
High Altitude Adaptation in Mammals
10
AMER. ZOOL., 13:447-456 (1973). High Altitude Adaptation in Mammals CLAUDE LENFANT National Heart and Lung Institute, National Institutes of Health, Bethesda, Maryland 20014 SYNOPSIS. The physiological, morphological, and biochemical characteristics of several species of mammals resident at high altitude are compared with those of their sea level counterparts. The differences noted in these characteristics are in a direction that facilitates the acclimatization of those living at high altitude. The differences among species point to the fact that the mechanism of adaptation to altitude (i.e., hypoxia) is still not understood. This review emphasizes that the adaptive process is complex and made up of several components, that these components are inter- related, and that neither the physiological nor morphological adaptations can fully account for the tolerance to hypoxia. Although only superficially studied as yet, the biochemical adaptations appear most important. INTRODUCTION "Altitude is a component of the physical environment to which animals show ad- justments" (Hall et al., 1936). Indeed, be- cause increasing altitude is characterized by decreasing oxygen availability in the en- vironment without major changes in tis- sue (i.e., metabolic) requirements, any ani- mal exposed to altitude must develop adaptive mechanisms if it is to maintain an adequate oxygen delivery to its tissue. Although it is recognized that numerous species of mammals have evolved at alti- tude or have adapted to it, there are rela- tively few studies of their adaptive mech- anisms in species other than man. The first comprehensive study of man's physio- logical response to altitude was in 1874 (Bert, 1874). The first studies of various other animals' response seem to be those of Muntz and of Viauk (Muntz, 1891; Viault, 1891). These studies, however, were very much limited since they were ex- clusively concerned with changes in hemo- globin concentration at altitude. "Com- parative physiology in high altitudes" (Hall et al., 1936) is probably the next landmark in our understanding of adaptation to al- titude. Hall's study stimulated considerable and increasing interest in a comparative approach to this important biological phenomenon (Chiodi, 1962, 1970; Reeves et al., 1963a; Banchero et al., 1971a; Lahiri et al., 1971). This presentation reviews and compares the physiological, morphological, and bio- chemical adaptations of various species of mammals to altitude. PHYSIOLOGICAL ADAPTATIONS Oxygen is transported from the environ- ment to the cells by means of four linked mechanisms (Lenfant and Sullivan, 1971): ventilation, pulmonary diffusion, circula- tion, and tissue diffusion. Upon ascent to, and during life at, altitude there are sev- eral physiological adjustments of these mechanisms that compensate for the de- crease in availability of environmental oxygen. Although all four mechanisms play an equally important role, not all of them have been examined in species other than man. For this comparative review, the physiological responses to altitude will be grouped into respiratory, circulatory, and hematological adaptation. Respiratory adaptations In all mammalian species, minute ven- tilation is determined by oxygen demand, and regulated by neural and chemical stim- uli. One of the latter is the partial pres- sure of oxygen, a decrease of which is char- acteristic of altitude. Hence, it is no sur- prise that in man (Kellogg, 1968), as well as in all other species of mammals that have been examined, there is a significant increase in ventilation upon acute ex- posure (Figs. 1, 2, 3). Figure 1 shows the in- 447 Downloaded from https://academic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of High Altitude Adaptation in Mammals

AMER. ZOOL., 13:447-456 (1973).
High Altitude Adaptation in Mammals
CLAUDE LENFANT
National Heart and Lung Institute, National Institutes of Health,Bethesda, Maryland 20014
SYNOPSIS. The physiological, morphological, and biochemical characteristics of severalspecies of mammals resident at high altitude are compared with those of their sealevel counterparts. The differences noted in these characteristics are in a directionthat facilitates the acclimatization of those living at high altitude. The differencesamong species point to the fact that the mechanism of adaptation to altitude (i.e.,hypoxia) is still not understood. This review emphasizes that the adaptive processis complex and made up of several components, that these components are inter-related, and that neither the physiological nor morphological adaptations can fullyaccount for the tolerance to hypoxia. Although only superficially studied as yet,the biochemical adaptations appear most important.
INTRODUCTION
"Altitude is a component of the physicalenvironment to which animals show ad-justments" (Hall et al., 1936). Indeed, be-cause increasing altitude is characterizedby decreasing oxygen availability in the en-vironment without major changes in tis-sue (i.e., metabolic) requirements, any ani-mal exposed to altitude must developadaptive mechanisms if it is to maintainan adequate oxygen delivery to its tissue.
Although it is recognized that numerousspecies of mammals have evolved at alti-tude or have adapted to it, there are rela-tively few studies of their adaptive mech-anisms in species other than man. Thefirst comprehensive study of man's physio-logical response to altitude was in 1874(Bert, 1874). The first studies of variousother animals' response seem to be thoseof Muntz and of Viauk (Muntz, 1891;Viault, 1891). These studies, however, werevery much limited since they were ex-clusively concerned with changes in hemo-globin concentration at altitude. "Com-parative physiology in high altitudes" (Hallet al., 1936) is probably the next landmarkin our understanding of adaptation to al-titude. Hall's study stimulated considerableand increasing interest in a comparativeapproach to this important biologicalphenomenon (Chiodi, 1962, 1970; Reeveset al., 1963a; Banchero et al., 1971a; Lahiriet al., 1971).
This presentation reviews and comparesthe physiological, morphological, and bio-chemical adaptations of various species ofmammals to altitude.
PHYSIOLOGICAL ADAPTATIONS
Oxygen is transported from the environ-ment to the cells by means of four linkedmechanisms (Lenfant and Sullivan, 1971):ventilation, pulmonary diffusion, circula-tion, and tissue diffusion. Upon ascent to,and during life at, altitude there are sev-eral physiological adjustments of thesemechanisms that compensate for the de-crease in availability of environmentaloxygen. Although all four mechanismsplay an equally important role, not all ofthem have been examined in species otherthan man. For this comparative review, thephysiological responses to altitude will begrouped into respiratory, circulatory, andhematological adaptation.
Respiratory adaptations
In all mammalian species, minute ven-tilation is determined by oxygen demand,and regulated by neural and chemical stim-uli. One of the latter is the partial pres-sure of oxygen, a decrease of which is char-acteristic of altitude. Hence, it is no sur-prise that in man (Kellogg, 1968), as wellas in all other species of mammals thathave been examined, there is a significantincrease in ventilation upon acute ex-posure (Figs. 1, 2, 3). Figure 1 shows the in-
447
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

448 CLAUDE LENFANT
VENTILATION CHANGE AT ALTITUDE(SOJOURNERSI
(MAN)
PROLONGED EXPOSURE|MORE THAN 7 DAYS)
ACUTE EXPOSUREILES5 THAN 2 DAVS)
CALF4900m
MAN4710m
LLAMA3420m
FIG. 1. Change in total ventilation after acuteand prolonged exposure to altitude. Data sourcesare as follows: man: Torrance et al. (1970/71) ,llama: Banchero et al. (1971a) ; goat: Lahiri et al.(1971) ; calf: Bisgard et al. (1969) ; lamb: Reeveset al. (1963d) .
crease in total ventilation noted in severalspecies upon acute exposure and after asojourn of at least several days. This figurealso shows that the magnitude of the in-crease varies among species and that it isgreatest immediately after ascent. Since thedemand for oxygen is the determining fac-tor in ventilation, it is possible that thespecies difference in ventilation increasemay in part be accounted for by a changein the oxygen requirement in some of thesespecies. The data shown in Figure 2 con-firm this possibility since the species differ-ences per unit of oxygen consumed areless than the species difference in total ven-tilation.
Whether the duration o£ exposure tohypoxia affects the magnitude of the vcn-
VE/VO2 CHANGE AT ALTITUDE(SOJOURNERS = MORE THAN 7 DAYS)
50 r
s; 25 -
4710m
3735m
LAMB
3900m
LLAMA
3420mn STEER
3900mnFIG. '2. Change in ventilatory equivalent at alti-tude. Data from same sources as in Figure 1, plusthat for steer from Grover et al. (1963).
M0R0C0CHA
LaOROYA J
SOJOURNERS13 DAYS)
•» NATIVES
2000 3000
ALTITUDE, METERS
4000 5000
FIG. 3. Ventilatory equivalent as a function of al-titude in natives and in sojourners. Data fromTorrance et al. (1970/71).
tilatory response to hypoxia stimuli hasbeen a question of great interest for manyyears (Rahn and Otis, 1949). It is now wellestablished that in man the duration ofresidence at altitude markedly affects theventilatory response (Lenfant and Sullivan,1971). Figure 3 shows the difference in re-sponse between sojourners and natives atvarious altitudes. The data of Weil et al.(1971) (Fig. 4) further demonstrate the at-tenuation of the hypoxic response in resi-dents at altitude as compared with thoseat sea level and in natives compared withsojourners. Some studies have examinedwhether a similar attenuation exists inother species of mammals (Brooks and Ten-ney, 1968; Hornbein and Sorensen, 1969;Lahiri et al., 1971). The results of thesestudies are presented in Figure 5 whichshows the increase in total ventilation when
30
E 20
I.
(MAN)
SOJOURNERS
NATIVES
60 80 100
ALVEOLAR OXYGEN TENSION. mmHg
120
FIG. 4. Total ventilation as a function of alveolaroxygen tension in sea level residents (two controls)and in altitude natives and sojourners. Data fromWeil et al. (1971).
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022
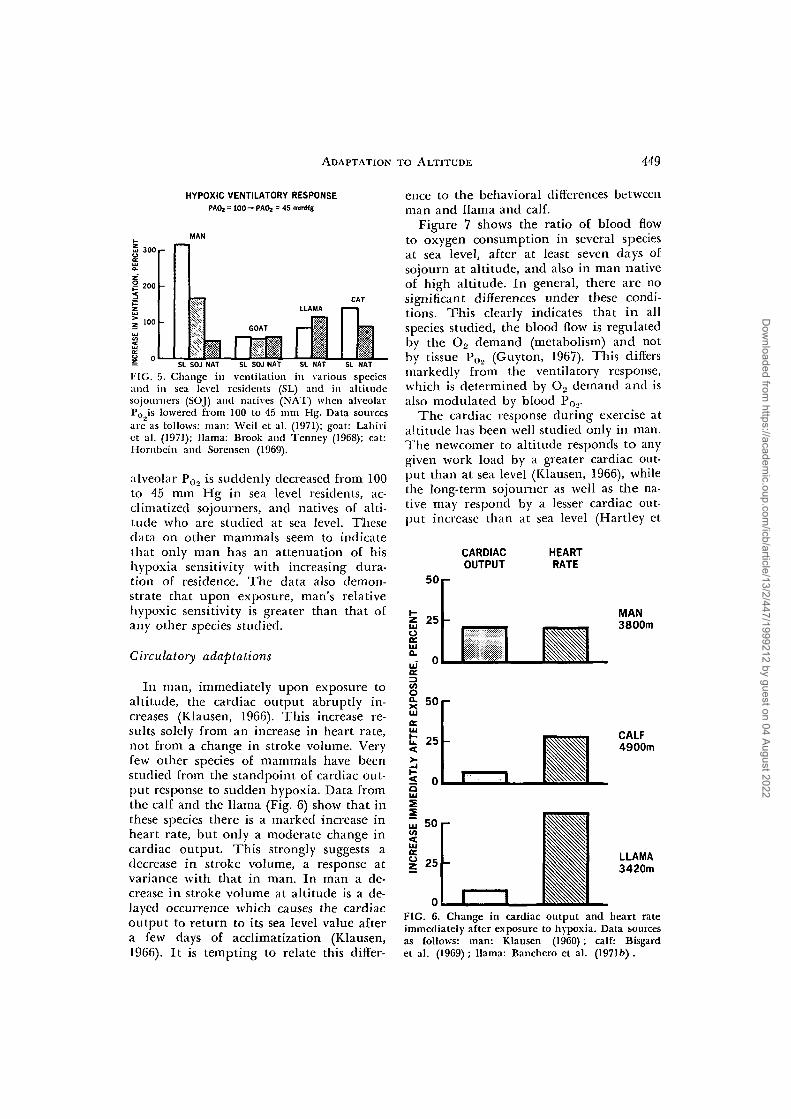
ADAPTATION TO ALTITUDE 449
HYPOXIC VENTILATORY RESPONSEPAOj = 100—PAO2 = 45 mmHg
SI SOJ NAT SL SOJ NAT SI NAT SL NAT
FIG. 5. Change in ventilation in various speciesand in sea level residents (SL) and in altitudesojourners (SOJ) and natives (NAT) when alveolarPo is lowered from 100 to 45 mm Hg. Data sourcesare as follows: man: Weil et al. (1971); goat: Lahirict al. (1971); llama: Brook and Tenney (1968); cat:Hornbein and Sorensen (1969).
alveolar Po, is suddenly decreased from 100to 45 mm Hg in sea level residents, ac-climatized sojourners, and natives of alti-tude who are studied at sea level. Thesedata on other mammals seem to indicatethat only man has an attenuation of hishypoxia sensitivity with increasing dura-tion of residence. The data also demon-strate that upon exposure, man's relativehypoxic sensitivity is greater than that ofany other species studied.
Circulatory adaptations
In man, immediately upon exposure toaltitude, the cardiac output abruptly in-creases (Klausen, 1966). This increase re-sults solely from an increase in heart rate,not from a change in stroke volume. Veryfew other species of mammals have beenstudied from the standpoint of cardiac out-put response to sudden hypoxia. Data fromthe calf and the llama (Fig. 6) show that inthese species there is a marked increase inheart rate, but only a moderate change incardiac output. This strongly suggests adecrease in stroke volume, a response atvariance with that in man. In man a de-crease in stroke volume at altitude is a de-layed occurrence which causes the cardiacoutput to return to its sea level value aftera few days of acclimatization (Klausen,1966). It is tempting to relate this differ-
ence to the behavioral differences betweenman and llama and calf.
Figure 7 shows the ratio of blood flowto oxygen consumption in several speciesat sea level, after at least seven clays ofsojourn at altitude, and also in man nativeof high altitude. In general, there are nosignificant differences under these condi-tions. This clearly indicates that in allspecies studied, the blood flow is regulatedby the O2 demand (metabolism) and notby tissue P02 (Guyton, 1967). This differsmarkedly from the ventilatory response,which is determined by O2 demand and isalso modulated by blood Po,.
The cardiac response during exercise ataltitude has been well studied only in man.The newcomer to altitude responds to anygiven work load by a greater cardiac out-put than at sea level (Klausen, 1966), whilethe long-term sojoumer as well as the na-tive may respond by a lesser cardiac out-put increase than at sea level (Hartley et
CARDIACOUTPUT
HEARTRATE
501-
5 25
§UJ
E 25<
MAN3800m
CALF4900m
50
u 25LLAMA3420m
FIG. 0. Change in cardiac output and heart rateimmediately after exposure to hypoxia. Data sourcesas follows: man: Klausen (1960) ; calf: Bisgardet al. (1969) ; llama: Banchero et al. (1971b) .
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

450 CLAUDE LENFANT
Q/VO2 CHANGE AT ALTITUDESOJOURNERS = MORE THAN 7 DAYS
30 1-
20
10
MAN4509m
LLAMA3420m
LAMB3900m
F10
CAT4300m
FIG. 7. Cardiac output to oxygen consumption ratioin various species and in sea level residents (SL)and in altitude sojourners (SOJ) and natives(NAT) . Sojourners are denned by more than 7
days stay. Data source as follows: man: Torranceet al. (1970/71); llama: Banchero et al. (1971b);lamb: Reeves et al. (1963a) ; steer: Grover et al.(1963) ; cat and rabbit: Reeves et al. (1963b).
al., 1967). A recent study on the llamaindicates that the cardiac output responseto a work load increases with increasingaltitude (Banchero et al., 19716).
The absence of an increase in restingcardiac output in long-term residents ataltitude is a distinct advantage in the sensethat there is no cardiac work increase. Theresult could, however, be a limitation inoxygen supply to those tissues that are sen-sitive to hypoxia. In man, this appears tobe prevented by a selective redistribution oflocal blood flow (Lenfant and Sullivan,1971). No similar observations have beenmade in other species.
The hemodynamics of pulmonary cir-culation in various species at altitude hasreceived considerable attention with regardto smooth muscle response to hypoxia. Inman, there is an increase in mean pul-monary artery pressure, the magnitude ofwhich is related to the altitude (Cruz-Jiboja et al., 1964). This results in a moreuniform distribution of blood flow to thelung at altitude (Fig. 8), and therefore, ina decrease in the ventilation to perfusionratio unevenness. This, in addition to theoverall increase of the mean alveolar ven-tilation to pulmonary capillary perfusionratio, enhances gas exchange in the lung
(Lenfant and Sullivan, 1971). Figure 9shows the pulmonary artery pressurechanges (or absence of change) in someother species of mammal. Only cattle seemto be experiencing a major increase inmean pulmonary artery pressure. Similarobservations have been made in cats, whilerabbits (Reeves et al., 19636), like the lamband llama shown in Figure 9, experience amuch milder degree of hypertension. Al-though some of this species difference is ex-plainable by smooth muscle distributionand thickness differences, some other fac-tors, such as autonomic regulatory mechan-isms, may play an important role (Wood-bury and Hamilton, 1941).
Hemalologic adaptations
The hematologic adaptations consideredin this review are blood oxygen-carryingcapacity and the hemoglobin affinity foroxygen. As noted earlier, the increase inoxygen-carrying capacity (i.e., increase inhemoglobin concentration) was first ob-served by Muntz (1891) and Viault (1891).This increase is the most consistent findingin all of the species of mammals living ataltitude. It is also to be found in speciesnot living at altitude but nevertheless ex-posed to severe hypoxia, such as the divingmammals whose adaptive characteristicsare extensively reported elsewhere (Len-fant et al., 1970; Kooyman, 1973).
Figure 10 illustrates the magnitude of
MAN
1000 2000 3000 4000
ALTITUDE. METERS
FIG. 8. Effect of altitude on mean pulmonary ar-tery pressure in relation to lung hilum. Data fromCruz-Jibaja et al. (1961) . (From Lenfant and Sulli-van, 1971. Reproduced with permission of NewEngland Journal of Medicine.)
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

ADAPTATION TO ALTITUDE 451
100
s 75
50
LAMB
...3?°.?.™ LLAMA
" " " 3 4 2 0 m "
1 2 3 4 5 6 7 8 9
TIME, WEEKS
FIG. 9. Mean pulmonary artery pressure as a func-tion of duration of stay at altitude. Data sourcesas follows: steer: Grover et al. (1963); lamb:Reeves et al. (1963a) ; llama: Banchero et al.(1971b) .
the increase in hemoglobin concentrationobserved in several species of mammals ac-climatized to altitude. In general, the rela-
20
15
10
5
10
5
- RABBIT
CHINCHILLA
ICAT
- STEER
-
M M
GOAT
SHEEP | —mmmm PH1
FIC. 10. Hemoglobin concentration in sea leveland altitude sojourner or native specimens of vari-ous species. Data sources as follows: rabbit andcat: Reeves et al. (19636) ; chinchilla: Hall (1965)and Chiodi (1962); steer: Grover et al. (1963);sheep: Hall et al. (1936); goat and llama: Chiodi(1970); dog: Lenfant (unpublished). Normal sealevel values for goat and dog are from Wintrobe(1949).
tive increase in hemoglobin concentrationis greater in natives of high altitude thanin sojourners. Figure 11 shows that in manit is closely related to altitude (or degreeof hypoxia) to which the subject is ex-posed. This figure also demonstrates thedifference between sojourners and natives.The increase in hemoglobin concentra-tion is undoubtedly a good index of im-paired oxygen delivery to the tissues. How-ever, it occurs quite slowly and, therefore,
25
NATIVES_
SOJOURNIRS
1000 2000 3000 4000
ALTITUDE, METERS
FIG. 11. Hemoglobin concentration as a functionof altitude in altitude native and sojourner man.Data from Torrance et al (1970/71) .
is not in contrast to the ventilatory andcirculatory responses—playing any role inthe adaptation to acute hypoxia states(Finch and Lenfant, 1971).
An increase in hemoglobin concentrationaugments the blood oxygen-carrying ca-pacity. However, in hypoxic hypoxia* therelative oxygen content of the blood andthe "loading" and "unloading" of oxygenare determined by the hemoglobin affinityfor oxygen. Hemoglobin affinity for oxy-gen is determined by the position of theoxygen dissociation curve; in turn, theposition of the curve is denned by the P50(7.40; 37 C) which is the partial pressure ofoxygen corresponding to 50% oxygensaturation of the hemoglobin at pH = 7.40and temperature = 37 C. Figure 12 showssome typical oxygen dissociation curvesfrom species of natives of high altitude andfrom other species of sea level residents.It can be seen that the latter have a mark-edly lower hemoglobin affinity for oxygen
* Hypoxic hypoxia is a state resulting from de-creased Po. in the alveolar air; for instance, at al-titude or in case of some respiratory diseases.
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

452 CLAUDE LENFANT
5 «o
NATIVESVICUNA
ALPACA
LLAMAGUANACO
SEA LEVELMAN
HORSEDOGRABBITPIG
PECCARYOX
SHEEP
pH = 7.4: T = 37 - 3 9 X
0 10 20 30 40 SO 60 70 80 90 100
OXYGEN TENSION. mniHi
FIG. 12. Typical oxygen dissociation causes of ani-mals native of high altitude and of animals resi-dent at sea level. Data from Hall et al. (1936) andChiodi (1970) .
(higher P50) than the former. The conse-quence of this difference in position of theoxygen dissociation curve is that at any al-titude (or Po,) the animal native to highaltitude has a higher oxygen saturationthan the sea level resident. This has beenconsidered by many as a distinct adaptivecharacteristic rather than a coincidence(Hall et al., 1936; Chiodi, 1962). This in-terpretation has been open to question bythe recent observation that human sea levelresidents, when exposed to altitude, exhibita rapid displacement of their oxygen dis-sociation curve to the right of the sea levelposition (Lenfant et al., 1969, 1970). Otherspecies, such as rat, rabbit, and dog, re-spond similarly, while still others, such assheep and goat, do not. These seeminglycontradictory observations provide evidencethat there is no simple way to achieve thebest adaptation to altitude, and also, thatno adaptive mechanism should be con-sidered independently of the other. This iswell demonstrated by Hall's findings (1966)that the hemoglobin affinity for O2 is re-lated to the critical P02 or the minimaltension at which utilizable oxygen can beremoved from the environment (Fig. 13).A low hemoglobin affinity for O2 may bethe best adaptive change in man who hassome specific physiological, morphological,and biochemical adaptive features. But ahigher affinity may be better for the vicuna,which has an entirely different set ofadaptive characteristics.
MORPHOLOGICAL ADAPTATIONS
There are relatively few studies con-cerned with the morphological adaptationto altitude. Three facets have, however,been considered: body morphology, lungdevelopment, and tissue capillary density.Although none of these facets appear toplay an essential role, they all seem tocontribute to the profile of adaptation toaltitude.
Body morphology
In man, gross morphological differenceshave been noted between sea level andhigh altitude adult populations of the sameethnic origin (Hurtado, 1932). The nativesof the high land have a significantly smallerbuild (size and weight) than the low-land-ers. Further studies have confirmed thesedifferences during childhood (Frisancho,1969) and have related them to measure-ments of lung volume and pulmonaryfunction. These latter studies have showna greater total lung capacity in childrenat high altitude.
Differences in size and rate of growthbetween sea level and high altitude speci-mens have also been observed in rabbits(Muntz, 1891), in rats (Timaras et al.,1957), and in guinea pigs (Delaquerrier-Richardson, 1965). However, these may notbe general as there are no differences inbody size between sheep born at sea leveland at altitude (Metcalfe et al., 1962).
" . 1 7
1. PRAIRIE DOG
2. ROUND TAIL GROUND SQUIRREL3. JACK RABBIT4. 13STRIPPED GROUND SQUIRREL5. CHINCHILLA6. HAMSTER7. GUINEA PIG8. ANTELOPE GROUND SQUIRREL9. COTTON TAIL RABBIT
10. GERB1L11. GRAV SQUIRREL12. ALBINO RAT13. COTTON RAT14. NORWAY RAT15. JIRD16. FLYING SQUIRREL17. MOUSEIB. KANGAROO RAT
10 20 30 40
CRITICAL P02, mmHg
FIG. 13. PM of several species of rodents as a func-tion of their critical Po>. (Reproduced from Hall,1966, with permission of the American Journal ofPhysiology.)
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

ADAPTATION TO ALTITUDE 453
The mechanism of these gross morpho-logical differences is not known; yet, anunderstanding of them is an essential stepfor the general understanding of adapta-tion to altitude.
Lung development
The development and morphometricevaluation of the lung have been the ob-ject of several studies. Studies on sheepand guinea pigs showed a slightly greatermean alveolar diameter, diffusing surfacearea to lung volume ratio, and total lungcapacity in the altitude population (Ten-ney and Remmers, 1966). Another study,by Bartlett (1970), showed that in hypoxicrats mean lung weight, alveolar surfacearea, and alveolar number were also slight-ly greater than in the normoxic animal.However, only the study of Burri andWeibel (1971) in rats demonstrated un-equivocally that specific alveolar and spe-cific capillary surface areas could be re-lated to the oxygen tension of the environ-ment in which the animals were born andraised (Fig. 14). Hence, it appears as ifhypoxic environment inhibits total bodygrowth, a situation which enhances thepulmonary gas exchange capacity. It isworth noting that in lower vertebrates anincrease in total gas exchange area can beinduced by a reduction of the environ-mental oxygen concentration (Bond, 1960).
Tissue capillary density
Several studies have been made to deter-mine whether the capillary density in-creases during chronic exposure to hypoxia.Such exposure would result in a decreaseof the distance between capillaries, a con-dition that compensates for the reductionat altitude of the oxygen tension differ-ence between the capillary blood and themitochondria. This increase in number ofcapillaries has been demonstrated for rab-bits (Cassin et al., 1966), guinea pigsValdivia, 1965-66), and rats (Tenney andOu, 1970). The increase in capillary densitymay result from the development of newcapillaries or from the opening of pre-existing ones. This latter possibility hasbeen shown to occur in some specialized
RATS BORN AT DEFINED PO,
E
OXYGENTENSION. mmHg 100 150 300
FIG. 14. Specific alveolar and capillary surface inrats born and raised at different environmentalPo.,. Data from Burri and Weibel (1971) .
tissue, such as myocardium (Martini, 1969;Poupa, 1971-72). It is quite evident thatthe role allegedly played by an increase incapillary density in the adaptation to alti-tude is far from being established. This is,however, an extremely important questionthat only further comparative studies willanswer. Such studies should attempt to in-tegrate this factor with others which arealso part of the adaptive process. For ex-ample, it has been shown that those specieswith a relatively higher metabolic rate al-so have the highest capillary density(Schmidt-Nielsen and Pennycuik, 1961). Ttis also known that these species have a lowhemoglobin affinity for oxygen (Schmidt-Nielsen and Larimer, 1958) which in turnhas been associated with relatively highcritical Po, (Hall, 1966). Hence, it istempting to postulate that species with arelatively high initial Po., also have a highcapillary density; such association is in-deed a distinct advantage.
BIOCHEMICAL ADAPTATIONS
Although there are only very few studiesaddressed to the biochemical processes dur-ing life at altitude, there is overwhelmingevidence that some significant subcellularchanges and alterations of the metabolicpathways are taking place.
Oxygen requirements
Whether the total oxygen demand is af-fected at altitude has not been extensively
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

454 CLAUDE LENFANT
studied. There is, however, some sparseevidence that hypoxic environments maycause a slight decrease in oxygen require-ments in mammals. This phenomenon,known as "respiratory dependence," is con-sistently observed in lower vertebrates; i.e.,the metabolic activity is—to a point—di-rectly related to the availability of oxygen.Figure 15 illustrates some of the data ob-tained in mammals. Results from the lambshow a significant decrease of the oxygenconsumption per minute and per kilo ofbody weight. In man, the results indicate atrend, but the difference is not statisticallysignificant. The data from the steer alsoshow a significant difference in total oxy-gen consumption at sea level and at alti-tude. However, as body weight sometimesdecreased at altitude, it is not certain thatthis significance would pertain if the datawere expressed per unit weight. As formany other aspects of the adaptive processto altitude, the question of whether oxygendemand changes is still unanswered. Yet,it is too important a question to be over-looked.
Myoglobin concentration
The role of myoglobin in facilitating thetransport of oxygen within the tissues isnow well established. A recent study in div-ing mammals has shown that they have ahigher myoglobin concentration than ter-restrial mammals. It was also shown thatthe concentration increases with the diving
OXYGEN CONSUMPTION
10.0
7.5
2.5
FIG. 15. Oxygen consumption, per kilo or total, atsea level (slatted blocks) and after acclimatizationat altitude (stippled blocks). Data sources as fol-lows: man: Tonante et al. (1970/71) ; lamb:Reeves ct al. (1936c) ; steer: Grover et al. (1963).
-
-
MAN4340m
1 1 11 II
UMB3420m
2.000
1.500cE
E 1.000
s500
- STEER3420m
-
HUMAN SARTORIUS MUSCLE
s 7-5i~
5 S3o
i
BEEF HEART TISSUE
= - 3 r£ 2
IDPNH
DEHYDROGENASE
SL NAT
DPNHcyto C
REDUCTASE
FIG. 16. Comparison of various "biochemical pa-rameters" at sea level (SL) and in. natives of highaltitude (NAT) . Data sources as follows: myoglo-bin concentration: Reynafarje (1962) ; mitochon-dria count and enzymes (measured by a change inoptical density) : Ou and Tenney (1970) .
ability, i.e., the tolerance to hypoxia (Len-fant et al., 1970). Two studies have shownan increase in myoglobin concentration inhigh altitude natives as compared to theirsea level counterparts: one study was con-cerned with humans (Reynafarje, 1962),and the other with the llama (Curaca-Pena, 1970). All these reports seem to in-dicate that myoglobin concentration is animportant component of the adaptation tochronic hypoxia.
Subcellular adaptations
Since oxygen is consumed at the subcel-lular level, i.e., by the mitochondria, it isimportant to determine whether themitochondria and some of the metabolicsubstrates are modified. There are, as yet,very few studies that have been concernedwith this aspect of high altitude adapta-tion. A recent review (Poupa, 1972) sum-marizes the major metabolic shifts in themammalian heart acclimated to high al-titude. Figure 16 shows some other dataon the mitochondria number and on en-zymes intervening in oxygen utilization.The increase in mitochondria number en-hances the intracellular diffusion processwhile the increase in en/yme concentrationsuggests a higher rate of oxygen utiliza-
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

ADAPTATION TO ALTITUDE 455
tion. Similar observations have been madein diving mammals and been related totheir diving capability (E. Robin, personalcommunication).
CONCLUSIONS
This review of the high altitude adaptivemechanisms in mammals leads to severalconclusions:
1) As a whole, the adaptive process isextremely complex, being made up of sev-eral components that are either physiologic,morphologic, or biochemical in nature.The respective importance of these com-ponents varies among species.
2) No single component can explain thecompleteness of the species adaptation,whether newcomer or native. Each com-ponent must be considered as an elementof an interdigitated system. It is clearlyevident that one form of adaptation in-fluences the next one. This is exemplifiedby the position of the oxygen dissociationcurve (physiological adaptation) that is cor-related to the capillary density (morpho-logical adaptation) and to the critical P02
(biochemical adaptation).3) It appears that neither physiological
nor morphological changes can fully ex-plain the mechanisms of adaptation to al-titude. Although our knowledge of thebiochemical alteration at altitude is stillsuperficial, there is clear evidence that themost potent basis of adaptation may be atthe subcellular level. This is illustrated bythe extreme tolerance of the diving mam-mals to severe hypoxia, even though theydo not have the benefit of the physiologi-cal adaptation found in residents at highaltitude.
REFERENCES
Banchero, N., R. F. Grover, and J. A. Will. 1971a.Oxygen transport in the llama (Lama glama) .Resp. Ph>siol. 13:102-115.
Banchero, N., R. F. Grover, and J. A. Will. 19716.High altitude-induced pulmonary artery hyper-tension in the llama (Lama glama) . Amer. J.Physiol. 220:422-427.
Bartlett, D. 1970. Postnatal growth of the mam-malian lung: influence of low and high oxygentension. Resp. Physiol. 9:58-64.
Bert, P. 1874. Recherches experimentales sur l'in-
fluence que les modifications de la pression baro-metrique exercent sur les phenomenes de la vie.Masson, Paris.
Bisgard, G. E., H. G. Alvarez, and R. F. Grover.1969. Decreased ventilatory response to hypoxiaduring acute polycythemia in the calf. Resp.Physiol. 7:369-382.
Bond, A. N. 1960. An analysis o£ the response ofsalamander gills to changes in the oxygen con-centration of the medium. Develop. Biol. 2:1-20.
Brooks, J. G., and S. M. Tenney. 1968. Ventilatoryresponse of llama to hypoxia at sea level andhigh altitude. Resp. Physiol. 5:269-278.
Burri, P. H., and E. R. Weibel. 1971. Morphometricestimation of pulmonary diffusion capacity. II:Effect of PO2 on the growing lung. Resp. Physiol.11:247-264.
Cassin, S., R. D. Gilbert, and E. M. Johnson. 1962.Capillary development during exposure tochronic hypoxia. USAF School of AerospaceMedicine SAM-TR-66-16.
Chiodi, H. 1962. Oxygen affinity of the hemoglobinof high altitude mamals. Acta Physiol. Latino-Amer. 12:208-209.
Chiodi, H. 1970. Comparative study of the bloodgas transport in high altitude and sea levelcamelidae and goats. Resp. Physiol. 11:84-93.
Cruz-Jibaja, J., N. Banchero, F. Sime, D. Penaloza,R. Gamboa, and E. Marticorena. 1964. Correla-tion between pulmonary artery pressure andlevel of altitude. Dis. Chest 46:446-451.
Curaca-Pena, A. A. 1970. Determination de mio-hemoglobina en musculo estriado de glama LamaPacos. Arch. Inst. Biol. Andina Lima 3:112-121.
Delaquerriere-Richardson, L., S. Forkes, and E.Valdivia. 1965. Effect of simulated high altitudeon the growth rate of the albino guinea pigs.J. Appl. Physiol. 20:1022-1025.
Finch, C. A., and C. Lenfant. 1972. Oxygen trans-port in man. N. Engl. J. Med. 286:407-415.
Frisancho, A. R. 1969. Human growth and pul-monary function of a high altitude PeruvianQuechua population. Hum. Biol. 41:365-379.
Crover, R. F., J. T. Reeves, D. H. Will, and S. G.Blount. 1963. Pulmonary vasoconstriction insteers at high altitude. 18:567-574.
Guyton, A. C. 1967. Regulation of cardiac output.N. Engl. J. Med. 277:805-812.
Hall, F. G. 1965. Blood gas transport in chinchillas.Proc. Soc. Exp. Biol. Med. 119:1071-1073.
Hall, F. G. 1966. Minimal utilized oxygen and theoxygen dissociation curve of blood of rodents.J. Appl. Physiol. 21:375-378.
Hall, F. G., D. B. Dill, and E. S. Guzman Barron.1936. Comparative physiology at high altitude.J. Cell Comp. Physiol. 8:301-313.
Hartley, L. H., J. K. Alexander, M. Madelski, andR. F. Grover. 1967. Subnormal cardiac outputat rest and during exercise in residents at 3,100m altitude. J. Appl. Physiol. 23:839-848.
Hornbein, T. F., and S. C. Sorensen. 1969. Ven-tilatory response to hypoxia and hypercapnia incats living at high altitude. J. Appl. Physiol. 27:834-836.
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022

456 CLAUDE LENFANT
Hurtado, A. 1932. Respiratory adaptation in theIndian natives of the Peruvian Andes; studies ofhigh altitude. Amer. J. Phys. Anthropol. 17:137-323.
Kellogg, R. H. 1968. Altitude acclimization, a his-torical introduction emphasizing the regulationof breathing. Physiologist 11:37-57.
Klausen, K. 1966. Cardiac output in man in restand work during and after acclimization to 3,800m. J. Appl. Physiol. 21:609-616.
Kooyman, G. L. 1973. Respiratory adaptations inmarine mammals. Amer. Zool. 13:457-468.
Lahiri, S., N. S. Cherniack, N. H. Edelman, andA. P. Fishman. 1971. Regulation of respirationin goat and its adaptation to chronic and lifelong hypoxia. Resp. Physiol. 12:388-403.
Lenfant, C, K. Johansen, and J. D. Torrance.1970. Gas transport and oxygen storage capacityin some pinnipeds and the sea otter. Resp.Physiol. 9:277-286.
Lenfant, C, and K. Sullivan. 1971. Adaptation toaltitude. N. Engl. J. Med. 284:1298-1309.
Lenfant, C., J. D. Torrance, E. English, C. A. Finch,C. Reynafarje, J. Ramos, and J. Faura. 1968. Ef-fect of altitude on oxygen binding by hemoglo-bin and on organic phosphate levels. J. Clin.Invest. 47:2652-2656.
Lenfant, C., J. D. Torrance, and C. Reynafarje.1971. Shift of the CvHb dissociation curve ataltitude: mechanism and effect. J. Appl. Physiol.30:625-631.
Martini, J., and C. R. Honig. 1969. Direct measure-ment of intercapillary distance in beating ratheart in situ and under various conditions ofO2 supply. Microvasc. Res. 1:244-256.
Metcalfe, J., G. Meschia, A. Hellegers, H. Prystow-sky, W. Huckabee, and D. H. Barron. 1962. Ob-servations on the growth rates and organ weightsof fetal sheep at altitude and sea level. Quart.J. Exp. Physiol. 47:305-313.
Muntz, M. A. 1891. De l'enrichissement du sang enhomoglobine, suivant les conditions d'existence.C. R. Acad. Sci. Paris 112:298-301.
Ou, L. C, and S. M. Tenney. 1970. Properties ofmitochondria from hearts acclimatized to highaltitude. Resp. Physiol. 8:151-159.
Poupa, O. 1971/72. Anoxic tolerance of the heartmuscle in different types of chronic hypoxia.Cardiology 56:188-196.
Poupa, O. 1972. Heart under unusual conditions:effects of gravitation and chronic hypoxia, p.779-802. In E. Bajusz and G. Rona [ed.], Myo-cardiology. Univ. Park Press, Baltimore.
Rahn, H., and A. B. Otis. 1949. Man's respiratory
response during and afler acclimatization to highaltitude. Amer. J. Physiol. 157:445-462.
Reeves, J. T., E. B. Grover, and R. F. Grover.1963a. Pulmonary circulation and oxygen trans-port in lambs at high altitude. J. Appl. Physiol.18:560-566.
Reeves, J. T., E. B. Grover, and R. F. Grover.1963b. Circulatory response to high altitude inthe cat and rabbit. J. Appl. Physiol. 18:575-579.
Reynafarje, B. 1962. Myoglobin content and en-zymatic activity of muscle and altitude adapta-tion. J. Appl. Physiol. 17:301-305.
Schmidt-Nielsen, K., and J. L. Larimer. 1958. Oxy-gen dissociation curves of mammalian blood inrelation to body size. Amer. j . Physiol. 195:424-428.
Schmidt-Nielsen, K., and P. Pennycuik. 1961. Capil-lary density in mammals in relation to body sizeand oxygen consumption. Amer. J. Physiol. 200:746-750.
Tenney, S. M., and L. C. Ou. 1970. Physiologicalevidence for increased tissue capillarity in ratsacclimatized to high altitude. Resp. Physiol.8:134-150.
Tenney, S. M., and J. E. Remmers. 1966. Alveolardimensions in the lungs of animals raised athigh altitude. J. Appl. Physiol. 21:1328-1330.
Timaras, P. S., A. A. Krum, and N. Pace. 1957.Body and oxygen weights of rats during ac-climatization to an altitude of 12,470 feet. Amer.J. Physiol. 191:598-604.
Torrance, J. D., C. Lenfant, J. Cruz, and E. Mar-ticorena. 1970/71. Oxygen transport mechanismsin residents at high altitude. Resp. Physiol. 11:1-15.
Valdivia, E. 1965/66. Observaciones sobre la redcapilar de los musculos estriados de cobayos de laaltura. Arch. Inst. Biol. Andina Lima 1:311-318.
Viault, M. 1891. Sur la quantite d'oxygene con-tenue dans le sang des animaux des hautsplateaux de l'Amerique du Sud. C. R. Acad.Sci. Paris 112:295-298.
Weil, J. V., E. Byrne-Quinn, I. E. Sodol, G. F.Filley, and R. F. Grover. 1971. Acquired altera-tion of chemoreceptor function in chronicallyhypoxic man at high altitude. J. Clin. Invest.50:186-195.
Wintrobe, M. 1949. Clinical hematology (Appen-dix A: Comparative physiology) . Lea and Febi-ger, Philadelphia.
Woodbury, R. A., and W. F. Hamilton. 1941. Theeffect of histamine on the pulmonary blood pres-sure of various animals with and without anes-thesia. J. Pharmacol. Exp. Ther. 71:293-301.
Dow
nloaded from https://academ
ic.oup.com/icb/article/13/2/447/1999212 by guest on 04 August 2022




















