
Henshilwood, C. & d’Errico, F. (editors). 2011. Homo symbolicus: The dawn of language,...
250
Homo Symbolicus
Transcript of Henshilwood, C. & d’Errico, F. (editors). 2011. Homo symbolicus: The dawn of language,...

Homo Symbolicus


Homo SymbolicusThe dawn of language, imagination and spirituality
Edited by
Christopher S. HenshilwoodUniversity of Bergen & University of the Witwatersrand
Francesco d’ErricoUniversity of Bordeaux I & University of Bergen
John Benjamins Publishing CompanyAmsterdam / Philadelphia

Library of Congress Cataloging-in-Publication Data
Homo symbolicus : the dawn of language, imagination and spirituality / edited by Christopher S. Henshilwood, Francesco d’Errico.
p. cm. Includes bibliographical references and index. 1. Symbolism (Psychology) 2. Human behavior. 3. Language and languages--Origin.
4. Psycholinguistics. 5. Biolinguistics. I. Henshilwood, Christopher Stuart. II. D’Errico, Francesco.
BF458.H65 2011 155.7--dc23 2011031012isbn 978 90 272 1189 7 (Hb ; alk. paper)isbn 978 90 272 8409 9 (Eb)
© 2011 – John Benjamins B.V.No part of this book may be reproduced in any form, by print, photoprint, microfilm, or any other means, without written permission from the publisher.
John Benjamins Publishing Co. · P.O. Box 36224 · 1020 me Amsterdam · The NetherlandsJohn Benjamins North America · P.O. Box 27519 · Philadelphia pa 19118-0519 · usa
The paper used in this publication meets the minimum requirements of American National Standard for Information Sciences – Permanence of Paper for Printed Library Materials, ansi z39.48-1984.
8 TM

Table of contents
Editors’ introduction vii
Acknowledgements xi
chapter 1Pan Symbolicus: A cultural primatologist’s viewpoint 1
William C. McGrew
chapter 2The evolution and the rise of human language: Carry the baby 13
E. Sue Savage-Rumbaugh & William M. Fields
chapter 3The origin of symbolically mediated behaviour: From antagonistic scenarios to a unified research strategy 49
Francesco d’Errico & Christopher S. Henshilwood
chapter 4Middle Stone Age engravings and their significance to the debate on the emergence of symbolic material culture 75
Christopher S. Henshilwood & Francesco d’Errico
chapter 5Complex cognition required for compound adhesive manufacture in the Middle Stone Age implies symbolic capacity 97
Lyn Wadley
chapter 6The emergence of language, art and symbolic thinking: A Neandertal test of competing hypotheses 111
João Zilhão
chapter 7The human major transition in relation to symbolic behaviour, including language, imagination, and spirituality 133
David Sloan Wilson
chapter 8The living as symbols, the dead as symbols: Problematising the scale and pace of hominin symbolic evolution 141
Paul Pettitt

vi Table of contents
chapter 9Biology and mechanisms related to the dawn of language 163
George F.R. Ellis
chapter 10The other middle-range theories: Mapping behaviour and the evolution of the mind 185
Benoît Dubreuil
chapter 11Metarepresentation, Homo religiosus, and Homo symbolicus 205
Justin L. Barrett
Index 225

Editors’ introduction
Few of us take the time to reflect on the role that symbols play in our everyday lives, both on a conscious and sub-conscious level, and how these symbols have become an intricate part of our humanity. Symbols can be inscribed on our bodies from birth through to later life, for example, in the form of circumcision, scarifica-tion, mutilation and tattoos. We also cover our bodies with symbols each day with the application of make-up, donning of clothing and accessories and the way we dress our hair. The artificial environments in which we live at home and at work are permeated with symbols and we attribute symbolic meanings to the natural world as well, including far away planets, stars and galaxies. A myriad of symbols are created and stored in our minds; we establish dialogues among them within our heads and are able to transmit these representations to others despite the fact they exist only in our imagination.
Symbols can act as powerful weapons in war and peace and we think here of insignia, flags and mottos. We recycle symbols used by others and give them a dif-ferent, possibly opposite meaning. Swastikas were important symbols in various ancient civilizations and remain widely in use in Indian religions. The adoption of the swastika by the Nazi party in Germany after 1920 led to an association of this symbol by some with fascism and white supremacists, the antithesis of its earlier meaning. Language, writing, religion, science, institutions and laws could not possibly exist without our capacity to master the creation, transmission, and disposal of symbols. We also take our symbols to the grave and after death we may ourselves become regarded by others as a symbol.
Some researchers believe that substantial progress has been made in under-standing the functioning of various categories of symbols and how they condi-tion our thoughts, speech and actions today, but our knowledge of how and when these aptitudes first emerged within our lineage is imprecise and speculative. If we adopt a Darwinian perspective, we could argue that the first innovative introduc-tion of symbols to mediate behaviour must stem from exaptation and long term selective pressures. For decades, Palaeolithic archaeologists have suggested that symbols first appeared in association with the material culture of Homo sapiens in Germany or France about 40 000 years ago. Recent archaeological discoveries in Africa support a much earlier emergence for the production and use of symbols. These finds have set the stage to re-open and re-examine theoretical perspectives and opened the door for constructive interdisciplinary collaborations. The more

viii Homo symbolicus
research data we receive, the more apparent it becomes that the use and applica-tion of symbols, at least to some degree, is a constitutive component of animal cog-nition. It is therefore counter-intuitive to suggest that exploring only how we and our ancestors used symbols can provide the key to understanding the very nature of this phenomenon. The net needs to be cast more widely. Early pre-adaptations to symbolic use must lie therefore within our own and other species and without these roots, the more recent imposing achievements associated with Homo sapiens could not have evolved.
Over the millennia, hominins have built on this potential to embark on a symbolically laden adventure, filled at times with ambushes and failures, but also successes. Only now are archaeologists starting to discover some of the trials of these early communities. It has been a to-and-fro trial with balances and coun-terbalances and the costs at times have exceeded the advantages, leading, in some cases, to the return of old ways and the maintenance of the same “solutions” for long periods of time. On occasion it has also led to the introduction of even more innovative solutions. How this process led in the end to the cultural complexity with which we are now familiar is one of the unanswered yet fascinating questions of our evolutionary history. In this volume, we offer an opportunity for interdis-ciplinary collaboration between scholars from a range of disciplines in an attempt at addressing this question and bridging the gap between raw archaeological evi-dence and human behaviour. We bring together new perspectives from the fields of archaeology, anthropology, philosophy, evolutionary biology, religion, palaeo-anthropology and linguistics with the aim of linking genetic, neural, cognitive, and behavioural development. We believe that this approach opens novel path-ways for understanding the ways in which the human mind evolved. In particular, we look at the emergence of symbolism, imagination and spirituality from diverse perspectives. The volume starts by examining the communication patterns and lives of apes in habitats that resemble those in which the last common ancestor of Homo and Pan resided, accepting that apes raised in a Pan/Homo environment acquire significant aspects of human language and human culture. Since language fluency was involved in archaic Homo sapiens becoming modern Homo sapiens, and as some members of Pan paniscus have also acquired symbolic language, they deserve the appellation Pan symbolicus. We subsequently explore the puzzling gap of more than 80,000 years that exists in the archaeological record between the first anatomical appearance of the species we call Homo sapiens sapiens and the appear-ance of behaviours which we associate with our own behaviour today. We evaluate archaeological evidence, including burial practices, the use of pigment and sym-bolic artefacts, the manufacture of complex adhesives and the innovations that set the stage for the later routine incorporation of material culture within a symbolic system that mediated human behaviour. In the volume, we discuss the adaptations

Editors’ introduction ix
that brought about the dispersal of modern humans out of Africa, into Asia, Australasia and Europe. We ask too, what are the main factors that account for the emergence of key behavioural innovations in Africa and Eurasia between 200,000 and 30,000 years ago. Do we see significant differences between modern humans in Africa and contemporaneous archaic populations living outside of this conti-nent? Did climatic changes influence the behavioural differences between Nean-dertals and Homo sapiens? A central focus of the volume is the development of the ability to deliberately consider, ponder, or evaluate mental representa-tions and how this may have lead to the development of language, spirituality and symbolic behaviour. To address this, we ask to what extent activities such as language, decoration, adornment, trade and exchange, and imagery are really predicated on the incorporation of symbols within a behavioural repertoire. At first sight the chapters in this book may appear eclectic, but with further reading it becomes clear that there is a subtle thread that links these ideas together – we believe that this approach of presenting highly diversified perspectives has pro-vided an insightful and innovative account into the whys and wherefores of later hominin evolution.


Acknowledgements
Financial support was provided to the editors of this volume by the European Research Council Advanced Grant TRACSYMBOLS (FP7 No. 249587), and the PROTEA France - South Africa exchange programme. Christopher S. Henshilwood was also supported by a National Research Foundation/Department of Science and Tech-nology funded Chair at the University of the Witwatersrand, South Africa and the Norwegian Research Council. The John Templeton Foundation and the Uni-versity of Bergen, Norway, have contributed to establishing the interdisciplinary research network that culminated in 2009 in a conference in Cape Town, South Africa titled ‘Homo symbolicus: The Dawn of Language, Imagination and Spiri-tuality’. The authors acknowledge the financial contributions of these two orga-nizations with special thanks to Mary Ann Meyers, Paul Wason, Anna Gro Vea Salvanes, Sigmund Grønmo, Svenn-Åge Dahl and Jan-Petter Myklebust. Apprecia-tion is extended to Laura van Niekerk and Petro Keene who worked on earlier and final versions of the texts. Anke de Looper and Patricia Leplae of John Benjamins NL kindly guided us through the publishing process.


chapter 1
Pan symbolicus
A cultural primatologist’s viewpoint
William C. McGrewDepartment of Archaeology and Anthropology, University of Cambridge
Traditional views of non-human primate communication held that it was hard-wired, involuntary, and emotionally expressive only. All of these conclusions have been proven wrong: chimpanzees use natural vocalisations to signal the nature and location of foodstuffs, that is, spontaneous referentiality. Gibbons signal anti-predator alarm by using song-elements in rule-governed combinations. Chimpanzees show tactical deception in their recruitment screams, tailoring these to the audience. Pant-hoots show local variation that if found in human language, would be called dialects. Males devise individualised courtship routines. Some of these signals become group-typical, and such gestures are neither iconic nor onomatopoeic, but instead are arbitrary. Primatological studies of apes behaving spontaneously show us repeatedly that their communicative and cognitive capacities exceed our anthropocentric expectations.
1. Introduction
Short of inventing a time-machine, we will never know directly the acts of our extinct ancestors and their ilk. So, we must model what is missing, by one of three methods: either we must use the discovered products of behaviour, that is, arte-facts, to infer the missing behaviour of their use in antiquity, via archaeology; or, we must seek to put flesh on the bones of the fossil record, and via functional morphology, infer the whole, behaving organism from palaeontology; or, we must scrutinise the behaviour of other living forms, on the grounds that their actions will be more informative of the evolutionary process that led to us, than are studies of ourselves, as the end-product (McGrew 2010).
Seeking such a proxy is not straightforward, and there is lively debate about how to choose such a model (Sayers & Lovejoy 2008; Lovejoy 2009). Some evo-lutionary anthropologists favour referential models, that is, choosing a living taxon (usually but not always another species), as the nearest approximation to an extinct hominin. Others favour non-referential models, that is, constructing a hypothetical model based on evolutionary and ecological principles, which is

2 William C. McGrew
not bounded by the limited choice available amongst extant taxa. Some favour some kind of combination of the two (McGrew 2004). (Not seriously considered here are more fanciful models based on speculative surmise, e.g. the Aquatic Ape Theory, Morgan, 1997. Such ‘evolutionarios’ may be good fun to play with, but unless they are testable, they take us nowhere scientifically).
One basis for choosing a referential model is phylogenetic proximity, that is, concentrating on humankind’s nearest living relative, on the grounds that maximally shared genes should be manifest in maximally similar behaviour, all other things being equal. This sends us to the two living species of the genus Pan: chimpanzee (P. troglodytes) and bonobo (P. paniscus). Each is equally related to anatomically and behaviourally modern Homo sapiens, as the two ape taxa separated after the Pan line separated from our own. Thus, these African apes are the living forms with whom we last shared common ancestry, albeit it about 7 million years ago. Genotypically, it is clear from the sequencing of the human and chimpanzee genomes, that we share more than 99% of our genes. Phenotypi-cally, there is more similarity than meets the eye (that is, hairy quadrupeds versus mostly hairless bipeds): Pan and Homo blood is virtually interchangeable, and any one of us could donate a life-saving transfusion to an ape, provided we correctly matched the ABO blood groups.
Another basis for choosing a referential model is ecological similarity. That is, we can focus on organisms that survive and thrive in environments similar to those in which humankind emerged. We should expect such creatures to show similar adaptations in response to similar natural selection pressures, whether these be from predators, prey, climate, vegetation, etc. Finally, we might choose a refer-ential model on the basis of behavioural performance. The archaeological record of the evolutionary origins of human technology usually is based on persisting lithics, as opposed to organic raw materials that perished long ago (McPherron et al. 2010). But we cannot ignore the elementary technology of capuchin mon-keys, especially Cebus libidinosus (for a recent review, see Ottoni & Izar 2008) or long-tailed macaques (Gumert et al. 2009). These monkey species are but distantly related to us by comparison with the African apes, but they do impressive things with stones. Capuchin monkeys use stones in communication, agonism, and sub-sistence, e.g. they are the only non-humans known to use stones as trowels to dig up the underground storage organs of plants (Moura & Lee 2004). Long-tailed macaques use different types of stone hammers for cracking open animal versus plant prey. When the monkeys show such percussive technology, the modified lithics are there to be recovered and analysed.
Here, in tackling the evolutionary origins of language, imagination and spiri-tuality, I will focus on Pan, and whenever possible, on apes living in habitats that resemble those in which the Last Common Ancestor of Homo and Pan resided.

Chapter 1. Pan symbolicus 3
For as much as possible, I will refer to observational studies of apes in nature, rather than experimental studies of apes in captivity. I can only hope that what is gained in socio-ecological validity from studying the organism in situ counter-balances what is lost in the absence of control of key variables. Why choose a refer-ential model? Because it gives us real behaviour in the real world. Why not choose between chimpanzee and bonobo? Because the fossil record for post-Miocene African apes is so sparse that we cannot say whether the Last Common Ancestor more closely resembled one kind of Pan or the other.
Finally, I make explicit, a methodological and epistemological bias: I am an ethologist, trained to record the behaviour of organisms and to try to infer their intentions from their actions. The rationale for this approach is the same, whether the subject of study is ant or aunt. Unobservable thoughts and feelings are thus opaque, in any species, including our own; only their ‘reflections’ can be noted, e.g. in autonomic responses, facial expressions, etc. Any such inference is an exer-cise in probability, based on the consequences of behaviour or other recordable phenomena, e.g. blushing. A person can no more readily penetrate the ‘black box’ that is the mind of a chimpanzee than it can that of a fellow human being. In fact, we are bound to be more sceptical of behavioural data (including verbal self-report) from human than from non-human animals, given the former’s capacity for deception and especially self-deception. This may make the remainder of this essay disappointingly conservative, but I hope not.
In tackling the prospect of Pan symbolicus, I make four basic assertions, striving to develop each with regard to our current state of knowledge regarding Pan:
1. Communication does not equal language.2. Language does not equal speech.3. Symbol-use does not equal language.4. Non-human language does not equal human language.
In all cases, I shall call upon the published literature in behavioural primatology, as manifest in anthropology, biology and psychology, based on natural or natura-listic processes. For that reason, experimental studies of ‘pongo-linguistics’ are omitted (Savage-Rumbaugh et al. 1998). Similarly, emerging studies of the neuronal substrates for communication, as studied comparatively (e.g. Taglialatela et al. 2008) cannot be done justice here.
2. Communication does not equal language
If communication is defined minimally by the sequence of ‘sender-message-receiver’, and if language is defined simply as a socially-acquired system of

4 William C. McGrew
semantic and syntactic processing, then the two phenomena obviously are not the same. Neither one is a necessary nor sufficient condition for the other. The two need not even overlap, that is, one can imagine an organism that communicates by pheromones, and thinks in terms of sentences but never transmits those thoughts. Lots of organisms, even one-celled ones, communicate, but few (apparently) use language to do so.
However, while language is undoubtedly useful in thinking, and most human thought is hard to imagine without it, in everyday human life, language in what-ever modality is a subset of communication. That is, humans habitually communi-cate verbally (words), but also non-verbally (facial expressions), and para-verbally (inflection, accent).
Chimpanzees communicate in all sensory modalities: Visually, they show a wide range of facial expression, gesture, posture, and locomotion. For example, invitation to play may take the simultaneous form of a ‘play-face’, slapping the ground, quadrupedal bouncing, and a syncopated gallop (Goodall 1986). Acousti-cally, chimpanzees drum with hands and feet on the resonating buttress roots of trees, and each individual has a distinctive drumming ‘signature’ (Arcadi et al. 2004). Olfactorally, cycling females signal their changing reproductive states over the menstrual cycle via perineal odour that is monitored by males. Tactilely, as well as visually, the relative positions of the hands in the grooming hand-clasp (see below) indicate the relative social dominance ranks of the two participants, with the subordinate providing the support for the upraised arms while the dominant is supported (McGrew 2004).
Is any of chimpanzee communication linguistic? Traditional views of nonhu-man primate communication held that it was instinctive, involuntary, and emo-tionally expressive only. Each of these conclusions since has been proven wrong: Chimpanzees use natural ‘rough grunt’ vocalisations to signal the nature and loca-tion of foodstuffs, that is, their communication shows spontaneous referentiality (Slocombe & Zuberbühler 2005). (This confirms, in everyday life, what Savage-Rumbaugh et al. 1978 have shown experimentally with an artificial language system.) Gibbons also show referential signalling in their songs that signal anti-predator alarm, by emitting song elements in rule-governed combinations (Clarke et al. 2006). These are the roots of semantics and syntax.
Are such vocal emissions intentional? Yes, even monkeys show situation-ally contingent vocal interaction: Langur monkey males give anti-predator alarm calls only if there is an audience; moreover they continue calling until all group members have responded, thus showing the ability to monitor and to recall the vocal replies of their group mates (Wich & de Vries 2006). More impressively, chimpanzees show tactical deception in their recruitment screams, when they are aggressed again: depending on the composition of the audience, that is, their

Chapter 1. Pan symbolicus 5
social rank relative to that of the persecutor, they may exaggerate their screams. This shows a working knowledge of so-called ‘triadic awareness’, indicating understanding of third-party relationships (Slocombe & Zuberbühler 2007; Zuberbühler 2006). Thus, the modern view of non-human primate communica-tion is that it is learned, voluntary, and intentionally referential and rule-governed.
Language as communication necessarily implies sociality, or in humans, collectivity. Hundreds of human languages exist that clearly demarcate societal boundaries and as a consequence, signal group identity, that is, ‘us versus them’. (In contrast, solitary creatures avoid interaction and may reduce communication to isolated ‘calling cards’, e.g. scent-marks left in the environment). To what extent is there evidence of group-specific communication in apes? Marshall et al. (1999) found that chimpanzees brought together in captivity from a variety of sources showed a wide range of variants in their performance of the species’ long-call vocalisation, the pant-hoot. However, over time these individual variants changed to converge on a single version that became characteristic of the group.
In nature, different chimpanzee populations perform pant-hoots that show local variation; the calls have the same elements but show nuanced differences that if found in human language, we would call dialect (Mitani et al. 1992).
3. Language does not equal speech
Speech (spoken language) is neither a necessary nor a sufficient condition for language. Signed language need not involve sound, and echolalic speech may be nonsense. Extensive study of standard sign languages (as used by the deaf) seems to indicate that it is not ‘second-class’ speech, and because of its visual format, it may be even more efficient or expressive than speech. Speech by talking parrots, mynah birds, etc. was once dismissible as mindless mimicry, but recent studies of an African gray parrot may shade this black and white distinction as gray (Pepperberg 1999).
More complicatingly, many vocalisations that are uttered are not verbal, whether these are sighs or cries, and so need not be linguistic. Their non-linguistic status is abundantly clear when they are involuntary, e.g. gasp, sob, yet they may be highly communicative and arresting in their attention-getting power. Furthermore, mechanical sounds may function linguistically, even if their perceived input is non-verbal, e.g. Morse code. Finally, in principle, other sensory-motor modalities than sound can function linguistically, but actual working examples are usually secondary and derived, e.g. visual printing, tactile Braille.
Non-vocal communication in apes varies at the individual or group level (Pollick & de Waal 2007). Individual male chimpanzees devise courtship ‘routines’

6 William C. McGrew
that communicate to females their desire to mate; these sometimes incorporate novel elements, e.g. an everted lower lip, that are added to behavioural sequences by association with more unmistakeable sexual signals, e.g. an erect penis. At Mahale, such signals are group-typical: males noisily tear to pieces leaf after leaf, seeking to draw the attention of females to their sexual arousal. Such gestures are not iconic or onomatopoeic; leaves have nothing to do with copulation per se (Nishida et al. 2010).
The grooming hand-clasp (GHC) is a striking behavioural variant in chim-panzee social grooming: Two individuals sit facing one another and each fully extends the right or left arm overhead, where they clasp hands (McGrew & Tutin 1978). This symmetrical, mutual posture, in ‘A-frame’ configuration, occurs in some populations of chimpanzees, but not in others. Detailed study shows inter-group variation in GHC, even within the same population, e.g. in some groups, the clasp is palm-to-palm, in others palm-to-wrist. In effect, the style of performance seems to be an indicator of natal group identity, and immigrants modify their GHC, but may retain it when grooming with others who originated in the same natal group (McGrew 2004). If we saw such signals in two neighbouring human groups, we would interpret these as ethnic markers.
4. Symbol-use does not equal language
If symbols are defined as mental (conceptual) representations of phenomena, whether concrete or abstract, then they must be distinguished from signs, which are more basic perceptual signals. The two are often confused. The colour of a traffic light (green or red) arbitrarily signifies permission to proceed or obligation to halt, but no mental representation is necessary for the system to function. Instead, a learned stimulus-response association will suffice. This distinction may be crucial in distinguishing between human and non-human language (see below).
Even if symbols were never used in communication, they seem to be essential to human thought. Despite the Sapir-Whorf hypothesis, that language determines culture, non-human cultures seem to exist without language (McGrew 2004; cf. Washburn & Benedict 1979). Mathematics is the prototypical symbol system, and even if some numerical abilities are widespread across species (Gelman & Gallistel 2004), some aspects of it, e.g. algebra, constitute invented symbol systems. Comparative data do not always show human superiority: chimpanzees are better at remembering strings of numerals presented briefly than are adult humans (Matsuzawa 2009). However, like many other forms of code, e.g. computer machine code, symbols may function non-linguistically, even if devised by linguistic creatures.

Chapter 1. Pan symbolicus 7
On the other hand, symbol-use seems to be a necessary condition for language, as defined above, even if it is not a sufficient one. All human languages seem to be symbolic, although for how long in human evolution this has been the case is hard to say, given that only written language persists.
5. Non-human language does not equal human language
The absent adjective ‘human’ is almost always assumed when the noun ‘language’ is used. This is understandable, given our species’ anthropocentrism, but it need not be the case. Decades ago, the dance language of honey bees (von Frisch 1927) was recognised to have many linguistic features (e.g. Hockett 1960). Does this mean that if a non-human language satisfied 9 of 10 criteria on some checklist of features, then that phenomena could be considered 90% human language? Such musing becomes more than speculation when contemplating the communicatory abilities of dolphins and whales, whether via echo-location or full-blown singing. Some of these issues are merely ones of sensory capacity, e.g. the vocal communication of bats (ultra-high-frequency) or of elephants (ultra-low-frequency). Even humbler creatures, that is, ones with relatively small brains, show unexpected complexity in their language-like signals, e.g. meerkats simultaneously encode referential and affective information in their alarm calls (Manser et al. 2002).
And what are we to do with human beings that are mewling infants or drooling elders? If they produce utterances, however unintelligible, must they be classed as language-users? Just because if they mature normally, the former will become linguistic, and the latter once were so? Or, being pre-verbal or post-verbal, either by immaturity or senility, are they in some way equivalent to the less-than-complete language produced by other species? (The preceding questions apply only to normal ontogeny, but become even more vexed when abnormality is involved; at what point on the scale does diminished linguistic ability become no different from absent ability?).
6. Pan symbolicus
To what extent can living Pan enlighten us about language, imagination, and spiri-tuality? Several studies have already been mentioned above, especially with regard to complex communication that may share some features with full-blown human language, and more continue to be added. For example, female chimpanzees manipulate their emission of copulation calls, increasing their calling frequency in the presence of high-ranking males (i.e. potential mates), while decreasing their

8 William C. McGrew
frequency in the presence of high-ranking females (i.e. competitors) (Townsend et al. 2008). Restrictive generalisations that limited non-human communication to involuntary expressions of emotion are no longer tenable.
Similarly, prior categorical dicta that had apes incapable of vocal imitation look increasingly shaky. In addition to earlier studies of bonobo vocalisations (see Savage-Rumbaugh et al. this volume), a recent report has an orang-utan showing spontaneously whistling in mimicry of its caretaker (Wich et al. 2008).
But what of symbolic communication in nature? Two reports stand out, but neither is conclusive. Boesch (1991) reported for the chimpanzees of the Tai For-est, Ivory Coast, symbolic drumming. That is, behavioural responses to drumming indicated that information about the drummer in time and space was being trans-mitted, e.g. ‘we are resting here for a certain period, then will move off in a particular direction’. However, once the main drummer died, the behaviour disappeared, and so the phenomenon could not be studied further. Savage-Rumbaugh et al. (1996) reported that bonobos at Wamba, Democratic Republic of Congo, left messages for others trailing behind, in the form of bent-over vegetation which communicated their direction of travel. However, these data were gathered over only two days, and so constitute only a pilot study that needs to be followed up.
What can be said about the imagination of apes in nature? The natural his-tory and ethnography of wild chimpanzee populations, especially reports from Gombe, Tanzania, are rich in content and potential. Goodall (1986) has variously described the ‘rain dance’, a wild and woolly social display, done especially by adult males, that occurs at the outset of thunder storms. This has been interpreted as some sort of statement of collective defiance at the sound and fury of nature, and has been likened to human rituals in response to sensorally overwhelming natural forces. However, rain dancing never has been analysed in detail, despite its exis-tence being known for more than 40 years.
Similarly, chimpanzees may show prolonged and acrobatic ‘waterfall displays’ at the 10-m-high waterfall at the head of Kakombe Valley (Goodall 1986). This is often done by solitary individuals, so that it can have no communicatory signifi-cance, and thus it has been likened to an individual expression of awe or perhaps superstition. What makes this more impressive than the rain dance is that it is done predictably, that is, the site and nature of the waterfall is constant and well-known. So, why should an ape be drawn to engage in such an energetic (and even dangerous, as it involves swinging on vines high above a rocky streambed) activity, as no reaction is possible from absent peers, predators, or prey? This mystery has suggested deeper motives, perhaps to do with the evolutionary roots of animism, to some commentators.
In fact, we have no more idea what is going on in the minds of ape rain dancers, or waterfall displayers, than we can know what motivated the individuals who

Chapter 1. Pan symbolicus 9
created startling depictions of animals on Upper Palaeolithic cave walls or crafted three-dimensional images of sumptuous women (Conard 2009). Even more tenta-tive are inferences drawn about symbol-use based on simpler artefacts, at least as it appears to this simple-minded primatologist. For example, from finding a con-centration of snail shells, each having a single, similar hole, it is inferred that these make a necklace, and that a necklace necessarily means deliberate ornamentation, which amounts to evidence of symbolic capacity. If we grant the existence of the still-hypothetical string that would be necessary to infer even the most minimal necklace, there are other reasons to string items, as any fisherman knows. (There is even an English vocational surname, Stringer, to remind us of this). Even if we grant the putative necklace status as a manufactured ornament, this still does not make it symbolic, as any ambitious bowerbird knows (Madden 2008). Perhaps there is still a place for Occam’s razor in interpreting the significance of artefacts, no matter how attractive the more complex explanation seems to be.
With the living apes, we can note certain individual variables, e.g. age, sex, rank, etc. of the performers, as well as some contextual ones, e.g. whether alone or in company, and if the latter, with whom; what activities both precede and follow the event; what is the state of arousal of the performer, either before, during or after the performance, etc. (Having said this, the published scientific literature is disappointing. Primatologists mostly have been content to report even provoca-tive events as anecdotes, but usually have not given them systematic study).
None of this tells us anything directly about the mental representations that may or may not underlie the performance. The behaviour may appear compelling, even ritualised, in terms of its form, tempo, and complexity, but this is true of many displays performed by a variety of animals, even invertebrates, and we do not seek to invoke imagination to explain their causation. In the uncontrolled circumstances of the field, I can think of no way to penetrate the ape’s ‘black box’, in order to ascertain the extent to which, if at all, imagination underlies such intriguing behaviour.
If imagination is a problem, what then to do with spirituality? If the latter phenomenon is notionally defined (crudely) as belief in supernatural forces, such a definition stumbles already on its non-operationality: belief is problematic enough to infer, but the supernatural by definition is inaccessible to science, as science is bounded by natural law. If, on the other hand, natural but yet unexplained forces of certain types are enough to infer spirituality, then we have to find a way to measure them. I confess to not being able to comprehend how spirituality can be validated even in Homo sapiens, given that verbal professions of faith are vulnerable to deception, either intentional or otherwise. The unintentional form of self-deception is especially problematic, even if one invokes the aid of psycho-physiology, e.g. polygraph. Much as I would like to present apes as models for the origin and

10 William C. McGrew
evolution of religion, I do not see an empirical way to do so. This is frustrating. (Of course, if apes gathered in a forest clearing and knelt collectively to face the setting sun, while vocalising rhythmically in unison, as it sank beneath the horizon, we might be tempted to infer something potentially interesting in this regard. Then we would have candidate behaviour to record and to analyse. Such a tableau is not as far-fetched as it seems, as anyone who has seen groups of ring-tailed lemurs pose with widespread arms to greet the morning sun can testify).
In summary, primatological studies of apes behaving spontaneously have shown us that their communicatory and cognitive capacities exceed what even recently was thought to be present. For example, a male chimpanzee surreptitiously made and stored stone missiles to be used later as projectiles (Osvath 2009); such acts done by humans would be labelled unhesitatingly as future planning. Fewer and fewer boundaries between human and non-human hold up to empirical scru-tiny, and the gap between linguistic and non-linguistic communication seems ever narrower. These questions, at least, are amenable to scientific investigation, in both field and laboratory. Questions of cognitive functioning, including imagination, would seem to be accessible to well-designed experiments (see Matsuzawa 2009), but not in nature, where too few variables can be controlled. Questions regarding spirituality take us into areas that primatologists are ill-equipped to tackle, at least with their current tools.
References
Arcadi, A.C., Robert, D. & Mugurusi, F. 2004. “A comparison of drumming by male chimpan-zees from two populations.” Primates 45: 135–139.
Boesch, C. 1991. “Symbolic communication in wild chimpanzees.” Human Evolution 6: 81–90.Clarke, E., Reichard, U. & Zuberbühler, K. 2006. “The syntax and meaning of wild gibbon
songs.” PLoS ONE 1 (1): e73.Conard, N.J. 2009. “A female figurine from the basal Aurignacian of Hohle Fels Cave in south-
western Germany.” Nature 459: 248–252.Gelman, R. & Gallistel, C.R. 2004. “Language and the origin of numerical concepts.” Science
306: 441–443.Goodall, J. 1986. The Chimpanzees of Gombe. Cambridge MA: Harvard University Press.Gumert, M.D., Kluck, M. & Malaivijitnond, S. 2009. “The physical characteristics and usage
patterns of stone axe and pounding hammers used by long-tailed macaques in the Andaman Sea region of Thailand.” American Journal of Primatology 71: 594–608.
Hockett, C.F. 1960. “The origin of speech.” Scientific American. 203: 88–96.Lovejoy, C.O. 2009. “Reexamining human origins in light of Ardipithecus ramidus.” Science 326
(5949): 74e1–74e8.Madden, J.R. 2008. “Do bowerbirds exhibit cultures?” Animal Cognition 11: 1–12.Manser, M.B., Seyfarth, R.M., & Cheney, D.L. 2002. “Suricate alarm calls signal predator class
and urgency.” Trends in Cognitive Sciences 6 (2): 55–57.

Chapter 1. Pan symbolicus 11
Marshall, A.J., Wrangham, R.W. & Arcadi, A. 1999. “Does learning affect the structure of vocal-izations in chimpanzees?” Animal Behaviour 58: 825–830.
Matsuzawa, T. 2009. “Symbolic representation of number in chimpanzees.” Current Opinion in Neurobiology 19: 1–7.
McGrew, W.C. 2004. The Cultured Chimpanzee. Reflections on Cultural Primatology. Cambridge: Cambridge University Press.
McGrew, W.C. 2010. “In search of the last common ancestor: new findings on wild chimpanzees.” Philosophical Transactions of the Royal Society B 365: 3265–3276.
McGrew, W.C. & Tutin, C.E.G. 1978. “Evidence for a social custom in wild chimpanzees?” Man 13: 234–251.
McPherron, S.P., Alemseged, Z., Marean, C.W., Wynn, J.G., Reed, D., Geraads, D., Bobe, R. & Bearat, H.A. 2010. “Evidence for stone-assisted consumption of animal tissues before 3.39 million years ago at Dakika, Ethiopia.” Nature 466: 857–860.
Mitani, J., Hasegawa, T., Gros-Louis, J., Marler, P., & Byrne, R.W. 1992. “Dialects in wild chimpanzees?” American Journal of Primatology 27: 233–243.
Moura, A.C.A. & Lee, P.C. 2004. “Capuchin stone tool use in a Caatinga dry forest.” Science 306: 1909.
Nishida, T., Zamma, K., Matsusaka, T., Inaba, A. & McGrew, W.C. 2010. Chimpanzee Behavior in the Wild. An Audio-Visual Encyclopedia. Tokyo: Springer.
Osvath, M. 2009. “Spontaneous planning for future stone throwing by a male chimpanzee.” Current Biology 19 (5): R190-191.
Ottoni, E.B. & Izar, P. 2008. “Capuchin monkey tool use: Overview and implications.” Evolutionary Anthropology 17: 171–178.
Pepperberg, I.M. 1999. The Alex Studies. Cognitive and Communicative Abilities of Grey Parrots. Cambridge, MA: Harvard University Press.
Pollick, A.S. & de Waal, F.B.M. 2007. Ape gestures and language evolution. Proceedings of the National Academy of Sciences 104: 8184–8189.
Savage-Rumbaugh, E.S., Rumbaugh, D.M. and Boysen, S. 1978. “Symbolic communication between two chimpanzees.” Science 201: 641–644.
Savage-Rumbaugh, S., Williams, S.L., Furuichi, T. and Kano, T. 1996. “Language perceived: Paniscus branches out.” In Great Ape Societies, W.C. McGrew, L. F. Marchant & T. Nishida (eds), 173–184. Cambridge: Cambridge University Press.
Savage-Rumbaugh, S., Shanker, S.G. and Taylor, T.J. 1998. Apes, Language, and the Human Mind. New York: Oxford University Press.
Sayers, K. & Lovejoy, C.O. 2008. “The chimpanzee has no clothes: A critical examination of Pan troglodytes in models of human evolution.” Current Anthropology 49: 87–114.
Slocombe, K.E. & Zuberbühler, K. 2005. “Functionally referential communication in a chim-panzee.” Current Biology 15: 1779–1784.
Slocombe, K.E. & Zuberbühler, K. 2007. Chimpanzees modify recruitment screams as a function of audience composition. Proceedings of the National Academy of Sciences 104: 17228–17233.
Taglialatela, J.P., Russell, J. L, Schaeffer, J.A. & Hopkins, W.D. 2008. “Communicative signalling activates Broca’s homolog in chimpanzees.” Current Biology 18: 343–348.
Townsend, S.W., Deschner, T. & Zuberbühler, K. 2008. “Female chimpanzees use copulation calls flexibly to prevent social competition.” PLoS ONE 3 (6): e2431. doi:10.1371/journal.pone.0002431.
von Frisch, K. 1927. (1954). The Dancing Bees. London: Methuen. (English translation).Washburn, S.L. & Benedict, B. 1979. “Non-human primate culture.” Man 14: 163–164.

12 William C. McGrew
Wich, S.A. & de Vries, H. 2006. Male monkeys remember which group members have given alarm calls. Proceedings of the Royal Society B 273: 735–740.
Wich, S.A., Swartz, K.B., Hardus, M.E., Lameira, A.R., Stromberg, E. & Shumaker, R.W. 2008. “A case of spontaneous acquisition of a human sound by an orang-utan.” Primates 50: 56–64.
Zuberbühler K. 2006. “Language evolution: The origin of meaning in primates.” Current Biology 16 (4): R123–125.

chapter 2
The evolution and the rise of human language
Carry the baby
E. Sue Savage-Rumbaugh & William M. FieldsGreat Ape Trust/Simpson College
Is it the case that a complete psyche discontinuity exists between ape and man, as a function of human consciousness and human grammatical abilities? The genetic evidence makes this seem unlikely. We offer an alternative explanation, based on the plasticity of neuronal development and the discontinuity between infant clinging and infant carriage in ape and human. Human infants not only fail to cling, they display innate motor patterns of rotational hand-waving and leg kicking. These patterns (absent in ape infants) prevent clinging. In their place, ape infants display an elaboration of the moro-reflex, which enables them to cling with all four limbs shortly after birth. The absence of clinging in the human infant is not due to loss of hair, but to the presence of motor patterns incompatible with clinging. The neuronal- developmental consequences of these contrasting innate patterns form the substrate for the emergence of human/ape differences.
1. Introduction
Although the classic man/animal dichotomy continues to undergird Western scientific thought, evidence from molecular genetics reveals that humans are more closely related to chimpanzees and bonobos than scientists could have surmised, from anatomical evidence, a few decades ago. The degree of DNA similarity between humans, bonobos and chimpanzees now classifies us as sibling species. This means that, as strange as it seems, we are more closely related to each other than either of us is, to a gorilla (Wildman et al. 2002). The behav-ioural discrepancies, and to a lesser degree the morphological ones, are larger than DNA differences readily explain. DNA differences of similar magnitude between other closely related species produce only minor changes in anatomi-cal and cognitive traits. These puzzling facts have led biologists, psychologists, primatologists, anthropologists and philosophers to assert that, at some point in the Homo lineage, a total discontinuity, at the psyche level, emerged between

14 E. Sue Savage-Rumbaugh & William M. Fields
ape and man. The epicenter of this discontinuity is taken to be the appearance of human language. Consequently great effort has been devoted to determining both the evolutionary and the genetic basis of language. Concomitant with this search are the following implicit assumptions regarding language: (a) that it is absent in all other life forms; (b) that it allows for the appearance of reason, self-reflection and moral agency; (c) that it makes possible complex planning; and (d) that material artefacts in the archaeological record which indicate art (such as ochre, markings that have the appearance of intentional symbols, carvings, etc.) are coincident with, as well as sufficient and necessary evidence for, the appearance of human language.
These genetic and philosophical issues will be considered below, in view of the following established facts: (a) bonobos reared in a complex bispecies Pan/Homo environment acquire, through observation, most components of human language; (b) they demonstrate complex self-reflection; (c) they are able to reason; (d) they exhibit moral agency; and (e) they comment upon the past, the future and events displaced in time and space. Importantly, they display these abilities in ways that are distinctly nonhuman, because they are reared – not in a human world as human children – but in a Pan/Homo world (Fields et al. 2007). Their world has provided an enculturation experience unique in evolutionary history. From the cultural fusion produced by the diverse Pan and Homo worlds around them, these bonobos have developed utterances which seem to function as a pigeon language (Savage-Rumbaugh et al. 2004). It is precisely the nonhuman categories and quali-ties of this language that render it difficult for un-acculturated humans to under-stand and accept the validity and the reality of their expressions. It is also these nonhuman qualities that lead to subtle differences in the Pan/Homo construction of moral agency, but not to the absence of moral agency, as often assumed. Fol-lowing human cultural norms, these bonobos make judgments regarding moral agency; but following bonobo cultural norms, they reliably take the welfare of the group into account. For example, if the caloric intake of some bonobos is restricted by staff at a facility, the other bonobos will hide their own food to give to those who are restricted when the staff is not watching.
The Great Ape Trust bonobos represent the only group of apes reared in a Pan/Homo world. Thus, it is logical to expect their linguistic and cultural abilities to differ from those of other living apes, captive or wild. The recurrent failures to find simple protohuman language skills in other apes (such as pointing) are understandable. The deficiency of nonverbal skills in other groups is a direct func-tion of rearing. Other apes have been reared (a) as human children (that is, in a completely human world); (b) as zoological specimens (where minimal cultural information is transmitted because apes are relocated for genetic reasons); or (c) as free-ranging apes. None of these rearing environments would lead to the mental

Chapter 2. The evolution and the rise of human language 15
construction of a Pan/Homo world, as experienced and as expressed by Kanzi and his family. Enculturation into a human world would lack the rich array of rapid sounds, movements and physical experiences present in a bonobo world. These presumably serve an important role in initiating and guiding the development of both the neurological “software” and “hardware” of the infant bonobo brain, enabling it to move toward its full functional capacity. The bicultural rearing of Kanzi and Panbanisha has affected their anatomical structure and their behaviour patterns. They are neither bonobo, nor human. They are neither protohuman, nor childlike. They are bicultural. They can, and do, shift strongly from one pole in their bicultural world, to another, depending upon their environmental circum-stances. The language they comprehend and employ is influenced by the culture Matata brought with her from Wamba.
The complex behaviours of great apes suggest that nonhuman forms of language are waiting to be revealed among free-ranging groups. They have likely gone unnoticed because scientists have searched only for utterances that possess a distinctly human flavour. That is, they have searched for human phonemes, human syntax, human sensibilities toward object names, a human sense of self-agency, a human tendency to evaluate the truth-value of propositional statements and a human kind of vocal fluency that reflects the safety afforded by a home base (Bickerton 2009). Were scientists to search instead for a quiet, symbolic, highly coded system, with intense cognitive loading, their results would, and will in the future, be different (Savage-Rumbaugh et al. 1996).
We suggest that human language is not special because of syntax or semantics, Lana demonstrated both abilities by 4 years of age (Savage-Rumbaugh & Fields, in prep). Human language is special because it portrays the world through linguistic devices that insist upon a fundamental distinction between I and me. These lin-guistic devices cause the human speaker to bifurcate his or her self-agency into two roles, that of the doer and the observer. Both of these conscious entities eventually come to co-reside in the adult human mind, at all times. This bifurcation of human consciousness into an ‘I’ – who becomes a separate entity from the ‘me’ – repre-sents the quintessential trait we call humanness. Through the structural vehicle of human language, our particularly human flavor of consciousness is thus passed on anew to each generation. Consequently, it is the structure of our language which significantly defines and delimits that which we identify as human conscious-ness, and that which we take to be distinct form animal consciousness. All human languages and all human cultures seem to encode this bifurcation between the ‘I’ and the ‘me’, though many indigenous cultures place far less emphasis on it than do modern cultures (Heider 2007, personal observation).
In this bifurcation of consciousness resides the ability of a postulated observer – living in the same body as the active doer, to self-reflect upon his

16 E. Sue Savage-Rumbaugh & William M. Fields
or her own actions, as though they were being performed by another. Out of this ever-present bifurcation and ensuing constant self-reflection arises the emer-gent ability (followed by a nascent sense of responsibility) to make judgments regarding the intent and the normativity of all of one’s own actions, as well as judgments about the actions of others. Also out of this ever-present bifurcation of the self, arises the sense of freewill. For as the observer begins to consider all the actions the doer might have done or might do – and thus to look forward and backward in time – that which might be done takes on a living visualized form in the consciousness of the observer. Thus does the self-reflective observer begin to assume increasingly greater control over the active (and sometimes thought-less) doer. Nonetheless, the doer cannot and does not go away easily, for most immediate action must occur without reflection, as the speed at which we engage our social and physical world demands a quickness that the reflective observer cannot match. The reflective mode constructs its own version of space and time (Castro-Tejerina & Rosa 2007; Calvin 1996, 2000). As a result, we may find we have considerable difficulty in explaining our actions, even to ourselves. However, even when we encounter anomalous circumstances, like those faced by split brain patients, in which the left hemisphere does not know what the right hemisphere sees, the observer is nonetheless able to deftly confabulate an acceptable justifica-tion for the action of the doer (Slunecko & Hengl 2007; Gillespie 2007), though it be completely false.
In addition, whenever we enter a state of hypnosis or heightened suggestibility, we abdicate our state of bifurcated consciousness, and give the ability of the observer (to evaluate and reflect) away to the hypnotist, or to others around us. As we do so, our perceived freewill vanishes, not by design or desire, but rather through our willingness to move into a conscious state which absolves us of the need to self-reflect, to choose our own course action, or to justify to ourselves and to others, the logical and normative (or moral) reasons for our actions.
It is the structural nature of the linguistic separation of the I from the me, coupled with the recursion embedded in human language, which makes possible the construction of questions such as “Why did I do that?” In linguistic construc-tions, the person raising the question is the ‘I’ and the person committing the action is viewed as the ‘me’, but it is still one person. This is a fundamental structural property of human language, and it is from this structural component of human language that the bifurcation of conscious into the primary-self and the reflective-self arises. This becomes the child’s way of perceiving causality as he or she acquires the social vehicle of language. Thus, does the underlying grammatical structure of human language frame our perception of causal reality. Or to put it more pointedly, the personal ego is constructed as we learn to speak because our language requires

Chapter 2. The evolution and the rise of human language 17
the construction of a personal ego. Since our language, once acquired, becomes our main vehicle of thought and reason, we have no ready means of conceptualizing non-ego based realities and/or languages, and we cannot recall what our life and/or reality was like prior to language acquisition.
Previous failures to find language in free-living apes, represent a failure to locate a specifically human language, that is one with recursive structure and the primary I/me division. Locked into our own perceptual frameworks, we fail to realize that our language is directly derived from, bounded by, and enmeshed within, our particular infant rearing patterns, our object gathering and arranging patterns and our home base spatial mapping patterns. These patterns define the basic categories of our world and our linguistic expressions of that world. They impose particular qualities upon the neurological substrate of our infants in ways we have not previously cognized. Yet the neural substrate that defines language is open to virtually any kind of organizational mapping, it only becomes defined to certain functions by the narrow constraints of human language.
Rearing behaviours are heavily cultural in both us and nonhuman apes. Therefore, human-like rearing patterns could quickly be brought into play, in living bonobos, with the appearance of infants less able to cling. Bonobos are prone to adopt bipedal gaits, both in the trees and on the ground. Infants less able to cling would derive rapid morphological adaptation for stable bipedalism, social reorganization with an increased dependence on the role of the male in child rearing, changes in diet and foraging patterns that would necessitate decreased dependence on arboreal feeding (because infants who don’t cling fall out of trees) and possibly the eventual adoption of a ground-based life style with multiple home-bases and inverted nests for protection from rain and predators. Bonobos, in contrast to chimpanzees, already display tendencies in these directions (Kano 1992; Savage-Rumbaugh et al. 1996).
The discovery of Ardipithecus suggests that bipedalism goes back further than previously thought (White et al. 2009). All apes have forms of locomotion distinctly unlike those of monkeys. Knuckle-walking is not the obvious “next step” away from the quadruped gait of monkeys, who employ the inner surface of their fingers, which are splayed backwards during travel. Gibbons (lesser apes who are presumed to be the first to have evolved away from the monkey lineage) are completely bipedal when they move on the ground. Yet they can travel only short distances before they must stop and rest and they never employ their hands for walking. Orangutans walk on their fists. Like the bipedal gibbon, they cannot travel long distances on the ground and they never employ their fingers for walk-ing. They travel too slowly on the ground to avoid predators, and are mostly arbo-real in the wild, unless they are reintroduced to the wild following captive rearing.

18 E. Sue Savage-Rumbaugh & William M. Fields
Only the African apes that have become specialized in the art of knuckle-walking, are able to travel long distances on the ground, with some speed.
Bonobo and chimpanzee infants are not initially adapted to walk upon their knuckles. The changes in the hand and wrist, required to support knuckle-walking, appear only with use. Some chimpanzee and bonobo infants try to become bipedal during the first few years of life. Wild bonobos walk bipedally in clearings, when travelling arboreally on large branches, in swamps and when the ground is wet. When ancient environmental conditions required a bipedal ape to travel farther and farther on the ground with some degree of rapidity, the ability of the infant to cling to a bipedal mother, without losing its grip, rapidly became the limiting factor for survival, not the efficiency of the gait. Any bipedal ape that could bend over, walk on its knuckles and employ its thighs to aid an infant trying to cling, would be able to travel further and faster. But if the infant were supported by the mother’s hands and her bipedal stride became proficient, then travel distance would be limited only by the ability of the mother, or others who might carry the infant, not by the strength of the infant to cling, with the intermittent aid of the mother’s thighs (Savage-Rumbaugh 1994).
If knuckle-walking and many other Homo traits are derived from a common bipedal ancestor, then the relationship we share with chimpanzees and bonobos is distinct from that which we share with other apes. Our DNA reveals that bonobos, chimpanzees and humans are sibling species; and more closely related to each other than to any other apes. At some point we must question whether it is scientifically appropriate to classify bonobos and chimpanzees as animals, and ourselves alone as “human.” This classification, once seemingly correct, does not fit comfortably with the new genetic knowledge. Our seeming differences may reflect the special lifestyles we adopted in the relatively recent past, and these lifestyles, in turn, affect our gene expression profiles. Most scientists view chimpanzees and bonobos as “relics” of an earlier time, long before traits of humanness appeared. We suggest that this view is the product of culturally biased ways of thinking about human/ape differences. Modern life is so far removed from life in the forest that scientists tend to overlook the deeper kinship, hidden by differences in gene expression. Human/bonobo rearing differences begin before birth and are a partial function of the epigenetic effects of arboreal locomotion and its effect on pre and postna-tal gene expression. In coming face-to-face with bonobos or chimpanzees while constructing shared linguistic lives on a daily basis, the superficial anatomical dif-ferences become meaningless, and we are left with the reality of similar minds encountering one another on essentially equal footing. When social interchanges takes place at this level, the human/animal boundary is an outmoded category. In our reality of shared social consciousness, members of both groups constantly experience a linguistically co-constructed continuity and flow of life.

Chapter 2. The evolution and the rise of human language 19
2. Biological humanness
The biological relationship between human and chimpanzee DNA, coupled with the chromosomal fusion or fission events that have occurred in the human/ape lineage, imply that factors other than selection for large brains, upright posture, hunting, throwing, tool manufacture, and the ability to plan for the future resulted in the appearance of the Homo lineage. The DNA data reveal that epigenetic factors, and seemingly random chromosomal rearrangements, affected and are continuing to affect the Pan and Homo lines in ways yet to be understood. For example, it is puzzling that the human genome is far less variable than that of living apes. This implies that apes evolved rapidly while Homo remained stable, and that all current living human beings descended from a relatively small population (Alon et al. 2009; Caswell et al. 2008).
When the chimpanzee genomes was first sequenced, it was thought that the greater number of duplications, inversions, and copy-number variations found, when better studied, would elucidate what made chimpanzees and bonobos different from ourselves. The opposite occurred. The more human genomes that have been sequenced, the more it has become apparent that the original view that only .01% of the human genome differing among humans was incorrect. Humans have copy number variations, duplications, inversions single nucleotide differences that vary from person to person to a considerable degree. Only as genomes of more apes and more humans are sequenced will anything like an accurate picture of evolution have the potential to emerge.
Reconciling the different lines of data produced by genetics and the fossil record is proving increasingly difficult with each new find in either area. The rise in discrepancies suggests that a complete revision of our view of human origins is drawing near. Let us take, by way of example, our most prized human trait, the large brain. According to interpretations of the fossil record, an increase in cranial size has been the major dimension of change and the driving factor of selection and specia-tion toward humanness. But DNA data suggest that the genes which have changed the fastest since the human/chimpanzee split are ones that code for inflammatory responses and cell proliferation (Perry et al. 2008), not brain size. Chimpanzees have improved their ability to resist microbial diseases and pathogenic organisms. Humans, on the other hand, have increased their susceptibility to cancer, an odd adaptation unless, as some speculate, by allowing apoptosis we are enabling the brain to grow larger as well. In addition, there are segments of the human genome that seem to be limiting change, while similar chimpanzee segments are continuing to evolve. Science cannot, at this time, tell us how precisely the genome is accessed or what kind of form will be created, as epigenetic factors act upon development from meiosis forward. Fossil data definitively reveal that many species, displaying

20 E. Sue Savage-Rumbaugh & William M. Fields
characteristics of both apes and humans, existed in the past but the clocks utilized by either approach are subject to error.
On the bases of a genome wide comparison of human and chimpanzee DNA, Cheng et al. (2005) suggest that most of the asymmetrical increase of duplicated DNA in the chimpanzee lineage has emerged as a mechanistic consequence of changes in the chromosome structure and is not due to selection. Most of the difference between human DNA and chimpanzee DNA is due to duplications. This view is echoed by Feuk et al. (2005) who note that the recent genetic evidence on deletions, inversions and copy number variations should ‘cause people to rethink their ideas about how species evolved’ (Howard Hughes Medical Institute, 2005).
The characteristics which taxonomists have traditionally employed as indica-tive of as morphological speciation, do not always correspond to the genetic differentiation being discovered as the genomes of ever more species become sequenced. Species which appear to have different morphologies but similar DNA, express their DNA differently, due to epigenetic factors. Many of these factors are not yet known, but they can include heat shock, population size, magnetic fields, dietary restrictions, and cultural variables. Epigenetic variables can cause changes in gene expression in a single generation. Copy number variations, deletions, insertions and transposons can similarly occur in a single generation, and at high rates. This ever-mushrooming new information regarding the mechanisms under-lying genetic selection and expression are quietly rewriting the Darwinian view of evolution and ‘natural selection.’ While there is no doubt that selection can and has occurred, it is no longer clear that natural selection is playing the major role in morphological change across time.
Darwin developed the hypothesis of natural selection, based on what had been achieved through the selective breeding of dogs and other domestic animals. He extended the idea of intentional selection to include what nature, operating blindly, could produce and reasoned that an intelligent designer was not required for a similar process of selection to operate. All that was required was a sufficiently constant criteria, held in place long enough by any means, to produce change across time (for example larger beaks, thinner beaks, thicker beaks, were the result of feeding constraints and food preferences of various species of bird, etc.). Dar-win extended this principle much further, suggesting that, given sufficient time, these cumulative changes would eventually lead to speciation events, brought about by selective pressures acting upon any new traits that proved valuable to the individual. It is remarkable that Darwin saw these kinds of connections even before there was an understanding of genetics. It is more remarkable that his views have stood the test of time, even as the field of genetics evolved.
But as science increasingly documents the external forces acting upon the genome, the theory of natural selection will face major challenges. To maintain

Chapter 2. The evolution and the rise of human language 21
that environmental events are both (a) causing genes to express particular traits and (b) also selecting for the genes that carry those traits, is not a theoretically viable concept. If flexible gene expression is a feature of selection over the long dure, then the organism would benefit by keeping its genome relatively intact across vast changes in geological time by simply coming up with a way to access its potential differently in new conditions. If the organism adopted such a strategy the genome would function more like a tool-kit than a map, in that it would contain an organism’s building blocks which could be employed in many different ways, depending upon the environmental conditions in place at the time. Some organisms with large genomes, such as corn, appear to have adopted the strategy of utilizing a very large genome to cope with environmental variation. Other organisms, with very small genomes seem to have acquired the ability to use the genes they have in new ways by adding epigenetic information. The latter process is not a matter of selection it is an intelligent biological response to rapid environmental change.
Support for this view comes from the observation that large-scale human alterations of the natural environment can result in extremely rapid changes in species characteristics. Unlike intentional selection, these events, such as habitat homogenization, are accidental, but nonetheless affect living organisms across the population. For example, in response to the building of dams, salmon have begun wintering over rather than migrating to the sea and their morphology has changed as well, in historical time. Also, when farmed salmon breed with wild salmon, hybridization interacts with the environment, in some manner other than selection, to produce a gene expression profile in the offspring which differs from both parents, more than the parents differ from each other. Such observations reveal that organisms have ways of responding to their environments, through gene expression, that are not the products of population selection. These mechanisms allow for rapid adaptive response to broad-scale habitat change. Traditionally, any genetic change not due to population selection has been attributed to random mutations. These ‘mutations’ theoretically became fixed in a population, only when they had a special survival value.
Often overlooked is the simple fact that the lifestyle choices of any mobile organism determine what it will need its genes to do. The assumption that genes can somehow unwittingly determine the organism’s lifestyle choices, while alluring, goes beyond the current biological explication of what genes actually do. While correlational data suggest the existence of genes for everything from ‘shopping behaviour’ to ‘language;’ the simple fact is that our current understanding of genes is limited to the proteins they code for and the developmental pathways that turn them off and on. We have no way to determine, given current scientific methods, how any gene would or could code for the complex behaviours represented in

22 E. Sue Savage-Rumbaugh & William M. Fields
the daily lifestyle choice made by primates. Correlational data which suggest that genes determine these kinds of complex behaviours look only at the relationship between a portion of the genome and behaviour, they do not explain anything at a mechanistic level.
The impact of the ‘new genetics’ upon our understanding of human origins, is just beginning to emerge. With the aid of modern molecular phylogenetics, we currently trace the origin of Homo sapiens sapiens to central Africa between 150,000 and 200,000 years ago (Liu et al. 2006). We also believe that our first ancestors moved out of Africa and into Asia and Europe between 60,000 and 80,000 years ago and that probably only a small group left Africa (possibly as few as 150 people) (Liu et al. 2006; Mancia et al. 2007). This suggests that the Homo erectus and Homo habilis populations found outside Africa are not direct ancestors of modern humans living in those locations today. After Homo sapiens arrived on the scene, more than 80,000 years elapsed (the time period between the anatomical appearance of the species we call Homo sapiens sapiens and the appearance of behaviours which are clearly associated with symbolic processes) before anything is found in the archaeological record to clearly indicate symbolic ability (Lock 2000; Mithen 1996; Yellen et al. 1995). If we trust the archaeological record and assume that this gap is real, rather than the result of inadequate preservation (or current lack of discovery of appropriate remains), then we must conclude that modern human language and symbolic behaviour arose between 40,000 and 100,000 years ago. Before that time, no examples of representational art, fossilized forms of bodily decoration (such as ochre, beadwork, clothing, etc.), hearths for cooked food or indications of widespread trading have been identified, at sites associated with Homo sapien remains. Thus, for a significant period of our existence, we probably lived in forest conditions similar to those inhabited by modern day bonobos. If so, we must have employed a dramatically distinct lifestyle from that which was to emerge later on. For this reason, the designation Homo symbolicus has arisen. Its usage implies the arrival of a unique kind of human entity on the scene – one which differed behaviourally, but not anatomically (at least as far as skeleton remains indicated) from the humans who preceded it.
It has been argued the change began with a single point mutation in one gene, the FoxP2, which enabled the appearance of human language (Enard et al. 2002; Varga-Khadem et al. 2005). When the FOXP2 gene undergoes mutation in modern humans, we manifest a language deficit known as developmental verbal dyspraxia (Vargah-Khadem et al. 2005). This is a disruption of the motor coordination and control required not for speech per se, but for fluent speech. Interestingly this deficit leaves cognition relatively intact. Mice that are genetically engineered to contain a copy of the human version of FOXP2 turn out to be more vocal than normal mice. They also display increased synaptic plasticity and dendrite length in the basal ganglia (Enard et al. 2009; Lieberman 2009) supporting the view that

Chapter 2. The evolution and the rise of human language 23
this gene acts to increase the tendency to make vocal noise. Whether or not this ‘noise’ becomes language depends upon many other factors. Further support for this hypothesis is provided by the discovery that the up-regulation of FOXP2 is related to song acquisition in birds (Haesler et al. 2004).
The chimpanzee version of the FOXP2 gene differs from the human version by only two amino acids. Such a small difference lends credence to the view that humans may have suddenly gained fluent speech (and thus become Homo sym-bolicus) with the incorporation of a single point mutation (Enard et al. 2002). However expression of the FOXP2 gene is regulated by other genes (Carroll 2005; King & Wilson 1975; Scharff & White 2004), which complicates the picture because we still do not understand how gene regulation leads to many human/ape differences. Nonetheless, the fact that such a small mutation could alter vocal fluency implies that the linguistic ability of chimpanzees and bonobos has prob-ably been underestimated, because their lack of speech possibly results from a motor inhibition of vocal behaviour (influenced by FOXP2), rather than a cogni-tive deficit as often assumed. It is logical that their vocal fluency would be kept in check by the high rate of infant predation they experience in the wild. Were this constraint lifted, as humans lifted it with the construction of home bases, infant apes that vocalized frequently and loudly could survive.
A troubling aspect of the Pan-Homo fossil record has always been the lack of fossil apes. This has caused White et al. (2009) to suggest that all modern apes are relics, cul de sacs of an earlier period that existed more or less unchanged for more than 6.5 million years. However, if this is so, then we would expect ape DNA to have changed far less than ours as well, but exactly the opposite is the case (Bakewell et al. 2007). Ape DNA has altered nearly twice as rapidly as ours, sug-gesting that they, not we, are the ones who have undergone environmental pressure for rapid change. But if White et al. (2009) are correct, then our ancestral form was much more apelike – and it is we, not they, who have changed morphologically and behaviourally. If we accept White’s view that all previous “ape-like” beings are protohominins not proto-apes, our DNA should have altered four or five times as rapidly as chimpanzee DNA. Many species of yeast differ by a greater genetic degree than do humans and chimpanzee, and most of the differences between human and chimpanzees lie in noncoding (Pollard 2009).
However it happened, what we do know is that a very special relationship exists between humans, bonobos, and chimpanzee and humans; for ‘there are widespread regions of the genome where chimpanzees and bonobos are less closely related to each other than any of them are to humans’ (Caswell et al. 2008). These findings do not fit well within in any version of current Darwinian Theory. This close relationship is manifest not only in our DNA, but also in the structural and organizational components of that DNA, as expressed in its location on the chromosomes themselves. The human chromosome number two is hypothesized

24 E. Sue Savage-Rumbaugh & William M. Fields
to have resulted from the fusion of chromosomes twelve and thirteen in an ances-tral ape, reducing the total number of human chromosomes from 48 to 46 (Yunis & Prakash 1982; Fan et al. 2002), as chromosome two is essentially identical to chromosomes twelve and thirteen in living chimpanzees. This fusion event, specu-lated to have occurred 15 mya ago, has resulted in either similar changes or no changes, in chromosome two for humans and chromosomes twelve and thirteen in apes and chimpanzees across 15 mya. Given the rapid fossil evolution docu-mented in just the last 6.5 mya for apes, it seems odd that changes on the chromo-somes which fused would remain essentially identical in the living representatives of both species (humans and chimpanzees) for 15 million years.
Inversion coupled with fusion is an uncommon, and typically unstable event, in primate evolution. The possibility exists that, rather than a fusion event, the common ancestral form contained 46 chromosomes and that the modern chim-panzee/bonobo, gorilla and orangutan are the result of at least one and possibly more chromosomal fission events. If so, this would explain why there are no fossil apes (McBrearty & Jablonski 2005), and why each of the great apes shares charac-teristics with humans that the others do not share, for example:
1. orangutans do not walk on their knuckles but African apes do.2. the teeth of orangutans are well enamelled as are human teeth, but not the
teeth of African apes.3. only chimpanzee and bonobo females exhibit sexual swellings.4. only bonobos and humans lack striking sexual dimorphism.5. only bonobo and human females are sexually receptive throughout their cycle.6. only chimpanzees, bonobos and humans live in large mixed social-sexual
groups.
Some of these traits, such as knuckle-walking, do not exist elsewhere in the order Primates. If knuckle-walking was an intermediary stage of locomotion between monkeys and Homo, it would likely be found in some monkeys, in gibbons, siamangs or orangutans, however, it is not. This makes it more reasonable to assume that knuckle-walking represents an adaptation away from bipedalism. Recent reports that chimpanzees in some areas use sticks for walking uphill also support this view (Ben Beck, personal communication, 2010) as does the recent report that an adult male gorilla in a zoo in England suddenly began walking upright.
Another clue to the mysterious origin of modern apes may come from a chromosomal rearrangement that is not uncommon in the human population, the trisomy of Down’s syndrome resulting in 47 chromosomes. The morphological and neurological changes which manifest as Down’s Syndrome include: short stature, decreased life span, speech and cognitive difficulties, larger than normal span

Chapter 2. The evolution and the rise of human language 25
between the first and second toes, a flat nasal bridge, shorter limbs, and differences in affective expression. Similar kinds of differences (short stature, decreased life span, speech and cognitive differences, larger than normal span between the first and second toes, shorter limbs and differences in affective expression) also exist between bonobos and humans, though they manifest differently, and no one would confuse a Down syndrome individual with an ape.
Still, the range of traits affected by Down syndrome reveals that fission events which result in the formation of two new chromosomes would affect a broad range of physical characteristics. Whether two of the 48 ape chromosomes fused to produce 46 human chromosomes, or one of the 46 human chromo-somes fissioned to produce 48 ape chromosomes, cannot be discerned from the fossil record, or from DNA. What is known is that once the lines leading to the African apes and humans began to separate, interbreeding between these lines continued for nearly 1.2 million years, (Patterson et al. 2009), because the X chromosome is younger that the rest of the genome. This suggests that either the differing number of chromosome (46 and 48) had no effect on interbreeding or that during the period of interbreeding the chromosomal number was the same and only changed later. If the latter were the case, then the current suggested date of the fusion at 15mya cannot be correct. If the former is the case, then interbreeding should currently be possible between chimpanzees and humans. Additionally, since the interbreeding that took place after the split lasted for 1.2 million years, ostensibly with Pan males interbreeding with Homo females, (since the X chromosome is younger than the rest of the genome), the question of how interbreeding could have occurred for 1.2 million years, between indi-viduals with different numbers of chromosomes has to be raised.
Should modern apes be the result of multiple fission events (rather than a single fusion event) it also becomes easier to understand (a) why their genes might have evolved at a more rapid rate than our own (to cope with the changes needed to adapt to forest life), (b) why there are no fossil apes, and (c) why there are anom-alies in all living apes, such as the presence or absence of knuckle-walking, tooth enamelisation, female sexual swelling, great sexual dimorphism, blue colouration of teeth in the orangutan, etc.
Additionally we might predict that if modern apes are derived from a human ancestral form, they could retain human cognitive characteristics, such as the capacity for some forms of language and self-awareness, but perhaps have lost the capacity for fluid control of the tongue and diaphragm. Although Lovejoy (2009) does not discuss living bonobos, the traits which are said to have placed Ardipithe-cus on its uniquely human journey (reduced canine size, increased bipedality, and decreased sexual dimorphism), are all extant in modern day bonobos, indicating that these features of hominin evolution are highly plastic, subject to epigenetic modification, and possibly recently derived.

26 E. Sue Savage-Rumbaugh & William M. Fields
3. Ape language and ape culture
Whether the explanation for the appearance of Homo symbolicus lies in the FOXP2 gene, the gradual appearance of our large brain, our capacity for joint regard and imitation, or in some puzzling recombination of chimpanzee genes, it remains the case that human language undergirds all the ways in which we think about ourselves, explain ourselves, plan for our future, conduct our science, devise our mathematics, construct our societies according to the rule of law, attempt to understand the reasons for our existence, fight our wars, plot against our fellow human beings and protect our property rights and individually accumulated wealth. Without human language we could do none of these things. Therefore, it is self-evident that any serious consideration of human origins must either place ape-language studies at the epicenter of that stage, or attempt to discredit them.
Robert Yerkes (1929) was among the first to speculate that, given the other obvious dimensions of chimpanzee intelligence, chimpanzees ought to be able to acquire human language. His views were put to the test by the work of the Kellogg’s in the 30’s with the co rearing experiment of Donald (their son) and Gua (his chimpanzee sister). The results were sufficiently startling as to cause Kellogg to state that the answer was clear even before Gua began to speak. The study was concluded early when Donald began demonstrating chimpanzee behav-iour, including delayed language onset, and Gua began demonstrating human behaviour, including comprehension of spoken language and increased bipedalism (Kellogg & Kellogg 1933).
In the 1970’s breakthroughs arrived with the use of non-speech modalities which produced the first widely heralded evidence for talking apes. These results were quickly disputed by claims that the chimpanzees did not know what they were doing. It was thought that they were producing behaviour that merely “aped language,” through means of imitation, cueing, conditioned discriminations, or chaining (Savage-Rumbaugh et al. 2009). But when two young chimpanzees (Sherman and Austin) began using symbols to communicate with one another, the realization arose that inculcating language also engendered properties of behaviour that altered the way apes communicated with other apes. For example, Sherman and Austin began employing gestures, glances and joint regard while communicating with symbols. They also negotiated outcomes and through such means, corrected symbolic mistakes. They shared food and objects calmly and deliberately. They formed novel two word utterances to communicate with the researchers and with each other. Their utterances, much like those employed by children, constantly reflected an understanding of the mutual knowledge shared by speaker and listener alike; knowledge which is inherent in every linguistic exchange, but which differs from one context to another (Greenfield & Savage-Rumbaugh 1984). None of these

Chapter 2. The evolution and the rise of human language 27
behaviours were shaped. Such observations awakened the scientific community to the fact that language was about something more than just symbols and grammar (Savage-Rumbaugh 1986). They suggested the existence of a deeper nature to language, one that arises out of the way in which it integrates social interaction into normative planned behaviour across time. The interpretations Sherman and Austin attributed readily to each other’s utterances implied the existence of a large body of shared knowledge about the history, and intent, of their past exchanges. To equate such behaviours with conditioned discriminations, as critics were initially prone to do, was to miss the essence of how it is that language builds new interpretations and creative combinations through shared exchanges across historical time.
This work was followed by a project designed to rear a bonobo (Kanzi) in a world lacking didactic language tutoring. In its place, language was constantly employed for normal communicative ends, as happens with human children. This world was intentionally composed of both bonobo and human beings. To rear a bonobo apart from others of its species is to deprive it of the normal social-ization experiences it requires to become a bonobo. Sherman and Austin’s world had been composed of apes and human beings, but Sherman and Austin did not have a chimpanzee mother present. Kanzi’s mother was present and participated in his rearing alongside human companions who permitted themselves to love and care for Kanzi in the same way that she did. Kanzi’s mother, Matata, was born and reared in the Congo. Any language or socio-cultural ways of being which she acquired while growing up were also components of Kanzi’s world. No previous ape language investigation attempted to include a mother as part of the social dynamic of the group, much less a wild-reared ape. This component of the research has not been replicated to date. Across time, the bonobo group expanded from Matata and Kanzi to a family of eight. As the size of the group grew, the complexity of the Pan/Homo world, which was established to rear Kanzi and Panbanisha as Pan/ Homo bonobos, also began to grow exponentially and the number of bonobos in the Pan/Homo community began to vastly outnumber the human members (Savage-Rumbaugh et al. 1986; Lyn and Savage-Rumbaugh 2000; Segerdahl et al. 2005; Savage-Rumbaugh et al. 2007).
The inclusion of bonobo infants into the true sociolinguistic dynamic of the human world as meaningful individuals with expectancies, rights and responsibilities rather than as subjects, was sufficient to result in the spontaneous comprehension of English grammar and the production of a proto-grammar at the lexical level (Savage-Rumbaugh et al. 1993; Greenfield & Lyn 2007; Greenfield et al. 2008). By nine years of age, they readily decoded novel spoken English sentences which included recursions, pronomials, and other syntax dependent utterances (Can you put the can of coke in the trash can, Give the dog a hotdog)

28 E. Sue Savage-Rumbaugh & William M. Fields
as well as concepts of metaphor (for example, Kanzi began to employ, on his own, the colour term ‘yellow’ for cowardly actions), moral agency, and pretense (Can you make the toy snake bite the toy dog?), (Lyn et al. 2008; Lyn et al. 2006; Savage-Rumbaugh et al. 1998). They also communicated representational information to one another through their own vocal channel on a voluntary basis (Taglialatela et al. 2003; Savage-Rumbaugh et al. 2004). Since apes are thought to be unable to vocalize except when the limbic system is emotionally aroused (Zuberbühler 2003), the existence of this ability in the Pan/Homo group implies that human language, acting as an epigenetic variable, has the capacity to override this neurological constraint.
This bicultural world of bonobos and human beings, sought to incorporate as many aspects of free-ranging bonobo culture as possible, therefore spoken language, the lexigram keyboard, and daily travel in a 50 acre forest environment constituted the world and mode of daily life for the experimental group – Kanzi, Panbanisha, Nathan, and Nyota. As infants and juveniles, these bonobos had no sense of being caged or confined, and no sense that bonobos and humans were members of different species. Their group was always composed of bonobos and humans living, interacting and travelling freely together as a unit in the forest. Close relatives of both species were always present. Conversations with humans (or other bonobos) were ubiquitous throughout the day and composed of contextually embedded normative gestural, vocal, and lexical communications about an unlim-ited range of topics directly relevant to all ongoing activities (Benson et al. 2002; Benson & Greaves 2005; Pederson & Fields 2009; Lyn et al. in press). By contrast a control group, consisting of Tamuli, Neema, Maisha, and Elykia were reared by the wild born parents, Matata and P-suke. The interactions of the control group with human beings were limited to those that took place behind wire, and humans and bonobos were clearly treated as different species in this case.
The experimental and control groups lived in different buildings but bono-bos of both groups were provided with frequent opportunities to interact and to form social bonds. Any vocal language which Matata might have brought with her from the wild was present in her behaviour, and thus available to the entire group. This may have played a key role in Kanzi’s acquisition of lexigrams and his understanding of human speech. To the degree that Matata’s vocal world was symbolic, Kanzi was a member of a bilingual world from birth. Children reared in bilingual homes pick up both languages. The early observation that Kanzi or Panbanisha would, when asked to “Tell Matata X or Y” – turn toward Matata and begin to vocalize – suggested an awareness of, and a competency for, a unique bilingual world. Upon moving to Iowa, both groups were housed together from 2006 to 2011. Together they were maintained in the manner originally accorded the control group and most human members of the bicultural group were absent. The effects of this common housing and the separation of the bonobos from the

Chapter 2. The evolution and the rise of human language 29
human members of their group were not investigated. The focus of the research turned toward determining whether or not students and investigators who were basically unknown to the bonobos could collect data about them.
The original rearing environment created a cultural group that was distinctly different from bonobos in zoos or in the wild. These bonobos employed referential gestures (Pederson et al. 2009) demonstrated ToM in the classic linguistic “Sally Ann” test setting (Savage-Rumbaugh 1997), engaged in conversations about com-plex events, including those removed in space and time, and were cognizant of moral rules of engagement and normative action (Savage -Rumbaugh & Fields 2007). They responded to questions regarding their social life and their captive conditions, and reported upon past events of their lives (Savage-Rumbaugh 1999; Savage-Rumbaugh & Fields 2000). They were aware of who they are and mani-fested internal personal and historical narratives of their lives. They functioned as moral agents within their world-view and behaved in socially appropriate and morally responsible ways as adults (Fields 2008).
Even though bonobos comprehend human language, they do not behave as human beings. Instead, they are constantly blending aspects of human culture with those of bonobo culture. To take a small example, Panbanisha began to regularly illustrate where she wanted to travel outdoors, by drawing lexigram-like con-structions on the floor with chalk, then adding a line that went from the lexigram directly toward the door, in the direction to be travelled in the forest. Human beings drew lexigrams on the floor, and in the wild bonobos drag branches in a line indicative of their travel plans. Panbanisha combined these two processes into one – of her own accord.
A high level of linguistic self-control has come only with adulthood. Adulthood has also brought with it an ever-increasing sense of moral agency and evermore reliable moral responsibility. Previous ape language studies never extended close human/ape relationships into adulthood, for fear of physical harm. Thus ape morality, from the standpoint of language related behaviour, was never investi-gated. Kanzi, Panbanisha and Nyota not only ever harm those human beings whom they have known all of their lives, but they protect them without fail. Given that other researchers have not replicated the Pan/Homo world which produced Kanzi, Panbanisha and Nyota, they should not expect other bonobos to display the cogni-tive and linguistic abilities reported for this group. Nor should they be skeptical of the results or assert that experimenters are over-interpreting the data – until they are willing to invest effort to determine the adequacy of those inter pretations from within the normative framework of a Pan/Homo culture. Language always requires interpretations that take into account cultural categories, common knowledge, and historical context. Therein lies the power of language. Speakers of any language, who lack the capacity to make rich interpretations of the utterances of others, are unable to engage in normal social dialogue.

30 E. Sue Savage-Rumbaugh & William M. Fields
A traditional scientific response would be an attempt to replicate the method which produced this unique group of bonobos, rather than to claim that the researchers are intentionally fraudulent (Sebeok & Umiker-Sebeok 1980; Wynn 2008); that the bonobos’ linguistic abilities lack some key aspect of human language (Tomasello 2008; de Waal 2002); or to confuse the issue with so called “language” studies that employ didactic training paradigms intentionally designed to eliminate communication (Matzuzawa 2001; Premack 2004; Povinelli & Vonk 2003; Shumaker & Beck 2003). The eagerness of scholars outside the field to accept superficial critiques as valid (Pinker 1994; Dunbar 2009), without addressing even minimal attention to the unique reality of a Pan/Homo world, illustrates the degree to which human egocentrics views act to maintain the politically correct view of apes as brutish, dull and incapable of behaviour worthy of serious scientific study (Lovejoy 2009; Savage-Rumbaugh et al. 2009).
4. Maternal infant carriage and interaction as substrate of human agency
Mammals, apart from primates, do not carry their offspring. Their infants either remain cached and in contact with siblings until they become mobile (wolves, lions, rodents, etc.) or they are able to walk within hours after birth (antelope, elephants, horses, etc.). Primates changed this basic pattern by bearing an infant that was immobile, but not cached. Their infants survived by clinging to moth-ers who travel constantly while foraging for fruit in trees. Humans changed the basic mammalian pattern yet again by bearing infants unable to cling or to remain immobile. The distinct Homo/Pan styles of infant care (carriage versus clinging) sets the development of the Homo/Pan infant neurological systems on very dif-ferent trajectories. As we shall see, minor changes in these initial set points, lead invariably to salient behavioural differences between humans and apes.
Before birth all mammals begin life as an integral part of the mother’s body. During pregnancy, every sound the mother hears, every move her body makes and every stress she experiences impinges upon, and has the potential to change, the developing embryo within. Truly the baby and the mother are one at this time. Although we know that epigenetic factors, such as drugs, can result in fetal malformation, we have yet to understand the myriad of internal and external prenatal factors that act to set the sensitivities of the nervous system during development. For example, it has only recently been determined that the language a baby experiences before birth, affects the pattern of its crying. The pattern of the infant’s cry also affects the mother and constitutes the first vocal communicative component of cultural exchange between the dyad (Wermke & Mende 2006; Wermke et al. 2007).

Chapter 2. The evolution and the rise of human language 31
Bonobo babies are always in full body contact with a mother or relative. Consequently they initially experience the world, with and through, the actions of the mother; rather than through their own actions as intentional free-willed agents. Such is true of human infants as well, but for a much briefer period. The amount of time human mothers spend carrying babies varies widely from indigenous to modern cultures. Cultures which extend this period tend toward greater co-dependence between adults. Trevarthen (1989) has characterized the experienced consciousness of the infant at this stage as ‘Mommy and I are one’ (Trevarthen & Hubley 1978). Human infants who do not achieve the state of joint consciousness (as well as those who fail to make a smooth transition out of this state) often become less dependent (Acquarone 2007).
For the first several weeks of life, bonobo infants are unable to support their own weight by clinging to the mother’s body for more than five to ten minutes. Bonobo mothers must therefore aid newborn babies by travelling with a crouched posture whilst supporting the baby with their thighs, or with one hand under as they move tripedally (Bolser & Savage-Rumbaugh 1989). The fact that a bonobo baby accounts for 2% or less of the mother’s body weight, makes these patterns of locomotion possible. However the human infant comprises 8 to 10% of the mother’s weight (Brakke & Savage-Rumbaugh 1990), and the mother must either support the baby with both hands or utilize a sling. If bonobo infants were to have body fat at birth they could not cling and the modes of transport utilized by their mothers would be untenable. By one month of age bonobo infants (who remain very thin and lightweight) are able to actively maintain contact with the mother by clinging on their own for up to 20 to 30 minutes. By 3 months of age they require only intermittent support as the mother travels. Bonobo infants cling reflexively from birth. The engagement of the clinging reflex inhibits the rotational hand-waving and foot kicking, so prevalent in human babies. Thus bonobo babies do not have any substantial periods of time during which they move their hands and feet, while observing themselves visually as they lie supine. Human babies tend to spend lots of time observing the movements of their own limbs and, as objects are placed in their hands, the causal effects of shaking and moving objects.
The need to remain with the mother – clinging as she travels – requires essentially all of the bonobo baby’s attention, as well as its constant mental and physical effort. It also ensures that the baby encounters its early world primarily through the lens of its mother’s bodily reactions to the events occurring around them both. The world of a bonobo baby is one of constant motion, constant sound, and constant reactivity to ongoing occurrences. In such a world, bonobos infants receive far less conscious visual attention than would a human baby. They do not cry or fuss for attention. They rarely vocalize except when separated, or to accompany sounds produced by their mothers. Mother-infant vocal dialogues do

32 E. Sue Savage-Rumbaugh & William M. Fields
not occur. Essentially all of the physical and emotional needs of the infant are mediated through the vehicle of constant contact.
Figure 1. Gorilla mother with newborn. This gorilla mother (left) is supporting her newborn infant with both hands, just as would a human mother. However, when she stands, she can no longer use her hands to support her infant, as do human mothers. Therefore her infant must quickly become able to support itself by clinging. To be able to do so it must mature rapidly at first and stay lightweight. Human infants (right) can afford to mature slowly and to add body fat because they do not need to cling to stay with the mother. The mother will keep them with her and will carry them as she travels
Figure 2. A human mother lays her infant down to free her hands. The chimpanzee infant clings to the mother, while being supported by her thighs, leaving the chimpanzee mother’s hands free

Chapter 2. The evolution and the rise of human language 33
Human infants encounter a radically different trajectory of maternal care. They are born unable to cling even if their mother had hair. They are too fat, and too weak. Their heads are too heavy and their necks are not able to support the head. They have reflex patterns of waving the hands and feet that interfere with clinging. Because of this the human mother, rather than the baby, must make every effort to hold the infant or to watch it carefully if she puts it down. The baby not only fails to support any of its own weight, it manifests behaviours which inter-fere with clinging, and it cries easily, which can attract the attention of predators. The added weight of the human infant results from the increase in head size and body fat, rather than an increase in overall body length. Body fat allows human infants to regulate their body temperature more effectively than ape infants, thereby enabling humans to occupy colder climates and to put their infants down. All human babies, regardless of climate, continue to show rapid increase in body fat after birth. They can easily afford to do so because they do not support their weight by clinging. They also need to do so because they are placed in positions where they are out of contact with the mother’s body and thus must regulate their body temperature on their own while still immobile. Even African climates can chill infants during the rainy season. The common view that the human baby is too “immature” to cling is incorrect. As the baby matures, it never reaches a point where it can support its own weight while the mother travels. Likewise, the com-mon view that the ape baby is more mature at birth is also incorrect. Ape babies do not cling because they are mature. They cling because they come equipped with a different set of reflexes, a small head and no body fat. If any of these factors are altered, ape babies do not cling, regardless of their level of ‘maturity.’
Because the human baby does not cling, the mother must decide what to do with it the moment it arrives. She must know how to support the large head, weak neck and mobile spine in a manner that fosters survival and she must either know how to keep the baby quiet or be in the midst of a protective group. Should human mothers wish to use their hands for something other than supporting the baby, they must either sit and put the baby in their lap or place it on a flat solid substrate, or in some object that will support its head and spine. Ape mothers do not have to ‘think about what the baby needs’ – they need only offer it brief support if they perceive it starting to fall.
The far-reaching effects of the lack of clinging upon the developing nervous system have been overlooked. When a non-mobile human infant is placed on a substrate away from the mother’s body, a very different visual, auditory, tactile, and kinesthetic world impinges upon it, than when it is in full body contact with the mother. Instead of looking at a body or a breast, such a baby is suddenly look-ing at the world around it. Often this includes mother’s face and a mouth which is moving in synchrony with the sound that is coming to its ears as she speaks (Figure 3).
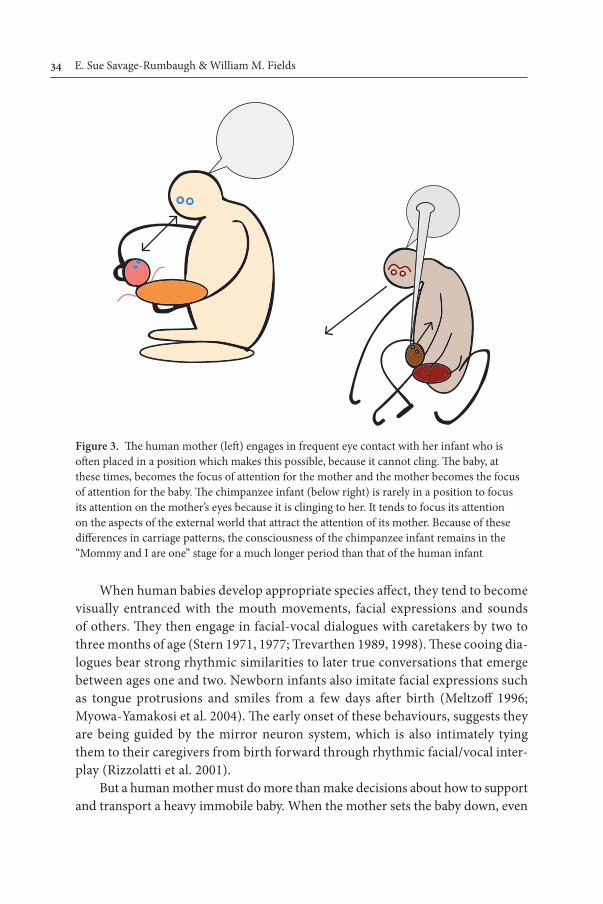
34 E. Sue Savage-Rumbaugh & William M. Fields
Figure 3. The human mother (left) engages in frequent eye contact with her infant who is often placed in a position which makes this possible, because it cannot cling. The baby, at these times, becomes the focus of attention for the mother and the mother becomes the focus of attention for the baby. The chimpanzee infant (below right) is rarely in a position to focus its attention on the mother’s eyes because it is clinging to her. It tends to focus its attention on the aspects of the external world that attract the attention of its mother. Because of these differences in carriage patterns, the consciousness of the chimpanzee infant remains in the “Mommy and I are one” stage for a much longer period than that of the human infant
When human babies develop appropriate species affect, they tend to become visually entranced with the mouth movements, facial expressions and sounds of others. They then engage in facial-vocal dialogues with caretakers by two to three months of age (Stern 1971, 1977; Trevarthen 1989, 1998). These cooing dia-logues bear strong rhythmic similarities to later true conversations that emerge between ages one and two. Newborn infants also imitate facial expressions such as tongue protrusions and smiles from a few days after birth (Meltzoff 1996; Myowa-Yamakosi et al. 2004). The early onset of these behaviours, suggests they are being guided by the mirror neuron system, which is also intimately tying them to their caregivers from birth forward through rhythmic facial/vocal inter-play (Rizzolatti et al. 2001).
But a human mother must do more than make decisions about how to support and transport a heavy immobile baby. When the mother sets the baby down, even

Chapter 2. The evolution and the rise of human language 35
for a short time – things can happen to the baby that could not happen to her. Human maternal care requires a continuous conscious awareness and monitoring of the difference between what is (or could be) happening to the mother herself, as contrasted with what is (or could be) happening to the baby. To take a simple example, if a human baby is exposed to sun, rain, smoke, insects, wind or preda-tors, it can be harmed quickly by events that would be of little consequence to the mother herself. The human mother must realize that heat from the sun or insects or predators, which will not harm her, can quickly harm the baby. She must monitor the baby’s world from the baby’s vantage point, in a way not required for nonliving objects. The ability of the human mother to create in her mind, a con-stant scenario of the baby’s needs and points of view is demanding for many new mothers.
Figure 4. The spine and head of the human infant require support during carriage which the mother must attend to carefully while the infant is very young because the weight of the infant is such that it cannot, until more fully developed, manage to keep the head and the spine aligned easily in all positions. The small head and lack of body fat enable the chimpanzee infant to manage its own postural adjustments of the head and spine from birth, freeing the mother’s attention to watch for predators without worrying about the infant
Their feeling about this effort, and the baby’s receptivity to it, affects the content and style of the linguistic dialogue which then accompanies the mothers actions toward the baby (Oh, you’re all wet, You are getting sleepy, Now smile, don’t be fussy, What is bothering you?). Such dialogue carries a quality of “aboutness” toward the baby and the degree of “aboutness” expressed corresponds closely to the degree of differentiation taking place between mother and infant (Brigaudiot et al. 1996; Savage-Rumbaugh 1990).
Clearly, in the order Primates, something unique happened when babies who could not cling appeared. What are the implications of this for the Pan/Homo

36 E. Sue Savage-Rumbaugh & William M. Fields
culture? Bonobo infants who are carried by human mothers quickly begin to use their hands for something other than clinging and they are exposed to languaged comments about their actions from birth forward. They reach for objects and manipulate them much sooner, and more frequently, than bonobo-only reared infants. In addition they habituate to being placed on the ground and do not cry or show distress when bodily contact is broken gently and slowly.
Human invention of support devices, preoccupation with objects andextension of consciousness enables baby to focus on self
Baby begins to check mother’smonitoring, cries for attention,
long before it can locomote.When mother’s consciousness is
constantly extended to thebaby, fussing is prevented
Once it locomotes, thebaby checks mother’s
position, cries only if mother isnot near. Baby does not fuss
as its consciousness isconstantly extended to the mother
Mother monitors baby’sworld as it cannot locomote
Mother monitors the worldof her group as �rst priority,
self and group overlap
Once baby can locomote itmonitors its own world
Figure 5. Differences in the bodily location of a newborn infant, relative to the mother, place specific constraints on the role of the mother. They also place specific demands on the infant. The mother must monitor any infant that is not in contact with her body. She must also monitor the external surround and any task she is doing. She must shift her attentional priorities between all three. A human infant that is not held (and that is also immobile) needs the ability to cry and to alert the mother that it is in pain or danger, in the event that she has placed her attention on other things. The chimpanzee infant, clinging tightly to its mother, experiences only the same dangers which might affect her. The ability to make noise must become available to the infant even before the infant has the cognitive ability to discern whether or not the situation is one in which a noise might attract a predator and thus prove to be dangerous for the group. The consciousness of the human infant thus becomes separated to communicate to the mother information about states that it experiences and she does not
When placed on a solid surface, they orient toward, and respond appropri-ately to the facial expressions and the rhythmic conversational engagements of

Chapter 2. The evolution and the rise of human language 37
the human mother. In bonding emotionally to a human mother, as well as to a bonobo mother, they begin a neurodevelopmental trajectory that carries them irrevocably along an extraordinary bicultural developmental path into human language. In such a rearing paradigm, human contact alone is not a sufficient variable. A mutual emotional attachment must take place as well. Without it, the bonding that is required to enable the infant to visually, and tactilely engage the rhythms of the human world, fails to take place. Ape infants raised with human contact in zoological nurseries are not the same beings as apes reared in a bicul-tural world. Ape infants raised in nurseries or as pets in human homes, typically manifest autistic behaviours such as rocking and visual detachment (personal observation). These infants experience human contact and care but lack true mothering, bonding and nurturing. They carry the scars of cognitive detachment throughout their lives.
From the different maternal rearing patterns, emerge the styles of conscious-ness which have long interested philosophers (Kitcher 2006; Lock 1690/1959; Savage-Rumbaugh et al. 2005). The consciousness of self-as-agent is emphasized early in ontogeny with the human pattern of rearing. Lengthy en face social engage-ment, visual/vocal exchanges, object manipulation with freed hands, and attuned linguistic overlay all emerge when babies do not cling, but are carried instead (Bullowa 1979; Halliday 1979; Lock 1978). The baby’s attention is focused early upon the results of its actions toward objects or the effects of its social facial/vocal displays toward others (Papousek 2007). Instead of ‘self-agency’ – consciousness of co-agency is emphasized early with the bonobo pattern of rearing. By the time the bonobo infant is locomoting on its own and beginning to develop an aware-ness of self-agency, its primary focus of attention has already become fixed upon group cohesion and group movement through the terrain and it is highly attentive to the slightest alteration in that terrain. When the wild-reared bonobo baby begins to manipulate objects in an exploratory manner, the cognitive information that is accrued is immediately linked into the previously mapped knowledge of the land-scape and how that landscape can be utilized in the service of survival. Thus the solid dense gnarled knobby handled sticks which bonobos employ for digging are not carried from place-to-place, but hidden at useful locations for later use (per-sonal observation, first author 1996). Wild bonobos are very quiet on the ground and extremely noisy in the tree-tops. Bonobo infants are unable to reliably discern which situations are safe for sound and which are not, thus they must be quiet at all times and are unable to cry and/or to make voluntary sounds. The ability to ‘speak at will’ does not appear in bonobos until adolescence or adulthood, therefore this skill, and that of object manipulation, fails to undergo the great freedom of expres-sion during the critical developmental period of childhood. The development of a bonobo baby is constrained by the need to cling and to be quiet.

38 E. Sue Savage-Rumbaugh & William M. Fields
Ape infants quickly loose their insistance on clinging if they arealways supported and put down only when comfortable
�ey begin to be self-aware andto attend to the gaze of the mother
and objects around them beforethey are able to locomote
Figure 6. When an ape infant is reared, as though it were a human infant, it begins to monitor its world and to vocalize for attention. It loses its seemingly constant need to cling and its attention begins to focus on objects at a much earlier age, while it is still relatively immobile. It is often in a position to engage in eye contact and facial gazing and the human mother’s face becomes the focus of its attention and the infant’s face and eyes become the focus of the human mother’s attention enabling the emergence of vocal/facial dialogues. Apes raised as pets, or in entertainment do not experience this kind of human maternal care. Additionally once apes reared as pets are able to cling they are often not supported by human caretakers. They are allowed to cling and support themselves, thereby eliminating, during a critical developmental period, the infant’s developing manual object manipulative skills, and vocal/facial exchanges, thereby forcing the chimpanzee to follow the same developmental path it would follow if it were being reared by a chimpanzee mother. Thus large differences in vocal/facial dialogue, object manipulatory skills and emotional development emerge between so called “human reared” apes
The human child’s attention is, by contrast, drawn toward object play and observation of social interactions. Its hands are free to manipulate objects even before it can sit up and in so doing its actions become landscape independent, and its visual and auditory system becomes human. Mothers provide babies with objects, such as rattles, that they can grasp and move, enabling babies to experience their own capacity to produce noise and movement, at an early age. Mothers over-lay the baby’s world with maternal comments which cast the baby in the role of self as agent and scaffold causal interactions between the baby and themselves (Wolff 1987). Even actions as simple as lifting the baby, offering it food and changing its position are overlaid with causal language (Why are you fussing? Can you hold

Chapter 2. The evolution and the rise of human language 39
still, Do you want some of this?). The highly patterned repetitive interactions of social exchange imprint the infant brain and form a trajectory that manifests (in its adult form) as the sociolinguistic interchange patterns of the group (Lock 2000; Lock & Peters 1996; Rogoff 1990; Vandell & Wilson 1988).
Most behaviours said to characterize the differences between humans and apes derive directly from these early mother-infant rearing patterns and the ensuing alterations in the hands, the gaze patterns, the locomotor patterns, the facial expressions, and the conversational style (Thibault 2004; Taglialatela et al. 2004). Because they occur so early in the development of mind, they set the form and tone for all future social-physical interaction in distinct ways. Sociality is a prerequisite for the emergence of language (Steels 2009), and the nature of that sociality defines the nature of the language which arises and rides upon it. The central dimension of all human solutions to infant rearing is that they are actions extended to the baby by the caregiver. Because it is not the baby’s job to maintain contact with the mother, but rather the mother’s job to maintain contact with the baby, self-agency becomes dimensionally different in ape and man.
5. Self-agency and the duality of consciousness
Most psychologists attribute some level of self-agency to apes, because of their capacity to recognize themselves in mirrors. We suggest that touching a mark on the body (the typical test for self recognition) can be accomplished by understanding that mirrors reflect images. However, awareness of the self, as distinct from being self-aware, requires a bifurcation of consciousness. When individuals seek out mirrors to investigate their appearance because they have intentionally self-altered that appearance, then consciousness has begun to bifurcate into an observer and a doer. Sherman, Austin, Kanzi, Panzee and Panbanisha all painted their faces, put on wigs, shawls, and monster masks, and tried to blow bubbles with bubble gum while using mirrors to check out their altered appearance. Sometimes they practiced display behaviours by adding fur capes while swaggering in front of a mirror. Only bonobos and chimpanzees who have achieved some degree of human language competency display mirror behaviour of this sort (Menzel et al. 1985; Savage-Rumbaugh 1986).
Human language is, after all, a system designed to maximize self-reflection. Its comprehension and use require increasing bifurcation of consciousness. This process is a natural extension of the discovery of self-agency which begins with early human infant care patterns. As the dualistic self-agency begins to operate, there emerges, within a single brain and body, the capacity to simultaneously sepa-rate the imaged self into a doer of actions and an observer of those same actions

40 E. Sue Savage-Rumbaugh & William M. Fields
(Bates 1990). This duality of consciousness permits the organism to think about what it is doing, the appearance of its actions, and how the actions will be perceived by others, at the same time. It is like a seed, which grows slowly, enabling steadily increased reflective control over the actions of the organism by the observer. This capacity to reflect inevitably results in a wish that certain actions had not been taken, either because they did not accomplish the organism’s goals, or because they could not be justified to others (Lewis 1991, 1995; Henriques 2004). With the realization that some actions are judged (by one’s companions) as more correct than others, moral agency obligatorily comes into focus.
When the bifurcation of consciousness is in its initial stages, the doer considers only the potential outcomes for himself and only for short spans of time ahead. With increased sociolinguistic experience, the doer and the observer (reflected linguistically as the ‘I’, who is an agent, and the ‘me’ who is the recipient of the agency of others) bifurcate more fully, and for longer periods of time (Smiley & Johnson 2006). Finally there arises a sense of permanence to the externally objectified conscious experience of, and narrative about, the self. Alternative outcomes of one’s actions can then become recognized as things which are inevitably shared by the group as a whole not just by the doer (Brownell & Carriger 1990). When this realization arises in the mind of the doer, it becomes possible to consciously and reflectively behave in a manner that is constructive or destructive for the larger social group in which one is enmeshed. Such is the sine quo none of human intelligence. While other organisms may benefit from altruism, only the human linguistic organism can consciously practice altruism with the full awareness that constructive action which benefits the group, inevitably benefits the self. From this conscious realization, all the greater principles of what is termed ‘religious awareness’ (for example, “do unto other as you would have them do unto you”) inevitably arise. Thus the human trajectory is, from birth forward, focused upon the growth of conscious awareness. The characteristics of the human infant, demand that it be cared for in ways unique within the order mammlia. All of its survival needs, when met appropriately, allow for the very early emergence of self-awareness. Self-awareness is quickly followed by other-awareness, language and bifurcation of consciousness into the “I” versus the “me” – as well as the “me” versus “you.” Much of the rest of each human beings life is spent learning how to resolve this bifurcation.
This capacity for dual consciousness lies at the root of Cartesianism. However, there is no need to reduce the bifurcation of consciousness to mind/body dualism. When human consciousness (as an emergent electronic property of the neural sys-tem) reaches a certain level of awareness of the self as causal agent (accomplished through the vehicle of human language) it bifurcates and gains the ability to reflect upon itself. As this occurs, consciousness can be thought of as splitting itself into

Chapter 2. The evolution and the rise of human language 41
two parts, rather like a soap bubble which divides in the middle. One half of the consciousness bubble specializes in guiding the immediate actions of the organ-ism and the other half specializes in reflecting upon those actions. The body and brain are quite competent to engage in many complex behaviours, that to all outer appearances, seem to be conscious intentional actions, but without any conscious evaluation of the adequacy of the action, much as occurs in states of dreaming or hypnosis. For shorter periods of time, we all hand over conscious evaluation of the adequacy of our actions to others during social encounters and self-agency becomes blurred.
6. Conclusion
Thus the more advanced properties of human language arise directly from the simple structural properties required to coordinate and order attention and to explore self-agency. These behaviours, which begin at birth, order our perceptions of time and space, our construction of reality, and the construction of our personal narrative. This organization of consciousness occurs so early that we lack any awareness that many aspects of ourselves could ever be fashioned differently. There is sufficient commonality among all human cultures, driven by the exigencies of caring for our immobile heavy infants, as to enable major dimensions of human behaviour to self-manifest in a comprehensible fashion across our different cultures.
Social exchanges occur, however, at a pace too rapid for any extensive conscious on-line self-reflection. Consequently, even highly self-reflective beings depend mainly upon the real-time sociolinguistic rules of engagement which are culturally co-constructed through social interaction during their childhood (Gladwell 2005; Wilson 2002). We know very little about the effects of how we actually interact, and less about what it is we really do during “en face” interactions, because these skills are acquired as we acquire language and function always in concert with it, except in written texts. Our means of social interactions are not merely imprinted upon our brains they become the structural substrate of our brains. This is as true for bonobos as it is for human beings, but the two species learn to do this differently. Both species take great infractions of expected rules of social engagement. This is because the bifurcated mind, once having accepted a distinction between the person who is acting, and the mind controlling the intention behind the action, forever after will always interpret the behaviour of other members of its species as manifestations of intentional acts. Consequently any lack of conformity to group norms is taken as an intentional affront to the larger group values (Shotter 1993). Every action must be considered intentional

42 E. Sue Savage-Rumbaugh & William M. Fields
and every action must be interpreted within the confines of the larger nomethetical context. This is the real dividing line we draw between human behaviour and animal behaviour. Human behaviour is deemed intentional and humans are held responsible for their acts. Animals are not. Childhood, among human beings, is about acquiring the knowledge and skills to intentionally control ones behaviour according to the normative standards of ones culture. Without human language this is not possible to do. One can however, imagine a language that has grammar and semanticity, but no I/Me distinction and which requires no explanation or justification of why one does or says any particular thing. Such a language seems to be the province of wild bonobos and is, we suggest, delimited by the fact that they do not “carry the baby.”
References
Acquarone, S. 2007. Signs of Autism in Infants: Recognition and Early Intervention. London: Karnac Books.
Alon, K., Mullikin, J.C., Patterson, N., Reich, D. 2009. “Accelerated genetic drift on chromosome X during the human dispersal out of Africa.” Nature Genetics 41: 6–70.
Bakewell, M., Shi, P. & Zhang, J. 2007. “More genes underwent positive selection in chimpanzee evolution than in human evolution.” Proceedings of the National Academy of Sciences 104: 7489–7494.
Bates, E. 1990. “Language about me and you: pronominal reference and the emerging concept of self.” In The Self in Transition, C. Cicchetti, and M. Beeghly (eds), 165–182. London: The University of Chicago Press.
Benson, J.D., Fries, P., Greaves, W.S., Iwamoto, K., Savage-Rumbaugh, S. & Taglialatela, J.P. 2002. “Confrontation and support in bonobo-human discourse.” Functions of Language 91: 1–38.
Benson, J.D. & Greaves, W.S. 2005. Functional Dimensions of Ape-Human Discourse. London: Equinox.
Bickerton, D. 2009. Adam’s Tongue: How Humans Made Language, How Language Made Humans. New York: Hill and Wang.
Bolser, L. & Savage-Rumbaugh, E.S. 1989. “Periparturitional behavior of a bonobo.” American Journal of Primatology 17: 93–103.
Bullowa, M. 1979. Before Speech: The Beginning of Interpersonal Communication. Cambridge: Cambridge University Press.
Brakke, K. & Savage-Rumbaugh, E.S. 1990. “Comparative motor and manipulatory development: Behavioral growth in infant human and apes.” Poster presented at the 11 Biennial Conference on Human Development Richmond Virginia.
Brigaudiot, M., Morgenstern, A. & Nicolas, C. 1996. “Guillaume i va pas gagner, c’et d’abord maman: Genesis of the first person pronoun.” In Children’s Language (9), C. Johnson and J. Gilbert (eds), 105–116. Mahwah, NJ: Lawrence Erlbaum Associates.
Brownell, C. & Carriger, M. 1990. “Changes in the cooperation and self-other differentiation during the second year.” Child Development 61: 1146–1174.
Calvin, W.C. 1996. The Cerebral Code: Thinking a Thought in the Mosaics of the Mind. Cambridge: MIT Press.

Chapter 2. The evolution and the rise of human language 43
Calvin, W.C. 2000. Lingua ex Machina: Reconciling Darwin and Chomsky with the Human Brain. Cambridge: MIT Press.
Carroll, S.B. 2005. “Evolution at Two Levels: On Genes and Form.” PLoS Biol. 3 (7): e245. doi:10.1371/journal.pbio.0030245. PMC 1174822. PMID 16000021.
Castro-Tejerina, J. & Rosa, A. 2007. “Psychology within Time: Theorizing about the Making of Socio-Cultural Psychology.” In The Cambridge Handbook of Sociocultural Psychology, J. Valsiner and A. Rosa (eds), 62–81. Cambridge: Cambridge University Press.
Caswell, J.L., Mallick, S., Richter, D. J, Neubauer, J., Schirmer, C., Gnerr, S. & Reich, D. 2008. Analysis of Chimpanzee History Based on Genome Sequence Alignments. PLoS Genet 4 (4): e1000057. doi:10.1371/journal.pgen.1000057.
Cheng, Z., Ventura, M., She, X., Khaitovich, P., Graves, T., Osoegawa, K., Church, D., DeJong, P., Wilson, R.K., Pääbo, S., Rocchi, M. & Eichler, E.E. 2005. A genome-wide comparison of recent chimpanzee and human segmental duplications. Nature 437: 88–93.
de Waal, F.B.M. 2002. Tree of Origin: What Primate Behavior Can Tell Us about Human Social Evolution. Cambridge, MA: Harvard University Press.
Dunbar, R. 2009. “Why only humans have language.” In The Prehistory of Language, R. Botha & C. Knight (eds), 12–35. Oxford: Oxford University Press.
Enard, W., Przeworski, M., Fisher, S., Lai, C., Wiebe, V., Kitano, T., Monaco, A. & Pääbo S. 2002. “Molecular evolution of FOXP2, a gene involved in speech and language.” Nature 418 (6900): 869–872.
Enard, W., Gehre, S., Hammerschmidt, K., Holter, S., Blass, T., Somel, M., Bruckner, M., Schreiweis, C., Winter, C., Sohr, R., Becker, L., Wiebe, V., Nickel, B., Giger, T., Müller, U., Groszer, M., Adler, T., Aguilar, A., Bolle, I., Calzada-Wack, J., Dalke, C., Ehrhardt, N., Favor, J., Fuchs, H., Gailus-Durner, V., Hans, W., Hölzlwimmer, G., Javaheri, A., Kalaydjiev, S., Kallnik, M., Kling, E., Kunder, S., Mobrugger, I., Naton, B., Racz, I., Rathkolb, B., Rozman, J., Schrewe, A., Busch, D., Graw, J., Ivandic, B., Klingenspor, M., Klopstock, T., Ollert, M., Quintanilla-Martinez, L., Schulz, H., Wolf, E., Wurst, W., Zimmer, A., Fisher, S., Morgenstern, R., Arendt, T., Hrabé de Angelis, M., Fischer, J., Schwarz, J. & Pääbo, S. 2009. “A humanized version of Foxp2 affects Cortico-Basal Ganlia Circuits in Mice.” Cell 137: 961–971.
Fan, Y., Newman, T., Linardopoulou, E. & Trask, B.J. 2002. “Gene content and function of the ancestral chromosome fusion site in human chromosome 2q13-2q14.1 and paraloguous regions.” Genome Research 12: 1663–1672.
Feuk, L., MacDonald, J.R., Tang, T., Carson, A.R., Li, M., Rao, G., Khaja, R. & Scherer, S.W. 2005. “Discovery of Human Inversion Polymorphisms by Comparative Analysis of Human and Chimpanzee DNA Sequence Assemblies.” PLoS Genet 1 (4). doi:10.1371/journal.pgen.0010056.
Fields, W.M., Segerdahl, P. & Savage-Rumbaugh, E.S. 2007. “The Materical Practices of Ape Language Research.” In The Cambridge Handbook of Sociocultural Psychology, J. Valsiner and A. Rosa (eds), 164–202. Cambridge: Cambridge University Press.
Fields, W.M. 2008. “Il Kanzi etnografico contro il Kanzi empirico: sulla distinzione tra “Casa” e “Laboratorio” nella vita delle antropomorfe acculturate.” In Rivista di Analisi del Testo Filosofico, Letterario e Figurativo 8, R. Mascialino (ed.), 171–207. Padova: Cleup.
Gillespie, A. 2007. “The Social Basis of Self-Reflection.” In The Cambridge Handbook of Sociocultural Psychology, J. Valsiner & A. Rosa (eds), 678–691. Cambridge: Cambridge University Press.
Gladwell, M. 2005. Blink: The Power of Thinking Without Thinking. New York: Little, Brown and Company.

44 E. Sue Savage-Rumbaugh & William M. Fields
Greenfield, P.M. & Savage-Rumbaugh, E.S. 1984. “Perceived variability and symbol use: A common language-cognition interface in children and chimpanzees.” Journal of Comparative Psychology 98: 201–218.
Greenfield, P.M. & Lyn, H. 2007. “Symbol Combination in Pan: Language, Action and Culture.” In Primate Perspectives on Behavior and Cognition, D. Washburn (ed.), 255–267. Washington D.C.: American Psychological Association.
Greenfield, P.M., Lyn, H. & Savage-Rumbaugh, E.S. 2008. “Protolanguage in ontongeny and phylogeny: Combining deixis and representation.” Interaction Studies 9: 34–35.
Halliday, M.A.K. 1979. “One child’s protolanguage.” In Before Speech: The Beginning of Interpersonal Communication, M. Bullowa (ed.), 171–190. Cambridge: Cambridge Univer-sity Press.
Haesler, S., Wada, K., Nshdejan, A., Morrisey, E.E., Lints, T., Jarvis, E.D. & Scharff, C. 2004. “FOXP2 expression in avian vocal learners and non-learners.” Neuroscience 24: 3164–3175.
Henriques, G.R. 2004. “The development of the unified theory and the future of psychotherapy.” Psychotherapy Bulletin 39: 16–21.
Kano, T. 1992. The Last Ape. Palo Alto: Stanford University Press.Kellogg, W.N. & Kellogg, L.A. 1933. The Ape and the Child: A Study of Environmental Influence
upon Early Behavior. New York: McGraw-Hill.King, M.C. & Wilson, A.C. 1975. “Evolution at two levels in humans and chimpanzees.” Science.
188 (4184): 107–16.Kitcher, P. 2006. “Two normative roles of consciousness.” In The Missing Link in Cognition, H.S.
Terrace & J. Metcalfe (eds), 174–187. Oxford: Oxford University Press.Lewis, M. 1991. “Ways of knowing.” Developmental Review 11: 231–243.Lewis, M. 1995. Shame, The Exposed Self. New York: The Free Press.Lieberman, P. 2009. FOXP2 and human cognition. Cell 137: 800–802.Liu, H., Prugnolle, F.A., Manica, A. & Balloux, F. 2006. “A geographically explicit genetic
model of worldwide human-settlement history.” American Journal of Human Genetics 79: 230–237.
Lock, A. 1978. Action, Gesture and Symbol: The Emergence of Language. London: Academic Press.Lock, A. 2000. “Phylogenetic Time and Symbol Creation: Where Do Zopeds Come From.”
Culture and Psychology 6: 105–129.Lock, A.J. & Peters, C. (eds). 1996. Handbook of Human Symbolic Evolution. Oxford: Clarendon
Press.Lock, J. 1690/1959. An Essay Concerning Human Understanding. 2 Vols. A.C. Fraser (ed.). Oxford
UK: Clarendon. Reprinted 1959. New York: Dover.Lovejoy, O.C. 2009. “Re-examining Human Origins in Light of Ardipithecus Ramidus.” Science
326: 74.Lyn, H. Greenfield, P.M. & Savage-Rumbaugh E. S. 2006. “The development of representational
play in chimpanzees and bonobos: Evolutionary implications, pretense and the role of inter-species communication.” Cognitive Development 21: 199–213.
Lyn, H. & Savage-Rumbaugh, E.S. 2000. “Observational word learning in two bonobos (Pan paniscus): ostensive and non-ostensive contexts.” Language & Communication 20: 255–273.
Lyn, H., Franks, B. & Savage-Rumbaugh, E.S. 2008. “Precursors of morality in the use of the symbols “good” and “bad” in two bonobos (Pan paniscus) and a chimpanzee (Pan troglodytes).” Language and Communication 28: 213–224.
Lyn, H., Greenfield, P.M. & Savage-Rumbaugh, E.S. in press. “Semiotic Combinations in Pan: A Comparison of Communication in a Chimpanzee and Two Bonobos.” First Language.

Chapter 2. The evolution and the rise of human language 45
Matsuzawa, T. 2001. Primate Origins of Human Cognition and Behavior. Tokyo: Springer-Verlag.McBrearty, S. & Jablonski, N.J. 2005. “First fossil chimpanzee.” Nature 437: 105–108.Meltzoff, A.N. 1996. “The human infant as imitative generalist: a 20 year progress report on
infant imitation with implications for comparative psychology.” In Social Learning in Animals: The Roots of Culture, C.M. Heyes and B. G. Galef (eds), 347–370. NY: Academic Press.
Menzel, E., Savage-Rumbaugh, E.S. & Lawson, J. 1985. “Chimpanzee (Pan troglodytes) spatial problem solving with the use of mirrors and televised equivalents of mirrors.” Journal of Comparative Psychology 99 (2): 177–185.
Mithen, S. 1996. The Prehistory of the Mind. New York: Thames and Hudson.Myowa-Yamakoshi, M., Tomonaga, M., Tanaka, M. & Matsuzawa, T. 2004. “Imitation in
neonatal chimpanzee (Pan troglodytes).” Developmental Science 7: 437–442.Patterson, N., Richter, D.J., Gnerre, S., Lander, E.S. & Reich, D. 2009. “Genetic evidence for
complex speciation of humans and chimpanzees.” Nature 441: 1103–1108.Papousek, M. 2007. “Communication in early infancy: An arena of intersubjective learning.”
Infant Behaviour and Development 30: 258–266.Pederson, J. & Fields, W.M. 2009. “Aspects of Repetition in Bonobo Human Conversation:
Creating Cohesion in a Conversation Between Species.” Integrative Psychological and Behavioral Science 43: 22–41.
Pederson, J., Segerdahl, P. & Fields, W.M. 2009. “Why apes point: Indexical pointing in spontaneous conversation of language-competent Pan/Homo bonobos.” In Primatology: Theories, Methods and Results, E. Potocki and J. Krasiñski (eds), 53–74. Hauppauge, NY: Nova Science Publishers.
Perry, G.H., Yang, F., Marques-Bonet, T., Murphy, C., Fitzgerald, T., Lee, A.S., Hyland, C., Stone, A.C., Hurles, M.E., Tyler-Smith, C., Eichler, E.E., Carter, N.P., Lee, C. & Redon, R. 2008. “Copy number variation and evolution in humans and chimpanzees.” Genome Research 18 (11): 1698–1710.
Pinker, S. 1994. The Language Instinct. New York: Morrow.Pollard, K. 2009. “What Makes Us Human.” Scientific American 300: 44–49.Povinelli, D.J. & Vonk. J. 2003. “Chimpanzee minds, suspiciously human?” Trends in Cognitive
Sciences 7 (4): 157–160.Premack, D. 2004. “Is language the key to human intelligence?” Science 303: 318–320.Rizzolatti, G., Fogassi, L. & Gallese, V. 2001. “Neurobiological mechanisms underlying the
understanding and imitation of action.” Nature Review Neuroscience 2: 661–700.Rogoff, B. 1990. Apprenticeship in thinking: Cognitive development in the social context. New
York: Oxford University Press.Savage-Rumbaugh, E.S. 1986. Ape Language: From Conditioned Response to Symbol. New York:
Columbia University Press.Savage-Rumbaugh, E.S. 1990. “Language as a Cause-Effect Communication System.”
Philosophical Psychology 90: 55–77.Savage-Rumbaugh, E.S. 1994. “Hominid evolution: Looking to modern apes for clues.” In
Hominid Culture in Primate Perspective, D. Quiatt & J. Itani (eds), 7–49. Niwot, CO: University Press of Colorado.
Savage-Rumbaugh, E.S. 1997. “Why are we afraid of apes with language?” In The Origin and Evolution of Intelligence, A.B. Scheibel & J. W. Schopf (eds), 43–69. Sudbury, MA: Jones & Bartlett.
Savage-Rumbaugh, E.S. 1999. “Ape Language: Between a rock and a hard place.” In The Origins of Language, Barbara King (ed.), 115–188. Sante Fe: School of American Research Press.

46 E. Sue Savage-Rumbaugh & William M. Fields
Savage-Rumbaugh, E.S., McDonald, K., Sevcik, R.A., Hopkins, W.D. & Rubert, E. 1986. “Sponta-neous symbol acquisition and communicative use by pygmy chimpanzees (Pan paniscus).” Journal of Experimental Psychology: General 115: 211–235.
Savage-Rumbaugh, E.S., Murphy, J., Sevcik, R.A., Brakke, K.E., Williams, S.L. & Rumbaugh, D.M. 1993. “Language Comprehension in Ape and Child.” Monographs of the Society for Child Development 58: 1–256.
Savage-Rumbaugh, E.S., Williams, S., Furuichi, T. & Kano, T. 1996. “Language Perceived, Pan paniscus branches out.” In Great Ape Societies, W.C. McGrew, L.F. Marchant, & T. Nishida (eds), 173–184. New York: Cambridge University Press.
Savage-Rumbaugh, E.S., Shanker, S.G. & Taylor, T.J. 1998. Apes, Language and the Human Mind Oxford: Oxford University Press.
Savage-Rumbaugh, E.S. & Fields, W.M. 2000. “Linguistic, Cultural and Cognitive Capacities of Bonobos (Pan paniscus).” Culture and Psychology 6: 131–153.
Savage-Rumbaugh, E.S., Fields, W.M. & Spircu, T. 2004. “The emergence of knapping and vocal expression embedded in Pan/Homo culture.” Biology and Philosophy 19: 541–575.
Savage-Rumbaugh, E.S., Segerdahl, P. & Fields, W.M. 2005. “Individual differences in language competencies resulting from unique rearing conditions imposed by different first epistemologies.” In Symbol Use and Symbolic Representation, L.L. Namy (ed.), 199–219. Mahwah: Lawrence Earlbaum Associates.
Savage-Rumbaugh, E.S., & Fields, W.M. 2007. “Rules and tools: Beyond anthropomorphism.” In The Oldowan: Case Studies into the Earlies Stone Age, N. Toth & K. Schick (eds), 223–241. Gosport: Stone Age Institute Press.
Savage-Rumbaugh, E.S., Wamba, K., Wamba, P. & Wamba, N. 2007. “Welfare of apes in a captive environment: Comments on and by, a specific group of apes.” Journal of Applied Animal Welfare Sciences 10: 7–19.
Savage-Rumbaugh, E.S., Rumbaugh, D. & Fields, W.M. 2009. “Empirical Kanzi: The Ape Language Controversy Revisited.” Skeptic 15 (1): 25–33.
Scharff, C. & White, S.A. 2004. “Genetic Components of vocal learning.” Annals of the New York Academy of Sciences 1016. 325–347.
Sebeok, T.A. & Umiker-Sebeok, J. 1980. Speaking of Apes: A Critical Anthology of Two-Way Communication with Man. New York: Plenum Press.
Segerdahl, P., Fields, W.M. & Savage-Rumbaugh, E.S. 2005. Kanzi’s Primal Language: The Cultural Initiation of Primates into Language. New York: Palgrave Macmillian.
Shotter, J. 1993. Conversational Realities: Constructing Life Through Language. London: Sage Publications.
Shumaker, R. & Beck, B. 2003. Primates in Question: The Smithsonian Answer Book. Washington, D.C.: Smithsonian Institution Press.
Sluneko, T. & Hengle, S. 2007. “Language, Cognition and Subjectivity: A Dynamic Constitution.” In The Cambridge Handbook of Sociocultural Psychology, J. Valsiner and A. Rosa (eds), 40–61. Cambridge: Cambridge University Press.
Smiley, P.A. & Johnson, R.S. 2006. “Self-referring terms, event transitivity and development of self.” Cognitive Development 21: 266–284.
Steels, L. 2009. “Is sociality a crucial prerequisite for language?” In The Prehistory of Language: Studies in the Evolution of Language, R. Botha and C. Knight, (eds), 36–57. Oxford: Oxford University Press.
Stern, D. 1971. “A micro-analysis of mother-infant interaction: Behavior regulation and social contact between a mother and her 3 1/2 month-old twins.” Journal of the American Academy of Child Psychiatry 12: 402–421.

Chapter 2. The evolution and the rise of human language 47
Stern, D. 1977. The First Relationship. London: Fontana/Open Books.Taglialatela, J., Savage-Rumbaugh, E.S. & Baker, L.A. 2003. “Vocal Production by a Language-
Competent Pan Paniscus.” International Journal of Primatology 24: 1–15.Taglialatela, J., Savage-Rumbaugh, E.S., Rumbaugh, D.M., Bensen, J. & Greaves, W. 2004.
“Language, Apes & Meaning Making.” In The Development of Language: Functional Perspectives on Species and Individuals, G. Lukin and A. Williams (eds), 91–111. New York: Continuum.
Thibault, P. 2004. Agency and Consciousness in Discourse: Self-other Dynamics as a Complex System. London: Continuum.
Tomasello, M. 2008. Origins of Human Communication. Cambridge, MA: MIT Press.Trevarthen, C. 1989. “Sharing makes sense: Intersubjectivity and the making of an infant’s
meaning.” In Language Topics: Essays in Honor of Michael Halliday, R. Steele and T. Thread-gold (eds), 177–191. Amsterdam: John Benjamins.
Trevarthen, C. 1998. “The concept and foundation of infant intersubjectivity.” In Intersubjectivity, Communication and Emotion in Early Ontogeny, S. Braten (ed.), 183–189. Cambridge: Cambridge University Press.
Trevarthen, C. & Hubley, P. 1978. “Secondary intersubjectivity: confidence, confiding and acts of meaning in the first year.” In Action, Gesture and Symbol: The Emergence of Language, A. Lock (ed.), 183–229. London: Academic Press.
Vandell, D.L. & Wilson, K.S. 1988. “Infants’ interactions with mother, sibling, and peer: Contrasts and relations between interaction systems.” Child Development 48: 176–186.
Vargah-Khadem, F., Gadian, D.G., Copp, A. & Mishkin, M. 2005. “FOXP2 and the neuroanatomy of speech and language.” Nature Review of Neuroscience 6: 131–138.
Wermke, K., Leising, D. & Stellzig-Eisenhauer, A. 2007. “Relation of melody in infant cries to language outcome in the second year of life.” Clinical Linguistics & Phonetics 21: 961–973.
Wermke, K. & Mende, W. 2006. “Melody as a primordial legacy from early roots of language.” Behavioral and Brain Sciences 29: 300.
White, T., Asfaw, B., Beyene, Y., Haile-Selassie, Y., Lovejoy, C.O., Suwa, G. & WoldeGabriel, G. 2009. “Ardipithecus ramidus and the Paleobiology of Early Hominids.” Science 236: 64.
Wildman, D., Grossman L. I. & Goodman, M. 2002. “Functional DNA in humans and chimpan-zees shows they are more similar to each other than either is to apes.” In Probing Human Origins, M. Goodman & A. S. Moffat (eds), 11–32. Cambridge, MA: American Academy of Arts and Sciences.
Wilson, T. 2002. Strangers to Ourselves: Discovering the Adaptive Unconscious. Cambridge, MA: Harvard University Press.
Wolff, P.H. 1987. The Development of Behavioral States and the Expression of Emotions: Early Infancy: New Proposals for Investigation. Chicago: University of Chicago Press.
Wynn, C. 2008. “Aping Language: A skeptical analysis of the evidence for nonhuman primate language.” Skeptic 13: 10–14.
Yellen, J.E., Brooks, A.S., Cornelissne, E., Mehlman, M.J. & Stewart, K. 1995. “A Middle Stone Age bone industry from Katanda, Upper Semliki Valley, Zaire.” Science 268: 553–557.
Yerkes, R. 1929. The Great Apes: A Study of Anthropoid Life. New Haven, CT: Yale University Press.Yunis, J.J. & Prakash, O. 1982. “The Origin of Man: A Chromosomal Pictorial Legacy.” Science
215: 1525–1530.Zuberbühler, K. 2003. “Referential signaling in non-human primates: Cognitive precursors
and limitations for the evolution of language.” In Advances in the Study of Behavior, P.P. Slater, J. Rosenblatt, C. Snowdon, T. Roper, & M. Naguib (eds), 265–307. New York: Academic Press.


chapter 3
The origin of symbolically mediated behaviour
From antagonistic scenarios to a unified research strategy
Francesco d’Errico1,2 & Christopher S. Henshilwood2,3
1CNRS-UMR 5199 PACEA, Université de Bordeaux/2Institute for Archaeology, History, Culture and Religion, University of Bergen/3Institute for Human Evolution, University of the Witwatersrand
The aim of this paper is to summarize what we do know about the origin of symbolic material cultures in Africa and Eurasia, and explore paths that could allow us to move from a situation in which the same evidence is accounted for by antagonistic scenarios to a research strategy that may produce, in the end, a unified theory for the emergence of this innovation. Our review highlights the need to develop an integrated research strategy in which assumptions on cognition based on taxonomic affiliation will play no role. The key tools to address this topic should be archaeology, palaeoenvironmental studies and new methods to integrate results from these disciplines. This appears to be one positive way to understand the mechanisms that governed cultural transmission and social learning between 160 and 30 ka.
1. Introduction
A fundamental change in human behaviour occurred when symbolism became inherent in material culture. This innovation, which demonstrates the ability for sharing, storing and transmitting coded information within and across groups, has played a crucial role in creating, maintaining and transmitting the social con-ventions, beliefs and identities that characterise all known human societies. The origin of symbolic material culture, long associated with a rapid cognitive change in Europe 40 ka ago, is now considered by many to have gradually emerged in Africa, in conjunction with the origin of our species. Others stress the use of sym-bolic artefacts by Neandertals, and their burial practices, to challenge the idea that symbolic behaviour is peculiar to our species.
The question of the emergence of symbolically mediated behaviour has stimulated heated debates in the last decade for a number of good reasons. The view one adopts on this issue has implications for our vision of the factors that have driven

50 Francesco d’Errico & Christopher S. Henshilwood
cultural evolution in our lineage (genetic or cultural drift, climate, demography), the way they have interacted, and the timing and mode of the emergence of other crucial human quintessential features such as modern cognition, working memory, language, religious beliefs, culturally transmitted social ranking systems, artificial memory systems and so forth. The aim of this paper is to summarize what we do know at present about the origin of symbolic material cultures in Africa and Eurasia, and explore paths that could allow us to move from a situation in which the same evidence is accounted for by antagonistic scenarios to a research strategy that may produce, in the end, a unified theory for the emergence of this innovation, and a better comprehension of the mechanisms that have stimulated the emergence, maintenance and transmission of this unique human adaptation.
We have long known that symbolic thinking, defined here as the capacity to attribute specific meaning to conventional signs, is not peculiar to our species (McGrew, this volume; Savage-Rumbaugh & Fields, this volume). We share that capacity with a growing number of primates and non-primate species. The key criterion is not the capacity for symbolic thought but the creation of symbolic material culture that unequivocally integrates socially shared symbolic meaning. It is through the existence of these symbols that humans are able to condition their and others’ actions and choices.
Until very recently the emergence of symbolism was considered coincident with the arrival of Homo sapiens in Europe at 40 ka. This view, commonly known as the “Human Revolution” scenario (Mellars 1996; Klein 2008), has been replaced in the last decade by the “out of Africa” model (McBrearty & Brooks 2000; Mellars 2006, 2007). This latter model argues that symbolically mediated behaviour developed in Africa and is linked with the emergence of Homo sapiens in that continent (Henshilwood & Marean 2003, 2006; Henshilwood & Dubreuil 2011). According to this view the appearance of modern cognitive behaviours (syntactical language, advanced cognition, symbolic thinking) in Eurasia, associated with Homo sapiens, is possibly the result of the dispersal of an already symbolic species at c. 60 ka or earlier. This spread would have led to rapid extinction of pre-modern human populations in Asia and Europe with little or no biological exchange and limited cultural interaction. However, an additional model that we may term the ‘Symbolic Neandertal’, suggests that the material culture associated with Neandertals in Europe before the first arrival of Homo sapiens was associated with behaviour that was mediated by symbols (Zilhão & d’Errico 1999; d’Errico 2003; Zilhão et al. 2010; d’Errico & Stringer 2011). If correct, then it seems possible that there was a parallel evolution towards behavioural modernity and the use of symbols on two continents by two or more distinct populations. At this stage the effect of genetic exchange between Homo sapiens and Neandertals on their respective evolution of material culture and

Chapter 3. The origin of symbolically mediated behaviour 51
innovations is not well understood (Green et al. 2010). The tenets of the parallel evolution model favours a scenario in which demographic changes triggered by environmental variation may have stimulated, but not necessarily synchronically, the emergence of innovations among both modern humans in Africa and Neandertals in Eurasia. Acquisition of innovations from a neighbouring population is also a possibility, though difficult to test outside the Near East and Europe, the areas where we know Homo sapiens and Neandertals came into direct contact. If demonstrated, the acquisition of innovation through contact would support rather than contradict the ‘Symbolic Neandertal’ scenario since it would show, as is the case in many historical instances, that both populations were able to assimilate cultural innovations created by other groups.
2. First instances of symbolic material culture and their implications
Deliberate inhumation and special treatment of the dead, such as secondary burial, cremations, intentional curation of the skeleton or selected body parts and the conscious use of human bone as a raw material to produce artefacts, are generally regarded as quintessential features of modern humanity (Pearson 1999; McBrearty & Brooks 2000; Henshilwood & Marean 2003; d’Errico et al. 2003). The oldest modern human burials date back to at least 100–130 ka (Grün et al. 2005). Neandertal burials are as old or might be older if one accepts the most ancient date for the Tabun C1 burial (Bar-Yosef & Callender 1999; Grün & Stringer 2000; Mercier et al. 2000).
The oldest burials in Africa, which may date to 60–76 ka, are those from Nazlet Kather (Vermeersch et al. 1984; Crevecoeur 2006) Qena, Egypt (Vermeersch et al. 1998) and Border Cave (Millard 2006; Bird et al. 2003). The intentional character and symbolic significance of burials prior to 30 ka, especially those of Neandertals, has been the subject of intense debate over the past decades. Most Neandertal burials were excavated long ago and available reports often lack the information now required to assess the anthropogenic origin of the inhumation and whether items interpreted as grave goods were deliberately interred. There is nevertheless enough information on Neandertal burials to accept that in some areas of Europe and the Near East these populations systematically buried their dead (Vandermeersch et al. 2008). Grave goods consisting of stone tools, bone retouchers, engraved bone and a rock slab engraved with cupules are also reported at La Ferrassie, La Chapelle-aux-Saint, Le Moustier in France, and Amud and Dederiyeh in the Near East.
The earliest abstract designs, engraved on bone and ochre, are found in South Africa (Figure 1) and are dated to ca 100 ka (Henshilwood et al. 2002, 2009; see
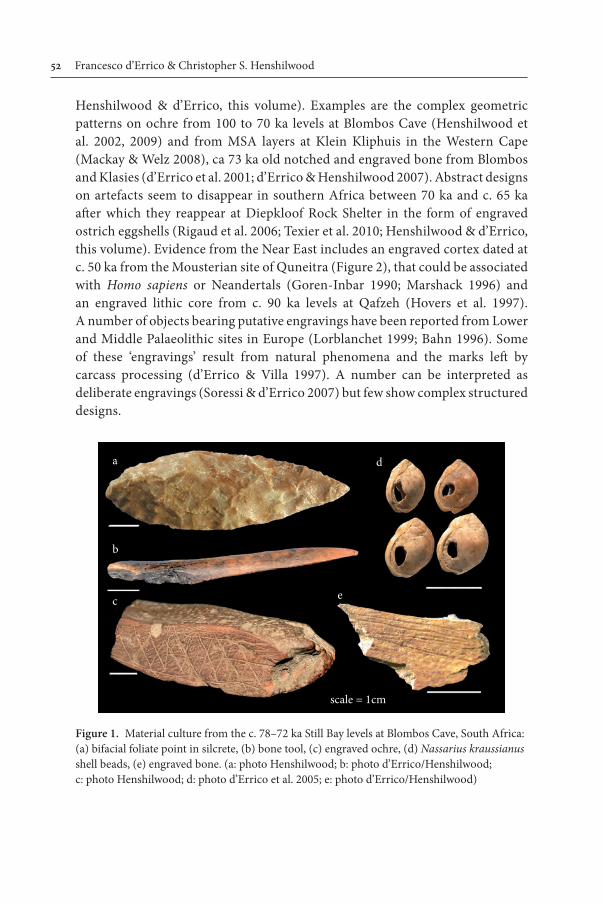
52 Francesco d’Errico & Christopher S. Henshilwood
Henshilwood & d’Errico, this volume). Examples are the complex geometric patterns on ochre from 100 to 70 ka levels at Blombos Cave (Henshilwood et al. 2002, 2009) and from MSA layers at Klein Kliphuis in the Western Cape (Mackay & Welz 2008), ca 73 ka old notched and engraved bone from Blombos and Klasies (d’Errico et al. 2001; d’Errico & Henshilwood 2007). Abstract designs on artefacts seem to disappear in southern Africa between 70 ka and c. 65 ka after which they reappear at Diepkloof Rock Shelter in the form of engraved ostrich eggshells (Rigaud et al. 2006; Texier et al. 2010; Henshilwood & d’Errico, this volume). Evidence from the Near East includes an engraved cortex dated at c. 50 ka from the Mousterian site of Quneitra (Figure 2), that could be associated with Homo sapiens or Neandertals (Goren-Inbar 1990; Marshack 1996) and an engraved lithic core from c. 90 ka levels at Qafzeh (Hovers et al. 1997). A number of objects bearing putative engravings have been reported from Lower and Middle Palaeolithic sites in Europe (Lorblanchet 1999; Bahn 1996). Some of these ‘engravings’ result from natural phenomena and the marks left by carcass processing (d’Errico & Villa 1997). A number can be interpreted as deliberate engravings (Soressi & d’Errico 2007) but few show complex structured designs.
a
b
c e
scale = 1cm
d
Figure 1. Material culture from the c. 78–72 ka Still Bay levels at Blombos Cave, South Africa: (a) bifacial foliate point in silcrete, (b) bone tool, (c) engraved ochre, (d) Nassarius kraussianus shell beads, (e) engraved bone. (a: photo Henshilwood; b: photo d’Errico/Henshilwood; c: photo Henshilwood; d: photo d’Errico et al. 2005; e: photo d’Errico/Henshilwood)

Chapter 3. The origin of symbolically mediated behaviour 53
Convincing evidence for the use of personal ornaments, consisting of perforated marine shell belonging, at each site, to a single species, is found at sites from South Africa (Henshilwood et al. 2004; d’Errico et al. 2005), North Africa (Bouzouggar et al. 2007; d’Errico et al. 2009), and the Near East (Vanhaeren et al. 2006) dated to between 100 and 70 ka. At Blombos Cave forty nine deliberately perforated Nassarius kraussianus shell beads (Figure 1) with clear evidence of use-wear and some bearing traces of ochre come from 75 ka old levels (Henshilwood. 2004; d’Errico et al. 2005). Possible beads made of Littorina africana marine shells were found in the upper Still Bay layers of Sibudu Cave (d’Errico et al. 2008). The perforated Conus shell from Border Cave, associated with the burial of a young individual may be as old as 76 ka according to the recent chronological attribution of this burial (Millard 2006). Perforated Nassa-rius gibbosulus shells (Figure 2) were recovered at Skhul from c. 100 ka levels that include eleven Homo sapiens burials (Vanhaeren et al. 2006). Perforated shells of the same species (Figure 2) showing traces of intentional modifications, possible deliberate heating to change the colour of the bead, use-wear, and traces of red ochre were recovered from the Aterian site of Oued Djebbana, Algeria, and the 80–70 ka levels at Grotte des Pigeons, Rhafas, Ifri n’Ammar, and Contrebandiers in Morocco (d’Errico et al. 2009).
Other marine shells interpreted as beads (Figure 2) come from the c. 90 ka Mousterian levels XXI–XIV at Qafzeh Cave in Israel (Bar-Yosef Mayer et al. 2009). These consist of ten naturally perforated Glycymeris insubrica shells, some with traces of red pigments. The only Neandertal site predating the arrival of modern human populations in Europe that have yielded possible evidence for the use of beads by Neandertals is Cueva de los Aviones, southern Spain (Zilhão et al. 2010). The Mousterian layers of this site, dated to ca 50–45 ka, contained marine shell assemblages including three valves of Acanthocardia and Glycymeris shells bearing natural perforations (Figure 2). One of the latter contained a residue of red pig-ment identified as haematite.
Beads disappear in Africa and the Near East between 70 ka and 40 ka (d’Errico & Vanhaeren 2007; d’Errico et al. 2009) and reappear almost every-where in Africa and Eurasia after this time span. In Europe 40 ka beads are associated with both Neandertals and Homo sapiens (d’Errico et al. 2003). These beads differ from their 100–70 ka antecedents in that they take the form of hundreds of discrete types that identify regional patterning (Vanhaeren & d’Errico 2006). At 40 ka, beads in Africa are made from ostrich eggshells and only later are a diverse range of raw materials introduced for bead manufacture (Figure 2).

54 Francesco d’Errico & Christopher S. Henshilwood
a
b c
d
ge
f
Scale = 1cm
Figure 2. (a) engraved cortex from the site of Quneitra (ca 50 ka), Israel; (b) Nassarius gibbosulus shell beads from Skhul, Israel (ca 100 ka); (c) Oued Djebbana, Algeria (>40 ka); (d) Grotte des Pigeons, Taforalt, Morocco (ca 82 ka) respectively; (e) perforated Glycimeris from Qafzeh, Israel (ca. 90 ka); (f) perforated Glycimeris from Cueva de los Aviones, Spain (ca 50 ka); (g) Ostrigh eggshell beads from Mumba cave (ca 40 ka). (a: photo d’Errico/Nowell; b–d: photo d’Errico/Vanhaeren; e: modified after Bar-Yosef et al. 2010; f: by courtesy of N. Conard, University of Tübingen)
In south east Asia, the oldest documented ornament is a perforated tiger shark tooth found in New Ireland, New Guinea at a site dated between 39.5 and 28 ka (Leavesley 2007). The earliest evidence for bead use in Australia comes from the site of Mandu Mandu, Cape Range of Western Australia, where 22 Conus shell beads were recovered in a layer dated to ca. 32 ka (Morse 1993). In addition, ten Dentalidae shell beads are reported from the 30 ka old layers of Riwi in the Kimberly region of Western Australia, a site located 300 km inland (Balme & Morse 2006).
The systematic use of red pigments (Figure 3) is evident in Africa at archae-ological sites dated to 160 ka (Marean et al. 2007) and possibly at sites dated

Chapter 3. The origin of symbolically mediated behaviour 55
to 280 ka (McBrearty & Brooks 2000). In the Near East the oldest evidence for systematic use of pigments dates to ca 100 ka (Hovers et al. 2003; d’Errico et al. 2010). Pigments, mostly black, have been sporadically used by Neandertals in Europe since 300 ka (Marshack 1981) but their use only becomes systematic after 60 ka (d’Errico 2003; Soressi & d’Errico 2007; Zilhão et al. 2010). The last Neandertals in France and Italy made intensive use of both black and red pigments just before, and at the moment of contact with Homo sapiens in Europe (Salomon 2009; Ronchitelli et al. 2009).
a
b
c
d
Scale = 1cm
Figure 3. Early instances of pigment use in the Near East, Africa, and Europe; (a) Skhul cave, Israel (100 ka); (b) Blombos Cave, South Africa, Still Bay layers, ca 75 ka; (c) Pech-de-L’ Azé I, Dordogne, France, Mousterian of Acheulean tradition, ca 50 ka; (d) Klasies River, South Africa, Howieson Poort layers, ca 60. ka. (a: photo d’Errico et al. 2010; b–d: photos F. d’Errico)

56 Francesco d’Errico & Christopher S. Henshilwood
Different interpretations have been placed on early pigment use. Some archaeologists have interpreted pigments as proof of a growing use of symbols (Knight et al. 1995; Watts 1999, 2002; McBrearty & Brooks 2000; Henshilwood et al. 2002; Henshilwood & Marean 2003; Hovers et al. 2003; d’Errico 2003; d’Errico et al. 2009; Zilhão et al. 2010; others stress the deliberate choice of pigments for their intense red hues (Watts 1999); a preference for pigments from far away sources (Zilhão et al. 2010); the deliberate heating of pigments to change their colour (Hovers et al. 2003); the deliberate use of pigment to stain one side only of an object (Zilhão et al. 2010); to colour shell beads (Henshilwood et al. 2004; d’Errico et al. 2005, 2009); and as decoration on bodies or on clothing. It is also suggested that early pigment use may have been utilitarian, for example as skin protection from sun or insects, as a medicine, for tanning hides, or as a binding agent to facilitate hafting (Velo 1984; Klein 1999; Wadley 2001; Klein & Edgar 2003; Wadley et al. 2004, 2009; Lombard 2007).
Figurative representations (Figure 4), consisting of painted, engraved and carved animals, appear much later, at c. 28 ka in Africa, at Apollo 11 shelter (Wendt 1976), Namibia, and at 32 ka in Europe, for example at Chauvet, Fumane and in southern Germany (Clottes 2003; Conard et al. 2004). The oldest carved musical instruments, consisting of flutes made of bird bone and mammoth ivory decorated with notches, are found in Europe and date to 32 ka (d’Errico et al. 2003; Conard et al. 2004). No convincing musical instruments are associated with Neandertals (d’Errico & Lawson 2006).
Inferring symbolic capacity from bone and stone tools is a notoriously tricky endeavour. If we accept, with some caveats (Wiessner 1983), the definition of style as an over - determination of form imposed by cultural standards that identifies ethnically bounded populations (Byers 1994, 1999; Chase 2006) then convincing instances of such behaviour can be found in southern Africa stone tool assemblages. Examples include the stone bifacial points from the c. 75 ka Still Bay levels at Blombos Cave (Villa et al. 2009), in the Near East Mousterian from 100 ka (Hovers & Belfer-Cohen 2006) and in Europe with regional traditions appearing in the Mousterian at c. 60 ka (Delagnes & Meignen 2005). Recent research has shown that such trends do not follow a progressive development. In South Africa, as in Europe, techno-complexes consisting of highly developed tool kits alternate with periods in which stylistic intent, if present, becomes difficult to perceive (Villa et al. 2009).
Fully shaped bone tools, some of which are decorated with engraved lines and notches (spear points, awls, spatulas, harpoons) occur in Africa from c. 75 ka and possibly at 90 ka (Henshilwood et al. 2001; d’Errico & Henshilwood 2007; Yellen et al. 1995; Backwell et al. 2008). The careful deliberate polishing of the ca 75 ka Blombos bone projectile points has no apparent functional purpose and, rather,

Chapter 3. The origin of symbolically mediated behaviour 57
seems a technique used to give a distinctive appearance and/or an ‘‘added value’’ to this category of artefacts. This may imply that symbolic meaning was attributed to bone tools. Bone tools disappear periodically after 70 ka in Africa and are only in common and continued use in the final periods of the Later Stone Age. Nean-dertals produced complex bone tools, some of which are decorated, only after 40 ka (d’Errico et al. 2003; d’Errico et al. 2011). From 35 ka, bone tools are ubiquitous in European tool kits associated with Homo sapiens.
a
f
b c
d
e g
h
i
j
k
Figure 4. First examples of figurative representations from Africa and Europe: (a) painted slab from Apollo 11 Cave, Namibia (ca 28 ka BP), (b) figurine from Straitzig/Krems-Rehberg, Austria (ca 32 ka BP), (c–d) wall fragments from Fumane, Italy (ca 32 ka BP), (e) wall paintings from Chauvet Cave, France (ca 32 ka BP), (f) human with a lion head from Hohlenstein-Stadel, Germany (ca 32 ka BP), (g–i) mammoth, lion and lion head from Vogelherd, Germany (ca 32 ka BP); (j) female figurine from Hohle Fels, Germany (ca 32 ka BP), (k) Sculpted vulvas from the Aurignacian layers of La Ferrassie rock shelter, France

58 Francesco d’Errico & Christopher S. Henshilwood
Our succinct review of the evidence for symbolic material culture (1) contradicts the idea that the production of symbolic material culture is the result of a sudden change in human cognition occurring in Europe or in Africa after 50–40 ka; (2) suggests the presence of symbolic material culture in Africa by at least 150 ka, in the Near East after 100 ka and probably by at least 60–50 ka in Europe; (3) indicates that engravings, pigments, personal ornaments, formal bone tools and burial practices do not appear in the archaeological record as a single package; (4) re-emphasizes, the equivocal status of pigment and more strongly of stone and bone tool stylistic trends as hallmarks of symbolic behaviour; (5) reveals that during c. 150 ka and 30 ka these material culture innovations appear, disappear and reappear in different forms suggesting major discontinuities in cultural transmission; (6) contradicts, if one accepts reported instances of symbolic artefacts associated with Neandertals, the assumption that ground breaking innovations can be associated only with Homo sapiens.
3. Demography as a triggering factor?
The complex patterns associated with innovations in material culture that we describe above cannot be explained solely by speciation. The evidence shows that it is discontinuous in time and space, and the commonalities found in both hemispheres indicate that once a cognitive threshold is crossed, local conditions must have played a role in the emergence, diffusion and the eventual disappearance of crucial innovations. These local conditions must have been closely linked to the size and organisation of cultural systems and ecological settings in which these populations evolved.
Can selective pressure eliminate cultural complexity and refined innovations when they are deemed unfit for use within a group or population? What are the conditions under which this can happen? Archaeological data and ethnographic and historical accounts show that cultural groups can lose innovations. At the beginning of the 20th century William Rivers (1912: 123) speculated on the rea-sons that led to the loss of seaworthy canoes in Oceania. His interpretation was that “social and magico-religious, as well as material and utilitarian factors should be taken into account”. Homo sapiens who colonised Tasmania at c. 35 ka, took with them a number of cultural innovations that they transmitted and implemented for 25 ka. After 10 ka, with the flooding of the Bass Strait, Tasmania was transformed into an island. The population was reduced to a few thousand and the technology employed by these people became very simple compared to that employed on the Australian mainland (Diamond 1978).
One research direction has been to try and create models of cultural transmission and then to link these models to models of genetic evolution. It is

Chapter 3. The origin of symbolically mediated behaviour 59
a tradition for which most archaeologists and palaeoanthropologists have paid little attention. Examples of this research are the seminal papers and book by Cavalli-Sforza and Feldman (1981), followed by Boyd and Richerson (Boyd & Richerson 1985; Richerson et al. 2005, 2009). Only recently have scholars tried to apply or adapt these models to account for the emergence and loss of cultural innovations. Recent papers (Shennan 2001; Henrich 2004; Powell et al. 2009; Premo & Kuhn 2010) have explored this approach by modelling the role of demography in the emergence and loss of cultural innovations. Shennan examines the vertical and oblique cultural transmission of craft-techniques in small size populations and highlights the crucial role of population size in the spread and maintenance of innovations. He supports the assumption that the emergence of modern human culture can be associated only with Homo sapiens and that even though there is sporadic evidence of this before 70 ka, symbolically mediated culture is prevalent only after 50 ka. Henrich’s model is based on the observation that humans have a propensity to imitate skilfull individuals. The combination of skill-biased transmission and learning errors would create a cumulative adaptive evolution. In this process the number of social learners appears as the key factor and this would account for the loss of technological innovations in Tasmania. Powell et al. (2009) extend Henrich’s model by partitioning, in two regions, different population densities, cultural exchange rates and migratory activities. Premo and Kuhn (2010) reach a comparable conclusion for cultural evolution in the Early Stone Age. Powell et al’s conclusion is that demography and migration are the key factors influencing cultural exchange. Theoretically their model accounts for the appearance and disappearance of innovations and for the delay between the origin of anatomically modern Homo sapiens and that of the first evidence for modern cultural traits. Their results are certainly significant as they provide a sound explanation for the emergence and loss of innovations without invoking speciation as the prime mover, but they nevertheless leave open a number of questions. Like Shennan (2001), these authors do not consider the possibility that their model accounting for the emergence of symbolic culture in Homo sapiens could be equally applied to Neandertals. Following the logic of Powell et al. (2009), one may argue that differences between Neandertals and Homo sapiens with regard to cultural modernity may have depended on variations in group size and the rate of cultural exchange rather than on hard wired differences in cognition.
Using a modelling approach to highlight the potential role of demographic factors in the emergence of cultural modernity does not provide absolute proof that this was the necessary path. Models are models and even when they seem to broadly account for reality they need to be tested against the empirical evidence. The question is how to test these models and what do we accept as proof that they account for past reality. Each appearance of a cultural innovation in the MSA, or of the spread of innovations associated with migratory activities such as the colonisation of Europe

60 Francesco d’Errico & Christopher S. Henshilwood
and Asia, are interpreted by Powell et al. (2009) as archaeological reflections of an increase in population size. Conversely, cultural discontinuities are interpreted as signs of demographic demises. Evidence supporting these links is however tenuous. Through genetics alone we are unable to reconstruct the timing of repeated bottle-necks or their impact on population dynamics, for example at c. 70 ka (Rampino & Self 1993; Ambrose 1998). Powell et al. (2009) evoke the climatic deterioration during Marine Isotope Stage (MIS) 4 as a possible factor leading to the decline, fragmentation and range contraction of human populations but they propose no clear mechanisms to explain this. Invoking climatic change as the prime mover equates with attributing these changes to a single speciation event. It creates a “deus ex machina” that answers the question without solving the problem.
4. Climate as a triggering factor
One paradox of research into the evolution of humans is the observation that climate change was the motor behind speciation and behavioural change. Only coarse tools however have been developed that integrate the relevant climatic data with human evolution, and the dialogue between archaeologists and palaeoclimatic researchers is limited. For example, what is the state of our current knowledge for the period 160–20 ka, the time during which symbolic material culture emerges?
Geological research records indicate that during the last million years the Earth underwent repeated changes between glacial periods, in which large continental ice sheets developed in the northern hemisphere, and interglacial periods with greatly reduced global ice volume. These changes link with changes in the amount of solar insolation received at high northern latitudes during the summer, with dominant cyclicities at 23, 41 and 100 ka reflecting shifts in the Earth’s orbital geometry, and the potential for the development of significant ice sheets (Imbrie 1992, 1993; Milankovitch 1969). Within this framework, major interglacials occur approximately every 100 ka, with periods of subtler warming, known as interstadials, corresponding to periods of increased northern summer insolation at c. 23 ka intervals. These large-scale cycles of climate change are well-documented in both northern and southern hemisphere polar ice cores (Dansgaard et al. 1993; Jouzel et al. 2007), and understood to define global climates during the late Pleistocene.
During the last 15 years, concerted research efforts have focused on rapid or millennial-scale climate changes occurring during the last glacial cycle (c. 80–10 ka to present). Detailed climate records (Figure 5) from the Greenland ice cores and North Atlantic marine cores reveal that the last glacial period was not uniformly cold, but was punctuated by abrupt atmospheric warming events,
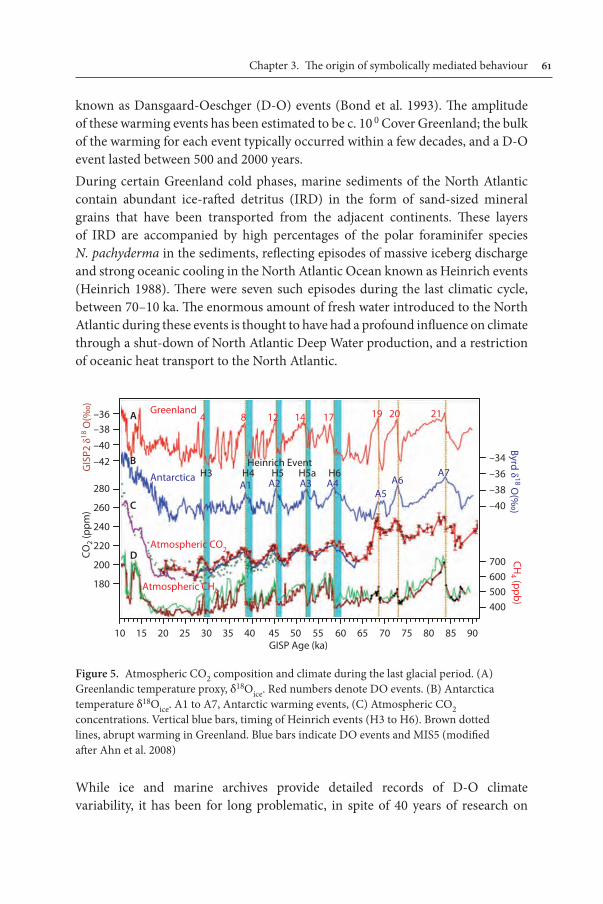
Chapter 3. The origin of symbolically mediated behaviour 61
known as Dansgaard-Oeschger (D-O) events (Bond et al. 1993). The amplitude of these warming events has been estimated to be c. 10 0 Cover Greenland; the bulk of the warming for each event typically occurred within a few decades, and a D-O event lasted between 500 and 2000 years.During certain Greenland cold phases, marine sediments of the North Atlantic contain abundant ice-rafted detritus (IRD) in the form of sand-sized mineral grains that have been transported from the adjacent continents. These layers of IRD are accompanied by high percentages of the polar foraminifer species N. pachyderma in the sediments, reflecting episodes of massive iceberg discharge and strong oceanic cooling in the North Atlantic Ocean known as Heinrich events (Heinrich 1988). There were seven such episodes during the last climatic cycle, between 70–10 ka. The enormous amount of fresh water introduced to the North Atlantic during these events is thought to have had a profound influence on climate through a shut-down of North Atlantic Deep Water production, and a restriction of oceanic heat transport to the North Atlantic.
Greenland
Antarctica
–36–38
–40
–42
280
260
240
CO2 (
ppm
)
–34–36–38–40
700600500400
CH4 (ppb)
GIS
P2 d
18 O
(‰)
Byrd d18 O
(‰)
220
200
180
10 15 20 25 30 35 40 45GISP Age (ka)
50 55 60 65 70 75 80 85 90
Atmospheric CO2
Atmospheric CH4
A
B
C
D
4
H3 H4A1 A2 A3 A4
A5A6
A7H5 H5a H6Heinrich Event
8 12 14 17 19 20 21
Figure 5. Atmospheric CO2 composition and climate during the last glacial period. (A) Greenlandic temperature proxy, δ18Oice. Red numbers denote DO events. (B) Antarctica temperature δ18Oice. A1 to A7, Antarctic warming events, (C) Atmospheric CO2
concentrations. Vertical blue bars, timing of Heinrich events (H3 to H6). Brown dotted lines, abrupt warming in Greenland. Blue bars indicate DO events and MIS5 (modified after Ahn et al. 2008)
While ice and marine archives provide detailed records of D-O climate variability, it has been for long problematic, in spite of 40 years of research on

62 Francesco d’Errico & Christopher S. Henshilwood
long pollen records, to link specific events in the ice and marine domains with vegetation changes, mostly because the duration of individual events is often less than the chronological uncertainties involved in the dating of these records. A major advance in this field has been introduced by the analysis of pollen and microcharcoal deposited in marine environments. Transported by winds and river currents into the ocean, these continental marine proxies sink rapidly through the water column as a part of the marine snow along with marine and ice volume climatic indicators, such as foraminifera on which stable isotope measurements are performed. In this way vegetation changes can be directly correlated with SST changes and ice dynamics. The analysis of cores strategically located along the Iberian and French margin has allowed researchers to document how vegetation and fire regime are affected by the millennial scale climatic variability of MOS 2 and 3 at different latitudes (Sanchez-Goñi et al. 2008).
Over the past decade and beyond, concentrated efforts by archaeologists, climatologists, geographers and environmental scientists are contributing to a fine tuning of the climatic variations in southern Africa after the end of the previous interglacial (Marine Isotope Stage 6/5e (MIS)), (e.g. Grindley 1969; Deacon & Lancaster 1988; Van Andel 1989; Thackeray 1992, 2007; Avery 1995; Partridge 1993, 1997; Lambeck & Chappell 2001; Blunier & Brook 2001; Bateman et al. 2004; Carr et al. 2006; Scholz et al. 2007; Roberts et al. 2008; Henshilwood 2008). One example is the IGCP 437 (International Geological Correlation Program) that examines coastal environmental change during sea-level high stands (Mastronuzzi et al. 2005). There remains, however, significant discordance and debate among records and researchers as to how regional environments have changed as a response to past cycles of global warming and cooling as registered in polar ice cores.
Attempts to understand the impact of environmental variability on human adaptation have used a variety of datasets and methods and have primarily focused on the archaeological records of Africa and Europe. In Africa, research has attempted to model the impact of environmental changes on hominin evolution, dispersal, and cultural evolution (Potts 1996; deMenocal 2004; McBrearty & Brooks 2000; Sepulchre et al. 2006; Henshilwood 2008). Some researchers have focused on the Toba super-eruption a c. 73 ka, and its possible role in creating a genetic bottleneck (Ambrose 1998; Rampino & Self 1993). Research projects have recently been targeted at South Africa during MIS 6–2 ( Henshilwood 2008; Henshilwood & Marean 2006; Jacobs et al. 2006; Marean et al. 2007; Jacobs & Roberts 2008). These pilot studies have reached differing conclusions concerning the influences of climatic variability on human behaviour, and it is clear that no consensus yet exists on how best to integrate the wide variety of datasets from multiple disciplines into a single analytical approach. This lack of consensus has led researchers to propose a direct link between ice core records, regional climates

Chapter 3. The origin of symbolically mediated behaviour 63
and population dynamics (e.g. Jacobs & Roberts 2008). This approach may identify general trends but it is clear that a better knowledge of the impact of climate on continental ecosystems at a regional scale is necessary to model those processes (c.f. Chase & Meadows 2007).
A similar situation exists in Europe where a variety of records have been used to examine climate human interaction. One of the better known European endeavours was the interdisciplinary Stage 3 Project (van Andel & Davies 2003), but its results were hampered by the fact that the scholars relied only on terrestrial climatic records and employed simplified palaeoclimatic reconstructions. Others have performed similar research at both continental (Bocquet-Appel & Demars 2000; Bocquet-Appel et al. 2005) and regional scales (Blockley et al. 2006; d’Errico & Sanchez-Goñi 2003; d’Errico et al. 2006; Finlayson 2004; Finlayson & Carrion 2007; Gamble et al. 2004; Stringer 2006; Sepulchre et al. 2007). In some cases these research projects have involved more precise and up-to-date climate models taking in due account the marine and terrestrial record (Sepulchre et al. 2007). In the framework of the ESF-funded RESOLuTION project, marine and terrestrial palaeoclimatic records have been examined in order to reconstruct palaeoenvironments more precisely (Ampel et al. 2008; Sanchez-Goñi et al. 2008; Fletcher & Sanchez-Goñi 2008) with an eye on how they impacted human populations, but such research has concentrated exclusively on MIS 3–2.
5. Discussion
We should now move forward and find a consensual way to make progress in this field. Our review highlights the need to develop an integrated research strategy in which assumptions on cognition based on taxonomic affiliation will play no role. The key tools should be archaeology, palaeoenvironmental studies and new methods to integrate results from these disciplines. This appears the only way to reconstruct the timing and mode of the emergence of key innovations in material culture in Europe and southern Africa, to identify whether and how climatic changes have influenced Neandertals and Homo sapiens’ distributions and behavioural patterns, and to understand the mechanisms that governed cultural transmission and social learning during this crucial time.
We argue that this requires (1) new archaeological excavations and dating of archaeological sites in Africa and Europe located in areas associated with the emergence and loss of cultural innovations. This need is more cogent in Africa where some cultural phases are only represented by a few sites but is also necessary in Europe where many key sites are undated; (2) the application of innovative methods to the analysis of early symbolic and complex material culture of Homo sapiens and

64 Francesco d’Errico & Christopher S. Henshilwood
Neandertals, including burials, abstract engravings, pigments, personal ornaments and stylised bone and stone tools. A large portion of this crucial archaeological material has never been properly analysed and one can reasonably predict that a closer analysis may strengthen the symbolic interpretation, for example in the case of pigments; (3) reconstruct climate, vegetation, fire regime and faunal changes in Europe and southern Africa by combining the analysis of multiple proxies from marine and terrestrial archives with high resolution palaeoclimatic simulations and up-to-date analyses of faunal assemblages; (4) bring together archaeological and palaeoenvironmental data, a coupled atmosphere-ocean General Circulation Model (GCM) and subsequently derived high resolution atmospheric simulations, and apply to them new tools that allows for the reconstruction, quantification and comparison of the ecological niches exploited by human populations within each climatic phase; (5) run realistic multi-agent based models to explore the impact of the reconstructed climatic changes on population dynamics at a regional scale.
Such an integrated approach is now within our reach. Recent advances in biodiversity studies have developed tools for estimating ecological niches of spe-cies and predicting their responses to environmental changes. It has been demon-strated that tools such as GARP (Genetic Algorithm for Rule Set Prediction) and Maxent have considerable potential for reconstructing eco-cultural niches of past human populations, defined as the potential range of environmental conditions within which a human adaptive system can exist, and identifying the environmen-tal parameters that define those niches (Banks et al. 2008, 2009). To do so, GARP identifies palaeoclimatic features shared by archaeological sites with contempo-raneous cultural levels, dated to a specific climatic event, and then finds other regions with the same palaeoclimatic parameters. Through hundreds of error-correcting iterations, it obtains an optimal eco-cultural niche reconstruction. GARP also allows for a reconstructed eco-cultural niche for one climatic phase to be projected onto the climatic conditions of a different one in order to evaluate whether an eco-cultural niche has remained stable, expanded, contracted, or even disappeared. This approach detects changes in niches and their respective ranges, and identifies the environmental and cultural mechanisms behind them. GARP has been already successfully applied for identifying and quantifying the mecha-nisms behind human-environment interactions in Europe during the Last Glacial Maximum and the Middle-to-Upper Palaeolithic transition (Banks et al. 2008).
If applied to the question at hand, this approach can potentially verify whether the disappearance of key cultural innovations was linked to environmental shifts. If so it may be possible to identify the underlying environmental factors and cul-tural mechanisms that lie behind stable or shifting niches. If climate turns out to have been a key factor in affecting the evolution and devolution of cultural

Chapter 3. The origin of symbolically mediated behaviour 65
innovations in the past this will serve to remind us that cultural innovations are not inscribed in our genes – climate change does not only affect what we can or cannot do but it also specifically defines who we were and what we have become.
Acknowledgements
Financial support was provided to the authors by the European Research Council Advanced Grant TRACSYMBOLS (FP7 No. 249587), and the PROTEA French-South Africa exchange programme. CSH was also supported by a National Research Foundation/Department of Sci-ence and Technology funded Chair at the University of the Witwatersrand, South Africa, the Norwegian Research Council and the University of Bergen.
References
Ahn, J. & Brook, E.J. 2008. “Atmospheric CO2 and Climate on Millennial Time Scales During the Last Glacial Period.” Science 322 (5898): 83–85.
Ambrose, S. 1998. “Late Pleistocene human population bottlenecks, volcanic winter, and differentiation of modern humans.” Journal of Human Evolution 34 (6): 623–651.
Ampel, L., Wohlfarth, B., Risberg, J. & Veres, D. 2008. “Paleolimnological response to millennia l and centennial scale climate variability during MIS 3 and 2 as suggested by the diatom record in Les Echets, France.” Quaternary Science Reviews 27: 1493–1504.
Avery, D.M. 1995. “Physical environment and site choice in South Africa.” Journal of Archaeo-logical Science 22: 343–353.
Backwell, L., d’Errico, F. & Wadley, L. 2008. “Middle Stone Age bone tools from the Howiesons Poort layers, Sibudu Cave, South Africa.” Journal of Archaeological Science 35: 566–1580.
Bahn, P. 1996. “New developments in Pleistocene art.” Evolutionary Anthropology 4: 204–215.Balme, J. & Morse, K. 2006. “Shell beads and social behaviour in Pleistocene Australia.” Antiquity
80 (310): 799–811.Banks, W.E., d’Errico, F., Townsend Peterson, A., Masa Kageyama, M., Sima, A. & Sanchez-Goñi,
M.F. 2008. “Neanderthal Extinction by Competitive Exclusion.” PLoS ONE 3 (12): 1–8.Banks, W., Zilhão, J., d’Errico, F., Kageyama, M., Sima, A. & Ronchitelli, A. 2009. “Investigating
Links between Ecology and Bifacial Tool Types in Western Europe during the Last Glacial Maximum.” Journal of Archaeological Science 36: 2853–2867.
Bar-Yosef Mayer, D., Vandermeersch, B. & Bar-Yosef, O. 2009. “Shells and ochre in Middle Paleolithic Qafzeh Cave, Israel: indications for modern behaviour.” Journal of Human Evolution 56: 307–314.
Bar-Yosef, O. & Callender, J. 1999. “The woman from Tabun: Garrod’s doubts in historical perspective.” Journal of Human Evolution 37: 879–885.
Bateman, M.D., Holmes, P.J., Carr, A.S., Horton, B.P. & Jaiswal, M.K. 2004. “Aeolianite and barrier dune construction spanning the last two glacial–interglacial cycles from the southern Cape coast.” South Africa Quaternary Science Reviews 23: 1681–1698.

66 Francesco d’Errico & Christopher S. Henshilwood
Bird, M.I., Fifield, L.K., Santos, G.M., Beaumont, P.B., Zhou, Y., di Tada, M.L. & Hausladen, P.A. 2003. “Radiocarbon dating from 40 to 60 ka BP at Border Cave, South Africa.” Quaternary Science Review 22: 943–947.
Blockley, S.M., Donahue, R.E., Lane, C.S., Lowe, J.J. & Pollard, A.M. 2006. “The chronology of abrupt climate change & Late Upper Palaeolithic human adaptation in Europe.” Journal of Quaternary Science 21: 575–584.
Blunier, T. & Brook, E.J. 2001. “Timing of millennial-scale climate change in Antarctica and Greenland during the Last Glacial period.” Science 291: 109–112.
Bocquet-Appel, J.-P. & Demars, P.-Y. 2000. “Population kinetics in the Upper Paleolithic of Western Europe.” Journal of Archaeological Science 27: 551–570.
Bocquet-Appel, J.-P., Demars, P.-Y., Noiret, L. & Dobrowsky, D. 2005. “Estimates of Upper Paleolithic Meta-Population Size in Europe from Archaeological Data.” Journal of Archaeo-logical Science 32: 1656–1668.
Bond, G., Broecker, W.S., Johnsen, S., McManus, J., Labeyrie, L.D., Jouzel, J. & Bonani, G. 1993. “Correlations between climate records from North Atlantic sediments and Greenland ice.” Nature 365: 143–147.
Bouzouggar, A., Barton, N., Vanhaeren, M., d’Errico, F., Collcutt, S., Higham, T., Hodge, E., Parfitt, S., Rhodes, E., Schwenninger, J.-L., Stringer, C., Turner, E., Ward, S., Moutmir, A. & Stambouli, A. 2007. 82 000-year-old shell beads from North Africa and implica-tions for the origins of modern human behaviour. Proceedings of the National Academy of Sciences 104: 9964–9969.
Boyd, R. & Richerson, P.J. 1985. Culture and the Evolutionary Process. Chicago: University of Chicago Press.
Byers, A.M. 1994. “Symboling and the Middle-Upper Palaeolithic transition: a theoretical and methodological critique.” Current Anthropology 35: 369–399.
Byers, A.M. 1999. “Communication and Material Culture: Pleistocene Tools as Action Cues.” Cambridge Archaeological Journal 9: 23–41.
Carr, A.S., Thomas, D.S.G. & Bateman, M.G. 2006. “Climatic and sea level controls on Late Quaternary aeolian activity on the Agulhas Plain.” South Africa Quaternary Research 65: 252- 263.
Cavalli-Sforza, L.L. & Feldman, M. 1981. Cultural Transmission and Evolution. Princeton: Princ-eton University Press.
Chase, B.M. & Meadows, M.E. 2007. “Late Quaternary dynamics of southern Africa’s winter rainfall zone.” Earth Science Reviews 84 (3–4): 103–138.
Chase, P.G. 2006. The Emergence of Culture: the Evolution of a Uniquely Human Way of Life. New York: Springer.
Clottes, J. 2003. Return to Chauvet Cave: Excavating the Birthplace of Art: the First Full Report. London: Thames & Hudson.
Conard, N.J., Malina, M., Münzel, S.C. & Seeberger, F. 2004. Eine Mammutelfenbeinflöte aus dem Aurignacien des Geissenklösterle: neue Belege für eine musikalische Tradition im Frühen Jungpaläolithikumauf der Schwäbischen Alb. Archäologisches Korrespondenzblatt 34: 447–462.
Crevecoeur, I. 2006. Etude anthropologique des restes humains de Nazlet Khater (Paléolithique supérieur, Egypte). Thesis (Ph.D.). University of Bordeaux 1.
d’Errico, F. 2003. “The invisible frontier. A multiple species model for the origin of behavioural modernity.” Evolutionary Anthropology 12: 182–202.
d’Errico, F. & Villa, P. 1997. “Holes and grooves: the contribution of microscopy and taphonomy to the problem of art origins.” Journal of Human Evolution 33: 1–31.

Chapter 3. The origin of symbolically mediated behaviour 67
d’Errico, F., Henshilwood C. & Nilssen, P. 2001. “An engraved bone fragment from 70,000 ka-year-old Middle Stone Age levels at Blombos Cave, South Africa: implications for the origin of symbolism and language.” Antiquity 75: 309–318.
d’Errico F. & Sanchez-Goñi M. F. 2003. “Neandertal extinction and the millennial scale climatic variability of the OIS 3.” Quaternary Science Reviews 22 (8–9): 769–788.
d’Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Soressi, M., Bresson, F., Tillier, A.-M., Maureille, B., Nowell, A., Backwell, L., Lakarra J. A. & Julien, M. 2003. “The search for the origin of symbolism, music and language: a multidisciplinary endeavour.” Journal of World Prehistory 17: 1–70.
d’Errico, F., Henshilwood, C., Vanhaeren, M. & van Niekerk, K. 2005. “Nassarius kraussianus shell beads from Blombos Cave: evidence for symbolic behaviour in the Middle Stone Age.” Journal of Human Evolution 48: 3–24.
d’Errico, F. & Lawson, G. 2006. “The sound paradox. How to assess the acoustic significance of archaeological evidence?” In Archaeoacoustics, C. Scarre & G. Lawson (eds), 41–57. Cambridge: McDonald Institute Monographs.
d’Errico, F., Sanchez-Goñi, M.F. & Vanhaeren, M. 2006. “L’impact de la variabilité climatique rapide des OIS3-2 sur le peuplement de l’Europe.” In L’homme face au climat, E. Bard (ed.), 265–282. Paris: Odile Jacob.
d’Errico, F. & Henshilwood, C. 2007. “Additional evidence for bone technology in the southern African Middle Stone Age.” Journal of Human Evolution 52: 142–163.
d’Errico, F. & Vanhaeren, M. 2007. “Evolution or revolution ? New evidence for the origin of symbolic behaviour in and out of Africa.” In Rethinking the Human Revolution, P. Mellars, K. Boyle, O. Bar-Yosef & C. Stringer (eds), 275–286. Cambridge: McDonald Institute Monographs.
d’Errico, F., Vanhaeren, M. & Wadley, L. 2008. “Possible shell beads from the Middle Stone Age layers of Sibudu Cave, South Africa.” Journal of Archaeological Science 35: 2675–2685.
d’Errico, F., Vanhaeren, M., Barton, N., Bouzouggar, A., Mienis, H. & Richter, D. 2009. “Additional evidence on the use of personal ornaments in the Middle Paleolithic of North Africa.” PNAS 106 (38): 16051–16056.
d’Errico, F., Salomon, H., Vignaud, C. & Stringer, C. 2010. “Pigments from the Middle Palaeolithic levels of Es-Skhul (Mount Carmel, Israel).” Journal of Archaeological Science 37: 3099–3110.
d’Errico, F., Borgia, V. & Ronchitelli, M.-G. 2011. “Uluzzian bone technology and its implications for the origin of behavioural modernity.” Quaternary International. doi: 10.1016/ j.quaint.2011.03.039.
d’Errico, F. & Stringer, C. 2011. “Evolution, Revolution or Saltation scenario for the emergence of modern cultures?” Philosophical Transactions of the Royal Society B: Biological Sciences 366: 1060–1069.
Dansgaard, W., Johnsen, S.J., Clausen, H.B., Dahl-Jensen, D., Gundestrup, N.S., Hammer, C.U., Hvidberg, C.S., Steffensen, J.P., Sveinbjörnsdottir, A.E., Jouzel, J. & Bond, G. 1993. “Evidence for general instability of past climate from a 250 kyr ice core record.” Nature 364: 218–220.
Deacon, J. & Lancaster, N. 1988. Late Quaternary palaeoenvironments of southern Africa. Oxford: Clarendon Press.
Delagnes, A. & Meignen, L. 2005. “Diversity of lithic production systems during the Middle Paleolithic in France.” In Transitions before the transition evolution and stability in the Middle Paleolithic and Middle Stone Age, E. Hovers and S. L. Kuhn (eds), 85–107. USA: Springer.

68 Francesco d’Errico & Christopher S. Henshilwood
deMenocal, P.B. 2004. “African climate change and faunal evolution during the Pliocene- Pleistocene.” Earth and Planetary Science Letters 220: 3–24.
Diamond, J. 1978. “The Tasmanians: the longest isolation, the simplest technology.” Nature 273: 185–6.
Finlayson, C. 2004. Neanderthals and Modern Humans. An Ecological and Evolutionary Perspective. Cambridge: Cambridge University Press.
Finlayson, C. & Carrión, J.S. 2007. “Rapid ecological turnover and its impact on Neanderthal and other human populations.” Trends in Ecology and Evolution 22: 213–222.
Fletcher, W.J. & Sánchez-Goñi, M.F. 2008. “Orbital- and sub-orbital climate impacts on vegetation of the western Mediterranean basin over the last 48,000 yr.” Quaternary Science Reviews 70: 451–464.
Gamble, C., Davies, W., Pettitt, P. & Richards, M. 2004. “Climate change and evolving human diversity in Europe during the last glacial.” Philosophical Transactions of the Royal Society of London B359: 243–254.
Goren-Inbar, N. 1990. Quneitra: A Mousterian Site on the Golan Heights. Jerusalem: Qedem 31.Green, R.E., Krause, J., Briggs, A., Maricic, T., Stenzel, U., Kircher, M., Patterson, N., Weiwei
Zhai, H., Hsi-Yang Fritz, M., Hansen, N., Durand, E., Malaspinas, A., Jensen, J., Marques-Bonet, T., Alkan, C., Prüfer, K., Meyer, M., Burbano, H., Good, J., Schultz, R., Aximu-Petri, A., Butthof, A., Höber, B., Höffner, B., Siegemund, M., Weihmann, A., Nusbaum, C., Lander, E., Russ, C., Novod, N., Affourtit, J., Egholm, M., Verna, C., Rudan, P., Brajkovic, D., Kucan, Z., Gušic, I., Doronichev, V., Golovanova, L., Lalueza-Fox, C., Rasilla, M., Fortea, J., Rosas, A., Schmitz, R., Johnson, P., Eichler, E., Falush, D., Birney, E., Mullikin, J., Slatkin, M., Nielsen, R., Kelso, J., Lachmann, M., Reich, D. & Pääbo, S. 2010. “A draft sequence of the Neandertal genome.” Science 328 (5979): 710–722.
Grindley, J. 1969. “Quaternary marine palaeoecology in South Africa.” South African Archaeo-logical Bulletin, Quaternary Studies in South Africa 24 (95/96): 151–157.
Grün, R. & Stringer, C.B. 2000. “Tabun revisited: revised ESR chronology and new ESR and U-series analyses of dental material from Tabun C1.” Journal of Human Evolution 39 (6): 601–612.
Grün, R., Stringer, C., McDermott, F., Nathan, R., Porat, N., Robertson, S., Taylor, L., Mortimer, G., Eggins, S. & McCulloch, M. 2005. “U-series and ESR analyses of bones and teeth relat-ing to the human burials from Skhul.” Journal of Human Evolution 49: 316–334.
Heinrich, H. 1988. “Origin and consequences of cyclic ice rafting in the NE Atlantic Ocean during the past 130,000 years.” Quaternary Research 29: 142–152.
Henrich, J. 2004. “Demography and cultural evolution: How adaptive cultural processes can produce maladaptive losses – the Tasmanian Case.” American Antiquity 69 (2): 197–214.
Henshilwood, C.S. 2008. “Winds of change: palaeoenvironments, material culture and human behaviour in the Late Pleistocene (c. 77–48 ka) in the Western Cape Province, South Africa.” South African Archaeological Society Goodwin Series 10: 35–5.
Henshilwood, C.S., d’Errico, F., Marean, C., Milo, W. & Yates, R. 2001. “An early bone tool industry from the Middle Stone Age at Blombos Cave, South Africa: implications for the origins of modern human behaviour, symbolism and language.” Journal of Human Evolution 41: 631–678.
Henshilwood, C.S., d’Errico, F., Yates, R., Jacobs, Z., Tribolo, C., Duller, G.A.T., Mercier, N., Sealy, J.C., Valladas, H., Watts I. & Wintle, A.G. 2002. “Emergence of modern human behavior: Middle Stone Age engravings from South Africa.” Science 295: 1278–1280.
Henshilwood, C.S. & Marean, C.W. 2003. “The origin of modern human behaviour: a review and critique of models and test implications.” Current Anthropology 44: 627–65.

Chapter 3. The origin of symbolically mediated behaviour 69
Henshilwood, C.S., d’Errico, F., Vanhaeren, M., Van Niekerk, K. & Jacobs, Z. 2004. “Middle Stone Age shell beads from South Africa.” Science 304: 404.
Henshilwood, C.S. & Marean, C.W. 2006. “Remodelling the origins of modern human behaviour.” In The prehistory of Africa: tracing the lineage of modern man, H. Soodyall (ed.), 31–46. Johannesburg: Jonathan Ball Publishers.
Henshilwood, C.S., d’Errico, F. & Watts. I. 2009. “Engraved ochres from the Middle Stone Age levels at Blombos Cave.” South Africa Journal of Human Evolution 57: 27–47.
Henshilwood, C.S. & Dubreuil, B. 2009. “Reading the artefacts: Gleaning language skills from the Middle Stone Age in southern Africa.” In The Cradle of Language, R. Botha & C. Knight (eds), 41–60. Oxford: Oxford University Press.
Henshilwood, C.S. & Dubreuil, B., 2011. “The Still Bay and Howiesons Poort, 77–59 ka: Perspective-taking and the evolution of the modern human mind during the African Middle Stone Age.” Current Anthropology.
Henshilwood, C.S. & d’Errico, F. this volume. “Middle Stone Age engravings and their significance to the debate on the emergence of symbolic material culture.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood & F. d’Errico (eds) 75–96. Amsterdam: John Benjamins.
Hovers, E., Vandermeersh, B. & Bar-Yosef, O. 1997. “A Middle Palaeolithic engraved artefact from Qafzeh Cave, Israel.” Rock Art Research 14: 79–87.
Hovers, E., Ilani, S., Bar-Yosef, O. & Vandermeersch, B. 2003. “An Early Case of Color Symbolism. Ochre Use by Modern Humans in Qafzeh Cave.” Current Anthropology 44 (4): 492–522.
Hovers, E. & Belfer-Cohen, A. 2006. “Now you see it, now you don’t – modern human behavior in the Middle Paleolithic.” In Transitions before the transition: evolution and stability in the Middle Paleolithic and Middle Stone Age, E. Hovers & S. L. Kuhn (eds), 205–304. New York: Springer.
Imbrie, J. 1992. “On the structure and origin of major glaciation cycles, 1. Linear responses to Milankovitch forcing.” Paleoceanography 7: 701–738.
Imbrie, J. 1993. “On the structure and origin of major glaciation cycles, 2. The 100,000-year cycle.” Paleoceanography 8: 669–735.
Jacobs, Z., Duller, G.A.T., Wintle A. G. & Henshilwood. C. 2006. “Extending the chronology of deposits at Blombos Cave, South Africa, back to 140 ka using optical dating of single and multiple grains of quartz.” Journal of Human Evolution 51: 255–273.
Jacobs, Z. & Roberts, R.G. 2008. “Testing times: old and new chronologies for the Howieson’s Poort and Still Bay Industries in environmental context.” South African Archaeological S ociety Goodwin Series 10: 9–34.
Jouzel, J., Masson-Delmotte, V., Cattani, O., Dreyfus, G., Falourd, S., Hoffmann, G., Minster, B., Nouet, J., Barnola, J.M., Chappellaz, J., Fischer, H., Gallet, J.C., Johnsen, S., Leuenberger, M., Loulergue, L., Luethi, D., Oerter, H., Parrenin, F., Raisbeck, G., Raynaud, D., Schilt, A., Schwander, J., Selmo, E., Souchez, R., Spahni, R., Stauffer, B., Steffensen, J.P., Stenni, B., Stocker, T.F., Tison, J.L., Werner, M. & Wolff, E.W. 2007. “Orbital and millennial Antarctic climate variability over the past 800,000 years.” Science 317: 793–797.
Klein, R.G. 1999. The human career. 2nd edition. Chicago (IL): Chicago University Press.Klein, R. 2008. “Out of Africa and the evolution of human behaviour.” Evolutionary Anthropology
17: 267–281.Klein, R. & Edgar, B. 2003. The Dawn of Human Culture. New York: John Wiley & Sons.Knight, C., Powers, C. & Watts, I. 1995. “The human symbolic revolution: a Darwinian account.”
Cambridge Archaeological Journal 5 (1): 75–114.

70 Francesco d’Errico & Christopher S. Henshilwood
Lambeck, K. & Chappell, J. 2001. “Sea level change through the Last Glacial cycle.” Science 292: 679–686.
Leavesley, M.G. 2007. “A shark-tooth ornament from Pleistocene Sahul.” Antiquity 81: 308–315.Lombard, M. 2007. “The gripping nature of ochre: the association of ochre with Howiesons
Poort adhesives and Later Stone Age mastics from South Africa.” Journal of Human Evolution 53: 406–419.
Lorblanchet, M. 1999. La naissance de l’art. Paris: Errance.Mackay, A. & Welz, A. 2008. “Engraved ochre from a Middle Stone Age context at Klein Kliphuis
in the Western Cape of South Africa.” Journal of Archaeological Science 35: 1521–1532.Marean, C.W., Bar-Matthews, M., Fisher, E., Goldberg, P., Herries, A.I. R., Jacobs, Z., Jerardino,
A., Karkanas, P., Minichillo, T., Nilssen, P.J., Thompson, E., Watts I. & Williams, H.M. 2007. “Early human use of marine resources and pigment in South Africa during the Middle Pleistocene.” Nature 449 (7164): 905–908.
Marshack, A. 1981. “On Paleolithic ochre and the early uses of color and symbol.” Current Anthropology 22: 188–191.
Marshack, A. 1996. “A Middle Palaeolithic symbolic composition from the Golan Heights: the earliest known depictive image.” Current Anthropology 37: 357–365.
Mastronuzzi, G., Sansò, P., Murray-Wallace, C.V. & Shennan, I. 2005. “Quaternary coastal morphology and sea-level changes – an introduction.” Quaternary Science Reviews 24 (18–19): 1963–1968.
McBrearty, S. & Brooks, A.S. 2000. “The revolution that wasn’t: a new interpretation of the origin of modern human behavior.” Journal of Human Evolution 39: 453–563.
McGrew, W.C. this volume. “Pan Symbolicus: A Cultural Primatologist’s Viewpoint” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds) 1–12. Amsterdam: John Benjamins.
Mellars, P. 1996. The Neandertal legacy: an archaeological perspective from western Europe. Princeton (NJ): Princeton University Press.
Mellars, P. 2006. “Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model.” Proceedings of the National Academy of Sciences 103: 9381–9386.
Mellars, P. 2007. “Rethinking the revolution: Eurasian and African perspectives” In Rethinking the revolution: new behavioural and biological perspectives on the origin and dispersal of modern humans, P. Mellars, K. Boyle, O. Bar-Yosef and C. Stringer (eds), 1–14. Cambridge: McDonald Institute Monographs.
Mercier, N., Valladas, H., Froget, L., Joron, J.L. & Ronen, A. 2000. “Datation par la thermolumi-nescence de la base du gisement paléolithique de Tabun (Mont Carmel, Israël).” Comptes Rendus de l’Académie des Sciences 330: 731–738.
Milankovitch, M.K. 1969. Canon of insolation and the Ice Age problem. Serbian Academy Beorg. Special Publication 132.
Millard, A.R. 2006. “Bayesian analysis of ESR dates, with application to Border Cave.” Quaternary Geochronology 1 (2): 159–166.
Morse, K. 1993. “Shell beads from Mandu Mandu Creek rock-shelter, Cape Range peninsula, Western Australia, dated before 30,000 ka BP.” Antiquity 67: 877–883.
Partridge, T.C. 1993. “Warming phases in Southern Africa during the last 150,000 years – an overview.” Palaeogeography, Palaeoclimatology, Palaeoecology l01: 237–244.
Partridge, T.C. 1997. “Cainozoic environmental change in southern Africa, with special empha-sis on the last 200 000 years.” Progress in Physical Geography 21 (1): 3–22.
Pearson, M.P. 1999. The Archaeology of Death and Burial. Phoenix Mill: Sutton.

Chapter 3. The origin of symbolically mediated behaviour 71
Potts, R. 1996. Humanity’s descent: the consequences of ecological instability. New York: William Morrow and Co.
Powell, A., Shennan, S. & Thomas, M. 2009. “Late Pleistocene demography and the appearance of modern human behavior.” Science 324 (5932): 1298–1301.
Premo, L. & Kuhn S. 2010. “Modeling Effects of Local Extinctions on Culture Change and Diversity in the Paleolithic.” PLoS ONE 5 (12): 15582. doi:10.1371/ journal.pone.0015582.
Rampino, M.R. & Self, S. 1993. “Climate – Volcanism feedback and the Toba eruption of ca. 74000 years ago.” Quaternary Research 40: 269–280.
Richerson, P., Bettinger, R. & Boyd, R. 2005. “Evolution on a Restless Planet: Were Environmental Variability and Environmental Change Major Drivers of Human Evolution?” In Handbook of Evolution, Vol. 2: The Evolution of Living Systems (Including Hominids), F. Wuketits and F. Ayala (eds), 223–242. Weinheim: Wiley.
Richerson, P., Boyd, R. & Bettinger, R. 2009. “Cultural Innovations and Demographic Change.” Human Biology 81 (2–3): 211–235.
Rigaud, J.-P., Texier, P.-J., Parkington, J. & Poggenpoel, C. 2006. “Le mobilier Stillbay et Howiesons Poort de l’abri Diepkloof: la chronologie du Middle Stone Age Sud-Africain et ses implications.” Comptes Rendus, Palevol 5: 839–849.
Rivers, W. 1912. “The disappearance of useful arts.” Festschrift Tillagnad Edvard Westermarck, Helsingfors 117–130.
Roberts, D.L., Bateman, M.D., Murray-Wallace, C.V., Carr, A.S. & Holmes, P.J. 2008. “Last inter-glacial fossil elephant trackways dated by OSL/AAR in coastal aeolianites, Still Bay, South Africa.” Palaeogeography, Palaeoclimatology, Palaeoecology 257 (3): 261–279.
Ronchitelli, A., Boscato, P. & Gambassini, P. 2009. “Gli ultimi neandertaliani in Italia: aspetti culturali.” In La lunga storia di neandertal. Biologia e comportamento, F. Facchini & M. Belcastro (eds), 257–287. Milano: Jaca Book.
Savage-Rumbaugh, E.S. & Fields, W.M. this volume. “The Evolution and the Rise of Human Language.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds) 13-47. Amsterdam: John Benjamins.
Salomon, H. 2009. Les matières colorantes au début du Paléolithique supérieur: sources, transformations et functions. Thesis (Ph.D.). University of Bordeaux.
Sanchez-Goñi, M.F., Landais, A., Fletcher, W.J., Naughton, F., Desprat, S. & Duprat, J. 2008. “Contrasting impacts of Dansgaard-Oeschger events over a western European latitudinal transect modulated by orbital parameters.” Quaternary Science Reviews 27: 1136–1151.
Scholz, C.A., Johnson, T.C., Cohen, A.S., King, J.W., Peck, J.A., Overpeck, J.T., Talbot, M.R., Brown, E.T., Kalindekafe, L., Amoako, P.Y., Lyons, R.P., Shanahan, T.M., Castañeda, I.S., Heil, C.W., Forman, S.L., McHargue, L.R., Beuning, K.R., Gomez, J. & Pierson, J. 2007. East African megadroughts between 135 and 75 thousand years ago and bearing on early-modern human origins. Proceedings of the National Academy of Sciences 104: 16416–16421.
Sepulchre, P., Ramstein, G., Fluteau, F., Schuster, M., Tiercelin, J-J. & Brunet, M. 2006. “Tectonic Uplift and Eastern Africa aridification.” Science 313 (5792): 1419–1423.
Sepulchre, P., Ramstein, G., Kageyama, M., Vanhaeren, M., Krinner, G., Sánchez-Goñi, M.F. & d’Errico, F. 2007. H4 abrupt event and late Neanderthal presence in Iberia. Earth and Planetary Science Letters 258: 283–292.
Shennan, S. 2001. “Demography and cultural innovation: a model and its implications for the emergence of modern human culture.” Cambridge Archaeological Journal 11: 5–16.

72 Francesco d’Errico & Christopher S. Henshilwood
Soressi, M. & d’Errico, F. 2007. “Pigments, gravures, parures: les comportements symboliques controversés des Néandertaliens.” In Les Néandertaliens. Biologie et cultures, B. Vandermeersch and B. Maureille (eds), 297–309. Paris: Éditions du CTHS.
Stringer, C. 2006. Homo Britannicus: The incredible story of human life in Britain. London: Penguin.
Texier, J.-P., Porraz, G., Parkington, J., Rigaud, J.-P., Poggenpoel, C., Miller, C., Tribolo, C., Cartwright, C., Coudenneau, A., Klein, R., Steele, T. & Vernai. C. 2010. A Howiesons Poort tradition of engraving ostrich eggshell containers dated to 60,000 years ago at Diepkloof Rock Shelter, South Africa. Proceedings of the National Academy of Sciences doi: 10.1073/pnas.0913047107.
Thackeray, J.F. 1992. “Chronology of Late Pleistocene deposits associated with Homo sapiens at Klasies River Mouth, South Africa.” Palaeoecology of Africa 23: 177–191.
Thackeray, J.F. 2007. Sea levels and chronology of Late Pleistocene coastal cave deposits at Klasies River in South Africa. Annals of the Transvaal Museum 44: 219–220.
Van Andel, T.H. 1989. “Late Pleistocene sea levels and the human exploitation of the shore and shelf of southern South Africa.” Journal of Field Archaeology 16: 133–154.
Van Andel, T.H. & Davies, W. (eds). 2003. Neanderthals and modern humans in the European landscape during the last glaciation: archaeological results of the Stage 3 Project. Cambridge: McDonald Institute for Archaeological Research, University of Cambridge.
Vandermeersch, B., Cleyet-Merle, J.-J., Jaubert, J., Maureille, B. & Turc, A. 2008. Première humanité, gestes funéraires des Néandertaliens. Paris: Réunion des Musées Nationaux.
Vanhaeren, M. & d’Errico, F. 2006. “Clinal distribution of personal ornaments reveals the ethno-linguistic geography of Early Upper Palaeolithic Europe.” Journal of Archaeological Science 33 (8): 1105–1128.
Vanhaeren, M., d’Errico, F., Stringer, C., James, S.L., Todd, J. & Mienis, H.K. 2006. “Middle Palaeolithic shell beads in Israel and Algeria.” Science 312 (5781): 1785–1788.
Velo, J. 1984. “Ochre as medicine: a suggestion for the interpretation of the archaeological record.” Current Anthropology 25 (5): 674.
Vermeersch, P.M., Gijselings, G. & Paulissen, E. 1984. “Discovery of the Nazlet Khater man, Upper Egypt.” Journal of Human Evolution 13: 281–286.
Vermeersch, P.M., Paulissen, E., Stokes, S., Charlier, C., Van Peer, P., Stringer, C.B. & Lislay, W. 1998. “A Middle Palaeolithic burial of a modern human at Taramsa Hill, Egypt.” Antiquity 72: 475–484.
Villa, P., Soressi, M., Henshilwood, C.S. & Mourre, V. 2009. “The Still Bay points of Blombos Cave (South Africa).” Journal of Archaeological Science 36 (2): 441–460.
Wadley, L. 2001. “What is cultural modernity? A general view and a South African perspective from Rose Cottage.” Cambridge Archaeological Journal 11 (2): 201–221.
Wadley, L., Williamson, B.S. & Lombard, M. 2004. “Ochre in hafting in Middle Stone Age southern Africa: a practical role.” Antiquity 78: 661–675.
Wadley, L., Hodgskiss, T. & Grant, M. 2009. Implications for complex cognition from the h afting of tools with compound adhesives in the Middle Stone Age, South Africa. Proceedings of the National Academy of Sciences 106: 9590–9594.
Watts, I. 1999. “The origin of symbolic culture.” In The evolution of culture, R. Dunbar, C. Knight and C. Power (eds), 113–146. Edinburgh: Edinburgh University Press.
Watts, I. 2002. “Ochre in the Middle Stone Age of southern Africa: ritualized display or hide preservative?” South African Archaeological Bulletin 57: 1–14.
Wendt, W.E. 1976. “‘Art mobilier’ from the Apollo 11 Cave, South West Africa: Africa’s oldest dated works of art.” South African Archaeological Bulletin 31: 1170–1199.

Chapter 3. The origin of symbolically mediated behaviour 73
Wiessner, P. 1983. “Style & social information in Kalahari San projectile points.” American Antiquity 48: 253–276.
Yellen, J.E., Brooks, A.S., Cornelissen, E., Mehlman, M. & Stewart, K. 1995. “A Middle Stone Age worked bone Industry from Katanda, Upper Semliki Valley, Zaire.” Science 268: 553–556.
Zilhão, J. & d’Errico, F. 1999. “The chronology and taphonomy of the earliest Aurignacian and its implications for the understanding of Neandertal extinction.” Journal of World Prehistory 13 (1): 1–68.
Zilhão, J., Angelucci, D., Badal-Garcíac, E., d’Errico, F., Daniel, F., Dayet, L., Douka, K., Higham, T., Martínez-Sánchez, M., Montes-Bernárdez, R., Murcia-Mascarósj, S., Pérez-Sirventh, C., Roldán-Garcíaj, C., Vanhaeren, M., Villaverde, V., Wood, R. & Zapata, J. 2010. “Symbolic use of marine shells and mineral pigments by Iberian Neandertals.” PNAS 107 (3): 1023–1028.


chapter 4
Middle Stone Age engravings and their significance to the debate on the emergence of symbolic material culture
Christopher S. Henshilwooda,b & Francesco d’Erricoc,a
aInstitute for Archaeology, History, Culture and Religion, University of Bergen/bInstitute for Human Evolution, University of the Witwatersrand/CCNRS-UMR 5199 PACEA, Université de Bordeaux
Archaeological evidence associated with modern cognitive abilities provides important insights into when and where modern human behaviour emerged. Modern human behaviour here means the thoughts and actions spontaneously shaped by minds equivalent to those of Homo sapiens today. Key among these is the use of symbols. Three models for the origins of behavioural modernity are current: (i) a late and rapid appearance at ~ 40–50 ka associated with the European Upper Palaeolithic and the Later Stone Age (LSA) of sub-Saharan Africa, (ii) an earlier and more gradual evolution rooted in the African Middle Stone Age (MSA ~ 300–40 ka), (iii) a discontinuous evolution rooted in both the African Middle Stone Age and the Mousterian of Eurasia. Material evidence for modern behaviour before 40 ka in Africa was, until a few years ago, relatively rare and often regarded as ambiguous compared to evidence from the Upper Palaeolithic or LSA. However, in sub-Saharan Africa archaeological evidence for changes in technology, economy and social organization and the emergence of symbolism in the Middle Stone Age contradicts the first model. Examples of these changes include standardized formal lithic tools, shaped bone implements, innovative subsistence strategies, evidence for personal ornaments and the deliberate engraving of abstract designs on ochre, ostrich eggshell and bone. In this chapter we review the earliest evidence for purposely made engravings from southern Africa and discuss their significance for arguments favouring early advances in human cognition.
1. Introduction
In past decades, anthropologists, archaeologists, linguists, and cognitive scien-tists assumed that the production of symbolic material culture, and its presumed involvement in the mediation of social behaviour, was a recent innovation in human history and the probable outcome of a stochastic event that took place after c. 40 ka. Instances of non-utilitarian behaviours older than 40 ka from Neandertal

76 Christopher S. Henshilwood & Francesco d’Errico
sites in Eurasia, and Homo sapiens sites in Africa were considered ambiguous with respect to their significance and/or their anthropogenic nature. Some archaeolo-gists now argue that ‘symbolically mediated behaviour’ (for definition see Wadley 2001; Henshilwood & Marean 2003, 2006; Henshilwood & Dubreuil 2009) evolved gradually in Africa, and link its origins with the emergence of Homo sapiens in that continent (e.g. Knight et al. 1995; Watts 1999, 2009; McBrearty & Brooks 2000; Henshilwood et al. 2002, 2004, 2009; Henshilwood & Dubreuil, 2011) define symbolically mediated culture as “one in which individuals understand that arte-facts are imbued with meaning and that these meanings are construed and depend on collectively shared beliefs. This criterion is crucial. It explains how human norms and conventions differ from the ritualized behaviours found in nonhuman primates”. The tenets of the ‘gradual emergence in Africa model’ are that the appearance of modern cognitive behaviours in Eurasia, associated with Homo sapiens, is the end result of an ‘out of Africa’ dispersal of an already symbolic species at c. 60 ka or earlier (e.g. Henshilwood & Marean 2003, 2006; Forster 2004; Mellars 2006). Other authors argue that Neandertal behaviour may have independently evolved along similar lines and favour demographic events triggered by climate change instead of a speciation event as the main factor accounting for the emergence of symbolic mediated behaviours in Africa and Eurasia (Zilhão 2001; d’Errico 2003; d’Errico et al. 2003; see d’Errico & Henshilwood, this volume).
New evidence for symbolic behaviour from MSA sites in Africa and Mousterian sites in Europe and the Near East is challenging these models. The evidence for the archaeological expression of symbolically mediated behaviours now includes geometric and iconographic representations, elaborate burials, and personal ornaments and, to some extent, pigment use and complex bone and stone tools (see d’Errico & Henshilwood, this volume for a review of the evidence). In this chapter we focus on the earliest evidence for deliberate engravings from southern Africa, explore their symbolic nature, and discuss their significance for the debate on the origins of cultural modernity. Before presenting the archaeo-logical evidence and its context, we address the question of why engravings are important for the symbol related debate and what information they may yield that is relevant to this debate.
In many respects engravings represent a special category of material culture. Pigments (colloquially called ochre by most archaeologists in Africa) found at archaeological sites are generally the by-product of a sequence of actions but recreating these sequences is problematic. It is one of the main reasons for the interpretation of pigments being a controversial topic. Different actions, for exam-ple grinding pigment, that were made with or without symbolic intent may well produce a similar end product. Personal ornaments found at archaeological sites, for example shell beads, could be described as the disposed elements of a string

Chapter 4. Middle Stone Age engravings and their significance 77
of signs that once was meaningful. How these ornaments were worn or arranged on the human body is now irretrievable. Perishable ornaments, perhaps in wood or on leather, have not survived to help us with our reconstruction of past mate-rial culture. Burials, virtually unknown for the MSA in Africa, may help with the reconstruction of funerary practices in the past (Duday 2009) but we know from ethnographic accounts of burials that archaeologists are only able to retrieve a fraction of the whole picture from site remains. Technological complexity and morphological standardisation in stone and bone tool production help with assessing the cognitive abilities of our ancestors, but again disentangling stylistic and symbolic elements from purely functional actions is often difficult.
An engraving is the end product of incising or pecking a design onto a material, such as pigment or stone with the aid of a tool. Engravings are perhaps the only category of potentially symbolic early material culture that still reflects the complete set of cutting actions performed by the artist. As is the case with drawing, engraving reflects deep unconscious feelings of self (Freeman & Cox 1985; Thomas & Silk 1990) while, at the same time, engravings “organise” a shared visual culture (Cox et al. 2001). Contrary to other early symbolic manifestations, engravings can be formally described, compared and their differences measured from a variety of perspectives (Bosinski et al. 2001). The microscopic analysis of the lines that make up engraved patterns, including junctions and crossings, provide information on the chronology of gestures employed, the use of a single vs multiple tools, engraver laterality, and possible changes in the orientation of the engraved surface during the production of incisions. It can also indicate the handedness and relative location of the engraver to the piece at the time of the incisions. Experimental reproduction of these processes helps to understand these past actions in fine detail (d’Errico 1996; Fritz 1999). By combining those data it is possible to evaluate the technical competence of the engraver, and, to some extent, to establish if the engraver followed a mental template during production.
A brief summary of the evidence for engravings in the MSA shows that overall, only a few isolated pieces are reported and that very few sites have more than one or two examples. An engraved stone with a cross-hatched pattern was recovered from presumed Howiesons Poort levels at the Kleinkliphuis River site, Western Cape (Mackay and Welz, 2008; Figure 1a); in 1963 an engraved pebble was found by Wolfgang Sydow at a site containing only MSA lithics near Palmenhorst, Namibia (Wendt 1975; Figure 1b). Parallel line engravings on stone are reported from Cape Hangklip (Heese n.d.; Figure 1c) and from an undated MSA context at Bushman Rock Shelter, Mpumulanga (Watts 1998). Dendritic/convergent line motifs were found in MSA II and Howiesons Poort (HP) contexts at Klasies, Eastern Cape (Knight et al. 1995; Watts 1998) and from c. 100 ka levels

78 Christopher S. Henshilwood & Francesco d’Errico
at Pinnacle Point, Western Cape (Watts, 2010). Similar parallel lines engraved on bone fragments are found at Klasies and at Blombos Cave (BBC) (d’Errico & Henshilwood 2007). A striking collection of engraved items comes from BBC and Diepkloof Rock Shelter (Figures 2, 3, 4 and 5). At the former site at least 14 pieces of ochre engraved with deliberate abstract patterns come from c. 75 ka and 100 ka levels (Henshilwood et al. 2002, 2009). At the latter site 270 engraved ostrich eggshell fragments were found in HP levels with an age of c. 60 ka (Texier et al. 2010). In this chapter we focus on these two sites.
a b c
scale=1cm
Figure 1. (a) engraved pebble from reportedly Howiesons Poort levels at Kleinkliphuis River, Western Cape, South Africa (after Mackay & Welz, 2008); (b) decorated pebble from MSA site near Palmenhorst, Namibia recovered by Wolfgang Sydow in 1963; (c) worked ochre pencil from the Still Bay site at Cape Hangklip, Western Cape described by Frans Malan (after Heese n.d., papers at the Iziko-South African Museum. The location of this artefact is currently unknown)
2. Howiesons Poort techno-tradition
2.1 Diepkloof Rock Shelter
2.1.1 Archaeological context and datingDiepkloof Rock Shelter (DRS) is a quartzitic sandstone shelter located 180 km north of Cape Town and 18 km from the Atlantic (Figure 2c). It overlooks the Verlorenvlei River. The site was excavated in the 1990s by a team from the University of Cape Town. A Franco-South African team have excavated the site since 1998. The DRS stratigraphic sequence consists of laminations of anthropo-genic material, mostly composed of burnt and non-burnt organic residues, ash associated with detrital sands and diagenetic components, and rich faunal and lithic assemblages (Texier et al. 2010; Figure 2b). The 2.5 m deep MSA sequence is classified, from top to bottom, as Post-Howiesons Poort, Howiesons Poort (HP), Still Bay (SB), and Pre-Still Bay. Thermoluminescence (TL) and optically

Chapter 4. Middle Stone Age engravings and their significance 79
Figu
re 2
. (a
) Blo
mbo
s Cav
e st
ratig
raph
y an
d ag
es; (
b) D
iepk
loof
Roc
k Sh
elte
r str
atig
raph
y; (c
) loc
atio
n of
Blo
mbo
s Cav
e (B
BC) a
nd D
iepk
loof
Roc
k Sh
elte
r (D
RS)
290
BP
2 ka
LSA M
SAph
ases
M1
c. 7
2 ka
M2
uppe
rc.
78
ka
M2
low
erc.
85
ka
M3
uppe
rc.
100
ka
Hia
tus
70 k
aD
UN CA CB CC CCC
CDCD
A
CFA
CFB/
CFC
CGA
ACG
AB
CGA
C
CH CI CJ
50 c
m
a
Blom
bos
Cave
b
Die
pklo
of R
ock
Shel
ter
30K6 50 60 70 80 90 100
110
120
130
140
150
160
170
180
50 60 70 80 90 100
110
120
130
140
150
160
170
180
4050
6070
8090
100
L6M
611
012
013
014
015
016
017
018
019
020
0
E O E S Z O N E
HP
P O S T H O W I E S O N S P O O R Tc
LSA
Clau
de
Dar
ryl
Deo
n
Este
rEs
ter
Eve
Fran
sFr
edFr
ank
Fox
Fox
Fion
aFi
ona
Je�
Je�
Joy
Gov
erno
r
John
Jack

80 Christopher S. Henshilwood & Francesco d’Errico
stimulated luminescence (OSL) techniques provide an age for these sequences that range from c. 130 ka to 45 ka.
TL dates of 74–60 ka (Parkington 1999) are provided for the HP at DRS but it was thought these dates may be too old (Rigaud et al. 2006). A subsequent set of ages for the HP levels, calculated by the TL method, range from c. 96 ka – 60 ka (Tribolo et al. 2009). These are not concordant with Jacobs et al.’s (2008) OSL ages of 58.1 ± 1.9 ka to 63.3 ± 2.2 ka (also Jacobs et al. 2008). The reason for the differ-ences in the ages reported by Jacobs et al. (2008) and Tribolo et al. (2009) has not been resolved. In summary, the most consistent ages for HP in southern Africa sites fall within the range of 65–59 ka.
3. Diepkloof engraved ostrich eggshells
Ostrich eggshell fragments are found in all the MSA levels although ostrich bones are absent, despite excellent preservation of faunal remains. Ostrich eggs are a regular dietary item at Diepkloof, as is the case at most other MSA sites in southern Africa (e.g. Henshilwood et al. 2001a; Klein et al. 2004). About 270 ostrich eggshell fragments from the upper levels of the HP (c. 60 ka) are engraved with abstract patterns (e.g. Figure 3) and three fragments have punctured open-ings on the apical part of the egg (e.g. Figure 3a). One of these has engraved lines that diverge from the aperture. The fragmentation of the eggshells into pieces 20–30 mm in size is attributed to post depositional processes. Reconstructing the original decoration on a whole egg is thus problematic but four distinct motifs, across the range of eggs, have been identified (Figure 3): (i) two parallel lines intersected at right angles by regularly spaced lines forming ladder-like patterns (Figure 3 b,c,d,e,f); (ii) a series of deeply engraved, straight, sub-parallel lines(Figure 3 g,h); (iii) slightly curved lines crossing a central line (Figure 3i); (iv) cross-hatched lines (Figure 3a).
The first of these motifs is recorded on 69 fragments that come only from the lower HP levels. The second motif is most common in the upper levels and the third and fourth motifs are present on one piece each. The third motif comes from a level in the middle of the HP and the provenance of the fourth motif is not reported. (Texier et al. 2010). Microscopic analysis of the crossing of the lines on the ladder motif pieces suggests to Texier et al. (2010) that the production of this pattern was standardised as the engraving of the long parallel lines was done first, then the shorter lines were made that cross over the longer lines. There is considerable variability in the spacing between the hatched lines and the angles of intersection.

Chapter 4. Middle Stone Age engravings and their significance 81
a b c
d e f
g h i
1 cm
Figure 3. Engraved ostrich eggshell from the c. 60 ka Howiesons Poort levels at Diepkloof Rock Shelter, southern Cape
Texier et al. (2010) compared the HP evidence for ostrich eggshell use with that of historical hunter-gatherers, mainly Khoesan, who used the eggs as flasks to store and transport water. Many of these were also engraved. The conclusion of the team’s study is that the engraved fragments from the HP levels probably came from eggs used as storage containers. Considering the diversity of motifs, their stylistic variability and the numbers of fragments, the authors suggest that the eggshell pieces came from at least 25 whole eggs. They argue that the signature of the designs on the shells is different to those on engraved ochre and bones from other MSA sites; specifically that the motifs served no technical purpose; that they are standardized and systematized; and that this

82 Christopher S. Henshilwood & Francesco d’Errico
favoured the development of specific geometric rules. The authors state further that as the engraved pieces come from 18 stratigraphic levels and as there are changes in engraved patterns across these levels, that the engraving of eggs can be interpreted as evidence for a tradition that lasted for several thousand years. They conclude that these engraved eggs provide compelling evidence for communication using symbols.
4. Still Bay techno-tradition
4.1 Blombos Cave
4.1.1 Archaeological context and datingBlombos Cave is situated on the southern Cape coast, 300 km east of Cape Town (Figure 2c). The MSA levels at the site are divided into three phases M1, M2 and M3 (Henshilwood et al. 2001a) (Figure 2a). The five uppermost layers below a sterile level named BBC Hiatus are assigned to the M1 phase. The M1 and Upper M2 phase lithics are typified by Still Bay type bifacial foliate points, the fossile directeur of the SB Industry (Goodwin and van Riet Lowe 1929; Villa et al. 2009) and end- and side- scrapers (Henshilwood et al. 2001a). Engraved ochre and bone (d’Errico et al. 2001; Henshilwood et al. 2002, 2009; Watts 2009), formal bone tools (Henshilwood et al. 2001b) and forty nine Nassarius shell beads (Henshilwood et al. 2004; d‘Errico et al. 2005) come from this phase. The lower M2 phase is typified by carbonised deposits, and includes shellfish and fauna, but these layers are generally low intensity deposits. High density shellfish deposits, hearths, faunal remains, many lithics and ochre pieces dominate most levels in the M3 phase.
The SB levels have been dated using a number of methods (Jones 2001; Jacobs et al. 2003a,b, 2006; Tribolo et al. 2006). The Hiatus level composed of undisturbed aeolian sand above the M1 phase is dated by OSL to 69 ± 5 ka and 70 ± 5 ka (Henshilwood et al. 2002: Jacobs et al. 2003 a,b; Jacobs et al. 2006) (Figure 2a) and provides a minimum age for the SB deposits at the site. An OSL age of 72.7 ±3.1 ka was obtained for the upper part of the SB M1 phase (Jacobs et al. 2003 a,b). Thermoluminescence (TL) dates for the M1 and Upper M2 phase indicate that 74 ± 5 and 78 ± 6 ka are the likely ages for these SB levels (Tribolo et al. 2006). OSL dates for the lower M2 phase fall between 84.6 ± 5.8 ka to 76.8 ± 3.1 ka (Jacobs et al. 2006) (Figure 2). Similar ages were obtained using the electron spin resonance method (Jones 2001). The lower M2 phase (CG levels) does not contain artefacts associated with the SB. The upper M2 phase does contain these SB markers and the inference drawn is that the age of 76.8 ± 3.1 for the CF level (Jacobs et al. 2006) (Figure 2a) should be regarded as the terminus

Chapter 4. Middle Stone Age engravings and their significance 83
post quem for the SB levels at BBC. Using single aliquots, an OSL age of 98.9 ± 5.5 ka was obtained for the upper portion of level CI, representing the upper M3 phase. The level below, CJ, has provided a date of 143.2 ± 5.5 ka by the same method (Jacobs et al. 2006); however, this came from a sub-unit of sterile sand within CJ and cannot, therefore, be treated as a reliable date for the archaeology. The stratigraphic integrity of artefacts recovered from these levels has been demonstrated and there is minimal evidence for movement of artefacts between the MSA phases (Jacobs et al. 2003 a, b; Henshilwood 2006; Henshilwood et al. 2009).
5. Blombos engraved ochre
In total 1,534 pieces of ochre ≥10mm in length with a net weight of 5,581 g have been recovered from the MSA levels at BBC (Henshilwood et al. 2009). Most of the ochre comes from the M3 phase (78.6% of pieces, 82% of mass) and over-all the quantity of ochre from BBC is considerably greater than that recorded for most southern African MSA sites (Henshilwood et al. 2001a, 2009; Watts 2009). Within an approximately fifty kilometre radius of BBC, the most likely sources of ochre are outcrops of Bokkeveld Group deposits (predominantly comprised of shale and siltstone) (Henshilwood et al. 2001a; Watts 2009). At present the nearest such outcrops are adjacent to the Goukou and Duiwenhoks Rivers (respectively 19 km east and 21 km west of BBC), but even a small drop in sea level (as occurred following the interglacial transgression of MIS 5e), may have exposed Bokkeveld shales, now buried under marine deposits and aeolian sands, within c. 3–5 km of the site (Watts 2009; see also Rogers 1988: 411).
Modification of ochre was primarily directed at producing a powder, either by grinding pieces across an abrasive surface or by scraping them using a stone tool. Grinding results in flat or slightly convex facets covered by multiple fusiform striations (d’Errico & Nowell 2000; d’Errico & Backwell 2003). Scraping produces parallel striations of different width and depth resulting from projections on the working surface of the implement. Repeated applications of the scraping tool, in the same area, result in wide grooved or concave areas with fringed ends corresponding to individual exit points.
Some pieces recovered at BBC bear striations inconsistent with a natural, incidental or “technical” interpretation and, we have recently argued (Henshilwood et al. 2009), represent instances of unequivocally intentional engraving (Figures 4 and 5; Table 1). None of the grooves present on the BBC pieces fit the criteria for geological or animal markings. Incidental markings left by trampling, common on the engraved ostrich eggshells from Diepkloof, result in randomly oriented

84 Christopher S. Henshilwood & Francesco d’Errico
straight or slightly curved individual striations and smoothing of the exposed areas. The incised BBC ochres are well preserved and show no evidence of taphonomic damage including striations indicative of natural abrasion. The incisions cannot be interpreted as the outcome of cutting motions. A flat stone used as a base on which to cut soft material will need to be of a size compatible with the task (Binford 1978). This requirement eliminates the likelihood that the pieces of ochre from BBC could have been used as cutting-boards. Lines left on cutting-boards are generally straight, sub-parallel, overlapping, and oriented on the long axis of the board, a feature not recorded at Blombos. Microscopic analysis of the BBC incisions shows that they were made by the point of a stone tool and not by a cutting edge (d’Errico 1995; Fritz 1999; d’Errico et al. 2001; Alvarez et al. 2001). The tip of a stone tool is not effective for repeated cutting as it blunts rapidly, yet it is effective to precisely engrave lines. Tool sharpening does not account for any of the incisions on the ochre pieces as the knapped stone tools from the MSA layers of BBC were not sharpened by grinding. The small size of the incisions on the ochre pieces indicates that they are not compatible with arrow/spear shaft straightening. Notched bones used to scrape soft material such as vegetable or hide are found in the MSA levels at Klasies River (d’Errico & Henshilwood 2007). However, none of the incised ochre pieces shows a wear pattern consistent with this kind of use. If ochre pieces were deeply incised or sawn to assist in splitting them there is no evidence of these techniques having been applied as the recorded lines are too shallow for this purpose.
Testing an ochre piece for powder quality or shade only requires one or two strokes, however while multiple lines are present on most of the incised BBC ochres. Further, in a number of cases incisions were made on a ground surface, making further testing unnecessary. To obtain a useable quantity of powder from an ochre piece requires grinding across an abrasive surface or scraping it repeatedly with a tool. On some of the pieces examined there is evidence of these modifications (e.g. M1/1, M1/3, M1/5, M1/6, M3/2, M3/6, M3/9, M3/3 and M3/8 conjoins: Figures 4 and 5), but these are not consistent with the production of ochre powder. Many incisions are narrow and superficial and would have resulted only in a tiny quantity of powder being produced. In some cases (M1/4, M3/1, M3/10: Figures 4 and 5) these narrow incisions are the only modification visible on the surface of the ochre, which contradict this interpretation. Further, many of the lines interpreted as deliberate engravings (M1/5, M1/6, M3/1, M3/6, M3/10: Figures 4 and 5) are different to those made when scraping ochre for powder extraction. The latter typically consists of a single wide deep groove with fringed ends, created by a relatively unrestrained back and forth motion, or multiple sub-parallel deep grooves produced by repeated motions in one direction within the same plane.

Chapter 4. Middle Stone Age engravings and their significance 85
a
1 cm
1 cm
1 cm
1 cm
1 cm
1 cm
b
c
d
e
f
Figure 4. Engraved ochre from the M1 phase (78–72 ka) at Blombos Cave

86 Christopher S. Henshilwood & Francesco d’Errico
a b
c d
g h
1 cm
1 cm
1 cm
1 cm
1 cm
e
f
Figure 5. Engraved ochre from the M2 and M3 phases (c. 85–100 ka) at Blombos Cave
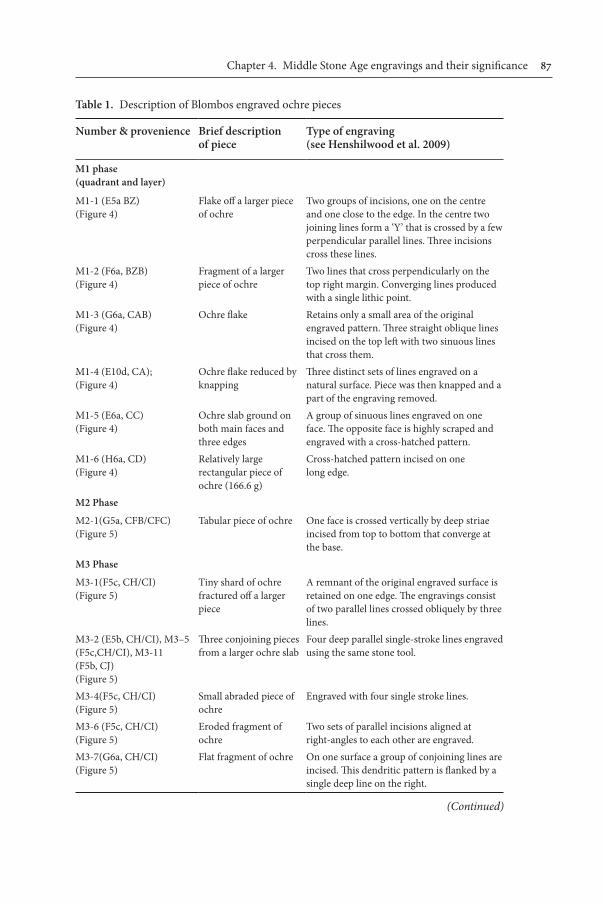
Chapter 4. Middle Stone Age engravings and their significance 87
Table 1. Description of Blombos engraved ochre pieces
Number & provenience Brief description of piece
Type of engraving (see Henshilwood et al. 2009)
M1 phase(quadrant and layer)
M1-1 (E5a BZ)(Figure 4)
Flake off a larger piece of ochre
Two groups of incisions, one on the centre and one close to the edge. In the centre two joining lines form a ‘Y’ that is crossed by a few perpendicular parallel lines. Three incisions cross these lines.
M1-2 (F6a, BZB)(Figure 4)
Fragment of a larger piece of ochre
Two lines that cross perpendicularly on the top right margin. Converging lines produced with a single lithic point.
M1-3 (G6a, CAB)(Figure 4)
Ochre flake Retains only a small area of the original engraved pattern. Three straight oblique lines incised on the top left with two sinuous lines that cross them.
M1-4 (E10d, CA);(Figure 4)
Ochre flake reduced by knapping
Three distinct sets of lines engraved on a natural surface. Piece was then knapped and a part of the engraving removed.
M1-5 (E6a, CC)(Figure 4)
Ochre slab ground on both main faces and three edges
A group of sinuous lines engraved on one face. The opposite face is highly scraped and engraved with a cross-hatched pattern.
M1-6 (H6a, CD)(Figure 4)
Relatively large rectangular piece of ochre (166.6 g)
Cross-hatched pattern incised on one long edge.
M2 Phase
M2-1(G5a, CFB/CFC)(Figure 5)
Tabular piece of ochre One face is crossed vertically by deep striae incised from top to bottom that converge at the base.
M3 Phase
M3-1(F5c, CH/CI)(Figure 5)
Tiny shard of ochre fractured off a larger piece
A remnant of the original engraved surface is retained on one edge. The engravings consist of two parallel lines crossed obliquely by three lines.
M3-2 (E5b, CH/CI), M3–5 (F5c,CH/CI), M3-11 (F5b, CJ)(Figure 5)
Three conjoining pieces from a larger ochre slab
Four deep parallel single-stroke lines engraved using the same stone tool.
M3-4(F5c, CH/CI)(Figure 5)
Small abraded piece of ochre
Engraved with four single stroke lines.
M3-6 (F5c, CH/CI)(Figure 5)
Eroded fragment of ochre
Two sets of parallel incisions aligned at right-angles to each other are engraved.
M3-7(G6a, CH/CI)(Figure 5)
Flat fragment of ochre On one surface a group of conjoining lines are incised. This dendritic pattern is flanked by a single deep line on the right.
(Continued)

88 Christopher S. Henshilwood & Francesco d’Errico
Number & Provenience Brief description of piece
Type of engraving (see Henshilwood et al. 2009)
M3-8 (E4, CH/CI), M3-3 (E5b, CH/CI)(Figure 5)
Three conjoined pieces of ochre that come from an originally thick tabular piece
Apparent crenellated pattern made with different lithic tools. It is difficult to interpret the result as having been made with deliberate intent.
M3-9 (E4, CH/CI)(Figure 5)
A relatively large rectangular slab of ochre
One edge is engraved with a cross-hatched pattern. There is a possible dendritic motif on one side.
M3-10 (E5a, CJ)(Figure 5)
Pinkish-grey rounded slab of siltstone
One surface displays a long pair of sub-parallel single stroke lines. Four shorter convergent single stroke lines are engraved on the left and follow the same direction. Two of these lines end at the base of a slight, but steep rise in the surface.
On a number of pieces (M1/2, M1/5, M1/6, M3/1, M3/6, M3/10; Figures 4 and 5) regularity in the profile and outline of incisions indicates precise neuro-motor control. Good examples are M1/6 and M3/1 (Figures 4 and 5). On the former the engraver filled in a blank space by incising two lines to complete the symmetry of the pattern; in the latter case the start and end points of some lines were placed to exactly conjoin with existing lines.
Clearly, the engraved lines that form the motifs were carefully controlled by the makers during their execution and this action now assists in gauging the intent of the artist. Microscopic analysis of the engraved patterns allows us to identify consistency in the engraving techniques, the direction in which the lines were incised, and their chronological order (M1/3, M1/5, M1/6, M3/1, M3/6, M3/10). In sum, although a degree of ambiguity is certainly implied in our interpretation of some pieces, the results indicate that clear evidence of unambiguous engraved patterns are present in the MSA levels at BBC. Conservatively, this conclusion can be extended to include at least seven pieces coming from different layers (M1/2, M1/4, M1/5, M1/6, M3/1, M3/6, M3/10, and possibly the conjoins M3/3, M3/8: Figures 4 and 5).
6. Contrasting the significance of early engravings
The discovery of tool made markings on ostrich eggshells and on ochre pieces at MSA sites raises a number of questions including: (i) were the markings on these objects produced with the intent of embodying material culture with sym-bolic meaning?; (ii) were the DRS and BBC engravings perceived as symbolic by
Table 1. Description of Blombos engraved ochre pieces (Continued)

Chapter 4. Middle Stone Age engravings and their significance 89
the inhabitants of these sites at the time of their production or perhaps just the artisan; (iii) what was the main purpose of these markings and how may they have functioned with a symbolically mediated society – if this was the case?
Definitions of symbols, and how they act as behavioural mediators when they are recorded on a variety of media, varies according to disciplines, epistemological stands, and individual scholars (see for example, Jung & De Laszlo 1958; Savage-Rumbaugh 1986; Savage-Rumbaugh et al. 1986; Savage-Rumbaugh, this volume; McGrew, this volume; Tomasello 2008). A ‘broad’ definition for a symbol is something that represents something else by association, resemblance, or convention. A more ‘strict’ definition, that we adopt here, denotes a sign that has no natural connection or resemblance to its referent (Peirce 1998).
Archaeologists are unable to formally demonstrate that in prehistoric human societies meaning was associated to what we now perceive to be abstract depictions (Hodder 1982; Stark et al. 1993; Robb 1998). Arguments in favour of a symbolic hypothesis for these depictions rely on, and often try to combine, observations and inferences of various natures and heuristic power. Evaluating the pertinence of those inferences is central to understanding what we can say (and cannot say) about possible early instances of symbolically mediated behaviours.
Various explanations are suggested to support the symbolic interpretation of early engravings: (i) an absence of obvious functional reasons behind the production of the engravings; (ii) consistencies in the media on which the engravings are made; (iii) the preparation of the surface prior to engraving; (iv) the degree of neuromotor control inferred from the analysis of each line; (v) the type of tool used; (vi) the use of the same tool for the production of the entire pattern; (vii) the consistent organisation of the sequence of motions articulating the marking action; (viii) the regularity of the resultant pattern; (ix) the presence of engravings on a number of objects rather than on a single one; (x) the repetition of the same motif on more than one object; (xi) variations within what is perceived as the same basic motif; (xii) the production of a variety of different motifs; (xiii), temporal continuity in the production of engravings on the same media; (xiv) persistence or change in the production of motifs through time; (xv), production of similar engravings on the same media at a number of sites; and (xvi), similarity in the media used for engraving by prehistoric, extant and/or historically known groups. Although none of these arguments proves that the engravings or the objects carried a specific meaning for past artisans and their associated band or group that may differ to our current interpretation, we can draw on these arguments to help rule out alternative hypotheses, highlight possible inconsistencies in our interpretations, and help in making balanced analyses.

90 Christopher S. Henshilwood & Francesco d’Errico
At BBC and DRS there is continuity, over time, in the engravings being made on the same types of raw material. At both sites the chaine opératoire employed for engraving on ochre or eggshell shows that the artisans were deliberately producing engraved designs – in other words they were not accidental. But just how deliberate these designs were, and how much planning went into executing them, and whether they conformed to personal or social templates, remains elusive. We previously argued against the “doodling” and the “notational” interpretation in our discussion of the BBC engravings on ochre and bone (Henshilwood et al. 2009). Doodling is broadly defined as unfocused drawing made while a person’s attention is otherwise occupied. Incising lines on hard ochre requires focused attention so that the right pressure is applied to keep the depth of the incision constant. Both hands are required to perform this action, one to hold the pieces, which are small in the case of BBC, and one to engrave. A “notation” can be defined as a marking system specifically conceived to record, store and recover information outside the physical body (d’Errico 1995, 2001). Such a system must allow for a clear distinction between the marks that carry specific information. This is not present in most of the BBC engravings as the individual marks cannot be visually identified as discrete signs. Also, no evidence exists of sequential markings produced by different tools that may be interpreted as a notation based on an accumulation of information over time. At both sites each engraving was made in a single session, the makers had a clear goal when producing an engraved pattern, and they showed interest in the end result of their design. For DRS the conclusions could be tempered by the limited number of published pieces and by the very few pieces that have been technologically or microscopically analysed.
At both sites templates are apparent on a number of engraved patterns, for example the BBC cross-hatched designs and the DRS ladder-like motifs. The latter, however, present a large range of morphological variation that apparently shows no obvious conformity. In some engravings the “rungs” only cover three quarters of the space between the “members” of the ladder leaving a large empty space, in others they overstep them, there are three or more “members” instead of two, or some have intermediate members between “ladders” with no rungs. Texier et al. (2010) attribute this diversity to personal variability in the production of the same motif. It is possible that these engravings belong either to a more discrete, still not categorised template, or that the template was not actually a single category in the minds of the artisans. This means there could have been a communication system in place in which different engravings did not function as distinct symbols. We suggest that the whole object could have conveyed a symbolic message, irrespective of the type of engraving.

Chapter 4. Middle Stone Age engravings and their significance 91
Choosing between alternative hypotheses is complicated by the few published images of the engraved pieces of eggshell and by inconsistencies in assigning motifs to pieces. For example, the more common pattern, the ladder motif, apparently appears only on 69 of the 270 engraved fragments (Texier et al. 2010: 3; SOM table S1–2). No count is given for the fragments with the second most frequent pattern, the parallel lines motif. As many of the pieces are very small, it may hamper attributing a particular motif to a specific piece. For the two remaining motifs, the line intersecting two sets of “s” shaped parallel lines and the criss-cross pattern, there is only one fragment for each, but the latter is not depicted in the publication.
A consensual exploration of the number and proportion of motifs depicted on eggshell containers requires comprehensive publication of all the engraved fragments from DRS, including their contextual information, photographs and technological analysis. This would also allow evaluation of some of the other arguments proposed by the authors of the study to support their symbolic interpretation. The apparently high number of engraved eggshells at DRS can be used to argue in favour of engraving being a consistent behaviour at the DRS site compared to the only 8 engraved ochre pieces from BBC. However, most DRS fragments are very small and a dozen or more could come from one egg. The method used by the authors for estimating that the recovered fragments come from at least 25 eggs is not explained. The surface of an ostrich egg (Paganelli & Olszowka 1974) and that of the recovered fragments can easily be calculated to establish the minimum number of eggs that produced the fragments. Even if 25 is the correct number for the number of ostrich eggs that were decorated in the HP levels then this sample size is not significantly different from that of the engraved ochres at BBC. The ‘consistent behaviour’ argument is therefore not, in our opinion, stronger for the DRS engraved shells than could be made for the engraved BBC ochres.
The similarity that the DRS engraved shells have with eggshell containers used by extant or historical Khoesan is cited by Texier et al. (2010) as strong support for symbolic meaning once being attached to the HP engraved eggs. But, it is also apparent that in recent contexts, engravings could also simply signal ownership and may not be symbolic. The dangers associated with using the ethnographic present to explain the prehistoric past abound. Binford (1978) invoked “Middle Range Theory” to provide explicitly scientific explanations for the patterning of faunal remains in the Nunamiut archaeological record. There is wide ranging agreement that Binford’s approach and particularly linking middle range theory to a body of general theory about human behaviour is problematic as there is no consensus for the latter within archaeology (e.g. Hodder, 1982; Gould and Yellen, 1987). Do the similarities of the Khoesan engraved eggs to

92 Christopher S. Henshilwood & Francesco d’Errico
those in the HP at DRS allow for an argument suggesting cultural continuity or symbolic links? We question this hypothesis.
Labels such as “ownership” or “context” have rich and complex meanings in ethnographic societies. Such meanings and their relationship with the production and use of the engravings need to be explored in depth before any generalisation is proposed. If the ownership function is correct then one would expect to find a variety of visually distinct markings identifying different owners. This is not the case at DRS if the categorisation of motifs is correct, since apart from two exceptions only two main categories of patterns are present. Perhaps the owner/maker of an egg could be identified by group members based on the variation within one large “motif ”, and also that a particular “motif ” reflected more the cultural affiliation of the group than of a particular individual. If correct, this supports our contention that the categories of motifs are too flexible to be meaningful with regard to ‘marks of ownership’.
The argument for continuity of the HP engraving traditions with those of the present Khoesan requires that engraved eggs are found at sites from 60 ka up to the ethnographic present. This is not the case as engraved eggshells are not found at southern African archaeological sites until at least 40 ka years later. In our view one explanation is that the engraved eggshell tradition identified at DRS could be regarded as a cultural innovation independent from more recent instances of engravings on the same media. In this respect the DRS evidence supports and shares a strong similarity with cultural innovations recorded at other MSA sites. An example is the marine shell beads found within layers older than 70 ka at archaeological sites in North and South Africa, and in the Near East (see d’Errico and Henshilwood, this volume).
In conclusion, formally demonstrating that the engravings from DRS and BBC were made with symbolic intent is problematic, but we highlight in this chapter a number of consistencies in these engraved pieces that point the evidence in that direction. The main difference between the engraved records from these two sites is that the DRS eggshells, contrary to the engraved ochres at BBC were clearly made for visual display and were probably recognized as such by the members of the group in which they were used, and perhaps by the regional bands or groups that shared these symbolic meanings. On the other hand the symbolic codes expressed by engravings on BBC ochre are discreet, almost secret, and may be better explained as being a part of communication that involved only few individuals – perhaps even only at the level of the exchange of a gift between two people. What is clear, as more archaeological evidence becomes available, is that methods of communication and the use of symbols to mediate behaviour in the southern African MSA was complex, with perhaps regional variation, and that our understanding of the symbolic behaviours of these early Homo sapiens is still rudimentary.

Chapter 4. Middle Stone Age engravings and their significance 93
Acknowledgements
Financial support was provided to the authors by the European Research Council Advanced Grant TRACSYMBOLS (FP7 No. 249587), and the PROTEA French-South Africa exchange programme. CSH was also supported by a National Research Foundation/Department of Science and Technology funded Chair at the University of the Witwatersrand, South Africa a joint Norwegian Research Council/South African National Research Foundation Grant and the University of Bergen.
References
Alvarez, M., Fiore, D., Favret, E. & Guerra C. R. 2001. “The use of lithic artefacts for making rock art engravings: observation and analysis of use-wear traces in experimental tools through optical Microscopy and SEM.” Journal of Archaeological Science 28: 457–464.
Binford, L.R. 1978. Nunamiut ethnoarchaeology. New York: Academic Press.Bosinski, G. d’Errico, F. & Schiller, P. 2001. Die gravierten Frauendarstellungen von Gonnersdorf
(Der Magdalonien-Fundplatz Gonnersdorf Vol. 8). Stuttgart: Franz Steiner.Cox, M.V., Koyasu, M., Hiranuma, H. & Perara J. 2001. “Children’s human figure drawings in the
UK and Japan: The effects of age, sex and culture.” British Journal of Developmental Psychology 19 (18): 275–292.
d’Errico, F. 1995. L’art gravé azilien. De la technique à la signification. XXXIème. Supplément à Gallia-Préhistoire. Paris: CNRS Editions.
d’Errico, F. 1996. “Image analysis and 3-D optical surface profiling of Upper Palaeolithic mobiliary art.” Microscopy and Analysis 39: 27–29.
d’Errico, F. 2003. “The invisible frontier. A multiple species model for the origin of behavioural modernity.” Evolutionary Anthropology 12: 182–202.
d’Errico F. & Nowell A. 2000. “A new look at the Berekhat Ram figurine: implications for the origins of symbolism.” Cambridge Archaeology Journal 10 (1): 123–167.
d’Errico F., Henshilwood, C.S. & Nilssen, P. 2001. “An engraved bone fragment from ca. 75 kyr Middle Stone Age levels at Blombos Cave, South Africa: implications for the origin of symbolism.” Antiquity 75: 309–18.
d’Errico, F. & Backwell, L.R. 2003. “Possible evidence of bone tool shaping by Swartkrans early hominids.” Journal of Archaeological Science (30) 12: 1559–1576.
d’Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Soressi, M., Bresson, F., Tillier, A.M., Maureille, B., Nowell, A., Backwell, L., Lakarra, J.A. & Julien, M. 2003. “The search for the origin of symbolism, music and language: a multidisciplinary endeavour.” Journal of World Prehistory 17 (1): 1–70.
d’Errico, F., Henshilwood, C., Vanhaeren, M. & van Niekerk, K. 2005. “Nassarius kraussianus shell beads from Blombos Cave: Evidence for symbolic behaviour in the Middle Stone Age.” Journal of Human Evolution 48: 3–24.
d’Errico, F. & Henshilwood, C. 2007. “Additional evidence for bone technology in the southern African Middle Stone Age.” Journal of Human Evolution 52 (2): 142–163.
d’Errico, F., Henshilwood, C.S. this volume “The origin of symbolically mediated behaviour: from antagonistic scenarios to a unified research strategy.” In ‘Homo Symbolicus’: The

94 Christopher S. Henshilwood & Francesco d’Errico
Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds), 49–73. Amsterdam: John Benjamins.
Duday, H. 2009. The Archaeology of the dead. Oxbow Books, Oxford.Forster, P. 2004. Ice ages and the mitochondrial DNA chronology of human dispersals: a review.
Philosophical Transactions of the Royal Society of London B 359, 255–264.Freeman, N.H. & Cox, M.V. 1985. Visual order: the nature and development of pictorial
representation. Cambridge: Cambridge University Press.Fritz, C. 1999. La Gravure Dans l’art Mobilier Magdalénien. Du Geste à la Représentation. Paris:
Documents d’Archéologie Française 75, Editions de la Maison des Sciences de l’Homme.Goodwin, A.J.H. & van Riet Lowe, C. 1929. The Stone Age cultures of South Africa. Annals of
the South African Museum 27: 1–289.Gould, R. & Yellen, J. 1987. “Man the hunted: determinants of household spacing in desert and
tropical foraging societies.” Journal of Anthropological Archaeology 6: 77–103.Heese, C.H.T. n.d. Collected papers of C. H. T. D. Heese, on file at Iziko, South African Museum,
Cape Town.Henshilwood, C.S. 2006. “Stratigraphic integrity of the Middle Stone Age levels at Blombos
Cave.” In From tools to symbols. From early hominids to modern humans, L. Backwell and F. d’Errico (eds), 441–458. Johannesburg: Witwatersrand University Press.
Henshilwood, C.S., Sealy, J.C., Yates, R.J., Cruz-Uribe, K., Goldberg, P., Grine, F.E., Klein, R.G., Poggenpoel, C., van Niekerk, K.L. & Watts, I. 2001a. “Blombos Cave, southern Cape, South Africa: Preliminary report on the 1992–1999 excavations of the Middle Stone Age levels.” Journal of Archaeological Science 28 (5): 421–448.
Henshilwood, C.S., d’Errico F., Marean, C., Milo, R. & Yates, R. 2001b. “An early bone tool industry from the Middle Stone Age at Blombos Cave, South Africa: implications for the origins of modern human behaviour, symbolism and language.” Journal of Human Evolution 41: 631–678.
Henshilwood, C.S., d’Errico, F., Yates, R., Jacobs, Z., Tribolo, C., Duller, G.A.T., Mercier N., Sealy, J.C., Valladas, H., Watts, I. & Wintle, A.G. 2002. “Emergence of Modern Human Behaviour: Middle Stone Age engravings from South Africa.” Science 295: 1278–1280.
Henshilwood, C.S. & Marean, C.W. 2003. “The origin of modern human behaviour: A review and critique of models and test implications.” Current Anthropology 44 (5): 627–651.
Henshilwood, C.S., d’Errico, F., Vanhaeren, M., van Niekerk, K. & Jacobs, Z. 2004. “Middle Stone Age shell beads from South Africa.” Science 384: 404.
Henshilwood, C.S. & Marean, C.W. 2006. “Remodelling the origins of modern human behaviour.” In The prehistory of Africa: tracing the lineage of modern man, H. Soodyall (ed), 31–46. Johannesburg: Jonathan Ball Publishers.
Henshilwood, C.S., d’ Errico, F. & Watts, I. 2009. “Engraved ochres from the Middle Stone Age levels at Blombos Cave, South Africa.” Journal of Human Evolution 57: 27–47.
Henshilwood, C.S. & Dubreuil, B. 2009. “Reading the artefacts: Gleaning language skills from the Middle Stone Age in southern Africa.” In The cradle of language, R. Botha and C. Knight (eds), 41–60. Oxford: Oxford University Press.
Henshilwood, C.S. & Dubreuil, B. June 2011. “The Still Bay and Howiesons Poort, 77–59ka: Symbolic Material Culture and the Evolution of the Mind during the African Middle Stone Age.” Current Anthropology (52) 3: 361–400.
Hodder, I. (ed.). 1982. Symbolic and Structural Archaeology. Cambridge: Cambridge University Press.
Jacobs, Z., Duller G.A.T. & Wintle, A.G. 2003b. “Optical dating of dune sand from Blombos Cave, South Africa: II – single grain data.” Journal of Human Evolution 44: 613–625.

Chapter 4. Middle Stone Age engravings and their significance 95
Jacobs, Z., Wintle, A.G. & Duller, G.A.T. 2003a. “Optical dating of dune sand from Blombos Cave, South Africa: I—Multiple Grain Data.” Journal of Human Evolution 44: 599–612.
Jacobs, Z., Duller, G.A.T., Henshilwood, C.S. & Wintle, A.G. 2006. “Extending the chronology of deposits at Blombos Cave, South Africa, back to 140 ka using optical dating of single and multiple grains of quartz.” Journal of Human Evolution 51: 255–273.
Jacobs, Z., Roberts, R.G., Galbraith, R.F., Deacon, H.J., Grun, R., Mackay, A., Mitchell, P., Vogel-sang, R. & Wadley, L. 2008. “Ages for the Middle Stone Age of Southern Africa: Implications for Human Behavior and Dispersal.” Science 322 (5902):733–735.
Jones, H.L. 2001. Electron spin resonance dating of tooth enamel at three Palaeolithic sites. M.Sc.dissertation, McMaster University.
Jung, C.G. & De Laszlo, V.S. 1958. Psyche and Symbol: A Selection from the Writings of C.G. Jung. New York: Doubleday.
Klein, R.G., Avery, G., Cruz-Uribe, K., Halkett, D., Parkington, J., Steele, T., Volman, T. & Yates, R. 2004. “The Ysterfontein 1 Middle Stone Age Site, South Africa, and Early Human Exploitation of Coastal Resources.” Proceedings of the National Academy of Sciences 101 (16): 5708–5715.
Knight, C., Powers, C. & Watts, I. 1995. “The human symbolic revolution: a Darwinian account.” Cambridge Archaeological Journal 5 (1): 75–114.
Mackay, A. & Welz, A. 2008. “Engraved ochre from a Middle Stone Age context at Klein Kliphuis in the Western Cape of South Africa.” Journal of Archaeological Science 35 (6): 1521–1532.
McBrearty, S. & Brooks, A.S. 2000. “The revolution that wasn’t: a new interpretation of the origin of modern human behavior.” Journal of Human Evolution 39: 453–563.
McGrew, W. C. this volume. “Pan symbolicus. A cultural primatologist’s viewpoint.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds), 1–12. Amsterdam: John Benjamins.
Mellars, P. 2006. “Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model.” Proceedings of the National Academy of Sciences 103 (25): 9381–9386.
Paganelli, C.V. & Olszowka, A.A. 1974. “The avian egg: surface, area, volume, and density.” The Condor 76: 319–325.
Parkington, J. 1999. “Western Cape landscapes.” Proceedings of the British Academy, Oxford: Oxford University Press.
Peirce, C.S., 1998. The essential Peirce: selected philosophical writings. Bloomington: Indiana University Press.
Rigaud, J–P., Texier, P–J., Parkington, J. & Poggenpoel, C. 2006. “South African Middle Stone Age Chronology: New Excavations at Diepkloof Rock Shelter; Preliminary Results.” Palaeovol 5: 839–849.
Robb, J. E.1998. “The archaeology of symbols.” Annual Review of Anthropology 27: 329–346.Rogers, J. 1988. “Stratigraphy and geomorphology of three generations of regressive sequences
in the Bredasdorp group, southern Cape Province, South Africa.” In Geomorphological studies in southern Africa, G.F. Dardis and B.P. Moon (eds); 407–433. Rotterdam: Balkema.
Savage-Rumbaugh, S. 1986. Ape language: from conditioned response to symbol. New York: Columbia University Press.
Savage-Rumbaugh, S., McDonald, K., Sevcik, R.A., Hopkins, W.D. & Rubert, E. 1986. “Spontaneous symbol acquisition and communicative use by pygmy chimpanzees (Pan paniscus).” Journal of Experimental Psychology: General 115 (3): 211–235.

96 Christopher S. Henshilwood & Francesco d’Errico
Savage-Rumbaugh, S. & Fields, W.M., this volume. “The evolution and the rise of human lan-guage.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds), 13–47. Amsterdam: John Benjamins
Stark, M.P., Yoffe, A. & Sherrat, A. 1993. Archaeological theory: who sets the agenda? Cambridge: Cambridge University Press.
Texier, J.-P., Porraz, G., Parkington, J., Rigaud, J.-P., Poggenpoel, C., Miller, C., Tribolo, C., Cartwright, C., Coudenneau, A., Klein, R., Steele, T. & Vernai, C. 2010. “A Howiesons Poort tradition of engraving ostrich eggshell containers dated to 60,000 years ago at Diep-kloof Rock Shelter, South Africa.” Proceedings of the National Academy of Sciences 107 (14): 6180–6185.
Thomas, G.V. & Silk, A.M.J. 1990. An introduction to the psychology of representation in children. New York: Harvester Wheatsheaf.
Tomasello, M. 2008. Origins of human communication. Cambridge, Mass.: MIT Press.Tribolo, C., Mercier, N., Selo, M., Joron, J–L., Reyss, J–L., Henshilwood, C., Sealy, J. & Yates,
R. 2006. “TL dating of burnt lithics from Blombos Cave (South Africa): further evidence for the antiquity of modern human behaviour.” Archaeometry 48 (2): 341–357.
Tribolo, C., N. Mercier, H. Valladas, J.L. Joron, P. Guibert, Y. Lefrais, M. Selo, P.J. Texier, J.P. Rigaud & G. Porraz. 2009. “Thermoluminescence dating of a Stillbay–Howiesons Poort sequence at Diepkloof Rock Shelter (Western Cape, South Africa).” Journal of Archaeological Science 36 (3): 730–739.
Villa, P., Soressi, M., Henshilwood, C.S. & Mourre, V. 2009. “The Still Bay points of Blombos Cave (South Africa).” Journal of Archaeological Science 36 (2): 441–460.
Wadley, L. 2001. “What is cultural modernity? A general view and a South African perspective from Rose Cottage Cave.” Cambridge Archaeological Journal 11: 201–221.
Watts, I. 1998. The origins of symbolic culture: The southern African Middle Stone Age and Khoisan ethnography. Ph.D. thesis: University of London.
Watts, I. 1999. “The origin of symbolic culture.” In The Evolution of Culture, R. Dunbar, C. Knight & C. Power (eds), 113–146. Edinburgh: Edinburgh University Press.
Watts, I. 2009. “Red ochre, body-painting, and language: interpreting the Blombos ochre.” In The Cradle of Language, R. Botha & C. Knight (eds), 62–92. Oxford: Oxford University Press.
Watts, I. 2010. “The pigments from Pinnacle Point Cave 13B, Western Cape, South Africa.” Journal of Human Evolution 59 (6): 641–656.
Wendt, W.E. 1975. “Notes on some unusual artefacts from South West Africa.” Cimbebasia Ser. B 2 (6), 180–185.
Zilhão, J. 2001. Anatomically archaic, behaviorally modern: the last Neanderthals and their destiny. Amsterdam: Amsterdams Archeologisch Centrum.

chapter 5
Complex cognition required for compound adhesive manufacture in the Middle Stone Age implies symbolic capacity
Lyn WadleyInstitute for Human Evolution and School of Geography, Archaeology and Environmental Studies, University of the Witwatersrand
Compound adhesives were made in the Middle Stone Age (MSA) of southern Africa using a complex process in order to attach stone tools to shafts. Glue-makers must switch attention between fire control, measuring ingredients, and assembling compound tools. No rehearsals or recipes guarantee success; artisans must multi-task (a characteristic of modern human minds involving cognitive fluidity) and think abstractly about properties of glue ingredients. Mental rotation, an ability implying advanced working memory capacity, is needed to place stone inserts in various positions to create novel weapons and tools. The manufacturing process is sufficiently intricate that the early artisans must have had minds with abilities that overlapped those of people living today; such minds are capable of symbolic thought and action.
1. Introduction
The name Homo symbolicus describes us well; almost everything we do has sym-bolic undertones. It is therefore not surprising that we expect people like us in the past to have depended equally on symbolism, which would have demanded the use of some form of language. It is an entirely reasonable assumption. Archaeolo-gists like to use symbolic material culture as a marker in the deep past for behav-iour that overlaps with that of people living today. Nonetheless, few archaeologists agree on definitions of symbolism, symbolic material culture or even of attributes that imply modern behaviour (Wadley 2001; Henshilwood & Marean 2003). The presence of art is often taken to be an indicator of symbolism in the past, yet art historians themselves are not agreed on what is or is not art. Personal ornaments are thought to be markers of symbolism because they imply marking of personal or group identity. Shell beads with ages of between ~80 and 70 ka have been found

98 Lyn Wadley
at three Middle Stone Age (MSA) sites from distant corners of Africa: Taforalt, Morocco (Bouzouggar et al. 2007), Sibudu, KwaZulu-Natal (d’Errico et al. 2008) and Blombos, Western Cape (d’Errico et al. 2005). Most archaeologists consider these beads as persuasive evidence for early symbolism, but Wynn et al. (2009) represent the perspective that these archaeological traces are not smoking guns for concept formation and do not require that their makers had language. Thus, notwithstanding the consensus view that symbolism is represented in items of material culture by at least 80 ka, this position remains controversial in archaeo-logical and other scientific circles.
While few archaeologists define symbolism as it applies to items of material culture, an even smaller number study the kind of mental architecture that is required for symbolic innovations (Coolidge & Wynn 2005). This omission creates serious interpretive problems for, as Barnard (2010a) points out, we cannot create behavioural theory without employing theories of mental architecture.
Here, I draw on the savoir faire of archaeologists, linguists, psychologists and cognitive scientists to create an original way to recognise minds that have some resemblance to our own in the deep past without relying on the traditionally accepted (and disputed) indicators for early symbolism. This does not mean that I do not think symbolism important; it is important, but in this paper I wish to avoid the contention that surrounds recognition of symbolism in material culture items found in MSA sites. I shall therefore approach symbolism indirectly by examining unequivocal evidence for behaviour in the past that can be linked to human cognition like our own. In borrowing theories from psychologists and cognitive scientists, I recognise that these can also be controversial and short-lived (Barnard 2010b), and that my interpretations based on such theories will, at best, be superseded in the near future.
2. What is complex cognition?
The type of cognition attributed to people who think like us must minimally involve language (in itself symbolic), abstract thought, and capacity for novel, sustained multilevel operations. Language requires not only symbolism, but also abstraction (the mental ability to recognise common attributes or regularity amongst diverse objects or behaviours). Yet language also permits people to think and talk about behavioural variation (Reuland 2010). All complex cognitive processes have abstraction as a common denominator; abstraction would not have been possible prior to the advent of people with thought processes much like ours (Barnard 2010a). Furthermore, Barnard suggests that only the most advanced mental architecture can simultaneously control multilevel operations

Chapter 5. Complex cognition required for compound adhesive manufacture 99
like talking, thinking and walking. The capacity for novel, sustained multilevel actions could have arisen from neural connectivity in part of the prefrontal cortex (Amati & Shallice 2007), a development that may have been favoured in modern humans by a relatively early increase in overall brain size (Aboitiz 2001). Mithen (1996) talks about ‘cognitive fluidity’; the term is compelling because it implies the ease with which modern humans generate and employ innovative thoughts. Many neurons are needed to resolve intricate behavioural challenges and the idea of a neural mutation giving rise to complex cognition of the kind used by modern people was introduced years ago (for example, Coolidge & Wynn 2005; Klein & Edgar 2002; Mithen 1996). Neurons need, and possess, numerous transmitters, maybe because many nerve terminals synapse on a single neuron (Deutch & Roth 2003). There are critical periods in animal and human maturation processes (required for bird song or human speech) that are greatly affected by the development of neural pathways. Neurons select their permanent range of inputs during childhood, when the capacity for adjustment is greater than is possible after the achievement of sexual maturity (Knudsen 2003). The opportunity for neural change tends to ‘close’ at sexual maturity, irrespective of experience obtained, thus long childhood and late onset of sex hormones allow for extended learning which is beneficial for the development of complex capabilities among humans (Knudsen 2003: 570).
3. What gave rise to complex cognition?
The prefrontal cortex seems to have a fundamental role in cognitive control. Prefrontal cortex neurons participate in cognitive control – they can sustain their activity to keep task-relevant information online during tasks (Miller & Wallis 2003). Neural storage in the form of ‘working memory’ can hold things in mind (in the sense discussed earlier) at the same time as manipulating information. Working memory capacity correlates with fluid intelligence (an ability to solve novel problems) and enhanced working memory is an important prerequisite for the attributes of modern cognition mentioned in the previous section: language, creation of abstract ideas and the planning required for manufacturing tools and other artefacts. The enhancement of working memory that occurred in the rela-tively recent human past, most likely after the first appearance of anatomically modern humans, may have marked the final stage in the evolution of reason and the development of language and culture (Coolidge & Wynn 2005). Enhanced working memory enables innovation by making thought experiments possible. These work through heuristic action and an ability to make predictions into the future. Relatively modern innovative technologies that provide good cases in point

100 Lyn Wadley
for enhanced working memory include alloying metals, and the production of kiln-fired ceramics (Coolidge & Wynn 2006; Wynn & Coolidge 2007a,b).
Working memory capacity can predict higher order cognitive tasks, such as attention, language comprehension and production, reasoning, and general and fluid intelligence (novel problem solving abilities) (Engle & Kane 2004; Kane & Engle 2002). One of the ways in which psychologists predict cognitive performance on higher order cognitive tasks is through reliable and valid psychometric tests involving complex span tasks (Kane et al. 2004). Amongst these tasks are spatial ones requiring a person, for example, to rotate a letter mentally or decide whether a figure is symmetrical around a vertical axis (Kane et al. 2004). Thus technology that requires mental rotation is a good indicator of complex cognition.
The appearance of enhanced working memory, most likely the result of a relatively simple, genetically inherited neurological advance, resulted in cognitive abilities called ‘executive functions’ that depend on frontal-lobe-linked abilities (Wynn & Coolidge 2003). Complex goal-directed actions, flexibility in problem-solving, innovative solutions to problems, analogical reasoning and planning over long distances or time are amongst executive functions (Wynn & Coolidge 2003). The ability to switch attention is a marker of the kind of thought process enabled by modern executive control. Switching attention requires people to move their focus of attention: it involves scheduling as well as inhibiting processes (Smith & Jonides 2003). One test for the ability to switch attention involves using the Wisconsin Card Sort Task in which subjects are asked to sort cards by colour. Then they are asked to switch attributes and sort cards, for instance, by the designs on them. People with frontal lobe damage cannot do this (Smith & Jonides 2003). A more complex task involves switching between objects in working memory (such as between equations and words). Here the subject needs to switch between updating working memory and some other task (Smith & Jonides 2003).
Generally I should agree with Wynn and Coolidge (2007b) that technology and subsistence are spheres of action that rely not on complex mental models, but rather on well-learned procedural routines that can be taught during apprenticeship. Most subsistence activities can be learned by observation and training in a principally non-verbal way and can be classified as belonging to the procedural type of long-term memory. Nevertheless, I suggest here that the capacity for novel, sustained multilevel operations can be predicted from some types of technology and I use compound adhesive manufacture as an example. In order to demonstrate that technology can be a proxy for complex cognition, I must make it clear that some cognitive executive steps cannot be taken without drawing on mental abilities such as abstraction, recursion, multilevel thought and cognitive fluidity.

Chapter 5. Complex cognition required for compound adhesive manufacture 101
4. The archaeological evidence
Pigments, including red ochre, have a long history of use in Africa, where they may even have been used by archaic Homo sapiens (d’Errico 2008). Thus the use of red ochre per se cannot be used as a marker of behaviour associated exclusively with anatomically modern humans. My own study was inspired by Middle Stone Age (MSA) stone tools called segments. Segments (sometimes known in the literature as crescents or lunates) are defined as a portion of a circle with a curved, abruptly-blunted back and a straight, sharp cord (Deacon 1984) (Figure 1). Each end of a segment is pointed. The abrupt blunting (which is faceted in a manner that archaeologists call backed retouch) on the convex edge of segments probably assists their hafting by creating an area of friction for the firm attachment of adhesives (Lombard 2007; Nuzhnyi 2000; Phillipson 1976). Residue analysis supports the suggestion that backed edges were designed to facilitate hafting because plant gum/resin, sometimes mixed with ochre or other substances, was found on the backed edges of segments and other backed tools from the Howiesons Poort Industry with ages of between 64 ka and 61 ka (Jacobs et al. 2008) at Rose Cottage (Gibson et al. 2004) and Sibudu (Delagnes et al. 2006; Lombard 2006, 2007, 2008) (Figure 1).
Ochre-stained segments were also found in Kenya at Enkapune Ya Muto, dated to between 50 ka and 40 ka (Ambrose 1998). At Sibudu, carefully-mapped positions of glue residues show that segments could have been hafted in a vari-ety of positions (Lombard 2007, 2008). By rotating a segment so that its cutting edge is horizontal, vertical or diagonal, it can be hafted to form one of a variety of tools, weapons such as arrowheads or spearheads, or weapon components, such as barbs that would be placed below the arrow tip or spearhead. Segments, because of their half-moon shape, with the straight cutting edge along the full length of the tool, cannot be bound with twine (the cutting edge would sever the twine) and the hafting of these stone inserts must have depended entirely on adhesive. Plant gum used alone is effective glue – Acacia gum, for example, is designed to seal wounds on the tree – though it often shatters like glass on direct impact. Such brittle glue is only advantageous when weapon tips are intended to shatter inside prey (Clark 1975) where the sharp, broken fragments of stone would cause severe haemorrhaging that would kill the animal and leave a trail for hunters to follow. Adding ochre or some other aggregate to plant gum creates an adhesive that is workable, plastic and with a good consistency that hardens to a robust product (Wadley 2005; Wadley et al. 2009). This is the kind of glue that would be needed to prevent the stone tip of a spear from splitting from its shaft during repeated thrusts at prey. Some glue on ancient stone tools contains red, yellow or orange ochre, while other specimens have no ochre at all. Instead, some glue seems to contain plant gum mixed with fat, and perhaps sand or charcoal (it is difficult to

102 Lyn Wadley
know whether charcoal and sand are post-depositionally incorporated into plant gum residues on stone tools). Thus, it looks as if artisans in the past intentionally selected task-appropriate recipes for their adhesives and that they were fully aware of the properties of the required ingredients. Replications of compound glues and their use with experimental tools demonstrate that chemical, mechanical and possibly electrostatic changes are affected by the addition of ochre to plant gum (Wadley et al. 2009), so it seems that ochre additions have technical applications, whether or not they also have symbolic meaning. People may have coloured their weapons with red ochre for symbolic purposes, for example, to signify the blood of prey and success in the hunt, but this explanation cannot be the only one for the mixing of ochre into glue. First, there are the technical issues already men-tioned. Then, the coloured stains are present only on those parts of stone tools once attached to hafts (Figure 1); MSA weapons were not entirely covered with ochre, as has been the case in recent ethnographic examples in Australia (Wallis & O’Connor 1998). Importantly, not all MSA adhesives contain red ochre and this would be expected if the colourant was considered essential for hunting fortune. Whether or not symbolism played a role in the placement of red ochre in the adhe-sives, the process of making compound adhesives is so complex that it could only have been undertaken by people whose minds had abilities that overlapped with our own. I shall demonstrate this point now.
Cutting edge
BackingOchre
10 mm
Figure 1. Segment from Sibudu Cave showing red ochre traces that were part of the compound adhesive used to attach it to a shaft or handle. The segment comes from a layer with an age of about 65 ka. The line drawing of another segment from Sibudu shows the relative positions of the backed and cutting edges

Chapter 5. Complex cognition required for compound adhesive manufacture 103
5. Replicated compound adhesive manufacture: Methods
To explore the efficacy of ochre-loaded glue, and the complex thought processes required by its manufacture, replicated tools were hafted with compound adhesive made from red ochre mixed with Acacia karroo gum (see Figure 2 for an example of one such tool). Only products and methods available in the MSA were used, except where synthetic iron oxide was attempted for comparison with the geologi-cal products. Yellow and red ochres were collected from a Snuffbox Shale quarry in the vicinity of Sibudu Cave, but one sample of red ochre came from the Waterberg, Limpopo. Ochre was powdered by rubbing nodules on coarse, flat, sandstone slabs (Sibudu roof-spall) (Figure 3) because such slabs with ochre stains have been found at Sibudu. Three main adhesive recipes were used: (1) Acacia gum alone, (2) Acacia gum mixed with either natural ochre powder or synthetic haematite powder and, (3) Acacia gum mixed with natural ochre powder and melted beeswax. All replica-tions were conducted with open wood fires whose temperatures varied depending on the type and amount of wood used, but did not exceed 726°C. Detailed method-ology is recorded elsewhere (Wadley 2005; Wadley et al. 2009).
1 cm
Figure 2. Replicated compound adhesive using Acacia gum and red ochre. The glue was used to attach a replicated stone flake to a wooden handle. The tool was subsequently successfully used to chop bark from branches
The partially dehydrated adhesive was used to mount the stone insert to its wooden haft (for example, Figure 2) and the hafted tool was placed near the fire (60oC–100oC) for four hours to dry and harden the adhesive. Each tool was rotated regularly to prevent adhesive dripping from the shaft, and each was moved closer

104 Lyn Wadley
or farther from the fire depending on its heat. The entire process was intricate and demanded full attention, requiring the manufacturer to hold many things in mind simultaneously. No set recipe or procedure could be used for any single tool or compound adhesive because of the variability amongst ingredients such as the plant gum. Natural products and wood fires have unpredictable variables. Artisans in the past were obliged to deal with these just as I was, and they had to compensate for the difficulties presented by their ingredients. For example, Acacia gums from different trees or different seasons have changeable viscosity, thereby requiring varying amounts of loading agent. Acacia gum that I worked with fluctuated between being highly viscous, and dry and crystalline, and its pH varied between 3.0 and 4.4. Thus, the chemical reaction between the gum and ochre differs between adhesive mixtures with different attributes. Ochre pieces can also be dissimilar in respect of their quantities of iron and other minerals and also in respect of particle size. Some ochre pieces grind into coarser powders than others, depending on their silica contents and also on the composition of the stone on which they were ground.
Ancient glues were almost certainly heated near a fire because, without dehydrating the replicated glues, they take six days or more to dry. Experimentally produced tools separated from their shafts when the glue was not properly dry. Camp fire temperatures depend on the wood type used, the amount of fuel, ambient humidity and wind speed, so artisans would have needed to control fire temperature to avoid spoiling the adhesives by burning them or causing them to boil. Thus, compound adhesives of the kind described here could not have been made using fixed recipes as we do for making cakes. On-going assessments and on-the-spot adaptations, requiring cognitive fluidity, must have been made during adhesive production. It is apparent that people in the MSA were sufficiently skilled to realise early in the amalgamation of ingredients whether adhesive had the right consistency and workability. If necessary, workability could be improved by adding more aggregate such as ochre, sand and/or small amounts of fat or beeswax. The early artisans must have made choices and measurements based on the ability of certain combinations of ingredients to create reliable adhesives. Successful compound adhesive manufacture using ochre and plant gum seems to have been achieved when there was (1) a chemical change from acid to less acid pH, (2) a transformation in mechanical properties to one of workability, and (3) slow dehydration of wet adhesives near open fires.
6. Compound adhesive manufacture as a proxy for modern cognition
Creating compound adhesive from disparate ingredients may have been regarded as symbolic in the past, but this is a difficult interpretation to make. From the evidence

Chapter 5. Complex cognition required for compound adhesive manufacture 105
presented here, it is easier to claim that people in the MSA were conversant with some aspects of chemistry and pyrotechnology. While they are unlikely to have known pH as we understand it, or have known how to gauge the percentages of iron or silica in rocks, they did seem to understand the importance of complexing iron minerals or colloids with acid plant gum and dehydrating them near controlled, open fires. Furthermore, some of the steps required for making compound adhesive seem impossible without the ability to abstract relevant features of the products they worked with. Qualities of gum, such as wet, sticky and viscous, must have been mentally abstracted and these meanings weighed against ochre powder properties such as dry, loose and dehydrating. It is not easy to imagine how an expert glue-maker could train an apprentice to make compound adhesives without language that could explain, in abstract terms, attributes and conditions such as stickiness, viscosity, workability, consistence, plasticity, texture, particle size, temperature, concretization, water-solubility, hydroscopic, dehydration, reversible process, irreversible process, shrinkage, homogeneity, creep and shrinkage. These concepts probably had to be explained using language as we appreciate it, in other words incorporating recursion, abstraction and concepts about the past and the future.
The glue makers needed to keep in mind work already done in order to carry out those tasks still needed for the successful completion of the glue and the mounting of the composite parts of weapons or tools. In addition, the artisans had to be equipped to alter ingredients and adjust fire temperatures on-the-spot; concurrently they had to think about the appropriate angles for placing stone inserts on their hafts. They were able to perform mental rotation of segments or other tools in order to create weapons or other implements in an assortment of designs for different objectives. They also needed to think abstractly about the qualities of their segments in order to envisage their use after they were rotated at different angles. As mentioned earlier, successful mental rotation requires advanced working memory capacity (Kane et al. 2004) and, in turn, complex cognition.
Some birds (such as swallows that build mud nests) and wasps also create forms of compound adhesives, but they do so instinctively with simply coded operational sequences, in which the distance between problem and solution is far smaller than that demonstrated by the human action of making a composite hunting weapon (Haidle 2010). One obvious difference in human manufacture of compound glue is the use of pyrotechnology. Temperature control depends on understanding wood types, their moisture or resin contents and their propensity to form long-lasting coals. Vigilance is essential because my adhesives burned, or boiled to form air bubbles, when they were too close to the fire. Over-dehydration caused loss of cohesiveness and a crumbly product, while adhesive that boiled lost homogeneity and consequently became unstable.

106 Lyn Wadley
Figure 3. An ochre nodule used to create red ochre powder by grinding it on a sandstone slab
The glue-maker needs to pay careful attention to the condition of ingredients before and during the procedure and must be able to switch attention between aspects of the methodology. To hold many courses of action in mind involves multi-tasking, which is one trait of modern human minds (Barnard et al. 2007) as well as mental flexibility. Capacity for multilevel operations, abstract thought and mental rotation are all required for the process of compound adhesive manufacture and hafting of tools. Although fully modern behaviour seems recognizable only relatively late in the MSA (Klein 2008), the circumstantial evidence provided here from experimental work with compound adhesives suggests that people who made them in the MSA shared some advanced behaviours with their modern successors. While compound adhesives have presently only been recorded on stone tools that are about 70 000 years old, this date is likely to be pushed farther into the past when residue analysis is conducted on older tools.
In conclusion, I contend that compound adhesive manufacture in the MSA required complex cognition of the kind that intersected with our own. People who make compound adhesives from natural products such as plant gum and geologically variable iron oxides must be able to multi-task because they must simultaneously think, mix glue (changing quantities at short notice) and maintain fire temperature. Since the intention is always to use the glue to create composite tools, the artisan must also be able mentally to rotate stone tools and visualise

Chapter 5. Complex cognition required for compound adhesive manufacture 107
the finished tool or weapon. Some of the stages involved in making compound adhesives, and their use with composite tools, entail not only multi-tasking, but also mental abstraction, switching attention, recursive behaviour and cognitive fluidity. Accordingly, it seems justifiable to attribute advanced mental abilities to people who lived in the African MSA. Although symbolism has not been studied directly in this paper, complex cognition of the kind described here is most likely to have co-occurred with a capacity for symbolic thought and behaviour. So many variables and unpredictable actions are involved in the production of compound adhesive and in the hafting of segments that the process seems to require language for instructing apprentices. I suggest that my circuitous route for finding symbolism in the archaeological record specifically that of the MSA of Africa is theoretically satisfying. Furthermore, the approach avoids making a leap of faith between ancient items of material culture and their meaning.
References
Aboitiz, F. 2001. “Size and complexity of the brain in human evolution.” In Humanity from African naissance to coming millenni, P. V. Tobias, M.A. Raath, J. Moggi-Cecchi & G.A. Doyle, (eds), 355–359. Firenze: Firenze University Press.
Amati, D. & Shallice, T. 2007. “On the emergence of modern humans.” Cognition 103: 358–385.
Ambrose, S.H. 1998. “Chronology of the Later Stone Age and food production in East Africa.” Journal of Archaeological Science 25: 377–392.
Barnard, P.J. 2010a. “From executive mechanisms underlying perception and action to the parallel processing of memory.” Current Anthropology 51 (1): S39–54.
Barnard, P.J. 2010b. “Current developments in inferring cognitive capabilities from the archaeological traces left by stone tools: caught between a rock and a hard inference.” In Stone tools and the evolution of human cognition, A. Nowell and I. Davidson (eds), 207–226. Boulder, CO: Colorado University Press.
Barnard, P.J., Duke, D.J., Byrne, R.W. & Davidson, I. 2007. “Differentiation in cognitive and emotional meanings: an evolutionary analysis.” Cognition and Emotion 21: 1155–1183.
Bouzouggar, A., Barton, N., Vanhaeren, M., d’Errico, F., Collcutt, S., Higham, T., Hodge, E., Parfitt, S., Rhodes, E., Schwenninger, J.-L., Stringer, C., Turner, E., Ward, S., Moutmir, A. & Stambouli, A. 2007. 82,000-year-old shell beads from North Africa and implications for the origins of modern human behaviour. Proceedings of the National Academy of Science, USA 104: 9964–9969.
Clark, J.D. 1975. “Interpretations of prehistoric technology from ancient Egyptian and other sources. Part II: Prehistoric arrow forms in Africa as shown by surviving examples of the traditional arrows of the San Bushmen.” Paléorient 3: 127–150.
Coolidge, F.L. & Wynn, T. 2005. “Working memory, its executive functions, and the emergence of modern thinking.” Cambridge Archaeological Journal 15: 5–26.
Coolidge, F.L. & Wynn, T. 2006. “Recursion, phonological storage capacity, and the evolution of modern speech.” A paper presented to the Cradle of Language Conference, November 2006, Stellenbosch, South Africa.

108 Lyn Wadley
Deacon, J. 1984. The Later Stone Age of southernmost Africa. Cambridge Monographs in African Archaeology 12. Oxford: British Archaeological Reports.
Delagnes, A., Wadley, L., Villa, P. & Lombard, M. 2006. “Crystal quartz backed tools from the Howiesons Poort at Sibudu Cave.” Southern African Humanities 18: 43–56.
d’Errico, F. 2008. “Le rouge et le noir: implications of early pigment use in Africa, the Near East and Europe for the origin of cultural modernity.” South African Archaeological Society Goodwin Series 10: 168–174.
d’Errico, F., Henshilwood, C., Vanhaeren, M. & van Niekerk, K., 2005. “Nassarius kraussianus shell beads from Blombos Cave: evidence for symbolic behaviour in the Middle Stone Age.” Journal of Human Evolution 48: 3–24.
d’Errico, F., Vanhaeren, M. & Wadley, L. 2008. “Possible shell beads from the Middle Stone Age layers of Sibudu Cave, South Africa.” Journal of Archaeological Science 35: 2675–2685.
Deutch, A.Y. & Roth, R.H. 2003. “Neurotransmitters.” In Fundamental Neuroscience, Second Edition, L.R. Squire, F.E. Bloom, S.K. McConnell, J.L. Roberts, N.C. Spitzer and M.J. Zigmond, (eds), 163–196. San Diego: Academic Press.
Engle, R.W. & Kane, M.J. 2004. “Executive attention, working memory capacity, and a two-factor theory of cognitive control.” In The psychology of learning and motivation, B. Ross (ed), 145–199. New York: Academic Press.
Gibson, N.E., Wadley, L. & Williamson, B.S. 2004. “Microscopic residues as evidence of hafting on backed tools from the 60 000 to 68 000 year-old Howiesons Poort layers of Rose Cottage Cave, South Africa.” Southern African Humanities 16: 1–11.
Haidle, M.N. 2010. “Working-Memory Capacity and the evolution of modern cognitive potential: Implications from animal and early human tool use.” Current Anthropology (51) 1: S149–166.
Henshilwood C. & Marean C. 2003. “The origin of modern human behavior: critique of the models and their test implications.” Current Anthropology 44: 627–651.
Jacobs, Z., Roberts, R.G., Galbraith, R.F., Deacon, H.J., Grün, R., Mackay, A.M., Mitchell, P., Vogelsang, R. & Wadley, L. 2008. “Ages for the Middle Stone Age of Southern Africa: Implications for human behavior and dispersal.” Science 322: 733–735.
Kane, M.J. & Engle, R.W. 2002. “The role of prefrontal cortex in working memory capacity, executive attention, and general fluid intelligence: An individual-differences perspective.” Psychonomic Bulletin and Review 9: 637–671.
Kane, M.J., Hambrick, D.Z., Tuholski, S.W., Wilhelm, O., Payne, T.W. & Engle, R.W. 2004. “The generality of working memory capacity: A latent-variable approach to verbal and visuospatial memory span and reasoning.” Journal of Experimental Psychology: General 133: 189–217.
Klein, R.G. & Edgar, B. 2002. The dawn of human culture. New York: Wiley and Sons.Klein, R.G. 2008. “Out of Africa and the evolution of human behavior.” Evolutionary Anthropology
17: 267–281.Knudsen, E.I. 2003. “Early experience and critical periods.” In Fundamental Neuroscience,
Second Edition. L. R. Squire, F.E. Bloom, S.K. McConnell, J.L. Roberts, N.C. Spitzer and M. J. Zigmond, (eds), 555–573. San Diego: Academic Press.
Lombard, M. 2006. “Direct evidence for the use of ochre in the hafting technology of Middle Stone Age tools from Sibudu Cave.” Southern African Humanities 18: 57–67.
Lombard, M. 2007. “The gripping nature of ochre: the association of ochre with Howiesons Poort adhesives and Later Stone Age mastics from South Africa.” Journal of Human Evolution 53: 406–19.

Chapter 5. Complex cognition required for compound adhesive manufacture 109
Lombard, M. 2008. “Finding resolution for the Howiesons Poort through the microscope: micro-residue analysis of segments from Sibudu Cave, South Africa.” Journal of Archaeo-logical Science 35: 26–41.
Miller, E.K. & Wallis, J.D. 2003. “The prefrontal cortex and executive brain functions.” In Fundamental Neuroscience, Second Edition. L.R. Squire, F.E. Bloom, S.K. McConnell, J.L. Roberts, N.C. Spitzer & M.J. Zigmond, (eds),1353–1376. San Diego: Academic Press.
Mithen, S. 1996. The prehistory of the mind. London: Thames and Hudson.Nuzhnyi, D.Y. 2000. “Development of microlithic projectile weapons in the Stone Age.” Anthro-
pologie et Prehistoire 111: 95–101.Phillipson, D.W. 1976. The prehistory of Eastern Zambia. Nairobi: Memoir 6 of the British
Institute in Eastern Africa.Reuland, E. 2010. “Imagination, planning and working memory: the emergence of language.”
Current Anthropology (51) 1: 99–110.Smith, E.E. & Jonides, J. 2003. “Executive control and thought.” In Fundamental Neuroscience,
Second Edition. L.R. Squire, F.E. Bloom, S.K. McConnell, J.L. Roberts, N.C. Spitzer & M.J. Zigmond, (eds), 1377–1394. San Diego: Academic Press.
Wadley, L. 2001. “What is cultural modernity? A general view and a South African perspective from Rose Cottage Cave.” Cambridge Archaeological Journal 11: 201–221.
Wadley, L. 2005. “Putting ochre to the test: replication studies of adhesives that may have been used for hafting tools in the Middle Stone Age.” Journal of Human Evolution 49: 587–601.
Wadley, L., Hodgskiss, T. & Grant, M. 2009. “Implications for complex cognition from the hafting of tools with compound adhesives in the Middle Stone Age, South Africa.” Proceedings of the National Academy of Science, USA 106: 9590–9594.
Wallis, L. & O’ Connor, S. 1998. “Residues on a sample of stone points from the west Kimberley” In A closer look: recent Australian studies of stone tools, R. Fullagar (ed), 149–178. Sydney: Sydney University Press.
Wynn, T. and Coolidge, F.L. 2003. “The role of working memory in the evolution of managed foraging.” Before Farming 2003/2 (1): 1–16.
Wynn, T. & Coolidge, F.L. 2007a. “Executive functions, working memory, and the evolution of the modern mind.” Paper presented to AAAS, March, 2007.
Wynn, T. & Coolidge, F.L. 2007b. “A Stone Age meeting of minds.” American Scientist 96: 44–51.Wynn, T., Coolidge, F.L. & Bright, M. 2009. “Hohlenstein-Stadel and the evolution of human
conceptual thought.” Cambridge Archaeological Journal 19: 73–83.


chapter 6
The emergence of language, art and symbolic thinking
A Neandertal test of competing hypotheses
João ZilhãoICREA Research Professor SERP – Department of Prehistory, Ancient History and Archaeology, University of Barcelona/Institució Catalana de Recerca i Estudis Avançats (ICREA)
There is a widespread understanding that the personal ornaments of the African Middle Stone Age and the animal and human figurines of the Aurignacian of southern Germany provide the earliest evidence of the possession of “modern” cognitive capabilities, ones that appeared for the first time in human evolution as a result of the speciation of Homo sapiens and that would explain its rapid expansion from Africa into Eurasia and the attendant extinction of coeval archaic humans (such as the Neandertals). The archaeological facts contradict this view, since there is abundant evidence for the existence of such “modern” capabilities in non-sapiens populations, and that language, “symbolic thinking” by definition, is probably as old as the human genus. Therefore, the explanation for the emergence of body ornamentation and figurative art must be sought not in the realm of cognition but in that of history, with demographic growth and the intensification of social interaction networks playing a primary role in the process.
1. Introduction
The last fifty years of scientific research established beyond reasonable doubt that the earliest human ancestors appeared in Africa some time around two million years ago. Soon after, these Homo erectus people expanded into Eurasia. By one and a half million years ago, they had already reached the Indonesian island of Java, but it would take a bit longer for Europe to be stably settled (Dennell & Roebroeks 2005).
The earliest evidence comes from Iberia, where the so-called Homo antecessor fossils from Atapuerca date to about one million years ago (Bermúdez de Castro et al. 2004; Carbonell et al. 2008). Coeval African fossils are scarce, but, altogether, the evidence suggests that a trend towards increased brain size and correlated changes in the shape of the skull was under way at this time throughout the entire

112 João Zilhão
Old World (McHenry & Coffing 2000; Lee & Wolpoff 2003). Geneticists have related these changes to a second Out-of-Africa expansion, represented, archaeo-logically, by the spread of the Acheulian technocomplex, whose iconic stone tool is the handaxe or biface (Templeton 2002, 2005).
Subsequent geographic isolation led to the differentiation of these Acheu-lian populations into two lineages. In Europe and western Asia, Homo erectus became Neandertal man sometime around 500 ka. At the same time, in Africa, Homo erectus became Homo sapiens (or “modern humans”), and, some 50 ka, in the framework of a third Out-of-Africa event, spread into Eurasia, Australia, and, eventually, the Americas (Trinkaus 2005).
Over the last quarter of a century, it has become clear that early African sapi-ens are ancestral to all present-day living humans but no agreement exists where Eurasian Neandertals are concerned. The level of their separation in taxonomy, the extent of their differences in biology, behaviour and culture, and their ultimate fate, remain to this day among the hottest topics in human evolution studies. That Nean-dertals are no more is uncontroversial, but when, why and how were they replaced?
These questions have fundamental implications for the understanding of the emergence of art, language and symbolic thinking in the human lineage. The long lasting geographical segregation of the two palaeontological taxa, Homo neanderalensis and Homo sapiens, and the ultimate replacement of the former by the latter are widely assumed to imply that they were truly different biological species. And, as textbook definitions require species to differ in behaviour as much as in morphology, the corollary expectation is that significant behav-ioural differences, with attendant cognitive implications, must have separated “anatomically modern” people from coeval “archaic” humans, namely the Neandertals (Henshilwood & Marean 2003).
The notion that such a separation existed at biospecies level dovetails with speculations that certain features of complex human culture, which are undoc-umented in the archaeological record of Homo erectus and other early humans such as art, or ritual burial, must have emerged as a by-product of the processes involved in the speciation of the African sapiens. Under this “Human Revolution” hypothesis (Mellars & Stringer 1989; Mellars et al. 2007), the absence of those features reflects the lack of the required cognitive capabilities. In this view, it is only after the acquisition of such capabilities by the first “modern humans” that the corresponding behavioural correlates could be and indeed were externalized in archaeologically visible ways.
Material evidence consistent with this paradigm – marine shell beads, inter-preted as personal ornaments and, in some instances, associated with skeletal remains of early modern humans – has been obtained over the last decade at archaeological sites dated to between 75 ka and 100 ka in Palestine, Morocco
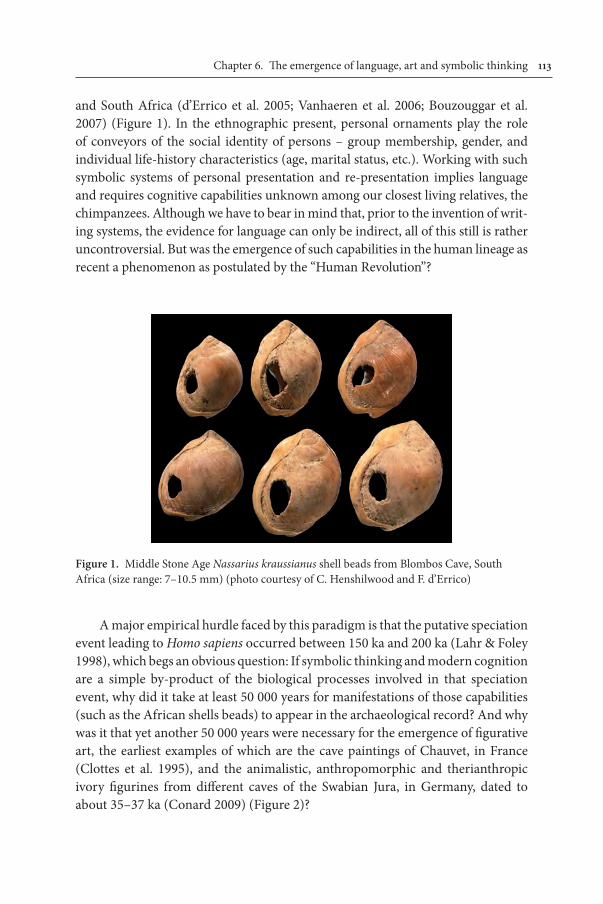
Chapter 6. The emergence of language, art and symbolic thinking 113
and South Africa (d’Errico et al. 2005; Vanhaeren et al. 2006; Bouzouggar et al. 2007) (Figure 1). In the ethnographic present, personal ornaments play the role of conveyors of the social identity of persons – group membership, gender, and individual life-history characteristics (age, marital status, etc.). Working with such symbolic systems of personal presentation and re-presentation implies language and requires cognitive capabilities unknown among our closest living relatives, the chimpanzees. Although we have to bear in mind that, prior to the invention of writ-ing systems, the evidence for language can only be indirect, all of this still is rather uncontroversial. But was the emergence of such capabilities in the human lineage as recent a phenomenon as postulated by the “Human Revolution”?
Figure 1. Middle Stone Age Nassarius kraussianus shell beads from Blombos Cave, South Africa (size range: 7–10.5 mm) (photo courtesy of C. Henshilwood and F. d’Errico)
A major empirical hurdle faced by this paradigm is that the putative speciation event leading to Homo sapiens occurred between 150 ka and 200 ka (Lahr & Foley 1998), which begs an obvious question: If symbolic thinking and modern cognition are a simple by-product of the biological processes involved in that speciation event, why did it take at least 50 000 years for manifestations of those capabilities (such as the African shells beads) to appear in the archaeological record? And why was it that yet another 50 000 years were necessary for the emergence of figurative art, the earliest examples of which are the cave paintings of Chauvet, in France (Clottes et al. 1995), and the animalistic, anthropomorphic and therianthropic ivory figurines from different caves of the Swabian Jura, in Germany, dated to about 35–37 ka (Conard 2009) (Figure 2)?

114 João Zilhão
From the ethnographic record, we also know that the visual display of objects conveying information on the personal and social identity of the indi-viduals carrying such objects is targeted at encounters with strangers or people infrequently met. There are two rather good reasons for this: firstly, without some prior experience of interaction, the meaning of the visual symbols would be opaque to the viewer; secondly, identifying one’s affiliation or identity to family and immediate acquaintances does not require material symbols (Kuhn et al. 2001). The possibility exists, therefore, that the appearance of ornaments in the archaeological record reflects the crossing of demographical thresholds, above which long-distance interaction networks involving alliance, exchange or mating were necessary.
Figure 2. Sculpted ivory figurines from the Late Aurignacian of southern Germany, dated to ~35–37 ka. Left: “Venus” pendant from Hohle Fels. Right: horse (top) and lion (bottom) from Vogelherd (photos by H. Jensen, courtesy of N. Conard, University of Tübingen)
If so, then the absence of evidence for “modern” cognition prior to about 100 ka would not be evidence for its absence among anatomically “modern” humans prior to that time; it would simply mean that, in those days, the social life of humans had not yet effected the “release from proximity” (Gamble 1998) that eventually gener-ated the need to have symbolic identities and ways of displaying it. But once we admit that the emergence of the earliest material evidence of “modern” cognition can relate to social, not biological processes, we have no choice but to ask ourselves whether the same does not apply as well to earlier humans, namely Homo erectus. Given that brains do not fossilise and that the evidence for language in Palaeolithic times is indirect, could it be that, cognitively, these earlier ancestors were also fully human, i.e. gifted with such behavioural features as language and all its palaeon-tologically invisible neurological correlates? Put another way, could it be that, as argued by Deacon (1997), language and symbolic thinking appeared in the earliest,

Chapter 6. The emergence of language, art and symbolic thinking 115
not the latest stages of the evolution of humans, but did not externalize in ways amenable to preservation in the material record of the prehistoric past until much more recently?
Given the genetic and palaeontological evidence that the European lineage leading to Neandertals had already branched off of the African stem by half a million years ago, the archaeology of the Neandertals provides the ideal test-ing ground for the different views of the emergence of language and “modern” cognition. If the “Human Revolution” is right, then neither personal ornaments nor art should be found among the Neandertals. If either is found, then the “Human Revolution” is refuted and we must look for alternative ways to explain the emergence of those behaviours in the archaeological record.
2. Neandertal-ness
Neandertals are named after a skeleton found in 1856 at the Kleine Feldhoffer cave, in the Neander valley, near Düsseldorf. Today’s scientists, however, are not the only group of people for whom “Neandertal” has a well-defined meaning. The word is also used in common language to disqualify dislikeable individuals, including political opponents. Opening any dictionary immediately brings up these alternative meanings. The Cambridge On-line, for instance, gives the follow-ing: (1) “relating to a type of primitive people who lived in Europe and Asia from about 150,000 to 30,000 years ago” (2) said “of people or beliefs very old-fashioned and not willing to change” (3) said “(of people) rude or offensive.”
In order to understand the Neandertals’ enduring bad reputation, we have to bear in mind that, in the mid-nineteenth century, Evolution was conceived in the framework of a progressivist mindset – the directional development of ever more complex and sophisticated forms of life from a simple, primitive common ancestor, with humans sitting at the top of the ladder. Evolution also implied, as Darwin eventually made explicit, that humankind had ape-like ancestors. In this context, two things were, in retrospect, entirely predictable: firstly, a predisposition to interpret any intermediate fossil forms as “part-ape/part-man” in both morphology and behaviour; secondly, because things are what they are only trough their opposition with what they are not, a predisposition to imagine the “animality” of those ancestors as consisting of features representing the exact opposite of “humanity” as Victorians perceived it.
To make things worse, progressivist preconceptions were compounded by scientific error (Trinkaus & Shipman 1993). One of the first Neandertal articulated skeletons to be found, 100 years ago now, was that from the cave site of La Chapelle-aux-Saints. The famous French physical anthropologist Marcelin Boule made a

116 João Zilhão
classical and in many respects paradigmatic description of this fossil. Unfortunately, he also mistook for the normal Neandertal condition numerous and major pathological, arthritic malformations developed late in the ontogeny of the elderly subject of his study. As a result, both popular and scientific opinion converged in considering Neandertals as a side branch, a dead-end of Evolution, both distinct from and inferior to true humans. As late as 1953, Neandertals were still portrayed as the archetypal “half-man/half-beast” of a famous Hollywood film (Figure 3).
Figure 3. The original advertisement for the 1953 movie “The Neanderthal Man” as reproduced on the cover of a recent DVD edition
In the 1960s, this prevailing view was challenged. Boule’s error was exposed, and greater emphasis was placed on the significance of skeletons found in Palestine in the 1930s. These fossils, recovered at two nearby cave sites in Mt. Carmel, seemingly displayed an intermediate anatomy, prompting suggestions that the Near East had functioned as a zone of admixture between European Neandertals and early African sapiens (McCown & Keith 1939). Moreover, there was growing recognition at this time that, archaeologically, the two groups had been doing

Chapter 6. The emergence of language, art and symbolic thinking 117
pretty much the same thing throughout the period between 100 ka and 50 ka. Their stone tools were indistinguishable, and they had buried their dead, a practice that implies world views and religious beliefs (Leakey & Lewin 1977). In sum, the two lineages behaved in ways whose level of complexity required the use of language and symbols, as should be expected from cranial capacity – Neandertal brains were in fact larger than ours.
These developments led many scholars to wonder whether Neandertals, instead of an unrelated side-branch, could have been a regional variant of a single evolving human species and, as such, the direct ancestors of today’s Europeans. In this view, called the Multiregional Hypothesis (Wolpoff 2002; Thorne & Wolpoff 2003), present racial diversity would be the outcome of a deep-rooted continuity between today’s populations and those of the remotest past. There would have been one and only one Out-of-Africa event, modern Asians and Europeans would be the descendants, through a series of convergent changes in morphology, of the first Homo erectus settlers, and features such as the big noses of Europeans would be an example of the persistence of Neandertal “blood” in living humans.
The 1980s saw the birth of an entirely new line of inquiry, human genetics, which eventually questioned this 1960s view of the Neandertals as fundamentally human. The study of variation in the mitochondrial DNA of extant people led to the conclusion that we are all very closely related, implying a very recent last common ancestor, one who would have lived in Africa some 150 ka (Cann et al. 1987). Because mtDNA is inherited through the maternal line, this genetic ances-tor was called Eve. In the Eve scenario, her children would subsequently replace the Neandertals and other coeval archaic forms of Eurasian humans, which would all become extinct without descent. This view was supported by genetic inferences derived from the fossil mtDNA successfully extracted from the original Neander-tal specimen, in 1997, followed, a decade later, by preliminary sequencing results for the entire nuclear genome of another Neandertal individual from the Croatian cave site of Vindija (Krings et al. 1997; Green et al. 2006; Noonan et al. 2006).
The weight of scientific opinion (e.g. Klein 2003) saw in these results support for the notion that the Neandertals were phylogenetically distant and belonged in an altogether different species. Unaffected by the “Human Revolution”, they must have lacked language, or have only had an exceedingly primitive version of it. Moreover, no division of labour and no form of social organization beyond that required by the group’s need to reproduce would have existed, and the so-called Neandertal burials probably were nothing more than simple body disposal without religious meaning. In these circumstances, the outcome of contact situations could only have been total replacement with no admixture. “Humans” would have seen “Neandertals” as unsuitable non-human mates, and the cognitive superiority of our ancestors meant that they would inevitably have prevailed in the competition for territory and resources.

118 João Zilhão
3. Paradigm lost
The empirical evidence generated by the last decade of research has falsified the behavioural tenets of the “Human Revolution”. Ironically, at the same time as archaeologists working in Africa were uncovering evidence supporting “modern” cognition tens of millennia prior to the Out-of-Africa of Eve’s children (and conceivably explaining it) (McBrearty & Brooks 2000), archaeologists working in Europe were also uncovering evidence that, contrary to the postulates of the “Human Revolution”, complex and sophisticated cognitive and intellectual capabilities were also apparent in the material culture of Eurasian Neandertals (d’Errico et al. 1998, 2003a; Zilhão and d’Errico 1999; Zilhão 2001, 2006a, 2006b, 2007; d’Errico 2003; Conard 2008).
For instance, excavations carried out between 1994 and 1997 in a German brown coal mine near Schöningen yielded three 400,000-year-old wooden artefacts of great significance (Thieme 1997). Long and pointed, they were made from the base of individual spruce trees, with the maximum thickness and weight at the front and long tails that taper towards the proximal end. In all these respects, they resemble the javelins of field-and-track competitions, suggesting use as projectile weapons rather than thrusting spears. They further imply that the laws of ballistics underlying the shape of modern javelins had already been mastered by the founding fathers of the Neandertal lineage.
Further evidence for sophisticated craftsmanship comes from Neumark-Nord, another German brown coal mining site. Chemical analysis of organic residue adhering to a flint flake recovered in levels dated to more than 100 ka showed it to be an extract of oak bark macerated in water, of a kind used until the ethnographic present in the tanning of hides for the manufacture of water-proof clothing and shoe wear (Meller 2003). In the 1930s, a nearby site, the Ilsenhöhle rock shelter, had already yielded a few bone awls from the time of the latest Neandertals, around 40 ka (Hülle 1977). Combined, this evidence suggested a long tradition of hide working for the manufacture of clothes and other equipment.
This should come as no surprise. Good-quality artificial insulation was a pre-requisite for survival in Ice Age central Europe, where, considering the wind-chill effect, average winter temperatures ranged between –20 and –30ºC. Thermoregulatory models (Aiello & Wheeler 2003) show that the minimum external temperature Neandertals would have been able to support if dressed in a modern business suit was –24ºC. In the absence of even such basic level of clothing, only a thickness of body fat below the skin in excess of 3 cm could have provided equivalent protection. The weight of such fat, however, would be of some 50 kg, an amount that would leave an 80 kg Neandertal very little left for muscle, bone and

Chapter 6. The emergence of language, art and symbolic thinking 119
other tissue; or, if added to the 80 kg of a lean, muscular body, would transform the average 1.65 m tall male Neandertal into the archetypal obese, unable to procure his own subsistence in a society that lacked cash, automobiles, and shopping malls. The implication is clear: like present-day subarctic peoples, Neandertals must have had good quality clothing as well as all the other gear without which survival in such environments is impossible.
Further and more telling evidence that Neandertals were quite good at Chemistry came from yet another German brown coal mining site, Königsaue. In 1963–64, salvage excavations yielded two fragments of birch bark pitch used for stone tool hafting, one of which still bore a fingerprint of the Neandertal that manipulated it. These items have since been directly dated by radiocarbon to more than 45 ka, and a study of their composition (Koller et al. 2001) showed that they are not unmodified natural products, such as the bitumen used in the Near East for the same hafting purposes since at least 200 ka. In fact, they are a synthetic raw material, the first ever in human history. They were produced through a several hour-long smouldering process requiring a strict manufacture protocol: under exclusion of oxygen, and at tightly controlled temperatures, between 340 and 400°C.
This evidence suggests that, in fact, Neandertals were cognitively as well endowed as we are. But what about their biology? Were they in fact a separate species? And was the reason for their eventual extinction somehow related to their biological separateness?
Careful consideration of the mtDNA evidence shows that such notions have little basis. Given that contemporary populations of chimpanzees are more diverse than all living humans and Neandertals put together (Gagneux et al. 1999), the parsimonious interpretation of the genetic evidence is that, by Primate standards, present-day humans are abnormally homogeneous, not that Neandertals were a different species. In fact, the most recent synthesis of the history of modern human dispersals based on mtDNA places the immigration of the bearers of the oldest extant European variants (haplotypes H, I and U) at some 30 ka (Forster 2004). Since anatomically modern people are documented in Europe since at least 40 ka (Zilhão et al. 2007), it follows that the mtDNA variants characteristic of those original settlers must be as extinct as those of the Neandertals. Obviously, this does not mean that such early European moderns belonged to a species different from our own, and one that went extinct without descent. By the same token, no such inferences can be made for the Neandertals on the basis of the absence of their mtDNA among living humans. The take-home message is that the mtDNA of present-day Europeans reflects recent demographic history, not the remote interactions (or lack thereof) between African sapiens and non-African archaics.

120 João Zilhão
In short: The pattern of mtDNA homogeneity among extant humans is consistent with a recent origin for modern humans, but it does not rule out the possibility that Neandertals and other archaic groups contributed to the gene pool of subsequent populations. That such contributions indeed occurred is otherwise apparent in the nuclear genome, where as much as 80% of its loci carry evidence of the assimilation of genetic material from non-African archaic people (Eswaran et al. 2005). One example is the microcephalin gene, involved in the control of brain size during development, whose adaptive allele, which occurs in 70% of today’s humans, seems to have introgressed from an archaic lineage, most probably the Neandertals, sometime around 37 ka (Evans et al. 2006). Most recently, the results of the Neandertal genome project also brought additional support to these findings by showing that Neandertals contributed something like 1–4% of the nuclear DNA of present-day humans, implying significant interbreeding at the time of contact (Green et al. 2010).
These developments contradict the notion that Neandertals were a different species and show that, even if they had been, they were so close that admixture at the time of contact was inevitable, and did happen. In fact, a recent study of species intersterility versus time of divergence (Holliday 2006) suggests that the whole debate concerning the taxonomic status of Neandertals is a lot like the proverbial Byzantine argument about the sex of the angels. The study shows that, among the many lineages of mammals for which fossil or molecular data are available, 1.4 million years is the minimum amount of time for reproductive separation to emerge between two branches splitting from a recent common ancestor. This minimum was observed among gazelles. Among hominins, however, the interval between generations is at least four times longer. The implication is one of no reproductive isolation between contemporary lineages of hominins separated by less than five to six million years of divergence. Such a length of time corresponds to the entire evolutionary life span of the hominin family, and is at least ten times the duration of the interval separating the Neandertal/ modern splitting event from the period of contact in Europe. By Mammal standards, therefore, Neandertals were not, and could not have been, a different biological species.
4. Paradigm found
Until about ten years ago, the presence of ornaments at late Neandertal sites was acknowledged by supporters of the “Human Revolution” but disregarded as a by-product of “imitation without understanding” of modern human behaviours observed in contact situations (Stringer & Gamble 1993). The following analogy

Chapter 6. The emergence of language, art and symbolic thinking 121
was famously proposed by a distinguished British archaeologist to make the point: “if a child puts on a string of pearls, she is probably doing this to imitate her mother, not to symbolize her wealth, emphasize her social status, or attract the opposite sex” (Mellars 1999).
Research carried out since 1998 on the Châtelperronian culture of France and northern Spain has dramatically changed the picture. At the Grotte du Renne, in France, the Châtelperronian levels yielded bone awls identical to those from the Ilsenhöle, but with three differences (d’Errico et al. 1998, 2003b): they came in larger numbers; some featured regular decorative patterns; and they were associated with body ornaments (Figure 4). These finds were published in the early 1960s (Leroi-Gourhan 1961, 1964), but their significance was impaired by doubts on the authorship of the Châtelperronian. In 1979, however, a Neandertal skeleton was found in a Châtelperronian context at the French site of St.-Césaire (Lévêque & Vandermeersch 1980), and, in 1996, the association was confirmed for the fragmentary remains recovered at the Grotte du Renne itself (Hublin et al. 1996). Eventually, it became clear that the Châtelperronian, with its suite of personal ornaments, was an independent Neandertal development predating modern human immigration by several millennia (Zilhão 2001, 2006b, 2007; d’Errico 2003). The conclusion that Neandertal society was symbolically organized is further strengthened by results from use-wear analyses of hundreds of chunks of black pigment from another and even earlier French cave site, the Pech de l’Azé. These analyses concluded that they were pencils used for body painting (Soressi & d’Errico 2007).
a bc
d
1 cm
Figure 4. Pierced and grooved pendants made of animal bone and teeth, the most common personal ornaments of Europe’s late Neandertals, all from basal Châtelperronian level X of the Grotte du Renne (Arcy-sur-Cure, France): (a-b) fox canines; (c) bison incisor; (d) lateral phalange of reindeer
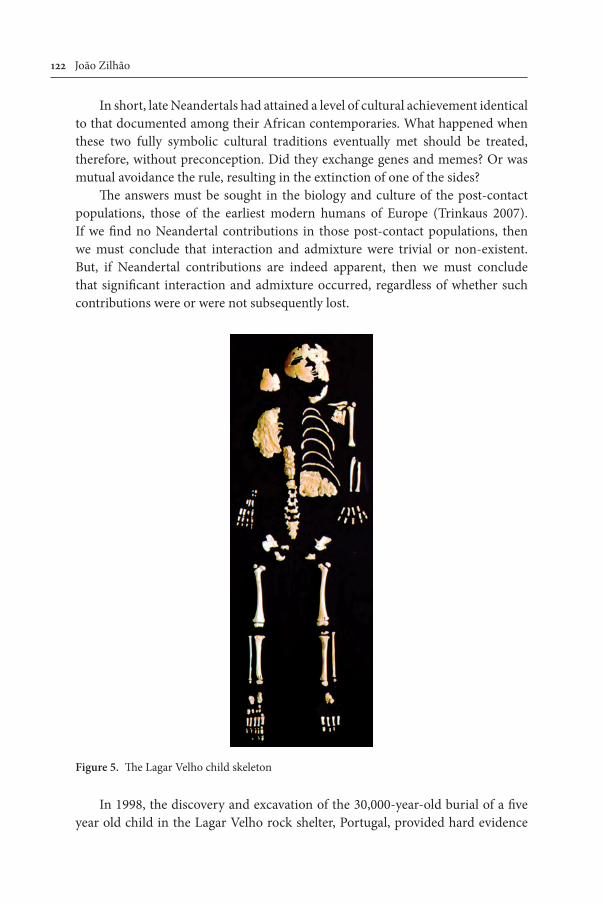
122 João Zilhão
In short, late Neandertals had attained a level of cultural achievement identical to that documented among their African contemporaries. What happened when these two fully symbolic cultural traditions eventually met should be treated, therefore, without preconception. Did they exchange genes and memes? Or was mutual avoidance the rule, resulting in the extinction of one of the sides?
The answers must be sought in the biology and culture of the post-contact populations, those of the earliest modern humans of Europe (Trinkaus 2007). If we find no Neandertal contributions in those post-contact populations, then we must conclude that interaction and admixture were trivial or non-existent. But, if Neandertal contributions are indeed apparent, then we must conclude that significant interaction and admixture occurred, regardless of whether such contributions were or were not subsequently lost.
Figure 5. The Lagar Velho child skeleton
In 1998, the discovery and excavation of the 30,000-year-old burial of a five year old child in the Lagar Velho rock shelter, Portugal, provided hard evidence

Chapter 6. The emergence of language, art and symbolic thinking 123
for a hypothesis that a group of scholars had been entertaining for many years: that populations of the African lineage spreading into Europe would have inter-bred with the local Neandertals, whose disappearance would have been largely a process of assimilation, not extinction without descent (Zilhão & Trinkaus 2002). In fact, this child (Figure 5) featured an anatomical mosaic mixing characteris-tics for the most part modern, such as a clear, prominent chin and a high cranial vault, with characteristics reminiscent or even distinctive of the Neandertals and other archaic Eurasian populations, such as the robusticity of the leg bones, the arctic, cold-adapted body proportions, and several minor features in the skull, the mandible and the dentition. These features are known to be genetically inherited, so their presence indicates a part-Neandertal ancestry for the child. Soon after the Lagar Velho discovery, in 2003–2005, the Romanian cave of Oase was to pro-vide additional evidence – the mandible of a young adult and the near complete cranium of an adolescent, dated to 40 ka and at present Europe’s earliest modern human fossils (Trinkaus et al. 2003; Rougier et al. 2007) – in support of this notion (Figure 6).
The archaeological evidence supports this scenario. The Protoaurignacian culture of western and central Europe is contemporary with the Oase fossils and, as such, the first cultural entity that reliably can be assigned to European early moderns. The personal ornaments of the Protoaurignacian are consistent with this notion. For the most part, they are made of the same small marine shell beads of diverse taxonomy but identical basket-shaped morphology found among modern human cultures of the Near East and Africa, where they go back to some 100 ka (cf. Figure 1). By comparison with these cultures, however, the Protoaurignacian also features some novelties, such as pierced animal teeth, namely of red deer and fox. These kinds of pendants are completely unknown in Africa and the Near East prior to the time of contact. But they are precisely the types of ornaments that characterize such pre-contact Neandertal-associated cultures as the Châtelperronian (cf. Figure 4). The parsimonious explanation for these elements of discontinuity with the African/Levantine tradition of personal ornamentation can only be that modern humans acquired them from the indigenous, Neandertal societies where such novel elements originally emerged (Zilhão 2007).
5. Conclusion
The implication of these finds is that, in their strict, original formulations, Multiregional Evolution and Mitochondrial Eve are now both obsolete views of the tempo and mode of human evolution. The palaeontological and archaeological evidence favours Assimilation models and refutes the notion that
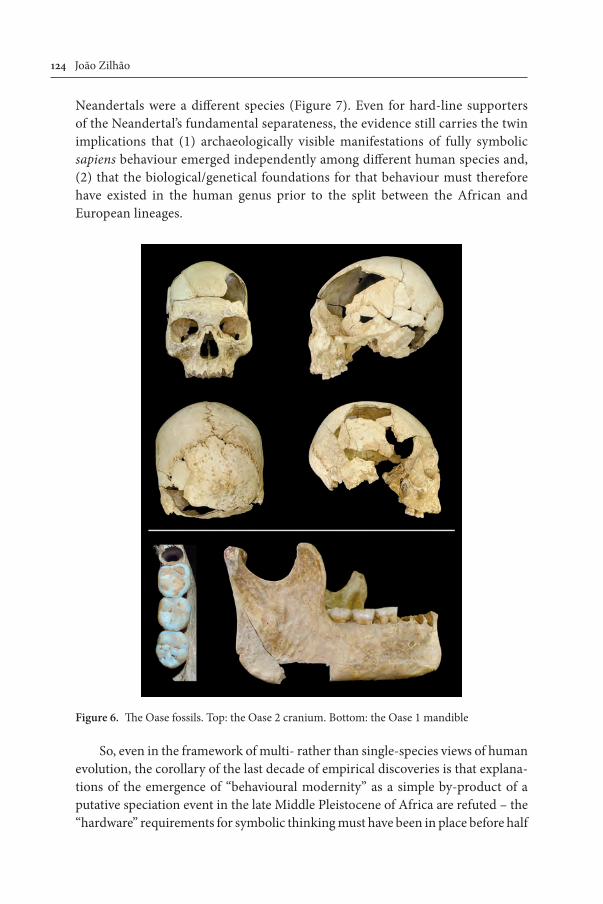
124 João Zilhão
Neandertals were a different species (Figure 7). Even for hard-line supporters of the Neandertal’s fundamental separateness, the evidence still carries the twin implications that (1) archaeologically visible manifestations of fully symbolic sapiens behaviour emerged independently among different human species and, (2) that the biological/genetical foundations for that behaviour must therefore have existed in the human genus prior to the split between the African and European lineages.
Figure 6. The Oase fossils. Top: the Oase 2 cranium. Bottom: the Oase 1 mandible
So, even in the framework of multi- rather than single-species views of human evolution, the corollary of the last decade of empirical discoveries is that explana-tions of the emergence of “behavioural modernity” as a simple by-product of a putative speciation event in the late Middle Pleistocene of Africa are refuted – the “hardware” requirements for symbolic thinking must have been in place before half
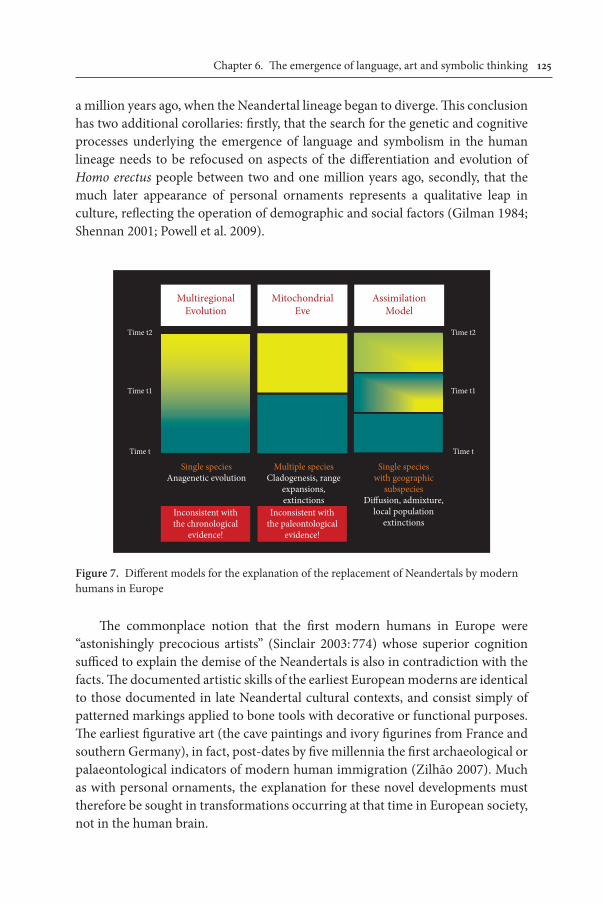
Chapter 6. The emergence of language, art and symbolic thinking 125
a million years ago, when the Neandertal lineage began to diverge. This conclusion has two additional corollaries: firstly, that the search for the genetic and cognitive processes underlying the emergence of language and symbolism in the human lineage needs to be refocused on aspects of the differentiation and evolution of Homo erectus people between two and one million years ago, secondly, that the much later appearance of personal ornaments represents a qualitative leap in culture, reflecting the operation of demographic and social factors (Gilman 1984; Shennan 2001; Powell et al. 2009).
Single specieswith geographic
subspeciesDi�usion, admixture,
local populationextinctions
Multiple speciesCladogenesis, range
expansions,extinctions
Single speciesAnagenetic evolution
Time t
Time t1
Time t2
Time t
Time t1
Time t2
Inconsistent withthe chronological
evidence!
Inconsistent withthe paleontological
evidence!
MultiregionalEvolution
MitochondrialEve
AssimilationModel
Figure 7. Different models for the explanation of the replacement of Neandertals by modern humans in Europe
The commonplace notion that the first modern humans in Europe were “astonishingly precocious artists” (Sinclair 2003: 774) whose superior cognition sufficed to explain the demise of the Neandertals is also in contradiction with the facts. The documented artistic skills of the earliest European moderns are identical to those documented in late Neandertal cultural contexts, and consist simply of patterned markings applied to bone tools with decorative or functional purposes. The earliest figurative art (the cave paintings and ivory figurines from France and southern Germany), in fact, post-dates by five millennia the first archaeological or palaeontological indicators of modern human immigration (Zilhão 2007). Much as with personal ornaments, the explanation for these novel developments must therefore be sought in transformations occurring at that time in European society, not in the human brain.

126 João Zilhão
The ethnographic record abundantly documents that rock art primarily functions as a way of embodying places with economic, ideological or social significance, and the thousands of open air petroglyphs of the Côa Valley, in Portugal (Zilhão et al. 1997; Baptista 2009), show that the same holds true for the Palaeolithic period (Figure 8).
Figure 8. Two examples of the 17 km-long Palaeolithic rock art complex of the Côa Valley, Portugal: top, Rock no. 6 of Penascosa, bottom, Rock no. 1 of Canada do Inferno (after Baptista 2009, photos by P. Guimarães, courtesy of Parque Arqueológico do Vale do Côa)

Chapter 6. The emergence of language, art and symbolic thinking 127
Thus, the parsimonious explanation for why art only appears in the archaeo-logical record around 35 ka is that only then did the need arise for systems of social identification/differentiation extending beyond the individual to include the land-scapes and resources claimed as territory by the different groups to whom people advertised their allegiance through the use of body ornaments. Sculpted figurines, in turn, are likely to have represented manifestations of the same phenomenon in the personal and domestic arenas of behaviour.
The need for such systems can easily be explained as a consequence of adaptive success, with technological innovation leading to demographic growth and implying both increased inter-group competition and increased regulation of that competition. In such a context, it is easy to understand the adaptive value of the emergence of ceremonial behaviours addressing issues of property and rights over resources, and of the development of myths and religious beliefs relating such rights to real or ideal ancestors. Therein lies the origins of art, not in an evolutionarily late mutation endowing modern humans with the capacity for symbolic thinking. The corresponding “hardware”, in fact, must have been in place as soon as the size and shape of the brain case entered modern ranges of variation and the cultural record documents behaviours that require language, i.e. symbolic thinking by definition. The palaeontological record concurs in suggesting that such a Rubicon had already been crossed by half a million years ago. The rest is History.
References
Aiello, L. & Wheeler, P. 2003. “Neandertal Thermoregulation and the Glacial Climate.” In Neanderthals and modern humans in the European landscape during the last glaciation, T. H. van Andel & W. Davies (eds), 147–166. Cambridge: McDonald Institute for Archaeo-logical Research.
Baptista, A.M. 2009. O Paradigma Perdido. O Vale do Côa e a Arte Paleolítica de Ar Livre em Portugal. Vila Nova de Foz Côa/Porto, Parque Arqueológico do Vale do Côa/Edições Afrontamento.
Bermúdez De Castro, J.M., Martinón-Torres, M., Carbonell, E., Sarmiento, S., Rosas, A., Van Der Made, J. & Lozano, M. 2004. “The Atapuerca Sites and Their Contribution to the Knowledge of Human Evolution in Europe.” Evolutionary Anthropology 13: 25–41.
Bouzouggar, A., Barton, N., Vanhaeren, M., d’Errico, F., Collcutt, S., Higham, T., Hodge, E., Parfitt, S., Rhodes, E., Schwenninger, J.-L., Stringer, C., Turner, E., Ward, S., Moutmir, A. & Stambouli, A. 2007. “82,000-year-old shell beads from North Africa and implications for the origins of modern human behavior.” Proceedings of the National Academy of Sciences 104: 9964–9969.
Cann, R.L., Stoneking, M. & Wilson, A.C. 1987. “Mitochondrial DNA and human evolution.” Nature 325: 31–36.

128 João Zilhão
Carbonell, E., Bermúdez De Castro, J.M., Parés, J.M., Pérez-González, A., Cuenca-Bescós, G., Ollé, A., Mosquera, M., Huguet, R., Van Der Made, J., Rosas, A., Sala, R., Vallverdú, J., García, N., Granger, D.E., Martinon-Torres, M., Rodríguez, X.P., Stock, G.M., Vergès, J.M., Allué, E., Burjachs, F., Caceres, I., Canals, A., Benito, A., Diez, C., Lozano, M., Mateos, A., Navazo, M., Rodríguez, J., Rosell, J. & Arsuaga, J.L. 2008. “The first hominin of Europe.” Nature 452: 465–470.
Clottes, J., Chauvet, J. -M., Brunel -Deschamps, E., Hillaire, Ch., Daugas, J. -P., Arnold, M., Cachier, H., Evin, J., Fortin, Ph., Oberlin, Ch., Tisnerat, N. & Valladas, H. 1995. “Les peintures paléolithiques de la Grotte Chauvet -Pont d’Arc (Ardèche, France): datations directes et indirectes par la méthode du radiocarbone.” Comptes Rendus de l’Académie des Sciences de Paris 320, IIa:1133 -1140.
Conard, N.J. 2008. “A Critical View of the Evidence for a Southern African Origin of Behavioural Modernity.” South African Archaeological Society Goodwin Series 10: 175–179.
Conard, N.J. 2009. “A female figurine from the basal Aurignacian of Hohle Fels Cave in southwestern Germany.” Nature 459: 248–252.
Deacon, T. 1997. The Symbolic Species: The Coevolution of Language and the Brain New York: W. W. Norton & Co.
d’ Errico, F. 2003. “The Invisible Frontier. A Multiple Species Model for the Origin of Behavioral Modernity.” Evolutionary Anthropology 12: 188–202.
d’Errico, F., Zilhão, J., Baffier, D., Julien, M. & Pelegrin, J. 1998. “Neanderthal Acculturation in Western Europe? A Critical Review of the Evidence and Its Interpretation.” Current Anthro-pology 39: 1 -44.
d’Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Tillier, A.-M., Soressi, M., Bresson, F., Maureille, B., Nowell, A., Lakarra, J., Backwell, L. & Julien, M. 2003a. “Archaeological Evidence for the Emergence of Language, Symbolism, and Music -An Alternative Multidis-ciplinary Perspective.” Journal of World Prehistory 17 (1): 1–70.
d’Errico, F., Julien, M., Liolios, D., Vanhaeren, M. & Baffier, D. 2003b. “Many awls in our argu-ment. Bone tool manufacture and use in the Châtelperronian and Aurignacian levels of the Grotte du Renne at Arcy-sur-Cure.” In The Chronology of the Aurignacian and of the Transitional Technocomplexes. Dating, Stratigraphies, Cultural Implications, J. Zilhão and F. d’Errico (eds), 247–270. Lisboa: Trabalhos de Arqueologia 33, Instituto Português de Arqueologia.
d’Errico, F., Henshilwood, C., Vanhaeren, M. & Van Niekerk, K. 2005. “Nassarius kraussianus shell beads from Blombos Cave: evidence for symbolic behaviour in the Middle Stone Age.” Journal of Human Evolution 48: 3–24.
Dennell, R. & Roebroeks, W. 2005. “An Asian perspective on early human dispersal from Africa.” Nature 438: 1099–1104.
Eswaran, V., Harpending, H. & Rogers, A.R. 2005. “Genomics refutes an exclusively African origin of humans.” Journal of Human Evolution 49 (1): 1–18.
Evans, P.D., Mekel-Bobrov, N., Vallender, E.J., Hudson, R.R. & Lahn, B.T. 2006. “Evidence that the adaptive allele of the brain size gene microcephalin introgressed into Homo sapiens from an archaic Homo lineage.” Proceedings of the National Academy of Sciences 103 (48): 18178–18183.
Forster, P. 2004. “Ice Ages & the mitochondrial DNA chronology of human dispersals: a review.” Philosophical Transactions of the Royal Society London B 359: 255–264.

Chapter 6. The emergence of language, art and symbolic thinking 129
Gagneux, P., Wills, C., Gerloff, U., Tautz, D., Morin, P.A., Boesch, C., Fruth, B., Hohmann, G., Ryder, O.A. & Woodruff, D.S. 1999. “Mitochondrial sequences show diverse evolutionary histories of African hominoids.” Proceedings of the National Academy of Sciences 96: 5077–5082.
Gamble, C. 1998. “Palaeolithic Society and the Release from Proximity: A Network Approach to Intimate Relations.” World Archaeology 29 (3): 426–449.
Gilman, A. 1984. “Explaining the Upper Palaeolithic Revolution.” In Marxist Perspectives in Archaeology, M. Spriggs (ed.), 115 -126. Cambridge: Cambridge University Press.
Green, R.E., Krause, J., Ptak, S.E., Briggs, A.W., Ronan, M.T., Simons, J.F., Du, L., Egholm, M., Rothberg, J.M., Paunovic, M. & Pääbo, S. 2006. “Analysis of one million base pairs of Neanderthal DNA.” Nature 444: 330–336.
Green, R.E., Krause, J., Briggs, A.W., Maricic, T., Stenzel, U., Kircher, M., Patterson, N., Li, H., Zhai, W., Fritz, M. H.-Y., Hansen, N.F., Durand, E.Y., Malaspinas, A.-S., Jensen, J., Marques-Bonet, T., Alkan, C., Prüfer, K., Meyer, M., Burbano, H.A., Good, J.M., Schultz, R., Aximu-Petri, A., Butthof, A., Höber, B., Höffner, B., Siegemund, M., Weihmann, A., Nusbaum, C., Lander, E.S., Russ, C., Novod, N., Affourtit, J., Egholm, M., Verna, C., Rudan, P., Brajkovic, D., Kucan, Ž., Gušic, I., Doronichev, V.B., Golovanova, L.V., Lalueza-Fox, C., De La Rasilla, M., Fortea, J., Rosas, A., Schmitz, R.W., Johnson, P.L. F., Eichler, E.E., Falush, D., Birney, E., Mullikin, J.C., Slatkin, M., Nielsen, R., Kelso, J., Lachmann, M., Reich, D. & Pääbo, S. 2010. “A Draft Sequence of the Neandertal Genome.” Science 328: 710–722.
Henshilwood, C. & Marean, C. 2003. “The Origin of Modern Human Behavior. Critique of the Models and Their Test Implications.” Current Anthropology 44 (5): 627–651.
Holliday, T.W. 2006. “Neanderthals and modern humans: an example of a mammalian syngameon?” In Neanderthals Revisited: New Approaches and Perspectives, K. Harvati and T. Harrison (eds), 289–306. New York: Springer.
Hublin, J.-J., Spoor, F., Braun, M., Zonneveld, F. & Condemi, S. 1996. “A late Neanderthal associated with Upper Palaeolithic artefacts.” Nature 381: 224–226.
Hülle, W.M. 1977. Die Ilsenhöhle unter Burg Ranis/Thüringen. Eine paläolitische Jägerstation. Stuttgart: Gustav Fischer.
Klein, R.G. 2003. “Whither the Neanderthals.” Science 299: 1525–1527.Koller, J., Baumer, U. & Mania, D. 2001. “High-tech in the middle Palaeolithic: Neandertal-
manufactured pitch identified.” European Journal of Archaeology 4 (3): 385–397.Krings, M., Stone, A., Schmitz, R.W., Krainitzki, H., Stoneking, M. & Pääbo, S. 1997. “Neandertal
DNA Sequences and the Origin of Modern Humans.” Cell 90: 19 -30.Kuhn, S.L., Stiner, M.C., Reese, D.S. & Güleç, E. 2001. “Ornaments of the earliest Upper
Paleolithic: New insights from the Levant.” Proceedings of the National Academy of Sciences 98 (13): 7641–7646.
Lahr, M.M. & Foley, R. 1998. “Towards a Theory of Modern Human Origins: Geography, Demog-raphy, and Diversity in Recent Human Evolution.” Yearbook of Physical Anthropology 41: 37–176.
Leakey, R. & Lewin, R. 1977. Origins. London: Mcdonald and Jane’s.Lee, S.-H. & Wolpoff, M.H. 2003. “The pattern of evolution in Pleistocene human brain size.”
Paleobiology 29 (2): 186–196.Leroi-Gourhan, A. 1961. “ Les fouilles d’Arcy-sur-Cure.” Gallia Préhistoire 4: 3–16.Leroi-Gourhan, A. 1964. Les religions de la Préhistoire. Paris: Presses Universitaires de France.

130 João Zilhão
Lévêque, F. & Vandermeersch B. 1980. “Découverte de restes humains dans un niveau castelperronien à Saint-Césaire (Charente-Maritime).” Comptes rendus de l’Académie des Sciences de Paris 291D: 187–189.
McBrearty, S. & Brooks, A. 2000. “The revolution that wasn’t: a new interpretation of the origin of modern human behavior.” Journal of Human Evolution 39: 453–563.
McCown, T.D. & Keith, A. 1939. The Stone Age of Mount Carmel. Vol. 2. The fossil human remains from the Levalloiso-Mousterian. Oxford: Clarendon Press.
McHenry, H.M. & Coffing, K. 2000. “Australopithecus-to-Homo: Transformations in Body and Mind.” Annual Reviews of Anthropology 29: 125–146.
Mellars, P.A. & Stringer, C.B. (eds). 1989. The Human Revolution. Edinburgh: Edinburgh University Press.
Mellars, P.A. 1999. “The Neanderthal Problem Continued.” Current Anthropology 40 (3): 341–350.Mellars, P. Boyle, K., Bar-Yosef, O. & Stringer, C. (eds). 2007. Rethinking the Human Revolution.
Cambridge: McDonald Institute for Archaeological Research.Meller, H. 2003. Geisteskraft. Alt- und Mittelpaläolithikum. Haale (Saale): Landesmuseum für
Vorgeschichte.Noonan, J.P., Coop, G., Kudaravalli, S., Smith, D., Krause, J., Alessi, J., Chen, F., Platt, D., Pääbo,
S., Pritchard, J.K. & Rubin, E.M. 2006. “Sequencing and Analysis of Neanderthal Genomic DNA.” Science 314: 1113–1118.
Powell, A., Shennan, S. & Thomas, M.G. 2009. “Late Pleistocene Demography and the Appearance of Modern Human Behavior.” Science 324: 1298–1301.
Rougier, H., Milota, S., Rodrigo, R., Gherase, M., Sarcină, L., Moldovan, O., Zilhão, J., Constantin, S., Franciscus, R.G., Zollikofer, C.P. E., Ponce De León, M. & Trinkaus, E. 2007. “Peşteracu Oase 2 and the cranial morphology of early modern Europeans.” Proceedings of the National Academy of Sciences 104 (4): 1165–1170.
Shennan, S. 2001. “Demography and Cultural Innovation: a Model and its Implications for the Emergence of Modern Human Culture.” Cambridge Archaeological Journal 11 (1): 5–16.
Sinclair, A. 2003. “Art of the ancients.” Nature 426: 774–775.Soressi, M. & d’Errico, F. 2007. “Pigments, gravures, parures: les comportements symboliques
controversés des Néandertaliens.” In Les Néandertaliens. Biologie et cultures, B. van der Meersch & B. Maureille (eds), 297–309. Paris: Documents Préhistoriques (23), Éditions du CTHS.
Stringer, C. & Gamble, C. 1993. Search of the Neanderthals. London: Thames and Hudson.Templeton, A. 2002. “Out of Africa again and again.” Nature 416: 45–50.Templeton, A. 2005. “Haplotype Trees and Modern Human Origins.” Yearbook of Physical
Anthropology 48: 33–59.Thieme, H. 1997. “Lower Palaeolithic hunting spears from Germany.” Nature 385: 807–810.Thorne, A. & Wolpoff, M. 2003. “The multiregional evolution of humans.” Scientific American
Special Issue 46–53.Trinkaus, E. 2005. “Early Modern Humans.” Annual Reviews of Anthropology 34: 207–230.Trinkaus, E. 2007. “European early modern humans and the fate of the Neandertals.” Proceedings
of the National Academy of Sciences 104 (18): 7367–7372.Trinkaus, E. & Shipman, P. 1993. The Neandertals. New York: Knopf.Trinkaus, E., Moldovan, O., Milota, S., Bîlgăr, A., Sarcina, L., Athreya, S., Bailey, S.E., Rodrigo,
R., Mircea, G., Higham, Th., Bronk Ramsey, C.H. & Plicht, J.V.D. 2003. “An early modern human from the Peştera cu Oase, Romania.” Proceedings of the National Academy of Sciences 100: 11231–11236.

Chapter 6. The emergence of language, art and symbolic thinking 131
Vanhaeren, M., d’Errico, F., Stringer, C., James, S.L., Todd, J.A. & Mienis, H.K. 2006. “Middle Paleolithic Shell Beads in Israel and Algeria.” Science 312: 1785–1787.
Wolpoff, M. 2002. Human Paleontology. Ann Arbor: University of Michigan.Zilhão, J. 2001. Anatomically Archaic, Behaviorally Modern: The Last Neanderthals and Their
Destiny. Amsterdam: Stichting Nederlands Museum voor Anthropologie en Praehistoriae.Zilhão, J. 2006a. “Genes, Fossils and Culture. An Overview of the Evidence for Neandertal-
Modern Human Interaction and Admixture.” Proceedings of the Prehistoric Society 72: 1–20.Zilhão, J. 2006b. “Neandertals and Moderns Mixed, and It Matters.” Evolutionary Anthropology
15: 183–195.Zilhão, J. 2007. “The emergence of ornaments and art: an archaeological perspective on the
origins of ‘behavioural modernity.’ ” Journal of Archaeological Research 15: 1–54.Zilhão, J., Aubry, Th., Carvalho, A.F., Baptista, A.M., Gomes, M.V. & Meireles, J. 1997. “The
Rock Art of the Côa Valley (Portugal) and its Archaeological Context.” Journal of European Archaeology 5 (1): 7 -49.
Zilhão, J. & d’Errico, F. 1999. “The chronology and taphonomy of the earliest Aurignacian and its implications for the understanding of Neanderthal extinction.” Journal of World Prehistory 13 (1): 1–68.
Zilhão, J. & Trinkaus, E. (eds). 2002. Portrait of the Artist as a Child. The Gravettian Human Skeleton from the Abrigo do Lagar Velho and its Archeological Context. Lisboa: Trabalhos de Arqueologia 22, Instituto Português de Arqueologia.
Zilhão J., Trinkaus, E., Constantin, S., Milota, S., Gherase, M., Sarcina, L., Danciu, A., Rougier, H., Quilès, J. & Rodrigo, R. 2007. “The Peştera cu Oase people, Europe’s earliest mod-ern humans.” In Rethinking the Human Revolution, P. Mellars, K. Boyle, O. Bar-Yosef and C. Stringer (eds), 249–262. Cambridge: McDonald Institute for Archaeological Research.


chapter 7
The human major transition in relation to symbolic behaviour, including language, imagination, and spirituality
David Sloan WilsonDepartments of Biology and Anthropology, Binghamton University
Human evolution can be described in terms of three C’s: Cognition, Culture, and Cooperation. Cognition includes the capacity for symbolic thought that lies at the heart of both language and spirituality. Culture includes the capacity to transmit information, both horizontally and vertically, leading to cumulative behavioural change and rapid adaptation to local environments. Cooperation includes the capacity to engage in prosocial behaviours far beyond one’s circle of genealogical relatives and narrow reciprocators. The three C’s all have precursors in nonhuman species, but they are vastly elaborated in our species.
In what sequence did the 3 C’s of human evolution arise and how are they related to each other? A commonly invoked scenario is that the first step was the evolution of advanced cognition, often called “theory of mind (ToM)”, which enabled widespread cooperation and culture (e.g. Tomasello 1999). More recently, a consensus is forming around a second scenario. The first step in human evolution was a major evolutionary transition, which enabled within-group cooperation to take place much more strongly than before (e.g. Boehm 1999; Wilson 2002, 2006, 2007; Tomasello et al. 2005). The major transition took place without a prior advance in cognitive ability and was a pre-requisite for the advanced forms of human cognition that we associate with language, symbolic thought, and spirituality. Moreover, much simpler adaptations were required as prerequisites for the advanced forms. The entire package of traits that make humans so distinctive are forms of teamwork that require interactions among trustworthy social partners. The first C to evolve was cooperation and the other two C’s are forms of cooperation.
1. What is a major evolutionary transition?
Until the 1970’s, natural selection was thought to take place entirely by small mutational change – individuals from individuals. Then, the cell biologist Lynn Margulis (1970) proposed that nucleated (eukaryotic) cells are highly integrated

134 David Sloan Wilson
communities of bacterial (prokaryotic) cells-individuals from groups. Her symbiotic cell theory was regarded as radical at the time but has since become widely accepted. Cell organelles such as mitochondria and chloroplasts are clearly derived from older species of microbes that led a more autonomous existence in the deep past.
In the 1990’s, John Maynard Smith and Eors Szathmary (1995, 1999) gener-alized Margulis’s theory to include other major events during the history of life, such as the first cells, multi-cellular organisms, social insect colonies, and even the origin of life itself as groups of cooperating molecular reactions. In each case, natural selection can take place within a given unit, which tends to favour traits that humans associate with cheating, free-riding, and exploitation. Or, natural selection can take place between units, favouring traits that humans associate with cooperation and prosociality. The balance between levels of selection is not static but can itself evolve. A major transition occurs when selection between units dom-inates selection within units, causing the unit to become a functionally adaptive “superorganism.”
As an example, consider a proto-cell containing genes that can replicate independently. Some genes are “solid citizens” that participate in the economy of the whole cell, such as gathering resources or defense, all of which would be recognized in human terms as public goods. Other cells “cheat” by using the common resources of the cell to replicate faster than the “solid citizen” genes. It is difficult to describe the scenario without resorting to the human lexicon of social behaviours! The “cheat” genes have the highest relative fitness within the cell. They will increase in frequency and will ultimately replace the “solid citizen” genes unless something more can be added to the story. That “something” is natural selection among cells. Cells with a high frequency of solid citizens will survive and divide faster than cells with a high frequency of cheats. What evolves in the total population depends upon the balance between opposing levels of selection, within and among groups.
Against this background, consider the evolution of chromosomes, which binds the genes within a cell into a single structure that must replicate as a unit. The chromosome prevents differential replication within groups, causing between-group selection to become the dominant evolutionary force. Natural selection now takes place primarily on the basis of teamwork within the cell, turning the cell into a superorganism. Similar scenarios can be described for the origin of life, multicel-lular organisms, and social insect colonies.
There are three hallmarks associated with a major evolutionary transition:
1. They are rare events in the history of life. Evidently, it is not easy for between-group selection to dominate within-group selection.

Chapter 7. The human major transition in relation to symbolic behaviour 135
2. Once a major transition occurs, the new “superorganisms” become ecologically dominant and diversify into many species. For example, eusociality in insects is estimated to have originated less than 20 times, but these gave rise to many thousands of eusocial insect species that account for over half of the biomass of all insects (Holldobler & Wilson 2008).
3. The transition is never complete. Within group “cheating” is only suppressed, never entirely eliminated. Even single cells and multicellular organisms, the paradigm of internally harmonious organisms, have a disturbing number of elements that benefit at the expense of the organism (Burt & Trivers 2006).
The concept of major evolutionary transitions is one of the most important devel-opments in the history of evolutionary theory. It is so recent that the generalized version didn’t appear in book form until the 1990’s!
2. Human evolution as a major transition
Although Maynard Smith and Szathmary were bold about expanding the concept of major transitions, they were timid about applying it to human evolution, restricting themselves to the genetic basis of language. Now it appears likely that human evolution was a full-fledged major transition. The reason that we are so unique among primates is because our ancestors became the primate equivalent of a single organism or a social insect colony (Boehm 1999; Wilson 2007; Wilson et al. 2008).
Recall that the key ingredient of a major transition is the suppression of fitness differences within groups, causing between-group selection to become the primary evolutionary force. In most primate species, including our closest ances-tors, intense within-group competition limits the opportunities for cooperation among members of the group. This is in contrast to extant human hunter-gatherer societies, which are fiercely egalitarian. What accounts for this shift and when did it occur in human evolution?
Humans are incomparably better at throwing projectiles than other primates, an ability that required whole-body anatomical changes and evolved early in the hominin lineage. Although the original purpose of throwing was presumably to deter predators and competing scavengers, it could also be used to suppress bullying and other domineering behaviour within-groups (Bingham 1999). This is a specific version of a more general hypothesis of guarded egalitarianism, advanced by Boehm (1993, 1999) on the basis of the egalitarian nature of most extant hunter-gatherer societies. However it was accomplished, guarded egalitarianism provides the key ingredient of an evolutionary transition.

136 David Sloan Wilson
In retrospect, human evolution has all the hallmarks of a major transition. It was a rare event, occurring only once among primates. It had momentous consequences; cooperation enabled our ancestors to spread over the planet, eliminating other hominins and many other species along the way. We also diver-sified to occupy all climatic zones and hundreds of ecological niches, although by cultural evolution rather than genetic evolution. The advent of agriculture enabled us to increase the scale of society by many orders of magnitude by a process of cultural multilevel selection (e.g. Turchin 2005). Finally, the transi-tion was not complete. Within-group selection still takes place and is merely suppressed compared to between-group selection.
3. Cognitive teamwork and simple forms that preceded more advanced forms
Physical forms of teamwork, such as hunting, gathering, defense, warfare, and childcare, have always been emphasized in scenarios of human evolution. It is somewhat less obvious that our mental capacities for symbolic thought on which language and spirituality are based, along with our capacity for cultural transmis-sion of traits, are forms of cognition cooperation that can only take place among trustworthy social partners. Moreover, the advanced forms of cognition and culture associated with modern humans required the prior evolution of much simpler adaptations. The reason that these adaptations did not evolve in other pri-mates is not because they were too complex, but because they were too helpful to be advantageous in groups compromised by within-group selection. Examples of such simple adaptations include the following.
1. Eyes as organs of communication. We are the only primate species with white sclera and other features that provide information to our social partners about direction of gaze, emotional state, and more. According to the “cooperative eye” hypothesis, this is because this degree of helpfulness was not favoured in any other primate species (reviewed by Tomasello et al. 2005).
2. Pointing. Even though pointing appears simple to us and appears early infancy, apes evidently do not point things out to each other or do anything comparable in natural environments. Apes raised in human social environments point to what they want but seldom point to help out their human social partners. What seems to be lacking is a sense of what others might want and/or the motivation to help by pointing. In another telling comparison, dogs comprehend pointing much better than wolves, even when wolves are raised in human social environments. The mental ability of dogs evidently evolved,

Chapter 7. The human major transition in relation to symbolic behaviour 137
by genetic evolution, over 100,000 years of domestication (Tomasello et al. 2005; Hare et al. 2002).
3. Shared social awareness in human infants. From an amazingly early age, human infants have an ability to adopt the perspective of others and to help by pointing, joining a task, or otherwise coordinating activities. In short, humans have instincts for teamwork that appear extremely early in life and are not manifested in apes at any age (Tomasello et al. 2005).
4. The evolution of laughter. Converging lines of evidence strongly suggest that our ancestors were laughing long before they were talking. Moreover, laughter has functions that are clearly related to teamwork, rapidly coordinating mood and facilitating playful activities during short periods of safety and satiety (Gervais & Wilson 2005).
The precursors for these relatively simple adaptations were present in our ape ancestors, and merely needed a more cooperative social environment to be favoured and elaborated by between-group selection.
4. Language, imagination, and spirituality
According to Terrence Deacon (1998) in his book The Symbolic Species, humans are unique in their capacity for symbolic thought. Yet, symbolic thought does not require a bigger brain, or even a different brain, than possessed by our closest ape relatives and even other non-human species. The reason that the capacity for symbolic thought is absent in most species is not because it is too complex, but because it isn’t adaptive. To understand the evolution of symbolic thought in humans, we must search for an environmental context that made it adaptive.
What distinguishes symbolic thought from other forms of cognition, such as associative learning? With associative learning, mental representations are learned when they correspond directly to the environment and are unlearned when the correspondence ceases (e.g. the sound of a bell when paired with food in Pavlovian learning). With symbolic thought, mental representations take on a life of their own, even when they don’t directly correspond to anything in the environment (e.g. the word and concept of “ghost”). Symbolic thought can be hugely adaptive because of the behaviours that they motivate in the real world, even though they don’t necessarily correspond directly to features of the real world.
One implication of Deacon’s thesis is that non-human species can be taught to think symbolically, more like us than their own kind. This retrospec-tive “prediction” is confirmed by examples such as Kanzi the bonobo and Alex the African Grey Parrot. The question is then to understand how the social

138 David Sloan Wilson
environment of proto-humans provided something equivalent to the lengthy training procedure that enabled Kanzi and Alex to think (and speak, in the case of Alex!) symbolically. It would need to be a cooperative social environment. Thus, Deacon’s thesis fits very nicely within the new paradigm of human evolu-tion as a major transition.
Language, imagination, and spirituality had at least three profound effects on human evolution. First, they enabled a higher degree of social control (e.g. through gossip), suppressing cheating and favouring teamwork of all kinds. Second, they provided a new source of behavioural variation, upon which natural selection can select. Third, they enabled the most successful behavioural alternatives to be transmitted, creating a new mechanism of inheritance. These abilities enabled our ancestors to achieve their ecological dominance and planetary diversity while remaining a single species.
5. Testable predictions
The scenario outlined above leads to numerous predictions that can be tested in the hominin fossil and early archeological record. The ability to throw stones requires many skeletal changes that can be dated during hominin evolution. Suppression of competition within groups includes competition among males for females, perhaps reducing the degree of sexual dimorphism (although between-group conflict can favour large males). Even the evolution of the whites of our eyes can potentially be genetically dated, once we identify the associated genes. Other testable predictions are possible with sufficient ingenuity.
References
Bingham, P.M. 1999. “Human Uniqueness: A general theory.” Quarterly Review of Biology 74: 33–169.
Boehm, C. 1993. “Egalitarian society and reverse dominance hierarchy.” Current Anthropology 34: 227–254.
Boehm, C. 1999. Hierarchy in the Forest: Egalitarianism and the Evolution of Human Altruism. Cambridge, Mass: Harvard University Press.
Burt, A., & Trivers, R. 2006. Genes in conflict. Cambridge, MA: Harvard University Press.Deacon, T.W. 1998. The Symbolic Species. New York: Norton.Gervais, M. & Wilson, D.S. 2005. “The Evolution and Functions of Laughter and Humor:
A Synthetic Approach.” Quarterly Review of Biology 80: 395–430.Hare, B., Brown, M., Williamson, C. & Tomasello, M. 2002. “The domestication of social
cognition in dogs.” Science 298: 1634–1636.Holldobler, B. & Wilson, E.O. 2008. The Superorganisms. New York: Norton.

Chapter 7. The human major transition in relation to symbolic behaviour 139
Margulis, L. 1970. Origin of Eukaryotic cells. New Haven: Yale University Press.Maynard Smith, J. & Szathmary, E. 1995. The major transitions of life. New York: W.H. Freeman.Maynard Smith, J. & Szathmary, E. 1999. The origins of life: from the birth of life to the origin of
language. Oxford: Oxford University Press.Tomasello, M. 1999. The cultural origins of human cognition. Cambridge, MA: Harvard Univer-
sity Press.Tomasello, M., Carpenter, J., Call, J., Behne, T. & Moll, H. 2005. “Understanding and sharing
intentions: the origins of cultural cognition.” Behavioral and Brain Sciences 28: 675–735.Turchin, P. 2005. War and Peace and War. New York: Pi Press.Wilson, D.S. 2002. Darwin’s Cathedral: Evolution, Religion and the Nature of Society. Chicago:
University of Chicago Press.Wilson, D.S. 2006. “Human groups as adaptive units: toward a permanent concensus.” In The
Innate Mind: Culture and Cognition, P. Carruthers, S. Laurence and S. Stich (eds), 78–90. Oxford: Oxford University Press.
Wilson, D.S. 2007. Evolution for Everyone: How Darwin’s Theory Can Change the Way We Think About Our Lives. New York: Delacorte.
Wilson, D.S., Van Vugt, M., & O’Gorman, R. 2008. “Multilevel selection and major evolutionary transitions: implications for psychological science.” Current Directions in Psychological Science 17: 6–9.


chapter 8
The living as symbols, the dead as symbolsProblematising the scale and pace of hominin symbolic evolution
Paul PettittDepartment of Archaeology, University of Sheffield
The ‘symbolic capacity’ has come to be seen as a core trait of anatomically modern humans, and probably separates them cognitively eand behaviourally from all other hominins. While archaeologists agree on what aspects of the archaeological record constitute evidence of symbolism, such as burials, use of pigments, and personal ornamentation, only generic concepts of ‘symbolism’ are invoked from these, resulting in a simplistic discourse about its origins. I try to problematise the concept of symbolism, using these archaeological categories, breaking each down into differing levels of symbolic sophistication. Following this, I try to link these to Dunbar’s levels of intention, and explore how one might identify these from the archaeological record. I conclude by making a necessarily coarse comparison of Neandertals and modern humans in terms of the expression of these characteristics.
1. Introduction: Living symbols, dead symbols
Palaeoanthropology is unique in providing insights into the long-term evolution of human behaviour. To a certain extent it should also provide unique insights into the cognitive capacities which underlie and facilitate certain behaviours. It should, at least in theory, provide middle range bridging between the modern human mind and that of our closest evolutionary relatives, the great apes. From what scholars can tell, a vast cognitive gulf separates the two; and at the heart of this difference apparently lies symbolism. If one accepts in a broad sense that religion is itself symbolic, then it is clear that a symbolic capacity and religious imperative is a fundamental part – perhaps an inevitability (Boyer 2008) – of being human. It is no surprise, therefore, that documenting the emergence of symbolism has in recent years become central to palaeoanthropology. One should add it to the major events of human social development that traditionally involved broad spectrum economies, agriculture, writing and state societies. In fact, it would be difficult to conceive of these without symbolic underpinning.

142 Paul Pettitt
Archaeologists have, however, approached the archaeological record in rela-tively simple ways, focussing on simple objects as being simply indicative of sym-bolism. Thus, the recovery of a used fragment of ochre becomes proxy evidence of symbolism. Such an approach does not take us very far, and certainly does not allow us to explore nuanced cognitive development among the later homini-nae. We should think of symbols not as just material cultural objects or things in the mind, but as ways of engaging with the world. Following on from Piercean semiotics, it becomes clear that what potentially makes signs symbolic is not necessarily inherent in the object itself, but is derived from its context, particularly how the sign and its signifier are regarded in relation (Schults 2009). Thus context is all-important, and, as Sloane Wilson (this volume) has noted, to understand the development of symbolism we must search for the context that made it adaptive. Here, I try to widen a contextual approach to the long-term evolutionary emer-gence of hominin symbol use, deconstructing what we mean by symbolism.
2. Problematising the archaeological debate: Symbolic revolutions that were or were not
The emergence of ‘symbolism’ over the course of hominin evolution has in the past few years become arguably the most important object of archaeological study in the quest for defining what makes us ‘behaviourally modern’ humans. Palaeolithic archaeologists seem to agree that the ‘symbolic capacity’ or ‘symbolically-mediated behaviour’ is a defining – perhaps the defining – behavioural trait of Homo sapiens, but debate has begun in the last few years as to whether ‘pre-modern’ hominins pos-sessed symbolic capacities and, if so, to what extent (see, for example, Mellars 1991, 1995; McBrearty & Brooks 2000; Wadley 2001; Henshilwood & Marean 2003; d’Errico et al. 2003 and particularly d’Errico 2003). Despite this, it is surprising that there is still no agreement on a definition of symbolism (Wadley, this volume). Instead, the debate so far has often centred upon a ‘trait list’ of behaviours that appear in the late Middle Pleistocene and Upper Pleistocene that were apparently novel to the hominin repertoire, although, as d’Errico (2003: 199) has succinctly observed, these traits “are no more than a list of the major archaeological features that characterise the Upper Palaeolithic in Europe”. Similarly, Henshilwood and Marean (2003) have rightly emphasised that the trait list is Eurocentric in origin and thus of questionable relevance to behaviours which apparently emerged first elsewhere, and have conse-quently proposed new traits with which to distinguish behavioural ‘modernity’. This has given the debate a new, at present Afrocentric, bias.
Whether African, European or, for that matter, Asian (a land mass far larger than Europe and the archaeologically explored areas of Africa put together, for

Chapter 8. The living as symbols, the dead as symbols 143
which relatively little is known) (Dennell 2008), implicit in all perspectives is the progressivist notion of an accumulation of novel behaviours that include symbolism. As with other aspects of the palaeoanthropological record (notably the geographical dispersal of hominins), archaeologists tend to assume that from their point of emergence, hominins and behaviours were present continuously from then on; dots signifying new behaviours are placed on time charts and the dots are then joined up, creating an impression of gradually increasing behavioural complexity. This is apparent, for example, in McBrearty and Brooks’ (2000) reorientation of the development of behavioural modernity to Africa and to a long-term gradualism. They see a “fitful expansion” that was “built incrementally” (ibid.: 531) over the long duration “since at least 250 ka” (ibid.: 532). With regard to symbolism, they discuss: special treatment of the dead (no evidence before the Upper Pleistocene and, in fact, no uncontroversial evidence for the African Middle Stone Age (MSA)); beads and ornaments (again no evidence before the Upper Pleistocene – see discussion below); and use of pigments (no figurative art until the Later Stone Age (LSA), the earliest date usually quoted being 26–28 ka BP for the Apollo 11 Cave plaques which are not reliably dated and may be much younger; but recovery of pigments from various African MSA sites spans the last 250,000 years). Leaving aside controversially dated examples, the evidence for early use of pigments comprises less than one dozen sites, far lower than the number of European Middle Palaeolithic sites with the same.
Of the African examples, however, a small group of sites cluster around 100 ka BP, a handful of other sites may furnish much older examples, and others post-date 80 ka BP. The earliest manifestations are few and are hampered by imprecise dating. ‘Red stained earth’, numerous haematite fragments, ochre and grinding stones occur in a context older than 250 ka BP at Kapthurin; natural stone balls coloured with manganese were recovered from Olorgesailie site BOK1 dating to >340 ka BP; and at site GOK1 a stone block coloured with ochre and bearing grinding marks was recovered from a context dating to 290–493 ka BP (Brooks & Yellen 2009). At least 150,000 years later than this group, a second comprises red ochre fragments at Klasies River Mouth >100 ka (although the majority of pieces are younger than 80 ka); haematite pencils “throughout the MSA sequence at Border Cave” (McBrearty & Brooks 2000: 528), the base of which has been estimated at >100 ka; grinding slabs (not pigment crayons) from Porc Epic, ESR dated to 121 ± 6 ka BP; and, most remarkable of the group by far, the numerous ochre fragments in all of the main (M1, M2 and M3) levels of Blombos Cave, South Africa, at least 18 of which bear engravings which can arguably be grouped into symbolic ‘traditions’ (Henshilwood et al. 2009). It was generally agreed in the Homo symbolicus workshop that it is difficult to interpret the Blombos examples as anything but symbolism. This may now be considered a robust archaeological record, but we may not be justified in drawing a continuous line from Kapthurin

144 Paul Pettitt
to the LSA. Well over 100,000 years separate Kapthurin from the earliest cluster of sites ~100 ka BP, and twenty thousand years or more separate this cluster from more numerous examples <80 ka BP (Henshilwood and d’Errico 2009). 240,000 years separate the use of pigments at Kapthurin and the earliest known appearance of figurative art after (possibly well after) 40,000 years in Africa. With a burgeoning number of well-excavated MSA sites, we cannot simply assume this is a factor of recovery, and I shall forward below a falsifiable hypothesis to account for this pattern.
‘Time depth’ is implicit in gradualist models of the emergence of human behaviour, but was it a significant aspect of symbolic behaviour? Perhaps we archaeologists implicitly assume it is. Once a novel behaviour appears, particularly one as profound as the symbolic capacity, it seems logically inconceivable to us that it might disappear again. Gaps in the chronological representation of these behav-iours are written off as deriving simply from the lack of excavated sites, justifying the drawing of solid lines. But why should the appearance of these behaviours have been cumulative? Broad surveys of the Eurasian Middle Palaeolithic and African MSA show how the appearance and chronological trajectories of many behaviours varied considerably, region to region (even within France, for example), often lacked clear trajectories, and in Europe did not inevitably lead to the Upper Palaeolithic. ‘Recursive change’, whereby novel traits appear in a region, rise in frequency, then disappear can be observed in the Levant and may have been widely characteristic of pre-modern behaviour (see papers in Hovers & Kuhn 2006). As d’Errico and Henshilwood (2009) have noted, there is no continuity in the expression of pigment use and personal ornamentation. As this is so, then why might a recursive nature not apply for symbolism or, for that matter, religion (Pettitt in press)? Chase (e.g. 1999, 2006) has specifically drawn attention to the recursive nature of symbolism, arguing that its manifestation should vary depending upon specific behavioural contexts, and thus that we should expect its appearance in the archaeological record to vary geographically and chronologically.
If one can challenge the ‘out of Africa 1’ model on palaeontological and archaeological grounds (Dennell & Roebroeks 2005) there is no reason why we should not be critical of the currently favoured African model for the origins of behavioural modernity (Pettitt 2007). If regional trajectories of change and ‘recursion’ are characteristic of pre-LSA/Upper Palaeolithic hunter-gatherers (see papers in Hovers & Kuhn 2006), then the African MSA may not have been so critical to the emergence of modern behaviour after all (Pettitt 2007). In this light, the multiregional, multispecies model for the emergence of modern behaviour proposed by d’Errico (2003) becomes highly feasible; ‘ “modern” [behavioural] traits may have appeared in different regions and among different groups of humans, much as happened later in history with the inventions of agriculture, writing, and state society’ (ibid. 200).

Chapter 8. The living as symbols, the dead as symbols 145
For the purposes of this paper I therefore assume a null hypothesis that there was no single centre of emergence of symbolism among hominin societies, or at least that such a single centre will not be recognisable archaeologically. Instead of trying in vain to identify origins, I shall instead attempt to deconstruct the notion of ‘symbolism’ as used by palaeoanthropologists, and suggest a more fragmented way in which it may have arisen among hominin groups, both in the long and short terms. I begin by elaborating an heuristic scheme using a relatively well documented and debated source of data – personal ornamentation – and then proceed to deconstruct another aspect of behaviour seen as behaviourally ‘mod-ern’ by many – the special treatment of the dead. Finally, I try to integrate these with Dunbar’s suggestions about the cognitive development of levels of intention over the course of hominin evolution to show how symbolism can occur at many organisational and cognitive levels.
3. Material culture symbols among the living
Symbols only function as such when both a writer and a reader are in accord. In archaeology, one tends to focus on the writer (i.e. through their non-perishable creations recovered through archaeology), and assume that all persons who came into contact with these material culture creations were informed readers (i.e. they could decode the intentional messages these were created to contain). This need not necessarily have been the case, and while an object can be considered as a symbol if it was created to function as such, it does not necessarily follow that it was widely or universally successful in that functioning. A shell pendant might therefore have functioned symbolically among the conspecifics of whichever erstwhile occupant of Blombos Cave made it around 80 ka BP (Henshilwood et al. 2004), but its status as a symbol may, or may not, have disappeared if it were viewed by other African Homo sapiens populations, or, for that matter, by an archaeologist eighty thousand years in the future. The only guide as to the efficiency of a symbol’s agency in the past might therefore be an abundance of that symbol, not only on one site, but among several sites of the same broad time period. Until we have such an archaeological record it might be rash to argue from a handful of sites of widely different ages that symbolism was widespread among groups and geographical regions, let alone endemic to the species.
This caution might be applied to a small group of artefacts that are often forwarded as potentially very early examples of symbolism. Three examples of these pierres figures are known; natural stone cobbles that fortuitously resemble the human form, a resemblance which was accentuated by restricted use of delib-erate engraving. Two derive from the Lower Palaeolithic (from the Levallois-rich site of Berekhat Ram, Israel, probably 350–500 ka BP; and from Acheulian deposits

146 Paul Pettitt
at Tan-Tan, Morocco, around 400 ka BP), and one from the Middle Palaeolithic (from the late Middle Palaeolithic cave site of La Roche Cotard, France ~32 ka [14C] BP). Respectively, these take the form of a pebble of basaltic tuff resembling a human torso and head, intentionally modified with grooves around its ‘neck’ and ‘sides’ (d’Errico & Nowell 2000); a quartzite cobble reminiscent of a human body modified with eight grooves and with the addition of red pigment (Bednarik 2003); and a flint beach cobble, around the periphery of which several flakes have been removed, through which a natural perforation runs and into which a bone splinter has been wedged – the overall effect resembling a human face (Marquet & Lorblanchet 2003). While a sample of three, widely spaced in time, is hardly grounds for robust interpretation of pierres figures as unambiguous indicators of early symbolism, we should not write them off as casual “lithic doodles,” as Dennell (2008: 285) has noted. Instead, he argues that, like the appearance of precocious lithic technologies in the Lower Palaeolithic such as end-scrapers and burins at Berekhet Ram, symbolism (and by extension, perhaps ritual) drifted in and out of use. In this case, “rather than dismissing these objects as non-symbolic that would be regarded as symbolic if found in later contexts, it might be advisable to consider instead why they are so rare, and under what circumstances they might occur” (ibid.: 285). Indeed, why are these figures not more common in the archaeological record? This cannot be due to recovery bias as one might argue for figurative art, so we may presumably conclude that their occurrence was genuinely rare, evi-dence of the regionally (or perhaps culturally) varied expression of early symbolic systems. But what kind of symbolism? The process begins with a reader, as the process is predicated on the initial recognition of the human form in a natural object. Thereafter the reader becomes the writer, making artificial modifications of the natural object to enhance its resemblance to the human form. The creative process therefore relates to the conscious removal of ambiguity in the symbol’s reading. Whether or not it was subsequently ‘read’ by its discoverer/creator alone or by others, the object must in any general use of the term be considered to be symbolic, because it carries within itself an explicit reference to the human body. But that is all; we can infer nothing further from its message: ‘I look like a human’. While we cannot, of course, rule out that the pierres figures symbolised a lot more (e.g. ‘I represent my creator, his agency in the group while he is not present, and the shared social norms that keep us together’) there is a considerable conceptual gap between the two forms of symbolism. The simplest way in which these pierres figures may have been used (if of course they were ‘used’ at all) cannot be regarded as cognitively sophisticated as the latter example. We need to deconstruct what we as archaeologists mean by symbolism, and I attempt to do so here with reference to personal ornamentation. I include pigment use in this argument, for which rea-sons should become apparent.

Chapter 8. The living as symbols, the dead as symbols 147
Whatever our opinion of the robustness of the earliest evidence for pigment use, some have seen the recovery of pigment ‘crayons’ from sites such as Twin Rivers (Zambia) and Kapthurin (Kenya) as “…convincing proof of the symbolic use of pigments…” by 200 ka years ago (d’Errico et al. 2003: 4 my emphasis). No criteria, however, have been proposed that allow us to identify from the archae-ological record exactly how pigments were used (see the useful discussion in Henshilwood et al. 2009 & Wadley 2005). The apparent selection of a small selec-tion of colours from a wider variety of those available (e.g. Barham 1998) and the selection of highly saturated reds in both South Africa (Watts 1999) and in Qafzeh Cave, Israel (Hovers et al. 2003) does at least suggest a symbolic func-tion, but the problem here is that archaeologists assume a broad interpretation of ‘symbolism’, namely that if pigments were in use, then whatever their specific use was it possessed a symbolic dimension. This is not simply a semantic problem; scales of symbolism vary from the simple to the complex, and archaeologists tend to assume only the latter. Simple ornaments, in the form of natural shells pierced for suspension, are known from secure contexts in Blombos Cave, South Africa for which a date of ~75 ka BP is usually cited (Henshilwood et al. 2004); Skhul Cave, Israel in a horizon dated to 100–135 ka BP; Qafzeh Cave, Israel around 90–100 ka BP (Vanhaeren et al. 2006); the Grotte des Pigeons, Taforalt, Morocco possibly around 82 ka BP (Bouzouggar et al. 2007, although one would like to see the OSL dates on which this is based backed up by other methods); Üçağizli Cave, Turkey around 39–41 ka BP and probably the same broad age at Ksar Akil, Lebanon (Kuhn et al. 2001); and >35 ka BP from Oued Djebbana, Algeria (Vanhaeren et al. 2006).1 Taking due consideration of chronometric imprecision, none of these need pre-date 100 ka BP. Taking the mean ages at face value, 30 000 years or more
1. The dates given are those generally cited in the literature. For Skhul, the ornaments were recovered from Layer B, for which ESR and U-series dates indicated an age range of 43–134 ka BP, ‘but recent ESR and U-series analyses, including direct dating of a [human] molar from the Skhul II skeleton, indicate ages between 100 and 135 ka’ (Vanhaeren et al. 2006, 786). There are, however, large errors associated with the coupled ESR/U-series dates from this level (including that on Skhul II at 116 +43/-24 ka BP) and the best estimate of the age of the Skhul II and IV is 98 +19/-10 ka BP (Grün et al. 2005) which could therefore be as young as 78 ka BP at 2 Sigma. Phase M1 at Blombos, from which 41 perforated tick shells derive, has been dated by OSL to 75.6 ± 3.4 ka BP and by TL to 77 ± 6 ka BP (Henshilwood et al. 2004, 304), thus could be younger than 70 ka BP at 2 Sigma. With one infinite conventional radiocarbon date for the open air site of Oued Djebbana, I consider it undated even though it has an Aterian attribution (Vanhaeren et al. 2006). Although the age of the ornaments from Üçağizli Cave are usually cited at 39–41 ka BP, the age range of five 14C dates for Layer H reported by Kuhn et al. (2001) could indicate an age as young as 35 ka BP at 2 Sigma, and ages for the relevant contexts at Ksar Akil are based on poor chronometric dates and estimated sedimentation rates.

148 Paul Pettitt
separate Blombos backwards to Skhul, and forwards to Ksar Akil and Üçağizli1. We should therefore be cautious about inferring that “the initial appearance of Upper Palaeolithic ornament technologies was essentially simultaneous on three continents” (Kuhn et al. 2001: 7641) or that these simple points in time reflect “a long-lasting and widespread beadworking tradition [that] existed in Africa and the Levant” (Vanhaeren et al. 2006: 1788). A simultaneous emergence and continuous tradition may, of course, eventually be demonstrated beyond reasonable doubt, but for now the archaeological record does not demonstrate this and, I suggest, we should conceive of a null hypothesis – there for elimination – that the appearance of traditions of personal ornamentation varied from region to region and, like the hominin populations themselves, were by no means continuous.
If discontinuity and regional variation in use was the rule, it follows that symbols need not have functioned in the same ways among different groups, or for that matter between different individuals. Assuming that pigment colourants and perforated shells and beads were used to ornament the body, one can conceive of different levels of use, from the simplest – what one might argue to be non-symbolic decoration – through to concept-mediated symbolism. Such a scheme of different symbol use need not be cumulative. I suggest, for example, that one or several of the following could be in operation at any one time or place:Decoration: the employment of colouring/ornamentation for visual effect with no associated symbolic meaning, or the uninformed reading of an otherwise sym-bolic code (‘I wear red because I like red’).Enhancement: the use of colouring/ornamentation/modification to bring out a simple (symbolic) message by enhancing existing clues (‘I wear red as I know you will read it as a sign of my strength or be impressed by it’).Accessorization: the use of colouring/ornamentation/modification to make a more subtle or specific statement than enhancement by acting as a material cultural accessory to message (‘I wear red as I know you will recognise it as the regalia of our clan and infer from it that were are culturally the same’).Full symbolism: the use of colouring/ornamentation/modification to make an explicit statement by acting as a full material cultural symbol from which a reader can decode complex messages from (‘I wear red as, like you, I am a successful hunter and have killed an adult eland; it is my right to wear this colour and I therefore command respect from all’).Time/space-factored symbolism: the incorporation of temporal and spatial dimen-sions into full symbolism, e.g. beliefs, myths and stories, object biographies and histories (‘I wear red only at a specific time, marking the time of the year when the ancestors created this land, in honour of the creation myths and to mark out that I am the bearer of this knowledge’).

Chapter 8. The living as symbols, the dead as symbols 149
A fanciful set of examples, but the problem is real: at which of these levels were the tick shells and engraved ochre fragments from Blombos Cave functioning? One, or multiple? As d’Errico and Henshilwood (this volume) have noted, pigment fragments are notoriously ambiguous as they ‘do not represent the direct outcome of past symbolic behaviours’. All one can do is apply a logical approach to intuitive interpretation. One might rule out simple decoration, given that one can observe redundancy in the selection of specific shells and creation of specific engravings (suggesting that each had specific meaning), but how might we confidently infer the full symbolism or time/space-factored symbolism usually assumed by archae-ologists from them? The problem is particularly acute on sites with the recovery of non-engraved pigment crayons. Leaving aside the often intractable arguments that pigments could be used for more prosaic purposes (well demonstrated by Wadley 2005, although cf. Watts 1999 & Hovers et al. 2003), we cannot eliminate the hypothesis that the recovery of pigment ‘crayons’ alone refers only to decoration or enhancement. On the other side of the coin of course a sceptic might argue that if one was to interpret Leonardo’s workshop on the grounds of his pigments alone, we might reduce him to decorated dunce, but one might propose that we should only confidently interpret pigment crayons alone as being indicative of full symbolism when they occur at times and places where figurative art or artificial memory systems are also known. In this case, context would be critical to the elucidation of specific symbolic systems. Interpreting the data presented by McBrearty and Brooks (2000) in this light, I suggest the following (I hope falsifiable) hypothesis for the emergence of these aspects of behavioural modernity in Africa:
1. Simple pigment crayons and pigment processing found before 100 ka BP and back, perhaps, to 250 ka BP or beyond, represent little more than decoration or enhancement. The recovery of large numbers of fragments and, particu-larly, engravings on fragments or other engraved objects in association with pigments, would allow us to reject this hypothesis.
2. The floruit of personal ornamentation after 100 ka BP, including traditions of engraved designs on ochre, suggests levels of accessorisation or full symbol-ism was in operation around 100 ka BP and after 80 ka BP.
3. True time/space-factored symbolism, in which figurative art often plays a role, did not emerge in Africa until after 40 ka years ago.
By this argument, I would also suggest that the Eurasian Middle Palaeolithic record (at least 40 Neandertal sites in Europe and a handful stretching back to the Lower Palaeolithic) (d’Errico 2003) shows that some groups of Homo neandertalensis and Homo heidelbergensis engaged in decoration or enhancement. Of interest here would be the items of personal ornamentation recovered from a Châtelperronian

150 Paul Pettitt
context in Layer X of the Grotte du Renne at Arcy-sur-Cure, given that the association of symbolic items with the body would be strong indication of symbol-ism of at least the level of enhancement (d’Errico & Henshilwood, this volume). It should at least be obvious by now that simply referring to these examples as ‘symbolic’ results in the meaningless attribution of ‘part behavioural modernity’ to the Neandertals. Such attributions take us nowhere; we need to define symbolism by deconstructing it; and we need to develop heuristic schemes that allow us to explore how we might identify levels of symbolism from the archaeological record (and, perhaps, primatological world – a difficult task, see McGrew, this volume).
4. The dead as symbols
The ‘trait list’ approach treats burial of the dead as an aspect of ‘modern’ behaviour, although proponents of the importance of burial do not define why it deserves to be on the list. How, for example, does placing a corpse in a shallow grave really differ from a female chimpanzee carrying around the body of her dead child for several days or, in one case, an entire month (e.g. Goodall 1986; Matsuzawa 2003; Pettitt 2011)? As with personal ornamentation, there is no reason why the treatment of the dead could not have differed in its symbolic function over the course of hominin evolution. Elsewhere, I argue that the social interaction of the living with the dead has a very long evolutionary history, beginning with the intellectual interest in the corpse (which I refer to as morbidity) that can be observed among extant primates. This became elaborated through the deliberate deposition (mortuary caching) of the dead at certain parts of the natural landscape, until features for caching were deliberately created (burials) and locales were given specific symbolic meaning as places for the dead (Pettitt 2011). Three examples widely separated in time and space serve as examples, the first perhaps most controversially.
The 3–3.5ma old locality AL-333/333w at Hadar (Ethiopia) lies on a steep hill slope, and yielded >200 hominin fossils, representing nine adults, two juveniles and two infants (MNI=13) assigned to Australopithecus afarensis within a small area (Aronson & Taieb 1981; Johanson et al. 1982). These stand out against a poor background of mammalian fauna at the site, and seem to have been covered by sediments fairly rapidly. The lack of palaeontology suggests that there was little activity in this point of the landscape. The site stands out from other Hadar localities as it is, as Johanson and Shreeve (1989: 87) note, “…just hominids littering a hillside”. The question as to how the hominin accumulation formed has attracted considerable debate. A dynamic event such as a flood can be ruled out on sedimentological grounds, and lack of carnivore modifications of the bones rules out predation; furthermore it is difficult to see how an entire group could

Chapter 8. The living as symbols, the dead as symbols 151
become bogged down on a wet plain to die together on a hill. To my knowledge, no one has advanced a hypothesis that sees australopithecines as the active agents of accumulation. I propose that at least thirteen dead individuals came to lie on the hill within a short space of time, because they had been deliberately placed there by their conspecifics. The locale seems to have been a relatively quiet area on a dynamic and dangerous landscape, perhaps given simple meaning as bodies could be placed in the long grass, minimizing the possibility that carnivores would scavenge from them. One needs invoke no specific meaning to this, further than the desire to protect corpses from scavenging, or even just to remove them from sight, but it is easy to see how, at some cognitive stage in hominin evolution, such places might begin to acquire meaning, and in such a case one might see this as relatively simple symbolism.
Secondly, the accumulation of the (complete) bodies of at least 32 individuals assigned to Homo heidelbergensis in the Sima de los Huesos (‘Pit of the Bones’) at Atapuerca, Spain, offers the earliest intriguing example of the use of a particular place for mortuary disposal. Between 400–500 ka BP thousands of bones accumu-lated in the 13m deep pit, mainly comprising bears (Ursus deningeri MNI=166), several felids and canids, and the hominins, the latter heavily skewed towards prime adults (Arsuaga et al. 1997; Bischoff et al. 2003). Lack of decent degrees of carnivore gnawing show that they were not responsible for the deposition of the hominin bodies, and degrees of articulation, and lack of damage and considerable mixing of the hominin bones, suggest that they are either in situ or have not moved far. The consensus seems to be that they were deliberately placed here, perhaps at the top of the shaft which may have been open to the air at the time (Arsuaga et al. 1997, 2003). The lack of any archaeology in the pit save for one Acheulian biface (ibid.) suggests, further, a non-prosaic nature of the accumulation. It is difficult to see this accumulation as anything other than the deliberate caching of the dead at this one place, and if this were so, then it must have been given specific mean-ing as a place of the dead, another example of simple association of a place in the landscape with death.
Finally, a parsimonious reading of the Middle Palaeolithic record shows that between 30–40 simple inhumations of Homo neandertalensis are known, with more inclusive estimates approaching 60 (see Pettitt 2002 and references therein). These burials, all without the inclusion of grave goods, span the period from 80–34 ka BP (possibly a little earlier), and overlap with the dates for the earliest burials of Homo sapiens at Qafzeh and Skhul in Israel and Mungo, Australia. Both young and old Neandertals were buried, and examples fall into distinct regional groups, notably SW France, Germany, and the Levant. Given the relatively low number of burials known despite a rich Middle Palaeolithic record, one should not simply conclude from this that ‘Neandertals buried their dead’; it may be more apposite

152 Paul Pettitt
to conclude that some Neandertals buried some of their dead, some of the time, (i.e. what we might call cultural variation). While it is unclear whether these simple inhumations were emplaced for prosaic reasons, the use of sites for multiple burials and the possible use of grave markers might suggest that some underlying belief accounts for the burials. The representation of multiple Neandertals among frag-mentary remains at several sites is intriguing: at least 25 individuals at Krapina; 20 at L’Hortus, France, among which young adults dominate; at least 22 at La Quina, France; seven at La Ferrassie of which two are juveniles and three foeti/neonates; at least seven at Shanidar cave, Iraq; and two in the Feldhoffer cave in the Neander Valley; and in Amud and Tabun caves, Israel. At La Ferrassie, several of the grave pits, those of children, seem to have been covered with large boulders, one of which bore ‘cup marks’ (Peyrony 1934). It is tempting to view the latter as specific grave markers, and if they are, they are at least simple symbols (message: the dead lie below here). In this light, Dunbar’s inclusion of Neandertals into the ‘fourth level intentionality’ may further support the notion that by the Late Middle Palaeolithic, at least an incipient ritual had emerged.
Three points in time, showing widely different funerary practices, which might have operated at different points on the following example scale:
1. Simple (non-symbolic) observation: little activity beyond morbidity; investiga-tion of the corpse, establishment that it is dead, and renegotiation of society now that a member has dropped out: (‘It is dead, I am confused’).
2. Emotive (non-symbolic) interaction: the living interact with the dead; their emotional response affects certain simple behaviours of disposal. (‘It is dead, I am mourning; hide the corpse away from activity’).
3. Associative (symbolic) interaction: the deceased is associated with a specific activity at a specific place; the place symbolises the dead. (‘He is dead; he must be disposed of at a recognized place’).
4. Time/Space-factored associative interaction: the agency of the dead is recog-nized in mortuary treatment (who gets special treatment, where and when) and mortuary activity is organized in time and space according to social rules. (‘He is dead; he was an elder in life and has earned the right to be buried at the place of the elders’).
One must of course remember that, presumably, most human mortuary activity in prehistory is invisible to archaeology, but this at least shows that even for archaeologically observable mortuary activity one cannot simply argue that it is ‘symbolic’ in any straightforward way. I propose that simple observation and morbidity has very deep evolutionary roots (at least back to Miocene apes); that emotive interaction with the dead might have originated in the earliest hominin

Chapter 8. The living as symbols, the dead as symbols 153
communities; that associative interaction with the dead (and thus a degree of symbolism) originated at least among Homo heidelberbergensis populations in the Middle Pleistocene and became more elaborate with Homo neandertalensis and early Homo sapiens, but that the earliest true time/space-factored associative interaction can as yet only be recognized among European Mid Upper Palaeolithic (Gravettian) burials, which represent a highly-redundant (and often pathological) sub-section of society in which social differentiation seems to have been one of the criteria governing the disposal of the dead (e.g. Zilhão & Trinkaus 2002; Zilhão 2003; Pettitt 2006; Formicola 2007).
5. The evolution of Homo symbolicus: Gradual, abrupt, or fragmentary?
Despite progressivist narratives the story of hominin evolution is in a sense largely one of failure: multiple dispersals from Africa (and one assumes elsewhere) when climatic and environmental circumstances allowed, most of which resulted in local extinctions as environments shut down in response to climatic downturns associated with Heinrich events. In the long-term context, as discussed above, behavioural repertoires of the MSA and Middle Palaeolithic waxed and waned, were situationally dependent, and, beyond drawing upon general repertoires, regionally independent (see Chase 1999, 2006 and papers in Hovers & Kuhn 2006). Such attenuated and regionally-differing dispersal and behavioural trajectories provide an appropriate context for the evolution of the symbolic capacity and its expression, which itself, I argue, should have had a recursive and interrupted developmental pattern. While I have concentrated upon specific examples to try to deconstruct what archaeologists mean by ‘symbolism’, a general evolutionary context has been provided by Dunbar, into which, I suggest, symbolic evolution might fit.
Dunbar (2003) has interpreted brain evolution in terms of intentional states – reflexive sequences of belief states – which range from one (‘I believe that…’), through typical human functioning of three, to the normal human limit of four. Although it became apparent in the Homo symbolicus workshop that there is considerable debate as to whether one can pigeonhole intentional states in this way, and whether such a classificatory system is of heuristic use for cognitive evolution, it at least forms a useful framework within which to conceptualise symbolic evolution. To Dunbar, Theory of mind, which in modern humans emerges between 4–5 years of age, requires level 2 intention (‘I believe that you believe…’). Requiring individuals to conform to social norms requires three levels of intention (‘I want you to believe that you must behave how we want’), whereas religion, at least as we conceive of it, requires level four intention (‘I have

154 Paul Pettitt
to believe that you suppose that there are supernatural beings who understand that you and I desire that things happen in a certain way’). Dunbar has suggested that levels of intentionality increased over the course of hominin evolution, and can be equated with increasing brain size, group size and grooming time. This would grant australopithecines approaching two levels of intentionality (thus a theory of mind), archaic Homo such as Homo erectus and Homo heidelbergensis three levels, and four levels to Neandertals and anatomically modern humans. In light of the latter it is perhaps not surprising that it is with both Neandertals and modern humans that burial of the dead was from time to time practised. Might Dunbar’s conclusions be relevant to the origins of symbolism?
While systems of decoration and enhancement I introduced above could function with two levels of intentionality (‘I know that you will be impressed’) ‘true’ symbolism should require three (for a full symbol to work ‘I need to know that you understand this symbol/place means this’). By extension, however, giving symbolic integration to people and places requires four (‘I need to know, that you under-stand, that this place gives meaning to this thing/person/act’). It follows that the evolution of the symbolic capacity (although not always accompanied by sym-bolic expression of that capacity) should have paralleled cognitive evolution, and it is the task of the archaeologist to identify the explicit way in which this might have occurred. In Table 1, I try to place the examples discussed above in Dunbar’s context of cognitive evolution.
Table 1. Scales of the evolution of symbolism using Dunbar’s (2003) concept of evolution of the social brain and personal ornamentation and mortuary activity as examples
Intentional level
Hominin grade
Dunbar’s example
Personal ornamentation
Mortuary activity
1 Pre-australopithecines & australopithecines
I believe that… Decoration Simple observation (I believe that you are dead)
2 Australopithecines I believe that you believe…
Enhancement (I know you will be impressed)
Emotive interaction (I empathise that you are dead) & simple mortuary caching
3 Archaic Homo I want you to believe that you must behave how we want
True symbolism (I know that you understand that this symbol means this…)
Associative symbolic caching (I know you must be deposited at a specific place)
(Continued)

Chapter 8. The living as symbols, the dead as symbols 155
Intentional level
Hominin grade
Dunbar’s example
Personal ornamentation
Mortuary activity
4 Homo neandertalensis, Homo sapiens
I have to believe that you suppose that there are supernatural beings who understand that you and I desire that things happen in a certain way
Time/space-factored symbolism (I know, that you understand, that this place gives meaning to this thing/person/act)
Time/space-factored associative symbolism (Because of your agency, you must be disposed of in this way, by this method, at this place, as recognised by our social rules)
The difficulty, of course, will be to develop specific predictions of the archaeological record that we have a chance of addressing. With an archaeological record that is still overwhelmingly poor, particularly for Africa, and largely non-existent for much of the Old World, all we can do for now is to adopt parsimony in our interpretations. Thus I suggest that, in situations where we have only pigment fragments, we need only infer decoration or enhancement; with simple burials perhaps only indicative of emotive interactions with the dead. As symbolic systems are elaborated, and in particular when they are employed in combination, one might reasonably infer more sophisticated forms of symbolism. I try to outline a set of archaeological predictions based on this notion of parsimonious interpretation in Table 2.
Table 2. Potential archaeological signatures of developing levels of symbolism, based on various combinations of pigment use, engraving, personal ornamentation, burial and art
Pigments Personal ornamentation
Figurative art
Burial Parsimonious symbolic function
Chronology
Ochre fragments/processing
Decoration Intermittent from the Middle Pleistocene
Personal ornamentation
Decoration Intermittent from >100 ka
Personal ornamentation (selection of restricted shell taxa & colouring by burning)
Enhancement, possibly accessorisation
Intermittent from >100 ka
Table 1. (Continued)
(Continued)
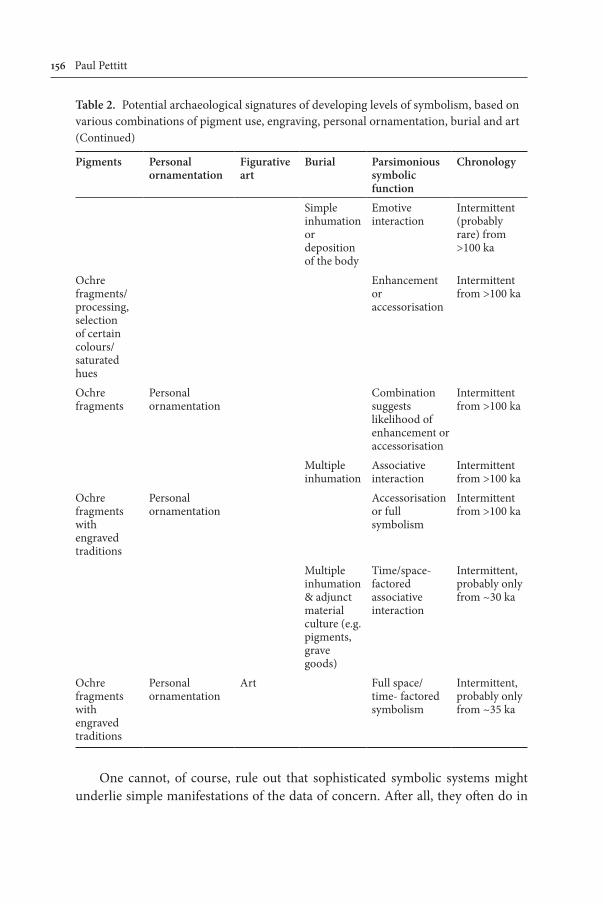
156 Paul Pettitt
Pigments Personal ornamentation
Figurative art
Burial Parsimonious symbolic function
Chronology
Simple inhumation or deposition of the body
Emotive interaction
Intermittent (probably rare) from >100 ka
Ochre fragments/processing, selection of certain colours/saturated hues
Enhancement or accessorisation
Intermittent from >100 ka
Ochre fragments
Personal ornamentation
Combination suggests likelihood of enhancement or accessorisation
Intermittent from >100 ka
Multiple inhumation
Associative interaction
Intermittent from >100 ka
Ochre fragments with engraved traditions
Personal ornamentation
Accessorisation or full symbolism
Intermittent from >100 ka
Multiple inhumation & adjunct material culture (e.g. pigments, grave goods)
Time/space-factored associative interaction
Intermittent, probably only from ~30 ka
Ochre fragments with engraved traditions
Personal ornamentation
Art Full space/time- factored symbolism
Intermittent, probably only from ~35 ka
One cannot, of course, rule out that sophisticated symbolic systems might underlie simple manifestations of the data of concern. After all, they often do in
Table 2. Potential archaeological signatures of developing levels of symbolism, based on various combinations of pigment use, engraving, personal ornamentation, burial and art (Continued)
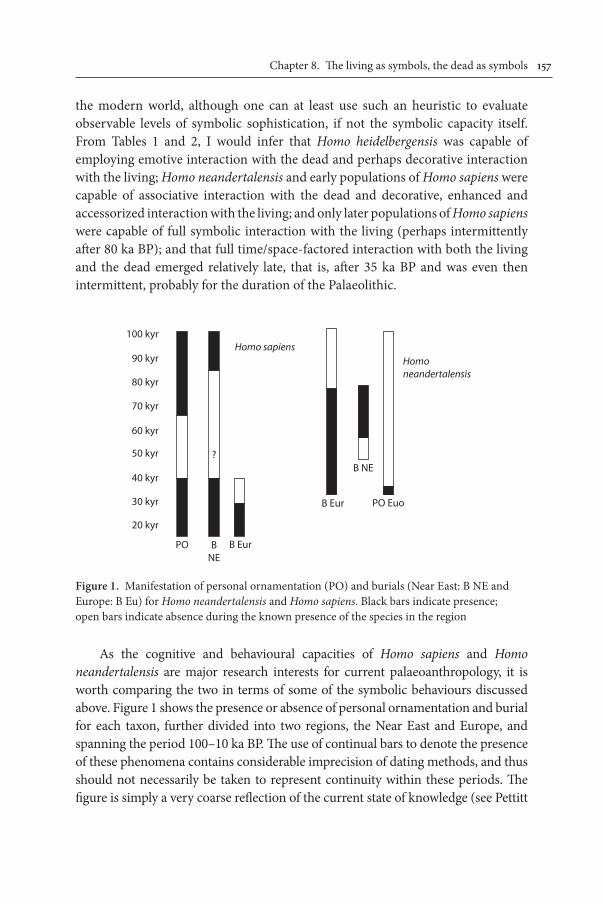
Chapter 8. The living as symbols, the dead as symbols 157
the modern world, although one can at least use such an heuristic to evaluate observable levels of symbolic sophistication, if not the symbolic capacity itself. From Tables 1 and 2, I would infer that Homo heidelbergensis was capable of employing emotive interaction with the dead and perhaps decorative interaction with the living; Homo neandertalensis and early populations of Homo sapiens were capable of associative interaction with the dead and decorative, enhanced and accessorized interaction with the living; and only later populations of Homo sapiens were capable of full symbolic interaction with the living (perhaps intermittently after 80 ka BP); and that full time/space-factored interaction with both the living and the dead emerged relatively late, that is, after 35 ka BP and was even then intermittent, probably for the duration of the Palaeolithic.
100 kyrHomo sapiens
Homoneandertalensis
PO
PO Euo
BNE
B NE
B Eur
B Eur
?
90 kyr
80 kyr
70 kyr
60 kyr
50 kyr
40 kyr
30 kyr
20 kyr
Figure 1. Manifestation of personal ornamentation (PO) and burials (Near East: B NE and Europe: B Eu) for Homo neandertalensis and Homo sapiens. Black bars indicate presence; open bars indicate absence during the known presence of the species in the region
As the cognitive and behavioural capacities of Homo sapiens and Homo neandertalensis are major research interests for current palaeoanthropology, it is worth comparing the two in terms of some of the symbolic behaviours discussed above. Figure 1 shows the presence or absence of personal ornamentation and burial for each taxon, further divided into two regions, the Near East and Europe, and spanning the period 100–10 ka BP. The use of continual bars to denote the presence of these phenomena contains considerable imprecision of dating methods, and thus should not necessarily be taken to represent continuity within these periods. The figure is simply a very coarse reflection of the current state of knowledge (see Pettitt

158 Paul Pettitt
2011 for a more contextualised discussion). A few very preliminary observations can be made from this. First, for Homo sapiens, there is a correlation between the presence/absence of burials in both Europe and the Near East and personal ornamentation; i.e. in periods where burial was practised, personal ornamentation was produced too, and this pattern holds for populations in both Europe and the Near East. The picture for the Neandertals is not as clear, although Near Eastern burials appear around the same time as those in Europe, but are truncated earlier, almost certainly because Neandertals became extinct in the region earlier than they did in Europe. A clear contrast with modern humans is the lack of personal ornamentation, the sole exception being the jewellery from the Grotte du Renne at Arcy-sur-Cure, assuming it was made by Neandertals. A final observation is that there is an inverse correlation between the two taxa: in the period that some Neandertals were burying some of their dead in Europe and the Near East, we have no examples for Homo sapiens, who are instead practising burial and personal ornamentation before and after the Neandertal practise of these phenomena. One would not yet want to place too much emphasis on this relatively poor record, but such regional trajectories may repay the effort of further study if and when the record improves.
6. Conclusion
This is by necessity a speculative paper, although I draw on the current state of knowledge for archaeological phenomena that are usually thought to be indicative of symbolic thought. It may be full of holes: the data I draw upon as examples are of course open to other interpretations; my reading of the existing archaeological record may be overly-critical, and the capacity for ‘symbolism’ may be heuristically divided in many other ways. It is also certain that the archaeological record will change as new discoveries are made, particularly in Africa and Asia. I hope at least, however, that the paper will stimulate discussion among Palaeolithic archaeologists as to how to develop our concepts of symbolism; after two decades of debate in which symbolism has emerged as the human capacity, we still use rudimentary concepts of what it is and how it is to be recognised archaeologically. These discussions began at the Cape Town workshop and, I hope, will continue to run. To be successful we have to problematise what we mean by symbolism. I’ve made a start here and throw down a friendly gauntlet.
Acknowledgements
I am much indebted to The Templeton Foundation, particularly Mary Ann Meyers, for providing the support for the Homo symbolicus symposium; to Christopher Henshilwood for being such a

Chapter 8. The living as symbols, the dead as symbols 159
pleasant and welcoming host, and Karen van Niekerk for her friendly organisation of the event. Christopher Henshilwood and Francesco d’Errico very kindly gave up time to show us Blombos Cave. The symposium provided a splendid opportunity to debate many issues ‘out of the box’ in pleasant surroundings with colleagues from very different disciplines. I very much benefited from discussion with all of the participants.
References
Aronson, J.L. & Taieb, M. 1981. “Geology and palaeography of the Hadar hominid site, Ethiopia.” In Hominid Sites: Their Geologic Settings, G. Rapp & C. F. Vondra (eds), 165–195. Westview: Boulder.
Arsuaga, J.L., Martínez, I., Gracia, A., Carretero, J.M., Lorenzo, C. & García, N. 1997. “Sima de los Huesos (Sierra de Atapuerca, Spain), the site.” Journal of Human Evolution 3: 109–127.
Arsuaga, J.L., Carbonell, E., & Bermúdez de Castro, J.M. 2003. The First Europeans: Treasures from the Hills of Atapuerca. New York: American Museum of Natural History/Junta de Castilla y León.
Barham, L. 1998. “Possible early pigment use in south-central Africa.” Current Anthropology 39: 703–710.
Bednarik, R. 2003. “A figurine from the African Acheulian.” Current Anthropology 44 (3): 405–412.Bischoff, J.L., Shamp, D.D., Aramburu, A., Arsuaga, J.L., Carbonell, E. & Bermudez de Castro, J.M.
2003. “The Sima de los Huesos hominids date to beyond U/Th equilibrium (>350Ka) and perhaps to 400–500 ka: new radiometric dates.” Journal of Archaeological Science 30: 275–280.
Boyer, P. 2008. “Religion: bound to believe?” Nature 455: 1038–1039.Bouzouggar, A., Barton, N., Vanhaeren, M., d’Errico, F., Collcutt, S., Higham, T., Hodge,
E., Parfitt, S., Rhones, E., Schwenninger, J.-L., Stringer, C., Turner, E., Ward, S., Moutmir, A. & Stambouli, A. 2007. “82,000 year-old shell beads from North Africa and implications for the origins of modern human behavior.” Proceedings of the National Academy of Sciences. 104 (24): 9964–9969.
Brooks, A. & Yellen, J. 2009. “Was Homo symbolicus also Homo sapiens? A palaeoanthropologi-cal perspective.” Pre-circulated paper for the Homo symbolicus symposium, Cape Town.
Chase, P.G. 1999. “Symbolism as reference and symbolism as culture.” In The Evolution of Culture, C. Power, R. Dunbar & C. Knight (eds), 34–49. Edinburgh: Edinburgh University Press.
Chase, P.G. 2006. The Emergence of Culture: the Evolution of a Uniquely Human Way of Life. New York: Springer.
Dennell, R. 2008. The Palaeolithic Settlement of Asia. Cambridge: Cambridge University Press.Dennell, R. & Roebroeks, W. 2005. “An Asian perspective on early human dispersal from Africa.”
Nature 438: 1099–1104.d’Errico, F. 2003. “The invisible frontier: a multiple species model for the origin of behavioural
modernity.” Evolutionary Anthropology 12: 188–202.d’Errico, F. & Nowell, A. 2000. “A new look at the Berrekhat Ram figurine: implications for the
origins of symbolism.” Cambridge Archaeological Journal 10 (1): 123–167.d’Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Tillier, A.-M., Soressi, M., Bresson,
F., Maureille, B., Nowell, A., Lakarra, J., Backwell, L. & Julien, M. 2003. “Archaeological evidence for the emergence of language, symbolism, and music – an alternative multidisci-plinary perspective.” Journal of World Prehistory 17 (1): 1–70.

160 Paul Pettitt
d’Errico, F. & Henshilwood, C. this volume. “The origin of symbolically mediated behaviour: from antagonistic scenarios to a unified research strategy.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds), 49–73. Amsterdam: John Benjamins.
Dunbar, R.I.M. 2003. “The social brain: mind, language, and society in evolutionary perspective.” Annual Review of Anthropology 32: 163–181.
Formicola, V. 2007. “From the Sunghir children to the Romito dwarf: aspects of the Upper Palaeolithic funerary landscape.” Current Anthropology 48 (3): 446–453.
Goodall, J. 1986. The Chimpanzees of Gombe: Patterns of Behaviour. Cambridge MA: Harvard University Press.
Grün, R., Stringer, C., McDermott, F., Nathan, R., Porat, M., Robertson, S., Taylor, L., Mortimer, G., Eggins, S. & McCulloch, M. 2005. “U-series and ESR analyses of bones and teeth relating to the human burials from Skhul.” Journal of Human Evolution 49 (3): 316–334.
Henshilwood, C. & Marean, C. 2003. “The origin of modern human behaviour: critique of the models and their test implications.” Current Anthropology 44 (5): 627–651.
Henshilwood, C.S., d’Errico, F., Vanhaeren, M., van Niekerk, K. & Jacobs, Z. 2004. “Middle Stone Age shell beads from South Africa.” Science 304: 404.
Henshilwood, C.S., d’Errico, F. & Watts. I. 2009. “Engraved ochres from the Middle Stone Age levels at Blombos Cave.” South Africa Journal of Human Evolution 57: 27–47.
Henshilwood, C. & d’Errico, F. this volume. “Middle Stone Age engravings and their significance to the debate on the emergence of symbolic material culture.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds), 75–96. Amsterdam: John Benjamins.
Hovers, E., Ilani, S., Bar-Yosef, O. & Vandermeersch, B. 2003. “An early case of colour symbolism: ochre use by modern humans in Qafzeh Cave.” Current Anthropology 44 (4): 491–522.
Hovers, E. & Kuhn, S. 2006. Transitions before the Transition: Evolution and Stability in the Middle Palaeolithic and Middle Stone Age. New York: Springer.
Johanson, D.C., Taieb, M. & Coppens, Y. 1982. “Pliocene hominids from the Hadar formation, Ehtiopia (1973–1977): stratigraphic, chonological, and paleoenvironmental contexts, with notes on hominid morphology and systematics.” American Journal of Physical Anthropology 57: 373–402.
Johanson. D. & Shreeve J. 1989. Lucy’s Child, The Discovery of a Human Ancestor. New York: Early Man Publishing, Inc.
Kuhn, S.L., Stiner, M.C., Reese, D.S. & Güleç, E. 2001. “Ornaments of the earliest Upper Paleolithic: New insights from the Levant.” Proceedings of the National Academy of Sciences. 98 (13): 7641–7646.
Marquet, J.-C. & Lorblanchet, M. 2003. “A Neanderthal face? The proto-figurine from La Roche-Cotard, Langeais (Indre-et-Loire, France).” Antiquity 77: 661–670.
Matsuzawa, T. 2003. Jokro: the Death of An Infant Chimpanzee (DVD film with associated leaflet). Kyoto: Primate Research Insititute.
McBrearty, S. & Brooks, A. 2000. “The revolution that wasn’t: a new interpretation of the origin of modern human behaviour.” Journal of Human Evolution 39: 453–563.
McGrew, W. this volume. “Pan symbolicus: a cultural primatologist’s view.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood & F. d’Errico (eds) 1–12. Amsterdam: John Benjamins.
Mellars, P. 1991. “Cognitive changes and the emergence of modern humans in Europe.” Cambridge Archaeological Journal (1): 63–76.

Chapter 8. The living as symbols, the dead as symbols 161
Mellars, P. 1995. The Neanderthal Legacy: An Archaeological Perspective from Western Europe. Princeton: Princeton University Press.
Pettitt, P.B. 2002. “The Neanderthal Dead: exploring mortuary variability in Middle Palaeolithic Eurasia.” Before Farming 2002/1 (http://www.waspress.co.uk/journals/beforefarming/journal_20021).
Pettitt, P.B. 2006. “The living dead and the dead living: burials, figurines and social performance in the European Mid Upper Palaeolithic.” In Social Archaeology of Funerary Remains, R. Gowland & C. Knüsel (eds), 292–308. Oxford: OxBow.
Pettitt, P.B. 2007. “Trajectories before the transition and revolutions that were or were not (lead review).” Journal of Human Evolution 53: 755–759.
Pettitt, P.B. 2011. The Palaeolithic Origins of Human Burial. Abingdon: Routledge.Pettitt, P.B. in press. “Religion and ritual in the Lower and Middle Palaeolithic.” In The Oxford
Handbook of the Archaeology of Ritual and Religion, T. Insoll and R. Maclean (eds). Oxford: Oxford University Press.
Peyrony, D. 1934. “La Ferrassie: Moustérien – Périgordien – Aurignacien.” Préhistoire 3: 1–92.Schults, F.L. 2009. Homo symbolicus: science, theology and the limits of semiotic engagement.
Pre-circulated paper for the Homo symbolicus symposium, Cape Town.Sloane Wilson, D. this volume. “The human major transition in relation to symbolic behaviour,
including language, imagination and spirituality.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds) 133–139. Amsterdam: John Benjamins.
Vanhaeren, M., d’Errico, F., Stringer, C., James, S.L., Todd, J.A. & Mienis, H.K. 2006. “Middle Palaeolithic shell beads in Israel and Algeria.” Science 312: 1185–1188.
Wadley, L. 2001. “What is cultural modernity? A general view and a South African perspective from Rose Cottage Cave.” Cambridge Archaeological Journal 11 (2): 201–221.
Wadley, L. 2005. “Putting ochre to the test: replication studies of adhesives that may have been used for hafting tools in the Middle Stone Age.” Journal of Human Evolution 49: 587–601.
Wadley, L. this volume. “Complex Cognition Required for Compound Adhesive Manufacture in the Middle Stone Age Implies Symbolic Capacity.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood & F. d’Errico (eds) 97–109. Amsterdam: John Benjamins.
Watts, I. 1999. “The origin of symbolic culture.” In The Evolution of Culture, R. Dunbar, C. Knight and C. Power (eds), 113–146. Edinburgh: Edinburgh University Press.
Zilhão, J. 2003. “Burial evidence for the social differentiation of age classes in the Early Upper Palaeolithic.” In Comportements des Hommes du Paléolithique Moyen et Supérieur en Europe: Territoires et Milieux, D. Vialou, J. Renault-Miskovsky and M. Patou-Mathis (eds), 231–241. Liège: ERAUL 111.
Zilhão, J. & Trinkaus, E. 2002. “Social implications.” In Portrait of the Artist as a Child: the Gravettian Human Skeleton from the Abrigo do Lagar Velho and its Archaeological Context, J. Zilhão & E. Trinkaus (eds), 519–541. Lisbon: Ministério da Cultura and Instituto Português de Arquelogia.


chapter 9
Biology and mechanisms related to the dawn of language
George F.R. EllisMathematics Department, University of Cape Town
The purpose of language is prediction and problem solving in the social and ecological context. Its implementation is enabled by the combination of bottom-up and top-down causation in the brain. Visual thinking develops basic abilities of pattern recognition and classification, but music and play can more effectively develop the key feature of recursion. The motivational impulse for language use starts with mother-child nurturing but moves on to the joy of imaginative play, which develops theories of other minds and basic symbolic abilities; it also easily includes music and song. Key steps towards language could be changes in neural connectivity allowing recursion to emerge, in the context of developing technological needs.
1. The functional and structural context
The emergence of language is partly a socially and environmentally based phenomenon, but equally it has a biological base. As well as social and functional needs, evolutionary arguments should take this link to biology into account. In biological terms, structure enables function, and hence study of language evolution must consider the neurological structure of the brain in relation to how it functions. Of course the nature of language itself is also central to this analysis, so key questions are what are the crucial features of language that enable its extraordinary representative and communicative abilities? And how are they realised in neurological terms?
Like all truly complex systems, language has both a modular hierarchical structure (Simon 2001; Booch 2007) and an evolutionary origin (Christiansen & Kirby 2005). It is clear that social development requires language: without communication society cannot exist. As social cooperation greatly increases survival prospects, evolutionary processes will favour capacities that lead to language development (Burling 2007). The purpose of it all is prediction and problem solving in the social and ecological context. The functional basis is the embodied brain and its biological context. Its mode is via the nature of semiotic

164 George F.R. Ellis
representation. The key functional element in representing complex aspects of reality is a modular hierarchical structure, with a class structure involving inheritance and the capacity for recursion. It is realised through grammar that is usage-based and embodied (Bergen & Chang 2003; Feldman 2006), rather than being rule-based.
The crucial biological feature whose emergence enabled the flowering of human language, separating us from all other animal species, is still unclear. A key question here is, was the advent of language essentially due to some change in brain structure, which then inevitably led to social changes? If so, what was that change? (Striedter 2005). Or was it rather basically due to ecological and social changes, which then led to changes in brain structure? If so, what were these key changes?
This chapter first looks at foundational issues underlying the discussion, and then at some evolutionary issues and possible archaeological implications. It should be emphasized that this is to some extent an incursion of mine into another field than my primary academic discipline; so the reader is asked to excuse some infelicities in presentation in exchange for some provocative ideas that may be productive.
2. The nature of language: Crucial features
Language has crucial features related both to its semiotic function and to its embodied nature.
2.1 An embodied symbolic system
Language is a symbolic system (Deacon 1997) with a semiotic function (Trask 2007): its purpose is to convey meaning, emotions, facts, and concepts in a social context through systematic use of symbols. It represents the world of objects, actions, feelings, and qualities, as well as relationships, ideas, and theories. This representational function involves naming, indexing, and use of metaphor (Trask 2007; Lakoff & Johnson 1980). Facts represented are both contingent (historical, geographical, and other specific features of the world and of narratives) and generic (universal patterns characterising the way it all works in general). The relation between these two features (concrete/specific and abstract/generic) is a key aspect of thought and of language, involving development of classes of entities and classification of specific instances.
While language has an abstract character, it is embodied via an equivalence class of physical representations; in particular it has spoken and written forms. Its existence enables the cumulative building up of understandings and ideas in

Chapter 9. Biology and mechanisms related to the dawn of language 165
individuals and in society, and (through technological means) their long-distance sharing in geographic terms as well as their storage and preservation in various media, enabling their propagation over long times. This storage of ideas in external media (ranging from diaries and memos to encyclopaedias and wikipedia) greatly facilitates our mental powers and underlies an exponential growth of knowledge (Clark 2008). Language attains its social power through enabling mediation of social interactions on a small scale, and through enabling the utility of mass media and communication systems on a large scale. These enable widespread dissemination of facts, ideas, and meanings, extending the cognitive web from local communication to a global system. Minds therefore cannot be understood on their own (Donald 2001): they are part of a society that is in turn part of a global intercommunication network.
2.2 Equivalence class of representations and embodiment
Physical realisation of language can be neural (in an individual’s brain), spoken (sound), written (visual), electronic (digital), or in visually transmitted sign patterns (sign languages). The same structural patterns are embodied in these different representations. They are all enabled by the physical structure of the brain, which is hierarchically structured so as to enable an interplay of sensory interpretation and prediction, based on pattern recognition, classification, memory, and extrapolation (Hawkins & Blakeslee 2004). Meaning is embodied in an equivalence class of such surface representations: it is independent of whether language is spoken, written or signed, and of language family, dialect/pronunciation, and font. A profound ability of the mind, underlying the flexibility of language usage, is to recognize them all as functionally equivalent. The concepts represented are recognized as entities that exist in their own right, which can be labeled and represented in many different ways.
2.3 Key features of language
The major function of language is its labelling of specific and generic objects and instances, as well as abstract entities, through use of indices and symbols. Via recursion (Deacon 2003; Trask 2007), this referential and representational nature can allow reference to itself, and hence disjunction from physical referents. Related key design features of language are (Trask 2007):Arbitrariness: the absence of any necessary connection between the form of a word and its meaning. This uncoupling of the signifier from the signified is crucial to representing objects symbolically (Striedter 2005). It allows equivalence classes of representations to exist: the same meaning can be expressed in different symbolic forms and systems.

166 George F.R. Ellis
Duality of patterning: a type of structure, encoded in a grammar with a particular syntactical structure, in which a small number of meaningless units are combined to produce a large number of meaningful (semantic) units. This introduces the crucial feature of discreteness of structural units (the quantum principle), which can then be repeatedly combined as recognised and named higher level units, giv-ing hierarchy and allowing arbitrary complexity of combinations to be built up through recursion. Its constituent structure is based on this principle (Trask 1999).Stimulus-freedom: our ability to say anything at all in any situation, so enabling discourse that is freed from the immediate situation and stimuli. This enables us to think off-line, i.e. without having to immediately act on what is thought about; hence it enables us to reflect on the past and plan for the future (Bickerton 2001). This facilitates the crucial feature of suppressing stimulus bound behaviours and replacing them with less determinate voluntary acts (Striedter 2005).Displacement: the ability to speak about things other than here and now: the future, the past, the possible, even the impossible. This enables us to analyse theoretical situations and so indulge in imagination and abstract analysis based in past mem-ories and future possibilities.Open-endedness: the ability of language to say new things, virtually without limit. This enables language to be a vehicle whereby creativity can emerge.Redundancy: the full message is entailed by part of the given text/message, hence one can determine the full message by partial information (if the context is known). This enables effective communication: we can understand the message in the presence of distortion or noise, and can predict what is meant on the basis of only a part of the text – an essential aspect of listening and reading.
A key feature of the way language functions is the use of metaphor, which plays a major role in cognition and meaning-making (Lakoff & Johnson 1980). This is enabled by the features listed above, and takes place in the context of conceptual schemas and cultural frames (Feldman 2006), which are the context of our understandings.
3. Hierarchical structuring
These features in turn are based in the functional structure of language, enabled by the physical structuring of the brain.
3.1 Functional structure
Language has a modular hierarchical structure (Booch 2007) that enables its com-pletely flexible representational and social function. This structure is bound by

Chapter 9. Biology and mechanisms related to the dawn of language 167
strict semiotic requirements (Deacon 2003), leading to a necessary set of implicit rules, but with a great variety of possible realisations (different languages/dialects). Its hierarchical character is enabled by a class structure with inheritance, embody-ing the crucial feature of recursion (the occurrence in a sentence of a syntactic category containing within it a smaller version of the same category, based on chunking and labelling). “The recognition of a suitable set of syntactic categories allows us to analyse all the sentences of a language as being built up, by means of a fairly small set of rules allowing recursion, from just these few categories” (Trask 1999: 288). This is a very general way of handling complexity: recursion occurs whenever you break up a complex task into simpler tasks that are completed first (Hofstadter 1980).
Reflecting and enabling the modular hierarchical structure of language, the brain also has a modular hierarchical structure (Beer 1981; Hawkins 2004), in turn based in the hierarchical structure of its physical constituents (Scott 1995), (see Table 1; for a more detailed description of this hierarchical structure, see http://www.mth.uct.ac.za/~ellis/cos0.html). Major issues arise in the linkages between different scales in the brain – the relation of molecules to neuronal activity; of neuronal connectivity to brain function; of brain regions to cognitive function; of the brain as a whole to individual psychology; and of the individ-ual to the social and ecological environment. A further major issue is how this physical hierarchical structure underlies and enables the hierarchical functional structure of language. This is part of the larger issue: how does brain structure underlie the complex functioning of the mind? (Koch 2004).
Table 1. The hierarchy of structure and causation. A simplified representation of the hierarchy of levels of causation in human beings, in terms of the corresponding academic subjects
Level 8 Sociology/Economics/PoliticsLevel 7 PsychologyLevel 6 PhysiologyLevel 5 Cell biologyLevel 4 BiochemistryLevel 3 ChemistryLevel 2 Atomic PhysicsLevel 1 Particle physics
3.2 Bottom-up and top-down causation
Emergent behaviour is enabled by the combination of bottom-up and top-down causation in the hierarchy of complexity and causation (Ellis 2008). This feature

168 George F.R. Ellis
occurs at all levels; in particular, between micro and macro aspects of the brain itself, in the relation of the brain to psychology, and in the relation of the individual to society (note that this applies to all animal brains, not just human). It is enabled by the existence of equivalence classes of lower level states that correspond to single higher level states (Auletta et al. 2008).
The interaction between these two types of causation in the functioning of the human cortex is emphasized by Feldman (2006) and Hawkins and Blakeslee (2004), while the way it takes place as regards individuals in the context of society is nicely illustrated in Berger (1963), Berger and Luckmann (1967), and Donald (2001). Top-down causation has crucial effects both on causal relations, and on the way we understand them. It is also a key feature in both language production and understanding, where holding context in mind is crucial to understanding both speech and writing (Smith 1976); key processes in language production, reception, and learning are therefore top-down driven. Indeed the way meaning is embodied in the hierarchical structure of language is context-dependent all the way down: the individual units at each level (sentences, phrases, words, phonemes) only attain their meaning and function, and even pronunciation, in the larger context of the whole meaningful situation (Krashen & Terrell 1983; Goodman 2005).
The key element allowing new ideas and information to come into being is adaptive selection, which is a specific form of top-down causation guided by higher-level selection criteria (Ellis 2008) which enable multi-level selection to take place (Okasha 2006). The top-down nature of this causation is crucial to this process, enabling new kinds of entities to come into being. Indeed, David Sloan Wilson (2011) remarks that the transition from bottom-up to top-down dominated causation in the relation of mind to the society in which it is imbedded is a major evolutionary transition in the historical development of humanity, resulting in the emergence of the social order as a higher level entity in its own right, and a consequent change in the nature of the evolutionary processes at work. However, adaptive selection also occurs on functional and developmental timescales.Production: Speaking and Writing: Speaking requires the physical ability to talk (vocal chords), and writing requires hands that can clasp and move delicately, plus a bipedal posture that allows them the needed freedom to operate. These are characteristically human features, although some animals do have some of these attributes (Striedter 2005). But one also needs the ability to form thoughts in the context of the current situation; this is where top-down causation occurs from that overall context to detailed thought processes. One then requires the ability to turn developing thoughts into a meaningful grammatical form, on the basis of past patterns experienced and usage-based language skills. This involves top-down causation from past experiences of linguistic patterns, as discussed below.

Chapter 9. Biology and mechanisms related to the dawn of language 169
Reception: Listening and Reading: Listening and reading are based on seeing and hearing, which are very ancient attributes in the animal kingdom, exquisitely developed in many higher animals as well as humans. Decoding syntax and grammar involves top-down causation from past experiences of linguistic patterns, similar to the processes involved in speech production. However, listening and reading are essentially based in processes of understanding, involving expectation and prediction in the face of partial information and noise (Smith 1976). These expectations are based on current context and memory, so this is also a form of top-down causation from those higher level concepts, based on similar processes to those occurring in general cognition (Hawkins & Blakeslee 2004).Learning: Language is learnt by experiential processes in a meaningful context (Smith 1976), supplemented by a combination of formal and informal instruction. This learning is an individual process of experimental development: hypothesis formation, trial and error, and imitation takes place in a social context of observation and experience. Like all human capacities, it is then embodied in a set of hierarchically structured automatized skills that are constructed in and constantly revised by consciousness (Donald 2001). This process is driven by the need to understand and predict (an example of top-down causation), in the overall context of the search for understanding and for meaning (Hawkins & Blakeslee 2004). Thus it is based – through experience and usage – firstly on intention-reading and cultural learning; secondly on schematization and analogy; thirdly on entrenchment and competition; and fourthly on functionally based distribution analysis (Tomasello 2003). This learning is enabled by Latent Semantic Analysis (Landauer et al. 1998), resulting in an Embodied Construction Grammar (Bergen & Chang 2003; Feldman 2006). Spoken and written language abilities are developed by variants of the same experiential process: meaningful experiences drive the process in a top-down way (Smith 1976). Formal teaching is needed to supplement and systematise this learning process, but cannot replace it.
4. Language modules and the development of language
The brain has developed through evolutionary processes that have led to the devel-opment of the conscious mind and language.
4.1 Evolutionary development
Evolutionary processes apply both to living beings, and to culture (Richerson & Boyd 2005; Mesoudi et al. 2006). Language develops over time through adaptation

170 George F.R. Ellis
in a social context, occurring via variation and then selection of language features according to communication utility (Burling 2007). The key point here is that according to standard genetic theory, nothing you understand or think influences the genes you pass on to your progeny, for they are fixed the day you are born. Hence any intellectual understandings you may develop cannot be directly selected for in biological terms, as they do not influence your genetic inheritance (although they do influence the likelihood of your passing on that unchanged genetic inheritance).
Rather, genetically based development results in general purpose intelligence (pattern recognition, classification, prediction) linked to memory (Hawkins & Blakeslee 2004), together with basic emotional modules (Panksepp 1998) that are effective in guiding the development in the individual of analytic ability in general, and language ability in particular (Greenspan & Shanker 2004). This takes place in his/her emotional context (Damasio 2000), with the social context crucially shaping individual minds in terms of understanding and language (Berger & Luckmann 1967; Donald 2001). That social context then reciprocally shapes the development of language itself: a culturally based dual evolutionary process (Richerson & Boyd 2005). This whole process favours genetic developments that lead to the propensity to develop language, but cannot lead to specific genetically determined language modules, for a variety of reasons I now briefly discuss.
4.2 Language modules
A large literature, largely based in linguistic analysis (for example, Pinker 1994), suggests that language processing is based in genetically determined language modules in the brain. Four problems make this extremely unlikely. First, it is developmentally unlikely that detailed connections in the cortex can be genetically determined, even though this does occur in other parts of the nervous system. Second, even if this was possible, there is not enough genetic information avail-able to carry out this task; essentially, this is a key finding of the human genome project. Thirdly, as just mentioned, one can’t directly select DNA to promote such connections because of the central dogma of molecular biology: the DNA one passes on to ones progeny is not affected by any of one’s intellectual activities. And fourthly, the kinds of specific issues involved in detailed language processing would have to compete with numerous other factors affecting survival, and it is highly unlikely these specific items would dominate over all the rest and so result in genetic processes determining specific language modules, as originally envis-aged by Chomsky.
An alternative view suggests language ability develops by conversion of genetically shaped general-purpose brain domains and capacities to language use.

Chapter 9. Biology and mechanisms related to the dawn of language 171
The way language is developed then is through the process of usage-based language acquisition (Tomasello 2003; Feldman 2006). As explained by these authors, these processes undermine the Poverty of Stimulus argument: sufficient depth of experience to carry out this process is provided by everyday life (Tomasello 2003). The universal underlying features of all languages (sometimes interpreted as resulting from the deep structure of a generic Universal Grammar) can then be understood as resulting from universal constrains intrinsic to their semiotic function (Deacon 2003).
Thus it is highly plausible that functional modular structures of language do not arise genetically, but rather have the restricted forms they do because of semiotic constraints. Then what is genetically determined in each individual is not specific language modules, but rather the capacity to develop language obeying these constraints. Hence in searching for the dawn of language, we do not have to explain genetically determined modules, but rather evolution of capacities that lead to the development of language. This has two aspects: the requisite intellectual capacities, and the needed emotional drivers; we discuss each below.
5. Patterns and symbols: Vision and music
What intellectual capacities and problem solving abilities underlie the develop-ment of language, and what hints of their presence could we hope to find in the archaeological record? Can we identify the intellectual capacities, and then pro-pose other ways they may display their presence?
5.1 Pattern recognition and classification
A first such key feature is the recognition of spatial and temporal patterns (Burling 2007: 82–84), and then categorisation: classifying and naming them, as discussed by Tomasello (2003), in the context of memory of past events and prediction of future happenings (Hawkins & Blakeslee 2004). Developing some form of con-ceptual structure is natural in higher animals (Burling 2007), indeed it is neces-sary for their functioning and survival, so they form proto-propositions (Hurford 2007). Classification skills allow organisms to categorize different aspects of their world into a manageable number of kinds of things and events (Tomasello 2003). Humans are able to go on to verbally name objects and events (labelling them symbolically), which develops naturally out of this process: the essence of lan-guage is its symbolic dimension, with grammar being derivative (Tomasello 2003).
It is suggested by Hawkins and Blakeslee (2004) that the basic purpose of intel-ligence is memory-based prediction: “the brain is not a computer, but a memory

172 George F.R. Ellis
based system that stores experiences in a way that reflects the true structure of the world, remembering sequences of events and their nested relationships and mak-ing predictions based on those memories. It is this memory-prediction system that forms the basis of intelligence, perception, creativity, and even consciousness”. The use of symbolism and language crucially helps these purposes, their utility being based in the core features of language discussed above. These abilities are based in the nature of local brain connections adapted to prediction and problem solving. How the core process of naming is implemented via neural connections is discussed by Hawkins and Blakeslee (2004); this is the basis of symbolic systems. A key question is why these kinds of inter-level connections came into being in the cortex. Understanding experiences, as mediated by the various senses, must have been the core driver, and in particular, interpreting visual images in conjunction with sounds may have been crucial in leading to their development.
5.2 Visual thinking
Vision is one of the first detailed sensory capacities to have been developed, and is so effective and important that there have been various independent origins of eyes (Conway Morris 2003; Parker 2004). It has been suggested in a provocative analysis by Rudolf Arnheim (1969) that visual thinking is primary in the development of intelligence, with abstract thinking being secondary to it. The description of his book states: “He shows that even the fundamental processes of vision involve mechanisms typical of reasoning … our perceptual response to the world is the basic means by which we structure events, and from which we derive ideas and therefore language”. He suggests the importance of the capacity to first represent relationships geometrically, identifying them as geometrical patterns, and then to express them abstractly, identifying them as abstract patterns. This is the start of symbolism, when one conjoins the ability to recognise both individual objects and classes of objects with the ability to associate some kind of symbol to each class of objects. This implies that the starting point for symbolic thinking is recognising patterns in space, hence the Blombos kind of pattern experiments are a starting point in such understanding. Additionally, vision involves the same kind of “filling in” of information that is a characteristic feature of listening and reading, as emphasized above (Koch 2004; Purves 2010).
Hence vision strongly develops core pre-language abilities. However these are shared with all the higher animals: it is not clear that human visual under-standing is greatly different from that of other animals. It is true that symbols are expressed in spatial or temporal relationships and patterns, and hence decoding them involves recognising such patterns; but it seems unlikely that such pattern recognition in humans is beyond that of many other higher animals, who through

Chapter 9. Biology and mechanisms related to the dawn of language 173
categorical perception can acquire a wide range of abstract concepts (Hauser et al. 2002). The progression from pictorial representation through icons to hierarchi-cally structured recursive symbol systems (see Figure 1) is, in intellectual terms, a natural progression (Burling 2007). Nevertheless visual thinking per se only weakly, if at all, develops the key feature of recursion: it only develops when writ-ing develops, long after spoken language has evolved. While fractal patterns occur in nature and in some art, such as Escher’s work, they are not a central feature of vision and visual understanding. Recursion is possible in sign language, but again is technically demanding and therefore not particularly natural and easy. Some-thing else must be involved in addition to vision.
→ Hierarchically structured symbolic systemsPictorial representations → icon → indices and symbols
→ Hierarchically structured recursive symbolic systems
Figure 1. Evolution of key characteristics of written language. A natural progression occurs in visual symbolic systems (Burling 2007); a parallel process should occur in the case of written words
5.3 Grammatical forms and music
Music and language each have complex hierarchical structures with an associated syntax. Together they are universal in human societies, and are unique to our spe-cies (Levitin 2007). They can be claimed to have interesting similarities in terms of their “syntactic architecture” (Patel 2008: 267), and to share a number of basic processing mechanisms with an overlap in the neural areas and operations that provide the resources for syntactic integration (Patel 2008). Singing and instru-mental playing help refine motor skills needed for speech production and writing. Listening to music involves processes of expectation, prediction, and perceptual completion (Levitin 2007), just as listening and reading do.
Thus music development would have encouraged the same intellectual abili-ties and motor skills as are needed for language, and may have played a more specific role in language development than vision. It would have had a deep effect on evolutionary psychological development because of the strong emotional power of music and dance, ensuring they played a major role in social bonding and cohesion, which thereby justifies its evolutionary importance. In particular, dance and music are associated with play, which is important for higher-level integrative processes (Levitin 2007) and specifically in language development (see the following section).

174 George F.R. Ellis
Most important of all, music commonly entails recursive structures (Patel 2008), which are key also in the structure of language, in both cases allowing its generative nature. The progression shown in Figure 1 is much more easily attained in vocal than visual form, because hierarchy and recursion are naturally developed through music and song.Development of Recursion: A key feature in the development of language is the emergence of recursion in symbolic systems (Deacon 2003), enabling abstract thought patterns to emerge. “Recursion is pervasive in the grammars of the lan-guages of the world, and its presence is the chief reason we are able to produce a limitless variety of sentences of unbounded length just by combining the same few building blocks” (Trask 1999: 244). Thus a crucial aspect of language is identify-ing as a single unit and naming compound experiences or concepts, thus allowing hierarchical structuring and recursion (building up patterns of patterns). This key ability for language (Hauser et al. 2002; Fitch et al. 2005; Burling 2007) and com-puter languages (Roberts 2009) must be based in specific aspects of local neural connections, involving specific kinds of connections enabling naming procedures to be applied to names themselves (perhaps developing specifically in Wernicke’s area: see Ramachandran 2011: 188–189). This type of neural connectivity, presum-ably involving links from higher levels of structure to lower levels, should be char-acterisable in the same kind of way that Hawkins and Blakeslee (2004) identify the neural bases of naming.
Consequently, one might envisage the key step in language development as being the development of neural connections allowing recursion, either directly as the result of some advantageous genetic mutation, or as a by-product of some other-directed advantageous mutation (Hauser et al. 2002; Fitch et al. 2005), per-haps related to the use of tools (Ramachandran 2011). In any event, as soon as the utility of this new ability was realised in developmental terms, perhaps first in relation to imaginative play, it would have become so important in mental life as to have demanded massive new physical resources, thus requiring major expan-sion of the cortex. Thus on this view, that crucial physical development (Striedter 2005) is regarded as being a result of, rather than the cause of, increased pro-cessing needs. A modest selective advantage in terms of allowing more effective cooperation between individuals will allow language and grammar to evolve by a conventional Darwinian process (Dunbar 2005). The analogue of Figure 1 can then occur in spoken language, and much later get embodied in written language.
Overall, I suggest that generic sensory interpretation, and visual thinking in particular, provided the broad basis for development of the abilities needed for language development and usage, with social interactions and play naturally developing the basis of symbolic reference through pointing and embryonic words.

Chapter 9. Biology and mechanisms related to the dawn of language 175
The specific capacities needed for language utility were greatly enhanced by the development of musical activities, evolving into singing and dancing. This provided a considerable part of the basis for language because of a deep connection between musical and linguistic syntax in the brain (Patel 2008), and in particular the use of recursion in musical syntax (Patel 2008).
If this view is true, it should have several testable consequences:First, the initial variation leading to recursive type structures would have been random (as there is no way it could have been directed), but it would have had a genetic basis, as otherwise it would not have been a heritable quality; hence there should be some determinable genetic linkage to this property. Either an identifi-able set of genes, or some identifiable epi-genetic processes, should be the key to making it possible.Second, it should result in some specific type of neural connectivity whereby recur-sive naming takes place, of the generic nature identified by Hawkins and Blakeslee (2004). This type of structure should eventually be identifiable.Thirdly, the resulting ability for recursive thought should then be manifested more or less simultaneously in various intellectual domains, particularly in language and play, but also in others such as patterns of music and art, and in technology.
6. The importance of emotions in this development
A key issue is what motivational structure drove these developments. Intelligent life is not a purely rational affair, as some analysts suggest (e.g. Gintis 2007). Rather emotional drivers largely determine to what use intellect is put (Damasio 1995).
6.1 The motivational impulse for language use
The motivation for language use is based in the functional importance of emotions at the psychological level, associated at the neuronal level with non-local neuronal connections such as the monoamine systems (Kingsley 2000). Particularly important here is the mother-child emotional connection, this is the key to early language development (Schore 1994; Greenspan & Shanker 2004), and the developmental significance of play in learning (Vygotsky 1978) and specifically in language (Paley 2004), a key to later language development. These features occur in the ambient social and ecological environment, where top-down action from the social level to individuals provides the integrated context for emergence of language, and plays a significant role in determining the outcome. The development of language is crucially dependent on the emotional drivers

176 George F.R. Ellis
that power the desire to communicate with others and assure the individual of their place in society. This leads to a key feature in language acquisition, namely intention reading (Tomasello 2003).
These drivers are based in the genetically determined primary emotional systems that are our evolutionary heritage from our distant ancestors (Panksepp 1998, 2008). According to Stevens and Price (2000), two key such systems are the Affiliation/Belonging system, and the Rank/Status system. These systems must therefore be important in communication, symbolism, and language development. This is confirmed by much evidence, the importance of the affiliation/belonging system in language development of children being emphasized by Greenspan and Shanker (2004), while that of the Rank/Status is emphasized by d’Errico and Henshilwood (this volume). Initial language development is based on the primary emotional bond of an infant with its mother/primary carer (Greenspan & Shanker 2004; Schore 1994) but then is developed through social interaction, particularly within the family context and peer group situations. These both underlie the development of joint attention and imitation, which are key features of human language development (Burling 2007).
A further key emotional system associated with language development is the Play system, discussed by Jaak Panksepp (1998, 2008). This system, with its associ-ated feelings of joy and fun and behavioural patterns of laughter, is a key founda-tion for learning and the development of imagination, which is why we share it with all our mammalian relatives, in particular the primates (Pellegrini & Smith 2005). However a probable major feature here was the change from rough and tumble play to imaginative play (Paley 2004; Smith 2005; Toronchuk & Ellis 2005), which probably played a significant role in developing theories of other minds. Evidence for this is the fact that weak functional and pretend play is a contributor to autism (Charman 2003).
6.2 The basis in developmental biology and evolution
By what kind of mechanism can brain plasticity lead to the formation of these effective modules in this adapted way? This has to be an adaptive process, of the kind labelled ‘Neural Darwinism’ by Gerald Edelman (1989), implemented by neuromodulators such as dopamine broadcast to the neocortex from the limbic system (thereby forming Edelman’s “value system” that guides the direction of plasticity). But the limbic system is the seat of emotional processes, so the fitness function guiding these adaptive processes is provided by the genetically determined primary emotional systems of the kind examined in detail by Panksepp (1998). Hence, evolutionary pressures in the ancestral environment developed various psychological traits that are experienced by us as emotions and

Chapter 9. Biology and mechanisms related to the dawn of language 177
feelings, which result in behaviour enhancing our evolutionary adaptation to the ancestral environment. Thus evolutionary processes enable brain development through genetically determined primary emotional modules (Panksepp 1998) that guide cortical development via a process of Neural Darwinism responding to daily experiences and events (Edelman 1989), with a crucial affective nature (Ellis & Toronchuk 1995). Note that this process in humans is just a further development of the same process occurring in vertebrates and primates. There is a discernable continuity of mechanism with our evolutionary predecessors that gives much evidential support for this proposal (Panksepp 1998).
What is inherited, then, are basic cognitive abilities rather than specific cog-nitive modules, plus the basic sensory and emotional systems that guide the use of cognition. Any effective cognitive modules that result develop from interac-tions of these systems with the social and physical environment, with the salience of reactions guided by the inherited emotional systems (Ellis 2008a) (Figure 2). Overall this process is of Darwinian rather than Lamarckian nature, because it does not propose genetically determined modules with specific cognitive con-tent, but rather genetically determined emotional systems that guide cognitive development. The behaviour that gets inbuilt in effective folk modules will be suitably tuned ab initio to the culture in which the individual lives, because they are created through interaction with that culture. This experiential shaping of these systems to fit the local environment is an aspect of the crucial feature of brain plasticity (Donald 2001).
Evolution → genetically determined emotional systemsEmotional systems + experience → effective folkpsychology behaviourEffective folkpsychology behaviour → learning effects
Figure 2. Evolution and effective psychological modules. The way emotion underlies the existence of effective psychological modules
7. Key steps towards language
Many of the basic cognitive capacities that allow language development (Toma-sello 2003) are shared between us and other higher mammals. The dramatic increase of brain size in relation to body weight (which may have been either the cause or the result of other changes) enables greatly increased memory and pro-cessing capacity. Together with physiological improvements in terms of speech production, these make speech physiologically possible. However this is not sufficient: intellectual development is crucial also. I suggest on the basis of the

178 George F.R. Ellis
above discussion that two crucial emergent features distinguishing human beings from other animals in both developmental and evolutionary terms are changes in the emotional system underlying language use, and changes in neural functional capacity so as to allow recursive thought.
7.1 Changes in physiology
The physiological changes mentioned above are crucial for language development, in particular, improved physical ability to form sounds and a greatly enlarged brain and associated greater memory capacity are certainly needed. But it is not clear if these are cause or effect. What caused them to happen? Brain size is crucial in terms of increasing both total computing power and memory capacity, and also because it necessarily promotes modularity (Striedter 2005), but this is not enough by itself (for there are other animals with larger brains!). We need something more than just brain size to make language fly.
7.2 Changes in the social and ecological context
Language development takes place in a social context where symbols are used for communication of social and environmental understandings so as to form a dis-tributed cognitive network (Donald 2001). Thus its development needs the right social context of a society with common needs and purposes. As emphasized by David Sloan Wilson (this volume), there will be particular environmental situa-tions where these will be stronger drivers for language development than others. Crucial drivers were the need for social communication to underlie coping and cooperation, enabled by development of a theory of mind and intention reading (Tomasello 2003), together with the child-caring and raring needs due to the long period between birth and adulthood. But no obvious change has been identified causing much greater need in these regards than in primates: there seems a con-tinuity with them rather than discontinuity, except perhaps in regard to develop-ment and use of technology.
It is certainly a possibility that development of technology was a major driver of the need for the kind of better communication afforded by language. According to Jacob Bronowski (1976: 64, 74) “The largest single step in the ascent of man is the change from nomad to village agriculture … settled agriculture creates a tech-nology from which all physics, all science takes off ”. But again it is not clear if that is the driver or the result of language change. The features of language listed above would have been very helpful in developing technology; perhaps one can make the case that language and technology co-evolved as equal partners, the rapid devel-opment at later times simply being the result of the essential nature of exponential growth. The idea that development of technology was the key contextual partner

Chapter 9. Biology and mechanisms related to the dawn of language 179
in language development seems a very viable proposal (see the articles in d’Errico & Backwell 2005 and particularly those by Tobias and by Parkington et al.). But what was the crucial threshold leading to the take-off of the language-technology partnership? It is argued above that this would have been when this co-evolution led to the discovery of the principle of recursion in symbolism.
Perhaps times of change associated with new environments are particular stressors demanding more effective communication, and so driving commu-nication and language development. Could the move out of Africa, between 50,000 and 100,000 years ago, perhaps have been one such stressor? The need to communicate is much greater in a strange environment than in a known environ-ment, so this move may have been the kind of change that strongly encouraged development of better communication abilities. This would have been greatly assisted by the associated freeing-up from debilitating diseases that resulted from the move out of Africa (Reader 1998), allowing the mind to flourish in an unprecedented way. This was of course recent in evolutionary terms, but perhaps gene-culture co-evolution (Richerson & Boyd 2005) could have occurred and allowed this event to become of evolutionary significance for the present day. Again this would have been related to development of technology, related both to travel and to adapting agriculture to new contexts. This might seem a promis-ing context for a leap forward in language.
7.3 Emotional development: The nature of play, other minds
It has been emphasized above that mother/child bonding is crucial ingredient in language development, but this is essentially the same as in other higher animals – apart from the use of language, which is what we are trying to explain! There is no obvious discontinuity here between pre-humans and humans. But the transition from rough-and-tumble play to imaginative play, based in the effects of non-local neural connections and resulting in displaced symbolic activity, could be a key development in terms of freeing up creativity and encouraging development of a theory of mind, as well as basic communication skills. This is a promising avenue for promoting language abilities, particularly because music and song are natural aspects of play. This kind of powerful joyful activity develops pre-language abilities in terms of listening, production of sounds, and theory of mind, as well as devel-oping basic grammatical skills.Intellectual development: Emergence of Recursion: It has been argued above (in agreement with Hauser et al. 2002; Fitch et al. 2005) that the single most important feature in the emergence of language is the development of recursion in symbolic systems, enabling abstract thought patterns to emerge. I suggested this is enabled by specific classes of neural connections, initially emerging through genetic chance

180 George F.R. Ellis
but then rapidly spreading because they provide a key aspect of the development of higher level thought and language.
One should recognise here the multimodal nature of meaning-making and cognition: each of the modes of communicating has a semiotic nature, and they support each other (Kress 2000; Kress & van Leeuwen 2001). However it has been suggested above that technology, music, and recursion may have been the links through to developing language capacity.Archaeological traces: Might any of the features discussed here perhaps leave some traces in the archaeological record? There are many such links to the above narra-tive, most already being explored; the point here is that this search can be related to aspects of how the brain underlies language, as outlined above, which might possibly help focus the search.
There are of course records of group membership, related to belonging/ cohe-sion, and symbols of social status, related to rank/power; these are a small step along the symbolic route. One might search further for records of music and danc-ing, argued above to be key players in the development of language, seeing if any traces of recursive patterning occur in these records. One can search for symbols and artefacts in the archaeological record associated with play: toys as symbols used in imaginative play, playing boards, scoreboards, and so on. Could any of the Blombos patterned artefacts be of this nature? And in particular do any show signs of recursive features, such as fractal-like patterning? One might ask if there is any way of identifying traces of recursive thought in any other artefacts, for example those that are purely decorative. And finally the link to technology is crucially important, as evidenced in other articles in this book. Perhaps recursive patterning can be found in tool design, implying it is embodied in manufacturing techniques. If so, that would be a significant discovery as regards thought patterns essential to language. However it has to be admitted they will individually be sug-gestive rather than decisive. A powerful case will only be made by a collection of evidence of all kinds, of such a nature as to provide a cumulative case.
References
Arnheim, R. 1969. Visual Thinking. Berkeley: University of California Press.Auletta, G., Ellis, G.F.R. & Jaeger, L. 2008. “Top-Down Causation: From a Philosophical Problem
to a Scientific Research Program.” J R Soc Interface 5: 1159–1172.Beer, S. 1981. Brain of the Firm. Chichester: John Wiley.Bergen, B.K. & Chang, N. 2003. “Embodied Construction Grammar in Simulation Based
Language Understanding.” In Construction Grammars(s): Cognitive and Cross-Language Dimensions, J–O. Östman & M. Fried (eds), 147–190. Amsterdam: John Benjamins.
Berger, P. 1963. Invitation to Sociology: A Humanistic Perspective. New York: Doubleday.

Chapter 9. Biology and mechanisms related to the dawn of language 181
Berger, P. & Luckmann, T. 1967. The Social Construction of Reality: A Treatise in the Sociology of Knowledge. New York: Anchor.
Bickerton, D. 2001. Language and Human Behaviour. Seattle: University of Washington Press.Booch, G. 2007. Object-Oriented Analysis and Design with Applications. Menlo Park:
Addison-Wesley.Bronowski, J. 1976. The Ascent of Man. London: The BBC.Burling, R. 2007. The Talking Ape: How Language Evolved. Oxford: Oxford University Press.Charman, T. 2003. “Theory of mind and the early diagnosis of autism.” In Understanding other
minds: Perspectives from developmental cognitive neuroscience, S. Baron Cohen, H. Tager-Flusberg & D. J. Cohen (eds), 422–441. Oxford: Oxford University Press.
Christiansen, M. & Kirby, S. (eds). 2005. Language Evolution. Oxford: Oxford University Press.Clark, A. 2008. Supersizing the Mind: Embodiment, Action, and Cognitive Extension. Oxford:
Oxford University Press.Conway Morris, S. 2003. Life’s Solution: Inevitable Human Beings in a Lonely Universe.
Cambridge: Cambridge University Press.Damasio, A. 1995. Descartes’ error: Emotion, reason, and the human brain. New York: Avon.Damasio, A. 2000. The feeling of what happens: Body, emotion and the making of consciousness.
London: Vintage.Deacon, T. 1997. The symbolic species: The co-evolution of language and the brain. London:
Penguin Books.Deacon, T. 2003. “Universal Grammar and Semiotic Constraints.” In Language Evolution,
M. Christiansen & S. Kirby (eds), 111–139. Oxford: Oxford University Press.d’Errico, F. & Backwell, L. (eds). 2005. From Tools to Symbols: From Early Hominids to Modern
Humans. Johannesburg: Witwatersrand University Press.d’Errico, F. & Henshilwood, C. this volume. “The origin of symbolically mediated behaviour:
from antagonistic scenarios to a unified research strategy.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds). 49–73. Amsterdam: John Benjamins.
Donald, M. 2001. A mind so rare. New York: Norton.Dunbar, R.I.M. 2005. “The origin and subsequent evolution of language.” In Language Evolution,
M. Christiansen & S. Kirby (eds), 219–234. Oxford: Oxford University Press.Edelman, G. 1989. Neural Darwinism: The theory of group neuronal selection. Oxford: Oxford
University Press.Ellis, G.F.R. 2008a. “Commentary on ‘An Evolutionarily Informed Education Sience’ by David C
Geary.” Educational Psychologist 43 (4): 1–8.Ellis, G.F.R. 2008. “On the nature of causation in complex systems.” Transactions of the Royal
Society of South Africa 63: 69–84.Ellis, G.F.R. & Toronchuk, J. 1995. “Neural development: Affective and immune system influ-
ences.” In Consciousness and emotion: Agency, conscious choice, and selective perception, R. D. Ellis & N. Newton (eds), 81–119. Amsterdam: John Benjamins.
Feldman, J. 2006. From Molecule to Metaphor: A Neural Theory of Language. Cambridge, Mass: MIT Press.
Fitch, W.T. Hauser, M.D. & Chomsky, N. 2005. “The evolution of the language faculty: clarifications and implications.” Cognition 97: 179–210.
Gintis, H. 2007. “A framework for the unification of the behavioral sciences.” Behavioral and brain sciences 30: 1–61.

182 George F.R. Ellis
Goodman, K. 2005. What’s Whole in Whole Language: 20th Anniversary Edition. Muskegon, MI: RDR Books.
Greenspan, S. & Shanker, S. 2004. The first idea: How symbols, language, and intelligence evolved from our primate ancestors to modern humans. Cambridge, Mass: Da Capo Press.
Hauser, M.D. Chomsky, N. & Fitch, W.T. 2002. “The faculty of language: What is it, Who has it, and How did it evolve?” Science 298: 1569–1579.
Hawkins, J. & Blakeslee, S. 2004. On Intelligence. New York: H Holt & Co.Hofstadter, D.R. 1980. Gödel, Escher, Bach: An Eternal Golden Braid. Harmandsworth: Penguin
books.Hurford, J.R. 2007. The Origins of Meaning. Language in the Light of Evolution. Oxford: Oxford
University Press.Kingsley, R. 2000. Concise text of neuroscience. Philadelphia: Lippinscott, Williams and Wilkins.Koch, C. 2004. The quest for consciousness: A Neurobiological approach. Englewood, Co: Roberts
and Company.Krashen, S.D. & Terrell, T.D. 1983. The natural approach: Language acquisition in the classroom.
San Francisco: Alemany Press.Kress, G. 2000. “Multimodality.” In Multiliteracies: Literacy and design of social futures, B. Cope &
M. Kalantzis (eds), 182–202. London: Routledge.Kress, G. & van Leeuwen, T. (2001). Multimodal discourse: The Modes and Media of Contemporary
Communication. London: Arnold.Lakoff, G. & Johnson, M. 1980. Metaphors we live by. Chicago: University of Chicago Press.Landauer, T.K., Foltz, P.W. & Laham, D. 1998. “An Introduction to Latent Semantic Analysis.”
Discourse Processes 25: 259–284.Levitin, D.J. 2007. This is your brain on music: the science of a human obsession. London: Plume.Mesoudi, A., Whiten, A. & Laland, K.N. 2006. “Towards a unified science of cultural Evolution.”
Behavioral and brain sciences 29: 329–383.Okasha, S. 2006. Evolution and the levels of selection. Oxford: Oxford University Press.Paley, V.G. 2004. A Child’s Work: the Importance of Fantasy Play. Chicago: University of Chicago
Press.Panksepp, J. 1998. Affective neuroscience: The foundations of human and animal emotions.
Oxford: Oxford University Press.Panksepp, J. 2008. “The Power of the Word May Reside in the Power of Affect.” Integr Psych
Behav 42: 47–55.Parker, A. 2004. In the blink of an eye: how vision kick-started the big bang of evolution. London:
Free Press.Patel, A.D. 2008. Music, Language, and the Brain. Oxford: Oxford University Press.Pellegrini, A.D. & Smith, P.K. 2005. The Nature of Play: Great Apes and Humans. New York: The
Guilford Press.Pinker, S. 1994. The Language Instinct. London: Penguin Books.Purves, D. 2010. Brains: How they seem to work. Saddle River, New Jersey: Pearson Education.Ramachandran, V.S. 2011. The Tell-Tale Brain: Unlocking the Mystery of Human Nature. London:
William Heinemann.Reader, J. 1998. Africa: A Biography of the Continent. London: Penguin Books.Richerson, P.J. & Boyd, R. 2005. Not by Genes Alone: how culture transformed human evolution.
Chicago: University of Chicago Press.Roberts, E.S. 2009. Java: An introduction to computer science. Boston: Addison Wesley.

Chapter 9. Biology and mechanisms related to the dawn of language 183
Schore, A.N. 1994. Affect regulation and the origin of the self: The neurobiology of emotional development. Hillsdale, NJ: Erlbaum.
Scott, A. 1995. Stairway to the mind. New York: Springer Verlag.Simon, H.A. 2001. The sciences of the artificial. Cambridge, Mass: MIT Press.Smith, F. 1976. Reading. Cambridge: Cambridge University Press.Smith, P.K. 2005. “Social and pretend play in children.” In The Nature of Play: Great Apes and
Humans, A.D. Pellegrini & P. K. Smith (eds), 173–209. New York: The Guilford Press.Stevens, A. & Price, J. 2000. Evolutionary psychiatry: A New beginning. New York: Taylor and
Francis.Striedter, G.F. 2005. Principles of Brain Evolution. Sunderland, Mass: Sinauer Associates.Tomasello, M. 2003. Constructing a language: A usage-based theory of language acquisition.
Cambridge, Mass.: Harvard University Press.Toronchuk, J.A. & Ellis, G.F.R. 2005. Affective neuronal group selection: The nature of the primary
emotional systems. Unpublished manuscript, University of Cape Town. Available at http://www.mth.uct.ac.za/~ellis/AND%20II.doc.
Trask, R. L 1999. Key Concepts in Language and Linguistics. Abingdon: Routledge.Trask, R.L. 2007. Language and Linguistics: The Key Concepts. Abingdon: Routledge.Vygotsky, L.S. 1978. Mind in Society: The development of higher psychological processes.
Cambridge, Mass: Harvard University Press.Wilson, D.S. this volume. “The human major transition in relation to symbolic behaviour,
including language, imagination, and spirituality.” In ‘Homo Symbolicus’: The Dawn of Language, Imagination, and Spirituality, C. Henshilwood and F. d’Errico (eds). Amsterdam: John Benjamins.


chapter 10
The other middle-range theories
Mapping behaviour and the evolution of the mind
Benoît DubreuilPhilosophy Department,Université du Québec à Montréal
Archaeologists have long been aware of the need to make explicit the middle-range theories that they use to bridge the gap between raw archaeological evidence and human behaviour. In this paper, I discuss the difficulties in proposing such theories when dealing with long-extinct hominin populations, among which significant biological differences may have existed. I emphasize how far we remain from a general theory that would link genetic, neural, cognitive, and behavioural evolution, and how difficult it is to go beyond trivial explanations of the evolution of the human mind and behaviour. I argue that students of human cognitive evolution should move away from general mental phenomena, which they have focused on in recent years, and turn their attention to increasingly precise cognitive mechanisms realized in specific neural structures.
1. Introduction
Lewis Binford’s theoretical work in archaeology is most famous for his call for the development of a middle-range theory, that is, for his emphasis on the need to develop procedures and frameworks to bridge the gap between raw archaeologi-cal evidence and dynamic human behaviour (Binford 1977, 1981). Over the years, archaeologists have followed this suggestion, developing diverse strategies to guide and constrain their interpretation of the archaeological record, and to reconstruct the behaviour of ancient human populations. To be sure, bridging the gap between material culture and behaviour is an unending endeavour. New technologies produce new kinds of data, which in turn create new problems of interpretation. Moreover, as archaeological data are by nature limited and more often than not compatible with various behavioural patterns, there is sometimes simply no way of arriving at a satisfactory middle-range theory, and large parts of our past are thus condemned to remain unexplained. These problems are well known and inescap-able. They represent the frontiers that archaeology faces as a discipline.

186 Benoît Dubreuil
Nevertheless, as an outsider to archaeology, I must admit that I have always been impressed by the amount of information that archaeologists can extract from scant evidence. Although they often disagree on their general interpretation of the archaeological record, they mostly agree on what can in principle be learned from various types of evidence, and how. This is true, at least, for what concerns recent human prehistory. The interpretation of older periods of human prehistory raises more acute problems. The reason is that the human species has evolved, and therefore the deeper we look into our past, the more likely we are to find a significant gap between the mind and behaviour of modern humans and that of extinct hominins. Human biological evolution thus adds several layers of complexity to our inquiry.
Anthropologists, sociologists, and other social scientists usually take for granted that any variation in behaviour between two human populations results from cultural or ecological factors. Sharing this assumption, students of our recent archaeological past have made extensive use of anthropological and sociological theories to constrain and regiment their interpretation of material culture. But expanding the scope of inquiry over hundreds of thousands of years makes this assumption questionable. The fact of human evolution dramatically alters our quest for a middle-range theory. Cognition and biology become additional vari-ables that must be taken into account as soon as we consider behaviours spanning into our distant past. For such an inquiry it is insufficient for archaeologists to take inspiration from anthropology and sociology; ethnology, cognitive neuroscience and evolutionary biology must kick in.
2. A research agenda for the next two centuries
As the number of disciplines and methods expand, however, the risk of students of human evolution being submerged by new theories and data becomes real and, to tell the truth, it is far from obvious that the challenges of interdisciplinary research will be met successfully. Indeed, working at several different explanatory levels at once is not only beyond the reach of most scholars, but inaccessible even to the best among them. There is simply too much information to process. A comprehensive theory of human evolution must start from the archaeological record and explain how the evolution of human behaviour relates to underlying cognitive, neural, and genetic evolution (see Figure 1). Fortunately, changes at one level are linked to changes at the other levels (e.g. genetic evolution is linked to neural evolution which, in turn, is linked to cognitive evolution). What we know at one level can thus in principle be used to guide and constrain hypotheses about what is going on at the other levels. Unfortunately, there is a wide gamut of ways in which the

Chapter 10. The other middle-range theories 187
various levels can relate to one another. What is worse, we know close to nothing about how genetic evolution affects neural evolution, or how neural evolution affects cognitive and behavioural evolution.
Material culture
Behaviour
Cognition
NeuronsNaturalenvironment
Culturalenvironment
Genes
Natural selection
Figure 1. Levels of explanations in human evolution
General theories covering the whole problem of evolution from natural selection to the archaeological record will probably remain beyond our reach for several generations. Moreover, given the intrinsic complexity of the biolog-ical world, the best we can hope is probably to produce patchy explanations, with piecemeal integration between different levels of explanation (Mitchell 2003; Craver 2007). In the case of human evolution, to my knowledge, the only hypothesis that ever attempted to cover the whole ladder of complexity was the so-called FOXP2 hypothesis. According to this hypothesis, the mutation of a specific gene (FOXP2), expressed in the basal ganglia, has been selected for in modern Homo sapiens because of its impact on language. The enhanced speech, motor control and recursive syntax made possible by this mutation would in turn explain the evolution of “modern behaviour,” including symbolic culture (Corballis 2004; Lieberman 2007). But nearly every step in this analysis can be questioned. FOXP2 apparently took its modern form long before the evolu-tion of modern humans (Krause et al. 2007) and its impact on language and cognition hardly explains the kinds of behaviours associated with Homo sapiens (Dubreuil 2008).

188 Benoît Dubreuil
Despite its limited plausibility, the FOXP2 hypothesis has generated much enthusiasm in the literature on human cognitive evolution. This is probably because it offered, for the first time ever, a complete explanatory scenario, drawing a connection between natural selection and the archaeological record, with at least some test implications. The FOXP2 hypothesis was ambitious, probably too ambitious, but it is definitely the kind of hypotheses that we would like to find. To understand how far we remain from such comprehensive theories, we should keep in mind how difficult it is to link only one level of explanation with another. Consider for instance the well-known difficulty in linking the archaeological record with behaviour, that is, of producing middle-range theories in Lewis Binford’s classical sense. We still have no consensus, for instance, on the behavioural implications of the use of symbolic artefacts by Neandertals, a question of the utmost importance for human cognitive evolution. Current evidence is compatible with various behavioural interpretations, each having different implications at the lower levels of cognitive, neural and genetic evolution (Mellars 2005; Zilhaõ 2006). Choosing among these interpretations is currently impossible and will remain so until new findings make it possible to reconstruct more precisely the evolution of symbolic behaviour in Eurasia and Africa.
As a philosopher working in the cognitive science and human behaviour, it is quite unlikely that I will contribute to these findings. Fortunately that is not my goal. The question that I would like to ask instead is the following one: is it possible to inquire into the lower levels of complexity before settling the most crucial issues at the higher levels? Can we discuss the genetic, neural, and cognitive foundations of symbolic behaviour before we determine if, and for what purpose, Neandertals created and used symbolic artefacts? In other words, can we devise middle-range theories linking behaviour to cognition, or cognition to neurons and genes, without first settling the hottest issues at the behavioural level?
If it is impossible, then specialists in cognitive neuroscience, genetics, and evolutionary biology might do well to avoid discussing the evolution of the human mind for some time, and to return to it once major debates among archaeologists have been settled. But the contribution of non-archaeologists might not be totally useless, even in this context of uncertainty. At the end of the day, archaeological data, fossils and artefacts, will be deciding, but this does not preclude going beyond the construction of middle-range theories in Lewis Binford’s classical sense, that is, theories linking the archaeological record to behaviour. We can also propose other middle-range theories at the lower level of genes, neurons, and cognitive functions. After all, our mind might have evolved following various scenarios, and yet all scenarios are not equally likely.

Chapter 10. The other middle-range theories 189
3. What middle-range theories for non-archaeologists?
A peculiar feature of non-archaeological research on human evolution is that the most advanced research has probably been achieved at the lowest level of complexity, that of natural selection, thanks to the work done over the years by evolutionary biologists. There are now very efficient theories on the market to model how specific cognitive or behavioural traits might have been selected for in our ancestral environment. This is truly helpful, especially since evolutionary biologists have learned to work hand in hand with evolutionary psychologists to determine how selective pressures might have led to the evolution of properly human dispositions.
The counterpart of this progress, obviously, is that it does not tell us when, and in which contexts, the specific features of our mind evolved in the human lineage. It is one thing to recognize that the modern mind has a genetic basis that has been selected for in our evolutionary past; it is another to determine what genes have been selected for, and when and what was the precise impact of this selection on our ancestors’ brains and behaviour. Very few genes can be directly and confidently linked to cognition and behaviour. In fact, higher-level cognitive processes and behaviours mostly have a polygenetic basis that makes a direct inquiry into the genetic foundations of the modern mind problematic (Schaffner 1998; Wynn & Coolidge 2007).
The other feature that I want to highlight is the difficulty that we have had until now in formulating nontrivial theories linking neural, cognitive, and behavioural evolution. Many proposals have not gone very far beyond what I would call the “bigger brain-greater intelligence hypothesis”. Encephalisation is beyond doubt the most noticeable fact in the evolution of the human brain. It would be astounding if this process were not causally linked specifically with human creativity and culture. But the “bigger brain – greater intelligence hypothesis” has no clear test implications, because there is an infinite variety of ways in which encephalization can be related to cognitive and behavioural evolution.
Science does not progress by explaining a complex phenomenon by an equally complex phenomenon. The creativity of modern Homo sapiens behaviour, for instance, remains entirely unexplained if it is simply related, with no further spec-ification, to the evolution of modern intelligence or language. The complexity of the explanans (modern intelligence or language) being equivalent to that of the explanandum (modern culture), it is not clear what such an explanation is doing apart from rephrasing the problem. I think that I am not too far from the truth if I say that archaeologists have sometimes been tempted by such trivial explanations. It is certainly not my objective to criticize them for that, since in the context of an

190 Benoît Dubreuil
interdisciplinary division of labour the construction of nontrivial cognitive and neural theories to account for behavioural evolution in extinct hominins is not strictly their responsibility.
The reason why archaeologists often have recourse to uninformative cognitive and biological explanations might be because such explanations are not at the centre of their interest, and often intervene in their accounts of past behaviour as an explanation of last resort. A behavioural change is explained biologically when other non-biological variables – such as culture, demography, or climate – have exhausted their explanatory potential. This is not totally unreasonable, because non-biological variables are likely to impact on the presence of the behaviours we are interested in, be it the production of symbolic artefacts or other innovations. It is thus reasonable to control for these variables before looking to biology for an explanation. But in the context of human evolution, using biology as a variable of last resort also creates some problems. Consider, for instance, the way Richard Klein frames the current alternatives in the explanation of modern sapiens behaviour:
“Most palaeoanthropologists now accept Out of Africa and argue mainly about the extent to which non modern Eurasians and modern African immigrants may have interbred. Most authorities further agree that a behavioural transformation underlay the modern African expansion, but they presently divide between a majority who believe that advanced behaviours accumulated gradually between perhaps 120 and 50 ka and a minority who believe they appeared abruptly about 50 ka. Those who favour gradual development usually attribute it to long-term social, demographic, or economic shifts, while those who perceive abrupt development, mainly the present writer, suggest that it was prompted by genetic change.” (Klein 2008: 267, emphasis added)
One problem with this proposition is that it is difficult to see how the value of the biological explanation can be weighed against the alternatives. To assess the explanatory value of genetic evolution, controlling for other non-biological vari-ables is certainly necessary but it is not on its own sufficient. Genetic or (more generally) biological changes are not all equally likely and they do not all explain equally well various behavioural changes. Thus, biology cannot simply enter as a “get out of jail free card” when other explanations become implausible. One also has to present scenarios that link plausible genetic, neural, and cognitive changes to the behavioural evolution to be explained. Those are precisely the nontrivial middle-range theories that we need. By contrast to Lewis Binford’s classical middle-range theories – linking the archaeological record to behaviour – they can only emerge from the cooperation of philosophers, psychologists, linguists, and social scientists with a strong background in cognitive neuroscience.

Chapter 10. The other middle-range theories 191
4. Current middle-range theories
Valuable proposals have already attempted to go beyond the “bigger brain-greater intelligence hypothesis”. In this section, I discuss the limitations of three influential and thought-provoking middle-range theories: (1) Derek Bickerton’s syntactic hypothesis, (2) Robin Dunbar’s social intelligence hypothesis, and (3) Frederick Coolidge and Thomas Wynn’s working memory hypothesis. In the following section, I argue that the common problem with these middle-range theories lies with their focus on complex mental phenomena, rather than on the cognitive and neural mechanisms that produce these phenomena.
Bickerton (2003) has gone beyond the trivial linguistic explanation by emphasizing the role of syntactic recursion. Recursion is a formal property of human language that makes it possible to embed clauses within clauses. There is no question that recursion (or something like it) is instrumental in expressing in language the kind of metarepresentations that humans are capable of. There are nevertheless important problems with this proposal. The first is that recursion is a formal property of language and not a cognitive mechanism. Unfortunately, there is no consensus on its cognitive (not to say neural) foundations in humans (Calvin & Bickerton 2000; Lieberman 2007; Wynn & Coolidge 2007). Another problem is that, while some contend that recursion is proper to humans, others doubt whether it is even a true property of natural human language (Hauser et al. 2002; Suddendorf & Corballis 2007; Marcus 2008). Another problem is that the behavioural implications of the evolution of linguistic recursion are far from clear. I have raised doubt elsewhere about the capacity of recursion to account for the diversity of behaviours that has been associated with modern sapiens culture and even to explain the emergence of symbolic behaviours (Dubreuil 2008).
Another challenging middle-range theory has been advanced by Robin Dunbar, who has argued over the years in favour of a “social brain hypothesis” (Dunbar 2003, 2007). Dunbar explains the evolution of modern sapiens culture by the evolution of “theory of mind” or the capacity to ascribe mental states to oneself and others. Psychologists still debate in what way(s) human theory of mind differs from that of nonhuman primates, but there is little doubt that differences exist and must have played a role in the evolution of language, culture, and imagination (Call & Tomasello 2008). One problem with the social brain hypothesis, however, is that the concept of “theory of mind”, although is it not as general as “language” or ‘intelligence”, is still very general. Many specific cognitive mechanisms might be responsible for the evolution of specifically human theory of mind. Dunbar (2004, 2007) points toward the existence of a correspondence between brain growth and the evolution of various levels of intentionality, but this move is not motivated at

192 Benoît Dubreuil
the cognitive or neurological levels. There are simply no data that could allow us to establish a correlation between brain size and levels of intentionality.
Psychologist Frederick Coolidge and archaeologist Thomas Wynn (Coolidge & Wynn 2001, 2005; Wynn & Coolidge 2007) have proposed a third hypothesis that I want to mention. Their proposal is similar to earlier ones that explain properly human behaviours by the evolution of the executive functions of the brain (Mithen 1996; Deacon 1997). Coolidge and Wynn’s hypothesis is centred on the concept of working memory, which is defined as the capacity to hold in mind recently processed information and to keep it available for further processing. They build on the model of working memory developed over the years by psychologist Alan Baddeley. According to Baddeley (Baddeley & Hitch 1974; Baddeley 2001), the system of working memory is composed of a central executive, responsible for the control and regulation of information, and of two “slave systems”, the phonological loop and the visuospatial sketchpad. The fact that modern humans perform better than nonhuman primates in different working memory tasks suggests that the system evolved in the human lineage (Read 2008). This evolution, according to Coolidge and Wynn, is likely to explain the appearance of more flexible behaviours in recent human evolution, as well as the difference between modern Homo sapiens and Neandertals (Coolidge & Wynn 2004; Wynn & Coolidge 2004). The central problem with the concept of working memory, even as specified in Baddeley’s model, is that it is still very general. At the neural level, working memory tasks activate widely distributed networks in the prefrontal, parietal, and temporal cortices. Moreover, it is difficult to establish exactly what is specific to the modern human working memory system. Although the human working memory system certainly differs from that of nonhuman primates, the different components of working memory are already present in our closest relatives. But if the gap between humans and nonhuman primates is more quantitative than qualitative, how are we to determine how much working memory is needed to produce modern sapiens behaviour? How much working memory was there in Neandertal or Homo heidelbergensis? Before coming back to these questions, I want to discuss what I see as a common limitation of these three hypotheses.
5. What’s wrong with current theories?
The middle-range theories that we need must offer a link between behaviour and cognitive evolution. This is basically what the hypotheses discussed above are doing. Recursive syntax, theory of mind, and working memory are cognitive concepts; that is, they refer to the ability of the human mind to process some sort of information. Their common limitation, however, comes from the fact that they

Chapter 10. The other middle-range theories 193
refer to complex cognitive tasks, which are realized by largely overlapping cognitive and neural mechanisms. This, I will argue, makes it impossible to draw distinct test implications from these hypotheses.
Let’s begin with working memory and theory of mind. At the cognitive level, they depend on similar cognitive functions. The capacity to inhibit prepotent responses, for instance, is needed to succeed in complex theory of mind tasks (Carlson et al. 2004), but it is also a function of the central executive in Baddeley’s model of working memory (Jurado & Rosselli 2007). The same can be said of selective attention and attention switching, which the central executive is thought to perform, but which are also essential to succeed in complex theory of mind tasks (Gerrans & Stone 2008). But if theory of mind and working memory depend on the same cognitive mechanisms (inhibitory control and selective attention), how are we to distinguish between a behavioural change related to (1) the evolution of theory of mind, (2) the evolution of working memory, and (3) the evolution of inhibitory control or selective attention?
A similar problem exists with the syntactic hypothesis. De Villiers and Pyers (2002), for instance, have shown that the mastery of embedded clauses in language development was predictive of success in complex theory of mind tasks, suggesting the presence of a common underlying mechanism. What then are the distinct implications of the syntactic hypothesis? Things risk getting even more confused when syntactic recursion is equated with other complex cognitive tasks. Suddendorf and Corballis (2007), for instance link recursive syntax not only to the evolution of theory of mind, but also to that of episodic memory and mental time travel. Coolidge and Wynn (2007), for their part, link the evolution of recursion to that of working memory and, more particularly of the phonological storage capacity. Here again, it is difficult to see how these various hypotheses have distinct test implications.
The picture does not get clearer if one shifts the focus from the cognitive to the neural level. Disentangling the different hypotheses remains difficult because syntax, theory of mind, and working memory activate overlapping brain areas. This is most clearly the case for theory of mind and working memory. Both of them, for instance, activate areas of the prefrontal cortex, associated with goal-maintenance and inhibition (Costa et al. 2008; Edin et al. 2009). Complex syntax, for its part, does activate brain areas that are not associated with theory of mind or working memory, such as Broca’s area in the inferior frontal gyrus. Nevertheless, the production of complex hierarchical sentences also depends on long-distance connections with more posterior regions of the brain such as Wernicke’s area (Friederici 2009), which is located in the temporoparietal junction, an area associated with both theory of mind and selective attention (Saxe & Kanwisher 2003; Decety & Lamm 2007).

194 Benoît Dubreuil
The presence of such overlapping areas creates an interesting conundrum for students of human evolution. If the evolution of language was related to a change in the temporoparietal junction, it might have coincided with the evolution of theory of mind and working memory. By contrast, if it was related to a change in Broca’s area, it is unlikely to have coincided with the evolution of these abilities, but it might have coincided with the evolution of other abilities that depend on hierarchical action sequencing (Koechlin & Jubault 2006; Stout & Chaminade 2009). Similarly, if the evolution of theory of mind was related to a change in the dorsal prefrontal cortex, it is likely to have coincided with a change in working memory, but not if it was related to the reorganization of a region that is not typically linked to working memory (such as the medial prefrontal cortex, for instance).
In sum, although different hypotheses have been proposed to account for the evolution of the modern mind, these hypotheses do not have distinct implica-tions because they refer to abilities realized by overlapping cognitive and neural mechanisms.
6. From mental phenomena to mental mechanisms
In a book on explanations in cognitive neuroscience, philosopher William Bechtel discusses epistemological problems that have appeared in research on memory and that, I suggest, are also relevant for research on the evolution of the human mind. Bechtel (2008) argues that psychologists working on memory first attempted to distinguish different types of memory. They established distinctions, for instance, between long-term and short-term memory, semantic and episodic memory, implicit and explicit memory, etc. Although they managed to identify distinct and robust phenomena at the psychological level, problems appeared when they tried to identify the cognitive and neural mechanisms that produced these phenomena. The different types of memory appeared not only to activate largely overlapping brain areas, but also to activate areas related to other processes, such as perception or motor control. Bechtel argues that the decomposition of memory processes have pointed toward a characterization of memory that is largely orthogonal to the phenomenal distinctions established by psychologists:
“Although the view that memory involves storing information in a location separate from where it might be processed (as references to memory stores suggest) seems compelling from the perspective of devising an artificial mind, it looks increasingly less plausible when considering the range of areas that seem to be activated in memory tasks as well as in other cognitive activities. An alternative view is that remembering is simply one aspect of various mental phenomena, that

Chapter 10. The other middle-range theories 195
as the mind performs the operations that generate those phenomena, it is altered in ways that alters future performance and in some cases allows it to remember what has happened to it.” (Bechtel 2008: 83)
What parallel can be drawn between research on memory and research on the evolution of the human mind? As with early memory research, the study of cognitive evolution has been largely centred on distinctive and well-studied mental phenomena. Recursive syntax, working memory, and theory of mind, I suggest, have received so much attention because they refer to well-studied cognitive tasks in which humans’ performance is robustly different from that of nonhuman primates. As in the case of memory, however, the clear distinction between these abilities is blurred when we consider how they are produced at the cognitive and neural levels. As explained above, recursive syntax, theory of mind, and working memory activate largely overlapping brain areas. To be sure, some areas are not activated by all three abilities, but these areas also perform operations that go well beyond the specific ability that the theories appeal to. Broca’s area, for instance, is involved in the production of complex sentences, but also in other behaviours that require hierarchical sequencing.
The lesson to be drawn from memory research is, I think, straightforward. Recursive syntax, theory of mind, and working memory refer to phenomena at a much too high level to be useful for explanation. We need to move toward more specific cognitive mechanisms as they are realized in specific brain areas. This objective, however, is not without its own difficulties, which we must be aware of. Memory research has been able to move toward finer-grained accounts of memory because it had access to the powerful tools of cognitive neuroscience. Above all, it could build on imaging techniques that make it possible to localize cognitive functions in brain structures. Students of human evolution must learn from cognitive neuroscience, but they also have to be aware that the architecture of the modern brain could have evolved along various pathways and that it is impossible to decide between them on the sole basis of what we know – or what we may know – of brain structures and cognitive functions in normal functioning modern Homo sapiens. Neuroimaging studies, for instance, are of no help in deciding if Broca’s or Wernicke’s area took their modern form in Homo erectus or Homo heidelbergensis. To produce such inferences we need different tools, tools which are more adapted to our object, but which come with their own limitations.
7. Toward palaeocognitive neurosciences
Students of human cognitive evolution will never directly benefit from the powerful imaging techniques that have revolutionized cognitive neurosciences. But that is

196 Benoît Dubreuil
not to say that there is no way to support specific inferences about the evolution of the brain. In this section, I present the three most relevant tools for our purpose.
7.1 Human and nonhuman primate comparative neuroscience
Although it is impossible to scan the brain of extinct hominins, it is still possible to compare cognitive functions and neural structures in humans and nonhuman primates. Comparative neuroscience can provide hints as to which cognitive and neural mechanisms evolved in the human lineage. Comparative lesion and imaging studies, for instance, indicate that visual working memory pathways in the parietal and temporal cortices have been displaced in the course of human evolution, leaving more space in- between for the parietotemporal areas dedicated to high-level cognitive abilities (Ungerleider et al. 1998). Comparative neuroanatomic studies have also yield interesting results. They have shown, for instance, that the human frontal lobe is not unexpectedly large for a primate of our brain size, and thus it did not benefit more form encephalization than other parts of the neocortex (Semendeferi et al. 2002). They have also highlighted the need for understanding the role of white matter in encephalization and its importance in establishing connections both between close and distant areas of the brain (Schenker et al. 2005; Schoenemann et al. 2005; Rilling et al. 2008). The obvious limitation of comparative neuroscience is that it says nothing about when different mechanisms might have evolved in the human lineage, or how they evolved. For this, we need to turn to other tools.
7.2 Developmental neuroscience
Development studies are both essential and dangerous for understanding the evolution of the human mind. On the one hand, it is often reasonable to assume that cognitive and neural mechanisms that develop later in childhood evolved more recently. This is especially the case when the later-developing mechanisms somehow depend on the normal functioning of the earlier-developing ones. It is not accidental, for instance, that the multimodal association areas develop after the different sensory cortical areas. Without an input from unimodal areas, there would simply be no multimodal information to process. On the other hand, there are well known caveats to the idea that ontogeny recapitulates phylogeny. One of them is that it is not strictly impossible for an early-developing trait or mechanism to evolve later, if it functions in a more or less modular fashion. Another is that, given the plasticity of the brain, later-evolving mechanisms can evolve first and then see their functioning altered by the later evolution of an early-developing mechanism. Despite these limitations, developmental neuroscience can function as a useful heuristic. It makes sense, for instance, to argue that the basic motivations

Chapter 10. The other middle-range theories 197
to communicate and to share attention with others evolved before more complex features of language, because they are at the foundation of language acquisition in modern humans (Tomasello 2008). At the same time, it also makes sense to argue that metalinguistic abilities evolved late in the human lineage, after recursive syntax, because they also develop later in human children (Dubreuil 2008). As for comparative neuroscience, however, developmental neuroscience is not likely to indicate when different mechanisms evolved in the human lineage.
7.3 Palaeoneurology
To address the when question, we have to turn to the study of endocasts. To be sure, cognitive functions are not written in endocasts, and inferences about brain evolution can be drawn only indirectly. Moreover, the paucity of the fossil record for crucial phases of human evolution (and especially for the evolution of modern Homo sapiens) often condemns researchers to base their interpretation on a few incomplete specimens. But we should not minimize the information that endocasts can convey. They are likely to provide indication about relative and absolute brain size, about the relative size and shape of different lobes, about gyrification, and, potentially, about the pace of brain growth in children. The study of endocasts alone, for instance, gives no principled way to decide if encephalization must be considered in absolute or relative terms (total brain mass or encephalization quotient). It indicates, however, that encephalization in early members of the genus Homo was more significant in absolute than in relative terms (given the larger body size of Homo ergaster and Homo erectus), while encephalization in Homo heidelbergensis was significant in both absolute and relative terms (Rightmire 2004). If encephalization was selected for its impact on cognition, as we can reasonably assume, then middle-range theories based on encephalization have more weight for Homo heidelbergensis than for early members of the genus Homo. The same principle applies to modern Homo sapiens. Different arguments have been made to link the evolution of the modern cranium with a relative expansion of the temporal and parietal lobes (Liebermann et al. 2002; Bruner et al. 2003). Although changes in behaviour are not yet unambiguously connected to the evolution of the cranium, palaeoneurology adds weight to explanations that link the evolution of the modern mind to the evolution of these lobes.
Comparative neuroscience, developmental neuroscience, and palaeoneurology are useful tools, but their real strength becomes apparent only when they are used together to explain behavioural transitions that can be documented in the archaeological record. Christopher Henshilwood and I (Henshilwood & Dubreuil 2009, 2011), for instance, have used these different tools to link the transition to “modern behaviours” to a change in the temporoparietal areas of the brain and

198 Benoît Dubreuil
to the evolution of attentional abilities underlying high-level perspective taking and theory of mind. This hypothesis provides a parsimonious account of the evolution of material culture (including the appearance of increased formalism, of symbolism, and style) and it can find support in the different tools described above. We know, for instance, that there are significant differences in theory of mind and perspective-taking abilities in humans and nonhuman primates and that there are structural and functional differences in the parietotemporal areas (comparative neuroscience). High-level theory of mind and perspective-taking abilities develop relatively late in infancy, significantly later than joint attention and syntactic recursion, and depend on the maturation of areas dedicated to cognitive control and multimodal association in the prefrontal, temporal, and parietal cortices (developmental neuroscience). Finally, the evolution of the modern cranium was not linked to a general encephalization process, but to a specific expansion of the temporal and parietal lobes (palaeoneurology).
Future research in archaeology and neuroscience may invalidate this hypothesis or parts of it, but it may also strengthen it or push us toward new refinements. As a middle-range theory, however, I would like to stress that it does not face the same difficulty as the three hypotheses discussed in the previous section. By contrast with them, it is not focused on a complex mental phenomenon such as theory of mind or working memory, but on an attentional mechanism that is involved in different tasks (including theory of mind, perspective taking, and categorisation tasks) and that is realized in a relatively precise area of the brain, one which is likely to have undergone recent evolution in the human lineage.
8. Conclusion and caveats
In this chapter, I have made the case that students of human cognitive evolution should move away from the mental phenomena that they have focused on in recent years and turn their attention to more precise cognitive mechanisms realized in specific neural structures. As became apparent in the last section, I am optimistic that significant progress can be made if we learn to make optimal use of the appropriate neurological tools. In conclusion, I want to mention two reasons for not being too optimistic about the kind of middle-range theories that can be proposed.
The first is that moving beyond coarse middle-range theories will appear as a lasting challenge. Archaeology and palaeoneurology, as well as comparative and developmental neuroscience provide useful guidance, but they are consis-tent with several different evolutionary histories in the fine-grained study of the cognitive and neural (not to say genetic) organisation of the brain. It is one thing,

Chapter 10. The other middle-range theories 199
for instance, to say that the temporoparietal areas are likely to have undergone evolutionary changes along with the evolution of modern Homo sapiens, but it is another to explain with sufficient detail what changes they underwent or how these areas were organized in closely related human populations such as Homo heidelbergensis or Neandertals. This problem, however, is not proper to human evolution. Even high-resolution neuroimaging techniques are often insufficient to detail the functional organization of specific brain areas (including the temporo-parietal junction). We have no alternative but to learn the optimal use of available tools, while remaining conscious of their limitations.
The second reason has to do with causality. Humans are complex organisms who live in rich and changing social environments. Because of this, it is not obvious how we are to determine the properly causal role of genetic, neural, and cognitive factors in human evolution. It is one thing to infer that significant neural or cognitive changes occur at different times in human evolution, but it is another to say that these changes caused behavioural evolution. The problem is obviously that, in Baldwinian evolutionary processes, changes in social environments are also likely to cause biological evolution. As a consequence, biology cannot be straightforwardly taken as the prime mover of human evolution. This problem must also be considered in the wider context of human neural and cognitive plasticity, which prevents us from concluding that causation goes from genes, to neurons, to cognition, and to behaviour. For the time being, and maybe forever, students of cognitive evolution should be content with establishing patchy (though reliable) correlations between neural, cognitive, and behavioural evolution, without claiming too much about causality.
To conclude on an additional note of caution, it should be remembered that the validity and the relevance of middle-range theories depend on their ability to fit archaeological data. Non-archaeologists can become quite enthusiastic about their evolutionary scenarios and models. This is understandable, given the kind of emotional processes that give us the motivation to engage in foundational research, but we have to bear in mind that the validity of our scenarios and models always depend on archaeological evidence, which is, and will for the most part remain, open to several conflicting interpretations.
Most authors who have dealt with the evolution of the human mind have tended to adopt very standard – not to say dominant – interpretations of the archaeological record. Most evolutionary scenarios have been based on the assumption that modern Homo sapiens is the only human population to have ever evolved modern-like cognition and culture, following a cognitive and bio-logical revolution that occurred about 200–50 ka in Africa, and was followed by modern humans’ expansion around the globe. Although this interpretation is supported by much of the data, we must remain conscious of the numerous

200 Benoît Dubreuil
uncertainties that remain to be clarified, including many that might have a deci-sive impact on the middle-range theories that we can devise in order to link neural, cognitive, and behavioural data. Did Neandertals use symbolic artefacts and in what context? Did “modern behaviour” in Africa appear as a package or gradually? Are early occurrences of ochre in the Middle Stone Age evidence of symbolic behaviour? Is the modernisation of Homo sapiens’ cranium concomi-tant with the emergence of modern sapiens behaviour, whatever that means? Although we all have our preferred answers to these questions, we must admit that they remain, for the most part, open. In this context, it might not be a bad idea to develop not one, but many middle-range theories to link neural, cogni-tive, and behavioural evolution, in order to test them afterwards against new archaeological findings.
Acknowledgements
I thank Luc Faucher, as well as the participants at the symposium Homo symbolicus in Cape Town, for comments on an earlier version of this chapter. I am grateful to the Templeton Foundation for the invitation to the symposium and to Chad Horne for comments and editorial support. This work was supported by a postdoctoral fellowship from the Fonds québécois de la recherche sur la société et la culture.
References
Baddeley, A.D. 2001. “Is working memory still working?” American Psychologist 56 (11): 851–864.
Baddeley, A.D. & Hitch, G. 1974. “Working memory.” In The psychology of learning and motivation: Advances in research and theory, G. H. Bower (ed.), 8: 47–89. New York: Academic Press.
Bechtel, W. 2008. Mental mechanisms: philosophical perspectives on cognitive neuroscience. New York: Routledge.
Bickerton, D. 2003. “Symbol and structure: A comprehensive framework for language evolution.” In Language evolution, M. H. Christiansen & S. Kirby (eds), 77–93. Oxford: Oxford University Press.
Binford, L.R. 1977. “Introduction.” In For Theory Building in Archaeology: Essays on Faunal Remains, Aquatic Resources, Spatial Analysis, and Systemic Modeling, L.R. Binford (ed.), 1–10. New York: Academic Press.
Binford, L.R. 1981. Bones: Ancient Men and Modern Myths. New York: Academic Press.Bruner, E.M. G., Manzi, G. & Arsuaga, J.L., 2003. “Encephalization and allometric trajectories in
the genus Homo: Evidence from the Neandertal and modern lineages.” Proceedings of the National Academy of Sciences 100: 15335–15340.
Call, J. & Tomasello, M. 2008. “Does the chimpanzee have a theory of mind? 30 years later.” Trends in Cognitive Science 12 (5): 187–192.

Chapter 10. The other middle-range theories 201
Calvin, W.H. & Bickerton D. 2000. Lingua ex machina: reconciling Darwin and Chomsky with the human brain. Cambridge, Mass.: MIT Press.
Carlson, S.M., Mandell, D.J. & Williams, L. 2004. “Executive function and theory of mind: stability and prediction from age 2 to 3.” Developmental Psychology 40: 1105–1122.
Coolidge, F.L. & Wynn, T. 2001. “Executive functions of the frontal lobes and the evolutionary ascendancy of Homo sapiens.” Cambridge Archaeological Journal 11: 255–260.
Coolidge, F.L. & Wynn, T. 2004. “A cognitive and neuropsychological perspective on the Chatelperronian.” Journal of Anthropological Research 60: 55–73.
Coolidge, F.L. & Wynn, T. 2005. “Working memory, its executive functions, and the emergence of modern thinking.” Cambridge Archaeological Journal 15 (1): 5–26.
Coolidge, F. & Wynn, T. 2007. “The working memory account of Neandertal cognition-How phonological storage capacity may be related to recursion and the pragmatics of modern speech.” Journal of Human Evolution 52 (6): 707–710.
Corballis, M.C. 2004. “The origins of modernity: Was autonomous speech the critical factor?” Psychological Review 111: 543–552.
Costa, A., Torriero, S., Olivieri, M. & Caltagirone, C. 2008. “Prefrontal and temporo-parietal involvement in taking others’ perspective: TMS evidence.” Behavioral Neurology 19 (1–2): 71–74.
Craver, C.F. 2007. Explaining the brain: mechanisms and the mosaic unity of neuroscience. New York: Oxford University Press; Oxford: Clarendon Press.
De Villiers, J.G. & Pyers, J.E. 2002. “Complements to cognition: a longitudinal study of the relationship between complex syntax and false-belief understanding.” Cognitive Development 17: 1037–1060.
Deacon, T.W. 1997. The symbolic species: the co-evolution of language and the brain. New York: W.W. Norton & Co.
Decety, J. & Lamm, C. 2007. “The Role of the Right Temporoparietal Junction in Social Interaction: How Low-Level Computational Processes Contribute to Meta-Cognition.” Neuroscientist 13: 580–593.
Dubreuil, B. 2008. “What do modern behaviors imply for the evolution of language?” In The Evolution of Language, A.D.M. Smith, K. Smith & R. Ferrer i Cancho (eds), 99–106. Proceedings of the 7th International Conference (Evolang 7), Singapore: World Scientific.
Dunbar, R.I.M. 2003. “The social brain: mind, language and society in evolutionary per-spective.” Annual Review of Anthropology 32: 163–181.
Dunbar, R.I.M. 2004. The human story: A new history of mankind’s evolution. London: Faber and Faber.
Dunbar, R.I.M. 2007. “The Social Brain and the Cultural Explosion of the Human Revolution.” In Rethinking the Human Revolution, P. Mellars, K. Boyle, O. Bar-Yosef and C. Stringer. (eds), 91–98. Cambridge: McDonald Institute Monographs.
Edin, F., Klingberg T., Johansson, P., McNab, F., Tegnér, J. & Compte, A. 2009. “Mechanism for top-down control of working memory capacity.” Proceedings of the National Academy of Sciences 97 (7): 3573–3578.
Friederici, A.D. 2009. “Pathways to language: fiber tracts in the human brain.” Trends in Cognitive Sciences 13 (4): 175–181.
Gerrans, P. & Stone, V.E. 2008. “Generous or Parsimonious Cognitive Architecture? Cognitive Neuroscience and Theory of Mind.” British Journal for the Philosophy of Science 59 (2): 121–141.

202 Benoît Dubreuil
Hauser, M.D., Chomsky, N. & Fitch, W.T. 2002. “The faculty of language: What is it, who has it, and how did it evolve?” Science 298: 1569–1579.
Henshilwood, C.S. & Dubreuil, B. 2009. “Reading the artifacts: Gleaning language skills from the Middle Stone Age in southern Africa.” In The Cradle of Language, Volume 2: African perspectives, R. Botha & C. Knight (eds), 41–61. Oxford: Oxford University Press.
Henshilwood, C.S. & Dubreuil, B., 2011. “The Still Bay and Howiesons Poort, 77–59 ka: Symbolic material culture and the evolution of the mind during the African Middle Stone Age.” Current Anthropology. 52 (3): 361–400.
Jurado, M.B. & Rosselli, M. 2007. “The elusive nature of executive functions: a review of our current understanding.” Neuropsychological Review 17 (3): 213–233.
Klein, R.G. 2008. “Out of Africa and the evolution of human behavior.” Evolutionary Anthropology 17 (6): 267–281.
Koechlin, E. & Jubault, T. 2006. “Broca’s area and the hierarchical organization of human behavior.” Neuron 50 (6): 963–974.
Krause, J., Lalueza-Fox, C., Orlando, L., Enard, W., Green, R., Burbano, H., Hublin, J–J., Ha¨nni, C., Fortea, J., de la Rasilla, M., Bertranpetit, J., Rosas, A. & Pa¨a¨bo, S. 2007. “The derived FOXP2 variant of modern humans was shared with Neandertals.” Current Biology 17: 1–5.
Lieberman, P. 2007. “The Evolution of Human Speech: Its Anatomical and Neural Bases.” Current Anthropology 48 (1): 39–66.
Lieberman, D.E., McBratney, B.M. & Krovitz, G. 2002. “The evolution and development of cranial form in Homo sapiens.” Proceedings of the National Academy of Sciences 99: 1134–1139.
Marcus, G.F. 2008. Kluge: The haphazard construction of the human mind. Boston: Houghton Mifflin.
Mellars, P. 2005. “The impossible coincidence. A single-species model for the origins of modern human behavior in Europe.” Evolutionary Anthropology 14: 12–27.
Mitchell, S.D. 2003. Biological complexity and integrative pluralism. Cambridge, UK: New York, N.Y.: Cambridge University Press.
Mithen, S. 1996. The Prehistory of the Mind. London: Thames & Hudson.Read, D.W. 2008. “Working memory: A cognitive limit to non-human primate recursive
thinking prior to hominid evolution.” Evolutionary Psychology 6 (4): 676–714.Rightmire, G.P. 2004. “Brain size and encephalization in Early to Mid-Pleistocene Homo.”
American Journal of Physical Anthropology 124: 109–123.Rilling, J.K., Glasser, M.F., Preuss, T.M., Ma, X., Zhao, T., Hu X. & Behrens, T.E. J. 2008. “The
evolution of the arcuate fasciculus revealed with comparative DTI.” Nature Neuroscience 11: 426–428.
Saxe, R. & Kanwisher, N. 2003. “People thinking about thinking people: the role of the temporoparietal junction in theory of mind.” NeuroImage 19: 1835–1842.
Schaffner, K. 1998. “Genes, behavior, and developmental emergentism: One process, ndivisible?” Philosophy of Science 65: 209–252.
Schenker, N.M., Desgouttes, A.M. & Semendeferi, K. 2005. “Neural connectivity and cortical substrates of cognition in hominoids.” Journal Human Evolution 49: 547–569.
Schoenemann, P.T. & Sheehan, M.J. 2005. “Prefrontal white matter volume is disproportionately larger in humans than in other primates.” Nature Neuroscience 8: 242–252.
Semendeferi, K., Lu, A. Schenker, N. & Damasio, H. 2002. “Humans and great apes share a large frontal cortex.” Nature Neuroscience 5 (3): 272–276.
Stout, D. & Chaminade, T. 2009. “Making tools and making sense: complex, intentional behaviour in human evolution.” Cambridge Archaeological Journal 19 (1): 85–96.

Chapter 10. The other middle-range theories 203
Suddendorf, T. & Corballis, M.C. 2007. “The evolution of foresight: What is mental time travel and is it unique to humans?” Behavioral and Brain Sciences 30 (3): 299–313.
Tomasello, M. 2008. Origins of human communication. Cambridge, Mass.: MIT Press.Ungerleider, L.G., Courtney, S.M. & Haxby, J.V. 1998. “A Neural System for Human Visual
Working Memory.” Proceedings of the National Academy of Sciences 95 (3): 883–890.Wynn, T. & Coolidge, F.L. 2004. “The skilled Neanderthal mind.” Journal of Human Evolution
46: 467–487.Wynn, T. & Coolidge, F.L. 2007. “Did a small but significant enhancement in working-memory
capacity power the evolution of modern thinking?” In Rethinking the Human Revolution, P. Mellars, K. Boyle, O. Bar-Yosef, & C. Stringer (eds), 79–90. Cambridge: McDonald Institute Monographs.
Zilhão, J. 2006. “Neandertals and Moderns Mixed, and It Matters.” Evolutionary Anthropology 15: 183–195.


chapter 11
Metarepresentation, Homo religiosus, and Homo symbolicus
Justin L. BarrettCentre for Anthropology & Mind Institute of Cognitive & Evolutionary Anthropology, University of Oxford
What cognitive adaptation enabled humans to become the distinctively symbolic species that we are? Drawing upon insights from the cognitive sciences and evolutionary psychology, research in cognitive science of religion (CSR) converges on the claim that the ability to form mental representations about mental representations (metarepresentation) is a key factor enabling and encouraging religious expression. Such metarepresentation may also be the key to symbolic behaviour – including linguistic expression – more generally. If so, then the same cognitive equipment that underwrites symbolism also gave rise to religion, and the two could have evolved concurrently.
1. Introduction
Cognitive science of religion (CSR) has drawn from evolutionary psychology and cognitive development to generate a number of theories regarding the cognitive architecture necessary for various religious ideas and practices. A review of these theories suggests that one common factor that propels proto-religious thoughts and impulses into the full-blown religion that we see in modern humans is metarepresentation – the ability to form mental representations of mental representations. For instance, it may be that basic belief in the afterlife is a byproduct of two different cognitive systems with different evolutionary and developmental histories – one that deals with bodies and one that deals with minds. But metarepresentation may be necessary for afterlife beliefs to transform into ancestor-spirit cults. Similarly, an agency detection device that is tuned to be hypersensitive may lead to the postulation of intentional beings with supernatural properties, but metarepresentation may be required to support beliefs and practices of interactions with these supernatural beings.
Metarepresentation may likewise serve as the critical factor in transforming other cognitive capacities into more sophisticated systems that can sustain cumu-lative cultural traditions including symbolic ones, and may be a distinctive feature of modern humans.

206 Justin L. Barrett
In this essay I begin by considering current theories of the evolution of religious cognition in humans and argue for the importance of metarepresentation as a critical capacity in religious thought. In the following section I extend the discussion to other cultural domains in which metarepresentation seems to play a key role, including cultural expression that appears to require symbolism. I conclude by discussing what the proposed account of religious and symbolic evolution might mean for archaeological studies.
Throughout I adopt the standard practice in CSR of treating ‘religion’ as ideas, practices, and commitments that are distributed across individuals, which pertain to the affirmed existence of at least one form of minded agency that deviates to some degree from the natural intuitions delivered by ordinary human cognitive systems. That is, religion concerns counterintuitive intentional agency. Such a definition is close to the common view that ‘religion’ has something to do with gods, spirits, ghosts, and the like. I recognize that such a definition carries the liability of being narrower than the myriad phenomena scholars may include under the ‘religion’ umbrella. It carries the strength, in this occasion, of picking out a set of phenomena that may be causally and explanatorily coherent from a cognitive perspective. I hope this working definition and its virtues in this context will become clearer below.
2. Metarepresentation and religious evolution
Metarepresentation, the ability to deliberately consider, ponder, or evaluate mental representations, that is, thinking about thoughts, may be the lynch-pin that transforms many other cognitive capacities into those attributes that consti-tute human distinctiveness. Current scholarship in evolutionary and cognitive studies of religion recurrently implicates or presumes a critical role for metare-presentation in explaining the cross-cultural recurrence of religious beliefs and practices.
Darwinian evolution raises at least four alternative possibilities concerning when and how religion might have arisen, when Homo might have become Homo religiosus: (1) the Standard View, that there is nothing in human nature that predisposes us or specially accounts for religious thought and behaviour; (2) the Adaptation Hypothesis that religion is an evolved trait on top of and later than the evolution of whatever it is that makes us behaviourally modern humans; (3) the Pre-Human Religion Hypothesis that first religion evolved as an adaptation and then later behaviourally modern humans evolved; and (4) the Concurrence

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 207
Hypothesis, that whatever it is that makes us behaviourally modern humans is the same thing that makes us religious.1
3. The Standard View
The Standard View is the view that there is nothing about human nature that makes us prone to be religious or to believe in gods. Religion is something of a historical-cultural accident analogous to the invention of modern science or the game of chess. Though we are capable of these forms of cultural expression, they might have never happened. Had someone not invented chess, there would not be any chess players. Had no one invented religion, people wouldn’t be religious.
On this view, religion does not have any special place in human nature and it surely is not an adaptation. The Standard View is rooted in the idea that human cognition provides a bias-free clean slate for experience – including cultural expe-riences – to write on unhindered. The environment largely dictates who and what we are. In some of their writings, Richard Dawkins (Dawkins 2006) and Nicholas Humphrey (Humphrey 1998) seem to express this Standard View, suggesting that if children were not indoctrinated to believe in religion it would disappear.2
I regard the Standard View as problematic. Its presumption that human minds can be likened to blank slates has been demonstrated to be erroneous many times over. It is simply not the case that humans can learn any information equally well, or are equally likely to acquire any ideas or practices they might be exposed to.
Such a simplistic view of human cognition and behaviour, though still common in neighbouring fields, has held no serious place in psychological sciences since the so-called cognitive revolution that began in the 1950s and by the 1970s had overthrown behaviourism’s empiricist regime. Though there is still lively disagreement about just what sorts of biases human minds naturally have, how they develop, and so forth, that ordinary human biological endowment plus ordinary features of the world yields a predictable, natural mental tool kit, is not
1. Strictly speaking, it could be that becoming religious and becoming modern humans occurred simultaneously due to the co-evolution of two different traits. That is, religion might be an adaptation that happened to evolve at the same time as other adaptations that make modern humans what they are. For the sake of this presentation, I consider such a possibility as a limiting case of both the Adaptationist and the Pre-Human Religion Hypotheses.
2. Dawkins (2006), Chapter 9, especially seems to suggest that the only critical disposition toward religious belief is evolved gullibility. In other parts of his book such as Chapter 5, however, he does take a more receptive stance toward religion having some natural anchors.

208 Justin L. Barrett
controversial. Rather than a sponge that soaks in anything or a blank slate ready for any scribbling, the human mind is more usefully likened to a Swiss Army knife possessing numerous functional units for solving particular problems.3
The Standard View also looks suspect given some striking divergences from other ‘arbitrary’ non-natural forms of cultural expression such as modern science or playing chess. Cultural expression that is largely unbuckled from natural biases or predilections tends to be extremely variable within and across cultures. So people and some cultures simply do not do modern science or play chess. In contrast, religious thought and action-perhaps particularly belief in some kind of supernatural agency – is ubiquitous across cultures (Atran 2002). But it isn’t just the greater recurrence of religious thought (as compared with chess playing or modern science) that suggests it is a different kind of cultural expression. We might also point to its relative ease of transmission even to very young children, and its resilience in the face of deliberate attempts to eliminate it.
Arbitrary cultural forms tend to be acquired later in life than the more naturally anchored sort of cultural expression. Doing algebra or modern science, playing chess or riding a bicycle all are forms of cultural expression that generally arrive late enough in our lifespan (if at all) that we remember not doing them. Walking, speaking, and entertaining religious thoughts, in contrast, can be picked up so early in childhood that we might not remember not doing them. Related, arbitrary cultural expression generally requires explicit instruction to acquire. Intensive training is usually required to do science, whereas doing religion appears to be almost caught like the common cold. We can just pick it up from observing those around us, much like how we just pick up our native language.
These sorts of considerations regarding the resilience, the ubiquity, and the ease with which even young children acquire religion has prompted many scholars to explore why it is that humans seem prone to religious expression (Atran 2002; Barrett 2000, 2004; Boyer 2003; Pyysiäinen 2001; Tremlin 2006). Why is it so common? Why is it so easy for children to acquire? In short, why does it seem that religion is a part of human nature? That scholars are making progress on these sorts of questions is evidence in itself that the Standard View is mistaken.
4. The Adaptation Hypothesis
The Adaptation Hypothesis is that religious belief and practice amount to an evolved propensity that arose after our ancestors had become behaviourally
3. Steven Pinker gives a good overview of this more content-rich view of the human mind (Pinker 1997).

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 209
modern humans. That is, religion is something extra added that was added because it helped our ancestors survive and reproduce better than competing modern humans lacking religion.
Adaptationist arguments tend to focus on intra-group cooperation in one of two ways: the ability for morally interested supernatural agents to police human action in social arrangements (Bering & Johnson 2005), and the role of rituals in stimulating intra-group trust (Alcorta & Sosis 2005; Wilson 2002).4 Both types of argument begin with some common premises. It is an advantage for members of a social group to share and trade. Sometimes our hunting, gathering, or farming is more successful than other times. If we share with each other then we level out boom-and-bust cycles for everyone. Likewise, if I am a great hunter but you are a great clothing maker, it might be best for both of us for me to give you meat in exchange for clothing. But such resource sharing typically requires a degree of trust. Will you really give me meat if I fail to find game when you are successful? Or will you hide what you caught and not tell me about it? Or knowing that you are better at gathering than I am and a generous sharer, why should I work so hard? Maybe I should do just enough work so you don’t think I am a free-loader. Cooperation has potential costs, so trust is important. I need to know that you won’t cheat me.
One way of handling the free-rider problem is to punish cheaters when they are caught. The downsides are that you have to expend energy catching and punishing cheaters, and cheaters could simply get better at not getting caught. Further, some people might not punish cheaters because it is too costly to them so then non-punishers would have to be punished for not punishing and so on. Enter the gods.
Dominic Johnson and Jesse Bering have suggested that gods help arrest this punishment regress (Bering & Johnson 2005). If gods do the punishing, we humans don’t have to do so much. Another up-side of gods policing human behaviour is that gods – by sake of being invisible, mind-reading, all-knowing, omnipresent, or able to see through walls – know with greater fidelity who is a cheat and who isn’t. They have what Pascal Boyer has called full access to strategic information about humans (Boyer 2001, 2003). People that believe in a morally interested, full-access agent such as a god are less likely to cheat on you.
4. Harvey Whitehouse has considered religious ritual participation as a means for forming coalitional commitment and trust with highly emotional, rarely performed rituals (such as initiation) leading to small, intensely cohesive groups, and repetitive, lower-arousal rituals leading to larger communities (Whitehouse 2000; Whitehouse 2004). Note, however, that Whitehouse does not regard these dynamics as evidence of religion being an adaptation, but emphasizes its role in cultural evolution.

210 Justin L. Barrett
So, they are more reliable sharing and trading partners and more people will want to share and trade with them. Hence, they have better access to resources than less trustworthy individuals. This resource advantage will make them fitter to survive and reproduce, and so, their genes that make them god-believers will be reinforced through natural selection.
Similarly, it may be that even aside from policing supernatural agents, the existence of a transcendent realm or of gods that our group recognizes can serve as the hub for ritual performances. These ritual performances, because they do not obviously have the direct utility of ordinary actions, can serve to signal commitment to the in-group. One’s participation in a ‘costly’ ritual display may signal to others that one is a true believer in the group’s values and practices (Alcorta & Sosis 2005; Ruffle & Sosis 2007; Sosis 2003). These signalling displays may then encourage trust and cooperation, leading to fitness benefits. The underlying gene-culture complex then would be selectively reinforced. A variation of the Adaptation Hypothesis has been advanced by David Sloan Wilson (Wilson 2002). He argues that religious groups out compete non-religious groups, and so the genes of groups with a propensity toward religious belief and action will be selectively promulgated.
Two sorts of evidence would be helpful in supporting variants of the Adapta-tion Hypothesis: evidence that there is a particular gene that promotes religiousness and, more importantly, evidence that being religious really is adaptive. Is there such evidence?
On the first score-is there a religion-or god-gene?-, sensational titles and headlines aside, the evidence to date is pretty thin. Certainly there appears some heritable component in what has been labelled ‘religiosity’, but whether this is a disposition toward believing in gods or ritualism or in-group conformity or some other feature of modern religiosity in contemporary western societies is unclear. In his book, The God Gene, Dean Hamer admits that the gene that he has identified that is related to some aspects of religious identification manages to explain only a very small fraction of the variance (Hamer 2004). That is, this ‘god gene’ is a long way from accounting for human religious propensities. So far then, evidence for a particular genetic adaptation that makes people religious (who otherwise would not be) is far from compelling.
The Adaptation Hypothesis does much better with regard to the second type of evidence. We do have reason to believe that being religious is adaptive, particularly that it may promote group cohesion and cooperation. Anthropologist Richard Sosis has produced several studies showing that religious communities often out-survive non-religious ones, and are marked by higher levels of cooperation. For instance, in one study Sosis compared religious and secular kibbutz communities in Israel. Members of religious kibbutzim had higher levels of trust on economic

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 211
games, particularly if they were regular participants in religious rituals (Sosis & Ruffle 2003). Likewise, religious communities appear to have greater survival as communities than comparable secular ones, particularly if there are ‘ritualized’ behaviours associated with membership (Sosis & Bressler 2003). Psychologists have recently shown that Belgian university students who were subliminally exposed to religious words acted with more pro-social intention (picking up more brochures to publicize a charitable organization) and were more forgiving of a harsh critic than students not so primed (Pichon et al. 2007; Saroglou et al. 2009). Suppose we accept that belief in gods and the actions that these beliefs motivate are adaptive, it does not follow that religion is an adaptation. Lots of things are or could be adaptive without being adaptations. Yoga may reduce stress and improve fitness, thereby promoting survival and reproductive success, but surely yoga is not a direct product of genetic selection. Having community fire brigades or rescue corps may improve the fitness of a community but they surely are not adaptations. Even something as old and as widespread as clothing making-clearly adaptive for living in cold climates – is an implausible candidate for an adaptation. The idea that behaviours such as clothing making are the result of a genetic mutation when our ancestors already possessed the ability to use tools, detect patterns, reason causally, and solve problems, seems implausible and unparsimonious. Rather, clothing-making was likely an innovative product of other capacities that then has been passed on through cultural transmission. Similarly, it seems unlikely that all that captured by ‘religion’ here is the result of a genetic adaptation. Consider what that would mean. Belief in gods, belief in the afterlife, reasoning as if gods are morally interested in the activity of humans and that they act on that interest to punish or reward, seeing natural phenomena as indicators of divine action, and socially organizing around these beliefs all would be products of the same genetic modification under the same selection pressure.
5. Possible but improbable
If the selective pressure was toward intra-group trust and cooperation for sharing resources, why not a much simpler mutation that made us all hyper-vigilant do-gooders? Or how about some kind of gentle paranoia that somehow whenever we are bad someone is bound to catch us? Introducing religion to solve this cooperation problem seems a little like trying to kill a gnat with a hand grenade. For these reasons, though religion may be adaptive, I suspect we cannot conclude that all of religion is an adaptation. Nevertheless, the possibility that a host of adaptations providing fragmentary preludes to religion were conjoined by a single modest adaptation remains plausible. I consider such a possibility below.

212 Justin L. Barrett
6. The Pre-human Religion Hypothesis
The Pre-human Religion Hypothesis is that at some point before becoming modern humans, our ancestors acquired religion as an adaptation through natural selection. Then, at a later date, they acquired additional adaptations that made them behaviourally modern humans. Some archaeologists argue that Neandertals were religious (Tattersall 1998). This claim is based on evidence that Neandertals occasionally buried their dead with grave goods in a way that suggested ritualized treatment of the remains and (perhaps) a belief in the afterlife. Maybe some Neandertals did have a belief in the afterlife that motivated the seemingly “ritualized” burial practices, or perhaps they simply regarded the bodies and possessions of deceased loved ones as somehow special and uncanny without any worked out ideas about what happens after death. Either way, we don’t have strong evidence that Neandertals had the kind of fully-developed religious beliefs and practices that modern humans have been expressing for thousands of years.
Another non-human species that has been cited as possibly having some kind of religious sentiment is the chimpanzee. Male chimpanzees have been known to make dominance displays of the sort they use in confrontation with other males in response to waterfalls, thunderstorms, and even motorized vehicles (Whiten et al. 2001). Does this mean chimpanzees are pan-omorphizing the natural world much like humans anthropomorphize the natural world in religious contexts? It is hard to know. It could be that chimpanzees have a behavioural routine (the dominance display) that usually gets triggered only in the presence of competing males but occasionally natural (or human made) sights and sounds satisfy the ‘input conditions’ to get these routines going. Likely, these ‘rain dances’ as they have been called, are more similar to horses running from harmless plastic grocery sacks blowing in the wind than to humans regarding hurricanes as divine judgement for misdeeds.
Further, as an adaptationist account, the Pre-human Religion Hypothesis also shares the same liabilities as the Adaptation Hypothesis, and so, for lack of evi-dence and plausibility, I suspect that the Pre-human Religion Hypothesis is false.
7. The Concurrence Hypothesis
The Concurrence Hypothesis suggests that whatever it was that made behav-iourally modern humans, happened to appear at the same time as whatever it was that gave humans a religious disposition. More precisely, those capacities that prompt us to be religious are the same capacities that make modern humans cognitively and behaviourally distinct. Becoming modern humans and becoming Homo religiosus are one and the same. On the surface, this seems an extremely

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 213
unlikely proposition, but let me try to persuade you that the Concurrence Hypoth-esis is a real possibility and perhaps even a more likely possibility than the other hypotheses I offered above.
To build my case, I will quickly sketch three accounts for why humans are naturally disposed to be religious developed by cognitive scientists of religion. What these accounts have in common is that instead of proposing special genetic mutations that give rise to religiousness, they observe that features of ordinary human cognitive systems that might have evolved for entirely different reasons in our ancestral past may encourage the religiousness of modern humans.
8. Anthropomorphism, agency detection, and gods
The cognitive area’s earliest account, developed by anthropologist Stewart Guthrie, is that because of the importance of detecting predators and prey, friends and foe, we have evolved a tendency to over-detect human-like beings in our environment. That is, we have a tendency to anthropomorphize objects and states of affairs in the natural world. Guthrie argues that such a tendency leads to postulating minded, non-human agents as being responsible for various events in the environment; beings more commonly known as ‘gods’.
Considerable research has been devoted to understanding the human Agency Detection Device (ADD) as it has sometimes been called (Barrett 2004). This ADD is regarded as being evolved to detect the presence of minded, intentional beings even under conditions in which evidence is scant or inconclusive. The evolutionary argument for this hair-trigger detection is that the cost of failing to detect a predator or prey, a friend or foe, would be so great that the ADD is better tuned to generate false positives-occasionally detecting agents when there are none-than false negatives-missing agents when they are around. And certainly experimental evidence suggests that infant and adult humans can be led to regard even geometric shapes as minded entities (Rochat et al. 1997). All it takes are certain seemingly non-inertial, goal-directed movements such as when an object launches itself toward another object. So the Agency Detection Device may be hypersensitive under certain conditions, hence it is sometimes called the Hypersensitive Agency Detection Device or HADD.
But this tendency to find goal-directed agents in the environment readily and rapidly is a trait shared by many non-human species. Mammals and birds can readily be tricked into acting as if a non-living object such as a blowing plastic bag or a remote-controlled vehicle is some kind of agent. Maybe they too have some kind of primitive HADD, but this kind of HADD, that only detects objects as potential agents, is insufficient for supporting a relationship with gods. HADD must also register states of affairs as the products of agency even when these states

214 Justin L. Barrett
are not the agents. That is, HADD must see sounds or traces or movement of objects as intentionally brought about.
If I wander through an allegedly haunted house and I hear chains rattling, I might regard the sound of chains as produced by a ghost. The chains are the prod-uct of intentional action but are not the agent. Without this capacity to perceive states of affairs as intentionally caused by an unseen agent, people could not regard events such as thunderstorms or good crops as the gift or curse of a god. Of course, modern humans do have such a capacity and take advantage of it to interact with other people who are not immediately present.
This sort of agency detection is much less clear in non-humans. Perhaps dogs regard the sound of their unseen human masters’ whistle as evidence of intentional agency, but perhaps they more simply develop a behavioural response to the whistle that brings reward. Perhaps animals are shy of human buildings and roads not because they understand them to be signs of human agency, but perhaps instead they just learn to associate these strange things with danger and do not entertain the contents of human mental states.5 Even if we give non-human animals the benefit of the doubt in these situations, humans can do something more complex still. Not only do we regard states of affairs as products of the intentional activity of an unseen agent, but then speculate what the intentions of the unseen agent might have been. Solid evidence that any non-humans have these cognitive capacities is yet to be found. Even if members of Pan and extinct Homo had such capacities, these are still insufficient to be counted as Homo religiosus. The ability to suppose that the thunderstorm was caused by a god and even perhaps because the god is angry is not quite sufficient for religion. Something else is needed. I’ll turn to the something else after sketching more cognitive accounts of religious predisposition.
9. Intuitive dualism and spirits
Another cognitive account is offered by Yale psychologist Paul Bloom (Bloom 2004; Bloom 2007). He notes that developmental psychology has produced evidence that from early in childhood we have two different conceptual systems that both
5. Perhaps the best evidence are reports that chimpanzees break branches or twigs to signal their travel route to other chimpanzees who then successfully use those signs. But even here it is possible to regard these behaviours on the part of chimpanzees in another way, however. Perhaps the following chimpanzees do not see the ‘signs’ as intentionally produced by other chimpanzees but have developed a behavioural routine such that they simply follow these markers to find other chimpanzees much like following the tracks or droppings of an animal or even an object that might have rolled away.

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 215
have to be used to make sense of the behaviour of humans. One system deals with the properties and motion of physical bodies. And one system, called ‘theory of mind’, tries to explain and predict behaviour based on mental states: beliefs, desires, goals, feelings, dispositions, and so forth. The theory of mind system (or ToM) is much newer in terms of both evolution and development. These two systems have to be held together to make sense of human behaviour, but their tenuous relationship leads to a sort of intuitive dualism. That is, we tacitly assume there is some minded part of an individual that is separable from the body. Given this intuitive dualism, the idea that some part of a person can continue existing and even acting after death is natural, says Bloom, and so the slightest encouraging evidence (perhaps provided by HADD) or corroboration from trusted others may be enough to get afterlife beliefs up and running.6 Similarly, Pascal Boyer has proposed that human corpses are perceived in a unique way because of conflicts between several specialized inference systems – notably the animacy system and the person-file that holds details pertaining to a specific known person. Seeing the corpse of a family member prompts uneasiness because the animacy system tells us that the person is not living but the person-file (with the aid of ToM) refuses to stop producing inferences about the dead person’s wants, needs, and beliefs (Boyer 2001). Bloom’s and Boyer’s accounts help explain why the idea that some human spirit survives death and continues to act in the world may be the most widespread and oldest religious concept.
But only believing that the ancestors can continue to act in the world and speculating about their goals, wants, and beliefs does not get us all the way to religion. Something else is still needed, but before I get to that something, allow me one more set of cognitive dynamics that may encourage belief in gods.
10. Naïve creationism and intuitive theism
Psychologist Deborah Kelemen has suggested that children may be intuitive theists because of a different set of cognitive dynamics (Kelemen 2004). She has conducted dozens of experiments with children and adults that show a strong tendency to see natural things such as rocks, plants, and animals as purposefully designed. Her research and that of others has also shown that young children assume that
6. Psychologist Jesse Bering has argued something similar, noting the great difficulty people have in imagining their own lack of all epistemic states. This inability to simulate not having any thoughts or consciousness at all may encourage belief in afterlife, argues Bering. He calls this account the simulation constraint hypothesis (Bering 2006).

216 Justin L. Barrett
purposeful design requires a designer – indeed, until Darwin essentially everyone assumed design requires a designer – and so children are prone toward believing in some kind of intelligent being or beings that account for the natural world. Children quickly learn that humans lack the ability to account for the apparent design in the natural world, so children are prepared to believe the suggestion that some kind of god or gods account for the natural world.
These three accounts-Guthrie’s anthropomorphism account, Bloom’s intuitive dualism account, and Kelemen’s intuitive theism account – each have evidence in their favour and they could be combined. For instance, intuitive dualism might lead us to suppose that some spirit continues after death, the Hypersensitive Agency Detection Device might then encourage us to detect evidence of that spirit’s activity, and the tendency to see the world as designed might encourage us to believe that the ancestor-spirits create and sustain the natural world. Other cognitive factors, too, have been scientifically developed that might buttress religiousness (Barrett 2007). These three accounts all make use of ordinary cognitive architecture that likely evolved for reasons entirely unrelated to religious belief and practice, and indeed, likely evolved, at least in large part, in pre-human species. We would not be too surprised if Homo erectus had a Hypersensitive Agency Detection Device, some kind of basic ToM system that enabled speculation about the mental states of others and leading to intuitive dualism, and even a bias to understand the natural world in terms of purpose and design, perhaps leading to speculations that someone intentionally caused elements of the natural world to be what they are.
All three cognitive accounts, as I have presented them so far, also lack the same extra factor that would produce what we identify as religion in humans. If Homo erectus did have the cognitive capacities identified above, Homo erectus was still not Homo religiosus.
11. Metarepresentation and Homo religiosus
Recall that to be Homo religiosus what is needed is to be able to have a joint rela-tionship with a god or gods. Homo religiosus must be able to act contingently in response to what they think the god’s mental states might be, and they must be able to believe that another shares a similar relation with the same god, otherwise we would have lots of individual, idiosyncratic beliefs and practices instead of faith communities.
The lynch-pin that makes this kind of relational religion possible is the cognitive capacity known as higher-order ToM or, as I prefer, metarepresenta-tional ToM: the ability deliberately and reflectively to think about the contents of other’s thoughts. I call this ‘metarepresentational ToM’ because it is the ability

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 217
to form mental representations about the contents of mental representations. One could think about another’s thoughts about thoughts, including my own thoughts (e.g. “She knows that I know she knows what I want”). Metarepresen-tational ToM opens new doors for thinking about and relating to gods above and beyond simpler ToM abilities. Consider the following:
a. The god has mental states (but I don’t know what they are) that motivate the god’s actions.
b. The god does not like my behaviour and expressed that dislike by sending illness.
c. The god does not like my behaviour and sent illness so that I would know that the god does not like my behaviour and I would change.
d. You know that the god does not like my behaviour and sent illness so that I would know that the god does not like my behaviour and would change.
Sentence (a) expresses a simple awareness that others have mental states that matter in some way to their actions. This represents the most basic mentalizing activity and seems to be a capacity that infants and perhaps non-human primates, dogs, and elephants may possess.
Sentence (b) expresses an understanding of the contents of another’s mental states and how those states are connected to an action. Two year olds and apes may have this ability.
Sentence (c) expresses awareness that other’s mental states may include representations of my mental states, as in ‘He wants me to know…”, and some sense that actions can be motivated by a desire to change other’s mental states and not merely their behaviours. Now we are in range of drawing complex relation-ships among mental states and their relations to actions. It is not clear that any non-human species alive today have this capacity or that humans exhibit it before age three or four.
Sentence (d) takes things one step further still. All that is expressed in (c) is now attributed as knowledge of another. This is very high order ToM but qualitatively may not differ from (c). If I can think about your thoughts about my thoughts, it may be that I have the capacity to think about your thoughts about my thoughts about your thoughts. I just need to be able to represent a thought-about-a-thought as a thought. Even if (d) represents an extension of the same cognitive capacity as in (c), it surely does require more conscious attention and effort. No wonder we have no good evidence that (d)-type thinking appears in any non-humans and is challenging even for adult humans in many situations.
It is (d)-type thinking, being able to understand that others share the same perspective that the gods want to change our thinking and behaving, that is required

218 Justin L. Barrett
for Homo religiosus. With this metarepresentational, ToM humans can acknowledge, worship, and serve the same god and know that they are worshipping the same god. Metarepresentational ToM, then, allows the cognitive accounts of religion to do more than explain why people postulate gods. They may begin to explain why people collectively and jointly can and do interact with the gods in relationship. Metarepresentational ToM would also allow for religious beliefs to facilitate intra-group cooperation and trust: we can know that each other both believe that certain behaviours are punishable by the gods (Dunbar 2004). Similarly, being able to speculate about the mental states, particularly the intentions, of another is essential for reading natural states of affairs as communicative acts. When people regard a famine or storm as an act of a god (perhaps requiring a ritual response), they are speculating that an unseen god has intended to act through these natural events for a particular reason. Such speculation is metarepresentational and skirts very near symbolism as discussed below. Given the pivotal role of metarepresentational theory of mind to religion, it is no surprise that ToM is cited as a critical cognitive component of religion by many cognitive scientists of religion (Barrett 2004; Boyer 2001; McCauley & Lawson 2002).
Metarepresentation in the domain of thinking about mental states may be the extra something that made our ancestors Homo religiosus. I regard metarep-resentational abilities as a more plausible adaptation than ‘religion’ as a whole, because metarepresentation is an incremental extension of regular, lower-order ToM capacities.
12. Metarepresentation and cumulative culture
The reason I regard this possibility as distinct from the Adaptation Hypothesis is that adding metarepresentation to the existing hominin cognitive tool kit would not only produce religion. Together with pre-existing capacities, metarepresenta-tion (particularly in the ToM realm) would yield many abilities and activities that we consider indicative of behaviourally modern humans. The ability to form rep-resentations of representations or thoughts about thoughts has been suggested as the secret ingredient, not just for religion, but cumulative culture more generally. By cumulative culture, I refer to socially-acquired ideas or practices that are read-ily expanded or modified by subsequent generations. Taking a useful tool such as a hand axe and hafting it to make it even more versatile is an example of cumulative culture or what Michael Tomasello calls the ‘ratchet effect’ because such cultural evolution seems to require a holding mechanism to prevent slippage ‘relatively faithful social transmission’ and then a mechanism for innovation and creativ-ity that moves the same goal forward (Tomasello 1999). The proposed holding

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 219
mechanism needed to facilitate strong, relatively error-free cultural transmission is joint attention, the ability for me to know that you know that we are both attend-ing to the same thing. ‘Attending’ here does not merely mean ‘oriented toward’ or even ‘looking at’ but something more like ‘holding in mind’. If we jointly hold in mind the same thing, then I can knowingly communicate information about it. Otherwise, I cannot be confident that any utterances or gestures I make in an effort to communicate about the thing to which I am attending will be understood as bearing on that thing. Likewise, unless you know you are attending to the same thing I am attending to, you would not be able to be confident of the target of my communication. For this reason, Tomasello argues that such joint attention is a prerequisite for human-type language. Note that this sort of joint attention is an instance of metarepresentation.
A more sophisticated form of joint attention may also be part of the mech-anism that moves cultural innovation forward: shared intentionality. By shared intentionality, I mean knowing that another is sharing a particular intention with you. If we both know that we both want to build the same hut, we have shared intentionality. This requires a more challenging form of joint attention because the object of our intention has to be imagined; it is not yet realized. This shared intentionality enables us to work together toward a single goal, vastly improving our capacity for cultural innovation. Here we are dealing with a form of metarep-resentation that may be absent in all non-humans.
Cognitive anthropologist Dan Sperber has similarly argued that the criti-cal adaptation for creative cultural evolution is a ‘module of metarepresentation’ (Sperber 1994). This mental device allows us to take a representation (e.g. as of a plant, animal, tool, or problem) and perform mental operations on it (e.g. what if my staff had a sharp end?). Cognitive archaeologist Steven Mithen similarly proposes that the modern human mind with its capacity for cumulative cultural evolution arose because of increased cognitive fluidity, made possible because of an adaptation enabling various specific mental devices to communicate with each other in a metarepresentational ‘space’ (Mithen 1996).
13. Metarepresentation and Homo symbolicus
The power of metarepresentation to transform preceding capacities into quali-tatively different abilities of cultural expression, including religion, may be clearer if I briefly consider the role of metarepresentation in symbolism and language.
Non-human primates, cetaceans, canids, honey bees, and other animals seem able to signal. Signals can be utterances, gestures, or complex dances. What

220 Justin L. Barrett
signalling does is prompt a behavioural reaction in the observers. For instance, cries may be uttered to prompt others to stop moving, to run, to climb, to draw near, or to attend to something or someone. Sophisticated animals can learn large numbers of signals and even combine context cues with signals to produce more nuanced signal-behaviour mappings. But signalling (as I am using the term here) falls short of symbolism.
I take symbolism to refer to the cognitive process whereby an object of perception triggers formation of a private mental representation that goes beyond generating an identification of the object and its ontological properties, and beyond the formation of a behavioural scheme for motor reaction to the object of perception. If I see a sphere, I may instantaneously represent it as a bounded physical object (with all the properties that go with such objects), as an artefact, and as having certain affordances for interaction (e.g. I might be able to pick it up and throw it). Such a set of representations would not amount to symbolism. To understand it as a symbol (say, of sovereign power), I would need to be able to represent the intentions of its maker or user. I would need to be able to wonder “what is it intended for?” much in the same way that religious thought often includes what signs and portents are intended for. Such cognitive activities are metarepresentational.
What metarepresentation seems to add is more flexible signalling still as well as symbolically triggering mental states (thoughts, ideas, affective states, etc.) instead of only triggering behavioural routines. When talking to a border collie (a very clever animal), a gesture toward ducks in a pond and the utterance ‘ducks’ will likely trigger the dog to attempt herding the ducks. When talking to a human child the same gesture and utterance is likely to trigger an epistemic state: ‘those things are called ducks,’ or ‘there are ducks in the pond.’ Because of metarepresen-tational abilities, humans can consider what mental state or intention is behind an utterance or gesture, including the possibility that the intention was to change a mental state, not just a behaviour. For these reasons cognitive scientist Michael Tomasello has argued that it is this higher-order ToM that makes human language possible. No metarepresentation, no true language. It may be that metarepresenta-tion (especially in the context of ToM) is the capacity that changes signalling into linguistic communication and symbolism more generally.
If this analysis is right, it may be that when metarepresentation was added to our ancestors’ cognitive tool kits, they quite suddenly became capable of cumula-tive culture, symbolism, and language all at the same time, all because of the addi-tion of one incremental change. If so, then the tool kit that makes Homo religiosus, makes Homo symbolicus and behaviourally modern humans as well. The three are identical and evolved concurrently.

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 221
14. On evidence for the evolution of metarepresentation, Homo religiosus and Homo symbolicus
If metarepresentation endows the possessor with just one of the adaptive capacities sketched above (cumulative culture, symbolism, language, or religion), then that ability could be sufficiently adaptive to drive the evolution of metarepresentation. If indeed metarepresentation has the whole suite of knock-off effects suggested above, then the selective advantage for the Homo that acquired it would be great. As metarepresentational ToM appears to be an extension of lower-level ToM, ToM is the likely evolutionary predecessor. But when did this metarepresentational variant of ToM emerge? Unfortunately, the picture is unclear.
One difficulty in determining when and how metarepresentational ToM and its alleged products such as religiousness might have evolved in humans is that they do not necessarily leave the kind of material traces that archaeologists use as evidence. I can believe in the existence and activity of forest spirits without leaving any material trace. If I pray devoutly five times a day, no distinctive mate-rial trace need be left behind. If I have a rich belief in afterlife and reincarnation that prompts me to burn dead bodies, the bodies and the evidence of the ritual is unlikely to be found. Symbolism presents similar difficulties. If I wear shell beads around my neck, I might be symbolizing my clan membership, my group rank, or my marital status. But I might just find them aesthetically pleasing and am encouraged to wear them by the positive attention they garner. If my symbolism is manifest through utterances or gestures, they may leave no material trace. Artefact evidence, then, may not be sufficient for determining the evolution of metarepre-sentation, symbolism, and religion.
Fortunately, other strategies are available. One way of estimating how old a cognitive mechanism is is to look for evidence in other species. If chimpanzees and gorillas share a particular cognitive mechanism with humans, it may be safe to suppose that our common ancestor with chimpanzees shared the mechanism. In the case of metarepresentational theory of mind, no non-human species have yet unambiguously exhibited it, with even the most generous of attributions being credibly limited to members of Pan (Call & Tomasello 2008; Penn & Povinelli 2007). Ignoring the possibility of convergent evolution, such evidence from comparative cognitive studies seems to set the bounds for the emergence of this higher-order theory of mind at the common ancestor of Pan and Homo, but more probably after the split.
Additionally, on a gross level, because certain cognitive mechanisms may be associated with changes in brain size or shape, cognitive evolution can be informed by skeletal remains. For instance, evolutionary anthropologist Robin Dunbar has

222 Justin L. Barrett
noted a linear relationship between brain cortex size and behavioural evidence of increasingly complex ToM in living species. Applying this linear relationship to the fossil record (treating cranial capacity as a proxy for brain size), Dunbar tells us we can be pretty confident that Homo erectus did not have enough brain-power to exhibit higher-order metarepresentational ToM (Dunbar 2004). Such an ability likely evolved later, likely not until (or after) the appearance of anatomically modern humans.
15. Conclusion
Though we cannot yet be certain when in our evolutionary history metarepresen-tation emerged and gave rise to religion and symbolism, if the analysis presented above is correct, we can be confident that when evidence of either symbolic or religious activity is discovered, capacity for the other is present. Further, evidence of cumulative cultural evolution suggests the presence of metarepresentation, the factor that makes both religion and symbolism possible. Metarepresentation transforms a host of disparate conceptual biases and predilections into a natural propensity toward religious thought and expression. Likewise, metarepresentation transforms gestures, utterances, and other signs into symbolism and language. If metarepresentation is the lynch-pin that holds together these forms of cultural expression, then its evolution, even if only a small modification on previous theory of mind capacities, could lead to a radical break in behavioural possibilities for its possessors.
References
Alcorta, C. & Sosis, R. 2005. “Ritual, emotion, and sacred symbols: The evolution of religion as adaptive complex.” Human Nature 16: 323–59.
Atran. S. 2002. In Gods We Trust: The evolutionary landscape of religion. Oxford: Oxford University Press.
Barrett, J.L. 2000. “Exploring the natural foundations of religion.” Trends in Cognitive Sciences 4: 29–34.
Barrett, J.L. 2004. Why would anyone believe in God? Walnut Creek, CA: AltaMira Press.Barrett, J.L. 2007. “Cognitive Science of Religion: What is it and why is it?” Religion Compass 1
(6): 768–786.Bering, J.M. 2006. “The folk psychology of souls.” Behavioral and Brain Sciences 29: 453–62.Bering, J.M. & Johnson. D.D.P. 2005. “’O Lord … You Perceive my Thoughts from Afar’:
Recursiveness and the Evolution of Supernatural Agency.” Journal of Cognition and Culture 5: 11842.

Chapter 11. Metarepresentation, Homo religiosus, and Homo symbolicus 223
Bloom, P. 2004. Descartes’ Baby: How child development explains what makes us human. London: William Heinemann.
Bloom, P. 2007. “Religion is natural.” Developmental Science 10: 147–51.Boyer, P. 2001. Religion Explained: The Evolutionary Origins of Religious Thought. New York:
Basic Books.Boyer, P. 2003. “Religious thought and behavior as by-products of brain function.” Trends in
Cognitive Sciences 7: 119–124.Call, J. & Tomasello, M. 2008. “Does the chimpanzee have a theory of mind? 30 years later.”
Trends in Cognitive Sciences 12: 187–192.Dawkins, R. 2006. The God Delusion. London: Bantam Press.Dunbar, R.I.M. 2004. The Human Story. London: Faber and Faber.Hamer, D. 2004. The God Gene: How faith is hardwired into our genes. New York: Doubleday.Humphrey, N. 1998. “What shall we tell the children?” Social Research 65: 777–805.Johnson, D.D.P. 2008. “Gods of War: The Adaptive Logic of Religious Conflict.” In The Evolution
of Religion: Studies, Theories, and Critiques, J. Bulbulia, R. Sosis, C. Genet, R. Genet, E. Harris & K. Wyman (eds), 111–117. Santa Margarita, CA: Collins Foundation Press.
Kelemen, D. 2004. “Are children “intuitive theists”? Reasoning about purpose and design in nature.” Psychological Science 15: 295–301.
McCauley, R.N. & Lawson, E.T. 2002. Bringing Ritual to Mind: Psychological Foundations of Cultural Forms. Cambridge: Cambridge University Press.
Mithen, S. 1996. The Prehistory of the Mind: A search for the origins of art, religion and science. London: Thames and Hudson Ltd.
Penn, D.C. & Povinelli, D.J. 2007. “On the lack of evidence that chimpanzees possess anything remotely resembling a ‘theory of mind.’” Philosophical Transactions of the Royal Society, B 362: 731–744.
Pichon, I., Boccato, G. & Saroglou, V. 2007. “Nonconscious influences of religion on prosociality: A priming study.” European Journal of Social Psychology, 37: 1032–1045.
Pinker, S. 1997. How the Mind Works. New York: W. W. Norton & Company.Pyysiäinen, I. 2001. How Religion Works: Towards a New Cognitive Science of Religion, Cognition
and Culture Book Series, 1. Leiden: Brill.Rochat, P., Morgan, R. & Carpenter, M. 1997. “Young infants’ sensitivity to movement infor-
mation specifying social causality.” Cognitive Development 12: 537–561.Ruffle, B. & Sosis, R. 2007. “Does it pay to pray? Costly ritual and cooperation.” The B. E. Journal
of Economic Analysis and Policy 7: 1–35 (Article 18).Saroglou, V., Corneille, O. & Cappellen, P. 2009. “ ‘Speak, Lord, your servant is listening’: Religious
priming activates submissive thoughts and behaviors.” International Journal for the Psychology of Religion 19: 143–154.
Sosis, R. 2003. “Why aren’t we all Hutterites? Costly signaling theory and religious behavior.” Human Nature 14: 91–127.
Sosis, R. & Bressler, E. 2003. “Cooperation and commune longevity: A test of the costly signaling theory of religion.” Cross-Cultural Research 37: 211–239.
Sosis, R. & Rouffle, B. 2003. “Religious Ritual and Cooperation: Testing for a Relationship on Israeli Religious and Secular Kibbutzim.” Current Anthropology 44: 713–722.
Sperber, D. 1994. “The modularity of thought and the epidemiology of representations.” In Mapping the Mind: Domain Specificity in Cognition and Culture, L. Hirschfeld & S. Gelman (eds), 39–67. Cambridge: Cambridge University Press.

224 Justin L. Barrett
Tattersall, I. 1998. Becoming Human: Evolution and Human Uniqueness. New York: Harcourt Brace.
Tomasello, M. 1999. The Cultural Origins of Human Cognition. Cambridge. MA: Harvard University Press.
Tremlin, T. 2006. Minds and Gods: The cognitive foundations of religion. New York: Oxford University Press.
Whitehouse, H. 2000. Arguments and Icons: Divergent modes of religiosity. Oxford: Oxford University Press.
Whitehouse, H. 2004. Modes of Religiosity: a cognitive theory of religious transmission. Walnut Creek, CA: AltaMira Press.
Whiten, A., Goodall, J., McGrew, W.C., Nishida, T., Reynolds, V., Sugiyama, Y., Tutin, C.E. G., Wrangham, R.W. & Boesch, C. 2001. “Charting cultural variation in chimpanzees.” Behaviour 138: 1481–1516.
Wilson, D.S. 2002. Darwin’s Cathedral: Evolution, Religion, and the Nature of Society. Chicago: University of Chicago Press.

Index
Aabnormality 7abstract 6, 51–52, 64, 75, 78,
80, 89, 98–99, 105–106, 164–166, 172–174, 179
abstract ideas 99abstract thinking 172Acacia 101, 103–104acanthocardia 53Acheulean 55Acheulian technocomplex 112acoustically 4adaptation 17, 19, 24, 50, 62, 133,
169, 177, 205–212, 218–219Adaptation Hypothesis 206,
208, 210, 212, 218adaptive 21, 59, 64, 120, 127,
134, 137, 142, 168, 176, 210–211, 221
adaptive allele 120adaptive selection 168ADD 32, 141, 213, 220adhesives 97, 101–102, 104–107adolescent 123advanced cognition 50, 133advanced working
memory 97, 105aeolian 82–83Affiliation/Belonging
system 176Africa 22, 49–58, 62–65, 75–78,
80, 92–93, 97–98, 101, 107, 111–113, 117–118, 123–124, 142–144, 147–149, 153, 155, 158, 179, 188, 190, 199–200
African 2–3, 5, 18, 24–25, 33, 75, 78, 83, 92–93, 107, 111–113, 115–116, 119–120, 122–124, 137, 142–145, 190
African apes 2–3, 18, 24–25African gray parrot 5African Grey Parrot 137African sapiens 112, 116, 119African/Levantine
tradition 123
Afrocentric 142afterlife 205, 211–212, 215, 221Agency Detection Device 205,
213, 216agriculture 136, 141, 144, 178–179alarm calls 4, 7Alex 137–138Algeria 53–54, 147allele 120alloying metals 100Americas 112Amud 51, 152analogical reasoning 100anatomical 13, 15, 18, 22,
123, 135anatomical changes 135anatomical structure 15anatomically modern
humans 99, 101, 141, 154, 222
ancestor-spirit 205ancient human
populations 185animacy 215animal 2, 13, 15, 18, 42, 83,
99, 101, 111, 121, 123, 164, 168–169, 214, 219–220
animal consciousness 15animal markings 83animalistic 113animality 115animism 8antagonistic scenarios 49–50Antarctic 61Antarctica 61anthropocentric 1anthropocentrism 7anthropogenic 51, 76, 78anthropological 186anthropologist 115, 210, 213,
219, 221anthropology 1, 3, 133, 186, 205anthropomorphic 113anthropomorphism 213, 216anthropomorphize 212–213
anti-predator alarm 1, 4ape 2, 8–9, 13–14, 18–19, 23–27,
29, 33, 37–39, 115, 137ape DNA 23ape infants 13, 33, 37–38ape language 26–27, 29ape like ancestors 115ape rain dancers apes 1–3, 5, 8–10, 14–15, 17–20,
23–28, 30, 37–39, 136–137, 141, 152, 217
Apollo 11, 56–57, 143Aquatic Ape Theory 2arboreal 17–18arboreal feeding 17arboreal locomotion 18archaic Homo sapiens 101archaics 119Arcy-sur-Cure 121, 150, 158Ardipithecus 10, 17, 25arrow tip 101arrowheads 101art 14, 18, 22, 97, 111–113, 115,
125–127, 143–144, 146, 149, 155–156, 173, 175
art historians 97arthritic malformations 116artificial language 4artisans 89–90, 97, 102,
104–105Asia 22, 50, 54, 60, 112, 115, 158Asian 142Assimilation models 123associative 137, 152–157associative (symbolic) 152associative interaction 152–153,
156–157associative learning 137Atapuerca 111, 151Aterian site 53Atlantic 60–61, 78Atomic Physics 167Aurignacian 57, 111, 114Australia 54, 102, 112, 151australopithecines 151, 154

226 Index
Australopithecus afarensis 150
Austria 57autism 176autonomic responses 3awls 56, 118, 121
Bbacked 101–102, 147bacterial (prokaryotic) cells 134Baddeley, Alan 192–193Barbs 101basal ganglia 22, 187basket-shaped
morphology 123Bass Strait 58bats 7BBC 78–79, 82–84, 88, 90–92BBC Hiatus 82beads 52–54, 56, 76, 82, 92,
97–98, 112–113, 123, 143, 148, 221
beadworking 148Bechtel, William 194behavioural 2–3, 6, 8, 13, 30,
50, 60, 63, 75, 89, 98–99, 112, 114, 118, 124, 133, 138, 142–144, 149–150, 153, 157, 176, 185, 187–191, 193, 197, 199–200, 212, 214, 220, 222
behavioural modernity 50, 75, 124, 143–144, 149–150
behavioural patterns 63, 176, 185
behavioural traits 144, 189behavioural
transformation 190behavioural variation 98, 138belief 9, 152–153, 205,
207–208, 210–212, 215–216, 221
bell 137Berekhat Ram 145between-group 134–138between-group conflict 138between-group selection
134–137Bickerton, Derek 191bicultural 15, 28, 37biface 112, 151bifacial 52, 56, 82big noses 117bigger brain-greater
intelligence 189, 191
Binford, Lewis 91, 185, 190biochemistry 167biodiversity 64biological 19, 21, 50, 112–114,
119–120, 124, 163–164, 170, 185–187, 190, 199, 207
biological differences 185biological species 112, 120biological/genetical
foundations 124biomass 135biospecies level 112bipedal 17–18, 168bipedalism 17, 24, 26bipeds 2birch bark 119bird bone 56birth 13, 18, 28, 30–31, 33–36,
40–41, 117, 178bison incisor 121bitumen 119Blakeslee 165, 168–172, 174–175Blombos Cave 52–53, 55–56,
78–79, 82, 85–86, 113, 143, 145, 147, 149, 159
blood 2, 102, 117blood groups 2Bloom, Paul 215-216body 15, 27, 30–33, 35–36,
39–41, 51, 77, 90–91, 111, 117–119, 121, 123, 127, 135, 146, 148, 150, 156, 177, 197, 215
body ornaments 121, 127body weight 31, 177Bokkeveld 83bond 61, 176bonding 37, 173, 179bone 51–52, 56–58, 64, 75–78,
82, 90, 118, 121, 125, 146bone awls 118, 121bone retouchers 51bone splinter 146bone tool 52, 56–58, 77, 82, 125bonobo 2–3, 8, 14–15, 18, 24,
27–29, 31, 36–37, 137Border Cave 51, 53, 143Boule, Marcelin 115-116Boyer, Pascal 209, 215Braille 5brain 15–16, 19, 26, 39, 41,
99, 111, 120, 125, 127, 137, 153–154, 163–172, 175–178, 180, 189, 191–199, 221–222
brain connections 172brain growth 191, 197brain plasticity 176–177brain size 19, 99, 111, 120, 154,
177–178, 192, 196–197, 221–222
brittle glue 101Broca’s area 193–195Brooks 50–51, 55–56, 62, 76,
118, 142–143, 149burial 49, 51, 53, 58, 112, 122,
150, 154–158, 212burial practices 49, 58, 212buried their dead 51, 117,
151, 212burning 104, 155Bushman Rock Shelter 77Byzantine 120
CCambridge On-line 115Canada do Inferno 126Cape Hangklip 77–78Cape Range 54Cape Town 78, 82, 158, 163, 200captive 14, 17, 29capuchin monkeys 2carbonised 82carnivore 150–151carved 56causal 16, 31, 38, 40, 168, 199causal reality 16causal relations 168cave paintings 113, 125Cebus libidinosus 2cell 19, 133–134, 167cell biologist 133cell biology 167cell organelles 134ceramics 100chaine opératoire 90charcoal 101–102Chase 56, 63, 144, 153Châtelperronian 121, 123, 149Chauvet 56–57, 113chemical analysis 118chemistry 105, 119, 167child skeleton 122childcare 136child-caring 178childlike 15chimpanzee 2–6, 8, 10, 18–20,
23–24, 26–27, 32, 34–36, 38, 150, 212

Index 227
chloroplasts 134Chomsky 170chromosomes 23–25, 134chunking 167climate 2, 33, 50, 60–61, 63–65,
76, 190climate change 60, 65, 76cling 13, 17–18, 30–35, 37–38clinging 13, 30–34, 36Clottes 56, 113Côa Valley 126coded information 49cognition 22, 49–50, 58–59,
63, 75, 97–100, 104–107, 111, 113–115, 118, 125, 133, 136–137, 166, 169, 177, 180, 186–189, 197, 199, 206–207
cognitive 1, 10, 13, 15, 23–25, 29, 36–37, 49–50, 58, 75–77, 97–100, 104, 107, 111–113, 117–118, 125, 133, 136, 141–142, 145, 151, 153–154, 157, 165, 167, 177–178, 185–200, 205–207, 213–221
cognitive abilities 75, 77, 100, 177, 196
cognitive adaptation 205 cognitive architecture 205, 216cognitive control 99, 198cognitive development 142,
145, 177, 205cognitive evolution 153, 154,
185–186, 188, 192, 195, 198–199, 221
cognitive fluidity 97, 99–100, 104, 107, 219
cognitive function 10, 167, 188, 193, 195–197
cognitive neuroscience 186, 188, 190, 194–195
cognitive science 188, 205cognitive science of
religion 205cognitive sciences 205cognitive systems 205–206, 213cold-adapted body
proportions 123colourant 102coloured stains 102common attributes 98communication 1–8, 10, 24, 30,
82, 90, 92, 136, 163, 165–166, 170, 176, 178–179, 219–220
communicative abilities 163comparative
neuroscience 196–198complex 7, 9, 14–15, 21–22, 29,
41, 52, 57–58, 63, 76, 92, 97–100, 102–103, 105–107, 112, 115, 118, 126, 136–137, 147–148, 163–164, 167, 173, 189, 191, 193, 195, 197–199, 210, 214, 217, 219, 222
complex capabilities 99complex cognition 97–100,
103, 107 complex cognitive
processes 98complex cognitive tasks 193complex hierarchical
sentences 193complex planning 14compound 97, 100, 102–107,
174compound adhesive
manufacture 97, 100, 103–104, 106
compound adhesives 97, 102, 104–107
compound tools 97computer languages 174computer machine code 6Conard 9, 54, 56, 113–114, 118conceptual 6, 146, 166, 171,
214, 222conceptual schemas 116conceptual structure 171Concurrence Hypothesis 206,
212–213Congo 8, 27conscious mind 169consciousness 13, 15–16, 18,
31, 34, 36–37, 39–41, 169, 172, 215
Conus 53–54convex edge 101Coolidge and Wynn 192–193Coolidge, Frederick 191–192cooperative eye hypothesis 136copulation calls 7core 52, 62, 141, 172cortex 52, 54, 99, 168, 170, 172,
174, 193–194, 222cortical development 177cortices 192, 196, 198counter intuitive intentional
agency 206
courtship 1, 5cracking open 2cranial capacity 117, 222cranium 123–124, 197–198, 200crayons 143, 147, 149creativity 166, 172, 179, 189, 218crescents 101cries 5, 33, 220Croatian cave site 117cross-cultural 206cross-hatched 77, 80,
87–88, 90CSR 205–206Cueva de los Aviones 53–54cultural 1, 14, 17, 20, 27, 29–30,
49–51, 56, 58–60, 62–65, 76, 92, 122–123, 125, 127, 136, 142, 148, 152, 166, 169, 186, 205–209, 211, 218–219, 222
cultural drift 50cultural evolution 50, 59,
62, 136, 209, 218–219, 222, 136
cultural expression 206–208, 219
cultural frames 166cultural learning 169cultural modernity 59, 76cultural multilevel
selection 136cultural systems 58cultural variation 152culture 6, 15, 26, 28–29,
36, 42, 49–52, 58–60, 63, 75–77, 88, 97–99, 107, 112, 118, 121–123, 125, 133, 136, 145, 156, 169, 177, 179, 185–187, 189–191, 198–200, 210, 218, 220–221
cumulative behavioural change 133
cup marks 152cupules 51cutting 77, 84, 101–102cutting edge 84, 101, 102
Dd’Errico, Francesco 159dance 7–8, 173dance language 7dancing 8, 175, 180Dansgaard-Oeschger 61

228 Index
Darwin 20, 115, 216Darwinian 20, 23, 174,
177, 206dating 62–63, 78, 82, 143,
147, 157dawn of language 163, 171Deacon’s thesis 137–138Dean, Hamer 210decorated 56–57, 78, 91, 149decoration 22, 56, 80, 148–149,
154–155decorative 121, 125, 157, 180decorative interaction 157decorative patterns 121Dederiyeh 51defense 134, 136deliberate heating 53, 56Democratic Republic of
Congo 8demographic changes 51demographical thresholds 114dendritic 77, 87–88Dentalidae 54dentition 123designs 51–52, 75, 81, 90, 100,
105, 149developmental 13, 21–22,
37–38, 153, 168, 174–176, 178, 196–198, 205, 214
developmental neuroscience 196–198
developmental verbal dyspraxia 22
dialects 1, 167Diepkloof Rock Shelter 52,
78–79, 81diet 17differences 5, 13–14, 18–19, 23,
25, 30, 34, 36, 38–39, 59, 77, 80, 112, 121, 135, 185, 191, 198
direction of gaze 136dispersal 50, 62, 76, 143, 153diversity 81, 90, 117, 138, 191DNA 13, 18–20, 23, 25, 117,
120, 170D-O 61dogs 20, 136, 214, 217dolphins 7domestication 137dominance 4, 138, 212domineering behaviour 135doodling 90dopamine 176
Dordogne 55dorsal prefrontal cortex 194Down’s syndrome 24drawing 19, 29, 77, 90, 100,
102, 143–144, 153, 188, 205, 217
DRS 78–80, 88, 90–92drumming 4, 8drumming signature 4duality of patterning 166Duiwenhoks 83Dunbar, Robin 141, 145,
152–154, 191, 221–222
EEarly Stone Age 59Eastern Cape 77echolalic speech 5echo-location 7eco-cultural 64ecological 1–3, 58, 64, 136,
138, 163–164, 167, 175, 178, 186
ecological dominance 138ecosystems 63egalitarian 135egalitarianism 135eggshell 54, 75, 78, 80–81,
90–92eggshell containers 91eggshells 52–53, 80, 83, 88,
91–92ego 16–17electron spin resonance 82electrostatic 102elephants 7, 30, 217Elykia 28embodied brain 163Embodied Construction
Grammar 169emergence of language 39, 111,
115, 125, 163, 175, 179emotional 32, 37–38, 136, 152,
170–171, 173, 175–179, 199, 209
emotional drivers 171, 175emotional power 173emotional state 136emotions 164, 175–176emotive 152, 154–157emotive (non-symbolic) 152emotive interaction 152,
154–157emotive non-symbolic 152
Enard 22–23encephalisation 189endocasts 197end-scrapers 146England 24engraved 51–52, 54, 56, 77–78,
80–83, 85–88, 90–92, 149, 156
engraved bone 51, 52engraved eggs 82, 91–92engraved lines 56, 80, 88engraved ochre 52, 81–83,
85–86, 88, 91, 92, 149engraved patterns 77, 82, 88,
90engraved pebble 77, 78 engraver 77, 88engraver laterality 77 engravings 52, 58, 64, 75–77,
84, 87–92, 143, 149enhanced speech 187Enkapune Ya Muto 101environmental variation 21,
51epigenetic 18–21, 25, 28, 30episodic memory 193–194epistemological 3, 89, 194ESR 143, 147Ethiopia 150ethnographic 58, 77, 91–92,
102, 113–114, 118, 126ethnographic present 91, 92,
113, 118ethnology 186ethologist 3eukaryotic 133eukaryotic cells 133Eurasia 49–51, 53, 75–76,
111–112, 188Eurasian 112, 117–118, 123, 144,
149Eurasian humans 117Eurasian Middle
Palaeolithic 144, 149Eurasian Neandertals 118, 112Eurocentric 142Europe 22, 49–53, 55–59,
62–64, 76, 111–112, 115, 118–123, 125, 142, 144, 149, 157–158
European 57, 63, 65, 75, 93, 115–116, 119, 123–125, 142–143, 153
eusocial insect species 135

Index 229
eusociality 135Eve 117–118, 123everted lower lip 6evolution 7, 10, 13, 19–20, 24–
25, 49–51, 58–60, 62, 64, 75, 97, 99, 111–112, 115–116, 123–125, 133–138, 141–142, 145, 150–151, 153–154, 163, 171, 173, 176–177, 179, 185–200, 206–207, 209, 215, 218–219, 221–222
evolution of reason 99evolutionary 1–2, 8, 14, 120,
133–135, 141–142, 150, 152–153, 163–164, 168–170, 173, 176–179, 186, 188–189, 198–199, 205–206, 213, 221–222
evolutionary biologists 189evolutionary biology 186, 188evolutionary development 169evolutionary psychological
development 173evolutionary psychologists 189evolutionary psychology 205evolutionary theory 135evolutionary transition 133–135,
168evolutionary transitions 135excavations 63, 118–119executive functions 100, 192experiential processes 169experimental studies 3explanandum 189explanandum (modern
culture) 189explanans 189explanans (modern
intelligence or language) 189
explicit memory 194extended learning 99extinct 1, 117, 119, 158, 185–186,
190, 196, 214extinction 50, 111, 119,
122–123eyes 34, 38, 136, 138, 172
Ffacial expressions 3–4, 34,
36, 39facial-vocal dialogues 34farming 209fat 31–33, 35, 101, 104, 118
fauna 82, 150faunal 64, 78, 80, 82, 91Feldhoffer 115, 152fetal malformation 30figurative art 111, 113, 125,
143–144, 146, 149, 155–156fingerprint 119fingers 17fire 62, 64, 97, 103–106, 211fire control 97fists 17fitness differences 135flake 87, 103, 118flakes 146flasks 81flint 118, 146flint flake 118flutes 56Foley 113foliate 52, 82foliate point 52food 14, 20, 22, 26, 38, 137foodstuffs 1, 4foraging patterns 17foraminifera 62forest 8, 10, 18, 22, 25, 28–29, 221formal 58, 75, 82, 169, 191formal bone tools 58, 82formal lithic tools 75fossil record 1, 3, 19, 23, 25,
197, 222fourth level intentionality 152fox canines 121FOXP2 22–23, 26, 187–188fractal patterns 173France 51, 55, 57, 113, 121, 125,
144, 146, 151–152Franco-South African 78free-ranging apes 14freewill 16frequency 7–8, 134, 144friction 101frontal 100, 193, 196frontal lobe 100, 196frontal lobe damage 100Fumane 56–57functional modular
structures 171funerary practices 77, 152
GGARP 64gaze 39, 136gazelles 120
GCM 64gender 113gene 18, 20–23, 26, 120, 179,
187, 210gene expression 18, 20, 21gene pool 120genealogical 133genealogical relatives 133General Circulation Model 64generic 141, 164–165, 171,
174–175genes 2, 19, 21–23, 25–26, 65,
122, 134, 138, 170, 175, 188–189, 199, 210
genetic 13–14, 18, 20–21, 23, 50, 58, 62, 64, 115, 117, 119–120, 125, 135–137, 170, 174–175, 179, 185–190, 198–199, 210–211, 213
Genetic Algorithm for Rule Set Prediction Gathering 64
genetic change 190, 21genetic evolution 58, 125, 136,
137, 186–188, 190genetic inheritance 170genetic selection 20, 211genetically inherited 100, 123geneticists 111genome 19–23, 25, 117,
120, 170genomes 2, 19–21geographic isolation 112geographical 112, 143, 145, 164geographical segregation 112geological 21, 60, 62, 83, 103geometric 52, 76, 82, 213geometrical pattern 172Germany 56–57, 111, 113–114,
125, 151gestures 1, 6, 26, 29, 77, 219,
221–222gibbons 1, 4, 17, 24glacial 60–61, 64global warming 62glue 97, 101–106glue makers 97, 105glue residues 101Glycimeris 54gnawing 151goal 90, 100, 188, 193, 213,
218–219goal directed actions 100goal maintenance 193god-gene 210

230 Index
gods 206–207, 209–211, 213, 215–218
Gombe 8Goodall 4, 8, 150gorillas 221Goukou 83grammatical 13, 16, 168,
173, 179grammatical abilities 13grammatical structure 16grave 51, 150–152, 156, 212grave goods 51, 151, 156, 212grave markers 152grave pits 152Gravettian 153Great Ape Trust 13–14Greenfield 26–27Greenland ice cores 60grinding 76, 83–84, 106, 143grinding marks 143grinding slabs 143grinding stones 143grooming 4, 6, 154grooming hand-clasp 4, 6grooved 83, 121grooved pendants 121Grotte des Pigeons 53–54, 147Grotte du Renne 121, 150, 158group 1, 4–6, 14, 22, 27–30,
33, 36–37, 39–41, 58–59, 83, 87, 89, 92, 97, 113, 115, 117, 123, 127, 133–138, 143, 145–146, 150, 154, 176, 180, 209–211, 218, 221
group identity 5, 6, 97group membership 113, 180group size 59group specific
communication 5grunt 4Gua 26guarded egalitarianism 135gum/resin 101
Hhabitat homogenization 21Hadar 150HADD 213–215haematite 53, 103, 143haematite pencils 143hafted 101, 103hafting 56, 101, 106–107,
119, 218hafts 102, 105
half-man/half-beast 116Hamer Dean 210hand 4, 6–7, 9, 13, 18–19,
31, 41, 64, 92, 189, 196, 211, 218
hand axe 112hand waving 13, 31haplotypes 119harpoons 56Hawkins 165, 167–172,
174–175Hawkins and Blakeslee 168,
171–172, 174–175hearths 22, 82heating 53, 56Heinrich events 61, 153hide 14, 84, 118, 152, 209hide working 118hides 56, 118hierarchical structuring 166,
174high 7–8, 20, 23, 29, 60–62, 64,
82, 91, 123, 134, 195–196, 198–199, 217
high cranial vault 123higher 100, 138, 166, 168–169,
171–174, 177, 179–180, 188–189, 210, 216, 220–222
higher level states 168higher order 100, 216, 220,
221, 222highly coded system 15high-resolution neuroimaging
techniques 199Hohle Fels 57, 114Hollywood film 116hominin 1, 25, 62, 120,
135, 138, 141–142, 145, 148, 150–155, 185, 218
hominin lineage 135homininae 142Homo 2, 9, 13–15, 18–19,
22–30, 35, 50–53, 55, 57–59, 63, 75–76, 92, 97, 101, 111–114, 117, 125, 142–143, 145, 149, 151, 153–155, 157–158, 187, 189, 192, 195, 197, 199–200, 205–206, 212, 214, 216, 218–222
Homo antecessor 111Homo bonobos 27
Homo erectus 22, 111, 112, 114, 117, 125, 154, 195, 197, 216
Homo ergaster 197Homo habilis 22Homo heidelbergensis 149,
151, 154, 157, 192, 197, 199Homo neandertalensis 149,
151, 153, 155, 157Homo religiosus 206, 212, 214,
216, 218, 220Homo sapiens 2, 9, 22,
50–53, 55, 57–59, 63, 75, 76, 92, 101–113, 128, 142, 145, 151, 153, 155, 157–158, 187, 189, 192, 195, 197, 199, 200
Homo sapiens sapiens 22Homo symbolicus 22, 23, 26,
97, 143, 153, 158, 200, 220hormones 99horse 114Howieson Poort 55HP 77–78, 80–81, 91–92hues 56, 156human 1–8, 10, 13–42,
49–51, 53, 57–60, 62–64, 75–77, 89, 91, 97–99, 105–106, 111–115, 117–121, 123–125, 133–138, 141, 144–147, 152–153, 158, 164, 167–170, 172–173, 176, 178, 185–192, 194–199, 206–210, 212–217, 219–221
human agency 213, 214human genome project 170human genus 111, 124human nature 206–208Human Revolution 50,
112–113, 115, 117–118, 120human species 117, 124, 133, 137,
186, 212, 213, 216, 217, 221human working memory
system 192human/ape differences 13,
18, 23hunt 102hunter 81, 135, 144, 148, 209hunter gatherer societies 135hunter gatherers 81, 144hunting 19, 102, 105, 136, 209Hypersensitive Agency
Detection Device 213, 216hypnosis 16, 41

Index 231
IIberia 111Iberian 62ice 60–62, 118Ice Age 118ice cores 60, 62ice sheets 60iceberg 61iconographic 76icons 173Ifri n’Ammar 53IGCP 437 62Ilsenhöhle rock shelter 118imaginative play 163, 174, 176,
179–180imitation 8, 26, 120,
169, 176implements 75, 105incising 77, 88, 90incisions 77, 84, 87–88incisor 121Indonesian island of Java 111infant 13, 15, 17–18, 23, 30–40,
176, 213infant carriage 13infant clinging 13, 36inferior frontal gyrus 193in-group conformity 210inheritance 138, 164, 167, 170inhibitory control 193inhumation 51, 156innate patterns 13innovation 49–51, 59, 75, 92,
99, 127, 218–219innovations 51, 58–59, 63–65,
92, 98, 190innovative technologies 99insects 35, 56, 135intellectual capacities 171intentionality 152, 154, 191–192,
219intention-reading 169interaction 4–5, 27, 30, 39,
41, 50, 63, 111, 114, 122, 150, 152–154, 156–157, 168, 176–177, 220
interbred 123, 190interbreeding 25, 120interglacial 60, 62, 83interglacial transgression 83inter-group competition 127inter-level connections 172International Geological
Correlation Program 62
intuitions 206intuitive dualism 214–216involuntary expressions 8Iraq 152iron oxide 103Israel 53–55, 145, 147,
151–152, 210Italy 55, 57ivory 8, 56, 113–114, 125Ivory Coast 8Iziko-South African
Museum 78
JJava 111javelins 118juveniles 28, 150, 152
KKakombe Valley 8Kanzi 15, 27–29, 39, 137–138Kanzi’s mother 27Kapthurin 143–144, 147Kenya 101, 147Khoesan 81, 91–92kiln-fired ceramics 100Kimberly 54kinship 18Klasies 52, 55, 77–78, 84, 143Klein Kliphuis 52Kleine Feldhoffer 115knapped 84, 87knapping 87knuckles 18, 24Königsaue 119Krapina 152Ksar Akil 147–148KwaZulu-Natal 98
LL’Hortus 152La Chapelle-aux-Saint 51La Chapelle-aux-Saints 115La Ferrassie 51, 57, 152La Quina 152La Roche Cotard 146ladder-like patterns 80Lagar Velho 122–123Lahr 113Lamarckian 177Lana 15language 1–7, 13–17, 21–23,
25–30, 37–42, 50, 97–100, 105, 107, 111–115, 117, 125,
127, 133, 135–138, 163–180, 187, 189, 191, 193–194, 197, 208, 219–222
language like signals 7language modules 169, 170, 171language technology 179 Langur monkey 4Last Common Ancestor 2–3,
117Last Glacial Maximum 64Late Aurignacian 114Late Middle Palaeolithic 146,
152Latent Semantic Analysis 169Later Stone Age 57, 75, 143laughter 137, 176Le Moustier 51leather 77Lebanon 147leg bones 123leg kicking 13Lemurs 10Leroi-Gourhan 121lesion 196Levallois-rich 145Levant 144, 148, 151limbic system 28, 176linguistic 4–7, 10, 14–18,
23, 26, 29–30, 35, 37, 40, 168–170, 175, 191, 205, 220
linguistic analysis 171linguistic constructions 16linguistic patterns 168, 169linguistic recursion 191lion 57, 114lithic 52, 75, 78, 87–88, 146lithic doodles 146Littorina Africana 53locomotion 4, 17–18, 24, 31long-tailed macaques 2Lovejoy 1, 10, 25, 30lower level states 168Lower Palaeolithic 145–146,
149LSA 75, 143–144lunates 101macaques 2
MMadden 9magico-religious 58Mahale 6Maisha 28

232 Index
malformation 30mammoth 56man/animal dichotomy 13mandible 123–124Mandu Mandu 54Manufacturing 97, 99, 180Manufacturing process 97Manufacturing tools 99mapping 17, 185Margulis, Lynn 134marine 53, 60–64, 83, 92,
112, 123marine cores 60marine deposits 83Marine Isotope Stage 60, 62marine shell 53marine shell beads 92, 112, 123markings 14, 83, 88–90,
92, 125Matata 15, 27-28mate 6material symbols 114maternal line 117mathematics 6, 26, 163mating 114Maxent 64Maynard Smith 134–135McBrearty 24, 50–51, 55–56,
62, 76, 118, 142–143, 149meaning-making 166, 180meat 209medial prefrontal cortex 194meerkats 7memes 122memory 50, 97, 99–100, 105,
149, 165, 169–172, 177–178, 191–196, 198
memory based prediction 171memory prediction system 172menstrual 4mental 6, 9, 14, 31, 77, 97–98,
100, 105–107, 136–137, 165, 174, 185, 191, 193–195, 198, 205–207, 214–220
mental architecture 98mental mechanisms 194mental phenomena 185, 191,
194, 195, 198mental powers 165 mental representations 9, 137,
205, 206, 217mental rotation 97, 100,
105, 106mental template 77
metalinguistic 197metarepresentation 205–206,
216, 218–222mice 22microcephalin gene 120microcharcoal 62Mid Upper Palaeolithic 153Middle 41, 52, 64, 75, 80, 91,
97–98, 101, 111, 113, 124, 141–144, 146, 149, 151–153, 155, 185–186, 188–192, 197–200
Middle Palaeolithic 52Middle Range Theory 91, 185,
186, 191, 198Middle Stone Age 75, 97–98,
101, 111, 113, 143, 200Middle to-Upper
Palaeolithic 64middle-range theories 185,
188–192, 197–200migration 59mimicry 5, 8mind tasks 193minded agency 206Miocene apes 152MIS 60, 62–63, 83Mithen 22, 99, 192, 219mitochondria 134mitochondrial DNA 117Mitochondrial Eve 123modern 2, 5, 15, 18, 22–25, 31,
50–51, 53, 59, 75–76, 97, 99–101, 104, 106, 111–115, 117–123, 125, 127, 136, 141–142, 144–145, 150, 153–154, 157–158, 186–187, 189–192, 194–195, 197–200, 205–210, 212–214, 218–220, 222
modern behaviour 65, 75, 97, 106, 144, 187, 197, 200
modern cognitive capabilities 111
modern cultural traits 59modern human 22, 50, 51,
53, 59modern human
immigration 121, 125modernity 50, 59, 75–76, 124,
142–144, 149–150modification 25, 83–84, 148,
211, 222modified lithics 2
modular hierarchical structure 163–164, 166–167
molecular 13, 22, 120, 134, 170molecular biology 170molecular data 120molecular genetics 13molecular phylogenetics 22molecules 167monoamine systems 175moral agency 14, 28–29, 40Morocco 53–54, 98, 112,
146–147moro-reflex 13mortuary 150–152, 154–155mortuary activity 152, 154, 155mortuary caching 150, 154MOS 2 62mother-child 163, 175mother-child nurturing 163motif 80, 88–92motor 5, 13, 22–23, 60, 88, 173,
187, 194, 220motor control 88, 89, 187, 194motor inhibition 23motor patterns 13, 39motor skills 173Mousterian 52–53, 55–56,
75–76Mpumulanga 77MSA 52, 59, 75–78, 80–84,
88, 92, 97–98, 101–107, 143–144, 153
MSA adhesives 102Mt. Carmel 116mtDNA 117, 119–120multicellular organisms 134–135multilevel 98–100, 106, 136multilevel actions 99multilevel operations 98, 100,
106multiple 17, 25, 62, 64, 77,
83–84, 149, 152–153, 156multiple burials 152multiple dispersals 153Multiregional 117, 123, 144Multiregional Evolution 123,
125Multiregional Hypothesis 117multispecies 144Mumba cave 54Mungo 151music 163, 171, 173–175, 179–180musical instruments 56

Index 233
mutation 22–23, 99, 127, 174, 187, 211
mutational change 133myths 127, 148
NN. pachyderma 61Namibia 56–57, 77–78Nassarius 52–54, 82, 113Nassarius gibbosulus 53, 54Nassarius kraussianus 52, 53natural 1–4, 8–9, 20–21, 39,
52–53, 83–84, 87, 89, 103–104, 106, 119, 133–134, 136, 138, 143, 145–147, 150, 171, 173, 179, 187–189, 191, 206–208, 210–213, 215–216, 218, 222
natural abrasion 84natural phenomena 52, 211natural selection 2, 20, 133,
134, 138, 187, 188, 189, 210, 212
Nazlet Kather 51Neander Valley 115, 152Neandertal 50–51, 53, 75–76,
111–112, 115–125, 149, 158, 192
Neandertal burials 51, 117Neandertal ness 115Neandertal skeleton 121Neandertals 49–53, 55–59,
63–64, 111–112, 115–125, 141, 150–152, 154, 158, 188, 192, 199–200, 212
Near East 51–53, 55–56, 58, 76, 92, 116, 119, 123, 157–158
necklace 9Neema 28neocortex 176, 196nervous system 30, 33, 170nests 17, 105Neumark-Nord 118neural 17, 40, 99, 163, 165,
172–179, 185–196, 198–200
neural connectivity 99, 163, 174–175
Neural Darwinism 176–177neural evolution 186, 187neural mechanisms 191, 193,
194, 196neural mutation 99neuroanatomic 196
neuroimaging 195, 199neurological structure 163neuromodulators 176neuromotor 89neuronal 3, 13, 167, 175neuronal activity 167neuronal connections 175neuronal connectivity 167neuronal developmental 13neurons 99, 188, 199neuroscience 186, 188, 190,
194–198New Guinea 54New Ireland 54nomad 178non-burnt 78non-human 1, 3, 5–8, 10, 117,
137, 212–214, 217, 219, 221non-human language 3, 6, 7non-human primates 217, 219non-human species 137, 212,
213, 217, 221non-linguistic 5, 10non-primate 50non-referential models 1non-religious 210non-utilitarian 75non-vocal communication 5North Africa 53North Atlantic 60–61notches 56nuanced cognitive
development 142nuclear genome 117, 120nucleated (eukaryotic)
cells 133numerical abilities 6Nunamiut 91
Ooak bark 118Oase 123–124Oceania 58ochre 14, 22, 51–53, 75–76,
78, 81–84, 86–88, 90–92, 101–106, 142–143, 149, 155–156, 200
ochre stained segments 101odour 4Old World 111, 155optically stimulated
luminescence 78orange ochre 101orang-utan 8
organic 2, 78, 118organic raw materials 2organic residue 78, 118organism 1, 3–4, 21, 40–41, 135organs of communication 136ornaments 53, 58, 64, 75–77,
97, 111–115, 120–121, 123, 125, 127, 143, 147
OSL 80, 82–83, 147ostrich 52–53, 75, 78, 80–81,
83, 88, 91ostrich eggshell 52, 53, 75, 78,
80, 81, 83, 88ostrich eggshells 52, 53, 80,
83, 88Oued Djebbana 53–54, 147out of Africa 22, 50, 76, 144,
179, 190
PP. paniscus 2P. troglodytes 2palaeoanthropology 141, 157palaeoclimatic 60, 63–64palaeocognitive 195palaeocognitive
neuroscience 195palaeoenvironmental 49, 63–64Palaeolithic 9, 52, 64, 75, 114,
126, 142–146, 148–149, 151–153, 157–158
palaeoneurology 197–198palaeontology 1, 150Palestine 112, 116Palmenhorst 77–78Pan symbolicus 1, 3, 7Pan/Homo 14–15, 27–30, 35Pan/Homo bonobos 27Panksepp 170, 176–177para-verbally 4Parkington 80, 179parrot 5, 137part- 115, 123part-ape/part-man 115part-Neandertal ancestry 123past behaviour 190pattern recognition 163, 165,
170–172patterning 53, 91, 166, 180patterns 13, 15, 17, 31, 33–34, 37,
39, 52, 58, 63, 77–78, 80, 82, 88, 90, 92, 121, 164–165, 168–169, 171–176, 179–180, 185, 211

234 Index
Pavlovian 137pecking 77Penascosa 126pencil 78pendant 114, 145perforated 53–54, 147–148perineal odour 4personal 15–17, 24, 29, 37, 41,
53, 58, 64, 75–76, 90, 97, 111–115, 121, 123, 125, 127, 141, 144–146, 148–150, 154–158
personal ego 16, 17personal ornamentation 123,
141, 144, 145, 146, 148, 149, 150, 154, 155, 156, 157, 158
personal ornaments 53, 58, 64personal presentation 113petroglyphs 126phalange 121pheromones 4phonemes 15, 168phylogenetic proximity 2phylogenetics 22phylogeny 196pictorial representation 173pierced 121, 123, 147pierced animal teeth 123pierres figures 145–146Pigeons 53–54, 147pigment 53, 55–56, 58, 76–77,
121, 143–144, 146–149, 155–156
pigment fragments 149, 155pilot study 8Pinnacle Point 78Pit of the Bones 151planetary diversity 138plant 2, 101–102, 104–106, 219plant gum 101, 102, 104, 105, 106plant prey 2plastic 25, 101, 212–213plasticity 13, 22, 105, 176–177,
196, 199play 2, 4, 17, 38, 49, 63, 113, 163,
173–176, 179–180, 206, 208play face 4Pleistocene 60, 124, 142–143,
153, 155pointing 14, 136–137, 174points 30, 35, 56, 82–83, 88, 98,
148, 152, 191polar ice cores 60, 62polishing 56
pollen 62polygenetic 189polygraph 9pongo-linguistics 3Porc Epic 143Portugal 122, 126post-contact populations 122Post-Howiesons Poort 78post-Miocene 3posture 4, 6, 19, 31, 168Poverty of Stimulus 171powder 83–84, 103,
105–106Pre australopithecines 154pre-contact 123predation 23, 150predators 2, 8, 17, 33, 35,
135, 213prefrontal cortex 99, 193–194Pre-Human Religion
Hypothesis 206, 212pre-language 172, 179pre-modern 50, 142, 144Pre-Still Bay 78primate 1, 4–5, 24, 50, 119,
135–136, 196primates 22, 24, 30, 35,
50, 76, 135–136, 150, 176–178, 191–192, 195–196, 198, 217, 219
primatological 1, 10, 150problem-solving 100prokaryotic 134prokaryotic cells 134prominent chin 123prosocial behaviours 133prosociality 134Protoaurignacian 123proto-cell 134protohuman language 14proto-religious 205P-suke 28psyche level 13psychological traits 176psychology 3, 167–168, 205, 214psychometric tests 100psycho-physiology 9punctured openings 80
QQafzeh 52–54, 147, 151Qena, Egypt 51quadruped gait 17
quadrupedal 4quadrupedal bouncing 4quadrupeds 2qualitative leap 125quantum principle 166quartzite 146quartzitic 78Quneitra 52, 54
Rracial diversity 117rain 8, 17, 35, 212rain dance 8Rank/Status system 176rapid expansion 111Reading 146, 148, 151, 158, 166,
169, 172–173, 176, 178, 209, 218
recruitment screams 1, 4recursive 17, 107, 144, 153,
173–175, 178, 180, 187, 192–193, 195, 197
recursive change 144recursive syntax 187, 192, 193,
195red 6, 53–56, 61, 101–103, 106,
123, 143, 146, 148red deer 123red ochre 53, 101, 102, 103,
106, 143red pigments 53, 54, 55red stained 143referential 1–5, 7, 29, 165referential models 1referential signalling 4reflections 3, 60reflective mode 16regional variant 117reindeer 121religion 10, 49, 75, 141, 144, 153,
205–216, 218–219, 221–222religiosity 210religious 40, 50, 58, 117, 127,
141, 205–216, 218, 220, 222religious beliefs 50, 117, 127,
206, 212, 218religious communities 210, 211religious expression 205, 208religious ritual 209, 211religious thought 205, 206,
208, 220, 222remains 22, 26, 34, 62, 77, 80,
82, 90–91, 98, 112, 121, 152, 189, 193, 211–212, 221

Index 235
reproductive separation 120residue 53, 101, 106, 118residue analysis 101, 106resin 101, 105retouch 101Rhafas 53rhythmic facial/vocal
interplay 34ring-tailed lemurs 10rituals 8, 209, 211Riwi 54rock art 126Romanian cave 123Rose Cottage 101rotational hand-waving 13, 31rough 4, 176, 179rough and-tumble 179rough grunt vocalisations 4rule-governed 1, 4–5
Ssandstone 78, 103, 106sapiens 2, 9, 22, 50–53, 55,
57–59, 63, 75–76, 92, 101, 111–113, 116, 119, 124, 142, 145, 151, 153, 155, 157–158, 187, 189–192, 195, 197, 199–200
Sapir-Whorf hypothesis 6scavenge 151scavengers 135scavenging 151scent-marks 5schematization 169Schöningen 118scrapers 82, 146screams 1, 4–5sculpted ivory figurines 114sea level 83segment 101–102selective 20, 58, 174, 189, 193,
211, 221selective attention 193selective pressures 20, 189self 3, 9, 14–16, 25–26, 29,
37–41, 60, 62, 77self agency 15self reflection 14, 16, 39, 41semantic 4, 147, 166, 169, 194semiotic 163–164, 167,
171, 180semiotic function 164, 171semiotic representation 5 sender-message-receiver 3
sensory 4–5, 7, 165, 172, 174, 177, 196
sensory motor modalities 5sex hormones 99sexual 6, 24–25, 99, 138sexual dimorphism 24, 138sexual maturity 99sexual signals 6sexual swellings 24shafts 97, 104shale 83, 103Shanidar cave 152shard 87shared beliefs 76sharing 49, 165, 186, 209–211,
219shell 52–54, 56, 76, 82, 92, 97,
112–113, 123, 145, 155, 221shell beads 52–54, 56, 92, 113,
123, 221shell pendant 145shellfish 82shelter 52, 56–57, 77–79, 81,
118, 122Sherman and Austin 26–27shoe wear 118short-term memory 194sibling species 13, 18Sibudu 53, 98, 101–103side branch 116side-scrapers 82sighs 5sign 5, 89, 142, 148, 165, 173sign language 5, 165, 173sign patterns 165signal 1, 4–5, 91, 210, 214,
219–220signalling 4, 210, 220signed language 5signifier 142, 165silcrete 52siltstone 83, 88Sima de los Huesos 151simple burials 155singing 7, 173, 175single 5, 9, 19–20, 22–23,
25, 39, 53, 58, 60, 62, 77, 83–84, 87–90, 99, 104, 117, 124, 134–135, 138, 145, 168, 174, 178–179, 211, 219
single cells 135 single package 58single species 53, 124, 125, 138skeletal remains 112, 221
skeleton 22, 51, 115, 121–122, 147Skhul 53–55, 147–148, 151skull 111, 123slave systems 192Smith Maynard 134-135smouldering process 119snail shells 9sob 5social 4–6, 8, 16–18, 24, 27–29,
37–41, 49–50, 58–59, 63, 75, 90, 111, 113–114, 117, 121, 125–127, 133–138, 141, 146, 150, 152–155, 163–170, 173–178, 180, 186, 190–191, 199, 209, 211, 218
social bonding 173social control 138social display 9social dominance 4social identification/
differentiation 127social identity 113, 114social insect colonies 134social insect colony 135social intelligence
hypothesis 191social interactions 38, 41,
165, 174social order 168 social rank 5, 50social reorganization 17social scientists 134sociality 5, 39societal boundaries 5socio-ecological 3solid citizen genes 134 song 1, 4, 23, 99, 163, 174, 179song elements 1, 4 Sosis Richard 210sounds 5, 15, 31, 34, 37, 172,
178–179, 212, 214South Africa 51–53, 55–56, 62,
65, 78, 92–93, 113, 143, 147south east Asia 54southern Africa 52, 56, 62–64,
75–76, 80, 97southern Cape 81–82Spain 53–54, 121, 151span tasks 100spatial 17, 100, 148, 171–172spatulas 56speaking 168, 207–208spear points 56spearheads 101

236 Index
speciation 19–20, 58–60, 76, 111–113, 124
species intersterility 120speech 3, 5, 22–26, 28, 99,
168–169, 173, 177, 187spirits 206, 214, 216, 221spirituality 2, 7, 9–10, 133,
136–138split brain patients 16spoken language 5, 26, 28,
173–174spontaneous referentiality 1, 4spruce trees 118SST 62St.-Césaire 121stable isotope 62Stage 3 Project 63stain 56Standard View 206–208state society 144Stewart Guthrie 213Still Bay 52–53, 55–56, 78, 82Still Bay techno-tradition 82stimulus 6, 166, 171stimulus bound behavior 166 stimulus freedom 166stimulus response 6stone 2, 10, 51, 56–59, 64,
75–77, 83–84, 87, 97–98, 101–106, 111–113, 117, 119, 143, 145, 193, 200
stone hammers 2stone inserts 97, 101, 105, 97,
101, 105,stone missiles 10 stone tool 51, 56, 64, 76, 84, 87,
101, 102, 106, 112, 117, 119stone tool hafting 119stone tools 51, 56, 64, 76, 84,
97, 101, 102, 106, 117storage containers 81Straitzig/Krems-Rehberg 57stratigraphic 78, 82–83striations 83–84Stringer 9, 50–51, 63, 112, 120style 6, 17, 35, 39, 56, 198stylistic variability 81sub-Saharan Africa 75subsistence 2, 75, 100, 119supernatural 9, 154–155, 205,
208–210superorganism 134suppression of fitness
differences 135
survival 18, 21, 33, 37, 40, 118–119, 163, 170–171, 211
SW France 151Swabian Jura 113Sydow Wolfgang 77symbiotic cell theory 134symbolic 7–9, 15, 22, 26, 28,
49–51, 56–60, 63–64, 75–77, 88–92, 97–98, 102, 104, 107, 111–114, 122, 124, 127, 133, 136–137, 141–144, 146–150, 152–158, 163–165, 171–174, 179–180, 187–188, 190–191, 200, 205–206, 222
symbolic ability 22symbolic artefacts 49, 58, 188,
190, 200 symbolic behaviour 22, 49, 58,
76, 92, 133, 144, 149, 157, 188, 200, 205
symbolic capacity 9, 56, 141, 142, 144, 153, 154, 157
symbolic culture 59, 187symbolic evolution 141, 153, 206 symbolic hypothesis 89symbolic innovations 98symbolic intent 76, 92 symbolic material 49–51, 58,
75, 97Symbolic Neandertal 50–51symbolic thinking 50, 111–113,
124, 127, 172 symbolically mediated
behaviour 49–50, 76Symbolism 49–50, 75, 97–98,
102, 107, 125, 141–151, 153–156, 158, 172, 176, 179, 198, 205–206, 218–222
symbols 6, 14, 26–27, 50, 56, 75, 82, 89–90, 92, 114, 117, 141–142, 145, 148, 150, 152, 164–165, 171–172, 178, 180
symbol-use 3, 6–7, 9syncopated gallop 4syntactic 4, 167, 173, 191, 193, 198syntactic architecture 173syntactic category 167 syntactic hypothesis 191, 193 syntactic integration 173 syntactic processing 4 syntax 4, 15, 27, 169, 173, 175,
187, 192–193, 195, 197synthetic iron oxide 103Szathmary 134–135
TTabun 51, 152Tabun C1 burial 51Taforalt 54, 98, 147Tai Forest 8talk 98, 168talking apes 26Tamuli 28tanning of hides 118Tan-Tan 146Tanzania 8Taphonomic 84task-appropriate recipes 102Tasmania 58–59taxonomic status 120teamwork 133–134, 136–138techno-complexes 56techno-tradition 78, 82teeth 24–25, 121, 123temporoparietal 193–194, 197,
199Terrence Deacon 137terrestrial 63–64Texier 52, 78, 80–81, 90–91The Symbolic Species 137theory of mind 133, 153–154,
178–179, 191–195, 198, 215, 218, 221–222
therianthropic 113therianthropic ivory
figurines 113 thermoluminescence 78, 82thermoregulatory models 118Thomas Wynn 191–192thought experiments 99tiger shark tooth 54time depth 144time/space-factored 148–149,
152–153, 155–157TL 78, 80, 82, 147Toba super-eruption 62Tobias 179ToM 29, 133, 215–218, 220–222Tomasello 30, 89, 133, 136–137,
169, 171, 176–178, 191, 197, 218–221
tool kit 207, 218, 220tools 10, 49, 51, 56–58, 60,
63–64, 75–77, 82, 84, 88, 90, 97, 99, 101–107, 117, 125, 174, 195–199, 211
tooth enamelisation 25top-down causation 163,
167–169

Index 237
toys 180traces 53, 98, 102, 180,
214, 221transition 31, 64, 133–136, 138,
168, 179, 197Trevarthen 31, 34triadic awareness 5Trinkaus 112, 115, 122–123, 153Tübingen 54, 114Turkey 147Twin Rivers 147twine 101
UUcagizli Cave 147Upper Palaeolithic 9, 64, 75,
142, 144, 148, 153Upper Pleistocene 142–143upraised arms 4Ursus deningeri 151usage-based language
skills 168use-wear 53, 121
VVarga-Khadem 22vegetable 84Venus 114
Venus pendant 114 verbal self-report 3Verlorenvlei River 78vertebrates 177Vindija 117vision 49, 171–173visual thinking 163, 172–174visuospatial 192vocal 4–5, 7–8, 15, 22–23, 28,
30–31, 34, 37–38, 168, 174vocal chords 168 vocal fluency 15, 23 vocal imitation 8 von Frisch 7
WWamba 8, 15warfare 136warming 60–62water-proof clothing 118weapons 97, 101–102,
105, 118Wernicke’s area 174, 193, 195Western Australia 54Western Cape 52, 77–78, 98whales 7whistling 8white 5, 17, 23, 136, 196
white matter 196 white sclera 136 Wisconsin Card Sort
Task 100within-group 133–136within-group
competition 135within-group
cooperation 134within-group selection 134,
136Wolfgang Sydow 77–78wolves 30, 136wooden artefacts 118working memory 50, 97,
99–100, 105, 191–196, 198working memory
hypothesis 191 writing 113, 141, 144, 168, 173Wynn and Coolidge 100
Yyellow 28, 101, 103Yerkes 26
ZZambia 147zoological specimens 14





















