
Hidalgo et al Climate Larvaldrift Prsb 2012
10
doi: 10.1098/rspb.2011.0750 , 275-283 first published online 15 June 2011 279 2012 Proc. R. Soc. B Stenseth M. Hidalgo, Y. Gusdal, G. E. Dingsør, D. Hjermann, G. Ottersen, L. C. Stige, A. Melsom and N. C. distributions reveals non-stationary climate effects on fish larvae A combination of hydrodynamical and statistical modelling Supplementary data tml http://rspb.royalsocietypublishing.org/content/suppl/2011/06/14/rspb.2011.0750.DC1.h "Data Supplement" References http://rspb.royalsocietypublishing.org/content/279/1727/275.full.html#ref-list-1 This article cites 33 articles, 6 of which can be accessed free Subject collections (142 articles) environmental science (945 articles) ecology Articles on similar topics can be found in the following collections Email alerting service here right-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top http://rspb.royalsocietypublishing.org/subscriptions go to: Proc. R. Soc. B To subscribe to This journal is © 2012 The Royal Society on December 23, 2011 rspb.royalsocietypublishing.org Downloaded from
-
Upload
havforskningsinstituttet -
Category
Documents
-
view
4 -
download
0
Transcript of Hidalgo et al Climate Larvaldrift Prsb 2012

doi: 10.1098/rspb.2011.0750, 275-283 first published online 15 June 2011279 2012 Proc. R. Soc. B
StensethM. Hidalgo, Y. Gusdal, G. E. Dingsør, D. Hjermann, G. Ottersen, L. C. Stige, A. Melsom and N. C. distributionsreveals non-stationary climate effects on fish larvae A combination of hydrodynamical and statistical modelling
Supplementary data
tml http://rspb.royalsocietypublishing.org/content/suppl/2011/06/14/rspb.2011.0750.DC1.h
"Data Supplement"
Referenceshttp://rspb.royalsocietypublishing.org/content/279/1727/275.full.html#ref-list-1
This article cites 33 articles, 6 of which can be accessed free
Subject collections
(142 articles)environmental science � (945 articles)ecology �
Articles on similar topics can be found in the following collections
Email alerting service hereright-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top
http://rspb.royalsocietypublishing.org/subscriptions go to: Proc. R. Soc. BTo subscribe to
This journal is © 2012 The Royal Society
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from

Proc. R. Soc. B (2012) 279, 275–283
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
*Author
Electron10.1098
doi:10.1098/rspb.2011.0750
Published online 15 June 2011
ReceivedAccepted
A combination of hydrodynamical andstatistical modelling reveals non-stationaryclimate effects on fish larvae distributions
M. Hidalgo1, Y. Gusdal2, G. E. Dingsør1,3, D. Hjermann1,
G. Ottersen1,4, L. C. Stige1, A. Melsom2 and N. C. Stenseth1,5,*1Centre for Ecological and Evolutionary Synthesis (CEES), Department of Biology, University of Oslo,
PO Box 1066 Blindern, 0316 Oslo, Norway2Norwegian Meteorological Institute, 0313 Oslo, Norway
3Institute of Marine Research, PO Box 1870 Nordnes, 5817 Bergen, Norway4Institute of Marine Research, Gaustadalleen 21, 0349 Oslo, Norway
5Institute of Marine Research, Flødevigen Marine Research Station, 4817 His, Norway
Biological processes and physical oceanography are often integrated in numerical modelling of marine fish
larvae, but rarely in statistical analyses of spatio-temporal observation data. Here, we examine the relative
contribution of inter-annual variability in spawner distribution, advection by ocean currents, hydrography
and climate in modifying observed distribution patterns of cod larvae in the Lofoten–Barents Sea.
By integrating predictions from a particle-tracking model into a spatially explicit statistical analysis, the
effects of advection and the timing and locations of spawning are accounted for. The analysis also
includes other environmental factors: temperature, salinity, a convergence index and a climate threshold
determined by the North Atlantic Oscillation (NAO). We found that the spatial pattern of larvae changed
over the two climate periods, being more upstream in low NAO years. We also demonstrate that spawning
distribution and ocean circulation are the main factors shaping this distribution, while temperature effects
are different between climate periods, probably due to a different spatial overlap of the fish larvae and
their prey, and the consequent effect on the spatial pattern of larval survival. Our new methodological
approach combines numerical and statistical modelling to draw robust inferences from observed distri-
butions and will be of general interest for studies of many marine fish species.
Keywords: ocean advection; particle-tracking models; spatially explicit analyses; fish larvae distribution;
spawning distribution; ecological climate effects
1. INTRODUCTIONSpatio-temporal variability in distributions of early life
stages (ELS; i.e. eggs and larvae) of marine fish is
shaped by complex interactions between the effects of
abundance, fecundity and behaviour of spawners, ocean
circulation patterns, and spatial and temporal patterns
of mortality (which are in turn dependent on availability
of food and predator aggregations). These interacting
processes operate across various scales to establish the
seemingly stochastic patterns observed [1]. Further,
changing climate regimes have the potential to disrupt
these interactions. Climate influences distributions, for
instance, by affecting larval dispersal pathways (through
speed and direction of currents) [2], through tempera-
ture-dependent processes (growth, food availability,
metabolic rates and energetic cost of larvae decisions)
[3], and by affecting spawner distribution and spawning
phenology [4].
Great progress has been made within the field of bio-
physical modelling of ELS of marine fish since its
humble beginnings some decades ago. The development
for correspondence ([email protected]).
ic supplementary material is available at http://dx.doi.org//rspb.2011.0750 or via http://rspb.royalsocietypublishing.org.
10 April 201127 May 2011 275
of computational tools allows linking high-spatial-
resolution numerical (hydrodynamic) models with indi-
vidual-based models (IBMs). This coupled approach
allows for simulating oceanographic transport and ELS
dynamics (i.e. growth, mortality and behaviour) to investi-
gate the processes shaping fish distributions (see reviews in
[5–8]). Independently, recent studies have demonstrated
the importance of including spatial aspects in statistical
analyses of inter-annual dynamics of plankton [9], ELS
[10,11] and fish juveniles [12]. Particle-tracking/IBMs of
ELS often use observational data to calibrate the
biological parameters of the models in order to be able to
qualitatively reproduce the observed patterns (e.g.
[13–15]). In some cases, particle-tracking estimates have
been used to throw light on the causes of inter-annual
variability in recruitment [16]. However, despite the valu-
able advances made within particle-tracking/IBMs of ELS,
the output is rarely analysed in unison with observational
data to draw quantitative inferences by means of statistical
methods. Here, we apply spatial statistics to link modelled
distributions with measured observations, and evaluate
which factors influence spatial and inter-annual variation
in larvae abundance.
Northeast Arctic (NEA) cod (Gadus morhua) in the
Lofoten–Barents Sea (figure 1a) is out used as our study sys-
tem. NEA cod population dynamics and the spatio-temporal
This journal is q 2011 The Royal Society

70° N
0
1200
(a) (b)
10° E 20° E 30° E 40° E 50° E 60° E2488
10°
W60
° N
0° grid
loca
tion
y (k
m)
grid location x (km)
80° N850
SSB
(×
103 k
g) 1110987654
700550400250100
1.5
infl
ow to
Bar
ents
Se
a (s
v)
ln (
0-gr
oup
num
bers
)
NA
O in
dex
4.5
Bar
ents
Sea
te
mpe
ratu
re (
°C)
4.0
3.5
3.0
1.00.50.0
–0.5–1.0
420
–2–4
1980 1983 1986 1989 1992year
1995 1998 2001
Figure 1. The Barents Sea–Lofoten ecosystem. (a) The inner black rectangle is the oceanographic model domain used as a
geographical reference in the whole study. Black contours are the isobaths for the oceanographic model domain with an intervalof 500 m. The dotted blue line indicates the spawning area (Lofoten archipelago; figure 2) and the dotted red line refers to thelarval area modelled (figure 3). The Norwegian Atlantic Current is shown with a yellow arrow and the Norwegian CoastalCurrent with a green arrow. (b) Biological (SSB; spawning stock biomass and 0-group abundance index; upper panel), hydro-graphical (winter inflow to the Barents Sea from southwest and mean value of sea temperature; adapted from Ottersen et al.[17]; middle panel) and climatic (NAO, North Atlantic Oscillation; lower panel) time series indicating the consistency ofshort-term periods (1986–1988 and 1989–1991) in the study period. Red and blue areas represent periods with the NAOindex above or below the mean value for the last 30 years, respectively. The inner rectangle of dashed lines refers to ourstudy period from 1986 to 1991, which comprises contrasting biological, hydrographical and climatic conditions.
276 M. Hidalgo et al. Embracing physics and biology
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
pattern of juvenile survival have been shown to change with
climate [18,19]. Biophysical modelling studies have also con-
firmed that NEA cod larvae drift routes and individual
growth can vary inter-annually with hydrographical regimes
[2,20,21]. NEA cod is thus a well-suited model organism
to test methods for determining the importance of biotic
factors and environmental conditions in modifying the spatial
distribution at the larval stage, and whether the effects differ
with climate.
We investigate the large-scale spatial patterns of larvae
advected from their spawning grounds by currents, and
how these patterns are affected by spawner distribution,
larval mortality and different environmental covariates of
the larval habitat. Our approach is to link the simulated dis-
tributions of virtual particles (hereafter named drifters) with
the observed distributions of larvae in July from 1986 to
1991. To do that, we incorporate drifter abundance for
each year and grid location as covariates in a statistical
spatial analysis, and include temperature, salinity and an
index of convergence/divergence (CD) activity as additional
covariates. Additionally, the study period comprises
contrasting conditions of the North Atlantic Oscillation
(NAO [22]). The NAO refers to inter-annual and inter-
decadal variability in atmospheric forcing, which affects
sea temperature, the strength of inflow to the Barents Sea
and the population dynamics of NEA cod [17,19,22]
(figure 1b). Consequently, the larvae experience both
spatially and inter-annually variable ocean climate. To inves-
tigate possible non-stationary effects of the investigated
predictor variables, we hypothesize that their functional
forms may change under different climate periods.
2. MATERIAL AND METHODS(a) Biological data
Abundance data for cod larvae were obtained from yearly
cruises carried out off northern Norway and in the western
Barents Sea during the months of June–July from 1977 to
Proc. R. Soc. B (2012)
1991 using a pelagic trawl, sampling a depth from 50 m to
the surface. The trawl net codend had a 4 m liner with
5 mm (stretched) hexagonal mesh [23]. Note that, for simpli-
city, we refer to all sampled young-of-the-year fish as larvae,
although an undetermined proportion of them had meta-
morphosed to a post-larval stage [24,25]. Their lengths
varied between 12 and 54 mm. Acoustic spawning aggre-
gation data were obtained during annual spring surveys
carried out from 1986 and onwards off the Lofoten archipe-
lago (in the centre of the blue box in figure 1a; figure 2a),
where spawning lasts from early March to mid-May (see
electronic supplementary material for details). The Lofoten
area was divided into three sub-areas (Vestfjorden, Yttersida
and Nord; figure 2a) based on Ellertsen et al. [24]. Centres of
gravity (CsG; abundance-weighted mean latitude and
longitude) of the spatial distribution of spawners were calcu-
lated for each sub-area and year. These CsG were used as
starting points for the release of drifters (i.e. eggs) in the
particle-tracking model (see details below).
(b) The ocean circulation model
Data on ocean currents, sea temperature and salinity were
obtained from a coupled model system consisting of an ocean
circulation model (MI-POM) and an ice model (MI-IM). MI-
POM is the Norwegian Meteorological Institutes version of
the Princeton Ocean Model [26]. The MI-IM model is
described by Røed & Debernard [27]. The coupled model
system has a nested set-up as illustrated by Melsom & Fossum
[28]. The coarse grid domain covers the Arctic Sea, Nordic
Seas and the North Atlantic Ocean, with a resolution of
20 km. The finer grid domain within this has a mesh size of
4 km, and covers the Barents Sea and most of the Norwegian
Sea (the inner black rectangle in figure 1a). Owing to storage
limitations, model results were interpolated to an 8 km grid.
Note that the 8 km grid is the geographical reference used in
this study and the basis for the statistical spatial analysis. More
information about the model initialization and data assimilation
is provided in the electronic supplementary material.
472
(a) (b)3.0
0.60 12.7
11.9
11.1
ln (
abun
danc
e)
10.3
9.5
0.55
0.50
0.45
0.40
0.35
0.30
%
0.25
0.20
0.15
0.10
0.05
1986 1987 1988year
1989 1990 1991
2.5
2.0
1.5
1.0
0.5
0
456 87 86
Nord
88 8789909186
8891
Yttersida
Vestfjorden
90
91878988 9086
89440
424
408
392
grid
loca
tion
y (k
m)
376
360
344
584 616 648 680 712 744 776 808
grid location x (km)
840 872 904 936 968 1008 1048
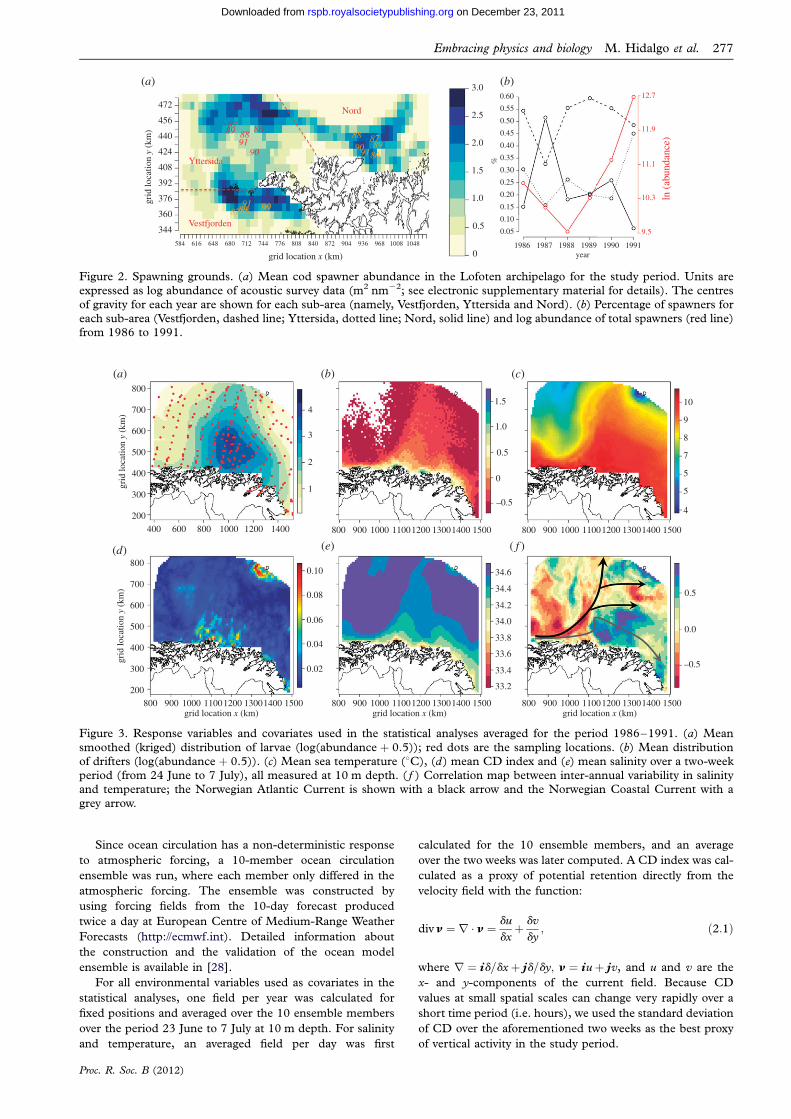
Figure 2. Spawning grounds. (a) Mean cod spawner abundance in the Lofoten archipelago for the study period. Units are
expressed as log abundance of acoustic survey data (m2 nm22; see electronic supplementary material for details). The centresof gravity for each year are shown for each sub-area (namely, Vestfjorden, Yttersida and Nord). (b) Percentage of spawners foreach sub-area (Vestfjorden, dashed line; Yttersida, dotted line; Nord, solid line) and log abundance of total spawners (red line)from 1986 to 1991.
800
(a) (b) (c)
(d) (e) ( f )
4
3
2
1
0.10 34.6
0.5
–0.5
0.0
34.4
34.2
34.0
33.8
33.6
33.4
33.2
0.08
0.06
0.04
0.02
1.5 10
9
8
7
5
5
4
1.0
0.5
–0.5
0
700
600
500
grid
loca
tion
y (k
m)
400
300
200
400 600 800 1000 1200 1400
800
700
600
500
grid
loca
tion
y (k
m)
grid location x (km)
400
300
200
800 900 1000 11001200 13001400 1500grid location x (km)
800 900 1000 11001200 13001400 1500grid location x (km)
800 900 1000 11001200 13001400 1500
800 900 1000 11001200 13001400 1500 800 900 1000 11001200 13001400 1500
Figure 3. Response variables and covariates used in the statistical analyses averaged for the period 1986–1991. (a) Mean
smoothed (kriged) distribution of larvae (log(abundance þ 0.5)); red dots are the sampling locations. (b) Mean distributionof drifters (log(abundance þ 0.5)). (c) Mean sea temperature (8C), (d) mean CD index and (e) mean salinity over a two-weekperiod (from 24 June to 7 July), all measured at 10 m depth. ( f ) Correlation map between inter-annual variability in salinityand temperature; the Norwegian Atlantic Current is shown with a black arrow and the Norwegian Coastal Current with a
grey arrow.
Embracing physics and biology M. Hidalgo et al. 277
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
Since ocean circulation has a non-deterministic response
to atmospheric forcing, a 10-member ocean circulation
ensemble was run, where each member only differed in the
atmospheric forcing. The ensemble was constructed by
using forcing fields from the 10-day forecast produced
twice a day at European Centre of Medium-Range Weather
Forecasts (http://ecmwf.int). Detailed information about
the construction and the validation of the ocean model
ensemble is available in [28].
For all environmental variables used as covariates in the
statistical analyses, one field per year was calculated for
fixed positions and averaged over the 10 ensemble members
over the period 23 June to 7 July at 10 m depth. For salinity
and temperature, an averaged field per day was first
Proc. R. Soc. B (2012)
calculated for the 10 ensemble members, and an average
over the two weeks was later computed. A CD index was cal-
culated as a proxy of potential retention directly from the
velocity field with the function:
divn ¼ r � n ¼ du
dxþ dv
dy; ð2:1Þ
where r ¼ id=dxþ jd=dy; n ¼ iuþ jv, and u and v are the
x- and y-components of the current field. Because CD
values at small spatial scales can change very rapidly over a
short time period (i.e. hours), we used the standard deviation
of CD over the aforementioned two weeks as the best proxy
of vertical activity in the study period.

278 M. Hidalgo et al. Embracing physics and biology
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
(c) The particle-tracking model and initialization
of virtual drifters
Egg/larvae drift was computed based on a semi-Lagrangian
advection model forced by currents from the ensemble
members. Particle trajectories were first computed based
on velocity fields from each of the ensemble members sep-
arately and then merged together to represent the average
particle concentration from the 10 realizations. These aver-
aged results were used in the statistical analysis. Owing to
the fine mesh size of 4 km for the model, high-resolution
current fields were simulated, which resolved ocean
eddies. However, there are processes at still finer scales
that were not resolved. These processes were parametrized
by adding Gaussian noise to the model results (see elec-
tronic supplementary material for details). Owing to a
potential overestimation of on-shore transport in the near-
shore areas, a padding zone of 0.4 km was included in the
particle-tracking model when an advection time step of
30 min was applied. Particles that were within this distance
from the coastline, and not stranded, were moved outside
the padding zone.
In the particle-tracking model, drifters were released
with a Gaussian distribution centred at the CsG with a
standard deviation of 4 km (0.5 grid units). For each
sub-area and for each year, 15 000 drifters were released
around the sub-area’s CsG for that year (figure 2a). To
account for variation in the timing of spawning, drifters
were released on 15 March, 1 April and 15 April. The
spatial transport of ELS of cod is also depth-dependent.
At 10–20 m depth, the Norwegian Coastal Current
(NCC) retains almost all the pelagic and passive-drifting
stages close to the coast. Above 10 m, ELS are probably
advected northwards by the Norwegian Atlantic Current
(NAC; figure 1a) [2,20]. Thus, to account for depth vari-
ation, drifters were released at depths of 10 and 20 m. As
there is little evidence of diel migration owing to 24 h
daylight during summer [20], drifters were passively trans-
ported at these fixed depths by the NCC and the NAC
based on results from the 10-member ocean model ensem-
ble. The resulting larvae distribution was recorded at the
time of the larval survey (around 1 July; i.e. after 90 days
of drifting). In total, 270 000 drifters were released each
year and standardized afterwards based on findings obtained
from Ellertsen et al. [24]. The standardization took into
account (see the electronic supplementary material): phenology
of spawning, depth distribution (assumed constant across
years), and inter-annual variation in the distribution of spaw-
ners between sub-areas and in total spawner abundance
(figure 2b).
(d) Statistical analysis of cod larvae distribution
Prior to analysis, the natural logarithm of observed larvae
abundances (plus a constant, k ¼ 0.5) was interpolated by
means of ordinary kriging over a regular grid. To allow
direct comparison of the different types of data, we used
the same standardized grid (8 � 8 km; figure 1a) for the kri-
ging as for the ocean circulation model. Kriged data for cod
larvae were regressed against co-located covariates using gen-
eralized additive models (GAMs) [29]. We used two types of
GAM formulations: (i) purely additive, assuming that the
effect of each covariate was stationary (that is, that the func-
tional form of the effect did not change over the years), and
(ii) threshold (non-additive), to test the hypothesis that the
effect of a covariate changed in relation to an external
Proc. R. Soc. B (2012)
threshold variable (e.g. different climate periods). The
additive formulation used was
Lt;ðx;yÞ ¼ at þ sðx; yÞt þ f ½Dt;ðx;yÞ� þ g½Tt;ðx;yÞ�þ h½St;ðx;yÞ� þ j½CDt;ðx;yÞ� þ 1t;ðx;yÞ; ð2:2Þ
where Lt,(x,y) is kriged cod larvae abundance (log-scale) in
year t at location x,y in the grid, at is a year-specific constant,
s is a two-dimensional non-parametric smoothing function
describing the geographical effect (thin plate regression
spline with maximally 27 degrees of freedom (28 knots)), f,
g, h and j are one-dimensional non-parametric smoothing
functions (cubic splines with maximally three degrees of free-
dom (four knots)), D is the natural logarithm of the density
of drifters (plus a constant, k ¼ 0.5) from the particle-
tracking model, T is temperature, S is salinity, CD is the
CD index calculated for the given grid location over two
weeks, and 1 is an error term.
To test the hypothesis of different effects of covariates
under different climatic periods based on the NAO index,
we fitted non-additive threshold GAMs (TGAMs) [30] to
the data. TGAM is a semi-parametric regression where the
shape of the exploratory function may change according to
whether an external covariate is below or above a threshold
value. The change in functional form may occur in one or sev-
eral of the terms of the TGAM (in our model, from s1 to s2,
from f1 to f2, from g1 to g2, from h1 to h2 and/or from j1 to j2):
Lt;ðx;yÞ ¼ at þ 1t;ðx;yÞ
þ
s1ðx; yÞt þ f1½Dt;ðx;yÞ� þ g1½Tt;ðx;yÞ�þh1½St;ðx;yÞ� þ j1½CDt;ðx;yÞ� if NAOt � n
s2ðx; yÞt þ f2½Dt;ðx;yÞ� þ g2½Tt;ðx;yÞ�þh2½St;ðx;yÞ� þ j2½CDt;ðx;yÞ� if NAOt . n:
8>>>><>>>>:
ð2:3Þ
We initially applied an NAO index threshold, n, on each
function separately (i.e. on effects of location, drifter abun-
dance, temperature, salinity and CD index), while the
other functions were identical in low and high NAO regimes.
We fitted series of GAMs and TGAMs with different levels of
complexity (i.e. from one to five covariates and with all poss-
ible combinations). We considered all threshold values of
NAO (¼0, 0.5, 1, 1.5 and 4) that could uniquely delimit
the potential temporal regimes from 1986 to 1991. Each
threshold value was individually applied to each of the covari-
ates for each model, resulting in a total of 114 models
considered. The best model, GAM or TGAM, was deter-
mined by computing the (genuine) cross-validation (CV;
a random sample of 10% of the data were used for these cal-
culations, but threshold values were kept fixed; see [30] for
details). Low CV values indicate the best model compromise
between model complexity and fit to the observed data. After
identifying the best models with only one threshold variable,
we considered selected models with up to two threshold
variables, as described in §3.
The residuals did not show temporal dependency but
were spatially correlated. This is unlikely to bias estimates
of the additive effects, but results in an overestimation of
the significance level of the covariates, thus invalidating the
standard p-values [12]. We used a non-parametric wild boot-
strap approach to get an accurate estimate of p-values and
confidence intervals of covariates [31]. For the best-fitted
model based on the CV minimization, the residuals were

Embracing physics and biology M. Hidalgo et al. 279
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
extracted, rescaled to the same variance as the estimated
scale parameter of the model and randomly replaced
among years (sampling with replacement), before being
used as response variable in the model (having the same set
of covariates as the original model; see details in [32]). As
an additional test of the significance of the threshold effects,
we used an alternative CV approach, in which we left out data
for whole years at a time (instead of a random 10% of the
data). We then confirmed that the final selected threshold
model (but without a year-specific intercept) had higher
predictive power for data cases (years) not used when fitting
the model compared with alternative formulations without
thresholds.
3. RESULTS(a) Spawning aggregations
Both the spatial distribution (CsG) and abundance within
sub-areas varied among years (figure 2a,b; see spawner
distribution and CsG for all the years in the electronic
supplementary material, figure S1). The CsG were gener-
ally more offshore from 1987 to 1989 compared with
1986, 1990 and 1991 in Yttersida and Nord. Addition-
ally, CsG were in the outer area of Vestfjorden from
1987 to 1989 and 1991. Pairwise correlations among
CsG showed inter-annual consistency in the spatial vari-
ation: x-axis positions (note spatial coordinates refer to
the oceanographic model domain; see figure 1a) were
highly and positively correlated between Vestfjorden and
Yttersida (Spearman, r ¼ 0.92, p , 0.05), while y-axis
positions were negatively correlated between Nord and
Vestfjorden (r ¼ 20.74, p , 0.05). With the exception
of 1987 and 1991, the relative contribution of each sub-
area to total annual cod abundance in Lofoten was quite
constant (figure 2b). In 1987, 51 per cent of the fish spawned
in the Nord sub-area, compared with 15 to 25 per cent for
most years. In 1991, the year with highest total spawner
abundance, only 6 per cent spawned in this sub-area.
(b) Particle-tracking model
Maps of simulated larvae distributions after 90 days of drift-
ing from the mean date of spawning (1 April) and from each
sub-area show only the predicted effects of inter-annual
variation in the current pattern (see electronic supplemen-
tary material, figure S2). By taking into account variation
in the relative contribution of each sub-area, the model pre-
dicted extensive spreading of drifters in the Barents Sea for
1987 (see electronic supplementary material, figure S3a).
After taking into account variation in total spawner abun-
dance, extensive spreading was also predicted for 1991
(electronic supplementary material, figure S3b). Higher
abundance of drifters in the near-shore areas is a general
pattern in the distributions obtained.
(c) Spatial analyses
We analysed the distribution of larvae estimated from
survey data (figure 3a; see all the years in the electronic
supplementary material, figure S4, and its inter-annual
variation in electronic supplementary material, figure S5)
using additive or non-additive models with one to five co-
variates. Besides drifter density estimates described above
(figure 3b; all years are shown in the electronic supplemen-
tary material, figure S3b), temperature, salinity and CD
index were used as covariates (figures 3c–e; all years are
Proc. R. Soc. B (2012)
shown in the electronic supplementary material, figure
S6). Results show that models with non-additive threshold
effects (TGAM) were more parsimonious (as given by CV
values) compared with purelyadditive models (see electronic
supplementary material, table S1). The most parsimonious
TGAMs generally identified a threshold of NAO ¼ 1, corre-
sponding to two short-term climatic periods: 1986–1988
(NAO , 1, ‘cold’ period) and 1989–1991 (NAO . 1,
‘warm’ period). The preliminary best models included all
five covariates and an NAO ¼ 1 threshold on either:
(i) spatial location, the best model (see electronic sup-
plementary material, table S1, model 80); (ii) temperature,
the second best model (see electronic supplementary
material, table S1, model 76); or (iii) drifter abundance,
the third best model (see electronic supplementary material,
table S1, model 78). These models were sequentially com-
bined, applying an NAO ¼ 1 threshold on two or three
covariates, and resulted in three models (models 81–83).
Two of these models included salinity, although salinity
was not significant (p ¼ 0.18) when applying a non-para-
metric bootstrap on model 81. Spatial heterogeneity in the
correlation between salinity and temperature as shown in
our study (figure 3f ) justifies the inclusion of both tempera-
ture and salinity in our models. The final model (see
electronic supplementary material, table S1, model 83;
figure 4) explained 84 per cent of the observed variance in
larvae abundance.
The final model is shown in figure 4. The partial effect of
year (p , 0.05; figure 4a) shows an increasing trend, partly
coinciding with the trend in spawner abundance (figure 1b).
The relationship between drifter abundance and observed
larval abundance is asymptotic (p , 0.05; figure 4b). Temp-
erature effects differed between the climatic periods (1986–
1988 and 1989–1991; the two effects p , 0.05; figure 4c).
Specifically, during the warm period, high abundances of
larvae were found in waters with temperatures around
9.58C, while in the cold period the effect of temperature
was stronger and the highest abundances were found in
temperatures above 108C. The effect of the geographical
location showed a clear and significant difference between
periods, suggesting positive anomalies in the southwestern
parts of the study area in the period 1986–1988 (p ,
0.05; figure 4d) and in the northeastern parts in the
period 1989–1991 (p , 0.05; figure 4e). Note that these
effects remain while accounting for local environmental co-
variates. Finally, there was some evidence for an effect of
CD activity (p¼ 0.073; figure 4f ). To assess the unique
contribution of each covariate, we calculated the difference
in percentage of variance explained between the full model
and a model with the given term removed. The spatial term
and the partial effect of year contributed the most (25 and
19 per cent, respectively), while temperature, drifter abun-
dance and CD activity explained 11.8, 7.2 and 1.8 per
cent, respectively.
4. DISCUSSIONOur study reveals how the abundance and distribution of
spawners, ocean circulation and hydrographical environ-
mental conditions in combination modify the spatial
distribution of ELS, and how the effects of these factors
differ between two periods with contrasting states of the
NAO. The best model obtained explains a major fraction
of the observed variance and includes drifter abundance

4(a)
(d) (e) ( f )
(b) (c)
3
2
1
4
3
2
1
0
–1
–2
–3
0
86
800
700
600
500
400
300
200800 900 1000 1100
grid location x (km)
grid
loca
tion
y (k
m)
800 2.0
1.5
1.0
0.5
–0.5
–1.0
0.0
700
600
500
400
300
200
grid
loca
tion
y (k
m)
1200 1300 14001500 800 900 1000 1100
grid location x (km)
1200 1300 1400 1500 0 0.02 0.04
convergence/divergence index
0.06 0.08 0.10
87
0.60.8
0.20.4
–0.4–0.2
–0.6
–0.2 0.20
0.2
–0.2
0.8 –0.8
–0.6
–0.2
0
1
0.6
–0.4
0.4
0.4
–0.4
–1
–0.8
–0.8
1
1
0
88 89year
90 91 –0.5 0 0.5 1.0drifters (ln [N/grid unit])
1.5 2.0 2.5
part
ial e
ffec
t of
fact
or
addi
tive
effe
ct
4
3
2
1
0
–1
–2
–3
5 6 7 8 9temperature (°C)
10 11 12
addi
tive
effe
ctad
ditiv
e ef
fect
–1
–2
Figure 4. Results of the best model using non-additive threshold effects: (a) partial effect of year factor, (b) drifter effect,(c) temperature effect for low and high NAO regimes (represented in blue and red, respectively), (d) partial spatial effectfor low NAO regime (less than 1, 1986–1988), (e) partial spatial effect for high NAO regime (greater than 1, 1989–1991)and ( f ) CD index effect. Whole and broken lines indicate fitted partial effects with two times standard deviation estimatedfrom bootstrapping, respectively. Confidence intervals for the spatial effects are shown in figure S7 in the electronic supplemen-
tary material. Points for (b), (c) and (f) represent the partial residuals for each smoothed term. Note that the scale of the y-axisfor CD index is different from (a–c).
280 M. Hidalgo et al. Embracing physics and biology
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
(which integrates effects of spawner abundance and dis-
tribution, and larval advection) and the CD index as
additive covariates, and temperature and geographical
location as non-additive covariates—their effects changing
between a high and low NAO index. We show that by com-
bining ocean circulation models and statistical techniques,
we can reproduce the spatio-temporal dynamics of NEA
cod larvae, and reveal the environmental factors shaping
their distribution. Thus, this study provides a novel and
promising methodological framework for integrating quan-
titative information on spawner aggregations with ocean
circulation and hydrography in statistical spatial analyses
of pelagic ELS.
(a) Implications of spawner distribution for
advection and survival
The starting position of drifters fundamentally controls
the directions and distance of their drift paths. Therefore,
the spatio-temporal distribution of spawners (when,
where and how many) is crucial for proper initialization
of particle-tracking models [8]. Although cod spawning
phenology and geographical location is a fine-tuned evol-
utionary strategy developed through generations to
maximize survival [33], large-scale inter-decadal displace-
ments of spawning aggregations have been described and
attributed to both climate [34] and fishing effects on size
structure [35]. From the mid-1980s, a warm period
during which cod populations were demographically
eroded, the Lofoten grounds were the most important
area for the spawner aggregations. Our results add to
this picture by emphasizing the effects of small-scale vari-
ation within the Lofoten archipelago. The inter-annual
variation in the CsG was consistent among sub-areas,
Proc. R. Soc. B (2012)
indicating a certain spatial synchrony in spawning aggre-
gation patterns. However, we cannot establish whether
the temporal variation of the fine-scale structure in the
spawning areas was related to climate, mating behaviour
or changes in the population density over time.
Our results confirm that spatio-temporal variation in
spawner aggregations within and among sub-areas
around Lofoten combined with varying circulation pat-
terns impact the resultant spatial distribution of eggs
and larvae. In 1987, for example, the higher contribution
of spawners at Nord (the more offshore sub-area)
facilitated a more extensive spreading of ELS. Such
spreading may ensure that a sufficient number of eggs
and larvae encounter environmentally favourable con-
ditions ([36] and references therein), and thus may have
contributed to the higher larval survival found that year
(figure 4a). High survival in 1991, when high spawner
abundance caused extensive spreading of drifters into
the Barents Sea, is consistent with this explanation.
(b) Spatial distribution of early life stages of cod
The asymptotic relationship observed between drifter
abundance and observed larval abundance could be
related to lower survival of ELS very close to the coast,
which is where the highest concentrations of drifters
occurred. Stranding of eggs onto the coast, higher preda-
tion during the drifting process and higher mortality at
high larval densities (e.g. because of food limitation) are
all factors that may have contributed to low survival.
However, it is worth noting that the unresolved currents
in the coastal areas owing to the confluence of multiple
drivers interacting at fine to meso scales near shore
could potentially overestimate drifters along the coast

Embracing physics and biology M. Hidalgo et al. 281
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
[6]. Although a padding zone was included in the
particle-tracking model to mitigate this problem, overesti-
mation of drifter density along the coast is certainly
possible. Nevertheless, the spatial pattern of our results
is consistent with other studies applying different ocean
models [2,20].
Having accounted for drifter abundance (i.e. the com-
bined effect of spawner distribution and larval advection),
the statistical model captures the potential effect of the
other covariates on larval distribution. The most novel
and striking result is the different contribution of
temperature and spatial location under two short-term
periods with contrasting climate conditions (1986–1988
and 1989–1991). Previous studies have shown that both
temperature and recruitment success of cod in the study
area are higher during NAO-positive years [36]. Our
analysis adds to this picture by showing that the generally
positive temperature–larval abundance relation was
weaker in years with high NAO values. During the gener-
ally warm (NAO-positive) 1989–1991 period, high
abundances of larvae were found in water masses with a
relatively wide range of temperatures. During the colder
1986–1988 period, high larval abundances were only
found in the warmest waters. The causal basis for this
difference is not clear, but we note that temperature is
positively associated with food availability and that there
is a better temporal match between the development of
cod larvae and their zooplankton prey in warm years
[24,37]. We speculate that in cold periods, zooplankton
of appropriate size is only available in sufficient quantity
in the warmest water masses, whereas in warm periods,
zooplankton conditions are favourable over larger areas
(and a wider temperature range). This is supported by
recent evidence showing that zooplankton biomass pre-
dicts survival of ELS of NEA cod [25], and how
prolonged temporal overlap between fish larvae and
their prey in warm years enhances cumulative growth
and survival [37].
Lastly, the partial effect of geographical location of
larvae shows a clear and significant difference between
the two periods. Based on earlier work, the southwestern
distribution of larvae could potentially be explained by
a reduction in transport during cold periods (NAO-nega-
tive years) [2,17]. However, as far as the drifters capture
the current transport, the spatial pattern summarizes
the variability left over by the selected environmental co-
variates, and therefore could indicate that the spatial
pattern in mortality differs between the two periods. Sev-
eral factors could cause such a shift in mortality, such as
changes in the distribution of predators or prey. Higher
zooplankton biomass in NAO-positive years has indeed
been shown for the ‘downstream’ [38] but not the
‘upstream’ (L. C. Stige 2011, unpublished data) part of
our study area, consistent with such a role of prey.
A potential role of predators is supported by the results
of Ciannelli et al. [18] suggesting that cannibalism may
modify the distribution pattern of 1-year-old cod in the
Barents Sea. On the other hand, the climatic influence
on the transport cannot be disregarded. For example,
during windy conditions (more frequent in NAO-positive
years) eggs are mixed deeper in the water column, where
NCC favours a more downstream distribution [20].
Further, our results also showed that CD activity could
affect the observed larval distribution. The highest CD
Proc. R. Soc. B (2012)
is found in the southern part of the shallow Barents
Sea, where the NAC is deflected northwards and the
NCC spreads out from the coast (figure 1a). This process
contributes to a front developing between the water
masses, creating a convergence zone with increased con-
centration of phytoplankton and zooplankton [39].
Finally, CD activity in this area could slow down the
northward transport of larvae and reinforce the southern
distribution observed in cold periods.
(c) Implications and future challenges for an
integrative approach
Disentangling the drivers of larval dispersal and the
consequences for the spatio-temporal patterns of ELS
distribution is an inherently biophysical problem. This
requires an interdisciplinary effort, integrating tools from
different scientific disciplines. This study provides a new
biophysical framework to link observational information
from spawner distributions to larval distributions, taking
into account variability in ocean circulation and the
hydrographical environmental conditions of the larval
habitat. Although we provide some methodological
advances, such as the use of a model ensemble to include
uncertainty into the numerically modelled larval
distributions and covariates, there is also room for
improvement. Key challenges for future studies include
improvement in the near-shore oceanographic modelling,
accounting for the size structure of the mature stock,
inclusion of larval behaviour linked to observations (e.g.
vertical migration linked to observations of food avail-
ability) and accounting for the lack of synopticity in the
ELS surveys.
5. CONCLUSIONSAlthough the circulation pattern is the main physical
factor shaping the spatial distribution of larvae, climate
indirectly modifies this distribution by affecting larval sur-
vival (e.g. [25,37]). It is worth noticing that inter-annual
variation in spatial distribution of spawners at scales of
less than 100 km can affect the spread of larvae into the
larval habitat. In essence, our study highlights that
larvae were distributed over a wider temperature range,
over a larger area and more downstream in warm periods
owing to a combination of (i) higher abundance of spaw-
ners, (ii) an increase of ocean transport, (iii) higher overall
survival (possibly because of better food availability and/
or escape from predation), and (iv) a likely change in
the spatial pattern in natural survival, hypothetically
caused by changes in spatial distribution of predators or
prey. ELS of many marine fish species experience differ-
ent environmental conditions throughout their
distribution ranges. The combination of hydrodynamical
and statistical modelling is a promising tool for integrat-
ing information about oceanographic processes, small-
scale ecological mechanisms and large-scale observations
to draw statistically robust inferences about the processes
shaping ELS distribution and survival. Our study shows
that such a tool is likely to provide novel knowledge,
even for highly well-studied species and systems.
M.H., Y.G., D.H., G.O. and L.C.S. received support fromthe Research Council of Norway (projects no. 173487 and178434). M.H. thanks M. Llope for the advice anddiscussions on an early version of this manuscript and for

282 M. Hidalgo et al. Embracing physics and biology
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
help in the data analysis, Lee Hsiang Liow and JasonD. Whittington for reviewing an advanced version, and twoanonymous reviewers for their helpful comments.
REFERENCES1 Cowen, R. K. & Sponaugle, S. 2009 Larval dispersal
and marine population connectivity. Annu. Rev. Mar.Sci. 1, 443–466. (doi:10.1146/annurev.marine.010908.163757)
2 Vikebø, F., Jørgensen, C., Kristiansen, T. & Fiksen, Ø.2007 Drift, growth, and survival of larval NortheastArctic cod with simple rules of behaviour. Mar. Ecol.Prog. Ser. 347, 207–219. (doi:10.3354/meps06979)
3 O’Connor, M. I., Bruno, J. F., Gaines, S. D., Halpern, B.S., Lester, S. E., Kinlan, B. P. & Weiss, J. M. 2007 Temp-erature control of larval dispersal and the implication for
marine ecology, evolution and conservation. Proc. NatlAcad. Sci. USA 104, 1266–1271. (doi:10.1073/pnas.0603422104)
4 Rindorf, A. & Lewy, P. 2006 Warm, windy winters drivecod north and homing of spawners keeps them there.
J. Appl. Ecol. 43, 445–453. (doi:10.1111/j.1365-2664.2006.01161.x)
5 Lett, C., Rose, K. A. & Megrey, B. A. 2010 Biophysicalmodels. In Climate change and small pelagic fish (edsD. Checkley, J. Alheit, Y. Oozeki & C. Roy), pp.
88–111. Cambridge, UK: Cambridge University Press.6 Werner, F. E., Cowen, R. K. & Paris, C. B. 2007 Coupled
biological and physical models: present capabilities andnecessary developments for future studies of populationsconnectivity. Oceanography 20, 54–69.
7 Fiksen, Ø., Jørgensen, C., Kristiansen, T., Vikebø, F. &Huse, G. 2007 Linking behavioural ecology and ocean-ography: larval behaviour determines growth, mortalityand dispersal. Mar. Ecol. Prog. Ser. 347, 195–205.
(doi:10.3354/meps06978)8 Gallego, A. & North, E. W. 2009 Biological processes. In
Manual of recommended practices for modelling physical–biological interactions during the fish early life (eds E. W.North, A. Gallego & P. Peptigas), pp. 20–59. ICES
Cooperative Research Report no. 295. Copenhagen,Denmark: ICES.
9 Boyce, D. G., Lewis, M. R. & Worm, B. 2010 Globalphytoplankton decline over the past century. Nature466, 591–596. (doi:10.1038/nature09268)
10 Fox, C. J., O’Brien, C. M., Dickey-Collas, M. & Nash,R. D. M. 2000 Patterns in the spawning of cod (Gadusmorhua L.), sole (Solea solea L.) and plaice (Pleuronectesplatessa L.) in the Irish Sea as determined by generalisedadditive modelling. Fish Oceanogr. 9, 33–49. (doi:10.
1046/j.1365-2419.2000.00120.x)11 Bacheler, N. M., Ciannelli, L., Bailey, K. M. & Duffy-
Anderson, J. T. 2010 Spatial and temporal patterns ofwalleye pollock (Theragra chalcogramma) spawning inthe eastern Bering Sea inferred from egg and larval dis-
tributions. Fish. Oceanogr. 19, 107–120. (doi:10.1111/j.1365-2419.2009.00531.x)
12 Ciannelli, L., Fauchald, P., Chan, K. S., Agostini, V. N. &Dingsør, G. E. 2008 Spatial fisheries ecology: recent pro-
gress and future prospects. J. Mar. Syst. 71, 223–236.(doi:10.1016/j.jmarsys.2007.02.031)
13 North, E. W., Schlag, Z., Hood, R. R., Li, M., Zhong,L., Gross, T. & Kennedy, V. S. 2008 Vertical swimmingbehavior influences the dispersal of simulated oyster
larvae in a coupled article-tracking and hydrodynamicmodel of Chesapeake Bay. Mar. Ecol. Prog. Ser. 359,99–115. (doi:10.3354/meps07317)
14 Brickman, D., Marteinsdottir, G. & Taylor, L. 2007 For-mulation and application of an efficient optimized
Proc. R. Soc. B (2012)
biophysical model. Mar. Ecol. Prog. Ser. 347, 275–284.(doi:10.3354/meps06984)
15 Hinrichsen, H. H., Mollmann, C., Voss, R., Koster, F. W. &
Kornilovs, G. 2002 Biophysical modelling of larval Balticcod (Gadus morhua L.) growth and survival. Can. J. FishAquat. Sci. 59, 1858–1873. (doi:10.1139/f02-149)
16 Baumann, H., Hinrichsen, H. H., Mollmann, C., Koster,F. W., Malzahn, A. M. & Temming, A. 2006 Recruitment
variability in Baltic sprat, Sprattus sprattus, is tightlycoupled to temperature and transport patterns affectingthe larval and early juvenile stages. Can. J. Fish Aquat.Sci. 63, 2191–2201. (doi:10.1139/f06-112)
17 Ottersen, G., Helle, K. & Bogstad, B. 2002 Do abioticmechanisms determine interannual variability in length-at-age of juvenile Arcto-Norwegian cod? Can. J. FishAquat. Sci. 59, 57–65. (doi:10.1139/f01-197)
18 Ciannelli, L., Dingsør, G. E., Bogstad, B., Ottersen, G.,
Chan, K. S., Gjøsæter, H., Stiansen, J. E. & Stenseth,N. C. 2007 Spatial anatomy of species survival: effects ofpredation and climate-driven environmental variability.Ecology 88, 635–646. (doi:10.1890/05-2035)
19 Dingsør, G. E., Ciannelli, L., Chan, K. S., Ottersen, G. &
Stenseth, N. S. 2007 Density dependence and density inde-pendence during the early life stages of four marine fishstocks. Ecology 88, 625–634. (doi:10.1890/05-1782)
20 Vikebø, F., Sundby, S., Adlandsvik, B. & Fiksen, Ø. 2005The combined effect of transport and temperature on
distribution and growth of larvae and pelagic juvenilesof Arcto-Norwegian cod. ICES J. Mar. Sci. 62,1375–1386. (doi:10.1016/j.icesjms.2005.05.017)
21 Kristiansen, T., Vikebø, F., Sundby, S., Huse, G. &
Fiksen, Ø. 2009 Modeling growth of larval cod (Gadusmorhua) in large-scale seasonal and latitudinal environ-mental gradients. Deep-Sea Res. II 56, 2001–2011.(doi:10.1016/j.dsr2.2008.11.011)
22 Hurrell, J. W. 1995 Decadal trends in the North Atlantic
Oscillation: regional temperatures and precipitation. Science169, 676–679. (doi:10.1126/science.269.5224.676)
23 Helle, K. & Pennington, M. 1999 The relation of the spatialdistribution of early juvenile cod (Gadus morhua L.) in theBarents Sea to zooplankton density and water flux during
the period 1978–1984. ICES J. Mar. Sci. 56, 15–27.(doi:10.1006/jmsc.1998.0427)
24 Ellertsen, B., Fossum, P., Solemdal, P. & Sundby, S.1989 Relation between temperature and survival ofeggs and first feeding larvae of northeast Arctic cod
(Gadus morhua L.). Rapports et Proces-verbaux desReunions Conseil International pour l’Exploration de la Mer191, 209–219.
25 Stige, L. C., Ottersen, G., Dalpadado, P., Chan, K.-S.,
Hjermann, D. Ø., Lajus, D. L., Yaragina, N. A. &Stenseth, N. C. 2010 Direct and indirect climate forcingin a multi-species marine system. Proc. R. Soc. B 277,3411–3420. (doi:10.1098/rspb.2010.0602)
26 Engedahl, H., Lunde, A., Melsom, A. & Shi, X. B. 2001
New schemes for vertical mixing in MI-POM and MICOM.Research report no. 118. Oslo, Norway: NorwegianMeteorological Institute.
27 Røed, L. & Debernard, J. 2004 Description of an integratedflux and sea-ice model suitable for coupling to an ocean andatmosphere model. Met. no. report 4/2004. Oslo, Norway:Norwegian Meteorological Institute.
28 Melsom, A. & Fossum, I. 2009 Validation of an oceanmodel ensemble for the Barents Sea and the northeasternNordic Seas. Met. no. Report 2/2009. Oslo, Norway:
Norwegian Meteorological Institute.29 Hastie, T. J. & Tibshirani, R. J. 1990 Generalized additive
models. London, UK: Chapman and Hall.30 Ciannelli, L., Chan, K. S., Bailey, K. M. & Stenseth,
N. C. 2004 Nonadditive effects of the environment on

Embracing physics and biology M. Hidalgo et al. 283
on December 23, 2011rspb.royalsocietypublishing.orgDownloaded from
the survival of a large marine fish population. Ecology 85,3418–3427. (doi:10.1890/03-0755)
31 Mammen, E. 1993 Bootstrap and wild bootstrap for high
dimensional linear models in resampling. Ann. Stat. 21,255–285. (doi:10.1214/aos/1176349025)
32 Stige, L. C., Ottersen, G., Brander, K., Chan, K.-S. &Stenseth, N. C. 2006 Cod and climate: effect of theNorth Atlantic Oscillation on recruitment in the North
Atlantic. Mar. Ecol. Prog. Ser. 325, 227–241. (doi:10.3354/meps325227)
33 Jørgensen, C., Dunlop, E. S., Opdal, A. F. & Fiksen, Ø.2008 The evolution of spawning migrations: state depen-
dence and fishing-induced changes. Ecology 89, 3436–3448. (doi:10.1890/07-1469.1)
34 Sundby, S. & Nakken, O. 2008 Spatial shifts in spawninghabitats of Arcto-Norwegian cod related to multidecadalclimate oscillations and climate change. ICES J. Mar. Sci.65, 953–962. (doi:10.1093/icesjms/fsn085)
35 Opdal, A. F. 2010 Fisheries change spawning ground dis-tribution of northeast Arctic cod. Biol. Lett. 6, 261–264.(doi:10.1098/rsbl.2009.0789)
Proc. R. Soc. B (2012)
36 Ottersen, G., Hjermann, D. O. & Stenseth, N. C.2006 Changes in spawning stock structure strengthenthe link between climate and recruitment in a
heavily fished cod (Gadus morhua) stock. FishOceanogr. 15, 230–243. (doi:10.1111/j.1365-2419.2006.00404.x)
37 Kristiansen, T., Drinkwater, K. F., Lough, R. G. &Sundby, S. 2011 Recruitment variability in North
Atlantic cod and match-mismatch dynamics. PLoSONE 6, e17456. (doi:10.1371/journal.pone.0017456)
38 Stige, L. C., Lajus, D. L., Chan, K. S., Dalpadado, P.,Basedow, S. L., Berchenko, I. & Stenseth, N. C. 2009
Climatic forcing of zooplankton dynamics is strongerduring low densities of planktivorous fish. Limnol.Oceanogr. 54, 1025–1036. (doi:10.4319/lo.2009.54.4.1025)
39 Skarðhamar, J., Slagstad, D. & Edvardsen, A. 2007
Plankton distributions related to hydrography andcirculation dynamics on a narrow continental shelf offNorthern Norway. Estuarine Coastal Shelf Sci. 75,381–392. (doi:10.1016/j.ecss.2007.05.044)




















