชบงช ?ความเขมขนของอาหารเหลว เชน น ?าเช0อม น ?าผลไมเขมขน จากการทดลองพบวา มะหวดตวอยางท ?ง 9 ตวอยาง มคาความเขมขนของน ?าผลไม อยในชวง 3.00-7.60 oBrix โดยท0อตราสวนท0เขมขน 1:1 เวลา 10 และ 15 นาท (T2 และ T3) แสดงคาความเขมขนสงท0สดท0 7.00 และ 7.60 oBrix ตามลาดบ ระดบความเขมขนท0ไดสามารถระบคณภาพของน ?ามะหวดตวอยางไดวามระดบความหวานนอยและลกษณะปรากฏของน ?ามะหวดอยในระดบใสมาก (extra-light) (Thai industrial standards institute (TISI), 2006)
การวเคราะหปรมาณแอนโทไซยานนรวม (TA) และปรมาณฟนอลกรวม (TPC)
จากผลการทดลองเหนไดวา ปรมาณของสารประกอบฟนอลกรวมพบวา น ?ามะหวดตวอยาง T2, T3, T5 และ T6 ใหผลท0สงกวาสารตวอยางอ0นๆ โดยปรมาณฟนอลกท0 ไ ดอย ในชวง 83-119 mg GAE/100 g crude extract ซ0งคาท0ไดแตกตางจากงานของ Sonchai (2009) ท0ไดศกษาปรมาณสารประกอบ ฟนอลกจากสารสกดมะหวดดวยตวทาละลายเอทานอล โดยพบวาปรมาณฟนอลกเทากบ 24.96 mg/L gallic acid equivalent สวนปรมาณแอนโทไซยานนรวมของน ?ามะหวดตวอยาง T2 และ T3 สงกวาในตวอยางอ0นๆ ท ?งน ?อาจเปนเพราะคา pH ของมะหวดตวอยาง T2 และ T3 ต0ากวาตวอยางอ0นๆ เลกนอย ซ0งจากการศกษาของ Cabrita, Fossen, & Andersen (2000); Pajareon (2019) บงช ?วาในสภาวะกรด (pH เทากบ 1 และ 4) จะมความคงตวของแอนโทไซยานนดกวา

RMUTSB Acad. J. 8(2) : 187-198 (2020) 195
สภาวะกรดออน หรอกลาง และจะไมคงตวท0 pH เทากบ 9 สวน อตราสวนระหวางน ?าหนกเน ?อมะหวดและระยะเวลาในการตมสงผลตอปรมาณสารตานอนมลอสระท ?ง 2 ชนด โดยพบวาระยะเวลาท0ใชในการตมเพ0มข ?นอาจสงผลใหไดปรมาณสารประกอบ ฟนอลกเพ0มข ?น โดย Roobha, Saravanakumar, Aravindhan, & Devi (2011) ไดอธบายวาอณหภมสงจะสงผลใหโครงสรางของแอนโทไซยานนถกเปล0ยนแปลงไปเน0องจากการเกดปฏกรยาไฮโดรไลเซชนของน ?าตาลท0เกาะอยกบโครงสรางของแอนโทไซยานนทาใหเกดเปนโครงสรางท0มความคงตวนอยและกลายเปนสารประกอบสน ?าตาลของสารฟนอลกในธรรมชาตท0ไมละลายน ?าและไมคงตว ท ?งน ?จากการทดลองคร ?งน ? ท0ใชอณหภมสงในการตมน ?ามะหวดท ?ง 9 ตวอยาง อาจกลาวไดวาการใชระยะเวลา ไมเกน 10-15 นาท อาจสงผลตอปรมาณแอนโทไซยานนมากกวาปรมาณสารประกอบฟนอลก
การวเคราะหฤทธ�ตานอนมลอสระของน �ามะหวด จากผลการทดลองความสามารถในการตาน
อนมลอสระดวยวธ DPPH และ ABTS แสดงใหเหนวาปรมาณแอนโทไซยานนและปรมาณฟนอลกรวมท0สงจะมความสามารถในการตานอนมลอสระสงดวย สอดคลองกบงานวจยของ Pimrote, Teekayu, & Sudprasert (2020) นอกจากน ?นการใชอณหภมสงและระยะเวลาในการตมนาน มผลตอปรมาณสารตานอนมลอสระ ซ0ง Pajareon (2019) พบวาท0อณหภม 90 องศาเซลเซยส มผลทาใหแอนโทไซยานนสลายตวอยางมนยสาคญทางสถต (p<0.05) และพบวาคาความสามารถในการตานอนมลอสระดวยวธ ABTS สงกวา DPPH ท ?งน ?เน0องมาจากความแตกตาง
ของปฏกรยาท0ใชในการวเคราะห แตมแนวโนม ไปในทางเดยวกน ซ0งสอดคลองกบการศกษาของ Wangcharoen, & Morasuk (2007) ความสมพนธระหวางฤทธ� ตานอนมลอสระปรมาณ ฟนอลกรวม และปรมาณแอนโทไซยานนรวม
เม0อนาผลของปรมาณของสารประกอบฟนอลก รวมมาเปรยบเทยบกบประสทธภาพในการตานอนมลอสระของน ?ามะหวดตวอยาง T1-T9 ท ?ง 2 วธ ซ0งพบวา ตวอยาง T3 มฤทธGการตานอนมลอสระสงท0สด รองลงมาไดแก T2, T6 และ T5 ตามลาดบ ซ0งผลการวเคราะหการตานอนมลอสระท ?งจากวธ DPPH และ ABTS สอดคลองกบปรมาณสารประกอบฟนอลกรวมท0พบในตวอยาง เน0องจากการท0สารประกอบฟนอลกมคณสมบตเปนกรดออนจงทาใหตวอยาง T3 และ T2 มประสทธภาพใ น ก า ร ต า น อ นมล อ สร ะ สง ก ว า ตว อ ย า ง อ0 น ๆ (Diaconeasa et al., 2015) แตเม0อพจารณาปรมาณแอนโทไซยานนรวมพบวาในตวอยาง T3 และ T2 มปรมาณสงสดเปนลาดบท0 1 และ 2 ตามลาดบ แตในตวอยางอ0นๆ มปรมาณแอนโทไซยานนใกลเคยงกน ท ?งน ?อาจเปนเพราะในสภาวะท0สารสกดม pH เปนกรดมากกวาจะมความคงตวของแอนโทไซยานนดกวา (Cabrita, Fossen, & Andersen, 2000; Hurtado, Morales, Gonzalez-Miret, & Escudero-Gilete, 2009) นอกจากน ?เม0อวเคราะหทางสถตดวยวธ Pearson correlation test (Kriengsak, Boonprakob, Crosby, Cisneros-Zevallos, & Byrne, 2006) พบวา ความสมพนธระหวางปรมาณฟนอลกรวมและปรมาณแอนโทไซยานนรวมตอฤทธGการตานอนมลอสระดวยวธ DPPH และ ABTS มความสมพนธก นในระดบสง โดยมคาสมประสทธGสหสมพนธ (r) ดงแสดงใน (Table 4)

196 ว.มทรส. 8(2) : 187-198 (2563)
ซ0งจากขอมลดงกลาวสามารถบอกไดวาฤทธGในการต า น อ นม ล อ ส ร ะ ม ค ว า ม สม พน ธ กบ ป ร ม า ณสารประกอบฟนอลกรวมมากกวาปรมาณแอนโท- ไซยานนรวม เน0องจากสารสกดท0ไดเปนสารสกดหยาบท0มสารออกฤทธGหลากหลายชนดซ0งแตละชนดมกลไกในการตานอนมลอสระท0แตกตางกน และการทดสอบการตานอนมลอสระแตละวธกมกลไกท0แตกตางกน โดยในการทดสอบวธ DPPH และ ABTS เปนการทดสอบความสามารถของสารตานอนมลอสระในการดกจบอนมลอสระ ซ0งมกจะใหผลท0สอดคลองกน Table 4 Pearson’s correlation coefficients of DPPH and ABTS assays versus total phenolic content (TPC) and total anthocyanin content (TAC).
variable correlation coefficient (r) DPPH ABTS
TPC 0.88* 0.72* TAC 0.82* 0.64* Note: * Means significance at p<0.05.
สรป
จากการศกษาปรมาณฟนอลกรวมดวยวธ Folin-Ciocalteu reagent ปรมาณแอนโทไซยานนรวมดวยวธ pH differential และฤทธGการตานอนมลอสระดวยวธ DPPH และ ABTS ในน ?ามะหวดท0อตราสวนของน ?าหนกเน ?อมะหวดสกตอน ?า เปน 1:1, 1:2 และ 1:3 ท0ผานการตมท0อณหภม 95 องศาเซลเซยส เปนเวลา 5, 10 และ 15 นาท รวม 9 ตวอยาง พบวาตวอยาง T3 คออตราสวน 1:1 ท0ผานการตม 15 นาท มปรมาณฟนอลกรวม ปรมาณแอนโทไซยานนรวม
และฤ ทธG กา ร ต า น อ น ม ล อ ส ร ะ ด ว ย ว ธ DPPH และ ABTS สงท0สด สวนตวอยาง T2, T5, T6 และ T1 เปนตวอยางท0มฤทธGการตานอนมลอสระอยในกลมท0สงกวาตว อ0นๆ ท ?ง น ?อาจเปนเพราะปจจยตางๆ ท0เก0ยวของ เชน คา pH ของตวอยาง และระยะเวลาท0ใชในการตม เปนตน ทาใหฤทธGการตานอนมลอสระมความแตกตางกน และเม0อวเคราะหความสมพนธทางสถตระหวางฤทธGตานอนมลอสระ ปรมาณฟนอลกรวม และปรมาณแอนโทไซยานนรวม พบวา ความสมพนธระหวางปรมาณฟนอลกรวมและปรมาณแอนโทไซ-ยานนรวมตอฤทธGการตานอนมลอสระดวยวธ DPPH และ ABTS มความสมพนธกนในระดบสง ท0ระดบนยสาคญ 0.05 อยางไรกตาม สาหรบการจะพฒนาตอยอดผลตภณฑน ?ามะหวดเพ0อผลตเปนเคร0องด0มน ?าผลไม จงยงมปจจยอกหลายประการในระหวางก ร ะ บ ว น ก า ร แ ป ร ร ป น ?า ผลไ ม ท0 ม อ ท ธ พ ลต อ ปรมาณสารตานอนมลอสระและฤทธGในการยบย ?งอนมลอสระ ไมวาจะเปนการเกบรกษาวตถดบ กระบวนการลาง การใหความรอนและระยะเวลาท0เหมาะสม การบรรจภณฑและการเกบรกษา ท0สงผลตอปรมาณสารตานอนมลอสระในน ?าผลไม ดงน ?น ในการศกษาวจยคร ?งตอไป จงควรศกษาถงการเปล0ยนแปลงปรมาณสารประกอบฟนอลกและฤทธG
ตานอนมลอสระในข ?นตอนตางๆ ของการผลตน ?าผลไม เพ0อใหเปนแนวทางในการพฒนาการผลตน ?าผลไมใหไดประโยชนของสารตานอนมลอสระสงท0สด
คาขอบคณ
งานว จย เ ร0 อง น ? ไ ด รบทนสนบสนนจากสานกงานคณะกรรมการวจยแหงชาต (วช.) และ

RMUTSB Acad. J. 8(2) : 187-198 (2020) 197
มหาวทยาลย เทคโนโลย ราชมงคลสวรรณภม ปงบประมาณ 2559 นอกจากน ?คณะผ วจยตองขอขอบพระคณสาขาวทยาศาสตร ในการเอ ?อเฟ?อสถานท0ทาการวจย และทรพยากรสนบสนนงานวจยอนเปนประโยชนอยางย0งตองานวจย
เอกสารอางอง Aninbon, C., Banterng, P., Sanitchon, J., & Songsri, P.
(2011). A study of anthocyanin content in black glutinous rice using HPLC and spectrophotometric methods. Khonkaen AGR. J.,
39(Special issue), 353-357. BGO Plant Databases, The Botanical Garden Organization.
(n.d.). Mahuad. Retrieved 20 January 2018, from http://www.qsbg.org/database/botanic_book20full% 20option/search_detail.asp?botanic_id= 1497 (in Thai)
Cabrita, L., Fossen, T., & Andersen, O. M. (2000). Colour and stability of the six common anthocyanidin 3-glucosides in aqueous solutions. Food Chem., 68, 101-107.
Chaovanalokit, A. (2011). Extraction and analysis of anthocyanin. SWU J Sci. & Tech., 6, 26-36. (in Thai)
Choudhury, A. K. R. (2014). Principles of colour and
appearance measurement. Cambridge: Woodhead. Diaconeasa, Z., Ranga, F., Rugina, D., Leopold, L., Pop,
O., Vodnar, D., Cuibus, L., & Socaciu, C. (2015). Phenolic content and their antioxidant activity in various berries cultivated in Romania. Bulletin
UASVM Food Sci. & Tech., 72(1), 99-103. Giusti, M. M., & Wrolstad, R. E. (2001). Characterization
and measurement of anthocyanins by UV-visible spectroscopy. Current Protocols in Food
Analytical Chemistry, 00(1), F1.2.1-F1.2.13.
Hurtado, N. H., Morales, A. L., Gonzalez-Miret, M. L., & Escudero-Gilete, M. L. (2009). Color, pH stability and antioxidant activity of anthocyanin rutinosides isolated from tamarillo fruit (Solanum betaceum Cav.). Food Chem., 117, 88-93.
Kriengsak, T., Boonprakob, U., Crosby, K., Cisneros-Zevallos, L., & Byrne, D. H. (2006). Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Comp. Anal., 19, 669-675.
Lee, J., Durst, R. W., & Wrolstad, R. E. (2005). Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J.
AOAC Int., 88(5), 1269-1278.
Lourith, N., Kanlayavattanakul, M., & Chanpirom, S. (2009). Free radical scavenging efficacy of tamarind seed coat and its cosmetics applications. J.
Health Res., 23, 159-162.
Pajareon, S. (2019). Stability of anthocyanin extract from riceberry rice bran encapsulated with rice bran protein concentrate under different pH and heating conditions. RMUTSB Academic Journal, 7(2), 205-215. (in Thai)
Pimrote, K., Teekayu, K., & Sudprasert, P. (2020). Antioxidant activity and inhibition effect on Pseudomonas aeruginosa of extracts from Pa-Yom (Shorea roxburghii G. Don). RMUTSB
Academic Journal, 8(1), 15-27. (in Thai)
Roobha, J. J., Saravanakumar, M., Aravindhan, K. M., & Devi, P. S. (2011). The effect of light, temperature, pH on stability of anthocyanin pigments in Musa
acuminata bract. Res. Plant Biol., 1, 5-12.

198 ว.มทรส. 8(2) : 187-198 (2563)
Shafazila, T. S., Pat, M. L., & Lee, K. H. (2010). Radical scavenging activities of extract and solvent-solvent partition fractions from Dendrobium
Sonia ‘Red Bom’ flower. In Proceeding of International Conference on Science and Social
Research (pp. 762-765). Kuala Lumpur, Malaysia: Universiti Teknologi MARA.
Sonchai, B. (2009). Study on the radical scavenging
activity of Lepisanthes rubiginosa (Roxb.) Leenh.
wine and Melodorum fruticosum Lour. Wine
(Master’s thesis). Ubon Ratchathani Rajabhat University, Ubon Ratchathani.
Thai industrial standards institute (TISI). (2006). Draft
Thai industrial standards (TIS 112-2549) fruit
juice: pine juice. Retrieved 20 January 2018, from http://appdb.tisi.go.th/tis_dev/p3_tis/p3tis.php (in Thai)
Wangcharoen, W., & Morasuk, W. (2007). Antioxidant capacity and phenolic content of holy basil. Songklanakarin J. Sci. Technol., 29(5), 1407-1415.
Wetwitayaklung, P., Charoenteeraboon, J., Limmatvapirat, C., & Phaechamud, T. (2012). Antioxidant activities of some Thai and exotic fruits cultivated in Thailand. RJPBCS, 3(1), 12-21.

70
EnvironmentAsia 13(3) 2020 70-79DOI 10.14456/ea.2020.43
ISSN 1906-1714; ONLINE ISSN: 2586-8861The international journal by the Thai Society of Higher Education Institutes on Environment
EnvironmentAsia
AbstractSome yeast species can be applied for treatment of oil-contaminated wastewater. Oil contamination is one of the most serious problems in agro-industrial countries. Fat, oil and grease (FOG) are waste products from household food preparation and industrial processing. The aims of this study are to isolate natural lipolytic yeasts from ripe pineapple and select for high lipase activity on Tween 80 agar. The result showed that yeast isolate No.27 had high lipase activity. It had oval-shaped cells with circular form, and the entire margin of the creamy colored colony had raised elevation and a rough surface. The isolate was identifi ed as Candida sp. using the ITS specifi c sequencing region and named Candida sp. strain RMUTSB-27. This isolation grew well on YP medium containing 2% cooking oil waste as well as 2% glucose at 32°C. For oil-contaminated treatment, 2% palm oil waste mixed with wastewater was added Candida sp. RMUTSB-27 showed signifi cantly reduction in pH, fat, oil and grease, chemical oxygen demand (COD), and total Kjeldahl nitrogen (TKN) for 4 days. Our results indicated that the yeast Candida sp. RMUTSB-27 is benefi cial for lipid uptake and degradation of oil-contaminated wastewater and could be utilized for future green environmental treatment.
Keywords: Ripe pineapple; Natural lipolytic yeast; Oil-contaminated wastewater treatment; Yeast Candida sp.
The Natural Lipolytic Yeast Candida sp. RMUTSB-27Isolated from Pineapple for Treatment of Cooking Oil
Contaminated WastewaterPitchya Tangsombatvichit1*, Kanjana Pisapak2, and Paweena Suksaard1
1Department of Science (Microbiology), Faculty of Science and Technology Rajamangala University of Technology Suvarnabhumi, Pranakhon Si Ayutthaya, Thailand
2Department of Science (Chemistry), Faculty of Science and Technology, RajamangalaUniversity of Technology Suvarnabhumi, Pranakhon Si Ayutthaya, Thailand
*Corresponding Author, E-mail: [email protected]: February 6, 2020; Revised: May 28, 2020; Accepted: June 24, 2020
1. IntroductionOil-contaminated wastewater is now
considered as the most serious problem in agro-industrial countries. Fat, oil and grease (FOG) are waste products from household preparation and industrial processing activities that impact the environment. Oils and fats are lipids and composed of fatty acids, triacylglycerols, and lipid-soluble hydrocarbons as one of the most important components of FOG (Husain et al., 2014). Oils and fats cause environmental damage to water and aquatic organisms. These substances form an oil layer on the water surface that reduces
biological activity and leads to decreased dissolved oxygen levels (Abd El-Gawad, 2014).
For oil-contaminated wastewater treatment, one of the conventional techniques to remove oil and grease is by using oil and grease traps and skimming tanks; however, the disadvantage of this method is the low efficiency of removal. The remaining oil causes clogging of the pipes in treatment units and these require periodical cleaning and replacement, leading to increased maintenance (Husain et al., 2014). An alternative method of oil-contaminated waste management involves using microorganisms that can produce lipase

P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
71
enzymes and accumulate oils and fats in their cells. Advantages of this method include environmental safety and low-cost in the production stage. Microbial lipase activity can eliminate the wastewater pretreatment process through the production of enzymes from microorganisms. Ibegbulam-Njoku et al., (2014) proposed a new approach to degrade organic matter using a commercial mixture of lipase enzymes to cleanse holding tanks, grease traps, and other systems. Pseudomonas aeruginosa T1 isolated from a hot spring in Hokkaido, Japan efficiently degraded different types of fats and oils, including edible oil wastes, with high capacity to utilize free fatty acids. When induced by salad oil and oleic acid, this Gram-negative, rod-shaped bacterium secretes extracellular lipase (Hasanuzzaman et al., 2004).
Yeasts are one of the alternative microorganisms for oil-contaminated wastewater treatment. Different yeast communities have diverse habitats and populations, and changes in their component species should reflect conditions within habitats. Trichosporon sp., Rhodotorula sp., Candida sp. and Cryptococcus sp. can be used as indicator fungi to measure water pollution. Yeasts also have effective degradability of some refractory substances and organic poisons (Qadir, 2019). Some yeast strains are used in wastewater treatment because they can uptake lipid, fat, and oil into cells and produce lipase enzymes. The acetyl-CoA metabolism in yeast is highly compartmentalized and occurs in the cytosol, mitochondrion, peroxisome and nucleus. In the peroxisome, acetyl-CoA can then be generated from β-oxidation of fatty acids. Peroxisomal acetyl-CoA can then be used as a substrate in the glyoxylate cycle (Krivoruchko et al., 2015). Some yeasts can convert organic matter into non-toxic and single cell protein, which has high efficiency for wastewater treatment (Yang and Zheng, 2014). Wastewater that has high concentrations of chemical oxygen demand (COD) and biochemical oxygen demand (BOD) causes serious environmental pollution. To solve this problem, yeast treatment technology can be used to quickly reduce the oil content in
wastewater from 10,000 mg/L to 100 mg/L as a green alternative to the pretreatment process (Hu et al., 2003). Mafakher et al., (2010) investigated the ability of yeasts Yarrowia lipolytica (M1 and M2) produce valuable products such as lipase and citric acid. These yeast strains produced maximum lipase levels of 11 U/mL (Y. lipolytica M1) and 8.3 U/mL (Y. lipolytica M2), on olive oil fermentation medium, and high levels of citric acid at 27 g/L (Y. lipolytica M1) and 8 g/L (Y. lipolytica M2), on citric acid fermentation medium. These abilities of yeasts can be used in oil waste treatments in many agro-industries. Moreover, the yeast Y. lipolytica is an oleaginous yeast that uses synthetic and food waste-derived volatile fatty acids as substrates (lipid degradation) and can adapt to survive under the high temperature and acidity involved in the biodiesel production process (Gao et al., 2017). Y. lipolytica is frequently isolated from different fermented food, soils, sewage, and natural environments. It also reduces organic acid products in olive mills, palm oil mills, and wastewater treatments. These applications involve yeast lipase activity. Production of lipase in Y. lipolytica is limited and its genetic modification might not be accepted; however, it can still benefit the environment and waste treatment through many processes and applications (Aloulou et al., 2007).
Nowadays, biological wastewater treatment methods are important for environmental safety. They are better than chemical treatment methods currently used in households and industries. Hence, the objectives of this experiment were to isolate and characterize lipolytic yeasts from ripe pineapple (Ananas comosus) fruit that is popular and cheap in Thailand. Yeasts were found from ripe pineapple in the course of natural fermentation. Next, one isolated yeast was selected that exhibited a high level of lipase activity. This isolated yeast was investigated for its cooking oil waste utilization efficiency. Finally, the potent isolated yeast was examined for its capability in oil-contaminated wastewater treatment compared to the well-known oleaginous yeast Yarrowia lipolytica TISTR5212.

P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
72
2. Materials and Methods
2.1 Yeast isolation and lipase activity test
R i p e ‘ P h u l a e ’ p i n e a p p l e f r o m Ay u t t h a y a M a r k e t , T h a i l a n d w a s peeled, cleaned with dist i l led water and fermented overnight for analysis. 10 g samples were cultured on a liquid medium containing 1% yeast 2% peptone (YP medium) and 2% oleic acid. The medium was shaken a t 150 rpm a t ambient temperature (32-33°C) for 24 hours. Next, the samples were serially diluted and spread on YP medium plates with 2% oleic acid containing 0.05 mg/mL of chloramphenicol to inhibit bacterial growth (30-300 colonies). The colonies were selected and streaked on YP medium plates with 2% oleic acid for five generations. The method was modified from Tan Gana et al., (2014) by selecting observed colonies on Tween 80 agar containing calcium chloride. Lipase activity in the colonies was compared to that in Saccharomyces cerevisiae (non-lipase producing strain). The plates were then incubated at temperature of 32°C for 3 days in triplicate. Lipase activity was indicated by the appearance of a visible precipitate resulting from the deposition of calcium crystal salts formed by fatty acid liberated by extracellular lipase.
2.2 Isolated yeast strain morphology, physiology and sequencing
The yeast strain was selected based on high lipase activity and determined by standard morphological and physiological methods. The strain was analyzed under an optical microscopy (Olympus) to identify the yeast strain colony as described in Tan Gana et al. (2014). Then, DNA in the selected yeast strain was extracted using a DNA extraction method modified from Tangsombatvichit et al. (2015) and purified. Finally, the internal transcribed spacer (ITS) region was identified using ITS1 5’ TCCGTAGGTGAACCTGCGG 3’ and ITS4 5’ TCCTCCGCTTATTGA TATGC 3’ (Macrogen sequencing).
2.3 Spot test of used household oil wastes
S. cerevisiae, isolated Candida sp. RMUTSB-27 and Y. lipolytica were grown in liquid yeast peptone dextrose (YPD medium) overnight. Cells were spun, suspended in distilled water and diluted to an optical density (OD600) of 0.1. Next, they were serially diluted at 104, 103, 102 and 101 cells and spotted on appropriated plates. Finally, growth assays were monitored on oil wastes such as palm oil, lard oil, soybean oil, sunflower seed oil, and rice bran oil compared to the growth on glucose at temperature of 32°C for 4 days. Growth assays were conducted in triplicate (modified from Tangsombatvichit et al., 2015).
2.4 The growth of selected yeast on used palm oil waste
Isolated Candida sp. RMUTSB-27 andY. lipolytica were grown in liquid yeast peptone dextrose (YPD medium) overnight. Cells were spun, suspended in distilled water and diluted to an optical density (OD600) of 0.1. Next, the yeast cells were inoculated in YP medium, adding 2% glucose or 2% palm oil waste with shaking at 150 rpm at ambient temperature (32-33°C) for 96 hours. One milliliter of each sample was taken and optical density was measured every 12 hours. All samples were performed in triplicate (Jarboui et al., 2012).
2.5 Utilization of selected yeast for oil contaminated wastewater treatment
Wastewater from the canteen at
Rajamangala University of Technology Suvarnabhumi (aerobic wastewater) kept in a refrigerator at 4°C was prepared for the oil wastewater treatment test. Two percent of palm oil waste from household cooking was added and homogenized with the wastewater in 3 L Erlenmeyer flask. Next, the isolated Candida sp. RMUTSB-27 and Y. lipolytica (lipase producing strain) were grown in liquid yeast peptone dextrose (YPD medium) overnight. Cells were spun, suspended in distilled water and diluted to an optical density (OD600), starting with 0.1. Each yeast
P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
73
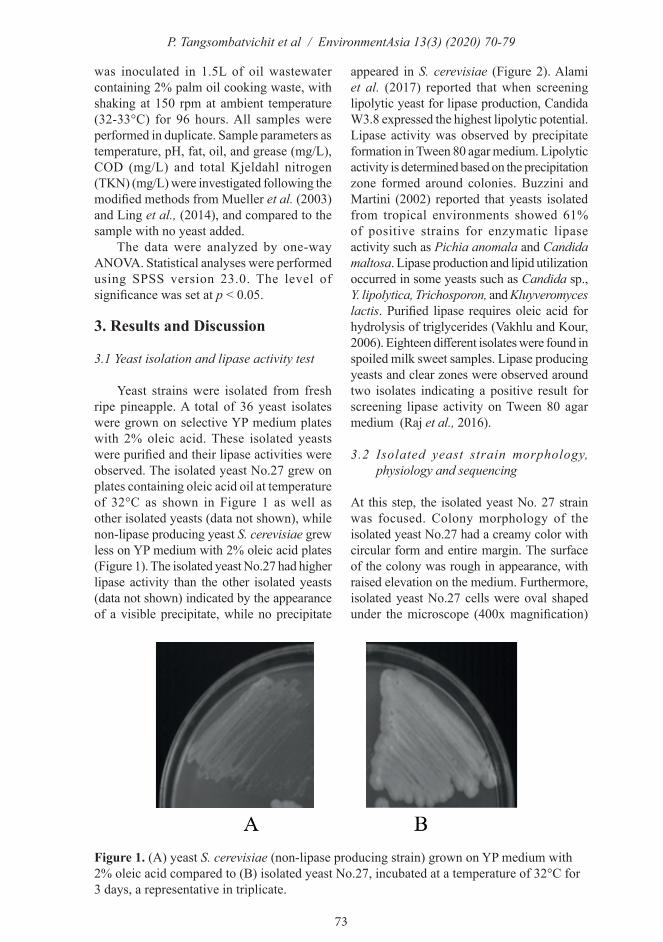
Figure 1. (A) yeast S. cerevisiae (non-lipase producing strain) grown on YP medium with 2% oleic acid compared to (B) isolated yeast No.27, incubated at a temperature of 32°C for 3 days, a representative in triplicate.
was inoculated in 1.5L of oil wastewater containing 2% palm oil cooking waste, with shaking at 150 rpm at ambient temperature (32-33°C) for 96 hours. All samples were performed in duplicate. Sample parameters as temperature, pH, fat, oil, and grease (mg/L), COD (mg/L) and total Kjeldahl nitrogen (TKN) (mg/L) were investigated following the modified methods from Mueller et al. (2003) and Ling et al., (2014), and compared to the sample with no yeast added.
The data were analyzed by one-way ANOVA. Statistical analyses were performed using SPSS version 23.0. The level of significance was set at p < 0.05.
3. Results and Discussion
3.1 Yeast isolation and lipase activity test
Yeast strains were isolated from fresh ripe pineapple. A total of 36 yeast isolates were grown on selective YP medium plates with 2% oleic acid. These isolated yeasts were purified and their lipase activities were observed. The isolated yeast No.27 grew on plates containing oleic acid oil at temperature of 32°C as shown in Figure 1 as well as other isolated yeasts (data not shown), while non-lipase producing yeast S. cerevisiae grew less on YP medium with 2% oleic acid plates (Figure 1). The isolated yeast No.27 had higher lipase activity than the other isolated yeasts (data not shown) indicated by the appearance of a visible precipitate, while no precipitate
appeared in S. cerevisiae (Figure 2). Alami et al. (2017) reported that when screening lipolytic yeast for lipase production, Candida W3.8 expressed the highest lipolytic potential. Lipase activity was observed by precipitate formation in Tween 80 agar medium. Lipolytic activity is determined based on the precipitation zone formed around colonies. Buzzini and Martini (2002) reported that yeasts isolated from tropical environments showed 61% of positive strains for enzymatic lipase activity such as Pichia anomala and Candida maltosa. Lipase production and lipid utilization occurred in some yeasts such as Candida sp., Y. lipolytica, Trichosporon, and Kluyveromyces lactis. Purified lipase requires oleic acid for hydrolysis of triglycerides (Vakhlu and Kour, 2006). Eighteen different isolates were found in spoiled milk sweet samples. Lipase producing yeasts and clear zones were observed around two isolates indicating a positive result for screening lipase activity on Tween 80 agar medium (Raj et al., 2016).
3.2 Isolated yeast strain morphology, physiology and sequencing
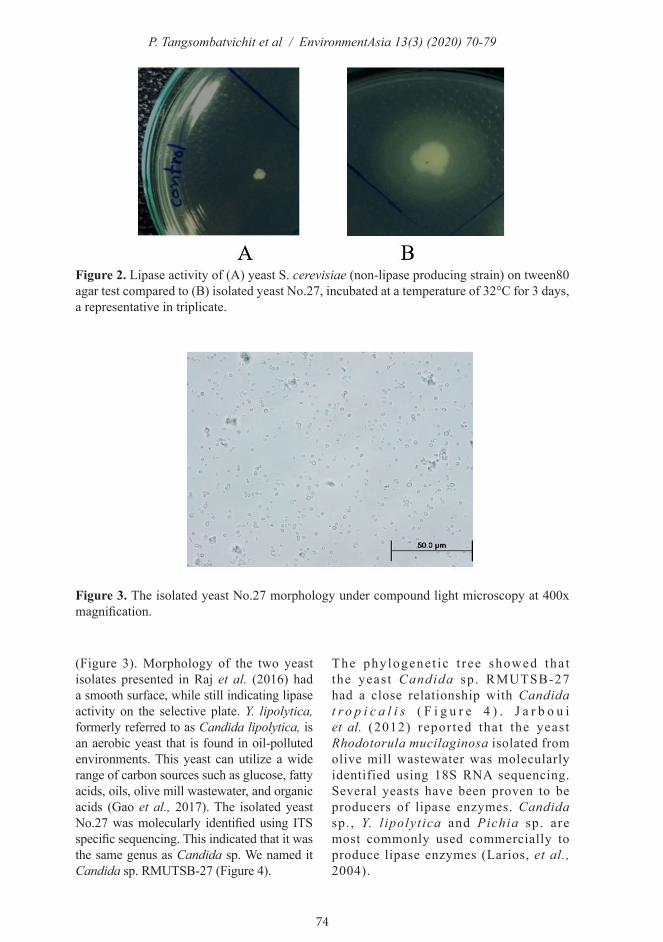
At this step, the isolated yeast No. 27 strain was focused. Colony morphology of the isolated yeast No.27 had a creamy color with circular form and entire margin. The surface of the colony was rough in appearance, with raised elevation on the medium. Furthermore, isolated yeast No.27 cells were oval shaped under the microscope (400x magnification)
P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
74
(Figure 3). Morphology of the two yeast isolates presented in Raj et al. (2016) had a smooth surface, while still indicating lipase activity on the selective plate. Y. lipolytica, formerly referred to as Candida lipolytica, is an aerobic yeast that is found in oil-polluted environments. This yeast can utilize a wide range of carbon sources such as glucose, fatty acids, oils, olive mill wastewater, and organic acids (Gao et al., 2017). The isolated yeast No.27 was molecularly identified using ITS specific sequencing. This indicated that it was the same genus as Candida sp. We named it Candida sp. RMUTSB-27 (Figure 4).
The phylogene t ic t ree showed tha t the yeast Candida sp . RMUTSB-27 had a close relationship with Candida t r o p i c a l i s ( F i g u r e 4 ) . J a r b o u i et al. (2012) reported that the yeast Rhodotorula mucilaginosa isolated from olive mill wastewater was molecularly identified using 18S RNA sequencing. Several yeasts have been proven to be producers of lipase enzymes. Candida sp. , Y. l ipolytica and Pichia sp. are most commonly used commercially to produce lipase enzymes (Larios, et al., 2004).
Figure 2. Lipase activity of (A) yeast S. cerevisiae (non-lipase producing strain) on tween80 agar test compared to (B) isolated yeast No.27, incubated at a temperature of 32°C for 3 days, a representative in triplicate.
Figure 3. The isolated yeast No.27 morphology under compound light microscopy at 400x magnification.

P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
75
Figure 4. The isolated yeast No.27 identified the phylogenetic tree using ITS specific sequencing as Candida sp.
3.3 Spot test of used household oil wastes
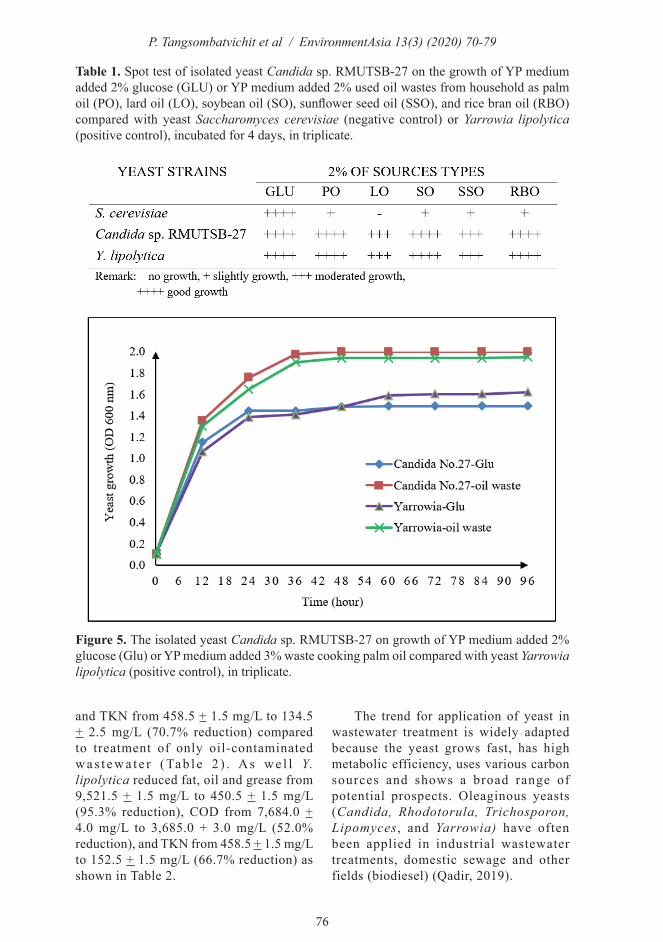
The isolated yeast No.27 was observed that it could utilize oil wastes from households. Lipid degradation was observed using the spot test method. The results indicated that the isolated yeast Candida sp. RMUTSB-27 could grow on a medium containing 2% of palm oil waste, lard oil waste, soybean oil waste, sunflower seed oil waste, and rice bran oil waste as well as 2% glucose (normal carbon source). Moreover, the isolated yeast Candida sp. RMUTSB-27 grew as well as Y. lipolytica (Table 1). In contrast, S. cerevisiae grew slightly or no grew on the cooking oil wastes, while it could grow on 2% glucose (Table 1). The oleaginous yeast Candida parapsilosis could uptake or synthesize fatty acid involved in OLE1 that leads to rapid accumulation of lipid droplets in the peroxisome. Deletion of Candida OLE1 (ole1Δ/Δ) results in impaired growth on oleic acid in yeast cells compared to that in wild-type cells since the OLE1 gene is important in the β-oxidation of fatty acid utilization (Nguyen and Nosanchuk, 2011).
3.4 The growth of selected yeast on used oil waste
The isola ted yeas t Candida sp . RMUTSB-27 could grow on 3% waste cooking palm oil at an optical density equal to 2.0 in 36 hours as well as the oleaginous yeast Y. lipolytica (Figure 5).
However, both yeast strains grew better on 3% waste cooking palm oil than 2% glucose, as shown in Figure 5. Progressive cell growth of yeast R. mucilaginosa increased the ability of the yeast to grow on olive mill wastewater as the sole carbon and energy source (Jarboui et al., 2012). Oleaginous yeasts can use lipids as substrates and accumulate lipid droplets in their biomass. These yeasts can also utilize a wide range of fatty acids for growth via a β-oxidation process in the peroxisome (Krivoruchko et al., 2015). Therefore, our result indicated that the natural isolated yeast Candida sp. RMUTSB-27 was more beneficial for lipid uptake and synthesis in cells for growth as well as Y. lipolytica and other oleaginous yeasts.
3.5 Utilization of selected yeast for oil contaminated wastewater treatment
All used oil-contaminated wastewater treatments indicated similar results at ambient temperature of 33°C (Table 2). The pH of oil wastewater treatment adding Candida sp. RMUTSB-27 isolate decreased from 8.6 to 6.9 as well as for Y. lipolytica (Table 2), while treatment of only oil-contaminated wastewater showed pH 8.6. Interestingly, our results presented that the isolated yeast Candida sp. RMUTSB-27 significantly reduced fat, oil and grease from 9,521.5 + 1.5 mg/L to 350.0 + 1.0 mg/L (96.3% reduction), COD from 7,684.0 + 4.0 mg/L to 3,841.5 + 1.5 (50.0% reduction),
P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
76
Table 1. Spot test of isolated yeast Candida sp. RMUTSB-27 on the growth of YP medium added 2% glucose (GLU) or YP medium added 2% used oil wastes from household as palm oil (PO), lard oil (LO), soybean oil (SO), sunflower seed oil (SSO), and rice bran oil (RBO) compared with yeast Saccharomyces cerevisiae (negative control) or Yarrowia lipolytica (positive control), incubated for 4 days, in triplicate.
Figure 5. The isolated yeast Candida sp. RMUTSB-27 on growth of YP medium added 2% glucose (Glu) or YP medium added 3% waste cooking palm oil compared with yeast Yarrowia lipolytica (positive control), in triplicate.
and TKN from 458.5 + 1.5 mg/L to 134.5 + 2.5 mg/L (70.7% reduction) compared to treatment of only oil-contaminated w a s t e w a t e r ( Ta b l e 2 ) . A s w e l l Y. lipolytica reduced fat, oil and grease from 9,521.5 + 1.5 mg/L to 450.5 + 1.5 mg/L (95.3% reduction), COD from 7,684.0 + 4.0 mg/L to 3,685.0 + 3.0 mg/L (52.0% reduction), and TKN from 458.5 + 1.5 mg/L to 152.5 + 1.5 mg/L (66.7% reduction) as shown in Table 2.
The trend for application of yeast in wastewater treatment is widely adapted because the yeast grows fast, has high metabolic efficiency, uses various carbon sources and shows a broad range of potential prospects. Oleaginous yeasts (Candida, Rhodotorula, Trichosporon, Lipomyces, and Yarrowia) have often been applied in industrial wastewater treatments, domestic sewage and other fields (biodiesel) (Qadir, 2019).

P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
77
Nine mixed yeast strains isolated from industrial wastewater have been used for biological processing treatment of soybean oil wastewater. From an influent concentration of COD, BOD and oil at 39,300 mg/L, 18,200 mg/L and 11,900 mg/L, respectively, the removal efficiencies of COD, BOD and oil-contaminated using the wastewater treatment process of yeasts were all more than 93% (Chigusa et al., 1996). The yeast R. mucilaginosa was studied for its ability to degrade phenolic compounds (highly toxic in the environment) and growth on olive mill wastewater. This isolated yeast reduced COD and phenols. At an initial COD concentration of 6,500 mg/L, the yeast strain decreased COD to 4,520 mg/L in 3 days of treatment (Jarboui et al., 2012). Nowadays, technological combinations between microalgae and yeasts are used for mixed wastewater treatment. Microalgae produce oxygen for the yeasts, while yeasts provide carbon dioxide for the microalgae. The yeasts uptake organic matter and the microalgae require nitrogen and phosphorus from the wastewater. These processes greatly improve both lipid production and removal of organic matter and nutrients from the wastewater. The lipid content and yield achieved were 63.45% and 4.60 g/L, while removal rates of COD, total nitrogen (TN), and total phosphorus (TP) were 95.34%, 51.18% and 89.29%, respectively (Ling et al., 2014). Thus, the yeast Candida sp. RMUTSB-27 can be combined with microalgae for more efficient oil wastewater treatment in the future.
4. Conclusion
The results from this study showed that the yeast strain isolated from ripe pineapple has the ability to degrade various cooking oil wastes. This yeast was identified as a Candida sp., using ITS specific sequencing. Cell growth was observed on used cooking palm oil, lard oil, soybean oil, sunflower seed oil and rice bran oil from households as the sole carbon source. The isolated yeast strain could be applied in used oil-contaminated wastewater treatment to reduce water pollution for domestic and industrial purposes. Furthermore, this strain could also be applied in lipid production for biodiesel to replace crude oil supplies. Yeast oils have advantages of being non-competitive with human and animal food. They have a short process cycle, and are independent of climatic factors compared to plant and animal oils.
References
Abd El-Gawad HS. Oil and grease removal from industrial wastewater using new utility approach. Advances in Environmental Chemistry 2014: 1-6.
Alami NH, Nasihah L, Umar RLA, Kuswytasari D, Zulaika E and Shovitri M. Lipase production in lipolytic yeast from Wonorejo mangrove area. Proceeding of international biology conference 2016: Biodiversity and Biotechnology for human welfare, AIP Conference Proceedings 1854, 020001 2017; https://doi.org/10.1063/ 1.4985392.
Table 2. Results of oil wastewater treatment using isolated yeast Candida sp. RMUTSB-27 compared with Yarrowia lipolytica. The wastewater was added 2% palm oil cooking waste from the household. Each parameter was analyzed in duplicate, for 4 days.

P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
78
Aloulou A, Puccinelli D, De Caro A, Leblond Y, Carriere F. A comparative study on two fungal lipases from Thermomyces lanuginosus and Yarrowia lipolytica shows the combined effects of detergents and pH on lipase adsorption and activity. Biochimica et Biophysica Acta 2007; 1771 (12): 1446–1456.
Buzzini P, Mart ini A. Extracel lular enzymatic activity profiles in yeast and yeast‐like strains isolated from tropical environments. Journal of Applied Microbiology 2002; 93 (6): 1020-1025.
Chigusa K, Hasegawa T, Yamamoto N, Watanabe Y. Treatment of wastewater from oil manufacturing plant by yeasts. Water Science and Technology 1996; 34 (11): 51-58.
Gao R, Li Z, Zhou X, Cheng S, Zheng L. Oleaginous yeast Yarrowia lipolytica culture with synthetic and food waste derived volatile fatty acids for lipid production. Biotechnology for Biofuels 2017; 10: 247-261.
Hasanuzzaman M, Umadhay-Briones KM, Zsiros SM, Morita N, Nodasaka Y, Yumoto I, Okuyama H. Isolation, identification, and characterization of a novel, oil-degrading bacterium, Pseudomonas aeruginosa T1. Current microbiology 2004; 49 (2): 108-114.
Hu JY, Ong SL, Ng WJ, Lu F, Fan XJ. A new method for characterizing deni t r i fying phosphorus removal bacter ia by using three different types of electron acceptors. Water R e s e a r c h 2 0 0 3 ; 3 7 ( 1 4 ) : 3 4 6 3 -3471.
Husain I, Alkhatib M, Jami MS, Mirghani ME, Bin Zainudin Z, Hoda A. Problems, control, and treatment of fat, oil, and grease (FOG): A review. Journal of Oleo Science 2014; 63 (8): 747-752.
I b e g b u l a m - N j o k u P N , A c h i O K , Chi j ioke-Osuj i CC. Use of palm oi l mi l l eff luent as fermentat ive m e d i u m b y l i p a s e p r o d u c i n g . International Journal of Scientific and Engineering Research 2014; 5 (2): 1631-1640.
Jarboui R, Baati H, Fetoui F, Gargouri A, Gharsallah N, Ammar E. Yeast performance in wastewater treatment: case study of Rhodotorula mucilaginosa. Environmental Technology 2012; 33 (7-9): 951-960.
Krivoruchko A, Zhang Y, Siewers V, Chen Y, Nielsen J. Microbial acetyl-CoA metabolism and metabolic engineering. Metabolic Engineering 2015; 25: 28-42.
Larios A, Garcia HS, Oliart RM, Valerio-Alfaro G. Synthesis of flavor and fragrance esters using Candida Antarctica lipase. Applied Microbiology and Biotechnology 2004; 65 (4): 373-376.
Ling JY, Nip S, Cheok WL, de Toledo RA, Shim H. Lipid production by a mixed
culture of oleaginous yeast and microalga from distillery and domestic mixed wastewater. Bioresource Technology 2014; 173: 132-139.
Mafakher L, Mirbafheri M, Darvishi F, Nahvi I, Zarkesh-Esfahani H, Emtiazi G. Isolation of lipase and citric acid producing yeasts from agro-industrial wastewater. New Biotechnology 2010; 27 (4): 337-340.
Mueller SA, Kim BR, Anderson JE, Gaslightwala A, Szafranski MJ, Gaines WA. Removal of oil and grease and chemical oxygen demand from oily automotive wastewater by adsorption after chemical de-emulsification. Practice Periodical of Hazardous Toxic and Radioactive Waste Management 2003; 7 (3): 156-162.
Nguyen LN, Nosanchuk JD. Lipid droplet formation protects against gluco/lipotoxicity in Candida parapsilosis: An essential role of fatty acid desaturase Ole1. Cell cycle 2011; 10 (18): 3159-3167.
Qadir G. Yeast a magical microorganism in the wastewater treatment. Journal of Pharmacognosy and Phytochemistry 2019; 8 (4): 1498-1500.
Raj A, Baby G, Dutta S, Sarkar A, Rao B. Isolation, characterization and antioxidant activity of lipase enzyme producing yeasts isolated from spoiled sweet sample. Scholars Research Library Der Pharmacia Lettre 2016; 8 (10): 129-135.

P. Tangsombatvichit et al / EnvironmentAsia 13(3) (2020) 70-79
79
Tan Gana NH, Mendoza BC, Monsalud RG. Isolation, screening and characterization of yeasts with amylolytic, lipolytic, and proteolytic activities from the surface of philippine bananas (Musa spp.). Philippine Journal of Science 2014; 143: 81-87.
Tangsombatvichit P, Semkiv MV, Sibirny AA, Jensen LT, Ratanakhanokchai K, Soontorngun N. Zinc cluster protein Znf1, a novel transcription factor of non-fermenta t ive metabol i sm in Saccharomyces cerevisiae. FEMS Yeast Research 2015; 15 (2): 1-16.
Vakhlu J, Kour A. Yeast lipases: enzyme purification, biochemical properties and gene cloning. Electronic Journal of Biotechnology 2006; 9 (1): 69-85.
Yang M, Zheng SK. Pollutant removal-oriented yeast biomass production from high-organic-strength industrial wastewater: A review. Biomass and Bioenergy 2014; 64: 356-362.

RMUTSB Acad. J. 8(2) : 165-175 (2020) 165
การคดแยกแบคทเรยท ผลตเอนไซมเซลลเลสจากตวอยางดนนาขาว ในจงหวดพระนครศรอยธยา
The isolation of cellulase producing bacteria from paddy field soil samples in Phra Nakhon Si Ayutthaya province
พรพรรณ รตนะสจจะ1* และ อษณย ทองดF
Pornpan Ruttanasutja1* and Usanee ThongdeeF
บทคดยอ การศกษาวจยในคร �งน �มวตถประสงคเพ!อคดแยกแบคทเรยท!มความสามารถในการผลตเอนไซมเซลลเลสจากตวอยางดน
นาขาวในจงหวดพระนครศรอยธยา ตวอยางดนนาขาวในเขตพ �นท! 5 อาเภอ ไดแก บางซาย บางบาล ทาเรอ ภาช และวงนอย นามาคดแยกแบคทเรยท!มความสามารถในการสรางเอนไซมเซลลเลสดวยวธการเจอจางแบบลาดบสวนบนอาหาร cellulose-degrading culture medium พบการเจรญของโคโลนแบคทเรยอยในชวง 1.06-6.87×106 CFU/g และสามารถคดแยกแบคทเรยไอโซเลทท �งหมด จานวน 26 ไอโซเลท ลกษณะทางสณฐานวทยาของแบคทเรยท!แยกไดสวนใหญเปนแบคทเรยแกรมบวกรปแทง (รอยละ 80.77) ผลจากการทดสอบความสามารถในการสรางเอนไซมเซลลเลสบนอาหาร carboxymethyl cellulose agar พบวามเพยง 4 ไอโซเลท (รอยละ 15.38) ท!สรางเอนไซมเซลลเลสเกดเปนวงใสรอบโคโลนขนาดมากกวาแบคทเรยควบคม Paenibacillus curdlanolyticus strain B-6 ซ!งไอโซเลท PC-C-1 มขนาดวงใสสงสด (2.56±1.03 เซนตเมตร) นอกจากน �เม!อนา ไอโซเลท PC-C-1 ไปทดสอบการยอยสลายฟางขาวดวยการเพาะเล �ยงในอาหาร nutrient broth ท!มการเตมฟางขาวลงไปรอยละ 5 ของปรมาตรอาหาร และจานวนแบคทเรยเร!มตนท! 1×108 เซลล/มลลลตร สามารถยอยฟางขาวไดสงถงรอยละ 64.80 จาก ผลการศกษาดงกลาวสามารถใชเปนขอมลเบ �องตนในการศกษาเพ!มเตมสาหรบการนาไปประยกตใชแบคทเรยในการยอยสลายฟางขาวจรง ซ!งเปนการลดวสดเหลอท �งทางการเกษตรตอไปในอนาคตได คาสาคญ: แบคทเรย เอนไซมเซลลเลส เอนไซมเอกซตราเซลลลาร อาหารทดสอบ carboxymethyl cellulose agar (CMC agar)
Abstract
The objective of this research was to isolate cellulase producing bacteria from rice paddy soil samples in Phra Nakhon Si Ayutthaya province. Soil samples of rice fields in five districts, namely Bangzai, Bangban, Tharuea, Phachi and Wangnoi, were used to screen for cellulase producing bacteria by serial ten-fold dilution method on cellulose-degrading culture medium. The bacterial colonization showed in the range of 1.06-6.87×106 CFU/g, and twenty-six isolates of bacteria were isolated. Morphological characterization of most isolates was gram-positive rod-
1 คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม 1 Faculty of Science and Technology, Rajamangala University of Technology Suvarnnabhumi * Corresponding author. E-mail: [email protected] Received: April 25, 2020; Revised: June 25, 2020; Accepted: August 25, 2020

166 ว.มทรส. 8(2) : 165-175 (2563)
shaped bacteria (80.77%). The results of cellulase production test on carboxymethyl cellulose agar were found that only four isolates (15.38%) produced cellulase, which a halo zone around the colony larger than the zone produced by positive control bacteria, Paenibacillus curdlanolyticus strain B-6. The isolate PC-C-1 had the highest halo zone size (2.56±1.03 centimeters). Moreover, the rice straw degradation test of isolate PC-C-1 in nutrient broth added 5% rice straw at 1×108 cell/mL showed digestion of rice straw up to 64.80%. These results can be used as preliminary data for further studies of the practical application in rice straw degradation by bacteria, which is an agricultural waste reduction in the future. Keywords: bacteria, cellulase, extracellular enzyme, carboxymethyl cellulose agar (CMC agar)
บทนา
จงหวดพระนครศรอยธยามการเพาะปลกขาวเปนอาชพหลกของเกษตรกรท �ง 16 อาเภอ โดยมผลผลตขาวท!ออกสตลาดในแตละปปรมาณมาก ซ!งหลงการเกบเก!ยวผลผลตขาวมกมวสดเหลอท �งทางการเกษตร คอ ฟางขาว ท!มรายงานขอมลทางสถตท!มสงถง 26.90 ลานตนตอป (Suphanburi Land Development Station, 2014) ท �งน �เกษตรกรมกระบวนการจดการฟางขาวในหลากหลายวธ ไดแก การแปรรปฟางขาวเปนอาหารสตว หรอพลงงานทดแทน การไถกลบตอซงฟางขาว การทาป ยหมกจากฟางขาว รวมถงการเผาฟางขาวหลงการเกบเก!ยว (Doungwongsa, & Losirikul, 2010) ซ!งเปนวธการท!เกษตรกรไมนอยท!นยมเลอกใช เน!องจากเปนวธการท!ทาไดงาย ใชระยะเวลาส �น และประหยดคาใชจาย แตผลเสยท!ตามมาคอเกดฝ นละอองขนาดเลกจานวนมาก สงผลเสยตอสขภาพตอผสดดม และเกดเปนปญหามลพษทางส!งแวดลอมท!รนแรง ดงน �นการหาแนวทางในการแกไขจงมความสาคญและจาเปนอยางมาก ซ!งจลนทรยท!อาศยอยในดนเปนอกทางเลอกหน!งท!มการศกษาและมบทบาทสาคญสาหรบเลอกใชในการกาจดฟางขาว โดยเฉพาะอยางย!งจลนทรยในกลมของแบคทเรย ท!พบมความหลากหลายและมจานวนมากท!สดในดน
และส!งแวดลอม ท �งน �มงานวจยมากมายท!แสดงถงความสามารถของแบคทเรยในการยอยสลายวสดเหลอท �งทางการเกษตรไดชนดตางๆ เชน ชานออย ซงขาวโพด ข �เล �อย เศษกระดาษ รวมถงฟางขาว (Supasinsathit, 2012) ซ!งการยอยฟางขาวน �นเกดจากเอนไซมจาเพาะปลอยออกสนอกเซลล (extracellular enzyme) ท!แบคทเรยสรางข �น ทาการยอยสวนประกอบของฟางขาวใหเปนสารโมเลกลเลกและนาเขาสเซลล เพ!อใชเปนแหลงของสารอาหาร โดยเอนไซมท!สาคญคอเอนไซมเซลลเลส (cellulase) ทาหนา ท!ยอยเซลลโลส (cellulose) (Manowattana, & Leksawasdi, 2014) ท!เปนองคประกอบหลกในฟางขาว (รอยละ 40-60) (Malherbe, & Cloete, 2002) ตวอยางของแบคทเรยในระดบจนสท!มรายงานการศกษาสามารถสรางเอนไซมเซลลเลส ไดแก Acetivibrio,
Bacillus, Bacteriodes, Cellulomonas, Clostridium, Erwinia, Paenibacillus, Ruminococcus, Thermonospora
และ Actinomycetes (Poodngam, 2014; Ahmed, Babalola, & McKay, 2017; Thanasrirangkul, Pinjai, & Vaithanomsat, 2018) ดงน �นการศกษาในคร �งน � จงทาการคดแยกแบคทเ รยจากตวอยางดนท!มความสามารถในการยอยสลายเซลลโลสท! เ ปนองคประกอบหลกของฟางขาวท!มประสทธภาพสง

RMUTSB Acad. J. 8(2) : 165-175 (2020) 167
วธการศกษา
1. การเกบตวอยางดน เกบตวอยางดนนาในเขตจงหวดพระนครศรอยธยา
จานวน 5 อาเภอ ท!พบมการเพาะปลกขาวมากท!สด ไดแก บางบาล บางซาย วงนอย ภาช และทาเรอ โดยทาการเกบตวอยางดนแบบสมบรเวณรอบรากพชของตอซง ขาวสายพนธ เดยวกน กาหนดพ �น ท! ของความกวาง×ยาว×ลก คอ 10×10×10 เซนตเมตร นาดนตวอยางมาศกษาลกษณะทางกายภาพของชนดดน 3 ประเภท ไดแก ดนรวน ดนเหนยวและ ดนทราย รวมถงคาความเปนกรด-ดาง (pH) ของดน
2. การคดแยกเช Uอแบคทเรยท มความสามารถในการยอยสลายเซลลโลส
ตวอยางดนน �าหนก 10 กรม ถกนามาทาการเจอจางดวยวธ serial ten-fold dilution ดวยสารละลาย 0.85 เปอรเซนต NaCl กอนนามาเพาะเล �ยงแบคทเรยดวยวธการ spread plate ลงบนอาหาร carboxymethyl cellulose agar (CMC agar) ตวอยางละ 3 ซ �า บมท!อณหภม 37 องศาเซลเซยส นาน 24-48 ช!วโมง สงเกตและนบจานวนโคโลนท!มการเจรญ เพ!อคานวณปรมาณแบคทเรย (CFU/g) และทาการจดกลมแบคทเรยท!มความเหมอนและแตกตางกนของสณฐานวทยาโคโลนท!สงเกตได เชน ส ขอบ ผวหนาโคโลน การยกตว เปนตน จากน �นเลอกตวแทนโคโลนแตละกลมมาทาใหเช �อบรสทธ� พรอมท �งศกษาลกษณะทางสณฐานวทยา การตดสแบบแกรม รปรางของเซลล (Liang, Zhang, Wu, Wu, & Feng, 2014)
3. การตรวจสอบความสามารถในการสรางเอนไซมเซลลเลส
แบคทเรยไอโซเลทท!คดแยกไดนาไปเพาะเล �ยงดวยวธการ point inoculation technique บนอาหาร
CMC agar ท!เตม 0.02 เปอรเซนต Congo red บมท!อณหภม 37 องศาเซลเซยสนาน 24-48 ช!วโมง สงเกตการสรางเอนไซมจากขนาดของวงใสท!เกดข �นเปรยบเทยบกบเช �อควบคมท!พบมการสรางเอนไซมเซลลเลส คอ Paenibacillus curdlanolyticus B-6 (positive control) (Pason, Kyu, & Ratanakhanokchai, 2006) 4. การทดสอบการยอยสลายฟางขาวของแบคทเรยไอโซเลทท มความสามารถสรางเอนไซมเซลลเลสสงท สด
นาฟางขาวท!ผ านการไปปรบสภาพดวยสารละลายโซเดยมไฮดรอกไซด (NaOH) ความเขมขนรอยละ 10 (ปรมาตรตอปรมาตร) เปนเวลา 48 ช!วโมง ลางดวยน �าจนคาความเปนกรด-ดาง มคาเปนกลาง (pH 7) และอบแหงท!อณหภม 60 องศาเซลเซยส ใสผสมลงในอาหารเหลว nutrient broth บมบนเคร!องเขยาความเรวรอบ 150 rpm อณหภม 37 องศาเซลเซยสเปนเวลา 7 วน (ดดแปลงจาก Tachaapaikoon et al., 2012) ทาการทดลอง 3 ซ �า โดยเปรยบเทยบปรมาณของฟางขาวท!ลดลงจากการวดน �าหนกเซลลแหงดวยการอบท!อณหภม 105 องศาเซลเซยส เปนเวลา 24 ช!วโมง
ผลการศกษาและอภปรายผล 1. คณสมบตทางกายภาพของดนตวอยาง
การ เ กบตวอยา งดนจากอา เภอท!ม การเพาะปลกขาวจานวน 5 อาเภอ (Table 1) ทาการเกบตวอยางดนจากบรเวณรอบรากตอซง ขาวจาวปทมธานสายพนธ RD31, RD41, RD57 และ RD61 โดยลกษณะทางกายภาพของดนตวอยางท �งหมดเปนชนดดนเหนยว และมคาความเปนกรด-ดางของดน (pH) อยในชวง 5.16-7.10 (Table 1) จะเหนไดวาจากคณสมบตทางกายภาพของดนตวอยางดงกลาว

168 ว.มทรส. 8(2) : 165-175 (2563)
สงผลใหน �าสามารถซมผานไดคอนขางชา การชะลางของสารอนทรยนอย จงทาใหมอนทรยวตถในดนคอนขางสง โดยจากการศกษากอนหนาน �พบวาคา
pH ของดนเหนยวอยในชวง 4.5-7.0 จลนทรยสามารถเจรญได (Hamarashid, Othman, & Hussain, 2010; Töysä, & Hänninen, 2017)
Table 1 Soil sample characteristics. soil samples soil physical characteristics
sampling location (sub-district/ district) rice varietie texture pH 1. Phra Khao/ Bang Ban (BB) Pathum Thani RD41 clay 5.49 2. Bang Sai/ Bang Sai (BC) Pathum Thani RD57 clay 5.16 3. Kra Chio/ Pha Chi (PC) Pathum Thani RD61 clay 5.45 4. Pak Tha/ Tha Ruea (TR) Pathum Thani RD57 clay 6.43 5. Lam Ta Sao/Wang Noi (WN) Pathum Thani RD31 clay 7.10
2. แบคทเรยไอโซเลทท มความสามารถในการยอยสลายเซลลโลส
จากการศกษานาตวอยางดนรอบตอซงขาว ท �ง 5 อาเภอ มาทาการคดแยกและเพาะเล �ยงแบคทเรย ดวยวธการ ten-fold dilution spread plate method บนอาหารแขงจาเพาะ CMC agar ซ!งมสวนผสมของเซลลโลสในรปของ carboxymethyl cellulose ซ!งจะไดขอมลการเจรญของจานวนโคโลนแบคทเรยและจานวนแบคทเรยไอโซเลทท!แยกไดจากแตละตวอยางดน (Table 2) โดยขอมลการเจรญของโคโลนแบคทเรยบนจานอาหารอยในชวง 1.06-6.87×106 CFU/g (Table 2) ซ!งจานวนของแบคทเรยท!พบมการเจรญบนจานอาหารเล �ยงเช �อซงท!แยกไดจากตวอยางดนนาขาวในแตละอาเภอน �นมคาใกลเคยงกน ท �งน �ปรมาณของโคโลนแบคทเรยท!แยกไดจากตวอยางดนในอาเภอทาเรอมปรมาณสงท!สดคอ 7.37×106 CFU/g และพบจานวนนอยท!สดในดนตวอยางจากอาเภอบางบาลท!มจานวนโคโลนเพยง 1.06×106 CFU/g แตเม!อทาการ
จดกลมของแบคทเรยตามลกษณะความเหมอน และความแตกตางของสณฐานของโคโลนท!ปรากฏ การเจรญบนจานอาหาร สามารถจดกลมและคดเลอกโคโลนท!เปนตวแทนแตละกลมไดจานวนรวม 26 ไอโซเลท (Table 2) ท �งน �พจารณาความหลากหลายของแบคทเรยไอโซเลทท!แยกไดจากตวอยางดนจากแตละอาเภอมความผกผนกบจานวนโคโลนท �งหมดท!นบได กลาวคอ แบคทเรยไอโซเลทท!แยกไดมากท!สดไดจากตวอยางดนอาเภอบางซาย จานวน 7 ไอโซเลท รองลงมาเปนอาเภอบางบาล (6 ไอโซเลท) อาเภอภาช (5 ไอโซเลท) อาเภอทาเรอและวงนอยไดแบคทเรยไดจานวนไอโซเลทท!เทากนคอ 4 ไอโซเลท ซ! งสอดคลองกบการวจยกอนหนาน �ของ Lu, & Mosier (2007) ท!กลาวถงชวงของคาความเปนกรด-ดางท!เหมาะสมในการสรางเอนไซมและการเกดกจกรรมการยอยของเอนไซมเซลลเลสบนอาหารท!มองคประกอบของเซลลโลส (CMC agar) ของแบคทเรยอยในชวง pH 4-8 (สอดคลองกบขอมลการศกษา

RMUTSB Acad. J. 8(2) : 165-175 (2020) 169
ท!ไดใน (Table 1) ท �งน �จะเปนการทางานของเอนไซมชนดท!สรางและปลอยออกสนอกเซลล ท!พบไดหลากหลายจากส!งแวดลอมตางๆ ท!มอนทรยวตถขององคประกอบเซลลโลส ในดนท!วๆ ไป (Thanonkeo,
1995) และ ตอ งมกา รระ บาย อากา ศขอ งดน ท!เหมาะสมกสงผลตอการเจรญของแบคทเรยท!มกจกรรมในการสรางเอนไซมดงกลาวได (Mawadza, Rahni, & Bo, 2000)
Table 2 Colony counting and isolates from soil samples. soil samples colony counting (CFU/g) isolates 1. Phra Khao/ Bang Ban (BB) 1.06×106 6 2. Bang Sai/ Bang Sai (BC) 1.95×106 7 3. Kra Chio/ Pha Chi (PC) 1.40×106 5 4. Pak Tha/ Tha Ruea (TR) 7.37×106 4 5. Lam Ta Sao/Wang Noi (WN) 6.87×106 4
เม!อศกษาการตดสยอมแบบแกรม (Gram’s
stain) และสงเกตรปรางของเซลลจากแบคทเรย ไอโซเลทท!แยกไดน �น (Table 1) ผลท!ไดสามารถแบงแบคทเรยได 3 กลมตามความแตกตางของลกษณะสณฐานวทยาของเซลล (Table 3) คอ (1) แบคทเรยแกรมบวก รปรางแทง (rod) จานวน 21 ไอโซเลท (คดเปนรอยละ 80.77), (2) แบคทเ รยแกรมลบ รปรางแทง จานวน 4 ไอโซเลท (คดเปนรอยละ 15.38) และ (3) แบคทเรยแกรมลบ รปรางกลม จานวน 1 ไอโซเลท (คดเปนรอยละ 3.85) ซ!งไอโซเลทท!แยกไดท �งหมดมลกษณะทางสณฐานวทยาของโคโลนท!แตกตางกน (Figure 1) ท �งน �ผลการศกษาท!ไดมความสอดคลองกบงานวจยตางๆ ท!สามารถพบแบคทเรยท!มความสามารถในการยอยสลายเซลลโลสดวยการสรางเอนไซมเซลลเลสออกมายอยท!พบไดท �งในแบคทเรยแกรมบวกและแกรมลบ และมลกษณะรปรางโคโลนท!แตกตางกน (Mawadza, Rahni, &
Bo, 2000; Chakdar et al., 2016; Prabhu, Rao, & Li, 2017; Thanasrirangkul, Pinjai, & Vaithanomsat, 2018) ท �งน �สวนใหญมกพบเปนแบคทเรยแกรมบวกมากกวาแบคทเรยแกรมลบ เน!องจากมผนงเซลลท!หนาและแขงแรงทาใหทน ตอสภาวะแวดลอมตางๆ ไดดกวาและมการปรบตวสรางสารบางชนดปลอยออกมาเพ!อยอยสารอาหารเ ข า ส ภา ย ใ น เ ซ ล ล อ ก ด ว ย ท! ร จ ก กน ด ไ ด แ ก Cellulomonas (Yin, Huang, & Lin, 2010; Prabhu, Rao, & Li, 2017) โดยเฉพาะอยางย!งแบคทเรย แกรมบวกท!สรางเอนโดสปอรท!ทาใหทนตอสภาวะ ไมเหมาะสมไดดและสามารถงอกเปนเซลลใหม มการเจรญและเพ!มจานวนไดดกวา เชน Bacillus และ Clostridium เปนตน (He, Ding, & Long, 1991; Warnick, Methe, & Leschine, 2002; Balasubramanian, Toubarro, Teixeira, & Simos, 2012; Prabhu, Rao, & Li, 2017)

170 ว.มทรส. 8(2) : 165-175 (2563)
Table 3 Morphological characteristics of bacteria isolated from soil samples. bacteria isolates cell shape Gram's stain BB-C-1, BB-C-2, BB- C-3, BB-C-4, BB-C-5, BB-C-6, BS-C-1, BS-C-2, BS-C-3, BS-C-4, BS-C-5, BS-C-6, BS-C-7, PC-C-1, PC-C-3, PC-C-4, PC-C-5 ,TR-C-2, TR-C-3, TR-C-4, WN-C-1
rod positive (+)
PC-C-2, TR-C-�,WN-C-2, WN-C-4 rod negative (-) WN-C-3 coccus negative (-)
Figure 1 Morphological characteristics of bacteria isolated PC-C-�; (A) morphological of colony, (B) Gram’s stain and cell shape.
3. การสรางเอนไซมเซลลเลสของแบคทเรย ไอโซเลทท แยกได
เม!อตรวจสอบความสามารถในการสรางเอนไซมเซลลเลสของแบคทเรยไอโซเลท ดวยการเพาะเล �ยงบนอาหาร CMC agar ท!เตม 0.02 เปอรเซนต Congo red พบวามการสรางเอนไซมเซลลเลสจากการปรากฏวงใสรอบโคโลน (Figure 2) เพยง 4 ไอโซเลท (คดเปนรอยละ 15.38) ท!สามารสรางเอนไซมเซลลเลสออกมายอยเซลลโลสบนอาหาร CMC agar (Table 4) ซ!งเม!อเปรยบเทยบปรมาณการสรางจากขนาดของวงใสกบเช �อควบคมท!พบมการรายงานวามสรางเอนไซม เซลลเลส คอ P. curdlanolyticus B-6 (positive control) (Pason, Kyu, & Ratanakhanokchai, 2006) น �น ยงพบปรมาณการสรางเอนไซมท!มากกวา (ขนาดของวงใส
รอบๆ โคโลนท!มากกวา) โดยแบคทเรยไอโซเลทท!ใหผลการสรางเอนไซมสงท!สด คอ PC-C-1 (ขนาด ของวงใส 2.56±1.03 เซนตเมตร) ท!แยกไดจากดนตวอยางในอาเภอภาช (Table 4) ซ!งผลท!ไดคดเปนรอยละ 3.84 ของแบคทเรยไอโซเลทท �งหมด
แบคทเรยไอโซเลทตอความสามารถในการสรางเอนไซมเซลลเลสท!ไดในการศกษาคร �งน �พบจานวนนอยน �น มผลท!ใกลเคยงกนเปรยบเทยบกบงานวจยของ Ahmed, Babalola, & McKay (2017) ท!รายงานผลการคดแยกแบคทเรยจากดนรอบรากพชในเมอง Mafikeng ทางตะวนออกเฉยงเหนอของประเทศแอฟรกาใต ไดจานวนแบคทเรยไอโซเลทจานวน 20 ไอโซเลท แตท �งน �มเพยง 10 ไอโซเลท ท!สามารถสรางเอนไซมเซลลเลสและไซลาเนสได ท �งน �ตวอยางดนท!นามาศกษามความแตกตางกนท �งทางดานลกษณะชนดของดนและคาความเปนกรด-ดางของดนตวอยาง รปแบบของการเพาะปลกขาวบนพ �นท!ดน ประวตของการใชป ย/สารเคมพ �นท! เ กบตวอยางดน และสภาพแวดลอมบรเวณท!ทาการเกบตวอยาง ลวนมผลตอจานวนและความหลากหลายของชนดแบคทเรยและปรมาณในการสรางเอนไซมเซลลเลสของแบคทเรยท!คดแยกไดใหผลแตกตางกน (Poodngam, 2014; Ahmed, Babalola, & McKay,
A B

RMUTSB Acad. J. 8(2) : 165-175 (2020) 171
2017) รวมถงพบวาผลการศกษาในคร �งน �มความแตกตางกบงานวจยของ Thanasrirangkul, Pinjai, & Vaithanomsat (2018) ท!รายงานผลการคดแยกแบคทเรยจากตวอยางตางชนดกน ไดแก ดนรอบรากชานออย ชานออย ทะลายปาลม และกอนเหด โดยแยกแบคทเรยไอโซเลทสงถง 100 ไอโซเลท และพบจานวน 73 ไอโซเลท (คดเปนรอยละ 73) ท!สามารถสรางเอนไซมเซลลเลสได ท �งน �ตวอยางท!นามาใชในการคดแยกแบคทเรยท!หลากหลายมผลตอจานวนของการคดแยกแบคทเรย ไอโซเลทและความสามารถในการสรางเอนไซมเซลลโลส รวมถงองคประกอบเซลลลโลสในตวอยางท!นามาศกษา และส!งแวดลอมในการเจ รญของแบคทเ รยยงมผลตอปรมาณ การสรางเอนไซมเซลลเลสอกดวย (Poodngam, 2014; Ahmed, Babalola, & McKay, 2017; Thanasrirangkul, Pinjai, & Vaithanomsat, 2018) แตท �งน �จากงานวจยของ Thanasrirangkul, Pinjai, & Vaithanomsat (2018) สามารถบงช �ถงการศกษาการสรางเอนไซมเซลลเลสดวยวธ Congo red (ในอาหาร CMC agar) เปนวธท!สามารถคดแยกแบคทเรยท!มความสามารถในการสรางเอนไซมเซลลเลสและ ทราบปรมาณการสรางเบ �องตน เน!องจากสามารถมองเหนวงไสบนอาหารเล �ยงเช �อ อนเน!องมาจาก การทาปฏกรยาของส Congo red ท!ผสมลงในอาหารโดยบรเวณท!มการผลตเอนไซมเซลลเลสออกมาน �น สาร CMC ท!ผสมอยในอาหารจะถกยอยสงผลใหเกดเปนโซนใสดงกลาว (Johnsen, & Krause, 2014) และเปนว ธการท! เหมาะสมในการศกษาความ สามารถเบ �องตนของแบคทเรยตอการสรางเอนไซมเซลลเลส
Table 4 Cellulase activity of bacteria isolates.
isolates clear zone (centimeters) ± standard deviation (S.D.)
control 2.10±0.14 PC-C-1 2.56±1.03
PC-C-5 2.22±0.95 TR-C-1 2.13±0.96 TR-C-2 2.45±1.51
Figure 2 Cellulase test on CMC agar and the halo zone around the bacteria colony.
จากผลการทดลองท!ไดช �ใหเหนวาแบคทเรยท!อาศยอยในดนท!มการเพาะปลกขาวมความสามารในการสรางเอนไซมชนดปลอยออกสนอกเซลล ออกมายอยสลายเซลลโลสท!เปนองคประกอบของฟางขาว ใหเปนสารประกอบอนทรยท!มโมเลกลเลกลงและละลายน �าไดและสามารถผานเขาเซลลได และนาไปใชเปนแหลงสารอาหารท!สาคญ (carbon source) ตอการเจรญเพ!มจานวนของแบคทเรยท!อาศยอยในดน โดยเฉพาะ อยางย!งสวนของตอซงขาวและฟางขาวท!อยใตพ �นดนท!มการสมผสกบแบคทเรยท!อาศยอยในดนโดยตรง แบคทเรยหลายกลมมความสามารถในการยอยสลายเซลลโลสได โดยกลไกการยอยเซลลโลสจะยอย

172 ว.มทรส. 8(2) : 165-175 (2563)
โครงสรางเซลลโลสท!เรยงตอกน (crystalline cellulose) ใหเปนสายส �นลงในรปเซลโลไบโอส (cellobiose) โดยอาศยเอนไซมเอนโดกลคาเนส (endoglucanase) และเอกโซกลคาเนส (exoglucanase) และไดผลผลตสดทายคอน �าตาลกลโคส (glucose) ซ!งยอยโดยเอนไซมเบตากลโคซ เดส (β-glucosidase) (Liang, Zhang, Wu, Wu, & Feng, 2014; Nafi, Usman, Abdullahi, Mustapha, & Maiturare, 2017) ซ!งสยอมCongo red จะสรางพนธะไฮโดรเจนอยางนอย 2 ตาแหนง โดยท!ระยะหางของหมท!สามารถสรางพนธะไฮโดรเจนซ!งระยะหางน �นจะใกลเคยงกบระยะหางของหม –OH ในเซลลโลส โมเลกลของสจะหนหมซลโฟเนต (sulfonate) ท!มประจลบในขณะท!ผวของเสนใยเซลลโลสกมประจลบเชนกนหนออกดานตรงขาม เน!องจากแรงผลกทางไฟฟา สงผลทาใหบรเวณท!มองคประกอบของเซลลโลสจะทาปฏกรยากบสยอมมองเหนเปนสแดง และสวนท!ไมมองคประกอบของเซลลโลสจะไมถกยอมดวยสมองเหนเปนบรเวณใส จงเปนวธการเหมาะสมในการศกษาความสามารถเบ �องตนของแบคทเรยตอการสรางเอนไซมเซลลเลสดงกลาว
4. ความสามารถยอยสลายฟางขาวของแบคทเรยไอโซเลท PC-C-1
ผลการทดสอบการยอยสลายฟางขาวของแบคทเรยไอโซเลทท!สรางเอนไซมเซลลโลสสงท!สดคอ PC-C-1 เม!อเพาะเล �ยงในอาหาร nutrient broth ท!มสวนผสมของฟางขาวครบ 7 วน ทาการเกบตวอยางฟางขาวดวยการกรองและนาไปอบแหงกอนทาการช!งหาน �าหนกแหงของฟางขาวท!เหลอในอาหารเล �ยงเช �อเปรยบเทยบกบชดควบคม (ไมมการเตมแบคทเรย) พบวา ไอโซเลท PC-C-1 มรอยละของการยอยฟางขาวท!
64.80 อาจกลาวไดวาแบคทเรย ไอโซเลท PC-C-1 มประสทธภาพในการยอยฟางขาวมากกวาชดควบคม (ท!มรอยละของการยอยสลายฟางขาวเทากบ 0) สาหรบยอยโครงสรางของผนงเซลลขาวสวนท!เปนเซลลโลส ซ!งสอดคลองกบงานวจยกอนหนาน �ท!พบวาแบคทเรยท!สรางเอนไซมเซลลเลสมการยอยสลายสารอนทรยท!เปนสวนประกอบของผนงเซลลพชสวนใหญ เพ!อนาไปใชเปนสารต �งตนในกระบวนการเมแทบอลซมและสรางพลงงานภายในเซลล (Gilbert, & Hazlewood, 1993; Subramaniyan, & Prema, 2000; Ahmed, Babalola, & McKay, 2017; Prabhu, Rao, & Li, 2017) แตท �งน �ในการนาไปประยกตใชจรงในการยอยฟางขาวในแปลงนาน �น ยงไมสามารถทาได เน!องจากในธรรมชาตของฟางขาวยงมสวนประกอบอ!นนอกเหนอจากเซลลโลส ไดแก เฮมเซลลโลส และลกนน (Malherbe, & Cloete, 2002) จงตองมการศกษาวจยเพ!มเตมของแบคทเรยไอโซเลท PC-C-1 ถงคณสมบตการสรางเอนไซมไซลาเนสท!สามารถยอยสลายไซเลน (ชนดสวนใหญของเฮมเซลลโลส) และลกนนเนสท!สามารถยอยสลายลกนนได (Ahmed, Babalola, & McKay, 2017; Thanasrirangkul, Pinjai, & Vaithanomsat, 2018) รวมถงการศกษาสภาวะท!เหมาะสมในการยอยสลายฟางขาวของแบคทเรยไอโซเลทท!มประสทธภาพในการยอยฟางขาวไดสงสดและการเกบขอมลเชงสถตของประสทธภาพการยอยสลายฟางขาวในระดบหองทดลอง และศกษาเกบขอมลผลการยอยฟางขาวในแปลงนาจรงเปนกระบวนการสดทาย จงจะสามารถประยกตใ ชประโยชนจากจลนทรยในกลมของแบคทเรยในการยอยสลายฟางขาวเพ!อลดปญหาการกาจดฟางขาวเหลอท �งทางการเกษตรใหแกเกษตรได

RMUTSB Acad. J. 8(2) : 165-175 (2020) 173
สรป
การเกบตวอยางดนรอบตอซงขาวสายพนธปทมธาน พ �นท! 5 อาเภอ ไดแก บางบาล บางซาย วงนอย ภาช และทาเรอ ในเขตจงหวดพระนครศรอยธยา ตวอยางดนท �งหมดเปนชนดดนเหนยว มคาความเปนกรด-ดางของดนท!เปนกรดเลกนอยจนถงคาเปนกลาง (pH 5.16-7.10) ซ!งเปนคาท!เหมาะสมตอการเจรญของจลนทรยในดน ซ!งพบการเจรญของโคโลนแบคทเรยท �งหมดท!สามารถยอยสลายเซลลโลสได อยในชวง 1.06-6.87×106 CFU/g และคดแยกแบคทเรยไดท �งหมด 26 ไอโซเลท โดยจาแนกจานวนพ �นท!เกบตวอยาง ดนรอบ คอ บางบาล บางซาย วงนอย ภาช และทาเรอ ไดแบคเทเรยไอโซเลทเทากบ 6, 7, 5, 4 และ 4 ไอโซเลท ตามลาดบ ท �งน �ผลการศกษาลกษณะสณฐานวทยาของเซลลและโคโลนของแบคทเรยไอโซเลท พบวาสวนใหญเปนแบคทเรยแกรมบวก รปรางแทง (รอยละ 80.77) และมลกษณะสณฐานวทยาของโคโลนท!แตกตางกน อกท �งผลการตรวจสอบการสรางเอนไซมเซลลเลสของแบคทเรยไอโซเลทท!แยกได พบเพยง 4 ไอโซเลท (คดเปนรอยละ 15.38) ท!มการสรางเอนไซมเซลลเลสเกดวงใสรอบโคโลนขนาดมากกวาแบคทเรยควบคม P. curdlanolyticus B-6 โดยไอโซเลทท!พบสรางเอนไซมเซลลเลสเกดวงใสรอบโคโลนสงท!สด คอ PC-C-1 (2.56±1.03 เซนตเมตร) จงนาไปทดสอบการยอยฟางขาวในอาหารเหลว nutrient broth ท!มการเตมฟางขาวลงไป และใหผลการยอยฟางขาวไดสงถงรอยละ 64.80 ในระยะเวลาการเพาะเล �ยง 7 วน ท �งน �ตองมการศกษาเพ!มเตมครอบคลมการจดจาแนกชนดของแบคทเรยดวยการใชเทคนคทางชววทยาโมเลกลเพ!มเตมสาหรบการบงช �ชนดของแบคทเรย
ไอโซเลท PC-C-1 ในระดบจนส และการหาสภาวะท!เหมาะตอการสรางและทางานของเอนไซมเซลลเลส ในการยอยฟางขาว ไดแก อณหภม คาความเปน กรด-ดาง สภาวะการทางานแบบมอากาศและไมมอากาศ เพ!อการนาไปพฒนาสาหรบใชประโยชนในการยอยสลายฟางขาวเหลอท �งทางการเกษตรใหแกเกษตรกรตอไปในอนาคต
คาขอบคณ
ขอขอบคณมหาวทยาลยเทคโนโลยราชมงคลสวรรณภมท!สนบสนนงบประมาณในการวจย ภายใตงบกองทนสงเสรมงานวจย ประจาปงบประมาณ 2561 และขอขอบคณสาขาวชาวทยาศาสตร (ชววทยา) คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ท!อนเคราะหสถานท!และสนบสนนดานวสด อปกรณ และเคร!องมอทางดานวทยาศาสตร สาหรบการศกษาวจยในคร �งน �
เอกสารอางอง Ahmed, A. A. Q., Babalola, O. O., & McKay, T. (2017).
Cellulase and xylanase producing bacterial isolates with the ability to saccharify wheat straw and their potential use in the production of pharmaceuticals and chemicals from lignocellulosic material. Waste and Biomass
Valorization, 9(5), 765-775. Balasubramanian, N., Toubarro, D., Teixeira, M., &
Simos, N. (2012). Purification and biochemical characterization of a novel thermos-stable carboxymethyl cellulose from Bacillus mycoides S122C. Applied Biochemistry and Biotechnology,
168(8), 2191-2204.

174 ว.มทรส. 8(2) : 165-175 (2563)
Chakdar, H., Kumar, M., Pandiyan, K., Singh, A., Nanjappan, K., Kashyap, P. L., & Srivastava, A. K. (2016). Bacterial xylanases: biology to biotechnology. 3 Biotech, 6(2),150.
Doungwongsa, I., & Losirikul, M. (2010). The rice straw
management for N, P and K element conservation
in the paddy field, Laos (Master’s thesis). Ubon Ratchathani University, Ubon Ratchathani. (in Thai)
Gilbert, H. J., & Hazlewood, P. (1993). Bacterial cellulases and xylanases. Journal of Microbiology, 139(1), 187-194.
Hamarashid, N, H., Othman, M. A., & Hussain, M. A. H. (2010). Effects of soil texture on chemical composition, microbial populations and carbon mineralization in soil. The Egyptian Society of
Experimental Biology, 6(1), 59-64.
He, Y. L., Ding, Y. F., & Long, Y. Q. (1991). Two cellulolytic Clostridium species: Clostridium cellulosi sp. nov. and Clostridium cellulofermentans sp. nov. International Journal of Systematic Bacteriology,
14(2), 306-309.
Johnsen, H. R., & Krause, K. (2014). Cellulase activity screening using pure carboxymethylcellulose: application to soluble cellulolytic samples and to plant tissue prints. International Journal of
Molecular Sciences, 15(1), 830-838. Liang, Y. L., Zhang, Z., Wu, M., Wu, Y., & Feng, J. X.
(2014). Isolation, screening, and identification of cellulolytic bacteria from natural reserves in the subtropical region of China and optimization of cellulase production by Paenibacillus terrae ME27-1. Journal of Biomedicine and Biotechnology,
2014(5), 1-13.
Lu, Y., & Mosier, N. S. (2007). Biomimetic catalysis for hemicellulose hydrolysis in corn stover. Biotechnology Progress, 23(1), 116-123.
Malherbe, S., & Cloete, T. E. (2002). Lignocellulose biodegradation: fundamentals and applications. Environmental Science and Bio/Technology, 1, 105-114.
Manowattana, A., & Leksawasdi, N. (2014). Kinetic modeling
enzyme xylanase (Master’s thesis). Chiang Mai University, Chiang Mai. (in Thai)
Mawadza, C. H., Rahni, R. Z., & Bo, M. (2000). Purification and characterization of cellulases produced by two Bacillus strains. Journal of Biotechnology, 83(3), 177-187.
Nafi, U. A., Usman, M. H., Abdullahi, B. Y., Mustapha, G., & Maiturare, H. M. (2017). Screening and identification of cellulase-producing bacteria isolated from rumen of camel in Sokoto main abattoir. Scientific
and Management, 5(7), 5826-5832.
Pason, P., Kyu, K. L., & Ratanakhanokchai, K. (2006). Paenibacillus curdanolyticus strain B-6 xylanolytic-cellulolytic enzyme system that degrades insoluble polysaccharides. Applied and
Environmental Microbiology, 72(4), 2483-2490. Prabhu, M., Rao, N., & Li, W. J. (2017). Chapter 4:
Microbial cellulose and xylanase: their sources and application. Advances in Biochemistry &
Applications in Medicine, 2017, 1-20. Poodngam, J. (2014). Isolation and selection of
Actinomycetes capability to degrade lignocelluloses compounds on soil at the Namtok Yong National Park. RMUTSB
Academic Journal, 2(2), 109-120. (in Thai)

RMUTSB Acad. J. 8(2) : 165-175 (2020) 175
Subramaniyan, S., & Prema, P. 2000. Cellulase-free xylanase from Bacillus and other microorganism. FEMS
Microbiology Letter, 183(1), 1-7. Supasinsathit, A. (2012). Energy from biomass with high
lignocellulose. Environmental Journal, 16(2), 36-43. (in Thai)
Suphanburi Land Development Station. (2014). The
incorporation of rice straw and crop residues
after harvesting. Retrieved 15 November 2019, from http://r01.ldd.go.th/spb/. (in Thai)
Tachaapaikoon, C., Kosugi, A., Pason, P., Waeonukul, R., Ratanakhanokchai, K, Kyu, K. L., Arai, T., Murata, Y., & Mori, Y. (2012). Isolation and characterization of a new cellulosome-producing Clostridium
thermocellum strain. Biodegradation, 23(1), 57-68. Thanasrirangkul, C., Pinjai, P., & Vaithanomsat, P. (2018).
Screening of cellulase producing bacteria and efficiency of lignocellulosic decomposition. King
Mongkut’s Agricultural Journal, 36(3), 1-12. (in Thai) Thanonkeo, P. (1995). Optimal conditions of cellulase
production from fungi isolated from Agave
sisalana Perrine plantations (Master’s thesis). Chaulalongkorn University, Bangkok. (in Thai)
Töysä, T., & Hänninen, O. (2017). Soil pH, Ca and Mg stability and pH association with temperature a groundwater silicon. Journal of Scientific &
Technical Research, 1(7), 1851-1853. Warnick, T. A., Methe, B. A., & Leschine, S. B. (2002).
Clostridium phytofermentans sp. nov., a cellulolytic mesophile from forest soil. International Journal
of Systematic and Evolutionary Microbiology,
52(4), 1155-1160. Yin, L. J., Huang, P. S., & Lin, H. H. (2010). Isolation of
cellulose-producing bacteria and characterization of the cellulose from the isolated bacterium
Cellulomonas sp. YJ5. Journal of Agricultural &
Food Chemistry, 58(17), 9833-9837.

RMUTSB Acad. J. 8(1) : 39-56 (2020) 39
การคดเลอกและการหาสภาวะท�เหมาะสมในการกาจดไนเตรทและฟอสเฟต ดวยสาหรายสเขยวท�คดแยกจากน !าเสยในโรงงานอตสาหกรรมผลตอาหารสาเรจรป Isolation and optimization of nitrate and phosphate removal by green algae
isolated from wastewater of food industrial
สามารถ ตายขาว1* Samart Taikhao1*
บทคดยอ
สาหรายสเขยวมความสามารถในการกาจดไนเตรทและฟอสเฟตออกจากน าเสย งานวจยน มวตถประสงคเพ(อคดแยกสาหรายสเขยวจากน าเสยจากโรงงานอตสาหกรรมผลตอาหารสาเรจรปในจงหวดพระนครศรอยธยา ท(มความสามารถสงในการกาจดไนเตรท และฟอสเฟตในน าเสย จากการทดลองพบวาสามารถคดแยกสาหรายสเขยวไดท งหมด 21 ไอโซเลท จากน นนาสาหรายสเขยวทกไอโซเลทมาเพาะเล ยงในน าเสยท(ผานการฆาเช อแลว (พเอช 7) ท(อณหภม 25 องศาเซลเซยส ความเขมแสง 30 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน และเพาะเล ยงในสภาวะเขยาท(ความเรวรอบ 120 รอบตอนาท เปนเวลา 5 วน กอนท(จะวดคณลกษณะตางๆ ทางเคมของน าเสยหลงการเพาะเล ยง จากการทดลองพบวาสาหรายสเขยวไอโซเลท FPI 18 มความสามารถมากท(สดในการกาจดไนเตรทและฟอสเฟต และยงมความสามารถสงสดในการกาจดคาบโอด และซโอด เม(อเปรยบเทยบกบสาหรายสเขยวไอโซเลทอ(นๆ จากการจาแนกสายพนธสาหรายสเขยวไอโซเลท FPI 18 โดยการศกษาลาดบ นวคลโอไทดบรเวณ 18S rDNA และศกษาแผนภมวฒนาการ พบวาสาหรายสเขยวไอโซเลท FPI 18 มความคลายคลงและใกลชดกบสาหรายสเขยวในจนส Tetraspora จงต งช(อสาหรายชนดน วา Tetraspora sp. Rmutsb FPI 18 การศกษาสณฐานวทยาภายใตกลองจลทรรศนพบวา Tetraspora sp. Rmutsb FPI 18 มรปรางกลม ขนาดเสนผานศนยกลาง 5-6 ไมโครเมตร นอกจากน เซลลมลกษณะเปนเซลลเดยว หรออยรวมกนเปนกลม 3-4 เซลลภายใตชทหมท(โปรงแสง ย(งไปกวาน นยงมการตรวจสอบพารามเตอรบางประการท(สงผลในการกาจดไนเตรทและฟอสเฟตออกจากน าเสยโดยสาหรายสเขยวสายพนธน เชน คาพเอช อณหภม และความเขมแสง จากการศกษาพบวา สภาวะท(ดท(สดในการกาจดไนเตรทและฟอสเฟต คอ น าเสยมคาพเอชเร(มตน 8 อณหภมในการเพาะเล ยง 30 องศาเซลเซยส และความเขมแสง 50 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน นอกจากน การลดลงของไนเตรทและฟอสเฟต ในสภาวะดงกลาวยงสอดคลองกบผลของชวมวลของ Tetraspora sp. Rmutsb FPI 18 ท(สงข นอกดวย เม(อนาสภาวะท(ดท(สดทกสภาวะ มารวมกนต งช(อวา “สภาวะท(เหมาะสม” โดยนาสภาวะท(เหมาะสมมาทดลองเพาะเล ยง Tetraspora sp. Rmutsb FPI 18 เทยบกบ สภาวะเพาะปกต (น าเสยพเอชเร(มตน 8 อณหภม 25 องศาเซลเซยส และความเขมแสง 30 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน) ผลการทดลองพบวา สาหรายสเขยว Tetraspora
1 คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ศนยนนทบร 1 Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi, Nonthaburi Campus * Corresponding author. E-mail: [email protected] Received: November 1, 2019; Revised: December 6, 2019; Accepted: January 14, 2020

40 ว.มทรส. 8(1) : 39-56 (2563)
sp. Rmutsb FPI 18 มประสทธภาพสงในการกาจดไนเตรทและฟอสเฟตออกจากน าเสยไดถงรอยละ 93.65 และ 96.65 ตามลาดบ เม(อเพาะเล ยงเซลลในสภาวะท(เหมาะสม คาสาคญ: สาหรายสเขยว การบาบดน าเสย ไนเตรท ฟอสเฟต น าเสยโรงงานอตสาหกรรมผลตอาหารสาเรจรป
Abstract Green algae are able to remove inorganic nitrogen and phosporus from wastewater. This work aimed to
isolate green algae from food processing industrial wastewater in Phra Nakhon Si Ayutthaya province that has highly efficient removal of nitrate and phosphate from wastewater. It was found that twenty-one green algae strains were isolated from wastewater samples. All green algal isolates were grown in sterile wastewater (pH 7) at 30 ºC under light intensity of 30 µmolphoton/m2/s for 18 h per day and shaking at 120 rpm for 5 days before measuring chemical characteristics of wastewater after cultivation. The results revealed that isolate FPI 18 showed the highest nitrate and phosphate removal and also showed the lowest BOD and COD when compared with other green algae isolates. The genetics characterization of green algal isolate FPI 18 was classified by using 18S rDNA squence and phylogenetics tree analysis. The results indicated that green algae isolate FPI 18 showed their similarities comparing to the green alge in the genus of Tetraspora. Thereby, this strain was named Tetraspora sp. Rmutsb FPI 18. In morphological characterization under light microscope, Tetraspora sp. Rmutsb FPI 18 has an oval shape and 5-6 µm of diameter. In addition, single cells and 3-4 cells surrounded by transparent sheath can be observed. Furthermore, some parameters for nitrate and phosphate removal by this strain such as pH, temperature and light intensity were also examined. The results showed that the best condition for nitrate and phosphate removal from wastewater by Tetraspora sp. Rmutsb FPI 18 were initial pH 8, temperature at 30 ºC and light intensity of 50 µmolphoton/m2/s for 18 h per day. In addition, the reduction of nitrate and phosphate under these conditions corresponded to the increasing of Tetraspora sp. Rmutsb FPI 18 biomass. The best of each condition was combined and designated as “optimal condition”. The optimal condition in wastewater was used for cultivation of Tetraspora sp. Rmutsb FPI 18 and compared with control condition (wastewater initial pH 7, 25 ºC and 30 µmolphoton/m2/s for 18 h per day). The results revealed that Tetraspora sp. Rmutsb FPI 18 showed high capability to remove nitrate and phosphate from wastewater with removal efficiency of 93.65 and 96.65%, respectively under cultivated cell in optimal condition. Keywords: green algae, wastewater treatment, nitrate, phosphate, food processing industrial wastewater
บทนา
ปจจบนปญหายโทรฟเคชน (Eutrophication) เปนปญหามลพษทางน าปญหาหน(งทมในทกทวป ท(วโลก (Walker, 1983) โดยทาใหเกดการบลม (bloom) ของแพลงคตอนพชในแหลงน าตางๆ เชน หนอง คลอง บง ทะเลสาบ และ อางเกบน า ซ(งสาเหตดงกลาวเกดจาก
ในแหลงน ามสารประกอบไนโตรเจนและฟอสฟอรสในปรมาณมาก โดยธาตอาหารไนโตรเจนและฟอสฟอรสอาจเกดจากกจกรรมตางๆ ของมนษย เชน การเกษตร อตสาหกรรม น าเสยจากบานเรอนประชาชน และน าเสยจากโรงงานอตสาหกรรม เปนตน โดยสารประกอบไนโตรเจนท(พบในแหลงน า เชน ไนเตรท (NO3
-) ไนไตรท (NO2-)

RMUTSB Acad. J. 8(1) : 39-56 (2020) 41
และแอมโมเนย (NH4+) สวนสารประกอบฟอสฟอรสท(
สามารถพบได เชน ฟอสเฟต (PO43-) หรอ โอโทฟอสเฟต
(Orthophosphate) (Abdel-Raouf, AI-Homaidan, & Ibraheem, 2012) เปนตน ซ(งสารประกอบดงกลาว แพลงคตอนพชจะนาไปใชในการเจรญ เตบโตอยางรวดเรว ทาใหแหลงน าน นไมสามารถนามาใชประโยชนได อกท งยงกอใหเกดการเนาเสย เน(องจากแหลงน าจะขาดออกซเจน รวมท งแพลงคตอนพชและไซยาโนแบคทเรยบางชนด เชน Microcycstis sp. ยงสรางสารพษในแหลงน าซ(งเปนอนตรายตอผบรโภคไดอกดวย (Florczyk, Lakomiak, Woêny, & Brzuzan, 2014) ถงแมวาน าเสยจากแหลงชมชน หรอน าเสยจากโรงงานอตสาหกรรม จะมการบาบดน าเสยโดยสามารถลดสารอนทรยและสารอนนทรยในน าเสยแลว แตยงมธาตอาหาร เชน ไนเตรท และฟอสเฟต หลงเหลออย ดงน นถามการปลอยน าเสยดงกลาวลงสแหลงน าธรรมชาตกอาจจะเกดปรากฏการณยโทรฟเคชนได (Taziki, Ahmadzadeh, Murry, & Lyon, 2015) การกาจดไนเตรทและฟอสเฟตในแหลงน าสามารถทาไดหลายวธ เชน ทางเคม ทางกายภาพ/เคม และทางชวภาพ โดยใชส(งมชวต เชน ไซยาโนแบคทเรยและสาหรายสเขยว
สาหรายสเขยวมความสามารถในการกาจดไนโตรเจนและฟอสฟอรสอนนทรยในน าเสยไดเปนอยางด (Boelee, Temmink, Janssen, Buisman, & Wijffels, 2011; Abinandan, & Shanthakumar, 2015) ซ(งการกาจดไนโตรเจนและฟอสฟอรสดงกลาวเปนกระบวนการท(เปนมตรกบส(งแวดลอม (Shi, Podola, & Melkonian, 2007; Christenson, & Sims, 2011) นอกจากน การเพาะเล ยงสาหรายสเขยวในน าเสยนอกจากจะสามารถกาจดไนโตรเจนและฟอสฟอรสไดแลว ยงลดคาบโอด (BOD: biochemical oxygen demand) ซโอด (COD; chemical oxygen
demand) เพ(มดโอ (DO; dissolved oxygen) และลดปรมาณคารบอนไดออกไซดในบรรยากาศ ผานกระบวนการสงเคราะหดวยแสงไดอกดวย (Oswald, & Golueke, 1960) ย(งไปกวาน นชวมวลท(ไดจากสาหรายสเขยวสามารถนาไปเพ(มมลคาทางเศรษฐกจได เชน อาหารเสรม อาหารสตว เคร(องสาอางค และเช อเพลงชวภาพ รวมถงกาซมเทนท(ผานการยอยสลายโดยแบคทเรยแบบไมใชออกซเจน (Spolaore, Joannis-Cassan, Duran, & Isambert, 2006; Ometto et al., 2014) ไดมรายงานการคดแยกสาหราย สเขยวจากน าเสยในแหลงตาง ๆเชน น าเสยจากฟารมสกร โดยสามารถคดแยกสาหราย Oocystis sp. (Riaño, Molinuevo, & García-González, 2011) และ Chlorella vulgaris YSW-04 (Ji et al., 2013) น าเสยโรงงานสรา สามารถคดแยกสาหราย Chlorella sp. (Farooq et al., 2013) และน าเสยจากเทศบาล สามารถคดแยกสาหราย Micractinium reisseri และ Scenedesmus obliquus (Abou-Shanab et al., 2014) โดยมความคาดหวงวาจะนาสาหรายสเขยวท(คดแยกไดมาใชใหเกดประโยชน เชน การบาบดน าเสย อาหารสตว และเปนพลงงานทดแทน
สาหรายสเขยวแตละสายพนธจะมความสามารถในการเจรญและกาจดไนโตรเจน และฟอสฟอรสในแหลงน าเสย ไดแตกตางกน อกท งสภาวะแวดลอมตางๆ ในการเพาะเล ยง เชน คาพเอช อณหภม และความเขมแสง ยงสงผลตอการเจรญและการกาจดไนโตรเจนและฟอสฟอรสในน าเสยอกดวย (Taziki, Ahmadzadeh, Murry, & Lyon, 2015) ไดมรายงานการศกษาสภาวะท(เหมาะสมในการกาจดไนโตรเจนและฟอสฟอรสในสาหรายสเขยวหลากหลายสายพนธ เชน สาหรายสเขยว Chlorella vulgaris สามารถกาจดไนเตรท และฟอสเฟตไดถงรอยละ 85 และ 77 ตามลาดบ เม(อเพาะเล ยงใน

42 ว.มทรส. 8(1) : 39-56 (2563)
น าเสยท(มคาพเอช 2 อณหภม 26.3 องศาเซลเซยส ท(ระยะเวลา 2 วน (Sabeti, Hejazi, & Karimi, 2019) และสาหรายสเขยว Scenedesmus obliquus สามารถกาจดไนเตรท และฟอสฟอรสรวมไดถงรอยละ 100 และ 60 ตามลาดบ เม(อเพาะเล ยงในน าเสยท(อณหภม 25 องศาเซลเซยส ความเขมแสง 152 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท ท(ระยะเวลา 2.1 วน (Ruiz-Marin, Mendoza-Espinosa, & Stephenson, 2010) เปนตน
งานวจยน จงมวตถประสงคเพ(อคดแยกสาหรายสเขยวจากน าเสยท(มประสทธภาพในการกาจดไนเตรท และฟอสเฟตจากน าเสยในโรงงานอตสาหกรรมอาหารสาเรจรป ซ(งมปรมาณในไตรทและฟอสเฟตในน าเสยปรมาณสง นอกจากน จะทาการ จาแนกสายพนธและระบสายพนธของสาหรายสเขยวท(มประสทธภาพดงกลาว ดวยการศกษาลาดบนวคลโอไทดบรเวณ 18S rDNA และการทาแผนภมววฒนาการ และหาสภาวะท(เหมาะสม ไดแก คาพเอช อณหภม และความเขมแสง ในการกาจดไนเตรทและฟอสเฟตในน าเสยโดยสาหรายสเขยวท(คดเลอก
วธการศกษา
การคดแยกสาหรายสเขยวจากน าเสย การเพาะเล ยง และการวดน าหนกเซลลแหงของสาหรายสเขยว
เกบตวอยางน าเสยในโรงงานผลตอาหารสาเรจรป จากบอเปดพกน าในกระบวนการบาบดน าเสยทางชวภาพ จงหวดพระนครศรอยธยา โดยเกบจากบอตกตะกอน (sedimentation tank) ท(พกน ามาแลว 3 วน โดยเกบตวอยาง น าเสยใสขวดเกบตวอยาง ต งท งไวท(อณหภม 25 องศาเซลเซยส ใหแสงท(ความเขม 30 ไมโครโมล-โฟตอนตอตารางเมตร ตอวนาท เปนเวลา 7 วน จากน น
คดแยกสาหรายโดยใชเทคนค single cell isolation techique บนอาหารแขง BG11 (Rippka, Deruelles, Waterbury, Herdman, & Stanier, 1979) เม(อไดสาหรายสเขยวท(บรสทธ�แลว นามาเพาะเล ยงในฟลาสก ขนาด 250 มลลลตร ท(บรรจอาหารเหลว tris-acetate-phospate (TAP) (Harris, 1989) 100 มลลลตร โดยเพาะเล ยงสาหรายท(ความเขมแสง 30 ไมโครโมลโฟตอน ตอตารางเมตรตอวนาท ท(อณหภม 25 องศาเซลเซยส ในสภาวะเขยาท(ความเรวรอบ 120 รอบตอนาทเปนเวลา 5 วน จากน นศกษาสณฐานวทยาภายใตกลองจลทรรศนแบบใชแสง (Olympus, BX51, Japan) สาหรบการวดน าหนกเซลลแหง ดาเนนการโดยกรองเซลลของสาหรายสเขยวปรมาตร 10 มลลลตร ผานกระดาษกรอง GF/C (เสนผานศนยกลาง 47 มลลเมตร) (Whatman, UK) นาไปอบท(อณหภม 85 องศาเซลเซยส และนาไปวางไวในโถดดความช น 1 ช(งโมง กอนนามาช(งน าหนก (Taikhao, & Phunpruch, 2017)
การวเคราะหองคประกอบในน าเสย
เกบตวอยางน าเสยกอนบาบดจากโรงงานอตสาหกรรมผลตอาหารสาเรจรป จงหวดพระนครศรอยธยา โดยทาการวดคาออกซเจนละลายน า (dissolved oxygen, DO) และอณหภมดวยเคร(อง dissolved oxygen meter (Digicon, DO-552SD, Japan) และวดคาพเอชดวยเคร(อง pH meter (Mettler Toledo, FiveEasy, Switzerland) จากน นนาน าเสยมากรองดวยกระดาษกรองเบอร 3 (เสนผานศนยกลาง 90 มลลเมตร) (Whatman, UK) จากน น นาน าเสยมาปรบพเอชเร(มตนใหเทากบ 7.0 ดวย NaOH ความเขมขน 6.0 โมลาร และนาไปน(งฆาเช อ เพ(อนาไปทดสอบการบาบดน าเสยดวยสาหรายสเขยว โดยฆาเช อท(อณหภม 121 องศาเซลเซยส เปนเวลา 15 นาท จากน นนาน าเสยท(ปราศจากเช อมาวดคาบโอด

RMUTSB Acad. J. 8(1) : 39-56 (2020) 43
(biochemical oxygen demand, BOD) ดวยเคร(อง BOD analyzer (Lovibond, Oxi-Direct, Germany) ตามวธการของ Taikhao, & Phunpruch (2017) วดคาซโอด (chemical oxygen demand, COD) ตามวธการของ American Public Health Association (APHA) 5220 C (1998) วดปรมาณไนเตรทและฟอสเฟตดวยเคร(อง ion chromatograph (IC) (Thermo Fisher Scientific, Dionex ICS-5000+DP, USA) ตามวธการของ Taikhao, & Phunpruch (2017)
การเพาะเล ยงสาหรายสเขยวในน าเสย
เพาะเล ยงสาหรายสเขยวในฟลาสกขนาด 250 มลลลตร ท(มน าเสยท(ผานการฆาเช อแลว (พเอช 7.0) ปรมาตร 100 มลลลตร โดยใหมคาการดดกลนแสงเร(มตนท(ความยาวคล(น 750 นาโนเมตร (Griffiths, Garcin, van Hille, & Harrison, 2014) เทากบ 0.1 เพาะเล ยงในสภาวะท(มความเขมแสง 30 ไมโครโมลโฟตรอน ตอตารางเมตรตอวนาทเปนเวลา 18 ช(วโมงตอวน ในสภาวะเขยาท(ความเรว 120 รอบตอนาท ท(อณหภม 25 องศาเซลเซยส เปนระยะเวลา 5 วน ทดสอบการบาบดน าเสยดวยสาหรายสเขยว โดยกรองเซลลสาหรายสเขยวออกจากน าเสยดวยกระดาษกรอง GF/C และนาน าเสยท(ไดไปวเคราะหองคประกอบตางๆ ไดแก คาพเอช ดโอ บโอด ซโอด ไนเตรท และฟอสเฟต จากน นคดเลอกสาหรายสเขยวท(มประสทธภาพสงในการกาจดไนเตรทและฟอสเฟตมาศกษาสายพนธ และทดลองหาสภาวะ ท(เหมาะสมในการกาจดไนเตรทและฟอสเฟตตอไป
การจาแนกสายพนธ สาหรายส เขยวโดยการศกษาลาดบนวคลโอไทดบรเวณ 18S rDNA
การจาแนกสายพนธสาหรายสเขยวท(คดเลอกได โดยการศกษาลาดบนวคลโอไทดบรเวณ 18S rDNA โดยเร( มจากข นตอนการสกดจโนมกดเอนเอของ
สาหรายสเขยวดวยชดสกด DNA wizard SV genomic DNA purification system (Promega, USA) จากน นนาจโนมกดเอนเอท(ไดไปเพ(มปรมาณยน 18S rDNA ดวยปฏกรยา ลกโซพอลเมอเรส (polymerase chain reaction, PCR) โดยใช forward ไพรเมอร คอ F-18S rDNA Green algae 5’ CTGCGAATGGCTCATTAAATC-3’ และ reverse ไพรเมอร คอ R-18S rDNA green algae 5’-AAGGCC AGGGACGTAATCAA-3’ ปฏกรยาพซอารและสภาวะท(ใชดาเนนการตามวธการของ Phunpruch, Taikhao, & Incharoensakdi (2016) จากน นนาผลตภณฑพซอารไปตรวจดวยเทคนคอะกาโรสเจลอเลกโตรโฟเรซส ท(ความเขมขนของอะกาโรสรอยละ 0.80 จากน นนาผลตภณฑพซอารมาทาใหบรสทธ�ดวยชด GenepHlowTM Gel/PCR (Geneaid, Taiwan) จากน นสงวเคราะหลาดบนวคลโอไทด (1st Base, Singapore) โดยใชไพเมอร ท(จาเพาะในทศทาง 5’ คอ F-18S rDNA green algae 5’-CTGCGAATGGCTCATTAAATC-3’ จากน นนาลาดบ นวคลโอไทดมาเปรยบเทยบในฐานขอมล National Center for biotechnology information (NCBI) ดวยโปรแกรม Basic local alignment search tool nucleotide (BLASTN) และศกษาแผนภมววฒนาการ (Phylogenetic tree) ดวยวธ neighbor-joining โดยใช โปรแกรม MEGA 7 (Choi et al., 2018)
การหาสภาวะท(เหมาะสมในการกาจดไนเตรทและฟอสเฟตโดยสาหรายสเขยว
เพาะเล ยงสาหรายสเขยวในน าเสยท(ผานการฆาเช อแลว ในกรณการแปรผนคาพเอชในน าเสย ปรบคาพเอชเร(มตนในน าเสยดวยกรดไฮโดรคลอรกความเขมขน 6.0 โมลาร หรอโซเดยมไฮดรอกไซดความเขมขน 6.0 โมลาร เพ(อใหคาพเอชมคาเทากบ

44 ว.มทรส. 8(1) : 39-56 (2563)
4, 5, 6, 7, 8 และ 9 เพาะเล ยงในสภาวะท(มแสง 30 ไมโคร-โมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน ท(อณหภม 25 องศาเซลเซยส ในกรณการแปรผนอณหภมใหแปรผน อณหภมในการเพาะเล ยงเทากบ 25, 30, 35 และ 40 องศาเซลเซยส โดยเพาะเล ยงในสภาวะท(มคาพเอชเร(มตน เทากบ 7 และความเขมแสง 30 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน ในกรณการแปรผนความเขมแสง ใหแปรผนความเขมแสงในการเพาะเล ยงเทากบ 15, 30, 50, 100 และ 200 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน ในสภาวะท(มคาพเอชเร(มตนเทากบ 7 และอณหภมในการเพาะเล ยง 25 องศาเซลเซยส เม(อทดลองเพาะเล ยงสาหรายสเขยวในสภาวะตางๆ เปนระยะเวลา 5 วน วดปรมาณชวมวลโดยการวดน าหนกเซลลแหงและนาน าเสยท(ไดจากการกรองเซลลออกมาวเคราะหปรมาณไนเตรทและ ฟอสเฟตท(เหลออยในน าเสย โดยทาการทดลอง 3 ซ า และวเคราะหผลทางสถตดวยโปรแกรม SPSS 13.0 (SPSS Inc.) โดยใช one-way ANOVA ดวยวธ Duncan (alpha=0.05)
ผลการศกษาและอภปรายผล
ผลการคดเลอกสาหรายสเขยวท(มประสทธภาพในการกาจดไนเตรทและฟอสเฟต
จากการคดแยกสาหรายสเขยวจากน าเสยในโรงงานอตสาหกรรมผลตอาหารสาเรจรป จงหวดพระนครศรอยธยา สามารถคดแยกสาหรายสเขยวไดท งหมด 21 ไอโซเลท โดยสวนใหญมรปรางกลม แตจะมขนาดท(แตกตางกนออกไป ต งแตขนาด 4 ถง 16 ไมโครเมตร (ไมไดแสดงขอมล) ไดมรายงานการคดแยกสาหรายสเขยว
จากน าเสยในแหลงตาง ๆเชน สาหรายสเขยว Oocystis sp. และ Chlorella vulgaris YSW-04 คดแยกไดจากน าเสยจากฟารมสกร (Riaño, Molinuevo, & García-González, 2011; Ji et al., 2013) Chlorella sp. คดแยกจากน าเสยโรงงานสรา (Farooq et al., 2013) Micractinium reisseri และ Scenedesmus obliquus คดแยกไดจากน าเสย เทศบาล (Abou-Shanab et al., 2014) โดยสาหรายสเขยวท(คดแยกไดน นถกนาไปใชเปนพลงงานทดแทน อาหารสตว และนาไปบาบดน าเสยได โดยการศกษาในคร งน สามารถคดแยกสาหรายสเขยวไดหลากหลายชนด ดงน นอาจมความเปนไปไดวาจะมสาหรายสเขยวท(มความสามารถในการกาจด ไนเตรท และฟอสเฟตในน าเสยได นอกจากน ในการทดลองยงไดมการวเคราะหน าเสยเบ องตนจากโรงงานอตสาหกรรมผลต อาหารสาเรจรปกอนท(จะนามาทดลองเพาะเล ยงสาหราย ผลการทดลองพบวาในน าเสยมอณหภมเทากบ 31.00±0.30 องศาเซลเซยส คาพเอชเทากบ 5.66±0.05 คาดโอเทากบ 0.12±0.01 มลลกรมออกซเจนตอลตร คาบโอดเทากบ 318.00±48.21 มลลกรมออกซเจนตอลตร คาซโอดเทากบ 1,813.00±36.95 มลลกรมออกซเจนตอลตร ไนเตรท เทากบ 2,566.65 ±15.89 ไมโครกรมตอลตร และฟอสเฟตเทากบ 963.95±9.23 ไมโครกรมตอลตร จากน +นนาน +าเสย ดงกลาวมาทดลองเพาะเล +ยงสาหรายสเขยวท +ง 21 ไอโซเลท ท0คดแยกได เทยบกบน +าเสยท0ปราศจากการเตมสาหราย (control) เปนระยะเวลา 5 วน เพ0อท0จะคดเลอกสาหรายสเขยวท0ประสทธภาพในการกาจดไนเตรท และฟอสเฟต ผลการทดลองพบวาสาหรายสเขยวไอโซเลท FPI 18 สามารถลดคาไนเตรทและฟอสเฟตไดสงท0สด โดยสามารถลดคาไนเตรทและฟอสเฟตในน +าเสยไดถงรอยละ 62.82 และ 69.52
RMUTSB Acad. J. 8(1) : 39-56 (2020) 45
ตามลาดบ (Table 1) และเม0อตรวจสอบน +าเสยท0 ปราศจากการเตมสาหรายสเขยว พบวาคาไนเตรทและฟอสเฟตลดลงเพยงเลกนอยเทาน +น (Table 1) เปนการแสดงใหเหนวาคาไนเตรทและฟอสเฟตท0ลดลงเปนผลมาจากการเจรญของสาหรายสเขยว
นอกจากน + สาหรายสเขยวไอโซเลท FPI 18 ยงสามารถลดคาบโอด และซโอดไดสงท0สด และสามารถเพ0ม คาดโอใหน +าอกดวย (Table 1) จงนาสาหรายสเขยว ไอโซเลท FPI 18 ไปศกษาสายพนธและหาสภาวะท0เหมาะสมในการกาจดไนเตรท และฟอสเฟตตอไป
Table 1 Chemical characteristics of wastewater after cultivation 21 green algal strains for 5 days.
green algal isolate
chemical characteristics
pH DO
(mgO2/L) BOD
(mgO2/L) COD
(mgO2/L)
NO3-
(µg/L)
PO43-
(µg/L)
NO3-
removal (%) PO4
3-
removal (%)
control* 5.82±0.04 1.23±0.05 315.00±27.11 1803.00±23.12 2560.16±11.72 959.15±8.12 0.25 0.49 FPI 1 8.42±0.14 9.53±0.35 77.66±2.30 1734.00±12.20 2403.86±11.20 484.80±6.52 6.34 49.71 FPI 2 7.87±1.00 9.74±0.46 72.30±45.20 970.00±36.95 2354.12±10.80 634.38±7.34 8.28 34.19 FPI 3 8.45±0.06 8.26±0.25 51.30±0.00 1261.00±18.47 2161.97±9.15 650.03±8.11 15.77 32.57 FPI 4 9.02±0.11 9.73±0.34 76.30±3.05 1400.00±0.00 2108.76±13.45 540.38±6.45 17.84 43.94 FPI 5 7.88±0.16 9.12±0.42 83.00±28.79 864.00±32.00 2055.45±10.81 692.98±4.56 19.92 28.11 FPI 6 7.93±0.19 6.91±0.02 90.00±34.77 1,002.00±18.47 2038.68±19.76 494.54±3.89 20.57 48.70 FPI 7 8.46±0.10 9.45±0.78 55.66±7.09 1397.30±18.47 2025.16±14.12 579.31±4.11 21.10 39.90 FPI 8 8.52±0.14 9.46±0.52 63.00±6.08 789.33±48.80 2071.12±19.45 451.15±5.98 19.31 53.20 FPI 12 7.76±0.12 9.04±0.44 33.60±8.73 1034.00±18.47 1786.14±12.45 554.17±6.15 30.41 42.51 FPI 15 8.16±0.08 8.70±0.24 69.30±5.85 972.00±18.48 2116.23±21.49 657.15±7.32 17.55 31.83 FPI 17 8.00±0.46 9.38±0.23 54.70±37.81 981.00±48.88 1908.19±12.65 697.17±3.47 25.65 27.68 FPI 18 8.50±0.34 9.57±0.09 26.30±1.52 405.00±18.47 954.29±8.41 293.77±1.22 62.82 69.52 FPI 21 8.31±0.18 8.97±0.00 45.66±0.57 725.00±22.00 2149.85±19.11 764.12±3.45 16.24 20.73 FPI 24 8.35±0.15 8.35±0.19 65.00±2.00 1376.00±32.00 1894.56±15.23 786.15±6.54 26.19 18.44 FPI 26 8.34±0.21 8.61±0.16 152.30±9.00 810.00±36.95 2184.60±19.23 765.76±6.42 14.89 20.56 FPI 27 8.36±0.07 8.13±0.18 114.60±8.14 1525.00±36.95 2237.66±11.21 729.81±7.43 12.82 24.29 FPI 29 9.32±0.26 9.32±0.14 42.30±5.50 1013.00±66.61 1921.96±12.18 454.12±6.87 25.12 52.89 FPI 30 8.18±0.26 8.42±0.18 33.30±4.16 1098.00±48.88 2094.35±17.45 524.56±5.67 18.40 45.58 FPI 31 8.30±0.51 8.85±1.29 72.00±6.65 1098.00±48.88 2184.68±15.14 689.76±4.98 14.88 28.44 FPI 32 8.30±0.09 9.70±0.06 48.00±6.08 853.00±36.95 2317.80±15.34 665.67±5.34 9.70 30.94 FPI 34 8.84±0.09 7.71±0.14 78.30±7.50 725.00±18.47 2201.64±12.13 698.97±6.51 14.22 27.49 * Control is wastewater without green algae.
ผลการศกษาสายพนธของสาหรายสเขยวไอโซเลท
FPI 18 จากการศกษาสายพนธของสาหรายสเขยวไฮโซเลท
FPI 18 โดยการศกษาลาดบนวคลโอไทดบรเวณ 18S
rDNA ดวยเทคนคปฏกรยาลกโซพอลเมอเรส และนามารนเจลอเลกโตรโฟเรซส ผลการทดลองพบวาไดผลตภณฑพซอาร ขนาดประมาณ 1,500 คเบส ซ(งตรงกบขนาดผลตภณฑพซอารท(คาดหวงไว (ไมไดแสดงขอมล) จากน น

46 ว.มทรส. 8(1) : 39-56 (2563)
นาผลตภณฑ พซอารมาทาใหบรสทธ� และสงวเคราะหลาดบนวคลโอไทด โดยใช forward ไพรเมอร ผลจากการสงวเคราะหลาดบ นวคลโอไทด ไดลาดบนวคลโอไทดเทากบ 1,048 นวคลโอไทด โดยจากการทดลองไดลาดบนวคลโอไทดเพยงบางสวนเทาน น ซ(งยงไมครบตามจานวน ผวจยจงยงไมไดขอเลข accession number จากฐานขอมล NCBI โดยในอนาคตผวจยไดวางแผนท(จะสงวเคราะหลาดบนวคลโอไทดจากการใช reverse ไพรเมอร เพ(อใหไดลาดบนวคลไทดท(สมบรณ และขอเลข accession number ตอไป และจากการนาลาดบนวคลโอไทดบางสวนท(ไดไปเปรยบเทยบกบฐานขอมล NCBI โดยใชโปรแกรม BLASTN ผลการทดลองพบวาสาหรายสเขยว FPI 18 มลกษณะใกลเคยงกบสาหรายสเขยว Tetraspora sp. CU2551 (KT984853.1) มากท(สดถงรอยละ 98.93 ใกลเคยงกบสาหรายสเขยว Coelastrum astroideum var. rugosum strain UTEX2442 (GQ375393.0) และ Asterarcys sp. YACCYB527 (MH683929.1) รอยละ 98.73 อยางไรกตามเม(อนาลาดบนวคลโอไทดไปทาแผนภมวฒนาการดวยวธ neighbor-joining (NJ) phylogenetic tree (คา bootstrap เทากบ 1,000 replicates) โดยคดเลอกสาหรายสเขยวจากฐานขอมล NCBI โดยใชจนสของสาหรายสเขยว ท(มความหลากหลาย และมรอยละความเหมอนของลาดบนวคลโอไทดเม(อเทยบกบไอโซเลท FPI 18 จากตาราง BLASTN อยในชวงรอยละ 98.54 ข นไป โดยใชสาหรายสเขยว Catena varidis clone KR 914 เปน out group จากการทดลองพบวา สาหรายสเขยว FPI 18 แยกกลมออกมาจากสาหรายสเขยวจนสอ(นๆ อยางชดเจน (Figure 1) อยางไรกตามลาดบ นวคลโอไทดของสาหรายสเขยว FPI 18 ท(นามาทาแผนภมววฒนาการเปนลาดบนวคลโทดเพยงบางสวนเทาน น
ดงน นถาไดลาดบนวคลโอไทดท(สมบรณมากกวาน อาจทาใหการจดกลมของสาหรายทาไดถกตองมากย(งข น นอกจากน เม(อศกษาสณฐานวทยาภายใตกลองจลทรรศนพบวาสาหรายสเขยว FPI 18 มลกษณะใกลเคยงกบ Tetraspora sp. CU2551 (Maneeruttanarungroj, Lindblad, & Incharoensakdi, 2010) แตเซลลมขนาดเลกกวา (Figure 2A และ 2B) โดยสาหรายสเขยว FPI 18 มรปรางกลม ขนาดประมาณ 5-6 ไมโครเมตร โดยจะเหนเปนเซลลเดยว หรอเหนเปน 3 และ 4 เซลล ภายใตชทหมท(โปรงแสง (Figure 2) ซ(งเปนลกษณะท(สาคญของสาหรายสเขยวในจนสน ดงน นผวจยจงต งช(อสาหรายสเขยวไอโซเลท FPI 18 น วา Tetraspora sp. Rmutsb FPI 18 และนาสาหรายสายพนธน ไปศกษาสภาวะท(เหมาะสมในการกาจดไนเตรทและฟอสเฟตในน าเสยตอไป
ผลของคาพเอชตอการกาจดไนเตรทและฟอสเฟตในสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 จากการเพาะเล ยงสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 ในน าเสยท(ผานการฆาเช อแลว และแปรผนคาพเอชในน าเสยเทากบ 4, 5, 6, 7, 8 และ 9 เพาะเล ยงเปนระยะเวลา 5 วน ผลการทดลองพบวา ในน าเสยท(มคาพเอชเร(มตนเทากบ 8 ใหคาการกาจดไนเตรทและฟอสเฟตดท(สดและแตกตางจากคาพเอชอ(นๆ อยางมนยสาคญทางสถต โดย Tatraspora sp. Rmutsb FPI 18 สามารถกาจดไนเตรทและฟอสเฟตไดรอยละ 79.74 และ 80.32 ตามลาดบ (Table 2) จากผลการทดลองท(ไดสอดคลองกบงานวจยของ Sabeti, Hejazi, & Karimi (2019) ท(สภาวะท(เหมาะสมในการกาจดไนเตรทและฟอสเฟตในน าเสยโดยสาหรายสเขยว Chlorella vulgaris ใหคากาจดไนเตรทและฟอสเฟตสงสดในน าเสยท(มคาพเอชเทากบ 8 นอกจากน ผลของการลดลงของไนเตรท

RMUTSB Acad. J. 8(1) : 39-56 (2020) 47
และฟอสเฟตยงสอดคลองกบชวมวล (วดจากน าหนกเซลลแหง) ท(เพ(มข น โดยท(คาพเอช 8 ใหคาชวมวลสงสด และแตกตางจากคาพเอชอ(นๆ อยางมนยสาคญทางสถต โดยมคาชวมวลเทากบ 0.693±0.045 กรมตอลตร (Table 2) นอกจากน ท(คาพเอช 8 ในการทดลองของ Li, Horsman, Wang, Wu, & Lan (2008) ทาใหสาหรายสเขยว Neochloris oleoabundans มการสะสมชวมวลสงสด และสามารถกาจดไนเตรทไดมากท(สดอกดวย โดยท(วไปแลวคาพเอชมผลตอการเจรญของสาหรายสเขยว โดยคาพเอชท(เหมาะสมตอการเจรญจะอยในชวง 6-8 (Larsdutter, 2006) คาพเอชท(สงหรอต(าเกนไปอาจสงผลตอการ เจรญเตบโต การทางานของออรแกเนลล เอนไซม และโปรตนตางๆ ภายในเซลลของสาหรายสเขยว (Jia, & Yuan, 2306) นอกจากน
คา พเอชยงสงผลตอการดดซมสารอาหาร เชน แอมโมเนย ไนเตรทและฟอสเฟตในแหลงน าไดอกดวย โดยปกตเม(อสาหรายมการตรงคารบอนไดออกไซด จะมการผลต OH- สงผลทาใหคาพเอชในแหลงน าสงข น ซ(งคาพเอชท(สงข นมแนวโนมตอการเพ(มการดดซมไนเตรท และฟอสเฟต ทาใหปรมาณไนเตรทและฟอสเฟตในแหลงน าลดลงได (Sayadi, Ahmadpour, Capoorchali, & Rezaei, 2016) ในการทดลองน ช ใหเหนวาคาพเอชเทากบ 8 เปนคาท(เหมาะสมในการเจรญและการกาจดไนเตรทและฟอสเฟตของสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 เปนไปไดวาท(คาพเอชดงกลาว ทาใหสาหรายมการดดซมไนเตรทและฟอสเฟตไปใชในการเจรญเตบโตมากข น ทาใหน ามไนเตรทและฟอสเฟตลดลงมากท(สด
Figure 1 Phylogenetic tree using the obtained 18S rDNA sequence of green algae isolate FPI 13 (underlined) and 17 other green algae species. The accession number available in the NCBI database shows in front of each name.

48 ว.มทรส. 8(1) : 39-56 (2563)
Figure 2 Morphological characterization of Tatraspora sp. CU2551 (A) (Maneeruttanarungroj, Lindblad, & Incharoensakdi, 2010) and Tatraspora sp. Rmutsb FPI 18 (B) under light microscope.
Table 2 Effect of initial pH on nitrate removal, phosphate removal and biomass under cultivated cell of Tatraspora sp. Rmutsb FPI 18 in wastewater for 5 days.
pH nitrate (NO3
-) phosphate (PO43-)
biomass (g/L)
NO3- concentration
(µg/L) NO3
- removal (%)
PO43- concentration
(µg/L) PO4
3- removal (%)
4 1186.87±6.56 53.76d 489.45±3.45 49.22d 0.273e±0.128 5 1045.36±8.83 59.27c 402.18±2.69 58.28c 0.347d±0.005 6 997.98±9.39 61.12c 376.78±3.90 60.91c 0.435c±0.017 7 954.29±8.41 62.82c 293.77±1.22 69.52b 0.517b±0.011 8 520.12±5.12 79.74a 189.75±2.23 80.32a 0.693a±0.045 9 852.12±6.09 66.80b 389.52±2.19 59.59c 0.491c±0.037 The lowercase letters in the same column represent significant differences according to Duncan’s test (alpha = 0.05).
ผลของอณหภมตอการกาจดไนเตรทและฟอสเฟต
ในสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 จากการเพาะเล ยงสาหรายสเขยว Tatraspora
sp. Rmutsb FPI 18 ในน าเสยท(ผานการฆาเช อแลว และ แปรผนอณหภมในการเพาะเล ยงเทากบ 25, 30, 35 และ 40 องศาเซลเซยส เพาะเล ยงเปนระยะเวลา 5 วน ผลการทดลองพบวา ในสภาวะการเพาะเล ยงท(อณหภม 30 องศาเซลเซยส ใหคาการกาจดไนเตรท
และฟอสเฟตดท(สด และแตกตางจากอณหภมอ(นๆ อยางมนยสาคญทางสถต โดย Tatraspora sp. Rmutsb FPI 18 สามารถกาจดไนเตรทและฟอสเฟตไดรอยละ 88.62 และ 89.08 ตามลาดบ (Table 3) นอกจากน ยงสอดคลองกบชวมวลท(มการสะสมมากท(สดเม(อเพาะเล ยง Tatraspora sp. Rmutsb FPI 18 ท(อณหภม 30 องศาเซลเซยส โดยใหคาชวมวลเทากบ 0.858±0.004 กรมตอลตร (Table 3) การกาจดไนเตรทและฟอสเฟต
A B
RMUTSB Acad. J. 8(1) : 39-56 (2020) 49
ไดดท(อณหภม 30 องศาเซลเซยส สอดคลองกบงานวจยของ Martínez, Sánchez, Jiménez, Yousfi, & Muñoz (2000) ท(เพาะเล ยงสาหรายสเขยว Scenedesmus
obliquus ในน าเสยชมชน และยงสอดคลองกบงานวจยของ Ogbonna, Yoshizawa, & Tanaka (2000) ท(เพาะเล ยงสาหรายสเขยว Chlorella sorokiniana และไซยาโน-แบคทเรย Spirulina platensis ในน าเสยท(อณหภม 30 องศาเซลเซยส สามารถกาจด ไนเตรทไดถงรอยละ 45.4 และ 100 ตามลาดบ โดยท(วไปอณหภมมผลตอการเจรญเตบโตของสาหรายสเขยว เน(องจากอณหภมมผลตอกจกรรมของเอนไซมตางๆ และมผลตอ แมแทบอลซมตางๆ ภายในเซลล (Taziki, Ahmadzadeh, Murry, & Lyon, 2015) โดยอณหภมท(เหมาะสมตอการเจรญเตบโตของสาหรายในเขตรอนช นจะอยท(ประมาณ
25 องศาเซลเซยส (Renaud, Thinh, Lambrinidis, & Parry, 2002) สาหรายสเขยวบางสายพนธ เม(ออณหภมในน าสงข น อตราการสงเคราะหดวยแสงจะสงข น ซ(งสงผลตอการดซมสารอาหาร รวมถงการนาไนเตรทและฟอสเฟตเขาสเซลลไดมากข น (Jiménez, & Niell, 1991) โดยในการศกษาคร งน ท(อณหภม 30 องศาเซลเซยส สาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 สามารถกาจดไนเตรทและฟอสเฟตไดสงกวาท(อณหภมต(า (25 องศาเซลเซยส) แตเม(อเพ(มอณหภมใหสงข นเปน 35 และ 40 องศาเซลเซยส การกาจดไนเตรทและฟอสเฟตกลบลดลง แสดงใหเหนวาสภาวะดงกลาวไมเหมาะสมตอการทางานของเอนไซม และ/หรอการสงเคราะหดวยแสงในสาหราย สเขยวสายพนธน
Table 3 Effect of temperature on nitrate removal, phosphate removal and biomass under cultivated cell of Tatraspora sp. Rmutsb FPI 18 in wastewater for 5 days.
temperature (oC) nitrate (NO3
-) phosphate (PO43-)
biomass (g/L) NO3
- concentration (µg/L) NO3- removal (%) PO4
3- concentration (µg/L) PO43- removal (%)
25 954.29±8.41 62.82d 293.77±1.22 69.52c 0.517c±0.011 30 292.11±2.99 88.62a 105.23±1.33 89.08a 0.858a±0.004 35 498.12±3.12 80.59b 250.12±2.14 74.05b 0.629b±0.045 40 692.15±3.43 73.03c 298.14±2.18 69.07c 0.522c±0.017 The lowercase letters in the same column represent significant differences according to Duncan’s test (alpha = 0.05).
ผลของความเขมแสงตอการกาจดไนเตรทและ
ฟอสเฟตในสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18
จ า ก ก า ร เ พ า ะ เ ล ย ง ส า ห ร า ย ส เ ข ย ว Tatraspora sp. Rmutsb FPI 18 ในน าเสยท(ผานการฆา เ ช อแ ลว และแปรผนความเ ขมแสงในการ
เพาะเล ยงเทากบ 15 30, 50, 100 และ 200 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน เพาะเล ยงเปนระยะเวลา 5 วน ผลการทดลองพบวาในสภาวะการเพาะเล ยงท(ความ เขมแสง 50 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท ใ ห ค า ก า ร ก า จด ไน เ ต ร ท แ ล ะ ฟ อ สเ ฟ ต ด ท( สด

50 ว.มทรส. 8(1) : 39-56 (2563)
และแตกตางจากความเขมแสงอ(นๆ อยางมนยสาคญทางสถต โดย Tatraspora sp. Rmutsb FPI 18 สามารถกาจดไนเตรทและฟอสเฟตไดรอยละ 77.40 และ 83.39 ตามลาดบ (Table 4) นอกจากน ยงสอดคลองกบชวมวลท(มการสะสมมากท(สดเม(อเพาะเล ยง Tatraspora sp. Rmutsb FPI 18 ท(ความเ ข ม แ ส ง ด ง ก ล า ว โ ด ย ใ ห ค า ช ว ม ว ล เ ท า ก บ 0.667±0.040 กรมตอลตร (Table 4) ซ(งสอดคลองกบการรายงานกอนหนา น ในสาห รายส เ ข ยว Haematococcus pluvialis (Kang, An, Park, & Sim, 2006) และ Scenedesmus accutus pvuw02 (Doria et al., 2012) ซ(งสามารถกาจดไนเตรทไดด เม(อเพาะเล ยงท(ความเขมแสง 50 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท และใกลเคยงกบการรายงานในสาหรายสเขยว Chlorella vulgaris ท(สามารถกาจดไนเตรทและฟอสเฟตจากน าเสยไดดเม(อเพาะเล ยงในสภาวะท(มความเขมแสง 58 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท (Lau, Tam, & Wong, 1995) โดยท(วไปแลวแสงเปนท(ปจจยสาคญในการสงเคราะหดวยแสงของสาหรายสเขยว ซ(งทาใหสาหรายสามารถเจรญเตบโตและเพ(มชวมวล อกท งยงเก(ยวของกบกระบวนการกาจดสารอาหารในน าเสย โดยการเพ(มความเขมแสงจะทาใหสาหรายสเขยวมการนาไนเตรทและฟอสเฟตเขาสเซลลมากข น สงผลใหไนเตรทและฟอสเฟตในแหลงน าลดลงได (Taziki, Ahmadzadeh, Murry, & Lyon, 2015) ซ(งสอดคลองกบการทดลองในคร งน เม(อเพ(มความเขมแสง (15-50 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท) การกาจดไนเตรทและฟอสเฟตของสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 เพ(มสงข น อยางไรกตามเม(อเพ(มความ
เขมแสงใหมากข นเปน 100-200 ไมโครโมลโฟตอน ตอตารางเมตรตอวนาท การกาจดไนเตรทและฟอสเฟตลดลง อาจเปนไปไดวาเม(อความเขมแสงสงข น เซลลอาจเกดกระบวนการ photoinhibition ซ(งทาใหการเจรญเตบโต และการสงเคราะหดวยแสงท(ลดลง สงผลใหการกาจดไนเตรทและฟอสเฟตลดลง (Markou, & Georgakakis, 2011)
จากการทดลองแปรผนคาพเอช อณหภม และความเขมแสง ทาใหทราบสภาวะท(เหมาะสมของ แตละปจจย จงนาสภาวะท(เหมาะสมของแตละปจจยมารวมกนเพ(อศกษาการกาจดไนเตรทและฟอสเฟตโดยสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 โดยสภาวะท(เหมาะสมประกอบดวย คาพเอชเร(มตนเทากบ 8 อณหภมเทากบ 30 องศาเซลเซยส และความเขมแสงเทากบ 50 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เรยกสภาวะดงกลาววา “สภาวะท(เหมาะสม“ หรอ “optimal condition“ โดยทาการทดลองเปรยบเทยบกบสภาวะปกต ท( ใ ช ในการเพาะเล ยงคอ คาพเอชเร(มตน 7 อณหภม 25 องศา-เซลเซยส และความเขมแสง 30 ไมโครโมลโฟตอน ตอตารางเมตรตอวนาท เ รยกสภาวะดงกลาววา “สภาวะควบคม” หรอ “control condition” ผลการทดลองแสดงดง (Figure 3) ซ(งพบวาเม(อเพาะเล ยงสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 ในน าเสยสภาวะท(เหมาะสม ใหคาการกาจดไนเตรทและฟอสเฟตสงกวาสภาวะควบคม โดยสามารถกาจดไนเตรทและฟอสเฟตไดถงรอยละ 93.65 และ 95.65 ตามลาดบ (Figure 3) โดยเม(อเปรยบเทยบการกาจดไนเตรทและฟอสเฟตของ Tatraspora sp. Rmutsb FPI 18 กบสาหรายสเขยวสายพนธตางๆ
RMUTSB Acad. J. 8(1) : 39-56 (2020) 51
ท(มในฐานขอมล (Table 5) พบวา Tatraspora sp. Rmutsb FPI 18 มประสทธภาพในการกาจดไนเตรทและฟอสเฟตใกลเคยงกบสาหรายสเขยวท(มรายงาน และยงมประสทธภาพในการกาจดไนเตรทและฟอสเฟตไดดกวาสาหรายส เขยวบางสายพนธ เชน Chlorella sorokiniana (Ogbonna, Yoshizawa, & Tanaka, 2000) และ Chlorella vulgaris (González, Cañizares, & Baena, 1997; Ruiz-Marin, Mendoza- Espinosa, & Stephenson, 2010; Sabeti, Hejazi, & Karimi, 2019) เปนตน เปนการแสดงใหเหนวาสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 มประสทธภาพในการกาจดไนเตรทและฟอสเฟตจากน าเสยได นอกจากน ผลพลอยไดจากการเพาะเล ยงสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 ในน าเสยคอชวมวล ซ(งสามารถนาไปใชประโยชน ในดานตางๆ ได เชน พลงงานทดแทน อาหารเสรม และ อ าห า ร สต ว อย า ง ไ ร ก ต า มก า ร ทด ลอ ง น ไมไดวดคา total Kjeldahl nitrogen (TKN) และ
total phosphorus ซ(งเปนคาพารามเตอรท(กาหนดโดยกรมโรงงานอตสาหกรรมและกรมควบคมมลพษ จงไมสามารถกลาว ไดวาน าเสยท(ผานการบาบด โดยสาหรายสเขยวน น ผานเกณฑมาตรฐานน าเสยตามท(กรมโรงงานอตสาหกรรมและกรมควบคมมลพษกาหนดไวหรอไม นอกจากน น า เสยท( ใ ช ในการทดลองเปนน าเสยท(ผานการฆาเช อแลว ซ(งเม(อนาไปใชจรงอาจมปญหาจากการปนเป อนแ บ ค ท เ ร ย แ ละ เ ช อ อ( น ๆ ซ( ง อ า จ ส ง ผลตอ ก า รเจรญเตบโตของสาหรายได ดงน นในการทดลอง คร งถดไปผ วจยไดวางแผนท(จะทดลองเพาะเล ยงสาหรายสเขยว Tatraspora sp. Rmutsb FPI 18 ในน า เสยท( ไมผานการฆา เ ช อ และวดคา TKN และ total phosphorus เพ(มเตม เพ(อท(จะจาลองสถานการณใหใกลเคยงกบความเปนจรง และ เ ทยบเกณฑมาตรฐานน า เสยกบกรมโรงงานอต ส า ห ก ร ร ม แ ล ะ ก ร ม ค ว บ คม ม ล พ ษ ใ น ก า ร บาบดน าเสยตอไป
Table 4 Effect of light intensity on nitrate removal, phosphate removal and biomass under cultivated cell of Tatraspora sp. Rmutsb FPI 18 in wastewater for 5 days.
light intensity (µmolphoton/m2/s)
nitrate (NO3-) phosphate (PO4
3-) biomass
(g/L) NO3
- concentration (µg/L)
NO3- removal (%)
PO43- concentration
(µg/L) PO4
3- removal (%)
15 1098.87±12.36 57.19d 389.12±2.33 59.63d 0.326d±0.023 30 954.29±8.41 62.82c 293.77±1.22 69.52c 0.517c±0.011 50 580.15±9.15 77.40a 160.11±1.14 83.39a 0.667a±0.040 100 750.22±6.71 70.77b 256.65±2.59 73.38b 0.598b±0.012 200 940.15±5.82 63.37c 390.11±2.45 59.53d 0.413c±0.015 The lowercase letters in the same column represent significant differences according to Duncan’s test (alpha = 0.05).
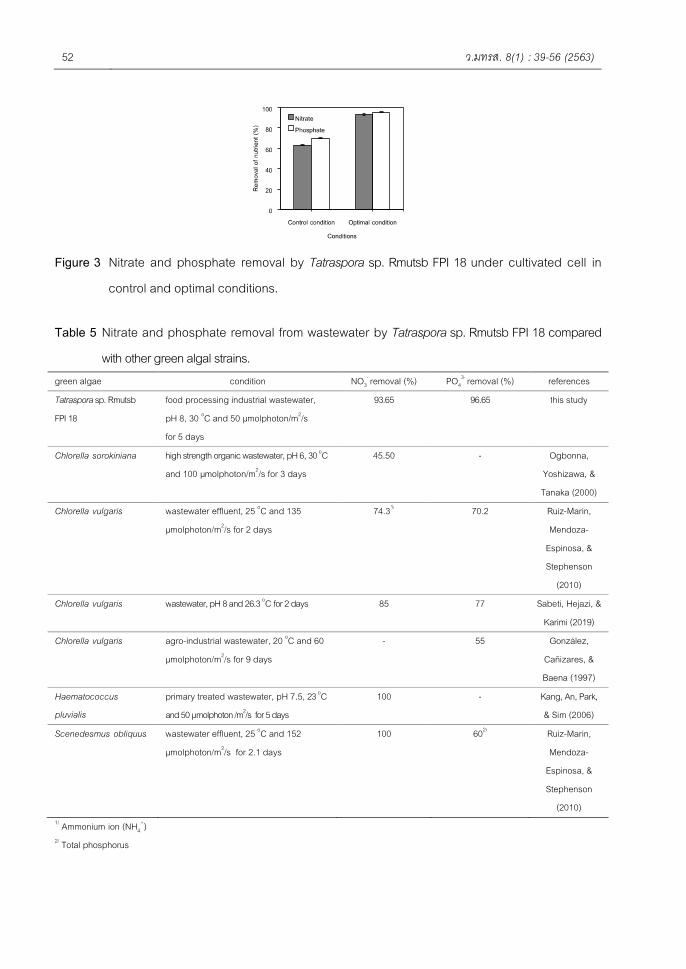
52 ว.มทรส. 8(1) : 39-56 (2563)
Figure 3 Nitrate and phosphate removal by Tatraspora sp. Rmutsb FPI 18 under cultivated cell in control and optimal conditions.
Table 5 Nitrate and phosphate removal from wastewater by Tatraspora sp. Rmutsb FPI 18 compared with other green algal strains. green algae condition NO3
- removal (%) PO43- removal (%) references
Tatraspora sp. Rmutsb FPI 18
food processing industrial wastewater, pH 8, 30 oC and 50 µmolphoton/m2/s for 5 days
93.65
96.65 this study
Chlorella sorokiniana high strength organic wastewater, pH 6, 30 oC and 100 µmolphoton/m2/s for 3 days
45.50 - Ogbonna, Yoshizawa, & Tanaka (2000)
Chlorella vulgaris wastewater effluent, 25 oC and 135 µmolphoton/m2/s for 2 days
74.31/ 70.2 Ruiz-Marin, Mendoza-
Espinosa, & Stephenson
(2010) Chlorella vulgaris wastewater, pH 8 and 26.3 oC for 2 days 85 77 Sabeti, Hejazi, &
Karimi (2019) Chlorella vulgaris agro-industrial wastewater, 20 oC and 60
µmolphoton/m2/s for 9 days - 55 González,
Cañizares, & Baena (1997)
Haematococcus
pluvialis
primary treated wastewater, pH 7.5, 23 oC and 50 µmolphoton /m2/s for 5 days
100 - Kang, An, Park, & Sim (2006)
Scenedesmus obliquus wastewater effluent, 25 oC and 152 µmolphoton/m2/s for 2.1 days
100 602/ Ruiz-Marin, Mendoza-
Espinosa, & Stephenson
(2010) 1/ Ammonium ion (NH4
+) 2/ Total phosphorus

RMUTSB Acad. J. 8(1) : 39-56 (2020) 53
สรป
จากการคดแยกสาหรายสเขยวในโรงงานอตสาหกรรมผลตอาหารสาเรจรป จงหวดพระนครศรอยธยา สามารถคดแยกสาหรายสเขยวได 21 ไอโซเลท จากน น นาท ง 21 ไอโซเลทมาศกษาการกาจดไนเตรทและฟอสเฟต ผลการทดลองพบวาสาหรายสเขยวไอโซเลท FPI 18 มความสามารถมากท(สดในการกาจดไนเตรทและฟอสเฟต และจากศกษาสายพนธ โดยการศกษาลาดบนวคลโอไทดบรเวณ 18S rDNA และศกษาแผนภมวฒนาการของสาหรายสเขยวไอโซเลท FPI 18 พบวาสาหรายสเขยวไอโซเลท FPI 18 มความใกลชดและจดอยในจนส Tetraspora sp. จงต งช(อสาหรายชนดน วา Tetraspora sp. Rmutsb FPI 18 โดย Tetraspora sp. Rmutsb FPI 18 มรปรางกลม ขนาด 5-6 ไมโครเมตร อยเปนเซลลเดยว หรออยรวมกนเปนกลม 3-4 เซลลภายใตชทห มท(โปรงแสง จากการศกษาสภาวะท(เหมาะสมในการกาจดไนเตรทและฟอสเฟต พบวาสภาวะท(เหมาะสมคอ คาพเอชเร(มตนในน าเสยมคาเทากบ 8 อณหภมในการเพาะเล ยงคอ 30 องศาเซลเซยส และใหความเขมแสงท( 50 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน และเม(อนาสภาวะดงกลาวทกสภาวะมารวมกน พบวาสาหรายสเขยว Tetraspora sp. Rmutsb FPI 18 สามารถกาจดไนเตรทและฟอสเฟตไดถงรอยละ 93.65 และ 96.65 ตามลาดบ และสงกวาสภาวะท(ใชเพาะเล ยงปกต (พเอชเร(มตนเทากบ 7 อณหภม 25 องศาเซลเซยส และความเขมแสง 30 ไมโครโมลโฟตอนตอตารางเมตรตอวนาท เปนเวลา 18 ช(วโมงตอวน) ท(ระยะเวลา 5 วน และเม(อนาคาการกาจดไนเตรทและฟอสเฟตของสาหรายสเขยว Tetraspora sp. Rmutsb FPI 18 ไปเทยบกบสาหรายสเขยวสายพนธ
อ(นๆ ในฐานขอมล พบวาสาหรายสเขยว Tetraspora sp. Rmutsb FPI 18 มความสามารถสงในการกาจดไนเตรทและฟอสเฟตใกลเคยงกบสาหรายท(เคยมรายงานไว เปนการแสดงใหเหนวาสาหรายสเขยว Tetraspora sp. Rmutsb FPI 18 มประสทธภาพ การกาจดไนเตรทและฟอสเฟตออกจากน าเสยได อกท งยงสามารถนาชวมวลของสาหรายสเขยว ท(ไดไปใชประโยชนในดานตางๆ เชน พลงงานทดแทน อาหารเสรมและอาหารสตว เปนตน
คาขอบคณ
งานวจย น ไ ด รบการสนบสนนจากคณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ขอขอบคณสาขาวชาวทยาศาสตร (ชววทยา) คณะวทยาศาสตรและเทคโนโลย ศนยพระนครศรอยธยา หนตรา ท(ใหความอนเคราะหสถานท(และเคร(องมอในการทางานวจย และขอขอบคณโรงงานอตสาหกรรมผลตอาหารสาเรจรป จงหวดพระนครศรอยธยา ท(ใหความอนเคราะหในการเขาไปเกบตวอยางน าเสยมาทางานวจยในคร งน
เอกสารอางอง
Abdel-Raouf, N., AI-Homaidan, A. A., & Ibraheem, I. B. M. (2012). Microalgae and wastewater treatment. Saudi Journal of Niological Science, 19, 257-275.
Abou-Shanab, R. A. I., El-Dalatony, M. M., EL-Sheekh, M. M., Ji, M. K., Salama, E. S., Kabra, A. N., & Jeon, B. H. (2014). Cultivation of a new microalga, Micractinium
reisseri, in municipal wastewater for nutrient removal, biomass, lipid, and fatty acid production. Biotechnology
Bioprocess, Engineering, 19, 510-518.

54 ว.มทรส. 8(1) : 39-56 (2563)
Abinandan, S., & Shanthakumar, S. (2015). Challenges and opportunities in application of microalgae (Chlorophyta) for wastewater treatment: a review. Renewable & Sustainable Energy Reviews, 52, 123-132.
American Public Health Association (APHA). (1998). Methods for biomass production. In Standard
Methods for the Examination of Water and
Wastewater. Baltimore MD: American Public Health Association.
Boelee, N. C., Temmink, H., Janssen, M., Buisman, C. J. N., & Wijffels, R. H. (2011). Nitrogen and phosphorus removal from municipal wastewater effluent using microalgal biofilms. Water Research,
45(18), 5925-5933. Choi, H. J., Joo, J. H., Kim, J. H., Wang, P., Ki, J. S., & Han,
M. S. (2018). Morphological characterization and molecular phylogenetic analysis of Dolichospermum
hangangense (Nostocales, Cyanobacteria) sp. nov. from Han River, Korea. Algae, 33(2), 143-156.
Christenson, L., & Sims, R. (2011). Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnology Advances, 29(6), 686-702.
Doria, E., Longoni, P., Scibilia, L., Iazzi, N., Cella, R., & Nielsen, E. (2012). Isolation and characterization of a Scenedesmus acutus strain to be used for bioremediation of urban wastewater. Journal of
Applied Phycology, 24(3), 375-83. Farooq, W., Lee, Y. C., Ryu, B. G., Kim, B. H., Kim, H. S.,
Choi, Y. E., & Yang. J. W. (2013). Two stage cultivation of two Chlorella sp. strains by simultaneous treatment of brewery wastewater and maximizing lipid productivity. Bioresource
Technology, 132, 230 -238.
Florczyk, M., Lakomiak, A., Woêny, M., & Brzuzan, P. (2014). Neurotoxicity of cyanobacterial toxins. Environmental Biotechnology, 10(1), 26-43.
González, L. E., Cañizares, R. O., & Baena, S. (1997). Efficiency of ammonia and phosphorus removal from a colombian agroindustrial wastewater by the microalgae Chlorella vulgaris and Scenedesmus dimorphus. Bioresource Technology, 60(3), 259-262.
Griffiths, M. J., Garcin, C., van Hille, R. P., & Harrison, S. T. (2011). Interference by pigment in the estimation of microalgal biomass concentration by optical density. Journal of Microbiological Methods, 85, 119-23.
Ji, M. K., Abou-Shanab, R. A. I., Kim, S. H., Salama, E. S., Lee, S. H., Kabra, N. A., Lee, Y. S., Hongf, S., & Jeon, B. H. (2013). Cultivation of microalgae species in tertiary municipal wastewater supplemented with CO2 for nutrient removal and biomass production. Ecological Engineering, 58, 142 -148.
Jia, H., & Yuan, Q. (2016). Removal of nitrogen from wastewater using microalgae and microalgae–bacteria consortia. Cogent Environmental Science,
2, 1-15. Jiménez, C., & Niell, F. X. (1991) Growth of Dunaliella
viridis teodoresco: effect of salinity, temperature and nitrogen concentration. Journal of Applied
Phycolog, 3(4), 319-327. Kang, C. D., An, J. Y., Park, T. H., & Sim, S. J. (2006).
Astaxanthin biosynthesis from simultaneous N and P uptake by the green alga Haematococcus
pluvialis in primary treated wastewater. Biochemical
Engineering Journal, 31(3), 234-238. Larsdotter, K. (2006). Wastewater treatment with microalgae:
a literature review. Journal of Water Management
and Research (Vatten), 62, 31-38.

RMUTSB Acad. J. 8(1) : 39-56 (2020) 55
Lau, P. S., Tam, N. F. Y., & Wong, Y. S. (1995). Effect of algal density on nutrient removal from primary settled wastewater. Environmental Pollution, 89(1), 59-66.
Li, Y., Horsman, M., Wang, B., Wu, N., & Lan, C.Q. (2008). Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Applied Microbiology
and Biotechnology, 81(4), 629-36. Maneeruttanarungroj, C., Lindblad, P., & Incharoensakdi,
A. (2010). A newly isolated green alga, Tetraspora sp. CU2551, from Thailand with efficient hydrogen production. International
Journal of Hydrogen Energy, 35, 13193-13199. Markou, G., & Georgakakis, D. (2011). Cultivation of
filamentous cyanobacteria (blue-green algae) in agro-industrial wastes and wastewaters: A review. Applied Energy, 88(10), 3389-3401.
Martínez, M. E., Sánchez, S., Jiménez, J. M. E. l., Yousfi, F., & Muñoz, L. (2000). Nitrogen and phosphorus removal from urban wastewater by the microalga Scenedesmus obliquus. Bioresource Technology,
73(3), 263-272. Ogbonna, J. C., Yoshizawa, H., & Tanaka, H. (2000).
Treatment of high strength organic wastewater by a mixed culture of photosynthetic microorganisms. Journal of Applied Phycology,
12(3-5), 277-284. Ometto, F., Pozza, C., Whitton R., Smyth, B., Torres, A. G.,
Henderson, R. K., Jarvis, P., Jefferson, B., & Villa, R. (2014). The impacts of replacing air bubbles with microspheres for the clarification of algae from low cell-density culture. Water
Research, 53, 168-179.
Oswald, W. J, & Golueke, C. G. (1960). Biological transformation of solar energy. Advances in
Applied Microbiology, 2, 223-262. Phunpruch, S., Taikhao, S., & Incharoensakdi, A. (2016).
Identification of bidirectional hydrogenase genes and their co-transcription in unicellular halotolerant cyanobacterium Aphanothece halophytica. Journal
of Applied Phycology, 28, 967-978. Renaud, S. M., Thinh, L. V., Lambrinidis, G., & Parry, D. L.
(2002). Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture, 211(1), 195-214.
Riaño, B., Molinuevo, B., & García-González, M. C. (2011). Treatment of fish processing wastewater with microalgae-containing microbiota. Bioresource
Technology, 102, 10829 -10833. Rippka, R., Deruelles, J., Waterbury, J. B., Herdman, M.,
& Stanier, R. Y. (1979). Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Journal of General
Microbiology, 111, 1-61. Ruiz-Marin, A., Mendoza-Espinosa, L. G., & Stephenson,
T. (2010). Growth and nutrient removal in free and immobilized green algae in batch and semi-continuous cultures treating real wastewater. Bioresource Technology, 101(1), 58-64.
Sabeti, M. B., Hejazi, M. A., & Karimi, A. (2019). Enhanced removal of nitrate and phosphate from wastewater by Chlorella vulgaris: Multi-objective optimization and CFD simulation. Chinese Journal of Chemical
Engineering, 27(3), 639-648. Sayadi, M. H., Ahmadpour, N., Capoorchali, M. F., &
Rezaei M. R. (2016). Removal of nitrate and phosphate from aqueous solutions by microalgae:

56 ว.มทรส. 8(1) : 39-56 (2563)
An experimental study. Global Journal of
Environmental Science and Management, 2(3), 257-264.
Shi, J., Podola, B., & Melkonian, M. (2007). Removal of nitrogen and phosphorus from wastewater using microalgae immobilized on twin layers: an experimental study. Journal of Applied Phycology,
19(5), 417-423. Spolaore, P., Joannis-Cassan, C., Duran, E., & Isambert, A.
(2006). Commercial applications of microalgae. Journal of Bioscience and Bioengineering, 101(2), 87-96.
Taikhao, S., & Phunpruch S. (2017). Biomass and biohydrogen production of unicellular green alga Chlorella vulgaris var. vulgaris TISTR 8261 in Frozen Food Industrial. In The 7
th International
Conference on Fermentation Technology for
Value Added Agricultural Products and The
12th Axian Biohydrogen & Biorefinery
Symposium (pp. 18-25). Khon Kaen: Khon Kaen University.
Taziki, M., Ahmadzadeh, H., Murry, M. A., & Lyon, S. R. (2015). Nitrate and nitrite removal from wastewater using algae. Current Biotechnology, 4, 1-15.
Walker, W. W., Jr. (1983). Significance of eutrophication in water supply reservoir. Journal of American
Water Work Association, 75(1), 38-42.



511 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
4ST-P06: ศกษาการปนเปอนแบคทเรยและการดอตอยาปฏชวนะของแบคทเรย ทคดแยกจากอาหารพรอมบรโภคในเขตอ าเภอพระนครศรอยธยา
The Study of Bacterial Contamination and Antibiotics Resistance in Bacteria isolated from Ready-to-Eat Foods in Phra Nakhon Si Ayutthaya district
กญญา กอแกว1* รฐธญา มาล1 และ ดวงหทย รตนสจธรรม1
Kanya Kokaew1*, Rattiya Malee1 and Dounghatai Rattanasatchatham1
บทคดยอ การศกษาคณภาพดานจลชววทยาของอาหารพรอมบรโภคทจ าหนายในเขตอ าเภอพระนครศรอยธยา โดยการเกบตวอยางอาหาร
จ านวน 20 ตวอยาง แบงเปน อาหารทผานความรอนมาก และอาหารทผานความรอนนอยหรอไมผานความรอน พบวามตวอยางอาหารไมผานเกณฑคณภาพทางจลชววทยาจ านวน 13 ตวอยาง คดเปนรอยละ 65 โดยพบวาอาหารประเภททผานความรอนนอยหรอไมผานความรอน มสดสวนไมผานเกณฑคณภาพทางจลชววทยามากกวาอาหารประเภททผานความรอนมาก ตวอยางอาหารทไมเปนไปตามเกณฑคณภาพดานจลชววทยาไดผลการตรวจสอบคณภาพ โดยพบจลนทรยทงหมด Escherichia coli Staphylococcus aureus และ Salmonella spp. ในตวอยางคดเปนรอยละ 50(10/20), 25(5/20), 20(4/20) และ 5(1/20) ตามล าดบ และจากการทดสอบความสามารถในการดอตอยาปฏชวนะของแบคทเรยทคดแยกจากอาหารพรอมบรโภค ประกอบดวย E. coli จ านวน 5 ไอโซเลท S. aureus จ านวน 4 ไอโซเลท และ Salmonella spp. จ านวน 1 ไอโซเลท มาทดสอบการตานทานตอยาปฏชวนะ 3 ชนด คอ Ampicillin, Ciprofloxain และ Norfloxacin พบวา เชอทกไอโซเลทไวตอยาปฏชวนะทง 3 ชนด ค าส าคญ: อาหารพรอมบรโภค แบคทเรยกอโรคในอาหาร แบคทเรยดอยา
Abstract The study of microbiological quality of ready-to-eat foods in Phra Nakhon Si Ayutthaya district. Two categories of
20 food samples were collected; 1) food using high heat and 2) food using low heat or not heat. The results showed that ready-to-eat foods 13 samples (65%) were not unacceptable according to microbiological standard of foods. The ratio of unacceptable microbiological quality in food using low heat or not heat was higher than food using high heat. The samples that not unacceptable according to microbiological quality criteria showed the total microbial count, Escherichia coli, Staphylococcus aureus and Salmonella spp. were found for 50%(10/20), 25%(5/20), 20%(4/20) and 5%(1/20) respectively. Investigation of antibiotics resistance in bacteria isolated from ready-to-eat foods, E. coli 5 isolates, S. aureus 4 isolates and Salmonella spp. 1 isolate were assessed for antimicrobial-susceptibility testing among three antibiotics including ampicillin, ciprofloxain and norfloxacin. Results suggested that all of isolates were susceptible to all three antibiotics. Keywords: Ready to eat-food, Foodborne pathogen, Antibiotic resistance
1 สาขาวชาวทยาศาสตร คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม 60 หม 3 ต.หนตรา พระนครศรอยธยา 13000 1 Science division, Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi, 60 Moo 3 Huntra district, Phra Nakhon Si Ayutthaya, 13000 Thailand. * Corresponding author. E-mail: [email protected]

512 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
บทน า ปจจบนประชากรสวนใหญในอ าเภอพระนครศรอยธยานยมเลอกรบประทานอาหารพรอมบรโภค (ready to
eat) มากขน เนองจากสะดวกหาซองาย มกจ าหนายในรปแบบอาหารรมถนน (street food) หรอจ าหนายในตลาดนดหรอในศนยอาหาร นอกจากนยงมราคาไมแพง แตการเลอกบรโภคอาหารประเภทนหากผบรโภคไมค านงถงหลกการเลอกบรโภคอาหารทถกสขลกษณะอาจเปนสาเหตของการท าใหเกดโรคอาหารเปนพษได จากรายงานส านกระบาดวทยา กรมควบคมโรค กระทรวงสาธารณสข ตงแตวนท 1 มกราคม พ.ศ.2560 ถง 19 มถนายน พ.ศ.2560 พบผ ปวยจากโรคอาหารเปนพษจ านวน 53,968 ราย จาก 77 จงหวด โดยพบผ ปวยทมสาเหตจากเชอ Staphylococcus จ านวน 98 ราย เชอ Salmonella spp. จ านวน 31 ราย และไมทราบเชอสาเหตอกประมาณ 50,000 ราย ซงในอาหารพรอมบรโภคทมจ าหนายในแถบภาคกลางตามตลาด และรานคาทวไปมรายงานพบการปนเปอนของเชอจลนทรยเกนมาตรฐาน โดยเฉพาะ Escherichia coli ซงเปนจลนทรยบงชสขลกษณะการผลตอาหาร และแบคทเรยทท าใหเกดโรค ไดแก Staphylococcus aureus เปนเชอกอโรคทมกพบปนเปอนในอาหารพรอมบรโภคมาก
ทสด และ Salmonella spp. (กมลวรรณ กนแตง, อจฉรา อยคง, และรชฎาพร สวรรณรตน, 2558; Ananchaipattana et al., 2016) ปจจบนกลมแบคทเรยทปนเปอนในอาหารพรอมบรโภคดอตอยาปฏชวนะเพมมากขน เชน S. aureus มความสามารถดอตอยากลม methicillin, clindamycin, ampicillin, chloramphenicole, Clocxacillin เชอ E. coli ดอยา ampicillin, chloramphenicole, norfloxacin สวนแบคทเรย Salmonella sp. ดอตอยา ampicillin และ chloramphenicol (Bunnueang et al., 2015; Eromo et al., 2016) หากแบคทเรยดอยาเหลานปนเปอนมากบอาหารพรอมบรโภคอาจสงผลตอการรกษาโรคทเกดจากอาหารเปนพษไมไดผล โดยเฉพาะแบคทเรยทสามารถดอตอยาปฏชวนะในกลมทโรงพยาบาลมกจายใหผ ปวย และหากเปนแบคทเรยสายพนธทสามารถดอตอยาปฏช วนะไดหลายชนดอาจสงผลใหไมสามารถใชยาใดๆ รกษาโรคทเกดจากแบคทเรยเหลานนได จากสถานการณดอยาของแบคทเรยกอโรคเปนปญหาทหนวยงานทวโลกตองเฝาระวงแบคทเรยกลมดงกลาวอยางอยางเครงครด
ดงนนเพอเปนขอมลดานความปลอดภยและประกอบการตดสนใจในการเลอกซออาหารพรอมบรโภค การศกษาครงนมวตถประสงคเพอตรวจสอบการปนเปอนของจลนทรยในอาหารพรอมบรโภคในเขตอ าเภอพระนครศรอยธยาทมกระบวนการปรงอาหาร โดยแบงตามลกษณะการสมผสความรอน ไดแก อาหารทผานความรอน และอาหารทผานความรอนนอยหรอไมผานความรอน และแนวโนมในการดอยาปฏชวนะของแบคทเรยกอโรคทคดแยกไดจากตวอยางอาหาร
วธการศกษา
การเกบตวอยาง สมเกบตวอยางอาหารพรอมบรโภคแบงออกเปน 2 ประเภท ประกอบดวย อาหารทผานความรอน จ านวน 7
ตวอยาง ไดแก แกงไขปลา แกงเปรอะ ตมจดแฟงตมจดเตาห ตมพะโล ลกชนทอด ปลาโอทอด และอาหารทผานความรอนนอย หรอไมผานความรอน จ านวน 13 ตวอยาง ไดแก ย าหมยอ ผดมกกะโรน ซช ลาบอสาน สลดโรล ผดหม ผดผก ไกยาง ตบยาง น าตก หมปงสมต า ไสกรอก รวมจ านวน 20 ตวอยาง ทจ าหนายบรเวณตลาดนดในเขตอ าเภอพระนครศรอยธยา เกบตวอยางระหวางเดอนพฤษภาคม พ.ศ.2560 – กมภาพนธ พ.ศ.2561 โดยเกบตวอยางอาหารพรอมบรโภคใสในถงรอนปราศจากเชอ ซงในระหวางการเดนทางตองเกบตวอยางไวในกลองน าแขงตลอด

513 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
ระหวางเกบตวอยาง เมอถงหองปฏบตการใหเกบตวอยางไวทอณหภม 4 องศาเซลเซยส จนกวาท าการทดสอบ จากนนท าการทดสอบตวอยางอาหารภายใน 24 ชวโมง หลงจากการเกบตวอยาง การตรวจวเคราะหคณภาพจลนทรย
การตรวจวเคราะหคณภาพทางจลชววทยาของอาหารพรอมบรโภค ไดแก การตรวจนบจลนทรยทงหมด จ านวนโคลฟอรมและ E. coli ปรมาณเชอ S. aureus และการปนเปอนเชอ Salmonella spp. ดงน
1) การตรวจนบปรมาณจลนทรยทงหมด ตามวธมาตรฐานของ BAM โดยท าการชงตวอยางอาหาร 25 กรม ใสในถงพลาสตกทปราศจากเชอ เตมน ากลน ปรมาตร 225 มลลลตร ผสมใหเปนเนอเดยวกนดวยเครองตปนอาหาร เจอจางตวอยางทระดบ 10-1-10-5 วเคราะหจลนทรยในตวอยาง โดยใชวธเทเพลท (pour plate method) บนอาหาร PCA น าไปบมทอณหภม 37 องศาเซลเซยส เปนเวลา 24-48 ชวโมง ตรวจนบจ านวนและค านวณปรมาณจลนทรยในหนวยโคโลนตอกรม (CFU/g)
2) การตรวจเชอ S. aureus ตามวธมาตรฐานของ BAM โดยชงตวอยางอาหาร 25 กรม ใสในถงพลาสตกปราศจากเชอ เตมน ากลน ปรมาตร 225 มลลลตร ผสมใหเปนเนอเดยวกน เจอจางตวอยางทระดบ 10-1-10-3 เพาะเลยงโดยวธ spread plate ลงในอาหาร Braid-Paker Egg yolk tellurite agar บมทอณหภม 35 องศาเซลเซยส นาน 24-48 ชวโมง สงเกตโคโลนเฉพาะของ S. aureus บนอาหาร โคโลนมสด า สรางตะกอนจากการยอยไขแดง และลอมรอบดวยวงใส คดเลอกโคโลนเพอน าไปทดสอบคณสมบตทางชวเคม และทดสอบการดอยาปฏชวนะตอไป
3) การตรวจนบปรมาณ Total Coliform และ E. coli ตามวธมาตรฐานของ BAM โดยชงตวอยางอาหาร 25 กรม ใสในถงพลาสตกทปราศจากเชอ เตมสารละลาย Phosphate buffer solution (PBS) ปรมาตร 225 มลลลตร ผสมใหเปนเนอเดยวกน เจอจางตวอยางทระดบ 10-1-10-3 วเคราะหโดยวธ MPN (ระบบ 3 หลอด) ในอาหารเหลว LST ทบรรจหลอดดกแกสความเจอจางละ 3 หลอด บมทอณหภม 35 องศาเซลเซยส เปนเวลา 24-48 ชวโมง สงเกตหลอดทใหผลบวกมความขนและเกดแกส น าไปตรวจสอบขนยนยน โดยถายเชอถายเชอทใหผลบวกใสในหลอดอาหารเหลว BGLB (ส าหรบตรวจวเคราะหโคลฟอรม) บมท 35 องศาเซลเซยส และในอาหารเหลว EC (ส าหรบตรวจวเคราะห E. coli) บมท 42 องศาเซลเซยส เปนเวลา 24-48 ชวโมง สงเกตผลการทดสอบเปนบวกตรวจพบความขนและเกดแกสในหลอดดกแกส ผลลบไมเกดแกส วเคราะหปรมาณ Coliform และ E. coli โดยเทยบคากบตาราง MPN และยนยนผล E. coli ดวยการทดสอบทางชวเคม
4) การตรวจเชอ Salmonella spp. ตามวธมาตรฐานของ ISO 6579 โดยชงตวอยางอาหาร 25 กรม ใสในถงพลาสตกทปราศจากเชอ เตมสารละลาย Peptone Water (BPW) ปรมาตร 225 มลลลตร ผสมใหเปนเนอเดยวกน บมท 37 องศาเซลเซยส เปนเวลา 24 ชวโมง ตรวจสอบเชอ Salmonella spp. โดยใชอาหารคดเลอก คอ Salmonella-Shigella (SS) agar และ Xylose Lysine Deoxycholate (XLD) Agar คดเลอกโคโลนเฉพาะของเชอทมลกษณะโคโลนใส มสด าตรงกลางจากการสรางกาซ H2S ท าใหเชอบรสทธแลวน าไปทดสอบทางชวเคม
5) แปรผลการตรวจวเคราะหคณภาพทางจลนทรยของตวอยางเปรยบเทยบกบเกณฑมาตรฐานอาหารพรอมบรโภคทวไปของกรมวทยาศาสตรการแพทย กระทรวงสาธารณสข พ.ศ.2560 (ตารางท 1)

514 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
ตารางท 1 Microbiological guidelines for ready to eat foods in general items, Department of medical science, Ministry of public health, 2017
Microbial criteria Standard of quality
Total viable count CFU/g 1 x 106
E. coli MPN/g 3
E. coli MPN/g* 10
S. aureus CFU/g 100
Salmonella spp./ 25 g Not detected * Food itemes; papaya slad (som-tum), spicy salad (Yum, Larb, slide grilled pork salad or beef)
การศกษาครงนวธตรวจวเคราะหจลนทรยในตวอยางอาหารจะใชวธการวเคราะหแบบดงเดม (conventional
method) ซงอาจมการวเคราะหหลายขนตอนและใชระยะเวลานานประมาณ 2-5 วน โดยในปจจบนนนมวธการตรวจวเคราะหแบบรวดเรว (Rapid method) ทสามารถตรวจหาเชอในอาหารโดยมขนตอนไมยงยากและใชเวลาในการวเคราะหไมเกน 1-2 วน เชน การตรวจโดยใชอาหารส าเรจรปในรปแบบทเคลอบบนแผนฟลม (Petrifilm) และการตรวจวเคราะหปรมาณชนสวนดเอนเอในหลอดทดลอง (Polymerase chain reaction, PCR assay) เปนตน ทดสอบการดอยาปฏชวนะ
คดเลอกแบคทเรย ไดแก E. coli, S. aureus และ Salmonella spp. ทคดแยกไดจากตวอยางอาหาร น ามาทดสอบการดอยาปฏชวนะ โดยเพาะเลยงเชอในอาหารเหลว TSB บมท 37 องศาเซลเซยส 24 ชวโมง เตรยมเชอเรมตนใหมความขนเทากบ 0.5 McFarland standard จากนน swab เชอลงบนอาหาร Mueller-Hinton agar วางแผนยาปฏชวนะ ไดแก แอมพซลน (ampicillin) 10 ไมโครกรม ซโพรฟลอกซาซน (ciprofloxacin) 5 ไมโครกรม และ นอรฟลอกซาซน (norfloxacin) 5 ไมโครกรม บนจานอาหารเลยงเชอ บมทอณหภม 37 องศาเซลเซยส นาน 18 ชวโมง (ดดแปลงจากวธของกฤษณา เลกยวง, สทธดา เนองกลยา, วรพรรณ เผาทองศข, และ รงรตน นลธเสน, 2560) ตรวจผลโดยสงเกตบรเวณทถกยบยง (Inhibition zone) วเคราะหการดอยาเปรยบเทยบกบตารางของ Clinical and Laboratory Standards Institute (CLSI) ซงแสดงผลออกมา 3 ลกษณะ ไดแก Susceptible (S) เชอมความไวตอสารตานแบคทเรย Intermediate (I) เชอมความไวปานกลางตอเชอทใชทดสอบ และ Resistant (R) เชอดอตอสารทใชทดสอบ ใชเชอมาตรฐาน E. coli ATCC25922 และ S. aureus ATCC29213 เปนชดควบคม
ผลการศกษาและอภปรายผล ผลการตรวจวเคราะหจลนทรยทงหมดในตวอยางอาหารพรอมบรโภคทง 2 ประเภท พบจลนทรยทงหมดอย
ในชวงระหวาง 5.70 x 103- 2.93 x 106 CFU/g เมอวเคราะหผลการทดสอบเปรยบเทยบกบเกณฑมาตรฐานของกรมวทยาศาสตรการแพทยอาหารพรอมควรมจลนทรยทงหมดไมเกน 106 CFU/g ผลการศกษาพบวาตวอยางอาหารทไมผานเกณฑจลนทรยทงหมดม 10 ตวอยาง ประกอบดวย อาหารประเภททผานความรอนมากจ านวน 3 ตวอยาง ไดแก แกงไขปลา แกงเปรอะ และลกชนทอด คดเปนรอยละ 4.8 จากตวอยางอาหารประเภทผานความรอนทงหมด ม

515 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
จลนทรยระหวาง 2.38 - 2.48 x 106 CFU/g และตวอยางอาหารประเภททผานความรอนนอยหรอไมผานความรอนจ านวน 7 ตวอยาง ไดแก ลาบอสาน สลดโรล ผดหม ผดผก ตบยาง น าตก และสมต า คดเปนรอยละ 53.8 จากตวอยางอาหารประเภทผานความรอนหรอไมผานความรอนทงหมด (table 2) มจลนทรยระหวาง 2.65 x 106 CFU/g จนถงปรมาณมากกวา 106 CFU/g ซงตวอยางสวนใหญมจลนทรยทงหมดเกนคาเกณฑมาตรฐานเพยงเลกนอย ยกเวนลาบอสานพบมการปนเปอนจลนทรยในปรมาณสงทสดเทากบ 1 x 107 CFU/g
การตรวจวเคราะหแบคทเรยดชน พบวาตวอยางอาหารมโคลฟอรมปนเปอนในชวงตงแต 3.6 จนถงมคามากกวา 1100 MPN/g จากการตรวจวเคราะหตวอยางอาหารบางชนดมการปนเปอนโคลฟอรมในปรมาณคอนขางมาก ไดแก แกงไขปลา แกงเปรอะ ตมจดเแฟง ซช ลาบหม เปนตน ซงโคลฟอรมแบคทเรยเปนจลนทรยดชนทแสดงถงสขลกษณะในการปรงอาหาร เชอกลมนพบไดทวไปในธรรมชาต มอและทางเดนอาหารของมนษยหรอสตว ใชวดความปลอดภยของอาหารได เนองจากมความสมพนธกบเชอกอโรคอาหารเปนพษ มกพบเชอจลนทรยนเสมอเมอตรวจพบจลนทรยกอโรค (บญศร จงเสรจตต, 2561) ดงนนหากมโคลฟอรมปนเปอนในอาหารมากเกนไปบงชถงกระบวนการปรงอาหารไมถกสขลกษณะ และยงมโอกาสปนเปอนเชอกอโรคไดดวย โคลฟอรมทใชเปนดชนการปนเปอนเชอในล าไสและอจจาระ คอเชอ E. coli จากผลการศกษาพบลกษณะของเชอทคาดวาเปน E. coli ปนเปอนในตวอยางอาหารเกนเกณฑมาตรฐาน จ านวน 5 ตวอยาง คดเปนรอยละ 25.0 (ตารางท 2) โดยพบในอาหารทผานความรอนมาก 2 ตวอยาง คดเปนรอยละ 28.5 ไดแก แกงเปรอะ ตมจดเตาห อาหารทผานความรอนนอยหรอไมผานความรอน 3 ตวอยาง คดเปนรอยละ 23.1 ไดแก ลาบอสาน ตบยาง และน าตก โดยเฉพาะตวอยางอาหาร ไดแก แกงเปรอะ ลาบอสาน และน าตก พบมการปนเปอน E. coli ในปรมาณมากกวา 1100 MPN/g ซงจากรายงานผลส ารวจอาหารพรอมบรโภคทจ าหนายในศนยอาหารและโรงอาหาร พบเชอ E. coli ในอาหารประเภทลาบ น าตก เกนเกณฑมาตรฐานเชนเดยวกน นอกจากนมกพบการปนเปอน E. coli ในอาหารประเภททผานความรอนนอย เชน ขาวมนไก ขาวหมแดงหมกรอบ ย า สมต า เปนตน (กมลวรรณ กนแตง, อจฉรา อยคง, และรชฎาพร สวรรณรตน, 2558; บญเลยง สพมพ, ปยะพงษ ชมศร, และอรทย ปารเพชร, 2560)
ผลการตรวจเชอแบคทเรยกอโรคอาหารเปนพษ พบ S. aureus เกนเกณฑมาตรฐานมจ านวน 4 ตวอยาง ไดแก ย าหมยอ ซช สลดโรล และผดผก จดอยในอาหารประเภททผานความรอนนอยหรอไมผานความรอน คดเปนรอยละ 25.0 จากตวอยางทงหมด ซงคลายคลงกบผลการศกษาการปนเปอนเชอ S. aureus ในอาหารพรอมบรโภคของ Shafizi et al. (2016) ทพบการปนเปอนเชอในอาหารดบหรอไมผานความรอนมากกวากลมอาหารปรงสก ซงมกเกดจากสขลกษณะในการประกอบอาหารทไมเหมาะสม เนองจาก S. aureus เปนเชอมกปนเปอนมากบผปรงอาหาร โดยเฉพาะมอของผปรงอาหาร อาหารทมการใชมอจบตกแตง เชน ซชจงมโอกาสปนเปอนเชอนได ซงโดยปกตเซลลจะถกท าลายดวยความรอนในระดบพลาสเจอรไรส แตไมเพยงพอตอการท าลายสารพษเอนเทอโรทอกซน (staphylococcal enterotoxins) ทเชอสราง ดงนนการประกอบอาหารทใชความรอนต าอาจท าใหเชอนรอดชวตได แตบางกรณการปนเปอนอาจเกดหลงจากการปรงสกแลวผปรงใชมอหยบจบอาหารหรอภาชนะทปนเปอนเชอ หากเชอทสรางสารพษปนเปอนในอาหารสงผลใหเกดอาการอาหารเปนพษ เชน ทองเสย มไข อาเจยน ได (บญศร จงเสรจตต, 2561) สวนผลการตรวจ Salmonella spp. พบปนเปอนจ านวน 1 ตวอยาง คอ น าตกหม ซงเปนอาหารประเภททผานความรอนนอย หรอไมผานความรอน คดเปนรอยละ 5.0 จากตวอยางทงหมด (ตารางท 2) โดยตามเกณฑมาตรฐานอาหารพรอมบรโภคก าหนดใหตองไมพบการปนเปอนของเชอ Salmonella spp. ตอตวอยางอาหาร 25 กรม เชอ Salmonella มกพบปนเปอนในอาหารทมเนอสตวเปนองคประกอบโดยเฉพาะเนอหมและเนอไก สาเหตทพบการปนเปอนเชอในตวอยางน าตกหมอาจเกดจากวตถดบโดยเฉพาะเนอหม เครองใน มการปนเปอนเชอ และขนตอนการ

516 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
ปรงอาหารโดยความรอนทใชไมเพยงพอในการท าลายเชอ เมอวเคราะหคณภาพทางจลชววทยาของอาหารพรอมบรโภคทจ าหนายในเขตอ าเภอพระนครศรอยธยา ทงหมดจ านวน 20 ตวอยาง มตวอยางผานทกเกณฑมาตรฐานทศกษา จ านวน 7 ตวอยาง คดเปนรอยละ 35.0 ไดแก ตมจดแฟง ตมพะโล ผดมกกะโรน ไกยาง ปลาโอทอด หมปง และไสกรอก และตวอยางทไมผานเกณฑมาตรฐาน จ านวน 13 ตวอยาง คดเปนรอยละ 65.0 เนองจากพบปรมาณจลนทรยเกนเกณฑมาตรฐานใดมาตรฐานหนง โดยพบตวอยางทไมผานเกณฑมาตรฐานมากทสด 3 เกณฑมาตรฐาน คอ น าตกหม โดยไมผานเกณฑดานจ านวนจลนทรยทงหมด E. coli และ Salmonella spp. ตวอยางลาบหม สลดโรล ผดหม ตบยาง และแกงเปรอะ ไมผาน 2 เกณฑมาตรฐาน และตวอยางสมต าไมผานเกณฑ 1 มาตรฐาน หากวเคราะหโดยภาพรวมตวอยางอาหารประเภทผานความรอนนอยหรอไมผานความรอนมแนวโนมไมผานเกณฑดานจลชววทยามากกวาอาหารทผานความรอนมาก ทงนอาจชใหเหนวากระบวนการปรงสกสามารถลดความเสยงในการปนเปอนเชอจลนทรยไดด ผบรโภคจงควรเลอกซออาหารพรอมบรโภคทผานกระบวนการปรงสกแลว อยางไรกตามแมวากลมอาหารพรอมบรโภคทผานกระบวนการปรงสกโดยใชความรอนต าหรอไมผานการปรงสกมโอกาสทจลนทรยจะรอดชวตในอาหารได แตผลการศกษาครงนพบวาตวอยางสมต าทเปนอาหารไมผานกระบวนการปรงสกกลบพบเฉพาะเชอจลนทรยเกนปรมาณทก าหนดแตไมพบปนเปอน E. coli และเชอกอโรค แสดงใหเหนวากลมอาหารดบพรอมบรโภคทไมไดผานการปรงสก เชน สมต า สลด หากเลอกใชวตถดบทสะอาดและผปรงอาหารมสขลกษณะทดท าใหอาหารนนผานเกณฑมาตรฐานจลนทรยและมความปลอดภยตอผบรโภคได ปจจยทมผลตอการปนเปอนเชอจลนทรยในอาหารพรอมบรโภค หรออาหารทจ าหนายรมถนน เชน การจดการฟารมไมดสงผลตอคณภาพของวตถดบได ซงวตถดบจ าพวกผกสด พบวาผกออรแกนคหรอผกปลกแบบอนทรยมโอกาสปนเปอนเชอ E. coli มากกวาผกทไมไดปลกแบบอนทรยเนองจากการใชป ยคอกทมการปนเปอนเชอจลนทรยประจ าถนในล าไสของมนษยและสตว (Szczech et al., 2018) การเกบรกษาวตถดบเครองปรงในการประกอบอาหารไมด โดยเฉพาะอาหารจ าพวกแปงหรอผกผลไมอบแหงอาจปนเปอนเชอรา และ Bacillus cereus ได กระบวนการปรงอาหารภาชนะตองสะอาด หรออณหภมทใชประกอบอาหารตองเหมาะสมทสามารถท าลายเซลลจลนทรยได วธการเกบรกษาอาหารระหวางการจ าหนายไมใหมการปนเปอน ซงอาหารพรอมบรโภคควรเกบทอณหภม 37-54 องศาเซลเซยส ไมเกน 4 ชวโมง เพราะอณหภมในชวงนเหมาะสมตอการเจรญของจลนทรย และปจจยทส าคญคอกระบวนการผลตทถกสขลกษณะซงตองเรมจากการท าความเขาใจตอผปรงอาหารใหเขาใจถงกระบวนการผลตอาหารทถกหลกอนามยซงจะสงผลใหมความปลอดภยตอผบรโภค (Alimi, 2016; Ananchaipattana et al., 2016)
ตารางท 2 Microbial quality of ready to eat foods in Phra Nakhon Si Ayutthaya district, Two categories food samples; food using high heat and food using low heat or not heat
Category of food Bacteria Number of positive*/
Total samples Ratio (%)
food using high heat
Total viable count 3/7 42.8
E. coli 2/7 28.5
S. aureus 0/7 0
Salmonella spp. 0/7 0
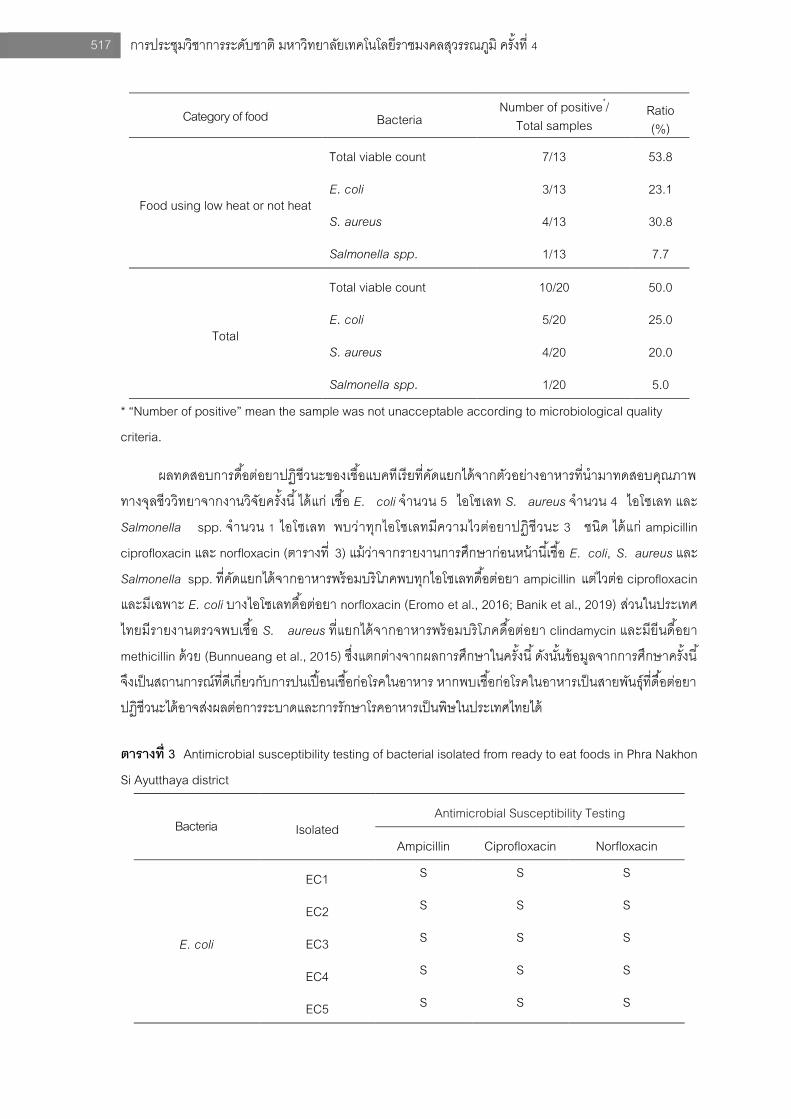
517 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
Category of food Bacteria Number of positive*/
Total samples Ratio (%)
Food using low heat or not heat
Total viable count 7/13 53.8
E. coli 3/13 23.1
S. aureus 4/13 30.8
Salmonella spp. 1/13 7.7
Total
Total viable count 10/20 50.0
E. coli 5/20 25.0
S. aureus 4/20 20.0
Salmonella spp. 1/20 5.0 * “Number of positive” mean the sample was not unacceptable according to microbiological quality criteria.
ผลทดสอบการดอตอยาปฏชวนะของเชอแบคทเรยทคดแยกไดจากตวอยางอาหารทน ามาทดสอบคณภาพทางจลชววทยาจากงานวจยครงน ไดแก เชอ E. coli จ านวน 5 ไอโซเลท S. aureus จ านวน 4 ไอโซเลท และ Salmonella spp. จ านวน 1 ไอโซเลท พบวาทกไอโซเลทมความไวตอยาปฏชวนะ 3 ชนด ไดแก ampicillin ciprofloxacin และ norfloxacin (ตารางท 3) แมวาจากรายงานการศกษากอนหนานเชอ E. coli, S. aureus และ Salmonella spp. ทคดแยกไดจากอาหารพรอมบรโภคพบทกไอโซเลทดอตอยา ampicillin แตไวตอ ciprofloxacin และมเฉพาะ E. coli บางไอโซเลทดอตอยา norfloxacin (Eromo et al., 2016; Banik et al., 2019) สวนในประเทศไทยมรายงานตรวจพบเชอ S. aureus ทแยกไดจากอาหารพรอมบรโภคดอตอยา clindamycin และมยนดอยา methicillin ดวย (Bunnueang et al., 2015) ซงแตกตางจากผลการศกษาในครงน ดงนนขอมลจากการศกษาครงน จงเปนสถานการณทดเกยวกบการปนเปอนเชอกอโรคในอาหาร หากพบเชอกอโรคในอาหารเปนสายพนธทดอตอยาปฏชวนะไดอาจสงผลตอการระบาดและการรกษาโรคอาหารเปนพษในประเทศไทยได
ตารางท 3 Antimicrobial susceptibility testing of bacterial isolated from ready to eat foods in Phra Nakhon Si Ayutthaya district
Bacteria Isolated Antimicrobial Susceptibility Testing
Ampicillin Ciprofloxacin Norfloxacin
E. coli
EC1 S S S
EC2 S S S
EC3 S S S
EC4 S S S
EC5 S S S

518 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
Bacteria Isolated Antimicrobial Susceptibility Testing
Ampicillin Ciprofloxacin Norfloxacin
S. aureus
SA1 S S S
SA2 S S S
SA3 S S S
SA4 S S S
Salmonella sp. SM1 S S S
“S” mean susceptible or sensitive.
สรป จากการสมตวอยางอาหารพรอมบรโภคทจ าหนายในเขตอ าเภอพระนครศรอยธยา จ านวน 20 ตวอยาง แบง
ตามลกษณะการผานความรอน 2 ประเภท ไดแก 1) อาหารทผานความรอนมาก และ 2) อาหารทผานความรอนนอยหรอไมผานความรอน ตรวจวเคราะหจลนทรย โดยเปรยบเทยบกบมาตรฐานทางจลชววทยาของอาหารและภาชนะสมผสอาหารเกณฑกรมวทยาศาสตรการแพทย ฉบบท 3 พ.ศ. 2560 พบตวอยางทปรมาณจลนทรยทงหมดเกนคามาตรฐาน จ านวน 10 ตวอยาง คดเปนรอยละ 50.0 พบเชอ E. coli เกนเกณฑมาตรฐาน จ านวน 5 ตวอยาง คดเปนรอยละ 25.0 ตวอยางอาหารทพบการปนเปอนของเชอกอโรคอาหารเปนพษ S. aureus และ Salmonella spp. เกนคามาตรฐาน จ านวน 4 ตวอยาง คดเปนรอยละ 20.0 และจ านวน 1 ตวอยาง คดเปนรอยละ 5.0 ของตวอยางทงหมดตามล าดบ เมอวเคราะหคณภาพอาหารทางจลชววทยาของตวอยางอาหารพรอมบรโภคทงหมด 20 ตวอยาง พบไมผานเกณฑมาตรฐาน จ านวน 13 ตวอยาง คดเปนรอยละ 65.0 โดยพบอาหารประเภททผานความรอนนอยหรอไมผานความรอน มสดสวนไมผานเกณฑคณภาพทางจลชววทยามากกวาอาหารประเภททผานความรอนมาก การทดสอบความสามารถในการดอตอยาปฏชวนะของแบคทเรยทคดแยกจากอาหารพรอมบรโภค ประกอบดวย E. coli จ านวน 5 ไอโซเลท เชอ S. aureus จ านวน 4 โซเลท และเชอ Salmonella spp. จ านวน 1 ไอโซเลท รวมทงหมด 10 ไอโซเลท ทดสอบโดยใชยาปฏชวนะ 3 ชนด คอ Ampicilin, Ciprofloxain และ Norfloxacin พบวาเชอทกไอโซเลทไวตอยาปฏชวนะทง 3 ชนด
การเลอกรบประทานอาหารพรอมบรโภคทจ าหนายในตลาดนดโดยเฉพาะแบบเปดโลง (open market) นน ผ บรโภคควรค านงถงสขลกษณะของการปรงอาหาร ควรเลอกอาหารทปรงสกใหม ใชวตถทสะอาด รวมถงสขลกษณะของพนทในการจ าหนายอาหารดวย เพอเปนทางเลอกในการบรโภคอาหารปลอดภยลดความเสยงในการเกดโรคอาหารเปนพษได
ค าขอบคณ ขอขอบคณสถาบนวจยและพฒนา มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ทไดมอบทนสนบสนนส าหรบ
การท าวจยครงน

519 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
เอกสารอางอง กฤษณา เลกยวง, สทธดา เนองกลยา, วรพรรณ เผาทองศข, และ รงรตน นลธเสน. (2560). การศกษาความไวตอยาปฏชวนะของเชอ
Salmonella spp. และ non-Salmonella ทแยกไดจากเนอไกสด. ใน การประชมวชาการระดบชาตวทยาศาสตรและเทคโนโลยระหวางสถาบนครงท 5 (น. 979-983). กรงเทพมหานคร: คณะวทยาศาสตร มหาวทยาลยสยาม.
กมลวรรณ กนแตง, อจฉรา อยคง, และรชฎาพร สวรรณรตน. (2558). ความปลอดภยทางจลชววทยาของอาหารพรอมบรโภคทจ าหนาย ณ สถานขนสงผ โดยสารในเขตกรงเทพมหานคร. วารสารกรมวทยาศาสตรการแพทย. 57(3), 269-278.
กรมวทยาศาสตรการแพทย กระทรวงสาธารณสข. (2560). เกณฑคณภาพทางจลชววทยาของอาหารและภาชนะสมผสอาหาร ฉบบท 3. สบคน 21 กมภาพนธ 2561, จาก http://bqsf.dmsc.moph.go.th/bqsfWeb/wp-content/uploads/2017/Publish/e-book/micro-ISBN60.pdf
บญเลยง สพมพ, ปยะพงษ ชมศร, และอรทย ปารเพชร. (2560). คณภาพดานจลชววทยาของอาหารปรงส าเรจในโรงอาหาร มหาวทยาลยราชภฏเลย. วารสารวทยาศาสตรและเทคโนโลย มหาวทยาลยอบลราชธาน, (ฉบบพเศษ), 72-81.
บญศร จงเสรจตต. (2561). จลชววทยาทางอาหาร. กรงเทพฯ: จฬาลงกรณมหาวทยาลย. ส านกระบาดวทยา กรมควบคมโรค กระทรวง สาธารณสข. (2560). รายงานงานโรคในระบบเฝาระวง “Food Poisoning”. สบคน 25 มถนายน 2560, จาก
http://www.boe.moph.go.th/boedb/surdata/506wk/y60/d03_2460.pdf Alimi, B. A. (2016). Risk factor in street food practices in developing countries: A review. Food Science and Human
Wellnes, 5, 141-148. Ananchaipattana, C., Bari, L. & Inatsu, Y. (2016). Bacterial Contamination into Ready-to-Eat Foods Sold in Middle Thailand.
Biocontrol Science, 21(4): 225-230. Banik, A., Abony, M., Datta, S. and Towhid, S.T. (2019). Microbiological Quality of Ready-to-Eat Food from Dhaka,
Bangladesh. Current Research in Nutrition and Food Science, 7(1), 161-168. Bunnueang, N., Kongpheng, S., Singkhamanan, K., Saengsuwan, P., Rattanachuay, P., Dangsriwan, S., and
Sukhumungoon, P. (2015). Methicillin-resistant Staphylococcus aureus from ready-to-eat foods in a hospital canteen, Southern Thailand: virulence characterization and genetic relationship. The Southeast Asian Journal of Tropical Medicine and Public Health, 46(1), 86-96.
Clinical and Laboratory Standard Institute (CLSI). (2015). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. Retrieved 20 March 2017, from https://www.facm.ucl.ac.be/intranet/CLSI/ CLSI-2015-M100-S25.unlocked.pdf
Eromo T., Tassew, H., Daka, D. and Kibru, G. (2016). Bacteriological Quality of Street Foods and Antimicrobial Resistance of Isolates in Hawassa, Ethiopia. Ethiopian Journal of Health Sciences, 26(6), 533-542.
Feng, P., Weagant, S.D., Grant, M.A. and Burkhardt, W. (2002). BAM 4: Enumeration of Escherichia coli and the Coliform Bacteria. Retrieved 17 March 2017, from https://www.fda.gov/food/laboratory-methods-food/bam-4-enumeration- escherichia-coli-and-coliform-bacteria
International Organization for Standardization (ISO). (2002). ISO 6579: 2002 Microbiology of food and Animal Feeding Stuffs-Horizontal Method for the Detection of Salmonella spp. Retrieved 20 March 2017, from https://www.iso.org/fr/standard/29315.html.
Maturin, L., and Peeler, J.T. (2001). Bacteriological Analytical Manual Chapter 3: Aerobic plate count. Retrieved 17 March 2017, from https://www.fda.gov/Food/FoodScienceResearch/LaboratoryMethods/ucm063346.htm
Shafizi, A.W., Mohammad Ridzuan, M.S., Ubong, A., New, C.Y., Mohhiddin, O., Toh, P.S., Chai, L.C., and Son, R. (2016). Assessing Staphylococcus aureus in ready to eat (RTE) food and risk assessment of food premises in Putrajaya. International Food Research Journal, 23(4), 1761-1766.

520 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
Szczech, M., Kowalska, B., Smolinska, U., Maciorowski, R., Oskiera, M., and Michalska, A. (2018). Microbial quality of organic and conventional vegetables from Polish farms. International Journal of Food Science, 286, 155-161.
Tallent, S., Hait, J., Bennett, R. W., and Lancette, G.A. (2016). Bacteriological Analytical Manual Chapter 12: Staphylococcus aureus. Retrieved 17 March 2017, from https://www.fda.gov/food/laboratory-methods-food/bam-staphylococcus-aureus
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/343923960
Nonomuraea antri sp. nov., an actinomycete isolated from cave soil in Thailand
Article in International Journal of Systematic and Evolutionary Microbiology · August 2020
DOI: 10.1099/ijsem.0.004413
CITATIONS
0READS
75
5 authors, including:
Some of the authors of this publication are also working on these related projects:
Melanogenic actinomycetes from rhizosphere soil — antagonistic activity against Xanthomonas oryzae and plant-growth-promoting traits View project
PLA degradation View project
Kenika Lipun
Kasetsart University
7 PUBLICATIONS 9 CITATIONS
SEE PROFILE
Wee Fei Aaron Teo
Kasetsart University
10 PUBLICATIONS 102 CITATIONS
SEE PROFILE
Paweena Suksaard
Rajamangala University of Technology Suvarnabhumi
11 PUBLICATIONS 42 CITATIONS
SEE PROFILE
Wasu Pathom-aree
Chiang Mai University
93 PUBLICATIONS 1,437 CITATIONS
SEE PROFILE
All content following this page was uploaded by Wee Fei Aaron Teo on 20 October 2020.
The user has requested enhancement of the downloaded file.

365 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
4ST-O09: เอนโดไฟตกแบคทเรยจากรากมนส าปะหลงและความสามารถ ในการสรางสารสงเสรมการเจรญของพช
Endophytic Bacteria from Cassava Root and Their Capacity to Produce Plant Growth Promoting Compounds
ปวณา สขสอาด1* พรรณราย ดอกไมเพง1 และ ดารานย รบเมอง1
Paweena Suksaard1*, Pannarai Dokmaipeng1 and Daranai Robmuang1
บทคดยอ การใชจลนทรยทมประโยชนทางการเกษตรเปนทางเลอกหนงในการลดการใชสารเคม เอนโดไฟตกแบคทเรยเปนจลนทรยท
อาศยในเนอเยอพชและใหประโยชนแกพชโดยไมกอโรค งานวจยนมวตถประสงคเพอศกษาเอนโดไฟตกแบคทเรยทสามารถสรางสารสงเสรมการเจรญของพชจากรากมนส าปะหลง และทนตอสารก าจดวชพชในการเพาะปลกได โดยคดแยกเอนโดไฟตกแบคทเรยจ านวน 30 ไอโซเลท พบแบคทเรยทมประสทธภาพในการละลายฟอสเฟตสงจ านวน 2 ไอโซเลท ไดแก CRP-22 และ CRP-13 มคาดชนการละลายฟอสเฟตท 1.24 และ 1.18 ตามล าดบ แบคทเรยไอโซเลททมความสามารถในการผลตอนโดลอะซตกแอซด ( IAA) สงมจ านวน 4 ไอโซเลท ไดแก CRP-12, CRP-19, CRP-15 และ CRP-29 ซงผลตได 428.7, 410.9, 302.6 และ 226.7 ไมโครกรมตอมลลลตร ตามล าดบ นอกจากนยงพบวาแบคทเรยทง 6 ไอโซเลท สามารถสรางสารสงเสรมการเจรญของพชไดทง 2 ชนดและทนตอสารก าจดวชพชทนยมใชในไรมนส าปะหลงได จากผลการทดลองแสดงใหเหนวามความเปนไปไดทจะน าเอนโดไฟตกแบคทเรยทมประสทธภาพดไปประยกตใชเพอสงเสรมการเจรญของมนส าปะหลงในพนททมการใชสารก าจดวชพชไดในอนาคต ค าส าคญ: การละลายฟอสเฟต สารก าจดวชพช อนโดลอะซตกแอซด เอนโดไฟตกแบคทเรย
Abstract The application of beneficial microbes in agriculture is an alternative way to reduce chemical inputs in agriculture.
Endophytic bacteria are thrive inside plants and improve plant growth without pathogenic effect. This research aims to study endophytic bacteria with plant growth promoting activities isolated from cassava roots which tolerance against herbicides. A total of 30 endophytic bacteria were isolated and two isolates, CRP-22 and CRP-13, are effectively solubilize phosphate and show the highest phosphate solubilization index at 1.24 and 1.18, respectively. There are 4 isolates, CRP-12, CRP-19, CRP-15 and CRP-29, were effectively produce IAA at 428.71, 410.89, 302.64 and 226.73 µg/ml, respectively. Moreover, all these 6 potent isolates are able to produce both of the plant growth promoting compound and resistant to herbicide that commonly used in cassava field, glyphosate and paraquat. These results suggest that the potent endophytic bacteria isolates probably be applied for promoting growth of cassava growing under herbicides accumulated field. Keywords: phosphate solubilization, herbicides, indole acetic acid, endophytic bacteria
1 คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม 60 หม 3 ถ.สายเอเซย ต.หนตรา อ.พระนครศรอยธยา จ.พระนครศรอยธยา 13000 1 Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi 60, Moo 3, Asian Highway Rd., Phranakhon Si Ayutthaya, 13000 * Corresponding author. E-mail: [email protected]

366 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
บทน า มนส าปะหลง (Manihot esculenta L. Crantz.) เปนพชเศรษฐกจส าคญและเปนหนงในสนคาสงออกหลกของ
ประเทศไทย ซงสภาพดนทใชในการเพาะปลกมนส าปะหลงนนมลกษณะดนชนบนเปนดนทราย มธาตอาหารพชตามธรรมชาตต า และยงประสบปญหาเกยวกบโรคทเกดจากแบคทเรย รา ราแปง แมลงศตรพช และวชพช สงผลใหตองมการใชสารเคมในปรมาณมาก เชน ยาฆาหญา ซงในการปลกมนส าปะหลงโดยทวไปมการใชสารก าจดวชพช เพอไมใหแยงธาตอาหารมนส าปะหลง เชน ไกลโฟเซต และพาราควอต ซงมผลแบบไมเลอกท าลายจงมผลตอมนส าปะหลงดวย ท าใหเกดการเนาและผลผลตมปรมาณลดลง สารเคมทใชยงสามารถตกคางในดนและถกดดซมโดยพชปลกได (มณทรา พรหมพทยายทธ, 2553; พวงรตน ขจตวชยานกล, 2556) แบคทเรยกลมสงเสรมการเจรญเตบโตของพช (Plant Growth Promoting Bacteria; PGPB) เปนแบคทเรยทเปนประโยชน สามารถสรางสารควบคมการเจรญเตบโตของพชได พบไดในบรเวณรอบรากพช (rhizosphere) และในเนอเยอพช (endophytic bacteria) ซงเอนโดไฟตกแบคทเรยจะใชชวตทงหมดหรอบางชวงอยในเนอเยอพช และใหประโยชนแกพชอาศยโดยไมท าอนตรายหรอกอใหเกดโรคแกพช (Hurek & Hurek, 1998) จากการศกษาเอนโดไฟตกแบคทเรยในพชหลายชนด พบวามความหลากหลายทางสปชสของเอนโดไฟตกแบคทเรย และสวนใหญมความสามารถในการสงเสรมการเจรญเตบโตของพช เชน การละลายฟอสเฟต การสรางฮอรโมนพช การตรงไนโตรเจน เปนตน ท าใหสารอาหารในดนเปลยนไปอยในรปทเปนประโยชนกบพช และกระตนการเจรญของพชเพมขน นอกจากนยงมการใชเอนโดไฟตกแบคทเรยเพอยบยงจลนทรยสาเหตโรคในมนส าปะหลง และพชอนๆ อกดวย (Nogueira et al., 2001) ในงานวจยนจงศกษาเอนโดไฟตกแบคทเรยในรากมนส าปะหลงทมความสามารถในการสรางสารสงเสรมการเจรญของพช และทนตอสารเคมก าจดวชพช เพอเปนขอมลพนฐานในการเพมโอกาสการน าจลนทรยมาประยกตใชสงเสรมการเจรญของมนส าปะหลงในพนททมการใชสารเคมก าจดวชพชไดในอนาคต
วธการศกษา
การแยกเอนโดไฟตกแบคทเรยทมความสามารถในการละลายฟอสเฟตจากตวอยางรากมนส าปะหลง
สมพนทเกบตวอยางแบบกระจายในพนทต าบลหนองปลาไหล อ าเภอบางละมง จงหวดชลบร เกบตวอยางรากมนส าปะหลงทมความสมบรณไมเปนโรคแปลงละ 5 ตนตอ 1 ตวอยาง โดยเกบจ านวน 5 ตวอยาง น าตวอยางรากมาฆาเชอบรเวณผวรากดวยแอลกอฮอลเขมขน 95 เปอรเซนต เปนเวลา 5 นาท แลวแชในสารละลายโซเดยมไฮโปคลอไรททผสม Tween20 น าไปเขยาทความเรวรอบ 150 รอบตอนาท เปนเวลา 10 นาท น าไปลางดวยน ากลน จากนนน าตวอยางรากไปบดในโกรง ผสมตวอยางกบสารละลายโซเดยมคลอไรดเขมขน 0.85 เปอรเซนต (ดดแปลงจาก Coombs & Franco, 2003) น าของเหลวทสกดไดไปเจอจางดวยสารละลายโซเดยมคลอไรดใหไดระดบความเจอจาง 10-3 ถง 10-5 และเกลยบนอาหาร Pikovskaya’s agar บมทอณหภม 30 องศาเซลเซยส เปนเวลา 3 วน นบจ านวนและคดแยกโคโลนของเอนโดไฟตกแบคทเรยทมวงใสรอบโคโลน (clear zone) และท าใหบรสทธ จากนนศกษาลกษณะเซลลภายใตกลองจลทรรศน โดยการยอมสแบบแกรม (Gram staining) และเกบรกษาไวในสารละลายกลเซอรอล ทอณหภม -20 องศาเซลเซยส
การทดสอบความสามารถในการละลายฟอสเฟต (Phosphate solubilization)
น าเอนโดไฟตกแบคทเรยแตละไอโซเลทไปเลยงในอาหาร nutrient broth บมแบบเขยาทอณหภม 30 องศาเซลเซยส เปนเวลา 24 ชวโมง และน ามาปรบความขนใหมคาการดดกลนแสงเทากบ 0.5 ทความยาวคลนแสง 530

367 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
นาโนเมตร หยดตวอยาง 10 ไมโครลตร ลงบนอาหาร Pikovskaya’s agar บมทอณหภม 30 องศาเซลเซยส เปนเวลา 5 วน สงเกตการเปลยนแปลงรอบโคโลน ถามการละลายฟอสเฟตจะเกดวงใสรอบโคโลน บนทกขนาดโคโลนและโซนใส เพอน าไปค านวณคาดชนการละลายฟอสเฟต ดงสมการตอไปน คาดชนการละลายฟอสเฟต = การทดสอบการผลตอนโดลอะซตกแอซด (Indole acetic acid หรอ IAA)
น าเอนโดไฟตกแบคทเรยทแยกไดไปเลยงในอาหาร nutrient broth ทผสม L-tryptophan เขมขน 0.2 เปอรเซนต บมแบบเขยาทอณหภม 30 องศาเซลเซยส เปนเวลา 48 ชวโมง และน ามาปรบความขนใหมคาการดดกลนแสงเทากบ 0.5 ทความยาวคลนแสง 530 นาโนเมตร น าไปปนเหวยงทความเรวรอบ 8,000 รอบตอนาท เปนเวลา 10 นาท เกบเอาสวนใส 1 มลลลตร ผสมกบ Salkowski’s reagent ปรมาตร 2 มลลลตร ตงไวในทมดเปนเวลา 30 นาท จากนนน ามาวดปรมาณการสราง IAA ดวยการวดคาการดดกลนคลนแสงทความยาวคลน 530 นาโนเมตร (Bric et al., 1991) น าคาทไดมาเทยบกบกราฟมาตรฐานความเขมขนของ IAA การทดสอบความสามารถในการทนตอสารก าจดวชพชเบองตน ทดสอบความสามารถของเอนโดไฟตกแบคทเรยในการทนตอสารก าจดวชพช ไดแก ไกลโฟเซต - ไอโซโพรพลแอมโมเนยม และพาราควอตไดคลอไรดบนอาหารทดสอบแบบลาดเอยง (gradient plate assay) ทมความเขมขนของสารก าจดวชพชตงแต 0 ถง 1 NRR น าสารแขวนลอยเอนโดไฟตกแบคทเรยทมคาการดดกลนแสง เทากบ 0.5 ทความยาวคลนแสง 530 นาโนเมตร มาลากบนผวหนาอาหารทดสอบเปนเสนตรงจากระดบความเขมขนสงไปหาความเขมขนต า น าไปบมทอณหภม 30 องศาเซลเซยส เปนเวลา 2 วน บนทกผลการเจรญ (ดดแปลงจากวนนทดา หมะมาน, 2541)
ผลการศกษาและอภปรายผล
การแยกเอนโดไฟตกแบคทเรยทมความสามารถในการละลายฟอสเฟตจากตวอยางรากมนส าปะหลง จากการเกบตวอยางรากมนส าปะหลงจากต าบลหนองปลาไหล อ าเภอบางละมง จงหวดชลบร จ านวน 25 จด จากแปลงปลกมนส าปะหลง 5 แปลง พบวาเอนโดไฟตกแบคทเรยทสามารถละลายฟอสเฟตไดจะเกดบรเวณใสขนรอบโคโลนบนอาหาร Pikovskaya’s agar จ านวนเอนโดไฟตกแบคทเรยทสามารถละลายฟอสเฟตไดอยในชวงระหวาง 105 ถง 106 โคโลนตอกรม (ตารางท 1) เมอคดเลอกลกษณะโคโลนทแตกตางกนของเอนโดไฟตกแบคทเรยทสามารถละลายฟอสเฟตได สามารถคดแยกไดทงหมด 30 ไอโซเลท โดยจดเปนแบคทเรยแกรมลบ 28 ไอโซเลท และแกรมบวก 2 ไอโซเลท ไอโซเลทสวนใหญมรปรางเซลลเปนทอนสน จากผลการคดแยกเอนโดไฟตกแบคทเรยจากบรเวณรากมนส าปะหลง พบแบคทเรยสวนมากเปนแบคทเรยแกรมลบ มรปรางเซลลเปนทอนสน อยในกลมทเจรญในททมอากาศหรอไมมกได (facultative anaerobe) ซงเปนสภาวะภายในเนอเยอของพช จงท าใหสวนใหญ เอนโดไฟตกแบคทเรยทคดแยกไดจงอยในกลมน เชนเดยวกบการศกษาเอนโดไฟตกแบคทเรยของพชตระกลถวในประเทศเมกซโกทพบแบคทเรยแกรมลบ มรปรางเซลลเปนทอนสนทงหมด (Wang et al., 2006)
ขนาดเสนผานศนยกลางวงใสรอบโคโลน ขนาดเสนผานศนยกลางโคโลน

368 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
ตารางท 1 The colony number of total bacteria and endophytic bacteria isolated from cassava root samples on Pikovskaya’s agar (PVK)
Sample Total colony (CFU/g)
Phosphate solubilizing colony (CFU/g)
Isolate number
Endophytic bacterial colony on PVK
1 4.9× 105 1.0× 105 4
2 1.3× 107 7.4× 106 7 3 1.5× 107 5.7× 106 10 4 9.1× 106 1.6× 106 5
5 1.5× 106 2.9× 105 4
การทดสอบความสามารถในการละลายฟอสเฟต (Phosphate solubilization)
จากการทดสอบความสามารถของเอนโดไฟตกแบคทเรยทมความสามารถในการละลายฟอสเฟตจ านวน 30 ไอโซเลท โดยการวดขนาดเสนผานศนยกลางโซนใสรอบโคโลนบนอาหาร Pikovskaya’s agar (PVK) เทยบกบขนาดเสนผานศนยกลางโคโลน เพอค านวณหาคาดชนการละลายฟอสเฟต พบวาเอนโดไฟตกแบคทเรยทคดเลอกได มคาดชนการละลายฟอสเฟตอยในชวงระหวาง 0.94 ± 0.06 ถง 1.24 ± 0.02 โดยมไอโซเลททมคาดชนการละลายฟอสเฟตมากกวา 1.0 จ านวน 27 ไอโซเลท นอกจากนมไอโซเลททมประสทธภาพในการละลายฟอสเฟตสงตงแตวนท 1 จ านวน 3 ไอโซเลท ไดแก CRP-13 CRP-16 และ CRP-22 (ตารางท 2) แสดงใหเหนวาเอนโดไฟตกแบคทเรยเหลาน มความสามารถในการละลายฟอสเฟตทไมละลายน าซงเปนองคประกอบหลกในอาหาร PVK ใหอยในรปทละลายน าได จงสงเกตการสรางไดจากบรเวณใสทจะเกดขนรอบโคโลน โดยอาจเกดจากกลไกการสรางกรดอนทรยหรอสารทสามารถแลกเปลยนประจกบสารประกอบฟอสเฟต ซงมการรายงานวาแบคทเรยและราทอาศยในดนสวนใหญสามารถละลายสารประกอบฟอสเฟตในดนไดดวยกลไกเหลาน (Whitelaw, 2000)
การทดสอบการผลตอนโดลอะซตกแอซด (Indole acetic acid หรอ IAA)
จากการคดเลอกเอนโดไฟตกแบคทเรย 30 ไอโซเลท ซงมความสามารถในการละลายฟอสเฟต มาทดสอบความสามารถในการสงเคราะห IAA โดยเลยงในอาหาร nutrient broth ทผสม L-tryptophan และทดสอบปฏกรยากบสาร Salkowski’s reagent พบวาเอนโดไฟตกแบคทเรยมการผลต IAA ไดอยในชวง 22.1 ถง 428.7 ไมโครกรมตอมลลลตร โดยมไอโซเลททสามารถผลต IAA ไดสงมากกวา 200 ไมโครกรมตอมลลลตรจ านวน 4 ไอโซเลท ไดแก CRP-12 CRP-15 CRP-19 และ CRP-29 (ตารางท 2) โดยจลนทรยทอาศยบรเวณรากพชทสามารถสงเสรมการเจรญของพชไดนนอาจผลตฮอรโมนพชไดหลายชนดรวมทง IAA ซงมผลตอการเพมพนทผวรากและจ านวนรากพช และสามารถใช tryptophan สรางฮอรโมนนไดหลายวธอกดวย (Spaepen et al., 2007) นอกจากน ยงมหลายรายงานทสนบสนนวาแบคทเรยทอาศยบรเวณรากพชหลายชนดมความหลากหลายสงและมความสามารถในการสรางฮอรโมนพชไดหลายชนดอกดวย (Bhattacharyya and Jha, 2012)
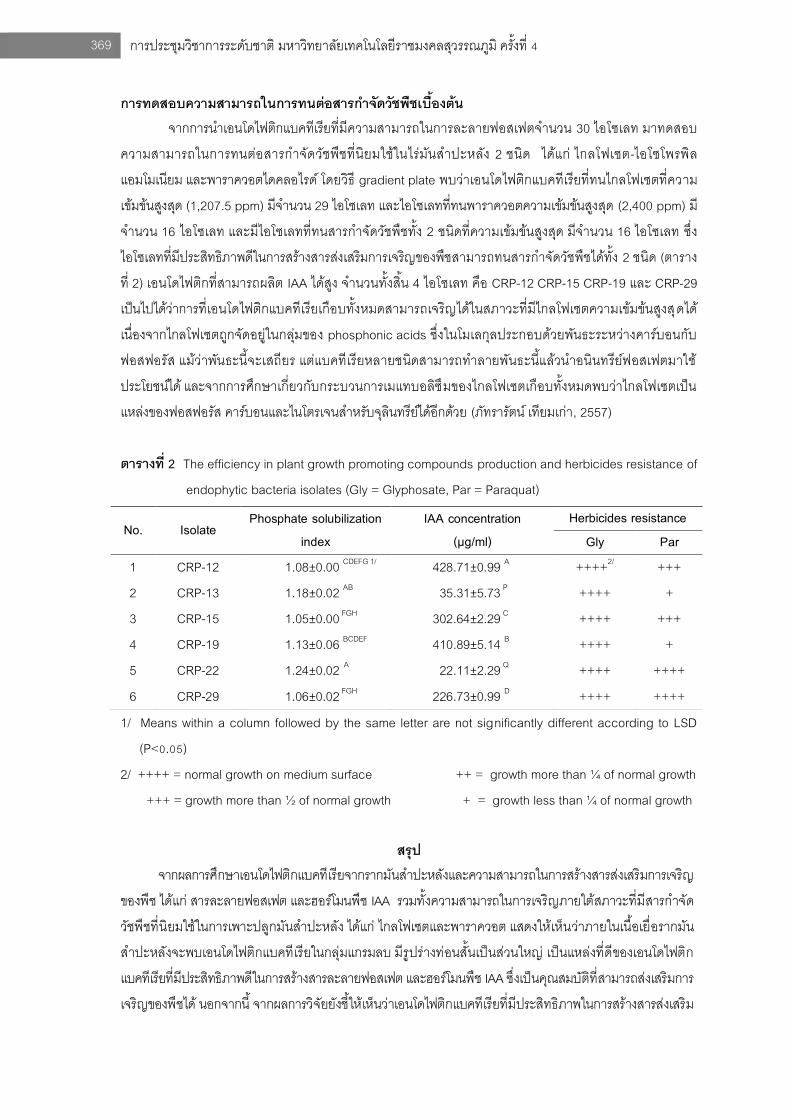
369 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
การทดสอบความสามารถในการทนตอสารก าจดวชพชเบองตน จากการน าเอนโดไฟตกแบคทเรยทมความสามารถในการละลายฟอสเฟตจ านวน 30 ไอโซเลท มาทดสอบ
ความสามารถในการทนตอสารก าจดวชพชทนยมใชในไรมนส าปะหลง 2 ชนด ไดแก ไกลโฟเซต-ไอโซโพรพลแอมโมเนยม และพาราควอตไดคลอไรด โดยวธ gradient plate พบวาเอนโดไฟตกแบคทเรยททนไกลโฟเซตทความเขมขนสงสด (1,207.5 ppm) มจ านวน 29 ไอโซเลท และไอโซเลทททนพาราควอตความเขมขนสงสด (2,400 ppm) มจ านวน 16 ไอโซเลท และมไอโซเลทททนสารก าจดวชพชทง 2 ชนดทความเขมขนสงสด มจ านวน 16 ไอโซเลท ซง ไอโซเลททมประสทธภาพดในการสรางสารสงเสรมการเจรญของพชสามารถทนสารก าจดวชพชไดทง 2 ชนด (ตารางท 2) เอนโดไฟตกทสามารถผลต IAA ไดสง จ านวนทงสน 4 ไอโซเลท คอ CRP-12 CRP-15 CRP-19 และ CRP-29 เปนไปไดวาการทเอนโดไฟตกแบคทเรยเกอบทงหมดสามารถเจรญไดในสภาวะทมไกลโฟเซตความเขมขนสงสดได เนองจากไกลโฟเซตถกจดอยในกลมของ phosphonic acids ซงในโมเลกลประกอบดวยพนธะระหวางคารบอนกบฟอสฟอรส แมวาพนธะนจะเสถยร แตแบคทเรยหลายชนดสามารถท าลายพนธะนแลวน าอนนทรยฟอสเฟตมาใชประโยชนได และจากการศกษาเกยวกบกระบวนการเมแทบอลซมของไกลโฟเซตเกอบทงหมดพบวาไกลโฟเซตเปนแหลงของฟอสฟอรส คารบอนและไนโตรเจนส าหรบจลนทรยไดอกดวย (ภทรารตน เทยมเกา, 2557)
ตารางท 2 The efficiency in plant growth promoting compounds production and herbicides resistance of endophytic bacteria isolates (Gly = Glyphosate, Par = Paraquat)
1/ Means within a column followed by the same letter are not significantly different according to LSD (P<0.05)
2/ ++++ = normal growth on medium surface ++ = growth more than ¼ of normal growth +++ = growth more than ½ of normal growth + = growth less than ¼ of normal growth
สรป จากผลการศกษาเอนโดไฟตกแบคทเรยจากรากมนส าปะหลงและความสามารถในการสรางสารสงเสรมการเจรญ
ของพช ไดแก สารละลายฟอสเฟต และฮอรโมนพช IAA รวมทงความสามารถในการเจรญภายใตสภาวะทมสารก าจดวชพชทนยมใชในการเพาะปลกมนส าปะหลง ไดแก ไกลโฟเซตและพาราควอต แสดงใหเหนวาภายในเนอเยอรากมนส าปะหลงจะพบเอนโดไฟตกแบคทเรยในกลมแกรมลบ มรปรางทอนสนเปนสวนใหญ เปนแหลงทดของเอนโดไฟตกแบคทเรยทมประสทธภาพดในการสรางสารละลายฟอสเฟต และฮอรโมนพช IAA ซงเปนคณสมบตทสามารถสงเสรมการเจรญของพชได นอกจากน จากผลการวจยยงชใหเหนวาเอนโดไฟตกแบคทเรยทมประสทธภาพในการสรางสารสงเสรม
No. Isolate Phosphate solubilization
index IAA concentration
(µg/ml) Herbicides resistance
Gly Par 1 CRP-12 1.08±0.00 CDEFG 1/
428.71±0.99 A ++++2/ +++ 2 CRP-13 1.18±0.02 AB 35.31±5.73 P ++++ + 3 CRP-15 1.05±0.00 FGH 302.64±2.29 C ++++ +++ 4 CRP-19 1.13±0.06 BCDEF 410.89±5.14 B ++++ + 5 CRP-22 1.24±0.02 A 22.11±2.29 Q ++++ ++++ 6 CRP-29 1.06±0.02 FGH 226.73±0.99 D ++++ ++++

370 การประชมวชาการระดบชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ครงท 4
การเจรญของพชทคดแยกไดจากไรมนส าปะหลงทมการใชสารก าจดวชพชมาเปนเวลานาน สามารถรอดชวตและเจรญเพมจ านวนได แมอยในสภาวะทมความเขมขนของสารก าจดวชพชสงสดกตาม ดงนน เอนโดไฟตกแบคทเรยเหลาน จงเปนตวเลอกทดในการน าไปศกษาตอเพอศกษาผลของการใชเอนโดไฟตกแบคทเรยรวมกบการปลกพชจรง ซงจะเปนประโยชนตอการน าไปประยกตใชเพอสงเสรมการเจรญของมนส าปะหลงหรอพชชนดอนๆ ทงในพนทเพาะปลกทมการใชสารก าจดวชพชหรอไมกตาม
ค าขอบคณ
ขอขอบคณคณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ศนยหนตรา ในการสนบสนนอปกรณ เครองมอ สารเคม และสถานทในการท าวจยในครงน
เอกสารอางอง
พวงรตน ขจตวชยานกล. (2556). การวจยเชงบรณาการเพอเสรมสรางศกยภาพหนวยงานทองถนในการจดการและปองกนการปนเปอนของสารพษบนพนทตนน านาน (รายงานผลการวจย). กรงเทพฯ: ส านกงานคณะกรรมการวจยแหงชาต.
ภทรารตน เทยมเกา. (2557). ความเปนพษของไกลโฟเซตและการยอยสลายโดยจลนทรย. วารสารเกษตรพระจอมเกลา. 32(3), 71–79. มณทรา พรหมพทยายทธ, (2553). ประสทธภาพการผลตมนส าปะหลง. กรงเทพฯ: สวนวจยเศรษฐกจพชไรนา ส านกวจยเศรษฐกจ
การเกษตร ส านกงานเศรษฐกจการเกษตร. วนนทดา หมะมาน. (2541). การตรวจหาแบคทเรยในดนทสามารถยอยสลายสารก าจดวชพชอะลาคลอรและออกซฟลออรเฟน
(วทยานพนธปรญญามหาบณฑต). มหาวทยาลยเชยงใหม, เชยงใหม. Bhattacharyya, P.N., & Jha, D.K. (2012). Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World
Journal of Microbiology and Biotechnology. 28, 1327–1350. Bric, J.M., Bostock, R.M., & Silversone, S.E. (1991). Rapid in situ assay for indole acetic acid production by bacteria
immobilization a nitrocellulose membrane. Applied and Environmental Microbiology. 57, 535–538. Coombs, J. T., & Franco, C.M.M. (2003). Isolation and identification of actinobacteria from surface-sterilized wheat roots.
Applied and Environmental Microbiology. 69, 5603–5608. Hurek, B.R., & Hurek, T. (1998). Life in grasses: diazotrophic endophytes. Trends in Microbiology. 6(4), 139–144. Nogueira, E.D.M., Vinagre, F., Masuda, H.P., Vargas, C., de Pádua, V.L.M., da Silva, F.R., dos Santos R.V., Baldani, J.I., Ferreira,
P.C.G., & Hemerley, A.S. (2001). Expression of sugarcane genes induced by inoculation with Gluconacetobacter diazotrophicus and Herbaspirillum rubrisubalbicans Genet. The Journal of Molecular Biology. 24, 199–206.
Spaepen, S., Vanderleyden, J., & Remans, R. (2007). Indole-3-acetic acid in microbial and microorganism- plant signaling. In Unden, F. (ed) FEMS microbiology review. (pp. 1–24) New York: Blackwell Publishing Ltd. Wang, E.T., Tan, Z.Y., Guo, X.W., Rodrí guez-Duran, R., Boll, G., & Martí nez, R.E. (2006). Diverse endophytic bacteria
isolated from a leguminous tree Conzattia multiflora grown in Mexico. Archives of Microbiology. 186, 251–259. Whitelaw, M.A. (2000). Growth promotion of plants inoculated with phosphate-solubilizing fungi. Advances
in Agronomy. 69, 99–151.

Agricultural Sci. J. 50 : 1 (Suppl.) : 306-312 (2019) ว. วทย. กษ. 50 : 1 (พเศษ) : 306-312 (2562)
306
ผลของสารสกดจากแหวหม ผกโขมหนาม และผกเสยนตอเชอรา Fusarium moniliforme
สาเหตโรคเหยวเนาแดงของออย
Effect of Cyperus rotundus, Amaranthus spinosus and Cleome gynandra Extracts on
Fusarium moniliforme Causing Red Rot Wilt Disease of Sugarcane
ดวงหทย รตนสจธรรม1* นฤมล ศรนวล1 และ กาญจนา พศาภาค1,2
Dounghatai Rattanasatchatham1*, Nareumol Srinual1 and Kanjana Pisapak1,2
160 หม 3 สาขาวชาวทยาศาสตร คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม พระนครศรอยธยา 13000 2หนวยวจยและพฒนาผลตภณฑธรรมชาต มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม พระนครศรอยธยา 13000 160 Moo 3 Division of Science, Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi
13000, Thailand 2Natural Product Research and Development Unit, Rajamangala University of Technology Suvarnabhumi 13000, Thailand *Corresponding author: [email protected]
บทคดยอ
ออย (Saccharum officinarum L.) จดเปนพชเศรษฐกจของประเทศไทย ปจจบนเกษตรกรใชสารเคมในการ
ควบคมศตรพช ซงสารเคมเหลานเปนพษตอสงแวดลอมและมนษย การศกษานจงมวตถประสงคเพอศกษาผลของสารสกดจาก
พชอบแหง ไดแก หวแหวหม (Cyperus rotundus L.) ดอกผกโขมหนาม (Amaranthus spinosus L.) และผกเสยนทงตน
(Cleome gynandra L.) ในการยบยงการเจรญของเชอรา Fusarium moniliforme ซงเปนสาเหตโรคเหยวเนาแดงของออย
โดยใชตวทาละลาย 2 ชนด ไดแก เอทานอลความเขมขน 95 เปอรเซนต และนากลน ทดสอบดวยวธ poisoned food
technique บนอาหาร potato dextrose agar (PDA) ทระดบความเขมขน 10 เปอรเซนต (ปรมาตร/ปรมาตร) ผลการศกษา
พบวาสารสกดจากพชทง 3 ชนด สามารถยบยงเชอรา Fusarium moniliforme ได โดยสารสกดจากผกโขมหนามและ
ผกเสยนทใชตวทาละลายเอทานอล ใหคาเปอรเซนตการยบยงเชอราสงทสดเทากบ 89.70 และ 88.82 เปอรเซนต ตามลาดบ
เมอนาไปศกษาระดบความเขมขนทเหมาะสม 5 ระดบ ไดแก 0.1, 0.5, 1, 5 และ 10 เปอรเซนต (ปรมาตร/ปรมาตร) และ
นามาทดสอบกบเชอราอกครงดวยวธเดยวกน พบวาสารสกดจากพชทง 2 ชนด ทระดบความเขมขน 10 เปอรเซนต (ปรมาตร/
ปรมาตร) ใหผลในการยบยงเชอรา Fusarium moniliforme ไดดทสด
คาสาคญ: สารสกดจากพช, การยบยงเชอรา, Fusarium sp.
ABSTRACT
Sugarcane ( Saccharum officinarum L. ) is an economic crop of Thailand. Currently, farmers use
chemicals to control pests which are toxic to the environment and humans. This study aim to test the
effects of crude extracts from dried plants, including tuber of nut grass ( Cyperus rotundus L. ) , flower of
spiny amaranth (Amaranthus spinosus L.) and whole parts of spider plant (Cleome gynandra L.) to inhibit
the growth of Fusarium moniliforme which cause red rot wilt disease. Two solvents, 95 percent ethanol
and distilled water were used as the extraction solvents. Antifungal properties were tested by using
poisoned food technique on potato dextrose agar (PDA) containing 10 percent (v/v) of the crude extract.

ผลของสารสกด ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
307
The results found that the crude extracts from these three plants could inhibit the growth of Fusarium
moniliforme. The ethanol crude extracts from Amaranthus spinosus L. and Cleome gynandra L. showed
the highest of percentage inhibition with 89.70 and 88.82, respectively. These crude extracts were diluted
to five concentrations level of 0.1, 0.5, 1, 5 and 10 percent (v/v) and tested by the same method. The best
inhibition of Fusarium moniliforme were found at 10 percent of both crude extracts.
Keywords: Plant extracts, Antifungal, Fusarium sp.
บทนา
ออย (sugarcane) เปนพชเศรษฐกจทสาคญในการสงออกนาตาลของประเทศไทย นอกจากนยงสามารถนาไปใช
ประโยชนอนๆ ได เชน ผลตเยอกระดาษและพลงงานทดแทน ปจจบนเกษตรกรชาวไรออยมกประสบปญหาโรคระบาดจาก
แมลงศตรพชและจลนทรย โดยโรคทสาคญ ไดแก โรคเหยวเนาแดง (red rod wilt disease) จากเชอรา Fusarium
moniliforme (Gawade et al., 2012) เชอราจะเขาทาลายระบบทอนาและทออาหารของพชสงผลใหไมสามารถสงตอนา
และสารอาหารไปทสวนบนของลาตนได รวมไปถงการสรางสารพษทาลายเนอเยอ จงทาใหพชแสดงอาการเหยวและเนา
บรเวณภายในลาตน โดยทวไปเกษตรกรจะใชสารเคมในการปองกนและกาจดเชอราทเปนสาเหตของโรค อยางไรกตามการใช
สารเคมในปรมาณทไมเหมาะสมและการใชสารเคมชนดเดมตดตอกนเปนเวลานาน อาจทาใหเชอราเกดการตานทานตอ
สารเคม อกทงยงสงผลใหเกดการตกคางของสารเคมในผลผลตและสงแวดลอม ซงสงผลเสยตอเกษตรกรและผบรโภค จาก
ปญหาดงกลาวการควบคมโรคพชโดยใชสารสกดจากธรรมชาต (natural extracts) จงเปนอกทางเลอกหนงทนาสนใจ จาก
รายงานวจยทผานมาพบวามพชหลายชนดทสามารถยบยงการเจรญของเชอราได เชน Rakphol (2012) พบวาสารสกดจาก
พช ไดแก ชองนาง รางจด และสรอยอนทนล มความสามารถยบยงเชอรา Fusarium oxysporum ทกอโรคเหยวในมะเขอ
เทศ นอกจากน Chuwatworakoon et al. (2017) ยงรายงานวาสารสกดหยาบจากอบเชยและขาทสกดดวยตวทาละลาย
เอทลอะซเตททความเขมขน 100,000 ppm มประสทธภาพในการยบยงเชอรา Penicillium citrinum, Aspergillus flavus
var. flavus และ Aspergillus tamari ไดด
ดงนนการศกษานจงไดนาพชทพบทวไปในทองถน ไดแก แหวหม ผกโขมหนาม และผกเสยน มาทาการสกดสารและ
ทดสอบการยบยงการเจรญของเชอรา Fusarium moniliforme ทเปนสาเหตโรคเหยวเนาแดงของออย โดยเลอกใชเฉพาะ
สวนหวแหวหม ดอกผกโขมหนาม และผกเสยนใชทงตน เนองจากมรายงานวาสารสกดจากสวนของพชดงกลาวมสารสาคญ
หลายชนด เชน สารกลมอลคาลอยด ซาโปนน และนามนหอมระเหย ซงมคณสมบตในการยบยงการเจรญของเชอราบางชนด
(Yusnawan, 2015; Al-Snafi, 2016; Imanirampa and Alele, 2016) ผลทไดจากการศกษานจะเปนขอมลเบองตนทสาคญ
ในการพฒนาและประยกตใชสารสกดจากพชทง 3 ชนด เปนสารในการควบคมโรคพชทมประสทธภาพ เพอลดการใชสารเคม
ทเปนผลเสยตอสงแวดลอมตอไป
อปกรณและวธการ
การเตรยมตวอยางพช
นาตวอยางพช 3 ชนด ไดแก หวแหวหม (Cyperus rotundus L.) ดอกผกโขมหนาม (Amaranthus spinosus L.)
และผกเสยนทงตน (Cleome gynandra L.) มาลางใหสะอาด ผงลมใหแหง ตดเปนชนเลกๆ แลวนาไปอบทอณหภม 55
องศาเซลเซยส นาน 48 ชวโมง จากนนนาตวอยางพชทอบแลวมาบดใหละเอยดและเกบในทแหง (Thongruk, 2010)
การสกดสารจากพช

ผลของสารสกด ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
308
ชงนาหนกพชอบแหงทบดแลวปรมาณ 200 กรม จากนนนาไปแชในตวทาละลาย 2 ชนด ไดแก เอทานอลความ
เขมขน 95 เปอรเซนต และนากลน ใหทวมตวอยางพช นาน 24 ชวโมง ทอณหภมหอง โดยไมเขยา จากนนกรองดวยกระดาษ
กรอง Whatman เบอร 1 แลวนาสารละลายทไดไประเหยตวทาละลายออกดวยเครองกลนระเหยสารแบบหมน (rotary
evaporator) ทอณหภม 50 องศาเซลเซยส ชงนาหนกของสารสกดหยาบ (crude extract) ทได แลวนาไปคานวณเปอรเซนต
ผลผลต (% yield) จากสตรดานลาง (Phrompittayarat et al., 2007) เกบสารสกดหยาบทไดในขวดสชา ทอณหภม 4
องศาเซลเซยส กอนนาไปทดสอบในขนตอนตอไป
นาหนกสารสกดหยาบทได
นาหนกตวอยางแหงเรมตน
เชอราทใชทดสอบ
เชอราทใชทดสอบ ไดแก Fusarium moniliforme TISTR3175 จากสถาบนวจยวทยาศาสตรและเทคโนโลยแหง
ประเทศไทย เมอตองการจะใชทดสอบใหนามาเพาะเลยงบนอาหาร potato dextrose agar (PDA) บมทอณหภม 25
องศาเซลเซยส นาน 7 วน
การทดสอบผลของสารสกดจากพชในการยบยงเชอรา Fusarium moniliforme
ทดสอบผลของสารสกดจากพชในการยบยงการเจรญของเชอรา Fusarium moniliforme ดวยวธ poisoned food
technique (Nuangmek and Aidunag, 2017) โดยตดเสนใยของเชอราบรเวณขอบโคโลนดวยทเจาะจกคอรก (cork borer)
ขนาดเสนผานศนยกลาง 0.5 เซนตเมตร วางบนอาหารเลยงเชอ PDA ทผสมสารสกดความเขมขน 10 เปอรเซนต
(ปรมาตร/ปรมาตร) และชดควบคม (control) ทไมผสมสารสกด ทาการทดลองทงหมด 3 ซา นาไปบมทอณหภม 25
องศาเซลเซยส นาน 7 วน จากนนวดขนาดเสนผานศนยกลางของโคโลนเชอราเปรยบเทยบกบชดควบคม คานวณคา
เปอรเซนตการยบยง (% inhibition) จากสตรดานลาง (Jamkratoke et al., 2004) จากนนนามาวเคราะหผลทางสถตดวย
Analysis of variance (ANOVA) และเปรยบเทยบคาเฉลยดวยวธ Duncan s new multiple range test
(A - B)
A
เมอ A คอ คาเฉลยเสนผานศนยกลางของโคโลนบนอาหารเลยงเชอชดควบคม
B คอ คาเฉลยเสนผานศนยกลางของโคโลนบนอาหารเลยงเชอทผสมสารสกด
การหาระดบความเขมขนทเหมาะสมของสารสกดจากพชในการยบยงเชอรา Fusarium moniliforme
นาสารสกดจากพชทใหคาเปอรเซนตการยบยงเชอราสงทสดมาทดสอบดวยวธ poisoned food technique โดย
ตดเสนใยของเชอราวางบนอาหาร PDA ทผสมสารสกดทระดบความเขมขนแตกตางกน ไดแก 0.1, 0.5, 1, 5 และ 10
เปอรเซนต (ปรมาตร/ปรมาตร) และชดควบคมทไมผสมสารสกด ทาการทดลองทงหมด 3 ซา บมทอณหภม 25 องศาเซลเซยส
นาน 7 วน คานวณคาเปอรเซนตการยบยง แลวนามาวเคราะหผลทางสถตดวย Analysis of variance (ANOVA) และ
เปรยบเทยบคาเฉลยดวยวธ Duncan s new multiple range test
ผลการทดลองและวจารณ
การเตรยมตวอยางพชและการสกดสารจากพช
จากการนาพช 3 ชนด ไดแก หวแหวหม ดอกผกโขมหนาม และผกเสยนทงตน ทอบแหงแลวมาบดใหละเอยด และ
แชในตวทาละลาย 2 ชนด ไดแก เอทานอลความเขมขน 95 เปอรเซนต และนากลน จากนนนาไประเหยตวทาละลายออกดวย
เครองกลนระเหยสารแบบหมน พบวาสารสกดหยาบทไดมลกษณะแตกตางกนขนอยกบชนดของพชและตวทาละลายทใช เมอ
นามาคานวณเปอรเซนตผลผลตของสารสกดพบวามคาอยในชวง 1.53–8.64 โดยแหวหมทใชตวทาละลายนามเปอรเซนต
เปอรเซนตผลผลต = X 100
เปอรเซนตการยบยง = X 100

ผลของสารสกด ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
309
ผลผลตสงทสด รองลงมาเปนผกเสยนทใชตวทาละลายนา โดยมเปอรเซนตผลผลตเทากบ 8.64 และ 7.98 ตามลาดบ สวน
แหวหมทใชตวทาละลายเอทานอลมเปอรเซนตผลผลตตาทสดเทากบ 1.53 (Table 1) จากผลการทดลองจะเหนไดวาพชทก
ชนดทสกดดวยตวทาละลายนาใหเปอรเซนตผลผลตมากกวาตวทาละลายเอทานอล แสดงใหเหนวาองคประกอบทางเคมทมอย
ในพชมความเปนขวสง จงสามารถละลายไดดในตวทาละลายนาซงมความเปนขวสงกวาตวทาละลายเอทานอล
การทดสอบผลของสารสกดจากพชในการยบยงเชอรา Fusarium moniliforme
การทดสอบสารสกดจากพชทระดบความเขมขน 10 เปอรเซนต (ปรมาตร/ปรมาตร) ในการยบยงเชอรา Fusarium
moniliforme ดวยวธ poisoned food technique พบวาสารสกดจากพชทง 3 ชนด สามารถยบยงการเจรญของเชอราได
โดยสารสกดจากพชทใชตวทาละลายเอทานอลมเปอรเซนตการยบยงเชอราสงกวาตวทาละลายนาอยางมนยสาคญทางสถต ซง
สอดคลองกบการทดลองของ Shanmuganathan and Kartikeyan (2016) ทพบวาสารสกดจากผกเสยนทใชตวทาละลาย
เอทานอลมผลในการยบยงเชอรา Fusarium solani ไดมากกวาการใชตวทาละลายนา นอกจากนผลการทดสอบยงพบวาสาร
สกดจากผกโขมหนามและผกเสยนทใชตวทาละลายเอทานอลมเปอรเซนตการยบยงเชอราสงทสดเทากบ 89.70 และ 88.82
ตามลาดบ ซงไมแตกตางกนอยางมนยสาคญทางสถต รองลงมาเปนผกโขมหนามทใชตวทาละลายนา แหวหมทใชตวทาละลาย
เอทานอล และแหวหมทใชตวทาละลายนามเปอรเซนตการยบยงเชอราเทากบ 73.16, 61.19 และ 53.33 ตามลาดบ ในขณะ
ทสารสกดจากผกเสยนทใชตวทาละลายนามเปอรเซนตการยบยงเชอราตาทสดเทากบ 9.59 (Figure 1) การทพชทง 3 ชนด
สามารถยบยงการเจรญของเชอราไดนนเนองมาจากสวนประกอบสาคญทมอยในพช โดยมรายงานวาหวแหวหมมนามนหอม
ระเหย (Al-Snafi, 2016; Bajpay et al., 2018) ดอกผกโขมหนามมอลคาลอยด ฟลาโวนอยด ซาโปนน และเทอรปนอยส
(Yusnawan, 2015) นอกจากนยงมรายงานวาผกเสยนมอลคาลอยด ฟลาโวนอยด ซาโปนน และแทนนน (Imanirampa and
Alele, 2016) ซงเปนสารทมความสามารถในการยบยงเชอราได ผลการทดลองทไดนสอดคลองกบ Bisht et al. (2011) และ
Jadhav and Biradar (2016) ทพบวาสารสกดจากแหวหมและผกโขมหนามสามารถยบยงเชอรา Fusarium oxysporum
และการทดลองของ Shanmuganathan and Kartikeyan (2016) ทพบวาสารสกดจากผกเสยนสามารถยบยงการเจรญของ
เชอรา Fusarium solani ได
การหาระดบความเขมขนทเหมาะสมของสารสกดจากพชในการยบยงเชอรา Fusarium moniliforme
จากการนาสารสกดจากพชทมเปอรเซนตการยบยงเชอราสงทสด ไดแก สารสกดผกโขมหนามและผกเสยนทใชตวทา
ละลายเอทานอล มาปรบความเขมขนเปน 5 ระดบ ไดแก 0.1, 0.5, 1, 5 และ 10 เปอรเซนต (ปรมาตร/ปรมาตร) แลวนาไป
ทดสอบประสทธภาพในการยบยงเชอราอกครง พบวาสารสกดจากพชทง 2 ชนดในทกระดบความเขมขนสามารถยบยงการ
เจรญของเชอราได โดยเชอรามการเจรญของเสนใยนอยลง เมอสารสกดมระดบความเขมขนสงขน จากผลการทดลองพบวา
สารสกดจากผกโขมหนามมเปอรเซนตการยบยงเชอราเทากบ 2.60, 11.40, 20.80, 40.00 และ 84.60 สวนสารสกดจาก
ผกเสยนมเปอรเซนตการยบยงเชอราเทากบ 7.69, 23.07, 28.85, 46.92 และ 84.03 ทระดบความเขมขน 0.1, 0.5, 1, 5 และ
10 เปอรเซนต (ปรมาตร/ปรมาตร) ตามลาดบ แสดงใหเหนวาสารสกดจากพชทง 2 ชนด ทระดบความเขมขนสงจะใหคา
เปอรเซนตการยบยงเชอราสงกวาการใชสารสกดทความเขมขนตา กลาวคอทระดบความเขมขน 10 เปอรเซนต ใหผลการ
ยบยงเชอราไดดทสด ซงมคาแตกตางจากระดบความเขมขนอนอยางมนยสาคญทางสถต (Figure 2) ผลการศกษาทไดน
สอดคลองกบผลการทดลองของ Shanmuganathan and Kartikeyan (2016) ทพบวาสารสกดจากผกเสยนทใชตวทาละลาย
เอทานอลใหผลการยบยงเชอรา Fusarium solani ไดดเมอมระดบความเขมขนสงขน
สรป
การทดสอบสารสกดจากพช 3 ชนด ไดแก หวแหวหม ดอกผกโขมหนาม และผกเสยนทงตน ทระดบความเขมขน 10
เปอรเซนต (ปรมาตร/ปรมาตร) ในการยบยงเชอรา Fusarium moniliforme พบวาสารสกดจากพชทง 3 ชนดมความสามารถ

ผลของสารสกด ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
310
ในการยบยงเชอราได โดยสารสกดจากผกโขมหนามและผกเสยนทใชตวทาละลายเอทานอลมเปอรเซนตการยบยงเชอราสง
ทสดเทากบ 89.70 และ 88.82 ตามลาดบ สวนสารสกดจากผกเสยนทใชตวทาละลายนามเปอรเซนตการยบยงเชอราตาทสด
เทากบ 9.59 เมอนาสารสกดจากผกโขมหนามและผกเสยนทใชตวทาละลายเอทานอลซงมเปอรเซนตการยบยงเชอราสงทสด
มาปรบความเขมขนเปน 5 ระดบ ไดแก 0.1, 0.5, 1, 5 และ 10 เปอรเซนต (ปรมาตร/ปรมาตร) แลวนามาทดสอบกบเชอรา
อกครงดวยวธเดยวกน พบวาสารสกดจากพชทง 2 ชนดในทกระดบความเขมขนสามารถยบยงเชอราได โดยทระดบความ
เขมขน 10 เปอรเซนต (ปรมาตร/ปรมาตร) ใหผลการยบยงเชอราไดดทสด จากผลการทดสอบจงมความเปนไปไดในการนา
สารสกดจากผกโขมหนามและผกเสยนทใชตวทาละลายเอทานอลไปพฒนาและประยกตใชในการควบคมโรคพช เพอลด
ปรมาณการใชสารเคม อยางไรกตามควรมการศกษาสารสกดจากพชในระดบความเขมขนทสงขนเพอเพมประสทธภาพในการ
ยบยงเชอรา รวมทงศกษาประสทธภาพในการควบคมโรคในสภาพพนทจรงตอไป
กตตกรรมประกาศ
ขอขอบคณคณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ศนยพระนครศรอยธยา
หนตรา ในการสนบสนนอปกรณ เครองมอ สารเคม และสถานทในการทาวจยครงน
เอกสารอางอง
Al-Snafi, A.E. 2016. A review on Cyperus rotundus A potential medicinal plant. IOSR. J. Pharm. 6: 32–48.
Bajpay, A., R.C. Nainwal, D. Singh and S.K. Tewari. 2018. Medicinal value of Cyperus rotundus Linn: An
updated review. Medicinal Plants. 10: 165–170.
Bisht, A., G.R.S. Bisht, M. Singh, R. Gupta and V. Singh. 2011. Chemical composition and antimicrobial
activity of essential oil of tubers of Cyperus rotundus Linn. collected from Dehradun
(Uttarakhand). International Journal of Research in Pharmaceutical and Biomedical Sciences. 2:
661–665.
Chuwatworakoon, T., S. Taveepanich, P. Suwanakood and S. Lumlong. 2017. Efficiency of herb crude
extracts to inhibit the growth of Penicillium citrinum Aspergillus flavus var. flavus and Aspergillus
tamarii isolated from the rubber sheet. KKU Science Journal. 45: 276–286. (in Thai)
Gawade, D.B., B.H. Pawar, S.J. Gawande and V.C. Vasekar. 2012. Antagonistic effect of Trichoderma against
Fusarium moniliformae the causal of sugarcane wilt. Am. Eurasian. J. Agric. Environ. Sci. 12: 1236–
1241.
Imanirampa, L. and P.E. Alele. 2016. Antifungal activity of Cleome gynandra L. aerial parts for topical
treatment of Tinea capitis: an in vitro evaluation. BMC. Complement. Altern. Med. 16: 1–8.
Jadhav, V. and S.D. Biradar. 2016. Evaluation of Antifungal Activity of Amaranthus spinosus L.
(Amaranthaceae). Int. J. Curr. Microbiol. Appl. Sci. 5: 38–43.
Jamkratoke, S., C. Rattanakreetakul, C. Leksomboon, N. Farungsang, K. Wanichkul and U. Farungsang. 2004.
Effects of Boesenbergia rotunda, Curcuma longa and Zingiber officinalis Extracts on Posthavest
Disease Fungi. Proceedings of 42nd Kasetsart University Annual Conference: Plants, Agricultural
Extension and Communication. Bangkok. 521–528. (in Thai)
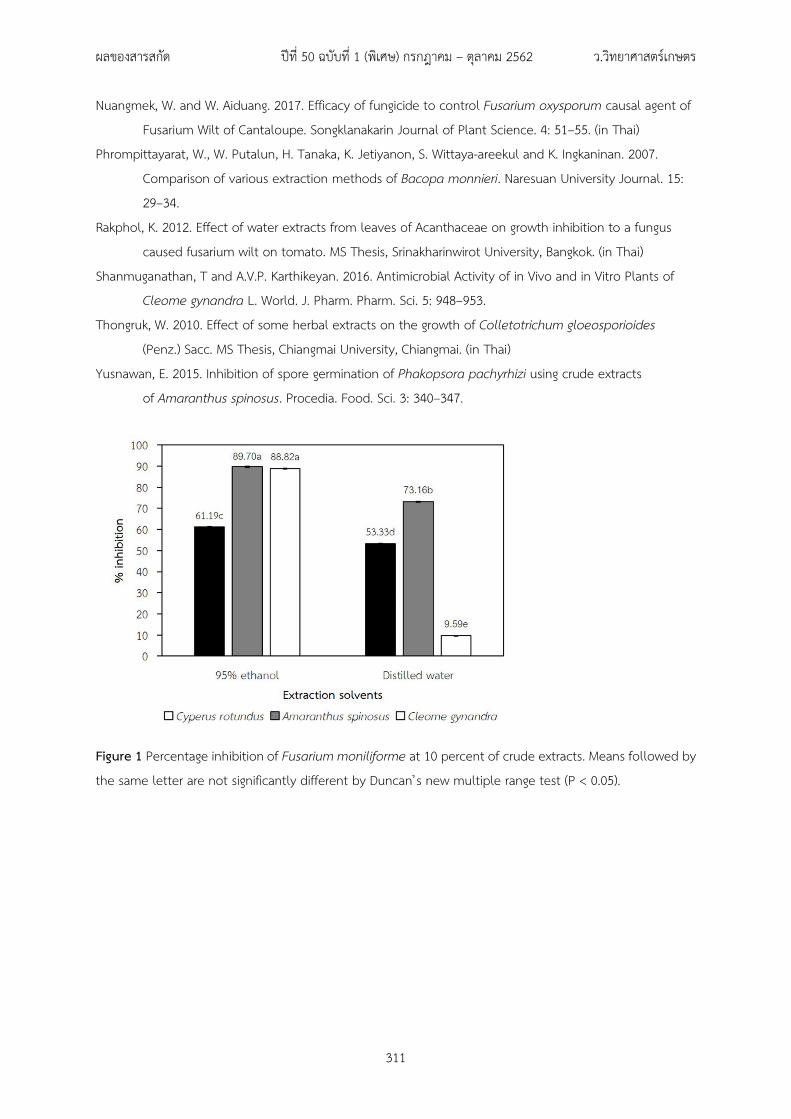
ผลของสารสกด ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
311
Nuangmek, W. and W. Aiduang. 2017. Efficacy of fungicide to control Fusarium oxysporum causal agent of
Fusarium Wilt of Cantaloupe. Songklanakarin Journal of Plant Science. 4: 51–55. (in Thai)
Phrompittayarat, W., W. Putalun, H. Tanaka, K. Jetiyanon, S. Wittaya-areekul and K. Ingkaninan. 2007.
Comparison of various extraction methods of Bacopa monnieri. Naresuan University Journal. 15:
29–34.
Rakphol, K. 2012. Effect of water extracts from leaves of Acanthaceae on growth inhibition to a fungus
caused fusarium wilt on tomato. MS Thesis, Srinakharinwirot University, Bangkok. (in Thai)
Shanmuganathan, T and A.V.P. Karthikeyan. 2016. Antimicrobial Activity of in Vivo and in Vitro Plants of
Cleome gynandra L. World. J. Pharm. Pharm. Sci. 5: 948–953.
Thongruk, W. 2010. Effect of some herbal extracts on the growth of Colletotrichum gloeosporioides
(Penz.) Sacc. MS Thesis, Chiangmai University, Chiangmai. (in Thai)
Yusnawan, E. 2015. Inhibition of spore germination of Phakopsora pachyrhizi using crude extracts
of Amaranthus spinosus. Procedia. Food. Sci. 3: 340–347.
Figure 1 Percentage inhibition of Fusarium moniliforme at 10 percent of crude extracts. Means followed by
the same letter are not significantly different by Duncan s new multiple range test (P < 0.05).
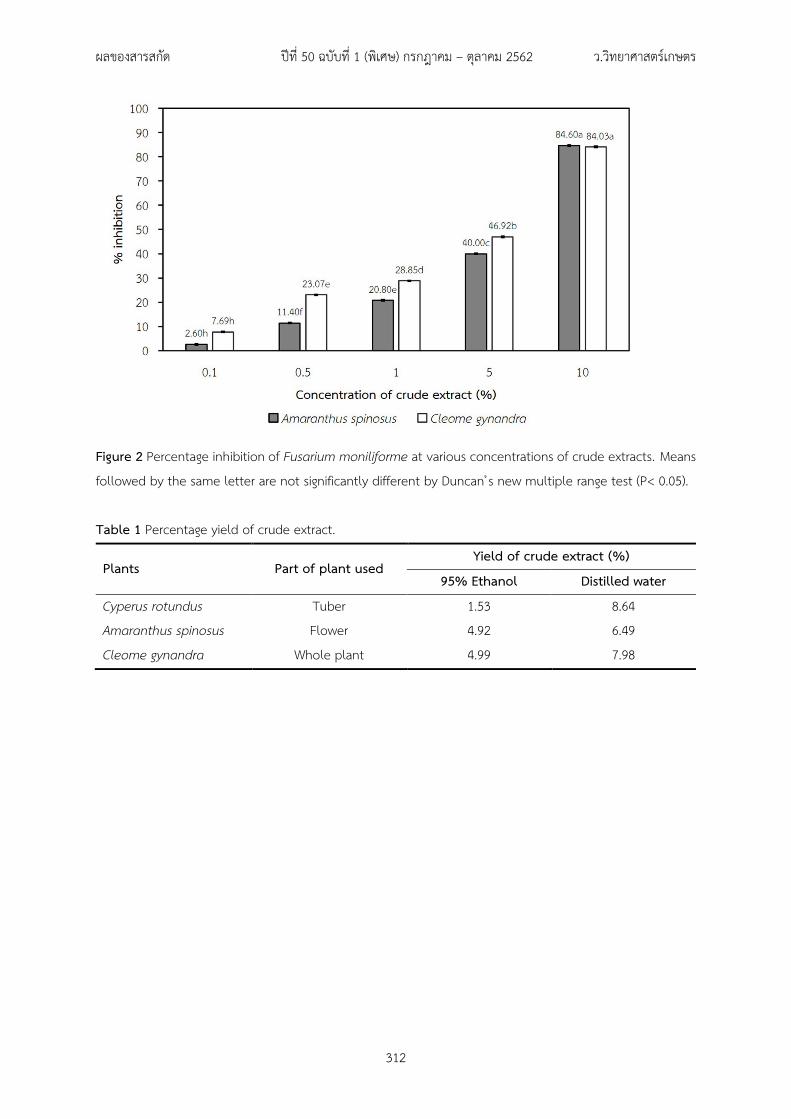
ผลของสารสกด ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
312
Figure 2 Percentage inhibition of Fusarium moniliforme at various concentrations of crude extracts. Means
followed by the same letter are not significantly different by Duncan s new multiple range test (P< 0.05).
Table 1 Percentage yield of crude extract.
Plants Part of plant used Yield of crude extract (%)
95% Ethanol Distilled water
Cyperus rotundus Tuber 1.53 8.64
Amaranthus spinosus Flower 4.92 6.49
Cleome gynandra Whole plant 4.99 7.98

1 สาขาสตวศาสตรและเทคโนโลยอาหารสตว คณะเกษตรศาสตร ทรพยากรธรรมชาตและสงแวดลอม มหาวทยาลยนเรศวร
Division of Animal Science and Feed Technology, Faculty of Agriculture Natural Resources and Environment, Naresuan University2 สถานวจยเพอความเปนเลศทางวชาการดานนวตกรรมการเกษตรและปศสตว คณะเกษตรศาสตร ทรพยากรธรรมชาตและสงแวดลอม
มหาวทยาลยนเรศวร
Center of Excellence for Agricultural and Livestock Innovations, Faculty of Agriculture Natural Resources and Environment
Naresuan University3 ภาควชาชววทยา คณะวทยาศาสตร มหาวทยาลยนเรศวร
Department of Biology, Faculty of Science, Naresuan University 4 สาขาวชาวทยาศาสตร คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม
Division of Science, Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi * Corresponding author: [email protected]
ผลกระทบเชงลบของใยอาหารชนดละลายน�าในอาหารกงบรสทธตอสมรรถภาพการผลต และจลนทรยในใสตงของไกเนอระยะแรก
Negative effects of soluble dietary fiber in semi-purified diets on performance and cecal microbe of young broiler chickens
ทศพร อนเจรญ1,2*, ขตตยา ลานแปง1, รองเดช ตงตระการพงษ3
และ พรพรรณ รตนะสจจะ4
Tossaporn Incharoen1,2*, Khattiya Lanpang1, Rongdej Tungtrakanpoung3
and Pornpan Ruttanasutja4
บทคดยอ: งานวจยชนนมวตถประสงคเพอประเมนผลของระดบใยอาหารชนดละลายน�าในอาหารกงบรสทธตอสมรรถภาพการผลต และจลนทรยในใสตงของไกเนอระยะแรก โดยใชกวกม (Guar gum) เปนแหลงของใยอาหารชนดละลายน�า ไกเนอ สายพนธ รอส 308 (Ross 308) เพศผ อาย 10 วน จ�านวน 80 ตว แบงถกออกเปน 4 กลมๆ ละ 4 ซ�าๆ ละ 5 ตว ไกแตละกลมไดรบอาหารทดลองเสรมกวกม ทระดบ 0 (T1), 3 (T2), 5 (T3) และ 7 (T4) เปอรเซนต ตามล�าดบ เมอสนสดการทดลอง ท อาย 24 วน พบวา น�าหนกตว (ทอาย 17 และ 24 วน) น�าหนกตวทเพมขนเฉลย และอตราการกนอาหาร มคาลดลง (P<0.05) ตามระดบใยอาหารชนดละลายน�าทเพมสงขน โดยเฉพาะอยางยงอตราการเปลยนอาหารเปนน�าหนกตวมคาแยลง (P<0.05) ในกลม T3 และ T4 เมอเปรยบเทยบกบกลม T1 ดานการประเมนทางจลชววทยา ไมพบเชอซลโมเนลลาในตวอยางสงเหลอของไสตงในไกทง 4 กลม ในขณะทสามารถตรวจพบเชออโคไลในปรมาณทไมแตกตางกนทางสถต (P>0.05) ดงนนการศกษานจงสามารถสรปไดวาอาหารไกเนอระยะแรกทมใยอาหารชนดละลายน�า ตงแตระดบ 3 เปอรเซนต ขนไป มผลท�าใหอตราการกนอาหาร และน�าหนกตวมคาลดลง รวมทงอตราการเปลยนอาหารเปนน�าหนกตวมคาแยลงอยางเหนไดชด (P<0.05) เมอระดบใยอาหารชนดละลายน�ามคาเพมสงขน ตงแตระดบ 5-7 เปอรเซนต อยางไรกตามระดบของใยอาหารชนดละลายน�าไมมผลตอการเปลยนแปลงของประชากรเชอซลโมเนลลาและอโคไลในไสตง ค�าส�าคญ: ใยอาหาร, อาหารกงบรสทธ, สมรรถภาพการผลต, ประชากรจลนทรยในใสตง, ไกเนอ
แกนเกษตร 47 ฉบบพเศษ 2 : (2562) KHON KAEN AGR. J. 47 SUPPL. 2 : (2019).

บทน�า
ใยอาหาร (Dietary fiber) ซงเปนองคประกอบหลกของผนงเซลลของพช สามารถจ�าแนกออกได 2 ชนด คอ ใยอาหารชนดละลายน�า (Soluble dietary fiber) และใยอาหารชนดไมละลายน�า (Insoluble dietary fiber) มความหนดต�า ความพองตวสงชวยเพมอตราการเคลอนทของอาหารในระบบทางเดนอาหารไดดขน ชวยกระตนการพฒนาโครงสรางของกระเพาะอาหารและล�าไส สอดคลองกบงานวจยกอนหนานของ Incharoen (2013) ทพบวาการเสรมใยอาหารชนดไมละลายน�าในอาหารไกเนอมผลชวยใหเกดการพฒนาของวลไลและเซลลดดซมของล�าไสเลกเพมขน สงผลใหประสทธภาพการยอยและดดซมดขน รวมไปถงสมรรถภาพการผลตทสงขน นอกจากนนแลวยงสงผลเชงบวกตอความสม�าเสมอฝงของไกไข รน (Incharoen and Maneechote, 2013) เปนไปในทศทางเดยวกนกบ Adibmoradi et al. (2016) ทรายงานวา การเสรมใยอาหารชนดไมละลายน�าจากแกลบขาวและเปลอกขาวบารเลย ทระดบ 12 เปอรเซนต ชวยเพมสมรรถภาพการเจรญเตบโตและการยอยไดของโปรตน รวมไปถงชวยกระตนสณฐานวทยาของวลไล แตในทางกลบกน ใยอาหารชนดละลายน�า มคณสมบตดดซบน�าไดด ท�าใหมความหนดเพมขน มลกษณะเปนเจล และไมสามารถยอยไดดวยเอม
ไซมในระบบทางเดนอาหาร แตแบคทเรยทอาศยในล�าไสใหญสามารถยอยและใชประโยชนได ใยอาหารชนดน ไดแก แพคตน (Pectin) กลโคแมนแนน (Glucomannan) กม (Gums) มวซเลจ (Mucilage) เปนตน Silva et al. (2013) พบวา อาหารไกเนอทเสรมดวยแพคตนใน ทระดบ 3-5 เปอรเซนต มผลตอการเพมความหนด และลดอตราการเคลอนทของอาหารในระบบทางเดนอาหาร รวมทงชวยเพมการใชประโยชนของพลงงานในอาหารใหดขนส�าหรบไกเนอระยะแรก แตไปมผลกระทบตออตราการยอยไดของโภชนะทลดลงในไกเนอระยะเจรญเตบโต Maisonnier et al. (2003) ท�าการทดลองเสรมกวกม (Guar gum) ในอาหารไกเนอ พบวา ความเขมขนและประสทธภาพในการหลงกรดเกลอลดลง และสงผลท�าใหอตราการยอยไดของไขมนลดลง สอดคลองกบงานวจยหลายชน ทรายงานวา ปรมาณใยอาหารชนดละลายน�าในอาหารไกทเพมสงขนมความสมพนธกบสมรรถภาพการเจรญเตบโตและอตราการยอยได ทลดลง (Maisonnier et al., 2001; Carre et al., 2002) ดวยเหตนโครงการวจยนจงมวตถประสงคเพอศกษาอทธพลของใยอาหารชนดละลายน�าในอาหารกงบ รสท ธตอสมรรถภาพการ เจ รญเตบโตและประชากรจลนทรยในใสตงของไกเนอระยะแรก โดยใชกวกม (Guar gum) เปนแหลงของใยอาหารชนดละลายน�า
648 แกนเกษตร 47 ฉบบพเศษ 2 : (2562)
ABSTRACT: The present research was to determine the effect of soluble dietary fiber in semi-purified diets on performance and cecal microbe of young broilers. Guar gum was used as source of soluble dietary fiber. Eighty 10-day-old Ross 308 chicks were used in the study. The birds were divided into 4 groups: each groups were provided with soluble dietary fiber at 0 (T1), 3 (T2), 5 (T3), and 7 (T4) % in semi-purified diets. At the end of experiment (24 d-old), body weight (17 and 24 d-old), weight gain and feed intake decreased (P<0.05) when increasing the level of soluble dietary fiber (3 to 7%). Compare to T1 group, poor feed conversion were found in T3 and T4 groups (P<0.05). For microbiological analysis, Salmonella spp. did not observed in all experimental groups, whereas similar amount of E. Coli were detected (P>0.05). In conclusion, since 3% level of soluble dietary fiber in grower diets (10 to 24 d-old) have a negative effects on feed intake and body weight. Moreover, poor feed conversion ratio was found when increasing 5-7% soluble dietary fiber in broiler diet. However, population of Salmonella spp. and E. Coli in cecal content was not affected by soluble dietary fiber.Keywords: dietary fiber, semi purified diets, performance, cecal microbial populations, broilers

KHON KAEN AGR. J. 47 SUPPL.2 : (2019). 649
วธการศกษา
การเตรยมสตวทดลองและออกแบบการทดลอง ซอลกไกเนอ สายพนธ รอส 308 (Ross 308) เพศผ อาย 1 วน จ�านวน 100 ตว จาก บรษท ซพเอฟ จ�ากด (มหาชน) ลกไกทกตวถกเลยงดภายในโรงเรอนทดลองระบบปด พนคอกปรองดวยแกลบมความหนาประมาณ 3-4 นว ปรบอณหภมในบรเวณพนทกก 32±1 องศาเซลเซยล ไกทกตวไดรบน�าสะอาดและอาหารระยะแรก (24% CP; 3,050 kcal/kg) ใหกนอยางเตมท เมออายครบ 10 วน ไกทกตวจะถกชงน�าหนก และคดเลอกไกทมน�าหนกใกลเคยงกน จ�านวน 80 ตว มาแบงออกเปน 4 กลมๆ ละ 4 ซ�าๆ ละ 5 ตว ไกแตละกลมไดรบอาหารทดลองเสรมกวกม ทระดบ 0 (T1), 3 (T2), 5 (T3) และ 7 (T4) เปอรเซนต ตามล�าดบ (Table 1) กวกมทใชในการทดลองครงนซอมาจาก หางหนสวนจ�ากด เอส พ กม แอนดเคมคอล ตลอดระยะเวลาการทดลองไกจะถกเลยงในคอกขนาด 0.72 ตารางเมตร ปรองพนดวยแกลบหนาประมาณ 2-3 นว ภายโรงเรอนระบบระเหยไอเยน (Evaporative cooling system) ควบคมอณหภมเฉลย 27-29 องศาเซลเซยส มถงใหอาหารและนปเปลใหน�าอตโนมต ควบคมโปรแกรมแสงสวางวนละ 17 ชวโมง ตลอดระยะเวลาการทดลองไกไดรบอาหารและน�าใหกนอยางเตมท (Ad libitum) ท�าการบนทกปรมาณอาหารทกน และชงน�าหนกตว ทกๆ สปดาห เพอน�าเอาขอมลทไดมาค�านวณหาสมรรถภาพการเจรญเตบโต เชน น�าหนกตวทเพมขน ปรมาณอาหารทกน และอตราการเปลยนอาหารเปนน�าหนกตว เปนตน เมอสนสดการทดลอง ทอาย 24 วน ท�าการสมไกจ�านวนกลมละ 4 ตว เพอการณยฆาต หลงจากนนท�าการเกบสงเหลอในไสตงใสในถงพลาสตก และเกบรกษาไวในถงน�าแขงระหวางการเคลอนยายตวอยางไปยงหองปฏบตการเพอน�าไปวเคราะหหาปรมาณจลนทรย การทดลองคร ง น ไ ดผานการรบรองจากคณะกรรมการก�ากบดแลการด�าเนนการตอสตวเพองานทางวทยาศาสตร มหาวทยาลยนเรศวร เอกสารรบรองเลขท 60 01 011
การตรวจหา Salmonella spp และ E. coli น� าตวอยาง ทผานการเ พมจ�านวนใน Lactose broth ไปท�าการเพมจ�านวนเฉพาะชนดแบคทเรย Salmonella spp. ในอาหารเหลว (Enrichment media) Tetrathionate (TT) broth บมเชอทอณหภม 35 ± 2.0°C นาน 24 ± 2 ชวโมง จากนนท�าการเพาะเลยงเพอตรวจนบจ�านวนบนอาหารจ�าเพาะดวยวธการ Dilution spread plate method บนอาหาร Salmonella agar โดยการทดสอบความเจอจางละ 3 ซ�า บมเชอทอณหภม 35°C นาน 24 ชวโมง และนบจ�านวนโคโลน ส�าหรบการตรวจหา E. coli โดยการน�าตวอยางทผานการเพมจ�านวนใน Lactose broth มาเพาะเลยงเชอดวยวธการเจอจางแบบล�าดบสวนและน�าไปเกลยบนจานอาหารเพาะเลยงเชอ (Dilution Spread plate method) ทจ�าเพาะ คอ Eosin Methylene Blue (EMB) agar โดยการทดสอบความเจอจางละ 3 ซ�า บมเพาะเชอทอณหภม 35°C ± 0.5 องศาเซลเซยส นาน 18-24 ชวโมง จ�านวนโคโลนของเชอทง 2 ชนดจะถกน�าไปค�านวณคาทนบไดในหนวยของ CFU/g
การวเคราะหทางสถต น�าขอมลสมรรถภาพการเจรญเตบโตและการวเคราะหจลนทรย มาวเคราะหความแปรปรวน (Analysis of variances) ตามแผนการทดลองแบบสมสมบรณ (Completely randomized design) และเปรยบเทยบความแตกตางระหวางคาเฉลยดวย Duncan’s New Multiple Range Test ทระดบนยส�าคญ P<0.05
ผลการศกษาและวจารณ
Table 2 แสดงอทธพลของใยอาหารชนดละลายน�าระดบตางๆ ในอาหารกงบรสทธตอสมรรถภาพการเจรญเตบโตของไกเนอระยะแรก จากผลการศกษาชใหเหนวา น�าหนกตว (ทอาย 17 และ 24 วน) น�าหนกตวทเพมขนเฉลย (ทอาย 10-24 วน) และอตราการกนอาหาร มคาลดลงอยางมนยส�าคญ (P<0.05) ตามระดบใยอาหารชนดละลาย

650 แกนเกษตร 47 ฉบบพเศษ 2 : (2562)
น�าทเพมสงขนตงแต 3 เปอรเซนตขนไป โดยเฉพาะอยางยงเมอใชใยอาหารชนดละลายน�า ทระดบ 5-7 เปอรเซนต พบวาอตราการเปลยนอาหารเปนน�าหนกตวมคาแยลงอยางมนยส�าคญ (P<0.05) เมอเปรยบเทยบกบกลมควบคม เปนไปในทศทางเดยวกนกบ Rahmatnejad and Saki (2016) ทไดท�าการทดสอบผลของเซลลโลสทมคณสมบตละลายน�าไดด และมความหนดสง ทเรยกวา คารบอกซเมทลเซลลโลส (Carboxymethyl cellulose) ในอาหารไกเนอ 2 และ 4 เปอรเซนต พบวา มผลท�าใหปรมาณอาหารทกน และน�าหนกตวทเพมขนมคาลดลงอยางมนยส�าคญ (P<0.05) ซงมความเปนไปไดวาผลดงกลาวมความสมพนธความหนดในระบบทางเดนอาหาร (Gastrointestinal system) ทเพมสงขน จงท�าใหอตราการไหลของอาหาร (Feed passage rate) ภายในทอทางเดนอาหารชาลง สตวจงลดอตราการกนอาหาร และสงผลใหอตราการเจรญเตบโตลดลงตามไปดวย (Shakouri et al., 2006) สอดคลองกบรายงานตางๆ ทกลาววา ปรมาณใยอาหารชนดละลายน�าในอาหารไกทเพมสงขนมความสมพนธกบสมรรถภาพการเจรญเตบโตและอตราการยอยไดทลดลง (Maisonnier et al., 2001; Carre et al., 2002) ยงไปกวานน Maisonnier et al. (2003) ไดบนทกไววา การเสรมกวกม (Guar gum) ในอาหารไกเนอ มผลท�าใหความเขมขนและประสทธภาพในการหลงกรดเกลอลดลง และสงผลท�าใหอตราการยอยไดของไขมนลดลงตามไปดวย นอกจากนนแลวจากผลการศกษาครงน คณะผวจยยงพบอกวา อาหารไกเนอกงบรสทธทใชในการทดลองมผลท�าใหสมรรถภาพการเจรญเตบโตของไกทกกลมมคาต�ากวามาตรฐานสายพนธ รอส 308 อยางเหนไดชด คาน�าหนกตวและอตราการกนอาหารของไกกลมควบคม มคาต�ากวามาตรฐานถง 40.0 และ 48.2 เปอรเซนต ตามล�าดบ ซงเปนททราบกนดอยแลววาหากปรมาณการกนอาหารลดลง มกจะสงผลกระทบโดยตรงตอน�าหนกตวทแยลงอยางหลกเลยงไมได ทงนมความเปนไปไดอยางยงวาสตวอาจสามารถใชประโยชนจากแปงขาวสาลทเปนองคประกอบหลกในอาหารกงบรสทธไดดกวาเมลดธญพชทใชประกอบสตรอาหารพน
ฐานทางการคาทวไป จงสงผลใหสตวไดรบพลงงานอยางเพยงพอและลดอตราการกนอาหารในทสด ซงคลายคลงกบการทดลองของ Rahmatnejad and Saki (2016) ทใชอาหารกงบรสทธเลยงไกเนอสายพนธ รอส 308 ทอาย 7-21 วน แลวพบวา อตราการกนอาหารมคาต�ากวามาตรฐาน คดเปน 52.0 เปอรเซนต ดานการประเมนทางจลชววทยา ไมพบเชอซลโมเนลลาในตวอยางสงเหลอของไสตงในไกทง 4 กลม ในขณะทสามารถตรวจพบเชออโคไลในปรมาณทไมแตกตางกนทางสถต (P>0.05) ดงขอมลทแสดงใน Table 3 จากผลการทดลองน ชใหเหนวา ใยอาหารชนดละลายน�าไมมผลตอการเปลยนแปลงของประชากรจลนทรยในไสตงของไกเนอระยะแรก ทงนอาจเปนผลมาจากไกอายนอยมโอกาสทไดรบเชอในสงแวดลอมไมมากนกเมอเทยบกบไกโต หรอไกทเลยงภายใตโรงเรอนระบบปดจะมโอกาสทไดรบเชอตามธรรมชาตนอยกวาไกทถกเลยงแบบปลอยตามธรรมชาต จงท�าใหผลการศกษานยงไมมความชดเจนเทาทควร แตอยางไรกตาม บทบาทหนาทของใยอาหารชนดละลายน�าเกยวของโดยตรงกบกระบวนการหมกในส�าไสสวนทายเพอผลตกรดไขมนสายสน (Van der Wielen et al., 2001) ในขณะทงานวจยในสกร ไดรายงานวา ถงแมการเพมระดบของใยอาหารชนดละลายน�าจะชวยเพมความเขมขนของกรดไขมนสายสนได แตยงมผลตอการเพมจ�านวนของเชออโคไลไดอยางรวดเรวเชนกน (Hopwood et al., 2004)
สรป
อาหารไกเนอระยะแรกทมใยอาหารชนดละลายน�า ตงแตระดบ 3 เปอรเซนต ขนไป มผลท�าใหอตราการกนอาหาร และน�าหนกตวมคาลดลง รวมทงอตราการเปลยนอาหารเปนน�าหนกตวมคาแยลงอยางเหนไดชด เมอระดบใยอาหารชนดละลายน�ามคาเพมสงขน ตงแตระดบ 5-7 เปอรเซนต อยางไรกตามระดบของใยอาหารชนดละลายน�าไมมผลตอการเปลยนแปลงของประชากรเชอซลโมเนลลาและอโคไลในไสตง
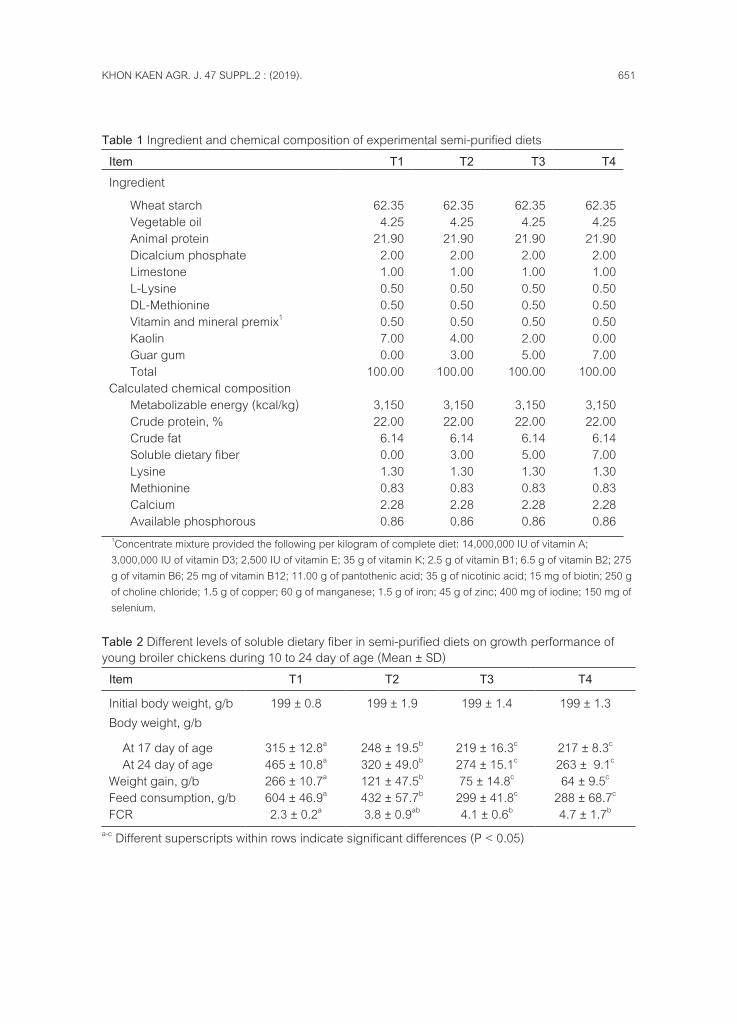
KHON KAEN AGR. J. 47 SUPPL.2 : (2019). 651
Table 1 Ingredient and chemical composition of experimental semi-purified dietsItem T1 T2 T3 T4Ingredient
Wheat starch 62.35 62.35 62.35 62.35Vegetable oil 4.25 4.25 4.25 4.25Animal protein 21.90 21.90 21.90 21.90Dicalcium phosphate 2.00 2.00 2.00 2.00Limestone 1.00 1.00 1.00 1.00L-Lysine 0.50 0.50 0.50 0.50DL-Methionine 0.50 0.50 0.50 0.50Vitamin and mineral premix1 0.50 0.50 0.50 0.50Kaolin 7.00 4.00 2.00 0.00Guar gum 0.00 3.00 5.00 7.00Total 100.00 100.00 100.00 100.00
Calculated chemical compositionMetabolizable energy (kcal/kg) 3,150 3,150 3,150 3,150Crude protein, % 22.00 22.00 22.00 22.00Crude fat 6.14 6.14 6.14 6.14Soluble dietary fiber 0.00 3.00 5.00 7.00Lysine 1.30 1.30 1.30 1.30Methionine 0.83 0.83 0.83 0.83Calcium 2.28 2.28 2.28 2.28Available phosphorous 0.86 0.86 0.86 0.86
1Concentrate mixture provided the following per kilogram of complete diet: 14,000,000 IU of vitamin A; 3,000,000 IU of vitamin D3; 2,500 IU of vitamin E; 35 g of vitamin K; 2.5 g of vitamin B1; 6.5 g of vitamin B2; 275 g of vitamin B6; 25 mg of vitamin B12; 11.00 g of pantothenic acid; 35 g of nicotinic acid; 15 mg of biotin; 250 g of choline chloride; 1.5 g of copper; 60 g of manganese; 1.5 g of iron; 45 g of zinc; 400 mg of iodine; 150 mg of selenium.
Table 2 Different levels of soluble dietary fiber in semi-purified diets on growth performance of young broiler chickens during 10 to 24 day of age (Mean ± SD)Item T1 T2 T3 T4
Initial body weight, g/b 199 ± 0.8 199 ± 1.9 199 ± 1.4 199 ± 1.3Body weight, g/b
At 17 day of age 315 ± 12.8a 248 ± 19.5b 219 ± 16.3c 217 ± 8.3c
At 24 day of age 465 ± 10.8a 320 ± 49.0b 274 ± 15.1c 263 ± 9.1c
Weight gain, g/b 266 ± 10.7a 121 ± 47.5b 75 ± 14.8c 64 ± 9.5c
Feed consumption, g/b 604 ± 46.9a 432 ± 57.7b 299 ± 41.8c 288 ± 68.7c
FCR 2.3 ± 0.2a 3.8 ± 0.9ab 4.1 ± 0.6b 4.7 ± 1.7b
a-c Different superscripts within rows indicate significant differences (P < 0.05)

652 แกนเกษตร 47 ฉบบพเศษ 2 : (2562)
ค�าขอบคณ
คณะผวจยขอขอบพระคณ กองการวจยและนวตกรรม มหาวทยาลยนเรศวร ทไดสนบสนนทนในการท�าวจยครงน ภายใตสญญารบทนอดหนนการวจยสญญาเลขท R2561B057 จากงบประมาณแผนดน (แบบปกต) มหาวทยาลยนเรศวร ประจ�าปงบประมาณ พ.ศ. 2561
เอกสารอางอง
Adibmoradi, M., B. Navidshad, and M.F. Jahromi. 2016. The effect of moderate levels of finely ground insoluble fibre on small intestine morphology, nutrient digestibility and performance of broiler chickens. Ital. J. Anim. Sci. 15:310–317.
Carre, B., A. Idi, S. Maisonnier, J. P. Melcion, F. X. Oury, J. Gomez, and P. Pluchard. 2002. Relationships between digestibilities of food components and characteristics of wheats (Triticum aestivum) introduced as the only cereal source in a broiler chicken diet. Br. Poult. Sci. 43:404–415.
Hopwood, D.E., D.W. Pethick, J.R. Pluske, and D.J. Hampson. 2004. Addition of pearl barley to a rice-based diet for newly weaned piglets increases the viscosity of the intestinal contents, reduces starch digestibility and exacerbates post-weaning colibacillosis. Br. J. Nutr. 92:419-427.
Incharoen, T. 2013. Histological adaptations of the gastrointestinal tract of broilers fed diets containing insoluble fiber from rice hull meal. Am. J. Anim. Vet. Sci. 8:79-88.
Incharoen, T., and P. Maneechote. 2013. The effects of dietary whole rice hull as
insoluble fiber on the flock uniformity of pullets and on the egg performance and intestinal mucosa of laying hens. Am. J. Agri. Biol. Sci. 8:323-329.
Maisonnier, S., J. Gomez, A. Bree, C. Berri, E. Baeza, and B. Carre. 2003. Effects of microflora status, dietary bile salts and guar gum on lipid digestibility, intestinal bile salts, and histomorphology in broiler chickens. Poult. Sci. 82:805–814.
Maisonnier, S., J. Gomez, and B. Carre. 2001. Nutrient digestibility and intestinal viscosities in broiler chickens fed on wheat diets, as compared to maize diets with added guar gum. Br. Poult. Sci. 42:102–110.
Rahmatnejad, E., and A.A. Saki. 2016. Effect of dietary fibres on small intestine histomorphology and lipid metabolism in young broiler chickens. J. Anim. Physiol. Anim. Nutr. 100:665–672.
Silva, V.K., V. de Souza Morita, and I.C. Boleli. 2013. Effect of pectin extracted from citrus pulp on digesta characteristics and nutrient digestibility in broilers chickens. R. Bras. Zootec. 42:575-583.
Shakouri, M., H. Kermanshahi, and M. Mohsenzadeh. 2006. Effect of different non starch polysaccharides in semi purified diets on performance and intestinal microflora of young broiler chickens. Int. J. Poult. Sci. 5:557–561.
Van der Wielen, P.W.J.J., S. Biesterveld, L.J.A. Lipman, and F.van Knapen. 2001. Inhibition of a glucose-limited sequencing fed-batch culture of Salmonella enterica serovar enteritidis by volatile fatty acids representative of the ceca of broiler chickens. Appl. Environ. Microbiol. 67:1979–1982.
Table 3 Different levels of soluble dietary fiber in semi-purified diets on cecal microbial populations of young broiler chickens at 24 day of age (Mean ± SD)
Item (cfu/g) T1 T2 T3 T4Salmonella spp. ND ND ND NDE. Coli 6.5 ± 0.6 6.2 ± 0.4 6.2 ± 0.6 6.3 ± 1.0
ND = not detected

Agricultural Sci. J. 50 : 1 (Suppl.) : 361-366 (2019) ว. วทย. กษ. 50 : 1 (พเศษ) : 361-366 (2562)
361
การคดแยกแอคตโนมยซทจากดนรอบรากขาวทสามารถยบยงเชอรากอโรคใบไหมในขาวโพด
Isolation of Rice Rhizospheric Actinomycetes Against Fungal Corn Leaf Blight Disease
นชนาถ เกตวงษ1 และ พรพรรณ รตนะสจจะ1*
Nutchanach Gadsuvong1 and Pornpan Ruttanasutja1*
160 หม 3 สาขาวชาวทยาศาสตร คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม พระนครศรอยธยา 13000 160 Moo 3 Major of Science, Faculty of Science and Technology, Rajamangala University of Technology Suvarnnabhumi
13000, Thailand *Corresponding author: [email protected]
บทคดยอ
การศกษาวจยในครงนมวตถประสงคเพอศกษาคดแยกแอคตโนมยซทจากดนบรเวณรอบรากขาวทมความสามารถใน
การยบยงเชอรากอโรคใบไหมในขาวโพด ในเขตพนทตาบลบานกระทมและหวเวยง อาเภอเสนา จงหวดพระนครศรอยธยา
แอคตโนมยซททงหมดจานวน 38 ไอโซเลท ถกคดแยกไดจากตวอยางดนในตาบลบานกระทม (จานวน 16 ไอโซเลท) และ
บานหวเวยง (จานวน 22 ไอโซเลท) ซงแอคตโนมยซททคดแยกทงหมดจดอยในสกล Streptomyces ทมลกษณะสณฐาน
วทยาและสรรวทยาทแตกตางกน เมอนาไอโซเลททงหมดมาทดสอบความสามารถในการยบยงเชอรากอโรคในขาวโพด
(Bipolaris maydis) ดวยวธ Dual culture มเพยงจานวน 9 ไอโซเลท (คดเปน 23.68 เปอรเซนต) สามารถยบยงการเจรญ
ของเชอราทดสอบได โดยมผลการยบยงอยในชวง 40 ถง 80 เปอรเซนต แอคตโนมยซทไอโซเลททมประสทธภาพดทสด
จานวน 3 ไอโซเลท คอ HVM3-2, HVM4-1 และ HVM5-1 โดยมผลการยบยงเชอราทดสอบเทากบ 80, 74 และ 68
เปอรเซนต ตามลาดบ
คาสาคญ: จลนทรยในดน, การควบคมโรคโดยชววธ, โรคใบไหมในขาวโพด
ABSTRACT
The objective of this research was to isolated actinomycetes from rice rhizosphere soil, with the
ability to inhibited fungal corn leaf blight disease, in Hua Wiang and Ban Krathum Sub-district, Sena District,
Phra Nakhon Si Ayutthaya Province. A total of 38 isolates were obtained from rice rhizosphere soil samples
in Ban Krathum (16 isolates) and Hua Wiang Sub-district (22 isolates). All of the isolates were a member in
genus Streptomyces with the difference in morphology and physiology characteristics. All the isolates were
tested for the efficiency to corn pathogenic fungi inhibition (Bipolaris maydis) by dual culture method. Only
9 isolates (23.68 percent) were showed the ability to pathogenic fungi inhibition, which the inhibition results
in the range of 40 to 80 percent. The three potent actinomycetes isolates were HVM3-2, HVM4-1 and HVM5-
1 with the inhibition results as 80, 74 and 68 percent, respectively.
Keywords: Soil microorganisms, bio-control, leaf blight disease in corn

การคดแยกแอคตโนมยซท ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
362
บทนา
ขาวโพด (Maize หรอ Corn) มชอทางวทยาศาสตร คอ Zea mays L. เปนกลมของพชอาหารทถกใชเปนแหลง
ของอาหารสตวเปนหลก ขอมลป 2561 ประเทศไทยมผลผลตขาวโพดเลยงสตวสงออก 82,428 ตน คดเปนมลคา 685.42
ลานบาท การเพาะปลกขาวโพดเลยงสตวภายในประเทศสงถง 6,579,194 ไร และใหผลผลตประมาณ 733 กโลกรมตอไร
(Centre for Agricultural Information, 2018) แตสถานการณการผลตขาวโพดเลยงสตวของโลกป 2562/63 คาดวาม
จานวนเพยง 1,099.19 ลานตน ลดลงจากป 2561/62 คดเปน 1.90 เปอรเซนต (Vorahan and Kumtong, 2019) ทงน
ปจจยทสงผลตอการเพาะปลกและปรมาณผลผลตขาวโพดทสาคญ คอโรคในขาวโพด อนมสาเหตหลกมาจากจลนทรยกอโรค
โดยเฉพาะอยางยงกลมเชอราสาเหตของโรคขาวโพดสาคญหลายชนด เชน โรครานาคาง (Downy mildew) โรคสมทหรอรา
เขมาสดา (Smut) โรคราสนม (Rust) โรคใบจด (Leaf spot) โรคใบไหมแผลใหญยาวร (Northern corn leaf blight)
รวมถงโรคใบไหมแผลเลก (Southern corn leaf blight) ทมสา เหตจากเ ชอรา Bipolaris maydis (ม ชอ เดมว า
Helminthosporium maydis) (Samphantharak, 1994) ซงพบการระบาดของโรคทาลายผลตผลขาวโพดเลยงสตวใน
ประเทศไทยทกๆ ป และมแนวโนมเพมมากขนในปจจบน ดงนนเกษตรกรมกใชวธควบคมโรคขาวดวยการใชสารเคม แตกพบ
ปญหาผลกระทบตอการตกคางของสารเคมในผลผลตและสขภาพของเกษตรกรผใช ทงยงมราคาแพงและเปนพษตอ
สงแวดลอม การควบคมโรคพชดวยชววธเปนอกหนงทางเลอกทถกนามาใชแกปญหา โดยเฉพาะอยางยงการใชจลนทรยในการ
ควบคมโรคในขาวโพดและพชทางการเกษตรอกหลายชนด
การศกษาวจยดานการควบคมโรคดวยการใชจลนทรยมมาอยางตอเนอง อาท การใชเชอราในยบยงรากอโรคพช การ
ใชแบคทเรยยบยงรากอโรคขาว (Rahman et al., 2018) รวมถงแบคทเรยกลมแอคตโนมยซททมความสามารถในการสราง
สารออกฤทธทางชวภาพไดหลายชนด ซงสงผลตอการยบยงจลนทรยกอโรคพชชนดอน รวมถงโรคพชทมสาเหตมาจากเชอรา
(Usuwan, 2018, Newitt et al., 2019) การศกษาวจยในครงนมงเนนการคดแยกแอคตโนมยซทจากตวอยางดนรอบรากขาว
ทมความสามารถในการยบยงจลนทรยกอโรคใบไหมในขาวโพดรวมกบการคดเลอกแอคตโนมยซททมประสทธภาพสงทสดใน
การยบยงการเจรญของจลนทรยกอโรคใบไหมในขาวโพดในระดบหองปฏบตการ เพอนาไปพฒนาและประยกตใชไดจรงใน
อนาคต
อปกรณและวธการ
การคดแยกแอคตโนมยซทจากตวอยางดนรอบรากขาว
เกบตวอยางดนรอบรากขาวในแปลงนาขาวในพนท 2 ตาบล ไดแก ตาบลบานกระทมและตาบลหวเวยง อาเภอเสนา
จงหวดพระนครศรอยธยา นามาผงใหแหงทอณหภมหองและคดแยกแอคตโนมยซทดวยวธการ Dilution spread plate บน
อาหาร Starch Casein Agar (SCA) บมทอณหภม 30 องศาเซลเซยส นาน 7–14 วน สงเกตการเจรญและนบจานวนโคโลน
แอคตโนมยซท ทาการจดกลมแอคตโนมยซทไอโซเลทและนามาทาใหบรสทธบนอาหาร SCA
การศกษาลกษณะสณฐานวทยาและการการจดจาแนกแอคตโนมยซท
ศกษาสณฐานวทยาของโคโลนและการสรางสของเสนใย 2 ชนดคอ เสนใยทเจรญในอาหาร (substrate mycelium)
และเสนใยชเหนอผวหนาอาหาร (aerial mycelium) บนอาหารเฉพาะ International Streptomycetes Project media
(ISP) ไดแก ISP2,ISP3 และ ISP6 บมทอณหภม 30 องศาเซลเซยส เปนเวลา 7 วน บนทกสเสนใยอาหารทง 2 ชนด และการ
สรางสเมลานน (Shirling and Gottlieb, 1996)
ศกษาลกษณะการสรางสปอรดวยวธการปกสไลด (immersed slide technique) บนอาหารแขง ISP2 (Holt, et
al., 1994) บมเชอทอณหภม 30 องศาเซลเซยส เปนเวลา 7 วน และนาสไลดทไดสองดรปแบบการสรางสปอรภายใต
กลองจลทรรศนแบบใชแสง (Light Microscope)

การคดแยกแอคตโนมยซท ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
363
ศกษาองคประกอบไอโซเมอรของกรด diaminopimelic (DAP) บนผนงเซลล โดยใชวธดดแปลงจาก Hasegawa
et.al. (1983) และนาไปทาการตรวจสอบดวยการทาโครมาโตกราฟฟแบบกระดาษ (paper chromatography) ในตวทา
ละลาย methanol : H2O : 10 N HCl : pyridine (80 : 17.5 : 2.5 : 10 (v/v)) และบนทกผลทไดกบสารละลายมาตรฐาน
การทดสอบความสามารถในการสรางสารยบยงเชอรากอโรคใบไหมในขาวโพด (Bipolaris maydis)
เพาะเลยงเชอราทดสอบ Bipolaris maydis บนอาหาร Potato Dextrose Agar (PDA) และเพาะเลยงแอคตโน-
มยซทบนจานอาหารแขง SCA เปนเวลา 7 วน ทอณหภม 30 องศาเซลเซยส จากนนทาการทดสอบการยบยงดวยวธ Dual
Culture บนอาหาร Nutrient Agar (NA) บมทอณหภม 30 องศาเซลเซยส นาน 5 วน บนทกผลและนามาคานวณเปอรเซนต
การยบยงการเจรญของเชอราทดสอบ
ผลการทดลองและวจารณ
การคดแยกแอคตโนมยซทจากตวอยางดนรอบรากขาว
ขอมลทางกายภาพตวอยางดนรอบรากขาวสวนใหญเปนดนเหนยว 75 เปอรเซนต และดนรวน 25 เปอรเซนต มคา
ความเปนกรด–ดาง (pH) อยในชวง 7.1–7.9 ตวอยางดนรอบรากพชทงหมดจานวน 16 ตวอยาง (Table 1) นบจานวนโคโลน
แอคตโนมยซทอยในชวง 3.0–5.4 x 105 CFU/g ซงจดกลมตามลกษณะสณฐานโคโลนทเจรญบนจานอาหารทงหมดรวม 38
ไอโซเลท โดยแบงเปน 1) ตาบลบานกระทมจานวน 16 ไอโซเลท และ2) ตาบลหวเวยง จานวน 22 ไอโซเลท (Table 1) ซง
ผลไอโซเลททแยกไดมจานวนนอยเมอเปรยบเทยบกบการศกษากอนหนานของ Usuwan (2018) ทแยกแอคตโนมยซท
ไอโซเลทจากดนในนาขาวจงหวดนครปฐม (107 ไอโซเลท) และจงหวดสพรรณบร (136 ไอโซเลท) อกทง Detraksa and
Surawattanakij (2018) ทแยกแอคตโนมยซทจากตวอยางดนนาขาวในจงหวดลพบรไดจานวน 116 ไอโซเลท เนองจากความ
แตกตางทางกายภาพของดนและสภาพแวดลอมทเกบมผลตอจานวนของแอคตโนมยซทนบไดและความหลากหลายของไอโซ
เลททแยกได (Punngram, et al., 2011) อกทงวธการทใชในการผงตวอยางดนใหแหง หรอการเลอกใชอาหารสาหรบเพม
จานวนแอคตโนมยซททตองการและลดจลนทรยกลมทไมตองการกอนนาไปเพาะเลยง รวมถงชนดของอาหารแขงทใชในการ
เพาะ เลยงลวนมผลตอการเจรญและจานวนแอคตโมมยซททแยกได (Dejnantarad, 2003, Wongcharone, 2004)
การศกษาลกษณะทางสณฐานวทยาและการศกษาการจดจาแนกแอคตโนมยซท
ผลของสณฐานวทยาโคโลนของแอคตโนมยซททง 38 ไอโซเลท การสรางสของเสนใย substrate และ aerial
mycelium บนอาหาร ISP2 และ ISP3 มความแตกตางกน และพบเพยง 6 ไอโซเลท ทมการสรางสารสเมลานนไดบนอาหาร
ISP6 คอ BTM4–1, BTM4–2, BTM7, BTM10–3, HVM6–2 และ HVM7–3 ทงนพบมการสรางสปอรตางกน 5 แบบ คอ
1) Open spirals (จานวน 6 ไอโซเลท) 2) closed spirals (จานวน 7 ไอโซเลท) 3) straight (จานวน 6 ไอโซเลท) 4) open
loops primitive spirals hooks (จานวน 5 ไอโซเลท) และ 5) flexous (จานวน 14 ไอโซเลท) อกทงผลการศกษา
องคประกอบของผนงเซลลพบวาทงหมดเปนชนด LL–DAP จากผลการศกษาลกษณะทางสณฐานวทยาดงกลาวสามารถจด
จาแนกเบองตนของแอคตโนมยซทไอเซเลททงหมดทแยกไดจดอยในสมาชกจนส Streptomyces (Holt et al., 1994)
การทดสอบความสามารถในการสรางสารยบยงเชอรากอโรคใบไหมในขาวโพด (Bipolaris maydis)
ผลทดสอบความสามารถในการยบยงเชอรากอโรคใบไหมในขาวโพด Bipolaris maydis ดวยวธ Dual culture พบ
จานวนเพยง 9 ไอโซเลท (23.68 เปอรเซนต) ทแสดงการยบยงเชอราทดสอบ (Table 2) ไดแก BTM1, BTM3–1, BTM4–2,
BTM5–1, HVM3–2, HVM4–1, HVM5–1, HVM5–3 และ HVM5–6 ซงมการยบยงอยในชวง 40–80 เปอรเซนต โดย ไอ
โซเลททมประสทธภาพในการยบยงสงสดจานวน 3 อนดบ คอ HVM3-2, HVM4-1 และ HVM5-1 มการยบยงเทากบ 80, 74
และ 68 เปอรเซนต ตามลาดบ เนองจากงานวจยเกยวกบการศกษาแอคตโนมยซททสามารถยบยงเชอรากอโรคใบใหมใน
ขาวโพดนนพบนอยมาก ดงนนจงเทยบผลการวจยกบงานวจยทใกลเคยงกน พบวาผลการยบยงดงกลาวสอดคลองกบงานวจย

การคดแยกแอคตโนมยซท ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
364
ของ Elamvazhuthi and Subramanian (2013) การคดแยกแอคตโนมยซทจากตวอยางดนนาขาวในประเทศอนเดยทม
ความสามารถในการยบยงเชอรากอโรคในพช 4 ชนด โดยพบมการยบยงเชอรา Helminthosporium oryzae คดเปน 33.33
เปอรเซนต และงานวจยของ Usuwan (2018) แอคตโนมยซททแยกไดจากดนนาขาวสามารถยบยงรากอโรคไหมในขาว
Pyricularia oryzae มคาการยบยงอยในชวง 54–94 เปอรเซนต โดยแอคตโนมยซททแยกไดจากดนรอบตนกลาเจรญเตมท
(20–30 วน) มคาการยบยงสงกวาระยะอน ทงนสอดคลองกบผลการทดลองในครงนในการแยกแอคตโนมยซทจากดนรอบ
รากขาวทมอาย 1 เดอน และใหผลการยบยงอยในชวงทใกลเคยงกน นอกจากน Detraksa and Surawattanakij (2018) พบ
แอคตโนมยซทเพยง 5 ไอโซเลท ทแยกไดจากดนนาขาวมการยบยงเชอรา Curvularia lunata สาเหตของโรคเมลดดางใน
ขาวทมากกวา 80 เปอรเซนตขนไป ซงผลดงกลาวแสดงใหเหนวาแอคตโนมยซททแยกไดจากตวอยางดนนาขาวม
ความสามารถยบยงรากอโรคไดหลายกลม โดยเฉพาะอยางยงแอคตโนมยซทจนส Streptomyces ทพบการยบยงแบคทเรย
และรากอโรคในพชหลายชนด (Lu et al., 2008) รวมถงรากอโรคในเมลดขาวโพดเลยงสตวทสาคญ (Bressan, 2003) โดยม
กลไกการยบยงดงกลาวในธรรมชาตทเกดขนในสภาวะการดารงชวตแบบแกงแยงแขงขนในการอยรอดของจลนทรย เชน การ
สรางสารบางชนดทมฤทธในการยบยงหรอทาลายจลนทรยกลมอน การแยงใชสารอาหารทจาเปนสงผลใหจลนทรยกลมอนไม
สามารถเจรญได เปนตน (Fangfuk, 2003, Zarandi et al., 2009) จากการศกษาวจยในครงนสามารถนาขอมลทไดไปพฒนา
ตอยอด เพอใชเปนแนวทางในการแกไขปญหาโรคใบไหมใหกบเกษตรกรผปลกขาวโพดและยงชวยลดการใชสารเคมในการ
กาจดศตรพชใหกบเกษตรกรชาวนาไดในอนาคต
สรป
การคดแยกแอคตโนมยซทจากตวอยางดนรอบรากขาวในเขตอาเภอบานกระทมและหวเวยง จงหวดพระนครศร-
-อยธยา ไดจานวนไอโซเลทรวม 38 ไอโซเลท ทมความแตกตางกนทงทางดานสณฐานโคโลน การสรางสารเมลานน รปแบบ
การสรางสปอร และองคประกอบของผนงเซลล ทงนสามารถจดจาแนกเบองตนอยในสมาชกจนส Streptomyces เมอนาไป
ทดสอบความสามารถในการยบยงรอกอโรคใบไหมในขาวโพดมเพยงจานวน 9 ไอโซเลท ทมการยบยงในชวง 40–80
เปอรเซนต และไอโซเลททมประสทธภาพในการยบยงสงสด 3 อนดบ ไดแก HVM3-2, HVM4-1 และ HVM5-1 มคาการ
ยบยงราทดสอบเทากบ 80, 74 และ 68 เปอรเซนต ตามลาดบ ซงผลทไดจากการศกษาในครงนจะเปนประโยชนตอการนาไป
ตอยอดและประยกตใชในการควบคมรากอโรคขาวโพด เพอลดความเสยหายของผลผลตขาวโพดและลดการใชสารเคมในการ
กาจดรากอโรค
กตตกรรมประกาศ
ขอขอบคณกรมวชาการเกษตรทอนเคราะหเชอรากอโรคใบไหมในขาวโพด และขอขอบคณสาขาวชาวทยาศาสตร
คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ทอนเคราะหสถานท และสนบสนนดานวสด
อปกรณ และเครองมอวทยาศาสตร สาหรบการศกษาวจยในครงน
เอกสารอางอง
Bressan, W. 2003. Biological control of maize seed pathogenic fungi by use of actinomycetes. BioControl.
48: 233-240.
Centre for Agricultural Information. 2018. Thailand foreign agricultural trade statistics 2018. Bangkok: Ministry
of Agriculture and Cooperatives, Office of Agricultural Economics.
Detraksa, J. and S. Surawattanakij. 2018. Isolation of actinomycetes with inhibitory activity against Curvularia
lunata causing dirty panicle disease in rice. Agricultural Sci. J. 49(2)(Suppl.): 201-204.

การคดแยกแอคตโนมยซท ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
365
Dejnantarad, T. 2003. Isolation of anti-fungal producing actinomycetes and studying the relationship by
polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) and ribotyping.
MS Thesis, Kasetsart University, Bangkok. (in Thai)
Elamvazhuthi and Subramanian. 2013. Antagonistic activity of actinomycetes from jeyporepaddy soils
against selective phytopathogenic fungi. J. Modern. Biotechnol. 2(3):66-72.
Fangfuk, W. 2003. Selection of Endophytic actinomycetes from rice for control of blast disease.
MS Thesis, Chiang Mai University, Chiang Mai. (in Thai)
Hasegawa, T., M. Takizawa, and S. Tanida. 1983. A rapid analysis for chemical grouping of aerobic
actinomycetes. J. Gen. Appl. Microbiol. 29: 319-322.
Holt, J.G., N.R. Krieg, P.H.A. Sneath, J.T. Staley and S.T. Williams. 1994. Bergey’s Manual of Determinative
Bacteriology 9th edition. Williams&Wilkins, A Wavery Company, USA.
Khamna, S., K. Yokata, J.F. Pebery and S. Lumyong. 2009. Antifungal activity of Streptomyces spp. Isolated
from rhizosphere of Thai medical plants. Int. J. Integr. Biol. 6: 143-147.
Lu, C.G., W.C. Liu, J.Y. Qiu, H.M. Wang, T. Liu and D.W. Liu. 2008. Identification of an antifungal metabolite
produced by a potential biocontrol Actinomyces strain A01. Braz. J. Microbiol. 39: 701-707
Newitt, J.T., S.M.M. Prudence, M.I. Hutchings and S.F. Worsley. 2019. Review: Biocontrol of cereal crop
disease using Streptomycetes. Pathogens. 8: 78-102.
Punngram, N., A. Thamchaipenet and K. Duangmal. 2011. Actinomycetes from rice field soil and their
activities to inhibit rice fungal pathogen. Proceedings of 49th Kasetsart University Annual Conference:
Science. (1-4 February 2011): 234-241.
Rahman, S.F.S.A., E. Singh, C.M.J. Pieterse and P.M. Schenk. 2018. Emerging microbial biocontrol
strategies for plant pathogen. Plant Sci. 267: 102-111.
Samphantharak, K. 1994. Agronomy. Thai Watana Panich Co., Ltd. Bangkok. (in Thai)
Shirling E.B. and D. Gottlieb. 1966. Methods for characterization of Streptomyces species. Int. J. Syst.
Bacteriol. 16(3): 313-340.
Usuwan, P. 2018. Biocontrol of rice blast disease by Streptomyces spp. Agricultural Sci. J. 49(1)(Suppl.): 645-
649.
Vorahan, V. and P. Kumtong. 2562. Maize. J. Agric. Econ. 65(752): 25.
Wongcharone, A. 2004. Polyphasic taxonomy of Streptomyces spp., antagonistic to phytopathogenic
bacteria. MS Thesis, Khon Kaen University, Khon Kaen. (in Thai)
Zarandi, M.E., G.H.S. Bonjar, F.P. Dehkaei, S.A.A. Moosavi, P.R. Farokhi and S. Aghighi. 2009. Biological control
of rice blast (Magnaporthe oryzae) by use of Streptomyces sindeneusis isolate 263 in
greenhouse. Am. J. Appl. Sci. 6(1): 194-199.
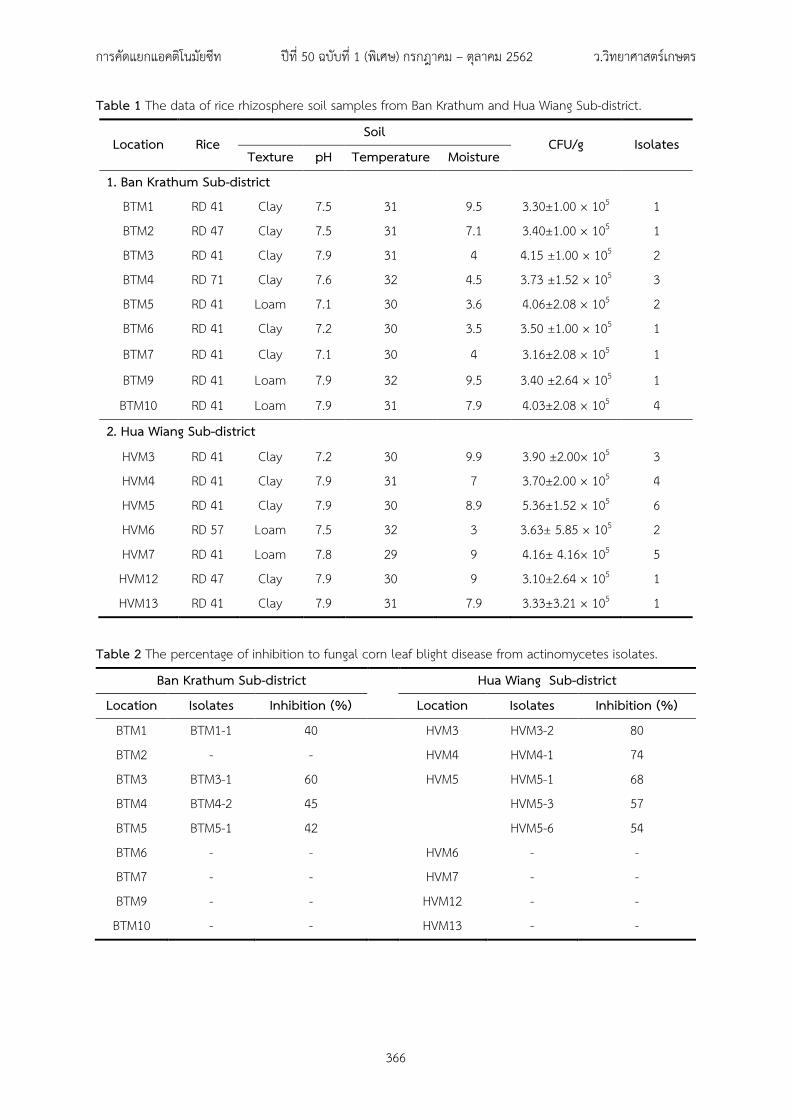
การคดแยกแอคตโนมยซท ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
366
Table 1 The data of rice rhizosphere soil samples from Ban Krathum and Hua Wiang Sub-district.
Location Rice Soil
CFU/g Isolates Texture pH Temperature Moisture
1. Ban Krathum Sub-district
BTM1 RD 41 Clay 7.5 31 9.5 3.30±1.00 × 105 1
BTM2 RD 47 Clay 7.5 31 7.1 3.40±1.00 × 105 1
BTM3 RD 41 Clay 7.9 31 4 4.15 ±1.00 × 105 2
BTM4 RD 71 Clay 7.6 32 4.5 3.73 ±1.52 × 105 3
BTM5 RD 41 Loam 7.1 30 3.6 4.06±2.08 × 105 2
BTM6 RD 41 Clay 7.2 30 3.5 3.50 ±1.00 × 105 1
BTM7 RD 41 Clay 7.1 30 4 3.16±2.08 × 105 1
BTM9 RD 41 Loam 7.9 32 9.5 3.40 ±2.64 × 105 1
BTM10 RD 41 Loam 7.9 31 7.9 4.03±2.08 × 105 4
2. Hua Wiang Sub-district
HVM3 RD 41 Clay 7.2 30 9.9 3.90 ±2.00× 105 3
HVM4 RD 41 Clay 7.9 31 7 3.70±2.00 × 105 4
HVM5 RD 41 Clay 7.9 30 8.9 5.36±1.52 × 105 6
HVM6 RD 57 Loam 7.5 32 3 3.63± 5.85 × 105 2
HVM7 RD 41 Loam 7.8 29 9 4.16± 4.16× 105 5
HVM12 RD 47 Clay 7.9 30 9 3.10±2.64 × 105 1
HVM13 RD 41 Clay 7.9 31 7.9 3.33±3.21 × 105 1
Table 2 The percentage of inhibition to fungal corn leaf blight disease from actinomycetes isolates.
Ban Krathum Sub-district Hua Wiang Sub-district
Location Isolates Inhibition (%) Location Isolates Inhibition (%)
BTM1 BTM1-1 40 HVM3 HVM3-2 80
BTM2 - - HVM4 HVM4-1 74
BTM3 BTM3-1 60 HVM5 HVM5-1 68
BTM4 BTM4-2 45 HVM5-3 57
BTM5 BTM5-1 42 HVM5-6 54
BTM6 - - HVM6 - -
BTM7 - - HVM7 - -
BTM9 - - HVM12 - -
BTM10 - - HVM13 - -

Agricultural Sci. J. 50 : 1 (Suppl.) : 313-318 (2019) ว. วทย. กษ. 50 : 1 (พเศษ) : 313-318 (2562)
313
การศกษาประสทธภาพของสารสกดหยาบจากพชในทองถนเพอยบยงแบคทเรยกอโรคพช
Xanthomonas campestris pv. campestris
Study on the Efficiency of Local Plant Crude Extract against Plant Pathogenic Bacteria
Xanthomonas campestris pv. campestris
กญญา กอแกว1* สรรตน ทรพยสวาง1 และ ดารานย รบเมอง1
Kanya Kokaew1*, Sureerat Subsawang1 and Daranai Robmuang1
160 หม 3 สาขาวชาวทยาศาสตร คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม พระนครศรอยธยา 13000 160 Moo 3 Science division, Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi 13000,
Thailand
*Corresponding author: [email protected]
บทคดยอ
งานวจยนเปนการศกษาประสทธภาพการยบยงเ ชอแบคทเรยกอโรคพช Xanthomonas campestris pv.
campestris ของสารสกดหยาบจากพชทองถน 4 ชนด ไดแก ขเหลก ยานาง บวบก และโคกกระออม ทสกดดวยตวทาละลาย
เอทานอล ความเขมขน 95 เปอรเซนต และนา ผลการทดสอบพบวาสารสกดหยาบของใบยานางทสกดดวยเอทานอล ใหผล
การยบยงไดดทสด ตามดวยสารสกดใบยานางดวยนา สารสกดดอกขเหลกดวยเอทานอล และสารสกดใบขเหลกดวยนา ม
คาเฉลยเสนผานศนยกลางบรเวณยบยงเทากบ 3.66, 3.00, 1.83 และ 1.05 มลลเมตร ตามลาดบ สวนสารสกดจากบวบกและ
โคกกระออมไมสามารถยบยงเชอทดสอบได เมอนาสารสกดใบยานางดวยเอทานอล ซงมประสทธภาพในการยบยงเชอกอโรค
ทดสอบไดดทสดมาหาคาความเขมขนตาสดทสามารถยบยงเชอได (MIC) พบวามคาเทากบ 6.25 และมคาความเขมขนตาสดท
สามารถฆาเชอได (MBC) เทากบ 6.25 มลลกรมตอมลลลตร จากผลการศกษาครงนพบวาสารสกดใบยานางทสกดดวยเอทา
นอลมประสทธภาพสงทสดในการยบยง Xanthomonas campestris pv. campestris ใบยานางจงเปนพชทสามารถนาไป
ศกษาพฒนาเพอใชสารสกดจากพชแทนการใชสารเคมในการกาจดโรคพชไดตอไป
คาสาคญ: แบคทเรยกอโรคพช, Xanthomonas campestris, สารสกดหยาบจากพช
ABSTRACT
This research was studied on the efficiency against plant pathogenic bacteria Xanthomonas
campestris pv. campestris of crude extracts from four local plants, Siamese cassia (Cassia siamea), Bamboo
grass ( Tiliacora triandra) , Asiatic pennywort ( Centella asiatica) and Heart pea ( Cardiospermum
halicacabum) with 95% ethanol and water as solvents. The result showed that ethanol crude extract from
leaves of Bamboo grass exhibited the highest activity against the bacteria, followed by water crude extract
from leaves of Bamboo grass, ethanol crude extract from flowers of Siamese cassia and water crude extract
from leaves of Siamese cassia with the inhibition zone of 3.66, 3.00, 1.83 and 1.05 mm, respectively while
crude extract from Asiatic pennywort and Heart pea had no effect. The crude extract with antagonistic
effect, the leaves of Bamboo grass extracted with ethanol, showed the MIC was 6.25 and the MBC was 6.25

การศกษาประสทธภาพ ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
314
mg/ ml. The results of this study showed that the ethanol crude extract from leaves of Bamboo grass
exhibited the highest activity against Xanthomonas campestris pv. campestris. The leaves of Bamboo grass
has potential to be used as plant pathogens biocontrol instead of some currently conventional chemical
agents.
Keyword: Plant pathogenic bacteria, Xanthomonas campestris, Plant crude extracts
บทนา
โรคเนาดา (black rot) เปนโรคทมกพบแพรระบาดในพชตระกลกะหลา (Cruciferae) เชน กะหลาปล คะนา
ผกกาดกวางตง ผกกาดขาว เปนตน อาการเรมจากรอยโรคมสเหลองรปตววบรเวณขอบใบลกลามไปจนถงเสนใบมสดา สาเหต
เกดจากการตดเชอแบคทเรย Xanthomonas campestris pv. campestris ซงมลกษณะเซลลรปทอนสน ตดสแกรมลบ
และโคโลนสเหลองบนอาหาร NA การเกดโรคสามารถพบการตดเชอไดทงโครงสรางเมลด ลาตน และใบ หากมการตดโรคจะ
สงผลใหแพรกระจายเชอไปทงแปลงเพาะปลก ทาใหผลผลตเสยหายไมสามารถจาหนายได สาหรบวธการควบคมโรคนนทาได
ยาก ซงการใชสารเคมในการควบคมโรคยงใหประสทธภาพตา ดงนนการแพรระบาดของโรคนยงคงเปนปญหาสาคญตอการ
เพาะปลกพชกลมน (Bila et al., 2013) โดยเฉพาะในปจจบนผบรโภคหนมาสนใจบรโภคสนคาทางการเกษตรทมการ
เพาะปลกแบบเกษตรอนทรยมากขน เนองจากตระหนกถงสารพษทตกคางในสนคา เกษตรกรผเพาะปลกจงตองใชปยชวภาพ
และสารกาจดศตรพชในรปแบบสารชวภาพจากธรรมชาต ดงนนการพฒนาสารสกดจากพชเพอนามาใชในการปองกนศตรพช
เรมมการศกษาเพมมากขน โดยมรายงานการศกษาสารสกดจากใบยหรา ใบกระเพรา ใบตาแยแมว และตนลกใตใบ ทสกดดวย
เอทานอล และนา มฤทธในการยบยงเชอ Xanthomonas campestris สาเหตโรคพชได (Britto et al., 2011; Kumar et
al., 2018) ซงพชเหลานเปนพชทพบโดยทวไปในไทย นอกจากนยงมพชสมนไพรอกหลายชนดในไทยทมรายงานพบสารมฤทธ
ในการยบยงเชอจลนทรยได เชน ยานาง ขเหลก บวบก และโคกกะกออม (Suresh et al.,2012; Abdulrasheed et al.,
2015; Nasution et al.,2018; Rahman et al., 2017) พชเหลานเปนพชสมนไพร และใชเปนพชอาหาร พบไดทวไปใน
ทองถน ดงนนการศกษาครงนมวตถประสงคศกษาฤทธสารสกดหยาบจากพชทองถน จ.พระนครศรอยธยา จานวน 4 ชนด
ไดแก ใบยานาง ใบและดอกขเหลก ใบบวบก และตนโคกกะกออม ในการยบยงแบคทเรย Xanthomonas campestris pv.
campestris สาเหตโรคเนาดา เพอนาไปใชเปนขอมลพนฐานในการพฒนาสารสกดใหมประสทธภาพทดแทนการใชสารเคม
ควบคมโรคพช
อปกรณและวธการ
การสกดสารจากตวอยางพช
เกบตวอยางพชในทองถน ไดแก 1)ขเหลก (Cassia siamea) ใชโครงสรางดอกและใบ 2)ใบยานาง (Tiliacora
triandra) ใชโครงสรางใบ 3)ใบบวบก (Centella asiatica) ใชโครงสรางใบ และ4)โคกกระออม (Cardiospermum
halicacabum) ใชทกสวนของพชในการสกดสาร วธการสกดดดแปลงจาก Burakorn and Praphruet (2012) โดยนาพชมา
ลางทาความสะอาด หนชนสวนใหมขนาดเลก อบแหงทอณหภม 50 องศาเซลเซยส ระยะเวลา 2 วน จากนนนามาปนให
ละเอยด สกดสารโดยใชพชอบแหง 100 กรม เตมตวทาละลาย 2 ชนด ไดแก เอทานอลความเขมขน 95 เปอรเซนต และนา
กลน ในอตราสวน 1:8 (พชตอตวทาละลาย) บมเขยาท 150 รอบตอนาท ระยะเวลา 3 วน กรองแยกสารสกดออกจากตวอยาง
พช ระเหยตวทาละลายออกดวยเครอง vacuum evaporator ทอณหภม 50 องศาเซลเซยส ชงนาหนกสารสกดแลวนาไป
คานวณหาเปอรเซนตสารสกดหยาบทได ละลายสารสกดดวยตวทาละลายทใชสกดใหไดความเขมขนเทากบ 100 มลลกรมตอ

การศกษาประสทธภาพ ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
315
มลลลตร ทาสารสกดใหปลอดเชอดวยการกรอง เกบสารสกดในขวดสชาทอณหภม 4 องศาเซลเซยส เพอนาไปศกษาฤทธใน
การยบยงเชอตอไป
การทดสอบประสทธภาพการยบยงการเจรญของแบคทเรยดวยวธ Agar disc diffusion (Srisukong et al., 2016)
เพาะเลยงเชอ X. campestris pv. campestris TISTR2065 (สถาบนวจยวทยาศาสตรและเทคโนโลยแหงประเทศ
ไทย) ในอาหาร NA บมทอณหภม 30 องศาเซลเซยส 24 ชวโมง จากนนถายเชอลงในอาหาร NB บมทอณหภม 30 องศา
เซลเซยส 24 ชวโมง ปรบคาความขนของเซลลเรมตน (OD600 = 0.1) เกลยเชอใหทวบนผวหนาอาหาร NA หยดสารสกด
ปรมาตร 50 ไมโครลตร ลงบน paper disc ปลอดเชอขนาดเสนผาศนยกลาง 6 มลลเมตร วางบนผวหนาอาหารทเกลยเชอไว
โดยใชตวทาละลายเปนชดควบคม ทาการทดลอง 3 ชา บมเชอทอณหภม 30 องศาเซลเซยส เปนเวลา 24 ชวโมง ตรวจผลการ
ยบยงเชอ โดยสงเกตวงใส (clear zone) หาคาเฉลยขนาดเสนผานศนยกลางบรเวณยบยง (inhibition zone)
การหาคาความเขมขนตาสดทสามารถยบยงการเจรญของแบคทเรย (Minimal inhibitory concentration, MIC) ดวย
วธ Macro broth dilution technique (Srisukong et al., 2016)
นาสารสกดทมฤทธยบยงเชอมาทาการเจอจางลดลงครงละสองเทา ไดความเขมขนของสารสกด 100, 50, 25, 12.5,
6.25, 3.12, 1.56, 0.78, 0.39, 0.19 และ 0.09 มลลกรมตอมลลลตร ตามลาดบสวน ใชอาหารเลยงเชอทไมมสารสกดเปนชด
ควบคม เตมเชอทปรบคาความขนของเซลลเรมตน (OD600 เทากบ 0.1) ลงในหลอดทมสารสกดแตละความเขมขน และหลอด
ควบคม ทาการทดลองละ 3 ซา จากนนบมทอณหภม 30 องศาเซลเซยส เปนเวลา 24 ชวโมง ตรวจผลโดยสงเกตจากความขน
หากพบสารสกดทยบยงการเจรญของเชอไดสารละลายจะมลกษณะใส บนทกคา MIC ในหนวยมลลกรมตอมลลเมตร
การหาความเขมขนตาสดทสามารถฆาแบคทเรยได (Minimal bactericidal concentration, MBC)
นาหลอดทไมพบความขนจากการทดสอบคา MIC มาทดสอบหาคา MBC โดยการเกลยบนอาหารแขง NA (หลอด
ละ 3 ชา) บมทอณหภม 30 องศาเซลเซยส เปนเวลา 24-48 ชวโมง หากสารสกดสามารถฆาเชอไดจะไมพบการเจรญของเชอ
บนอาหาร บนทกคา MBC ในหนวยมลลกรมตอมลลลตร (Srisukong et al., 2016)
ผลการทดลองและวจารณ
การสกดสารจากตวอยางพช
สารสกดจากพชดวยตวทาละลาย 2 ชนด จานวน 10 ตวอยาง ไดปรมาณสารสกดรอยละ 4.84-29.29 โดยดอก
ขเหลกทสกดดวยเอทานอลไดปรมาณสารสกดมากทสด และใบบวบกทสกดดวยนาใหปรมาณสารสกดมากทสด (table 1) ใน
งานวจยครงนเลอกตวทาละลายนา และเอทานอลมาใชสกดสารจากพชเนองจากเปนตวทาละลายทใหปรมาณรอยละสารสกด
ไดด เมอเทยบกบตวทาละลายอน เชน คลอโรฟอรม และ อะซโตน (Viji et al., 2010) จากรายงานของ Troung et al.
(2019) พบวาตวทาละลายนาสกดสารจากพชไดปรมาณสารสกดมากกวาตวทาละลายเอทานอล แตไดปรมาณสารออกฤทธ
ทางชวภาพ (Bioactive compound) เชน เทอรพนอยด (Terpenoids) และ แอลคาลอยด (alkaloids) นอยกวาการสกด
ดวยเอทานอล
การทดสอบประสทธภาพการยบยงการเจรญของแบคทเรย
ทดสอบการยบยงเชอ X. campestris pv. campestris พบวา มสารสกดจากพช 4 ตวอยางทสามารถยบยงเชอได
ไดแก สารสกดใบยางนางดวยเอทานอลและนา ตามดวยสารสกดใบขเหลกดวยตวทาละลายนา และสารสกดดอกขเหลกดวย
ตวทาละลาย เอทานอล พบคาเฉลยเสนผานศนยกลางบรเวณยบยงเทากบ 3.66, 3.00, 1.05 และ 1.83 มลลเมตร ตามลาดบ
(table 1) จากผลการทดสอบสารสกดใบยานางใหผลการยบยงเชอไดดทสด ซงสอดคลองกบคณสมบตทางเคมในใบยานางท
ประกอบดวย ฟลาโวนอยด (Flavonoids) เทอรพนอยด และ แอลคาลอยด ซงมฤทธตานเชอแบคทเรย และเชอราได
(Rahman et al., 2017) สารสกดจากดอกและใบขเหลกมประสทธภาพในการยบยงเชอรองลงมาจากสารสกดใบยานาง ทงน

การศกษาประสทธภาพ ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
316
ขเหลกประกอบดวยสารแอลคาลอยด และฟลาโวนอยด คลายกบทพบในยานาง และแทนนน (Tannins) ทมคณสมบตในการ
ตานเชอจลนทรยเชนกน (Abdulrasheed et al., 2015) สวนสารกดจากใบบวบก และตนโคกกะออมดวยตวทาละลายทง
สองชนด จากการทดลองครงนไมพบความสามารถในการยบยงเชอแบคทเรยทดสอบ ซงใหผลการทดสอบแตกตางจากรายงาน
ของ Nasution et al. (2018) ทมการทดสอบสารสกดจากพชในกลมใบบวบก (Centella asiatica) ทสกดดวยนา และเอทา
นอล สามารถยบยงเชอแบคทเรยไดทงแกรมบวกและแกรมลบ โดยเฉพาะสารสกดจากรากสามารถยบยงเชอไดดกวาสารสกด
จากใบ เชนเดยวกบรายงานของ Suresh et al. (2012) สกดสารจากลาตนของพชในกลมเดยวกบโคกกะออม
(Cardiospermum halicacabum) ดวยเอทานอล พบสามารถยบยงเชอแบคทเรยไดทงแกรมบวกและแกรมลบ
ความเขมขนตาสดทสามารถยบยงการเจรญของแบคทเรย
จากผลทดสอบของสารสกดทสามารถยบยงเชอ X. campestris pv. campestris ได เมอนามาศกษาคา MIC พบวา
สารสกดทสามารถยบยงเชอได มลกษณะสารสารละลายใส เมอเปรยบเทยบกบชดควบคมจะพบลกษณะสารละลายขน
เนองจากการเจรญของเชอ ผลการทดสอบสารสกดมฤทธยบยงการเจรญของเชอไดทความเขมขนตงแต 3.12-100 มลลกรม
ตอมลลลตร สารสกดใบยานางดวยเอทานอล และนา มคา MIC เทากบ 6.25 และ 25 มลลกรมตอมลลลตร ตามลาดบ สาร
สกดดอกขเหลกดวยเอทานอล และสารกดใบขเหลกดวยนา มคา MIC เทากบ 3.12 และ 12.5 มลลกรมตอมลลลตร ตามลาดบ
(table 2)
ความเขมขนตาสดทสามารถฆาแบคทเรยได
เมอนาหลอดทความเขมขนของสารสกดสามารถยบยงเชอไดมาทดสอบหาคา MBC แสดงใน table 2 พบวา สาร
สกดใบยานางดวยเอทานอล และสารสกดใบขเหลกดวยนา มคา MBC เทากบ 6.25 และ 50 มลลกรมตอมลลลตร ตามลาดบ
เนองจากไมพบการเจรญของเชอบนอาหาร NA ในขณะทสารสกดใบยานางดวยนา และสารสกดดอกขเหลกดวยเอทานอลยง
พบการเจรญของเชอบนผวหนาอาหารทความเขมขน 25-100 และ 3.12-100 มลลกรมตอมลลลตร ตามลาดบ แตทงนการ
เจรญของเชอบนผวหนาอาหารลดลงเมอเพมความเขมขนสารสกดมากขน คาดวาสารสกดทงสองตวอยางนาจะสามารถฆาเชอ
ทดสอบไดทความเขมขนมากกวา 100 มลลกรมตอมลลลตร ดงนนหากตองการทราบคา MBC ของสารสกดทงสองชนด ควร
เพมความเขมขนของสารสกดใหมคามากกวา 100 มลลกรมตอมลลลตร จากผลการศกษาฤทธของสารสกดหยาบในพช 4 ชนด
พบวาสารสกดจากใบยานางดวยตวทาละลายเอทานอล และสารสกดใบขเหลกดวยตวทาละลายนาใหประสทธภาพในการฆา
เชอ X. campestris pv. campestris ไดด
สรป
จากการสกดสารดวยตวทาละลายเอทานอล และนาจากพชในทองถน 4 ชนด ไดแก ใบยานาง ใบและดอกขเหลก ใบ
บวบก และตนโคกกะออม พบวาสารสกดจากใบยานางดวยเอทานอล และนา สามารถยบยงเชอ X. campestris pv.
campestris ไดดทสด รองลงมาคอสารสกดดอกขเหลกดวยเอทานอล และสารสกดใบขดวยนา เมอนาสารสกดสกดใบยานาง
ดวยเอทานอล และนา ทดสอบ MIC มคาเทากบ 6.25 และ 25.00 มลลกรมตอมลลลตร ตามลาดบ และสารสกดใบยานาง
ดวยเอทานอลมคา MBC เทากบ 6.25 มลลกรมตอมลลลตร สวนสารสกดจากดอกขเหลกดวย และใบขเหลกดวยนา มคา MIC
เทากบ 3.12 และ 12.5 มลลกรมตอมลลลตร ตามลาดบ และสารสกดใบขเหลกมคา MBC เทากบ 50.00 มลลกรมตอมลลลตร
ในขณะทสารสกดจากใบบวบกและโคกกระออม ในตวทาละลายทงสองชนด ไมสามารถยบยงเชอทดสอบได จากผลการศกษา
ครงนสรปวาใบยานางมประสทธภาพในการยบยงเชอแบคทเรย X. campestris pv. campestris สาเหตโรคเนาดาไดดทสด
ซงควรศกษาพฒนาสารสกดจากใบยานางตอเพอใหสามารถใชกาจดโรคพชในแปลงผกไดจรงทดแทนการใชสารเคม

การศกษาประสทธภาพ ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
317
กตตกรรมประกาศ
ขอขอบคณ คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม ทใหความอนเคราะหใน
การทางานวจยครงน
เอกสารอางอง
Abdulrasheed, M., Ibrahim Hussein Isiaka and Ismail Ahmed Siddan. 2015. Determining the
phytochemical constituents and the antimicrobial activity of ethanolic extract of Acassia Leaf
(Senna Siamea) on some Enterobacteriaceae. IOSR J. Pharm. 6(5): 18-22.
Bila, j., C. N. Mortensen, M. Andresen, J. G. Vicente and E. G. Wulff. 2013. Xanthomonas campestris pv.
campestris race 1 is the main causal agent of black rot of Brassicas in Southern Mozambique. Afr.
J. Biotechnol. 12(6): 602-610.
Britto, A. J. D., D. H. Sh. Gracelin and S. R. Sebastian. 2011. Antibacterial activity of a few medicinal plants
against Xanthomonas campestris and Aeromonas hydrophila. Journal of Biopesticides. 4(1): 57-60.
Burakorn, J. and R. Praphruet. 2012. Antibacterial activities of seven indigenous vegetables. J. Thai Trad.
Alt. Med. 10: 11-22.
Kumar, N. K., V.B. S. Kumar, S. E. Manjunatha, A. S Padmaja, N. S. Pankaja and Venkatesh. 2018.
Antimicrobial activity of medicinal plants against Xanthomonas campestris pv. campestris causing
black rot of cabbage. Int. J. Chem. Stud. 6(1): 1871-1874.
Nasution Mhd. Y., M. Restuati, A. S. S. Pulungan, N. Pratiwi and D. S. Diningrat. 2018. Antimicrobial activities
of Centella asiatica leaf and root extracts on selected pathogenic micro-organisms. J. Med. Sci.
18(4): 198-204.
Rahman, Md. M., Md. Shamsuzzaman, M. Khatun, Md. M. Rahman, , A. S. M. S. Hossain, A. S. M. K. Alam,
A. Mosaddik, and M. I. I. Wahed. 2017. Phytochemical and antimicrobial properties of Tiliacora
triandra stem bark. Bayero Journal of Pure and Applied Sciences. 17(2): 1-9.
Srisukong, A., K. Jantree and S. Hanpakphum. 2016. The Study of antibacterial in weed extracts.
VRUResearch and Development Journal Science and Technology. 11(1): 69-82.
Suresh, S. N., S. Rathishkumar, V. Rajeshwari, P. Sagadevan, S. Gayathri and D. Vithya Eswari. 2012.
Phytochemical analysis and antibacterial potential of Cardiospermum halicacabum Linn.
(Sapindaceae). Int. J. of Pharm. & Life Sci. 12(3): 2209-2212.
Truong, D. H., D. H. Nguyen, N. T. Anh-Ta, A. V. Bui, T. Ha-Do and H. C. Nguyen. 2019. Evaluation of the use
of different solvents for phytochemical constituents, antioxidants, and in vitro anti- Inflammatory
activities of Severinia buxifolia. Journal of Food Quality. DOI10.1155/2019/8178294.
Viji, M. , M. Sathiya and S. Murugesan. 2010. Phytochemical analysis and antibacterial activity of medicinal
plant Cardiospermum haicacabum linn. Pharmacologyonline. 2: 445-456.
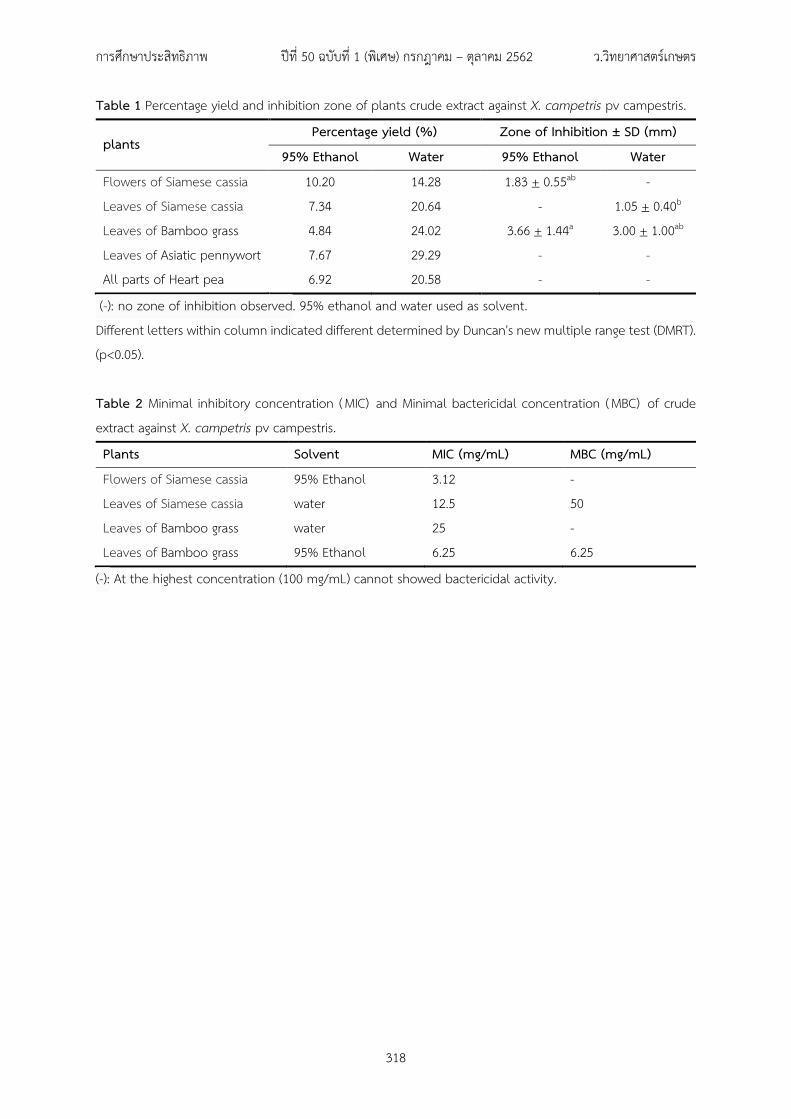
การศกษาประสทธภาพ ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
318
Table 1 Percentage yield and inhibition zone of plants crude extract against X. campetris pv campestris.
plants Percentage yield (%) Zone of Inhibition ± SD (mm)
95% Ethanol Water 95% Ethanol Water
Flowers of Siamese cassia 10.20 14.28 1.83 + 0.55ab -
Leaves of Siamese cassia 7.34 20.64 - 1.05 + 0.40b
Leaves of Bamboo grass 4.84 24.02 3.66 + 1.44a 3.00 + 1.00ab
Leaves of Asiatic pennywort 7.67 29.29 - -
All parts of Heart pea 6.92 20.58 - -
(-): no zone of inhibition observed. 95% ethanol and water used as solvent.
Different letters within column indicated different determined by Duncan's new multiple range test (DMRT).
(p<0.05).
Table 2 Minimal inhibitory concentration ( MIC) and Minimal bactericidal concentration ( MBC) of crude
extract against X. campetris pv campestris.
Plants Solvent MIC (mg/mL) MBC (mg/mL)
Flowers of Siamese cassia 95% Ethanol 3.12 -
Leaves of Siamese cassia water 12.5 50
Leaves of Bamboo grass water 25 -
Leaves of Bamboo grass 95% Ethanol 6.25 6.25
(-): At the highest concentration (100 mg/mL) cannot showed bactericidal activity.

170 วารสารวจยราชภฏพระนคร สาขาวทยาศาสตรและเทคโนโลย ปท 14 ฉบบท 2 (กรกฎาคม - ธนวาคม 2562)
ปยหมกมลไสเดอนดน : เทคโนโลยชววถเพอการอนรกษดนและการจดการขยะอนทรยในประเทศไทย
VERMICOMPOSTING: BIOLOGICAL TECHNOLOGY FOR SOIL CONSERVATION AND ORGANIC WASTES MANAGEMENT
IN THAILAND
พชญ ตงสมบตวจตร1* และ อทาน บรณศกดศร2
1*สาขาวชาวทยาศาสตร (ชววทยา) คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม
สพรรณบร 721302สาขาวชาวทยาการคอมพวเตอร คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม
สพรรณบร 72130
Pitchya Tangsombatvichit1* and Utharn Buranasaksee2
1*Department of Science (Biology) Faculty of Science and Technology, Rajamangala University of Technology
Suvarnabhumi, Suphanburi, 721302Department of Computer Science Faculty of Science and Technology, Rajamangala University of Technology
Suvarnabhumi, Suphanburi, 72130
*E-mail: [email protected]
Received: 2018-10-30
Revised: 2018-12-07
Accepted: 2019-01-10
บทความวชาการ
บทคดยอ
การท�าปยหมกมลไสเดอนในประเทศไทยเปนเทคโนโลยทยงยนส�าหรบการก�าจด
ของเสยทางการเกษตรและสารอนทรย ซงเปนกระบวนการทางธรรมชาตในการรไซเคล
สารอนทรยอยางรวดเรวเพอเปลยนเปนปยหมกทอดมดวยสารอาหารโดยใชไสเดอนสายพนธ
Eudrilus eugeniaeภายใตสภาวะมอากาศปยหมกมลไสเดอนเปนผลตภณฑปลอดสารพษ
ทเปนมตรกบสงแวดลอมมตนทนต�าใชเครองจกรนอยใชงานงายและใชพลงงานต�าในระหวาง
กระบวนการกระบวนการนถกน�ามาใชเพอปรบปรงโครงสรางดนทเสยใหดขนดงนนเทคนคน
จงเปนเทคโนโลยทางเลอกทดและงายปยหมกมลไสเดอนจากวสดเหลอทงทางการเกษตร
ถกน�ามาทดสอบเพอคนหาวสดทางการเกษตรทดทสดส�าหรบผลตปยหมกมลไสเดอนทม

171Phranakhon Rajabhat Research Journal (Science and Technology)Vol.14 No.2 (July - December 2019)
สารอาหารทจ�าเปนส�าหรบพชปรมาณสง และมจลนทรยทดทชวยในการเจรญเตบโตของพช
โดยการเพมฮอรโมนสดทายกระบวนการท�าปยหมกมลไสเดอนชวยเกษตรกรยคใหมในการ
บรหารจดการของเสยอนทรยและวสดเหลอทงทางการเกษตร อกทงเพมความปลอดภยของ
ผลผลตทางการเกษตรและสขภาพของผบรโภคในประเทศไทย
ค�าส�าคญ:เทคโนโลยชววถปยหมกมลไสเดอนดนขยะอนทรยเปนมตรกบธรรมชาต
ABSTRACT
In Thailand, vermicomposting is an innovative sustainable technology
for organic and agricultural waste treatment which is a natural process of quick
recycling of organic material into nutrient-rich compost using earthworms such as
Eudrilus eugeniae under aerobic condition. Vermicompost is an eco-friendly non-toxic
product, low cost, less mechanized, easy operation and uses low energy input during
processing etc. This process is adopted for the improvement of soil damage. Thus,
this technique has become a simpler alternative technology. The vermicompost from
variousorganicwasteswasevaluatedtofindout thebestorganicrawmaterial for
quality vermicompost production that has high essential nutrients for plants and contains
good microorganisms that help to grow the plant as a hormones enhancement. Finally,
vermicompostingprocessbenefitstheorganicandagriculturalwastesmanagement
for smart farmer while improving agricultural safety and consumer health in Thailand.
Keywords: Biological technology, Vermicomposting, Organic wastes, Nature-friendly
บทน�า
ประเทศไทยเปนประเทศเกษตรกรรม ท�าการเพาะปลกพช ไมวาจะเปนพชอาหาร
พชประดบพชน�ามน ดวยประเทศไทยเปนประเทศทอยในเขตรอนชนมชวงฝนตก อากาศ
อบอนและมดนทอดมสมบรณเหมาะแกการเจรญเตบโตของพชจงท�าใหมความหลากหลาย
ทางสายพนธพชเปนอยางมากพชเปนสงมชวตทตองการอาหารเพอชวยในการสรางพลงงาน
จากกระบวนการสงเคราะหแสง เนองจากพชมรงควตถทมชอวา คลอโรฟลล ดงนน อาหาร
ของพชจะไดจากดนเมอมการปลกพชอยางตอเนองเปนเวลานานหลายปตดตอกนสภาพดน
ทเคยมธาตอาหารทจ�าเปนตอการเจรญเตบโตของพชกจะถกน�าไปใชและลดลงตามล�าดบ
ดงนนมนษยจงคดทจะเพมธาตอาหารในดนเพอชดเชยสวนทถกใชไปในรปของสงทเรยกวา

172 วารสารวจยราชภฏพระนคร สาขาวทยาศาสตรและเทคโนโลย ปท 14 ฉบบท 2 (กรกฎาคม - ธนวาคม 2562)
“ปย”เปนวสดทเพมธาตอาหารพชในดนจากความฉลาดทไมเฉลยวของมนษยมนษยไดผลต
ปยเคมซงเกดจากการสงเคราะหองคประกอบของสารดวยโครงสรางทางเคมมใชสงทมาจาก
ธรรมชาต ชวงแรกทมการใชจะปรากฎผลการเจรญเตบโตของพชทดและรวดเรว เนองจาก
ปยเคมถกสรางใหละลายน�าไดงายแตผลเสยทเกดขนคอปยเคมจะอดชองวางของอากาศ
ภายในดนท�าใหการระบายน�ามปญหาและเมอสะสมไปนานๆจะเกดเปนสารตกคางในดน
และเปนอนตรายตอสงมชวต และระบบนเวศ ดงนน ในปจจบนมนษยจงเหนความส�าคญ
ของทรพยากรดนมากขน ไดมการคดผลตปยทมาจากธรรมชาต เพอกลบสธรรมชาต เรยกวา
ปยอนทรยอาทเชนปยมลไสเดอนดนกเปนปยอนทรยชนดหนงทมกระบวนการคดใชคณสมบต
ของไสเดอนดนรวมกบวธการผลตปยหมก ปยทผลตไดจากวสดทางธรรมชาตจะเปนมตรตอ
สงแวดลอมการผลตปยหมกมลไสเดอนดนจงเปนอกหนงวธ ทจะชวยพลกฟนสภาพดนใหม
คณภาพดจดเปนเทคโนโลยสเขยวเปนมตรตอมนษยและสงแวดลอม การผลตป ยหมก
มลไสเดอนดน เพอชวยก�าจดขยะอนทรยและไดป ยคณภาพดใชในการท�าเกษตรอนทรย
การผลตปยมลไสเดอนดนเกดขนในประเทศตาง ๆ ไมวาจะเปน เวยดนาม จนสหรฐอเมรกา
อนเดยแคนาดารวมทงประเทศไทยดวย
เทคโนโลยการผลตปยมลไสเดอนดน
ป ยมลไสเดอนดนเปนกระบวนการผลตปยหมกซงเปนปยอนทรย เกดจากการ
เปลยนวสดอนทรยทเปนสวนเหลอทงมาผานขนตอนการหมกตามธรรมชาตดวยจลนทรย
ทมประโยชนรวมกบการท�างานของไสเดอนดนสายพนธทเหมาะสมของแตละสภาพพนทของ
ประเทศตางๆเพอผลตปยอนทรยทมคณภาพส�าหรบการปลกพชชนดตางๆปยทไดถกน�าไป
ใชในการเจรญเตบโตของพช ปรบปรงโครงสรางของดนใหมชองวางของอากาศ การระบาย
น�าด รวมทงเพมจ�านวนจลนทรยทมประโยชนในดน เชนจลนทรยปฏปกษกบเชอกอโรคพช
ในขณะทตวไสเดอนดนสามารถน�าไปใชเปนอาหารสตวเนองจากมโปรตนสง (Sinha et al.,
2010)จากการศกษาธรรมชาตของไสเดอนดนและมนกวจยสนใจศกษาจงมการน�าไสเดอนดน
มาเลยงและศกษาสภาพเลยงใหเหมาะสมเลยนแบบธรรมชาต เชน การเลยงในลนชกไม
ลนชกพลาสตกกลองพลาสตกถงพลาสตกรวมทงกะละมงพลาสตกภายใตโรงเรอนปดปองกน
การโดนแสงแดดจดใหความชนและออกซเจนทพอเหมาะ(Sharmaetal.,2005)

173Phranakhon Rajabhat Research Journal (Science and Technology)Vol.14 No.2 (July - December 2019)
ไสเดอนดน
ไสเดอนดนเปนสงมชวตทจดอยในอาณาจกรสตว ไฟลมแอนเนลดา เปนสตวไมม
กระดกสนหลงล�าตวกลมยาวมขอปลองมชอเรยกทวไปวาEarthwormสวนชอวทยาศาสตรนน
มความหลากหลายของสายพนธมากถง 8,000 สายพนธ แหลงทพบไสเดอนดน มกเปน
กองปยหมก กองมลสตว บรเวณดนทตาง ๆ ทมแหลงอาหารและความชนเพยงพอตอการ
ด�ารงชวตของไสเดอนดน (Pathma and Sakthivel, 2012) ไสเดอนดนเปนสตวทม 2 เพศ
ในตวเดยวเวลาผสมพนธจะมการจบคกนของตวผตวหนงและตวเมยจากอกตวหนงการจบคกน
และผสมพนธจะเกดขนกบไสเดอนดนทมการสราง ไคลเทลลมชดเจนทบรเวณสวนหว เวลา
ชวงกลางคนไมมแสงและมความชนสงจะเปนชวงเวลาทเหมาะสมตอการผสมพนธหลงจาก
ผสมพนธกนเรยบรอยแลวจะเกดการสรางถงไข(cocoon)เปนทบรรจตวออนของไสเดอนดน
เปนจ�านวนมาก ในชวงฤดฝนทมความชนสงจะมการสรางถงไขไดมากกวาชวงฤดอน ๆ
เมอไสเดอนดนฟกออกจากถงไข ล�าตวจะมลกษณะใสตอมาจะเรมเจรญเตบโตและพฒนา
สผวเปนสน�าตาลแดงสแดง เปนระยะวยกอนเจรญพนธ แตกตางกนไปตามสายพนธ และ
เรมมการสรางไคลเทลลมชดเจน ซงเปนสญญาณบอกใหรวาเปนไสเดอนดนระยะตวเตมวย
พรอมผสมพนธและขยายพนธตอไปเกดขนเปนวฏจกรวงจรชวต(Edwards,2004)
ไสเดอนดนสามารถจดกลมได 3 ลกษณะตามการอยอาศยในระดบความลกของ
ชนดนวดจากระดบผวดนไดแกกลมไสเดอนดนผวดน(0-5เซนตเมตร)กลมไสเดอนดนในดน
(0-25เซนตเมตร)และกลมไสเดอนดนในชนดนลก(0-100เซนตเมตร)(ตารางท1)ไสเดอนดน
ในกลมผวดน(Epigeics)เปนไสเดอนดนทเลอดมรงควตถจะเหนไดวาไสเดอนดนชนดนจะม
สแดงสแดงน�าตาลเหลอบสน�าตาลแดงล�าตวมขนาดเลกถงปานกลางยาวประมาณ2-5นวชอบอย
บรเวณระดบผวดนไมลกมากมความชนสงตลอดป ไมมการขดร ชอบกนอนทรยวตถตาง ๆ
กนในปรมาณมาก แพรพนธ ขยายพนธไดรวดเรว ไสเดอนดนในกลมนไดแก ไสเดอนดน
สายพนธ ขตาแร แอฟรกนไนท คอลเลอร บลเวรม ไทเกอรเวรม และไสเดอนดนสแดง
เปนสายพนธทพบในประเทศไทยและถกน�ามาใชในการก�าจดขยะอนทรย ในกระบวนการ
ผลตปยหมกมลไสเดอนดน(PathmaandSakthivel,2012)

174 วารสารวจยราชภฏพระนคร สาขาวทยาศาสตรและเทคโนโลย ปท 14 ฉบบท 2 (กรกฎาคม - ธนวาคม 2562)
บทบาทไสเดอนดนทางการเกษตร
ปกตไสเดอนดนถกพบในดนหรอกองมลสตวใชเปนตววดความอดมสมบรณของดน
และตรวจสอบการปนเปอนของสารเคมในดนเรยกวาดชนชวดทางชวสงแวดลอม(Bio-index)
ดนทมความอดมสมบรณและไมมสารเคมตกคางในดนจะมความหลากหลายของสงมชวต
รวมทงไสเดอนดนดวยเนองจากไสเดอนดนมไขมนทสามารถดดซบสารเคมทใชก�าจดศตรพช
หรอโลหะหลกทปนเปอนในดน(Paolettietal.,1991)สายพนธไสเดอนดนบางชนดยงมความ
ทนทานตอการไดรบสารเคมเขาไปภายในเซลลอกดวยไสเดอนดนมคณสมบตชอบชอนไชท�าให
ดนรวนซย เพมชองวางในดน ใหอากาศถายเทการระบายน�าและการอมน�าดขน การละลาย
ของธาตอาหารเกดขนไดงายท�าใหรากพชดดซมสารอาหารไดสะดวกการเคลอนทของไสเดอน
ดนจะชวยพลกดนกลบจากดานลางขนมาดานบนท�าใหดนดานบนลงไปดานลางแทนซงสวนดน
ดานบนจะมมลไสเดอนดนอย เปนสวนทมธาตอาหารตางๆ ของพช เปนการผสมคลกเคลา
แรธาตอาหารในดนเปนประโยชนตอรากพชในชนใตดนไดดดธาตอาหารไปใช ดวยธรรมชาต
ของไสเดอนดนทสามารถยอยสลายอนทรยวตถในดนซากพชซากสตวและสารอนทรยตางๆ
จงเปนประโยชนตอการสรางธาตอาหารทมโมเลกลเหมาะสมตอการทพชจะน�าไปใชไดเลย
ทนท (Edwards and Bohlen, 1996; Edwards, 2004) รวมทงการผลตสารควบคมการ
เจรญเตบโตหรอฮอรโมนทพชตองการ(PGRs)เชนออกซนจบเบอเรลลนไซโตไคนนเอทลน
และกรดแอบไซซกวตามนทจ�าเปนจะไดมาพรอมกบมลไสเดอนดนซงชวยสงเสรมการเจรญ
ตารางท 1 การจดจ�าแนกชนดของไสเดอนดนตามความชอบอยในแหลงทอยอาศยในระดบ
ความลกของชนดน(GajalakshmiandAbbasi,2004)
คณลกษณะ
ขนาดรปราง
ความทนทานตอแสง
การเคลอนทในดน
ความชนของผว
การสรางเมดส
ปรมาณไขตอรอบ
การแพรพนธ
ชวงอาย
เลก
นอย
คลองแคลว
มการสรางเมอกชน
ม
มาก
เรว
ปานกลาง
ใหญ
มาก
เชองชา
มการสรางเมอกชน
ไมม
นอย
ชา
นานหลายป
ปานกลาง
ปานกลาง
ปานกลาง
ไมสรางเมอกชน
มนอย
นอย
ชา
นานหลายป
ชนดของไสเดอนดน (Earthworm’s ecological type)
ผวดน (Epigeics) ในดน (Endogeics) ชนดนลก (Anecics)
175Phranakhon Rajabhat Research Journal (Science and Technology)Vol.14 No.2 (July - December 2019)
เตบโตของพช (Edwards et al., 2011)นอกจากนดวยระบบการยอยอาหารของไสเดอนดน
ทมแบคทเรยและจลนทรยหลายชนดอยเปนเชอประจ�าถน เมอไสเดอนดนยอยสลายอนทรย-
วตถแลวไดเปนมลไสเดอนดนออกมาจะมการปลดปลอยเชอจลนทรยทมประโยชนออกมา
บรเวณรากพชดวยปองกนการเขาท�าลายของเชอกอโรค(ตารางท2)(PathmaandSakthivel,
2012)
ชนดของไสเดอนดน
Lumbricus rubellus
L. terrestris
Eisenia foetida
E. foetida
Eudrilus sp.,
Eudrilus eugeniae
Rhizobium japonicum
Pseudomonas putida
Bradyrhizobium japonicum
Bacillus sp.
B. megaterium
B. pumilus
B. subtilis
Proteobacteria
Bacteroidetes
Verrucomicrobia
Actinobacteria
Firmicutes
Free-living N2fixers
Azospirillum
Azotobacter
Autotrophic
Nitrosomonas
Nitrobacter
Ammonifyingbacteria
กระตนการสรางฮอรโมนพชเพอการ
เจรญเตบโต
สงเสรมการสรางปมปมทราก
ของถวเหลอง
ชวยยบยงการเจรญเตบโตของ
จลนทรยกอโรคเชน
Enterococcus faecalis
Staphylococcus aureus
ชวยยบยงการเจรญเตบโต
ของเชอรากอโรคเชน
Colletotrichum coccodes
R. solani
P. ultimum, P. capsici
F. moliniforme
กระตนการสรางฮอรโมนพช
เพอการเจรญเตบโต
ชวยตรงไนโตรเจน
ชวยในการละลายของฟอสเฟส
ตานทานตอการเขาท�าลายของเชอ
กอโรคพช
Madsen and
Alexander(1982)
Rouelle(1983)
Vaz-Moreira et al.
(2008)
Yasiretal.(2009)
Gopaletal.(2009)
การใชประโยชนจากจลนทรย อางองชนดของจลนทรย
ทไดจากมลไสเดอนดน
ตารางท 2 ความหลากหลายของจลนทรยทพบในปยมลไสเดอนดนทผลตจากไสเดอนดน
สายพนธตางๆและการใชประโยชนของจลนทรย(ดดแปลงจากPathmaand
Sakthivel,2012)

176 วารสารวจยราชภฏพระนคร สาขาวทยาศาสตรและเทคโนโลย ปท 14 ฉบบท 2 (กรกฎาคม - ธนวาคม 2562)
วสดอนทรยทเหมาะน�ามาผลตปยมลไสเดอนดน
กระบวนการผลตปยมลไสเดอนดนเปนวธผลตปยทใชตนทนต�าไดธาตอาหารและ
ฮอรโมนตาง ๆ ทจ�าเปนตอการเจรญเตบโตของพชในปรมาณทพชตองการ เปนมตรกบ
สงแวดลอมรวมทงชวยลดปรมาณขยะอนทรยทมาจากแหลงชมชนตลาดแหลงอตสาหกรรม
แหลงสถานศกษาหนวยงานราชการตาง ๆ และจากการเกษตร เราสามารถน�าขยะอนทรย
มาใชเปนวสดเรมตนในกระบวนการผลต โดยใชคณสมบตของไสเดอนดนและจลนทรย
ทเกยวของเปนตวขบเคลอนกระบวนการสรางปยมลไสเดอนดนใหเกดเรวขนใชเวลาในการผลต
เปนปยสนขนเมอเทยบกบการผลตปยหมก (Dominguez andEdwards, 2004; Edwards
etal.,2011)เนองจากในกระเพาะอาหารของไสเดอนดนจะมจลนทรยทสามารถผลตเอนไซม
หลายชนดชวยยอยขยะอนทรยตางๆ(Lazcanoetal.,2008)ไดแกมลสตวทไดจากการท�า
การเกษตร เชนมลวว มลไก หรอมลสกร วสดเหลอทงทางการเกษตรตาง ๆ เชนฟางขาว
ชานออย ซงขาวโพดกากผลไม เปลอกผลไมหลากหลายชนด วสดเพาะเหดททงแลว วชพช
เศษผกททง เปนตน รวมทงกระดาษททงแลวกสามารถน�ามาผลตป ยมลไสเดอนดนได
(Tangsombatvichit et al., 2016) ปยมลไสเดอนดน จดเปนปยอนทรย ซงมาตรฐานของ
ปยอนทรยคอมความชนนอยกวาหรอเทากบ30%ความเปนกรด-ดางอยในชวง5.5-8.5
คาการน�าไฟฟา(EC)นอยกวาหรอเทากบ10dS/mการละลายสมบรณของปย(GI)มากกวา80%
อนทรยวตถ (OM)มากกวาหรอเทากบ 20% รวมทงธาตอาหารหลกของพช คอ ไนโตรเจน
ทงหมดมากกวา1.0%ฟอสฟอรสทงหมดมากกวา0.5%และโพแทสเซยมทงหมดมากกวา
0.5%(Prasitketetal.,2005)
วสดเหลอทงทางการเกษตรวชพชเศษผกผลไมถกน�ามาผลตเปนปยหมกเปรยบเทยบ
กบปยหมกทใชไสเดอนดนชวยท�างานพบวาปยมลไสเดอนดนมคณสมบตชวยสงเสรมสภาพดน
ใหด มธาตอาหารส�าหรบพชมากกวาปยหมกอนทรยธรรมดา ในระยะเวลาทใชในการหมก
เทากนการผลตปยมลไสเดอนดนจากขยะอนทรยแลวน�ามาใชในการปลกพชพบวาชวยสงเสรม
ใหดนมความโปรงรวนซย เพมชองวางชวยกกเกบน�าในดนท�าใหจลนทรยดนบรเวณรากพช
ท�างานไดด(BansalและKapoor,2000;Sinhaetal.,2010)การน�ามลแพะฟางขาววสด
เพาะเหด ขเลอยมาเปนวสดเรมตนในการผลตปยมลไสเดอนดน โดยใชไสเดอนดนสายพนธ
Lumbricus rubellus เปนสายพนธทพบในประเทศมาเลเซยพบวาคณภาพปยมลไสเดอนดน
ทผลตไดใหคาธาตอาหารหลกของพชเชนN,P,K,CuและZnรวมทงคาC:Nratioมคาสง
เมอเทยบกบคามาตรฐานทก�าหนด (Sailila et al., 2010) ในขณะท Klangkongsub and
Sohsalam(2013)ไดน�าไสเดอนดนสายพนธEudrilus eugenine มาชวยในการยอยเศษวสด
เหลอทงจากแปลงปลกมะเขอเทศ เพอผลตปยมลไสเดอน ปยทผลตไดเมอน�าไปวเคราะห

177Phranakhon Rajabhat Research Journal (Science and Technology)Vol.14 No.2 (July - December 2019)
ใหคาธาตอาหารหลกN,PและKสงกวาคาทก�าหนดของปยอนทรยมาตรฐานและสามารถใช
ปลกผกกาดขาวจนและบานชนไดคณภาพดกวาการใชปยเคม หรอแมกระทงวชพชอยาง
ผกตบชวากสามารถน�ามาผลตปยมลไสเดอนดนทมคณภาพดได โดยZirbes et al. (2011)
ไดน�ามลสกรมาผสมกบผกตบชวาในอตราสวน 3:1 ผานการท�างานของไสเดอนดนสายพนธ
Perionyx excauvatus ซงเปนสายพนธทพบในประเทศเวยดนาม เมอวเคราะหคณภาพ
ของปย พบวา ปยมลไสเดอนดนทผลตได ชวยสงเสรมการงอกของเมลดขาว เนองจากม
ธาตอาหารพชไดแกN,P,K,CaและMgอยในปรมาณเพยงพอตอการเจรญเตบโตของพช
หรอผกตบชวาทผานกระบวนการยอยจากไสเดอนดนสายพนธEudrilus engeniaeกใหผลผลต
ของปยอนทรยทมคณภาพด เหมาะแกการเจรญเตบโตของพช (Gajalakshmi et al., 2001)
วสดเพาะเหดทตองการทงแลวกสามารถน�ามาผลตปยมลไสเดอนดนไดดวยผานกระบวนการ
ท�างานของไสเดอนดนสายพนธEisenia foetida and E. andrei (Tajbakhshetal.,2008)
รวมทงขยะทเปนกระดาษทตองการทง กสามารถน�ามาเปนวสดเรมตนในการผลตปยมลไสเดอน
ไดเชนกนโดยผานการท�างานของไสเดอนดนสายพนธชนดนลกLampito mauritii(Gajalakshmi
et al., 2002)
ปจจยทสงผลตอการผลตปยมลไสเดอนดน
กระบวนการผลตปยหมกมลไสเดอนดนปจจยส�าคญทตองค�านงถงในการน�าไสเดอน
มาชวยในการยอยขยะอนทรยคอความชนทพอเหมาะตอการด�ารงชวตของไสเดอนดนในอาหาร
หลากหลายชนดทเราเลอกมาสราง bedding เนองจากไสเดอนดนใชผวหนงและความชน
ในกระบวนการหายใจเพอปรบสภาพใหมชวตอยไดในสภาวะทรอนหรอหนาวเกนไปซงความชน
ทเหมาะสมตอการด�ารงชวตของไสเดอนดนมคาอยท 45-60%แตหากเปนความชนโดยรวม
ของ bedding ส�าหรบไสเดอนดนในการผลตปยมลไสเดอนดนจะตองอยในชวง 70-90%
เปนคาความชนทเหมาะสมตอการเจรญเตบโตและการขยายพนธของไสเดอนดนอณหภม
ของสภาพอากาศโดยรวม ในการเลยงไสเดอนดนกควรอยในชวง 27 - 35 องศาเซลเซยส
ขนอยกบสายพนธของไสเดอนดนดวยแสงแดดและความรอนมผลอยางมากตอความเปนอย
ของไสเดอนดนเนองจากตามธรรมชาตไสเดอนดนชอบอยในทมแสงนอยมากดงนนหากตองการ
ผลตปยมลไสเดอนดนจะตองหาสถานททไมมแสงแดดสองถง และความรอนกมผลตอการ
สญเสยน�าและความชนในตวไสเดอนดน (Sinhaet al., 2010; Tangsombatvichit et al.,
2016) คาความเปนกรด-ดาง (pH) ของวสดอนทรยทใชเปนสวนผสมใน bedding ส�าหรบ
ใหไสเดอนดนอย กเปนอกปจจยหนงทมผลตอการมชวตรอดของไสเดอนดน เนองจาก pH
ทไสเดอนดนสามารถอยรอดไดตามธรรมชาตอยในชวง6-8(Edwardsetal.,2011)

178 วารสารวจยราชภฏพระนคร สาขาวทยาศาสตรและเทคโนโลย ปท 14 ฉบบท 2 (กรกฎาคม - ธนวาคม 2562)
สรป
ประโยชนจากการผลตปยมลไสเดอนดน
ในประเทศไทยปจจบนขยะอนทรยมปรมาณมากนอกจากการฝงกลบในดนแลว
กยงถกน�ามาผลตปยหมกอนทรย กระบวนการท�าปยหมกยงคงใหธาตอาหารทจ�าเปนตอ
การเจรญเตบโตของพชนอยและอาจมการปนเปอนของเชอจลนทรยกอโรคในปย เมอมการ
วจยการผลตปยหมกโดยการขบเคลอนของไสเดอนดน กลบพบความมหศจรรยวา ปยหมก
มลไสเดอนดนนน สามารถลดปรมาณการปนเปอนของเชอจลนทรยกอโรคได เชนEscherichia
coli fecal coliform และSalmonella sp. เปนตน (Dominguez and Edwards, 2004)
นนหมายความวา ปยหมกทไดจากเจาไสเดอนดนนน ไมเพยงแตลดปรมาณขยะอนทรย
จ�านวนมหาศาลแลวยงสงผลใหปยลดการปนเปอนของเชอโรคเมอน�าปยไปใชกบพชพชกจะ
ไมมการตดเชอจลนทรยกอโรคทางเดนอาหารทอาจสงผลกระทบตอผบรโภค เจาไสเดอนดน
ยงมความสามารถในการกระตน การท�างานของแบคทเรยในดนทท�าหนาทยอยสลายซากพช
(Decomposer) ไดอกดวย (Sinha et al., 2010) และประโยชนหลกของปยหมกทผานการ
ท�างานของเจาไสเดอนดนนนคอไดธาตอาหารทจ�าเปนตอการเจรญเตบโตของพชในปรมาณสง
อาทเชน N P และ K เปนตน ซงสงผลตอการเจรญเตบโตของตนพชอยางเหนผลชดเจน
(Tangsombatvichit et al., 2016) รวมทงปยมลไสเดอนดนยงมธาตอาหารรองและธาต
อาหารเสรมทพชตองการปรงปรงโครงสรางทางกายภาพของดนคอมความรวนซยมชองวาง
ของอากาศน�าสามารถไหลผานไดดรากพชสามารถชอนไชดดธาตอาหารและน�าไดดมากขน
อกดวยดงนน การผลตปยมลไสเดอนดนจงเหมาะตอบรบทของประเทศไทยซงเปนประเทศท
รายไดหลกมาจากการเกษตรปลกพชผกตาง ๆการผลตปยชนดนยงลดตนทนการผลตไดอก
วสดอนทรยหรอขยะอนทรยหาไดฟรตามครวเรอน(PathmaandSakthivel,2012)เทคโนโลย
ปยมลไสเดอนดนไมเพยงแคประโยชนในการผลตปยไวใชในการเพาะปลกปรบปรงโครงสรางดน
ใหดขนหรอลดปรมาณขยะอนทรย วสดเหลอทงทางการเกษตรในประเทศเทานนการใชปย
มลไสเดอนดนยงสงเสรมเกษตรกรใหผลตพชผกปลอดภย สงผลตอความปลอดภยดาน
สขภาพของผผลต ผบรโภคและสงแวดลอมสนบสนนการเปนเกษตรกรยคดจทลเพอพฒนา
ประเทศไทยตอไปในอนาคต

179Phranakhon Rajabhat Research Journal (Science and Technology)Vol.14 No.2 (July - December 2019)
เอกสารอางอง
BansalS,KapoorK.K. (2000).Vermicompostingofcrop residuesandcattledung
withEiseniafoetida.Bioresour Technol.73,95–98.
DominguezJ.&EdwardsC.A.(2004).Vermicomposting organic wastes: A review.
In: ShakirHannaS.H.,MikhailW.Z.A. (eds) Soil zoology for sustainable
development in the 21stcentury.,Cairo,pp.369-395.
EdwardsC.A.andBohlenP.J. (1996).Biology and Ecology of Earthworms. Third
editionLondon:Chapman&Hall.
EdwardsC.A.(2004).Earthworm ecology.SecondeditionTheUnitedStatesofAmerica:
CRCPress.
Edwards C.A., Arancon N.Q. & Sherman R. (2011).Vermiculture technology:
Earthworms, organic wastes and environmental management.TheUnited
StatesofAmerica:TaylorandFrancisGroup.
GajalakshmiS.,RamasamyE.V.&AbbasiS.A. (2001).Assessmentofsustainable
vermiconversionofwaterhyacinthatdifferentreactorefficienciesemploying
EudrilusengeniaeKingburg.Bioresour Technol.80,131–135.
GajalakshmiS.,RamasamyE.V.&AbbasiS.A. (2002).Vermicompostingofpaper
wastewiththeanecicearthwormLampitomauritiiKingburg.Indian J Chem
Technol.9,306–311.
GajalakshmiS.&AbbasiS.A. (2004).Earthwormsandvermicomposting. Indian J.
of Biotechnology.3,486-494.
GopalM.,GuptaA.,SunilE.&ThomasV.G.(2009).Amplifacationofplantbeneficial
microbial communities during conversion of coconut leaf substrate to
vermicompostbyEudrilussp.Current Microbiology.59,15-20.
KlangkongsubS.&SohsalamP.(2013).Vermicompostproductionbyusingtomato
residue and yard waste. Journal of Medical and Bioengineering.2,270-273.
Lazcano C., Gomez-BrandonM. & Dominguez J. (2008). Comparison of the
effectiveness of composting and vermicomposting for the biological
stabilization of cattle manure. Chemosphere.72,1013–1019.
Madsen E.L. &AlexanderM. (1982). Transport of Rhizobium andPseudomonas
through soil. Soil Science Society of America.46,557-560.

180 วารสารวจยราชภฏพระนคร สาขาวทยาศาสตรและเทคโนโลย ปท 14 ฉบบท 2 (กรกฎาคม - ธนวาคม 2562)
PaolettiM.G., FaverettoM.R., Stinner B.R., Purrington F.F. & Bater J.E. (1991).
Invertibrates as bioindicators of soil use. Agriculture Ecosystem Environment.
34,341-362.
Pathma J. & Sakthivel N. (2012).Microbial diversity of vermicompost bacteria
that exhibit useful agricultural traits and waste management potential.
SpringerPlus.26,1-19.
Prasitket J., Chuvorrivate N., RuengnabM., Padung T., Choulvanapong P. &
ReanjareangS.(2005).Organic fertilizer: Production, utilization and quality.
DepartmentofAgriculture.pp.87.
RouelleJ.(1983).Introduction of an amoeba and Rhizobium Japonicum into the gut
of Eisenia fetida (Sav.) and Lumbricus terrestris L. In: Satchel1 J.E. (ed)
Earthworm Ecology: From Darwin to Vermiculture.NewYork:Chapman
andHall.
SaililaN.,BakarA.A.,MahmoodN.Z.,SilivaJ.T.,AbdullahN.&Jamaludin.(2010).
Nutrient elements of different agricultural wastes from vermicomposting
activity. Dynamic Soil, Dynamic Plant.4,155-158.
Sharma,S.,Pradham,K.,Satya,S.&Vasudevan,P.(2005).Potentialityofearthworms
forwastemanagementandinotherUse-AReview.Journal American Science.
1,4-16.
Sinha,R.K.,Agarwai,S.,Chauhan,K.&Valani,D.(2010).Thewondersofearthworms
&itsVermicompostinfarmproduction:CharlesDarwin’sfriendsoffarmers’
with potential to replace destructive chemical fertilizers from agriculture.
Agricultural Sciences.1,76-94.
Sinha,R.K.,Agarwal,S.,Chauhan,K.,Chandran,V.&Soni,B.K.(2010).Vermiculture
technology:Reviving thedreamsof SirCharlesDarwin for scientific use
of earthworms in sustainable development programs. Technology and
Investment.1,155-172.
TajbakhshJ,AbdoliMA,MohammadiGoltapehE,AlahdadiI,MalakoutiMJ(2008).
Trend of physico-chemical properties change in recycling spent mushroom
compostthroughvermicompostingbyepigeicearthwormsEiseniafoetida
andE.Andrei.J Agric Technol.4,185–198.

181Phranakhon Rajabhat Research Journal (Science and Technology)Vol.14 No.2 (July - December 2019)
Tangsombatvichit P., Chupong S., Ketrot D. & Boonlerthirun K. (2016). The
management of organic wastes produced vermicompost using earthworm
Eudrilus eugeniae and effects of vermicompost on growth of Helianthus
annuus.5th Internationalconferenceon food,AgriculturalandBiological
Sciences(ICFABS-2016),Dec25-26,2016,Thailand.
Vaz-Moreira I.,MariaE.,SilvaC.M.,ManaiaOlgaC.&Nunes. (2008).Diversityof
bacterial isolates from commercial and homemade composts. Microbial
Ecology.55,714–722.
YasirM.,AslamZ.,KimS.W.,LeeS.W.,JeonC.O.&ChungY.R.(2009).Bacterial
community composition and chitinase gene diversity of vermicompost with
antifungal activity. Bioresour Technol.100,4396–4403.
Zirbes, L.,Renard,Q.,Dufey, J., Tu, P.K.,Duyet,H.N., Lebailly, P., Francis, F.&
Haubruge,E.(2011).Valorisationofawaterhyacinthinvermicomposting
using an epigenic earthworm Perionyx excavatus in central Vietnam.
Biotechnol. Agron. Soc. Environ.15,85-93.

182 วารสารวจยราชภฏพระนคร สาขาวทยาศาสตรและเทคโนโลย ปท� 14 ฉบบท� 2 (กรกฎาคม - ธนวาคม 2562)

Agricultural Sci. J. 50 : 1 (Suppl.) : 319-327 (2019) ว. วทย. กษ. 50 : 1 (พเศษ) : 319-327 (2562)
319
การคดเลอกและการหาสภาวะทเหมาะสมในการบาบดนาเสยโดยสาหรายสเขยวทคดแยกได
จากนาเสยในโรงงานอตสาหกรรมอาหาร
Screening and Optimization of Wastewater Treatment by Green Algae Isolated
from Food Industrial Wastewater
สามารถ ตายขาว1* และ ปรชาต แซออ1
Samart Taikhao1* and Parichard Saeu1
160 หม 3 สาขาวชาวทยาศาสตร คณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม พระนครศรอยธยา 13000 160 Moo 3 Division of Science Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi,
13000, Thailand *Corresponding author: [email protected]
บทคดยอ
สาหรายสเขยวมความสามารถในการกาจดสารอาหาร เชน สารอนทรย ไนเตรท และฟอสเฟตในนาเสยได งานวจย
นมวตถประสงคเพอคดแยกสาหรายสเขยวจากนาเสยในโรงงานอตสาหกรรมอาหารในจงหวดพระนครศรอยธยา ทมความ
สามารถสงในการบาบดนาเสย จากการทดลองสามารถคดแยกสาหรายสเขยวได 76 ไอโซเลท จากนาเสยในโรงงาน
อตสาหกรรมอาหาร 3 แหง และสามารถแบงกลมตามลกษณะสณฐานวทยาได 11 กลม จากการศกษาการเจรญเตบโตดวย
การวดคาการดดกลนแสงทความยาวคลน 750 นาโนเมตร (OD750) และปรมาณคลอโรฟลลภายใตการเพาะเลยงในอาหาร
Tris Acetate Phophate (TAP) พบวาสาหรายสเขยวไอโซเลท FFP 13 มการเจรญสงสด (OD750 = 3.87±0.02) และม
ปรมาณคลอโรฟลลสงสด (216.17±0.120 ไมโครกรมคลอโรฟลลตอลตร) หลงจากเพาะเลยงเปนเวลา 96 ชวโมง จากนน นา
ไอโซเลท FFP 13 มาตรวจสอบการบาบดนาเสยโดยแปรผนความเจอจางของนาเสย พเอช และอณหภม พบวาสภาวะท
เหมาะสมในการบาบดนาเสย คอการใชนาเสยทไมเจอจาง คาพเอชเทากบ 8 และอณหภมเทากบ 30 องศาเซลเซยส ภายใต
สภาวะดงกลาว ไอโซเลท FFP 13 มประสทธภาพสงในการกาจดคา บโอด ซโอด ไนเตรท และ ฟอสเฟต ดวยคาประสทธภาพ
ในการกาจดเทากบรอยละ 78.57 77.39 62.56 และ 75.16 ตามลาดบ นอกจากนนาเสยทไดจากการบาบดยงผานเกณฑ
มาตรฐานนาทงโรงงานอตสาหกรรมอกดวย
คาสาคญ: สาหรายสเขยว, การบาบดนาเสย, นาเสยในโรงงานอตสาหกรรมอาหาร
ABSTRACT
Green algae are capable of removing nutrients such as organic, nitrate and phosphate from
wastewater. This work aimed to isolate green algae from food industrial wastewater in Phra Nakhon Si
Ayutthata province that has highly efficient wastewater treatment. Seventy-Six isolate of green algae were
isolated from three food industrial wastewater samples. All green algae isolates were divided into 11 groups
by morphological characterization. The growth by measuring optical density at 750 nm (OD750) and
chlorophyll content under cultivated cell in Tris Acetate Phosphate (TAP) medium were determined. The
results found that the green algae isolate FFP 13 showed the highest growth (OD750 = 3.87±0.02) and the

การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
320
highest chlorophyll content (216.17±0.120 μgChl L-1) after 96 h of cultivation. Then, isolate FFP 13 were
also examined for wastewater treatment under various the dilutions of wastewater, pH and temperature.
The optimal conditions for wastewater treatment were undiluted wastewater, pH 8, and temperature of 30
˚C. Under these conditions, the isolate FFP 13 showed high capability to remove BOD, COD, nitrate and
phosphate from wastewater with removal efficiency of 78.57, 77.39, 62.56 and 75.16%, respectively. In
addition, the treated wastewater also passed the criteria of industrial effluent standards.
Keywords: green algae, wastewater treatment, food industrial wastewater
บทนา
สาหรายสเขยวเปนจลนทรยทเจรญไดงายในสภาวะทหลากหลาย เชน สภาวะโฟโตออโตโทรป (Photoautotroph)
โฟโตเฮเทอโรโทรป (Photoheterotroph) และโฟโตมกซโซโทรป (Photomixotroph) ในปจจบนไดมการนาสาหรายสเขยว
มาเพาะเลยงในนาเสยเพอลดปรมาณสารอนทรย เชน ไนโตรเจน และ ฟอสฟอรส ซงเปนการบาบดนาเสยวธหนงทประหยด
ตนทนและมประสทธภาพ (Aslan et al., 2006) นอกจากนยงมรายงานการศกษาการคดแยกสาหรายสเขยวจากนาเสย เพอ
คดเลอกสาหรายสเขยวสายพนธใหมๆ ทมประสทธภาพในการบาบดนาเสย เชน Oocystis sp. คดแยกไดจากนาเสย จาก
ฟารมสกร (Riano et al., 2011) Chlorella sp. คดแยกจากนาเสยโรงงานสรา (Farooq et al., 2013) Chlorella vulgaris
YSW–04 คดแยกจากนาเสยในฟารมสกร (Ji et al., 2013), Micractinium reisseri และ Scenedesmus obliquus คด
แยกไดจากนาเสยเทศบาล (Abou–Shana et al., 2014) เปนตน โดยสาหรายสเขยวแตละชนดจะมความ สามารถในการ
บาบดนาเสยทแตกตางกน และสภาวะทใชในการเพาะเลยง เชน ความเขมขนของนาเสย พเอช และอณหภม ยงสงผลตอการ
เจรญและการบาบดนาเสยในสาหรายสเขยวอกดวย (Molinuevo–Salces et al., 2019)
งานวจยนจงมจดมงหมาย เพอคดเลอกและหาสภาวะทเหมาะสมในการบาบดนาเสยโดยสาหรายสเขยวทคดแยกได
จากนาเสยในโรงงานอตสาหกรรมอาหารในเขตจงหวดพระนครศรอยธยา โดยทาการคดแยกสาหรายสเขยวจากนาเสย จนได
เชอบรสทธ จากนนศกษาลกษณะสณฐานวทยา การเจรญ และคดเลอกสาหรายทมการเจรญเตบโตทดมาศกษาการบาบด นา
เสย ตลอดจนหาสภาวะทเหมาะสมในการบาบดนาเสยโดยสาหรายทคดเลอกได
อปกรณและวธการ
การเกบตวอยางและการคดแยกสาหรายสเขยวจากนาเสย
เกบตวอยางสาหรายสเขยวจากนาเสยในบอบาบดนาเสย จากโรงงานอตสาหกรรมอาหารทมกระบวนการบาบดนา
เสยทางชวภาพ (Biological Treatment) ในเขตจงหวดพระนครศรอยธยา จานวน 3 โรงงาน คอ โรงงานผลตผลตภณฑนม
โรงงานผลตอาหารแชแขง และโรงงานผลตแปงพซซา จากนนนาตวอยางนามาวางไวในททมแสง เปนระยะเวลาประมาณ 3-5
วน จากนนแยกเชอสาหรายโดยวธการ Spread plate method บนอาหาร BG11 (Rippka et al., 1979) และนาไวในทม
แสงสวางทความเขมแสง 30 ไมโครโมลไอนสไตนตอตารางเมตรตอวนาท อณหภม 25 องศาเซยลเซยส จากนนแยกเชอให
บรสทธดวยวธ Streak plate method บนอาหาร BG11 และนาเซลลสาหรายสเขยวมาศกษาลกษณะทางสณฐานวทยา และ
จาแนกหมวดหม โดยตรวจสอบลกษณะรปราง และการจดเรยงตวของเซลล ภายใตกลองจลทรรศนแบบใชแสง
การเพาะเลยงสาหรายสเขยวและการวดการเจรญเตบโต
เพาะเลยงสาหรายสเขยวแตละไอโซเลทในฟลาสกขนาด 250 มลลลตร ทมอาหารเหลว Tris Acetate Phosphate
(TAP) (Harris, 1989) ปรมาตร 100 มลลลตร โดยใหมคา OD750 เรมตนเทากบ 0.1 โดยใชเครองสเปคโตรโฟโตมเตอร

การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
321
(Biochrom, Libra S70, US) เลยงในสภาวะเขยาทความเรวรอบ 120 รอบตอนาท ทอณหภม 25 องศาเซลเซยส และให
ความเขมแสง 30 ไมโครโมลไอนสไตนตอตารางเมตรตอวนาท เปนเวลา 18 ชวโมงตอวน วดการเจรญเตบโตโดยวดคา OD750
และวดปรมาณคลอโรฟลลตามวธการของ Riemann and Ernst (1982) ทก 24 ชวโมงเปนระยะเวลา 96 ชวโมง
การเกบตวอยาง การวดองคประกอบของนาเสย และการเตรยมนาเสยสาหรบเพาะเลยงสาหราย
นาเสยทใชในการทดลองนเกบมาจากนาเสยในโรงงานอตสาหกรรมอาหารแชแขง จงหวดพระนครศรอยธยา โดยนา
นาเสยมาวดคาพเอช อณหภม และคาออกซเจนละลายนา (Dissolved Oxygen, DO) ทนท ดวยเครอง multi-parameter
meter (HACH, HQ40D, USA) จากนนนานาเสยมากรองดวยกระดาษกรองเบอร 3 (เสนผานศนยกลาง 90 มลลเมตร)
(Whatman, UK) จากนนนานาเสยมาปรบพเอชเทากบ 7 และนาไปนงฆาเชอเพอนาไปเพาะเลยงสาหราย จากนนนาเสย ท
ปราศจากเชอสวนหนงมาวดคาบโอด (Biochemical Oxygen Demand, BOD) ดวยเครอง BOD analyzer (Lovibond,
Oxi-Direct, Germany) วดคาซโอด (Chemical Oxygen Demand, COD) ตามวธการของ American Public Health
Association (APHA) 5220 C (1998) และวดปรมาณไนเตรท และฟอสเฟต ดวยเครอง Ion chromatograph (IC) (Thermo
Fisher Scientific, Dionex ICS-5000+ DP, USA)
การเพาะเลยงสาหรายสเขยวในนาเสย
ทาการเจอจางนาเสยทปราศจากเชอดวยนากลนทปราศจากเชอ ใหนาเสยมความเขมขนรอยละ 100 90 80 70
และ 60 (ปรมาตรตอปรมาตร) ในกรณการแปรผนคาพเอชในนาเสย ปรบคาพเอชในนาเสยดวยกรดไฮโดรคลอรกความ
เขมขน 6 โมลาร หรอโซเดยมไฮดรอกไซดความเขมขน 6 โมลาร เพอใหคาพเอชมคาเทากบ 5 6 7 8 และ 9 จากนนเพาะเลยง
สาหรายสเขยวในฟลาสกขนาด 250 มลลลตร ทมนาเสยทเตรยมไวปรมาตร 100 มลลลตร โดยใหมคา OD750 เรมตนเทากบ
0.1 เพาะเลยงในสภาวะเขยาทความเรว 120 รอบตอนาท ทอณหภม 25 องศาเซลเซยส และใหความเขมแสง 30 ไมโครโมล
ไอนสไตนตอตารางเมตรตอวนาท เปนเวลา 18 ชวโมงตอวน เปนระยะเวลา 5 วน ในกรณการแปรผนอณหภม ใหแปรผน
อณหภมในการเพาะเลยงเทากบ 25 30 35 และ 40 องศาเซลเซยส เมอเพาะ เลยงสาหรายสเขยวครบ 5 วน กรองสาหรายส
เขยวดวยกระดาษกรอง GF/C (เสนผานศนยกลาง 47 มลลเมตร) (Whatman, UK) จากนนนานาเสยทไดไปวเคราะห
องคประกอบตางๆ โดยทาการทดลอง 3 ซาและวเคราะหผลทางสถตดวยโปรแกรม SPSS 13.0 (SPSS Inc.) โดยใช one-way
ANOVA ดวยวธ Duncan (alpha = 0.05)
ผลการทดลองและวจารณ
ผลการคดแยกสาหรายสเขยวจากนาเสย
ผลการคดแยกสาหรายสเขยวจากนาเสยทเกบจากโรงงานอตสาหกรรมอาหาร ในเขตจงหวดพระนครศรอยธยา
จานวน 3 โรงงาน ไดแก โรงงานผลตแปงพซซา โรงงานผลตอาหารแชแขง และโรงงงานผลตภณฑนม สามารถคดแยก
สาหรายสเขยวไดทงหมด 76 ไอโซเลท เมอศกษาลกษณะสณฐานวทยาภายใตกลองจลทรรศน สามารถจดกลมสาหรายสเขยว
ไดทงหมด 11 กลม โดยมเซลลทมรปรางกลม รปรางร รปรางรเรยงตดกน และรปรางพระจนทรเสยว โดยในชวงหลายปท
ผานมามการศกษาการคดแยกสาหรายสเขยวจากแหลงนาเสยตางๆ เชน Oocystis sp. คดแยกไดจากนาเสยฟารมสกร
(Riano et al., 2011) Chlorella sp. คดแยกจากนาเสยโรงงานสรา (Farooq et al., 2013) Chlorella vulgaris YSW-04
คดแยกจากนาเสยฟารมสกร (Ji et al., 2013) Micractinium reisseri และ Scenedesmus obliquus ซงคดแยกไดจาก
นาเสยเทศบาล (Abou-Shana et al., 2014) เพอนาสาหรายเหลานไปใชประโยชนในดานตางๆ เชน ผลตเปนพลงงาน
ทดแทน อาหารสตว และใชสาหรบบาบดนาเสย เปนตน โดยในการทดลองนสามารถแยกสาหรายสเขยวไดหลากหลาย สาย
พนธทาใหมความเปนไปไดทจะมสาหรายสายพนธทมความสามารถในการบาบดนาเสยได

การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
322
ผลการศกษาการเจรญเตบโตของสาหรายสเขยวทคดแยกได
จากการนาสาหรายสเขยวทกไอโซเลทมาทาการศกษาการเจรญเตบโตโดยวดคา OD750 และวดปรมาณคลอโรฟลล
เมอเพาะเลยงในอาหาร TAP ผลการทดลองพบวาสาหรายสเขยวไอโซเลท FFP 13 ทคดแยกไดจากนาเสยในโรงงาน
อตสาหกรรมอาหารแชแขงมคาการเจรญจากวดคา OD750 สงสดเทากบ 3.87±0.02 และมปรมาณคลอโรฟลลสงสดเทากบ
216.17±0.120 ไมโครกรมคลอโรฟลลตอลตร หลงจากเพาะเลยงเปนเวลา 96 ชวโมง (ไมไดแสดงขอมล) โดยสาหรายสเขยว
FFP 13 มลกษณะเซลลรปรางกลม ขนาดเสนผานศนยกลางประมาณ 3 ไมโครเมตร ไมมโครงสรางทใชในการเคลอนท
(Figure 1) เมอเปรยบเทยบรปรางและขนาดของเซลลตามอนกรมวธานของ Leliaert et al. (2012) พบวาสาหราย สเขยวไอ
โซเลท FFP 13 จดอยในจนส Chlorella sp. ทงนในอนาคตทมวจยจะศกษาสายพนธของสาหรายสเขยว FFP 13 ดวยวธการ
ศกษายน 18S rDNA ตอไป นอกจากนจากการทสาหรายสเขยว FFP 13 มการเจรญเตบโตทด และมปรมาณ คลอโรฟลลสง
จงคดเลอกสาหรายสเขยวไอโซเลท FFP 13 มาศกษาการบาบดนาเสยตอไป
ผลการวดองคประกอบของนาเสย
จากการทดลองนานาเสยจากโรงงานผลตอาหารแชแขงมาทาการศกษาการบาบดนาเสยดวยสาหรายสเขยว FFP 13
เนองจากเปนสาหรายทคดแยกไดจากนาเสยในโรงงานอตสาหกรรมแหงน และคาดวาสาหรายสเขยวสายพนธนจะสามารถ
เจรญและบาบดนาเสยได โดยจากการวดองคประกอบของนาเสยทนทหลงจากเกบตวอยาง พบวานาเสยมอณหภม
30.50±0.15 องศาเซลเซยส คาพเอชเทากบ 8.10±0.12 และมคาดโอเทากบ 1.20±0.10 มลลกรมออกซเจนตอลตร จากนน
นานาเสยมากรองปรบพเอชเทากบ 7 นงฆาเชอและวดคาดโอ บโอด ซโอด ไนเตรท และฟอสเฟต พบวานาเสยมคา ดโอ บโอด
และซโอดเทากบ 1.39±0.01 84.00±1.00 และ 522.00±3.00 มลลกรมออกซเจนตอลตร คาไนเตรท และฟอสเฟต เทากบ
61.70±1.25 และ 5,195.00±3.50 ไมโครกรมตอลตร จากผลการทดลองชใหเหนวานาเสยดงกลาวมความเหมาะสม ตอการ
นาไปใชเพาะเลยงสาหรายเพอบาบดนาเสย โดยสารอาหารในนาเสย เชน คารบอนอนทรย ไนโตรเจน และฟอสฟอรส สามารถ
เปนแหลงสารอาหารทสาคญตอการเจรญของสาหรายได (Pittman et al., 2011)
อทธพลของความเขมขนนาเสยตอการบดบดนาเสยโดยสาหรายสเขยวไอโซเลท FFP 13
จากการเพาะเลยงสาหรายสเขยวในนาเสยทแปรผนความเขมขนตางๆ คอ รอยละ 100 90 80 70 และ 60 เปน
ระยะเวลา 5 วน พบวานาเสยความเขนขนรอยละ 100 สาหรายสเขยว FFP 13 มคาการเจรญเตบโต และมปรมาณ ชวมวลสง
ทสด (ไมไดแสดงขอมล) และยงสามารถลดคาบโอดจาก 84.00±1.00 เหลอ 62.00±2.00 มลลกรมออกซเจนตอลตร ลดคาซ
โอดจาก 522.00±3.00 เหลอ 208.00±2.00 มลลกรมออกซเจนตอลตร ลดคาไนเตรทจาก 61.70±1.25 เหลอ 35.85 ±2.12
ไมโครกรมตอลตร และลดคาฟอสเฟตจาก 5,192.00±3.50 เหลอ 1,374.00±3.20 ไมโครกรมตอลตร นอกจากนยง สามารถ
เพมคาดโอจาก 1.39±0.01 เปน 6.80±0.12 มลลกรมออกซเจนตอลตร (Table 1) ชใหเหนวานาเสยทไมไดเจอจาง จะม
สารอนทรยคารบอน ไนเตรท และฟอสเฟต ในปรมาณสง ซงอาจเปนไปไดวาสาหรายสเขยว FFP 13 มความตองการสาร
อาหารเหลานในปรมาณมากเพอนาไปใหในการเจรญเตบโต ประกอบกบเซลลของสาหราย FFP 13 มขนาดเลกจงเพมจานวน
ไดอยางรวดเรว ทาใหชวยลดปรมาณสารอนทรย ไนเตรท และฟอสเฟตในนาเสยได
อทธพลของคาพเอชตอการบาบดนาเสยโดยสาหรายสเขยวไอโซเลท FFP 13
จากการแปรผนคาพเอชในนาเสยทไมเจอจาง โดยใหมคาพเอชเทากบ 5 6 7 8 และ 9 และทดลองเพาะเลยง
สาหราย FFP 13 เปนเวลา 5 วน ผลการทดลองแสดงดง Table 2 จากการทดลองพบวาเมอเพาะเลยงสาหรายสเขยว FFP 13
ในนาเสยทไมเจอจางทคาพเอช 8 สาหรายสเขยว FFP 13 สามารถบาบดนาเสยไดดทสด และแตกตางจากคาพเอช อนอยางม
นยสาคญทางสถต โดยสามารถลดคาบโอดจาก 84.00±1.00 เหลอ 55.00±2.00 มลลกรมออกซเจนตอลตร ลดคาซโอดจาก
522.00±3.00 เหลอ 170.00±1.00 มลลกรมออกซเจนตอลตร ลดคาไนเตรทจาก 61.70±1.25 เหลอ 30.85±1.12 ไมโครกรม
ตอลตร และลดคาฟอสเฟตจาก 5,192.00±2.50 เหลอ 1,340.00±1.30 ไมโครกรมตอลตร โดยทวไปแลวคาพเอชมผลตอการ
เจรญและการบาบดนาเสยในสาหรายสเขยว โดยคาพเอชทเหมาะสมสวนใหญอยในชวง 6-8 และสาหรายแตละสายพนธจะ

การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
323
เจรญไดดในชวงพเอชทแตกตางกน (Larsdutter, 2006) ซงการทดลองนชใหเหนวาทคาพเอช 8 เปนคาพเอชทเหมาะสมใน
การบาบดนาเสยโดยสาหรายสเขยว FFP 13
อทธพลของอณหภมตอการบาบดนาเสยโดยสาหรายสเขยวไอโซเลท FFP 13
จากการใชนาเสยทไมเจอจาง ทพเอช 8 เปนสภาวะทเหมาะในการบาบดนาเสย จงเลอกสภาวะดงกลาวมาแปรผน
อณหภมในการเพาะเลยง โดยแปรผนท 25 30 35 และ 40 องศาเซลเซยส เพาะเลยงเปนเวลา 5 วน ผลการทดลองแสดงดง
Table 3 จากการทดลองพบวาเมอเพาะเลยงสาหรายสเขยว FFP 13 ในนาเสยทไมเจอจางทคาพเอช 8 และอณหภม 30 องศา
เซลเซยส สาหรายสเขยว FFP 13 สามารถบาบดนาเสยไดดทสด และแตกตางจากอณหภมอนอยางมนยสาคญทางสถต โดย
สอดคลองกบงานวจยของ Wang and Lan (2011) ทเพาะเลยงสาหรายสเขยว Neochloris oleoabundans ในนาเสย
เทศบาลทอณหภม 30 องศาเซลเซยส ทาใหสาหรายมการเจรญและสามารถบาบดนาเสยไดดทสด โดยในการทดลองน สาหราย
สเขยว FFP 13 สามารถลดคาบโอดจาก 84.00±1.00 เหลอ 18.00±3.00 มลลกรมออกซเจนตอลตร และลดคาซโอด จาก
522.00±3.00 เหลอ 118.00±2.00 มลลกรมออกซเจนตอลตร ซงคดเปนคาประสทธภาพในการกาจดคาบโอด และซโอด
เทากบรอยละ 78.57 และ 77.39 ตามลาดบ โดยนาเสยทไดจากการบาบดมคาบโอด และซโอด ผานเกณฑมาตรฐานนาทง
โรงงานอตสาหกรรม (เกณฑมาตรฐานนาทงโรงงานอตสาหกรมมคาบโอดและซโอดตองไมเกน 20 และ 120 มลลกรม
ออกซเจนตอลตร ตามลาดบ) นอกจากนสภาวะดงกลาวสาหรายสเขยว FFP 13 ยงสามารถลดคาไนเตรทจาก 61.70±1.25
เหลอ 23.10±1.53 ไมโครกรมตอลตร และลดคาฟอสเฟตจาก 5,195.00±2.50 เหลอ 1,290±1.30 ไมโครกรมตอลตร ซงคด
เปนคาประสทธภาพในการกาจดไนเตรทและฟอสเฟตไดรอยละ 62.56 และ 75.16 ตามลาดบ ผลการทดลองดงกลาว
สอดคลองกบการรายงานกอนหนานในสาหรายสเขยวทมความสามารถสงในการกาจดไนเตรท และฟอสเฟตออกจากนาเสย
เชน Chlorella sorokiniana ทเพาะเลยงในนาเสยฟารมสกร (Hernández et al., 2013) และ Coelastrum microporum
ทเพาะเลยงในนาเสยเทศบาล (Lee et al., 2015) โดยปกตแลวสภาวะตางๆ ในนาเสย เชน ความเขมขนของ นาเสย คาพเอช
และอณหภม มผลอยางมากตอการเจรญและการบาบดนาเสยในสาหรายสเขยว โดยสาหรายแตละสายพนธ ตองการสภาวะท
เหมาะสมในการบาบดนาเสยทแตกตางกน (Molinuevo–Salces et al., 2019) จากการทดลองชใหเหนวา การใชนาเสยทไม
เจอจาง คาพเอช 8 และเพาะเลยงทอณหภม 30 องศาเซลเซยส มความเหมาะสมตอการบาบดนาเสยโดย สาหรายสเขยว FFP
13 อยางไรกตามการทดลองนใชนาเสยทผานการฆาเชอแลว ดงนนในอนาคตทมวจยจะทดลองเพาะ เลยงสาหรายสเขยวในนา
เสยทไมผานการฆาเชอเพอจาลองสภาวะจรงในการบาบดนาเสยตอไป
สรป
จากการคดแยกสาหรายสเขยวในโรงงานอตสาหกรรมอาหาร 3 แหงสามารถคดแยกสาหรายสเขยวได 76 ไอโซเลท
และสามารถแบงกลมตามลกษณะสณฐานวทยาได 11 กลม จากการศกษาการเจรญเตบโตพบวาสาหรายสเขยวไอโซเลท FFP
13 มการเจรญและมปรมาณคลอโรฟลลสงสด จากนนนาไอโซเลท FFP 13 มาตรวจสอบการบาบดนาเสย โดยศกษาอทธพล
ของความเขมขนนาเสย พเอช และอณหภม พบวาสภาวะทเหมาะสมในการบาบดนาเสย คอการใชนาเสยทไมเจอจาง คาพเอช
เทากบ 8 และอณหภม 30 องศาเซลเซยส ภายใตสภาวะดงกลาวสาหรายสเขยวไอโซเลท FFP 13 มประสทธภาพสงในการ
กาจดคาบโอด ซโอด ไนเตรท และฟอสเฟต โดยนาเสยทไดจากการบาบดผานเกณฑมาตรฐานนาทงโรงงานอตสาหกรรม แสดง
ใหเหนวาสามารถนาสาหรายสเขยว FFP 13 ไปบาบดนาเสยจากโรงงานอตสาหกรรมอาหารไดในอนาคต

การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
324
กตตกรรมประกาศ
งานวจยนไดรบการสนบสนนจากคณะวทยาศาสตรและเทคโนโลย มหาวทยาลยเทคโนโลยราชมงคลสวรรณภม และ
ขอขอบคณสาขาวชาวทยาศาสตร (ชววทยา) คณะวทยาศาสตรและเทคโนโลย มหาวยาลยเทคโนโลยราชมงคลสวรรณภม ศนย
นนทบร และศนยพระนครศรอยธยาหนตรา ทใหความอนเคราะหสถานทและเครองมอในการทางานวจย
เอกสารอางอง
Abou-Shanab, R.A.I., M.M. El-Dalatony, M.M. EL-Sheekh, M.K. Ji, E.S. Salama, A.N. Kabra and B.H. Jeon. 2014.
Cultivation of a new microalga, Micractinium reisseri, in municipal wastewater for nutrient removal,
biomass, lipid, and fatty acid production. Biotechnol. Bioprocess. Eng. 19: 510 – 518.
American Public Health Association (APHA). 1998. Methods for biomass production. In: Standard Methods
for the Examination of Water and Wastewater. American Public Health Association, Baltimore MD.
Aslan, S. and I.K. Kapdan. 2006. Batch kinetics of nitrogen and phosphorus removal from synthetic
wastewater by algae. Ecol. Eng. 28: 64 – 70.
Farooq, W., Y.C. Lee, B.G. Ryu, B.H. Kim, H.S. Kim, Y.E. Choi and J.W. Yang. 2013. Two stage cultivation of
two Chlorella sp. strains by simultaneous treatment of brewery wastewater and maximizing lipid
productivity. Bioresour. Technol. 132: 230 – 238.
Harris E.H. 1989. The Chlamydomonas sourcebook: a comprehensive guide to biology and laboratory use.
San Diego: Academic Press.
Hernández, D., B. Riaño, M. Coca and M.C. García-González. 2013. Treatment of agro-industrial wastewater
using microalgae–bacteria consortium combined with anaerobic digestion of the produced biomass.
Bioresour. Technol. 135: 598 – 603.
Ji, M.K., R.A.I. Abou-Shanab, S.H. Kim, E.S. Salama, S.H. Lee, N.A. Kabra, Y.S. Lee, S. Hongf and B,H, Jeon.
2013. Cultivation of microalgae species in tertiary municipal wastewater supplemented with
CO2 for nutrient removal and biomass production. Eco. Eng. 58: 142 – 148.
Larsdotter, K. 2006. Wastewater treatment with microalgae: a literature review. Vatten. 62: 31 – 38.
Lee, C.S., S.A. Lee, S.R. Ko, H.M. Oh and C.Y. Ahn. 2015. Effects of photoperiod on nutrient removal,
biomass production, and algal-bacterial population dynamics in lab-scale photobioreactors
treating municipal wastewater. Water. Res. 68: 680 – 691.
Leliaert, F., D.R. Smith, H. Moreau, M.D. Herron, H. Verbruggen, C.F. Delwiche and O.D. Clerck. 2012.
Phylogeny and molecular evolution of the green algae. CRC. Crit. Rev. Plant. Sci. 31: 1 – 46.
Molinuevo-Salces, B., B. Riaño, D. Hernández and M.C. García-González. 2019. Microalgae and wastewater
treatment: advantages and disadvantages. In: Alam M., Wang Z. (eds) Microalgae biotechnology for
development of biofuel and wastewater treatment. Springer, Singapore. pp. 505 – 533.
Pittman, J.K., A.P. Dean and O. Osundeko. 2011. The potential of sustainable algal biofuel production using
wastewater resources. Bioresour. Technol. 102: 17 – 25.
Riaño, B., B. Molinuevo and M.C. García-González. 2011. Treatment of fish processing wastewater with
microalgae-containing microbiota. Bioresour. Technol. 102: 10829 – 10833.

การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
325
Riemann, B. and D. Ernst. 1982. Extraction of chlorophylls a and b from phytoplankton using standard
extraction techniques. Freshwater. Biol. 12: 217 – 223.
Rippka, R., J. Deruelles, J.B. Waterbury, M. Herdman and R.Y. Stanier. 1979. Generic assignments, strain
histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 111: 1 – 61.
Wang, B. and C.Q. Lan. 2011. Biomass production and nitrogen and phosphorus removal by the green alga
Neochloris oleoabundans in simulated wastewater and secondary municipal wastewater effluent.
Bioresour. Technol. 102: 5639 – 5644.
Figure 1 Cell morphology of green algae isolate FFP 13 observed under light microscope.
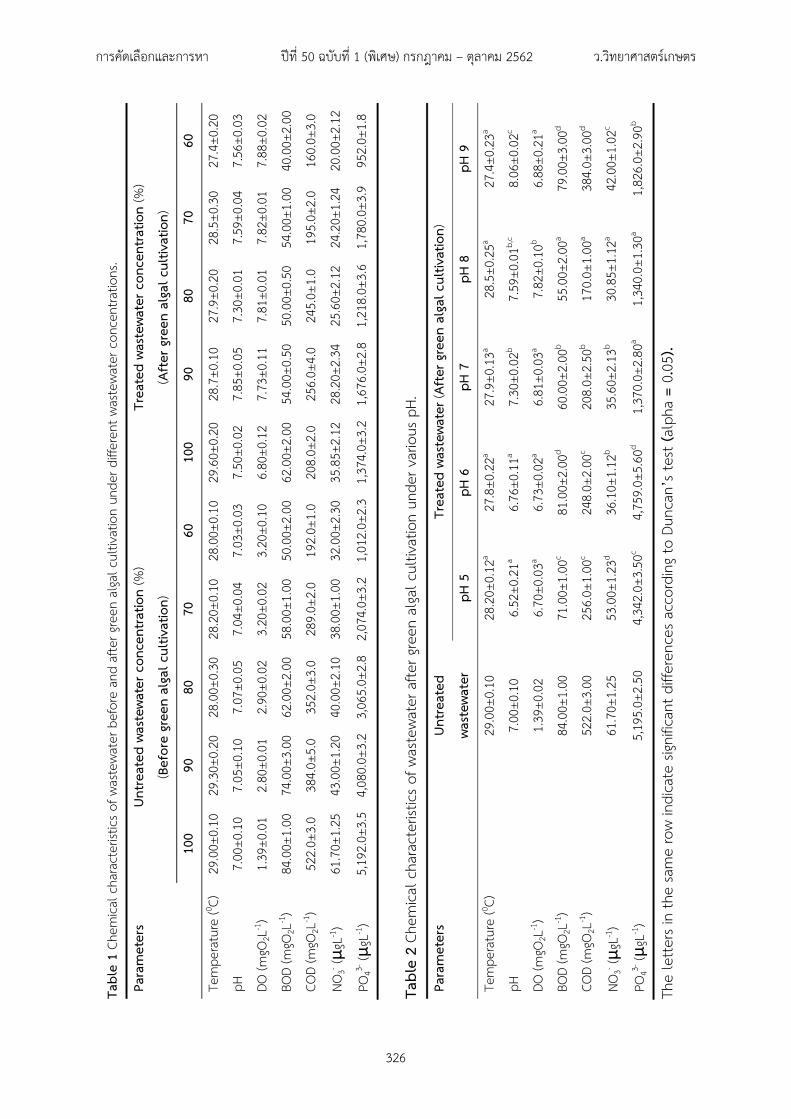
การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
326
Tabl
e 1
Chem
ical
cha
ract
erist
ics
of w
aste
wat
er b
efor
e an
d af
ter g
reen
alg
al c
ultiv
atio
n un
der d
iffer
ent w
aste
wat
er c
once
ntra
tions
.
Para
met
ers
Unt
reat
ed w
aste
wat
er c
once
ntra
tion
(%)
(Bef
ore
gree
n al
gal c
ultiv
atio
n)
Trea
ted
was
tew
ater
con
cent
ratio
n (%
)
(Aft
er g
reen
alg
al c
ultiv
atio
n)
100
90
80
70
60
100
90
80
70
60
Tem
pera
ture
(0 C)
29.0
0±0.
10
29.3
0±0.
20
28.0
0±0.
30
28.2
0±0.
10
28.0
0±0.
10
29.6
0±0.
20
28.7
±0.1
0 27
.9±0
.20
28.5
±0.3
0 27
.4±0
.20
pH
7.00
±0.1
0 7.
05±0
.10
7.07
±0.0
5 7.
04±0
.04
7.03
±0.0
3 7.
50±0
.02
7.85
±0.0
5 7.
30±0
.01
7.59
±0.0
4 7.
56±0
.03
DO (m
gO2L
-1)
1.39
±0.0
1 2.
80±0
.01
2.90
±0.0
2 3.
20±0
.02
3.20
±0.1
0 6.
80±0
.12
7.73
±0.1
1 7.
81±0
.01
7.82
±0.0
1 7.
88±0
.02
BOD
(mgO
2L-1)
84.0
0±1.
00
74.0
0±3.
00
62.0
0±2.
00
58.0
0±1.
00
50.0
0±2.
00
62.0
0±2.
00
54.0
0±0.
50
50.0
0±0.
50
54.0
0±1.
00
40.0
0±2.
00
COD
(mgO
2L-1)
522.
0±3.
0 38
4.0±
5.0
352.
0±3.
0 28
9.0±
2.0
192.
0±1.
0 20
8.0±
2.0
256.
0±4.
0 24
5.0±
1.0
195.
0±2.
0 16
0.0±
3.0
NO3- (μ
gL-1)
61.7
0±1.
25
43.0
0±1.
20
40.0
0±2.
10
38.0
0±1.
00
32.0
0±2.
30
35.8
5±2.
12
28.2
0±2.
34
25.6
0±2.
12
24.2
0±1.
24
20.0
0±2.
12
PO43-
(μgL
-1)
5,19
2.0±
3.5
4,08
0.0±
3.2
3,06
5.0±
2.8
2,07
4.0±
3.2
1,01
2.0±
2.3
1,37
4.0±
3.2
1,67
6.0±
2.8
1,21
8.0±
3.6
1,78
0.0±
3.9
952.
0±1.
8
Tabl
e 2
Chem
ical c
hara
cter
istics
of w
aste
wat
er a
fter g
reen
alg
al c
ultiv
atio
n un
der v
ario
us p
H.
Para
met
ers
Unt
reat
ed
was
tew
ater
Trea
ted
was
tew
ater
(Aft
er g
reen
alg
al c
ultiv
atio
n)
pH 5
pH
6
pH 7
pH
8
pH 9
Tem
pera
ture
(0 C)
29.0
0±0.
10
28.2
0±0.
12a
27.8
±0.2
2a 27
.9±0
.13a
28.5
±0.2
5a 27
.4±0
.23a
pH
7.00
±0.1
0 6.
52±0
.21a
6.76
±0.1
1a 7.
30±0
.02b
7.59
±0.0
1b,c
8.06
±0.0
2c
DO (m
gO2L
-1)
1.39
±0.0
2 6.
70±0
.03a
6.73
±0.0
2a 6.
81±0
.03a
7.82
±0.1
0b 6.
88±0
.21a
BOD
(mgO
2L-1)
84.0
0±1.
00
71.0
0±1.
00c
81.0
0±2.
00d
60.0
0±2.
00b
55.0
0±2.
00a
79.0
0±3.
00d
COD
(mgO
2L-1)
522.
0±3.
00
256.
0±1.
00c
248.
0±2.
00c
208.
0±2.
50b
170.
0±1.
00a
384.
0±3.
00d
NO3- (μ
gL-1)
61.7
0±1.
25
53.0
0±1.
23d
36.1
0±1.
12b
35.6
0±2.
13b
30.8
5±1.
12a
42.0
0±1.
02c
PO43-
(μgL
-1)
5,19
5.0±
2.50
4,
342.
0±3.
50c
4,75
9.0±
5.60
d 1,
370.
0±2.
80a
1,34
0.0±
1.30
a 1,
826.
0±2.
90b
The
lett
ers
in th
e sa
me
row
indi
cate
sign
ifica
nt d
iffer
ence
s ac
cord
ing
to D
unca
n’s
test
(alp
ha =
0.0
5).
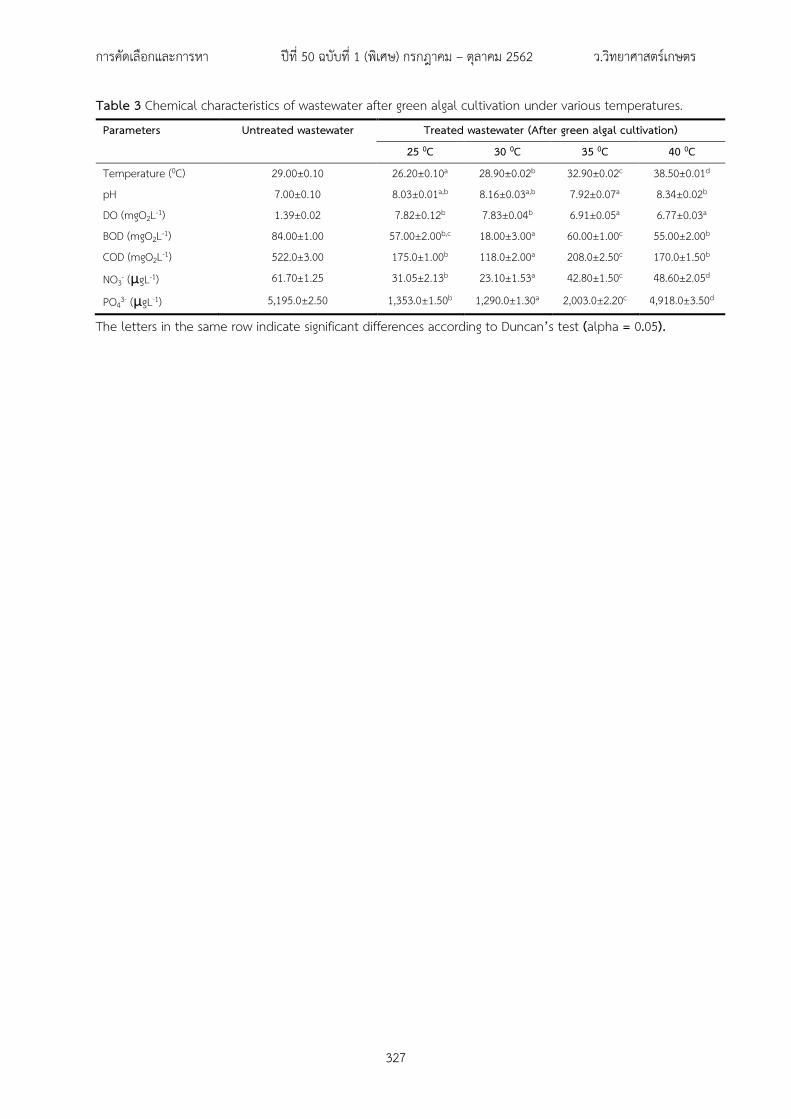
การคดเลอกและการหา ปท 50 ฉบบท 1 (พเศษ) กรกฎาคม – ตลาคม 2562 ว.วทยาศาสตรเกษตร
327
Table 3 Chemical characteristics of wastewater after green algal cultivation under various temperatures.
Parameters Untreated wastewater Treated wastewater (After green algal cultivation)
25 0C 30 0C 35 0C 40 0C
Temperature (0C) 29.00±0.10 26.20±0.10a 28.90±0.02b 32.90±0.02c 38.50±0.01d
pH 7.00±0.10 8.03±0.01a,b 8.16±0.03a,b 7.92±0.07a 8.34±0.02b
DO (mgO2L-1) 1.39±0.02 7.82±0.12b 7.83±0.04b 6.91±0.05a 6.77±0.03a
BOD (mgO2L-1) 84.00±1.00 57.00±2.00b,c 18.00±3.00a 60.00±1.00c 55.00±2.00b
COD (mgO2L-1) 522.0±3.00 175.0±1.00b 118.0±2.00a 208.0±2.50c 170.0±1.50b
NO3- (μgL-1) 61.70±1.25 31.05±2.13b 23.10±1.53a 42.80±1.50c 48.60±2.05d
PO43- (μgL-1) 5,195.0±2.50 1,353.0±1.50b 1,290.0±1.30a 2,003.0±2.20c 4,918.0±3.50d
The letters in the same row indicate significant differences according to Duncan’s test (alpha = 0.05).

International Conference on Power, Energy and Innovation (ICPEI 2019) October 16-18, 2019, Pattaya, THAILAND
High Lipid Content in Unicellular Green Alga Asteracys sp. FFP40 Isolated from Food Industrial Wastewater in Phra Nakhon Si Ayutthaya Province
Samart Taikhao Division of Science (Biology) Faculty
of Science and Technology Rajamangala University of Technology
Suvarnabhumi Nonthaburi, Thailand
Suntara Fueangfung Division of Science (Chemistry) Faculty
of Science and Technology Rajamangala University of Technology
Suvarnabhumi Phra Nakhon Si Ayutthaya, Thailand
Wachiraporn Srisong Division of Microbiology Faculty of
Science and Technology Rajamangala University of Technology
Suvarnabhumi Phra Nakhon Si Ayutthaya, Thailand
Abstract—Green algae are photoautotrophic and photoheterotrophic microorganisms that are capable of lipid production. This study aims to isolate green algae from food industrial wastewater in Phra Nakhon Si Ayutthaya province. Thirty isolates of green algae were isolated from wastewater of four food industrial plants. Lipid content under cultivated cells in N-deprivation Tris Acetate Phosphate TAP(-N) medium was measured in order to select the green algae that had the highest lipid production. The results found that among 30 green algal strains, the green alga Asteracys sp. FFP40 showed the highest lipid content of 2.884±0.064 % (w/w) under cultivated cells in TAP(-N) medium for 3 days. During photoheterotrophic cultivation in TAP medium, Asteracys sp. FFP40 was rapidly grown to the stationary growth phase after 48 h. The biomass of Asteracys sp. FFP40 was continuously increased and reached to 0.966±0.115 mg ml-1 after 120 h of cultivation. Finally, Asteracys sp. FFP40 also provided the highest lipid content of 58.446±0.652 % (w/w) under cultivated cells in TAP(-N) medium for 28 days. These results indicated that Asteracys sp. FFP40 could be a source of highly efficient lipid production for biodiesel evolution in the future.
Keywords— Lipid content, Green algae, Food industrial wastewater
I. INTRODUCTION
Nowadays, the world population has rapidly increased, resulting in rising demand for fossil fuels. However, they are not long-term sustainable energy resources. It is because their consumption can cause the emission of greenhouse gases, and resulting in global warming [1]. Therefore, many researchers currently are concentrating on environment-friendly biofuels production for alternative energy sources [2].
Not only green algae can produce biofuels but also capture CO2 from the atmosphere, indirectly decreasing one of the primary greenhouse gases. Green algae can be used as a raw material source to produce biofuels in various forms such as ethanol [3], bioelectricity [4], hydrogen [5-6], methane [7] and biodiesel [8-9].
Currently, the production of biodiesel from green algae shows a good advantage because of their higher amount of biomass and lipid yields comparing to traditional oilseed crops [10-12]. Furthermore, green algae can perform both roles of wastewater treatment and lipid-containing biomass production [13]. Several researchers have found that some
978-1-7281-5266-0/19/$31.00 ©2019 IEEE
green algal species isolated form wastewater treatment plant sites can be better suited to local environments than non-native species [14-15]. Many green algae isolated from wastewater have been shown to produce lipid such as Coelastrum sp. isolated from cattle manure leachate [16], Coelastrella sp., Chlorella vulgaris and Desmodesmus sp. isolated from Swedish fresh and wastewater sources [17] and Micractinium reisseri isolated from municipal wastewater mixed with agricultural drainage [18].
In Thailand, many studies have been focused on lipids production of natural freshwater green algae such as Botryococcus sp. [19], Chlorella vulgaris [20], Chlorococcum humicola [21], Coelastrum microsporum [22] and Scenedesmus acutus [21]. Since there is large amount wastewater from many food industrial plants in Phra Nakhon Si Ayutthaya province, it is possible to discover diverse kinds of green algae and one of them might show a high lipid content.
The aims of this research are to screen high lipid content of green algae isolated from food industrial wastewater in Phra Nakhon Si Ayutthaya province, to investigate its strain, growth and biomass under the photoheterotrophic cultivation, and to examine its lipid content under various cultivation time period.
II. MATERIALS AND METHODS
A. Sampling, purification and cultivation of green algaeThirty green algal strains were isolated from wastewater
in open pond of four food industry plants in Phra Nakhon Si Ayutthaya province. They were purified by single cell isolation techique [23]. The purified strains were grown in a 250-mL Erlenmeyer flask containing 100 mL of Tris-Acetate-Phospate (TAP) medium (pH 7.2) [24]. The algalcells were cultivated at 30 0C under white-light illuminationof 30 µmol photon m-2 s-1 and shaking speed of 120 rpm for5 days.
B. Growth and dry cell weight determinationThe optical density of 750 nm (OD750) was used to
monitor the algal growth. After that, dry cell weight was determined by filtering 5 mL of cell culture through a filter paper No 1 (55 mm diameter) (Whatman, UK). The cells were washed twice in distilled water, then dried at 85 0C in an oven for 16 h before weighting.
36

International Conference on Power, Energy and Innovation (ICPEI 2019) October 16-18, 2019, Pattaya, THAILAND
C. Pepararing cell for lipid accumulationAll green algal strains were grown in TAP mdeium for 3
days to accumulate biomass. After that, 3 days old cell were harvested by centrifugation at 5,000 rpm for 5 min at 4 0C, washed twice and resuspended in 100 mL of N-free TAP (TAP-N) medium and further incubated for 3 days before harvesting cell to determine lipid content.
D. Lipid extractionGreen algal cells were harvested by cetrifugation at 5,000
rpm for 5 min at 4 0C, rinsed with distilled deionized water, then dried in an oven at 70 0C for 24 h. The extraction method of Shin and co-workers (2018) was used to obtain the total lipid content [25]. Approximately 1.0 g of dry cells was extracted in triplicate with 20 mL hexanes at room temperature for 24 h. Then, the extract solution was filtered to separate cell debris, and transferred to round bottom flask. The solution was evaporated by rotary evaporator (Buchi, Rotavapor R-3, Switzerland) to yield the lipid content. Finally, the data were analyzed with SPSS 13.0 (SPSS Inc.) using one-way ANOVA.
III. RESULTS AND DISCUSSIONS
A. Selection of high lipid-producing green algal strainsThirty green algal strains were isolated from wastewater
of four food industrial plants in Phra Nakhon Si Ayutthaya province, Thailand. The water was retrieved from convenience food production industry (CFP), dairy products production industry (DPP), frozen food production industry (FFP) and pizza dough production industry (PDP). All green algal cells were grown in TAP medium for 3 days, then harvested by centrifugation, washed twice, and resuspended in 100 mL of TAP(-N) medium and further cultivated by shaking at 30 0C under light for the next 3 days. The algal cells were harvested, dried and their lipid content was measured. The results revealed that all green algal strains could possibly lipid (Table I). Among them, the green algal isolate CFP13, FFP9 and FFP40 showed significantly high lipid content under nitrogen deprivation conditions (Table I). The highest lipid content was found in isolate FFP40 with 2.884±0.064 % (w/w) under cultivated cells in TAP(-N) medium for 3 days (Table I). From previous literature, there are a number of lipid-containing green algae isolated from different sources of wastewater. For example, Coelastrum sp. was isolated from cattle manure leachate [16]. Coelastrella sp., Chlorella vulgaris and Desmodesmus sp. were isolated from Swedish fresh and wastewater sources [17]. Micractinium reisseri was isolated from municipal wastewater mixed with agricultural drainage [18]. Typically, these green algae isolated from wastewater were used for wastewater treatment. Therefore, wastewater would be served as medium which could reduce costs for biomass and and lipid production. Additionally, COD, nitrate and phosphate from wastewater were also removed by those green algae.
TABLE I. LIPID PRODUCTION BY 30 GREEN ALGAL STRAINS ISOLATED FROM FOOD INDUSTRIAL WASTEWATER IN PHRA NAKHON SI
AYUTTHAYA PROVINCE, THAILAND
Green algal strains
Origin of industrial wastewater
Lipid content % (w/w)
CFP3 Convenience food production 0.912± 0.023hi CFP4 Convenience food production 0.367± 0.037n CFP5 Convenience food production 1.690± 0.063e CFP6 Convenience food production 1.025 ± 0.067h CFP7 Convenience food production 0.863± 0.052ij CFP8 Convenience food production 0.665±0.045kl CFP9 Convenience food production 0.564±0.050lm
CFP10 Convenience food production 0.427±0.062mn CFP13 Convenience food production 2.727±0.038b CFP15 Convenience food production 0.427±0.067mn CFP21 Convenience food production 0.818±0.056ijk CFP22 Convenience food production 0.794±0.048ijk CFP23 Convenience food production 0.729±0.059jk CFP25 Convenience food production 0.760±0.048ijk CFP27 Convenience food production 1.410±0.047f CFP29 Convenience food production 0.775±0.054ijk CFP30 Convenience food production 1.649±0.068e CFP31 Convenience food production 0.287±0.051no CFP33 Convenience food production 0.209±0.042op DPP14 Dairy products production 1.191±0.117g FFP4 Frozen food production 0.788±0.036ijk FFP9 Frozen food production 2.435±0.055c
FFP13 Frozen food production 0.211±0.019op FFP40 Frozen food production 2.884±0.059a FFP46 Frozen food production 0.301±0.018no FFP48 Frozen food production 0.115±0.024p FFP50 Frozen food production 0.384±0.048n
FFP52.2 Frozen food production 1.962±0.093d PDP3 Pizza dough production 0.680±0.076kl PDP4 Pizza dough production 0.191±0.006op
Different letters indicate significant differences according to Duncan’s test (alpha = 0.05).
B. Growth and biomass of Asteracys sp. FFP40 underheterotophic conditionsThe morphological and genetics characterization of
green alga FFP40 were classified by using 18S rRNA sequence analysis. The results showed that the isolate FFP40 showed their similarities comparing to the green algae in the genus Asteracys. Asteracys sp. FFP40 was a unicellular green alga and has oval shape with an approximate 8-9 µm cell diameter. No motile flagella were found (Fig.1).
Fig. 1 Cell morphology of Asteracys sp. FFP40 observed under alight microscope.
The algal cells of Asteracys sp. FFP40 were grown in 100 mL of TAP medium and shaken under the light for 5 days. Their growth and biomass were also determined. The results illustrated that the cells was rapidly grown and reached to stationary growth phase after 48 h (Fig. 2A). Meanwhile, the biomass of Asteracys sp. FFP40 was
37
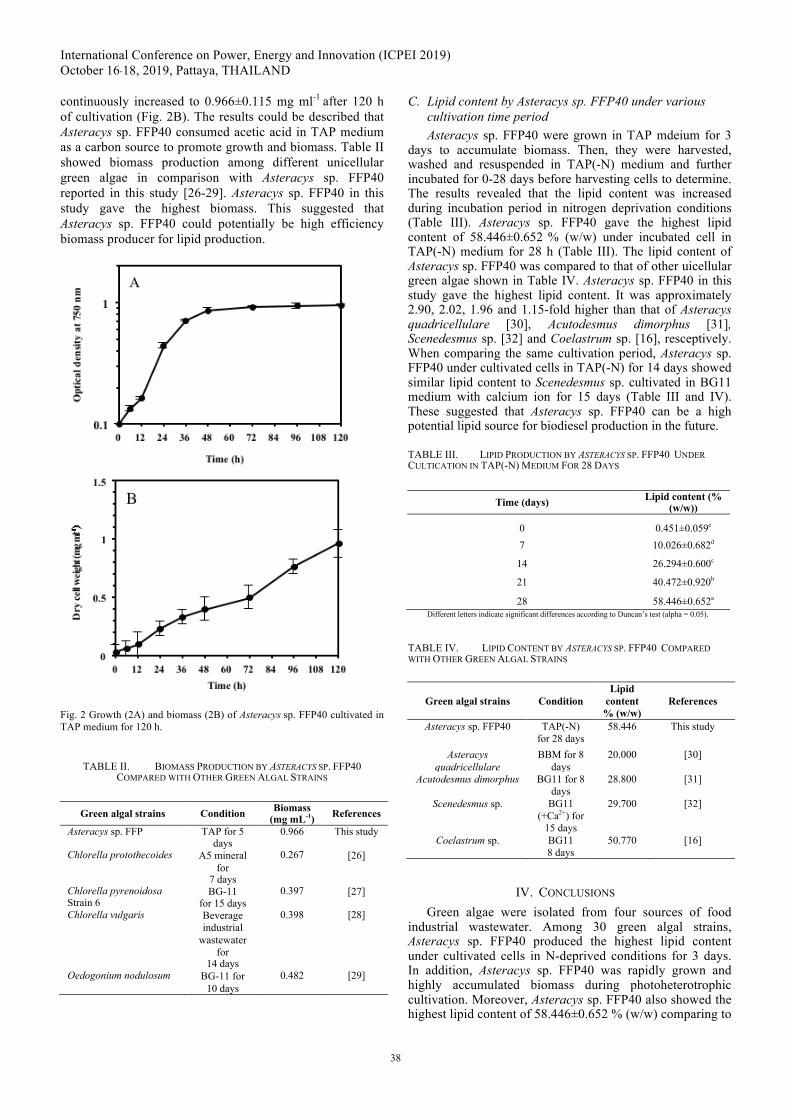
International Conference on Power, Energy and Innovation (ICPEI 2019) October 16-18, 2019, Pattaya, THAILAND
continuously increased to 0.966±0.115 mg ml-1 after 120 h of cultivation (Fig. 2B). The results could be described that Asteracys sp. FFP40 consumed acetic acid in TAP medium as a carbon source to promote growth and biomass. Table II showed biomass production among different unicellular green algae in comparison with Asteracys sp. FFP40 reported in this study [26-29]. Asteracys sp. FFP40 in this study gave the highest biomass. This suggested that Asteracys sp. FFP40 could potentially be high efficiency biomass producer for lipid production.
Fig. 2 Growth (2A) and biomass (2B) of Asteracys sp. FFP40 cultivated in TAP medium for 120 h.
TABLE II. BIOMASS PRODUCTION BY ASTERACYS SP. FFP40 COMPARED WITH OTHER GREEN ALGAL STRAINS
C. Lipid content by Asteracys sp. FFP40 under variouscultivation time periodAsteracys sp. FFP40 were grown in TAP mdeium for 3
days to accumulate biomass. Then, they were harvested, washed and resuspended in TAP(-N) medium and further incubated for 0-28 days before harvesting cells to determine. The results revealed that the lipid content was increased during incubation period in nitrogen deprivation conditions (Table III). Asteracys sp. FFP40 gave the highest lipid content of 58.446±0.652 % (w/w) under incubated cell in TAP(-N) medium for 28 h (Table III). The lipid content of Asteracys sp. FFP40 was compared to that of other uicellular green algae shown in Table IV. Asteracys sp. FFP40 in this study gave the highest lipid content. It was approximately 2.90, 2.02, 1.96 and 1.15-fold higher than that of Asteracys quadricellulare [30], Acutodesmus dimorphus [31], Scenedesmus sp. [32] and Coelastrum sp. [16], resceptively. When comparing the same cultivation period, Asteracys sp. FFP40 under cultivated cells in TAP(-N) for 14 days showed similar lipid content to Scenedesmus sp. cultivated in BG11 medium with calcium ion for 15 days (Table III and IV). These suggested that Asteracys sp. FFP40 can be a high potential lipid source for biodiesel production in the future.
TABLE III. LIPID PRODUCTION BY ASTERACYS SP. FFP40 UNDER CULTICATION IN TAP(-N) MEDIUM FOR 28 DAYS
Different letters indicate significant differences according to Duncan’s test (alpha = 0.05).
TABLE IV. LIPID CONTENT BY ASTERACYS SP. FFP40 COMPARED WITH OTHER GREEN ALGAL STRAINS
IV. CONCLUSIONS
Green algae were isolated from four sources of food industrial wastewater. Among 30 green algal strains, Asteracys sp. FFP40 produced the highest lipid content under cultivated cells in N-deprived conditions for 3 days. In addition, Asteracys sp. FFP40 was rapidly grown and highly accumulated biomass during photoheterotrophic cultivation. Moreover, Asteracys sp. FFP40 also showed the highest lipid content of 58.446±0.652 % (w/w) comparing to
Green algal strains Condition Biomass (mg mL-1) References
Asteracys sp. FFP TAP for 5 days
0.966 This study
Chlorella protothecoides A5 mineral for
7 days
0.267 [26]
Chlorella pyrenoidosa Strain 6
BG-11 for 15 days
0.397 [27]
Chlorella vulgaris Beverage industrial
wastewater for
14 days
0.398 [28]
Oedogonium nodulosum BG-11 for 10 days
0.482 [29]
Time (days) Lipid content (% (w/w))
0 0.451±0.059e 7 10.026±0.682d
14 26.294±0.600c
21 40.472±0.920b
28 58.446±0.652a
Green algal strains Condition Lipid
content % (w/w)
References
Asteracys sp. FFP40 TAP(-N) for 28 days
58.446 This study
Asteracys quadricellulare
BBM for 8 days
20.000 [30]
Acutodesmus dimorphus BG11 for 8 days
28.800 [31]
Scenedesmus sp. BG11 (+Ca2+) for
15 days
29.700 [32]
Coelastrum sp. BG11 8 days
50.770 [16]
38

International Conference on Power, Energy and Innovation (ICPEI 2019) October 16-18, 2019, Pattaya, THAILAND
several green algae. From these results, it is possible to use biomass of green alga Asteracys sp. FFP40 as a raw material for biodiesel production in the future.
Acknowledgment
Financial support form Ministry of Higher Education, Science, Research and Innovation and National Science and Technology Development Agency (NSTDA) are gratefully acknowledged.
REFERENCES
[1] J. Hill, E. Nelson, D. Tilman, S. Polasky, and D. Tiffany,“Environmental, economic, and energetic costs and benefits ofbiodiesel and ethanol biofuels,” PNAS. vol. 103, pp. 11206–11210,
[2] 2006.[3] J. K. Pittman, A. P. Dean, and O. Osundeko, “The potential of
sustainable algal biofuel production using wastewater resources,”Bioresour. Technol. vol. 102, pp. 17–25, 2011.
[4] F. Shirai, K. Kunii, C. Sato, Y. Teramoto, E. Mizuki, S. Murao, andS. Nakayama, “Cultivation of microalgae in the solution from thedesalting process of soy sauce waste treatment and utilization of the algal biomass for ethanol fermentation,” J. Microbiol. Biotechnol.vol. 14, pp. 839–842, 1998.
[5] E. E. Powell, M. L. Mapiour, R. W. Evitts, and G. A. Hill, “Growthkinetics of Chlorella vulgaris and its use as a cathodic half cell,”Bioresour. Technol. vol. 100, pp. 269–274, 2009.
[6] M. L. Ghirardi, “Hydrogen production by photosynthetic greenalgae,” Indian. J. Biochem. Biophys. vol. 43, pp. 201–210, 2006.
[7] A. Hemschemeier, A. Melis, and T. Happe, “Analytical approaches tophotobiological hydrogen production in unicellular green algae,”Photosynth. Res. vol. 102, pp. 523–540, 2009.
[8] S. Stucki, F. Vogel, C. Ludwig, A. G. Haiduc, M. Brandenberger,“Catalytic gasification of algae in supercritical water for biofuelproduction and carbon capture,” Energ. Environ. Sci. vol. 2, pp. 535–541, 2009.
[9] A. Converti, A. A. Casazza, E. Y. Ortiz, P. Perego, and B. M. Del,“Effect of temperature and nitrogen concentration on the growth andlipid content of Nannochloropsis oculata and Chlorella vulgaris forbiodiesel production,” Chem. Eng. Prog. vol. 48, pp. 1146–1151,2009.
[10] C. F. Gao, Y. Zhai, Y. Ding, and Q. Y. Wu, “Application of sweetsorghum for biodiesel production by heterotrophic microalgaChlorella protothecoides,” Appl. Energy. vol. 87, pp. 756–761, 2010.
[11] Y. Chisti, “Biodiesel from microalgae,” Biotechnol. Adv. vol. 25, pp.294–306, 2007.
[12] Y. Li, Y. F. Chen, P. Chen, M. Min, W. Zhou, B. Martinez, J. Zhub,and R. Ruan, “Characterization of a microalga Chlorella sp. welladapted to highly concentrated municipal wastewater for nutrientremoval and biodiesel production,” Bioresour. Technol. vol. 102, pp.5138–5144, 2011.
[13] T. Takeshita, “Competitiveness, role, and impact of microalgalbiodiesel in theglobal energy future,” Appl. Energy. vol. 88, pp.3481–3491, 2011.
[14] I. Rawat, R. R. Kumar, T. Mutanda, and F. Bux, “Dual role ofmicroalgae: phycoremediation of domestic wastewater and biomassproduction for sustainable biofuels production,” Renew. Energy. vol.88, pp. 3411–3424, 2011.
[15] M. V. J. Pérez, P. S. Castillo, O. Romera, D. F. Moreno, and C. P.Martínez, “Growth andnutrient removal in free and immobilizedplanktonic green algae isolated from pig manure,” Enzyme. Microb.Technol. vol. 34, pp. 392–398, 2004.
[16] M. Odlare, E. Nehrenheim, V. Ribe, E. Thorin, M. Gavare, and M.Grube, “Cultivation of algae with indigenous species – potentials forregional biofuel production,” Appl. Energy. vol. 88, pp. 3280–3285,2011.
[17] S. Mousavi, G. D. Najafpour, M. Mohammadi, and M. H. Seifi,“Cultivation of newly isolated microalgae Coelastrum sp. inwastewater for simultaneous CO2 fixation, lipid production andwastewater treatment,” Bioproc. Biosyst. Eng. vol. 41, pp. 519-530,2018.
[18] L. Ferro, F. G. Gentili, and C. Funk, “Isolation and characterization ofmicroalgal strains for biomass production and wastewater reclamationin Northern Sweden,” Agal. Res. vol. 32, pp. 45-53, 2018.
[19] R. Abou-Shanab, M. M. El-Dalatony, M. M. EL-Sheekh, M. K. Ji, E.S. Salama, A. N. Kabra, and B. H. Jeon, “Cultivation of a newmicroalga, Micractinium reisseri, in municipal wastewater fornutrient removal, biomass, lipid, and fatty acid production,”Biotechnol. Bioprocess. Eng. vol. 19, pp. 510-518, 2014.
[20] C. Yeesang, and B. Cheirsilp. “Effect of nitrogen, salt, and ironcontent in the growthmedium and light intensity on lipid productionby microalgae isolated from fresh water sources in Thailand,”Bioresour. Technol. vol. 102, pp. 3034–3040, 2011.
[21] R. Ramaraj, Y. Unpaprom, and N. Dussadee, “Cultivation of greenmicroalga, Chlorella vulgaris for biogas purification,” Int. J. New.Technol. Res. vol. 2, pp. 117-122, 2016.
[22] S. Chaichalerm, P. Pokethitiyook, W. Yuan, M. Meetam, K. Sritong,W. Pugkaew, K. Kungvansaicho, M. Kruatrachue, and P.Damrongphol, “Culture of microalgal strains isolated from naturalhabitats in Thailand invarious enriched media,” Appl. Energ. vol. 89,pp. 296-302, 2012.
[23] T. Prasertsin, and Y. Peerapornpisal, “Distribution and isolation ofmicroalgae for lipid production in selected freshwater reservoirs ofnorthern Thailand,” Biodiversitas. vol. 19, pp. 343-350, 2018.
[24] R. W. Hoshaw, J. R. Rosowksi, “Methods for microscopic algae,” inHandbook of phycological methods culture methods and growthmeasurements, J. Stein, Edi. New York: Cambridge University Press,1973, pp. 53-67.
[25] E. H. Harris, “The Chlamydomonas sourcebook: a comprehensiveguide to biology and laboratory use,” San Diego: Academic Press,1989.
[26] H. Y. Shin, S. H. Shim, Y. J Ryu, J. H. Yang, S. M. Lim, and C. G.Lee, “Lipid extraction from Tetraselmis sp. microalgae for biodieselproduction using hexane-based solvent mixtures,” Biotechnol.Bioprocess. Eng. vol. 23, pp. 16-22, 2018.
[27] W. Xiong, X. F. Li, J. Y. Xiang, and Q. Y. Wu, “High-densityfermentation of microalga Chlorella protothecoides in bioreactor formicrobio-diesel production,” Appl. Microbiol. Biotechnol. vol.78, pp.29-36, 2008.
[28] A. Y. Liu, W. Chen, L. L. Zheng, and L. R. Song, “Identification ofhigh-lipid producers for biodiesel production from forty-three greenalgal isolates in China,” Prog. Nat. Sci-Mater. vol. 21, pp. 269-276,2011.
[29] C. Y. Chen, and H. Y. Chamg, “Engineering strategies for enhancingC. vulgaris ESP-31 lipid production using effluents of coke-makingwastewater. J. Biosci. Bioeng. vol. 125, pp. 710-716, 2018.
[30] W. Zhang, Y. Zhao, B. Cui, H. Wang, and T. Liu, “Evaluation offilamentous green algae as feedstocks for biofuel production,”Bioresour. Technol. vol.16, pp. 407-413, 2016.
[31] O. Oliveira, S. Gianesella, V. Silva, T. Mata, and N. Caetano, “Lipidand carbohydrate profile of a microalga isolated from wastewater,”Energy. Procedia. vol. 136, pp. 468–437, 2017.
[32] H. J. La, G. G. Choi, C. Cho, S. H. Seo, A. Srivastava, B. H. Jo, J. Y.Lee, Y. S. Jin, and H. M. Oh, “Increased lipid productivity ofAcutodesmus dimorphus using optimized pulsed electric field. J.Appl. Phycol. vol. 28, pp. 931–938, 2016.
[33] H. Y. Ren, B. F. Liu, F. Kong, L. Zhao, G. J. Xie, and N. Q. Ren,.“Enhanced lipid accumulation of green microalga Scenedesmus sp. bymetal ions and EDTA addition,” Bioresour. Technol. vol 169, pp.763-767, 2014.
39

Amycolatopsis suaedae sp. nov., an endophytic actinomyceteisolated from Suaeda maritima roots
Tanatorn Chantavorakit,1 Paweena Suksaard,2 Atsuko Matsumoto3 and Kannika Duangmal1,*
Abstract
An actinomycete strain, designated 8-3EHSuT, was isolated from surface-sterilised roots of Suaeda maritima, collected from
Petchburi province, Thailand. Taxonomic position of strain 8-3EHSuT was studied using a polyphasic approach. Phylogenetic
determination based on the 16S rRNA gene sequence similarity showed that strain 8-3EHSuT belongs to the genus
Amycolatopsis, with the highest sequence similarity to Amycolatopsis jiangsuensis KLBMP 1262T (96.9%). The colony of strain 8-
3EHSuT was yellowish white. Long straight mycelium breaking down into fragments were observed. Growth occurred at
temperatures 15–45�C and at pH 6.0–10.0. The strain contained meso-diaminopimelic acid, arabinose, galactose and mannose
in whole-organism hydrolysates. The predominant menaquinone was MK-9(H4). The major fatty acids were C16 : 0, iso-C16 : 0 and
iso-C15 : 0. Polar lipids consisted of diphosphatidylglycerol, phosphatidylethanolamine, hydroxyphosphatidylethanolamine, two
unknown amino lipids and an unknown lipid. The G+C content of genomic DNA was 71.8mol%. A phylogenetic tree based on 16S
rRNA sequences showed that the strain 8-3EHSuT formed a distinct evolutionary linage within the genus Amycolatopsis. Based
on analysis results of physiological, biochemical and chemotaxonomic data, strain 8-3EHSuT represents a novel species of the
genus Amycolatopsis, for which the name Amycolatopsis suaedae sp. nov. is proposed. The type strain is 8-3EHSuT (=TBRC 8488T
=NBRC 113449T).
The genus Amycolatopsis is a member of the family Pseudo-
nocardiaceae. The genus was first proposed by Lechevalier
et al. [1] and the genus description was emended by Lee in
2009 [2]. Generally, members of the genus Amycolatopsis
are aerobic to facultatively anaerobic, Gram-stain-positive,
non-acid-fast, non-motile actinomycetes that form branch-
ing substrate hyphae which fragment into squarish and rod-
shaped subunits [3]. The typical chemotaxonomic charac-
teristics of the genus Amycolatopsis are meso-diaminopi-
melic acid, and arabinose and galactose contained in whole
cell hydrolysates with MK-9(H4) as the major menaqui-
none, and lack of mycolic acids [1, 4]. Members of the genus
have G+C content of the genomic DNA ranges from 66 to
73mol%. At the time of writing, the genus Amycolatopsis
contained 76 recognised species and four subspecies (LPSN:
http://www.bacterio.net/amycolatopsis.html). Members of
the genus Amycolatopsis are well known as an important
source of bioactive compounds [5] and have been found to
be widely distributed in natural habitats, especially in soils
[6–10]. In this study, a novel Amycolatopsis strain, isolated
from surface-sterilised Suaeda maritima roots, is described.
During the study of endophytic actinomycetes from man-grove plants, strain 8-3EHSuT was isolated from surface-sterilised roots of sea-blite (Suaeda maritima) collectedfrom Petchburi province, Thailand. The plant roots werewashed thoroughly with tap water and surface-sterilisedby shaking in 75% (v/v) ethanol for 5min followed by asolution of 1% (w/v) sodium hypochlorite (NaOCl) and10% (w/v) sodium thiosulfate (Na2S2O3) for 5 and10min, respectively. The roots were then rinsed with ster-ile distilled water twice and soaked in 10% (w/v) sodiumbicarbonate (NaHCO3) solution for 10min. Surface-steri-lised roots were crushed and suspended in sterile normalsaline solution, then the suspension was spread ontohumic acid-vitamin (HV) agar [11] supplemented withnalidixic acid (25 µgml�1) and nystatin (50 µgml�1). Afterincubation of the plates at 28
�C for 14 days, the strain
was transferred and purified on glucose-yeast extract agar(GYE agar; containing (w/v) 1.0% glucose, 1.0 % yeast
Author affiliations:1Department of Microbiology, Faculty of Science, Kasetsart University, Chatuchak, Bangkok 10900, Thailand; 2Department of
Science, Faculty of Science and Technology, Rajamangala University of Technology Suvarnabhumi, Phranakhon Si Ayutthaya 13000, Thailand; 3
Kitasato Institute for Life Sciences, Kitasato University, 5-9-1 Shirokane, Minato-ku, Tokyo 108-8641, Japan.*Correspondence: Kannika Duangmal, [email protected]: actinomycete; Amycolatopsis; Suaeda maritima; polyphasic taxonomy; endophyte.Abbreviations: GLC, gas-liquid chromatography; MIDI, Microbial Identification System; ML, maximum-likelihood ; MP, maximum-parsimony.The Genbank accession number for the 16S rRNA gene sequence of strain 8-3EHSuT is MG757753. The Genbank accession number for the wholegenome sequence of strain 8- 3EHSuT is SFCC00000000.Three supplementary figures and two supplementary tables are available with the online version of this article.
TAXONOMIC DESCRIPTION
Chantavorakit et al., Int J Syst Evol Microbiol 2019;69:2591–2596
DOI 10.1099/ijsem.0.003546
003546 ã 2019 IUMS
2591
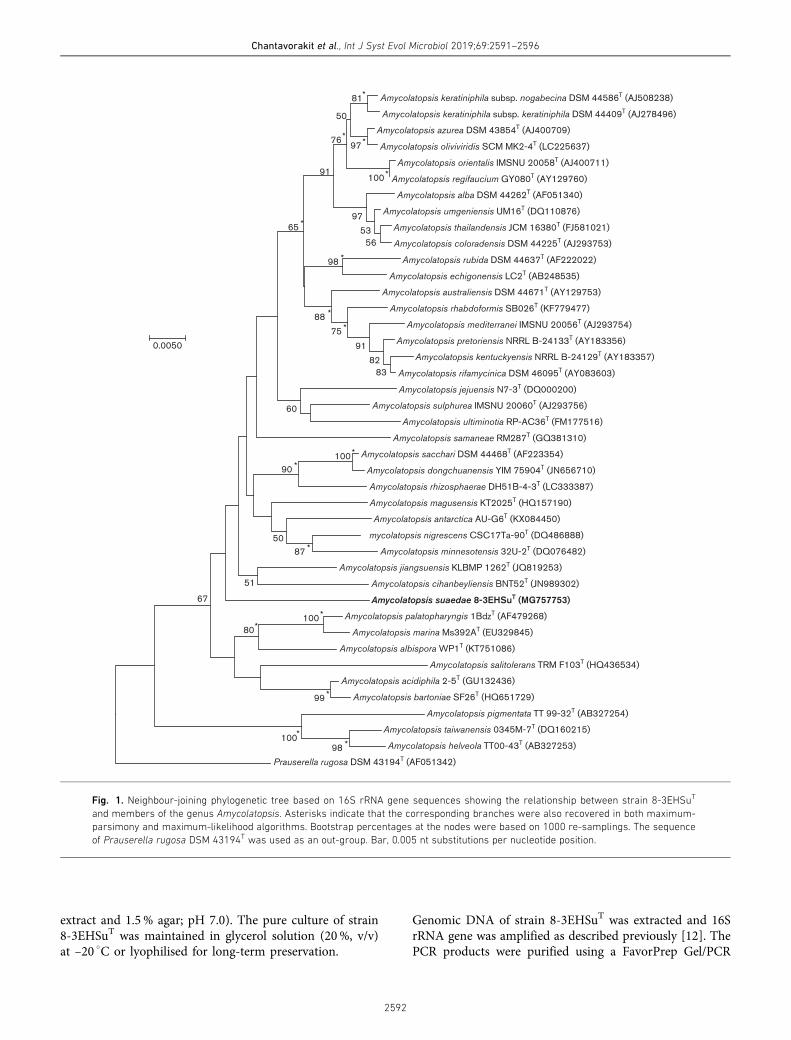
extract and 1.5% agar; pH 7.0). The pure culture of strain8-3EHSuT was maintained in glycerol solution (20%, v/v)at –20
�C or lyophilised for long-term preservation.
Genomic DNA of strain 8-3EHSuT was extracted and 16SrRNA gene was amplified as described previously [12]. ThePCR products were purified using a FavorPrep Gel/PCR
Amycolatopsis keratiniphila subsp. nogabecina DSM 44586T (AJ508238)
Amycolatopsis keratiniphila subsp. keratiniphila DSM 44409T (AJ278496)
Amycolatopsis azurea DSM 43854T (AJ400709)
Amycolatopsis oliviviridis SCM MK2-4T (LC225637)
Amycolatopsis orientalis IMSNU 20058T (AJ400711)
97*
81*
50
76*
91Amycolatopsis regifaucium GY080T (AY129760)
Amycolatopsis alba DSM 44262T (AF051340)
Amycolatopsis umgeniensis UM16T (DQ110876)
Amycolatopsis thailandensis JCM 16380T (FJ581021)
Amycolatopsis coloradensis DSM 44225T (AJ293753)
Amycolatopsis rubida DSM 44637T (AF222022)
Amycolatopsis echigonensis LC2T (AB248535)
Amycolatopsis australiensis DSM 44671T (AY129753)
100*
56
53
97
98 *
65 *
Amycolatopsis rhabdoformis SB026T (KF779477)
Amycolatopsis mediterranei IMSNU 20056T (AJ293754)
Amycolatopsis pretoriensis NRRL B-24133T (AY183356)
Amycolatopsis kentuckyensis NRRL B-24129T (AY183357)
Amycolatopsis rifamycinica DSM 46095T (AY083603)
Amycolatopsis jejuensis N7-3T (DQ000200)
Amycolatopsis sulphurea IMSNU 20060T (AJ293756)
83
82
91
75 *88 *
60
Amycolatopsis ultiminotia RP-AC36T (FM177516)
Amycolatopsis samaneae RM287T (GQ381310)
Amycolatopsis sacchari DSM 44468T (AF223354)
Amycolatopsis dongchuanensis YIM 75904T (JN656710)
Amycolatopsis rhizosphaerae DH51B-4-3T (LC333387)
Amycolatopsis magusensis KT2025T (HQ157190)
Amycolatopsis antarctica AU-G6T (KX084450)
100*
90 *
mycolatopsis nigrescens CSC17Ta-90T (DQ486888)
Amycolatopsis minnesotensis 32U-2T (DQ076482)
Amycolatopsis jiangsuensis KLBMP 1262T (JQ819253)
Amycolatopsis cihanbeyliensis BNT52T (JN989302)
Amycolatopsis suaedae 8-3EHSuT (MG757753)
Amycolatopsis palatopharyngis 1BdzT (AF479268)
Amycolatopsis marina Ms392AT (EU329845)
100*
80*
67
87 *50
51
Amycolatopsis albispora WP1T (KT751086)
Amycolatopsis salitolerans TRM F103T (HQ436534)
Amycolatopsis acidiphila 2-5T (GU132436)
Amycolatopsis bartoniae SF26T (HQ651729)
Amycolatopsis pigmentata TT 99-32T (AB327254)
Amycolatopsis taiwanensis 0345M-7T (DQ160215)
Amycolatopsis helveola TT00-43T (AB327253)
Prauserella rugosa DSM 43194T (AF051342)
98 *100*
99 *
0.0050
Fig. 1. Neighbour-joining phylogenetic tree based on 16S rRNA gene sequences showing the relationship between strain 8-3EHSuT
and members of the genus Amycolatopsis. Asterisks indicate that the corresponding branches were also recovered in both maximum-
parsimony and maximum-likelihood algorithms. Bootstrap percentages at the nodes were based on 1000 re-samplings. The sequence
of Prauserella rugosa DSM 43194T was used as an out-group. Bar, 0.005 nt substitutions per nucleotide position.
Chantavorakit et al., Int J Syst Evol Microbiol 2019;69:2591–2596
2592

Purification Mini Kit (Favorgen Biotech Corp.) followingthe manufacturer’s protocol and sequencing of 16S rRNAgene was performed using primers 27F (5¢- AGAGTTTGATCMTGGCTCAG-3¢), 1492R (5¢-TACGGYTACCTTGTTACGACTT-3¢), Mg4F (5¢-AATTCCTGGTGTAGCGGT-3¢) and 782R (5¢-ACCAGGGTATCTAATCCTGT-3¢).The resultant 16S rRNA gene sequence of the novel strain(1491 nt) was compared with available 16S rRNA genesequences in EzBioCloud database (https://www.ezbiocloud.net) [13] and aligned with related species of the genus Amy-colatopsis. Evolutionary trees were inferred using the maxi-mum-parsimony [14], maximum-likelihood [15] andneighbour-joining [16] algorithms from the MEGA version7.0 software [17], evaluated by bootstrap analysis [18] of1000 replications. The phylogenetic closest type strains ofthe genus Amycolatopsis [Amycolatopsis jiangsuensis NBRC108679T (96.9%), Amycolatopsis sacchari JCM 11272T
(96.3%) and Amycolatopsis sulphurea NBRC 13270T
(96.3%)] were used for morphological, physiological, che-motaxonomic and molecular taxonomic studies.
Cultural characteristics of strain 8-3EHSuT were determinedafter incubation at 28
�C for 7–21 days on International
Streptomyces Project (ISP) media 2, 3, 4, 5, 6 and 7 [19] andGYE agar. The NBS/ISCC Colour System charts were usedto determine the designations of substrate mycelia, aerialmycelia and soluble pigment colours [20]. Cell morphologyof the strain grown on GYE agar for 14 days at 28
�C was
examined using scanning electron microscopy (Quanta 450FEI). Growth was tested at different temperatures using atemperature gradient incubator model TN–3, Toyo KagakuSangyo (Tokyo, Japan) between 15 and 50
�C on ISP 2 agar.
NaCl tolerance was studied on ISP 2 agar containing NaClat final concentration of 0–12% (w/v), at intervals of 1%.The pH range (pH 4.0–12.0, at intervals of 1 pH units) for
growth was studied using ISP 2 agar adjusted with the buffersolutions: 0.1 M citric acid/0.1 M sodium citrate (pH 4.0–5.0); 0.1 M KH2PO4/0.1 M NaOH (pH 6.0–8.0); 0.1 MNaHCO3/0.1 M Na2CO3 (pH 9.0–12.0) after sterilisationand incubated at 28
�C for 14 days. Utilisation of sugar and
sugar alcohol as sole carbon sources at a final concentrationof 1% (w/v) were tested on ISP 9 agar [19]. Enzyme activityprofiles were tested by using API ZYM system (bioM�erieux)according to the manufacturer’s instructions. Urease activityand hydrolysis of adenine, aesculin, arbutin, casein(skimmed milk), carboxymethyl cellulose (CMC), gelatin,guanine, hypoxanthine, starch, Tween 20, Tween 80, L-tyro-sine, xanthine and xylan were evaluated using the media asdescribed by Gordon et al. [21] and Williams et al. [22].Catalase and oxidase activities were performed as describedby Gerhardt et al. [23]. Nitrate reduction was also studiedfollowing standard methods [24].
Freeze-dried biomass for chemotaxonomic analyses wasobtained after growing the strain 8-3EHSuT in ISP 2 brothby shaking at 150 r.p.m., 28
�C for 4 days. Cells were washed
three times in distilled water and freeze-dried. Standardprocedure was used to determine the isomer of diaminopi-melic acid [25, 26]. Whole-cell sugars were analysed bythin-layer chromatography (TLC) as described by Staneckand Roberts [27]. Menaquinones were extracted and puri-fied using the method of Collins et al. [28], and isopreneunits were subsequently analysed by LC/MS (JMS-T100LP,JEOL) with CAPCELL PAK C18 UG120 using methanol/2-propanol (7 : 3). The cellular fatty acid analysis was deter-mined using gas-liquid chromatography (GLC) accordingto the instructions of the Microbial Identification System(MIDI), Sherlock Version 6.2B [29] using the RTSBA6 data-base, which was performed at Thailand Institute of Scien-tific and Technological Research (TISTR). Polar lipids wereanalysed using the method described by Minnikin et al.[30]. Mycolic acids were investigated by one-dimensionalTLC according to the method of Tomiyasu [31]. The DNAG+C content was determined by HPLC according to themethod of Tamaoka and Komagata [32].
The nearly complete 16S rRNA gene sequence of strain 8-3EHSuT (1491 nt) was determined and compared with thecorresponding sequences in EzBioCloud database. Thestrain 8-3EHSuT shared the highest 16S rRNA genesequence similarity with Amycolatopsis jiangsuensis KLBMP1262T (96.9%), followed by Amycolatopsis sacchari DSM44468T (96.3%) and Amycolatopsis sulphurea DSM 46092T
(96.3%). Phylogenetic analysis of 16S rRNA gene sequenceof strain 8-3EHSuT generated with the neighbour-joiningalgorithm indicated that the strain formed a distinct phy-letic line within the genus Amycolatopsis (Fig. 1). This rela-tionship was also supported by the results obtained withmaximum-likelihood and maximum-parsimony algorithms(Figs S1 and S2, available in the online version of this arti-cle). The relatively low levels of 16S rRNA gene sequencesimilarity between strain 8-3EHSuT and its nearest neigh-bours clearly differentiated strain 8-3EHSuT from its closest
Fig. 2. Scanning electron micrographs of strain 8-3EHSuT grown on
GYE agar at 28�C for 14 days. Bar, 1 µm.
Chantavorakit et al., Int J Syst Evol Microbiol 2019;69:2591–2596
2593
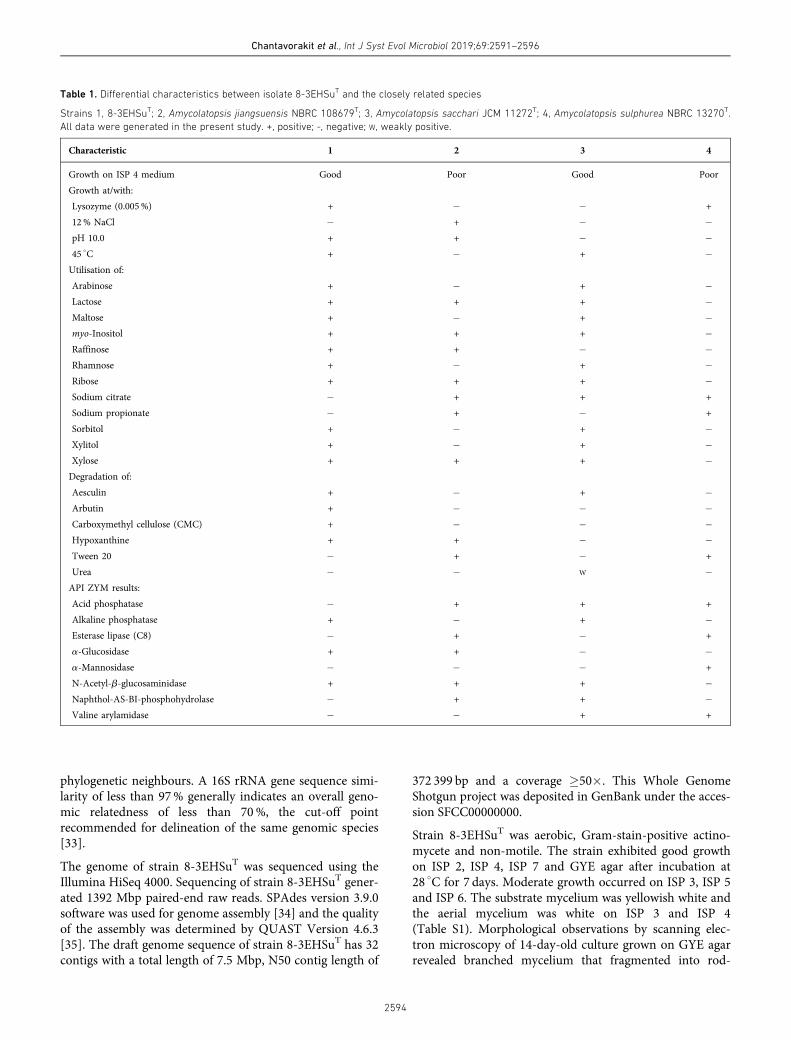
phylogenetic neighbours. A 16S rRNA gene sequence simi-larity of less than 97% generally indicates an overall geno-mic relatedness of less than 70%, the cut-off pointrecommended for delineation of the same genomic species[33].
The genome of strain 8-3EHSuT was sequenced using theIllumina HiSeq 4000. Sequencing of strain 8-3EHSuT gener-ated 1392 Mbp paired-end raw reads. SPAdes version 3.9.0software was used for genome assembly [34] and the qualityof the assembly was determined by QUAST Version 4.6.3[35]. The draft genome sequence of strain 8-3EHSuT has 32contigs with a total length of 7.5 Mbp, N50 contig length of
372 399 bp and a coverage �50�. This Whole GenomeShotgun project was deposited in GenBank under the acces-sion SFCC00000000.
Strain 8-3EHSuT was aerobic, Gram-stain-positive actino-mycete and non-motile. The strain exhibited good growthon ISP 2, ISP 4, ISP 7 and GYE agar after incubation at28
�C for 7 days. Moderate growth occurred on ISP 3, ISP 5
and ISP 6. The substrate mycelium was yellowish white andthe aerial mycelium was white on ISP 3 and ISP 4(Table S1). Morphological observations by scanning elec-tron microscopy of 14-day-old culture grown on GYE agarrevealed branched mycelium that fragmented into rod-
Table 1. Differential characteristics between isolate 8-3EHSuT and the closely related species
Strains 1, 8-3EHSuT; 2, Amycolatopsis jiangsuensis NBRC 108679T; 3, Amycolatopsis sacchari JCM 11272T; 4, Amycolatopsis sulphurea NBRC 13270T.
All data were generated in the present study. +, positive; -, negative; W, weakly positive.
Characteristic 1 2 3 4
Growth on ISP 4 medium Good Poor Good Poor
Growth at/with:
Lysozyme (0.005%) + � � +
12% NaCl � + � �
pH 10.0 + + � �
45�C + � + �
Utilisation of:
Arabinose + � + �
Lactose + + + �
Maltose + � + �
myo-Inositol + + + �
Raffinose + + � �
Rhamnose + � + �
Ribose + + + �
Sodium citrate � + + +
Sodium propionate � + � +
Sorbitol + � + �
Xylitol + � + �
Xylose + + + �
Degradation of:
Aesculin + � + �
Arbutin + � � �
Carboxymethyl cellulose (CMC) + � � �
Hypoxanthine + + � �
Tween 20 � + � +
Urea � � W �
API ZYM results:
Acid phosphatase � + + +
Alkaline phosphatase + � + �
Esterase lipase (C8) � + � +
a-Glucosidase + + � �
a-Mannosidase � � � +
N-Acetyl-b-glucosaminidase + + + �
Naphthol-AS-BI-phosphohydrolase � + + �
Valine arylamidase � � + +
Chantavorakit et al., Int J Syst Evol Microbiol 2019;69:2591–2596
2594

shaped elements (Fig. 2). The temperature range for growthwas between 15–45
�C with optimum growth at 28–37
�C.
The strain was able to grow at pH 6.0–10.0, with optimalgrowth at pH 7.0–8.0. Growth was observed at 11% NaCl(w/v). Differentiation of biochemical and physiologicalproperties of strain 8-3EHSuT and closest members of thegenus Amycolatopsis are shown in Table 1. For example,degradation of arbutin and carboxymethyl cellulose werepositive in strain 8-3EHSuT, whereas the other type strainsshowed negative results. In carbon utilisation, strain 8-3EHSuT was unable to use sodium citrate as sole carbonsource, whereas the other type strains were positive. Strain8-3EHSuT was able to grow at 45
�C and resistant to lyso-
zyme but not for A. jiangsuensis. Moreover, strain 8-3EHSuT showed good growth on ISP 4 medium, whereaspoor growth was observed in A. jiangsuensis.
The strain 8-3EHSuT exhibited typical characteristics of thegenus Amycolatopsis. The whole-cell hydrolysates containedmeso-diaminopimelic acid and arabinose, galactose andmannose as whole-cell sugars. MK-9(H4) (94.2%) wasfound as major menaquinone with small amount of MK-10(H4) (5.8%). Mycolic acids were not found. The polar lipidswere diphosphatidylglycerol, phosphatidylethanolamine,hydroxyphosphatidylethanolamine, two unknown aminolipids and unknown lipid (Fig. S3). The major cellular fattyacids were C16 : 0 (17.0 %), iso-C16 : 0 (16.4 %) and iso-C15 : 0
(12.2%) (Table S2). The G+C content of the DNA was71.8mol%.
In conclusion, it is evident on the basis of phenotypic andgenotypic data that strain 8-3EHSuT is clearly distinguishedfrom previously described species and represented a novelspecies of the genus Amycolatopsis, for which the nameAmycolatopsis suaedae sp. nov. is proposed.
DESCRIPTION OF AMYCOLATOPSIS SUAEDAE
SP. NOV.
Amycolatopsis suaedae (su.ae’dae. N.L. gen. n. suaedae ofSuaeda, referring to the isolation of the type strain fromroots of Suaeda maritima).
Aerobic, Gram-stain-positive, non-motile, catalase-positiveactinobacterium. Forms extensively branched substrate andaerial mycelia that fragment into rod-shaped elements. Thesubstrate mycelium is yellowish white and the aerial myce-lium is white on ISP 3 and ISP 4. Diffusible pigments arenot produced. Good growth occurs on ISP 2, ISP 4, ISP 7and GYE agar; moderate growth ISP 3, ISP 5 and ISP 6.Temperature range for growth is at 15–45
�C with optimum
growth at 28–37�C. The NaCl range for growth is 0–11%
(w/v). Growth occurs at pH 6.0–10.0. Grows in the presenceof lysozyme (0.005%). Catalase is positive. Oxidase is nega-tive. H2S production and nitrate reduction are positive, buturease is negative. Degradation of aesculin, arbutin, carbox-ymethyl cellulose (CMC), casein, gelatin, hypoxanthine, L-tyrosine, Tween 80 and xanthine are positive but not ade-nine, guanine, starch, Tween 20 and xylan. Positive for
utilisation of L(+)arabinose, cellobiose, fructose, galactose,glucose, lactose, maltose, mannitol, mannose, myo-inositol,raffinose, rhamnose, ribose, sorbitol, sucrose, trehalose, xyli-tol and D(+)xylose. Sodium citrate and sodium propionateare not utilised. Enzyme assay with the API ZYM are posi-tive for alkaline phosphatase, a–glucosidase, leucine aryla-midase and N–acetyl–b–glucosaminidase whereas negativefor acid phosphatase, a-chymotrypsin, cystine arylamidase,esterase (C4), esterase lipase (C8), a-fucosidase, a-galactosi-dase, b-galactosidase, b-glucosidase, b-glucuronidase, lipase(C14), a–mannosidase, naphthol–AS–BI–phosphohydro-lase, trypsin and valine arylamidase. The cell wall containsmeso-diaminopimelic acid with arabinose, galactose andmannose as the whole-cell sugars. The predominant mena-quinone is MK-9(H4). Mycolic acids are absent. Polar lipidsare diphosphatidylglycerol, phosphatidylethanolamine,hydroxyphosphatidylethanolamine, two unknown aminolipids and an unknown lipid. The major fatty acids areC16 : 0, iso-C16 : 0 and iso-C15 : 0. The G+C content of the typestrain is 71.8mol%.
The type strain, 8-3EHSuT (=TBRC 8488T=NBRC 113449T), was isolated from Suaeda maritima roots, collectedfrom Petchburi province, Thailand. The Genbank accessionnumber for the 16S rRNA gene sequence of strain 8-3EHSuT is MG757753.
Funding information
This research has been supported by Center of Excellence on Biodiver-sity (BDC), Office of Higher Education Commission (Project Code BDC-PG1-160003) and supported in part by the Graduate Program Scholar-ship from The Graduate School, Kasetsart University to TanatornChantavorakit.
Acknowledgements
The authors are grateful to Professor Dr Savitree Limtong of KasetsartUniversity, the director of the research program and Jutapohn Tong-pan for isolation of the strain. We are grateful to the King’s Royally Ini-tiated Laem Phak Bia Environmental Research and Developmentproject for samples collection.
Conflicts of interest
The authors declare that there are no conflicts of interest.
References
1. Lechevalier MP, Prauser H, Labeda DP, Ruan J-S. Two new gen-era of nocardioform actinomycetes: Amycolata gen. nov. and Amy-
colatopsis gen. nov. Int J Syst Bacteriol 1986;36:29–37.
2. Lee SD. Amycolatopsis ultiminotia sp. nov., isolated from rhizo-sphere soil, and emended description of the genus Amycolatopsis.Int J Syst Evol Microbiol 2009;59:1401–1404.
3. Tan GYA, Goodfellow M. Genus Amycolatopsis. In: Goodfellow M,K€ampfer P and Busse HJ (editors). Bergey’s Manual of Systematic
Bacteriology, 2nd ed. New York: Springer; 2012. pp. 1334–1358.
4. Lechevalier MP, De Bievre C, Lechevalier H. Chemotaxonomy ofaerobic actinomycetes: phospholipid composition. Biochem Syst
Ecol 1977;5:249–260.
5. Oyuntsetseg B, Cho SH, Jeon SJ, Lee HB, Shin KS et al. Amycola-
topsis acidiphila sp. nov., a moderately acidophilic species isolatedfrom coal mine soil. Int J Syst Evol Microbiol 2017;67:3387–3392.
6. Lee SD, Kinkel LL, Samac DA. Amycolatopsis minnesotensis sp.nov., isolated from a prairie soil. Int J Syst Evol Microbiol 2006;56:265–269.
Chantavorakit et al., Int J Syst Evol Microbiol 2019;69:2591–2596
2595

7. Tamura T, Ishida Y, Otoguro M, Suzuki K. Amycolatopsis helveola
sp. nov. and Amycolatopsis pigmentata sp. nov., isolated from soil.Int J Syst Evol Microbiol 2010;60:2629–2633.
8. Nie GX, Ming H, Li S, Zhou EM, Cheng J et al. Amycolatopsis dong-
chuanensis sp. nov., an actinobacterium isolated from soil. Int JSyst Evol Microbiol 2012;62:2650–2656.
9. Camas M, Sahin N, Sazak A, Spröer C, Klenk HP et al. Amycola-
topsis magusensis sp. nov., isolated from soil. Int J Syst Evol
Microbiol 2013;63:1254–1260.
10. Souza WR, Silva RE, Goodfellow M, Busarakam K, Figueiro FS
et al. Amycolatopsis rhabdoformis sp. nov., an actinomycete iso-lated from a tropical forest soil. Int J Syst Evol Microbiol 2015;65:1786–1793.
11. Hayakawa M, Nonomura H. Humic acid-vitamin agar, a newmedium for the selective isolation of soil actinomycetes. J Ferment
Technol 1987;65:501–509.
12. Mingma R, Pathom-aree W, Trakulnaleamsai S, Thamchaipenet
A, Duangmal K. Isolation of rhizospheric and roots endophyticactinomycetes from Leguminosae plant and their activities toinhibit soybean pathogen, Xanthomonas campestris pv. glycine.World J Microbiol Biotechnol 2014;30:271–280.
13. Yoon SH, Ha SM, Kwon S, Lim J, Kim Y et al. Introducing EzBio-Cloud: a taxonomically united database of 16S rRNA gene sequen-ces and whole-genome assemblies. Int J Syst Evol Microbiol 2017;67:1613–1617.
14. Fitch WM. Toward defining the course of evolution: minimumchange for a specific tree topology. Syst Zool 1971;20:406–416.
15. Felsenstein J. Evolutionary trees from DNA sequences: a maxi-mum likelihood approach. J Mol Evol 1981;17:368–376.
16. Saitou N, Nei M. The neighbor-joining method: a new method forreconstructing phylogenetic trees. Mol Biol Evol 1987;4:406–425.
17. Kumar S, Stecher G, Tamura K. MEGA7: Molecular evolutionarygenetics analysis version 7.0 for bigger datasets. Mol Biol Evol2016;33:1870–1874.
18. Felsenstein J. Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 1985;39:783–791.
19. Shirling EB, Gottlieb D. Methods for characterization of Streptomy-
ces species. Int J Syst Bacteriol 1966;16:313–340.
20. Mundie D. The NBS/ISCC Color System/David A. Mundie Pittsburgh,PA: Polymath Systems 535.6 dc-20; 1995.
21. Gordon RE, Barnett DA, Handerhan JE, Pang CH-N. Nocardia coe-
liaca, Nocardia autotrophica, and the nocardin strain. Int J Syst
Bacteriol 1974;24:54–63.
22. Williams ST, Goodfellow M, Alderson G, Wellington EM, Sneath
PH et al. Numerical classification of Streptomyces and related
genera. J Gen Microbiol 1983;129:1743–1813.
23. Gerhardt P, Murray RGE, Krieg NR, Wood WA. Methods for General
and Molecular Bacteriology American Society for Microbiology;1994.
24. Gordon RE, Mihm JM. A comparative study of some strainsreceived as nocardiae. J Bacteriol 1957;73:15–27.
25. Becker B, Lechevalier MP, Lechevalier HA. Chemical composition
of cell-wall preparations from strains of various form-genera of
aerobic actinomycetes. Appl Microbiol 1965;13:236–243.
26. Hasegawa T, Takizawa M, Tanida S. A rapid analysis for chemicalgrouping of aerobic actinomycetes. J Gen Appl Microbiol 1983;29:319–322.
27. Staneck JL, Roberts GD. Simplified approach to identification ofaerobic actinomycetes by thin-layer chromatography. Appl
Microbiol 1974;28:226–231.
28. Collins MD, Pirouz T, Goodfellow M, Minnikin DE. Distribution of
menaquinones in actinomycetes and corynebacteria. J Gen
Microbiol 1977;100:221–230.
29. Sasser M. Identification of Bacteria by Gas Chromatography of
Cellular Fatty Acids, MIDI Technical Note 101. Newark, DE: MIDIInc; 1990.
30. Minnikin DE, Patel PV, Alshamaony L, Goodfellow M. Polar lipid
composition in the classification of Nocardia and related bacteria.
Int J Syst Bacteriol 1977;27:104–117.
31. Tomiyasu I. Mycolic acid composition and thermally adaptative
changes in Nocardia asteroides. J Bacteriol 1982;151:828–837.
32. Tamaoka J, Komagata K. Determination of DNA base composition
by reversed-phase high-performance liquid chromatography.
FEMS Microbiol Lett 1984;25:125–128.
33. Wayne LG, Moore WEC, Stackebrandt E, Kandler O, Colwell RR
et al. Report of the ad hoc committee on reconciliation of
approaches to bacterial systematics. Int J Syst Evol Microbiol
1987;37:463–464.
34. Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M et al.
SPAdes: a new genome assembly algorithm and its applications
to single-cell sequencing. J Comput Biol 2012;19:455–477.
35. Gurevich A, Saveliev V, Vyahhi N, Tesler G. QUAST: qualityassessment tool for genome assemblies. Bioinformatics 2013;29:1072–1075.
Chantavorakit et al., Int J Syst Evol Microbiol 2019;69:2591–2596
2596
Five reasons to publish your next article with a Microbiology Society journal
1. The Microbiology Society is a not-for-profit organization.
2. We offer fast and rigorous peer review – average time to first decision is 4–6 weeks.
3. Our journals have a global readership with subscriptions held in research institutions aroundthe world.
4. 80% of our authors rate our submission process as ‘excellent’ or ‘very good’.
5. Your article will be published on an interactive journal platform with advanced metrics.
Find out more and submit your article at microbiologyresearch.org.




















