
Improving and Enhancing Telephone-based Disease Surveillance Systems in Local Health Departments
Upload
independentCategory
view
0download
0
GRIM-19 in Health and Disease
Valdemar Maximo, PhD,*w Jorge Lima, PhD,* Paula Soares, PhD,*w Andre Silva, MSc,*Ines Bento, MSc,* and Manuel Sobrinho-Simoes, MD, PhD*w z
Abstract: GRIM-19, a gene associated with retinoid interferon-
induced mortality, was originally identified as a critical
regulatory protein for interferon-b and retinoic acid-induced
cell death. It was also demonstrated that GRIM-19 is involved
in mitochondrial metabolism, as an integrant component of
complex I of the mitochondrial respiratory chain. GRIM-19
appears, therefore, as a dual function protein involved in cell
death and mitochondrial metabolism. GRIM-19 knock out
leads to Complex I assembly disruption and embryonic lethality
in mice, showing that it is a crucial component of the
mitochondrial respiratory chain essential for early embryonic
development. Recently, mutations in GRIM-19 were described
in Hurthle cell (mitochondrion-rich) tumors of the thyroid and
down-regulation or loss of its expression were found in renal cell
carcinomas, suggesting a role for GRIM-19 in tumorigenesis. As
GRIM-19 binds and inhibits the signal transducer and activator
of transcription-3 (STAT3), which has been shown to be
activated in several human tumors it is tempting to advance
that GRIM-19 may function as a tumor suppressor gene in
tumors in which STAT3 plays a major role.
Key Words: GRIM-19, cell death, mitochondria, cancer, STAT3
(Adv Anat Pathol 2008;15:46–53)
Commentary on: Alchanati I, Nallar SC, Sun P, Gao L, Hu J,
Stein A, Yakirevich E, Konforty D, Alroy I, Zhao X, Reddy SP,
Resnick MB, Kalvakolanu DV. A proteomic analysis reveals the
loss of expression of the cell death regulatory gene GRIM-19 in
human renal cell carcinomas. Oncogene. 2006;25:7138–7147.
CONDENSATION
Purpose of StudyRenal cell carcinoma is a common malignant tumor
with uncertain molecular pathogenesis. This study identifiesdown-regulation of a cell death regulatory gene, GRIM-19,in renal cell carcinoma by proteomic analysis. The authorsfurther attempt to delineate the mechanism of its down-
regulation and explore the potential role of this protein as atumor suppressor via STAT-3 regulatory pathway.
Major Findings
Loss of GRIM-19 Protein Expressionin Renal Cell Carcinoma
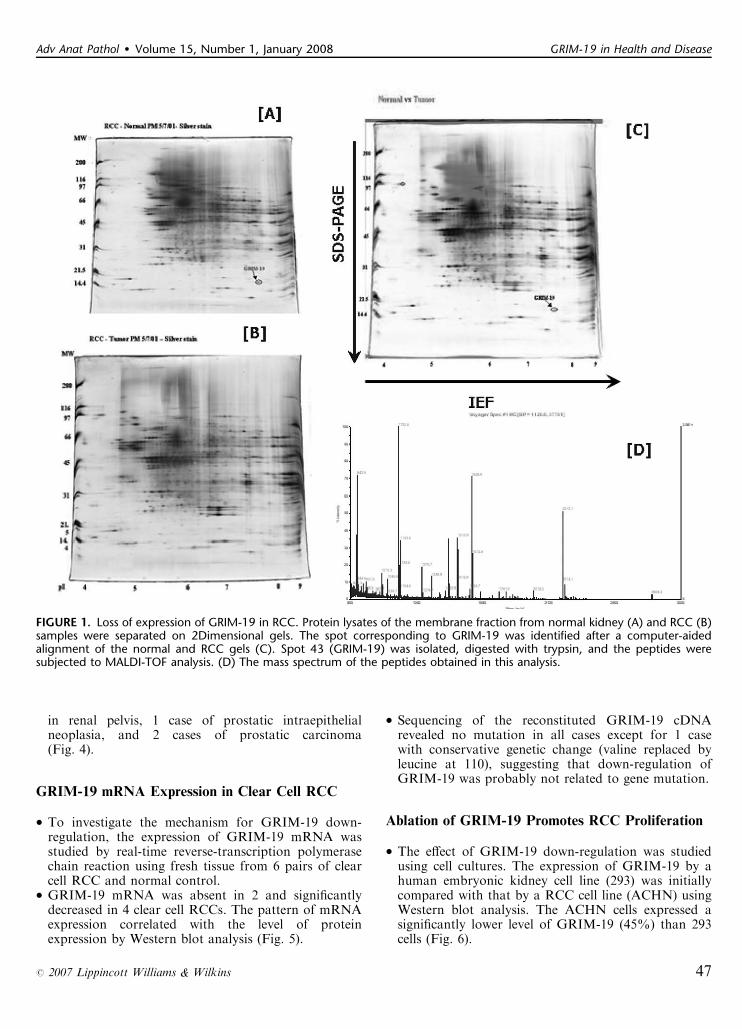
� Five pairs of normal kidney and clear cell renal cellcarcinoma (RCC) tissues were initially analyzed by2-dimensional mass spectrometry. About 50 proteinswere differentially expressed between the normal kidneyand RCC. These protein spots were identified andselected by the Z3 software (Compugen Inc, San Jose,CA) (Fig. 1).� Peptides recovered from spot number 43, which wasconsistently present in normal tissue but absent intumor, were chosen for matrix-assisted laser desorptionionization-time of flight (MALDI-TOR) analysis. Themass fingerprints of the peptides were identified asGRIM-19 from the ProFound database, with aMALDI score of 1.4E07 covering about 56% of theGRIM-19 protein (Fig. 1).� Western blot analysis using mouse monoclonalantibodies against GRIM-19 protein was then appliedto confirm this finding in 11 additional clear cell RCCs.GRIM-19 protein was completely absent in 4, veryweak (10% to 12% of normal) in 6, and moderatelydecreased (50% of normal) in 1 clear cell RCCscompared with normal control (Fig. 2).� Immunostaining for GRIM-19 was also performedin 29 cases of various types of RCCs (20 clear cell, 5chromophobe, and 4 papillary) on paraffin-embeddedmaterials. Intense nuclear membrane and cytoplasmicstaining was observed in proximal tubular epithelium innormal kidney. Expression of GRIM-19 was absent in27 cases (93%), with only very weak staining observedin 1 case of chromophobe RCC and 1 case of clear cellRCC (Fig. 3).
Expression of GRIM-19 in Other Urogenital Tumors
� GRIM-19 expression in other urogenital tumors (6transitional cell carcinomas and 17 prostatic carcinomas)was studied by immunohistochemistry.� GRIM-19 staining was found in normal bladderurothelium and prostatic epithelium. Loss of GRIM-19was found in only 1 case of transitional cell carcinomaCopyright r 2007 by Lippincott Williams & Wilkins
From the *IPATIMUP-Institute of Molecular Pathology and Immunol-ogy of the University of Porto; wDepartment of Pathology, MedicalFaculty of Porto; and zDepartment of Pathology, Hospital SaoJoao, Porto, Portugal.
Funded by The late David and Esther Bernstein Halpern fund.Reprints: Dr Manuel Sobrinho-Simoes, MD, PhD, IPATIMUP, Rua
Dr Roberto Frias s/n, 4200-465 Porto, Portugal (e-mail: [email protected]).
46 Adv Anat Pathol � Volume 15, Number 1, January 2008
COMMENTARY
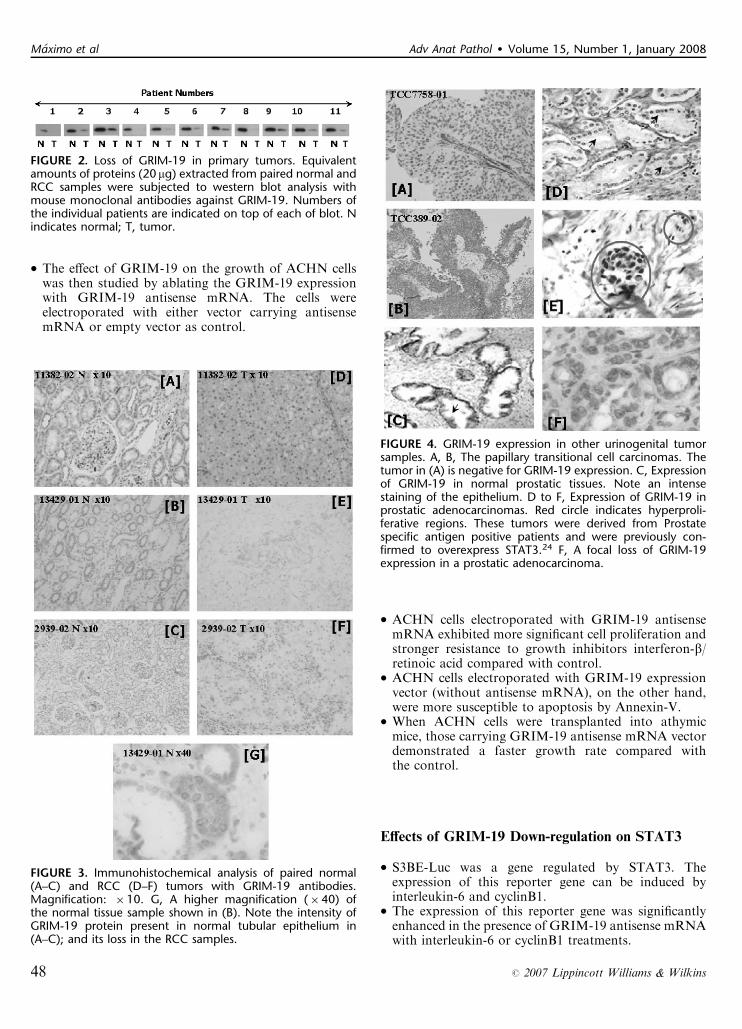
in renal pelvis, 1 case of prostatic intraepithelialneoplasia, and 2 cases of prostatic carcinoma(Fig. 4).
GRIM-19 mRNA Expression in Clear Cell RCC
� To investigate the mechanism for GRIM-19 down-regulation, the expression of GRIM-19 mRNA wasstudied by real-time reverse-transcription polymerasechain reaction using fresh tissue from 6 pairs of clearcell RCC and normal control.� GRIM-19 mRNA was absent in 2 and significantlydecreased in 4 clear cell RCCs. The pattern of mRNAexpression correlated with the level of proteinexpression by Western blot analysis (Fig. 5).
� Sequencing of the reconstituted GRIM-19 cDNArevealed no mutation in all cases except for 1 casewith conservative genetic change (valine replaced byleucine at 110), suggesting that down-regulation ofGRIM-19 was probably not related to gene mutation.
Ablation of GRIM-19 Promotes RCC Proliferation
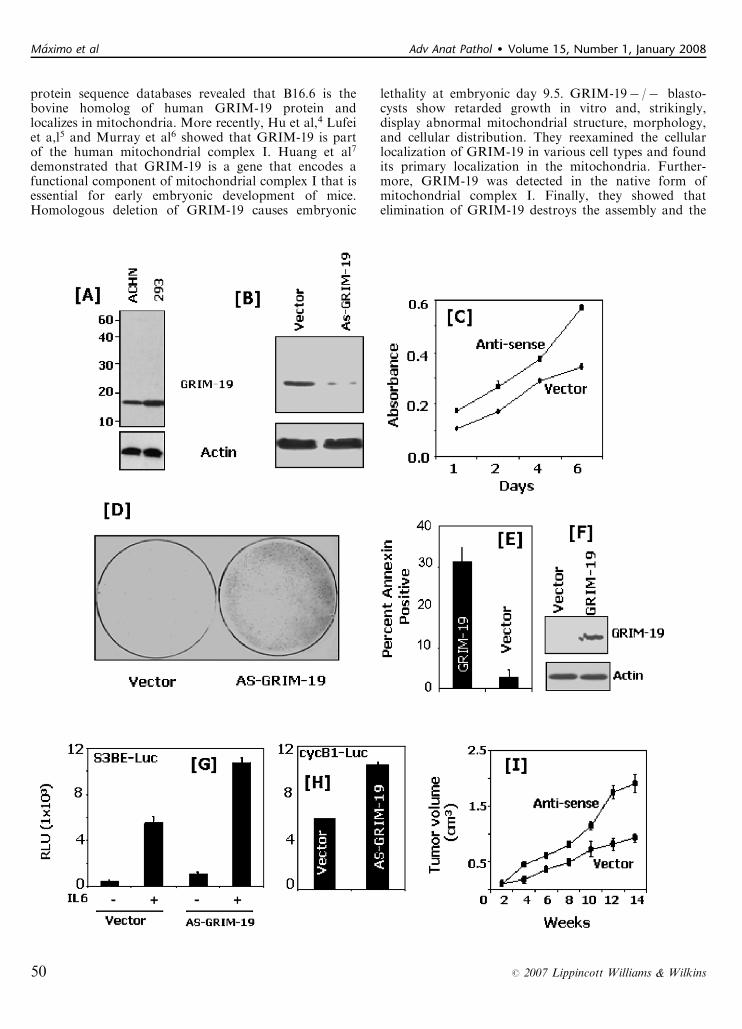
� The effect of GRIM-19 down-regulation was studiedusing cell cultures. The expression of GRIM-19 by ahuman embryonic kidney cell line (293) was initiallycompared with that by a RCC cell line (ACHN) usingWestern blot analysis. The ACHN cells expressed asignificantly lower level of GRIM-19 (45%) than 293cells (Fig. 6).
FIGURE 1. Loss of expression of GRIM-19 in RCC. Protein lysates of the membrane fraction from normal kidney (A) and RCC (B)samples were separated on 2Dimensional gels. The spot corresponding to GRIM-19 was identified after a computer-aidedalignment of the normal and RCC gels (C). Spot 43 (GRIM-19) was isolated, digested with trypsin, and the peptides weresubjected to MALDI-TOF analysis. (D) The mass spectrum of the peptides obtained in this analysis.
Adv Anat Pathol � Volume 15, Number 1, January 2008 GRIM-19 in Health and Disease
r 2007 Lippincott Williams & Wilkins 47
� The effect of GRIM-19 on the growth of ACHN cellswas then studied by ablating the GRIM-19 expressionwith GRIM-19 antisense mRNA. The cells wereelectroporated with either vector carrying antisensemRNA or empty vector as control.
� ACHN cells electroporated with GRIM-19 antisensemRNA exhibited more significant cell proliferation andstronger resistance to growth inhibitors interferon-b/retinoic acid compared with control.� ACHN cells electroporated with GRIM-19 expressionvector (without antisense mRNA), on the other hand,were more susceptible to apoptosis by Annexin-V.� When ACHN cells were transplanted into athymicmice, those carrying GRIM-19 antisense mRNA vectordemonstrated a faster growth rate compared withthe control.
Effects of GRIM-19 Down-regulation on STAT3
� S3BE-Luc was a gene regulated by STAT3. Theexpression of this reporter gene can be induced byinterleukin-6 and cyclinB1.� The expression of this reporter gene was significantlyenhanced in the presence of GRIM-19 antisense mRNAwith interleukin-6 or cyclinB1 treatments.
FIGURE 3. Immunohistochemical analysis of paired normal(A–C) and RCC (D–F) tumors with GRIM-19 antibodies.Magnification: �10. G, A higher magnification (�40) ofthe normal tissue sample shown in (B). Note the intensity ofGRIM-19 protein present in normal tubular epithelium in(A–C); and its loss in the RCC samples.
FIGURE 2. Loss of GRIM-19 in primary tumors. Equivalentamounts of proteins (20 mg) extracted from paired normal andRCC samples were subjected to western blot analysis withmouse monoclonal antibodies against GRIM-19. Numbers ofthe individual patients are indicated on top of each of blot. Nindicates normal; T, tumor.
FIGURE 4. GRIM-19 expression in other urinogenital tumorsamples. A, B, The papillary transitional cell carcinomas. Thetumor in (A) is negative for GRIM-19 expression. C, Expressionof GRIM-19 in normal prostatic tissues. Note an intensestaining of the epithelium. D to F, Expression of GRIM-19 inprostatic adenocarcinomas. Red circle indicates hyperproli-ferative regions. These tumors were derived from Prostatespecific antigen positive patients and were previously con-firmed to overexpress STAT3.24 F, A focal loss of GRIM-19expression in a prostatic adenocarcinoma.
Maximo et al Adv Anat Pathol � Volume 15, Number 1, January 2008
48 r 2007 Lippincott Williams & Wilkins

Conclusions
� GRIM-19 protein expression is significantly decreasedin renal cell carcinoma, but not in other urogenitaltumors such as transitional cell carcinoma or prostaticcarcinoma.
� The down-regulation of GRIM-19 protein is probablyowing to inhibition of transcription rather than genemutation.� Down-regulation of GRIM-19 is associated withenhanced cell proliferation, which may act viauninhibited STAT3 regulatory pathway.� GRIM-19 may represent a tumor suppressor geneinvolved in the pathogenesis of renal cell carcinoma.
COMMENTARYThis study shows another group of human tumors
in which mitochondrial alterations in general, and ofGRIM-19 in particular, may play a major pathogenicrole. GRIM-19 is a mitochondrial protein localized incomplex I of the mitochondrial respiratory chain (MRC)that is encoded by a nuclear gene. Together with a largeamount of information on the gene, and on its mRNAand protein, very interesting data on the relationshipbetween GRIM-19 and human tumorigenesis have beenrecently published.
The GRIM-19 GeneUsing an antisense knockout technique to screen for
genes required for interferon-b (IFN-b)—and retinoicacid (RA)—nduced cell death in a mammary carcinomacell line, followed by classic cDNA library screening,Angell et al1 cloned GRIM-19. Northern blot analysisdetected GRIM-19 in all adult and fetal tissues examined,with highest levels in adult heart, skeletal muscle, liver,kidney, and placenta, and in fetal liver, spleen, and bonemarrow. GRIM-19 expression seems to be higher intissues with high-energy requirements and may also reflectthe number of mitochondria per cell. By Northern andWestern blot analyzes, Angell et al1 found that IFN-b andRA induced GRIM-19 mRNA and protein expressionin breast carcinoma cell lines. Expression of antisenseGRIM-19 conferred resistance against IFN-b/RA-in-duced cell death, and overexpression of GRIM-19produced cells hypersensitive to IFN-b/RA-induced celldeath. Immunofluorescence microscopy localized GRIM-19 to the nucleoplasm of transfected HeLa cells, withsome punctate cytoplasmic staining. Database analysisidentified GRIM-19 homologs in mammalian, plant,nematode, and bacterial cDNA libraries.
Also in 2000, Chidambaram et al2 described thelocalization of GRIM-19 to human chromosome 19p13.1to 13.2 and based in its death-inductive role in the IFN-retinoid-regulated pathways, proposed that GRIM-19might be a novel tumor suppressor.
In 2001, Fearnley et al3 separated the subunits ofbovine mitochondrial complex I and subcomplex Il bysodium dodecyl sulfate polyacrylamide gel electrophoresisand analyzed them by peptide mass fingerprinting. Theyconfirmed the presence of 14 known components ofcomplex Il and observed a further band that did notcorrespond to any known complex I subunit that inaccordance with the nomenclature for bovine mitochon-drial complex I has been named B16.6. Comparison with
FIGURE 5. Expression of GRIM-19 mRNA in normal andtumor cells. A, reverse-transcription polymerase chain reaction(RT-PCR) analysis of the GRIM-19 mRNA expression. Total RNAwas extracted from isogenic pairs of normal and clear cell RCCtumor samples and RT-PCR analysis was performed usinggene-specific-primers. N indicates Normal; T, tumor. B and C,Real-time RT-PCR analysis of the tumor and normal tissuederived GRIM-19 mRNAs. In each pair data were normalizedto that of RP-L32. Individual patient numbers are indicatedbelow each pair of samples. D, Results of a western blotanalysis of the selected normal and RCC tumor samplesprobed with GRIM-19 and actin (internal control) specificantibodies. Normal and tumor samples from the same patientwere used for this analysis.
Adv Anat Pathol � Volume 15, Number 1, January 2008 GRIM-19 in Health and Disease
r 2007 Lippincott Williams & Wilkins 49
protein sequence databases revealed that B16.6 is thebovine homolog of human GRIM-19 protein andlocalizes in mitochondria. More recently, Hu et al,4 Lufeiet a,l5 and Murray et al6 showed that GRIM-19 is partof the human mitochondrial complex I. Huang et al7
demonstrated that GRIM-19 is a gene that encodes afunctional component of mitochondrial complex I that isessential for early embryonic development of mice.Homologous deletion of GRIM-19 causes embryonic
lethality at embryonic day 9.5. GRIM-19� /� blasto-cysts show retarded growth in vitro and, strikingly,display abnormal mitochondrial structure, morphology,and cellular distribution. They reexamined the cellularlocalization of GRIM-19 in various cell types and foundits primary localization in the mitochondria. Further-more, GRIM-19 was detected in the native form ofmitochondrial complex I. Finally, they showed thatelimination of GRIM-19 destroys the assembly and the
Maximo et al Adv Anat Pathol � Volume 15, Number 1, January 2008
50 r 2007 Lippincott Williams & Wilkins

electron transfer activity of complex I and also influencesthe other complexes in the MRC.
GRIM-19 and PathologyGRIM-19 has been found to fulfill 2 roles within the
cell: as a member of the IFN-b and RA-induced pathwayof cell death1 and as part of the mitochondrial complex Iassembly.3,7 These 2 seemingly disparate functions maybe linked, through the involvement of mitochondria inapoptotic cell death. Subsequent studies have shown thatsome oncogenic proteins, such as the viral interferonregulatory factor 1 of human herpes virus-8 and the E6oncogenes of high-risk human papilloma viruses, bind toGRIM-19 and inactivate it.8 Similarly, GW112, a proteinfound in certain human esophageal cancers, has been shownto inhibit the death activating function of GRIM-19.9
By yeast 2-hybrid screening of a human bonemarrow cDNA expression library, Zhang et al10 foundthat GRIM-19 interacted directly with STAT3. STAT3bound GRIM-19 in unstimulated human breast carcino-ma cells, and the amount of STAT3 that coimmunopre-cipitated with GRIM-19 increased after IFN/RAtreatment. No strong interaction was detected betweenGRIM-19 and other STAT proteins abundantly ex-pressed in the carcinoma cell lines. GRIM-19 bindingblocked transcription driven by STAT3 activation, but itdid not inhibit ligand-induced activation of STAT3 norblocked its ability to bind DNA. Mutation analysisindicated that GRIM-19 bound to the transactivationdomain of STAT3, and binding required ser727 withinthe transactivation domain. As constitutively activeSTAT3 up-regulated antiapoptotic genes to promotetumor survival, Zhang et al10 predicted that GRIM-19inhibition of STAT3 would be antioncogenic. Lufei et al5
found that mouse STAT3 interacted with human GRIM-19 when coexpressed in several mammalian cell lines.Coexpression of GRIM-19 inhibited epidermal growth
factor-induced nuclear translocation of STAT3. GRIM-19 repressed STAT3 transcriptional activity and its targetgene expression, and it also suppressed cell growth inSrc-transformed cells. Lufei et al5 concluded that GRIM-19is a negative regulator of STAT3.
Using the same approach, Barnich et al11 found thatGRIM-19 interacted directly with nucleotide-bindingoligomerization domain containing 2 (NOD2). Theantibacterial effects of NOD2, and also those of IFN-band RA, were dependent on GRIM-19 expression.Barnich et al11 also found that expression of GRIM-19was significantly reduced in affected mucosa from Crohndisease and ulcerative colitis patients. Noninvolvedpatient mucosa showed GRIM-19 mRNA expressioncomparable with that in control patients. It is importantto refer that the disparate data concerning GRIM-19localization —nuclear, mitochondrial, and/or cytoplas-mic— may reflect the different roles of GRIM-19 in cellbiology, as part of the MRC and of the apoptoticpathways induced by IFN-b/RA, and also its interactionswith cytoplasmic proteins such as STAT3 andNOD2.1,5,10,11 Summing up, there is enough evidence toclaim that GRIM-19 exerts different cellular functions,which are not yet fully understood.
GRIM-19 Mutations and Human TumorsRecently, we described mutations in the human
GRIM-19 coding region in Hurthle cell thyroid carcino-mas.12 The GRIM-19 mutations we have detected are thefirst nuclear gene mutations specific to Hurthle celltumors to be reported to date, and we proposed thatsuch mutations could be involved in the genesis ofsporadic or familial Hurthle cell tumors through the dualfunction of GRIM-19 in mitochondrial metabolism andcell death.12
RCCs are common malignant tumors that areclassified into clear cell, papillary, chromophobe, and
FIGURE 6. Down-regulation of GRIM-19 in RCC cell line ACHN promotes cell growth. A, A comparison of the GRIM-19 levels inhuman 293 (embryonic kidney cell line) and ACHN cells using western blot analysis. Blots were probed with the indicatedantibodies (B) Antisense mediated knockdown of GRIM-19 expression in ACHN cells. Total cellular protein (50 mg) was used forwestern blot. C, Growth rates of empty vector (pTKO1) and antisense GRIM-19 (AS-GRIM-19) expressing ACHN cells. D, Cellsexpressing AS-GRIM-19 are resistant to the growth inhibitory effects of IFN-b (750 U/mL) and RA (2 mM) combination. Equalnumbers of cells (104) expressing vector or AS-GRIM-19 were selected for 3 weeks in the presence of IFN-b/RA along withhygromycin B (75 mg/mL). Fresh drugs prepared in growth medium were added to the cultures on every third day. At the end ofthird week, plates were fixed with trichloroacetic acid (10%) and then stained with Sulphorhodamine B for 1 hour to visualize thegrowing colonies. The plates were photographed. E, Overexpression of GRIM-19 induces apoptosis. ACHN cells wereelectroporated with 2 mg of empty expression vector (pCXN2-myc) or the same vector carrying a myc-eptiope tagged GRIM-191
and cell death was measured using FACS after staining with FITC-labeled Annexin-V. F, Expression of GRIM-19 in the transfectedcells. Protein extracts from the transfected cells (70 mg) were subjected to western blot analysis using the indicated antibodies.G, and H, Antisense GRIM-19 promotes the STAT3-dependent transcriptional induction of the luciferase reporter driven by aSTAT3-binding element (G) and cyclin B1 promoter (H). Luciferase reporters were transfected along with the b-actin-b-galactosidase vector (internal control for transfection) and the indicated GRIM-19 expression vector into ACHN cells. In (G), cellswere treated with interleukin (IL)-6 (500 U/mL) for 16 hours for stimulating STAT3 activity. The cyclinB1-reporter does not requireIL-6 treatment. Luciferase activity was normalized to that of b-galactosidase and plotted. Each bar represents mean luciferaseactivity [measured as relative light units (RLU)] ± SE of triplicates. I, In vivo growth rates of ACHN tumors expressing AS-GRIM-19or the empty vector. Athymic nude mice (n = 10/group) were transplanted with the indicated cells in 0.1 mL of phosphate-buffered saline. Mean tumor volume ± SE was measured for several weeks and plotted. Mice were killed at the end of 14th week.
Adv Anat Pathol � Volume 15, Number 1, January 2008 GRIM-19 in Health and Disease
r 2007 Lippincott Williams & Wilkins 51

collecting duct types.13,14 Mutations in the von Hippel-Lindau (VHL) tumor suppressor gene or amplificationand activation of the tyrosine kinase c-Met, have beenshown to play critical roles in the development of someRCCs.13,14 Some syndromes associated with hereditaryrenal carcinomas have been described; VHL diseaseassociated with clear cell RCC (susceptibility gene-VHL); hereditary papillary RCC associated with type 1papillary RCC (susceptibility gene-MET); hereditaryleiomyomatosis RCC associated with type 2 papillaryRCC (susceptibility gene-FH); and Birt-Hogg-Dubesyndrome associated with chromophobe renal carcinomasand oncocytomas (susceptibility gene-FLCN).15 Despitethese advances, the full spectrum of molecular changesthat occur in such tumors is far from clear.
The study of Alchanati et al16 provides extremelyinteresting data on the molecular alterations underneathRCC. Using mass spectrometry, they discovered thatexpression of GRIM-19 is lost or severely depressed in anumber of primary RCC and in some urogenital tumors.16
The authors further attempted to delineate the mechanismof GRIM-19 down-regulation and explore the potential roleof this protein as a tumor suppressor via STAT3 regulatorypathway. Using an RCC cell line, the authors have shownthat down-regulation of GRIM-19 promotes tumor growthvia an augmentation of STAT3-dependent gene expression.16
The report by Alchanati et al16 represents animportant step forward in the molecular understandingof the etiopathogenesis of RCCs. Each type of RCC inhereditary syndrome seems to be due to mutations/alterations in different genes.13,14 The study of Alchanatiet al16 demonstrates that in sporadic RCCs, regardlessof the type of tumor, GRIM-19 apparently plays animportant role from the tumorigenesis standpoint.
GRIM-19 alterations have been described, for thefirst time, by Maximo et al12 in Hurthle cell tumors of thethyroid. Mutations in 4/26 (15.4%) of Hurthle cell tumorsand no mutations in 20 non-Hurthle cell tumors, nor in 96blood sample donors, suggest the involvement of GRIM-19alterations in the etiopathogenesis of mitochondrion-richcell tumors of the thyroid. The mitochondrial and the cellsurvival activity of GRIM-19, coupled with the presenceof abnormal mitochondrial structures in Hurthle celltumors, suggest that GRIM-19 mutations may beimportant for Hurthle cell tumor pathogenesis.
In the study by Maximo et al,12 GRIM-19 altera-tions (mutations) were found only on mitochondrion-richtumors of the thyroid, thus leading the authors to suggestthat, because GRIM-19 is part of the MRC, mutationsor deregulation in GRIM-19 protein can explain theaccumulation of mitochondria in tumor cells as acompensatory effect, similar to that observed when thereare mutations in mitochondrial DNA.17 In one case witha GRIM-19 mutation, Maximo et al12 found a ret/PTC-1rearrangement, suggesting that GRIM-19 mutation mayserve as a predisposition alteration for the occurrence oftumors with cell oxiphilia, and that other alterations, suchas ret/PTC rearrangements, may be necessary for theacquisition of a malignant phenotype.12
The results of Alchanati et al,16 raise the hypothesisthat, at least in RCCs, GRIM-19 alterations have nothingto do with mitochondrion-rich tumor cells and may beimportant in every RCC histotypes; nevertheless, it will beinteresting to known if those tumors with down-regula-tion of GRIM-19 present a higher number of mitochon-dria in their cells. The study of oncocytomas may alsoprovide useful data for the clarification of the role ofGRIM-19 alterations in the acquisition of high amountsof mitochondria. The results of Alchanati et al16 areparticularly interesting, because at variance with thevarious genetic alterations of the different types ofRCC,13,14 the GRIM-19 down-regulation was observedin all types of RCC studied, thus suggesting that GRIM-19 may act as a central molecule in RCC.
Alchanati et al16 described a GRIM-19 mutation in1out of 6 (16.7%) RCCs, a low frequency, which is similarto that detected in Hurthle cell tumors of the thyroid.12
These findings indicate that the down-regulation ofGRIM-19 protein is probably because of inhibitionof transcription rather than to gene mutation(s). Theanalysis of GRIM-19 mutations, and also of methylationand loss of transcription factor engagement with thepromoter, in a large series of samples from different typesof RCCs may clarify this issue.
Alchanati et al16 revealed, in RCCs, a severe tocomplete loss of GRIM-19 an IFN-b and RA -inducedcell death-associated protein. IFNs are family cytokinesinvolved in the regulation of innate immunity againstinfectious pathogens, neoplastic growth, and adaptiveimmunity and are used for treatement of several cancers,including RCC, in addition to viral and neurodegenera-tive diseases.18–20 Previous studies showed that the IFN/RA combination synergistically suppresses tumor growthin preclinical and clinical settings.20,21 The recentdiscovery that GRIM-19 interacts directly with NOD211
and that the antibacterial effects of NOD2, and also thoseof RA and IFN-b, were dependent on GRIM-19expression, and on the interaction of GRIM-19 withSTAT3,10 specially after IFN/RA treatment, and, finally,the discovery that GW112, a protein found in certainesophageal cancers, inhibits the death activating functionof GRIM-19,9 suggest that GRIM-19 plays a pathogenicrole in those different disorders. Apparently, the action ofIFN/RA is exerted via GRIM-19, suggesting a putativerole of GRIM-19 in disorders as different as viral andneurodegenerative diseases and cancer.9–12,16,18–23 Thesefindings highlight, once more, the dual role of GRIM-19:as a member of the interferon-b and retinoic acid-inducedpathway of cell death1 and as part of the mitochondrialcomplex I assembly.3,7 In contrast with the embryoniclethal phenotype observed in GRIM-19 knockout mice, adrastic or even complete loss of GRIM-19 expression intumor cells does not apparently interfere with theirviability; indeed, tumors seem to grow perfectly well inthe absence of GRIM-19.1,7 This paradox can beexplained by the so-called Warburg’s effect25: apparentlycancer cells show a decrease in MRC activity and higherlevels of glycolysis. The loss of GRIM-19 may not be
Maximo et al Adv Anat Pathol � Volume 15, Number 1, January 2008
52 r 2007 Lippincott Williams & Wilkins

important regarding energy demands of cancer cells andmay have a positive effect in the decreased apoptoticcapacity of the cells, thus turning the loss of GRIM-19 anadvantage for cancer cells.
ConclusionsThe study of Alchanati et al16 strongly suggests that
GRIM-19 may represent a tumor suppressor geneinvolved in the pathogenesis of RCC and others in whichSTAT3 plays a important role.12,16 It remains to beclarified if the data provided by Alchanati et al16 may beused for practical proposes either in the diagnosis/prog-nosis of RCC in the therapy selection for such tumors.
ACKNOWLEDGMENTThe authors thank Dr Dhan Kalvakolanu, for having
provided the original pictures and for scientific support.Suggested References: Kalvakolanu et al, 2001; Ikeda
et al, 2002; Moore et al, 1994; Kalvakolanu, 2004; Angell,2000; Lufei et al, 2003; Zhang et al, 2003; Buettner et al,2002; Yang et al, 2005; Bromberg et al, 1999; Xu, 2005;Fearnley et al, 2001.
REFERENCES1. Angell JE, Lindner DJ, Shapiro PS, et al. Identification of GRIM-
19, a novel cell death-regulatory gene induced by the interferon-betaand retinoic acid combination, using a genetic approach. J BiolChem. 2000;275:33416–33426.
2. Chidambaram NV, Angell JE, Ling W, et al. Chromosomallocalization of human GRIM-19, a novel IFN-beta and retinoicacid-activated regulator of cell death. J Interferon Cytokine Res.2000;20:661–665.
3. Fearnley IM, Carroll J, Shannon RJ, et al. GRIM-19, a cell deathregulatory gene product, is a subunit of bovine mitochondrialNADH: ubiquinone oxidoreductase (complex I). J Biol Chem.2001;276:38345–38348.
4. Hu J, Angell JE, Zhang J, et al. Characterization of monoclonalantibodies against GRIM-19, a novel IFN-beta and retinoic acid-activated regulator of cell death. J Interferon Cytokine Res. 2002;22:1017–1026. [Erratum in: J Interferon Cytokine Res. 2002;22:1150].
5. Lufei C, Ma J, Huang G, et al. GRIM-19, a death-regulatory geneproduct, suppresses Stat3 activity via functional interaction. EMBOJ. 2003;22:1325–1335.
6. Murray J, Zhang B, Taylor SW, et al. The subunit composition ofthe human NADH dehydrogenase obtained by rapid one-stepimmunopurification. J Biol Chem. 2003;278:13619–13622.
7. Huang G, Lu H, Hao A, et al. GRIM-19, a cell death regulatoryprotein, is essential for assembly and function of mitochondrialcomplex I. Mol Cell Biol. 2004;24:8447–8456.
8. Seo T, Lee D, Shim YS, et al. Viral interferon regulatory factor 1 ofKaposi’s sarcoma-associated herpesvirus interacts with a cell death
regulator, GRIM-19, and inhibits interferon/retinoic acid-inducedcell death. J Virol. 2002;76:8797–8807.
9. Zhang X, Huang Q, Yang Z, et al. GW112, a novel antiapoptoticprotein that promotes tumor growth. Cancer Res. 2004;64:2474–2481.
10. Zhang J, Yang J, Roy SK, et al. The cell death regulator GRIM-19is an inhibitor of signal transducer and activator of transcription 3.Proc Natl Acad Sci U S A. 2003;100:9342–9347.
11. Barnich N, Hisamatsu T, Aguirre JE, et al. GRIM-19 interacts withnucleotide oligomerization domain 2 and serves as downstreameffector of anti-bacterial function in intestinal epithelial cells. J BiolChem. 2005;280:19021–19026.
12. Maximo V, Botelho T, Capela J, et al. Somatic and germlinemutation in GRIM-19, a dual function gene involved in mitochon-drial metabolism and cell death, is linked to mitochondrion-rich(Hurthle cell) tumors of the thyroid. Br J Cancer. 2005;92:1892–1898.
13. Cohen HT, McGovern FJ. Renal-cell carcinoma. N Engl J Med.2005;353:2477–2490.
14. Sudarshan S, Linehan WM. Genetic basis of cancer of the kidney.Semin Oncol. 2006;33:544–551.
15. Eng C, Kiuru M, Fernandez MJ, et al. A role for mitochondrialenzymes in inherited neoplasia and beyond. Nat Rev Cancer.2003;3:193–202.
16. Alchanati I, Nallar SC, Sun P, et al. A proteomic analysis reveals theloss of expression of the cell death regulatory gene GRIM-19 inhuman renal cell carcinomas. Oncogene. 2006;25:7138–7147.
17. Maximo V, Soares P, Lima J, et al. Mitochondrial DNA somaticmutations (point mutations and large deletions) and mitochondrialDNA variants in human thyroid pathology: a study with emphasison Hurthle cell tumors. Am J Pathol. 2002;160:1857–1865.
18. Kalvakolanu DV, Choi K, Borden EC. Interferons and hemato-poietic cytokines. In: Mendelsohn J, Howley PM, Israel MA, et al,eds. The Molecular Basis of Cancer. W.B. Philadelphia: SaundersCompany; 2001:503–534.
19. Ikeda H, Old LJ, Schreiber RD. The roles of IFN gamma inprotection against tumor development and cancer immunoediting.Cytokine Growth Factor Rev. 2002;13:95–109.
20. Kalvakolanu DV. The GRIMs: a new interface between cell deathregulation and interferon/retinoid induced growth suppression.Cytokine Growth Factor Rev. 2004;15:169–194.
21. Moore DM, Kalvakolanu DV, Lippman SM, et al. Retinoic acidand interferon in human cancer: mechanistic and clinical studies.Semin Hematol. 1994;31:31–37.
22. Buettner R, Mora LB, Jove R. Activated STAT signaling in humantumors provides novel molecular targets for therapeutic interven-tion. Clin Cancer Res. 2002;8:945–954.
23. Bromberg JF, Wrzeszczynska MH, Devgan G, et al. Stat3 as anoncogene. Cell. 1999;98:295–303. [Erratum in: Cell. 1999;99:239].
24. Gao L, Zhang L, Hu J, et al. Down-regulation of signal transducerand activator of transcription 3 expression using vector-based smallinterfering RNAs suppresses growth of human prostate tumor invivo. Clin Cancer Res. 2005;11:6333–6341.
25. Warburg O. On the origin of cancer cells. Science. 1956;123:309–314.
Adv Anat Pathol � Volume 15, Number 1, January 2008 GRIM-19 in Health and Disease
r 2007 Lippincott Williams & Wilkins 53
Copyright © 2022 FDOKUMEN




















