
Growth and ethanol fermentation ability on hexose and pentose sugars and glucose effect under...
14
BIOENERGY AND BIOFUELS Growth and ethanol fermentation ability on hexose and pentose sugars and glucose effect under various conditions in thermotolerant yeast Kluyveromyces marxianus Nadchanok Rodrussamee & Noppon Lertwattanasakul & Katsushi Hirata & Suprayogi & Savitree Limtong & Tomoyuki Kosaka & Mamoru Yamada Received: 31 December 2010 / Revised: 13 February 2011 /Accepted: 24 February 2011 /Published online: 8 April 2011 # Springer-Verlag 2011 Abstract Ethanol fermentation ability of the thermotolerant yeast Kluyveromyces marxianus, which is able to utilize various sugars including glucose, mannose, galactose, xylose, and arabinose, was examined under shaking and static conditions at high temperatures. The yeast was found to produce ethanol from all of these sugars except for arabinose under a shaking condition but only from hexose sugars under a static condition. Growth and sugar utilization rate under a static condition were slower than those under a shaking condition, but maximum ethanol yield was slightly higher. Even at 40°C, a level of ethanol production similar to that at 30°C was observed except for galactose under a static condition. Glucose repression on utilization of other sugars was observed, and it was more evident at elevated temper- atures. Consistent results were obtained by the addition of 2-deoxyglucose. The glucose effect was further examined at a transcription level, and it was found that KmGAL1 for galactokinase and KmXYL1 for xylose reductase for galac- tose and xylose/arabinose utilization, respectively, were repressed by glucose at low and high temperatures, but KmHXK2 for hexokinase was not repressed. We discuss the possible mechanism of glucose repression and the potential for utilization of K. marxianus in high-temperature fermen- tation with mixed sugars containing glucose. Keywords Kluyveromyces marxianus . Ethanol production . Glucose effect . High-temperature fermentation Introduction Kluyveromyces marxianus is a close relative of Kluyveromy- ces lactis, a model Crabtree-negative yeast that has been extensively investigated (Gonzalez-Siso et al. 2000, 2009; Schaffrath and Breunig 2000; van Ooyen et al. 2006). The major common feature of K. lactis and K. marxianus is the capacity to assimilate lactose as a carbon source, a feature that is absent in Saccharomyces cerevisiae. In contrast to K. lactis, there is no complete genome sequence, no type strain adopted as a reference for basic research purposes, and little accumulation of information for K. marxianus, though some heterologous protein production systems in the yeast become available (Lane and Morrissey 2010). Nevertheless, K. marxianus has a number of advantages over K. lactis or S. cerevisiae, including a broad substrate spectrum, thermotol- erance, and high growth rate (Fonseca et al. 2008), and is thus an alternative to S. cerevisiae as an ethanol producer. Because of the increasing concern regarding petroleum costs and global warming, worldwide has been shown the production of bioethanol as a renewable energy source, especially from renewable resources (Hahn-Hagerdal et al. 2006). Nowadays, the most commonly utilized renewable fuel is ethanol mainly from sugar cane and starch, and thus, the consumption of these biomasses has dramatically N. Rodrussamee : K. Hirata : Suprayogi : M. Yamada Graduate School of Medicine, Yamaguchi University, 1-1-1 Minami-Kogushi, Ube, Yamaguchi 755-8505, Japan N. Lertwattanasakul : T. Kosaka : M. Yamada (*) Faculty of Agriculture, Yamaguchi University, 1677-1 Yoshida, Yamaguchi-shi, Yamaguchi 753-8515, Japan e-mail: [email protected] S. Limtong Faculty of Science, Kasetsart University, 50 Paholyothin Road, Bangkok 10900, Thailand Appl Microbiol Biotechnol (2011) 90:1573–1586 DOI 10.1007/s00253-011-3218-2
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Growth and ethanol fermentation ability on hexose and pentose sugars and glucose effect under...

BIOENERGY AND BIOFUELS
Growth and ethanol fermentation ability on hexoseand pentose sugars and glucose effect under variousconditions in thermotolerant yeast Kluyveromyces marxianus
Nadchanok Rodrussamee & Noppon Lertwattanasakul &Katsushi Hirata & Suprayogi & Savitree Limtong &
Tomoyuki Kosaka & Mamoru Yamada
Received: 31 December 2010 /Revised: 13 February 2011 /Accepted: 24 February 2011 /Published online: 8 April 2011# Springer-Verlag 2011
Abstract Ethanol fermentation ability of the thermotolerantyeast Kluyveromyces marxianus, which is able to utilizevarious sugars including glucose, mannose, galactose,xylose, and arabinose, was examined under shaking andstatic conditions at high temperatures. The yeast was foundto produce ethanol from all of these sugars except forarabinose under a shaking condition but only from hexosesugars under a static condition. Growth and sugar utilizationrate under a static condition were slower than those under ashaking condition, but maximum ethanol yield was slightlyhigher. Even at 40°C, a level of ethanol production similar tothat at 30°C was observed except for galactose under a staticcondition. Glucose repression on utilization of other sugarswas observed, and it was more evident at elevated temper-atures. Consistent results were obtained by the addition of2-deoxyglucose. The glucose effect was further examined ata transcription level, and it was found that KmGAL1 forgalactokinase and KmXYL1 for xylose reductase for galac-tose and xylose/arabinose utilization, respectively, wererepressed by glucose at low and high temperatures, but
KmHXK2 for hexokinase was not repressed. We discuss thepossible mechanism of glucose repression and the potentialfor utilization of K. marxianus in high-temperature fermen-tation with mixed sugars containing glucose.
Keywords Kluyveromyces marxianus . Ethanolproduction . Glucose effect . High-temperature fermentation
Introduction
Kluyveromyces marxianus is a close relative of Kluyveromy-ces lactis, a model Crabtree-negative yeast that has beenextensively investigated (Gonzalez-Siso et al. 2000, 2009;Schaffrath and Breunig 2000; van Ooyen et al. 2006). Themajor common feature of K. lactis and K. marxianus is thecapacity to assimilate lactose as a carbon source, a featurethat is absent in Saccharomyces cerevisiae. In contrast to K.lactis, there is no complete genome sequence, no type strainadopted as a reference for basic research purposes, and littleaccumulation of information for K. marxianus, though someheterologous protein production systems in the yeast becomeavailable (Lane and Morrissey 2010). Nevertheless, K.marxianus has a number of advantages over K. lactis or S.cerevisiae, including a broad substrate spectrum, thermotol-erance, and high growth rate (Fonseca et al. 2008), and isthus an alternative to S. cerevisiae as an ethanol producer.
Because of the increasing concern regarding petroleumcosts and global warming, worldwide has been shown theproduction of bioethanol as a renewable energy source,especially from renewable resources (Hahn-Hagerdal et al.2006). Nowadays, the most commonly utilized renewablefuel is ethanol mainly from sugar cane and starch, and thus,the consumption of these biomasses has dramatically
N. Rodrussamee :K. Hirata : Suprayogi :M. YamadaGraduate School of Medicine, Yamaguchi University,1-1-1 Minami-Kogushi,Ube, Yamaguchi 755-8505, Japan
N. Lertwattanasakul : T. Kosaka :M. Yamada (*)Faculty of Agriculture, Yamaguchi University,1677-1 Yoshida,Yamaguchi-shi, Yamaguchi 753-8515, Japane-mail: [email protected]
S. LimtongFaculty of Science, Kasetsart University,50 Paholyothin Road,Bangkok 10900, Thailand
Appl Microbiol Biotechnol (2011) 90:1573–1586DOI 10.1007/s00253-011-3218-2

increased. The utilization of corn starch for bioethanolproduction may cause a serious problem in competition withcorn starch as food. In the near future, it is expected that asource for low-cost ethanol production will be lignocellulosicbiomass such as agricultural and forestry residues. Lignocel-lulosic materials are composed of three main polymers:cellulose (~47% of dry weight), hemicelluloses (~30% ofdry weight), and lignin (~25% of dry weight; Aristidou andPenttila 2000). Cellulose is a homopolymer of glucose (Glc),while hemicellulose is composed of hexose sugars—glucose,mannose (Man), and galactose (Gal)—and pentose sugars—xylose (Xyl) and arabinose (Ara; Saha 2003). Hemicelluloseis easily hydrolyzed to its constituent monosaccharidescompared to cellulose (Aristidou and Penttila 2000; Perezet al. 2002; Zaldivar et al. 2001).
Considering sugar constituents in hemicellulose, thedevelopment of microbes that can utilize and convert a varietyof sugars into ethanol is required. Besides, thermotolerantmicrobes applicable for high-temperature fermentation thatefficiently produce ethanol at elevated temperatures areexpected to have a potential for reducing cooling costs (Banatet al. 1998). High-temperature fermentation also has advan-tages of efficient simultaneous saccharification and fermen-tation, a continuous shift from fermentation to distillation,reducing risk of contamination, and suitability for applica-tion in tropical countries (Anderson et al. 1986; Banat et al.1998; Limtong et al. 2007). Additionally, glucose effect thatreduces synthesis of enzymes for catabolism of alternativecarbon sources should be considered. A significant reductionof a large number of cellular functions as the glucose effecthas been reported in S. cerevisiae (Gancedo 1998). Theregulatory mechanism for D-galactose utilization is at leastpartially conserved between K. lactis and S. cerevisiae,though the molecular interactions between regulators aredifferent (Venkat et al. 2010; Rubio-Texeira 2005). Severalglucose-repressed genes are known to be controlled byMig1p, a transcription factor involved in glucose repression,directly or indirectly via repression of a transcriptionactivator (Gancedo 1998). On the other hand, the regulationof genes for a D-xylose-inducible pathway, including anNAD(P)H-linked xylose reductase (Xyl1p) and an NAD-linked xylitol dehydrogenase (Xyl2p), and for anL-arabinose-inducible pathway partially overlapping withthe D-xylose-inducible pathway (Fig. 1) has not beenelucidated in yeast.
K. marxianus DMKU3-1042 is a microorganism thatpossesses the capability for assimilation of many differentkinds of sugars at high temperatures (Nonklang et al. 2008).Nevertheless, there are no reports on glucose repression at ahigh temperature in K. marxianus.
In this study, the potential of K. marxianus DMKU3-1042 for utilization of and ethanol production from sugarspresent in hemicellulose hydrolysate was for the first time
examined under shaking and static conditions at hightemperatures. This strain can grow and produce ethanol ata high temperature (Limtong et al. 2007) and is the mostthermotolerant among strains available (Abdel-Banat et al.2010; Nonklang et al. 2008). We therefore consider thisstrain to be a suitable candidate for high-temperatureethanol fermentation. Glucose repression on assimilationof other sugars was also tested not only by the addition ofglucose but also by the addition of 2-deoxyglucose(2-DOG). To further consider the repression, the expressionof genes responsible for utilization of mannose, galactose,xylose, or arabinose was analyzed. This is the first study toreveal that the extent of glucose repression is different atdifferent temperatures and on different sugars in K. marx-ianus. This study also provides valuable information forapplication of K. marxianus and indicates its usefulness forhigh-temperature ethanol fermentation.
Materials and methods
Strain and media
The yeast strain used in this study was K. marxianusDMKU3-1042 strain, one of the isolates from soil andwater samples obtained around sugar cane plantations andsugar factories in Thailand (Limtong et al. 2007). Culturewas carried out in YPD medium (10 g/l yeast extract, 20g/l peptone, and 20 g/l glucose) as used for preparation ofthe inoculum. Different carbon sources were used, singly orin combination with 20 g/l of glucose. For investigating theutilization of an individual sugar, YP medium (10 g/l yeastextract and 20 g/l peptone) supplemented with 20 g/l ofGlc, Man, Gal, Xyl, or Ara was used. These media weredesignated as YPD, YPMan, YPGal, YPXyl, and YPAra,respectively. The YP medium with 20 g/l glucose incombination with 20 g/l of Man, Gal, Xyl, or Ara wasused for experiments with mixed sugars that were namedYPDMan, YPDGal, YPDXyl, and YPDAra, respectively.To examine the effect of antimycin A, 5 μM antimycin Awas added.
Cultivation conditions
For the inoculum preparation, the yeast strain was cultivat-ed in 20 ml of YPD medium in a 100-ml Erlenmeyer flaskat 30°Cwith orbital shaking at 160 rpm for 18 h. The preculturewas inoculated into a 300-ml Erlenmeyer flask containing100 ml fresh medium of YP containing Glc, Man, Gal, Xyl, orAra or containing Glc combined with others sugars at initialOD660 values of 0.1 and 1 for shaking and static conditions,respectively. Incubation conditions for the shaking conditionwere standardized on the rotary shaker with 160 rpm at 30°C,
1574 Appl Microbiol Biotechnol (2011) 90:1573–1586

40°C, and 45°C, whereas for the static condition, flasks wereconstantly stood at 30°C and 45°C.
Analytical methods
Cell density was measured turbidimetrically at 660 nm. Todetermine sugar and ethanol concentrations in culturemedia, cultures were sampled and subjected to a low-speed centrifugation. The supernatant was frozen and keptat −20°C until analyzed. Quantitative analysis of sugar andethanol was performed by high-performance liquid chro-matography (Hitachi, Japan). A Gel pack column GL-C610-S (Hitachi, Japan) was used together with a refractiveindex detector (Model L-2490, Hitachi) at 60°C with0.3 ml/min eluent of deionized water.
Analysis of glucose repression by using glucose analog2-DOG
Cells grown in YPD medium for 18 h were collected. Afterwashing the cells with deionized water, the suspended cells (l×107 cells/ml) were 10-fold sequentially diluted and then
spotted onto agar plates of YPMan, YPGal, YPXyl, andYPAra with or without 0.01% 2-DOG. YPD plates were usedas a control. These plates were incubated at 30°C, 40°C, and45°C for 48 h.
RT-PCR analysis
Total RNA from cells, which had been grown in variousmedia as described below, was isolated by the hot phenolmethod (Aiba et al. 1981). Isolated RNA was treated withRNase-free DNase I (Qiagen, Japan) for 15 min, and thenDNase I was inactivated at 75°C for 10 min. For eachexperimental condition, the three genes KmHXK2,KmGAL1, and KmXYL1 for hexokinase, galactokinase,and xylose reductase, respectively, were used to determinethe effect of glucose repression. KmACT1 for actin was usedas an internal control. Cells were first grown in YPD mediumuntil the exponential phase at 30°C, washed two times andresuspended with YP medium, and subsequently inoculated at5% into YPD, YPMan, YPGal, YPXyl, or YPAra for a singlesugar condition or into YPDMan, YPDGal, YPDXyl, orYPDAra for a mixed sugars condition and further incubated
ATP GUT
G3P
1,3DPG
ZWF NADPH
NADP+ CO2
G6P
Rul5P
NADH
NAD+
TDH
PDC
GPD
NADH NAD+
PYR
PYK PEP
ENO
GPM
PGK
TPI
PFK FBP
F6P
PGI
6PG
ADH
D-glucose
FDP
DHAP
3PG
2PG
ATP
ATP
ATP
Acetaldehyde EtOH
Glycerol 3P
Glycerol
GPP
FBA
CO2
D-xylose
Xu5P
RI5P
G3P
S7P
E4P
NADPH NADP+
XYL3 REP RKI
TKL
TKL
TAL
NADH NAD+
GND
NAD(P)H
NAD(P)+
ATP
Xylulose
XYL2
Xylitol NAD+
NADH
ATP
HXK2
Mannnose-6-P
HXK2
ATP
XYL1
L-arabinose
L-arabinitol
XYL1
galactose-1-P
glucose-1-P
ATP
UDP-glucose
UDP-galactose
GAL1 (Leloir pathway)
GAL7 GAL10
GAL5
Pentose phosphate pathway
PMI
NAD(P)H
NAD(P)+
NAD+
NADH NAD(P)H
NAD(P)+
b a c
D-mannose D-galactose
L-xylulose
Fig. 1 Schematic diagram representing sugar metabolisms to ethanol inyeast and selected genes, GAL1, HXK2, and XYL1 for expressionanalysis. Three genes were involved in galactose, mannose/glucose, andxylose/arabinose catabolic pathways, respectively. a Galactose wasconverted to galactose-1-phosphate by galactokinase encoded by GAL1and then subsequently converted to glucose-6-phosphate via the Leloirpathway, which consists of four catalytic steps. b After glucose ormannose phosphorylation by hexokinase encoded by HXK2, glucosewas converted to be glucose-6-phosphate, an intermediate in glycolysis,whereas mannose was converted to be mannose-6-phosphate and then
was isomerized to fructose-6-phosphate by phosphomannose isomerase.c D-Xylose and L-arabinose metabolisms in yeast share a partial overlapbetween their catabolic pathways. Xylose metabolism proceeds mostlyvia two oxidative–reductive reactions. Xylose is firstly reduced toxylitol by an NAD(P)H-linked xylose reductase encoded by XYL1,followed by xylitol oxidation to xylulose by an NAD+-linked xylitoldehydrogenase encoded by XYL2. Arabinose is also converted toL-arabinitol by XYL1. This metabolism pathway is quite similar to thatof the xylose metabolism and consists of two NAD+-linked oxidativereactions and two NAD(P)H-linked reductive reactions
Appl Microbiol Biotechnol (2011) 90:1573–1586 1575

for 6 h (mid-exponential phase) at 30°C or 45°C. Theconcentration of RNA was estimated spectrophotometricallyat 260 nm. RT-PCR analysis was performed using an mRNASelective RT-PCR kit (Takara, Japan) with 0.1 μg of total RNAas a template and the primer sets for RT-PCR (Table 1). Fordetection of an internal control, 0.01 μg of total RNA wasused as a template. RT-PCR was performed on a Takara PCRthermal cycler MP (Takara Biomedicals, Japan). After RTreaction had been performed at 40°C for 15 min, PCRconsisting of denaturing at 82°C for 1 min, annealing for1 min at a fixed temperature, 5° lower than Tm, which wascalculated by the rule of thumb method, and extension at72°C for 1 min was carried out using two primers for eachgene. The PCR products after 20, 25, 30, and 35 cycles or 20,23, 26, 29, and 32 cycles for each gene were analyzed by0.9% agarose gel electrophoresis. Intensity of bands of RT-PCR products was quantitatively determined using the UN-SCAN-IT gel™ automated digitizing system (Silk Scientific,USA). Under our conditions, the RNA-selective RT-PCR wasable to specifically detect mRNA because no band wasobserved when reverse transcriptase was omitted.
Results
Cell growth and ethanol production in YPmedium containinga single sugar under a shaking conditionat different temperatures
To examine the potential for utilization of sugars found inhemicellulose hydrolysate in K. marxianus DMKU3-1042,cells were grown in a medium containing Glc, Man, Gal,Xyl, or Ara under a shaking condition at different temper-atures, and cell growth and concentrations of sugar andethanol in the medium were monitored (Fig. 2; Table 2). Cellgrowth was observed as sugar reduced on all sugars tested at30°C, 40°C, and 45°C, indicating that the strain can usethese sugars for growth at high temperatures. In YPD,YPMan, and YPGal, ethanol was increased roughly in anti-parallel with decrease in sugar at the beginning ofcultivation. The growth was gradually decreased withincrease in temperature, but ethanol accumulation was nearly
the same except for a delay at 45°C. In YPXyl, ethanol andxylitol were accumulated. No ethanol accumulation, however,was detected in YPAra. YPXyl and YPAra showed a longerlag phase and a lower maximum growth rates.
At 30°C, Glc, Man, and Gal were completely consumedat 12 h, while Xyl and Ara utilization was completed at 72and 96 h, respectively. The maximum ethanol yield was thehighest in YPD and YPGal followed by that in YPMan. InYPXyl, the largest amount of ethanol production was 2.5g/l at 72 h, and xylitol production was 4.3 g/l at 48 h. Themaximum ethanol yield in YPXyl was 3.6 times lower thanthose in YPD and YPGal. The maximum sugar utilizationrate and the maximum growth rate were the highest inYPGal, and the specific growth rate and the specific sugarutilization rate at 6 h were the highest in YPD.
At 40°C, Glc, Man, and Gal utilization was nearlycompleted at 12 h, whereas Xyl was completely consumedwithin 72 h. Ara was not completely utilized within 96 h.The amount of ethanol production was the largest at 12 h inYPD, YPMan, and YPGal. In YPXyl, xylitol accumulationwas 7.0 g/l, which was higher than the amounts at 30°C and45°C. The maximum ethanol yield was the highest inYPGal, which was higher than those at 30°C and 45°C. Thespecific growth and sugar utilization rates at 6 h wereslightly lower than those at 30°C in YPMan and YPGal.
At 45°C, Glc and Man were completely utilized at 24 h,whereas Gal was completely utilized at 48 h. Xyl and Arawere not completely utilized within 96 h. Interestingly, theethanol level in YPGal was stable until the end offermentation, whereas the level in other media at alltemperatures was gradually reduced with incubation. Thespecific growth rate and sugar utilization speed were greatlyreduced in YPD, YPMan, and YPGal compared to those at30°C or 40°C.
Cell growth and ethanol production in YP mediumcontaining mixed sugars with Glc under a shakingcondition at different temperatures
There have been a number of extensive studies on glucoserepression in S. cerevisiae and K. lactis, which were mainlyfocused on sucrose, lactose, or galactose utilization (Venkat
Name Gene Sequence 5′→3′ Length (bp)
RT-KmHXK2-F KmHXK2 AAGAAGCCACCAGCCAGA 542RT-KmHXK2-R ACATCGTGGCCTTCGACA
RT-KmGAL1-F KmGAL1 AGGTCGCCAGGTAGAGTG 529RT-KmGAL1-R CGTCCTTACCGCAGATAG
RT-KmXYL1-F KmXYL1 CTCGCACCAACAGTTACC 571RT-KmXYL1-R AGGCGACTGGCTTGATAC
RT-KmACT1-F KmACT1 ACGTTGTTCCAATCTACGCC 491RT-KmACT1-R AGAAGATGGAGCCAAAGCAG
Table 1 Primers used for RT-PCR in this study
1576 Appl Microbiol Biotechnol (2011) 90:1573–1586
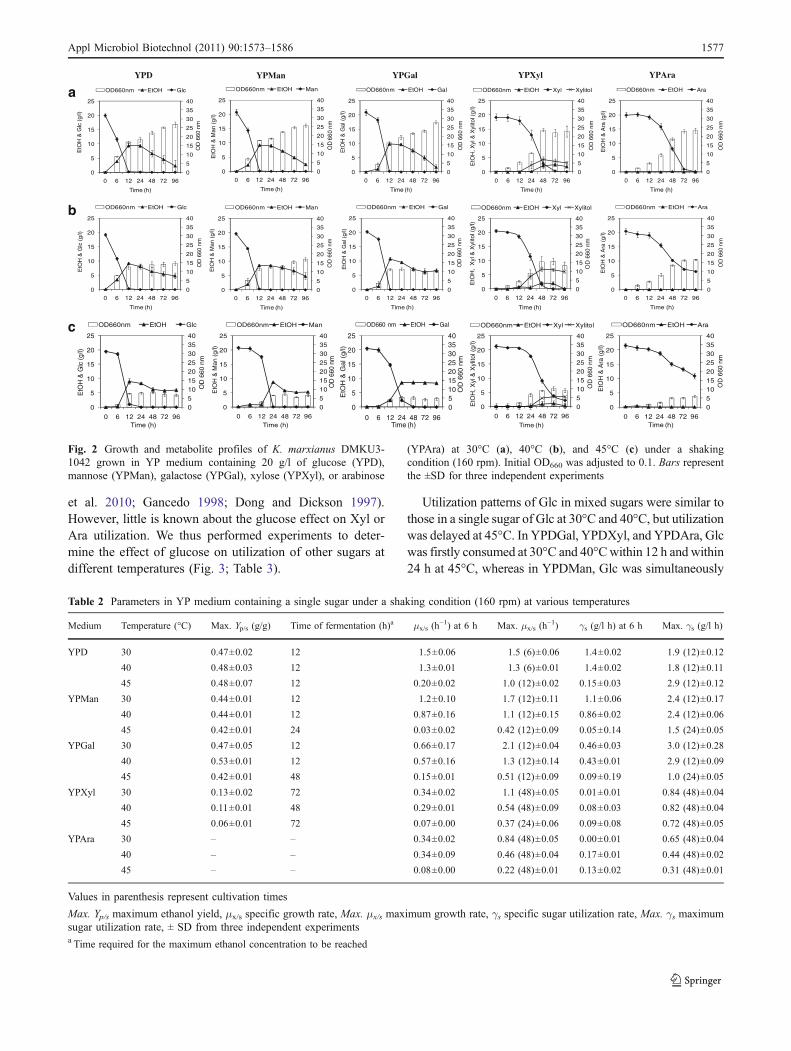
et al. 2010; Gancedo 1998; Dong and Dickson 1997).However, little is known about the glucose effect on Xyl orAra utilization. We thus performed experiments to deter-mine the effect of glucose on utilization of other sugars atdifferent temperatures (Fig. 3; Table 3).
Utilization patterns of Glc in mixed sugars were similar tothose in a single sugar of Glc at 30°C and 40°C, but utilizationwas delayed at 45°C. In YPDGal, YPDXyl, and YPDAra, Glcwas firstly consumed at 30°C and 40°Cwithin 12 h and within24 h at 45°C, whereas in YPDMan, Glc was simultaneously
a
c
YPD YPAra YPXyl YPMan YPGal
b
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Glc
(g/
l)
Time (h)
OD660nm EtOH Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Gal
(g/
l)
Time (h)
OD660nm EtOH Gal
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Man
(g/
l)
Time (h)
OD660nm EtOH Man
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, X
yl &
Xyl
itol (
g/l)
Time (h)
OD660nm EtOH Xyl Xylitol
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Glc
(g/
l)
Time (h)
OD660nm EtOH Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Man
(g/
l)
Time (h)
OD660nm EtOH Man
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Gal
(g/
l)
Time (h)
OD660nm EtOH Gal
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H,
Xyl
& X
ylito
l (g/
l)
Time (h)
OD660nm EtOH Xyl Xylitol
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Glc
(g/
l)
Time (h)
OD660nm EtOH Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Man
(g/
l)
Time (h)
OD660nm EtOH Man
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Gal
(g/
l)
Time (h)
OD660 nm EtOH Gal
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, X
yl &
Xyl
itol (
g/l)
Time (h)
OD660nm EtOH Xyl Xylitol
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara
Fig. 2 Growth and metabolite profiles of K. marxianus DMKU3-1042 grown in YP medium containing 20 g/l of glucose (YPD),mannose (YPMan), galactose (YPGal), xylose (YPXyl), or arabinose
(YPAra) at 30°C (a), 40°C (b), and 45°C (c) under a shakingcondition (160 rpm). Initial OD660 was adjusted to 0.1. Bars representthe ±SD for three independent experiments
Table 2 Parameters in YP medium containing a single sugar under a shaking condition (160 rpm) at various temperatures
Medium Temperature (°C) Max. Yp/s (g/g) Time of fermentation (h)a μx/s (h−1) at 6 h Max. μx/s (h
−1) γs (g/l h) at 6 h Max. γs (g/l h)
YPD 30 0.47±0.02 12 1.5±0.06 1.5 (6)±0.06 1.4±0.02 1.9 (12)±0.12
40 0.48±0.03 12 1.3±0.01 1.3 (6)±0.01 1.4±0.02 1.8 (12)±0.11
45 0.48±0.07 12 0.20±0.02 1.0 (12)±0.02 0.15±0.03 2.9 (12)±0.12
YPMan 30 0.44±0.01 12 1.2±0.10 1.7 (12)±0.11 1.1±0.06 2.4 (12)±0.17
40 0.44±0.01 12 0.87±0.16 1.1 (12)±0.15 0.86±0.02 2.4 (12)±0.06
45 0.42±0.01 24 0.03±0.02 0.42 (12)±0.09 0.05±0.14 1.5 (24)±0.05
YPGal 30 0.47±0.05 12 0.66±0.17 2.1 (12)±0.04 0.46±0.03 3.0 (12)±0.28
40 0.53±0.01 12 0.57±0.16 1.3 (12)±0.14 0.43±0.01 2.9 (12)±0.09
45 0.42±0.01 48 0.15±0.01 0.51 (12)±0.09 0.09±0.19 1.0 (24)±0.05
YPXyl 30 0.13±0.02 72 0.34±0.02 1.1 (48)±0.05 0.01±0.01 0.84 (48)±0.04
40 0.11±0.01 48 0.29±0.01 0.54 (48)±0.09 0.08±0.03 0.82 (48)±0.04
45 0.06±0.01 72 0.07±0.00 0.37 (24)±0.06 0.09±0.08 0.72 (48)±0.05
YPAra 30 – – 0.34±0.02 0.84 (48)±0.05 0.00±0.01 0.65 (48)±0.04
40 – – 0.34±0.09 0.46 (48)±0.04 0.17±0.01 0.44 (48)±0.02
45 – – 0.08±0.00 0.22 (48)±0.01 0.13±0.02 0.31 (48)±0.01
Values in parenthesis represent cultivation times
Max. Yp/s maximum ethanol yield, μx/s specific growth rate, Max. μx/s maximum growth rate, γs specific sugar utilization rate, Max. γs maximumsugar utilization rate, ± SD from three independent experimentsa Time required for the maximum ethanol concentration to be reached
Appl Microbiol Biotechnol (2011) 90:1573–1586 1577
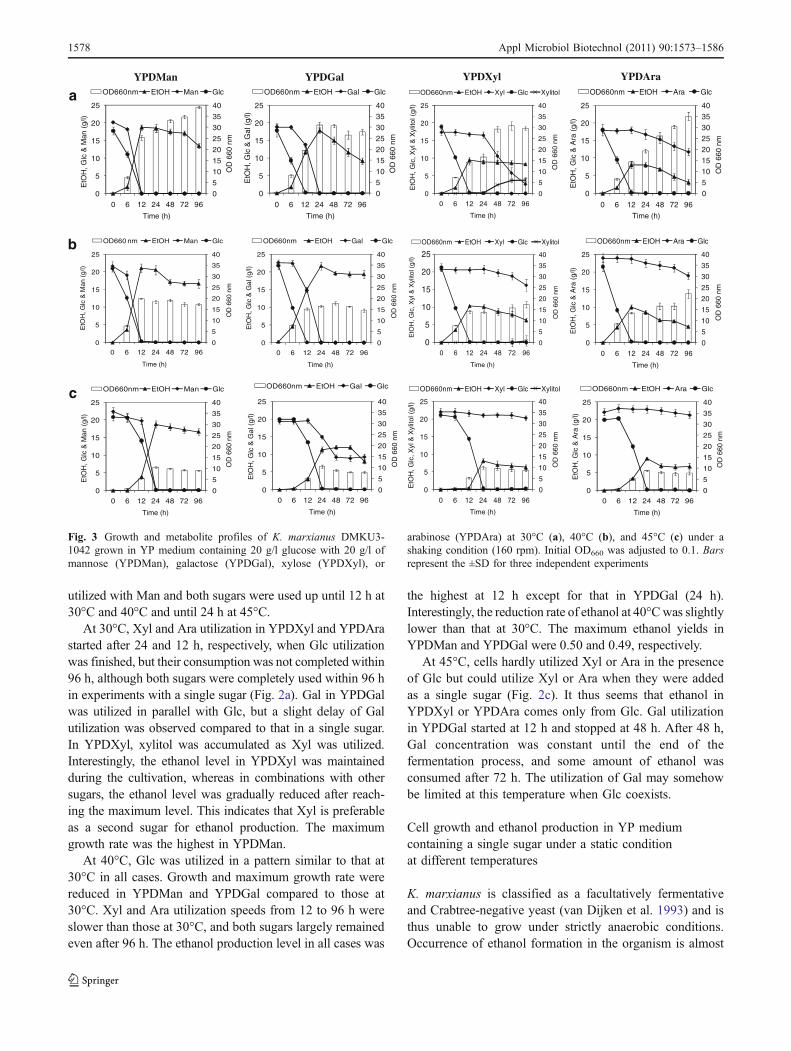
utilized with Man and both sugars were used up until 12 h at30°C and 40°C and until 24 h at 45°C.
At 30°C, Xyl and Ara utilization in YPDXyl and YPDArastarted after 24 and 12 h, respectively, when Glc utilizationwas finished, but their consumption was not completed within96 h, although both sugars were completely used within 96 hin experiments with a single sugar (Fig. 2a). Gal in YPDGalwas utilized in parallel with Glc, but a slight delay of Galutilization was observed compared to that in a single sugar.In YPDXyl, xylitol was accumulated as Xyl was utilized.Interestingly, the ethanol level in YPDXyl was maintainedduring the cultivation, whereas in combinations with othersugars, the ethanol level was gradually reduced after reach-ing the maximum level. This indicates that Xyl is preferableas a second sugar for ethanol production. The maximumgrowth rate was the highest in YPDMan.
At 40°C, Glc was utilized in a pattern similar to that at30°C in all cases. Growth and maximum growth rate werereduced in YPDMan and YPDGal compared to those at30°C. Xyl and Ara utilization speeds from 12 to 96 h wereslower than those at 30°C, and both sugars largely remainedeven after 96 h. The ethanol production level in all cases was
the highest at 12 h except for that in YPDGal (24 h).Interestingly, the reduction rate of ethanol at 40°Cwas slightlylower than that at 30°C. The maximum ethanol yields inYPDMan and YPDGal were 0.50 and 0.49, respectively.
At 45°C, cells hardly utilized Xyl or Ara in the presenceof Glc but could utilize Xyl or Ara when they were addedas a single sugar (Fig. 2c). It thus seems that ethanol inYPDXyl or YPDAra comes only from Glc. Gal utilizationin YPDGal started at 12 h and stopped at 48 h. After 48 h,Gal concentration was constant until the end of thefermentation process, and some amount of ethanol wasconsumed after 72 h. The utilization of Gal may somehowbe limited at this temperature when Glc coexists.
Cell growth and ethanol production in YP mediumcontaining a single sugar under a static conditionat different temperatures
K. marxianus is classified as a facultatively fermentativeand Crabtree-negative yeast (van Dijken et al. 1993) and isthus unable to grow under strictly anaerobic conditions.Occurrence of ethanol formation in the organism is almost
a
c
b
YPDAra YPDGal YPDXyl YPDMan
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Man
(g/
l)
Time (h)
OD660nm EtOH Man Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc, X
yl &
Xyl
itol (
g/l)
Time (h)
OD660nm EtOH Xyl Glc Xylitol
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Man
(g/
l)
Time (h)
OD660 nm EtOH Man Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Gal
(g/
l)
Time (h)
OD660nm EtOH Gal Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc, X
yl &
Xyl
itol (
g/l)
Time (h)
OD660nm EtOH Xyl Glc Xylitol
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Man
(g/
l)
Time (h)
OD660nm EtOH Man Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Gal
(g/
l)
Time (h)
OD660nm EtOH Gal Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc, X
yl &
Xyl
itol (
g/l)
Time (h)
OD660nm EtOH Xyl Glc Xylitol
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara Glc
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Gal
(g/
l)
Time (h)
OD660nm EtOH Gal Glc
Fig. 3 Growth and metabolite profiles of K. marxianus DMKU3-1042 grown in YP medium containing 20 g/l glucose with 20 g/l ofmannose (YPDMan), galactose (YPDGal), xylose (YPDXyl), or
arabinose (YPDAra) at 30°C (a), 40°C (b), and 45°C (c) under ashaking condition (160 rpm). Initial OD660 was adjusted to 0.1. Barsrepresent the ±SD for three independent experiments
1578 Appl Microbiol Biotechnol (2011) 90:1573–1586
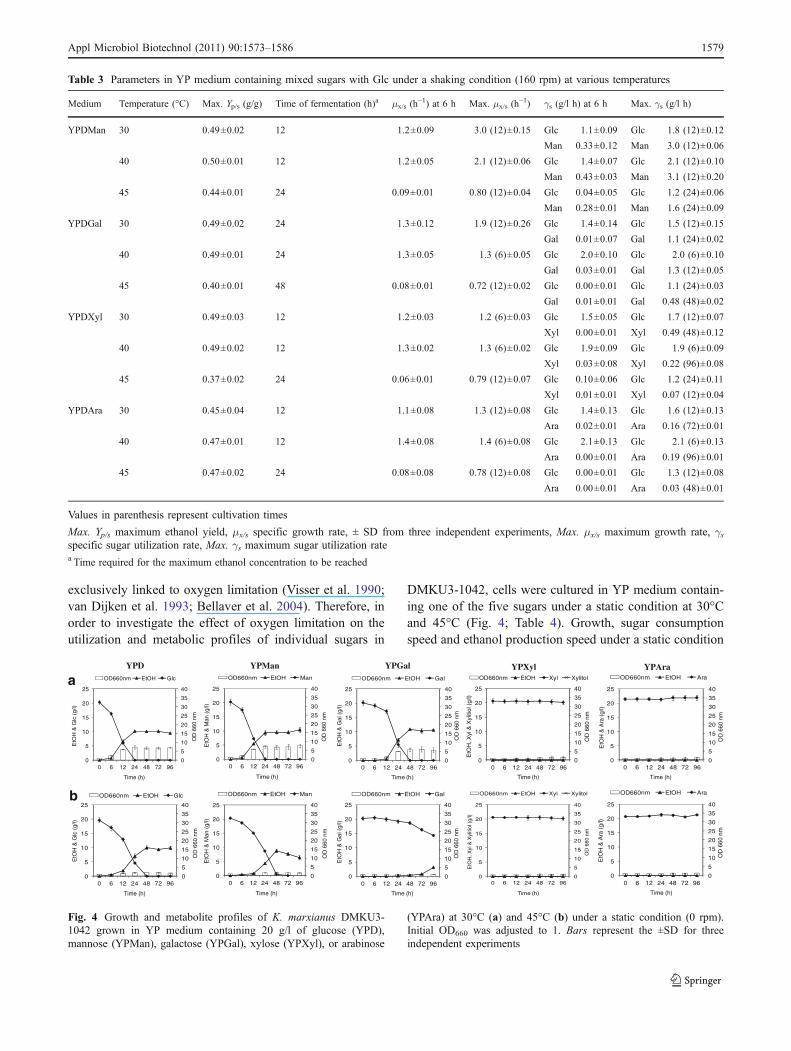
exclusively linked to oxygen limitation (Visser et al. 1990;van Dijken et al. 1993; Bellaver et al. 2004). Therefore, inorder to investigate the effect of oxygen limitation on theutilization and metabolic profiles of individual sugars in
DMKU3-1042, cells were cultured in YP medium contain-ing one of the five sugars under a static condition at 30°Cand 45°C (Fig. 4; Table 4). Growth, sugar consumptionspeed and ethanol production speed under a static condition
a
b
YPD YPAra YPXyl laGPYnaMPY
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Glc
(g/
l)
Time (h)
OD660nm EtOH Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Man
(g/
l)
Time (h)
OD660nm EtOH Man
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Gal
(g/
l)
Time (h)
OD660nm EtOH Gal
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, X
yl &
Xyl
itiol
(g/
l)
Time (h)
OD660nm EtOH Xyl Xylitol
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Glc
(g/
l)
Time (h)
OD660nm EtOH Glc
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Man
(g/
l)
Time (h)
OD660nm EtOH Man
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Gal
(g/
l)
Time (h)
OD660nm EtOH Gal
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H, X
yl &
Xyl
itiol
(g/
l)
Time (h)
OD660nm EtOH Xyl Xylitol
0
5
10
15
20
25
30
35
40
0
5
10
15
20
25
0 6 12 24 48 72 96
OD
660
nm
EtO
H &
Ara
(g/
l)
Time (h)
OD660nm EtOH Ara
Fig. 4 Growth and metabolite profiles of K. marxianus DMKU3-1042 grown in YP medium containing 20 g/l of glucose (YPD),mannose (YPMan), galactose (YPGal), xylose (YPXyl), or arabinose
(YPAra) at 30°C (a) and 45°C (b) under a static condition (0 rpm).Initial OD660 was adjusted to 1. Bars represent the ±SD for threeindependent experiments
Table 3 Parameters in YP medium containing mixed sugars with Glc under a shaking condition (160 rpm) at various temperatures
Medium Temperature (°C) Max. Yp/s (g/g) Time of fermentation (h)a μx/s (h−1) at 6 h Max. μx/s (h
−1) γs (g/l h) at 6 h Max. γs (g/l h)
YPDMan 30 0.49±0.02 12 1.2±0.09 3.0 (12)±0.15 Glc 1.1±0.09 Glc 1.8 (12)±0.12
Man 0.33±0.12 Man 3.0 (12)±0.06
40 0.50±0.01 12 1.2±0.05 2.1 (12)±0.06 Glc 1.4±0.07 Glc 2.1 (12)±0.10
Man 0.43±0.03 Man 3.1 (12)±0.20
45 0.44±0.01 24 0.09±0.01 0.80 (12)±0.04 Glc 0.04±0.05 Glc 1.2 (24)±0.06
Man 0.28±0.01 Man 1.6 (24)±0.09
YPDGal 30 0.49±0.02 24 1.3±0.12 1.9 (12)±0.26 Glc 1.4±0.14 Glc 1.5 (12)±0.15
Gal 0.01±0.07 Gal 1.1 (24)±0.02
40 0.49±0.01 24 1.3±0.05 1.3 (6)±0.05 Glc 2.0±0.10 Glc 2.0 (6)±0.10
Gal 0.03±0.01 Gal 1.3 (12)±0.05
45 0.40±0.01 48 0.08±0.01 0.72 (12)±0.02 Glc 0.00±0.01 Glc 1.1 (24)±0.03
Gal 0.01±0.01 Gal 0.48 (48)±0.02
YPDXyl 30 0.49±0.03 12 1.2±0.03 1.2 (6)±0.03 Glc 1.5±0.05 Glc 1.7 (12)±0.07
Xyl 0.00±0.01 Xyl 0.49 (48)±0.12
40 0.49±0.02 12 1.3±0.02 1.3 (6)±0.02 Glc 1.9±0.09 Glc 1.9 (6)±0.09
Xyl 0.03±0.08 Xyl 0.22 (96)±0.08
45 0.37±0.02 24 0.06±0.01 0.79 (12)±0.07 Glc 0.10±0.06 Glc 1.2 (24)±0.11
Xyl 0.01±0.01 Xyl 0.07 (12)±0.04
YPDAra 30 0.45±0.04 12 1.1±0.08 1.3 (12)±0.08 Glc 1.4±0.13 Glc 1.6 (12)±0.13
Ara 0.02±0.01 Ara 0.16 (72)±0.01
40 0.47±0.01 12 1.4±0.08 1.4 (6)±0.08 Glc 2.1±0.13 Glc 2.1 (6)±0.13
Ara 0.00±0.01 Ara 0.19 (96)±0.01
45 0.47±0.02 24 0.08±0.08 0.78 (12)±0.08 Glc 0.00±0.01 Glc 1.3 (12)±0.08
Ara 0.00±0.01 Ara 0.03 (48)±0.01
Values in parenthesis represent cultivation times
Max. Yp/s maximum ethanol yield, μx/s specific growth rate, ± SD from three independent experiments, Max. μx/s maximum growth rate, γsspecific sugar utilization rate, Max. γs maximum sugar utilization ratea Time required for the maximum ethanol concentration to be reached
Appl Microbiol Biotechnol (2011) 90:1573–1586 1579
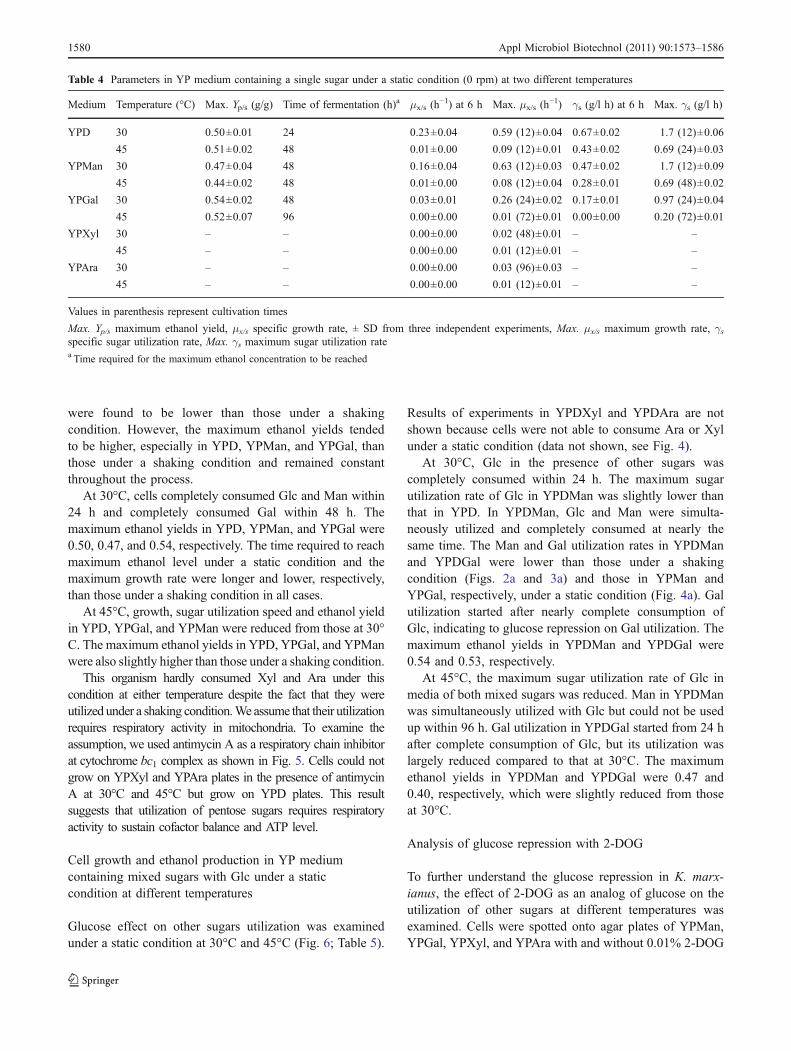
were found to be lower than those under a shakingcondition. However, the maximum ethanol yields tendedto be higher, especially in YPD, YPMan, and YPGal, thanthose under a shaking condition and remained constantthroughout the process.
At 30°C, cells completely consumed Glc and Man within24 h and completely consumed Gal within 48 h. Themaximum ethanol yields in YPD, YPMan, and YPGal were0.50, 0.47, and 0.54, respectively. The time required to reachmaximum ethanol level under a static condition and themaximum growth rate were longer and lower, respectively,than those under a shaking condition in all cases.
At 45°C, growth, sugar utilization speed and ethanol yieldin YPD, YPGal, and YPMan were reduced from those at 30°C. The maximum ethanol yields in YPD, YPGal, and YPManwere also slightly higher than those under a shaking condition.
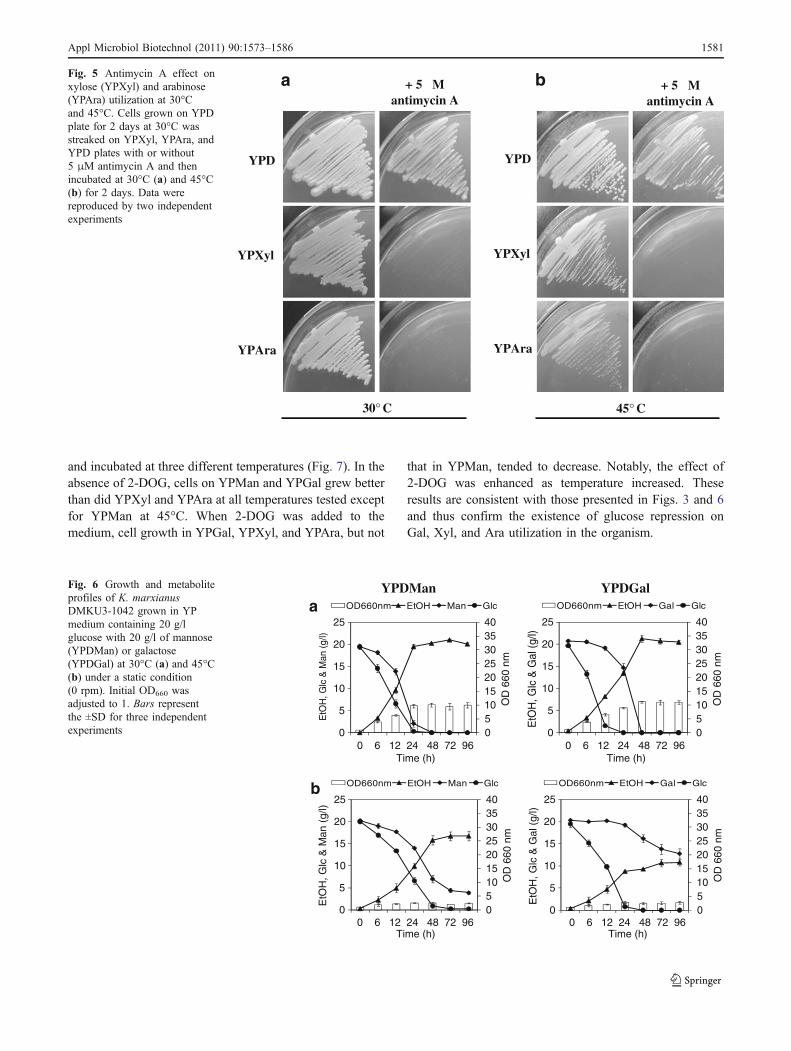
This organism hardly consumed Xyl and Ara under thiscondition at either temperature despite the fact that they wereutilized under a shaking condition.We assume that their utilizationrequires respiratory activity in mitochondria. To examine theassumption, we used antimycin A as a respiratory chain inhibitorat cytochrome bc1 complex as shown in Fig. 5. Cells could notgrow on YPXyl and YPAra plates in the presence of antimycinA at 30°C and 45°C but grow on YPD plates. This resultsuggests that utilization of pentose sugars requires respiratoryactivity to sustain cofactor balance and ATP level.
Cell growth and ethanol production in YP mediumcontaining mixed sugars with Glc under a staticcondition at different temperatures
Glucose effect on other sugars utilization was examinedunder a static condition at 30°C and 45°C (Fig. 6; Table 5).
Results of experiments in YPDXyl and YPDAra are notshown because cells were not able to consume Ara or Xylunder a static condition (data not shown, see Fig. 4).
At 30°C, Glc in the presence of other sugars wascompletely consumed within 24 h. The maximum sugarutilization rate of Glc in YPDMan was slightly lower thanthat in YPD. In YPDMan, Glc and Man were simulta-neously utilized and completely consumed at nearly thesame time. The Man and Gal utilization rates in YPDManand YPDGal were lower than those under a shakingcondition (Figs. 2a and 3a) and those in YPMan andYPGal, respectively, under a static condition (Fig. 4a). Galutilization started after nearly complete consumption ofGlc, indicating to glucose repression on Gal utilization. Themaximum ethanol yields in YPDMan and YPDGal were0.54 and 0.53, respectively.
At 45°C, the maximum sugar utilization rate of Glc inmedia of both mixed sugars was reduced. Man in YPDManwas simultaneously utilized with Glc but could not be usedup within 96 h. Gal utilization in YPDGal started from 24 hafter complete consumption of Glc, but its utilization waslargely reduced compared to that at 30°C. The maximumethanol yields in YPDMan and YPDGal were 0.47 and0.40, respectively, which were slightly reduced from thoseat 30°C.
Analysis of glucose repression with 2-DOG
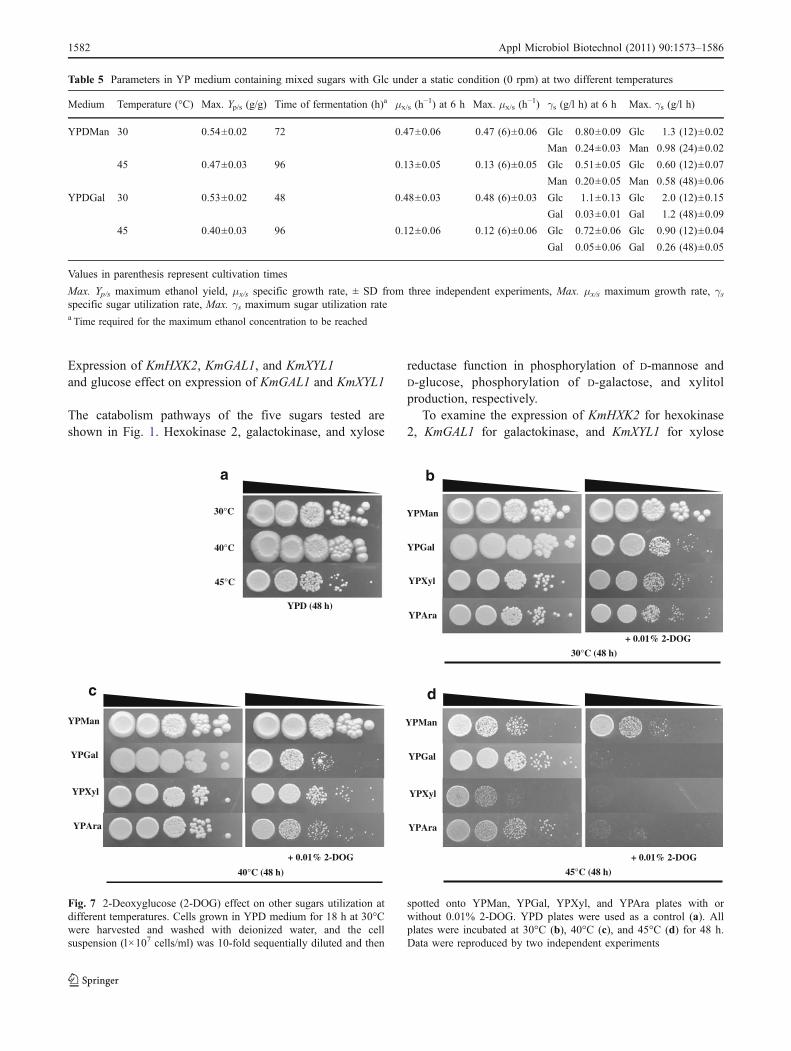
To further understand the glucose repression in K. marx-ianus, the effect of 2-DOG as an analog of glucose on theutilization of other sugars at different temperatures wasexamined. Cells were spotted onto agar plates of YPMan,YPGal, YPXyl, and YPAra with and without 0.01% 2-DOG
Table 4 Parameters in YP medium containing a single sugar under a static condition (0 rpm) at two different temperatures
Medium Temperature (°C) Max. Yp/s (g/g) Time of fermentation (h)a μx/s (h−1) at 6 h Max. μx/s (h
−1) γs (g/l h) at 6 h Max. γs (g/l h)
YPD 30 0.50±0.01 24 0.23±0.04 0.59 (12)±0.04 0.67±0.02 1.7 (12)±0.06
45 0.51±0.02 48 0.01±0.00 0.09 (12)±0.01 0.43±0.02 0.69 (24)±0.03
YPMan 30 0.47±0.04 48 0.16±0.04 0.63 (12)±0.03 0.47±0.02 1.7 (12)±0.09
45 0.44±0.02 48 0.01±0.00 0.08 (12)±0.04 0.28±0.01 0.69 (48)±0.02
YPGal 30 0.54±0.02 48 0.03±0.01 0.26 (24)±0.02 0.17±0.01 0.97 (24)±0.04
45 0.52±0.07 96 0.00±0.00 0.01 (72)±0.01 0.00±0.00 0.20 (72)±0.01
YPXyl 30 – – 0.00±0.00 0.02 (48)±0.01 – –
45 – – 0.00±0.00 0.01 (12)±0.01 – –
YPAra 30 – – 0.00±0.00 0.03 (96)±0.03 – –
45 – – 0.00±0.00 0.01 (12)±0.01 – –
Values in parenthesis represent cultivation times
Max. Yp/s maximum ethanol yield, μx/s specific growth rate, ± SD from three independent experiments, Max. μx/s maximum growth rate, γsspecific sugar utilization rate, Max. γs maximum sugar utilization ratea Time required for the maximum ethanol concentration to be reached
1580 Appl Microbiol Biotechnol (2011) 90:1573–1586
and incubated at three different temperatures (Fig. 7). In theabsence of 2-DOG, cells on YPMan and YPGal grew betterthan did YPXyl and YPAra at all temperatures tested exceptfor YPMan at 45°C. When 2-DOG was added to themedium, cell growth in YPGal, YPXyl, and YPAra, but not
that in YPMan, tended to decrease. Notably, the effect of2-DOG was enhanced as temperature increased. Theseresults are consistent with those presented in Figs. 3 and 6and thus confirm the existence of glucose repression onGal, Xyl, and Ara utilization in the organism.
a
b
YPDGal YPDMan
0510152025303540
0
5
10
15
20
25
0 6 12 24 48 72 96
0 6 12 24 48 72 96
0 6 12 24 48 72 96
OD
660
nm
EtO
H, G
lc &
Man
(g/l)
Time (h)
Time (h)0 6 12 24 48 72 96
Time (h)
Time (h)
OD660nm EtOH Man Glc
0510152025303540
0
5
10
15
20
25
OD
660
nm
EtO
H, G
lc &
Gal
(g/l)
OD660nm EtOH Gal Glc
0510152025303540
0
5
10
15
20
25
OD
660
nm
EtO
H, G
lc &
Gal
(g/
l)
OD660nm EtOH Gal Glc
0510152025303540
0
5
10
15
20
25
OD
660
nm
EtO
H, G
lc &
Man
(g/
l)
OD660nm EtOH Man Glc
Fig. 6 Growth and metaboliteprofiles of K. marxianusDMKU3-1042 grown in YPmedium containing 20 g/lglucose with 20 g/l of mannose(YPDMan) or galactose(YPDGal) at 30°C (a) and 45°C(b) under a static condition(0 rpm). Initial OD660 wasadjusted to 1. Bars representthe ±SD for three independentexperiments
YPD
YPXyl
YPAra
+ 5 µM antimycin A
30° C
+ 5 µM antimycin A
45° C
a b
YPD
YPXyl
YPAra
Fig. 5 Antimycin A effect onxylose (YPXyl) and arabinose(YPAra) utilization at 30°Cand 45°C. Cells grown on YPDplate for 2 days at 30°C wasstreaked on YPXyl, YPAra, andYPD plates with or without5 μM antimycin A and thenincubated at 30°C (a) and 45°C(b) for 2 days. Data werereproduced by two independentexperiments
Appl Microbiol Biotechnol (2011) 90:1573–1586 1581
Expression of KmHXK2, KmGAL1, and KmXYL1and glucose effect on expression of KmGAL1 and KmXYL1
The catabolism pathways of the five sugars tested areshown in Fig. 1. Hexokinase 2, galactokinase, and xylose
reductase function in phosphorylation of D-mannose andD-glucose, phosphorylation of D-galactose, and xylitolproduction, respectively.
To examine the expression of KmHXK2 for hexokinase2, KmGAL1 for galactokinase, and KmXYL1 for xylose
Table 5 Parameters in YP medium containing mixed sugars with Glc under a static condition (0 rpm) at two different temperatures
Medium Temperature (°C) Max. Yp/s (g/g) Time of fermentation (h)a μx/s (h−1) at 6 h Max. μx/s (h
−1) γs (g/l h) at 6 h Max. γs (g/l h)
YPDMan 30 0.54±0.02 72 0.47±0.06 0.47 (6)±0.06 Glc 0.80±0.09 Glc 1.3 (12)±0.02
Man 0.24±0.03 Man 0.98 (24)±0.02
45 0.47±0.03 96 0.13±0.05 0.13 (6)±0.05 Glc 0.51±0.05 Glc 0.60 (12)±0.07
Man 0.20±0.05 Man 0.58 (48)±0.06
YPDGal 30 0.53±0.02 48 0.48±0.03 0.48 (6)±0.03 Glc 1.1±0.13 Glc 2.0 (12)±0.15
Gal 0.03±0.01 Gal 1.2 (48)±0.09
45 0.40±0.03 96 0.12±0.06 0.12 (6)±0.06 Glc 0.72±0.06 Glc 0.90 (12)±0.04
Gal 0.05±0.06 Gal 0.26 (48)±0.05
Values in parenthesis represent cultivation times
Max. Yp/s maximum ethanol yield, μx/s specific growth rate, ± SD from three independent experiments, Max. μx/s maximum growth rate, γsspecific sugar utilization rate, Max. γs maximum sugar utilization ratea Time required for the maximum ethanol concentration to be reached
YPD (48 h)
40°C
45°C (48 h)
YPMan
+ 0.01% 2-DOG
YPXyl
YPAra
YPGal
30°C (48 h)
+ 0.01% 2-DOG
+ 0.01% 2-DOG
30°C
45°C
40°C (48 h)
YPMan
YPXyl
YPAra
YPGal
a b
c d
YPMan
YPXyl
YPAra
YPGal
Fig. 7 2-Deoxyglucose (2-DOG) effect on other sugars utilization atdifferent temperatures. Cells grown in YPD medium for 18 h at 30°Cwere harvested and washed with deionized water, and the cellsuspension (l×107 cells/ml) was 10-fold sequentially diluted and then
spotted onto YPMan, YPGal, YPXyl, and YPAra plates with orwithout 0.01% 2-DOG. YPD plates were used as a control (a). Allplates were incubated at 30°C (b), 40°C (c), and 45°C (d) for 48 h.Data were reproduced by two independent experiments
1582 Appl Microbiol Biotechnol (2011) 90:1573–1586

reductase, we performed RT-PCR with total RNAs fromcells grown until the mid-exponential phase in YP mediumcontaining different sugars or different sugars with Glc attwo different temperatures under shaking conditions(Fig. 8). The band intensity was converted to relativevalues by comparison with that of KmACT1 as an internalcontrol. The values thus reflect the expression level of eachgene tested.
The expression profiles indicated that in YPD, YPMan,and YPDMan, KmHXK2 was similarly expressed at both30°C and 45°C, but its expression was slightly lower at45°C in YPD. Expressional regulation of the gene as theglucose repression was not observed. KmGAL1 was down-regulated in YPD at both temperatures and induced about2-fold in the presence of Gal. Glucose repression onKmGAL1 was observed in YPDGal at 30°C (Fig. 8a, c)and was more evident at 45°C (Fig. 8b, c). KmXYL1
expression in YPD at 30°C and 45°C could not be detected,but it was clearly induced in the presence of Xyl or Ara atboth temperatures. Its induction levels in YPXyl and YPAraat 30°C were about 2.8 and 3.8 times higher, respectively,than those at 45°C. The ratios of KmXYL1 expression inYPXyl versus that in YPDXyl and in YPAra versus that inYPDAra at 30°C were reduced by 5.7- and 6.8-fold,respectively, and those at 45°C were reduced by 2.4- and5.1-fold, respectively. Therefore, these results suggest thatKmGAL1 and KmXYL1 are subjected to glucose repression.
Discussion
In this study, we evaluated the capability of K. marxianusDMKU3-1042 for utilization of various sugars as constit-uents of hemicellulose or ethanol fermentation at high
c
0.0
1.5
3.0
4.5
KmHXK2
Rel
ativ
e ba
nd in
tens
ity
YPD30
YPMan30
YPDMan30
YPD45
YPMan45
YPDMan45
KmACT1
KmXYL1
KmACT1
KmXYL1
YPAra YPDAra
YPXyl YPDXyl 20 25 30 35
YPD 45° C
YPGal YPDGal
b
KmGAL1
KmACT1
KmHXK2
KmXYL1
KmGAL1
KmACT1 KmHXK2
KmACT1
YPMan
YPDMan
20 25 30 35
20 25 30 35 KmHXK2
KmACT1
YPXyl YPDXyl
YPAra YPDAra
KmACT1
KmXYL1
KmACT1
KmXYL1
YPGal YPDGal
YPMan
YPDMan
KmGAL1
KmACT1
KmHXK2 KmXYL1
KmGAL1
KmACT1 KmHXK2
KmACT1
a 30° C YPD
20 25 30 35 20 25 30 35
20 25 30 35
20 23 26 29 32 20 23 26 29 32
20 23 26 29 32 20 23 26 29 32
20 23 26 29 32 20 23 26 29 32
20 23 26 29 32 20 23 26 29 32
20 23 26 29 32 20 23 26 29 32
20 23 26 29 32 20 23 26 29 32
KmHXK2
KmACT1
0.0
1.5
3.0
4.5
KmGAL1
Rel
ativ
e ba
nd in
tens
ity
YPD30
YPGal30
YPDGal30
YPD45
YPGal45
YPDGal45
0.0
1.5
3.0
4.5
KmXYL1
Rel
ativ
e ba
nd in
tens
ity
YPD30
YPAra30
YPDAra30
YPD45
YPAra45
YPDAra45
0.0
1.5
3.0
4.5
KmXYL1
Rel
ativ
e ba
nd in
tens
ity
YPD30
YPXyl30
YPDXyl30
YPD45
YPXyl45
YPDXyl45
Fig. 8 Expression of KmHXK2, KmGAL, and KmXYL1 genes in YPmedium containing a single sugar or a sugar in combination with Glc.Cells grown in YPD medium for 18 h at 30°C were inoculated intobatch culture, which was conducted in 100-ml Erlenmeyer flasks in30-ml YP medium containing 20 g/l of glucose (YPD), mannose(YPMan), galactose (YPGal), arabinose (YPAra), or xylose (YPXyl)or 20 g/l of glucose with 20 g/l of mannose (YPDMan), galactose(YPDGal), xylose (YPDXyl), or arabinose (YPDAra). Cells werefurther cultivated at 30°C (a) or 45°C (b) for 6 h under a shakingcondition (160 rpm). Total RNA was then isolated and subjected to
RT-PCR. RT-PCR was performed with primers specific to KmGAL1,KmHXK2, and KmXYL1. After reverse transcriptase reaction, PCRproducts of 20, 25, 30, and 35 cycles or 20, 23, 26, 29, and 32 cycleswere subjected to agarose gel electrophoresis and stained withethidium bromide. c Band intensity was analyzed by using UN-SCAN-IT gel™ automated digitizing system (Silk Scientific). Relativevalues of intensity of bands in a and b were determined as a ratio ofintensity of each band for each gene to that for KmACT1. Barsrepresent the ±SD for three independent experiments
Appl Microbiol Biotechnol (2011) 90:1573–1586 1583

temperatures and under different aeration conditions. Addition-ally, its glucose effect on utilization of other sugars wasexamined because glucose coexists with other sugars in variousbiomasses. It was found that the yeast was efficiently capable ofassimilating Glc, Man, and Gal up to 45°C under shaking andstatic conditions except for Gal at high temperatures under astatic condition. Xyl and Ara were also utilized under a shakingcondition, though the utilization speed was slower than thosefor the three hexoses and gradually decreased as temperatureincreased. These pentoses, however, could not be utilized undera static condition. These findings indicate high capabilities ofthe organism for utilization of various sugars at relatively hightemperatures and of hexoses under a static condition.
The inability of the organism to utilize the two pentosesunder a static condition is possibly due to incapability ofNADH oxidation by respiratory chain in mitochondria.Consistent with this, antimycin A hampered cell growth(Fig. 5), and disruption of components or synthesis ofcomponents of the respiratory chain caused defectivegrowth in YPXyl or YPAra (unpublished data). Theutilization of Gal at 45°C under a static condition was greatlyreduced compared to that at 30°C. We have no clearexplanation for this phenomenon, but the initial stepsincluding the import of Gal and its conversion to glucose6-phosphate may require more energy than those for Glc orMan (De Bruijne et al. 1988; Van Leeuwen et al. 1991) or theATP level may influence the activity of the H+-galactosidesymporter (Van den Broek et al. 1987).
Ethanol yield from Gal was higher than those from Glcand Man (Fig. 2; Table 2), and slight glucose repressionwas observed in Gal utilization even at 40°C (Figs. 3 and 6;Tables 3 and 5). Although Xyl and Ara utilization wasretarded in the presence of Glc, especially at 45°C,alleviation of the retardation was observed at 40°C (Figs. 3and 6). Unlike S. cerevisiae, this strain shows no glucoserepression in sucrose utilization (unpublished data). Inaddition, reduction of ethanol level at elevated temperaturesunder a shaking condition was lower than that at 30°C(Figs. 2 and 3). This phenomenon is possibly due to thereduction of expression of KmADH4 for alcohol dehydro-genase 4, which is induced by ethanol and thought to beinvolved in ethanol degradation (Lertwattanasakul et al.2007). KmADH4 is expressed in the stationary phase, inwhich glucose is completely consumed and ethanolconcentration becomes the highest. The transcription levelof KmADH4, however, is low at 45°C compared to that at30°C. Ethanol evaporation is another factor to be consid-ered, though it had a minor effect under this conditionbecause efficient evaporation occurs under the conditions ofa higher concentration of ethanol and higher temperature(Abdel-Banat et al. 2010).
It is likely that Man and Glc uptake in K. marxianus isperformed by the same low-affinity glucose transporter
(Gasnier 1987). When the two sugars were present together,only slight delay of Man utilization was observed (Figs. 3and 6). This may be due not to glucose repression but to theKm value for Man being higher than that for Glc as in thecase of S. cerevisiae (Reifenberger et al. 1997). Consistentwith this, no growth retardation was observed in YPMan inthe presence of 2-DOG (Fig. 7).
Glucose repression was observed as a delay of utilizationof Gal, Xyl, and Ara when Glc was present. One of thepossible targets responsible for the repression is the processfor sugar uptake. Three symporters in K. marxianus appearto be sensitive to carbon catabolite inactivation (De Bruijneet al. 1988), but no detailed report on Xyl and Ara transportsystems in K. marxianus is available. Hexose transporterssensitive to glucose repression as in S. cerevisiae andPichia stipitis (Hamacher et al. 2002; Lee et al. 2002;Sedlak and Ho 2004; Kilian and van Uden 1988; Agbogboand Coward-Kelly 2008) would function in the transport ofthese pentoses. These sugars were utilized under a shakingcondition but not under a static condition (Fig. 4), which isknown as the Kluyver effect (Fukuhara 2003). Because of acofactor imbalance in Xyl and Ara metabolic pathways(van Maris et al. 2006; Dien et al. 1996) and low ATPproductivity compared to that of hexose, ethanol productionwas not observed in YPAra and was retarded in YPXyl withaccumulation of xylitol (Fig. 2).
KmGAL1 as a representative gene in Gal utilization wasexamined as a target for the glucose repression. KmGAL1had no canonical sequence for Mig1p but had five putativebinding sequences for Gal4p within a 1-kb upstreamregion from its translation initiation site (data not shown).On the other hand, ScGAL1 in S. cerevisiae and KlGAL1 inK. lactis possess one and four canonical sequences forMig1p and Gal4p, respectively (Rubio-Texeira 2005).KmGAL1 was repressed in YPDGal, the repression beingsignificant at a high temperature (Fig. 8), indicating thepossibility that KmGAL1 has a Mig1p-binding site(s) that isa sequence different from those for Mig1p in S. cerevisiae.The significant effect at a high temperature might be due tostrong binding of Mig1p compared to that at a lowtemperature or an unknown factor(s) might be involved.
When cells were harvested for expression analysis at 6 h,when Xyl and Ara had hardly been utilized in any of themedia tested (Figs. 2 and 3), KmXYL1 was strongly inducedand glucose repression upon its induction was observed(Fig. 8). KmXYL1, however, has no Mig1p-bindingsequence at its upstream region, suggesting the involvementof another glucose repression factor in its repression.Jeffries and Van Vleet (2009) reported that PsXYL1,PsXYL2, and PsXYL3 involved in P. stipitis Xyl metabo-lism, which are upregulated by Xyl under aerobic oroxygen-limited conditions, are downregulated by Glc.However, there are no data for their expression regulation
1584 Appl Microbiol Biotechnol (2011) 90:1573–1586

under the condition with mixed sugars. Bicho et al. (1988)reported that Glc represses activities of xylose reductase andxylitol dehydrogenase in P. stipitis and Pachysolen tanno-philus in a medium containing Glu and Xyl. However, it isnot clear whether glucose repression occurs at the transcrip-tion level or at the post-transcription level. Therefore, themechanism of glucose repression of genes responsible forXyl or Ara metabolism in yeast is still not clear. It is possiblethat there are other regulatory pathways instead of Mig1p forthe glucose repression of KmXYL1.
Our findings allow us to conclude that K. marxianusDMKU3-1042 has high potential for utilization of hexosesand pentoses derived from hemicellulose. This study hasrevealed for the first time the extent of glucose repression ondifferent sugars at different temperatures and has providedvaluable information for application of K. marxianus and forhigh-temperature ethanol fermentation, which will help toimprove the efficiency of its utilization for biotechnologicalpurposes. Its relatively weak glucose repression also encour-ages us to apply it for conversion of biomass containingvarious sugars.
Acknowledgments We thank K. Matsushita and T. Yakushi forhelpful discussion. This work is supported by the Program forPromotion of Basic Research Activities for Innovative Biosciences,NEDO and the Special Coordination Funds for Promoting Science &Technology, Ministry of Education, Culture, Sports, Science &Technology. This work was performed as a collaborative research inthe Asian Core Program between Yamaguchi University and KhonKaen University, which was supported by the Scientific CooperationProgram agreed by the Japan Society for the Promotion of Science andthe National Research Council of Thailand.
References
Abdel-Banat BM, Hoshida H, Ano A, Nonklang S, Akada R (2010)High-temperature fermentation: how can processes for ethanolproduction at high temperatures become superior to the tradi-tional process using mesophilic yeast? Appl Microbiol Biotech-nol 85(4):861–867
Agbogbo FK, Coward-Kelly G (2008) Cellulosic ethanol productionusing the naturally occurring xylose-fermenting yeast, Pichiastipitis. Biotechnol Lett 30(9):1515–1524
Aiba H, Adhya S, de Crombrugghe B (1981) Evidence for twofunctional gal promoters in intact Escherichia coli cells. J BiolChem 256(22):11905–11910
Anderson PJ, McNeil K, Watson K (1986) High-efficiency carbohy-drate fermentation to ethanol at temperatures above 40 degrees Cby Kluyveromyces marxianus var. marxianus isolated from sugarmills. Appl Environ Microbiol 51(6):1314–1320
Aristidou A, Penttila M (2000) Metabolic engineering applications torenewable resource utilization. Curr Opin Biotechnol 11(2):187–198
Banat IM, Nigam P, Singh D, Marchant R, McHale AP (1998) Ethanolproduction at elevated temperatures and alcohol concentrations:part I—yeasts in general. World J Microbiol Biotechnol 14(6):809–821
Bellaver LH, de Carvalho NM, Abrahao-Neto J, Gombert AK (2004)Ethanol formation and enzyme activities around glucose-6-phosphate in Kluyveromyces marxianus CBS 6556 exposed toglucose or lactose excess. FEMS Yeast Res 4(7):691–698
Bicho PA, Runnals PL, Cunningham JD, Lee H (1988) Induction ofxylose reductase and xylitol dehydrogenase activities in Pachy-solen tannophilus and Pichia stipitis on mixed sugars. ApplEnviron Microbiol 54(1):50–54
De Bruijne AW, Schuddemat J, Van den Broek PJ, Van Steveninck J(1988) Regulation of sugar transport systems of Kluyveromycesmarxianus: the role of carbohydrates and their catabolism.Biochim Biophys Acta 939(3):569–576
Dien BS, Kurtzman CP, Saha BC, Bothast RJ (1996) Screening for L-arabinose fermenting yeasts. Appl Biochem Biotechnol 57–58(1):233–242
Dong J, Dickson RC (1997) Glucose represses the lactose–galactoseregulon in Kluyveromyces lactis through a SNF1 and MIG1-dependent pathway that modulates galactokinase (GAL1) geneexpression. Nucleic Acids Res 25(18):3657–3664
Fonseca GG, Heinzle E, Wittmann C, Gombert AK (2008) The yeastKluyveromyces marxianus and its biotechnological potential.Appl Microbiol Biotechnol 79(3):339–354
Fukuhara H (2003) The Kluyver effect revisited. FEMS Yeast Res 3(4):327–331
Gancedo JM (1998) Yeast carbon catabolite repression. Microbiol MolBiol Rev 62(2):334–361
Gasnier B (1987) Characterization of low- and high-affinity glucosetransports in the yeast Kluyveromyces marxianus. BiochimBiophys Acta 903(3):425–433
Gonzalez-Siso MI, Freire-Picos MA, Ramil E, Gonzalez-DominguezM, Rodriguez Torres A, Cerdan ME (2000) Respirofermentativemetabolism in Kluyveromyces lactis: insights and perspectives.Enzyme Microb Technol 26(9–10):699–705
Gonzalez-Siso MI, Garcia-Leiro A, Tarrio N, Cerdan ME (2009)Sugar metabolism, redox balance and oxidative stress responsein the respiratory yeast Kluyveromyces lactis. Microb Cell Fact8:46
Hahn-Hagerdal B, Galbe M, Gorwa-Grauslund MF, Liden G, ZacchiG (2006) Bio-ethanol—the fuel of tomorrow from the residues oftoday. Trends Biotechnol 24(12):549–556
Hamacher T, Becker J, Gardonyi M, Hahn-Hagerdal B, Boles E(2002) Characterization of the xylose-transporting properties ofyeast hexose transporters and their influence on xylose utiliza-tion. Microbiology 148(Pt 9):2783–2788
Jeffries TW, Van Vleet JR (2009) Pichia stipitis genomics, tran-scriptomics, and gene clusters. FEMS Yeast Res 9(6):793–807
Kilian SG, van Uden N (1988) Transport of xylose and glucose in thexylose fermenting yeast Pichia stipitis. Appl Microbiol Bio-technol 27(5–6):545–548
Lane MM, Morrissey PJ (2010) Kluyveromyces marxianus: a yeastemerging from its sister’s shadow. Fungal Biol Rev 24(1–2):17–26
Lee WJ, Kim MD, Ryu YW, Bisson LF, Seo JH (2002) Kinetic studieson glucose and xylose transport in Saccharomyces cerevisiae.Appl Microbiol Biotechnol 60(1–2):186–191
Lertwattanasakul N, Sootsuwan K, Limtong S, Thanonkeo P, YamadaM (2007) Comparison of the gene expression patterns of alcoholdehydrogenase isozymes in the thermotolerant yeast Kluyver-omyces marxianus and their physiological functions. BiosciBiotechnol Biochem 71(5):1170–1182
Limtong S, Sringiew C, Yongmanitchai W (2007) Production of fuelethanol at high temperature from sugar cane juice by a newlyisolated Kluyveromyces marxianus. Bioresour Technol 98(17):3367–3374
Nonklang S, Abdel-Banat BM, Cha-aim K, Moonjai N, Hoshida H,Limtong S, Yamada M, Akada R (2008) High-temperatureethanol fermentation and transformation with linear DNA in the
Appl Microbiol Biotechnol (2011) 90:1573–1586 1585

thermotolerant yeast Kluyveromyces marxianus DMKU3-1042.Appl Environ Microbiol 74(24):7514–7521
Perez J, Munoz-Dorado J, de la Rubia T, Martinez J (2002)Biodegradation and biological treatments of cellulose, hemicel-lulose and lignin: an overview. Int Microbiol 5(2):53–63
Reifenberger E, Boles E, Ciriacy M (1997) Kinetic characterization ofindividual hexose transporters of Saccharomyces cerevisiae andtheir relation to the triggering mechanisms of glucose repression.Eur J Biochem 245(2):324–333
Rubio-Texeira M (2005) A comparative analysis of the GAL geneticswitch between not-so-distant cousins: Saccharomyces cerevisiaeversus Kluyveromyces lactis. FEMS Yeast Res 5(12):1115–1128
Saha BC (2003) Hemicellulose bioconversion. J Ind Microbiol 30(5):279–291
Schaffrath R, Breunig KD (2000) Genetics and molecular physiologyof the yeast Kluyveromyces lactis. Fungal Genet Biol 30(3):173–190
Sedlak M, Ho NW (2004) Characterization of the effectiveness ofhexose transporters for transporting xylose during glucose andxylose co-fermentation by a recombinant Saccharomyces yeast.Yeast 21(8):671–684
Van den Broek PJ, De Bruijne AW, Van Steveninck J (1987) The roleof ATP in the control of H+-galactoside symport in the yeastKluyveromyces marxianus. Biochem J 242(3):729–734
van Dijken JP, Weusthuis RA, Pronk JT (1993) Kinetics of growth andsugar consumption in yeasts. Antonie Leeuwenhoek 63(3–4):343–352
Van Leeuwen CC, Postma E, Van den Broek PJ, Van Steveninck J(1991) Proton-motive force-driven D-galactose transport inplasma membrane vesicles from the yeast Kluyveromyces marx-ianus. J Biol Chem 266(19):12146–12151
van Maris AJ, Abbott DA, Bellissimi E, van den Brink J, Kuyper M,Luttik MA, Wisselink HW, Scheffers WA, van Dijken JP, PronkJT (2006) Alcoholic fermentation of carbon sources in biomasshydrolysates by Saccharomyces cerevisiae: current status. Anto-nie Leeuwenhoek 90(4):391–418
van Ooyen AJ, Dekker P, Huang M, Olsthoorn MM, Jacobs DI,Colussi PA, Taron CH (2006) Heterologous protein production inthe yeast Kluyveromyces lactis. FEMS Yeast Res 6(3):381–392
Venkat RP, Sharad B, Kareenhalli VV (2010) Experimental andsteady-state analysis of the GAL regulatory system in Kluyver-omyces lactis. FEBS J 277(14):2987–3002
Visser W, Scheffers WA, Batenburg-van der Vegte WH, van Dijken JP(1990) Oxygen requirements of yeasts. Appl Environ Microbiol56(12):3785–3792
Zaldivar J, Nielsen J, Olsson L (2001) Fuel ethanol production fromlignocellulose: a challenge for metabolic engineering and processintegration. Appl Microbiol Biotechnol 56(1–2):17–34
1586 Appl Microbiol Biotechnol (2011) 90:1573–1586




















