
Habitat characteristics and macroinvertebrate assemblages in boreal forest streams: relations to...
13
Hydrobiologia 474: 239–251, 2002. © 2002 Kluwer Academic Publishers. Printed in the Netherlands. 239 Habitat characteristics and macroinvertebrate assemblages in boreal forest streams: relations to catchment silvicultural activities Petri Liljaniemi 1.∗ , Kari-Matti Vuori 2 , Boris Ilyashuk 3 & Hannu Luotonen 2 1 University of Jyväskylä, Department of Biological and Environmental Science, P.O. Box 35, 40351 Jyväskylä, Finland Tel.: +358-400-665143. Fax: +358-14-2602321. E-mail: [email protected].fi 2 North Karelia Regional Environment Centre, P.O. Box 69, 80101 Joensuu, Finland 3 Institute of North Industrial Ecology Problems, Kola Science Centre, Russian Academy of Sciences, 14 Fersman St., INEP, Apatity, Murmansk Region, 184200, Russia ( ∗ Author for correspondence) Received 14 December 2000; in revised form 28 January 2002; accepted 6 February 2002 Key words: headwater streams, macroinvertebrates, habitat structure, silviculture, woody debris Abstract We compared the stream habitat characteristics and macroinvertebrate assemblages of boreal headwater streams in both the Finnish and the Russian parts of a single river basin, the Koitajoki River. Over the last 50 years, the Finnish side of the catchment has been managed using modern forestry techniques, whereas Russian side has remained nearly unexploited and is near to its natural state. Differences in silvicultural activities were observed to contribute to differences in habitat structure. The channel habitats were in fairly natural state in the Russian reference streams, whereas the impacted Finnish sites were cleared and straightened. In comparison with the impacted channels, the abundance of coarse woody debris (CWD) was 10–100-fold higher in the reference streams. Implications on the forestry-induced deterioration of water quality were also observed. On the contrary, only small differences in macroinvertebrate assemblages were detected. Despite the lower amount of retentive structures (CWD), sig- nificantly higher relative abundance of shredders was observed in the forestry-impacted streams. Otherwise the zoobenthic communities were quite similar in the two subcatchments. We suggest that several mechanisms may explain this similarity: (1) community structure is controlled by naturally acidic conditions, (2) the adverse impacts of forestry on habitat structure and water quality of streams may be compensated by increased input of deciduous litter and organic compounds from drained, structurally young riparian forests and (3) macroinvertebrate species have flexible feeding habits and may thus readily adapt to changing conditions. Introduction Silviculture is a major form of land use contributing to environmental changes in many boreal headwater catchments. Clear-cutting and drainage, scarification and fertilization of forest soil have long-term effects on the discharge, water quality, temperature and sub- strate composition of streams (Golladay et al., 1989; Garman & Moring, 1991; Ahtiainen, 1992; Vuori & Joensuu, 1996; Holopainen & Huttunen, 1998; Vuori et al., 1998). Furthermore, forestry practices have been observed to alter the water balance, geomorphology and vegetation cover of the riparian zone (Osborne & Kovacic, 1993; Huggenberger et al., 1998). Straight- ening and clearing of streams to enable log floating, and drying of peatlands to enhance wood production have homogenized the channel morphology and de- creased the retention time of allochthonous material, thus decreasing the amount of food and habitats avail- able for stream biota (Golladay et al., 1989; Petersen & Petersen Jr., 1991; Maser & Sedell, 1994; Haapala & Muotka, 1998; Laasonen et al., 1998). A major problem in assessing the impact of water- shed land use on aquatic ecosystems is often the lack of reliable reference conditions. In Western Europe and North America, it is difficult to find catchment
Transcript of Habitat characteristics and macroinvertebrate assemblages in boreal forest streams: relations to...

Hydrobiologia 474: 239–251, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
239
Habitat characteristics and macroinvertebrate assemblages in borealforest streams: relations to catchment silvicultural activities
Petri Liljaniemi1.∗, Kari-Matti Vuori2, Boris Ilyashuk3 & Hannu Luotonen2
1University of Jyväskylä, Department of Biological and Environmental Science, P.O. Box 35,40351 Jyväskylä, FinlandTel.: +358-400-665143. Fax: +358-14-2602321. E-mail: [email protected] Karelia Regional Environment Centre, P.O. Box 69, 80101 Joensuu, Finland3Institute of North Industrial Ecology Problems, Kola Science Centre, Russian Academy of Sciences,14 Fersman St., INEP, Apatity, Murmansk Region, 184200, Russia(∗Author for correspondence)
Received 14 December 2000; in revised form 28 January 2002; accepted 6 February 2002
Key words: headwater streams, macroinvertebrates, habitat structure, silviculture, woody debris
Abstract
We compared the stream habitat characteristics and macroinvertebrate assemblages of boreal headwater streams inboth the Finnish and the Russian parts of a single river basin, the Koitajoki River. Over the last 50 years, the Finnishside of the catchment has been managed using modern forestry techniques, whereas Russian side has remainednearly unexploited and is near to its natural state. Differences in silvicultural activities were observed to contributeto differences in habitat structure. The channel habitats were in fairly natural state in the Russian reference streams,whereas the impacted Finnish sites were cleared and straightened. In comparison with the impacted channels,the abundance of coarse woody debris (CWD) was 10–100-fold higher in the reference streams. Implications onthe forestry-induced deterioration of water quality were also observed. On the contrary, only small differencesin macroinvertebrate assemblages were detected. Despite the lower amount of retentive structures (CWD), sig-nificantly higher relative abundance of shredders was observed in the forestry-impacted streams. Otherwise thezoobenthic communities were quite similar in the two subcatchments. We suggest that several mechanisms mayexplain this similarity: (1) community structure is controlled by naturally acidic conditions, (2) the adverse impactsof forestry on habitat structure and water quality of streams may be compensated by increased input of deciduouslitter and organic compounds from drained, structurally young riparian forests and (3) macroinvertebrate specieshave flexible feeding habits and may thus readily adapt to changing conditions.
Introduction
Silviculture is a major form of land use contributingto environmental changes in many boreal headwatercatchments. Clear-cutting and drainage, scarificationand fertilization of forest soil have long-term effectson the discharge, water quality, temperature and sub-strate composition of streams (Golladay et al., 1989;Garman & Moring, 1991; Ahtiainen, 1992; Vuori &Joensuu, 1996; Holopainen & Huttunen, 1998; Vuoriet al., 1998). Furthermore, forestry practices have beenobserved to alter the water balance, geomorphologyand vegetation cover of the riparian zone (Osborne &
Kovacic, 1993; Huggenberger et al., 1998). Straight-ening and clearing of streams to enable log floating,and drying of peatlands to enhance wood productionhave homogenized the channel morphology and de-creased the retention time of allochthonous material,thus decreasing the amount of food and habitats avail-able for stream biota (Golladay et al., 1989; Petersen& Petersen Jr., 1991; Maser & Sedell, 1994; Haapala& Muotka, 1998; Laasonen et al., 1998).
A major problem in assessing the impact of water-shed land use on aquatic ecosystems is often the lackof reliable reference conditions. In Western Europeand North America, it is difficult to find catchment

240
basins in such a natural state that they could be usedas a reference for comparison with variously affectedcatchments (Benke, 1990; Zwick, 1992; Muhar etal.,1995). For Finnish streams and rivers, however,many neighboring catchments in Northwestern Rus-sia offer an opportunity for such a comparison, be-cause they have largely remained unexploited since the1930s (Vuori et al., 1999).
The aim of our study was to compare the habitatcharacteristics and composition of the benthic fauna innatural and silviculturally modified tributaries of theKoitajoki River, a brown water, acidic river systemlocated on both sides of the Finnish–Russian border.We hypothesized that the structure and function ofbenthic communities may reflect the impacts of long-term silvicultural activities, presumably as changes inthe relative abundances of functional feeding groupsand an overall impoverishment of benthic diversity.
Study area
The study was conducted in the catchment area of theKoitajoki River, located in North Karelia on both sidesof the Finnish–Russian border (Fig. 1). The drainagearea of the studied headwaters is 2262 km2 (522 km2
in Finland) and lake covering 5.9%. The bedrock ofthe drainage area is mainly gneiss–granite. The soilconsists mostly of peat and moraine. More than 50%of the land area is covered by peatland, the rest beingmainly forests. The Koitajoki River is a part of theVuoksi drainage basin.
Forestry is the main form of land use in the head-water areas of the Koitajoki River. Before the 1940s,when the whole catchment was part of Finland, thesilvicultural activities in this area were quite limitedin intensity and spatial coverage. After the SecondWorld War, since 1944, the eastern parts of catchmentarea were connected to Russia, and major differencesin silviculture started to appear. The period of mostintensive forestry in the Finnish side of the studyarea began in the 1960s. Large scale clear cuttings,scarifications and forest road constructions have beenconducted until the late 1990s. In addition, almost allof the wetlands have been drained, and only some pro-tected bogs have been left untouched. The drainageswere mainly done in the 1960s, but continued untilthe 1990s. The forests are dominated by saplings andyoung stands of pine (Pinus sylvestris). The riparianwoodlands are mainly covered by mixture of saplingsand young stands of coniferous and deciduous trees.
Figure 1. Study area and sampling sites. Codes of the qualitat-ive samples are in italics. Sites R10 and R14 are located in theHaapajoki River, 5 km north of the Koitajoki River.
The stream channels, including the first-order brooks,were cleared and straightened during the first half ofthe 19th century. Fertilizations of the forest soil anddrained peatlands have been carried out over the last30 years.
On the Russian side practically all wetlands are innatural state. Apart from some small scale loggingsand scarification, silvicultural activities and other hu-man impacts in the Russian side of the catchmenthave been minimal, especially during the last 50years. Some indications of stream channel modifica-tions, made at the beginning of the 20th century tofacilitate log floating, can still be seen, although theaccumulated woody material has often restructured thechannels. Forests and riparian areas are mainly dom-inated by coniferous forests in late successional stage.Compared to the Finnish areas, the Russian study sitescan be regarded as being close to their natural state.
241
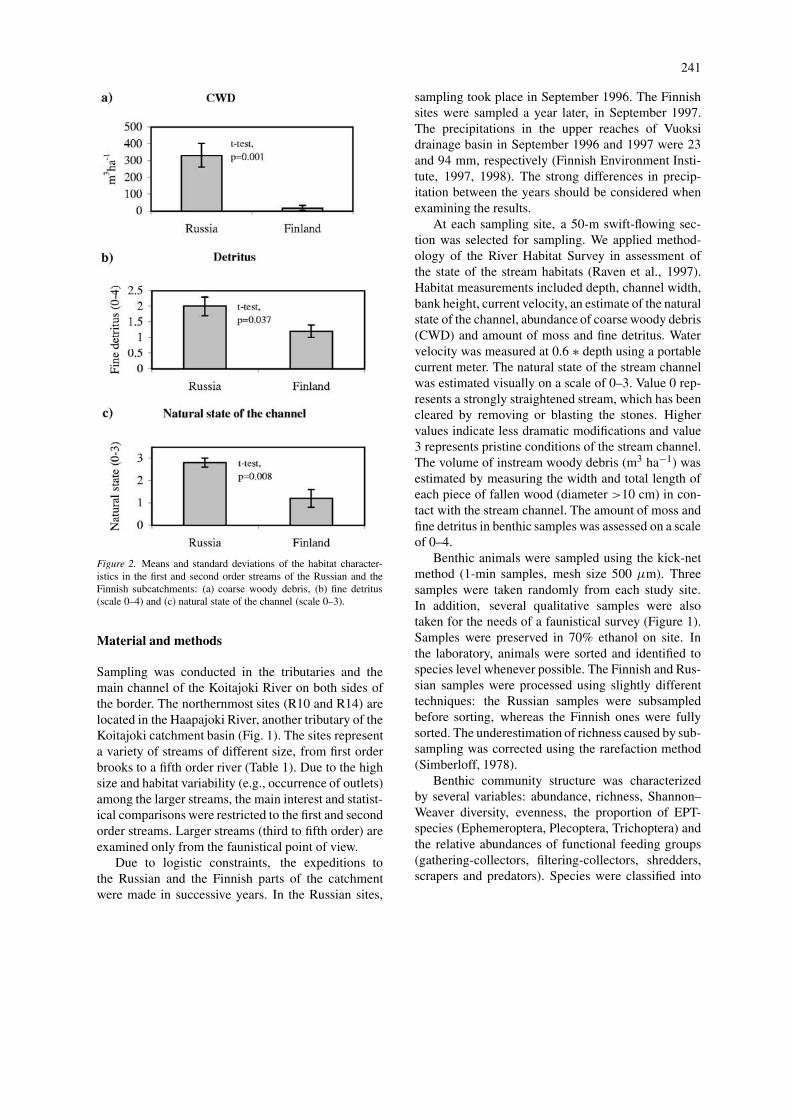
Figure 2. Means and standard deviations of the habitat character-istics in the first and second order streams of the Russian and theFinnish subcatchments: (a) coarse woody debris, (b) fine detritus(scale 0–4) and (c) natural state of the channel (scale 0–3).
Material and methods
Sampling was conducted in the tributaries and themain channel of the Koitajoki River on both sides ofthe border. The northernmost sites (R10 and R14) arelocated in the Haapajoki River, another tributary of theKoitajoki catchment basin (Fig. 1). The sites representa variety of streams of different size, from first orderbrooks to a fifth order river (Table 1). Due to the highsize and habitat variability (e.g., occurrence of outlets)among the larger streams, the main interest and statist-ical comparisons were restricted to the first and secondorder streams. Larger streams (third to fifth order) areexamined only from the faunistical point of view.
Due to logistic constraints, the expeditions tothe Russian and the Finnish parts of the catchmentwere made in successive years. In the Russian sites,
sampling took place in September 1996. The Finnishsites were sampled a year later, in September 1997.The precipitations in the upper reaches of Vuoksidrainage basin in September 1996 and 1997 were 23and 94 mm, respectively (Finnish Environment Insti-tute, 1997, 1998). The strong differences in precip-itation between the years should be considered whenexamining the results.
At each sampling site, a 50-m swift-flowing sec-tion was selected for sampling. We applied method-ology of the River Habitat Survey in assessment ofthe state of the stream habitats (Raven et al., 1997).Habitat measurements included depth, channel width,bank height, current velocity, an estimate of the naturalstate of the channel, abundance of coarse woody debris(CWD) and amount of moss and fine detritus. Watervelocity was measured at 0.6 ∗ depth using a portablecurrent meter. The natural state of the stream channelwas estimated visually on a scale of 0–3. Value 0 rep-resents a strongly straightened stream, which has beencleared by removing or blasting the stones. Highervalues indicate less dramatic modifications and value3 represents pristine conditions of the stream channel.The volume of instream woody debris (m3 ha−1) wasestimated by measuring the width and total length ofeach piece of fallen wood (diameter >10 cm) in con-tact with the stream channel. The amount of moss andfine detritus in benthic samples was assessed on a scaleof 0–4.
Benthic animals were sampled using the kick-netmethod (1-min samples, mesh size 500 µm). Threesamples were taken randomly from each study site.In addition, several qualitative samples were alsotaken for the needs of a faunistical survey (Figure 1).Samples were preserved in 70% ethanol on site. Inthe laboratory, animals were sorted and identified tospecies level whenever possible. The Finnish and Rus-sian samples were processed using slightly differenttechniques: the Russian samples were subsampledbefore sorting, whereas the Finnish ones were fullysorted. The underestimation of richness caused by sub-sampling was corrected using the rarefaction method(Simberloff, 1978).
Benthic community structure was characterizedby several variables: abundance, richness, Shannon–Weaver diversity, evenness, the proportion of EPT-species (Ephemeroptera, Plecoptera, Trichoptera) andthe relative abundances of functional feeding groups(gathering-collectors, filtering-collectors, shredders,scrapers and predators). Species were classified into
242
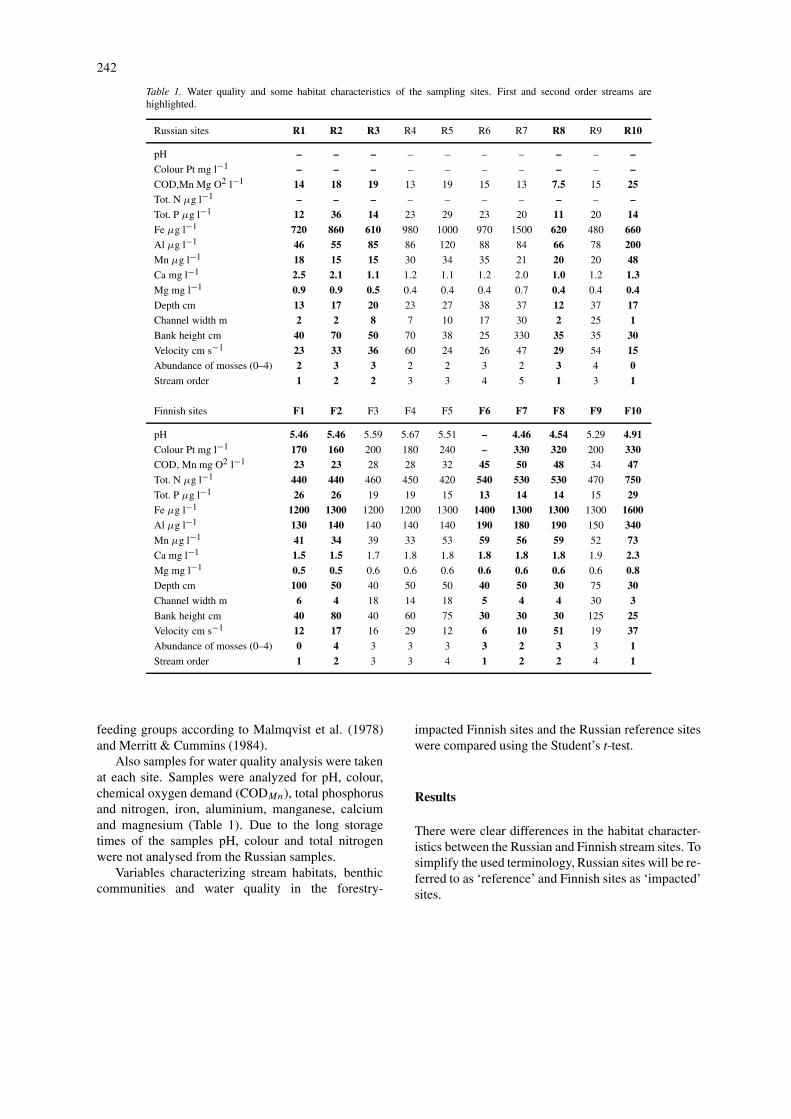
Table 1. Water quality and some habitat characteristics of the sampling sites. First and second order streams arehighlighted.
Russian sites R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
pH – – – – – – – – – –Colour Pt mg l−1 – – – – – – – – – –COD,Mn Mg O2 l−1 14 18 19 13 19 15 13 7.5 15 25Tot. N µg l−1 – – – – – – – – – –Tot. P µg l−1 12 36 14 23 29 23 20 11 20 14Fe µg l−1 720 860 610 980 1000 970 1500 620 480 660Al µg l−1 46 55 85 86 120 88 84 66 78 200Mn µg l−1 18 15 15 30 34 35 21 20 20 48Ca mg l−1 2.5 2.1 1.1 1.2 1.1 1.2 2.0 1.0 1.2 1.3Mg mg l−1 0.9 0.9 0.5 0.4 0.4 0.4 0.7 0.4 0.4 0.4Depth cm 13 17 20 23 27 38 37 12 37 17Channel width m 2 2 8 7 10 17 30 2 25 1Bank height cm 40 70 50 70 38 25 330 35 35 30Velocity cm s−1 23 33 36 60 24 26 47 29 54 15Abundance of mosses (0–4) 2 3 3 2 2 3 2 3 4 0Stream order 1 2 2 3 3 4 5 1 3 1
Finnish sites F1 F2 F3 F4 F5 F6 F7 F8 F9 F10
pH 5.46 5.46 5.59 5.67 5.51 – 4.46 4.54 5.29 4.91Colour Pt mg l−1 170 160 200 180 240 – 330 320 200 330COD, Mn mg O2 l−1 23 23 28 28 32 45 50 48 34 47Tot. N µg l−1 440 440 460 450 420 540 530 530 470 750Tot. P µg l−1 26 26 19 19 15 13 14 14 15 29Fe µg l−1 1200 1300 1200 1200 1300 1400 1300 1300 1300 1600Al µg l−1 130 140 140 140 140 190 180 190 150 340Mn µg l−1 41 34 39 33 53 59 56 59 52 73Ca mg l−1 1.5 1.5 1.7 1.8 1.8 1.8 1.8 1.8 1.9 2.3Mg mg l−1 0.5 0.5 0.6 0.6 0.6 0.6 0.6 0.6 0.6 0.8Depth cm 100 50 40 50 50 40 50 30 75 30Channel width m 6 4 18 14 18 5 4 4 30 3Bank height cm 40 80 40 60 75 30 30 30 125 25Velocity cm s−1 12 17 16 29 12 6 10 51 19 37Abundance of mosses (0–4) 0 4 3 3 3 3 2 3 3 1Stream order 1 2 3 3 4 1 2 2 4 1
feeding groups according to Malmqvist et al. (1978)and Merritt & Cummins (1984).
Also samples for water quality analysis were takenat each site. Samples were analyzed for pH, colour,chemical oxygen demand (CODMn), total phosphorusand nitrogen, iron, aluminium, manganese, calciumand magnesium (Table 1). Due to the long storagetimes of the samples pH, colour and total nitrogenwere not analysed from the Russian samples.
Variables characterizing stream habitats, benthiccommunities and water quality in the forestry-
impacted Finnish sites and the Russian reference siteswere compared using the Student’s t-test.
Results
There were clear differences in the habitat character-istics between the Russian and Finnish stream sites. Tosimplify the used terminology, Russian sites will be re-ferred to as ‘reference’ and Finnish sites as ‘impacted’sites.

243
The amount of coarse woody debris was 10–100-fold higher in the reference streams and differedsignificantly from the impacted sites (t-test, d.f.=9,p<0.001). The average amount of CWD was 331.6 m3
ha−1 and 16.8 m3 ha−1 in reference and impactedsites, respectively. Also the amount of fine detrituswas significantly higher in reference streams (t-test,d.f.=9, p=0.037) (Fig. 2). The average classificationscores indicating the natural state of the stream chan-nels were 2.8 in Russia and 1.2 in Finland. TheRussian scores for the natural state of the stream chan-nel were significantly higher (t-test, d.f.=9, p=0.008)(Fig. 2). All stream sites in the Finnish subcatchmenthad clear marks of recent, intensive silvicultural activ-ities, whereas only few past, small-scale loggings werevisible in Russian sites. Furthermore, loggings anddrainage ditches frequently extended up to the banktopin the Finnish streams.
According to the water analyses the Finnish partsof the Koitajoki River catchment area are acidic andhumic (pH 4.46–5.67, colour: 160–330 Pt mg/l). Stud-ies carried out by Russian scientists in the vicinityof our sampling sites have also obtained similar pH(4.50–6.31) and colour (100–350 Pt mg/l) values (A.Litvinenko, Northern Water Problems Institute, Pet-rozavodsk, in litt.). The concentrations of major nu-trients indicate oligo-mesotrophic conditions (Tot. P:11–36 µg/l, Tot. N: 420–750 µg/l) (Dodds et al.,1998) (Table 1).
The greatest differences in water quality betweenreference and impacted streams were found in CODand in the concentrations of iron, aluminium andmanganese, which were significantly higher in theimpacted sites (t-test, d.f.=9, COD: p=0.006, Fe:p=0.000, Al: p=0.036 and Mn: p=0.006) (Table 1).
Zoobenthic measures did not exhibit major dif-ferences between the subcatchments. A total of 201taxa were identified from the benthic samples, 141taxa from the Russian streams and 166 from theFinnish parts of the catchment (Appendix). The meanmacroinvertebrate abundances were 1026 and 296 in-dividuals per sample in the reference and impactedstreams, respectively. However, this difference was notstatistically significant due to the high variance amongreference sites (Table 2).
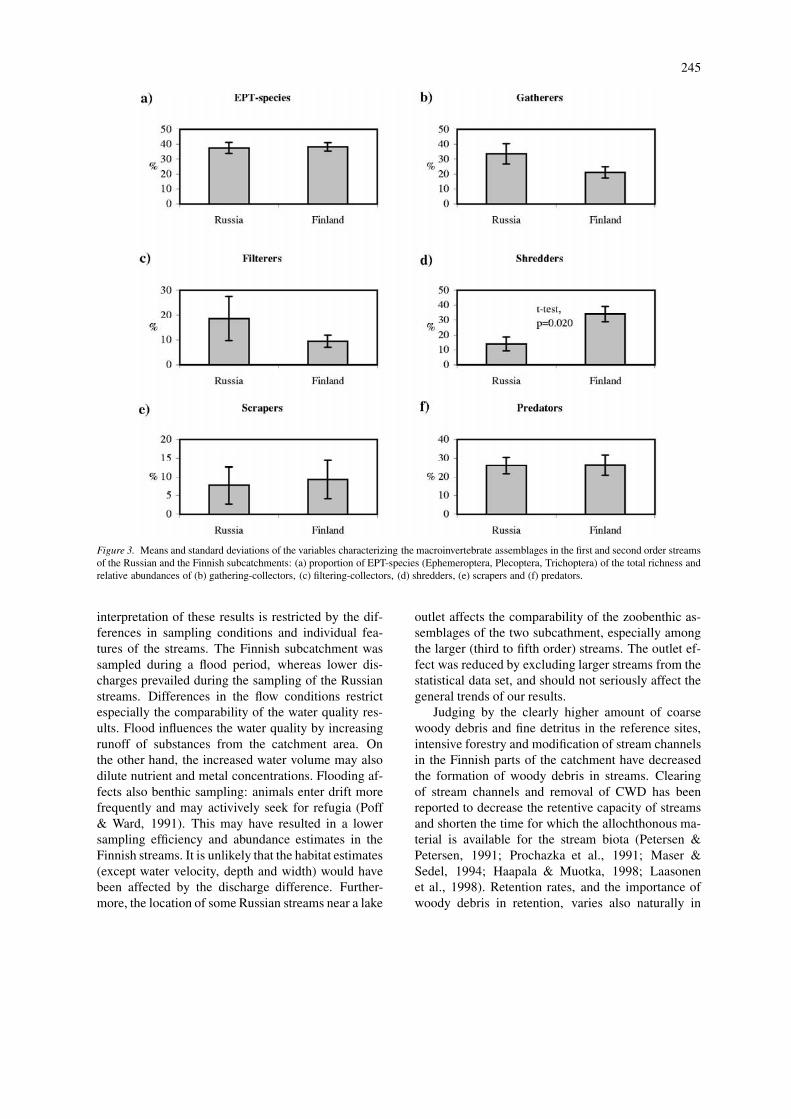
Total richness and diversity did not differ signi-ficantly between the reference and impacted streams.Further, only minor distinctions appeared in the rel-ative numbers of EPT-species (37.5 in reference and38.3 in impacted sites. Difference between the sub-catchments was unsignificant). More apparent differ-
ence was found when the relative abundances of thefeeding groups were compared. The relative abund-ance of shredders was significantly higher in theimpacted streams (13.9 in reference and 33.9 in im-pacted sites, t-test, d.f.=9, p=0.020) (Fig. 3). Dif-ferences between the subcatchments regarding therelative abundances of other feeding groups werenot significant despite the prominently high relat-ive abundance of filter-feeders in reference streams.Relative abundances of gathering-collectors, filtering-collectors, scrapers and predators were 33.6, 18.6, 7.7and 26,0, and 21.1, 9.5, 9.3 and 26.2 in Russia andFinland, respectively (Figure 3).
Although the predominant species were abundantin both reference and forestry-affected areas, therewere some differences in the relative abundance of themost common species. Especially the shredding ple-copterans, like Taeniopteryx nebulosa and Nemouraavicularis, were more common in silviculturally mod-ified parts of the catchment (Table 2). In larger streams(third to fifth order), the relative abundances offiltering-collector species, like Hydropsyche siltalai,H. pellucidula, Neureclipsis bimaculata, simuliids andchironomids of the Tanytarsinae-group (Tanytarsusspp., Micropsectra spp.), were higher in the Rus-sian streams, whereas shredders Lepidostoma hirtumand Micrasema gelidum were more abundant in theFinnish sites (Table 2).
The occurrence of some common species were re-stricted to a few sample sites. For example, Nemurellapictetii and Nemoura cinerea were found in great num-bers from small brooks in Russia (sites R12 and R13in Fig. 1). Nemoura flexuosa was abundant in theFinnish site F10, a small brown-water stream withhigh iron and aluminium content. The occurrence ofthe trichopteran predator Rhyacophila fasciata wasconfined almost exclusively to small brooks in theRussian parts of the catchment. Species regardedas endangered (Aphelocheirus aestivalis, Arctopsycheladogensis and Semblis atrata) by the Finnish environ-mental authorities occurred scarcely in both countries(Committee for the Monitoring of Threatened Animalsand Plants in Finland, 1992).
Discussion
Our results indicate clear forestry-induced differencesin stream habitat characteristics, while distinctionsbetween zoobenthic assemblages of the subcatch-ments were less obvious. We are aware that the

244
Tabl
e2.
Mea
nab
unda
nce
ofal
lani
mal
san
dre
lativ
eab
unda
nces
ofth
edo
min
ants
peci
es(r
elat
ive
abun
danc
e>
10%
).Fi
rsta
ndse
cond
orde
rst
ream
sar
ehi
ghlig
hted
Site
R1
R2
R3
R4
R5
R6
R7
R8
R9
R10
F1
F2
F3F4
F5F
6F
7F
8F9
F10
Tota
labu
ndan
ce10
2051
924
2610
0819
1437
618
672
612
1044
130
844
114
7255
811
522
422
827
742
929
6E
phem
erop
tera
Lep
toph
lebi
am
argi
nata
22.2
9.5
1.1
2.0
4.8
2.2
0.2
2.6
1.6
12.7
25.7
0.3
3.0
0.7
11.9
2.4
1.2
3.6
6.0
1.0
Plec
opte
ra
Taen
iopt
eryx
nebu
losa
0.5
1.5
0.1
1.9
4.4
0.4
24.0
0.5
2.5
0.1
–10
.84.
80.
91.
212
.218
.44.
96.
80.
8N
emou
raav
icul
aris
0.1
0.7
–0.
10.
1–
–0.
10.
10.
11.
50.
10.
40.
15.
28.
54.
731
.31.
4–
N.fl
exuo
sa–
––
––
––
––
––
–0.
3–
––
–0.
1–
10.9
Prot
onem
ura
mey
eri
12.3
––
0.9
–0.
40.
4–
0.1
0.1
–0.
20.
20.
1–
0.1
–7.
9–
–L
euct
radi
gita
ta6.
921
.0–
0.2
0.1
–1.
88.
81.
90.
6–
4.3
0.5
3.5
0.3
4.3
2.2
13.1
5.1
16.1
Col
eopt
era
Elm
isae
nea
8.9
1.3
0.1
1.7
0.6
1.7
9.7
12.5
6.2
––
7.2
4.8
7.5
6.4
0.1
0.1
0.5
5.0
0.1
Oul
imni
ustu
berc
ulat
us0.
7–
––
0.1
–0.
913
.40.
1–
21.8
2.6
0.2
1.2
2.6
0.1
–0.
51.
21.
4T
rich
opte
ra
Hyd
rops
yche
pellu
cidu
la–
––
1.2
–0.
715
.82.
76.
7–
–0.
15.
80.
9–
––
0.1
0.7
–H
.silt
alai
––
–13
.00.
744
.81.
10.
516
.8–
–0.
118
.0–
––
––
––
Neu
recl
ipsi
sbi
mac
ulat
a–
–19
.934
.226
.015
.1–
–6.
90.
51.
20.
21.
4–
––
––
–0.
6Po
lyce
ntro
pus
flavo
mac
ulat
us11
.03.
44.
11.
62.
84.
1–
3.4
1.4
6.0
4.4
1.9
0.6
0.3
0.3
20.3
25.4
9.6
3.7
17.3
Mic
rase
ma
gelid
um2.
7–
––
––
––
0.1
––
–3.
04.
215
.1–
––
20.2
0.1
Lep
idos
tom
ahi
rtum
––
––
0.1
–0.
70.
76.
7–
1.2
5.1
11.8
6.4
8.4
–0.
4–
4.7
0.1
Dip
tera
Sim
uliid
ae7.
413
.4–
–0.
61.
92.
21.
21.
00.
2–
0.9
0.3
8.6
1.4
0.4
0.6
0.1
0.3
1.1
Chi
rono
mid
ae
Con
chap
elop
iasp
p.2.
113
.717
.73.
56.
41.
80.
71.
20.
73.
60.
90.
40.
30.
32.
68.
07.
72.
53.
912
.4E
ukie
ffer
iella
devo
nica
gr.
0.4
1.6
–0.
10.
1–
0.4
–0.
5–
–12
.80.
55.
6–
19.8
1.5
11.2
0.1
–T
vete
nia
pauc
unca
3.4
0.5
0.2
1.2
0.3
1.4
0.4
0.7
10.7
1.2
–6.
00.
90.
8–
0.1
0.3
0.8
0.4
0.3
Mic
rops
ectr
asp
p.4.
21.
523
.31.
10.
50.
5–
4.6
0.3
48.5
0.2
––
––
0.1
0.3
0.1
–5.
1Ta
nyta
rsus
chin
yens
isgr
.1.
00.
420
.43.
34.
3–
0.2
0.8
3.3
10.9
0.6
1.7
2.5
0.3
–5.
827
.33.
21.
29.
8
245
Figure 3. Means and standard deviations of the variables characterizing the macroinvertebrate assemblages in the first and second order streamsof the Russian and the Finnish subcatchments: (a) proportion of EPT-species (Ephemeroptera, Plecoptera, Trichoptera) of the total richness andrelative abundances of (b) gathering-collectors, (c) filtering-collectors, (d) shredders, (e) scrapers and (f) predators.
interpretation of these results is restricted by the dif-ferences in sampling conditions and individual fea-tures of the streams. The Finnish subcatchment wassampled during a flood period, whereas lower dis-charges prevailed during the sampling of the Russianstreams. Differences in the flow conditions restrictespecially the comparability of the water quality res-ults. Flood influences the water quality by increasingrunoff of substances from the catchment area. Onthe other hand, the increased water volume may alsodilute nutrient and metal concentrations. Flooding af-fects also benthic sampling: animals enter drift morefrequently and may activively seek for refugia (Poff& Ward, 1991). This may have resulted in a lowersampling efficiency and abundance estimates in theFinnish streams. It is unlikely that the habitat estimates(except water velocity, depth and width) would havebeen affected by the discharge difference. Further-more, the location of some Russian streams near a lake
outlet affects the comparability of the zoobenthic as-semblages of the two subcathment, especially amongthe larger (third to fifth order) streams. The outlet ef-fect was reduced by excluding larger streams from thestatistical data set, and should not seriously affect thegeneral trends of our results.
Judging by the clearly higher amount of coarsewoody debris and fine detritus in the reference sites,intensive forestry and modification of stream channelsin the Finnish parts of the catchment have decreasedthe formation of woody debris in streams. Clearingof stream channels and removal of CWD has beenreported to decrease the retentive capacity of streamsand shorten the time for which the allochthonous ma-terial is available for the stream biota (Petersen &Petersen, 1991; Prochazka et al., 1991; Maser &Sedel, 1994; Haapala & Muotka, 1998; Laasonenet al., 1998). Retention rates, and the importance ofwoody debris in retention, varies also naturally in

246
different stream systems depending on channel gradi-ent, geomorphology and characteristics of the riparianvegetation (Cummins et al., 1989; Berg et al., 1998).
Studied headwaters appear to be naturally acidicand humic. The silvicultural activities may be thereason for the elevated concentrations of metals andsuspended solids (indicated by the higher COD) inforestry-impacted streams, although impact of flood-ing can not be ruled out. However, results of ametal contamination study conducted in our samplingsites lean support to our hypothesis on the forestry-induced deterioration of the water quality (Vuori et al.2002, unpublished data). In this study, aquatic mosssamples (Fontinalis antipyretica) were sampled in au-tumn 1996 and significantly higher metal concentra-tions were measured in the Finnish forestry-impactedstreams as compared to the Russian streams. Suchforestry-induced changes in water quality have alsobeen observed in other studies (Garman & Moring,1991; Vuori & Joensuu, 1996; Vuori et al., 1998).The modification of catchment soil and forests havebeen reported to cause long-term changes in the wa-ter quality of streams. Studies conducted in Nurmes,located in the same ecoregion as the Koitajoki River,reported a drastic increase in nutrient content, suspen-ded solids and colour of water following clear-cuttingand draining of the catchment. The levels of nutrients,suspended solids and water colour were still elevated10 years after catchment modifications (Holopainen &Huttunen, 1998; Ahtiainen & Huttunen, 1999).
Despite the differences in habitat characteristicsand water quality between the reference and impactedstreams, benthic community structure and species as-semblages appeared relatively similar. Richness anddiversity may be useful indicators of acute and intensepollution (e.g., toxic pollutants, heavy organic pollu-tion), but are quite ineffective indicators of forestry-induced disturbances that only cause shifts in rel-ative abundances (Perkins, 1983; Stone & Wallace,1998). Although the abundance of benthic macroin-vertebrates did not differ significantly between theFinnish and the Russian parts of the catchment, meanabundances were notably higher in several referencestreams. This may be a sampling artifact caused bydifferences in sampling time, discharge conditions andsample processing techniques between the subcatch-ments. However, high macroinvertebrate density andbiomass associated with rich CWD has been reportedin several studies (Smock et al., 1989; Wallace et al.,1995; Friberg & Larsen, 1998).
According to several studies, streams with a higherretention capacity, and thus more abundant litter,
support higher densities of shredding invertebrates(Molles, 1982; Prochazka et al., 1991; Dobson et al.,1995; Friberg, 1997; Murphy & Giller, 2000). Des-pite the higher amount of CWD and fine detritus inour reference sites, relative abundance of shredderswas higher in the impacted streams. This can be ex-plained by several factors. First, abundant CWD instream channels increase the proportional area of poolhabitats and slows down the current speed, thus in-creasing the sedimentation of fine particles. This mayresult in increased relative abundance of collectorsand filterers and corresponding decrease in shreddersand scrapers (Lemly & Hilderbrand, 2000). Second,forest succession affects the quality and quantity of al-lochthonic material entering the streams. Compared toold, conifer-dominated forest, litter originating froma young, mixed forest is more readily and effect-ively conditioned and decomposed by the stream biota(Cummins et al., 1989). Studies concerning the long-term effects of clear-cutting have reported increasedshredder abundances in forestry-affected streams andsuggested that the reason for the increase was the moreprofitable litter delivered by the young riparian forest(Stout et al., 1993; Stone & Wallace, 1998). Thismight also be the case in the Koitajoki River, wherethe riparian tree stands in Finnish forestry-impactedareas are considerably younger than their Russiancounterparts. Third possible explanation might be theopportunistic feeding habits of detritivorous inverteb-rates (Williams & Hynes, 1973; Friberg & Jacobsen,1994; Dobson et al., 1995). The increased run-off ofallochthonous material caused by the drainage of thecatchment area may compensate and even enrich thefood resources lost due to the lowered retentive capa-city of the stream channels. Shredding invertebratesmight be able to utilize these alternative food sourceswhen leaf litter is scarce. Furthermore, increased inputof nutrients from silvicultural activities may accel-erate decomposition and conditioning of low-qualitydetritus and make it more usable for detritivores (Vuori& Muotka, 1999).
The high relative abundance of filtering-collectorsin the reference subcatchment is a result of onesampling site (R3) located near a lake outlet. However,the difference between the impacted and referencestreams was not statistically significant. Lake outletswith stable substrata are known to be favourable hab-itats for filter-feeders (Hildrew & Townsend, 1976;Crichton et al., 1978; Boon, 1979; Brönmark & Malm-qvist, 1984; Stone & Wallace, 1998). The outlet effectexplains also the high proportion of filterers in lar-ger (third to fifth order) Russian streams, which were

247
excluded from the statistical analysis. The species as-semblages of these outlet streams closely resemble thefauna of similar habitats in the silviculturally affectedstreams of Eastern Finland (Hämäläinen & Huttunen,1990).
Allthough the relative abundances of functionalfeeding groups revealed some differences, silvicultur-ally modified and reference areas were faunisticallyquite similar. It is possible that the impacts of forestryon benthic assemblages of humic, naturally acidicstreams are overridden by natural constraints like lowpH. In the forestry-affected parts of the catchmentthe increased run-off of humic substances and nutri-ents probably buffers the harmful effects of acidityand increased metal concentrations. Compared to clearwaters, many species have been observed to toleratelower pH and higher metal concentrations in humicstreams (Gerhardt, 1993; Hämäläinen & Huttunen,1996, 1998).
Restricted distributions of species like Nemurellapictetii, Nemoura cinerea and N. flexuosa can beexplained by the individual characteristics of thestreams. Although inhabiting a variety of differentwater bodies, the acid-tolerant Nemurella pictetii andNemoura cinerea are most abundant in small, cool,acidic brooks and appear to be quite scarce in lar-ger streams (Dobson & Hildrew, 1992; Hämäläinen& Huttunen, 1996; Vuori et al., 1998). Accordingto Lillehammer (1988), the occurrence of Nemouraflexuosa is restricted to small streams, which is alsoevident in our material. Interspecific competition hasbeen suggested to be an important factor affecting thedistribution of Rhyacophila species (Cereghino et al.,1997). This might also be the case with Rhyacophilaspecies in the Koitajoki River: Rhyacophila nubilawas found most frequently in larger streams, while theoccurrence of R. fasciata was restricted to small first-order brooks. It is also possible that impacts of forestryhas affected the distribution of R. fasciata, since thespecies was found in any greater numbers only in thenatural parts of the catchment area.
In conclusion, several mechanisms may explainthe faunal similarity in the streams with markedlydifferent catchment land use, habitat structure andwater quality: (1) community structure is controlledby naturally acidic conditions, (2) increased flux oforganic matter and nutrients from the silviculturallymodified catchments may compensate for the adverseimpacts of simplified habitat structure, acidity andmetal contamination and (3) flexible feeding habits ofthe macroinvertebrate species enable adaption to thechanging environmental conditions.
Acknowledgments
We wish to thank the Northern Water Problems Insti-tute (NWPI) of the Russian Academy for co-operationin organizing the Russian expedition, especially Alex-ander Freindling for his help in the field and AlexanderLitvinenko for data concerning the water quality ofthe Russian sites. Antero Koikkalainen is gratefullyacknowledged for his efforts in the field and laborat-ory. Olavi Sandman, Reijo Lähteenmäki and rest ofthe field team from the South Savo Regional Environ-ment Centre provided us with lots of logistic supportduring the Russian expedition. We also thank the En-vironmental Laboratory of the North Karelia RegionalEnvironment Centre for conducting the water analysesand Heikki Hämäläinen and Timo Muotka for theircomments during the writing of the article.
References
Ahtiainen, M., 1992. The effects of forest clear-cutting and scar-ification on water quality of small brooks. Hydrobiologia 244:465–473.
Ahtiainen, M. & P. Huttunen, 1999. Long-term effects of forestrymanagements on water quality and loading in brooks. BorealEnv. Res. 4: 101–114.
Benke, A. C. 1990. A perspective on America’s vanishing streams.J. north am. benthol. Soc. 9: 77–88.
Berg, N., A. Carlson & D. Azuma, 1998. Function and dynamicsof woody debris in stream reaches in the central Sierra Nevada,California. Can. J. Fish. aquat. Sci. 55: 1807–1820.
Boon P. J. 1979. Studies on the spatial and temporal distributionof larval Hydropsychidae in North Tyne river system (NorthernEngland). Arch. Hydrobiol. 85: 336–359.
Brönmark, C. & B. Malmqvist, 1984. Spatial and temporal patternsof lake outlet benthos. Verh. Int. Ver. Limnol. 22: 1986–1991.
Cereghino, P., T. Boutet & P. Lavandier, 1997. Abundance, bio-mass, life history and growth of six Trichoptera species undernatural and hydropeaking conditions with hypolimnetic releasesin a Pyrenean stream. Arch. Hydrobiol. 138: 307–328.
Committee for the monitoring of Threatened animals and Plantsin Finland, 1992. Uhanalaisten eläinten ja kasvien seur-antatoiminkunnan mietintö (Report on the monitoring ofthreatened animals and plants in Finland, in Finnish). Ministryof the Environment, Committee report 1991: 30, 328 pp.
Crichton, M. I., D. Fisher & I. P. Woivod, 1978. Life historiesand distribution of British Trichoptera, excluding Limnephilidaeand Hydroptilidae, based on Rothamsted Insect Survey. Holarct.Ecol. 1: 31–45.
Cummins, K. W., M. A. Wilzbach, D. M. Gates, J. B. Perry & W. B.Taliaferro, 1989. Shredders and riparian vegetation. BioScience39: 24–30.
Dobson, M. & A. G. Hildrew, 1992. A test of resource limitationamong shredding detritivores in low order streams in southernEngland. J. Anim. Ecol. 61: 69–77.
Dobson, M., A. G. Hildrew, S. Orton & S. J. Ormerod, 1995.Increasing litter retention in moorland streams: ecological andmanagement aspects of a field experiment. Freshwat. Biol. 33:325–337.

248
Dodds, W. K., J. R. Jones & E. B. Welch, 1998: Suggested classific-ation of stream trophic state: Distributions of temperate streamtypes by chlorophyll, total nitrogen, and phosphorus. Wat. Res.32: 1455–1462.
Finnish Environment Institute, 1997. Hydrological Review 1996.Print Oy, Helsinki, pp. 3–4.
Finnish Environment Institute, 1998. Monthly hydrological report.Hydrological Review 1997. Print Oy, Helsinki, pp. 3–4.
Friberg, N., 1997. Benthic invertebrate communities in six Danishforest streams: Impact of forest type on structure and function.Ecography 29: 19–28.
Friberg, N. & D. Jacobsen, 1994. Feeding plasticity of twodetritivore-shredders. Freshwat. Biol. 32: 133–142.
Friberg, N. & S. E. Larsen, 1998. Microhabitat selection by streaminvertebrates: importance of detritus aggregations. Verh. int. Ver.Limnol. 26: 1016–1020.
Garman, G. C. & J . R. Moring, 1991. Initial effects of deforestationon physical characteristics of a boreal river. Hydrobiologia 209:29–37.
Gerhardt, A., 1993. Review of impact of heavy metals on streaminvertebrates with special emphasis on acid conditions. Wat. AirSoil Pollut. 66: 289–314.
Golladay, S. W., J. R. Webster, J. R. & E. F. Benfield, 1989. Changesin benthic organic matter following watershed disturbance. Hol-arct. Ecol. 12: 96–105.
Haapala, A. & T. Muotka, 1998. Seasonal dynamics of detritusand associated macroinvertebrates in channelized boreal stream.Arch. Hydrobiol. 142: 171–189.
Hildrew, A. G. & C. R. Townsend, 1976. The distribution of twopredators and their prey in an iron rich stream. J. Anim. Ecol. 45:41–57.
Holopainen, A. L. & P. Huttunen, 1998. Impact of forestry practiceson ecology of algal communities in small brooks in the Nurmesexperimental forest area, Finland. Boreal Environ. Res. 3: 63–73.
Huggenberger, P., E. Hoehn, R. Beschta & W. Woessner, 1998.Abiotic aspects of channels and floodplains in riparian ecology.Freshwatat. Biol. 40: 407–425.
Hämäläinen, H. & P. Huttunen, 1990. Estimation of acidity instreams by means of benthic invertebrates: evaluation of twomethods. In: Kauppi, P., P. Anttila & K. Kenttämies (eds),Acidification in Finland. Springer, Berlin, pp. 1051–1070.
Hämäläinen, H. & P. Huttunen, 1996. Inferring the minimum pHof streams from macroinvertebrates using weighted averagingregression and calibration. Freshw. Biol. 36: 697–709.
Hämäläinen, H. & P. Huttunen, 1998. Macroinvertebrate-inferredstream-water acidity in North Eastern Finland along a sulphurdeposition gradient. Water, Air, and Soil Pollution 104: 223–236.
Laasonen, P., T. Muotka & I. Kivijärvi, 1998. Recovery of macroin-vertebrate communities from stream habitat restoration. AquaticConserv: Mar. Freshw. Ecosyst. 8: 101–113.
Lemly, A. D. & R. H. Hilderbrand, 2000. Influence of largewoody debris on stream insect communities and benthic detritus.Hydrobiologia 421: 179–185.
Lillehammer, A., 1988. Stoneflies (Plecoptera) of Fennoscandia andDenmark. Fauna Ent. Scand. 21: 1–165.
Malmqvist, B., L. M. Nilsson & B. S. Svensson, 1978. Dynamics ofdetritus in a small stream in southern Sweden and its influenceon the distribution of the bottom animal communities. Oikos 31:3–16.
Maser, C. & J. R. Sedell, 1994. From the Forest to the Sea. TheEcology of Wood in Streams, Rivers, Estuaries, and Oceans. St.Lucie Press, U.S.A, 200 pp.
Merritt, R. W. & K. W. Cummins (eds), 1984. An Introduction to theAquatic Insects of North America. 2nd edn. Kendall/Hunt Publ.Co., Dubuque, IA, U.S.A, 722 pp.
Molles, M. C. 1982. Trichopteran communities of streams associ-ated with aspen and conifer forests: long-term structural change.Ecology 63: 1–6.
Muhar, S., S. Schmutz & M. Jungwirth, 1995. River restorationconcepts -goals and perspectives. Hydrobiologia 303: 183–194.
Murphy, J. F. & P. S. Giller, 2000. Seasonal dynamics of macroin-vertebrate assemblages in the benthos and associated with de-tritus packs in two low-order streams with different riparianvegetation. Freshwat. Biol. 43: 617–631.
Osborne, L. L. & D. A. Kovacic, 1993. Riparian vegetated buf-fer strips in water-quality restoration and stream management.Freshwat. Biol. 29: 243–258.
Perkins, J. L., 1983. Bioassay evaluation of diversity and com-munity comparison indexes. J. Wat. Pollut. Cont. Fed 55: 522–530.
Petersen L. B. M. & R. C. Petersen R. C. Jr., 1991. Short term reten-tion of channelized and natural streams. Verh. int. Ver. Limnol.24: 1756–1759.
Poff, N. L. & J. V. Ward, 1991. Drift responses of benthic inver-tebrates to experimental streamflow variation in a hydrologicallystable stream. Can. J. Fish. aquat. Sci. 48: 1926–1936.
Prochazka, K., B. A. Stewart, & R. D. Bryan, 1991. Leaf litterretention and its implications for shredder distribution in twoheadwater streams. Arch. Hydrobiol. 120: 315–325.
Raven, P. J., P. Fox, M. Everard, N. T. H. Holmes & F. H. Dawson1997. River Habitat Survey: a new system for classifying riversaccording to their habitat quality. In Boon, P. J. & D. L. How-ell (eds), Freshwater Quality: Defining the Indefinable? TheStationery Office, Edinburgh: 215–234.
Simberloff, D. S., 1978. Use of rarefaction and related methods inecology. In: Dickinson, K. L., J.Cairns & R. J. Livingston (eds),Biological Data in Water Quality Assesment: Quantitative andStatistical Analysis. Am. Soc. Testing Materials: 150–156.
Smock, L. A., G. M. Metzler & J. A. Gladden, 1989. Role of debrisdams in the structure and functioning of low-gradient headwaterstreams. Ecology 70: 764–775.
Stone, M. K. & J. B. Wallace, 1998. Long-term recovery of amountain stream from clear-cut logging: the effects of forest suc-cession on benthic invertebrate community structure. Freshwat.Biol. 39: 151–169.
Stout, B. M., E. F. Benfield & J. R. Webster, 1993. Effects of a forestdisturbance on shredder production in southern Appalachianheadwater streams. Freshwat. Biol. 29: 59–69.
Vuori, K.-M. & I. Joensuu, 1996. Impact of forest drainage on themacroinvertebrates of a small boreal headwater stream: do bufferzones protect lotic biodiversity? Biol. Conserv. 77: 87–95.
Vuori, K.-M., I. Joensuu, J. Latvala, E. Jutila & A. Ahvonen,1998. Forest drainage: a threat to benthic biodiversity of borealheadwater streams? Aquat. Conserv: Mar. Freshwat. Ecosyst. 8:745–759.
Vuori, K.-M., H. Luotonen & P. Liljaniemi, 1999. Benthic macroin-vertebrates and aquatic mosses in pristine streams of the Tolva-järvi region, Russian Karelia. Boreal Env. Res. 4: 187–200.
Vuori, K.-M. & T. Muotka, 1999. Benthic communities in humicstreams. In: Keskitalo, J. & P. Eloranta (eds), Limnology ofHumic Waters. Backhuys Publishers, Leiden, The Netherlands:193–207.
Wallace, J. B., J. R. Webster & J. L. Meyer, 1995. Influence oflog additions on physical and biotic characteristics of a mountainstream. Can. J. Fish. aquat. Sci. 52: 2120–2137.
Williams, N. E. & H. B. N. Hynes, 1973. Microdistribution andfeeding of the net-spinning caddisflies (Trichoptera) of a Cana-dian stream. Oikos 24: 73–84.
Zwick, P., 1992. Stream habitat fragmentation – a threat to biod-iversity. Biodiv. Conserv. 1: 80–97.

249
Appendix. Macroinvertebrates collected from the Koitajoki River; +++, >10%, ++, >5% and +, <5% of the total abundance of thesampling site. Functional feeding groups: gg, gathering collectors; fg, filtering collectors; sh, shredders; sc, scrapers; pr, predators
Russia Finland Russia Finland
Turbellaria pr + – Plecoptera
Nematoda pr + + Diura nanseni (Kempny) pr + +
Gastropoda Isoperla grammatica (Poda) pr + –
Gyraulus albus (Müll.) sc – + I. obscura (Zett.) pr ++ +
Lymnea sp. sc – + Taeniopteryx nebulosa (L.) sh +++ +++
Lamellibranchiata Amphinemura borealis (Mort.) sh + +
Pisidium casertanum Poli fc + + Nemoura avicularis Mort. sh + +++
P. hibernicum Westerlund fc ++ ++ N. cinerea (Retz.) sh ++ –
P. obtusale Pfeiffer fc +++ + N. flexuosa Aubert sh – +++
P. pseudosphaerium Favre fc + – Nemurella pictetii Klap. sh +++ +
P. supinum A. Schmidt fc – + Protonemura meyeri (Pict.) sh +++ ++
Pisidium spp. fc ++ +++ Capnopsis schilleri (Rost.) sh + +
Oligochaeta Xanthoperla apicalis Newman sh + +
Aelosoma sp. gc – + Leuctra digitata Kempny sh +++ +++
Eiseniella tetraedra (Savigny) gc – + L. nigra (Oliv.) sh – +
Stylodrilus heringianus Clap. gc + + Odonata
Lumbriculidae gc + + Calopteryx virgo (L.) pr + +
Chaetogaster diaphanus Platycnemis pennipes (Pallas) pr – +
(Gruithuisen) gc + + Cordulegaster boltonii (Donovan) pr + +
Arcteonais lomondi (Mart.) gc – + Somatochlora metallica (Lind.) pr + +
Nais alpina Sperber gc + + Heteroptera
N. communis Piq. gc – + Aphelocheirus aestivalis (Fabr.) pr – +
Ripistes parasita (O. Schmidt) gc – + Coleoptera
Slavina appendiculata (Ud.) gc + + Gyrinidae pr – +
Stylaria lacustris (L.) gc – + Agabus sp. pr + –
Vejdovskyella comata (Vej.) gc + + Hydroporus obscurus Sturm pr + –
Spirosperma ferox (Eisen) gc + + Macroplea (?) sp. sh + –
Tubifex tubifex (Müll.) gc + + Hydrophilidae pr + +
Enchytraeidae gc + + Elmis aenea (Müll.) sc +++ ++
Hirudinea Oulimnius tuberculatus (Müll.) sc +++ +++
Glossiphonia complanata (L.) pr + + Limnius volckmari (Panzer) sc ++ +
Helobdella stagnalis (L.) pr + – Megaloptera
Erpobdella octoculata (L.) pr + + Sialis lutaria (L.) pr + –
E. testacea (Savigny) pr – + S. fuliginosa Pict. pr + +
Isopoda S. sordida Klingst. pr – +
Asellus aquaticus (L.) sh + – Lepidoptera
Hydracarina pr + + Cataclysta lemnata (L.) sh – +
Ephemeroptera Trichoptera
Centroptilum sp. gc + – Arctopsyche ladogensis (Kolenati) fc + –
Baetis niger (L.) gc +++ + Hydropsyche angustipennis (Curt.) fc ++ –
B. rhodani (Pict.) gc + – H. pellucidula (Curt.) fc +++ ++
B. subalpinus Bgtss. gc + – H. silfvenii (Ulmer) fc – +
B. vernus Curt. gc + – H. siltalai (Döhl. fc +++ +++
Heptagenia fuscogrisea (Retz.) sc + ++ Neureclipsis bimaculata (L.) fc +++ +
H. sulphurea (Müll.) sc + + Plectrocnemia conspersa (Curt.) pr + +
Leptophlebia marginata (L.) gc +++ +++ Polycentropus
Ephemerella ignita flavomaculatus (Pict.) pr +++ +++
(lactataform) (Poda) gc + – P. irrotatus (Curt.) pr + +
Ephemerella mucronata (Bgtss.) gc ++ + Lype reducta (Hagen) sc + –
Continued on p. 250

250
Appendix. Continued
Russia Finland Russia Finland
Ithytrichia lamellaris Eaton sc + + Prodiamesa olivacea (Mg.) gc – +
Oxyethira tristella Klap. gc + + Brillia longifurca K. sh + –
Rhyacophila fasciata Hagen pr + + Chaetocladius piger gr. gc + +
R. nubila (Zett.) pr + + C. vitellinus gr. gc – +
Rhyacophila sp. pr + + Corynoneura coronata Edw. gc + +
Agapetus ochripes Curt. sc + + C. lobata Edw. gc + –
Chaetopteryx villosa (Fabr.) sh + – Cricotopus (Cricotopus)
Glyphotaelius pellucidus (Retz.) sh – + pulchripes Verr. sh – +
Limnephilus subcentralis Brauer sh – + C. (Isocladius) laricomalis gr. sh ++ +++
Limnephilus sp. sh + + C. (Nostococladius) sp. sh – +
Limnephilidae sh + ++ Diplocladius cultriger K. gc – +
Potamophylax nigricornis (Pict.) sh + – Eukiefferiella claripennis gr. gc ++ +
Potamophylax sp. sh + + E. devonica gr. gc + +++
Brachycentrus subnubilus Curt. fc + + Eukiefferiella sp. gc – +
Micrasema gelidum McLach. sh ++ +++ Heterotanytarsus apicalis (K.) gc + +
Lepidostoma hirtum (Fabr.) sh ++ +++ Heterotrissocladius marcidus gr. gc ++ +
Agrypnia varia (Fabr.) pr – + Metriocnemus sp. gc + –
Oligostomis reticulata (L.) pr + – Nanocladius (Nanocladius)
Semblis atrata (Gmelin) pr + – bicolor gr. gc + +
Molanna angustata Curt. sc – + N. (Nanocladius) rectinervis gr. gc – +
Molannodes tinctus (Zett.) gc – + Orthocladius (Orthocladius) spp. gc +++ +
Molannidae sc – + Paralimnophyes sp. gc + +
Sericostoma personatum (Spence) sh – + Psectrocladius
Oecetis testacea (Curt.) pr + + (Monopsectrocladius) sp. gc + –
O. notata (Rambur) pr + – P. (Psectrocladius) psilopterus gr. gc + +
Athripsodes cinereus (Curt.) gc + + P. (Psectrocladius) sordidellus gr. gc + +
A. commutatus (Rostock) gc + – Psectrocladius sp. gc – +
Ceraclea senilis (Burmeister) gc – + Rheocricotopus fuscipes (K.) gc ++ +
Ceraclea sp. gc + – Rheocricotopus sp. gc – +
Mystacides azurea (L.) gc + + Symposiocladius lignicola (K.) sh + –
Triaenodes sp. gc + + Thienemanniella clavicornis (K.) gc + +
Diptera Thienemanniella (species A) sp. gc – +
Tipulidae sh + + Tvetenia discoloripes (G.) gc – +
Pediciidae T. paucunca (Sæther) gc +++ +++
Dicranota spp. pr +++ + Tvetenia (species A) sp. gc ++ +
Limoniidae sh + + Orthocladiinaegc – +
Tabanidae pr + – Chironomus sp. gc – +
Simuliidae fc +++ ++ Cryptochironomus defectus gr. pr + +
Chironomidae Demicryptochironomus
Ablabesmyia longistyla Fitt. pr + ++ vulneratus (Zett.) gc – +
A. monilis (L.) pr – + Microtendipes pedellus (Geer) gc + +
Conchapelopia spp. pr +++ +++ Pagastiella orophila (Edw.) gc – +
Helopelopia spp. pr + + Parachironomus arcuatus gr. pr + +
Macropelopia nebulosa gr. pr + – P. frequens gr pr – +
M. notata gr. pr + + Paracladopelma doris gr. gc – +
Natarsia spp. pr + + Phaenopsectra flavipes (Mg.) sc + +
Nilotanypus (?) spp. pr – + Phaenopsectra sp. sc – +
Procladius (Holotanypus) sp. pr + + Polypedilum gr. bicrenatum sh – +
Rheopelopia spp. pr + – P. laetum agg. sh – +
Zavrelimyia spp. pr ++ + P. nubeculosum gr. sh – +
Tanypodinae pr + – P. pedestre agg. sh – +
Potthastia longimana (K.) gc + + P. pullum gr. sh + ++
Continued on p. 251

251
Appendix. Continued
Russia Finland
Polypedilum sp. sh – +
Robackia demeijerei (Kruseman) gc + +
Sergentia coracina (Zett.) sc – +
Stenochironomus sp. sh + +
Stictochironomus
rosenschoeldi (Zett.) gc - +
Xenochironomus xenolabis (K.) pr – +
Cladotanytarsus vanderwulpi gr. gc + +
Cladotanytarsus sp. gc – +
Constempellina brevicosta (Edw.) gc – +
Micropsectra spp. gc +++ ++
Rheotanytarsus spp. fc + ++
Stempellina bausei (K.) gc – +
Stempellinella brevis (Edw.) gc + +
S. minor (Edw.) gc – +
Tanytarsus chinyensis gr. fc +++ +++
Tanytarsus spp. fc + +
Ceratopogonidae
Bezzia spp. pr + +
Palpomyia spp. pr + –
Empididae pr + +
Muscidae pr + +
Psychodidae gc – +
Rhagionidae
Atherix ibis (Fabr.) pr + +




















