
Hepatitis B virus-related insertional mutagenesis occurs frequently in human liver cancers and...
10
Hepatitis B virus-related insertional mutagenesis implicates SERCA1 gene in the control of apoptosis Mounia Chami 1,7 , Devrim Gozuacik 1,7 , Kenichi Saigo 1,2,7 , Thierry Capiod 3 , Pierre Falson 4 , Herve´ Lecoeur 5 , Tetsuro Urashima 2 , Jack Beckmann 6 , Marie-Lyse Gougeon 5 , Michel Claret 3 , Marc le Maire 4 , Christian Bre´chot 1 and Patrizia Paterlini-Bre´chot* ,1 1 U-370 INSERM, Necker Institute, 75015 Paris, France; 2 Second Department of Surgery, Chiba University, Chiba 263-8852, Japan; 3 U-442 INSERM, Universite´ Paris Sud, 91405 Orsay, France; 4 URA CNRS 2096, CEA Saclay, 91191 Gif sur Yvette, France; 5 Pasteur Institute, 75015 Paris, France; 6 Centre National Ge ´notypage, 91057 Evry, France We have used the Hepatitis B Virus DNA genome as a probe to identify genes clonally mutated in vivo, in human liver cancers. In a tumor, HBV-DNA was found to be integrated into the gene encoding Sarco/Endoplas- mic Reticulum Calcium ATPase (SERCA), which pumps calcium, an important intracellular messenger for cell viability and growth, from the cytosol to the endoplasmic reticulum. The HBV X gene promoter cis-activates chimeric HBV X/SERCA1 transcripts, with splicing of SERCA1 exon 11, encoding C-terminally truncated SERCA1 proteins. Two chimeric HBV X/SERCA1 proteins accumulate in the tumor and form dimers. In vitro analyses have demonstrated that these proteins localize to the ER, determine its calcium depletion and induce cell death. We have also shown that these biological eects are related to expression of the SERCA, rather than of the viral moiety. This report involves for the first time the expression of mutated SERCA proteins in vivo in a tumor cell proliferation and in vitro in the control of cell viability. Oncogene (2000) 19, 2877 – 2886. Keywords: SERCA1; Hepatitis B Virus; calcium; apoptosis; cancer Introduction Chronic HBV infection is a major etiologic factor of hepatocellular carcinoma (HCC) (Chang et al., 1997; Hildt et al., 1996). Although no transforming viral oncogene has so far been identified, HBV-induced chronic liver inflammation is thought to play an important role in carcinogenesis (Buendia, 1992; Paterlini and Bre´chot, 1994). In addition, the expres- sion of HBV X, PreS2/S (Kekule´, 1994), and PreS1/ PreS2/S (Chisari et al., 1989) viral proteins has been associated with the development of HCC. In particular, the 17 kDa HBV X encoded protein has been shown to activate in trans a wide variety of cellular and viral genes (reviewed in Weil et al., 1999). HBV X activates signal transduction pathways such as Ras/Raf/MAP kinase (Benn and Schneider, 1994), Jak1-STAT (Lee and Yun, 1998) and NF-kB signaling (Chirillo et al., 1996). It has also been suggested that it directly interacts with cellular proteins controlling cell growth (p53 (Truant et al., 1995; Wang et al., 1994a)), DNA repair (ERCC (Wang et al., 1994b) and UVDDB (Lee et al., 1995; Sitterlin et al., 1997)), senescence (Sun et al., 1998), NFkB activity (Weil et al., 1999), basal transcription (RBP5 (Cheong et al., 1995) and RMP (Dorjsuren et al., 1998)), signal transduction (ATF/ CREB (Williams and Andrisani, 1995)) and with proteasome (Sirma et al., 1998). Depending on the experimental conditions, HBV X has also been reported to accelerate (Koike et al., 1994) or inhibit cell cycle progression (Benn and Schneider, 1995), and to induce (Chirillo et al., 1997; Sirma et al., 1999; Terradillos et al., 1998) or inhibit apoptosis (Wang et al., 1995). Furthermore, HBV X induces HCC in certain transgenic mice (Kim et al., 1991). However, in a dierent genetic context, HBV X transgenic mice exhibit only increased susceptibility to chemical carcinogenes (Slagle et al., 1996), or an accelerated development of c-myc induced HCC (Terradillos et al., 1997). In addition to these mechanisms, the clonal integra- tion of HBV-DNA into the host cell genome has been found in more than 90% of HBV-related HCCs (Paterlini and Bre´ chot, 1994). HBV-DNA integration may directly promote genetic instability or lead to cis- activation of growth-related genes (Paterlini and Bre´ chot, 1994). In certain isolated cases, the viral genome was found to be integrated into the Retinoic acid receptor b gene (Dejean et al., 1986), and Cyclin A2 gene (Wang et al., 1990), leading to the expression of chimeric transcripts and proteins, the transforming activity of which was subsequently proved (Berasain et al., 1998; Garcia et al., 1993). Reports have also been made of HBV-DNA integration in the gene encoding mevalonate kinase in the PLC/PRF/5 cell line (Graef et al., 1994), and in the Carboxypeptidase N locus in a human HCC (Pineau et al., 1996). In both instances, chimeric transcripts were identified in the tumor cells, but the eect of their in vitro expression on cell phenotype has not been investigated so far. On the other hand, a number of studies have failed to identify HBV-DNA integration into cellular DNA coding sequences, so that insertional mutagenesis has been considered as a rare event in HBV-related carcinogen- esis (Koike, 1998). In contrast, in the woodchuck animal model, Woodchuck hepadnavirus (WHV) integration into the Oncogene (2000) 19, 2877 – 2886 ª 2000 Macmillan Publishers Ltd All rights reserved 0950 – 9232/00 $15.00 www.nature.com/onc *Correspondence: P Paterlini-Bre´chot, Unite´ INSERM 370, 156 rue de Vaugirard, 75015 Paris, France 7 The first three authors contributed equally to this work Received 12 November 1999; revised 3 April 2000; accepted 4 April 2000
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Hepatitis B virus-related insertional mutagenesis occurs frequently in human liver cancers and...

Hepatitis B virus-related insertional mutagenesis implicates SERCA1 genein the control of apoptosis
Mounia Chami1,7, Devrim Gozuacik1,7, Kenichi Saigo1,2,7, Thierry Capiod3, Pierre Falson4,Herve Lecoeur5, Tetsuro Urashima2, Jack Beckmann6, Marie-Lyse Gougeon5, Michel Claret3,Marc le Maire4, Christian Bre chot1 and Patrizia Paterlini-Bre chot*,1
1U-370 INSERM, Necker Institute, 75015 Paris, France; 2Second Department of Surgery, Chiba University, Chiba 263-8852,Japan; 3U-442 INSERM, Universite Paris Sud, 91405 Orsay, France; 4URA CNRS 2096, CEA Saclay, 91191 Gif sur Yvette,France; 5Pasteur Institute, 75015 Paris, France; 6Centre National GeÂnotypage, 91057 Evry, France
We have used the Hepatitis B Virus DNA genome as aprobe to identify genes clonally mutated in vivo, inhuman liver cancers. In a tumor, HBV-DNA was foundto be integrated into the gene encoding Sarco/Endoplas-mic Reticulum Calcium ATPase (SERCA), which pumpscalcium, an important intracellular messenger for cellviability and growth, from the cytosol to the endoplasmicreticulum. The HBV X gene promoter cis-activateschimeric HBV X/SERCA1 transcripts, with splicing ofSERCA1 exon 11, encoding C-terminally truncatedSERCA1 proteins. Two chimeric HBV X/SERCA1proteins accumulate in the tumor and form dimers. Invitro analyses have demonstrated that these proteinslocalize to the ER, determine its calcium depletion andinduce cell death. We have also shown that thesebiological e�ects are related to expression of theSERCA, rather than of the viral moiety. This reportinvolves for the ®rst time the expression of mutatedSERCA proteins in vivo in a tumor cell proliferation andin vitro in the control of cell viability. Oncogene (2000)19, 2877 ± 2886.
Keywords: SERCA1; Hepatitis B Virus; calcium;apoptosis; cancer
Introduction
Chronic HBV infection is a major etiologic factor ofhepatocellular carcinoma (HCC) (Chang et al., 1997;Hildt et al., 1996). Although no transforming viraloncogene has so far been identi®ed, HBV-inducedchronic liver in¯ammation is thought to play animportant role in carcinogenesis (Buendia, 1992;Paterlini and Bre chot, 1994). In addition, the expres-sion of HBV X, PreS2/S (Kekule , 1994), and PreS1/PreS2/S (Chisari et al., 1989) viral proteins has beenassociated with the development of HCC. In particular,the 17 kDa HBV X encoded protein has been shown toactivate in trans a wide variety of cellular and viralgenes (reviewed in Weil et al., 1999). HBV X activatessignal transduction pathways such as Ras/Raf/MAPkinase (Benn and Schneider, 1994), Jak1-STAT (Lee
and Yun, 1998) and NF-kB signaling (Chirillo et al.,1996). It has also been suggested that it directlyinteracts with cellular proteins controlling cell growth(p53 (Truant et al., 1995; Wang et al., 1994a)), DNArepair (ERCC (Wang et al., 1994b) and UVDDB (Leeet al., 1995; Sitterlin et al., 1997)), senescence (Sun etal., 1998), NFkB activity (Weil et al., 1999), basaltranscription (RBP5 (Cheong et al., 1995) and RMP(Dorjsuren et al., 1998)), signal transduction (ATF/CREB (Williams and Andrisani, 1995)) and withproteasome (Sirma et al., 1998). Depending on theexperimental conditions, HBV X has also beenreported to accelerate (Koike et al., 1994) or inhibitcell cycle progression (Benn and Schneider, 1995), andto induce (Chirillo et al., 1997; Sirma et al., 1999;Terradillos et al., 1998) or inhibit apoptosis (Wang etal., 1995). Furthermore, HBV X induces HCC incertain transgenic mice (Kim et al., 1991). However, ina di�erent genetic context, HBV X transgenic miceexhibit only increased susceptibility to chemicalcarcinogenes (Slagle et al., 1996), or an accelerateddevelopment of c-myc induced HCC (Terradillos et al.,1997).
In addition to these mechanisms, the clonal integra-tion of HBV-DNA into the host cell genome has beenfound in more than 90% of HBV-related HCCs(Paterlini and Bre chot, 1994). HBV-DNA integrationmay directly promote genetic instability or lead to cis-activation of growth-related genes (Paterlini andBre chot, 1994). In certain isolated cases, the viralgenome was found to be integrated into the Retinoicacid receptor b gene (Dejean et al., 1986), and CyclinA2 gene (Wang et al., 1990), leading to the expressionof chimeric transcripts and proteins, the transformingactivity of which was subsequently proved (Berasain etal., 1998; Garcia et al., 1993). Reports have also beenmade of HBV-DNA integration in the gene encodingmevalonate kinase in the PLC/PRF/5 cell line (Graef etal., 1994), and in the Carboxypeptidase N locus in ahuman HCC (Pineau et al., 1996). In both instances,chimeric transcripts were identi®ed in the tumor cells,but the e�ect of their in vitro expression on cellphenotype has not been investigated so far. On theother hand, a number of studies have failed to identifyHBV-DNA integration into cellular DNA codingsequences, so that insertional mutagenesis has beenconsidered as a rare event in HBV-related carcinogen-esis (Koike, 1998).
In contrast, in the woodchuck animal model,Woodchuck hepadnavirus (WHV) integration into the
Oncogene (2000) 19, 2877 ± 2886ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00
www.nature.com/onc
*Correspondence: P Paterlini-Bre chot, Unite INSERM 370, 156 ruede Vaugirard, 75015 Paris, France7The ®rst three authors contributed equally to this workReceived 12 November 1999; revised 3 April 2000; accepted 4 April2000

N-myc2 and c-myc oncogenes is a frequent event,occurring in more than 70% of tumors, and itstransforming e�ect has been well established (Buendia,1992). Interestingly, in some tumors, WHV-DNAintegration occurs in a locus called win, which issituated at more than 150 Kb from the N-myc2 gene(Fourel et al., 1994).
In HBV-related, human HCC, however, the actualimpact of insertional mutagenesis has not yet beenexplored exhaustively. This is due to technical limita-tions of the genomic libraries previously used foridentifying HBV-DNA integration sites, which pre-cluded large scale screening.
Using an original, Alu-PCR-based approach devel-oped in our laboratory (Minami et al., 1995), we haveinitiated a screening program of HBV-related HCC,searching for HBV/cellular DNA junctions. In thepresent study, we report on a liver tumor cellproliferation carrying the HBV-DNA integration intothe SERCA1 gene.
SERCA proteins play a pivotal role in regulatingcellular calcium (Pozzan et al., 1994), which, in turn,acts as an intracellular messenger involved in a broadrange of specialized and basic cellular activities,ranging from muscle contraction, transmission ofneuronal signal, cell proliferation and death (Berridgeet al., 1998). Three distinct SERCA genes encode anumber of di�erentially expressed isoforms (Wu et al.,1995). SERCA1a and 1b are mainly found in fast-twitch skeletal muscle, SERCA2a in slow-twitchskeletal and cardiac muscle, SERCA2b is ubiquitousand SERCA3 which also displays isoforms is expressedin several tissues, including hemopoietic, endothelialand secretory epithelial cells. We show here that HBVintegration into the SERCA1 gene in a human livertumor cis-activates SERCA1 chimeric transcriptsencoding chimeric proteins. We also provide evidencethat the in vitro expression of these transcripts and oftheir SERCA moiety induces ER calcium depletion andapoptosis. Our study therefore highlights the implica-tion of SERCA1 gene mutation in clonal cell expansionand the control of cell viability.
Results
HBV-DNA integration into the SERCA1 gene drives theexpression of chimeric transcripts
By using an HBV X speci®c and an Alu-speci®c primer(Minami et al., 1995), we have isolated a PCRfragment of 498 bp from a liver tumorous DNA(T86). In this fragment, the HBV X gene was fusedin frame with SERCA1 gene exon 3 (67 bp) (Zhang etal., 1995) and intron 3 (247 bp) (MacLennan, personalcommunication) (Figure 1A).
A Northern blot analysis performed on tumorous(T86) and non tumorous mRNAs showed a 3.2 kbtranscript, hybridizing with both HBV and SERCA1probes, and expressed in T86 but not in adjacent, non-tumorous liver tissue (Figure 1B). A cDNA library,constructed with poly(A)+ mRNAs from T86, yieldedeight recombinant clones (Figure 1C). Their 5' endsshowed the same HBV X sequence, starting at nt 1309HBV (adr4 subtype) (Raney and McLachlan, 1991),64 bp before the HBV X ATG, up to nucleotide 1819
(446 in Figure 1A), fused in frame to SERCA1 cDNAnucleotide 153 in exon 3. Interestingly, SERCA1cDNA was characterized by the splicing of exon 11,leading to a 22 codon frameshift and a premature stopcodon in exon 12. Three of eight clones exhibited anadditional splicing of SERCA1 exon 4. These twochimeric cDNAs and encoded proteins will be referredto as Xt/St+4 and Xt/St74: Xt for HBV X truncatedsequence fused to truncated SERCA1 (St), with orwithout exon 4.
Chimeric HBV X/SERCA1 proteins predicted structureprevents calcium pumping
According to the cDNA sequence, the chimericproteins have their SERCA1 N-terminal 51 aminoacidsreplaced by 148 aminoacids of the HBV X protein andan additional residue encoded by the viral/cellular inframe junction (Figure 2A). Xt/St+4 protein lacks sixputative SERCA1 transmembrane segments (M5-M10),which include ®ve of the six Ca2+ binding residues(Glu-771, Asn-796, Thr-799, Asp-800, and Glu-908)(MacLennan et al., 1997) as well as the cytoplasmicloop between transmembrane segments 6 and 7 thatalso controls Ca2+ binding (Falson et al., 1997). Amutated peptide of 22 aminoacids is encoded at the C-terminal end. In addition to that, Xt/St-4 protein lacksa peptide (aa. 74 ± 108) encoding the second putativetransmembrane segment (M2) and the last ®ve C-terminal residues of M1. The expected size of the Xt/St+4 and Xt/St74 chimeric proteins is 56 and53 kDa, respectively.
Thus, although these proteins retain the putativecalcium binding residue Glu 309 and the phosphory-latable Asp-351, consistent with previously reportedmutants (MacLennan et al., 1998), they cannotfunction as calcium pumps.
Chimeric proteins are expressed and form dimers in thetumor
We then looked for these chimeric and truncatedproteins in the tumorous tissue (T86) by Western blot.A chimeric protein (53/56 kDa), reacting with bothanti-HBV X (anti-X) and anti-SERCA1 antibodies(Figure 2B), was clearly detectable in the tumoroustissue. High resolution analysis demonstrated twoproteins, of 53 and 56 kDa (Figure 2C), which sizewas consistent with the cDNA sequences. We con-sistently observed a weaker upper band (56 kDa), ascompared to the strongest lower band (53 kDa). Thiscontrasted with the higher number of clones (5/8)encoding the 56 kDa protein. This discrepancy may bedue to the non representative number of clones or todi�erent conformation and reactivity to the antibodyof the two proteins.
Oligomerization of SERCA proteins has beenalready reported (Martonosi, 1996). Therefore, in thissetting, we looked for heterodimers of endogenous liverSERCA isoform (SERCA2b) and chimeric HBV X/SERCA1 proteins in the tumorous tissue (Figure 2C).Anti-SERCA1 antibody showed absence of hetero-logous aggregates of SERCA2b with chimeric proteins(predicted size: 163 ± 166 kDa) and, interestingly re-vealed prominent aggregates of chimeric proteins(106 kDa), possibly corresponding to chimeric dimers,
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2878
Oncogene

in non-heated tumor sample, whereas SERCA1 innormal fast skeletal muscle (SR) underwent only slightaggregation (220 kDa) under the same conditions. Theexpression of SERCA2b was sustained in T86 at a level
which was not signi®cantly lower than that found inthree, di�erent, normal livers (Figure 2D) and cell linestransfected with the Xt/St constructs (data not shown).SERCA3 mRNA was expressed at very low and similar
Figure 1 Integration of HBV into SERCA1 gene in a tumorous tissue (T86) and expression of chimeric transcripts. (A) Structureof the PCR product showing the HBV integration in frame into the third exon of SERCA1 gene. HBV speci®c primer (MD26C),and Alu speci®c primer (AluTag 5) were used to amplify the HBV/cellular DNA junction: 8 indicates the 1819th nucleotide of theHBV genome (adr4 genotype), * indicates the 153 nucleotide of SERCA1 cDNA sequence. The amino acid mutated by the HBVintegration is underlined. The SERCA1 gene structure is depicted below. Exonic sequences are represented as black boxes. (B)Expression of chimeric HBV X/SERCA1 mRNA in T86. The same Northern blot of poly(A)+ RNA (10 mg) from various tissues(M, muscle (positive control for SERCA1 probe); T86, tumorous tissue T86; NT, adjacent non tumorous tissue NT86; NL, normalliver and HepG2
+, HepG2 cells transfected with HBV (positive control for HBV probe)) was hybridized with SERCA1 and HBVprobe, and then normalized with the GAPDH probe. A 3.2 kb transcript hybridizing with both HBV and SERCA1 probe is presentin T86 (arrow), but not in NT. In T86 the HBV probe also shows the expression of the 2.1 kb HBV surface transcript. (C) HybridcDNAs cloned from T86. Xt/St+4, clones with splicing of SERCA1 exon 11 (-ex11) leading to a frameshift (black box) and a stopcodon in exon 12. Xt/St74, clones with exon 11 and exon 4 (-ex4) splicing. X, HBV X sequence
Oncogene
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2879

levels in both tumorous and corresponding non-tumorous tissues (data not shown).
Chimeric SERCA proteins are localized to the ER
We then investigated whether chimeric proteinslocalize to the ER. According to their predictedstructure, they lack 51 (Xt/St+4) or 51+34 (Xt/St74) N-terminal SERCA1 residues, which includethe 28 N-terminal amino acids sequence previouslyreported as the minimal domain for SERCA proteinsretention in the ER (Guerini et al., 1998). Constructsexpressing chimeric proteins, their HBV X andSERCA1 moieties, as well as wild type HBV X andSERCA1 proteins (Figure 3) were cloned in expression
vectors with or without fusion to GFP or Myc tagsequences.
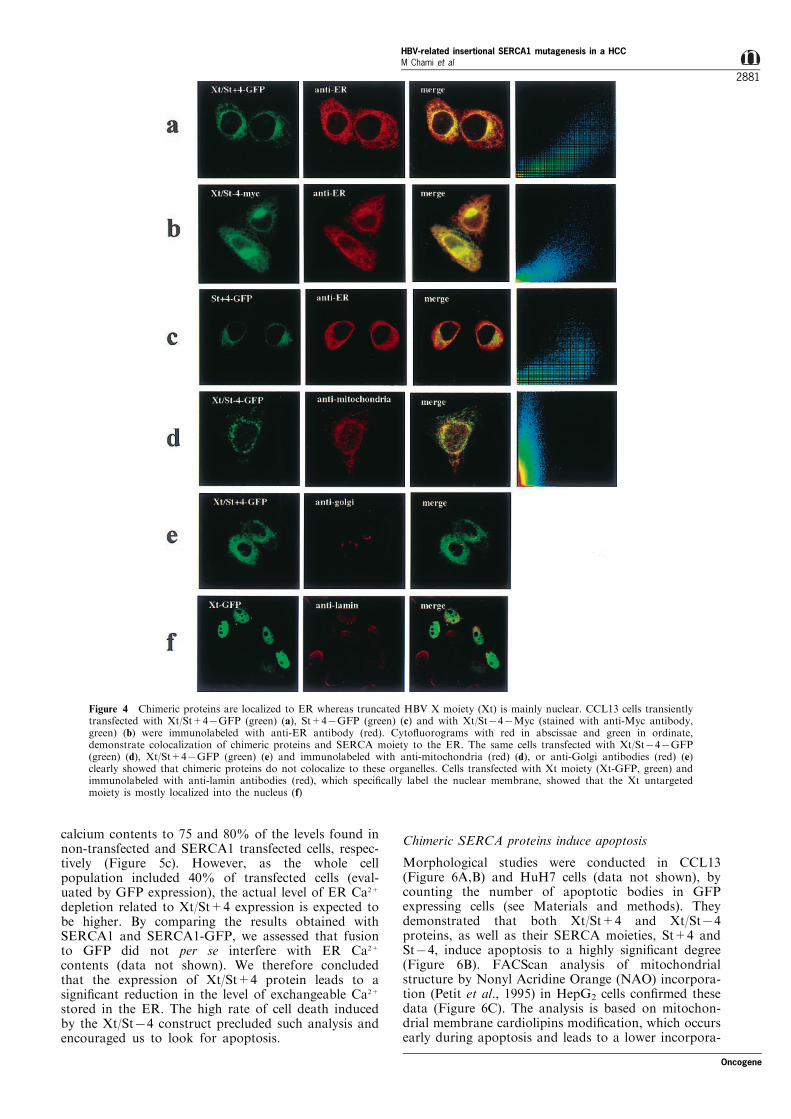
The subcellular distribution of the encoded proteinswas studied in transiently transfected CCL13 cells,using immuno¯uorescence and scanning confocalmicroscopy (Figure 4). Xt/St+4 and Xt/St74 pro-teins, as well as their SERCA moieties (St+4 andSt74), fused to GFP (Figure 4a,c) or to the Mycpeptide (Figure 4b), colocalized with ER (Figure 4a ±c), but not with mitochondria and golgi (Figure 4d,e).The untargeted Xt moiety was mostly nuclear (Figure4f), without any ER localization, indicating that theXt part of the chimeric proteins is targeted to the ERby the SERCA moiety. This result led us to clone theXt sequence downstream to a prolactin cassette inorder to obtain its targeting to the ER (Figure 3:Xtpro).
Chimeric SERCA proteins induce depletion of ER Ca2+
pools
We ®rstly tried to analyse the rate of calcium uptakeon isolated microsomal fractions. We tested whetherwe might use transiently transfected cells, sorted byFACS to obtain equivalent amounts of microsomesfrom cells transfected with di�erent constructs.However, we noticed a massive cell death in cellstransfected with chimeric cDNAs after sorting,avoiding to perform the test. We then investigatedwhether chimeric proteins induce, upon in vitroexpression, modi®cations of the ER and cytosolicCa2+ pools (Figure 5). Indeed, if the SERCA pumpactivity is reduced, the ER free calcium content willreach a new equilibrium at lower Ca2+ levels. Thecytosolic free [Ca2+]i concentration was studied in cellstransiently expressing Xt/St+4-GFP or SERCA1-GFPconstructs and in non-transfected cells (NTC). Nosigni®cant di�erence was found in resting cytosolic[Ca2+]i in Xt/St+4 transfected cells (64+4 nM, n=16)when compared to controls (NTC: 61+4 nM, n=18and SERCA1: 66+5 nM, n=22) (Figure 5b). Incontrast, Xt/St+4 expression decreased the ER
Figure 2 Chimeric proteins are expressed and form dimers in thetumorous tissue (T86). (A) A schematic representation of chimericproteins: numbers below drawings indicate transmembranedomains. D351: phosphorylatable aspartic acid 351. (.) trans-membrane and (o) cytoplasmic calcium binding residues. TheHBV X is depicted as broken line, the peptide created byframeshift is shown as a dotted line and the peptide encoded byexon 4 and deleted in Xt/St74 is in gray. (B) Chimeric protein(X/SERCA) is expressed in T86. The same Western blot (14%SDS±PAGE) was successively hybridized with anti-HBV X andanti-SERCA1 antibodies. X Ag (21 kDa), puri®ed recombinantHBV X protein (a: 100 ng, b: 50 ng, c: 50 ng mixed with 10 mg ofnormal liver microsomes); T86, T86 microsomes (50 mg); SR,SERCA1 from rabbit muscle microsomes (50 ng). (C) Westernblot (7% SDS±PAGE) of heated (h) and non-heated (nh)microsomes under mildly denaturing conditions (NL and T86:50 mg; SR: 20 ng). A band of 106 kDa corresponding to chimericproteins dimers is present in the T86 non heated sample. (D)SERCA2b protein expression in T86 compared to three normallivers (NL1-3). Western blot analysis was performed on the totalprotein and normalized with anti-tubulin antibody. TheSERCA2b accumulation was not signi®cantly di�erent in T86as compared to normal livers
Figure 3 Schematic representation of the di�erent clonedconstructs. Xt/St+4 and Xt/St74: chimeric clones correspondingto chimeric cDNAs; St+4 and St74, SERCA moieties ofchimeric clones; Xt, HBV X moiety; Xtpro, HBV X moietydirected to the ER membrane by means of the prolactin cassette
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2880
Oncogene
calcium contents to 75 and 80% of the levels found innon-transfected and SERCA1 transfected cells, respec-tively (Figure 5c). However, as the whole cellpopulation included 40% of transfected cells (eval-uated by GFP expression), the actual level of ER Ca2+
depletion related to Xt/St+4 expression is expected tobe higher. By comparing the results obtained withSERCA1 and SERCA1-GFP, we assessed that fusionto GFP did not per se interfere with ER Ca2+
contents (data not shown). We therefore concludedthat the expression of Xt/St+4 protein leads to asigni®cant reduction in the level of exchangeable Ca2+
stored in the ER. The high rate of cell death inducedby the Xt/St74 construct precluded such analysis andencouraged us to look for apoptosis.
Chimeric SERCA proteins induce apoptosis
Morphological studies were conducted in CCL13(Figure 6A,B) and HuH7 cells (data not shown), bycounting the number of apoptotic bodies in GFPexpressing cells (see Materials and methods). Theydemonstrated that both Xt/St+4 and Xt/St74proteins, as well as their SERCA moieties, St+4 andSt74, induce apoptosis to a highly signi®cant degree(Figure 6B). FACScan analysis of mitochondrialstructure by Nonyl Acridine Orange (NAO) incorpora-tion (Petit et al., 1995) in HepG2 cells con®rmed thesedata (Figure 6C). The analysis is based on mitochon-drial membrane cardiolipins modi®cation, which occursearly during apoptosis and leads to a lower incorpora-
Figure 4 Chimeric proteins are localized to ER whereas truncated HBV X moiety (Xt) is mainly nuclear. CCL13 cells transientlytransfected with Xt/St+47GFP (green) (a), St+47GFP (green) (c) and with Xt/St747Myc (stained with anti-Myc antibody,green) (b) were immunolabeled with anti-ER antibody (red). Cyto¯uorograms with red in abscissae and green in ordinate,demonstrate colocalization of chimeric proteins and SERCA moiety to the ER. The same cells transfected with Xt/St747GFP(green) (d), Xt/St+47GFP (green) (e) and immunolabeled with anti-mitochondria (red) (d), or anti-Golgi antibodies (red) (e)clearly showed that chimeric proteins do not colocalize to these organelles. Cells transfected with Xt moiety (Xt-GFP, green) andimmunolabeled with anti-lamin antibodies (red), which speci®cally label the nuclear membrane, showed that the Xt untargetedmoiety is mostly localized into the nucleus (f)
Oncogene
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2881

tion of NAO. The percentage of apoptotic cells wasestimated according to lower NAO incorporationassociated to smaller cell size. These results led us toconclude that both chimeric constructs induce apopto-sis in three di�erent cell lines. In an independentexperiment we also tested, by FACS analysis, whethertransient transfection of HepG2 cells with a constructencoding a truncated SERCA protein having a normalN-terminal end and a truncated C-terminal end(SERCA1+4711) was associated to apoptosis. Ascompared with the empty vector, the SERCA1+4711construct induced apoptosis at a statistically signi®cantlevel (22% vs 6%).
Targeting of Xt to the ER abrogates its proapoptotic andtransactivating property
By using both morphological counting of apoptoticbodies (Figure 6B) and FACScan analysis of mito-chondrial membrane by NAO incorporation (Figure6C), we observed that both the wild type (Xwt) and thetruncated untargeted HBV X moiety (Xt) induceapoptosis in three di�erent cell lines. In contrast withthese ®ndings, the ER targeted HBV X moiety (Xtpro)barely induced apoptosis, reaching non signi®cant(Figure 6B) or border-line (Figure 6C) levels. Thusthe proapoptotic e�ect of chimeric constructs is mainlyrelated to their SERCA moiety. Since the proapoptotice�ect of HBV X has been associated to its transactivat-ing property, we analysed the transactivating e�ect ofXtpro as compared to Xwt and to the Xt moiety. Inour system (see Materials and methods), Xwt displayedtransactivating activity on NFkB reporter gene (Sirmaet al., 1999). In contrast with these ®ndings, we werenot able to evidence a signi®cant transactivatingactivity of Xtpro and Xt/St+4 in triplicate experiments(Figure 7).
Discussion
Our work reports on a new case of cis-activation of acellular gene upon HBV-DNA integration in a humanhepatocellular carcinoma. It also identi®es for the ®rsttime SERCA1 as a potential target for mutation in ahuman cancer.
SERCA2b is the endogenous SERCA in livertissues, the SERCA1 gene expression being detectableonly by RT±PCR. In the liver tumor we studied, theviral HBV X gene promoter cis-activates two chimericHBV X/SERCA1 transcripts, characterized by exon 11splicing and a premature stop codon in exon 12.These transcripts encode chimeric proteins with theirC-terminal end truncated and their N-terminal endreplaced by the C-terminally truncated HBV Xprotein.
Figure 5 Chimeric proteins signi®cantly reduce the ER Ca2+
pool. (a) Estimation of ER and non-ER Ca2+ contents (seeMaterials and methods). (b) The size of the TBuBHQ-sensitiveER Ca2+ pool (ER [Ca2+]) was signi®cantly reduced in Xt/St+4-transfected cells compared to non-transfected cells (NTC) andSERCA1 transfected cells. No signi®cant change in the non-ERCa2+ pool was detected under any of the three experimentalconditions. (c) The ER Ca2+ pool/total mobilizable Ca2+ ratiowas 51+3% in NTC cells, 38+2% in Xt/St+4 transfected cellsand 47+3% in SERCA1 transfected cells. Results are expressedas mean+s.e.m. *, P50.001; **, P50.02 (Student t-test)
Figure 6 Chimeric proteins induce apoptosis in transfected cells.(A) Apoptotic bodies (arrows) were visualized by confocalmicroscopy (scale bars: 10 mm) 48 h after transfection of CCL13cells. (B) Percentage of apoptotic cells recorded in at least 300transfected CCL13 cells; values are the mean of two independentexperiments. *, P50.001; **, P50.0001; NS, not signi®cant,calculated vs GFP transfected cells value. GFP, cells transfectedwith the empty pEGFP-N1 vector. (C) FACScan analysis ofapoptosis in HepG2 transfected cells by mitochondrial incorpora-tion of NAO (Nonyl-Acridine Orange) (ordinate) and cell size(abscissae). The percentage of apoptotic cells (black dots), withlower NAO incorporation associated to a smaller cell size, isshown in each case. NTC, non transfected cells, pcDNA-3, cellstransfected with empty pcDNA3.1 vector
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2882
Oncogene

When compared with the functional structure of wildtype SERCA1, HBV X/SERCA1 proteins have lostcalcium binding residues and the ATP binding domain.They are therefore unable to pump calcium. Further,they have also lost 51 N-terminal residues, includingthe 28 residues previously identi®ed as the minimalSERCA1 ER targeting signal (Guerini et al., 1998).Using immuno¯uorescence and confocal analysis, wehave shown that these chimeric proteins still localize tothe ER, and do not colocalize with mitochondria orGolgi. Since previous reports (Bayle et al., 1997)showed that the ®rst four predicted SERCA transmem-brane domains, which are retained in the chimericprotein, act as signal anchor and stop transfersequences, chimeric proteins are presumed to beinserted into the ER membrane. We cannot, however,rule out misfolding of these proteins into the ERlumen.
With respect to HBV X localization, our results raisetwo interesting points. Wild type HBV X shows apredominant cytoplasmic localization (Sirma et al.,1998) and it is unclear whether a small fractiontranslocates or not to the nucleus (Weil et al., 1999).It has been recently shown that large C-terminaldeletions promote HBV X nuclear localization (Sirmaet al., 1999). Although localization signals have notbeen mapped to date, our study shows that deletion ofonly ®ve C-terminal residues is su�cient to drive HBVX to the nucleus. Moreover, we have shown that, inthe chimeric proteins, the HBV X moiety is driven tothe ER by the SERCA moiety.
Cells transiently transfected with the chimericconstruct display a signi®cant depletion of ER calcium.Several di�erent, non-exclusive mechanisms mayaccount for this depletion. Chimeric proteins maydownregulate endogenous SERCA2b expression. How-ever, we failed to demonstrate a signi®cantly decreasedexpression of SERCA2b in the tumorous tissue (T86)as compared to normal livers and in cells transfectedwith the chimeric constructs. SERCA proteins havebeen shown to dimerize. Chimeric proteins may form
heterodimers or aggregates with endogenous SERCA2bproteins and thus alter their biological activity.However, heterodimers were not detectable by Westernblot, even under weak denaturing conditions. Instead,dimers of chimeric proteins were found in thetumorous tissue. HBV X may have favored thisdimerization in our model, since in vitro expressedHBV X has indeed been shown to dimerize (Lin andLo, 1989). These results suggest that chimeric proteinsmay determine calcium leakage from the ER byembedding the ER membrane (ER overload) (Pahland Baeuerle, 1997) or by forming a cation pore upondimerization of the two M4 transmembrane segments,which contain a calcium binding residue (Rice andMacLennan, 1996). Finally, calcium release may alsofollow the accumulation of unfolded proteins in theER, leading to an unfolded protein response (Kauf-man, 1999). The chimeric SERCA proteins areexpected, however, to be inserted into the ERmembrane. Overexpression of the SERCA moietiesinduce apoptosis, while overexpression of the Xtproconstruct, which is also inserted into the ERmembrane, does not. In agreement with this result,we have also observed that overexpression of the Pro-GFP construct does not induce a decreased ERcalcium content (data not shown). Finally, overexpres-sion of truncated SERCA proteins (having the normalSERCA1 N-terminal end, and therefore presumablycorrectly folded) induce apoptosis in the same propor-tions as compared with the chimeric SERCA proteins.Therefore, even if we cannot formally rule out a minorcontribution of ER overload and/or unfolding proteinresponse to the ER calcium depletion and apoptosis,our data demonstrate that mutated SERCAs areinvolved in the control of apoptosis.
ER calcium depletion has been shown to causerepression of protein synthesis and folding (Brostromand Brostrom, 1990) which can induce apoptosis(Nicotera and Orrenius, 1998). SERCA genes, includ-ing SERCA1, have consistently been shown to controlInositol 1,4,5-triphosphate (IP3)-mediated calciumwaves (Camacho and Lechleiter, 1993; Morgan andJacob, 1998) and it has been recently shown thatcalcium wave amplitude and duration di�erentiallyactivate certain transcription factors (Dolmetsch et al.,1997). In this setting, modi®cation of the SERCAfunction, in the tumor we studied, could havederegulated cell viability and growth. In fact, thetransient expression of chimeric proteins induced celldeath in three HCC-derived cell lines. In line withprevious results from our and other laboratories, wealso found that the wild type HBV X protein inducesapoptosis. The HBV X moiety, which has lost only ®veC-terminal residues as compared to the wild type form,also demonstrated a proapoptotic e�ect, but only in itsfree form. Namely, apoptotic activity was lost bytargeting the HBV X moiety to the ER through aprolactin cassette. As with previously reported data,showing that the proapoptotic e�ect of HBV Xcorrelates with its transactivation activity, we foundthat targeting the HBV X moiety to the ERsigni®cantly reduced its transactivating property.
Finally, transient transfection of the SERCA moi-eties massively induced apoptosis. Taken together,these data demonstrate that the SERCA moietiesaccount for most of the apoptotic activity due to
Figure 7 Targeting of HBV X moiety (Xt) to the ER decreasesits transactivating activity. CCL13 cells were co-transfected withpcDNA3.1 empty vector (pcDNA-3), wild type HBV X (Xwt),Xt/St+4 chimeric construct, and Xtpro (Xt targeted to the ER)expression plasmids and with reporter vector p-NF-kB-LUC andp-b-actin-b-galactosidase (see Materials and methods). A de-creased transactivating activity was observed with Xtpro and thechimeric construct Xt/St+4, both targeting the Xt to the ER, ascompared to the wild type HBV X protein (Xwt)
Oncogene
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2883

chimeric proteins. In agreement with our results, Ma etal. (1999) have recently provided evidence for a directimplication of SERCA2a in apoptotic cell death.
This is the ®rst report demonstrating that directmutation of the SERCA1 gene a�ects the control ofcell death. SERCA1 gene mutations have beenreported in some patients with Brody disease (Mac-Lennan et al., 1998) characterized by autosomalrecessive inheritance of SERCA1 (ATP2A1) genemutations on chromosome 16p12.1 ± 12.2. In thesepatients, muscle relaxation is impaired because of theexpression of mutated SERCA1 proteins, which areunable to pump calcium. Recently, SERCA2 genemutations have been reported in Darier-White disease(Sakuntabhai et al., 1999), a rare dermatosis withdominant inheritance characterized by focal areas ofseparation between suprabasal epidermal cells, andabnormal keratinocytes di�erentiation. Darier-Whitedisease SERCA2 mutations therefore highlight the factthat aberrations of intracellular calcium levels may leadto a distinct program of altered cell growth anddi�erentiation (Peacocke and Christiano, 1999).
Our work constitutes the ®rst report on an in vivoSERCA1 gene mutation occurring selectively intumorous cells and not in adjacent, non tumorousliver tissue. Chromosomal aberrations involving the16p12 region, which contains the SERCA1 gene, havebeen detected in several tumors, including pancreaticendocrine tumors, colorectal carcinomas and semino-mas (Chung et al., 1998; Ottesen et al., 1997; Pirc-Danoewinata et al., 1996). Thus, depending on thecellular environment, mutations of SERCA1 mightderegulate cell viability and growth.
Our data provide new information concerning theimpact of HBV-related insertional mutagenesis. Thismechanism has been considered as a rare event inhuman liver cancer. The number of tumors in whichHBV integrants were studied in detail has remained,however, low (around 30 in a 20-year period). We havetherefore developed an HBV-Alu PCR based techniqueto rapidly isolate viral/cellular junctions (Minami et al.,1995). This approach has allowed us to isolate 18HBV/cellular DNA junction fragments; among these,new cellular genes have been recently identi®ed (workin progress). Thus, our results suggest that the actualprevalence of HBV-related cis-activation of cellulargenes should be reassessed.
In conclusion, our results stimulate further analysisof SERCA gene mutations in cancer, and reinforce theconcept that the pharmacological modulation ofintracellular calcium may have an impact in cancertreatment (Kohn and Liotta, 1995).
Materials and methods
Tissues
Tumorous (T86) and non tumorous (NT86) frozen livertissues were obtained at surgery from a 65-year-old Japanese,HBsAg negative, HBV-DNA positive, anti-HCV positivepatient with primary liver cancer. The tumorous tissue wasclassi®ed as HCC Edmonson's grade 2. Chronic activehepatitis without cirrhosis was found in the non tumoroustissue. Other tumors used for this study were diagnosed asprimary liver cancer after pathological examination.
PCR and RT±PCR protocols and primer sequences
DNA was extracted from tumorous and non tumoroustissues as previously described (Paterlini et al., 1990). In orderto amplify HBV/cellular DNA junctions in the tumoroustissue, Alu-PCR was performed as previously reported(Minami et al., 1995).
RNA isolation and Northern blot analysis
Total cellular RNA was extracted using Trizol reagent (LifeTechnologies). mRNA isolation was done using Dynabeadsoligo (dT) (Dynal). Northern blot was performed asdescribed (Church and Gilbert, 1984). Full-length SERCA1cDNA and full-length HBV cDNA were used as speci®cprobes. Blots were normalized with a full-length GAPDHcDNA probe.
cDNA cloning
Double strand cDNA was synthesized using the MarathonTM cDNA Ampli®cation kit (Clontech) starting from 1 mg ofpoly(A)+ RNA from T86. The cDNA 5' and 3' ends wereampli®ed and sequenced with adaptor primer AP1 (Mara-thon kit) and HBV-junction primers Hsh-s (5'-accagcaccatg-caagctggtgatagagca-3' and Hsh-r (5'-accagcaccatgcaagctgg-tgatagagca-3'). Full-length hybrid cDNAs were ampli®edwith HBV X-s (5'-tttccatggctgctaggctgtactgccaa-3') andSERCA1 ex23-r (5'-cacagctctgcctgaagatgtgtcact-3') primersand cloned into pMos vector (Gibco ±BRL).
Isolation of microsomal fraction and immunoblotting
Liver microsomes were isolated as described (Heilmann et al.,1985). Proteins were treated with concentrated urea (20 mg/50 mg protein), SDS 1.8% and b-mercaptoethanol 9%. AfterSDS ±PAGE (14%), proteins were electrotransferred ontoPVDF membranes (Immobilon). For immunodetection, weused rabbit polyclonal antibody raised against HBV Xprotein (gift from P Arbuthnot), guinea-pig polyclonalantibody 79 B raised against rabbit fast twitch muscleSERCA1 (gift from AM Lompre ), mouse monoclonalantibody raised against human SERCA2 (anti-SERCA2,clone IID8, Tebu) and mouse monoclonal antibody raisedagainst human b-tubulin (Boehringer Mannheim). Followingreaction with a horseradish peroxydase-coupled secondaryantibody (goat anti rabbit or anti-guinea-pig or anti-mouse),immunodetection was visualized by the super signal kit(Pierce). Dimers or aggregates were sought with 7% SDS±PAGE in non-heated samples under mildly denaturingconditions (without urea) (Soulie et al., 1996).
Plasmid constructs and transient transfection
The constructs shown in Figure 3 were subcloned intopcDNA3.1, pcDNA3.1/Myc-His (Invitrogen) and pEGFP-N1(Clontech) and were used to transfect hepatoma derived celllines (CCL13, HuH7 and HepG2) by the calcium phosphatecoprecipitation method (10 mg/dish). The Xtpro constructwas obtained by subcloning Xt downstream the ERmembrane targeting cassette (prolactin peptide signal, blactamase sequence and IgM stop transfer signal, gift of VLingappa). In addition to that, we also cloned the prolactincassette upstream to the GFP sequence (ProGFP) and theSERCA1+4711 constructs.
Immunofluorescence
Cells were ®xed in 4% paraformaldehyde and permeabilizedfor 30 s in acetone/methanol (1 : 1). Non-speci®c binding siteswere blocked with 3% bovine serum albumin. Immunostain-ing was performed with anti-ER polyclonal antibody (giftfrom D Louvard), anti-myc monoclonal antibody (9E10,
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2884
Oncogene

Santa Cruz), anti-mitochondria polyclonal antibody (giftfrom C Marsac), anti-Golgi monoclonal antibody (gift fromM Bornens), and anti-lamin B antibody. Immunodetectionwas carried out using a ¯uorescein isothiocyanate (FITC)(green), texas red or Cy5 (red)-conjugated second antibody.Coverslips were analysed using confocal laser scanningmicroscopy (Zeiss, LSM 510).
Cell calcium contents
CCL13 cells were loaded with the ¯uorescent Ca2+ indicatorfura2-AM (1.2 mM) for 40 min at 378C in loading bu�er thatdoes not contain Ca2+. Successive applications of EGTA(0.1 mM), 2,5-Di-tert-butylhydroquinone (TBUBHQ, 50 mM),speci®cally inhibiting SERCA and thus releasing the ERcalcium, and of ionomycin (IONO, 2 mM), which is a non-speci®c calcium ionophore, enabled us to determine theincrease in [Ca2+]i due to the ER and non-ER Ca2+ pools,respectively (Figure 5a). The cytosolic-free Ca2+ concentra-tion ([Ca2+]i) was determined by the ratio of 340/380 nm¯uorescence intensities using a Kd for Ca2+-dye of 225 nM,and the maximum and minimum ¯uorescence responses weredetermined as described (Grynkiewicz et al., 1985).
Apoptosis
Transfected cells were analysed 48 h after transfection withGFP-fusion constructs. The percentage of apoptotic nucleiwas evaluated in GFP-positive cells using the DNA-dye 7-amino actinomycin D (7-AAD). Alterations to mitochondrialstructure were assessed following Nonyl Acridine Orange
(NAO) incorporation (Petit et al., 1995). Adherent and¯oating cells were incubated for 30 min at 378C in PBScontaining 4 mM NAO and harvested cells were then analysedin the FACScan ¯ow cytometer. Data from 20 000 eventswere recorded on a logarithmic scale and analysed using cellQuest software.
Transactivation assay
CCL13 cells were cotransfected with 2 mg of each of reportervectors: p-NF-kB-LUC, (containing the luciferase gene underthe control of the NF-kB promoter) and p-b-actin-b-galactosidase (b-galactosidase gene under control of b-actinpromoter) and 5 mg of the di�erent activators (wild type X,Xtpro, Xt/St+4 and the empty expression plasmidpcDNA3). Luciferase and b galactosidase analyses wereperformed according to standard protocols (Sirma et al.,1999). Luciferase activity was normalized against b galacto-sidase activity. Data presented are the mean values oftransfections done in triplicate experiments.
AcknowledgmentsWe thank P Champeil for critical reading of the paper andmost helpful discussion. This work was supported bygrants from INSERM, LNC no 8981 and ARC no 9327(France). M Chami, D Gozuacik and K Saigo are recipientof fellowships from LNC (France), TUBITAK (Turkey)and FRM (France), respectively.
References
Bayle D, Weeks D, Hallen S, Melchers K, Bamberg K andSachs G. (1997). J. Recept. Signal. Transduct. Res., 17,29 ± 56.
Benn J and Schneider R. (1994). Proc. Natl. Acad. Sci. USA,91, 10350 ± 10354.
Benn J and Schneider R. (1995). Proc. Natl. Acad. Sci. USA,92, 11215 ± 11219.
Berasain C, Patil D, Perera E, Huang S, Mouly H andBre chot C. (1998). Oncogene, 16, 1277 ± 1288.
Berridge MJ, Bootman MD and Lipp P. (1998). Nature, 395,645 ± 648.
Brostrom CO and Brostrom MA. (1990). Annu. Rev.Physiol., 52, 577 ± 590.
Buendia MA. (1992). Adv. Cancer Res., 59, 167 ± 226.Camacho P and Lechleiter JD. (1993). Science, 260, 226 ±
229.Chang MH, Chen CJ, Lai MS, Hsu HM, Wu TC, Kong MS
et al. (1997). New Engl. J. Med., 336, 1855 ± 1859.Cheong JH, Yi M, Lin Y and Murakami S. (1995). EMBO J.,
14, 143 ± 150.Chirillo P, Falco M, Puri PL, Artini M, Balsano C, Levrero
M and Natoli G. (1996). J. Virol, 70, 641 ± 644.Chirillo P, Pagano S, Natoli G, Puri P, Burgio V, Balsano C
and Levrero M. (1997). Proc. Natl. Acad. Sci. USA, 94,8162 ± 8167.
Chisari FV, Klopchin K, Moriyama T, Pasquinelli C,Dunsford HA, Sell S, Pinkert CA, Brinster RL andPalmiter RD. (1989). Cell, 59, 1145 ± 1156.
Chung DC, Brown SB, Graeme-Cook F, Tillotson LG,Warshaw AL, Jensen RT and Arnold A. (1998). CancerRes., 58, 3706 ± 3711.
Church GM and Gilbert W. (1984). Proc. Natl. Acad. Sci.USA, 81, 1991 ± 1995.
Dejean A, Bougueleret L, Grzeschik KH and Tiollais P.(1986). Nature, 322, 70 ± 72.
Dolmetsch RE, Lewis RS, Goodnow CC and Healy JI.(1997). Nature, 386, 855 ± 858.
Dorjsuren D, Lin Y, Wei W, Yamashita T, Nomura T,Hayashi N and Murakami S. (1998). Mol. Cell. Biol., 18,7546 ± 7555.
Falson P, Menguy T, Corre F, Bouneau L, de Gracia AG,Soulie S, Centeno F, Moller JV, Champeil P and le Maire,M. (1997). J. Biol. Chem., 272, 17258 ± 17262.
Fourel G, Couturier J, Wei Y, Apiou F, Tiollais P andBuendia MA. (1994). EMBO J., 13, 2526 ± 2534.
Garcia M, De The H, Tiollais P, Samarut J and Dejean A.(1993). Cell Biol., 90, 89 ± 93.
Graef E, Caselmann W, Wells J and Koshy R. (1994).Oncogene, 9, 81 ± 87.
Grynkiewicz G, Poenie M and Tsien RY. (1985). J. Biol.Chem., 260, 3440 ± 3450.
Guerini D, Garcia-Martin E, Zecca A, Guidi F and CarafoliE. (1998). Acta. Physiol. Scand. Suppl., 643, 265 ± 273.
Heilmann C, Spamer C and Gerok W. (1985). J. Biol. Chem.,260, 788 ± 794.
Hildt E, Hofschneider P and Urban S. (1996). Virology, 7,333 ± 347.
Kaufman RJ. (1999). Genes Dev., 13, 1211 ± 1233.Kekule AS. (1994). Primary liver cancer: Etiological and
progression factors, Vol. 13. Bre chot C. (ed). CRC Press:London.
Kim C, Koike K, Saito I, Miyamura T and Jay G. (1991).Nature, 351, 317 ± 320.
Kohn E and Liotta L. (1995). Cancer Res., 55, 1856 ± 1862.Koike K. (1998). Hepatitis B virus: molecular mechanisms in
disease and novel strategies for therapy, Vol. 3. Koshy Rand Caselmann WH (eds). Imperial College Press:London, pp. 133 ± 160.
Koike K, Moriya H, Yotsuyanagi H, Iino S and KurokawaK. (1994). J. Clin. Invest., 94, 44 ± 49.
Lee TH, Elledge SJ and Butel JS. (1995). J. Virol., 69, 1107 ±1114.
Lee YH and Yun Y. (1998). J. Biol. Chem., 273, 25510 ±25515.
Oncogene
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2885

Lin MH and Lo SC. (1989). Biochem. Biophys. Res.Commun., 164, 14 ± 21.
Ma TS, Mann DL, Lee JH and Gallinghouse GJ. (1999). CellCalcium, 26, 25 ± 36.
MacLennan DH, Rice WJ and Green NM. (1997). J. Biol.Chem., 272, 28815 ± 28818.
MacLennan DH, Rice WJ, Odermatt A and Green NM.(1998). Acta. Physiol. Scand. Suppl., 643, 55 ± 67.
Martonosi AN. (1996). Biochim. Biophys. Acta., 1275, 111 ±117.
Minami M, Poussin K, Bre chot C and Paterlini P. (1995).Genomics, 29, 403 ± 408.
Morgan AJ and Jacob R. (1998). J. Physiol. (Lond)., 513,83 ± 101.
Nicotera P and Orrenius S. (1998). Cell Calcium, 23, 173 ±180.
Ottesen AM, Kirchho� M, De-Meyts ER, Maahr J, GerdesT, Rose H, Lundsteen C, Petersen PM, Philip J andSkakkebaek NE. (1997). Genes Chromosomes Cancer, 20,412 ± 418.
Pahl HL and Baeuerle PA. (1997). Trends Biochem. Sci., 22,63 ± 67.
Paterlini P and Bre chot C. (1994). Primary liver cancer:Etiological and progression factors, Vol. 12. Bre chot C.(ed). CRC Press: London, pp. 167 ± 190.
Paterlini P, Gerken G, Khemeny D, Franco D, D'Errico A,Grigioni W, Wands J, Kew M, Pisi E, Tiollais P andBre chot C. (1990). Viral hepatitis and liver disease.Hollinger FB, Lemon SM and Margolis HS (eds).Williams and Wilkins: Baltimore, pp. 556 ± 559.
Peacocke M and Christiano AM. (1999). Nat. Genet., 21,252 ± 253.
Petit PX, Lecoeur H, Zorn E, Dauguet C, Mignotte B andGougeon ML. (1995). J. Cell. Biol., 130, 157 ± 167.
Pineau P, Marchio A, Terris B, Mattei MG, Tu ZX, TiollaisP and Dejean A. (1996). J. Virol., 70, 7280 ± 7284.
Pirc-Danoewinata H, Bull JP, Okamoto I, Karner J,Breiteneder S, Liebhardt A, Budinsky A and Marosi C.(1996). Wien Klin. Wochenschr., 108, 752 ± 758.
Pozzan T, Rizzuto R, Volpe P and Meldolesi J. (1994).Physiol. Rev., 74, 595 ± 635.
Raney AK and McLachlan A. (1991). Molecular biology ofHepatitis B Virus. McLachlan A. (ed). CRC Press. pp. 1 ±26.
Rice WJ and MacLennan DH. (1996). J. Biol. Chem., 271,31412 ± 31419.
Sakuntabhai A, Ruiz-Perez V, Carter S, Jacobsen N, BurgeS, Monk S, Smith M, Munro CS, O'Donovan M,Craddock N, Kucherlapati R, Rees JL, Owen M, LathropGM, Monaco AP, Strachan T and Hovnanian A. (1999).Nat. Genet., 21, 271 ± 277.
Sirma H, Giannini C, Poussin K, Paterlini P, Kremsdorf Dand Bre chot C. (1999). Oncogene, 18, 4848 ± 4859.
Sirma H, Weil R, Rosmorduc O, Urban S, Israel A,Kremsdorf D and Bre chot C. (1998). Oncogene, 16,2051 ± 2063.
Sitterlin D, Lee TH, Prigent S, Tiollais P, Butel JS andTransy C. (1997). J. Virol., 71, 6194 ± 6199.
Slagle B, Lee T, Medina D, Finegold M and Butel J. (1996).Mol. Carcinog., 15, 261 ± 269.
Soulie S, Moller JV, Falson P and le Maire M. (1996). Anal.Biochem., 236, 363 ± 364.
Sun BS, Zhu X, Clayton MM, Pan J and Feitelson MA.(1998). Hepatology, 27, 228 ± 239.
Terradillos O, Billet O, Renard C, Levy R, Molina T, BriandP and Buendia M. (1997). Oncogene, 14, 395 ± 404.
Terradillos O, Pollicino T, Lecoeur H, Tripodi M, GougeonML, Tiollais P and Buendia MA. (1998). Oncogene, 17,2115 ± 2123.
Truant R, Antunovic J, Greenblatt J, Prives C and CromlishJ. (1995). J. Virol., 69, 1851 ± 1859.
Wang J, Chenivesse X, Henglein B and Bre chot C. (1990).Nature, 343, 555 ± 557.
Wang X, Forrester K, Yeh H, Feitelson M, Gu J and HarrisC. (1994a). Proc. Natl. Acad. Sci. USA, 91, 2230 ± 2234.
Wang X, Gibson M, Vermeulen W, Yeh H, Forester K,Stuà rzbecher H, Hoeijmekers J and Harris C. (1995).Cancer Res., 55, 6012 ± 6016.
Wang XW, Forrester K, Yeh H, Feitelson MA, Gu JR andHarris CC. (1994b). Proc. Natl. Acad. Sci. USA, 91,2230 ± 2234.
Weil R, Sirma H, Giannini C, Kremsdorf D, Bessia C,Dargemont C, Bre chot C and Israel A. (1999). Mol. Cell.Biol., 19, 6345 ± 6354.
Williams JS and Andrisani OM. (1995). Proc. Natl. Acad.Sci. USA, 92, 3819 ± 3823.
Wu KD, Lee WS, Wey J, Bungard D and Lytton J. (1995).Am. J. Physiol., 269, C775 ±C784.
Zhang Y, Fujii J, Phillips M, Chen H-S, Karpati G, Yee W-C, Schrank B, Cornblath D, Bobylan K and MacLennanD. (1995). Genomics, 30, 415 ± 424.
HBV-related insertional SERCA1 mutagenesis in a HCCM Chami et al
2886
Oncogene




















