Hormonal regulation of behavioural development in the honey bee is based on changes in the rate of...
9
HORMONAL REGULATION OF BEHAVIOURAL DEVELOPMENT IN THE HONEY BEE IS BASED ON CHANGES IN THE RATE OF JUVENILE HORMONE BIOSYNTHESIS ZHI-YONG HUANG,’ GENE E. ROBINSON,’ STEPHEN S. Toes~ KOICHIRO J. YAOI, COLETTE STRAMBI,’ ALAIN STRAMBI~ and BARBARA STAY’ ‘Department of Entomology, University of Illinois, Urbana, IL 61801, U.S.A., ‘Department of Zoology, University of Toronto, Toronto, Ontario, Canada MSS Ml, ‘Centre National Recherche Scien$%pc, Laboratoire de Neurabiologie, B.P. 71, 13402 Marseille C&x 9, France and 4Department af Biology, University of lowa, Iowa City, 1.4 52242, U.S.A. (Received *5 April ,991; reuised 13 June 1991) Abstract-In the adult worker honey bee (Apis meNifera L.), increases in the haemolymph titre ofjuvenile hormone underlie behavioural development, from nest duties to foraging. However, the physiological basis of juvenile hormone titre regulation was unknown. Using a radiochemi- cal assay for juvenile hormone biosynthesis in vifro, we demonstrate that differences in juvenile hormone titres among bees performing different age-dependent tasks are a consequence of changes in rates of hormone synthesis by the corpora allata. Rates of juvenile hormone biosynthesis were low in newly emerged bees, 7-9.day-old nurse bees, and 14-15-day-old bees collected from the nest periphery, and high in foragers. Rates of biosynthesis were highly correlated with haemolymph titres of juvenile hormone measured in the same individuals. Corpora allata contained mostly methyl farnesoate, the immediate precursor of juvenile hormone, and released principally juvenile hormone III into the incubation medium, indicating no appreciable hormone storage. We also report similarities and differences in parameters of juvenile hormone biosynthesis between nurse bees and foragers that were found during the course of a detailed characterization of the radiochemical assay for adult worker honey bees. These results, coupled with the fact that it is possible to measure rates of juvenile hormone biosynthesis from individual bees, indicate that the radiochemical assay will be useful in further studies of hormonal regulation of bee behaviour. Key Word Index: Apis mellifera; corpora allata; division of labour; honey bee: juvenile hormone; social insects Division of labour in honey bee colonies is based, in part, on worker age. Bees change jobs as they age, a form of behavioural development called “age polyethism” (reviewed by Winston, 1987). Juvenile hommne is involved in the control of age polyethism in adult worker honey bees (Jaycox ef al., 1974; Jaycox, 1976; Robinson, 1985, 1987% b; reviewed by Robinson, in press). Haemolymph titres of juvenile hormone increase as the bee ages (Fluri ef al., 1982; Robinson ef al., 1987). Low titres are associated with the performance of tasks in the nest such as brood care during the first 1-3 weeks of the bee’s 4-7-week adult life, whereas a higher titre at about 3 weeks of age is associated with foraging. Treatment with juvenile hormone (Jaycox, 1976), juvenile hormone mimic (Jaycox ef al., 1974), or juvenile hormone aualogue (Robinson, 1985, 1987a; Sasagawa et al., 1985) induces precocious foraging. The division of labour in honey bee colonies is also flexible, because workers can respond to changing colony needs by accelerating, retarding, or reversing their behavioural development (e.g. RBsch, 1930; Milojevic, 1940; Robinson et al., unpublished; reviewed by Robinson, in press), Flexibility in age- based division of labour in honey bee colonies is also regulated by juvenile hormone, because the titre of this hormone is modulated by changes in environmental conditions (Robinson et al., 1989, unpublished), These findings are consistent with the well-supported belief that the corpora allata, the glands that produce juvenile hormone, occupy a key position in the neuroendocrine response of insects to environmental factors (e.g. De Wilde et al., 1959; Tobe and Stay, 1985). It is not known how the juvenile hormone titre is regulated in adult honey bees, and this information is important for two reasons. First, the relationship 133
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Hormonal regulation of behavioural development in the honey bee is based on changes in the rate of...

HORMONAL REGULATION OF BEHAVIOURALDEVELOPMENT IN THE HONEY BEE IS BASED
ON CHANGES IN THE RATE OF JUVENILEHORMONE BIOSYNTHESIS
ZHI-YONG HUANG,’ GENE E. ROBINSON,’ STEPHEN S. Toes~ KOICHIRO J. YAOI,COLETTE STRAMBI,’ ALAIN STRAMBI~ and BARBARA STAY’
‘Department of Entomology, University of Illinois, Urbana, IL 61801, U.S.A., ‘Department of Zoology,University of Toronto, Toronto, Ontario, Canada MSS Ml, ‘Centre National Recherche Scien$%pc,Laboratoire de Neurabiologie, B.P. 71, 13402 Marseille C&x 9, France and 4Department af Biology,
University of lowa, Iowa City, 1.4 52242, U.S.A.
(Received *5 April ,991; reuised 13 June 1991)
Abstract-In the adult worker honey bee (Apis meNifera L.), increases in the haemolymph titreofjuvenile hormone underlie behavioural development, from nest duties to foraging. However,the physiological basis of juvenile hormone titre regulation was unknown. Using a radiochemi-cal assay for juvenile hormone biosynthesis in vifro, we demonstrate that differences in juvenilehormone titres among bees performing different age-dependent tasks are a consequence ofchanges in rates of hormone synthesis by the corpora allata. Rates of juvenile hormonebiosynthesis were low in newly emerged bees, 7-9.day-old nurse bees, and 14-15-day-old beescollected from the nest periphery, and high in foragers. Rates of biosynthesis were highlycorrelated with haemolymph titres of juvenile hormone measured in the same individuals.Corpora allata contained mostly methyl farnesoate, the immediate precursor of juvenilehormone, and released principally juvenile hormone III into the incubation medium, indicatingno appreciable hormone storage. We also report similarities and differences in parameters ofjuvenile hormone biosynthesis between nurse bees and foragers that were found during thecourse of a detailed characterization of the radiochemical assay for adult worker honey bees.These results, coupled with the fact that it is possible to measure rates of juvenile hormonebiosynthesis from individual bees, indicate that the radiochemical assay will be useful in furtherstudies of hormonal regulation of bee behaviour.
Key Word Index: Apis mellifera; corpora allata; division of labour; honey bee: juvenilehormone; social insects
Division of labour in honey bee colonies is based,in part, on worker age. Bees change jobs as theyage, a form of behavioural development called “agepolyethism” (reviewed by Winston, 1987). Juvenilehommne is involved in the control of age polyethismin adult worker honey bees (Jaycox ef al., 1974;Jaycox, 1976; Robinson, 1985, 1987% b; reviewed byRobinson, in press). Haemolymph titres of juvenilehormone increase as the bee ages (Fluri ef al., 1982;Robinson ef al., 1987). Low titres are associated withthe performance of tasks in the nest such as broodcare during the first 1-3 weeks of the bee’s 4-7-weekadult life, whereas a higher titre at about 3 weeksof age is associated with foraging. Treatment withjuvenile hormone (Jaycox, 1976), juvenile hormonemimic (Jaycox ef al., 1974), or juvenile hormoneaualogue (Robinson, 1985, 1987a; Sasagawa et al.,1985) induces precocious foraging.
The division of labour in honey bee coloniesis also flexible, because workers can respond tochanging colony needs by accelerating, retarding, orreversing their behavioural development (e.g. RBsch,1930; Milojevic, 1940; Robinson et al., unpublished;reviewed by Robinson, in press), Flexibility in age-based division of labour in honey bee colonies isalso regulated by juvenile hormone, because thetitre of this hormone is modulated by changes inenvironmental conditions (Robinson et al., 1989,unpublished), These findings are consistent with thewell-supported belief that the corpora allata, theglands that produce juvenile hormone, occupy a keyposition in the neuroendocrine response of insects toenvironmental factors (e.g. De Wilde et al., 1959;Tobe and Stay, 1985).
It is not known how the juvenile hormone titre isregulated in adult honey bees, and this informationis important for two reasons. First, the relationship
133

734 Zm-YONG HUANG et al.
between juvenile hormone titre and rate of biosyn-thesis is not well understood in insects, becausemeasurements of both variables have been made forrelatively few species to date (Lanzrein et al., 1978;De Kort et al., 1982; Tobe et al., 1985; Rachinsky andHartfelder, 1990). Second, if changes in juvenilehormone titre are due to changes in rate of juvenilehormone biosynthesis in honey bees, then analysesof corpora all&a activity may be useful in studiesof hormonal regulation of bee behaviour. Forexample, an important question in the study ofdivision of labour is how colony needs are perceivedby workers. The recent discovery that plasticityin honey bee age polyethism is mediated by juvenilehormone (Robinson et al., 1989, unpublished)suggests that it may be possible to probe sensitivelyfor the effects of environmental stimuli on division oflabour by measuring juvenile hormone biosynthesis.In other insects it is known that juvenile hormonesynthesis by the corpora allata is controlled by neuro-hormones from the brain (reviewed by Tobe andStay, 1985; Feyereisen, 198%; Khan, 1988).
Using a radiochemical assay of juvenile hormonebiosynthesis (Tobe and Pratt, 1974; Pratt and Tobe,1974) that is widely used in studies of insect neuro-endoctinology (see Tobe and Stay, 1985), we reportthat there is a strong correlation between rates ofjuvenile hormone biosynthesis and haemolymphtitres ofjuvenile hormone in adult worker honey bees,suggesting that titres are regulated primarily throughchanges in biosynthesis. We also describe similaritiesand differences in parameters of juvenile hormonebiosynthesis between bees displaying two differentage-dependent behaviours, brood care or foraging.These findings are supported by the results of adetailed characterization of the radiochemical assayfor adult worker honey bees, also presented here, thatis based on an established protocol for using thisassay on other species (Feyereisen, 1985b).
Bees were obtained from colonies maintainedaccording to standard techniques at the Universityof Illinois Bee Research Facility, Urbana, III. Theywere typical of North American populations of Apismel/ifera [a mix of predominantly European sub-species (Phillips, 1915; Pellett, 1938)].
Bees of known age were obtained by removingcombs containing developing pupae from a colonyand placing them in an incubator (33°C). Emergingadults (N = 800) were marked on the abdominaldorsum with a spot of paint (Testor’s PLA) andintroduced to a different colony with a populationof about 40,000 workers, occupying two Langstrothhive bodies.
Individuals engaged in either brood care (“nurse”bees) or foraging were sampled, as in previous studiesof hormonal regulation of division of labour
(Robinson et a/., 1989). Focusing on these two taskssimplifies physiological analyses of division of labourbecause they are associated with easily detectabledifferences in worker age, behaviour, and juvenilehormone titre. A nurse bee was identified as a workerwith her head in a cell containing a larva (e.g.Sakagami, 1953; Robinson, 1987a). Foragerswere obtained by temporarily obstructing the hiveentrance with a piece of S-mesh hardware cloth. Onlyreturning bees with pollen loads in their corbiculaewere collected. In one experiment we also sampledl-day-old bees upon emergence from combs in theincubator and 14-15-day-old workers from thenest periphery. Workers were collected and stored invials without food for 3&60 min at room tempera-ture. They were then immobilized on ice for 5 minto 3 h until dissected.
Incubation medium
A medium specially formulated for honey beetissue maintenance and culture (Kaatz et a/., 1985)was modified as follows: methionine and bovineserum albumin were omitted, and I mM leucine andFicoll(20 mg/ml) added. The pH of the medium wasadjusted to 7.0 (with a mixture of I.5 NaOH: I KOH,v:v) and the osmolarity adjusted to 52OmOsmol/l(with sucrose) to make it similar to honey bee haemo-lymph (Florkin and Jeuniaux, 1974). The mediumwas sterilized by filtration (Millipore, 0.22 pm). Pre-liminary studies indicated that use of this medium(“bee medium”) resulted in significantly higher ratesof juvenile hormone biosynthesis than either Grace’sinsect medium (Gibco), or bee medium with 5%bovine serum albumin (F = 7.77, P < 0.01, N = 36).Rachinsky and Hartfelder (1990) also reported thattheir modified bee medium is superior to other mediafor juvenile hormone biosynthesis in honey beelarvae, presumably because it has a Na:K ratio thatis more similar to that of honey bee haemolymph.
After removing the head, an incision was madeposteriorly and the corpora all&a-corpora cardiacacomplex removed. The complex was then placed ina disposable culture tube (Kimble 10 x 5Omm,pre-baked for 3 h at SOOOC) containing 25 @I non-radioactive bee medium. Corpora alla&corporacardiaca were incubated together in all experimentsreported here; a pilot study revealed that corporaallata-corpora cardiaca complexes showed similarrates of juvenile hormone biosynthesis to corporaall&a incubated without corpora cardiaca (t = 0.31,P > 0.3, N = 12) (corpora allata-corpora cardiacacomplexes are henceforth referred to as “corporaallata”). Dissections were conducted in a dissectionsaline with a Na:K ratio similar to that of thebee medium (55mM N&l, 35 mM KCI, 3 mMCaCl,~2 H,O, 10 mM MgQ.6 H,O, 55.5mM glu-cose, 55.5 mM fructose, 14.6 mM sucrose). Rates ofjuvenile hormone biosynthesis for glands dissected in

Juvenile hormone biosynthesis in honey bees 735
this saline were not significantly different than forglands dissected with bee medium (f = 0.30, P > 0.3,N = 12). The size of one corpus allatum per bee wasmeasured immediately after dissection with an ocularmicrometer. Corpus allatum volume was calculatedwith the formula V = n.(L x W’)/6, where L isthe larger diameter and W the smaller diameter,measured when the corpora alla& were oriented sideby
RL
..1” ---.“...~ .- led-1.111.1.procedures (Pratt and Tobe, 1974; Tobe and Pratt,1974; modified by Feyereisen and Tobe, 1981) and issummarized as follows. Incubations began after alldissections were complete (l-3 h) by adding to eachtube an additional 25 ~1 bee medium that containedL-[lH-methyl]methionine (NEN, specific activity7.4 GBq/mmol). The optimal final concentration ofmethionine was determined to be 100-150 PM (seeResults) and was used in all experiments reported inthis paper. Corpora allata were incubated for 3 h indarkness at 35°C. Glands incubated at this tempera-ture exhibited the highest rates of juvenile hormonebiosynthesis among the four temperatures tested (20,30, 35 and 40°C). This temperature is also closest tothe temperature maintained in the centre of a honeybee nest during the summer (see Seeley, 1985). Cor-pora allata were not shaken during incubationbecause we found that rates of juvenile hormonebiosynthesis were not significantly affected by shak-ing (t = 0.57, P > 0.3, N = 12).
Juvenile hormone was extracted according to themethod of Feyereisen and Tobe (1981). Glands wereremoved following the incubation period, and 250 ~1isooctane added to each tube of medium (isooctaneand all other solvents were HPLC grade, from FisherScientific), A 200.PI-aliquot of isooctane extract wasremoved after thorough vortexing and centrifugation(10 min, 17OOg), and radioactivity determined byliquid scintillation spectrometry.
To determine the time course of juvenile hormonebiosynthesis, corpora allata from nurses and foragers(N = 3 and 4, respectively) were incubated for 9 h,but were transferred to new medium after I, 2, 4 and6 h of incubation.
The effect of farnesoic acid (a juvenile hormoneprecursor) on juvenile hormone biosynthesis wasdetermined by incubating corpora allata from for-agers in medium that contained either 0, 20, 40, 80,160, 320, 640 or 1000 PM farnesoic acid (N = 6 perconcentration).
Two experiments were performed to determinethe precise rates of juvenile hormone biosynthesis.We measured the ratio of incorporation of theS-methyl moiety of radiolabelled methionine into thecarbon skeleton of the juvenile hormone molecule.In the first experiment, the medium was modifiedby omitting fructose and glucose, and adding[‘Hlmethionine (36 PM, 7.4 GBq/mmol, NEN) and
[2-“Q&ate (3.6 mM, 0.074 GBq/mmol, NEN).In the second experiment, [“CJmethionine (100 NM,2.07 GBq/mmol, NEN) and [‘H]famesoic acid(160~M, 1.01 GBq/mmol, courtesy of G. D. Prest-with) was used in standard bee medium. Analyseswere performed by thin-layer chromatography (TLC)and liquid scintillation spectrometry.
Identification of radiolabelled compounds synthesizedby the corporn aliata
Two experiments were performed with high per-formance liquid chromatography (HPLC). In Exper-iment I, 4-S pairs of corpora allata from nurse beesor foragers were incubated per incubation tube(100~1 medium) for 15 h (6 tubes each for nurse beesand foragers). Each sample (N = 5) was prepared bypooling isooctane extracts of medium from either twotubes (for foragers), or three tubes (for nurse bees).In Experiment 2, corpora allata from individual bees(N = 46 nurse bees and 41 foragers) were incubatedseparately for 3 h. Each sample (N = 6) was preparedby pooling medium extracts from l&22 individualbees. In Experiment 1, juvenile hormone was ex-tracted from the samples immediately, whereas inExperiment 2 the isooctane-medium mixtures werestored at -70°C for 1-2 days before extraction.
Juvenile hormone was extracted according toWeaver et al. (1980), except isooctane was usedinstead of chloroform. Samples were each passedthrough a Na,SO, column and loaded onto apre-washed silica cartridge (SepPak), rinsed withn-pentane, and &ted with n-pentane:diethyl ether(1:I). They were then evaporated under nitrogenand redissolved in IOfll hexane. HPLC analysiswas performed on a 5 pm Brownlee Spheri-5 Silicacolumn (250 cm x 4.6 mm) with water-saturated re-distilled ethyl ether (10%) in hexane as solvent. Flowrate was 1 ml/min at a pressure of 16-18 atm anddetector wavelength was set at 219nm. Fractionswere collected at 12-s intervals and radioactivitydetermined by liquid scintillation spectrometry. Syn-thetic juvenile hormone III (Chemical DynamicsCorp.) and methyl farnesoate (courtesy of F. C.Baker, Sandoz Crop Protection Corp.) were used asstandards.
Thin-layer chromatography
TLC was performed to provide independentidentification of radiolabelled products of the COT-pora allata and to quantify the relationship betweenthe amount of juvenile hormone released into theincubation medium fiuvenile hormone “release”) andthe total amount ofjuvenile hormone biosynthesized.This information is necessary to determine whethermeasuring juvenile hormone release alone providesan accurate estimate of total juvenile hormone bio-synthesis (Tobe and Stay, 1985). For this experimentonly, juvenile hormone biosynthesis is defined as theamount of juvenile hormone in the medium plusthe amount present in the glands at the conclusion
736 Zm-YONO HUANO ef nl
of the incubation period. Juvenile hormone wasextracted by first adding methanol and disodiumethylene-diamine-tetraacetate, and then partitioningagainst chloroform (Weaver et al., 1980). TLC wasperformed with a solvent system of xylenejethylacetate (3: l), with juvenile hormone III (Sigma) andmethyl farnesoate as internal markers.
Correlation between juvenile hormone fibe and rateof biosynthesis
Both variables were measured in the same individ-uals: l-day-old bees, 7-9.day-old nurses, 14-15.day-old bees located on the periphery of the nest, and21-24-day-old foragers (N = 9 or 10 per age group).Bees were chilled (0°C for 5 min) and immobilizedwith plasticine in a dissecting dish. Haemolymph(2.5-15fil) was removed from each bee, pooled(21.9-41.7~1 for each age group), and placed in150 ~1 methanol. The methanol-haemolymph mix-ture was partitioned against hexane three times andthe hexane phase, containing the juvenile hormone,removed. Samples were coded for blind analyses andstored at -20°C. Immediately after haemolymph wastaken from a bee, its corpora alla& were removed andplaced in incubation medium for the radiochemicalassay. Additional marked bees from the samecolony were collected for titre determinations only, toyield a total of 5-8 measurements per age group ofworkers.
Juvenile hormone titres were determined by radio-immunoassay (RIA) (Strambi et al., 1984). Thistechnique provides consistent results that agree withanalyses made by gas chromatography/mass spec-trometry (CC/MS) (De Kort et al., 1985). Inaddition, juvenile hormone titre determinationsfor honey bees with this RIA (Robinson ef al.,1987, 1989, unpublished) agree with measurementsmade with the Galleria bioassay (Fluri ef a/., 1982)and a GC/MS assay (Hagenguth and Rembold,1978).
Mean rates of biosynthesis (+ SEM) were calcu-lated based on measurements of individually incu-bated corpora allata (i.e. a gland pair). Sample sizesthus refer to the number of individual bees assayed,unless otherwise indicated. Statistical tests used arereported with the results.
RESULTS
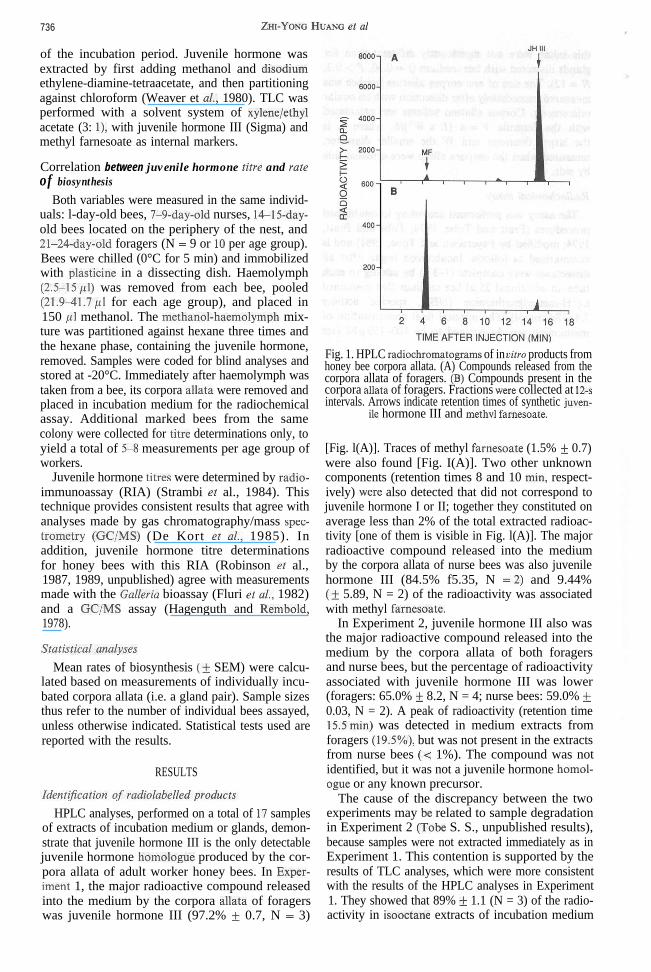
HPLC analyses, performed on a total of 17 samplesof extracts of incubation medium or glands, demon-strate that juvenile hormone III is the only detectablejuvenile hormone homologue produced by the cor-pora allata of adult worker honey bees. In Exper-iment 1, the major radioactive compound releasedinto the medium by the corpora all&a of foragerswas juvenile hormone III (97.2% + 0.7, N = 3)
Fig. 1. HPLC radiochromatograms of in vitro products fromhoney bee corpora allata. (A) Compounds released from thecorpora allata of foragers. (El) Compounds present in thecorpora alla& of foragers. Fractions were collected at 12-sintervals. Arrows indicate retention times of synthetic juven-
ile hormone III and methvl famesaate.
[Fig. l(A)]. Traces of methyl farnesoate (1.5% f 0.7)were also found [Fig. I(A)]. Two other unknowncomponents (retention times 8 and 10 min, respect-ively) were also detected that did not correspond tojuvenile hormone I or II; together they constituted onaverage less than 2% of the total extracted radioac-tivity [one of them is visible in Fig. l(A)]. The majorradioactive compound released into the mediumby the corpora allata of nurse bees was also juvenilehormone III (84.5% f5.35, N =2) and 9.44%(? 5.89, N = 2) of the radioactivity was associatedwith methyl famesoate.
In Experiment 2, juvenile hormone III also wasthe major radioactive compound released into themedium by the corpora allata of both foragersand nurse bees, but the percentage of radioactivityassociated with juvenile hormone III was lower(foragers: 65.0% k 8.2, N = 4; nurse bees: 59.0% f0.03, N = 2). A peak of radioactivity (retention time15.5min) was detected in medium extracts fromforagers (19.5%), but was not present in the extractsfrom nurse bees (< 1%). The compound was notidentified, but it was not a juvenile hormone homol-ague or any known precursor.
The cause of the discrepancy between the twoexperiments may be related to sample degradationin Experiment 2 (Tobe S. S., unpublished results),because samples were not extracted immediately as inExperiment 1. This contention is supported by theresults of TLC analyses, which were more consistentwith the results of the HPLC analyses in Experiment1. They showed that 89% f 1.1 (N = 3) of the radio-activity in isooctane extracts of incubation medium
Juvenile hormone biosynthesis in honey bees
con&rated with the unlabelled juvenile hormoneIII internal standard. Although the percentage ofjuvenile hormone III produced was lower in Exper-iment 2 relative to Experiment 1,87% of the radioac-tivity in Experiment 2 was still associated with eitherjuvenile hormone or methyl farnesoate, a juvenilehormone precursor. Thus both HPLC and TLCanalyses indicate that juvenile hormone III is theprincipal product of synthesis and release in adultworker honey bees.
HPLC analyses revealed that extracts of glandsfrom both foragers and nurse bees contained princi-pally methyl farnesoate (89.3% + 4.9, N = 6), andonly 4.9% f I.9 juvenile hormone III [Fig. l(B)].No other juvenile hormone homologues besidesjuvenile hormone III were detected from eithermedium or gland extracts.
Experiments performed with [2-“C]acetate and[3H]methionine revealed that honey bee corporaallata did not incorporate acetate into juvenile hor-mone, either in the presence or absence of fructoseand glucose. When corpora allata from foragers wereincubated in the normal medium (i.e. with glucoseand fructose), “C from acetate accounted for only0.81% + 0.31 (N = 4) of the total radioactivity in thesynthesized juvenile hormone. Similarly, when COT-pora allata were incubated in a medium withoutglucose and fructose, only 0.15% k 0.1 I (N = I I) ofthe radioactivity was from acetate. We thus failedto observe the predicted molar ratio of 9:1 “C:‘H[which is based on the incorporation of nine acetateunits from [2-‘4C]acetate and one methyl group fromL-methionine into each juvenile hormone III molecule(see Feyereisen ef al., 1984; Smith et al., 1989)].
Experiments performed with @C]methionineand [‘H]farnesoic acid showed that methionine, rela-tive to farnesoic acid, is incorporated into juvenilehormone in a I:1 molar ratio (l:l.O6 +O.O6molL-methionine: farnesoic acid, N = 22). A I : I molarratio is expected theoretically because one mol-ecule of farnesoic acid and one methyl group fromL-methionine are incorporated into each juvenilehormone III molecule (Tobe and Pratt, 1974; Prattand Tobe, 1974). This result demonstrates that honeybee corpora all&a use methionine as the exclusivemethyl donor to produce juvenile hormone III (Tobeand Stay, 1985). Accordingly, we calculated rates ofjuvenile hormone biosynthesis based on the specificradioactivity of methionine throughout this paper.
Differences in juvenile hormone biosynthesis between“w-se bees and foragers
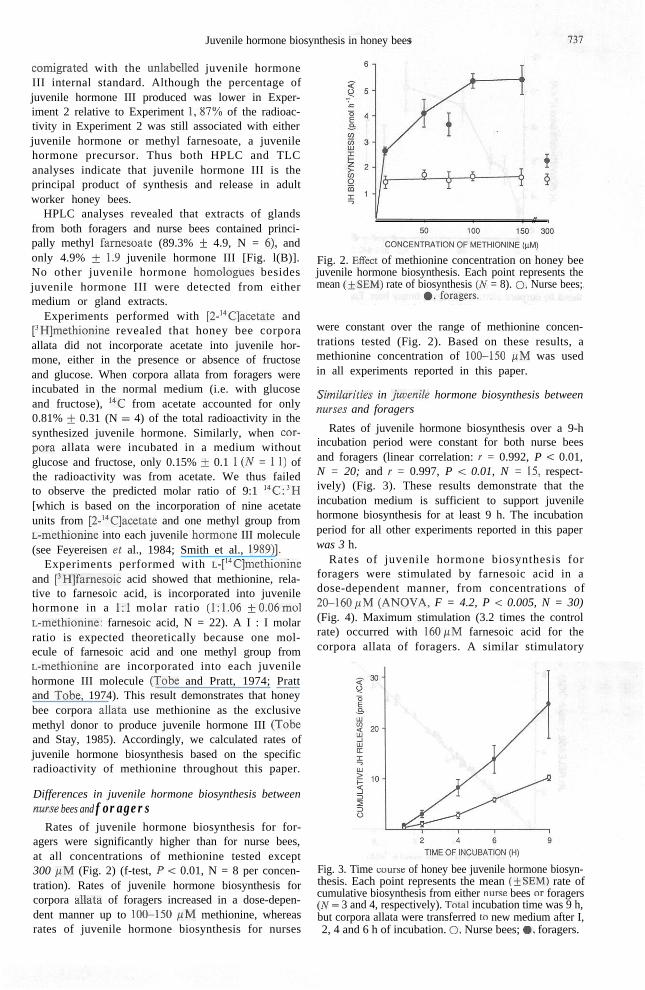
Rates of juvenile hormone biosynthesis for for-agers were significantly higher than for nurse bees,at all concentrations of methionine tested except300 pM (Fig. 2) (f-test, P < 0.01, N = 8 per concen-tration). Rates of juvenile hormone biosynthesis forcorpora alla& of foragers increased in a dose-depen-dent manner up to 100-150 PM methionine, whereasrates of juvenile hormone biosynthesis for nurses
Fig. 2. E&t of methionine concentration on honey beejuvenile hormone biosynthesis. Each point represents themean (iSEM) rate of biosynthesis (N = 8). 0, Nurse bees;
were constant over the range of methionine concen-trations tested (Fig. 2). Based on these results, amethionine concentration of lOSl50 PM was usedin all experiments reported in this paper.
Similarifies in juvenile hormone biosynthesis betweennurses and foragers
Rates of juvenile hormone biosynthesis over a 9-hincubation period were constant for both nurse beesand foragers (linear correlation: I = 0.992, P c 0.01,N = 20; and I = 0.997, P < 0.01, N = 15, respect-ively) (Fig. 3). These results demonstrate that theincubation medium is sufficient to support juvenilehormone biosynthesis for at least 9 h. The incubationperiod for all other experiments reported in this paperwas 3 h.
Rates of juvenile hormone biosynthesis forforagers were stimulated by farnesoic acid in adose-dependent manner, from concentrations of20-l60pM (ANOVA, F = 4.2, P < 0.005, N = 30)(Fig. 4). Maximum stimulation (3.2 times the controlrate) occurred with 160pM farnesoic acid for thecorpora allata of foragers. A similar stimulatory
Fig. 3. Time wurse of honey bee juvenile hormone biosyn-thesis. Each point represents the mean (iSEM) rate ofcumulative biosynthesis from either nurse bees or foragers(N = 3 and 4, respectively). Totai incubation time was 9 h,but corpora allata were transferred to new medium after I,2, 4 and 6 h of incubation. 0, Nurse bees; ., foragers.

0 80 160 320 640 loo0
CONCENTRATlON OF FmNESOIC ACD (L&4,
Fig. 4. Effect of famesoic acid on juvenile hormone biosyn-thesis by corpora allata of foraging honey bees. Each pointrepresents the mean (+SEM) rate of biosynthesis (N = 6).
effect occurred for the corpora allata of nurse beesat a farnesoic acid concentration of 160 PM (3.4 vs1.2pmol h-‘/corpora a l la ta , N = 12 , da ta notshown). Corpora all&a of nurse bees and foragersthus showed similar responses to farnesoic acid de-spite the previously mentioned differences in rates ofjuvenile hormone biosynthesis.
There is a linear relationship between juvenilehormone release and juvenile hormone biosynthesisfor both nurse bees and foragers (r = 0.99, P < 0.001,N = 15; and I = 0.997, P < 0.001, N = 32, respect-ively) (Fig. 5). Regression analysis of the combineddata yielded a slope of 0.95 i 0.01 (N = 47), indicat-ing that 95% of the biosynthesized juvenile hormonewas released into the medium. These results areconsistent with the results of HPLC analyses pre-sented above. Isooctane extraction of the mediumthus provides an accurate estimate of juvenile hor-mone biosynthesis in adult worker honey bees.
Fig. 5. Relationship between honey bee juvenile hormonebiosynthesis and release. For this experiment only, biosyn-thesis is defined as the total amount of juvenile hormonereleased into the medium and that present in the corpora
allata. A, Nurse bees (N = 15); ., foragers (N = 32).
Fig. 6. Correlation between rate ofjuvenile hormone biosyn-thesis and haemolymph titre in honey bees. (A) Mean rateof biosynthesis (+SEM; N = 10); (B) 0, titre determinedfrom pooled haemolymph from the same bees for whichrates of biosynthesis were measured; ., mean (*SEMItitres from 5-8 groups of bees (5-9 bees per group),
including those depicted with 0.
Correlation between juvenile hormone biosynthesis andimenile hormone live
There is a significant correlation between juvenilehormone biosynthesis and juvenile hormone titre,based on measurements of both variables in the sameindividuals (r = 0.96, P < 0.05) (Fig. 6). In foragers,juvenile hormone biosynthesis rates and haemolymphtitre were significantly higher than l-day old bees,7-9-day-old nurse bees and 14-15-day-old nes tperiphery bees (ANOVA: biosynthesis: P < 0.001,N = 38; titre: P < 0.001, N = 26).
Corpora allatum volumes ranged from 0.27to 1.16~10”~m’. There is a significant correlationbetween juvenile hormone biosynthesis and corporaallata volume (Fig. 7). However, only 13% of thevariation in juvenile hormone biosynthesis can beexplained on the basis of volume alone (r’ = 0.13).Separate analyses of the data for nurse bees andforagers revealed that a significant correlation existsonly for the former, but not for the latter. In addition,the range of corpora allatum volumes is smallerthan the range of rates of juvenile hormone biosyn-thesis. Mean volumes for nurse bees and foragerswere 0.62 and 0.82. IO6 pm’, respectively, whereas

Juvenile hormone biosynthesis in honey bees 739
CORP”S AUATUM VOLUME (10’ x pmy
Fig. 7. Relationship between rates of juvenile hormonebiosynthesis and corpus allatum volume in honey bees
1.23 and 4.37 pmol h-‘/corpora allata, respectively(N = 70).
Differences in juvenile hormone ties betweenadult worker honey bees performing different age-dependent tasks appear to be regulated primarilyby changes in rates of biosynthesis rather than bychanges in rates of hormone release or degradation.This conclusion is supported by two lines of evidence.First, almost all juvenile hormone in the corporaallata is released upon synthesis. Second, there is ahigh correlation between juvenile hormone biosyn-thesis and juvenile hormone titre. There are only twoprevious studies in which measurements were made ofboth juvenile hormone titres and rates of biosynthesisfrom the same individuals, and in both the two werehighly correlated (see Tobe er al., 1985), as in ourstudy. Our results are consistent with an apparentabsence of juvenile hormone-specific esterases inthe haemolymph of adult worker bees (Strambi C.er al., unpublished results). Rachinsky and Hartfelder(1990) reported that juvenile hormone ties in honeybee queen and worker larvae also are regulated bychanges in rates ofjuvenile hormone biosynthesis (seealso Tobe ef al., 1985). Only very small amounts ofjuvenile hormone III estemse are found in honeybee larvae, about 1000.fold less than in Coloradopotato beetle (De Kort et al., 1977), and its specificityis uncertain. Degradation of juvenile hormone never-theless is probably important in regulating thehaemolymph titre in worker honey bees, becausethe rate of biosynthesis in vitro is relatively highcompared with the haemolymph titre. However, thehigh correlation of titre and rate of biosynthesis inboth nurses and foragers suggests that degradationdoes not appear to play a major role in regulating
differences in titres that are associated with differentage-dependent behavioural states.
The in vitro rates of juvenile hormone biosynthesisreported here are similar to those reported byHildebrandt and Kaatz (1990) for adult worker bees.They are much higher than those reported by Biihleret al. (1983) (<0.5pmol h-‘/corpora allata) perhapsbecause in that study glands were incubated in asuboptimal medium, as a medium formulated specifi-cally for honey bees (Kaatz ef a/., 1985) was not yetavailable. Our detection of only juvenile hormone IIIand our measurements of juvenile hormone titresare consistent with previous analyses, using theGderia bioassay (Fluri er a/., 1982), a GC/MSassay (Hagenguth and Rembold, 1978), and RIA(Robinson et al., 1987, 1989). High titres in foragersand low titres in nurse bees also agree with exper-iments demonstrating that foraging behaviour canbe introduced by treatment with juvenile hormone(Jaycox, 1976), juvenile hormone mimic (Jaycox efal., 1974), or juvenile hormone analogue (Robinson,1985, 1987a; Sasagawa ef al., 1985).
Juvenile hormone titres and rates of juvenile hor-mone biosynthesis for 14-15-day-old bees from thenest periphery were similar to those for 7-9-day-oldnurse bees. Bees from the nest periphery are in abehavioural phase that is in transition from nursingto foraging, and they are involved in nest mainten-ance and food storage tasks (see Seeley, 1982; andWinston, 1987). Our results suggest that there arediscrete states of corpora allata activity associatedwith nursing and foraging, but not with the interven-ing behavioural phase(s). Robinson ef al. (1987)reported no differences in juvenile hormone titresbetween bees aged 5, 10 or 15 days in one experiment,which is consistent with our findings. They did,however, detect an age-dependent increase in juvenilehormone titres in a second experiment, whichsuggests continuous changes in corpora allata activitythroughout behavioural development. It is clear thatlow rates of juvenile hormone biosynthesis and lowjuvenile hormone titres are associated with nursingbehaviour and high rates and titres with foraging,but additional studies are needed to determine therelationship between juvenile hormone and otherworker behaviours.
Measurements of corpus allatum volumes, whichagreed with those in previous studies (Gast, 1967;Breed, 1983; Biihler et al., 1983), indicate that it is notpossible to use such measurements to estimate accu-rately levels of corpora allata activity in honey bees.Similar results have been obtained for other insectspecies (see Tobe and Pratt, 1975). For example, thevolume of a corpus allatum in the wasp Po/isfesgallicus was correlated with juvenile hormone bio-synthesis in females undergoing egg maturation, butnot in overwintering or parasitized females (R&&rer al., 1980).
Our results reveal two intriguing differences inparameters ofjuvenile hormone biosynthesis between

nurse bees and foragers, aside from the large differ-ences in rates. Rates ofjuvenile hormone biosynthesisapparently reached maximal levels in nurse bees ata concentration d 10 PM methionine, but increasedsteadily in foragers up to a concentration of IOOfiM.This difference may reflect differences between nursebees and foragers in transport of methionine into thecorpora allata and/or differences in the formationof S-adenosyl methionine. Insect species show awide range of optimal L-methionine concentrationsfor juvenile hormone biosynthesis, from as low as5pM in termites (Greenberg and Tobe, 1985) to20&300pM in locusts (Tohe and Pratt, 1974).
The second difference between nurse bees andforagers is that the rate of juvenile hormone biosyn-thesis, when stimulated with fames& acid, is muchlower for nurse bees than for foragers. This suggeststhat there also may be differences between nurse beesand foragers in the activity of some rate-limitingenzyme(s) prior to the formation of fames& acid(see Tobe and Stay, 1985). For both nurse bees andforagers, the degree of stimulation by farnesoic acidwe observed (three times the spontaneous rate) is nearthe lower end of the range reported for other species[1.2-10 times the spontaneous rate (see Tobe andStay, 19X5)]. Relatively low stimulation by farnesoicacid in our results suggests that corpora allata weremaintained in relatively optimal conditions for juven-ile hormone biosynthesis in our experiments (Tobeand Stay, 1985). Alternatively, low stimulation byfarnesoic acid may be a consequence of the inacti-vation or saturation of o-methyl transferase, theenzyme responsible for formation of methyl farne-soate from fames& acid, as in last instar Diploptera(Yagi ef al., in press).
Our results also suggest one difference in juvenilehormone biosynthesis between honey bees and otherspecies. Corpora allata from honey bees apparentlydo not use acetate as a substrate for juvenile hormonebiosynthesis, in contrast to all other insects studied todate (see Tobe and Stay, 1985). Incorporation of[“C]acetate and [lH]methionine into adult bee cor-pora allata did not exhibit the expected 9: 1 “C:‘Hincorporation ratio. Corpora allata from honey beelarvae also fail to show the expected ratio (RachinskyA. and Tobe S. S., unpublished results). Alterna-tively, perhaps corpora allata from honey bees douse acetate as a substrate for juvenile hormonebiosynthesis, but the sucrose present in the incu-bation medium may have been broken down tosimple sugars and used as a carbon source insteadof acetate.
The strong correlation between juvenile hormonebiosynthesis and titre for nurse bees and foragersindicates that changes in biosynthetic rates appearto accurately presage changes in ham&mph titresthat have been determined to mediate behaviouralontogeny in worker honey bees (Robinson er al.,1989, unpublished). The radiochemical assay ofjuvenile hormone biosynthesis provides a reliable and
sensitive method for the study of hormonal regu-lation of honey bee behavioural development. It ispossible to measure rates ofjuvenile hormone biosyn-thesis in individual bees, whereas measurements ofjuvenile hormone titres (Robinson et al., 1987, 1989)require pooled samples from at least several individ-uals. The ability to measure rates of juvenile hormonebiosynthesis for individual bees will increase theprecision of studies relating juvenile hormone andbehaviour.
Acknowledgements-We thank 1. C. Kuehn for technicalassistance; H. H. Katz and A. Rachinsky for advice on theincubation medium and the stoichiometry experiments,respectively; F. C. Baker and 0. D. Prestwich for generouslyproviding methyl famesoate and famesoic acid, respectively;P. M. Best, J. Nardi and J. H. Willis for use of their facilitiesfor medium preparation, C. Schal for graciously andpromptly providing [‘H]methionine during an unforeseendelay in commercial deliveries, and two anonymous review-ers who offered helpful comments. Supported by grantsfrom NSF (BSR-88W227), NIMH (MH47374.01) andthe University of Illinois (a Beckman Award and NIH-Biomedical Research Support Grant PHS-507.RR-07030)to G.E.R., USPHS Grant Al 15230 to B.S. and an NSERCOperating Grant to S.S.T.
Breed M. D. (1983) Correlations between aggressivenessand corpora alla& volume, social isolation, age anddietary protein in worker honeybees. Inseczes Sot. 30,482495.
Biihler A., Lanzrein B. and Wille H. (1983) Influence oftemperature and carbon dioxide concentration on juvenilehormone titre and dependant parameters of adult workerhoney bees (Rpir me/Ii/em L.). J. Inset! Physio,. 29,885-893.
De Kort C. A. D., Wieten, M., Kramer S. I. and GoewieE. (1977) Juvenile hormone degradation and carrier pro-teins on honey bee lame. Proc. K. ned. Aknd. Wet. 8OC,297-301.
De Kart C. A. D., Bergot B. A. and Schooley D. A. (1982)The natnre and titre of juvenile hormone in the ColoradoBeetle, Leplinofarsa decemlineata. J. Imeet Physiol. 28,47,474.
De Kort C. A. D., Koopmanschap A. B., Strambi C. andStrambi A. (1985) The application and evaluation of aradioimmunoassay for measuring juvenile hormone titresin Colorado beetle haemolymph. Insect Biochem. 15,n - 7 7 5 .
De Wilde J., Duintjer C. S. and Mook L. (1959) Physiologyof diapause in the adult Colorado beetle (Leprinomrsadeeemlineara Say)-I. The photoperiod as a controllingfactor. J. Inseel Physiol. 3, 75-85.
Feyereisen R. (l985a) Regulation of juvenile hormonetiter: synthesis. In Comprehensive Insect Physiology,Biochemistry and Pharmacology (Eds Kerkut G. A. andGilbert L. I.), Vol. 7, pp. 391429. Pergamon Press,Oxford.
Feyereisen R. (1985b) Radiochemical assay for juvenilehormone III biosynthesis in oiwo. Merh. Enzym. 111,530-539.
Feyereisen R. and Tobe S. S. (1981) A rapid partition assayfor routine analysis of juvenile hormone release by insectcorpora allata. Analyr. Biochem. 111, 372-375.
Feyereisen R., Ruegg R. P. and Tabe S. S. (1984) Juvenilehormone III biosynthesis; stoichiometric incorporation of(2-V] acetate and effects of exogenous famesol andfamesoic acid. I~eer Biochem. 14, 657461.

Juvenile hormone biosynthesis in honey bees 741
Florkin M. and Jenniaux C. (1974) Hemolymph compo-sition. In Physiology of Inseem (Ed. Rockstein M.),Vol. 5, pp. 225-307. Academic Press, New York.
Fluri P., L&her M., Wille H. and Gerig L. (1982) Changesin weight of pharyngeal gland and haemolymph titres ofjuvenile hormone, protein and vitellogenin in workerhoney bees. J. Insecr Physio,. 28, 6148.
Gas, V. (1967) “ntersuchungen iiber den &Russ derKbniginnensubstanz auf die Entwicklung der endokrinenDriisen bei der Arbeiterin der Honigbiene (&is me&a).Insectes sot. 14, l-12.
Greenberg S. and Tobe S. S. (1985) Adaptation of aradiochemical assay for juvenile hormone biosynthesis tostudy caste differentiation in a primitive termite. J. Insec,Pkysio,. 31, 347-352.
Hagenguth H. and Rembold H. (1978). Identit%ation ofjuvenile hormone 3 as the only juvenile hormone horn&gin all developmental stages of the honey bee. Z. NarurfPC, 847-850.
Hildebrandt H. H. and Katz H.-H. (1990) impact ofqueen pheromone an the physiological status of workerhoney bees (&is me,,ife,u L.). In Social Inreets andthe Eneironmenr (Eds Veeresh G. K., Mallik B. andViraktamath C. A.), pp. 74&741. IBH, New Delhi.
Jaycox E. R. (1976) Behavioral changes in worker honeybees (&is mellifera L.) after injection with syntheticjuvenile hormone (Hymenoptera, Apidae). J. Kahns. Em.sot. 49, 165-170.
Jaycox E. R., Skowrenek W. and Gwynn G. (1974) Behav-ioral changes in a worker hone” bee iA& melli~era)induced by-injections of a juvenile ho&& mimic.‘Ann:en!. Sot. Am. 67, 529-534.
Katz H.-H., Hagedorn H. H. and Engels W. (1985) Cultureof honeybee organs: development of a new medium andthe importance of tracheadan. In Yiwo Gel/. De”. Biol. 21,141L2<7_ ., __-.
Khan M. (1988) Brain-controlled synthesis of juvenilehormone in adult insects. Enromologia exp. a&. 46,3-17.
Lanzrein B., Gentinetta V. and L&her M. (1978)Correlation between haemolymph juvenile hormonetitre, corpus allatum volume, and corpus allaturn in uivoand in oifm activity during oocyte maturation in acockroach (Naupheota ebwea). Gen. eomp. Endow 36,339-345.
Milojevic B. D. (1940) A new interpretation of the sociallife of the honeybee. Bee W/d 21, 39-41.
Pellet F. C. (1938) Hisrory of American Beekeeping.Collegiate Press, Ames, IA.
Phillips E. F. (1915) Beekeeping. Macmillan, New York.Pratt G. E. and Tobe S. S. (1974) Juvenile hormones
radiobiosynthesized by corpora allata of adult femalelocusts m “iW0. Life sci. 14, 575-586.
Rachinsky A. and Hartfelder K. (1990) Corpora allataactivity, a prime regulating element for caste-specificjuvenile hormone titre in honey bee larvae (,Qir mellr/erncamico). .I Imecl Physiol. 36, 189-194.
Robinson G. E. (1985) Effects of a juvenile hormoneanalague on the ontogeny of honey bee foragingbehaviour and alarm pheromone production. J. InseerPhysiol. 31, 277-282.
Robinson G. E. (1987a) Regulation of honey bee agepolyethism by juvenile hormone. Behov. Eeol. Sociobiol.120, 329-338.
Robinson G. E. (3987b) Alarm pheromone perception inthe honey bee, evidence for division of labor based onhonnonally modulated response thresholds. J. camp.physiol. XOA, 613++19.
Robinson G. E. (1992) Regulation of division of labor ininsect societies. A. Rev Em. In press.
Robinson G. E., Strambi A., Strambi C., Pauline-SimBesZ. L., Toreto S. 0. and Barbosa J. M. N. (1987) Juvenilehormone titers in Africanized and European honey beesin Brazil. Gen. camp. Endocr. 66, 457459.
Robinson G. E., Page R. E., Strambi C. and Strambi A.(1989) Hormonal and genetic control of behavioral inte-gration in honey bee colonies. Science 246, 109-112.
Robinson G. E., Page R. E., Strambi C. and Strambi A.Colony integration in honey bees: mechanisms ofbehavioural reversion. Submitted.
R&h G. A. (1930) Untersuchungen iiber die Arbeitsteilungim Bienenstaat-2. Die Titigkeiten der Arbeitsbienenunter experimentelI verinderten Bedingungen. Z. vergl.Physiol. 12, l-71.
Rdseler P. F., RBseler 1. and Strambi A. (1980) Theactivity of corpora alla& in dominant and subordinatedfemales of the wasp F’oltires gallicus. Insares Sot. 27,97-107.
Sakagami S. F. (1953) Untersuchungen iiber die Arbeit-&lung in einem Zwergvalk der Honigbiene: BeitrriigeZUT Biologic des Bienenvolkes, &is mdfera L. Jop. J.Zoo,. 11, 117-185.
Sasagawa H., Sasaki M. and Okada I. (1985) Experimentalinduction of the division of labor in worker &is melliferaL. by juvenile hormone (JH) and its analog. Mrh Inf.Congr. Apimondio, Japan, pp. 13&132.
Seeley T. D. (1982) Adaptive significance of the agepolyethism schedule in honeybee colonies. Behov. Ecol.Sociobiol. 11, 287-293.
Seeley T. D. (1985) Honeybee Ecology. Princeton UniversityPress, Princeton, N.J.
Smith A. F., Yagi K. Tobe S. S. and Schal C. (1989) In v&ojuvenile hormone biosynthesis in adult virgin and matedfemale brown-branded cockroaches, Sup& longipalpa.J. Insect Physiol. 35, 781-785.
Strambi C., Strambi A., De Reggi M. L. and Del&ageM. A. (1984) Radioimmunoassays of juvenile hormones:state of the methods and recent data on validation. InBiosynrhesis, Metabolism and Mode of Refion of Inverr-ebrare Hormones (Eds HoiTmann I. and Porchet M.),pp. 356362. Springer, Berlin.
Tabe S. S. and Pratt G. E. (1974) The influence of substrateconcentrations on the rate if insect juvenile hormonebiosynthesis by corpora allata of the desert locust in vitro.Biochem. J. 144, 107-133.
Tabe S. S. and Pratt G. E. (1975) The synthetic activity andglandular volume of the corpus allaturn during ovarianmaturation in the desert locust S’chistocerco gregoria.Life Sci. 17, 417-422.
Tabe S. S. and Stay B. (1985) Structure and regulation ofthe CO~DUS allaturn. Ah. Insecr Phvsiol. IS. 306432.
Tabe S. <, Ruegg R. P., Stay B., B&r F. C.,‘Miller C. A.and Schooley D. A. (1985). Juvenile hormone titer andregulation in the cockroach Diploprern puncnmto. Experi-enria 41, 1028-1034.
Weaver R. J., Pratt G. E., Hamnett A. F. and JenningsR. C. (1980) The influence of incubation conditions on therates of juvenile honnone biosynthesis by corpora alla&isolated from adult females of the beetle Tenebrio molitor.Insect Biochem. 10, 245-254.
Winston M. L. (1987) The Biology of the Honey Bee.Harvard University Press, Cambridge, Mass.
Yagi K., Konz K. G., Stay B. and Tobe S. S. (1991)Production and utilization of famesoic acid in the juvenilehormone biosynthetic pathway by corpora allata of larvalDiploplera punctnto. Cm mmp. Endow. In press.