
HnRNP L-mediated regulation of mammalian alternative splicing by interference with splice site...
10
This article was downloaded by: [5.56.12.10] On: 16 July 2015, At: 03:43 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: 5 Howick Place, London, SW1P 1WG RNA Biology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/krnb20 HnRNP L-mediated regulation of mammalian alternative splicing by interference with splice site recognition Monika Heiner, Jingyi Hui, Silke Schreiner, Lee-Hsueh Hung & Albrecht Bindereif Published online: 01 Jan 2010. To cite this article: Monika Heiner, Jingyi Hui, Silke Schreiner, Lee-Hsueh Hung & Albrecht Bindereif (2010) HnRNP L-mediated regulation of mammalian alternative splicing by interference with splice site recognition, RNA Biology, 7:1, 56-64, DOI: 10.4161/rna.7.1.10402 To link to this article: http://dx.doi.org/10.4161/rna.7.1.10402 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
Transcript of HnRNP L-mediated regulation of mammalian alternative splicing by interference with splice site...

This article was downloaded by: [5.56.12.10]On: 16 July 2015, At: 03:43Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: 5 Howick Place,London, SW1P 1WG
RNA BiologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/krnb20
HnRNP L-mediated regulation of mammalianalternative splicing by interference with splice siterecognitionMonika Heiner, Jingyi Hui, Silke Schreiner, Lee-Hsueh Hung & Albrecht BindereifPublished online: 01 Jan 2010.
To cite this article: Monika Heiner, Jingyi Hui, Silke Schreiner, Lee-Hsueh Hung & Albrecht Bindereif (2010) HnRNP L-mediatedregulation of mammalian alternative splicing by interference with splice site recognition, RNA Biology, 7:1, 56-64, DOI:10.4161/rna.7.1.10402
To link to this article: http://dx.doi.org/10.4161/rna.7.1.10402
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

RNA Biology 7:1, 56-64; January/February 2010; © 2010 Landes Bioscience
ReseARch pApeR
56 RNA Biology Volume 7 Issue 1
*Correspondence to: Albrecht Bindereif; Email: [email protected]: 10/21/09; Accepted: 10/21/09Previously published online: www.landesbioscience.com/journals/rnabiology/article/10402
Introduction
Alternative mRNA splicing in the human system mainly con-tributes to the generation of complex proteomes from a compara-tively low number of genes. It is currently estimated that more than 90% of all human protein-coding genes undergo alterna-tive splicing.1 Besides the canonical splice site signals, additional sequence elements are required to define genuine exon-intron boundaries since cryptic splice sites that loosely match the con-sensus sequence are very common in introns. Splicing enhancer and silencer are cis-regulatory elements with the information nec-essary for either stimulating or repressing splicing. They function through binding of trans-acting factors, which can be divided into two major groups, SR (serine-arginine-rich) proteins and hnRNPs, typically representing splicing activators or repressors, respectively.2-6
We have focused on hnRNP L, an abundant nuclear RNA-binding protein, which we initially identified as a splicing activator of the human eNOS gene.7 Besides splicing regula-tion several functions had been previously documented, where hnRNP L is involved, such as export of intronless mRNAs,8,9 IRES-mediated translation,10,11 mRNA stability,12-14 and regu-lation of VEGFA expression through a novel RNA switch.15 Determination of the binding specificity by in vitro SELEX revealed that hnRNP L preferentially binds to CA repeats and certain CA-rich sequences.16 Considering that CA-repetitive
hnRNp L-mediated regulation of mammalian alternative splicing by interference
with splice site recognitionMonika heiner,1 Jingyi hui,2 silke schreiner,1 Lee-hsueh hung1 and Albrecht Bindereif1,*
1Institute of Biochemistry; Justus Liebig University of Giessen; Giessen, Germany; 2State Key Laboratory of Molecular Biology; Institute of Biochemistry and Cell Biology; Shanghai Institutes for Biological Sciences; Chinese Academy of Sciences; Shanghai, China
Keywords: splicing, alternative splicing, silencer, hnRNP, snRNP
ReseARch pApeR
sequences represent the most common simple sequence repeat in the human genome,17 it was likely that hnRNP L is involved in a global splicing network. Both by motif-based database searches as well as by splice-sensitive microarray analysis we have identi-fied a set of hnRNP L targets, where hnRNP L participates in different types of alternative splicing regulation, acting either as an activator or repressor.16,18 HnRNP L was also reported to interact with an exonic splicing silencer in exon 4 of the human CD45 gene, mediating repression of the corresponding exon.19,20 Recently, we have characterized as an unusual target of hnRNP L protein the hnRNP L pre-mRNA itself, where a poison exon is activated, resulting in NMD-mediated autoregulation of hnRNP L protein levels.21
Based on two representative target genes we have addressed here the mechanisms of hnRNP L’s repressor activity in alterna-tive splicing regulation. First, for the TJP1 gene we demonstrate by mutational analysis that an intronic CA-rich cluster close to the regulated 3' splice site represents an hnRNP L-dependent intronic splicing silencer. UV crosslinking and immunoprecipita-tion experiments showed that hnRNP L interferes with 3' splice site recognition. Second, the SLC2A2 gene contains an intronic CA repeat splicing silencer near the 5' splice site of the regulated exon. In this case, we demonstrate that hnRNP L interferes with 5' splice site recognition by the U1 snRNP. In sum, our data pro-vide new insights into the mechanisms of how hnRNP L—bound to intronic sites—can repress exon recognition.
heterogeneous nuclear ribonucleoprotein (hnRNp) L can regulate alternative mRNA splicing in diverse ways, binding to exonic or intronic sites and acting as either an activator or repressor. To investigate the mechanistic basis of hnRNp L-regulated alternative splicing, we focus here on two specific cases of hnRNP L-dependent splice site recognition. First, in the case of TJP1 our microarray data had suggested that exon 20 inclusion is regulated by hnRNp L as a repressor. here we demonstrate by mutational analysis that exon skipping is mediated by a short silencer sequence consisting of three hnRNp L high-score binding motifs located upstream of the 3' splice site of the regulated exon. UV crosslinking and immu-noprecipitation experiments showed that hnRNp L binding interferes with 3' splice site recognition by U2AF65. second, SLC2A2 contains a cA-repeat sequence close to the 5' splice site of the regulated exon 4. Using psoralen crosslinking, we demonstrate that hnRNp L represses splicing by preventing 5' splice site recognition of the U1 snRNp. In sum, our data provide new insights into the mechanisms of how hnRNp L—bound to intronic sites—regulates exon recognition.
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5

www.landesbioscience.com RNA Biology 57
ReseARch pApeR ReseARch pApeR
SDS-PAGE analysis (lanes 3 and 4). Both wildtype and mutant derivatives were used (mutations as shown in Fig. 1A). To control for specificity, UV crosslinking was also done in the absence of protein (lanes 1 and 2). As a result, hnRNP L could be cross-linked only to TJP1 wildtype RNA (lane 3), but not detectably to the mutant derivative nor in the absence of protein (lanes 1, 2 and 4). We conclude that hnRNP L interacts directly with the CA-rich silencer element of TJP1.
HnRNP L binding to intronic splicing silencer impairs 3' splice site recognition. How does hnRNP L mediate TJP1 exon 20 skipping? Since the hnRNP L binding sites reside within the polypyrimidine tract of the regulated exon, we tested the hypothesis that hnRNP L binding may interfere with recognition of the polypyrimidine tract by U2AF65. The 65 kDa subunit of U2AF directly contacts the polypyrimidine tract followed by recruitment of the U2 snRNP to the branch site.2
First, we analyzed hnRNP L binding to the polypyrimi-dine tract of TJP1 exon 20 by bandshift assays, using recom-binant hnRNP L protein and a 32P-labeled TJP1 RNA of 84 nucleotides with branch site, polypyrimidine tract, 3' splice site (Fig. 2B) and 27 nucleotides of exon 20. Already with a two-fold molar excess of hnRNP L protein over RNA (0.2 μM; lane 2), we could observe complex formation; a second complex appeared with higher protein concentrations (0.4, 0.8 μM; lanes 3 and 4), most likely due to multiple hnRNP L binding sites. With U2AF65 on the other hand, complex formation was detect-able only at comparatively high protein concentrations. Even at the highest concentration (20 μM; lane 8), which represents a 640-fold molar excess of protein over RNA, complex formation was inefficient, most likely due to the relatively poor quality of this polypyrimidine tract (low score for this 3' splice site of 5.2; determined by MaxEntScan23). As a control for the bind-ing activity of the recombinant U2AF65, we detected complex formation for MINX pre-mRNA, a well characterized in vitro splicing substrate, already at an 8-fold molar excess of U2AF65 (0.3 μM; lane 10), and the addition of increasing amounts of U2AF65 shifted all free RNA into complexes (lanes 11 and 12). Note that MINX splicing substrate contains a much stronger polypyrimidine tract than TJP1 intron 19 (MaxEntScore of 3' splice sites: 12.47 vs. 5.21).
To investigate whether hnRNP L interferes with U2AF65 binding at the polypyrimidine tract of TJP1 exon 20, we per-formed UV crosslinking experiments, using 32P-labeled wildtype TJP1 RNAs of 318 nucleotides containing the full-length exon 20 with its complete 3' splice site region as well as with its 5' splice site (Fig. 3A). After incubation with either GST-U2AF65 (see leg-end), hnRNP L, or both proteins, UV crosslinking was done, fol-lowed by RNase A treatment and analysis of crosslinked proteins by SDS-PAGE (lanes 2–4); as a negative control, crosslinking was carried out in the absence of proteins (lane 1). After incuba-tion with either protein alone, we detected a specific crosslinked band for U2AF65 and hnRNP L, respectively (lanes 2 and 3). On the other hand, incubation with both proteins resulted in an almost exclusive hnRNP L crosslink, with insignificant amounts of U2AF65 crosslinked (lane 4), suggesting that hnRNP L pre-vents binding of U2AF65 to TJP1 RNA.
Results and Discussion
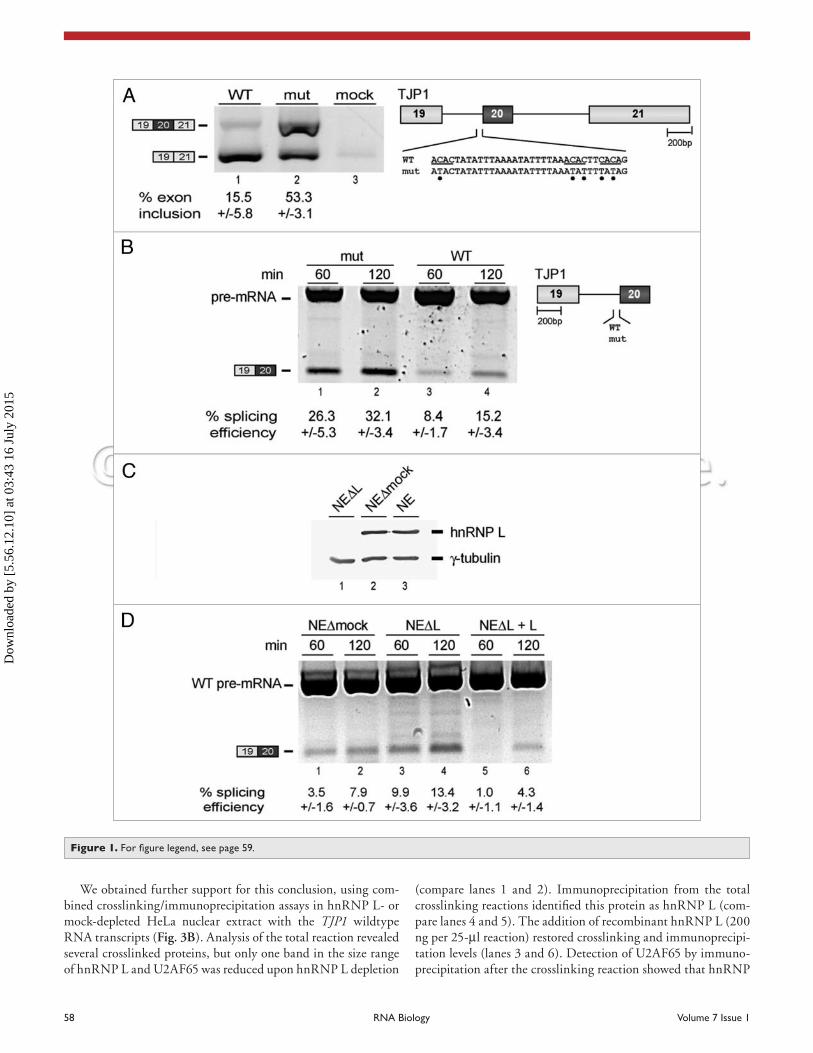
Identification and characterization of an intronic splicing silencer in the TJP1 gene. Based on a combined microarray and RNAi analysis, we have recently identified TJP1 as an hnRNP L target gene, with hnRNP L acting as a repressor on alterna-tive exon 20.18 TJP1 encodes the tight junction protein 1 of 225 kDa, which is located on the surface of cytoplasmic membranes in tight junctions and most likely is involved in signal transduc-tion. Interestingly, Willot and co-workers reported an alternative splicing event, involving exon 20 and resulting in two distinct isoforms, which may contribute to tight junction diversity.22
We focussed our mutational analysis on a small cluster of three high-score hnRNP L-binding motifs16 close to the 3' splice site of the regulated exon, using minigene constructs of the alter-native exon 20, the flanking exons 19 and 21, and the full-length introns (Fig. 1A). Five point mutations (C→T) were intro-duced to abolish hnRNP L binding. After transient transfection into HeLa cells, alternative splicing was analyzed by RT-PCR. Splicing of the TJP1 wildtype construct resulted predominantly in exon 19–21 mRNA with exon 20 skipped (lane 1), whereas the mutant derivative included exon 20 to more than 50% (lane 2). The mock control, done in the absence of transfected DNA, gave almost no products, demonstrating the specificity of the RT-PCR assay (lane 3). Taken together, we conclude that the CA-rich clus-ter in TJP1 intron 19 represents an intronic splicing silencer.
In addition, we analyzed the effect of the same mutation on TJP1 in vitro splicing in two-exon constructs comprised of exons 19 and 20 (Fig. 1B). With over 30%, the splicing efficiency after 120 min was significantly higher with the mutant deriva-tive (compare lanes 2 and 4) than with the wildtype (~15%). A control for DNA contamination (in the absence of reverse tran-scriptase) yielded no visible products (data not shown). In sum, these data support the in vivo splicing results and suggest that the CA-rich cluster in TJP1 intron 19 functions as an intronic splicing silencer.
To demonstrate that the effect observed directly depends on hnRNP L we used hnRNP L- or mock-depleted HeLa nuclear extract (Fig. 1C). HnRNP L was depleted from HeLa nuclear extract with a 5'-biotinylated (CA)
32 RNA oligonucleotide (lane
1);7 as control, a mock depletion was done in the absence of RNA oligonucleotide (lane 2). Western blot analysis using an hnRNP L-specific antibody confirmed efficient depletion from nuclear extract. TJP1 in vitro splicing was analyzed by RT-PCR (Fig. 1D). Compared to mock-depletion (lanes 1 and 2) the splic-ing efficiency for the two-exon TJP1 construct was significantly higher in hnRNP L-depleted nuclear extract (lanes 3 and 4). Adding back recombinant hnRNP L (200 ng per 13.25-μl reac-tion) strongly reduced splicing activity, establishing the repressor activity of hnRNP L in this case (lanes 5 and 6).
Next we used UV crosslinking to analyze hnRNP L binding to the CA-rich cluster (Fig. 2A). Baculovirus-expressed recom-binant hnRNP L was incubated with a short 32P-labeled TJP1 RNA containing the 3'-terminal 57 nucleotides of intron 19 (including branch point and CA-rich region) and 27 nucleotides of exon 20, followed by UV crosslinking, RNase A digestion, and
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5
58 RNA Biology Volume 7 Issue 1
(compare lanes 1 and 2). Immunoprecipitation from the total crosslinking reactions identified this protein as hnRNP L (com-pare lanes 4 and 5). The addition of recombinant hnRNP L (200 ng per 25-μl reaction) restored crosslinking and immunoprecipi-tation levels (lanes 3 and 6). Detection of U2AF65 by immuno-precipitation after the crosslinking reaction showed that hnRNP
We obtained further support for this conclusion, using com-bined crosslinking/immunoprecipitation assays in hnRNP L- or mock-depleted HeLa nuclear extract with the TJP1 wildtype RNA transcripts (Fig. 3B). Analysis of the total reaction revealed several crosslinked proteins, but only one band in the size range of hnRNP L and U2AF65 was reduced upon hnRNP L depletion
Figure 1. For figure legend, see page 59.
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5

www.landesbioscience.com RNA Biology 59
with the TJP1 mutant transcript showed no significant change in U2AF65 crosslinking efficiency upon addition of hnRNP L (data not shown). Taken together, our data indicate that hnRNP L impairs binding of U2AF65 to the polypyrimidine tract of TJP1
L depletion is associated with increased crosslinking of U2AF65 (compare lanes 7 and 8). With the addition of recombinant hnRNP L, this effect could be reversed, that is U2AF65 inter-action was reduced (lane 9). The same experiment carried out
Figure 1. cA-rich sequence acts as an hnRNp L-dependent splicing silencer on TJP1 exon 20 inclusion. (A) TJP1 alternative splicing in vivo. TJP1 mini-gene constructs were transfected into heLa cells (WT, wildtype, lane 1; mut, mutant, lane 2; see schematic of minigenes on the right, with the hnRNp L high-score motifs underlined and mutated nucleotides marked by dots); a control transfection (mock, lane 3) was carried out in the absence of DNA. Alternative splicing was analyzed by RT-pcR (positions of pcR products indicated on the left). percentages of exon inclusion with standard deviations (n = 4) are given below the corresponding lanes. (B) TJP1 splicing in vitro. TJP1 two-exon pre-mRNAs (WT, wildtype, lanes 3 and 4; mut, mutant, lanes 1 and 2; see schematic of pre-mRNAs on the right) were spliced in heLa nuclear extract for 60 or 120 min (as indicated), followed by RT-pcR analysis. Positions of PCR products are shown on the left. Splicing efficiencies with standard deviations (n = 3) are given below the corresponding lanes. (c) hnRNp L depletion. hnRNp L-depleted (NeΔL; lane 1), mock-depleted (NeΔmock; lane 2), and untreated nuclear extract (Ne; lane 3) were ana-lyzed by western blot with antibodies against hnRNp L, using γ-tubulin as an internal standard. (D) TJP1 splicing in vitro in hnRNp L-depleted extract. Wildtype (WT) TJP1 two-exon pre-mRNA was spliced in mock-depleted (NeΔmock; lanes 1 and 2), hnRNp L-depleted nuclear extract (NeΔL; lanes 3 and 4), or in hnRNp L-depleted extract complemented with recombinant hnRNp L (NeΔL + L; 200 ng per 13.25-μl reaction; lanes 5 and 6) for 60 or 120 min (as indicated), followed by RT-PCR analysis. Positions of PCR products are shown on the left. Splicing efficiencies with standard deviations (n = 3) are given below the corresponding lanes.
Figure 2. TJP1 pre-mRNA interacts with hnRNP L and U2AF65 with different efficiencies. (A) 32p-labeled TJP1 wildtype (WT) and mutant (mut) RNAs (84 nucleotides; schematically shown below) were UV-crosslinked in the absence or presence of recombinant hnRNp L (-/+hnRNp L; lanes 1–4), followed by sDs-pAGe. The arrow indicates the position of crosslinked hnRNp L. Molecular size markers (in kDa) are given on the left. (B) 32p-labeled TJP1 wildtype RNA (84 nucleotides) was incubated with increasing amounts of recombinant hnRNp L (0, 0.2, 0.4 and 0.8 μM; left, lanes 1–4) or U2AF65 (0, 5, 10 and 20 μM; middle, lanes 5–8). 32p-labeled MINX pre-mRNA was incubated with U2AF65 (0, 0.3, 1 and 4 μM; right; lanes 9–12). complexes were separated by native polyacrylamide gel electrophoresis. positions of RNA-protein complexes and free RNAs are indicated.
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5

60 RNA Biology Volume 7 Issue 1
intronic silencers upstream or downstream of the regulated exon (reviewed in ref. 26). Several models for PTB-dependent splicing repression have been proposed, depending on the position of its binding sites. For example, in the α-actinin and β-tropomyosin genes PTB represses use of an alternative exon by directly inter-fering with U2AF65 binding, leading to skipping of the regulated exon. These models are based on the presence of PTB binding sites within the polypyrimidine tract.27-32 Here we have demonstrated for the first time that hnRNP L can mediate splicing repression by a similar mechanism. With binding sites located within the polypyrimidine tract of the regulated exon, hnRNP L interferes with binding of U2AF65, resulting in impaired recruitment of the U2 snRNP and exon skipping. Although dispensable for splicing of AG-dependent introns that contain a strong polypyrimidine tract, it has been shown that the small subunit of U2AF is essen-tial for splicing of introns with weak polypyrimidine tracts.33,34
exon 20. Since binding sites for both proteins reside in the TJP1 3' splice site region, we suggest that silencing of TJP1 exon 20 is mediated by hnRNP L competing for U2AF65 binding; as a result, the U2 snRNP would not be recruited, and TJP1 exon 20 skipped.
Several cases are known where splicing regulators interfere with binding of U2AF to the 3' splice site, leading to exon skipping. The sex determination pathway in Drosophila melanogaster pro-vides a well studied example:2,24 In female flies, Sex lethal protein (Sxl) binds to the 3' splice site of exon 2 in the transformer (tra) pre-mRNA, promoting use of an alternative, female-specific 3' splice site by blocking U2AF65 recruitment. Further examples for interference with 3' splice site recognition are given by the polypy-rimidine tract binding protein (PTB/hnRNP I) which is closely related to hnRNP L:25 PTB is involved in the repression of several tissue-specific exons by preferentially binding to pyrimidine-rich
Figure 3. hnRNp L interferes with binding of U2AF65 to TJP1 pre-mRNA. (A) 32p-labeled TJP1 wildtype RNA (318 nucleotides, see schematic representation below) was UV-crosslinked in the absence of protein (lane 1), or in the presence of either recombinant U2AF65 (lane 2), recombinant hnRNp L (lane 3), or both proteins (lane 4). After RNase A digestion, crosslinked proteins were analyzed by sDs-pAGe. The positions of crosslinked proteins are indicated on the right. Molecular size markers (in kDa) are given on the left. (B) 32p-labeled TJP1 RNA (318 nucleotides, as in A) was UV-crosslinked (XL) in mock-depleted (NeΔmock; lane 1), hnRNp L-depleted nuclear extract (NeΔL; lane 2), or after complementation of hnRNp L-depleted extract with recombinant hnRNp L (NeΔL + L; 200 ng per 25 μl; lane 3), followed by sDs-pAGe. In addition, UV crosslinking was combined with immunoprecipitations (IP), using anti-hnRNP L (lanes 4–6) or anti-U2AF65 antibodies (lanes 7–9). Note that the crosslinked hnRNP L is slightly larger than the endogenous protein (compare lanes 4 and 6), due to the his-tag present in the recombinant protein. The position of the hnRNp L crosslink is marked by an arrow. Molecular size markers (in kDa) are given on the left.
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5

www.landesbioscience.com RNA Biology 61
Considering the suboptimal polypyrimidine tract of TJP1 intron 19, impaired U2AF35 recruitment to the 3' splice site may also contribute to hnRNP L-mediated splicing repression.
HnRNP L-dependent splicing silencer mediates skipping of SLC2A2 exon 4. As one of our first hnRNP L target genes we had identified SLC2A2, where in intron 4 a CA-repeat element starting at position +7 acts as splicing silencer of the variable exon 4 in vivo.16 SLC2A2 (solute carrier family 2) encodes a glucose transporter-like protein, which is an integral glycoprotein of the plasma membrane, mediating facilitated bidirectional transfer of glucose.35,36 Due to its low affinity to glucose, the protein may be part of a glucose-sensing mechanism in pancreatic islet β-cells. To study the hnRNP L mechanism in this case, we established an in vitro alternative splicing assay, based on an SLC2A2 three-exon pre-mRNA containing the variable exon 4 and the flank-ing exons 3 and 5, with both introns shortened, in a wildtype (WT ) and mutated version (mut; CA
19 substituted by unspecific
sequence; see Fig. 4A). SLC2A2 WT and mut pre-mRNAs were spliced in vitro, followed by RT-PCR assays. After 240 min, we detected around 89% exon inclusion for the mutant derivative (lane 6), but only around 42% for the wildtype pre-mRNA (lane 3), indicating a silencer function for the intronic CA
19 ele-
ment and consistent with our previous results based on minigene transfections.16
To investigate whether splicing repression of SLC2A2 exon 4 depends on hnRNP L, we performed depletion/complementa-tion experiments in the in vitro system. SLC2A2 wildtype pre-mRNA was spliced in hnRNP L-depleted or mock-depleted HeLa nuclear extract, and alternative splicing was analyzed by RT-PCR (Fig. 4B). In mock-depleted nuclear extract, exon 4 was included to levels of about 52%, which increased to over 70% in hnRNP L-depleted extract (lanes 1 and 2). The addition of hnRNP L protein (100, 200, 300 ng per 25-μl splicing reaction) to the depleted extract restored exon inclusion levels in a dose-dependent manner (lanes 3–5). Note that 200 ng of hnRNP L per 25-μl splicing reaction correspond to the normal level in HeLa nuclear extract.18 We conclude that the CA-repeat splicing silencer function in SLC2A2 is mediated by hnRNP L, which acts as a repressor of exon 4 inclusion.
HnRNP L binding to intronic splicing silencer inter-feres with 5' splice site recognition of SLC2A2 exon 4. Next, we addressed the mechanism of splicing repression by hnRNP L. Because of the close proximity of the CA-repeat silencer element to the regulated 5' splice site, we tested by psoralen crosslinking whether hnRNP L might interfere with 5' splice site recognition by the U1 snRNP (Fig. 4C). We used an SLC2A2 RNA of 101 nucleotides comprised of part of exon 4, its 5' splice site and the CA-repeat element, in comparison with a mutant derivative in which the CA
19 sequence was substituted. After incubation in
HeLa nuclear extract under splicing conditions, UV crosslink-ing was induced, followed by RNA preparation and denaturing PAGE analysis. U1/SLC2A2 RNA crosslinks were identified by RNase H cleavage, comparing RNA from untreated extract (lanes 5 and 6), or from extract after RNase H cleavage by a U1-specific (ΔU1; lanes 9 and 10), and as a control, a U6-specific DNA oligonucleotide (ΔU6; lanes 7 and 8). As an additional
control, HeLa nuclear extract was used, in which U1 snRNA had been cleaved by RNase H; no crosslinked bands were observed in this case (lanes 3 and 4).
Significantly, wildtype and mutant SLC2A2 RNAs differed in their crosslinking efficiencies (compare lanes 5 and 6, or 7 and 8, or 9 and 10): We consistently observed more efficient crosslinking with the mutant derivative than with the wildtype RNA, supporting the idea that hnRNP L interferes with 5' splice site recognition by the U1 snRNP. Furthermore, crosslinking of wildtype SLC2A2 RNA in mock- or hnRNP L-depleted extract confirmed that the observed effect is hnRNP L-dependent: The U1/SLC2A2 RNA crosslink was more prominent in hnRNP L-depleted extract than in mock-depleted extract (compare lanes 11 and 12). We conclude that hnRNP L bound to the 5' splice site-associated silencer in SLC2A2 intron 4 represses the recog-nition of that 5' splice site, thereby regulating exon 4 inclusion. Regulation of alternative splicing is often mediated by promo-tion or inhibition of early events in spliceosome assembly such as 5' or 3' splice site recognition. Another member of the hnRNP protein family, hnRNP A1, was previously shown to interfere with U1 snRNP binding through competition for binding to the pre-mRNA.37 In contrast, PTB represses splicing in the major-ity of cases by interacting with multiple binding sites flanking the regulated exon.26 Current models suggest that PTB proteins can either multimerize across long regions of silencing, or alter-natively, can in its monomeric state bridge flanking intronic binding sites, thereby interacting across the regulated exon.38-40 As shown for the c-src alternative N1 exon, PTB does not affect U1 snRNP binding, but instead interferes with the 5' splice site-dependent recruitment of U2AF65.41 For hnRNP L-mediated splicing regulation, we could demonstrate here that hnRNP L specifically functions on either the 5' or the 3' splice site of a regulated exon, thereby showing versatile regulatory actions like other members of the hnRNP family. In contrast to some models proposed for splicing regulation by PTB, we did not observe any cross-exon interaction of hnRNP L proteins in the cases of TJP1 and SLC2A2.
Taken together, we gained new insights into the mechanism of hnRNP L-dependent splicing repression, using two target genes as examples. In these cases hnRNP L specifically interferes with early events in spliceosome assembly, namely 5' or 3' splice site recognition, depending on the intronic position of the hnRNP L binding sites relative to the regulated exon.
Materials and Methods
Minigene constructs. TJP1 and SLC2A2 minigene constructs consist of three exons and two introns. The TJP1 genomic sequence for the minigene constructs was amplified using genomic DNA isolated from primary HUVEC cells as template (kindly provided by Karl Stangl, Charité, Berlin) and oligonucleotides TJP-C2/2, followed by cloning into pcDNA3 (Invitrogen). A TJP1 mutant derivative was constructed by PCR, using oligonu-cleotides T7 and TJP-C1, which introduced five point mutations (see Fig. 1A). The PCR product was digested with HindIII, fol-lowed by cloning into the wildtype minigene, thereby substituting
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5

62 RNA Biology Volume 7 Issue 1
Figure 4. hnRNp L-dependent splicing repression of SLC2A2 exon 4 is based on impaired 5' splice site recognition. (A) Intronic cA-repeat as silencer element for SLC2A2 exon 4 repression. The three-exon SLC2A2 pre-mRNAs (WT, wildtype; mut, mutant; see schematic of minigenes on the left) were spliced in vitro for the times indicated (0, 120, 240 min), followed by RT-pcR assays. The positions of pcR products corresponding to pre-mRNA and splice variants are schematically shown on the right. The percentages of exon inclusion with standard deviations (n = 3) are given below the cor-responding lanes. (B) hnRNp L requirement for SLC2A2 exon 4 repression. The three-exon SLC2A2 pre-mRNA was spliced in vitro for 240 min in mock-depleted (NeΔmock; lane 1), in hnRNp L-depleted nuclear extract (NeΔL; lane 2), or in hnRNp L-depleted nuclear extract complemented with recombinant hnRNp L (NeΔL + L; 100, 200 and 300 ng hnRNp L per 25 μl-splicing reaction; lanes 3–5). Alternative splicing was assayed by RT-pcR, with designation of pcR products and quantitation as described for (A). (c) 32p-labeled SLC2A2 wildtype (WT) or mutant (mut) RNAs (101 nucle-otides) were incubated without heLa nuclear extract (input; lanes 1 and 2), under splicing conditions in U1-inactivated nuclear extract (NeΔU1; lanes 3 and 4), or in normal nuclear extract (NE, lanes 5 and 6); to identify crosslinked products, purified RNAs were treated with RNase H and a DNA oligonucleotide complementary to U1 snRNA (ΔU1; lanes 9 and 10), or as control, to U6 snRNA (ΔU6; lanes 7 and 8). In addition, crosslinking was done in mock-depleted (NeΔmock; lane 11) or hnRNp L-depleted extract (NeΔL; lane 12). RNAs were analyzed by denaturing pAGe. The positions of free RNAs and crosslinked species (RNA/U1) are indicated. In addition, the experimental outline is schematically represented on the right.
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5

www.landesbioscience.com RNA Biology 63
DNA oligonucleotide at 50 ng/μl in a 20-μl reaction. DNA oli-gonucleotides used for RNase H treatment: U1 1–14, U1 124–140, U6 26–43, directed against nucleotides 1–14 or 124–140 of U1 snRNA, or nucleotides 26–43 of U6 snRNA. RNAs were analyzed by denaturing polyacrylamide electrophoresis and autoradiography.
Bandshift assay. Increasing amounts of recombinant His-hnRNP L (0.2, 0.4, 0.8 μM; Fig. 2B, left) or GST-U2AF65 (5, 10, 20 μM; Fig. 2B, middle; kindly provided by Juan Valcárcel, Barcelona) were incubated with 32P-labeled TJP1 RNAs (100 nM for His-hnRNP L or 30 nM for GST-U2AF65, respectively). In addition, 30 nM of 32P-labeled MINX pre-mRNA44 was incu-bated with GST-U2AF65 (0.3, 1, 4 μM; Fig. 2B, right). The reactions were adjusted to a final volume of 10 μl with buffer D (20 mM HEPES pH 8.0, 100 mM KCl, 100 mM MgCl
2, 0.5 M
EDTA, 20% (v/v) glycerol, 1 mM DTT) and contained 300 ng/μl tRNA and 2 U/μl RNase inhibitor. After incubation on ice for 30 min, reactions were separated on a 6% native gel (6% acrylam-ide/bisacrylamide 80:1, 1xTBE), followed by autoradiography.
UV crosslinking and immunoprecipitation. 32P-labeled TJP1 RNA (20,000 cpm) was incubated at 4°C for 30 min in a final volume of 10 μl buffer D containing 1.5 mM ATP, 4.1 mM MgCl
2, 30 mM creatine phosphate, 300 ng/μl tRNA, and 600
ng His-hnRNP L or GST-U2AF65 (Fig. 3A). As negative control, RNA was incubated alone without the addition of recombinant protein. UV crosslinking was carried out at 254 nm, followed by incubation with RNase A (1.5 μg/μl; 30 min at 37°C), SDS-PAGE, and autoradiography.
For UV crosslinking in nuclear extract, 32P-labeled TJP1 RNA (200,000 cpm) was incubated in a 25-μl reaction under splicing conditions (Fig. 3B). For complementation, 200 ng of recom-binant hnRNP L per 25-μl reaction was added to the hnRNP L-depleted extract. The splicing reactions were incubated at 30°C for 30 min and then irradiated with UV light (254 nm). Samples were treated with 60 ng/μl RNase A for 30 min at 37°C and then subjected to immunoprecipitation with either anti-hnRNP L monoclonal antibody (4D11; Sigma) or anti-U2AF65 mono-clonal antibody (MC3; Sigma), followed by SDS-PAGE and phosphorimager visualization (Bio-Rad).
Acknowledgements
We thank Juan Valcárcel (CRG, Barcelona) for gifts of U2AF65 antibodies and GST-U2AF65 protein, Joao Paulo Tavanez (CRG, Barcelona) for essential help with the UV crosslink-ing and immunoprecipitation procedure, and members of our group for discussions. This work was supported by grants from the Deutsche Forschungsgemeinschaft (DFG grants Bi 316/10 and 316/12, to A.B.), the Federal Ministry for Education and Research (BMBF NGNF-2 program, to A.B.), the European-Commission-funded Network of Excellence EURASNET, to A.B.), and from the Shanghai Municipal Council for Science and Technology (09PJ1411000, to J.H.).
the HindIII fragment. Construction of the SLC2A2 wildtype minigene construct was described by Hui et al.16 The SLC2A2 mutations were introduced by a series of PCRs, using oligonucle-otides SLC2A2-C1 and -C2 in combination with M-SLC2A2 fwd and rev. The whole fragment, including restriction sites, was obtained by a second-step PCR, using the previous PCR prod-ucts as templates, and inserted into pcDNA3. As a result, the CA repeat was replaced by a random sequence: 5'-GCA TTA TTG CAT TTC ATT CTG AAG CAG TCC AAT GAC TA-3'.
Cell transfection, RNA isolation, RT-PCR analysis. One day before transfection, 6 x 105 HeLa cells were seeded in 6-cm dishes. For transfection, cells were less than 80% confluent. 8 μg of the respective minigene construct was transfected by the calcium phosphate method. Two days after transfection cells were harvested and total RNA was isolated, using guanidium thiocyanate. Reverse transcriptions were carried out by Expand RT (Roche), using vector-specific BGH rev primer. The resulting cDNA was amplified by PCR using oligonucleotides against the flanking exons. In the case of TJP1, the BGH rev primer was also used for PCR.
In vitro transcription and splicing. pcDNA3-SLC2A2 minigenes were cut with XhoI, and transcribed by T7 RNA polymerase. TJP1 templates were obtained by PCR. 10 ng of pre-mRNA was incubated in a 25-μl reaction containing 60% HeLa nuclear extract.42 Splicing was analyzed by RT-PCR as described before using gene-specific primers. DNA templates for 32P-labeled in vitro transcription were obtained by PCR using pcDNA3-SLC2A2 plasmid DNAs as template and oligonucle-otides T7-SLC2A2s fwd and SLC2A2s rev. The shorter TJP1 PCR product for T7 transcription of an RNA of 84 nucleotides was obtained by the oligonucleotide combination T7-TJP-1 fwd/TJP-22 rev, whereas for the longer product of 318 nucle-otides TJP-33 rev was used as reverse primer. Transcription reac-tions were carried out as described above, except that [α-32P]ATP was used.
Depletion of hnRNP L from HeLa nuclear extract. HnRNP L was depleted from nuclear extract with a 5'-biotinylated (CA)
32
RNA oligonucleotide described by Hui et al.7 Depletion of hnRNP L was analyzed by western blot using anti-hnRNP L polyclonal peptide antibody (D-17; Santa Cruz Biotechnology) and anti-γ-tubulin monoclonal antibody (GTU-88; Sigma) as a loading control. As control, a mock depletion was done in the absence of (CA)
32 oligonucleotide. Depleted extracts were com-
plemented by baculovirus-expressed hnRNP L.7
Psoralen crosslinking and RNase H cleavage. Psoralen cross-linking was done according to Wassarman,43 with the follow-ing modifications: 32P-labeled splicing substrate SLC2A2 RNA (100,000 cpm) was incubated in 60% HeLa nuclear extract under splicing conditions (Fig. 4C). Psoralen was added to a final concentration of 20 μg/ml and crosslinking carried out at 350 nm. RNA was purified after psoralen crosslinking or directly from HeLa nuclear extract, followed by RNase H digestion with
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5

64 RNA Biology Volume 7 Issue 1
32. Spellman R, Rideau A, Matlin A, Gooding C, Robinson F, McGlincy N, et al. Regulation of alternative splicing by PTB and associated factors. Biochem Soc Trans 2005; 33:457-60.
33. Pacheco TR, Coelho MB, Desterro JMP, Mollet I, Carmo-Fonseca M. In vivo requirement of the small subunit of U2AF for recognition of a weak 3' splice site. Mol Cell Biol 2006; 26:8183-90.
34. Wu S, Romfo CM, Nilsen TW, Green MR. Functional recognition of the 3' splice site AG by the splicing fac-tor U2AF35. Nature 1999; 402:832-5.
35. Fukumoto H, Seino S, Imura H, Seino Y, Eddy RL, Fukushima Y, et al. Sequence, tissue distribution, and chromosomal localization of mRNA encoding a human glucose transporter-like protein. Proc Natl Acad Sci USA 1988; 85:5434-8.
36. Thorens B, Sarkar HK, Kaback HR, Lodish HF. Cloning and functional expression in bacteria of a novel glucose transporter present in liver, intestine, kidney and beta-pancreatic islet cells. Cell 1988; 55:281-90.
37. Eperon IC, Makarova OV, Mayeda A, Munroe SH, Caceres JF, Hayward DG, et al. Selection of alternative 5' splice sites: role of U1 snRNP and models for the antagonistic effects of SF2/ASF and hnRNP A1. Mol Cell Biol 2000; 20:8303-18.
38. Chou MY, Underwood JG, Nikolic J, Luu MH, Black DL. Multisite RNA binding and release of polypyrimi-dine tract binding protein during the regulation of c-src neural-specific splicing. Mol Cell 2000; 5:949-57.
39. Oberstrass FC, Auweter SD, Erat M, Hargous Y, Henning A, Wenter P, et al. Structure of PTB bound to RNA: specific binding and implications for splicing regulation. Science 2005; 309:2054-7.
40. Spellman R, Smith CW. Novel modes of splicing repression by PTB. Trends Biochem Sci 2006; 31:73-6.
41. Sharma S, Falick AM, Black DL. Polypyrimidine tract binding protein blocks the 5' splice site-dependent assembly of U2AF and the prespliceosomal E complex. Mol Cell 2005; 19:485-96.
42. Bindereif A, Green MR. An ordered pathway of snRNP binding during mammalian pre-mRNA splicing com-plex assembly. EMBO J 1987; 6:2415-24.
43. Wassarman DA. Psoralen crosslinking of small RNAs in vitro. Molecular Biology Reports 1993; 17:143-51.
44. Zillmann M, Zapp ML, Berget SM. Gel electropho-retic isolation of splicing complexes containing U1 small nuclear ribonucleoprotein particles. Mol Cell Biol 1988; 8:814-21.
17. Waterston RH, Lindblad-Toh K, Birney E, Rogers J, Abril JF, Agarwal P, et al. Initial sequencing and com-parative analysis of the mouse genome. Nature 2002; 420:520-62.
18. Hung LH, Heiner M, Hui J, Schreiner S, Benes V, Bindereif A. Diverse roles of hnRNP L in mammalian mRNA processing: a combined microarray and RNAi analysis. RNA 2008; 14:284-96.
19. House AE, Lynch KW. An exonic splicing silencer represses spliceosome assembly after ATP-dependent exon recognition. Nat Struct Mol Biol 2006; 13:937-44.
20. Rothrock CR, House AE, Lynch KW. HnRNP L represses exon splicing via a regulated exonic splicing silencer. EMBO J 2005; 24:2792-802.
21. Rossbach O, Hung L-H, Schreiner S, Grishina I, Heiner M, Hui J, et al. Auto- and cross-regulation of the hnRNP L proteins by alternative splicing. Mol Cell Biol 2009; 29:1442-51.
22. Willott E, Balda MS, Heintzelman M, Jameson B, Anderson JM. Localization and differential expression of two isoforms of the tight junction protein ZO-1. Am J Physiol Cell Physiol 1992; 262:1119-24.
23. Yeo G, Burge CB. Maximum entropy modeling of short sequence motifs with applications to RNA splic-ing signals. Journal of Computational Biology 2004; 11:377-94.
24. Valcarcel J, Singh R, Zamore PD, Green MR. The pro-tein Sex-lethal antagonizes the splicing factor U2AF to regulate alternative splicing of transformer pre-mRNA. Nature 1993; 362:171-5.
25. Kim JH, Hahm B, Kim YK, Choi M, Jang SK. Protein-protein interaction among hnRNPs shuttling between nucleus and cytoplasm. J Mol Biol 2000; 298:395-405.
26. Wagner EJ, Garcia-Blanco MA. Polypyrimidine tract binding protein antagonizes exon definition. Mol Cell Biol 2001; 21:3281-8.
27. Amir-Ahmady B, Boutz PL, Markovtsov V, Phillips ML, Black DL. Exon repression by polypyrimidine tract binding protein. RNA 2005; 11:699-716.
28. Lin CH, Patton JG. Regulation of alternative 3' splice site selection by constitutive splicing factors. RNA 1995; 1:234-45.
29. Matlin AJ, Southby J, Gooding C, Smith CWJ. Repression of alpha-actinin SM exon splicing by assisted binding of PTB to the polypyrimidine tract. RNA 2007; 13:1214-23.
30. Sauliere J, Sureau A, Expert-Bezancon A, Marie J. The polypyrimidine tract binding protein (PTB) represses splicing of exon 6B from the beta-tropomyosin pre-mRNA by directly interfering with the binding of the U2AF65 subunit. Mol Cell Biol 2006; 26:8755-69.
31. Singh R, Valcarcel J, Green MR. Distinct bind-ing specificities and functions of higher eukaryotic polypyrimidine tract-binding proteins. Science 1995; 268:1173-6.
References1. Wang ET, Sandberg R, Luo S, Khrebtukova I, Zhang L,
Mayr C, et al. Alternative isoform regulation in human tissue transcriptomes. Nature 2008; 456:470-6.
2. Black DL. Mechanism of alternative pre-messenger RNA splicing. Annual Review of Biochemistry 2003; 72:291-336.
3. Blencowe BJ. Alternative splicing: new insights from global analyses. Cell 2006; 126:37-47.
4. Cartegni L, Chew SL, Krainer AR. Listening to silence and understanding nonsense: exonic mutations that affect splicing. Nat Rev Genet 2002; 3:285-98.
5. Modrek B, Lee C. A genomic view of alternative splic-ing. Nat Genet 2002; 30:13-9.
6. Smith CWJ, Valcárcel J. Alternative pre-mRNA splic-ing: the logic of combinatorial control. Trends in Biochemical Sciences 2000; 25:381-8.
7. Hui J, Stangl K, Lane WS, Bindereif A. HnRNP L stimulates splicing of the eNOS gene by binding to variable-length CA repeats. Nat Struct Biol 2003; 10:33-7.
8. Guang S, Felthauser AM, Mertz JE. Binding of hnRNP L to the pre-mRNA processing enhancer of the her-pes simplex virus thymidine kinase gene enhances both polyadenylation and nucleocytoplasmic export of intronless mRNAs. Mol Cell Biol 2005; 25:6303-13.
9. Liu X, Mertz JE. HnRNP L binds a cis-acting RNA sequence element that enables intron-dependent gene expression. Genes Dev 1995; 9:1766-80.
10. Hahm B, Kim YK, Kim JH, Kim TY, Jang SK. Heterogeneous nuclear ribonucleoprotein L interacts with the 3' border of the internal ribosomal entry site of Hepatitis C virus. J Virol 1998; 72:8782-8.
11. Hwang B, Lim JH, Hahm B, Jang SK, Lee S-W. HnRNP L is required for the translation mediated by HCV IRES. Biochemical and Biophysical Research Communications 2009; 378:584-8.
12. Hamilton BJ, Wang XW, Collins J, Bloch D, Bergeron A, Henry B, et al. Separate cis-trans pathways post-transcriptionally regulate murine CD154 (CD40 ligand) expression: a novel function for CA repeats in the 3'-untranslated region. J Biol Chem 2008; 283:25606-16.
13. Hui J, Reither G, Bindereif A. Novel functional role of CA repeats and hnRNP L in RNA stability. RNA 2003; 9:931-6.
14. Shih SC, Claffey KP. Regulation of human vascular endothelial growth factor mRNA stability in hypoxia by heterogeneous nuclear ribonucleoprotein L. J Biol Chem 1999; 274:1359-65.
15. Ray PS, Jia J, Yao P, Majumder M, Hatzoglou M, Fox PL. A stress-responsive RNA switch regulates VEGFA expression. Nature 2009; 457:915-9.
16. Hui J, Hung LH, Heiner M, Schreiner S, Neumuller N, Reither G, et al. Intronic CA-repeat and CA-rich elements: a new class of regulators of mammalian alter-native splicing. EMBO J 2005; 24:1988-98.
Dow
nloa
ded
by [
5.56
.12.
10]
at 0
3:43
16
July
201
5




















