
Host haemocyte inactivation by an insect parasitoid: transient expression of a polydnavirus gene
10
Downloaded from www.microbiologyresearch.org by IP: 107.22.17.133 On: Sat, 13 Aug 2016 03:42:00 Journal of General Virology (1996), 77, 2653-2662. Printed in Great Britain Host haernocyte inactivation by an insect parasitoid: transient expression of a polydnavirus gene Sassan Asgari, Marianne Hellers and Otto Schmidt The University of Adelaide, Waite Campus, Department of Crop Protection, Glen Osmond, SA 5064, Australia Polydnaviruses produced by the hymenopteran endoparasitoid Cotesia rubecula are deposited together with the egg into the lepidopteran host Pieris rapae and are apparently involved in the suppression of the host's defence system. Around 6 h post-parasitization host haemocytes change their surface properties, actin cytoskeleton struc- ture and adhesion properties. Here we show that a single polydnavirus gene is expressed inside the caterpillar haemocytes in a transient fashion be- tween 4 to 8 h post-parasitization. Introduction Solitary wasps that deposit eggs into other insects and subsequently develop within the host, which is invariably killed, have adopted effective physiological devices to over- come the defence system of their habitual host. For example, most endoparasitoid wasps within the hymenopteran families carry polydnaviruses which are co-injected together with the egg into the host insect and are believed to be involved in the manipulation of host physiology. Polydnaviruses are unusual in that the production of virus particles and replication of viral DNA are not detected outside the ovary of the female wasp. Instead, polydnavirus DNA is an integrated part of the wasp genome and is transmitted chromosomally from one wasp generation to the next (Fleming, 1992; Fleming & Summers, 1986; Stoltz et al., 1986). Polydnaviruses infect host tissues (Stoltz & Vinson, 1979), and transcribe specific parts of the segmented, circular DNA molecules in virus-infected cells (Blissard et al., 1986a, b; Fleming et al., 1983 ; Stoltz et al., 1988; Theilmann & Summers, 1988). A number of cellular and physiological effects in the parasitized host are directly related to the expression of polydnavirus DNA, including hormonal (Beckage, 1993; Dover et aI., 1987, 1988) and developmental changes (Tanaka et al., 1987), suggestive of virus involvement in host ma- nipulation by the parasitoid. Host immune suppression by polydnaviruses was first demonstrated in caterpillars of Heliothis virescens with purified Author for correspondence: Otto Schmidt. Fax + 61 8 379 4095. e-mail [email protected] The GenBank/EMBL accession number of the DNA sequencereported in this paper is U55279. polydnaviruses from the ichneumonid Campoletis sonorensis (Edson et al., 1981; Theilmann & Summers, 1986). Subsequent studies in this and other systems showed that polydnavirus virions enter the haemocytes of the host insect changing cellular properties that are crucial for defence reactions against foreign objects, like the intruding parasite (Davies et a]., 1987; Guzo & Stoltz, 1987; Stoltz & Guzo, 1986; Strand, 1994; Strand & Pech, 1995a). Essential structural and functional aspects of haemocytes are affected by polydnaviruses including abolition of cell adhesion properties (Davies et al., 1987; Strand & Pech, 1995a), disruption of actin filaments (Webb & Luckhart, 1994) and induction of cell death (Strand & Pech, 1995 b). However, the identification of specific virus gene products that are involved in specific haemocyte changes is hampered by the large size of the viral genome and the complexity of viral gene expression in the host. Moreover, in some parasitoid systems the polydnavirus-mediated inactivation of the host immune response is complicated by a separate action of calyx gland proteins (Webb & Luckard, 1996) and venom gland secretions (Kitano, 1986; Tanaka, 1987) involved in the inactivation or enhancement of virus-mediated inactivation (Stoltz et al., 1988). In the braconid parasitoid Cotesia rubecula a combination of passive protection of the egg surface against immune rec- ognition (Asgari & Schmidt, •994) is combined with an active abrogation of the cell-mediated defence system of the host caterpillar Pieris rapae. Here we show that the inactivation of host haemocytes is caused by C. rubecula polydnavirus (CrV), which is expressed in host haemocytes as a single transcript in a transient fashion. Cloning and sequencing of the coding DNA provides an experimental basis for the study of virus- mediated haemocyte changes in parasitized host insects. !65! 0001-3866 © 1996 SGM
Transcript of Host haemocyte inactivation by an insect parasitoid: transient expression of a polydnavirus gene

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
Journal of General Virology (1996), 77, 2653-2662. Printed in Great Britain
Host haernocyte inactivation by an insect parasitoid: transient expression of a polydnavirus gene
Sassan Asgari, M a r i a n n e H e l l e r s a n d O t t o Schmidt
The University of Adelaide, Waite Campus, Department of Crop Protection, Glen Osmond, SA 5064, Australia
Polydnaviruses produced by the hymenopteran endoparasitoid Cotesia rubecula are deposited together with the egg into the lepidopteran host Pieris rapae and are apparently involved in the suppression of the host's defence system. Around 6 h post-parasitization host haemocytes change
their surface properties, actin cytoskeleton struc- ture and adhesion properties. Here we show that a single polydnavirus gene is expressed inside the caterpillar haemocytes in a transient fashion be- tween 4 to 8 h post-parasitization.
In t roduct ion
Solitary wasps that deposit eggs into other insects and subsequently develop within the host, which is invariably killed, have adopted effective physiological devices to over- come the defence system of their habitual host. For example, most endoparasitoid wasps within the hymenopteran families carry polydnaviruses which are co-injected together with the egg into the host insect and are believed to be involved in the manipulation of host physiology. Polydnaviruses are unusual in that the production of virus particles and replication of viral DNA are not detected outside the ovary of the female wasp. Instead, polydnavirus DNA is an integrated part of the wasp genome and is transmitted chromosomally from one wasp generation to the next (Fleming, 1992; Fleming & Summers, 1986; Stoltz et al., 1986).
Polydnaviruses infect host tissues (Stoltz & Vinson, 1979), and transcribe specific parts of the segmented, circular DNA molecules in virus-infected cells (Blissard et al., 1986a, b; Fleming et al., 1983 ; Stoltz et al., 1988; Theilmann & Summers, 1988). A number of cellular and physiological effects in the parasitized host are directly related to the expression of polydnavirus DNA, including hormonal (Beckage, 1993; Dover et aI., 1987, 1988) and developmental changes (Tanaka et al., 1987), suggestive of virus involvement in host ma- nipulation by the parasitoid.
Host immune suppression by polydnaviruses was first demonstrated in caterpillars of Heliothis virescens with purified
Author for correspondence: Otto Schmidt.
Fax + 61 8 379 4095. e-mail [email protected]
The GenBank/EMBL accession number of the DNA sequence reported in this paper is U55279.
polydnaviruses from the ichneumonid Campoletis sonorensis (Edson et al., 1981; Theilmann & Summers, 1986). Subsequent studies in this and other systems showed that polydnavirus virions enter the haemocytes of the host insect changing cellular properties that are crucial for defence reactions against foreign objects, like the intruding parasite (Davies et a]., 1987; Guzo & Stoltz, 1987; Stoltz & Guzo, 1986; Strand, 1994; Strand & Pech, 1995a). Essential structural and functional aspects of haemocytes are affected by polydnaviruses including abolition of cell adhesion properties (Davies et al., 1987; Strand & Pech, 1995a), disruption of actin filaments (Webb & Luckhart, 1994) and induction of cell death (Strand & Pech, 1995 b).
However, the identification of specific virus gene products that are involved in specific haemocyte changes is hampered by the large size of the viral genome and the complexity of viral gene expression in the host. Moreover, in some parasitoid systems the polydnavirus-mediated inactivation of the host immune response is complicated by a separate action of calyx gland proteins (Webb & Luckard, 1996) and venom gland secretions (Kitano, 1986; Tanaka, 1987) involved in the inactivation or enhancement of virus-mediated inactivation (Stoltz et al., 1988).
In the braconid parasitoid Cotesia rubecula a combination of passive protection of the egg surface against immune rec- ognition (Asgari & Schmidt, •994) is combined with an active abrogation of the cell-mediated defence system of the host caterpillar Pieris rapae. Here we show that the inactivation of host haemocytes is caused by C. rubecula polydnavirus (CrV), which is expressed in host haemocytes as a single transcript in a transient fashion. Cloning and sequencing of the coding DNA provides an experimental basis for the study of virus- mediated haemocyte changes in parasitized host insects.
!65! 0001-3866 © 1996 SGM

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
Methods • Insect cultures and parasitization. The parasitoid wasp C. rubecula was reared in the laboratory on the host caterpillar R rapae, which was grown on cabbage plants. Insects were maintained under a 14 h light-10 h dark photoperiodic regime at 25 °C. Adult wasps and butterflies were fed with 100% and 25 % honey solutions, respectively. To obtain parasitized caterpillars, fourth instars were placed in small containers along with adult female wasps. The caterpillars were collected at a series of time periods post-parasitization (p.p.) and kept frozen at --80 °C for further analysis.
• Southern blot hybridization. Polydnaviruses were extracted from dissected ovaries of female C. rubecula wasps and DNA was isolated from virus preparation as described previously (Asgari & Schmidt, 1994).
DNA was electrophoretically separated on I% agarose gels and transferred onto nylon membranes (Amersham) as described (Sambrook et al., 1989). The filters were incubated for 2 h in prehybridization solution (in 5 x SSC, 5 x Denhardt's solution, 0'5% SDS, 0'2 mg/ml herring sperm DNA). The labelled probe was added and hybridization was carried out overnight at 65 °C. The filters were washed twice in 2xSSC--O'1% SDS and 0"2 x SSC--0'1% SDS for 20min at 65 °C, respectively, and autoradiographed.
• Northern blot hybridization. Total RNA was extracted from P. rapae caterpillars or dissected fat body and haemocytes as described (Chomczynski & Sacchi, 1987).
RNA samples were electrophoretically separated under denaturing conditions on low-formaldehyde 1"2 % agarose gel as described (Ausubel et al., 1993). RNA samples were transferred to a nylon membrane for hybridization (Sambrook et al., 1989). The filter was prehybridized for 3 h (in 5 x SSC, 5 x Denhardt's solution, 0"5% SDS, 0'2 mg/ml herring sperm DNA). The labelled probe was then added and hybridization was carried out overnight at 65 °C. The membrane was washed twice in 2 x SSC-O-1% SDS for 20 min at 65 °C and twice in 0"2 x SSC-0"1% SDS, 20 min at 65 °C under continuous rotation and then auto- radiographed.
• cDNA library construction and screening. A cDNA library was constructed from P. rapae caterpillars that were parasitized 6 h previously. Total RNA was extracted and poly(A)+-containing RNA was isolated from total RNA using PolyATtract mRNA Isolation System IV (Promega, Technical Manual). DNA strands complementary to the RNA were synthesized using RiboClone cDNA Synthesis System (Promega) in two reactions. EcoRI adaptors were ligated to blunt-ended cDNA molecules followed by a kinase reaction, using RiboClone EcoRI Adaptor Ligation System (Promega). cDNA fragments were ligated to 2gtl0 arms and packaged using Packagene extracts (Promega).
About 5 x 108 recombinant phages were screened using stringent hybridization conditions (Ausubel et al., 1993). The filters were hybridized (in 5 x SSC, 5 x Denhardt's solution, 0"5% SDS, 0"2 mg/ml herring sperm DNA) overnight at 65 °C, using ~P-labelled CrV-DNA as a probe. The filters were washed twice with 2 x SSC, O" 1% SDS and twice with 0"2xSSC, 0'1% SDS, 20rain each time at 65°C, and then autoradiographed.
Positive plaques were picked with a Pasteur pipette and incubated in suspension medium (lOOmM-NaC1; 8 mM-MgSO4; 50 mM-Tris--HCl, pH 7"0; 0"01% gelatine). The eluted phages were titrated, plated and rescreened to obtain single phage lysates. Specific primers located adjacent to the EcoRI cloning sites of 2gtl0 were used to amplify the insert by PCR. Amplified DNA bands were cloned into pBluescript II KS(+) and sequenced with an Applied Biosystems 373A DNA Sequencing System.
• Labelling of DNA probes, cDNA fragments used as hybridization
probes for Southern and Northern blots were labelled with [32PldCTP using the Megaprime DNA labelling system (Amersham). CrV-DNA was cut with EcoRI, HindIII and AluI before the labelling reaction.
DNA probes for in situ hybridization to tissue sections were digoxigenin (DIG)-labelled, using a 2gtl0-specific reverse primer and a CrVI-specific primer (5' CTGAGGCACCTCGGAAGC 3') which ampli- fied a 330 bp fragment. A DIG-labelling kit (Boehringer Mannheim) was used with the components and thermocycling conditions of the PCR reaction adopted from manufacturer's protocol.
• In situ hybridization of tissue sections. Hybridization of DIG- labelled DNA to tissue sections was performed as described previously (TheopoId et al., 1995). In short, the caterpillars were cut into 8 ~am sections and mounted on Silane-prep slides (Sigma), fixed with para- formaldehyde, treated with proteinase K, and acetylated. Following prehybridization, 5 gl of a DIG-labelled probe was mixed with 50 gl prehybridization solution (4 x SSC, 50% formamide, I x Denhardt's solution, I mg/ml tRNA, 10 mM-DTT and 5% dextran sulphate), denatured at 70 °C for 5 min, cooled on ice for 5 min and applied onto each slide, covered with a coverslip and sealed with rubber cement. Hybridization was at 42 °C in a moist chamber. After extensive washing, 50 gl of phosphatase-conjugated anti-DIG antibody solution (1 : 1000 in PBS with 2% BSA, 0'3% Triton X-100) was applied onto each slide and incubated for 1 h at room temperature. To visualize the hybridized probes, 50 gl colour solution (50 gl Nitro Blue Tetrazolium, 2'5 gl 5- bromo-4-chloro-3-indolyl phosphate in I ml (100 mM-Tris-HC1, pH 9"5, i00 mM-NaC1, 100 mM-MgC12) was applied onto each slide, covered with a coverslip and incubated ovemight at room temperature. The slides were washed in Tris-EDTA (pH 8"0) for 10 rain and in double distilled water for 2 min.
• F-actin and lectin staining of haemocytes. FITC-conjugated phalloidin was used to stain F-actin in P. rapae haemocytes after attachment to glass slides (Webb & Luckhart, 1994). Helix pomatia lectin conjugated with FITC was applied onto haemocytes as described (Theopold et al., I996).
Results Haemocyte changes after polydnavirus infection
H a e m o l y m p h from parasi t ized and unparas i t ized caterpillars was analysed at different t ime-per iods p.p. N o differences were found until 6 h p.p., when the h a e m o l y m p h became less viscous and haemocy te s s tar ted to change the compos i t ion of cell surface componen t s and adhesive proper - ties. Similar results were ob ta ined after virus inject ion into caterpillars (not shown) but on ly effects from parasi t ized caterpillars are r epor ted here due to the ease in t iming and minimal effects of w o u n d i n g in parasi t ized caterpillars.
Haemocy te s f rom caterpillars that have been parasi t ized for more than 6 h agg rega t ed less f requent ly and granulocytes a t tached to the glass surface spread less readily. To test whe ther cell surface proper t ies are changed in parasi t ized haemocytes , t hey were a l lowed to at tach to a glass surface and t rea ted wi th FITC-conjugated Helix pomatia lectin, which specifically recognizes a Drosophila melanogaster mucin-like g lycoprote in , located on the haemocy te surface and invo lved in immune induct ion (Theopold et al., 1996). In haemocy tes f rom unparasi t ized caterpillars a large p ropor t ion of haemocytes , mainly granulocytes , were heavi ly labelled b y the lectin,
~_65~

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
O~
LFI
Ol
Fig.
I.
Lect
in-s
tain
ing
of p
aras
itize
d an
d un
para
sitiz
ed h
aem
ocyt
es f
rom
P. r
apae
. (a
) Ha
emoc
ytes
fro
m a
n un
para
sitiz
ed c
ater
pilla
r un
der
phas
e co
ntra
st;
(b)
hem
ocyt
es s
tain
ed w
ith F
ITC-
conj
ugat
ed H
. pom
atia
lec
tin u
nder
indi
rect
UV
light
; (c
) ha
emoc
ytes
fro
m a
cat
erpi
llar
para
sitiz
ed 2
4 h
prev
ious
ly u
nder
pha
se
cont
rast
; (d
) ha
emoc
ytes
fro
m a
par
asiti
zed
cate
rpill
ar s
tain
ed w
ith F
ITC-
conj
ugat
ed H
. pom
atia
lec
tin u
nder
UV
light
.
iiiii iiiii
ii ii
ill iii|iiii
iiiii~iiiiii
ii: ili~
il/~iiiii
ili ::::
::::::::
::::::::
:
ii iiiiiiii
iilil 'i
lililil ~
:,:.::,:
,::,:,:,
:,: :~:
:+::~:
,::+:
:+:.:+
:~:+:~
:~:
ii!i!i!i!i
!ili!iiiii!
i
iiiiiiiiiiii
iiiiiiiiii~
Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
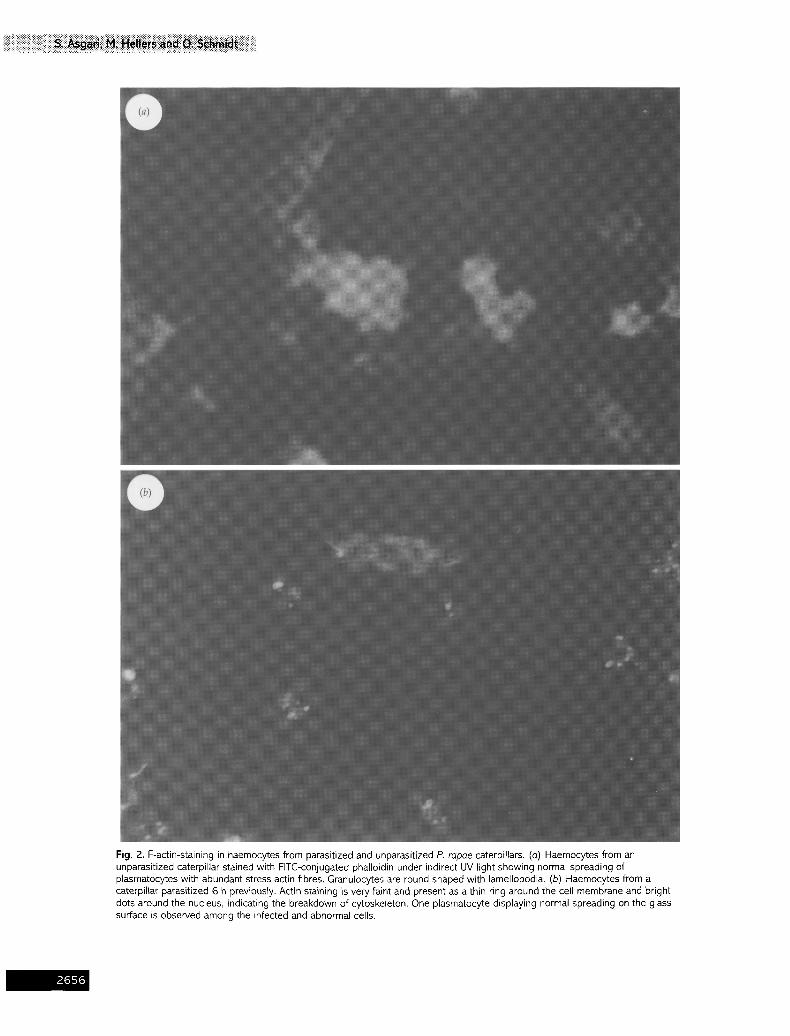
Fig. 2. F-actin-staining in haemocytes from parasitized and unparasitized P. rapae caterpillars. (a) Haemocytes from an unparasitized caterpillar stained with FITC-conjugated phalloidin under indirect UV light showing normal spreading of plasmatocytes with abundant stress actin fibres. Granulocytes are round shaped with lamellopodia. (b) Haemocytes from a caterpillar parasitized 6 h previously. Actin staining is yew faint and present as a thin ring around the cell membrane and bright dots around the nucleus, indicating the breakdown of cytoskeleton. One plasmatocyte displaying normal spreading on the glass surface is observed among the infected and abnormal cells.
-~65{

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
iiiiiiiiiiiiiiiiiiiiiii i i iiii iii ii iiiiiiiiiiiiiiiiii ii!i!iiii i i i i i i i iiiii i iii ii
(a) (b)
2 4 6 8 12 24 48 C C 2 4 6 8 12 24 48
: : . : ' i : : . . . .
1.4 kb
Fig. 3. Hybridization to Northern blots containing total RNA isolated from P. ropae caterpillars at 2, 4, 6, 8, 12, 24 and 48 h after parasitization by C rubecula. RNA from unparasitized caterpillars was used as a control (lanes C). Each RNA fraction was spectroscopically measured and equal aliquots of RNA (2 p,g) were applied to each slot, The RNA was electrophoretically separated under denaturing conditions and transferred onto a filter. (a) The filter was probed with 32p-labelled C rubecula viral DNA comprising the complete polydnavirus genome. (b) The filter was probed with 32p-labelled CrVl-cDNA fragment.
1 2 3 4 5 6
1.4 kb
1.1 kb
Fig. 4. Hybridization to a Northern blot containing (lane 1 ) mRNA isolated from 6 h parasitized P. rapae caterpillars (2 ~tg per lane), (lane 2) mRNA isolated from unparasitized caterpillars, (lane 3) total RNA from the fat body of 6 h parasitized caterpillars, (lane 4) total RNA from the fat body of unparasitized caterpillars, (lane 5) total RNA isolated from haemocytes of 6 h parasitized caterpillars and (lane 6) total RNA isolated from haemocytes of unparasitized caterpillars. The filter was probed with 32p_ labelled total virus genomic DNA and exposed to X-ray film for 30 min for haemocytes and 5 h for the fat body. CrV1 was the major virus gene expressed in the fat body and haemocytes. Minor expression of CrV2 was detected in the fat body but not in haemocytes even under overexposed conditions.
whereas in haemocytes from parasitized caterpillars changes in lectin-labelling became visible 6 h p.p. when the lectin-label on the haemocyte surface starts to decrease and after 24 h it had completely disappeared (Fig. 1). This is probably due to the failure of granulocytes from parasitized caterpillars to attach to
the glass surface, reducing the number of granulocytes present. However, under conditions where haemocytes could be inspected without the need to attach to a glass surface, the lectin-label has nevertheless disappeared from the cell surface of the granulocytes. Using Hoechst staining, on haemocytes from parasitized caterpillars, no evidence for apoptosis was found, in contrast to the observations in a related braconid system (Strand & Pech, 1995 b).
Since it is known that mucin-like components are con- tinuously internalized and recycled to the surface (Theopold et al., 1996), we investigated whether haemocytes from parasitized caterpillars display structural alterations of actin filaments. Using FITC-conjugated phalloidin as an indicator of F-actin (Webb & Luckhart, 1994), changes in the cytoplasmic actin filaments were detected in haemocytes around 6 h p.p. (Fig. 2). The changes in actin filaments are comparable to those seen in H. virescens haemocytes caused by Campoletis sonorensis parasitization (Webb & Luckhart, 1994). Since the effects in H. virescens are already visible 30 min p.p., they are probably not caused by C. sonorensis polydnavirus expression in host haemocytes, but are due to components in the ovarian fluid that are co-injected into the host together with the egg and the virus (Webb & Luckhart, 1994). In contrast, the effects in parasitized P. rapae haemocytes are not visible within the first 4 h p.p. and coincide with the detection of the polydnavirus transcript, suggesting that the effects in this system are probably caused by virus gene expression.
Temporal expression of CrV genes in the host
Total RNA extracted from P. rapae caterpillars at various times p.p. was analysed on Northern blots using radioactively labelled C. rubecula viral DNA (CrV-DNA), comprising the complete virus genome, as a hybridization probe. Hybridization results suggest that the virus RNA is expressed
165~

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
Fig. 5. Expression of polydnavirus transcript in tissue sections of parasitized P. rapae caterpillars using DIG-labelled CrVl -cDNA as a probe. After in situ hybridization the localization of labelled probe was visualized using phosphatase-conjugated antibodies as prescribed in Methods. Examples of stained cells are indicated by arrowheads.
in host tissues within a distinct time-period, starting from 4 h p.p. to 8 h p.p. (Fig. 3 a). Two major RNAs were detected in caterpillars and the peak of gene expression in both RNAs was observed 6 h p.p. RNA from unparasitized caterpillars did not cross-hybridize to the probe. To establish whether the expressed RNA constitutes protein-coding RNA, a poly(A) +- RNA fraction from caterpillars was tested on a Northern blot (Fig. 4, lane I). The hybridization experiment suggests that the virus DNA hybridized to two transcripts with an estimated size of 1-4 kb (CrVI) and 1"i kb (CrV2).
To establish whether there is tissue-specific expression of virus DNA in the host caterpillar, RNA from the fat body and haemocytes was tested on Northern blots using total CrV- DNA as a probe. Under these conditions only a single transcript (CrV1) was detected in haemocytes (Fig. 4, lane 5), even under overexposed conditions, whereas in the fat body both CrVI and CrV2 were detected, similar to the signals in total mRNA (Fig. 4, lane 3). The two transcripts CrVI and CrV2 are probably encoded by unrelated virus genes (see below).
The transcript of the polydnavirus gene CrV1 was located to small patches within the fat body of parasitized caterpillars by in situ hybridization to tissue sections (Fig. 5). The transcripts were detectable between 4 and 8 h p.p. No attempts were made to establish possible structural and functional changes in infected fat body cells from parasitized caterpillars.
Molecular cloning of the CrV1 gene
To isolate cDNA fragments from virus encoded mRNA, a
figtl0 cDNA library was constructed from P. rapae caterpillars that were parasitized 6 h previously. The library was screened using radioactively labelled CrV-DNA as a probe. Positive recombinant phages were analysed by PCR, using 2gtl0 specific primers, and a 1"4 kb clone was chosen for subcloning into a plasmid vector. To establish the identity of the cDNA fragment, the insert was used as a probe to test a Northern blot of total RNA from parasitized and unparasitized caterpillars (Fig. 3 b). The cDNA fragment hybridized to the CrVI band, indicative that the cloned cDNA fragment represented the expressed CrV transcript. The hybridization pattern to cater- pillar RNA from various time-periods p.p. coincided with the CrV1 band labelled with total CrV-DNA as a probe (Fig. 3 a), and a similar short time-window of expression with virtually no transcripts visible except for the period 4-8 h p.p.
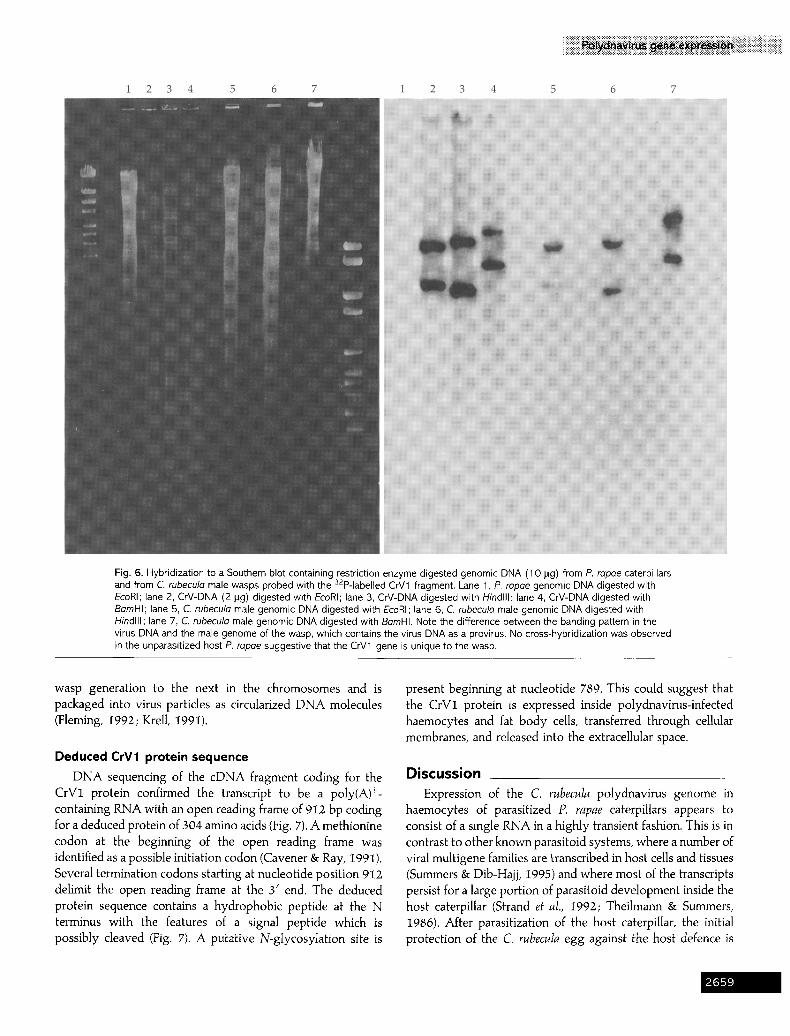
To verify the virus origin of the CrV1 gene, a Southern blot containing genomic DNA from male wasps and CrV-DNA from isolated virus particles was tested using the cDNA fragment as a probe (Fig. 6). No cross-hybridization to P. rapae genomic DNA was detectable, indicating that the cDNA is wasp-specific (Fig. 6, lane 1). When the pattern of hybridization between wasp genomic DNA and virus DNA was compared, a difference in one of the two BamHI restriction fragments was observed. Whereas EcoRI and HindIII digests are not different for this particular gene, one of the BamHI restriction fragments is larger in the wasp genome. This supports the notion that the CrV1 gene is present as a circular DNA and is integrated into the wasp genome. As in other braconid and ichneumonid parasitoids, the polydnavirus DNA is transmitted from one
_~65~
Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
iiiiiiiiiiiiiiiiiiiii i i iiii ii i iiiiiiiiiiiiiiiiiiiiiiiiiii i iiiiiiiiiiiiiiiiiiiiiiiiiiii
1 2 3 4 5 6 1 2 3 4 5 6 7
Fig. 6. Hybridization to a Southern blot containing restriction enzyme digested genomic DNA (1 0 pg) from P. rapae caterpillars and from C. rubecule male wasps probed with the 32p-labelled CrV1 fragment. Lane 1, P. repoe genomic DNA digested with EcoRI; lane 2, CrV-DNA (2 lag) digested with EcoRI; lane 3, CrV-DNA digested with Hindlll; lane 4, CrV-DNA digested with BamHI; lane 5, C. rubecula male genomic DNA digested with EcoRI; lane 6, C. rubeculo male genomic DNA digested with Hindlll; lane 7, C. rubecula male genomic DNA digested with BemHI. Note the difference between the banding pattern in the virus DNA and the male genome of the wasp, which contains the virus DNA as a provirus. No cross-hybridization was observed in the unparasitized host P. rapae suggestive that the CrV1 gene is unique to the wasp.
wasp generation to the next in the chromosomes and is packaged into virus particles as circularized DNA molecules (Fleming, 1992; Krell, 1991).
Deduced CrV1 protein sequence
DNA sequencing of the cDNA fragment coding for the CrV1 protein confirmed the transcript to be a poly(A) +- containing RNA with an open reading flame of 912 bp coding for a deduced protein of 304 amino acids (Fig. 7). A methionine codon at the beginning of the open reading frame was identified as a possible initiation codon (Cavener & Ray, 1991). Several termination codons starting at nucleotide position 912 delimit the open reading frame at the 3' end. The deduced protein sequence contains a hydrophobic peptide at the N terminus with the features of a signal peptide which is possibly cleaved (Fig. 7). A putative N-glycosylation site is
present beginning at nucleotide 789. This could suggest that the CrV1 protein is expressed inside polydnavirus-infected haemocytes and fat body cells, transferred through cellular membranes, and released into the extracellular space.
Discussion Expression of the C. rubecula po lydnav i rus genome in
haemocytes of parasitized P. rapae caterpillars appears to consist of a single RNA in a highly transient fashion. This is in contrast to other known parasitoid systems, where a number of viral multigene families are transcribed in host cells and tissues (Summers & Dib-Hajj, 1995) and where most of the transcripts persist for a large portion of parasitoid development inside the host caterpillar (Strand et al., 1992; Theilmann & Summers, 1986). After parasitization of the host caterpillar, the initial protection of the C. rubecuIa egg against the host defence is
'.65!

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
M s c v z Ip . ~ v L t. S A L L S O S C T O ^ Y Pt A T G T C A C r C G T E A A A AGTUCGTL--rGTGCTGCTC TCA OCT C I ~ C ] ~ A G T C A A T C A T G ' r A C G CAAGCI 'TATCCA
9 18 27 36 45 54 63 72
S E Y Q W E R I Q N I P P A D V A S L S P L T D TCC GAA TAC CAA TGG GAA AGA AI"Y CAA AAT ATY Cr.A T r r GCr GAc GTr G c r TCC c-'rTAGTCU-X]CTGACCGAT
81 90 99 1(]6 117 126 135 144
Y F S P K S D S T A E R L A P S T N D F D E S D TAT "FrCTL~CCA AAA TCA GATTCT ACA GOU GAAAGACTTGCG CCATCA ACA AAC GACTTC GATGAA TCT GAT
153 162 171 180 189 198 207 216
M N N R E F L M I P K S Y S E A P R K H F D V E ATG AAC/,,AT AGA GAA T r T T T G ATG ATY CCG AAG'ru-'Y TATTCT GAG GCA CCT CGG AAG C A T T Y r GAT GTT GAA
225 234 243 252 261 270 279 288
Y F L K H H P tl I K P G H S T R H R P A F E I R TAT Trl" c r r AAG CAT CAC CCT CAT ArC AAG C C r GGA CAC AGT ACA AGA CAT AGA CCT GCG T I C GAA ATA AGA
297 306 315 324 333 342 351 360
P F V G P I S E L S S G $ S G I O Q L O S S V A cc'r TYC GTA GGA CC'T ATY AGT GAA TTA TCC AGT GGA AGT AGC GGT ATC GG'T CAA CTr GGA TCG A~ ~ ~
369 378 387 396 403 414 423 432
D E I F S G L K N F i G N V K F T Q P E L F D T GAT GAA ATA ' r r T TCC GGA CTr AAA A A T ' r r c A T r GGA AAT GTG AAA T r r A C t C.AA CCT GAA TTA T r r GAC ACA
441 450 459 568 477 486 495 504
S K T H Q Y E Y E S E T S E K R H I T R E D L L TCC AAA ACC CAC CAG TAT GAA TAT GAA T c r GAA ACA TCW GAA AAA CGC CAC Al"r ACC CU-'T GAA GAT 1'TG c r r
513 522 531 .540 549 55g 56"7 5"76
S E L H A I K E A L Q N L K S A V I R V E N E l TCT GAA CTA CAT GCA ATA AAA GAG GCA c T r CAA AAT C'rA AAA TCT Gc ' r GTC A T r CGG GTA GAA AAT GAG ATC
585 594 603 612 62t 630 639 648
S F N K E G V T Y P S V A I S P Q P L V L 1 N P A G C ' I T r A A C A A A G A A G G A G T r A C T T A C C C C T C T G T G G C A A T C T C A O L ' G C A A C c r c ' r A G ' I ' r C r G A T r AATCCT
657 666 675 684 693 702 711 720
N G A S E Q G P I E A I T E E A V T H P T V T N AAC GGA GCA 'I3L'T GAA CAG GGA ~ ATA GAA GU'W ATT ACC GAA GAA GCA G ' r r AG'r CAT COU ACT GTG ACA AAC
729 738 747 756 765 774 783 792
S S Q P P V L I N P N G A S E Q G P I E A N N E TCA TCG CAA CCT CCA GTr CTG AT r AAT C~-'r AAC GGA GCA TCr GAA CAG GGA CCC ATA G ~ ~ ~ T ~ C G ~
801 810 819 828 837 I;46 855 864
E G V T Y P S D T T P S Q T F F G A A G G A G T r A C ' r T A T C C C A G T G A T A C A A C C C C A T C G C A A A C r T T I ' T r r T G A TCC CTA T A T G T A G C A T C T G A A
873 882 891 900 909 918 927 936
C'Tr GAA C'TC AGT AGA GTC TAC ' r r A AGA AAG TAT GTA TAA AA.A TAC TGC TAC AAA TGA AGC CAT ATG ATA A r c 945 954 963 972 981 990 999 10~
GAC COU 'rAG AGA ACA CAC TAT CTA AAA (Tl"r A'l 'r TCC CAT AAT "ITF "ITr AGG AGA AAG AAC AAG CGT TC'I" TAA 1017 1026 1035 1044 1053 1062 1071 1080
AGT ACT GAA CAT ATC GAA "lTr CI'I" GTC ATC G r c AAA TAT GTA TAT AAT AAC "F1A GAA AAC CTT GCA ATC TAA 1089 1098 1107 1116 1125 1134 1143 1152
ATG T I C TAG CTI- TAA GTA AGT'ITI" GAT CGA AGA AAA ACG CCA A T r T I T ATA GTG TAA ACA G'lr'r TTG TAT T I T t 16I 1170 I 179 1188 1197 1206 1215 1224
GAT GAA TAA AT(} AAT ATA Tl~ TCC AAT "ITA ~ AAA AAA AAA AAA AAA AAA AAA AAA AAA AA
1233 1242 1251 1260
Fig. 7. DNA nucleotide sequence and deduced amino acid sequence of the eDNA fragment in pCrV1. The hydrophobic region at the N terminus, representing a putative signal peptide is boxed. A computer predicted proteolytic cleavage site is indicated by an arrowhead. A putative N-glycosylation site is underlined.
achieved by a passive coating of the egg surface (Asgari & published results). The virus-related haemocyte changes in Schmidt, 1994). The haemocyte changes observed 4 h p.p. are parasitized caterpillars are only temporary and disappear after probably caused exclusively by CrVs, since haemocytes are 2-3 d p.p. (S. Asgari, unpublished results). At this time the not visibly altered by virus-free calyx fluid (S. Asgari, un- emerged wasp larva appears to be protected against the
!66(

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
iiiiiiiiiii ii iiii iii i iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii
recovered host defence by an unknown mechanism. This could indicate that the initial protective strategy of C. rubecula eggs and early larvae relies on two types of molecules, one providing protection against host immune recognition (Asgari & Schmidt, 1994), the other altering immune competence of host haemocytes in a transient fashion. Given that host haemocytes are probably changed by a short pulse of CrV1 gene activity, the C. rubecula endoparasitoid may constitute an ideal experimental system to study the transformation of haemocyte properties in the parasitized host.
The disappearance of cell surface glycoproteins recognized by the H. pomatia lectin may constitute a cellular indicator for changes in the adhesive properties, since other lectin-binding properties, like wheat-germ agglutinin did not change after parasitization. In Drosophila, the H. pomatia Iectin specifically binds to a mucin-like glycoprotein which is involved in immune induction of haemocytes (Theopold et al., 1996). In P. rapae haemocytes, the H. pomatia lectin binds predominantly to granulocytes, which are known in lepidopterans to initiate the immune response (Ratcliffe et al., 1993). In granulocytes from parasitized caterpillars the lectin-labelling changes between 4 and 8 h p.p. and has completely disappeared by 24 h p.p. The disappearance of lectin-labelling is probably due to the reduction in the amount of glycoprotein on granulocytes caused by an unknown mechanism, and coincides with changes in adhesive properties of the cells. Polydnavirus infected haemocytes do not attach to each other or to bacteria and no phagocytic activity is discernible in these haemocytes. Thus, the observed changes in the corresponding haemocyte types are probably due to the reduction in adhesive properties, as indicated by altered attachment and spreading behaviour of granulocytes from parasitized caterpillars on a glass surface (Davies ef aI., 1987). Other haemocytes, like plasmatocytes, lack a mucin-like glycoprotein, but it is possible that surface properties of all haemocytes from parasitized caterpillars are changed, resulting in a dramatic shift of adhesive properties. Both granulocytes and plasmatocytes from parasitized cater- pillars differ from non-parasitized haemocytes in the cyto- plasmic structure of actin filaments, suggestive of a disruption of cell surface and cytoplasmic connections.
How are polydnaviruses able to change haemocyte proper- ties? Two possible modes of virus action are possible: virus coat or associated proteins might interact directly with the haemocyte, or the virus DNA is transcribed inside the host cell exerting cellular changes through viral gene expression. We suspect that the virus protein pCrV1, coded by the C. rubecula polydnavirus DNA, is responsible for the observed haemocyte changes, although an (additional) involvement of particle proteins is difficult to rule out at this stage. Although two polydnavirus-specific transcripts are produced in P. rapae fat body tissues, only a transcript of 1"4 kb in size is detected in haemocytes. Moreover, the timing of cellular changes and the production of the polydnavirus transcript inside haemocytes coincide between 4-8 h p.p. If the observed haemocyte
changes are caused by the interaction of particle proteins with infected haemocytes some of the changes should be detectable before a 4 h period p.p. Infection of haemocytes with UV- irradiated viruses will clarify the mode of CrV action.
Previous results suggest that C. rubecula eggs are passively protected against the host's immune recognition by a calyx fluid protein, which is found on the surface of eggs and polydnaviruses (Asgari & Schmidt, I994) precluding immune recognition and inactivation of the egg immediately after parasitization. Haemocyte inactivation, starting 4 h p.p., may then provide protection for the emerging parasitoid larva at a later stage of wasp development inside the caterpillar. This is obviously different to the protection strategy observed in the ichneumonid C. sonorensis, where a passive protection of wasp eggs has been ruled out (Edson et at., I98I). Instead the wasp egg appears to be protected by the po]ydnavirus or virus- related proteins (Theilmann & Summers, I986; Webb & Luckhart, 1996). Early haemocyte alterations in the host H. virescens that precede virus expression appear to be caused by ovarian fluid proteins that are similar to C. sonorensis virus particle proteins and are co-injected together with the eggs into the caterpillar (Webb & Luckhart, I994). It will be interesting to uncover the differences and similarities of the two systems when the corresponding genes in C. sonorensis have been cloned.
At this stage we can only speculate on the possible function of the C. rubecula polydnavirus protein coded by the CrV1 gene. The presence of a signal peptide suggests that the protein is secreted through cellular membranes and probably proteolytically cleaved to be secreted. However, the mode of interaction with cellular proteins and the identity of possible cellular receptors for the CrV1 protein are not known at this stage.
It is of interest to note the steep decline of polydnavirus gene expression in haemocytes around 6 h p.p. The quick shut- off in gene expression is apparently not affected by the initial quantity of polydnavirus or subsequent addition of virus through super-parasitization, suggesting that the decrease in virus gene expression is not caused by a depletion of virus DNA template. The observation that all infected cells dis- continue transcription at the same time suggests that regu- latory events initiated by polydnavirus gene expression cause a dramatic change in differentiation status which may lead to the temporary inactivation of these cells.
We would like to thank Dr Uli Theopold for help and advice during the experiments and Dr Neil Shirley and Dr Jing Li for help with the sequencing analysis. This study was supported by a scholarship from the Ministry of Culture and Higher Education of Iran to S. Asgari and an ARC grant to O. Schmidt.
References Asgari, S. & Schmidt, O. (1994). Passive protection of eggs from the parasitoid, Cotesia rubecula, in the host Pieris rapae. Journal of Insect Physiology 40, 789-795.
!66

Downloaded from www.microbiologyresearch.org by
IP: 107.22.17.133
On: Sat, 13 Aug 2016 03:42:00
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A. & Struhl, K. (1993). Current Protocols in Molecular Biology. New York: John Wiley.
Beckage, N. E. {1993). Endocrine and neuroendocrine host-parasite relationships. Receptor 3, 233-245.
Blissard, G.W., Fleming, J. G. W., Vinson, S. B. & Summers, M.D. (1986 a). Campoletis sonorensis virus : expression in Heliothis virescens and identification of expressed sequences. Journal of Insect Physiology 32, 351-359.
Blissard, G. W., Vinson, S. B. & Summers, M. D. (1986 b). Identification, mapping, and in vitro translation of Campoletis sonorensis virus mRNAs from parasitized Heliothis v irescens larvae, Journal of Virology 57, 318-327.
Cavener, R. C. & Ray, S. C. (1991 ). Eukaryotic start and stop translation sites. Nucleic Acids Research 19, 3185-3192.
Chomczynski, P. & Sacchi, N. (1987). Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol~hloroforrn extrac- tion. Analytical Biochemistry 162, 156--159.
Davies, D.H., Strand, M.R. & Vinson, S.B. (1987). Changes in differential haemocyte count and in vitro behaviour of plasmatocytes from host Heliothis virescens caused by Campoletis sonorensis polydnavirus. Journal of Insect Physiology 33, 143-153.
Dover, B. A., Davies, D. H., Strand, M. R., Gray, R. S., Keeley, L. L. & Vinson, S. B. (1987). Ecdysteroid-titre reduction and developmental arrest of last instar Heliothis virescens larvae by calyx fluid from the parasitoid Campoletis sonorensis. Journal of Insect Physiology 33, 333-338.
Dover, B. A., Davies, D. N. & Vinson, S. B. (1988). Dose-dependent influence of Campoletis sonorensis polydnavirus on the development and ecdysteroid titers of last instar Heliothis virescens larvae. Archives of Insect Biochemistry and Physiology 8, 113-126.
Edson, K. M., Vinson, S. B., Stoltz, D. B. & Summers, M. D. (1981). Virus in a parasitoid wasp: suppression of the cellular immune response in the parasitoid's host. Science 211, 582-583.
Fleming, J,-A. (1992). Polydnaviruses: mutualists and pathogens. Annual Review of Entomology 37, 401-423.
Fleming, J. G. & Summers, M. D. {1986). Campoletis sonorensis endo- parasitic wasps contain forms of C. sonorensis virus DNA suggestive of integrated and extrachromosomal polydnavirus DNAs. Journal of Virology 57, 552-562.
Fleming, J. G. W., Blissard, G.W., Summers, M.D. & Vinson, S.B. (1983). Expression of Campoletis sonorensis virus in the parasitized host, Hdiothis virescens. Journal of Virology 48, 74-78.
Guze, D. & Stoltz, D. B. (1987). Observation on cellular immunity and parasitism in the tussock moth. Journal of Insect Physiology 33, 19-31.
Kitano, H. (t986). The role of Apanteles glomeratus venom in the defensive response of its host, Pieris rapae crucivora. Journal of Insect Physiology 32, 369-370.
Krell, P. J. (1991 ). The polydnaviruses: multipartite DNA viruses from parasitic Hymenoptera. In Atlas of Invertebrate Viruses, pp. 141-177. Edited by J. Adams & J. R. Bonami. Boca Raton, Fla. : CRC Press.
Ratcliffe, N. A. (1993). Cellular defense responses of insects: unresolved problems. In Parasites and Pathogens of Insects, pp. 267-304. Edited by N. E. Beckage, S. N Thompson & B. A. Federici. San Diego: Academic Press.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual, 2nd edn. New York: Cold Spring Harbor Laboratory.
Stoltz, D. B. & Guzo, D. (1986). Apparent haemocytic transformation associated with parasitoid-induced inhibition of immunity in Malacosoma disstria larvae. Journal of Insect Physiology 32, 377-388.
Stoltz, D. B. & Vinson, S. B. (1979). Viruses and parasitism in insects. Advances in Virus Research 24, 125-171.
Stoltz, D. B., Guzo, D. & Cook, D. (1986). Studies on polydnavirus transmission. Virology 155, 120--131.
Stoltz, D. B., Guzo, D., Belland, E. R., Lucarotti, C. J. & MacKinnon, E. A. (1988). Venom promotes uncoating in vitro and persistence in vivo of DNA from a braconid polydnavirus. Journal of General Virology 69, 903-907.
Strand, M.R. (1994). Microplitis demolitor polydnavirus infects and expresses in specific morphotypes of Pseudoplusia includens haemocytes. Journal of General Virology 75, 3007-3020.
Strand, H. R. & Pech, L. L. (19950). Immunological basis for com- patibility in parasitoid-host relationships. Annual Review of Entomology 40, 31-56.
Strand, M. R. & Pech, L. L. (1995 b). Microplitis demolitor polydnavirus induces apoptosis of a specific morphotype in Pseudoplusia includens. Journal of General Virology 76, 283-291.
Strand, fvl. R., McKenzie, D. i., Grassl, V., Dover, B. A. & Aiken, J. M. (1992). Persistence and expression of Microplitis demolitor polydnavirus in Pseudoplusia includens. Journal of General Virology 73, 1627-1635.
Summers, M.D. & Dib-Haii, S.D. (1995). Po[ydnavirus-facilitated endoparasite protection against host immune defenses. Proceedings of National Academy of Sciences, USA 92, 29-36,
Tanaka, T. (1987). Calyx and venom fluids of Apante]es kariyai (Hymenoptera: Braconidae) as factors that prolong larval period of the host, Pseudaletia separata (Lepidoptera: Noctuidae). Annals of the Ento- mological Society of America 80, 530--533.
Tanaka, T., Agui, N. & Hiruma, K. {1987). The parasitoid Apanteles kariyai inhibits pupation of its host, Pseudaletia separata, via disruption of prothoracicotropic hormone release. General and Comparative Endo- crinology 67, 364-374.
Theilmann, D.A. & Summers, M.D. (1986). Molecular analysis of Campoletis sonorensis virus DNA in the lepidopteran host Heliothis virescens. Journal of General Virology 67, 1061-1969.
Theilmann, D.A. & Summers, M. D. (1988). Identification and com- parison of Campoletis sonorensis virus transcripts expressed from four genomic segments in the insect hosts Campoletis sonorensis and Heliothis virescens. Virology 167, 329-341.
Theopold, U., Pinter, M., Daffre, S., Tryselius, Y., Friedrich, P., N~ssel, D. R. & Hultmark, D. (1995). CalpA, a Drosophila calpain homolog specifically expressed in a small set of nerve, midgut and blood cells. Molecular Cell Biology 15, 824-834.
Theopold, U., Samakovlis, C., Tempst, P., Dillon, N., Axelsson, B., Schmidt, O. & Hultmark, D. (1996). A Drosophila mucin involved in immune induction. Journal of Biological Chemistry 271, 12708-12715.
Webb, B.A. & Luckhart, S. {1994). Evidence for an early immune- suppressive role for related Campoletis sonorensis venom and ovarian proteins in Heliothis virescens. Archives of Insect Biochemistry and Physiology 26, 147-163.
Webb, B. A. & Luckhart, S. (1996). Factors mediating short- and long- term immune suppression in a parasitoid insect. Journal of Insecl: Physiology 42, 33-40,
Received 10 January 1996; Accepted 31 May 1996
!66:"




















