
Human ILT2 receptor associates with murine MHC class I moleculesin vivo and impairs T cell function
15
Human ILT2 receptor associates with murine MHC class I molecules in vivo and impairs T cell function Siyuan Liang*, Wei Zhang* and Anatolij Horuzsko Center for Molecular Chaperone/Radiobiology and Cancer Virology, Department of Medicine, Medical College of Georgia, Augusta, USA Immunoglobulin-like transcript 2 (ILT2/LILRB1/LIR1/CD85j) is an inhibitory receptor broadly expressed on leukocytes and antigen-presenting cells that recognizes HLA-class I and human cytomegalovirus UL18 proteins. The function of this receptor is to generate negative signals or to inhibit positive signals. Here, we demonstrate the model to study the function of ILT2 in vivo using a newly generated transgenic mouse expressing the human inhibitory receptor on T, B, NK, and NKT cells. ILT2 expression affects thymocyte development and targets the proximal TCR signaling pathway, resulting in long-term survival or acceptance of skin allografts. The phenotype and constitutive tyrosine phosphorylation of ILT2 in transgenic mice illustrate the possible existence of a murine ligand. We report here that H-2D b , a murine MHC class I molecule, associates with human ILT2 in vivo. This engagement with ILT2 directs effects on thymocyte development, negative regulation of TCR signaling, T cell activation, and alloimmune responses. Our finding provides support for an important inhibitory function of ILT2 in T cells in vivo and opens up strategies for targeting proximal TCR signaling for prevention of allograft rejection. Supporting information for this article is available at http://www.wiley-vch.de/contents/jc_2040/2006/36031_s.pdf Introduction Immunoglobulin-like transcript 2 (ILT2) is an inhibitory receptor broadly expressed on leukocytes and antigen- presenting cells and recognizes HLA-class I and human cytomegalovirus UL18 proteins. ILT2 is a transmem- brane protein with an Ig-like extracellular domain [1, 2]. This receptor is structurally related to several other inhibitory Ig superfamily members, including human killer cell immunoglobulin-like receptors (KIR) and Fca receptors [3, 4]. The cytoplasmic region of ILT2 contains four ITIM-like sequences. On tyrosine phosphorylation, ITIM recruits the tyrosine phosphatases, SHP-1 and C-terminal Src kinase (Csk), that can dephosphorylate molecules involved in ITAM-induced signaling pathways [4, 5]. ILT2 can bind to a variety of HLA-class I molecules and human cytomegalovirus UL18 protein through two membrane-distal extracellular domains, D1 and D2 [6–9]. The binding site for ILT2 within the a3 region of human MHC class I is highly conserved between classical and nonclassical MHC class I molecule and similar to the murine MHC class I molecule. This similarity provides the potential for the recognition of human ILT2 by murine MHC class I molecules. Trans-species recogni- tion is well known for TCR [10, 11]. For instance, the mouse TCR repertoire is capable of recognizing transgenic human HLA molecules [10, 11]. Its extra- Immunomodulation * The first two authors contributed equally to this work. Correspondence: Dr. Anatolij Horuzsko, Center for Molecular Chaperone/Radiobiology and Cancer Virology, Medical College of Georgia, 1410 Laney Walker Boulevard, Augusta, GA 30912, USA Fax: +1-706-721-0101 e-mail: [email protected] Received 2/3/06 Revised 31/5/06 Accepted 28/6/06 [DOI 10.1002/eji.200636031] Key words: Allograft survival Inhibitory receptor Ligand Negative regulation T cell activation Abbreviations: DN: double-negative DP: double-positive ILT: immunoglobulin-like transcript MST: mean of survival time PD-1: programmed death-1 SP: single-positive Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2457 f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Human ILT2 receptor associates with murine MHC class I moleculesin vivo and impairs T cell function

Human ILT2 receptor associates with murine MHCclass I molecules in vivo and impairs T cell function
Siyuan Liang*, Wei Zhang* and Anatolij Horuzsko
Center for Molecular Chaperone/Radiobiology and Cancer Virology, Department ofMedicine, Medical College of Georgia, Augusta, USA
Immunoglobulin-like transcript 2 (ILT2/LILRB1/LIR1/CD85j) is an inhibitory receptorbroadly expressed on leukocytes and antigen-presenting cells that recognizes HLA-classI and human cytomegalovirus UL18 proteins. The function of this receptor is to generatenegative signals or to inhibit positive signals. Here, we demonstrate the model to studythe function of ILT2 in vivo using a newly generated transgenic mouse expressing thehuman inhibitory receptor onT, B, NK, and NKTcells. ILT2 expression affects thymocytedevelopment and targets the proximal TCR signaling pathway, resulting in long-termsurvival or acceptance of skin allografts. The phenotype and constitutive tyrosinephosphorylation of ILT2 in transgenic mice illustrate the possible existence of a murineligand. We report here that H-2Db, a murine MHC class I molecule, associates withhuman ILT2 in vivo. This engagement with ILT2 directs effects on thymocytedevelopment, negative regulation of TCR signaling, T cell activation, and alloimmuneresponses. Our finding provides support for an important inhibitory function of ILT2 inT cells in vivo and opens up strategies for targeting proximal TCR signaling forprevention of allograft rejection.
Supporting information for this article is available athttp://www.wiley-vch.de/contents/jc_2040/2006/36031_s.pdf
Introduction
Immunoglobulin-like transcript 2 (ILT2) is an inhibitoryreceptor broadly expressed on leukocytes and antigen-presenting cells and recognizes HLA-class I and humancytomegalovirus UL18 proteins. ILT2 is a transmem-brane proteinwith an Ig-like extracellular domain [1, 2].This receptor is structurally related to several other
inhibitory Ig superfamily members, including humankiller cell immunoglobulin-like receptors (KIR) and Fcareceptors [3, 4]. The cytoplasmic region of ILT2 containsfour ITIM-like sequences. On tyrosine phosphorylation,ITIM recruits the tyrosine phosphatases, SHP-1 andC-terminal Src kinase (Csk), that can dephosphorylatemolecules involved in ITAM-induced signaling pathways[4, 5]. ILT2 can bind to a variety of HLA-class I moleculesand human cytomegalovirus UL18 protein through twomembrane-distal extracellular domains, D1 and D2[6–9]. The binding site for ILT2 within the a3 region ofhumanMHC class I is highly conserved between classicaland nonclassical MHC class I molecule and similar to themurine MHC class I molecule. This similarity providesthe potential for the recognition of human ILT2 bymurine MHC class I molecules. Trans-species recogni-tion is well known for TCR [10, 11]. For instance, themouse TCR repertoire is capable of recognizingtransgenic human HLA molecules [10, 11]. Its extra-
Immunomodulation
* The first two authors contributed equally to this work.
Correspondence: Dr. Anatolij Horuzsko, Center for MolecularChaperone/Radiobiology and Cancer Virology, Medical Collegeof Georgia, 1410 Laney Walker Boulevard, Augusta, GA 30912,USAFax: +1-706-721-0101e-mail: [email protected]
Received 2/3/06Revised 31/5/06
Accepted 28/6/06
[DOI 10.1002/eji.200636031]
Key words:Allograft survival
� Inhibitory receptor� Ligand � Negativeregulation � T cell
activation
Abbreviations: DN: double-negative � DP: double-positive �ILT: immunoglobulin-like transcript � MST: mean of survivaltime � PD-1: programmed death-1 � SP: single-positive
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2457
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

ordinary binding of HLA-G, a human nonclassical MHCmolecule, raises the possibility that ILT2 may speciallycontribute to functional interactions between cellsexpressing ILT2 and cells expressing HLA-G [12, 13].The expression of HLA-G occurs on thymic epithelialcells, fetal cytotrophoblast cells, cells infected withcytomegalovirus, cancer cells and transplanted tissues[14–18]. ILT2 may influence thymocyte development byinteracting with HLA-G and classical MHC molecules onthymic epithelial cells. Another possibility includes thatILT2 may be involved in modulation of T cell signalingduring allorecognition of transplanted tissues or organsas HLA-G ligand is expressed by accepted allografts [19].Indeed, HLA-G directly up-regulates ILT2 expression onT cells before antigenic stimulation [20]. Thus, thefunction of this receptor is to generate negative signalingor inhibition of positive signaling. Several experimentsin vitro show that cross-linking of ILT2 can effectivelyinhibit T cell activation and proliferation [21]. Since theinhibitory nature of ILT2 was previously observed forTcell clones and Tcell lines [21, 22], not much is knownabout the function of this immunoinhibitory molecule invivo. Of particular interest is the determination of thecapacity of ILT2 to affect T cell responses duringalloantigen recognition for modulation of graft rejec-tion. Here, we demonstrate the function of ILT2 in vivousing a newly generated transgenic mouse modelexpressing human inhibitory receptor on T, B, NK,and NKT cells. The expression of ILT2 significantlyaffects thymocyte development and down-modulatesT cell function. The target has been determined as theproximal TCR signaling pathway, resulting in long-termskin graft survival or acceptance of allografts. Thephenotype and constitutive tyrosine phosphorylation ofILT2 in transgenic mice illustrate the possible existenceof a murine cell-surface ligand. In an attempt todetermine the potential ligand(s), ILT2 transgenic micehave been crossed with MHC class I-deficient (b2m–/–)mice, which resulted in the rescue of compromised Tcellfunctions and decreased phosphorylation of ILT2. Here,we demonstrate that H-2Db, a murine b2 m-associatedMHC class I molecule is a cell-surface ligand for humanILT2, and the engagement of ILT2 directs the effects onthymocyte development and diminished Tcell functions.Our finding provides new insight into the nature of thenegative regulation of TCR signaling in vivo by ILT2 andopens up several strategies for targeting proximal TCRsignaling to improve the outcome of solid organ or tissuetransplantation.
Results
Expression of ILT2 in transgenic mice
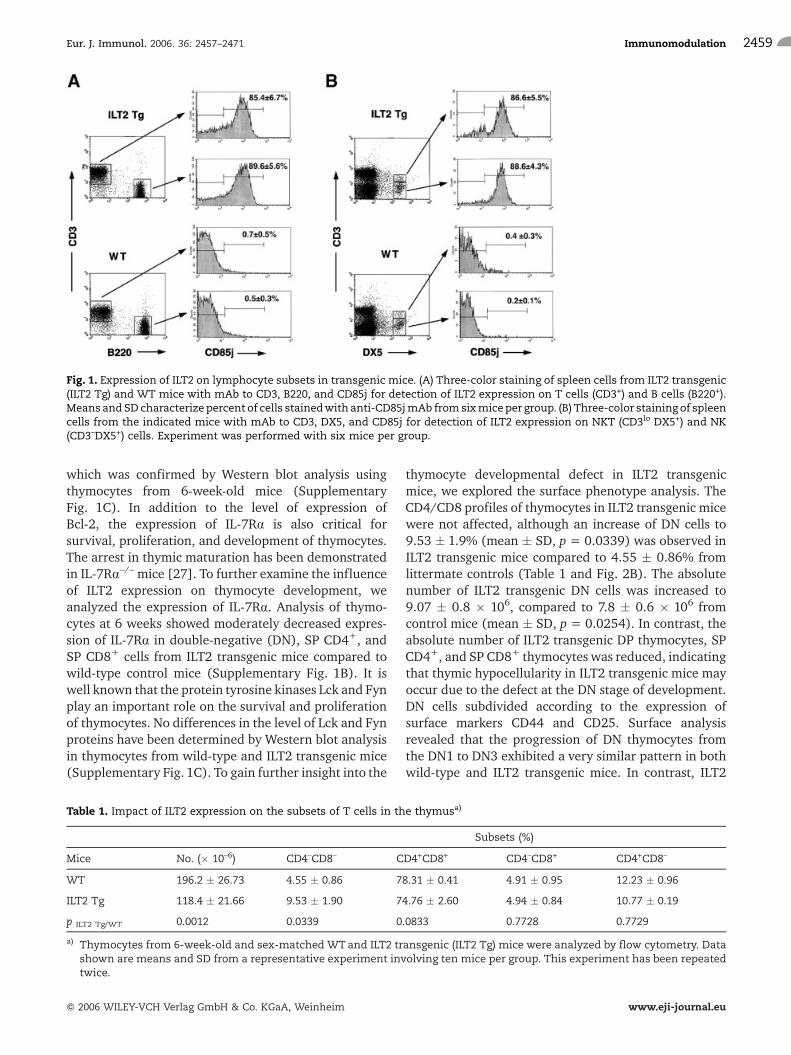
To generate an experimental model for the study offunctions of immune inhibitory receptor ILT2 in vivo, wedeveloped transgenic mice expressing ILT2 under thecontrol of a human CD2 promoter. To analyze the ILT2expression pattern in transgenic mice, we stained spleencells, lymph node cells, and thymocytes with mAbspecific to CD85j, which confirmed transgene expres-sion in vivo. As predicted from the activity of the humanCD2 promoter, ILT2 protein was expressed at high levelson almost all peripheral T and B cells (Fig. 1A). The cellsurface expression of ILT2 was also detected on themajority of NK and NKT cells and thymocytes (Fig. 1Band data not shown). Two independent lines of ILT2transgenic mice have been established. No differences inthe pattern and the level of expression of ILT2 proteinhave been determined in two lines of transgenic mice.The ILT2 transgene was expressed in a similar pattern tothat observed in humans [22, 23].
Abnormal T cell development in ILT2 transgenicmice
Tcell inhibitory receptors have the potential to affect thedevelopment of T cells. To investigate the functionalconsequences of ILT2 expression on T cell development,the thymuses were analyzed at the different time pointsin young mice. The absolute numbers of thymocytes in6-week-old ILT2 transgenic mice are lower than in theage-matched control mice (118.4 � 21.66 � 106 cells intransgenic versus 196.2 � 26.73 � 106 in wild- type;mean� SD, p= 0.0012, Table 1). Decreased numbers ofthymocytes could be due to reduced production or toincreased elimination. To determine the level ofapoptosis of thymocytes, cells were stained withAnnexin V-FITC and propidium iodide. The profile ofviable, early apoptotic, and late apoptotic cells showedno difference between thymocytes from ILT2 transgenicand control mice (Fig. 2A). To gain further insight intothe thymocyte developmental defect in ILT2 transgenicmice, we examined the pattern of expression of Bcl-2,which is known to be tightly controlled in the thymus.Bcl-2 expression normally appears highest in thymocyteprecursors, drops to low levels in double-positive (DP)cells, and is sharply up-regulated during or immediatelyafter positive selection [24–26]. FACS analysis of theintracellular expression of Bcl-2 showed that ILT2expression does not have any role in the modulationof Bcl-2 expression during positive selection; both DPand single-positive (SP) CD4+ and SP CD8+ thymocytesfrom ILT2 transgenic mice contained similar amounts ofintracellular Bcl-2 protein (Supplementary Fig. 1A),
Siyuan Liang et al. Eur. J. Immunol. 2006. 36: 2457–24712458
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
which was confirmed by Western blot analysis usingthymocytes from 6-week-old mice (SupplementaryFig. 1C). In addition to the level of expression ofBcl-2, the expression of IL-7Ra is also critical forsurvival, proliferation, and development of thymocytes.The arrest in thymic maturation has been demonstratedin IL-7Ra–/– mice [27]. To further examine the influenceof ILT2 expression on thymocyte development, weanalyzed the expression of IL-7Ra. Analysis of thymo-cytes at 6 weeks showed moderately decreased expres-sion of IL-7Ra in double-negative (DN), SP CD4+, andSP CD8+ cells from ILT2 transgenic mice compared towild-type control mice (Supplementary Fig. 1B). It iswell known that the protein tyrosine kinases Lck and Fynplay an important role on the survival and proliferationof thymocytes. No differences in the level of Lck and Fynproteins have been determined by Western blot analysisin thymocytes from wild-type and ILT2 transgenic mice(Supplementary Fig. 1C). To gain further insight into the
thymocyte developmental defect in ILT2 transgenicmice, we explored the surface phenotype analysis. TheCD4/CD8 profiles of thymocytes in ILT2 transgenic micewere not affected, although an increase of DN cells to9.53 � 1.9% (mean � SD, p = 0.0339) was observed inILT2 transgenic mice compared to 4.55 � 0.86% fromlittermate controls (Table 1 and Fig. 2B). The absolutenumber of ILT2 transgenic DN cells was increased to9.07 � 0.8 � 106, compared to 7.8 � 0.6 � 106 fromcontrol mice (mean � SD, p = 0.0254). In contrast, theabsolute number of ILT2 transgenic DP thymocytes, SPCD4+, and SP CD8+ thymocytes was reduced, indicatingthat thymic hypocellularity in ILT2 transgenic mice mayoccur due to the defect at the DN stage of development.DN cells subdivided according to the expression ofsurface markers CD44 and CD25. Surface analysisrevealed that the progression of DN thymocytes fromthe DN1 to DN3 exhibited a very similar pattern in bothwild-type and ILT2 transgenic mice. In contrast, ILT2
Table 1. Impact of ILT2 expression on the subsets of T cells in the thymusa)
Subsets (%)
Mice No. (� 10–6) CD4–CD8– CD4+CD8+ CD4–CD8+ CD4+CD8–
WT 196.2 � 26.73 4.55 � 0.86 78.31 � 0.41 4.91 � 0.95 12.23 � 0.96
ILT2 Tg 118.4 � 21.66 9.53 � 1.90 74.76 � 2.60 4.94 � 0.84 10.77 � 0.19
p ILT2 Tg/WT 0.0012 0.0339 0.0833 0.7728 0.7729
a) Thymocytes from 6-week-old and sex-matched WTand ILT2 transgenic (ILT2 Tg) mice were analyzed by flow cytometry. Datashown are means and SD from a representative experiment involving ten mice per group. This experiment has been repeatedtwice.
Fig. 1. Expression of ILT2 on lymphocyte subsets in transgenic mice. (A) Three-color staining of spleen cells from ILT2 transgenic(ILT2 Tg) and WT mice with mAb to CD3, B220, and CD85j for detection of ILT2 expression on T cells (CD3+) and B cells (B220+).Means andSD characterize percent of cells stainedwith anti-CD85jmAb fromsixmiceper group. (B) Three-color staining of spleencells from the indicated mice with mAb to CD3, DX5, and CD85j for detection of ILT2 expression on NKT (CD3lo DX5+) and NK(CD3–DX5+) cells. Experiment was performed with six mice per group.
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2459
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
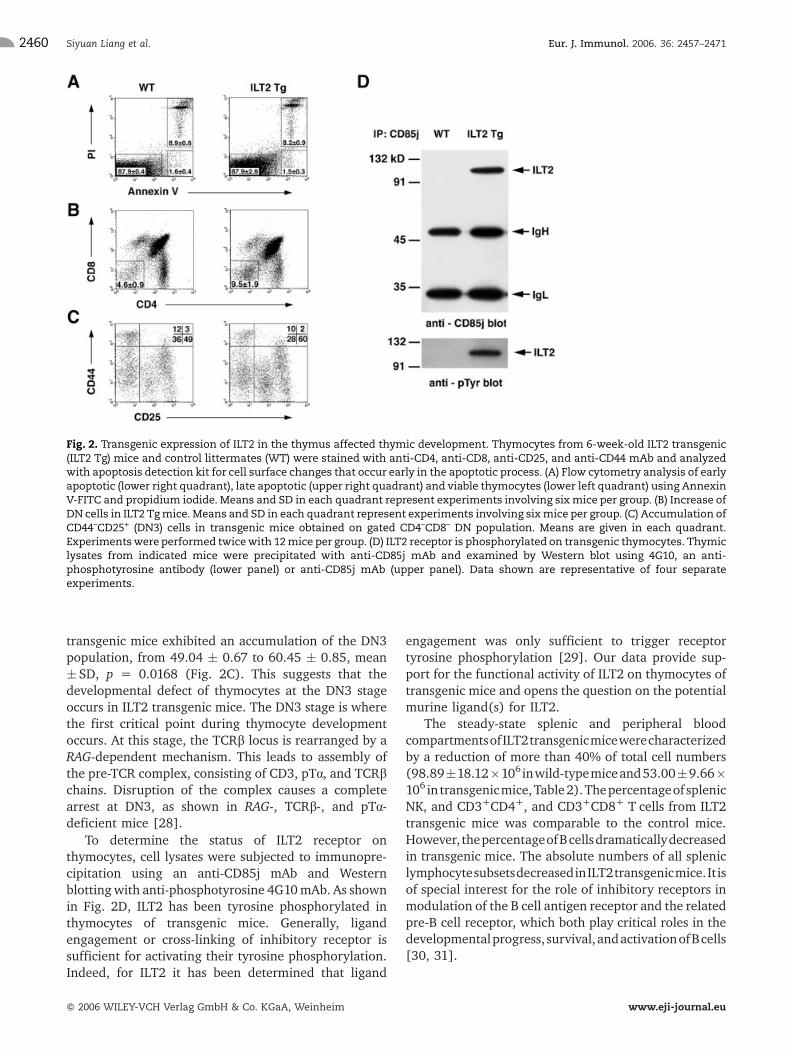
transgenic mice exhibited an accumulation of the DN3population, from 49.04 � 0.67 to 60.45 � 0.85, mean� SD, p = 0.0168 (Fig. 2C). This suggests that thedevelopmental defect of thymocytes at the DN3 stageoccurs in ILT2 transgenic mice. The DN3 stage is wherethe first critical point during thymocyte developmentoccurs. At this stage, the TCRb locus is rearranged by aRAG-dependent mechanism. This leads to assembly ofthe pre-TCR complex, consisting of CD3, pTa, and TCRbchains. Disruption of the complex causes a completearrest at DN3, as shown in RAG-, TCRb-, and pTa-deficient mice [28].
To determine the status of ILT2 receptor onthymocytes, cell lysates were subjected to immunopre-cipitation using an anti-CD85j mAb and Westernblottingwith anti-phosphotyrosine 4G10mAb. As shownin Fig. 2D, ILT2 has been tyrosine phosphorylated inthymocytes of transgenic mice. Generally, ligandengagement or cross-linking of inhibitory receptor issufficient for activating their tyrosine phosphorylation.Indeed, for ILT2 it has been determined that ligand
engagement was only sufficient to trigger receptortyrosine phosphorylation [29]. Our data provide sup-port for the functional activity of ILT2 on thymocytes oftransgenic mice and opens the question on the potentialmurine ligand(s) for ILT2.
The steady-state splenic and peripheral bloodcompartmentsofILT2transgenicmicewerecharacterizedby a reduction of more than 40% of total cell numbers(98.89�18.12�106inwild-typemiceand53.00�9.66�106 intransgenicmice,Table2).Thepercentageof splenicNK, and CD3+CD4+, and CD3+CD8+ T cells from ILT2transgenic mice was comparable to the control mice.However, thepercentageofBcellsdramaticallydecreasedin transgenic mice. The absolute numbers of all spleniclymphocytesubsetsdecreasedinILT2transgenicmice.Itisof special interest for the role of inhibitory receptors inmodulation of the B cell antigen receptor and the relatedpre-B cell receptor, which both play critical roles in thedevelopmentalprogress, survival,andactivationofBcells[30, 31].
Fig. 2. Transgenic expression of ILT2 in the thymus affected thymic development. Thymocytes from 6-week-old ILT2 transgenic(ILT2 Tg) mice and control littermates (WT) were stained with anti-CD4, anti-CD8, anti-CD25, and anti-CD44 mAb and analyzedwith apoptosis detection kit for cell surface changes that occur early in the apoptotic process. (A) Flow cytometry analysis of earlyapoptotic (lower right quadrant), late apoptotic (upper right quadrant) and viable thymocytes (lower left quadrant) using AnnexinV-FITC and propidium iodide. Means and SD in each quadrant represent experiments involving sixmice per group. (B) Increase ofDN cells in ILT2 Tgmice. Means and SD in each quadrant represent experiments involving sixmice per group. (C) Accumulation ofCD44–CD25+ (DN3) cells in transgenic mice obtained on gated CD4–CD8– DN population. Means are given in each quadrant.Experimentswere performed twicewith 12mice per group. (D) ILT2 receptor is phosphorylated on transgenic thymocytes. Thymiclysates from indicated mice were precipitated with anti-CD85j mAb and examined by Western blot using 4G10, an anti-phosphotyrosine antibody (lower panel) or anti-CD85j mAb (upper panel). Data shown are representative of four separateexperiments.
Siyuan Liang et al. Eur. J. Immunol. 2006. 36: 2457–24712460
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Impaired T cell function in ILT2 transgenic mice
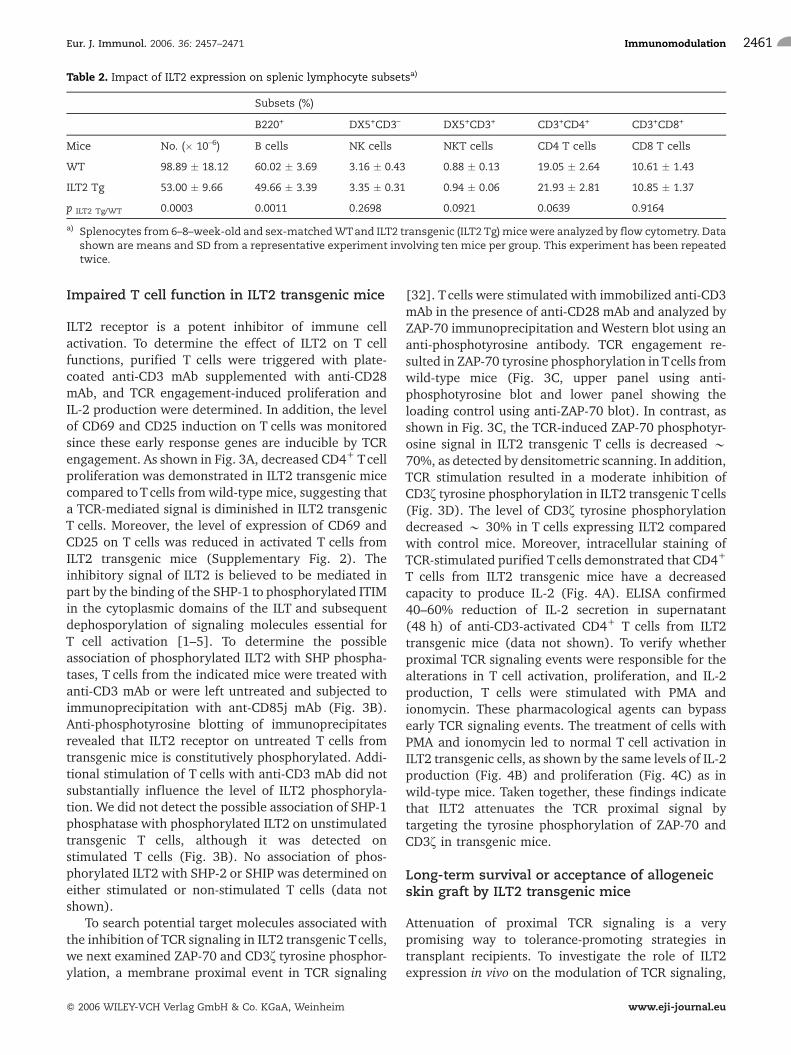
ILT2 receptor is a potent inhibitor of immune cellactivation. To determine the effect of ILT2 on T cellfunctions, purified T cells were triggered with plate-coated anti-CD3 mAb supplemented with anti-CD28mAb, and TCR engagement-induced proliferation andIL-2 production were determined. In addition, the levelof CD69 and CD25 induction on T cells was monitoredsince these early response genes are inducible by TCRengagement. As shown in Fig. 3A, decreased CD4+ Tcellproliferation was demonstrated in ILT2 transgenic micecompared toTcells from wild-type mice, suggesting thata TCR-mediated signal is diminished in ILT2 transgenicT cells. Moreover, the level of expression of CD69 andCD25 on T cells was reduced in activated T cells fromILT2 transgenic mice (Supplementary Fig. 2). Theinhibitory signal of ILT2 is believed to be mediated inpart by the binding of the SHP-1 to phosphorylated ITIMin the cytoplasmic domains of the ILT and subsequentdephosporylation of signaling molecules essential forT cell activation [1–5]. To determine the possibleassociation of phosphorylated ILT2 with SHP phospha-tases, T cells from the indicated mice were treated withanti-CD3 mAb or were left untreated and subjected toimmunoprecipitation with ant-CD85j mAb (Fig. 3B).Anti-phosphotyrosine blotting of immunoprecipitatesrevealed that ILT2 receptor on untreated T cells fromtransgenic mice is constitutively phosphorylated. Addi-tional stimulation of T cells with anti-CD3 mAb did notsubstantially influence the level of ILT2 phosphoryla-tion. We did not detect the possible association of SHP-1phosphatase with phosphorylated ILT2 on unstimulatedtransgenic T cells, although it was detected onstimulated T cells (Fig. 3B). No association of phos-phorylated ILT2 with SHP-2 or SHIP was determined oneither stimulated or non-stimulated T cells (data notshown).
To search potential target molecules associated withthe inhibition of TCR signaling in ILT2 transgenic Tcells,we next examined ZAP-70 and CD3f tyrosine phosphor-ylation, a membrane proximal event in TCR signaling
[32]. T cells were stimulated with immobilized anti-CD3mAb in the presence of anti-CD28 mAb and analyzed byZAP-70 immunoprecipitation and Western blot using ananti-phosphotyrosine antibody. TCR engagement re-sulted in ZAP-70 tyrosine phosphorylation inTcells fromwild-type mice (Fig. 3C, upper panel using anti-phosphotyrosine blot and lower panel showing theloading control using anti-ZAP-70 blot). In contrast, asshown in Fig. 3C, the TCR-induced ZAP-70 phosphotyr-osine signal in ILT2 transgenic T cells is decreased *70%, as detected by densitometric scanning. In addition,TCR stimulation resulted in a moderate inhibition ofCD3f tyrosine phosphorylation in ILT2 transgenic Tcells(Fig. 3D). The level of CD3f tyrosine phosphorylationdecreased * 30% in T cells expressing ILT2 comparedwith control mice. Moreover, intracellular staining ofTCR-stimulated purified Tcells demonstrated that CD4+
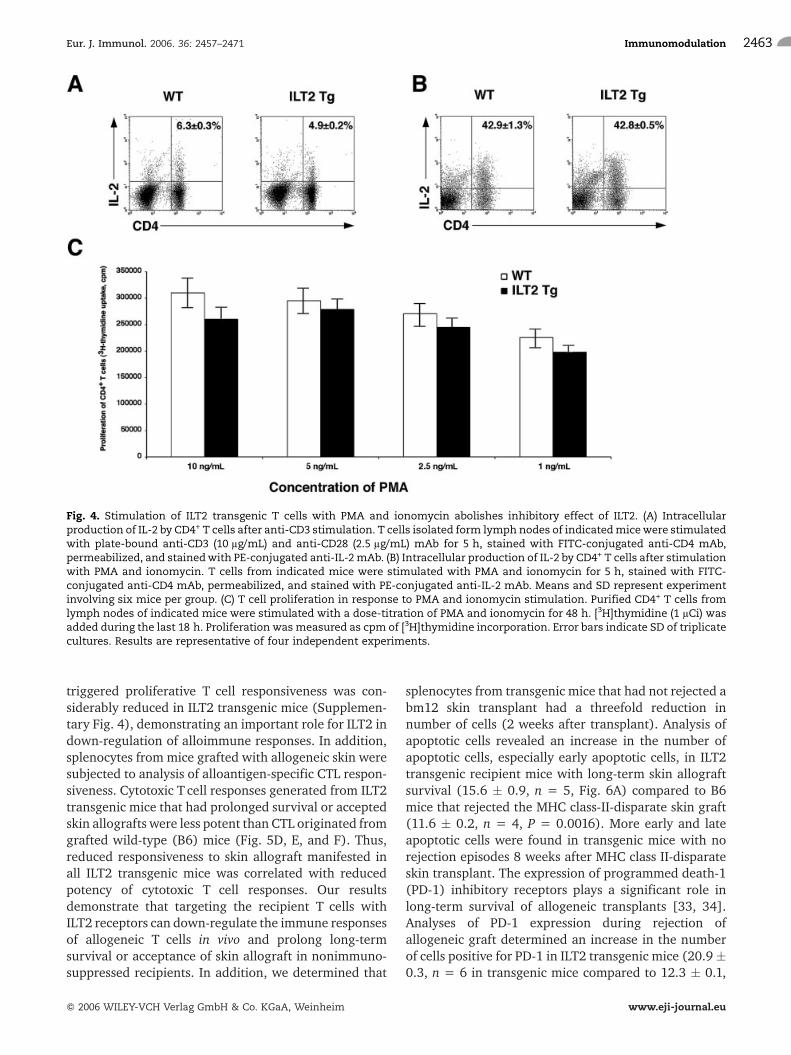
T cells from ILT2 transgenic mice have a decreasedcapacity to produce IL-2 (Fig. 4A). ELISA confirmed40–60% reduction of IL-2 secretion in supernatant(48 h) of anti-CD3-activated CD4+ T cells from ILT2transgenic mice (data not shown). To verify whetherproximal TCR signaling events were responsible for thealterations in T cell activation, proliferation, and IL-2production, T cells were stimulated with PMA andionomycin. These pharmacological agents can bypassearly TCR signaling events. The treatment of cells withPMA and ionomycin led to normal T cell activation inILT2 transgenic cells, as shown by the same levels of IL-2production (Fig. 4B) and proliferation (Fig. 4C) as inwild-type mice. Taken together, these findings indicatethat ILT2 attenuates the TCR proximal signal bytargeting the tyrosine phosphorylation of ZAP-70 andCD3f in transgenic mice.
Long-term survival or acceptance of allogeneicskin graft by ILT2 transgenic mice
Attenuation of proximal TCR signaling is a verypromising way to tolerance-promoting strategies intransplant recipients. To investigate the role of ILT2expression in vivo on the modulation of TCR signaling,
Table 2. Impact of ILT2 expression on splenic lymphocyte subsetsa)
Subsets (%)
B220+ DX5+CD3– DX5+CD3+ CD3+CD4+ CD3+CD8+
Mice No. (� 10–6) B cells NK cells NKT cells CD4 T cells CD8 T cells
WT 98.89 � 18.12 60.02 � 3.69 3.16 � 0.43 0.88 � 0.13 19.05 � 2.64 10.61 � 1.43
ILT2 Tg 53.00 � 9.66 49.66 � 3.39 3.35 � 0.31 0.94 � 0.06 21.93 � 2.81 10.85 � 1.37
p ILT2 Tg/WT 0.0003 0.0011 0.2698 0.0921 0.0639 0.9164
a) Splenocytes from 6–8–week-old and sex-matchedWTand ILT2 transgenic (ILT2 Tg)micewere analyzed by flow cytometry. Datashown are means and SD from a representative experiment involving ten mice per group. This experiment has been repeatedtwice.
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2461
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
we used a skin allograft model. Skin graft survival from afully allogeneic (CBA) donor was prolonged in ILT2transgenic mice (mean of survival time ( MST) = 16.5, n= 6, p = 0.0012, Fig. 5A). Wild-type C57BL/6 (B6) micestrongly recognized the MHC class I-disparate mutantmouse, B6.C-H-2bm1 (bm1) carrying the MHC class Ialloantigen (MST = 17.5, n = 16, Fig. 5B). In contrast,skin graft survival from bm1 donors was prolonged oraccepted in ILT2 transgenic mice (MST = 96.5, n = 22,p< 0.0001, Fig. 5B, Supplementary Fig. 3A). Moreover,50% of ILT2 transgenic mice (n = 11) never rejectedallogeneic skin from an MHC class I-mismatched donor.In another model, B6 mice more strongly recognized theMHC class II-disparate mutant mouse, B6.C-H-2bm12
(bm12) carrying the I-Abm12 alloantigen (MST = 11.3,n=12, Fig. 5C). Increased prolongation or acceptanceof skin graft from differences in MHC class II alloantigenoccurred in ILT2 transgenic mice (MST = 86.4, n = 16,p<0.0001, Fig. 5C and Supplementary Fig. 3B).Furthermore, 44% of ILT2 transgenic mice (n = 7)never rejected the allogeneic MHC class II-disparateallografts. To understand the mechanisms of long-termskin allograft survival in ILT2 transgenic mice, weanalyzed their alloantigen-triggered T cell proliferativeand CTL responsiveness. Allogeneic combinationsbetween responder cells from ILT2 transgenic mice(H-2b) and stimulator cells from CBA (H-2k), BALB/c(H-2d) mice have been used in MLR. Alloantigen-
Fig. 3. Transgenic expression of ILT2 inhibits early T cell activation. (A) Purified CD4+ T cells were isolated from 6- to 8-week-oldILT2 transgenic (ILT2 Tg) mice and control littermates (WT) and stimulated with varying amounts of plate-bound anti-CD3 mAband 2.5 lg/mL of anti-CD28 mAb and cultured for 48 h. [3H]thymidine (1 lCi) was added during the last 18 h. Proliferation wasmeasured as cpm of [3H]thymidine incorporation. Error bars indicate SD of triplicate cultures. Results represent four independentexperiments. (B) Phosphorylation of ILT2 receptor on transgenic T cells and recruitment of SHP-1 after CD3 stimulation. T cellswere isolated from lymph nodes of indicatedmice and stimulated for 30 minwith anti-CD3mAb (2.5 lg/mL) or left unstimulated,then subjected to immunoprecipitation andWestern blot analysis as indicated. Cell lysateswereprecipitatedwith anti-CD85jmAband examined byWestern blot using 4G10, an anti-phosphotyrosine antibody (middle panel) orwith anti-CD85jmAb (upper panel)or anti-SHP-1 Ab (lower panel). Data shown are representative of four separate experiments. (C) Reduced ZAP-70 phosphorylationin ILT2 Tgmice after T cell activation. T cells from indicatedmicewere stimulatedwith anti-CD3mAb for 30 min. Cell lysateswereimmunoprecipitated with anti-ZAP-70 mAb and subjected to Western blot analysis with anti-phosphotyrosine antibody 4G10(upper panel) or anti-ZAP-70mAb (lower panel). Densitometry analysiswas performed. (D) Reduced level of CD3f phosphorylationin ILT2 Tg mice after activation of T cells. T cells from indicated mice were stimulated with anti-CD3 mAb for 30 min. Cell lysateswere immunoprecipitated with anti-CD3f mAb and subjected to Western blot analysis with anti-phosphotyrosine antibody 4G10(upper panel) or anti-CD3f mAb (lower panel). Densitometry analysis was performed.
Siyuan Liang et al. Eur. J. Immunol. 2006. 36: 2457–24712462
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
triggered proliferative T cell responsiveness was con-siderably reduced in ILT2 transgenic mice (Supplemen-tary Fig. 4), demonstrating an important role for ILT2 indown-regulation of alloimmune responses. In addition,splenocytes from mice grafted with allogeneic skin weresubjected to analysis of alloantigen-specific CTL respon-siveness. Cytotoxic T cell responses generated from ILT2transgenic mice that had prolonged survival or acceptedskin allografts were less potent than CTL originated fromgrafted wild-type (B6) mice (Fig. 5D, E, and F). Thus,reduced responsiveness to skin allograft manifested inall ILT2 transgenic mice was correlated with reducedpotency of cytotoxic T cell responses. Our resultsdemonstrate that targeting the recipient T cells withILT2 receptors can down-regulate the immune responsesof allogeneic T cells in vivo and prolong long-termsurvival or acceptance of skin allograft in nonimmuno-suppressed recipients. In addition, we determined that
splenocytes from transgenic mice that had not rejected abm12 skin transplant had a threefold reduction innumber of cells (2 weeks after transplant). Analysis ofapoptotic cells revealed an increase in the number ofapoptotic cells, especially early apoptotic cells, in ILT2transgenic recipient mice with long-term skin allograftsurvival (15.6 � 0.9, n = 5, Fig. 6A) compared to B6mice that rejected the MHC class-II-disparate skin graft(11.6 � 0.2, n = 4, P = 0.0016). More early and lateapoptotic cells were found in transgenic mice with norejection episodes 8 weeks after MHC class II-disparateskin transplant. The expression of programmed death-1(PD-1) inhibitory receptors plays a significant role inlong-term survival of allogeneic transplants [33, 34].Analyses of PD-1 expression during rejection ofallogeneic graft determined an increase in the numberof cells positive for PD-1 in ILT2 transgenic mice (20.9�0.3, n = 6 in transgenic mice compared to 12.3 � 0.1,
Fig. 4. Stimulation of ILT2 transgenic T cells with PMA and ionomycin abolishes inhibitory effect of ILT2. (A) Intracellularproduction of IL-2 by CD4+ T cells after anti-CD3 stimulation. T cells isolated form lymph nodes of indicatedmicewere stimulatedwith plate-bound anti-CD3 (10 lg/mL) and anti-CD28 (2.5 lg/mL) mAb for 5 h, stained with FITC-conjugated anti-CD4 mAb,permeabilized, and stainedwith PE-conjugated anti-IL-2mAb. (B) Intracellular production of IL-2 by CD4+ T cells after stimulationwith PMA and ionomycin. T cells from indicated mice were stimulated with PMA and ionomycin for 5 h, stained with FITC-conjugated anti-CD4 mAb, permeabilized, and stained with PE-conjugated anti-IL-2 mAb. Means and SD represent experimentinvolving six mice per group. (C) T cell proliferation in response to PMA and ionomycin stimulation. Purified CD4+ T cells fromlymph nodes of indicated mice were stimulated with a dose-titration of PMA and ionomycin for 48 h. [3H]thymidine (1 lCi) wasadded during the last 18 h. Proliferation was measured as cpm of [3H]thymidine incorporation. Error bars indicate SD of triplicatecultures. Results are representative of four independent experiments.
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2463
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
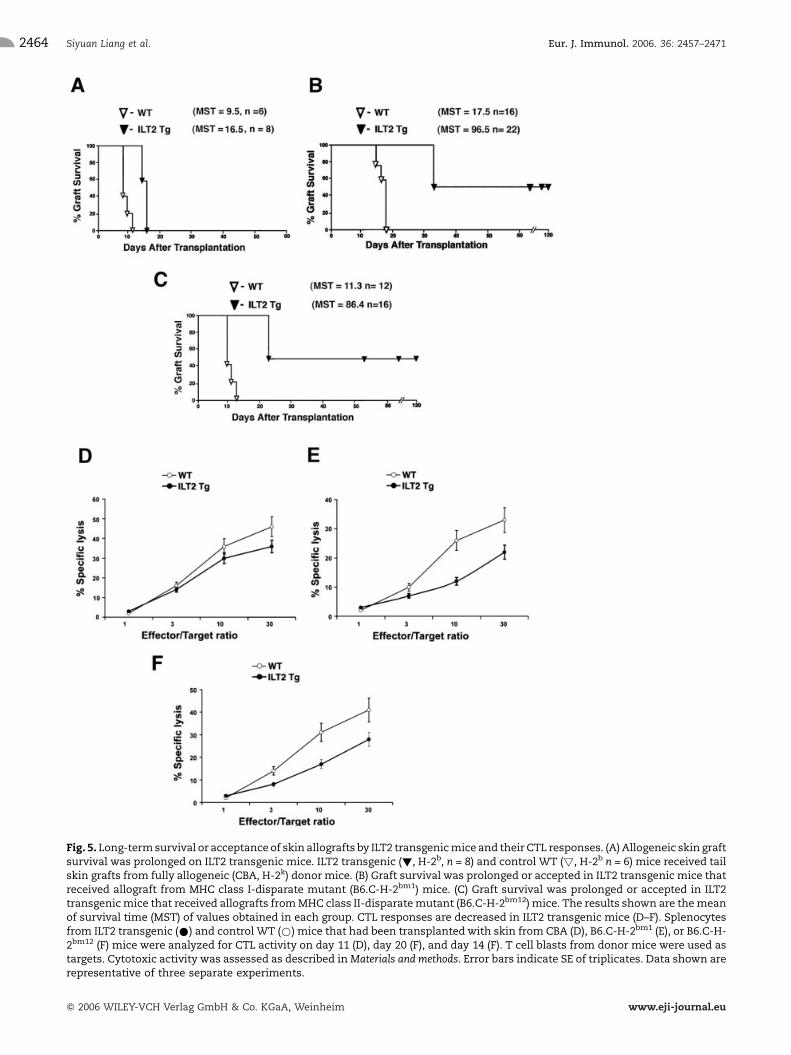
Fig. 5. Long-termsurvival or acceptance of skin allografts by ILT2 transgenicmice and their CTL responses. (A) Allogeneic skin graftsurvival was prolonged on ILT2 transgenic mice. ILT2 transgenic (!, H-2b, n = 8) and control WT (!, H-2b n = 6) mice received tailskin grafts from fully allogeneic (CBA, H-2k) donor mice. (B) Graft survival was prolonged or accepted in ILT2 transgenic mice thatreceived allograft from MHC class I-disparate mutant (B6.C-H-2bm1) mice. (C) Graft survival was prolonged or accepted in ILT2transgenicmice that received allografts fromMHC class II-disparatemutant (B6.C-H-2bm12) mice. The results shown are themeanof survival time (MST) of values obtained in each group. CTL responses are decreased in ILT2 transgenic mice (D–F). Splenocytesfrom ILT2 transgenic (*) and control WT (*) mice that had been transplantedwith skin from CBA (D), B6.C-H-2bm1 (E), or B6.C-H-2bm12 (F) mice were analyzed for CTL activity on day 11 (D), day 20 (F), and day 14 (F). T cell blasts from donor mice were used astargets. Cytotoxic activity was assessed as described inMaterials and methods. Error bars indicate SE of triplicates. Data shown arerepresentative of three separate experiments.
Siyuan Liang et al. Eur. J. Immunol. 2006. 36: 2457–24712464
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
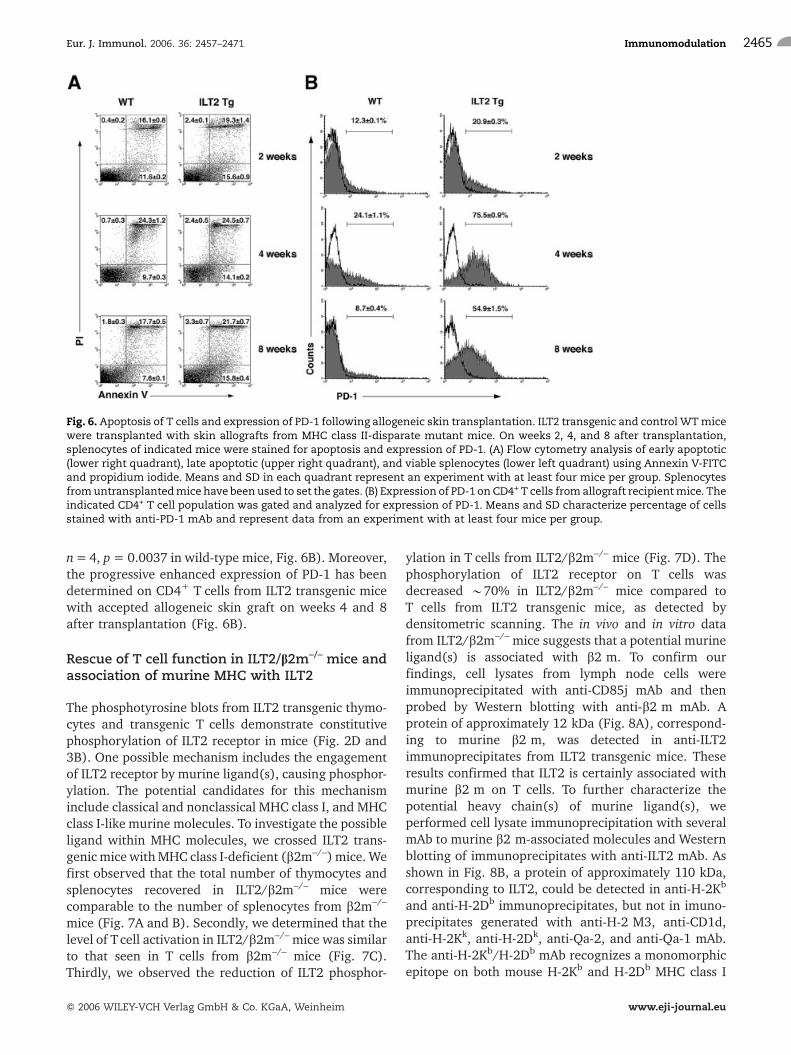
n=4, p= 0.0037 in wild-type mice, Fig. 6B). Moreover,the progressive enhanced expression of PD-1 has beendetermined on CD4+ T cells from ILT2 transgenic micewith accepted allogeneic skin graft on weeks 4 and 8after transplantation (Fig. 6B).
Rescue of T cell function in ILT2/b2m–/– mice andassociation of murine MHC with ILT2
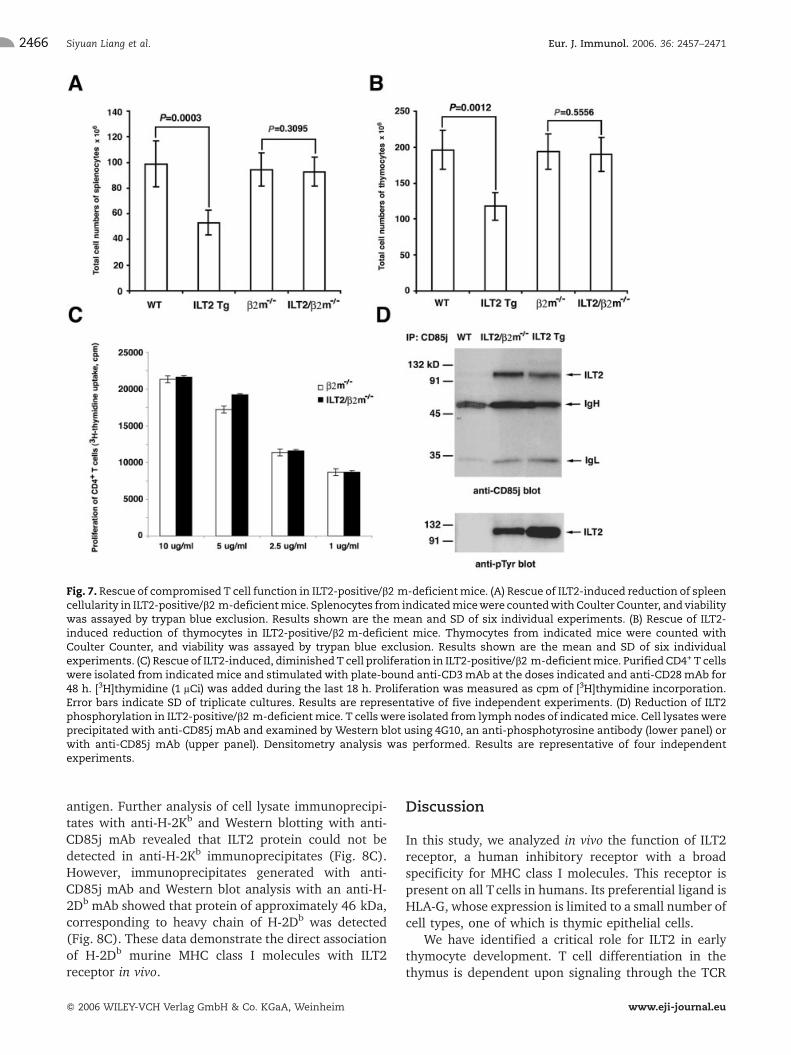
The phosphotyrosine blots from ILT2 transgenic thymo-cytes and transgenic T cells demonstrate constitutivephosphorylation of ILT2 receptor in mice (Fig. 2D and3B). One possible mechanism includes the engagementof ILT2 receptor by murine ligand(s), causing phosphor-ylation. The potential candidates for this mechanisminclude classical and nonclassical MHC class I, and MHCclass I-like murine molecules. To investigate the possibleligand within MHC molecules, we crossed ILT2 trans-genicmice withMHC class I-deficient (b2m–/–) mice. Wefirst observed that the total number of thymocytes andsplenocytes recovered in ILT2/b2m–/– mice werecomparable to the number of splenocytes from b2m–/–
mice (Fig. 7A and B). Secondly, we determined that thelevel of Tcell activation in ILT2/b2m–/– mice was similarto that seen in T cells from b2m–/– mice (Fig. 7C).Thirdly, we observed the reduction of ILT2 phosphor-
ylation in T cells from ILT2/b2m–/– mice (Fig. 7D). Thephosphorylation of ILT2 receptor on T cells wasdecreased *70% in ILT2/b2m–/– mice compared toT cells from ILT2 transgenic mice, as detected bydensitometric scanning. The in vivo and in vitro datafrom ILT2/b2m–/– mice suggests that a potential murineligand(s) is associated with b2 m. To confirm ourfindings, cell lysates from lymph node cells wereimmunoprecipitated with anti-CD85j mAb and thenprobed by Western blotting with anti-b2 m mAb. Aprotein of approximately 12 kDa (Fig. 8A), correspond-ing to murine b2 m, was detected in anti-ILT2immunoprecipitates from ILT2 transgenic mice. Theseresults confirmed that ILT2 is certainly associated withmurine b2 m on T cells. To further characterize thepotential heavy chain(s) of murine ligand(s), weperformed cell lysate immunoprecipitation with severalmAb to murine b2 m-associated molecules and Westernblotting of immunoprecipitates with anti-ILT2 mAb. Asshown in Fig. 8B, a protein of approximately 110 kDa,corresponding to ILT2, could be detected in anti-H-2Kb
and anti-H-2Db immunoprecipitates, but not in imuno-precipitates generated with anti-H-2 M3, anti-CD1d,anti-H-2Kk, anti-H-2Dk, anti-Qa-2, and anti-Qa-1 mAb.The anti-H-2Kb/H-2Db mAb recognizes a monomorphicepitope on both mouse H-2Kb and H-2Db MHC class I
Fig. 6. Apoptosis of T cells and expression of PD-1 following allogeneic skin transplantation. ILT2 transgenic and control WTmicewere transplanted with skin allografts from MHC class II-disparate mutant mice. On weeks 2, 4, and 8 after transplantation,splenocytes of indicated mice were stained for apoptosis and expression of PD-1. (A) Flow cytometry analysis of early apoptotic(lower right quadrant), late apoptotic (upper right quadrant), and viable splenocytes (lower left quadrant) using Annexin V-FITCand propidium iodide. Means and SD in each quadrant represent an experiment with at least four mice per group. Splenocytesfromuntransplantedmice have been used to set the gates. (B) Expression of PD-1 on CD4+ T cells fromallograft recipientmice. Theindicated CD4+ T cell population was gated and analyzed for expression of PD-1. Means and SD characterize percentage of cellsstained with anti-PD-1 mAb and represent data from an experiment with at least four mice per group.
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2465
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
antigen. Further analysis of cell lysate immunoprecipi-tates with anti-H-2Kb and Western blotting with anti-CD85j mAb revealed that ILT2 protein could not bedetected in anti-H-2Kb immunoprecipitates (Fig. 8C).However, immunoprecipitates generated with anti-CD85j mAb and Western blot analysis with an anti-H-2Db mAb showed that protein of approximately 46 kDa,corresponding to heavy chain of H-2Db was detected(Fig. 8C). These data demonstrate the direct associationof H-2Db murine MHC class I molecules with ILT2receptor in vivo.
Discussion
In this study, we analyzed in vivo the function of ILT2receptor, a human inhibitory receptor with a broadspecificity for MHC class I molecules. This receptor ispresent on all T cells in humans. Its preferential ligand isHLA-G, whose expression is limited to a small number ofcell types, one of which is thymic epithelial cells.
We have identified a critical role for ILT2 in earlythymocyte development. T cell differentiation in thethymus is dependent upon signaling through the TCR
Fig. 7. Rescue of compromised T cell function in ILT2-positive/b2 m-deficientmice. (A) Rescue of ILT2-induced reduction of spleencellularity in ILT2-positive/b2 m-deficientmice. Splenocytes from indicatedmicewere countedwith Coulter Counter, and viabilitywas assayed by trypan blue exclusion. Results shown are the mean and SD of six individual experiments. (B) Rescue of ILT2-induced reduction of thymocytes in ILT2-positive/b2 m-deficient mice. Thymocytes from indicated mice were counted withCoulter Counter, and viability was assayed by trypan blue exclusion. Results shown are the mean and SD of six individualexperiments. (C) Rescue of ILT2-induced, diminishedT cell proliferation in ILT2-positive/b2 m-deficientmice. Purified CD4+ T cellswere isolated from indicated mice and stimulated with plate-bound anti-CD3mAb at the doses indicated and anti-CD28mAb for48 h. [3H]thymidine (1 lCi) was added during the last 18 h. Proliferation was measured as cpm of [3H]thymidine incorporation.Error bars indicate SD of triplicate cultures. Results are representative of five independent experiments. (D) Reduction of ILT2phosphorylation in ILT2-positive/b2 m-deficientmice. T cellswere isolated from lymph nodes of indicatedmice. Cell lysateswereprecipitated with anti-CD85j mAb and examined by Western blot using 4G10, an anti-phosphotyrosine antibody (lower panel) orwith anti-CD85j mAb (upper panel). Densitometry analysis was performed. Results are representative of four independentexperiments.
Siyuan Liang et al. Eur. J. Immunol. 2006. 36: 2457–24712466
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

and is characterized by the resulting changes inexpression patterns of CD4 and CD8 surface co-receptormolecules. Thymocytes from ILT2 transgenic miceshowed an impaired ability to transit from the DN3 toDN4 stage of development, suggesting that ILT2 has animpact on this stage of development. In addition, thymiccellularity was reduced in ILT2 transgenic mice. The firstcritical checkpoint, which is called b-selection, takesplace during thymocyte differentiation and ensures thatonly thymocytes that have generated a functional TCR-bchain become selected to differentiate to the DP stage[35–37]. Although the TCR- b gene locus is presentduring the DN1 and DN2 stages of development, theb-selection occurs only when the TCR-b gene iscovalently coupled to the non-polymorphic pre-Ta(pTa) chain and CD3 molecules to form the pre-TCRcomplex, which is expressed on DN3 thymocytes. TheILT2 receptor is expressed in transgenic mice on allstages of DN thymocytes (data not shown). In addition,it is well known that classical MHC class I molecules areexpressed at all stages of DN thymocytes as well. Thisindicates that pre-TCR-negative DN1 and DN2 cells are
not affected by the ILT2-ligand interaction. In contrast,the expression of ILT2 affects the development ofthymocytes at the DN3 stage, which are pre-TCR-positive cells, suggesting that ILT2 alters the formationor function of the pre-TCR complex. It is of specialinterest to elucidate the possible molecular and signal-ing mechanisms of negative regulation of ILT2 on theformation and function of the pre-TCR complex, a highlycomplex and multifaceted process that ensures thesurvival and differentiation of cells expressing afunctional TCR-b chain [28, 37]. It is still an openquestion if ILT2 affects individual components of thepre-TCR complex or inhibits the signals induced by thepre-TCR-composed complex. It is possible that ILT20smajor target during thymocyte development is theproximal signaling events activated following pre-TCRcomplex formation. Further analysis is required toclarify the role of ILT2 on the pre-TCR complex.
As the major target of ILT2 is TCR signaling, it isimportant to find out which signaling molecules aremodulated by ILT2 in vivo for potential in preventingallogeneic graft rejection. Here, we demonstrate that on
Fig. 8.Association of ILT2withmurine b2 mand heavy chain of H-2DbMHC class Imolecule. (A)Murine b2 massociatedwith ILT2.T cellswere isolated from lymph nodes of ILT2 transgenicmice. Cell lysateswere precipitatedwith anti-CD85jmAb and examinedby Western blot using an anti-CD85j mAb (upper panel) or with anti-b2 m mAb (lower panel). Results are representative of fiveindependent experiments. (B) Association of H-2KbDb molecules with ILT2. T cells were isolated from lymph nodes of ILT2transgenic mice. Cell lysateswere precipitatedwith anti-CD1, anti-H-2 M3, anti-H-2KbDb, anti-H-2Kk, anti-H-2Dk, anti-Qa-2, anti-Qa-1, and anti-CD85jmAb and examined byWestern blot using an anti-CD85jmAb. Results are representative of four independentexperiments. (C) Heavy chain of murine H-2Kb molecules does not associate with ILT2. T cells were isolated from lymph nodes ofindicated mice. Cell lysates were precipitated with anti-CD85j mAb (two left lanes) or with anti-H-2Kb mAb (two right lanes) andexamined by Western blot using anti-CD85j mAb. Results are representative of four independent experiments. (D) Murine heavychain of H-2Db molecule associates with ILT2. T cells were isolated from lymph nodes of indicated mice. Cell lysates wereprecipitated with anti-CD85j mAb and examined by Western blot using an anti-H-2Db mAb. Results are representative of fiveindependent experiments.
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2467
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

mature T lymphocytes, ILT2 targets the proximal eventsin TCR signaling. Targeting the early TCR signals is avery promising strategy for blocking T cell activation inalloimmune responses to prevent allograft rejection. Incontrast, molecules that become activated late in thesignaling cascade are found in several signaling path-ways and inmany different cell types. Themodulation ofthese molecules should target several cell types and leadto severe side effects [38]. We demonstrate that inmature Tcells, ILT2 targets ZAP-70 and CD3f, which areessential molecules for early TCR-mediated T cellactivation. Studies have shown that targeting ZAP-70and CD3f might be useful for preventing graft rejection[38, 39]. In our model, the long-term survival ofallogeneic graft and acceptance of allografts in ILT2transgenic mice were correlated with considerablyreduced allogeneic T cell and CTL responsiveness,demonstrating an important role for ILT2 in theregulation of alloantigen-specific T cells and CTLresponses. The interaction between TCR and alloanti-gens leads to the phosphorylation of tyrosine residues inthe ITAM of several signaling molecules. It is most likelythat targeting of CD3f and ZAP-70 by ILT2 duringallorecognition attenuates TCR signals, which are notsufficient to fully activate allogeneic T cells and lead toT cell unresponsiveness. This study indicates thattargeting multiple signaling molecules of TCR proximalevents by ILT2 might be useful for preventing graftrejection.
In addition, in our model, the long-term survival ofallogeneic graft and acceptance of allografts in ILT2transgenic mice have been correlated with high expres-sion of the PD-1 receptor, a homologue of CD28 onT cells, which represents a new costimulatory pathway.PD-1 receptor is expressed by activated, but notunstimulated, T cells and plays an important role inmaintenance of peripheral tolerance to self-antigens andprevention of allograft rejection [33]. It is of interest toelucidate the exact mechanisms of regulation ofalloimmune responses in vivo by ILT2. Data from ourstudies suggest that these mechanisms involve both theattenuation of TCR signaling and promotion or up-regulation of negative costimulatory signals involvingthe PD-1 pathway. Further studies will be required todetermine themolecular basis for association of the ILT2and PD-1 pathways and the time points when thisassociation occurs following alloantigenic challenge.The progressive increased expression of PD-1 on T cellsfrom ILT2 transgenic mice with accepted allogeneic skingraft and moderate increase of apoptotic T cells suggestthe important role of PD-1 in maintenance of the long-term survival of allografts, probably due to an effect oncell-cycle arrest rather than cell death. This function ofPD-1 has been demonstrated previously [33, 34].
We have identified at least one of the murine MHCclass I molecules, H-2Db, that interact with human ILT2.Cross-species recognition is well known for severalmolecules onTcells [10,11], dendritic cells [40], and NKcells [41] and may create the major barrier forxenotransplantation. In contrast, cross-species compat-ibility can be used for the induction of xenografttolerance in humans in the context of pig-to-humanxenotransplantation. Warrens et al. [42] demonstratedthe molecular interactions of human ICAM-1 (CD54)with its ligands on porcine cells, which will benefit thesuccess of porcine bone-marrow-induced tolerance inhumans. Humans and mice share CD94/NKG2 receptorson NK cells, suggesting that this NK recognition systemarose before the divergence of primate and mouseancestors [41]. Buslepp et al. [10] provide a basis forcross-species cross-reactivity, elucidated by the crystalstructure of a complex involving a mouse TCR and ahuman HLA-A2 molecule. While the polymorphic a1and a2 domains of human MHC class I heavy chain forma complex for antigen-specific TCR or allele-specifickiller inhibitory receptors, a3 and b2m form the bindingsite for ILT2. It is possible that in the murine model, thebinding of ILT2 receptor is more dependent on theinteraction with b2m, which facilitates the activation ofreceptor. Our data with ILT2 transgenic mice on a b2mknockout background support this hypothesis. Inparticular, the phenotype and Tcell function are rescuedin ILT2/b2m–/– transgenic mice. This applies only forCD4+ T cells since b2m–/– mice have defective CD8+
Tcells. However, immunoprecipitation andWestern blotanalysis demonstrate a link of ILT2 receptor withmurineb2m-associated MHC class I molecules. We do notexclude the possibility that another classical or non-classical MHC class I or MHC class I-like molecule caninteract with human ILT2 immunoreceptor to potentiatethe inhibitory effects in transgenic mice. To betterunderstand how surface ILT2 receptors and H-2Db
mediate the intracellular signaling pathways, furtheranalyses of their binding affinities and the determina-tion of whether they act in cis or trans interactions arenecessary. To analyze the type of interaction, thechimeric mouse was generated where bone marrowfrom the ILT2/b2m–/– mice was placed into an irradiatedC57BL/6 mouse. There were no differences in the levelof T cell activation between the chimeric mouse and theILT2/b2m–/– mouse (data not shown), suggesting that acis interaction of H-2Db with ILT2 was important.
Here, we provide an in vivo model in which tyrosinephosphorylation of ILT2 by its ligand affects thymicdevelopment, interferes with the proximal TCR signal-ing, inhibits T cell function, and down-regulatesalloimmune responses. Our finding provides an oppor-tunity to evaluate the importance of the interactionbetween ILT2 and its ligands in terms of its ability to
Siyuan Liang et al. Eur. J. Immunol. 2006. 36: 2457–24712468
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

regulate immune responses during maternal-fetal tol-erance, survival of allografts, tumor-escape mechan-isms, and development of cytomegalovirus infectionwhen both ILT2 and its ligands are expressed. Furtherstudies will not only clarify themechanisms of inhibitoryfunctions of ILT2 and their ligands, but also providenovel approaches to achieve a reproducible transplanta-tion tolerance.
Materials and methods
Generation of ILT2 transgenic mice
To direct the expression of ILT2 on all T cells, we used animproved version of a human CD2 minigene-based vector,provided by Dr. D. Kioussis at the National Institute for MedicalResearch, London, UK. This cassette (11.2 kb) includesflanking sequences of the promoter, a unique transcriptioninitiation site, and the locus control region of the human CD2gene. This expression cassette (VA hCD2) has been used todrive the T cell-specific expression of various transgenes invivo. ILT2 cDNA (2.0-kb fragment, provided byDr.M. Colonna,Washington University School of Medicine, St. Louis, MO) wassubcloned into the SmaI site of the expression vector. Afterconfirming the nucleotide sequence of the ILT2 expressionvector, the 13.2-kb plasmid-free fragment was microinjectedinto the pronuclei of fertilized oocytes of C57BL/6 mice. Theexpression of ILT2 receptor on murine cells has beendetermined by flow cytometry using an anti-CD85j mAb (BDBiosciences, San Diego, CA). Two independent lines of ILT2transgenic mice have been established and maintained on aC57BL/6 background as transgenic heterozygotes. MHC classI-deficient (b2m–/–) mice were maintained on a C57BL/6background. Experiments were conducted in accordance withinstitutional guidelines for animal care and use.
Preparations of cell suspensions
Thymuses, spleens, and lymph nodes from 6- to 8-wk-old micewere removed and disrupted between glass slides and washedtwice with IMDM medium (Life Technologies, Gaithersburg,MD). Cells debris was depleted by passage through cellstrainer. Splenocytes and thymocytes were treated withammonium chloride-potassium bicarbonate solution(150 mM NH4Cl and 10 mM KHCO3) to lyse RBC. Forexperiments requiring isolation of Tcells, CD4+ Tcells, spleniccell preps were depleted of other cell types by the use of T cellisolation kit, and of CD4+ T cell isolation kit (Miltenyi Biotec,Auburn, CA). All cells were counted with a Coulter Counter(Coulter, Miami, FL), and viability was assayed by trypan blueexclusion.
Immunoprecipitation and Western blot analysis
The pellets from splenocytes, thymocytes, and purified T cells(5 � 106) were lysed in 1 mL radioimmunoprecipitation(RIPA) buffer and precleared several times with Sepharose-coupled protein G. Proteins were immunoprecipitated with
anti-CD85j (ILT2) mAb (VMP55, Santa Cruz Biotechnology,Santa Cruz, CA), anti-ZAP-70 mAb (eBioscience, San Diego,CA), anti-CD3f mAb (6B10, Zymed Laboratories, San Fran-cisco, CA), anti-CD1 (M-276, Santa Cruz Biotechnology), anti-H-2 M3 (130, BD Biosciences), anti-H-2Kb/Db (2G5, Serotec,Raleigh, NC), anti-H-2Kk (11–4.1, eBioscience), anti-H-2Dk
(15-5-5, eBioscience), anti-Qa-1 (6A8.6F10, Santa CruzBiotechnology), anti-Qa-2 (69H1-9-9, eBioscience), or anti-H-2Kb (AF6–88.5, Biolegend, San Diego, CA), and protein GSepharose (Amersham Pharmacia Biotech, Piscataway, NJ).Immunoblotting was performed with anti-phosphotyrosineantibody (4G10; Upstate USA, Charlottesville, VA), anti-CD85jmAb, ant-ZAP-70 mAb, anti-SHP-1 (Upstate), anti-SHP-2(Upstate), anti-SHIP (Upstate), anti-CD3f, anti-b2m (M-20,Santa Cruz Biotechnology) or anti-H-2Db mAb (KH95,Biolegend). Signals were developed using the ECL system(Amersham Pharmacia Biotech). Cell extracts were alsosubjected to Western blot analysis with antibodies againstLck (3A5), Bcl-2 (C-2), Fyn (clone 15), and b-actin (AC-15).These antibodies were from Santa Cruz Biotechnology, exceptanti-b-actin (Sigma). In some cases, densitometric scans ofbands were obtained to determine the extent of ILT2-mediatedsignal inhibition.
T cell activation
Activation of T cells via TCR/CD3 complex was assessed bystimulation of 4 � 105 purified CD4+ T cells with gradedconcentrations of plate-bound anti-CD3 mAb (eBioscience)plus 2.5 lg/mL of anti-CD28 mAb (eBioscience) or gradedconcentration of PMA (Sigma) plus 200 ng/mL of ionomycin(Sigma) in complete medium for 48 h in 96-well, flat-bottomplates. During the last 18 h of culture, 1 lCi of [3H]hymidinewas added to each well, and cellular incorporation wasdetermined with a liquid scintillation counter. All experimentswere performed in triplicate.
CTL assay
Effector cells were generated in MLR and tested in a standard51Cr-release assay. Briefly, spleen cells (5 � 106/mL) from WTor ILT2 transgenic mice that had been transplanted withallogeneic skin grafts were co-cultured with irradiated (20 Gy)allogeneic splenocytes (5 � 106/mL) for 5 days. Cytotoxicitytests were performed in triplicates at E:T ratios of 30:1, 10:1,3:1, and 1:1. Target cells were incubated for 90 min at 37�Cwith 100 lCi 51Cr (GE Healthcare Bio-Sciences, Piscataway,NJ) before being mixed with effector cells. After 4 h at 37�C,25 lL of supernatant was collected and counted (Betaplate1205, Wallac, Turku, Finland). T cells blasts were generated bycultured splenocytes with 2 lg/mL of Con A (Sigma) and usedas targets. Allospecific CTL activity was determined as % ofspecific lysis by standard formula.
Antibodies and flow cytometry
Cells were stained with FITC-, PE-, CyChrome- or allophyco-cyanin-conjugated mAb specific for CD4 (RM4–5), CD8a(53–6.7), CD44 (IM7), B220 (RA3–6B2), CD49b (DX5),CD127 (A7R34), Bcl-2 (10C4), PD-1 (J43), and isotype
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2469
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

controls rat IgG2a, PE-rat IgG2b and FITC-rat IgM were fromeBioscience. CD3-CyChrome- (145–2C11), CD25-FITC- (7D4),CD69-PE- (H1.2F3)-conjugated mAb were from BD Bios-ciences. Cells were incubated with primary antibodies in PBScontaining 2% BSA for 30 min at 4�C and washed twice. Whennecessary, conjugated secondary antibodies were used. A BDBiosciences FACSCalibur (Mountain View, CA) was used fordata acquisition, and CellQuest software was used for analysis.
Intracellular staining of proteins and cytokines,detection of apoptotic process
106 lymphocytes were stimulated with PMA (50 ng/mL) andionomycin (500 ng/mL) for 5 h in complete medium in thepresence of BD GolgiPlugTM (BD Biosciences). Cells wereharvested and stained with allophycocyanin-conjugated anti-CD4 mAb for 30 min at 4�C. Cells were washed followed byincubation with cytofix/cytoperm solution (BD Biosciences).Permeabilized cells were stained with PE-conjugated anti-IL-2mAb (JES6–5H4, eBioscience) or with anti-Bcl-2 mAb for30 min at 4�C. For some experiments, cells were stimulatedwith anti-CD3 and anti-CD28 mAb. For detection earlyapoptotic, late apoptotic, necrotic and viable cells, thymocytesand lymphocytes were incubated with 5 lL of Annexin V and5 lg/mL of PI in 100 lL Annexin V binding buffer for 15 minat room temperature (BD Biosciences). The cells were analyzedusing FACSCalibur with CellQuest Analysis software.
Skin grafting
Specific pathogen-free C57BL/6 (H-2b) mice and ILT2transgenic mice (H-2b) (8–10 weeks of age) were used asskin graft recipients throughout the study. Donor skin wasfrom fully allogeneic CBA (H-2k), MHC class I-disparate B6.C-H-2bm1 (bm1, H-2b), or MHC class II-disparate B6.C-H-2bm12
(bm12, H-2b) mice. Allogeneic skin grafts were performed bystandard methods as described previously [40].
Statistical analysis
Where applicable, values were compared by theMann-WhitneyU test using Stata 7.0 software.
Acknowledgements:We thank Dr. Dimitris Kioussis forthe original gift of the VA hCD2 minigene-based vector,and Drs. Marina Cella and Marco Colonna for ILT2cDNA. We thank Drs. Demetrius Moskophidis, NahidMivechi, Rhea-Beth Markowitz for advice and criticalcomments on the manuscript. Research support wasprovided by NIH grant RO1 AI 055923.
References
1 Colonna, M., Navarro, F., Bellon, T., Llano, M., Garcia, P., Samaridis, J.,Angman, L. et al., A common inhibitory receptor for major histocompat-ibility complex class I molecules on human lymphoid and myelomonocyticcells. J. Exp. Med. 1997. 186: 1809–1818.
2 Cosman, D., Fanger, N., Borges, L., Kubin, M., Chin, W., Peterson, L. andHsu, M. L., A novel immunoglobulin superfamily receptor for cellular andviral MHC class I molecules. Immunity 1997. 7: 273–282.
3 Brown, D., Trowsdale, J. and Allen, A., The LILR family: modulators ofinnate and adaptive immune pathways in health and disease. Tissue Antigens2004. 64: 215–225.
4 Veillette, A., Latour, S. and Davidson, D., Negative regulation ofimmunoreceptor signaling. Annu. Rev. Immunol. 2002. 20: 669–707.
5 Long, E. O., Regulation of immune responses through inhibitory receptors.Annu. Rev. Immunol. 1999. 17: 875–904.
6 Chapman, T. L., Heikema, A. P. and Bjorkman, P. J., The inhibitoryreceptor LIR-1 uses a common binding interaction to recognize class I MHCmolecules and the viral homolog UL18 . Immunity 1999. 11: 603–613.
7 Willcox, B. E., Thomas, L. M., Chapman, T. L., Heikema, A. P., West, A. P.Jr. and Bjorkman, P. J., Crystal structure of LIR-2 (ILT4) at 1.8 A:differences from LIR-1 (ILT2) in regions implicated in the binding of thehuman cytomegalovirus class I MHC homolog UL18. BMC Struct. Biol. 2002.2: 6.
8 Chapman, T. L., Heikema, A. P.,West, A. P. Jr. and Bjorkman, P. J., Crystalstructure and ligand binding properties of the D1D2 region of the inhibitoryreceptor LIR-1 (ILT2) . Immunity 2000. 13: 727–736.
9 Willcox, B. E., Thomas, L.M. and Bjorkman, P. J.,Crystal structure of HLA-A2 bound to LIR-1, a host and viral major histocompatibility complexreceptor. Nat. Immunol. 2003. 4: 913–919.
10 Buslepp, J., Wang, H., Biddison, W. E., Appella, E. and Collins, E. J., Acorrelation between TCR V alpha docking on MHC and CD8 dependence:implication for T cell selection. Immunity 2003. 19: 595–606.
11 Housset, D. and Malissen, B., What doTCR-pMHC crystal structures teachus about MHC restriction and alloreactivity? Trends Immunol 2003. 24:429–437.
12 Clements, C. S., Kjer-Nielson, L., Kostenko, L., Hoare, H. L., Dunstone,M.A., Moses, E., Freed, K. et al., Crystal structure of HLA-G: a nonclassicalMHC class I molecule expressed at the fetal-maternal interface. Proc. Natl.Acad. Sci. USA 2005. 102: 3360–3365.
13 Shiroishi, M., Tsumoto, K., Amano, K., Shirakihara, Y., Colonna, M.,Braud, V. M., Allan, D. S. et al., Human inhibitory receptors Ig-liketranscript 2 (ILT2) and ILT4 compete with CD8 for MHC class I binding andbind preferentially to HLA-G. Proc. Natl. Acad. Sci. USA 2003. 100:8856–8861.
14 Morales, P. J., Pace, J. L., Platt, J. S., Phillips, T. A., Morgan, K.,Fazleabas, A. T. and Hunt, J. S., Placental cell expression of HLA-G2isoforms is limited to the invasive trophoblast phenotype. J. Immunol. 2003.171: 6215–6224.
15 Fisher, S., Genbacev, O., Maidji, F. and Pereira, L., Human cytomega-lovirus infection of placental cytotrophoblasts in vitro and in utero:implications for transmission and pathogenesis. J. Virol. 2000. 74:6808–6820.
16 Lefebvre, S., Antoine, M., Uzan, S., McMaster, M., Dausset, J., Carosella,E. D. and Paul, P., Specific activation of the non-classical class Ihistocompatibility HLA-G antigen and expression of the ILT2 inhibitoryreceptor in human breast cancer. J. Pathol. 2002. 196: 266–274.
17 Paul, P., Rouas-Freiss, N., KhalIL-Daher, I., Moreau, P., Riteau, B., Le Gal,F.A ., Avril, M. F. et al., HLA-G expression in melanoma: a way for tumorcells to escape from immunosurveillance. Proc. Natl. Acad. Sci. USA 1998.95: 4510–4515.
18 Berg, L., Riise, G. C., Cosman, D., Bergstrom, T., Olofsson, S., Karre, K.and Carbone, E., LIR-1 expression on lymphocytes, and cytomegalovirusdisease in lung-transplant recipients. Lancet 2003. 361: 1099–1101.
19 Lila, N., Amrein, C., Guillemain, R., Chevalier, P., Latremouille, C.,Fabiani, J. N., Dausset, J. et al., Human leukocyte antigen-G expressionafter heart transplantation is associated with a reduced incidence ofrejection. Circulation 2002. 105: 1949–1954.
20 LeMaoult, J., Zafaranloo, K., Le Danff, C. and Carosella, E. D., HLA-G up-regulates ILT2, ILT3, ILT4 and KIR2DL4 in antigen presenting cells, NK cellsand T cells. FASEB J. 2005. 6: 662–664.
Siyuan Liang et al. Eur. J. Immunol. 2006. 36: 2457–24712470
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

21 Dietrich, J., Cella, M. and Colonna,M., Ig-like transcript 2 (ILT2) leukocyteIg-like receptor 1 (LIR1) inhibits TCR signaling and actin cytoskeletonreorganization. J. Immunol. 2001. 166: 2514–2521.
22 Saverino, D., Fabbi, M., Ghiotto, F., Merlo, A., Bruno, S., Zarcone, D.,Tenca, C. et al., The CD85/LIR-1/ILT2 inhibitory receptor is expressed by allhuman T lymphocytes and down-regulates their functions. J. Immunol.2000. 165: 3742–3755.
23 Belkin, D., Torkar, M., Chang, C., Barten, R., Tolaini, M., Haude, A.,Allen, R. et al., Killer cell Ig-like receptor and leukocyte Ig-like receptortransgenic mice exhibit tissue- and cell-specific transgene expression. J.Immunol. 2003. 171: 3056–3063.
24 Linette, G. P., Grusby, M. J., Hedrick, S. M., Hansen, T. H., Glimcher, L. H.and Korsmeyer, S. J., Bcl-2 is upregulated at the CD4+ CD8+ stage duringpositive selection and promotes thymocyte differentiation at several controlpoints. Immunity 1994. 1: 197–205.
25 Chao, D. T. and Korsmeyer, S. J., BCL-2 family: regulators of cell death.Annu. Rev. Immunol. 1998. 16: 395–419.
26 Lantz, O., Grandjean, I., Matzinger, P. and Di Santo, J. P., Gamma chainrequired for na�ve CD4+ Tcell survival but not for antigen proliferation. Nat.Immunol. 2000. 1: 54–58.
27 Crompton, T., Outram, S. V., Buckland, J. and Owen, M. J., Distinct rolesof the interleukin-7 receptor alpha chain in fetal and adult thymocytedevelopment revealed by analysis of interleukin-7 receptor alpha-deficientmice. Eur. J. Immunol. 1998. 28: 1859–1866.
28 Palacios, E. H. and Weiss, A., Function of the Src-family kinases, Lck andFyn, in T-cell development and activation. Oncogene 2004. 23: 7990–8000.
29 Bellon, T., Kitzig, F., Sayos, J. and Lopez-Botet, M.,Mutational analysis ofimmunoreceptor tyrosine-based inhibition motifs of the Ig-like transcript 2(CD85j) leukocyte receptor. J. Immunol. 2002. 168: 3351–3359.
30 Lam, K. P., Kuhn, R. and Rajewsky, K., In vivo ablation of surfaceimmunoglobulin on mature B cells by inducible gene targeting results inrapid cell death. Cell 1997. 90: 1073–1083.
31 Kraus, M., Alimzhanov, M. B., Rajewsky, N. and Rajewsky, K., Survival ofresting mature B lymphocytes depends on BCR signaling via the Ig alpha/beta heterodimer. Cell 2004. 117: 787–800.
32 Germain, R. N., The T cell receptor for antigen: signaling and liganddiscrimination. J. Biol. Chem. 2001. 276: 35223–35226.
33 Sandner, S. E., Clarkson, M. R., Salama, A. D., Sanchez-Fueyo, A.,Domenig, C., Habicht, A., Najafian, N. et al., Role of the programmeddeath-1 pathway in regulation of alloimmune responses in vivo. J. Immunol.2005. 174: 3408–3415.
34 Latchman, Y., Wood, C. R., Chernova, T., Chaudhary, D., Borde,Chernova, M. I., Iwai, Y. et al., PD-L2 is a second ligand for PD-1 andinhibits T cell activation. Nat. Immunol. 2001. 2: 261–268.
35 Dudley, E. C., Petrie, H. T., Shah, L. M., Owen, M. J. and Hayday, A. C.,T cell receptor beta chain gene rearrangement and selection duringthymocyte development in adult mice. Immunity 1994. 1: 83–93.
36 von Boehmer, H., Aifantis, I., Feinberg, J., Lechner, O., Saint-Ruf, C.,Walter, U., Buer, J. and Azogui, O., Pleiotropic changes controlled by thepre-T-cell receptor. Curr. Opin. Immunol. 1999. 11: 135–142.
37 Michie, A. M. and Zuniga-Pflucker, J. C., Regulation of thymocytedifferentiation: pre-TCR signals and beta-selection. Semin. Immunol. 2002.14: 311–323.
38 Hamawy, M. M., Targeting proximal T cell receptor signaling intransplantation. Transplantation 2003. 75: 1921–1927.
39 Fernandez, L. A., Torrealba, J., Yagci, G., Ishido, N., Tsuchida, M., TaeKim, H., Dong, Y. et al., Piceatannol in combination with low doses ofcyclosporine A prolongs kidney allograft survival in a stringent rattransplantation model. Transplantation 2002. 74: 1609–1617.
40 Liang, S., Baibakov, B. and Horuzsko, A., HLA-G inhibits the functions ofmurine dendritic cells via the PIR-B immune inhibitory receptor. Eur. J.Immunol. 2002. 32: 2418–2426.
41 Miller, J. D., Weber, D. A., Ibegbu, C., Pohl, J., Altman, J. D. and Jensen,P. E., Analysis of HLA-E peptide-binding specificity and contact residues inbond peptide required for recognition by CD94/NKG2. J. Immunol. 2003.171: 1369–1375.
42 Warrens, A. N., Simon, A. R., Theodore, P. R., Sachs, D. H. and Sykes, M.,Cross-species compatibility of intercellular adhesion mmolecule-1 (CD54)with its ligand. Transplantation 2000. 69: 394–399.
Eur. J. Immunol. 2006. 36: 2457–2471 Immunomodulation 2471
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu




















