
Gm-TX, a new toxic protein from soybean ( Glycine max) seeds with potential for controlling insect...
7
Gm-TX, a new toxic protein from soybean (Glycine max) seeds with potential for controlling insect pests Hermogenes D. Oliveira a , Daniele O.B. Sousa a , Jose T.A. Oliveira a, *, Celia R. Carlini b , Henrique P. Oliveira a , Mirella L. Pereira a , Raquel O. Rocha a , Janne K.S. Morais a , Eneas Gomes-Filho a , Ilka M. Vasconcelos a, * a Departamento de Bioquı´mica e Biologia Molecular, Universidade Federal do Ceara ´, Fortaleza 60451-970, CE, Brazil b Departamento de Biofı´sica, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brazil 1. Introduction Plant defence mechanisms against pathogens and herbivores involve a variety of responses. They include production of reactive oxygen species, programmed cell death, synthesis of antifungal [1,2], insecticidal [3,4] and nematicidal compounds, and cell wall fortification [5,6]. In soybean, little attention has been paid to the physiological and biochemical basis underlying its defence mechanisms in response to pathogen and herbivore attack [7].A recent investigation on the proteomics of seed filling in soybean showed that >600 proteins are expressed during five key stages of seed development. However, most of them, including 7% involved in plant defence, have not been purified and their biological properties evaluated experimentally [8]. In soybean seed coat, a 41-kDa peroxidase [9] and a 32-kDa class I chitinase [10] have been identified, but their functions were not established. On the other hand, Choi et al. [11] showed that SE60, a member of the g-thionin family in soybean seeds, confers resistance to transgenic tobacco plants against Pseudomonas syringae and might function as a defence chemical against invading pathogens. These studies reveal that soybean seeds have defence- related proteins that, once identified and with functions estab- lished, could be expressed in transgenic plants for improved resistance to pathogens and herbivores. Vasconcelos et al. [12] purified and characterised a single-chain acidic (pI 4.4–4.6) protein of 21 kDa from soybean seeds, named soyatoxin (SYTX), that produces acute neurotoxic effects when administered intraperitoneally into mice, with convulsions pre- ceding death (LD 50 7–8 mg/kg mouse body weight). A basic toxic protein (pI 8.2), of 44 kDa, composed of two polypeptide chain (27 and 17 kDa, respectively), linked by disulphide bonds and designated soybean toxin (SBTX), was also isolated from G. max seeds. SBTX showed a LD 50 of 5.6 mg/kg mouse body weight and its toxic effects after intraperitoneal administration in mice [13] were similar to those of SYTX. Despite distinct molecular and physicochemical properties, rabbit antibodies against SYTX cross-reacted with SBTX, suggesting the presence of common antigenic determinants in these proteins. Moreover, in contrast to SBTX, SYTX has a blocked N-terminal amino acid sequence. Nevertheless, SBTX inhibited the mycelial mass development of Cercospora sojina and the N-termini of its 27 and 17 kDa subunits Process Biochemistry 45 (2010) 634–640 ARTICLE INFO Article history: Received 24 August 2009 Received in revised form 1 December 2009 Accepted 16 December 2009 Keywords: Glycine max Soybean Plant protein Insecticidal activity Plant defence Biocontrol ABSTRACT A novel toxic protein from Glycine max seeds, named Gm-TX, was purified by combination of differential precipitation with ammonium sulphate, DEAE-cellulose, anhydrotrypsin-sepharose 4B and Superdex 200 HR fast-protein liquid chromatography. Gm-TX is composed of one polypeptide chain of 28 kDa (SDS-PAGE), with pI 5.1–5.2, contains <0.5% neutral sugar, has N-terminal sequence comprised of KTISSEDSPFFNCREK, maximum absorption spectrum at 280 nm and molar extinction coefficient of 16.9. It exhibited ribonuclease activity (1821.42 3.34 UA/h/mgP) toward yeast RNA and promoted rabbit erythrocyte agglutination (120.4 HU/ mgP) mediated by anti-Gm-TX IgG, which was inhibited by mannose. It was devoid of trypsin inhibitory, urease and chitinase activities. Gm-TX was lethal to mice by intraperitoneal route with LD 50 of 4.5 mg/kg. Moreover, it showed dose-dependent insecticidal activity on the major economically important pests Callosobruchus maculatus and Dysdercus peruvianus, producing ca. 50% and 56% mortality when incorporated into artificial diets at a level of 1.16% and 0.25%, respectively. These data indicate that Gm-TX is a candidate protein that could be expressed in genetically transformed plants for improved resistance to insect pests. ß 2009 Elsevier Ltd. All rights reserved. * Corresponding authors. Tel.: +55 85 3366 9822; fax: +55 85 3366 9789. E-mail addresses: [email protected] (Jose T.A. Oliveira), [email protected] (I.M. Vasconcelos). Contents lists available at ScienceDirect Process Biochemistry journal homepage: www.elsevier.com/locate/procbio 1359-5113/$ – see front matter ß 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.procbio.2009.12.012
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Gm-TX, a new toxic protein from soybean ( Glycine max) seeds with potential for controlling insect...

Gm-TX, a new toxic protein from soybean (Glycine max) seeds with potentialfor controlling insect pests
Hermogenes D. Oliveira a, Daniele O.B. Sousa a, Jose T.A. Oliveira a,*, Celia R. Carlini b,Henrique P. Oliveira a, Mirella L. Pereira a, Raquel O. Rocha a, Janne K.S. Morais a,Eneas Gomes-Filho a, Ilka M. Vasconcelos a,*a Departamento de Bioquımica e Biologia Molecular, Universidade Federal do Ceara, Fortaleza 60451-970, CE, Brazilb Departamento de Biofısica, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brazil
1. Introduction
Plant defence mechanisms against pathogens and herbivoresinvolve a variety of responses. They include production of reactiveoxygen species, programmed cell death, synthesis of antifungal[1,2], insecticidal [3,4] and nematicidal compounds, and cell wallfortification [5,6]. In soybean, little attention has been paid to thephysiological and biochemical basis underlying its defencemechanisms in response to pathogen and herbivore attack [7]. Arecent investigation on the proteomics of seed filling in soybeanshowed that >600 proteins are expressed during five key stages ofseed development. However, most of them, including 7% involvedin plant defence, have not been purified and their biologicalproperties evaluated experimentally [8].
In soybean seed coat, a 41-kDa peroxidase [9] and a 32-kDaclass I chitinase [10] have been identified, but their functions werenot established. On the other hand, Choi et al. [11] showed thatSE60, a member of the g-thionin family in soybean seeds, confers
resistance to transgenic tobacco plants against Pseudomonassyringae and might function as a defence chemical against invadingpathogens. These studies reveal that soybean seeds have defence-related proteins that, once identified and with functions estab-lished, could be expressed in transgenic plants for improvedresistance to pathogens and herbivores.
Vasconcelos et al. [12] purified and characterised a single-chainacidic (pI 4.4–4.6) protein of 21 kDa from soybean seeds, namedsoyatoxin (SYTX), that produces acute neurotoxic effects whenadministered intraperitoneally into mice, with convulsions pre-ceding death (LD50 7–8 mg/kg mouse body weight). A basic toxicprotein (pI 8.2), of 44 kDa, composed of two polypeptide chain (27and 17 kDa, respectively), linked by disulphide bonds anddesignated soybean toxin (SBTX), was also isolated from G. maxseeds. SBTX showed a LD50 of 5.6 mg/kg mouse body weight and itstoxic effects after intraperitoneal administration in mice [13] weresimilar to those of SYTX. Despite distinct molecular andphysicochemical properties, rabbit antibodies against SYTXcross-reacted with SBTX, suggesting the presence of commonantigenic determinants in these proteins. Moreover, in contrast toSBTX, SYTX has a blocked N-terminal amino acid sequence.Nevertheless, SBTX inhibited the mycelial mass development ofCercospora sojina and the N-termini of its 27 and 17 kDa subunits
Process Biochemistry 45 (2010) 634–640
A R T I C L E I N F O
Article history:Received 24 August 2009Received in revised form 1 December 2009Accepted 16 December 2009
Keywords:Glycine maxSoybeanPlant proteinInsecticidal activityPlant defenceBiocontrol
A B S T R A C T
A novel toxic protein from Glycine max seeds, named Gm-TX, was purified by combination of differentialprecipitation with ammonium sulphate, DEAE-cellulose, anhydrotrypsin-sepharose 4B and Superdex200 HR fast-protein liquid chromatography.
Gm-TX is composed of one polypeptide chain of 28 kDa (SDS-PAGE), with pI 5.1–5.2, contains <0.5%neutral sugar, has N-terminal sequence comprised of KTISSEDSPFFNCREK, maximum absorptionspectrum at 280 nm and molar extinction coefficient of 16.9. It exhibited ribonuclease activity(1821.42 ! 3.34 UA/h/mgP) toward yeast RNA and promoted rabbit erythrocyte agglutination (120.4 HU/mgP) mediated by anti-Gm-TX IgG, which was inhibited by mannose. It was devoid of trypsin inhibitory,urease and chitinase activities. Gm-TX was lethal to mice by intraperitoneal route with LD50 of 4.5 mg/kg.Moreover, it showed dose-dependent insecticidal activity on the major economically important pestsCallosobruchus maculatus and Dysdercus peruvianus, producing ca. 50% and 56% mortality when incorporatedinto artificial diets at a level of 1.16% and 0.25%, respectively. These data indicate that Gm-TX is a candidateprotein that could be expressed in genetically transformed plants for improved resistance to insect pests.
! 2009 Elsevier Ltd. All rights reserved.
* Corresponding authors. Tel.: +55 85 3366 9822; fax: +55 85 3366 9789.E-mail addresses: [email protected] (Jose T.A. Oliveira), [email protected]
(I.M. Vasconcelos).
Contents lists available at ScienceDirect
Process Biochemistry
journa l homepage: www.e lsev ier .com/ locate /procbio
1359-5113/$ – see front matter ! 2009 Elsevier Ltd. All rights reserved.doi:10.1016/j.procbio.2009.12.012

share, respectively, high similarities to SC24, an abundant 24-kDaprotein from soybean seed hull, and to cyclophilins, to whichprotective function was attributed [14,15]. Altogether, these datasuggest that SYTX and SBTX might be part of the general defensivestrategy of soybean against pathogens [13]. In our presentinvestigation on soybean proteins we found a novel single-chain28-kDa neurotoxic protein, named G. max toxin (Gm-TX), distinctfrom SYTX and SBTX. In this study, a protocol of purification of Gm-TX, some of its physicochemical properties and its biological effectson two economically important insect pests, the cowpea weevilCallosobruchus maculatus and the cotton stainer bug Dysdercusperuvianus, are presented.
2. Materials and methods
2.1. General
Mature seeds of soybean, Glycine max (L.) Merr. cv. BR-10, adapted to Brazilianlow latitudes were developed and supplied by Empresa Brasileira de PesquisaAgropecuaria (Embrapa – Meio Norte), Piauı, Brazil. Three-month-old New Zealandrabbits were provided by the Departamento de Zootecnia, Universidade Federal doCeara (UFC), Fortaleza, Brazil, and used for production of anti-Gm-TX antibodies andas donors of blood cells for the assay of haemagglutinating activity. C. maculatus(Coleoptera: Bruchidae) and D. peruvianus (Hemiptera: Pyrrhocoridae) wereobtained from the permanent mass-reared populations maintained by theentomology section of Departamento de Biologia, UFC, Fortaleza, Brazil, andDepartamento de Biofısica, Universidade Federal do Rio Grande do Sul, Porto Alegre,Brazil, respectively. The chemicals used were of reagent grade.
2.2. G. max toxin purification
Mature seeds were ground in a coffee grinder fitted with a 1-mm-mesh screenand the resulting flour treated with petroleum ether (1:10, w/v). Proteins wereextracted from defatted soybean by homogenising 1 g of meal in 5 ml of 0.025 MTris–HCl/0.005 M dithiothreitol (DTT), pH 7.5, containing 1.0 " 10#6 M pepstatinand 1.0 " 10#4 M phenylmethylsulfonyl fluoride. The suspension was stirred for 3 hat 4 8C and then filtered through cheesecloth. The residue was re-extracted underthe above conditions and the supernatant collected by centrifugation at 10,000 " gand 4 8C for 30 min. The clear supernatant was precipitated by slowly adding solidammonium sulphate [(NH4)2SO4], with stirring, to 0–20, 20–55 and 55–85%saturation, centrifuged as above, and the precipitates dissolved in and dialysed (cut-off MW 12,000) exhaustively against 0.025 M Tris–HCl/0.005 M DTT, pH 7.5. The20–55% fraction was applied to a DEAE-cellulose (DE-52, Whatman Biochemicals)column previously equilibrated and eluted with the extracting buffer untilcomplete removal of non-retained proteins. Bound proteins were eluted initiallywith 0.15 M NaCl and then with a linear gradient of NaCl (0.15–0.7 M) prepared inthe extracting buffer. To remove any contaminating Kunitz soybean trypsininhibitor (KSTI), samples with intraperitoneal (i.p.) toxic activity to mice werepooled and applied into a anhydrotrypsin-sepharose 4B (GE Healthcare) columnpreviously equilibrated with 0.025 M Tris–HCl containing 0.15 M NaCl, pH 7.5,without DTT. The non-retained, KSTI-free proteins were washed out with theequilibrium buffer, DTT was immediately added to each tube to give a 0.005 M finalconcentration, and the toxic activity to mice assayed. The retained proteinscontaining KSTI were eluted with 0.2 M NaCl prepared in 0.05 M glycine-HCl buffer,pH 2.2. After concentration of the KSTI-free toxic proteins with 90% solid (NH4)2SO4
and dialysis against 0.025 M Tris–HCl containing 0.5 M NaCl, pH 7.5, they wereapplied on a Superdex 200 HR 10/30 column previously equilibrated with the abovebuffer. The fractions displaying toxicity to mice were pooled, concentrated with 90%solid (NH4)2SO4 and dialysed against 0.025 M Tris–HCl, pH 7.5, to which DTT at0.005 M final concentration was added. This purified toxin will hereafter be referredto as Gm-TX (G. max toxin).
2.3. Molecular mass and isoelectric point determination
Native molecular mass (Mr) of Gm-TX (3 mg) was determined by gel-filtrationchromatography on a Superdex 200 HR 10/30 column, equilibrated with 0.025 MTris–HCl buffer, containing 0.5 M NaCl, pH 7.5, coupled to a liquid chromatographysystem (FPLC, Pharmacia-LKB Biotechnology, Sweden), at a constant flow rate of0.5 ml/min. Gm-TX was centrifuged at 10,000 " g at 4 8C, for 10 min and thesupernatant filtered through a Pro-XTM filter unit (0.22-mm hydrophilic celluloseacetate membrane) and applied to the column previously calibrated with proteinsof known molecular mass (bovine serum albumin, 66 kDa; egg albumin, 45 kDa;carbonic anhydrase, 29 kDa; soybean trypsin inhibitor, 20.1 kDa; cytochrome c,12.4 kDa; and aprotinin, 6.5 kDa). The void volume (Vo) was estimated with bluedextran.
Molecular mass estimation under denaturing conditions was carried out bydiscontinuous polyacrylamide gel electrophoresis (SDS-PAGE) using a verticalsystem [16]. A 12.5% polyacrylamide gel prepared in 0.025 M Tris–HCl buffer, pH
8.9, containing 1% sodium dodecyl sulfate (SDS) was used. Samples (20 mg) wereprepared in 0.5 M Tris–HCl buffer, pH 6.8, containing 1% SDS, in the presence orabsence of 1% 2-mercaptoethanol and heated at 100 8C for 10 min, beforeelectrophoresis. Molecular mass markers were used as mentioned above. Proteinbands were stained with colloidal coomassie [17].
Bidimensional electrophoresis (2D) of Gm-TX was carried out according to Gorget al. [18] with minor modifications. Gm-TX (100 mg) was solubilised in 200 ml re-hydration buffer [19], loaded directly on immobilised pH gradient gel strip (11 cm,pH 3–10, GE Healthcare) and incubated for 12 h. Focusing was performed in astepwise mode: 500 V, 30 min; 1000 V, 30 min; 3500 V, 30 min; and 3500 V, 5.5 husing Multiphor II Electrophoresis System (GE Healthcare). SDS-PAGE (12.5%;18 cm " 24 cm " 0.15 cm) was conducted [16] in a vertical system (Hoefer SE 600,GE Healthcare) under 50 mA, 250 V and 20 8C. Protein spots were stained aspreviously [17]. The molecular mass of Gm-TX was estimated using the sameprotein markers above plus lactalbumin (14.2 kDa). The gel was scanned (LabScansoftware) and analysed using the program Image Master TM 2D Platinum v. 7.0 (GEHealthcare).
2.4. Amino acid sequence analysis
The N-terminal amino acid sequence of Gm-TX was analysed on a Shimadzu PPSQ-10 Automated Protein Sequencer performing Edman degradation. Sequences weredetermined from Gm-TX blotted on polyvinylidene fluoride membrane after SDS-PAGE. Phenylthiohydantoin-amino acids were detected at 269 nm after separation ona reversed phase C18 column (4.6 mm" 2.5 mm) under isocratic conditions,according to the manufacturer’s instructions. The sequence obtained was submittedto automatic alignment, performed using the NCBI-BLAST search system [20].
2.5. Absorption spectra
Absorption spectra were recorded on a Spectronic 7 spectrophotometer(Pharmacia-LKB) with the wavelengths of 220–360 nm, using a 1-cm path-lengthcuvette (1 ml volume). Solutions of Gm-TX were prepared by dissolving 1 mg of thetoxin in double distilled water, with and without 0.005 M DTT
2.6. Protein and carbohydrate contents
Protein content was determined using the method described by Bradford [21]with bovine serum albumin as standard. Absorbance at 280 nm was also used todetermine the protein content of column eluates. The neutral sugar content of Gm-TX was estimated by the phenol–sulphuric acid method with reference to D-glucose[22]. The presence of carbohydrate was also evaluated by specific staining of Gm-TXband after SDS-PAGE using the Glycoprotein Detection Kit (Sigma).
2.7. Production of anti-Gm-TX antibodies
Rabbits were immunized with Gm-TX as in Rios et al. [23]. Polyclonal antibodies,including anti-Gm-TX IgG, were purified from rabbit serum by saturation to 33%(NH4)2SO4 [24] and chromatography on Protein A-Agarose, following themanufacturer’s instructions (Sigma).
2.8. Direct and indirect haemagglutination assay
The haemagglutinating activity (direct activity) of Gm-TX was measured by usingfresh rabbit erythrocytes [25]. A serial 2-fold dilution of Gm-TX in a microtitre U-plates (100 ml) was mixed with 100 ml of a 2% suspension of erythrocytes inphosphate-buffered saline (pH 7.2) at 22 8C for 1 h. After this time, haemagglutinationwas checked in comparison to a blank in which Gm-TX had been omitted. Indirecthaemagglutinating activity was performed as described by Carlini and Guimaraes[26]. Briefly, Gm-TX was previously incubated with the rabbit red blood cells (2%) for1 h, at 22 8C, followed by addition of an equal volume of anti-Gm-TX IgG (0.5 mg/ml),for a further 1 h, and haemagglutination checked as above. Red cells incubated onlywith anti-Gm-TX IgG or treated only with Gm-TX were used as negative controls. Thehaemagglutination titre, defined as the reciprocal of the highest dilution exhibitingvisible agglutination at naked eyes, was considered as 1 haemagglutination unit (HU).Specific activity was expressed as HU per mg protein (HU/mg).
2.9. Inhibition of Gm-TX-induced haemagglutination by carbohydrates
Inhibition of indirect haemagglutination activity by simple sugars was carriedout by incubating 100 ml of D-glucose, D-mannose, D-galactose and N-acetyl-D-glucosamine, at 0.1 M final concentration, with 100 ml of a solution of Gm-TX with4 HU. The mixture was allowed to stand for 30 min at 22 8C followed by incubationwith 2% suspension of erythrocytes and anti-Gm-TX IgG as described before.Haemagglutination titre was evaluated as before.
2.10. Trypsin inhibitory activity
Trypsin inhibitor activity was determined by a slight modification of the methodoriginally described by Kakade et al. [27,28]. The samples were tested for their
H.D. Oliveira et al. / Process Biochemistry 45 (2010) 634–640 635
inhibitory effect on the hydrolytic activity of trypsin upon benzoyl-L-arginine-p-nitroanilide (BApNA). The release of p-nitroaniline was measured at 410 nm.
2.11. Urease assay
Urease assay was carried out according to Kaplan [29] with minor modifications[25]. The enzyme activity was estimated calorimetrically at 625 nm and calculatedfrom a calibration curve using crystalline urease (Sigma 41H7008, 870,000 units/g).
2.12. Chitinase assay (CHI, EC 3.2.1.14)
Chitinase activity was measured colorimetrically by release of N-acetyl-D-glucosamine produced by the hydrolytic action of chitinases upon colloidal chitin asa substrate [30–32]. Colloidal chitin was prepared following the protocol describedby Molano et al. [33], except that non-radioactive acetic anhydride was used toacetylate the crab shell chitosan obtained from Sigma Chem. Co.
2.13. Ribonuclease activity
This was determined by modification of the method described by Tuve andAnfinsen [34,35]. Assays were carried out at 40 8C in 2.5 ml of 0.1 M sodium acetatebuffer, pH 5.8, containing 0.2% yeast RNA type IV (Sigma Chemical Co.) substrateand 350 mg Gm-TX. The blank consisted of the same reaction mixture as the testsample, except that Gm-TX was not added. The reaction was terminated after 1 h byaddition of 0.75% uranyl acetate in HClO4 at 25%. Acid-soluble nucleotideproduction was determined spectrophotometrically at 260 nm. One unit of activity(UA) was defined as a 0.01 difference in absorbance (DA260).
2.14. Toxicity activity in mice
This study was performed in accordance with the federal law regarding theprotection of animals, reviewed and approved by the Animal Ethics Committee(CEPA) of UFC, Brazil. Toxic activity was defined as mortality observed in Swiss micewithin 24 h after i.p. or intravenous injections of Gm-TX [12]. One LD50 unit wastaken as the amount of protein (mg protein/kg mouse body weight) producingconvulsions and death of 50% tested animals (six doses; six mice per dose).
2.15. Insecticidal effect of Gm-TX on C. maculatus
The stock culture of C. maculatus was maintained on susceptible commercialcowpea beans (Vigna unguiculata cv. Epace 10) purchased at local stores. Rearing ofinsects and experiments were carried out in an incubator at a constanttemperature of 27 ! 2 8C and 60–70% relative humidity. The insecticidal effect ofGm-TX on C. maculatus in artificial seeds was assessed following the previouslydescribed method by Macedo et al. [4]. Artificial seeds (ca. 400 mg each) wereprepared with finely ground cowpea seed flour (cv. Epace 10) using a cylindrical brassmal and a hand press. Artificial seeds containing Gm-TX at 0.2, 0.4, 0.8 and 1.6% (w/w)final concentrations were manufactured by thoroughly mixing the protein withcowpea seed meal and pressing as described above. After a 48-h adjustment period inthe growth chamber, the seeds were offered to nine 2–3-day-old fertilised females for24 h. The control was cowpea seed meal. After oviposition, the number of eggs wasmanually reduced to 6/seed (n = 15), and adult emergence observed over 8 weeks.Control and test groups consisted of 15 artificial seeds per vial, with four replicates.The percentage of adult emergence, developmental time and body weight werecalculated.
2.16. Insecticidal effect of Gm-TX on D. peruvianus
The D. peruvianus were reared at 26 8C, 70–80% relative humidity and 16-h-light/8-h-dark cycle. Groups of second instar insects were fed on cotton seed meal [36]mixed or not (control) with freeze-dried Gm-TX at concentrations of 0.25, 0.025 and0.0025% (w/w). The insects were observed for 28 d for lethality (survival rate) andbody weight. Control and test groups consisted of 20 insects per vial containing fiveartificial seeds, with four replicates.
2.17. Statistical analysis
Data were examined using one-way analysis of variance (ANOVA). Tukey’s testwas used to identify significance differences amongst means (P < 0.05).
3. Results and discussion
3.1. Purification of Gm-TX
Gm-TX was successfully purified by a combination of (NH4)2SO4
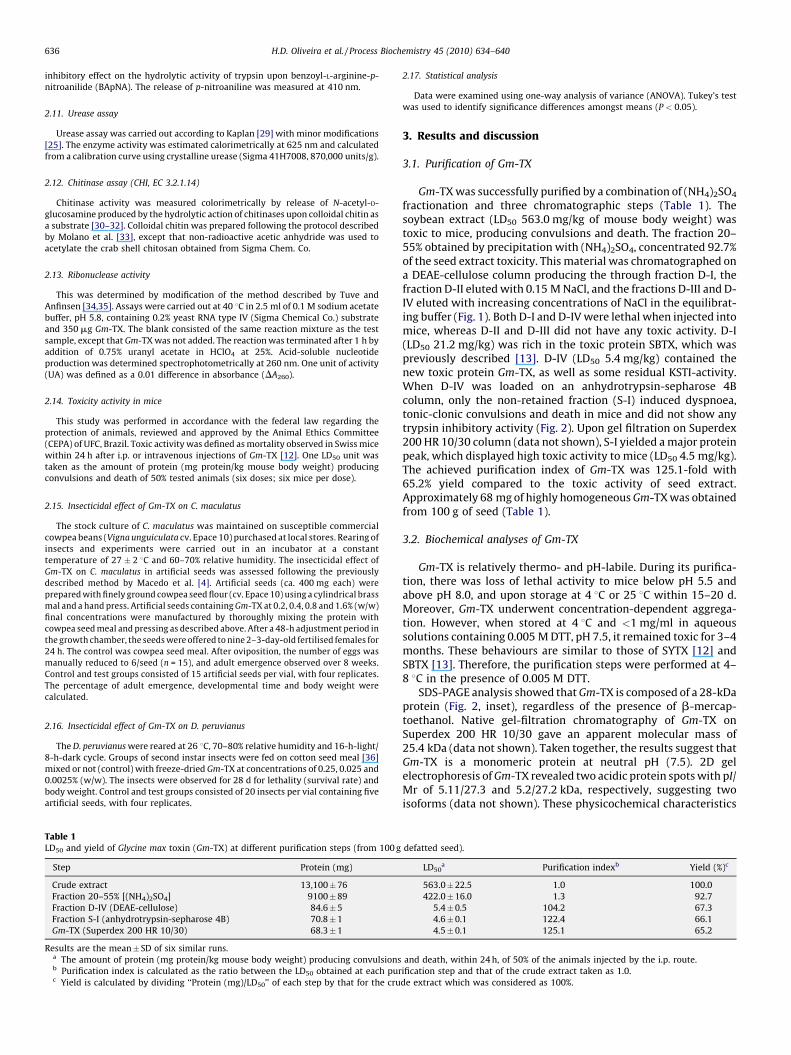
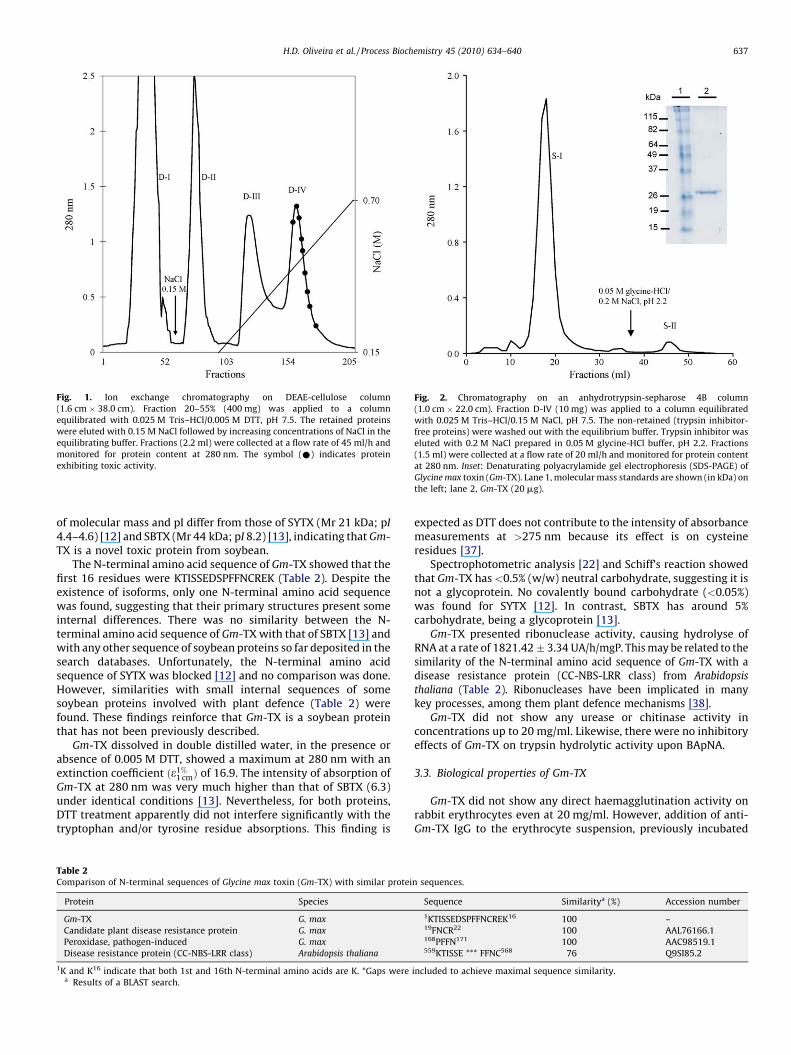
fractionation and three chromatographic steps (Table 1). Thesoybean extract (LD50 563.0 mg/kg of mouse body weight) wastoxic to mice, producing convulsions and death. The fraction 20–55% obtained by precipitation with (NH4)2SO4, concentrated 92.7%of the seed extract toxicity. This material was chromatographed ona DEAE-cellulose column producing the through fraction D-I, thefraction D-II eluted with 0.15 M NaCl, and the fractions D-III and D-IV eluted with increasing concentrations of NaCl in the equilibrat-ing buffer (Fig. 1). Both D-I and D-IV were lethal when injected intomice, whereas D-II and D-III did not have any toxic activity. D-I(LD50 21.2 mg/kg) was rich in the toxic protein SBTX, which waspreviously described [13]. D-IV (LD50 5.4 mg/kg) contained thenew toxic protein Gm-TX, as well as some residual KSTI-activity.When D-IV was loaded on an anhydrotrypsin-sepharose 4Bcolumn, only the non-retained fraction (S-I) induced dyspnoea,tonic-clonic convulsions and death in mice and did not show anytrypsin inhibitory activity (Fig. 2). Upon gel filtration on Superdex200 HR 10/30 column (data not shown), S-I yielded a major proteinpeak, which displayed high toxic activity to mice (LD50 4.5 mg/kg).The achieved purification index of Gm-TX was 125.1-fold with65.2% yield compared to the toxic activity of seed extract.Approximately 68 mg of highly homogeneous Gm-TX was obtainedfrom 100 g of seed (Table 1).
3.2. Biochemical analyses of Gm-TX
Gm-TX is relatively thermo- and pH-labile. During its purifica-tion, there was loss of lethal activity to mice below pH 5.5 andabove pH 8.0, and upon storage at 4 8C or 25 8C within 15–20 d.Moreover, Gm-TX underwent concentration-dependent aggrega-tion. However, when stored at 4 8C and <1 mg/ml in aqueoussolutions containing 0.005 M DTT, pH 7.5, it remained toxic for 3–4months. These behaviours are similar to those of SYTX [12] andSBTX [13]. Therefore, the purification steps were performed at 4–8 8C in the presence of 0.005 M DTT.
SDS-PAGE analysis showed that Gm-TX is composed of a 28-kDaprotein (Fig. 2, inset), regardless of the presence of b-mercap-toethanol. Native gel-filtration chromatography of Gm-TX onSuperdex 200 HR 10/30 gave an apparent molecular mass of25.4 kDa (data not shown). Taken together, the results suggest thatGm-TX is a monomeric protein at neutral pH (7.5). 2D gelelectrophoresis of Gm-TX revealed two acidic protein spots with pI/Mr of 5.11/27.3 and 5.2/27.2 kDa, respectively, suggesting twoisoforms (data not shown). These physicochemical characteristics
Table 1LD50 and yield of Glycine max toxin (Gm-TX) at different purification steps (from 100 g defatted seed).
Step Protein (mg) LD50a Purification indexb Yield (%)c
Crude extract 13,100!76 563.0!22.5 1.0 100.0Fraction 20–55% [(NH4)2SO4] 9100!89 422.0!16.0 1.3 92.7Fraction D-IV (DEAE-cellulose) 84.6!5 5.4! 0.5 104.2 67.3Fraction S-I (anhydrotrypsin-sepharose 4B) 70.8!1 4.6! 0.1 122.4 66.1Gm-TX (Superdex 200 HR 10/30) 68.3!1 4.5! 0.1 125.1 65.2
Results are the mean! SD of six similar runs.a The amount of protein (mg protein/kg mouse body weight) producing convulsions and death, within 24 h, of 50% of the animals injected by the i.p. route.b Purification index is calculated as the ratio between the LD50 obtained at each purification step and that of the crude extract taken as 1.0.c Yield is calculated by dividing ‘‘Protein (mg)/LD50’’ of each step by that for the crude extract which was considered as 100%.
H.D. Oliveira et al. / Process Biochemistry 45 (2010) 634–640636
of molecular mass and pI differ from those of SYTX (Mr 21 kDa; pI4.4–4.6) [12] and SBTX (Mr 44 kDa; pI 8.2) [13], indicating that Gm-TX is a novel toxic protein from soybean.
The N-terminal amino acid sequence of Gm-TX showed that thefirst 16 residues were KTISSEDSPFFNCREK (Table 2). Despite theexistence of isoforms, only one N-terminal amino acid sequencewas found, suggesting that their primary structures present someinternal differences. There was no similarity between the N-terminal amino acid sequence of Gm-TX with that of SBTX [13] andwith any other sequence of soybean proteins so far deposited in thesearch databases. Unfortunately, the N-terminal amino acidsequence of SYTX was blocked [12] and no comparison was done.However, similarities with small internal sequences of somesoybean proteins involved with plant defence (Table 2) werefound. These findings reinforce that Gm-TX is a soybean proteinthat has not been previously described.
Gm-TX dissolved in double distilled water, in the presence orabsence of 0.005 M DTT, showed a maximum at 280 nm with anextinction coefficient $e1%
1 cm% of 16.9. The intensity of absorption ofGm-TX at 280 nm was very much higher than that of SBTX (6.3)under identical conditions [13]. Nevertheless, for both proteins,DTT treatment apparently did not interfere significantly with thetryptophan and/or tyrosine residue absorptions. This finding is
expected as DTT does not contribute to the intensity of absorbancemeasurements at >275 nm because its effect is on cysteineresidues [37].
Spectrophotometric analysis [22] and Schiff’s reaction showedthat Gm-TX has <0.5% (w/w) neutral carbohydrate, suggesting it isnot a glycoprotein. No covalently bound carbohydrate (<0.05%)was found for SYTX [12]. In contrast, SBTX has around 5%carbohydrate, being a glycoprotein [13].
Gm-TX presented ribonuclease activity, causing hydrolyse ofRNA at a rate of 1821.42 ! 3.34 UA/h/mgP. This may be related to thesimilarity of the N-terminal amino acid sequence of Gm-TX with adisease resistance protein (CC-NBS-LRR class) from Arabidopsisthaliana (Table 2). Ribonucleases have been implicated in manykey processes, among them plant defence mechanisms [38].
Gm-TX did not show any urease or chitinase activity inconcentrations up to 20 mg/ml. Likewise, there were no inhibitoryeffects of Gm-TX on trypsin hydrolytic activity upon BApNA.
3.3. Biological properties of Gm-TX
Gm-TX did not show any direct haemagglutination activity onrabbit erythrocytes even at 20 mg/ml. However, addition of anti-Gm-TX IgG to the erythrocyte suspension, previously incubated
Fig. 2. Chromatography on an anhydrotrypsin-sepharose 4B column(1.0 cm " 22.0 cm). Fraction D-IV (10 mg) was applied to a column equilibratedwith 0.025 M Tris–HCl/0.15 M NaCl, pH 7.5. The non-retained (trypsin inhibitor-free proteins) were washed out with the equilibrium buffer. Trypsin inhibitor waseluted with 0.2 M NaCl prepared in 0.05 M glycine-HCl buffer, pH 2.2. Fractions(1.5 ml) were collected at a flow rate of 20 ml/h and monitored for protein contentat 280 nm. Inset: Denaturating polyacrylamide gel electrophoresis (SDS-PAGE) ofGlycine max toxin (Gm-TX). Lane 1, molecular mass standards are shown (in kDa) onthe left; lane 2, Gm-TX (20 mg).
Fig. 1. Ion exchange chromatography on DEAE-cellulose column(1.6 cm " 38.0 cm). Fraction 20–55% (400 mg) was applied to a columnequilibrated with 0.025 M Tris–HCl/0.005 M DTT, pH 7.5. The retained proteinswere eluted with 0.15 M NaCl followed by increasing concentrations of NaCl in theequilibrating buffer. Fractions (2.2 ml) were collected at a flow rate of 45 ml/h andmonitored for protein content at 280 nm. The symbol (*) indicates proteinexhibiting toxic activity.
Table 2Comparison of N-terminal sequences of Glycine max toxin (Gm-TX) with similar protein sequences.
Protein Species Sequence Similaritya (%) Accession number
Gm-TX G. max 1KTISSEDSPFFNCREK16 100 –Candidate plant disease resistance protein G. max 19FNCR22 100 AAL76166.1Peroxidase, pathogen-induced G. max 168PFFN171 100 AAC98519.1Disease resistance protein (CC-NBS-LRR class) Arabidopsis thaliana 559KTISSE *** FFNC568 76 Q9SI85.2
1K and K16 indicate that both 1st and 16th N-terminal amino acids are K. *Gaps were included to achieve maximal sequence similarity.a Results of a BLAST search.
H.D. Oliveira et al. / Process Biochemistry 45 (2010) 634–640 637
with Gm-TX, promoted indirect agglutination with 2048 HU/mland 120.4 HU/mg protein. Moreover, this indirect haemagglutina-tion activity of Gm-TX was specifically inhibited by 0.1 M D-mannose, but not by D-glucose, D-galactose and N-acetyl-D-glucosamine. Thus, according to Peumans and Damme [39], Gm-TX could be classified as a merolectin since it has apparently a solecarbohydrate-binding site in its structure.
Gm-TX was highly toxic to mice (LD50 4.5 mg/kg) by i.p. route,producing dyspnoea, convulsions and death. These clinicalsymptoms were similar to those observed for SYTX (LD50 7–8 mg/kg) [12] and SBTX (LD50 5.6 mg/kg) [13]. Gm-TX is morepotent than the other soybean neurotoxins either because of itslower LD50 or the higher number and intensity of convulsions itprovoked. Conversely, there were no acute symptoms when Gm-TXwas administered intragastrically to mice, even at doses 10-foldhigher than the LD50 obtained by i.p. route.
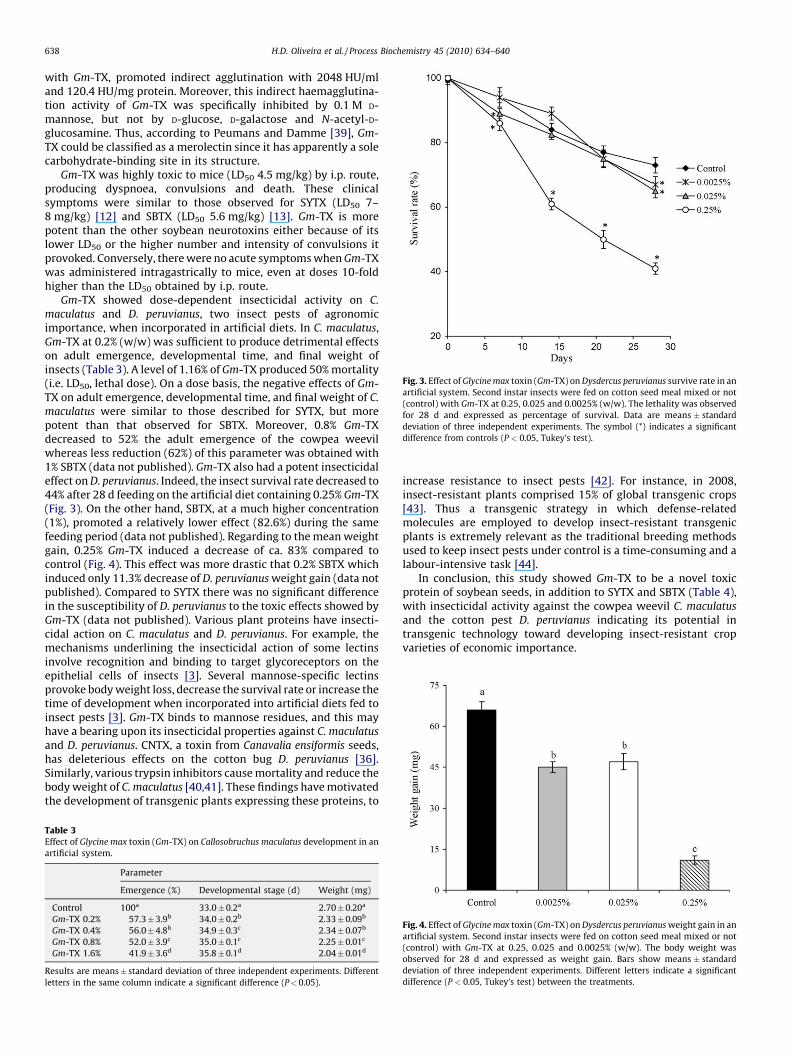
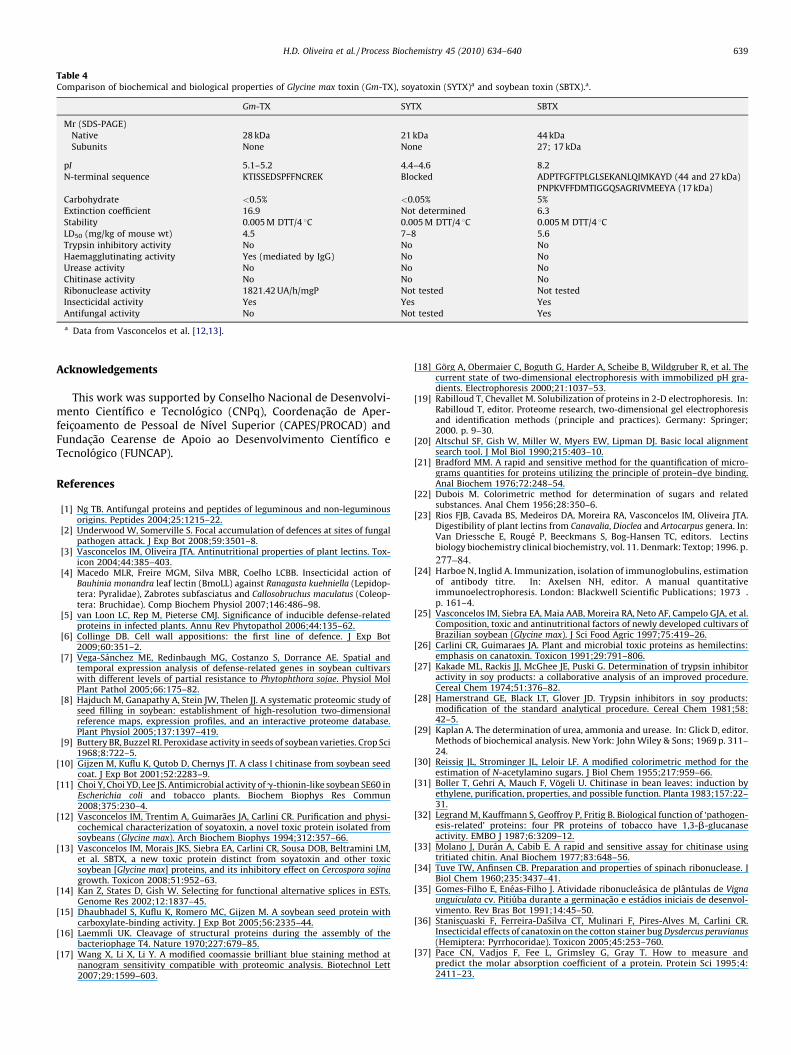
Gm-TX showed dose-dependent insecticidal activity on C.maculatus and D. peruvianus, two insect pests of agronomicimportance, when incorporated in artificial diets. In C. maculatus,Gm-TX at 0.2% (w/w) was sufficient to produce detrimental effectson adult emergence, developmental time, and final weight ofinsects (Table 3). A level of 1.16% of Gm-TX produced 50% mortality(i.e. LD50, lethal dose). On a dose basis, the negative effects of Gm-TX on adult emergence, developmental time, and final weight of C.maculatus were similar to those described for SYTX, but morepotent than that observed for SBTX. Moreover, 0.8% Gm-TXdecreased to 52% the adult emergence of the cowpea weevilwhereas less reduction (62%) of this parameter was obtained with1% SBTX (data not published). Gm-TX also had a potent insecticidaleffect on D. peruvianus. Indeed, the insect survival rate decreased to44% after 28 d feeding on the artificial diet containing 0.25% Gm-TX(Fig. 3). On the other hand, SBTX, at a much higher concentration(1%), promoted a relatively lower effect (82.6%) during the samefeeding period (data not published). Regarding to the mean weightgain, 0.25% Gm-TX induced a decrease of ca. 83% compared tocontrol (Fig. 4). This effect was more drastic that 0.2% SBTX whichinduced only 11.3% decrease of D. peruvianus weight gain (data notpublished). Compared to SYTX there was no significant differencein the susceptibility of D. peruvianus to the toxic effects showed byGm-TX (data not published). Various plant proteins have insecti-cidal action on C. maculatus and D. peruvianus. For example, themechanisms underlining the insecticidal action of some lectinsinvolve recognition and binding to target glycoreceptors on theepithelial cells of insects [3]. Several mannose-specific lectinsprovoke body weight loss, decrease the survival rate or increase thetime of development when incorporated into artificial diets fed toinsect pests [3]. Gm-TX binds to mannose residues, and this mayhave a bearing upon its insecticidal properties against C. maculatusand D. peruvianus. CNTX, a toxin from Canavalia ensiformis seeds,has deleterious effects on the cotton bug D. peruvianus [36].Similarly, various trypsin inhibitors cause mortality and reduce thebody weight of C. maculatus [40,41]. These findings have motivatedthe development of transgenic plants expressing these proteins, to
increase resistance to insect pests [42]. For instance, in 2008,insect-resistant plants comprised 15% of global transgenic crops[43]. Thus a transgenic strategy in which defense-relatedmolecules are employed to develop insect-resistant transgenicplants is extremely relevant as the traditional breeding methodsused to keep insect pests under control is a time-consuming and alabour-intensive task [44].
In conclusion, this study showed Gm-TX to be a novel toxicprotein of soybean seeds, in addition to SYTX and SBTX (Table 4),with insecticidal activity against the cowpea weevil C. maculatusand the cotton pest D. peruvianus indicating its potential intransgenic technology toward developing insect-resistant cropvarieties of economic importance.
Table 3Effect of Glycine max toxin (Gm-TX) on Callosobruchus maculatus development in anartificial system.
Parameter
Emergence (%) Developmental stage (d) Weight (mg)
Control 100a 33.0! 0.2a 2.70!0.20a
Gm-TX 0.2% 57.3!3.9b 34.0! 0.2b 2.33!0.09b
Gm-TX 0.4% 56.0!4.8b 34.9! 0.3c 2.34!0.07b
Gm-TX 0.8% 52.0!3.9c 35.0! 0.1c 2.25!0.01c
Gm-TX 1.6% 41.9!3.6d 35.8! 0.1d 2.04!0.01d
Results are means! standard deviation of three independent experiments. Differentletters in the same column indicate a significant difference (P< 0.05).
Fig. 3. Effect of Glycine max toxin (Gm-TX) on Dysdercus peruvianus survive rate in anartificial system. Second instar insects were fed on cotton seed meal mixed or not(control) with Gm-TX at 0.25, 0.025 and 0.0025% (w/w). The lethality was observedfor 28 d and expressed as percentage of survival. Data are means ! standarddeviation of three independent experiments. The symbol (*) indicates a significantdifference from controls (P < 0.05, Tukey’s test).
Fig. 4. Effect of Glycine max toxin (Gm-TX) on Dysdercus peruvianus weight gain in anartificial system. Second instar insects were fed on cotton seed meal mixed or not(control) with Gm-TX at 0.25, 0.025 and 0.0025% (w/w). The body weight wasobserved for 28 d and expressed as weight gain. Bars show means ! standarddeviation of three independent experiments. Different letters indicate a significantdifference (P < 0.05, Tukey’s test) between the treatments.
H.D. Oliveira et al. / Process Biochemistry 45 (2010) 634–640638
Acknowledgements
This work was supported by Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq), Coordenacao de Aper-feicoamento de Pessoal de Nıvel Superior (CAPES/PROCAD) andFundacao Cearense de Apoio ao Desenvolvimento Cientıfico eTecnologico (FUNCAP).
References
[1] Ng TB. Antifungal proteins and peptides of leguminous and non-leguminousorigins. Peptides 2004;25:1215–22.
[2] Underwood W, Somerville S. Focal accumulation of defences at sites of fungalpathogen attack. J Exp Bot 2008;59:3501–8.
[3] Vasconcelos IM, Oliveira JTA. Antinutritional properties of plant lectins. Tox-icon 2004;44:385–403.
[4] Macedo MLR, Freire MGM, Silva MBR, Coelho LCBB. Insecticidal action ofBauhinia monandra leaf lectin (BmoLL) against Ranagasta kuehniella (Lepidop-tera: Pyralidae), Zabrotes subfasciatus and Callosobruchus maculatus (Coleop-tera: Bruchidae). Comp Biochem Physiol 2007;146:486–98.
[5] van Loon LC, Rep M, Pieterse CMJ. Significance of inducible defense-relatedproteins in infected plants. Annu Rev Phytopathol 2006;44:135–62.
[6] Collinge DB. Cell wall appositions: the first line of defence. J Exp Bot2009;60:351–2.
[7] Vega-Sanchez ME, Redinbaugh MG, Costanzo S, Dorrance AE. Spatial andtemporal expression analysis of defense-related genes in soybean cultivarswith different levels of partial resistance to Phytophthora sojae. Physiol MolPlant Pathol 2005;66:175–82.
[8] Hajduch M, Ganapathy A, Stein JW, Thelen JJ. A systematic proteomic study ofseed filling in soybean: establishment of high-resolution two-dimensionalreference maps, expression profiles, and an interactive proteome database.Plant Physiol 2005;137:1397–419.
[9] Buttery BR, Buzzel RI. Peroxidase activity in seeds of soybean varieties. Crop Sci1968;8:722–5.
[10] Gijzen M, Kuflu K, Qutob D, Chernys JT. A class I chitinase from soybean seedcoat. J Exp Bot 2001;52:2283–9.
[11] Choi Y, Choi YD, Lee JS. Antimicrobial activity of g-thionin-like soybean SE60 inEscherichia coli and tobacco plants. Biochem Biophys Res Commun2008;375:230–4.
[12] Vasconcelos IM, Trentim A, Guimaraes JA, Carlini CR. Purification and physi-cochemical characterization of soyatoxin, a novel toxic protein isolated fromsoybeans (Glycine max). Arch Biochem Biophys 1994;312:357–66.
[13] Vasconcelos IM, Morais JKS, Siebra EA, Carlini CR, Sousa DOB, Beltramini LM,et al. SBTX, a new toxic protein distinct from soyatoxin and other toxicsoybean [Glycine max] proteins, and its inhibitory effect on Cercospora sojinagrowth. Toxicon 2008;51:952–63.
[14] Kan Z, States D, Gish W. Selecting for functional alternative splices in ESTs.Genome Res 2002;12:1837–45.
[15] Dhaubhadel S, Kuflu K, Romero MC, Gijzen M. A soybean seed protein withcarboxylate-binding activity. J Exp Bot 2005;56:2335–44.
[16] Laemmli UK. Cleavage of structural proteins during the assembly of thebacteriophage T4. Nature 1970;227:679–85.
[17] Wang X, Li X, Li Y. A modified coomassie brilliant blue staining method atnanogram sensitivity compatible with proteomic analysis. Biotechnol Lett2007;29:1599–603.
[18] Gorg A, Obermaier C, Boguth G, Harder A, Scheibe B, Wildgruber R, et al. Thecurrent state of two-dimensional electrophoresis with immobilized pH gra-dients. Electrophoresis 2000;21:1037–53.
[19] Rabilloud T, Chevallet M. Solubilization of proteins in 2-D electrophoresis. In:Rabilloud T, editor. Proteome research, two-dimensional gel electrophoresisand identification methods (principle and practices). Germany: Springer;2000. p. 9–30.
[20] Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignmentsearch tool. J Mol Biol 1990;215:403–10.
[21] Bradford MM. A rapid and sensitive method for the quantification of micro-grams quantities for proteins utilizing the principle of protein–dye binding.Anal Biochem 1976;72:248–54.
[22] Dubois M. Colorimetric method for determination of sugars and relatedsubstances. Anal Chem 1956;28:350–6.
[23] Rios FJB, Cavada BS, Medeiros DA, Moreira RA, Vasconcelos IM, Oliveira JTA.Digestibility of plant lectins from Canavalia, Dioclea and Artocarpus genera. In:Van Driessche E, Rouge P, Beeckmans S, Bog-Hansen TC, editors. Lectinsbiology biochemistry clinical biochemistry, vol. 11. Denmark: Textop; 1996. p.
277–84.[24] Harboe N, Inglid A. Immunization, isolation of immunoglobulins, estimation
of antibody titre. In: Axelsen NH, editor. A manual quantitativeimmunoelectrophoresis. London: Blackwell Scientific Publications; 1973 .p. 161–4.
[25] Vasconcelos IM, Siebra EA, Maia AAB, Moreira RA, Neto AF, Campelo GJA, et al.Composition, toxic and antinutritional factors of newly developed cultivars ofBrazilian soybean (Glycine max). J Sci Food Agric 1997;75:419–26.
[26] Carlini CR, Guimaraes JA. Plant and microbial toxic proteins as hemilectins:emphasis on canatoxin. Toxicon 1991;29:791–806.
[27] Kakade ML, Rackis JJ, McGhee JE, Puski G. Determination of trypsin inhibitoractivity in soy products: a collaborative analysis of an improved procedure.Cereal Chem 1974;51:376–82.
[28] Hamerstrand GE, Black LT, Glover JD. Trypsin inhibitors in soy products:modification of the standard analytical procedure. Cereal Chem 1981;58:42–5.
[29] Kaplan A. The determination of urea, ammonia and urease. In: Glick D, editor.Methods of biochemical analysis. New York: John Wiley & Sons; 1969. p. 311–24.
[30] Reissig JL, Strominger JL, Leloir LF. A modified colorimetric method for theestimation of N-acetylamino sugars. J Biol Chem 1955;217:959–66.
[31] Boller T, Gehri A, Mauch F, Vogeli U. Chitinase in bean leaves: induction byethylene, purification, properties, and possible function. Planta 1983;157:22–31.
[32] Legrand M, Kauffmann S, Geoffroy P, Fritig B. Biological function of ‘pathogen-esis-related’ proteins: four PR proteins of tobacco have 1,3-b-glucanaseactivity. EMBO J 1987;6:3209–12.
[33] Molano J, Duran A, Cabib E. A rapid and sensitive assay for chitinase usingtritiated chitin. Anal Biochem 1977;83:648–56.
[34] Tuve TW, Anfinsen CB. Preparation and properties of spinach ribonuclease. JBiol Chem 1960;235:3437–41.
[35] Gomes-Filho E, Eneas-Filho J. Atividade ribonucleasica de plantulas de Vignaunguiculata cv. Pitiuba durante a germinacao e estadios iniciais de desenvol-vimento. Rev Bras Bot 1991;14:45–50.
[36] Staniscuaski F, Ferreira-DaSilva CT, Mulinari F, Pires-Alves M, Carlini CR.Insecticidal effects of canatoxin on the cotton stainer bug Dysdercus peruvianus(Hemiptera: Pyrrhocoridae). Toxicon 2005;45:253–760.
[37] Pace CN, Vadjos F, Fee L, Grimsley G, Gray T. How to measure andpredict the molar absorption coefficient of a protein. Protein Sci 1995;4:2411–23.
Table 4Comparison of biochemical and biological properties of Glycine max toxin (Gm-TX), soyatoxin (SYTX)a and soybean toxin (SBTX).a.
Gm-TX SYTX SBTX
Mr (SDS-PAGE)Native 28 kDa 21 kDa 44 kDaSubunits None None 27; 17 kDa
pI 5.1–5.2 4.4–4.6 8.2N-terminal sequence KTISSEDSPFFNCREK Blocked ADPTFGFTPLGLSEKANLQIMKAYD (44 and 27 kDa)
PNPKVFFDMTIGGQSAGRIVMEEYA (17 kDa)Carbohydrate <0.5% <0.05% 5%Extinction coefficient 16.9 Not determined 6.3Stability 0.005 M DTT/4 8C 0.005 M DTT/4 8C 0.005 M DTT/4 8CLD50 (mg/kg of mouse wt) 4.5 7–8 5.6Trypsin inhibitory activity No No NoHaemagglutinating activity Yes (mediated by IgG) No NoUrease activity No No NoChitinase activity No No NoRibonuclease activity 1821.42 UA/h/mgP Not tested Not testedInsecticidal activity Yes Yes YesAntifungal activity No Not tested Yes
a Data from Vasconcelos et al. [12,13].
H.D. Oliveira et al. / Process Biochemistry 45 (2010) 634–640 639

[38] Sels J, Mathys J, Coninck BMA, Cammue BPA, Bolle MFC. Plant pathogenesis-related (PR) proteins: a focus on PR peptides. Plant Physiol Biochem 2008;46:941–50.
[39] Peumans WJ, Damme EJM. Lectin as plant defense proteins. Plant Physiol1995;109:347–52.
[40] Araujo CL, Bezerra IWL, Oliveira AS, Moura FT, Macedo LLP, Gomes CEM,et al. In vivo bioinsecticidal activity toward Ceratitis capitata (fruit fly) andCallosobruchus maculatus (cowpea weevil) and in vitro bioinsecticidal ac-tivity toward different orders of insect pests of a trypsin inhibitor purifiedfrom tamarind tree (Tamarindus indica) seeds. J Agric Food Chem 2005;53:4381–7.
[41] Gomes CEM, Barbosa AEAD, Macedo LLP, Pitanga JCM, Moura FT, Oliveira AS,et al. Effects of a trypsin inhibitor from Crotalaria pallida seeds on Calloso-bruchus maculatus (cowpea weevil) and Ceratitis capitata (fruit fly). PlantPhysiol Biochem 2005;43:1095–102.
[42] Babu RM, Sajeena A, Seetharaman K, Reddy MS. Advances in geneticallyengineered (transgenic) plants in insect management—an overview. Crop Prot2003;22:1071–86.
[43] James C. Global status of commercialized biotech/GM crops: 2008. ISAAABriefs; 2008. p. 39.
[44] Kos M, van Loon JJA, Dicke M, Vet LEM. Transgenic plants as vital componentsof integrated pest management. Trends Biotechnol 2009;27:621–7.
H.D. Oliveira et al. / Process Biochemistry 45 (2010) 634–640640




















