
Gestione e sfruttamento delle risorse faunistiche nei siti di Vallone Inferno (PA) e Case Bastione...
165
UNIVERSITÀ DEGLI STUDI DI FERRARA UNIVERSITÀ DEGLI STUDI DI MODENA E REGGIO EMILIA Corso di Laurea Magistrale in QUATERNARIO, PREISTORIA E ARCHEOLOGIA (Classe LM 2 –Archeologia) Gestione e sfruttamento delle risorse faunistiche nei siti di Vallone Inferno (PA) e Case Bastione (EN). Relatore: Dott. ssa URSULA THUN HOHENSTEIN Correlatori: Dott. ssa DARIA PETRUSO Dott. ssa VINCENZA FORGIA Prof. OSCAR BELVEDERE Laureando: GIOVANNI DI SIMONE ___________________________________ Anno Accademico 2013-2014
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Gestione e sfruttamento delle risorse faunistiche nei siti di Vallone Inferno (PA) e Case Bastione...

UNIVERSITÀ DEGLI STUDI DI FERRARA
UNIVERSITÀ DEGLI STUDI DI MODENA E REGGIO
EMILIA
Corso di Laurea Magistrale in QUATERNARIO, PREISTORIA E
ARCHEOLOGIA (Classe LM 2 –Archeologia)
Gestione e sfruttamento delle risorse faunistiche nei siti di Vallone Inferno
(PA) e Case Bastione (EN).
Relatore: Dott. ssa URSULA THUN HOHENSTEIN Correlatori: Dott. ssa DARIA PETRUSO Dott. ssa VINCENZA FORGIA Prof. OSCAR BELVEDERE
Laureando: GIOVANNI DI SIMONE
___________________________________
Anno Accademico 2013-2014


1
INDICE
INTRODUZIONE p. 7
Capitolo 1 “Il riparo sotto roccia di Vallone Inferno”
1.1 Introduzione p. 9
1.2 Inquadramento geografico, geologico e geomorfologico p. 9
1.3 Notizie storiche della frequentazione dell’area p. 10
1.4 Le campagne di scavo p. 11
1.4.1 La sezione indagata p. 11
1.5 La fauna a mammiferi delle campagne 2008-2011 p. 15
1.6 I resti umani p. 17
1.7 Ceramica p. 17
1.8 Industria litica p. 19
1.9 Archeobotanica p. 20
Capitolo 2 “Il villaggio di Case Bastione”
2.1 Introduzione p. 21
2.2 Inquadramento geografico, geologico e geomorfologico p. 21
2.3 Le campagne di scavo p. 22
2.4 Le aree indagate p. 22
2.5 La fase del Bronzo antico dell’Area β p. 24
2.6 La fauna a mammiferi dell’età del Bronzo p. 27
2.7 Ceramica p. 29
Capitolo 3 “Metodologia: l’analisi archeozoologica”
3.1 Introduzione p. 31
3.2 L’identificazione anatomica e tassonomica p. 31
3.2.1 Classificazione per categoria p. 33
3.2.2 Classificazione per taglia p. 34

2
3.2.3 Classificazione per porzione e parte p. 35
3.3 Analisi quantitative p. 36
3.3.1 NR “Numero dei Resti” p. 37
3.3.2 NME “Numero minimo degli Elementi” p. 37
3.3.3 NEE “Numero Minimo degli Elementi Aspettati” p. 38
3.3.4 NMI “Numero Minimo di Individui” p. 38
3.4 Calcolo dell’età di morte p. 38
3.4.1 Calcolo dell’età di morte degli ovicaprini p. 39
3.4.2 Calcolo dell’età di morte dei suini p. 40
3.4.3 Calcolo dell’età di morte del Cervus elaphus p. 42
3.5 Misure osteometriche p. 44
Capitolo 4 “Metodologia: l’analisi tafonomica”
4.1 Introduzione p. 47
4.2 Tracce di origine naturale p. 47
4.2.1 Alterazione prodotta da carnivori p. 48
4.2.2 Alterazioni prodotte da roditori p. 48
4.2.3 Calpestio p. 49
4.2.4 Alterazione prodotta da radici p. 50
4.2.5 Ossido di manganese p. 51
4.2.6 Weathering p. 51
4.3 Tracce di origine antropica p. 52
4.3.1 Le tracce di macellazione p. 52
4.3.2 Fratture p. 57
4.3.3 Bollitura p. 58
4.3.4 Combustione p. 59
Capitolo 5 “Registrazione dei dati”
5.1 Introduzione p. 63

3
5.2 Utilizzo di un database p. 63
5.3 Registrazione dei dati p. 63
Capitolo 6 “Il campione faunistico di Vallone Inferno”
6.1 Introduzione p. 69
6.2 La fauna analizzata p. 69
6.3 Ovicaprini p. 73
6.4 Ovis aries Linnaeus, 1758 & Capra hircus Linnaeus, 1758 p. 78
6.5 Sus domesticus Erxleben, 1777 p. 79
6.6 Bos taurus Linnaeus, 1758 p. 80
6.7 Cervus elaphus Linnaeus, 1758 p. 82
6.8 Carnivori p. 83
Capitolo 7 “Analisi tafonomiche di Vallone Inferno”
7.1 Introduzione p. 85
7.2 Tracce di origine naturale p. 85
7.2.1 Tracce di radici p. 86
7.2.2 Tracce di ossido di manganese p. 87
7.2.3 Tracce di rosicature di carnivori e roditori p. 87
7.2.4 Calpestio p. 88
7.2.5 Tracce di weathering p. 89
7.3 Tracce di origine antropica p. 89
7.3.1 Tracce di combustione p. 90
7.3.2 Tracce di bollitura p. 91
7.3.3 Fratture antropiche p. 92
7.3.4 Tracce di macellazione p. 92
Capitolo 8 “Il campione faunistico di Case Bastione”
8.1 Introduzione p. 95

4
8.2 La fauna analizzata p. 95
8.3 Ovicaprini p. 99
8.4 Sus domesticus Erxleben, 1777 p. 104
8.5 Bos taurus Linnaeus, 1758 p. 105
8.6 Canis familiaris Linnaeus, 1758 p. 106
8.7 Cervus elaphus Linnaeus, 1758 p. 107
Capitolo 9 “Analisi tafonomiche di Case Bastione”
9.1 Introduzione p. 109
9.2 Tracce di origina naturale p. 109
9.2.1 Tracce di radici p. 110
9.2.2 Tracce di ossido di manganese p. 111
9.2.3 Tracce di rosicature di carnivori e roditori p. 111
9.2.4 Calpestio p. 113
9.2.5 Tracce di weathering p. 113
9.3 Tracce di origine antropica p. 114
9.3.1 Tracce di combustione p. 115
9.3.2 Tracce di bollitura p. 116
9.3.3 Fratture antropiche p. 116
9.3.4 Tracce di macellazione p. 117
Capitolo 10 “Analisi archeozoologiche, Vallone Inferno e Case
Bastione a confronto”
10.1 Introduzione p. 119
10.2 I campioni faunistici a confronto p. 119
10.2.1 Taxa riconosciuti e strategie economiche a confronto p. 122
Capitolo 11 “Analisi tafonomiche, Vallone Inferno e Case
Bastione a confronto”
11.1 Introduzione p. 125

5
11.2 Tracce di origine naturale, confronto tra VI e CBS p. 125
11.3 Tracce di origine antropiche, confronto tra VI e CBS p. 126
CONCLUSIONI p. 129
BIBLIOGRAFIA p. 133
APPENDICE p. 141
TABELLE OSTEOMETRICHE p. 143
TAVOLE p. 150
RINGRAZIAMENTI p. 163

6

7
INTRODUZIONE
Nel presente elaborato è stato affrontato lo studio archeozoologico e tafonomico dei resti
faunistici provenienti dai livelli dell’età del Bronzo Antico del riparo sotto roccia di
Vallone Inferno (PA) e dal villaggio di Case Bastione (EN), entrambi situati nella regione
Sicilia. La finalità principale di questo lavoro è quello di mettere a confronto i due
campioni faunistici, appartenenti a due siti cronologicamente simili ma tipologicamente
diversi, infatti Vallone Inferno è un riparo sotto roccia e Case Bastione, essendo un
villaggio, costituisce un contesto all’aperto. A tale scopo sono state ricostruite le modalità
di gestione e sfruttamento delle faune domestiche e selvatiche, tramite le analisi
archeozoologiche e tafonomiche.
Nei Capitoli 1 e 2 vengono forniti gli inquadramenti generali dei due siti archeologici
oggetto di studio di questo lavoro, ponendo l’attenzione sugli aspetti geografici, geologici,
geomorfologici, frequentazione storica dell’area, le campagne di scavo, le sezioni e le aree
indagate. Inoltre, verrà effettuata anche una descrizione delle varie tipologie di reperti
archeologici rinvenuti (reperti archeofaunistici, ossa umane, industria litica, ceramica e
resti archeobotanici).
Nel Capitolo 3 sono state esposte le metodologie che riguardano le analisi
archeozoologiche, utilizzate per lo studio dei due insiemi faunistici (riconoscimento
anatomico e tassonomico, le analisi quantitative e il calcolo dell’età di morte).
Il Capitolo 4 è stato dedicato alle metodologie delle analisi tafonomiche, ponendo
l’attenzione sulle tracce di origine naturale e antropiche che sono state riconosciute durante
lo studio dei reperti.
Il Capitolo 5 riguarda le modalità di registrazione dei dati. Per questo lavoro è stato
utilizzato il database realizzato tramite il programma FileMaker Pro Advanced, adoperato
dal gruppo di ricerca dell’IPHES/Università Rovina i Virgili di Tarragona (Spagna). Inoltre
è stata sottolineata l’importanza dell’utilizzo di un database per l’analisi di reperti
archeofaunistici.
I Capitoli 6 e 7 sono dedicati alla presentazione dei delle analisi archeozoologiche e
tafonomiche del campione faunistico di Vallone Inferno; nel capitolo 6 i taxa riconosciuti
vengono presentati per ordine di abbondanza.
I Capitoli 8 e 9 sono stati riservati alla presentazione dei dati delle analisi archeozoologiche
e tafonomiche del sito di Case Bastione, anche in questo caso nel capitolo 8 i taxa sono
stati presentati per ordine di abbondanza.

8
I Capitoli 10 e 11 sono stati dedicati invece ai confronti dei due insiemi faunistici per
evidenziare le differenze nella gestione e nello sfruttamento delle risorse faunistiche.
Infine vengono poste le conclusioni, per riassumere le interpretazioni e le considerazioni
riscontrate nel corso dell’analisi dei reperti.

9
Capitolo 1 “Il riparo sotto roccia di Vallone Inferno”
1.1 Introduzione
Il presente capitolo fornisce un inquadramento esaustivo del sito preistorico di Vallone
Inferno presso Scillato (PA). Per meglio comprendere il contesto ambientale in cui il sito è
ubicato viene data una breve descrizione topografica del territorio circostante nonché della
morfologia e litologia delle rocce affioranti nell’area. Infine, viene fornita una descrizione
stratigrafica del deposito e dei materiali archeologici e paleobiologici ivi recuperati,
durante le campagne di scavo condotte dal 2008 al 2014, alle quali ha partecipato anche lo
scrivente.
1.2 Inquadramento geografico, geologico e geomorfologico
Il riparo sotto roccia di Vallone Inferno si trova nel territorio del comune di Scillato (PA),
situato nella porzione settentrionale della catena montuosa delle Madonie. Tale area è
compresa topograficamente nel Parco delle Madonie, che si estende tra le valli dei fiumi
Torto ad Ovest e del fiume Pollina ad Est; a Sud vi è il confine amministrativo con il
comune di Caltanissetta, mentre a Nord confina con il Mar Tirreno. Il sito, in particolare,
ricade nella tavoletta I.G.M. 1: 25.000 Scillato foglio 259 quadrante I SE ed è posto ad un
altitudine di circa 770 m s.l.m.
Dal punto di vista topografico il sito si trova all’interno di una valle di formazione carsico-
fluviale, chiamata appunto Vallone dell’Inferno o Fosso Inferno, che si apre nel pendio
nord di Monte dei Cervi, il cui andamento segue il margine nord-ovest di Monte Fanusi
(Figg. 1.1-1.2). Il riparo si trova in una posizione strategica, infatti collega la valle
alluvionale del fiume Imera alla cima della montagna e risulta essere una naturale via di
comunicazione tra le basse e le alte Madonie; è proprio la sua posizione strategica che deve
aver attirato in passato le popolazioni umane che ivi si stanziarono utilizzandolo come
crocevia per i pascoli (Forgia et al 2012).
Il variegato aspetto geomorfologico dell’area si deve alla differente litologia degli
affioramenti costituiti da rocce prevalentemente carbonatiche di età Mesozoica che
costituiscono i massicci montuosi di Monte dei Cervi - Monte Fanusi, e da argille e
arenarie del Flysh Numidico di età Cenozoica che formano i prospicenti dolci pendii.
(Forgia et al 2012).

10
Il riparo si trova nel lato sinistro del Vallone Inferno (Fig 1.2) e si sviluppa per un’altezza
di poco più di 10 m, per una ampiezza di 6 m ed una profondità di 5 m; al di sopra del
deposito la volta del riparo aggetta per altri 4 m (Forgia et al 2012).
Fig. 1.1: Veduta del Vallone dell’Inferno. Documentazione fotografica di scavo per gentile concessione
dott.ssa Vincenza Forgia.
Fig. 1.2: Veduta schematica dei Monti dei Cervi e Fanusi con la localizzazione del sito (Forgia et al. 2012,
pag. 3, fig. 1).
1.3 Notizie storiche della frequentazione dell’area
Il territorio delle Madonie è stato esplorato da gruppi umani sin dall’epoca preistorica. Le
ricerche e le fonti paleoantropologiche e archeologiche hanno mostrato che l’area delle
Madonie è stata frequentata a partire dal Paleolitico Superiore, anche se attraverso notizie
di tipo sporadiche (Minà Palumbo, 1869; Baldini et al., 1976). Una frequentazione più
certa si ha a partire dalla fine del Neolitico al Bronzo antico, attestata dai ritrovamenti

11
presso la grotta del Vecchiuzzo situata a 800 m s.l.m. (Bovio Marconi, 1979). La prima
antropizzazione di questa grotta risale al Neolitico Tardo ed è testimoniata da frammenti
della facies Diana (4000-3400 BC). L’abbandono della grotta sembra coincidere con
l’inizio dell’Antica Età del Bronzo, attestata da pezzi sporadici della facies di Castelluccio
(2200-1400 BC) (Tusa, 1999). Reperti di età del Rame e del Bronzo sono stati invece
documentati nelle grotte di Chiusilla, del Fico e dell’Abisso del Vento, nella valle del
fiume Isnello (Giuffrida Ruggeri, 1901-1902; 1905; Mannino, 1989).
1.4. Le campagne di scavo
Il sito archeologico di Vallone Inferno è stato scoperto nel 2007 nell’ambito di un progetto
di ricognizione di emergenze preistoriche portato avanti dalla dottoressa Vincenza Forgia e
dal prof. Oscar Belvedere dell’Università degli Studi di Palermo in collaborazione con il
prof. Andreu Ollé e il prof. Josep Maria Vergès dell’Iphes/Università Rovina i Virgili di
Tarragona (Spagna). Il sito, al momento del suo ritrovamento ha mostrato subito grandi
potenzialità; lungo la sezione del deposito di circa 5 m, esposto dall’erosione del torrente
Inferno, spiccavano alla vista alcuni frammenti ceramici (ceramica d’impasto preistorica),
resti faunistici e due denti umani. Come conseguenza dal 2008 sono iniziate le campagne
di scavo del suddetto gruppo di ricerca che si sono protratte fino al 2014, in collaborazione
con la Soprintendenza di Palermo.
Dai dati raccolti durante la prima campagna di scavo si è evinto che il deposito documenta
una frequentazione che va dall’età preistorica, in particolare dal Neolitico Medio, come
testimoniano i reperti di ceramica tricromica, fino all’epoca moderna. Negli anni successivi
lo scavo è stato esteso coprendo tutta la superficie del riparo ed i risultati, oltre a
confermare la frequentazione neolitica, hanno esteso la frequentazione ad altri periodi
preistorici (età del Bronzo antico come attestato da ceramica castellucciana) e storici (sulla
base di frammenti ceramici del VII sec. d.C.) (Forgia et al 2012). La cronologia relativa di
queste due nuove fasi sono confermate dalle datazioni al radiocarbonio riassunte in tabella
1.1.
1.4.1 La sezione indagata
La stratigrafia è stata riconosciuta e dettagliata dal prof. Diego E. Angelucci, del
Dipartimento di Filosofia, Storia e Beni Culturali dell’Università degli Studi di Trento.
L’intero giacimento è stato suddiviso in 4 complessi stratigrafici principali e taluni di
questi a loro volta distinti in sub-complessi, seguendo un criterio di tipo litologico-

12
sedimentario. Sono stati utilizzati dei numeri per codificare i complessi e i sub-complessi e
delle lettere per indicare le suddivisioni minori. I quattro complessi qui di seguito elencati,
dal più alto al più basso, sono distinti utilizzando le diverse caratteristiche litologiche e
datati relativamente sulla base dell’analisi dei reperti culturali (Forgia et al 2012):
Complesso 1: struttura o muro di pietra datato in epoca moderna; probabilmente un
piccolo riparo provvisorio di un pastore (Figg. 1.3 – 1.4);
Fig. 1.3 - Veduta della parte
superiore del deposito in corso di
scavo. Sulla destra il livello di
frequentazione sub-attuale - 1.2 - con
resti di struttura a secco. Sulla sinistra
il saggio esplorativo del 2008.
Documentazione fotografica per
gentile concessione dott.ssa Vincenza
Forgia.
Complesso 2: grosso accumulo di sedimento, spesso 2.5 m, affiora nel lato est
magnetico del sito, costituito prevalentemente da brecce calcaree, non presenta nessuna
evidenza archeologica, si definisce perciò sterile. In cima sono presenti grossi massi
scarsamente stratificati, in basso frammenti spigolosi di pietra calcarea, a cui si aggiungono
occasionali elementi di selce e sedimento argilloso (Fig. 1.4);
Complesso 3: questo complesso ha restituito la maggior parte del materiale
archeologico rinvenuto, ed è caratterizzato dall’alternanza di strati a granulometria fine e
grossolana. La successione archeologica del Complesso 3 si divide in 4 sub-complessi, che
vengono di seguito elencati dall’alto verso il basso (Fig. 1.4):
- Sub-complesso 3.1, costituito da cenere e da strati termo-alterati, alternati da strati
con pietre grossolane e soprattutto con materiali archeologici di età tardo-antica;
- Sub-complesso 3.2, include materiale archeologico di età tardo-antica;
- Sub-complesso 3.3, costituito da strati organici e termo-alterati, alternati con strati
grossolani, inoltre contiene reperti di età tardo-antica;
- Sub-complesso 3.4, è costituito da un alternanza di strati grossolani e fini,
restituisce reperti archeologici risalenti all’età del Bronzo antico datato al 2200-1400 a.C.
(Tusa, 1999) e al Neolitico Medio compreso tra il 4500 e il 4000 a.C. (Tusa, 1999). La

13
commistione di reperti ceramici risalenti al Bronzo antico (facies Castelluccio), e di reperti
caratteristici del Neolitico Medio (ceramica tricromica) si spiega analizzando questo sub-
complesso dal punto di vista delle dinamiche di sedimentazione ed erosione. La più antica
deposizione doveva risalire alla frequentazione neolitica del riparo, cui segue una fase
erosiva che ha asportato la maggior parte di questo livello e anche la parte superiore del
sottostante Complesso 4; l’unico lembo testimone di questa frequentazione neolitica (sub-
complesso 3.4. N; Fig. 1.5) si trova nella parete di fondo del riparo. Infine un nuovo
episodio di sedimentazione testimonia la frequentazione durante il Bronzo antico (sub-
complesso 3.4.) a cui si uniscono i reperti neolitici portati a giorno dall’erosione (Fig. 1.5).
Complesso 4: tale complesso, spesso circa 2 m, si presenta scarsamente stratificato.
E’ costituito da brecce calcaree, moderatamente cementate da carbonato di calcio, formate
da frammenti decimetrici a spigoli vivi immersi entro una matrice argilloso-limosa di
colore marrone. Nonostante siano stati trovati resti di micromammiferi e campioni
pollinici, questo complesso non è ancora stato scavato, e non ci sono altre informazioni
oltre le datazioni riportate nella tabella 1.1.
Sono stati selezionati cinque campioni dall’intera sequenza archeologica destinati alle
datazioni al radiocarbonio: quattro campioni provengono dal complesso 3 e sono stati
analizzati al Circe Innova Lab; il quinto campione proviene dal Complesso 4 ed è stato
analizzato da Beta analytic Inc (Tab. 1.1). Le calibrazioni per tutti i campioni sono state
calcolate usando il database INTCAL09 (Forgia et al. 2012).
Strato
archeologico
Campione Età Radiocarbonio
Convenzionale
(BP)
Età Radiocarbonio
Calibrata (AD/BC) (2
sigma, 95%)
3.1.l Resto vegetale 1260±34 669-828 AD
3.2.C Carbone di
legna
1332±26 649-714 AD
3.4.B Seme 3244±42 1616-1433 BC
3.4.G Osso umano 3948±35 2570-2310 BC
4.2 Carbone di
legna
9450 ± 50 9110 to 9090 BC
9040 to 9030 BC
8840 to 8620 BC
Tab. 1.1: Schema delle datazioni effettuate in alcuni campioni del deposito (Forgia et al. 2012).

14
Fig.1.4: Sezione indagata che evidenzia i 4 complessi litologici ed i sub-complessi (Forgia et al. 2012, pag.
4, fig. 2).
Fig. 1.5: Sezione schematica del deposito con interpretazione delle fasi di erosione e deposizione. In alto
orientazione del riparo e quadrettatura. Da (Forgia et al. 2012, pag. 5, fig. 3).
15
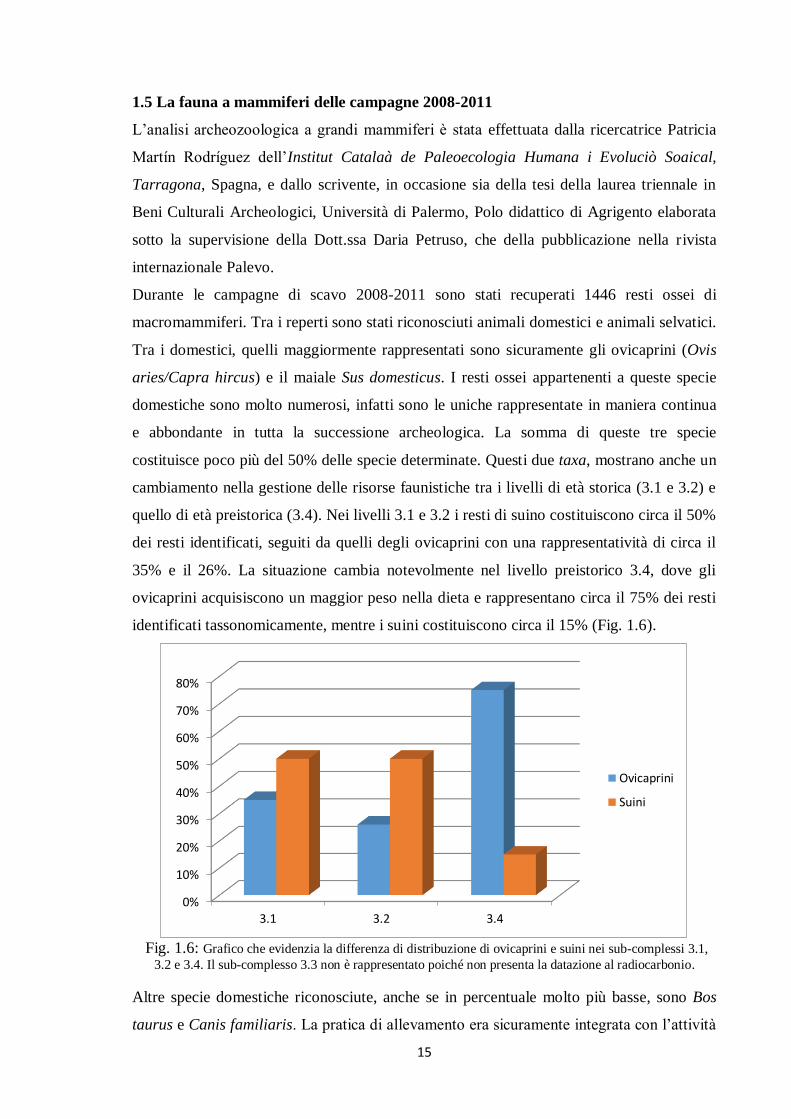
1.5 La fauna a mammiferi delle campagne 2008-2011
L’analisi archeozoologica a grandi mammiferi è stata effettuata dalla ricercatrice Patricia
Martín Rodríguez dell’Institut Catalaà de Paleoecologia Humana i Evoluciò Soaical,
Tarragona, Spagna, e dallo scrivente, in occasione sia della tesi della laurea triennale in
Beni Culturali Archeologici, Università di Palermo, Polo didattico di Agrigento elaborata
sotto la supervisione della Dott.ssa Daria Petruso, che della pubblicazione nella rivista
internazionale Palevo.
Durante le campagne di scavo 2008-2011 sono stati recuperati 1446 resti ossei di
macromammiferi. Tra i reperti sono stati riconosciuti animali domestici e animali selvatici.
Tra i domestici, quelli maggiormente rappresentati sono sicuramente gli ovicaprini (Ovis
aries/Capra hircus) e il maiale Sus domesticus. I resti ossei appartenenti a queste specie
domestiche sono molto numerosi, infatti sono le uniche rappresentate in maniera continua
e abbondante in tutta la successione archeologica. La somma di queste tre specie
costituisce poco più del 50% delle specie determinate. Questi due taxa, mostrano anche un
cambiamento nella gestione delle risorse faunistiche tra i livelli di età storica (3.1 e 3.2) e
quello di età preistorica (3.4). Nei livelli 3.1 e 3.2 i resti di suino costituiscono circa il 50%
dei resti identificati, seguiti da quelli degli ovicaprini con una rappresentatività di circa il
35% e il 26%. La situazione cambia notevolmente nel livello preistorico 3.4, dove gli
ovicaprini acquisiscono un maggior peso nella dieta e rappresentano circa il 75% dei resti
identificati tassonomicamente, mentre i suini costituiscono circa il 15% (Fig. 1.6).
Fig. 1.6: Grafico che evidenzia la differenza di distribuzione di ovicaprini e suini nei sub-complessi 3.1,
3.2 e 3.4. Il sub-complesso 3.3 non è rappresentato poiché non presenta la datazione al radiocarbonio.
Altre specie domestiche riconosciute, anche se in percentuale molto più basse, sono Bos
taurus e Canis familiaris. La pratica di allevamento era sicuramente integrata con l’attività
0%
10%
20%
30%
40%
50%
60%
70%
80%
3.1 3.2 3.4
Ovicaprini
Suini

16
di caccia, infatti gli animali selvatici recuperati sono soprattutto cervidi (cervi o daini) e
conigli.
Di seguito è riportata la Tabella (Tab. 1.2) con tutte le specie di macromammiferi che sono
state ritrovate all’interno del Complesso 3, poiché è quello che ha restituito il materiale
archeologico più significativo (Forgia et al 2012).
Macromammiferi 3.1 3.2 3.3 3.4
Bos taurus X X X
Sus sp. X X X X
Ovis aries/Capra hircus X X X X
Cervus elaphus X X X X
Dama dama X X X
LagomorphaIV X X
Canis familiaris X X X
Canis lupus X
Carnivori X X
Tab. 1.2: Lista delle specie di macromammiferi trovate nel Complesso 3. Da (Forgia et al. 2012).
I resti dei piccoli vertebrati sono stati studiati dal ricercatore Juan Manuel López-García
dell’Institut Catalaà de Paleoecologia Humana i Evoluciò Soaical, Tarragona in Spagna e
dallo studente Enrico Pagano per quanto riguarda il materiale recuperato dal 2008 al 2011,
e sono oggetto della tesi di laurea triennale dello stesso studente in Beni Culturali
Archeologici del Polo didattico di Agrigento sotto la supervisione della dott.ssa Daria
Petruso per quanto riguarda il materiale del 2012.
Il materiale del 2008-2011 ammonta a circa 300 frammenti, appartenenti a resti di anuri
(rane e raganelle), rettili squamati (serpenti), insettivori, chirotteri (pipistrelli) e roditori,
per un totale di 19 taxa (Tab. 1.3). I dati più importanti che consentono di effettuare una
ricostruzione paleo-ambientale più puntuale, provengono dal sub-complesso 3.4.
L’abbondanza del topo campagnolo Microtus (Terricola) savii suggerisce l’aspetto
degradato dell’ambiente. Il topo selvatico Apodemus sylvaticus, è una forma ubiquitaria
per cui non fornisce particolari indicazioni climatiche, mentre la presenza del ghiro
comune Myoxus glis e dei rettili squamati Vipera gr. V. Aspis indicano la presenza di una
copertura forestale.

17
Micromammiferi 3.1 3.2 3.3 3.4
Erinaceus europaeus X
Crocidura cf. C. sicula X X
Suncus etruscus X
Myotis gr.myotis-blythii X
Arvicola cf. A. terrestris X
Microtus (Terricola) savii X X
Apodemus sylvaticus X X
Rattus norvegicus X
Eliomys quercinus X X
Glis glis X X X
Muscardinus avellanarius X
Avifauna
Aves indet. X X X
Erpetofauna
Discoglossus cf. D. pictus X
cf. Bufo bufo X
Tarentola sp. X
Lacerta gr. L. viridis X X
cf. Podarcis X X
Testudo sp X
Elaphe cf. E. quatuorlineata X X
Vipera gr. V. aspis X
Tab. 1.3 Lista dei taxa di microvertebrati trovate nel Complesso 3 (Forgia et al. 2012).
1.6 I resti umani
All’interno del deposito archeologico di Vallone Inferno, sono stati trovati 33 frammenti
ossei e dentari appartenenti ad Homo sapiens (materiale recuperato dal 2008 al 2011)
Questi reperti, studiati dal Prof. Luca Sineo del Dipartimento di Biologia Animale
dell’Università degli studi di Palermo, provengono dal sub-complesso 3.4. Il 76% dei resti
ritrovati sono denti, in particolare molari e premolari; in alcuni di questi sono presenti delle
lesioni, mentre solo in uno è presente una semplice carie. I denti possono essere ascritti ad
almeno 2 individui adulti e 1 molto giovane. I resti non dentari appartengono, invece, alle
ossa delle mani e dei piedi (Forgia et al 2012).
1.7 Ceramica
I frammenti ceramici, studiati dalle Dott.sse Vincenza Forgia e Giuseppa Scopelliti del
Dipartimento di Beni Culturali dell’Università di Palermo, recuperati in tutto il giacimento
sono 520. Tali frammenti provengono dai sub-complessi 3.1 e 3.2 e sono attribuibili al
periodo tardo-antico. Si tratta di frammenti di brocche utilizzate per la conservazione ed il
trasporto, ma anche dolia, anfore, e diversi frammenti di ceramica da cucina. È stato
rinvenuto anche materiale da costruzione come frammenti di tegole pettinate (Wilson

18
1979) (Fig. 1.7). Particolarmente interessante è un anfora di tipo Keay LXI A (Keay 1984)
(Fig. 1.8) trovata nel complesso 3.2, anfora che risale al VII sec. d.C., cronologia
perfettamente coerente con la datazione al radiocarbonio (Tab. 1.1).
Fig.1.7: frammento di tegolo pettinato tardo-antico
(Forgia et al 2012, particolare di pag. 6 fig. 4) Fig. 1.8: anfora di tipo Keayda LXI A (Forgia et al
2012, particolare di pag. 6 fig. 4)
La ceramica proveniente dal sub-complesso 3.3, è costituita in gran parte da frammenti di
anfore.
Come abbiamo visto nel paragrafo 1.4.1, all’interno del sub-complesso 3.4 ritroviamo
reperti riferiti al Bronzo antico e al Neolitico Medio. Per quanto riguarda la ceramica del
Bronzo antico, questa è rappresentata dalla facies di Castelluccio. Questa facies (dal sito
eponimo presso Noto), definita in seguito agli scavi di Paolo Orsi (Orsi, 1893), ha
un’ampia distribuzione geografica, infatti copre tutta la Sicilia centro-meridionale ed
orientale, ed è inquadrabile tra la fine del III millennio e gli inizi del II millennio a. C. Essa
è considerata contemporanea a quella eoliana di Capo Graziano, in base alla presenza di
ceramica di questa facies in alcuni contesti funerari siciliani (Procelli, 2013). La
produzione ceramica castellucciana è caratterizzata da forme tipiche, come la coppa su alto
piede o a “clessidra” e l’anfora-brocca con anse sopraelevate. La decorazione è dipinta,
presenta un colore bruno – nerastro su fondo rosso o giallo paglierino, con motivi
geometrici e lineari (Tusa, 1994).
I reperti risalenti al Neolitico Medio sono rappresentati dalla facies Tricromica. La
ceramica tipica di questa facies presenta un impasto molto fine, depurato, infatti viene
chiamata anche ceramica figulina; la decorazione è dipinta e presenta i tre colori
caratteristici (giallo-nero-rosso), con motivi geometrici, in particolare bande o fiamme con

19
i bordi neri, campite di rosso, su fondo giallo (Tusa, 1999). Di seguito vi è la figura del
frammento ceramico più rappresentativo (Fig. 1.9).
Fig. 1.9: Frammento di ceramica tricromica p roveniente dallo strato 3.4.N. (Forgia et al. 2012, particolare
di pag. 6 fig. 4)
1.8 Industria litica
L’industria litica di Vallone Inferno è rappresentata da 107 campioni, per lo più si tratta di
industria su scheggia e lama studiati dalla Dott.ssa Vincenza Forgia del Dipartimento di
Beni Culturali dell’Università degli studi di Palermo (Fig. 1.10). L’industria più
significativa proviene dal sub-complesso 3.4, per un totale di 79 pezzi, soprattutto in selce
e ossidiana. La selce è rappresentata da 49 pezzi (61%), principalmente è di provenienza
locale. L’ossidiana, probabilmente proveniente da Lipari, è rappresentata da 18 pezzi
(22%), comprende frammenti e strumenti non ritoccati (Forgia et al 2012).
Fig. 1.10: Manufatti di selce e ossidiana. Per gentile concessione della Dott.ssa Vincenza Forgia.

20
1.9 Archeobotanica
Le analisi archeobotaniche si sono concentrate prevalentemente sullo studio dei resti
carpologici, carboni di legna e resti pollinici studiati dalla Prof.ssa Ethel Allué dell’Institut
Catalaà de Paleoecologia Humana i Evoluciò Soaical, Tarragona in Spagna. Per quanto
riguarda i resti macrobotanici, sono stati recuperati 114 reperti, provenienti dal Complesso
3. Sono stati identificati 21 taxa che comprendono cereali, frutti e colture erbacee. La
maggior parte dei semi sono carbonizzati, tranne 14 resti di Vitis sp. (vitigno). I semi
provengono soprattutto da piante coltivate e frutti. Nell’ambito dei cereali sono stati
riconosciuti semi di grano, in quantità maggiori, e di farro. I semi di leguminose
(lenticchie, fave e piselli) sono molto scarsi (meno del 4%). I resti dei frutti hanno
permesso di riconoscere 6 taxa di cui la specie più abbondante è il frutto della quercia
(ghianda). Un dato interessante proviene dall’analisi dei germogli (10.53%) probabilmente
appartenenti alla famiglia delle Rosacee, che possono essere stati utilizzati come foraggio
per il bestiame.
Tra i carboni di legna sono stati identificati un totale di 19 taxa; tra questi quello più
significativo, visto che è stato trovato in tutta la sequenza è quello della quercia.
Nell’analisi dei resti microbotanici costituiti dai pollini, tra i diversi campioni prelevati
solamente 8 hanno restituito dati più significativi e sono quelli provenienti dai sub-
complessi 3.1, 3.2, 3.4. Il risultato delle analisi dimostra che nei sub-complessi 3.2 e 3.4, è
presente una grossa percentuale di taxa di tipo arboreo (intorno al 20 e al 50%), mentre nel
3.1 i taxa di tipo arboreo diminuiscono sensibilmente e si aggirano tra il 7 e il 20%. In
linea generale, le analisi dei resti pollinici, sembrano restituire un paesaggio formato da
una foresta di querce miste (Forgia et al 2012).

21
Capitolo 2 “Il villaggio di Case Bastione”
2.1 Introduzione
Il presente capitolo è dedicato alla descrizione del villaggio preistorico di Case Bastione,
Villarosa, (EN). Anche qui verrà brevemente descritto l’inquadramento geografico,
geologico e geomorfologico dell’area. Verrà fatta una descrizione generale delle aree di
scavo, ponendo una particolare attenzione nei confronti dell’Area β, in quanto oggetto di
studio di questo lavoro. Verranno presentati anche i materiali faunistici e ceramici
rinvenuti nelle tre campagne di scavo.
2.2 Inquadramento geografico, geologico e geomorfologico
Il villaggio di Case Bastione è situato nella parte occidentale dell’altopiano degli Erei, un
gruppo montuoso della Sicilia centrale, a circa 13 km a Nord della città di Enna, lungo la
valle del fiume Morello, un affluente dell’Imera Meridionale. In particolare ricade nel
foglio IGM 268 I NO, Calascibetta, scala 1:25.000, a 610 m s.l.m. (Fig. 2.1).
Fig. 2.1 Posizione di Case Bastione (foglio IGM 268 I NO – Calascibetta, scala 1:25.000); a sinistra un
modello 3D del sito (Giannitrapani et alii, in cds, pag. 182, fig. 1).
L’insediamento è grande circa 2 ha, ed è situato ai piedi di una ripida parete rocciosa, che
chiude a sud l’area del Lago Stelo, un bacino artificiale drenato nel 1930. Sulla superficie
di questa parete di roccia, costituita principalmente da calcarenite, sono state individuate
diverse tombe a grotticella artificiale, probabilmente risalenti all’Età del Bronzo.

22
Nell’area affiorano i conglomerati arenaceo marnosi della Formazione Terravecchia
risalenti al Miocene superiore – Tortoniano inferiore e litotipi della Formazione Gessoso-
Solfifera (gessi, sali potassico-magnesiaci e zolfo) del Messiniano (fine del Miocene
Superiore) insieme a sedimenti alluvionali come arenarie e marne del Pliocene Medio e
Superiore. (Giannitrapani et alii, in cds).
2.3 Le campagne di scavo
Il sito è stato scoperto nel 2000, grazie ad una ricognizione archeologica all’interno della
Valle del Morello. La prima campagna di scavo è stata nel 2007, inclusa nel progetto
“Interventi di recupero, tutela, restauro ed aumento della fruibilità delle aree
archeologiche della valle del Morello” del POR Sicilia 2000-2006, realizzato dai comuni
di Villarosa e Calascibetta. La seconda campagna è avvenuta nel 2009, grazie alla
collaborazione tra la Soprintendenza per i Beni Culturali di Enna e il Centro di Studi di
Archeologia Mediterranea di Enna. Le ricerche a Case Bastione sono riprese nel 2013,
tramite una nuova partnership tra la Soprintendenza di Enna e la soc. coop. Arkeos, con la
collaborazione scientifica della Università inglese di Newcastle . Il team di ricerca che ha
partecipato alle tre campagne di scavo è stato diretto dagli Archeologi Enrico
Giannitrapani, Filippo Iannì e da I. Gennuso, quest’ultima come responsabile di settore; il
team comprende anche Claudia Speciale come esperta di intonaci, M. Baldi come
specialista di industria litica, Salvatore Chilardi come archeozoologo, M. Arnone come
esperto di geomorfologia, Andrea Dolfini come esperto in archeometallurgia e L.
Anguilano per le analisi di laboratorio (Giannitrapani et alii, in cds).
2.4 Le aree indagate
Durante le prime due campagne di scavo, sono state distinte due aree, chiamate
rispettivamente Area α e Area β. La prima è localizzata nella parte nord del sito, mentre la
seconda 15 m a sud della prima, in corrispondenza di una grande concentrazione di
materiale ceramico rinvenuto in superficie. Nel 2013, grazie ad una ricognizione
geomagnetica, effettuata sull’intera superficie del sito, insieme ad una serie di analisi
chimiche e fisiche su alcuni campioni raccolti, è stata individuata una nuova area di scavo,
chiamata Area γ, situata immediatamente a sud dell’Area α (Fig. 2.2).

23
Fig. 2.2 Ortofoto di Case Bastione (Giannitrapani et alii, in cds, pag. 183, fig. 2).
Questa nuova area è stata indagata effettuando delle trincee esplorative, in corrispondenza
dei punti in cui sono state individuate le anomalie geomagnetiche (Giannitrapani et alii, in
cds).
Di seguito viene riportata una breve descrizione delle tre aree indagate:
Area α: capanna visibile solo in minima parte, la tecnologia costruttiva permette di
confrontarla con la Capanna 1 di Tornambè (Iannì 2013); al di sopra è stata trovata un’area
artigianale, connessa con attività metallurgiche, databile al Rame Finale (2700-2300 a.C.);
il materiale ceramico è stato attribuito alla facies di Malpasso S’Ippolito, al Bicchiere
Campaniforme e alla facies eoliana di Capo Graziano. Dopo un breve periodo di
abbandono l’area è stata rioccupata alla fine del III e l’inizio del II mill. a.C.; a questo
periodo sono attribuite due capanne, la Capanna 2 e 3. Il materiale ceramico trovato
appartiene alla facies di Castelluccio.
Area β: resti della Capanna 1 risalente al Bronzo antico (vedi paragrafo successivo)
Area γ: nella parte centrale è stata trovata una struttura pseudo-circolare, con un
diametro di circa 4 m, che è stata parzialmente danneggiata da recenti arature. La struttura
risale alla fase finale dell’Età del Rame.
Grazie ai numerosi frammenti ceramici è stato possibile documentare la lunga occupazione
umana di Case Bastione, datata a partire dal Neolitico Tardo, fino all’Età del Bronzo (IV –
II mill. cal. B.C). Sono state riconosciute le culture di Diana, San Cono-Piano Notaro,
Serraferlicchio insieme a ceramica dell’Età del Rame e del Bronzo antico. Durante un

24
lungo periodo di abbandono, il sito è stato rioccupato durante l’età Tardoantica e l’età
Bizantina (VI – IX sec. d. C) (Giannitrapani et alii, in cds).
La sequenza stratigrafica è stata suddivisa in varie US, in base alla composizione
litologica, sedimentologica e al materiale archeologico ritrovato. Tutta l’area è stata
suddivisa in quadrati di 4x4 m, che vengono indicati con una lettera e numeri arabi. Il
quadrato di 4 m è stato suddiviso in quattro quadrati da 2x2 m, indicati con un numero
romano (I-II-III-IV), partendo dal quadrato in alto a destra; a sua volta il quadrato di 2 m, è
stato ulteriormente diviso, a seconda della problematica archeologica incontrata, in quattro
quadrati da 1x1 m, indicati con i punti cardinali, NE-SE-SW-NW, partendo sempre dal
quadrato in alto a destra.
2.5 La fase del Bronzo antico dell’Area β
I livelli riferiti al Bronzo antico (2200-1400) (Tusa, 1999), sono stati scavati e analizzati
nell’Area β, dove è stata trovata una capanna, chiamata Capanna 1 (Fig. 2.3), datata al III
mill. cal. B.C..
Fig.2.3 Pianta dell’Area β (Giannitrapani et alii, in cds, pag. 187, fig. 4).
Tutti i livelli più tardi sono stati completamente rimossi a causa di lavori agricoli di epoca
recente; inoltre, durante il Medioevo è stata scavata una grande fossa di forma ellittica,

25
usata per scaricare grossi blocchi in pietra calcarea, probabilmente come risultato di una
frana dalla ripida parete rocciosa che chiude a nord il sito.
La Capanna 1, occupa la parte meridionale dell’Area β, ha pianta ovale, presenta un
orientamento Est-Ovest e le sue dimensioni sono di 10 x 6 m. I due lati corti sono absidati
(Giannitrapani et alii, in cds). La tecnica di costruzione ricorda quella della Capanna 9 del
villaggio di Manfria a Gela (Orlandini 1962), infatti è incassata nel terreno e presenta un
muro perimetrale con pietre di media grandezza, che doveva rappresentare la base per
l’alzato ligneo. Durante lo scavo della capanna sono state riconosciute due fasi di
costruzione, come mostrato dai due battuti pavimentali sovrapposti. La fase più recente (II
fase) è stata danneggiata da lavori agricoli di età moderna, infatti si conservano alcuni tratti
del battuto pavimentale, il crollo dell’intonaco e la copertura di argilla concotta. La fase
più antica (I fase), è meglio conservata, infatti troviamo il battuto pavimentale che è posto
circa 50-60 cm al piano d’uso esterno; inoltre sul piano di calpestio sono stati trovati i resti
di due focolari costituiti da piastre di argilla, mentre nell’abside occidentale è stato trovato
un forno con copertura in argilla. Entrambe le fasi possono essere collocate tra la fine del
III e l’inizio del II mill. cal. B.C. Durante la I fase, c’è stato un incendio che ha portato alla
distruzione e al conseguente abbandono della capanna; nella II fase viene ricostruita. Tale
incendio ha portato alla cottura dello strato di intonaco in argilla steso sulle pareti e sul
tetto della struttura, infatti è stata rinvenuta una grande quantità di concotto su cui sono
evidenti le impronte di pali che dovevano sostenere la copertura, inoltre sono stati ritrovati
molti elementi faunistici in uno stato di calcinazione dovuto all’esposizione ad alte
temperature (Giannitrapani et alii, in cds). All’interno della struttura troviamo anche
ceramica appartenente alla facies di Castelluccio, una grande quantità di pesi da telaio, di
fuseruole e anche reperti faunistici; questo dimostra che parte delle attività quotidiane,
dalla cottura dei cibi alla tessitura, dovevano avvenire all'interno dell'abitazione (Iannì
2013).
In questo lavoro verrà analizzato la fauna a mammiferi proveniente dalle seguenti UU.SS.
(Area β, Capanna 1, I fase), perché considerate più importanti per la ricostruzione storica
del sito, coeve anche alla cronologia del sito di Vallone Inferno (Figg. 2.4-2.5):
• US 1057: crollo Capanna 1 I fase; da questo strato, sembra che gli incendi, siano
dovuti a un evento di tipo traumatico. Questo è testimoniato dai resti di crollo della
capanna e dalla grande quantità di materiale ritrovato, in particolare ceramica e litica,
dovuto quindi ad un abbandono improvviso del sito. Da questo strato provengono anche
due datazioni assolute (Tab. 2.1).

26
• US 2001: crollo interno I fase tra 1057 e piano pavimentale.
• US 2002: strato sopra il probabile focolare della capanna. Si tratta di un livello di
bruciato che presenta fauna e ceramica.
• US 2003: cordolo in concotto del focolare
• US 2079: livello di crollo tra 1064 e 1057
• US 2083: crollo I fase
• US 2087: crollo I fase
• Us 2089: crollo I fase
Contesto Materiale Cad. Lab Data BP Data cal 1⌠ BC Data cal 2 ⌠ BC
US 1057 Charcoal LTL-3657° 3699 ± 45 2190 – 2030 2205 - 1951
US 1057 Fauna Rome – 2055 3645 ± 35 2120 – 1950 2135 – 1921
Tab. 2.1 Lista delle datazioni al 14C, calibrate con OxCal v4.1.7 (Giannitrapani et alii, in cds).
Fig. 2.4 Particolare della pianta dell’Area β (Giannitrapani et alii, in cds, pag. 187, fig. 4).

27
Fig. 2.5 Particolare del
Matrix delle campagne di
scavo 2007, 2009, 2013, con datazioni radiometriche
(Giannitrapani et alii, in cds,
pag. 192, fig. 8).
2.6 La fauna a mammiferi dell’età del Bronzo
I resti dei grandi mammiferi studiati dal Prof. Salvatore Chilardi provengono dai livelli
dell’età del Bronzo ammontano a 790; nell’Area α il numero dei resti (NR) equivale a 426,
mentre nell’ Area β a 364, come indicato nella Tabella 2.2.
Area α Area β
Taxon NR NR% NR NR%
Ovis/Capra
Sus domesticus
Cervus elaphus
Bos taurus
Canis familiaris
Vulpes vulpes
234
108
42
34
7
1
54,9
25,4
9,9
8,0
1,6
0,2
240
67
24
29
2
2
66,0
18,4
6,6
8,0
0,5
0,5
TOTALE 426 100,0 364 100,0
Tab 2.2 NR dei taxa riconosciuti nell’Area α e Area β (Giannitrapani et alii, in cds).
In entrambe le aree sono state rinvenute prevalentemente faune domestiche, anche se in
minor misura, quelle selvatiche sembrano ben rappresentate. In particolare il cervo rosso,
Cervus elaphus, nell’Area α presenta una percentuale di NR pari a circa il 10 %. Il taxon

28
dominante è rappresentato dagli ovicaprini, la cui percentuale è tra il 55% e il 66%, seguito
dal maiale e dal cervo. Una discreta percentuale annovera anche Bos taurus (8%) in
entrambe la aree, mentre gli unici carnivori sono rappresentati da bassissime percentuali
dei canidi, Canis familiaris e Vulpes vulpes. All’interno della Capanna 1 sono stati
rinvenuti pochissimi resti faunistici, il che sembra testimoniare una certa cura nel
mantenimento della pulizia dello spazio interno della struttura; probabilmente i rifiuti
venivano gettati fuori dalla capanna. L’abbondanza di reperti di cervo, utilizzato non solo
per la sua carne ma anche per le sue materie prime come pelli e palchi per la produzione di
manufatti, fa pensare che l’ambiente intorno al sito doveva presentare vaste estensioni di
boschi (Giannitrapani et alii, in cds).
Le analisi archeozoologiche effettuate sui reperti di grandi mammiferi sono il calcolo
dell’età di morte e la determinazione del sesso. Per quanto riguarda la determinazione
dell’età di morte, è stata eseguita sulla base dell’analisi del grado di fusione delle epifisi
delle ossa lunghe, delle ossa costituenti distretti scheletrici come i coxali ed il cranio, e in
parte attraverso le osservazioni compiute sullo stato di usura dei denti. I risultati migliori
sono stati ottenuti dallo studio dei profili di mortalità degli ovicaprini, ottenuti a partire dai
dati di eruzione ed usura dentaria. Pecore e capre venivano abbattute soprattutto fra 1 e 2
anni di età e fra 3 e 4 anni di età; questo evidenzia un chiaro sfruttamento di questi animali
per la produzione di carne, in quanto in questa età vi è un apporto carneo maggiore. Circa
il 38 % è stato abbattuto a circa 4 anni di età, il che testimonia anche un notevole interesse
per la produzione della lana (Giannitrapani et alii, in cds).
Per quanto riguarda i maiali è stato trovato un probabile feto e alcuni resti di un individuo
neonatale, con premolari decidui senza tracce di usura. Tuttavia, la maggior parte dei
reperti ritrovati appartengono ad individui adulti, il che testimonia una pratica di
allevamento mirata all’ottenimento della carne. All’interno del campione faunistico sono
stati ritrovati alcuni canini che hanno dato la possibilità di effettuare la discriminazione fra
maschi e femmine. Da queste analisi, sembra essere testimoniata una tendenza più
conservativa nei confronti delle scrofe, in quanto utili per la riproduzione. Per quanto
riguarda i cervi i dati sull’età di abbattimento non sono numerosi, in ogni caso si può
ipotizzare che c'era una certa attenzione nella scelta delle prede, infatti molti reperti
appartengono ad individui adulti o senili, mentre pochissimi ad individui giovani,
testimoniando così le pratiche di una caccia selettiva Giannitrapani et alii, in cds).

29
2.7 Ceramica
La produzione ceramica del Bronzo antico, riscontrata nella I fase della Capanna 1
dell’Area β, si può riferire con una certa sicurezza alla facies di Castelluccio (vedi Cap. 1,
par. 1.6). La ceramica ritrovata all’interno della Capanna 1 rappresenta l’intero repertorio
di questa facies: coppe su piede, anfore, brocche, giare e dolia cordonati. Questo materiale
mostra una stretta somiglianza stilistica e formale, con il materiale del santuario della
Muculufa (Holloway R. R. et al 1995), posto a nella media valle del Salso a circa 20 km
dalla sua foce, nei pressi di Licata in provincia di Agrigento, con la diffusa presenza della
decorazioni a spina di pesce e metopali, presentando la superficie del vaso divisa in vari
riquadri (Fig 2.6). Questo stile è stato recentemente definito come “Stile di Muculufa”
(Iannì F. 2009).
Fig 2.6, 337 coppa su alto piede, decorazione interna ed esterna, sul piede bande verticali campite da un
motivo a reticolo alternato con probabili motivi vegetali e zoomorfi; 398 coppa, decorazione metopale
campita con motivi radiali; 397 boccale, decorazione con bande orizzontali e triangoli, campiti con motivo a
reticolo; 241 coppa su alto piede, frammento decorazione metope affiancate e sovrapposte, campite con linee
tremule; 425 Anfora di tipo Muculufa, decorazione metope affiancate e sovrapposte, campite con linee
tremule; 259 Anfora di tipo Muculufa, decorazione con metope orientate verticalmente, campite da motivo a
spina di pesce; 359 Anfora di tipo Muculufa, decorazione con metope affiancate e sovrapposte, campite da
motivo a reticolo, linee tremule e spina di pesce (Giannitrapani et alii, in cds, pag. 197, fig. 9).

30

31
Capitolo 3 “Metodologia: l’analisi archeozoologica”
3.1 Introduzione
Nel presente capitolo verranno esposte le metodologie archeozoologiche utilizzate, per
l’analisi dei campioni faunistici oggetto di studio di questa tesi. Nell’ottica di rendere la
mia analisi fruibile ad altri utenti e i dati confrontabili con altri siti, tutti i dati relativi ai
reperti analizzati (stratigrafici, tassonomici, anatomici, tracceologici) sono stati inseriti in
un database i cui contenuti verranno discussi nel capitolo 5. Proprio a questo scopo i dati
sono stati oggetto di varie classificazioni e abbreviazioni che saranno descritte nei
paragrafi successivi.
3.2 L’identificazione anatomica e tassonomica
Il riconoscimento anatomico e tassonomico dei reperti scheletrici, è il primo passo da
effettuare prima di qualsiasi analisi archeozoologica. Per questo lavoro, il riconoscimento è
stato reso possibile grazie alla collezione di confronto del Dipartimento di Scienze della
Terra e del Mare della Facoltà di Scienze Naturali dell’Università di Palermo. A tale fine
sono stati utilizzati anche vari atlanti osteologici come Schmid (1972), Barone (1976),
Hillson (1986), Wilkens (2003). In particolare per gli erbivori è stato utilizzato l’atlante di
Pales & Lambert (1971).
In alcuni casi, è stata effettuata la discriminazione tra specie e generi osteologicamente
vicini come pecora e capra, e la distinzione tra la forma selvatica e quella domestica, come
tra cinghiale e maiale. Per quanto riguarda la distinzione tra pecora e capra, è stata
effettuata sia grazie ai quarti premolari decidui, sia agli astragali. Per quanto riguarda i
quarti premolari decidui, è stato utilizzato il metodo di Payne (1985) (Fig. 3.1). Tale
metodo fa riferimento alla presenza/assenza del tubercolo tra i lobi mediale e distale del
dente, presente nella capra e assente nella pecora, e alla conformazione tra corona e radice,
che nella capra sembra avere un orientamento orizzontale mentre nella pecore un
orientamento obliquo, sia nella parte mediale che nella parte distale.

32
Fig 3.1. Differenze fra capra e pecora visibile nel dP4 (Payne 1985).
Tale discriminazione è stata effettuata anche grazie all’astragalo, utilizzando il criterio 1 di
Zeder & Lapham (2010) (Fig. 3.2). Tale criterio è basato sull’orientamento della cresta
articolare mediale presente nel lato anteriore, in Ovis risulta essere orientata
orizzontalmente, mentre in Capra presenta un angolo obliquo.
Fig. 3.2 Distinzione Ovis/Capra attraverso l’astragalo tramite Criterio 1 di Zeder & Lapham (2010).
Per quanto riguarda la discriminazione tra cinghiale e maiale, è stata effettuata grazie alla
presenza di un canino, che date le dimensioni, è stato attribuito alla forma selvatica Sus
scrofa.

33
3.2.1 Classificazione per categoria
Per le ossa che non sono state riconosciute a livello anatomico, dato l’alto grado di
frammentazione, è stato possibile inserirle all’interno delle categorie osso lungo, osso
piatto e osso articolare (Fig. 3.3):
Osso lungo: la dimensione maggiore è la lunghezza, costituiscono le ossa degli arti
come il femore, l’omero, il radio, l’ulna, la tibia, la fibula, i metapodi e le falangi; hanno
una sezione più o meno cilindrica, la loro funzione è quella di sopportare il peso del corpo.
Anatomicamente si distinguono in tre parti, la parte centrale del corpo dell’osso detta
diafisi, le estremità chiamate epifisi e il punto di raccordo (dove si trova la parte di
collagene nelle ossa in crescita dei soggetti giovani) tra diafisi ed epifisi, metafisi. Ognuna
di queste parti ha una composizione di tessuto osseo differente, infatti la diafisi è un
cilindro cavo costituito da tessuto osseo compatto, al cui interno si trova la cavità
midollare. Il massimo spessore è al centro e va diminuendo verso le estremità. Le epifisi
sono costituite da tessuto osseo spugnoso, il cui reticolo di trabecole comunica
direttamente con la cavità midollare della diafisi (De Grossi Mazzorin 2008).
Ossa piatte: non hanno cavità midollare, sono pertinenti allo scheletro assile
(vertebre e coste), allo scheletro craniale (cranio e mandibola) e i cinti scapolari e pelvici.
Sono costituite da due tavolati, uno interno e l’altro esterno, di tessuto osseo compatto, che
a loro volta circondano uno strato più interno di tessuto spugnoso. Queste ossa hanno
essenzialmente due funzioni, proteggono i tessuti molli sottostanti e la loro larga superficie
è importante per l’attacco dei muscoli (De Grossi Mazzorin 2008).
Ossa articolari: ossa che costituiscono le articolazioni e comprendono carpali e
tarsali, la patella e le epifisi delle ossa lunghe, che a causa della frammentazione, non è
stato possibile riconoscerle a livello anatomico (Caceres 2002).

34
Fig. 3.3 Classificazione delle ossa per tipo di osso (Blasco, 2006).
3.2.2 Classificazione per taglia
Le ossa così inserite all’interno di una delle tre categorie sopra elencate e quelle
riconosciute a livello anatomico sono state anche distinte sulla base della taglia; la
categoria di taglia è stata dedotta sulla base dello spessore del tessuto corticale e delle
dimensioni generali del frammento osseo. In particolare, sono state introdotte quattro
categorie di taglia (taglia grande, taglia media, taglia piccola e taglia molto piccola), create
a partire dalla classe di età e dalle dimensioni degli animali domestici e selvatici attuali,
prendendo anche in considerazione i campioni faunistici presenti nei siti archeologici
oggetto di studio di questo lavoro (Tab. 3.1). Occorre sottolineare che a questa distinzione
sono state escluse le ossa di animali troppo giovani perché ovviamente non hanno
raggiunto la taglia finale.
Osso lungo
Osso piatto
Osso articolare

35
Taglia Peso (Kg) Specie
Taglia grande 1000-300 Bos taurus adulto
Taglia media 300-100 Sus scrofa adulto; Cervus elaphus adulto; Bos taurus giovane; Dama
dama adulto
Taglia piccola 100-10 Sus scrofa giovane; Cervus elaphus
giovane; Ovis/Capra; Dama dama
giovane; Canis familiaris; Canis lupusi; piccoli carnivori
Taglia molto piccola <10 Lagomorpha; Ovis/Capra giovane;
Canis giovane
Tab. 3.1 Categorie di taglia.
3.2.3 Classificazione per porzione e parte
Allo scopo di migliorare la valutazione del campione faunistico analizzato i reperti
scheletrici riconosciuti a livello tassonomico, anatomico e, ove possibile, quelli inseriti
all’interno delle categorie osso lungo, piatto e articolare, sono stati distint i, sulla base della
porzione conservata. Le ossa lunghe ad esempio sono costituite da una diafisi, la porzione
centrale dell’osso, da due epifisi, le estremità articolari, e da due metafisi, la giunzione tra
la prima e le seconde. In questo lavoro, per identificare queste porzioni è stato assegnato
un numero progressivo ad ogni porzione dell’osso; per le ossa lunghe la numerazione
progredisce a partire dalla porzione prossimale verso quella distale (Saladiè, 1998; Rosell,
2001; Blasco, 2006; Fig. 20), mentre nelle ossa piatte come il cranio e la mandibola dalla
parte anteriore verso quella posteriore e la scapola dal margine vertebrale verso quello
craniale (Figg. 3.4-3.5):
Porzione 1: epifisi prossimale
Porzione 2: metafisi prossimale
Porzione 3: diafisi
Porzione 4: metafisi distale
Porzione 5: epifisi distale
Fig. 3.4 Schema delle porzioni in un osso lungo

36
Fig. 3.5 Porzioni delle ossa piatte
Per questo lavoro è stata classificata anche quale parte dell’osso si è conservata. Per le ossa
riconosciute tassonomicamente e anatomicamente è stata catalogata la parte conservata con
la sigla ant per indicare la parte craniale, post per quella caudale, int mediale ed ext per
quella laterale. Per le ossa che non sono state riconosciute a livello anatomico, quindi
inserite nelle categorie osso lungo, piatto e articolare, le parti conservate sono state indicate
con dei numeri, 1 quando vi è solo una parte conservata, 2 quando vi sono due parti
conservate e così via (Fig. 3.6).
Fig. 3.6 Schema delle parti che compongono l’osso.
3.3 Analisi quantitative
Una volta aver effettuato il riconoscimento anatomico e la eventuale determinazione della
specie e aver calcolato l’età di morte sulle ossa (fusione epifisi-diafisi, fusione ossa del

37
cranio) o sui denti (eruzione-sostituzione dentaria, usura superficie occlusale) e solo in rari
casi essere riusciti a determinare il sesso, si procede al conteggio dei reperti. L’analisi
quantitativa dei reperti permette di capire quale specie avesse più importanza all’interno
del contesto archeologico oggetto di studio, in pratica quali animali venivano più cacciati e
quali più allevati. Inoltre stabilire la quantità numerica di ciascun taxon, permette di
effettuare la ricostruzione paleo-ambientale e individuare eventuali cambiamenti nelle
strategie di approvvigionamento animale (Wilkens, 2003). Accostando queste informazioni
con i dati stratigrafici è possibile dare a questi cambiamenti una connotazione di tipo
temporale e spaziale. Per questo lavoro sono stati utilizzati il NR (numero dei resti), il
NME (numero minimo degli elementi), il NMI (numero minimo degli individui) e il NEE
(numero minimo degli elementi aspettati).
3.3.1 NR “Numero dei Resti”
Il NR consiste nel conteggio di tutti i frammenti ossei e dentali presenti all’interno del
campione faunistico. Questo indice può essere rapportato ad una singola specie, quindi
vengono contate le ossa riconosciute totalmente (NRDt, “numero dei resti determinati
totalmente”), può essere rivolto a quelle riconosciute solo a livello anatomico (NRDa,
“numero dei resti determinati anatomicamente”), e viene anche utilizzato per il conteggio
delle ossa indeterminate (NRindet). Per questo lavoro il NR è stato utilizzato anche per
quantificare le ossa inserite all’interno delle categorie di taglia. Questo metodo di
quantificazione permette di valutare la quantità dei resti e la frammentazione del campione
faunistico.
3.3.2 NME “Numero minimo degli Elementi”
Il NME consiste nel conteggio degli elementi anatomici per i quali è stato riconosciuto il
taxon di appartenenza, l’età, la porzione, e il lato. Differisce dal NRDt, in quanto
quest’ultimo computa tutti gli elementi anatomici rinvenuti, anche se non è stato possibile
distinguere l’età e il lato. Questo metodo di quantificazione serve anche come fase
preliminare per il calcolo del NMI. Dove è stato possibile, in questo lavoro il NME è stato
messo in rapporto con il NRDt, per calcolare l’indice di frammentazione di ogni elemento
anatomico di ogni singola specie riconosciuta.

38
3.3.3 NEE “Numero Minimo degli Elementi Aspettati”
Questo metodo di quantificazione, chiamato anche tasso di sopravvivenza scheletrica
(Lyman, 1994) serve per valutare l’abbondanza degli elementi scheletrici all’interno del
campione faunistico, cioè quale distretto scheletrico è maggiormente rappresentato. Questo
metodo si applica mettendo in rapporto il NME, con il numero degli elementi aspettati, ed
è basato sul numero minimo di individui precedentemente calcolato.
3.3.4 NMI “Numero Minimo di Individui”
Il NMI quantifica le carcasse alle quali l’uomo ha avuto accesso, determinando di
conseguenza la specie più rappresentata all’interno del deposito, basandosi naturalmente
sugli elementi scheletrici recuperati (Wilkens, 2003). Si tratta sempre di una stima
orientativa, infatti grazie ai processi tafonomici di origine antropica e naturale, non sarà
mai possibile calcolare il numero iniziale di individui (NI). Non esiste un unico metodo per
calcolare il NMI. Per questo lavoro è stato calcolato attraverso il conteggio del NME, cioè
si considera l’elemento anatomico più numeroso di ogni taxon, prendendo in
considerazione anche l’età e il lato.
3.4 Calcolo dell’età di morte
Per questo lavoro, l’età di morte è stata calcolata principalmente attraverso lo stato di
eruzione e di usura dei denti, in quanto molto numerosi all’interno dei due campioni
faunistici analizzati, e anche perché hanno restituito dei dati più attendibili. È stato preso in
considerazione anche lo stato di fusione delle epifisi delle ossa lunghe. Inoltre, l’aspetto
poroso o compatto delle ossa ha permesso di attribuire il reperto a soggetti giovani anche
quando questo era in cattive condizioni di conservazione. In tal senso sono state create
semplicemente tre classi di età: alla classe neonatale sono state attribuite quelle ossa dotate
di un elevato grado di porosità oltre che di una taglia piccola rapportata alla specie e alla
parte anatomica di appartenenza; alla classe giovanile sono state attribuite quelle ossa le
cui epifisi non sono saldate alle diafisi; la classe adulta invece è rappresentata dalle ossa le
cui epifisi, sono saldate e che hanno raggiunto la taglia finale in rapporto alla specie e
all’elemento scheletrico di attribuzione. Unendo i dati ricavati dallo stato di eruzione e di
usura dei denti, e quelli ricavati dallo stato di fusione delle epifisi sono state create le
seguenti classi d’età:
Neonato: 1 mese
Infantile: 6-8 mesi

39
Giovane: 9-18 mesi
Sub-adulto: 18-24 mesi
Adulto: più di 24 mesi
3.4.1 Calcolo dell’età di morte degli ovicaprini
Per gli ovicaprini sono state utilizzate le tabelle che indicano lo stato di eruzione dei denti
secondo Habermehl (1961), Barone (1980) e Silver (1969), (Tab. 3.2). Queste tabelle sono
derivate da studi su animali viventi, sulle quali si può trovare il momento della sostituzione
dei singoli denti, sia per gli animali domestici che per quelli selvatici.
Tab. 3.2 Stato di eruzione dei denti degli ovicaprini secondo Habermehl (1961), Barone (1980) e Silver
(1969).
Per quanto riguarda lo stadio di usura dei denti è stato utilizzato il metodo Wilkens (2003),
basato sia sullo stato di eruzione che su quello di usura, in particolare sulla perdita degli
infundiboli dei molari inferiori, prendendo in considerazione anche l’altezza della corona
dentaria (Tab. 3.3).
Capra hircus
Ovis aries
Habermehl
1975
Habermehl
1975
Barone
1976
Barone
1976
Silver
1969
Silver
1969
pecora capra pecora capra pecora capra
Id1-Id2 nascita nascita 1 giorno 1 giorno entro 1° sett. nascita
Id3 nascita nascita 1 giorno 1 giorno entro 2°sett. nascita
Cd/(Id4) 1 sett. 15-25
giorni
entro 3°sett. 1-3 sett.
Pd tutti 4 sett. 3 mesi 1 mese 1-3 mesi entro 6°sett. 3 mesi
I1 12-20 mesi 14-16 mesi 15 mesi 15 mesi 12-15/18m. 15 mesi
I2 18-27 mesi 19-22 mesi 21 mesi 21 mesi 18-24/30m. 21/27 mesi
I3 27-36 mesi 21-26 mesi 28 mesi 28 mesi 27-36/42m. 27/36 mesi
C (I4) 36-46 mesi 29-36 mesi 3 anni 3 anni 33-48/50m. 36/40 mesi
P2 - P3 24 mesi 17-20 mesi >20 m. 20 mesi 21-24/30m. 17-20/30
m.
P4 24 mesi 17-20 mesi > 20 m. 20 mesi 21-24/40m. 17-20/30
m.
M1 3 mesi 5½ mesi 3 mesi dopo 3 m. 5/6 mesi 5-6 mesi
M1 3 mesi 5½ mesi 3 mesi dopo 3 m. 3/6 mesi 5-6 mesi
M2 9 mesi 9-10 mesi 9 mesi dopo 9 m. 9-12/18m. 8-10/12 m.
M2 9 mesi 8-9 mesi 9 mesi dopo 9 m. 9-12/18m. 8-10/12 m.
M3 18 mesi 18-24 mesi 18 mesi dopo 18
m.
18-24m/3-4a 18-24/30
m.

40
1 Pd4 senza usura 1-3 mesi
2 Eruzione e inizio usura di M1 3-5 mesi
3 M1 in uso, gemma di M2
4 M2 in eruzione, si perde l’infundibolo del primo elemento di Pd4 8-9 mesi
5 M2 in uso, gemma di M3
I Eruzione di M3 e dei premolari 18-24 mesi
II
Usura sul primo e secondo elemento
di M3, inizio di usura sul terzo
elemento
III Terzo elemento di
M3 in uso
IV
Si perde
l’infundibolo
del primo
elemento di
M1. M3 con
radici in
genere aperte
V
M1 senza
infundiboli.
Corona di M3
poco più alta
che lunga, tendente al
quadrato
VI
Si perde
l’infundibolo
del primo
elemento di
M2. Corona di
M3 più lunga
che alta
VII
M2 senza
infundiboli.
Altezza della
corona di M3 uguale o
minore
dell’altezza
delle radici
Tab. 3.3 Stadio di usura dei denti degli ovicaprini secondo Wilkens (2003)
3.4.2 Calcolo dell’età di morte dei suini
Anche per i suini sono state utilizzate le tabelle che indicano lo stato di eruzione dei denti,
secondo Habermehl (1961), Barone (1980) e Silver (1969), (Tab. 3.4).

41
Sus
scrofa
Habermehl
1975
Habermehl
1975
Barone
1976
Silver
1969
cinghiale maiale maiale maiale
Id1 1° mese 1-4 sett. 2-3 sett. 1-3s/4-14g
Id1 1° mese 1-4 sett. 2 sett. 1-3s/4-14g
Id2 12-14 sett. 2-3 mesi 10-14/6-12s
Id2 10 sett. 6-10 sett. 2 mesi 10-14/6-12s
Id3 prenatale prenatale prenatale nascita
Cd prenatale prenatale prenatale nascita
Pd2 10 sett. 5-7 sett. 1½ mese 7-10 sett.
Pd3 1°mese 6-8 giorni 1-2 sett. 1-3/1-5 s.
Pd3 2-4 sett. 2-5 sett. 1-3/1-5 s.
Pd4 1° mese 6-8 giorni 2-3 sett. 1-4/2-7 s.
Pd4 2-4 sett. 4-8 sett. 1-4/2-7 s.
P1 6, 6½ mesi 5 mesi 4-6 mesi 3½-6½ mesi
I1 15-18 mesi 11-14 mesi 12-15 mesi 12-17 mesi
I2 20 mesi 16-20 mesi 18 mesi 17-20 mesi
I3 12 mesi 6-9 mesi 9-12 mesi 8-12 mesi
C 10 mesi 6-9 mesi 9-12 mesi 8-12 mesi
P 12-20 mesi 12-15 mesi 12-15 mesi 12-16 mesi
M1 5-6 mesi 4-6 mesi 4-6 mesi 4-6 mesi
M2 12-18 mesi 8-12 mesi 8-12 mesi 7-13 mesi
M3 21-24 mesi 18-20 mesi 18-22 mesi 17-22 mesi
Tab.3.4 Stadio di eruzione dei denti di maiali e cinghiali secondo Habermehl (1961), Barone (1980) e
Silver (1969).
Per l’usura dentaria, anche in questo caso è stata utilizzata la tabella ideata dalla Wilkens
(2003), basato soprattutto sullo stadio di usura dei molari (Tab. 3.5)
1 Pd4 senza usura 2-4 settimane
2 Eruzione e inizio usura di M1 4-6 mesi
3 M1 in uso, gemma di M2
4 M2 in eruzione Maiale 8-12 mesi
Cinghiale 12-18 mesi
5 M2 in uso, gemma di M3
I M3 senza usura, radici aperte Maiale 18-20 mesi
Cinghiale 21-24 mesi

42
II
Usura di M3 solo sullo smalto. Dentina non visibile
o visibile solo in piccoli punti sulle cuspidi
principali
III
Dentina rotonda sulle cuspidi principali e su alcune
secondarie
IV
La dentina di cuspidi diverse si unisce
V
Nel terzo elemento l’usura è arrivata al colletto e lo
smalto scompare. Dentina visibile su tutto il piano
masticatorio
Tab. 3.5 Stadio di usura dei denti dei suini secondo Wilkens (2003)
3.4.3 Calcolo dell’età di morte del Cervus elaphus
Per il Cervus elaphus è stato utilizzato il metodo di Brown & Chapman (1991), basato
sullo stadio eruzione dei denti permanenti, osservato sulle radiografie di un campione di
esemplari di maschi e femmine attuali. In dettaglio sono stati analizzati i denti relativi alla
mandibola, ma gli stessi principi possono essere attribuiti ai denti mascellari (Brown &
Chapman,1991) (Fig. 3.7):

43
Fig. 3.7 Metodo Brown & Chapman (1991), per calcolare l’età di morte del Cervus elaphus.
Di seguite vengono descritte le classi di età illustrate in fig. 3.7 :
(a) 5 mesi: il primo molare ha iniziato a svilupparsi nell’utero e a 5 mesi è erotto più
della metà dell’altezza della corona. Tutte le cuspidi del secondo molare sono
mineralizzate anche se gli infundiboli non sono ancora visibili. Non c’è nessuna evidenza
dei premolari permanenti.
(b) 10 mesi: il primo molare ha raggiunto la piena altezza della corona e le radici sono
aperte. Il secondo molare ha la corona completa. È presente la cripta del terzo molare.
(c) 13 mesi: il terzo molare è mineralizzato. Sono evidenti le cripte del secondo, terzo e
quarto premolare permanente. Ha inizio la mineralizzazione del quarto premolare
permanente.
(d) 15 mesi: tutti i premolari sono mineralizzati nelle loro cripte, sotto i rispettivi denti
decidui.

44
(e) 18 mesi: le corone del terzo molare e di tutti i premolari sono formate. Le radici dei
premolari decidui sono in riassorbimento, il che implica che l’eruzione dei sottostanti denti
permanenti è iniziata.
(f) 27 mesi: il terzo molare sta formandola radice posteriore. I premolari sono tutti in
eruzione per andare a sostituire gli ultimi denti decidui.
(g) 50 mesi: i molari sono completamente formati, a volte fra i 28 e i 40 mesi.
3.5 Misure osteometriche
In questo lavoro sono state prese le misure osteometriche, sulle ossa di animali adulti,
attraverso il metodo descritto da Von Der Driesch (1976) al fine di stimare la statura degli
animali riconosciuti nel campione faunistico. Questa misurazione è correlata alla
lunghezza delle ossa degli arti, per cui è possibile valutarne l’altezza al garrese, cioè la
zona del corpo che corrisponde alle prime vertebre toraciche (De Grossi Mazzorin, 2008).
Tale altezza è stimata moltiplicando l’altezza dell’osso per alcuni coefficienti (Von Den
Driesch, Boessneck, 1974) (Tab. 3.6). Questi calcoli possono presentare un margine di
errore, in quanto le misure calcolate sugli astragali, possono essere riferite ad animali che
non hanno ancora raggiunto l’età adulta, ma anche il pregio di fornire un’immagine delle
dimensioni dell’animale immediatamente accessibile anche ai non specialisti del settore
(De Grossi Mazzorin, 2008). Questa stima permette anche di confrontare le variazioni
dimensionali tra contesti archeologici differenti.

45
Aut. MATOLCSI
1970 TEICHERT
1975 SCHRAMM
1967* TEICHERT
1969 HARCOURT
1974 GODYNOCKI
1965 Taxon Bos taurus Ovis aries Capra hircus Sus scrofa Canis
familiaris Cervus elaphus
Scapola (Sc) 4,22 3,8 Omero (Hu) 4,14 4,28 3,86 4,05 3,43 Gl –
26,54
Radio (Ra) 4,3 4,02 3,98 5,26 3,18 GL + 19,51
Hu+Ra 1,65 GL – 4,32
Ul 3,32 3,97 2,78 Gl + 6,21 Mc III 10,72 Mc IV 10,53 Mc 6,18 4,89 5,75 4,55
Mc maschile 6,33 4,67
Mc
femminile
6,03 4,48
Pelvi (Pv) 3,04
Femore (Fe) 3,23 3,53 3,45 3,65 3,14 Gl – 12,96
Tibia (Ti) 3,45 3.01 2,97 3,92 2,92 Gl + 9,41
Fe + Ti 1,52 – 2,47
Fibula (Fi) 4,23
Calcagno 11,40
Astragalo 22,68
Tarsale (Tl) 17,9
Carpale (Cc) 9,34
Mt III 9,34
Mt IV 8,84
Mt 5,47 4,54 5,34 4,08
Mt maschile 5,62 4,19
Mt femminile 5,33 4,01
Tab. 3.6 Coefficienti per stimare l’altezza al garrese del bue, della pecora, del cane e del maiale (Von Den
Driesch, Boessneck, 1974).

46

47
Capitolo 4 “Metodologia: l’analisi tafonomica”
4.1 Introduzione
La tafonomia, dal greco τάφος (tàfos)= tomba-sepoltura, e νόμος (nòmos)= legge, è la
scienza che studia i processi che generano, modificano e distruggono i depositi fossili
(Efremov 1940). Tali processi comprendono il periodo in cui l’animale è ancora in vita
fino all’arrivo dei reperti scheletrici sul tavolo dell’archeozoologo (Davis 1978). Alla
morte dell’animale segue la distruzione della sua spoglia operata da agenti esogeni chimici
(ossigeno, acqua, acidità del suolo), fisici (alte temperature, correnti turbolente, effetti
gelo-disgelo) e biologici (per opera di radici, carnivori, roditori) che se si protrae in tempi
lunghi e non viene interrotta dal seppellimento, può portare alla completa distruzione. Se
una coltre sedimentaria di opportuno spessore si deposita sulla spoglia, comincia la
fossilizzazione vera e propria che in tempi geologici porta alla litificazione. I processi
tafonomici sono determinati anche dall’azione dell’uomo, quindi alla componente naturale,
viene a sommarsi e ad intersecarsi quella antropica. L’uomo con la sua attività di caccia e/o
di allevamento opera un processo di selezione di specie (solo quelle cacciate o allevare), un
processo di concentrazione di resti scheletrici e di selezione di alcune ossa più ricche in
carne. Inoltre, concorre alla morte dell’animale e sulle ossa vi lascia le tracce della sua
attività di macellazione. L’individuazione e il riconoscimento di queste tracce permette di
ricostruire le modalità di formazione del campione faunistico, inoltre permette di ottenere
dati importanti sulle pratiche culturali del gruppo umano in esame (Solari et al. 2013).
L’individuazione e l’analisi delle tracce tafonomiche, sono state effettuate nel
Dipartimento di Scienze della Terra e del Mare della Facoltà di Scienze Naturali
dell’Università di Palermo, tramite il microscopio Leica zoom 2000 con ingrandimenti da
10.5 a 45, e nel Laboratorio di Archeozoologia e Tafonomia del Dipartimento di Biologia
ed Evoluzione dell’Università degli Studi di Ferrara, tramite il microscopio Leica S6D con
ingrandimenti da 0.63 a 4.0, con fotocamera Leica EC3. Le fotografie dei reperti sono state
effettuate tramite la macchina fotografica reflex Canon 600D con obiettivo Canon EF-S 60
mm f/2.8 Macro USM. Le analisi al SEM sono state effettuate al TekneHub.
4.2 Tracce di origine naturale
Molti fattori biostratinomici di origine naturale possono agire sulle carcasse e ossa di
animali, nel periodo che concorre tra la morte dell’individuo e il seppellimento dei resti.
Alcuni di questi fattori sono dati da deteriorazioni chimiche e meccaniche, per

48
l’esposizione in un contesto sub-aereo o di superficie (weathering) (Lyman 1994), da
alterazioni prodotte da carnivori e roditori, dal calpestio da parte di altri animali,
dall’azione delle radici e alterazioni chimiche che portano all’accumulo di ossidi di
manganese. Anche le tracce di origine naturale possono portare ad un ulteriore
fratturazione del campione archeofaunistico.
4.2.1 Alterazione prodotta da carnivori.
L’azione determinata dei carnivori sulle ossa, è finalizzata all’estrazione dei tessuti molli
rimasti, e al consumo del midollo. Queste azioni portano ad avere particolari tipi di tracce e
a volte ad un ulteriore fratturazione dell’osso. Le tracce di rosicatura prodotte da carnivori
sono molto variabili, infatti diversi autori ne hanno individuato le varie tipologie, (Binford
1981, Blumenschine y Selvaggio 1988, Bunn 1981, Haynes 1980, 1983, Maguire et al.
1980, Shipman, 1981; Stiner, 1994). Per questo lavoro sono state individuate e classificate
i seguenti tipi di tracce:
Solchi: tracce prodotte dallo sfregamento dei canini e dei molari, per lacerare le
fibre muscolari. Sono strie dalla lunghezza e profondità variabile, con una superfice ruvida
e irregolare. Corrisponde al termine inglese scoring.
Depressioni: si originano per la pressione esercitata dai molari sulla superficie
corticale dell’osso; presentano una morfologia circolare o ovale.
Impronta: corrisponde alle tracce lasciate da più cuspidi dello stesso dente o di una
serie dentaria.
Un altro tipo di alterazione della superficie dell’osso dovuta all’azione da parte di
carnivori, sono le tracce di morsicatura. Questa azione viene indicata con il termine inglese
pitting (Binford 1981), presenta abbondanti depressioni di piccole dimensioni e poco
profonde, solitamente localizzate lungo le estremità dell’osso; inoltre grazie anche
all’azione degli enzimi salivari, le estremità risultano essere arrotondate e lucide (Caceres
2002).
4.2.2 Alterazioni prodotte da roditori
All’interno del campione faunistico analizzato in questo lavoro, sono state trovate delle
ossa che presentano tracce di rosicatura prodotte da roditori. Questi animali rosicchiano le
ossa, probabilmente non per scopo nutritivo, ma bensì per limare gli incisivi che sono in
continua crescita (Caceres 2002). Alcuni autori pensano invece che ci possa essere una
componente alimentare, in quanto ottengono dei minerali che sono carenti nella loro dieta

49
(Laudet & Fosse 2001). In questo caso l’osso, al momento dell’attività prodotta dai
roditori, si presenta secco, cioè senza periostio e grasso. Questo tipo di tracce si presentano
come delle scanalature parallele tra di loro, non molto profonde, di lunghezza e larghezza
variabili. Si possono trovare in unico punto oppure su tutta la superficie dell’osso (Sutcliffe
y Collings, 1972; Maguire et al, 1980, Brain, 1981).
4.2.3 Calpestio
Il calpestio, termine inglese trampling, è un processo meccanico determinato dal passaggio
di animali e uomini, che porta allo sfregamento tra la superficie dell’osso e il substrato
(Caceres 2002). Behrensmeyer (1986, 1989) e Fiorillo (1989), hanno dimostrato
sperimentalmente che il calpestio, può produrre delle strie sulla superficie delle ossa. Da
questi studi si è evinto che le strie determinate attraverso il trampling, sono molto simili
morfologicamente a quelle ottenute attraverso uno strumento litico, principalmente
incisioni e raschiature (Oliver 1989), ma a differenza di queste, presentano un
orientamento casuale e multidirezionale (Lyman 1994). Le tracce da calpestio, sono meno
profonde rispetto alle tracce da macellazione e presentano una sezione a U o piana
(Andrews & Cook 1985, Olsen & Shipman 1988), hanno una lunghezza e larghezza
variabile e si possono trovare isolate e raggruppate. Inoltre, mentre le tracce di
macellazione si possono trovare in punti ben precisi, ad esempio all’altezza delle
articolazioni e inserzioni muscolari, le tracce da calpestio non hanno nessuna relazione con
la struttura anatomica del resto osseo, pertanto la loro distribuzione sulla superficie sarà
casuale (Gifford-González et al 1985, Gifford-González 1989 a, 1989b, Bunn & Kroll
1986, Agenbroad 1989, Irving et al. 1989, Oliver 1989). In questo lavoro le tracce
determinate dal trampling, sono state catalogate a seconda della loro distribuzione sulla
superficie dell’osso: generalizzate (Gen) se si trovano su tutta la superficie; concentrate
(Conc) se sono numerose ma tutte nello stesso punto; isolata (Isol) se è presente solo una
stria; disperso (Disp) se si trovano più di una stria ma distanziate fra di loro.
Il trampling, oltre a determinare le strie, può portare alla frammentazione del campione
faunistico, producendo, schegge e bordi irregolari (Agenbroad, 1989; Gifford-González,
1989b; Irving et al., 1989). La frammentazione per calpestio, avviene con più facilità per
quelle ossa che sono state già alterate da altri processi tafonomici; mentre, nel caso di ossa
fresche è più facile che si vengano a formare le strie precedentemente descritte. La
frammentazione dipende anche dalla morfologia dell’elemento scheletrico, un osso piatto

50
può andare più facilmente incontro a frammentazione rispetto a un osso articolare (Yellen,
1991).
Infine bisogna sottolineare, che il calpestio, può portare allo spostamento o al movimento
delle ossa, all’interno del contesto archeologico in cui sono state ritrovate (Andrews y
Cook, 1985). Questo movimento può essere orizzontale o verticale. Per quanto riguarda
quello orizzontale, dipende molto dallo stato di compattezza del suolo, infatti uno strato
duro, non favorendo l’interramento delle ossa, aumenta la possibilità di una distribuzione
orizzontale lungo la superficie (Gifford-González et al. 1985, Olsen & Shipman 1988).
Anche il movimento verticale dipende dallo struttura e dalla compattezza del suolo.
Quando questo si presenta particolarmente friabile, provoca un interramento verticale
piuttosto rapido (Olsen y Shipman, 1988).
4.2.4 Alterazione prodotta da radici
Questo tipo di alterazioni, lascia delle tracce che si vengono a formare quando vi è il
contatto tra le radici delle piante e la superfice dell’osso. Le radici tendono ad aggredire gli
elementi scheletrici in quanto questi contengono dei minerali che ne possono garantirne la
crescita; quando avviene l’assorbimento delle sostanze nutritive, le radici secernano acido
umico che è il responsabile della formazione di questi tipi di tracce (Behrensmeyer 1978).
Alcuni autori hanno suggerito che i solchi possono essere causati da acidi prodotti da
determinati funghi associati alla decomposizione delle piante. Questi funghi possono
decomporre, non solo la materia organica in condizioni relativamente secche, ma possono
anche produrre una grande quantità di acidi organici durante tale processo (Morlan 1980,
Grayson 1988). Queste alterazioni appaiono sia sotto forma di solchi che si ramificano
lungo la superficie dell’osso, di lunghezza, larghezza e profondità variabile, presentando
una sezione a U, sia sotto forma di perforazioni (Caceres 2002).
In questo lavoro, le alterazioni prodotte da radici sono state catalogate, distinguendo se si
tratta di ramificazioni o perforazioni. Per quanto riguarda le ramificazioni sono state
indicate a seconda della loro distribuzione sulla superficie dell’osso: generalizzate (Gen) se
si trovano su tutta la superficie; concentrate (Conc) se sono numerose ma tutte nello stesso
punto; isolata (Isol) se è presente solo una stria; disperso (Disp) se si trovano più di una
stria ma distanziate fra di loro.

51
4.2.5 Ossido di manganese
Le alterazioni prodotte da ossido di manganese, portano le superfici ad avere una
colorazione nera, che si può manifestare su tutta la superficie dell’osso o concentrata in un
singolo punto. Questo tipo di alterazione si manifesta principalmente in contesti di grotta o
ripari sotto roccia. L’ossido di manganese si trova infatti in determinati contesti ambientali,
per esempio nelle rocce calcaree (López-González et al. 2006). I composti solubili di
manganese sono mobilitati dall’azione dell’acqua che disintegra la roccia calcarea e ne
permette la percolazione nel sedimento, andandosi a depositare sulla superficie delle ossa
(Burriel et al 1989, Cotton 1993). Hill (1982) sostiene che esistono tre processi che
possono portare alla formazione di depositi di manganese. Il primo è che il manganese in
grotta può derivare dalla circolazione dell’acqua o da piccole quantità di manganese
presente nella roccia calcarea circostante; il secondo processo è determinato dalla
dissoluzione del manganese da parte delle falde acquifere in presenza del CO2, grazie alla
formazione del bicarbonato solubile Mn (HCO3)2. Il terzo è che in contesti di grotta
avviene la perdita di diossido di carbonio, ossidazione ed evaporazione che concentrano il
manganese in soluzioni, dopo che questo è precipitato. In questo lavoro le tracce da
manganese sono state catalogate a seconda della sua distribuzione sulla superficie
dell’osso: generalizzato (Gen), concentrato (Conc), isolato (Isol) e disperso (Disp).
4.2.6 Weathering
Dopo la morte dell’animale e la putrefazione delle parti organiche, vi è un periodo, più o
meno lungo, in cui gli elementi scheletrici sono esposti ad alterazioni dovute
all’esposizione sub-aerea, prima di essere eventualmente inglobati nel sedimento (Caceres
2002). Questo tipo di alterazione chimico-fisica viene indicato con il termine inglese
weathering. Behrensmeyer (1978) lo definisce come il processo attraverso il quale, le
componenti microscopiche organiche ed’ inorganiche originali presenti nell’osso, vengono
progressivamente separate l’una dall’altra e distrutte ad opera degli agenti chimici e fisici
operanti nel sito. Le variazioni di temperatura e umidità sono le principali responsabili di
questo fenomeno che caratterizza alcuni reperti dalla superficie fino alle parti più profonde.
Altri autori indicano che il weathering si riferisce agli effetti sulle ossa dovuti alla
saturazione, essiccazione e cambiamenti della temperatura (Miller 1975).
Per catalogare queste alterazioni, sono stati utilizzati gli stadi indicati da Behrensmeyer
(1978). Lui ha definito sei stadi di weathering sulle ossa di mammiferi > di 5 kg di peso

52
corporeo, utilizzando criteri macroscopici per effettuare con facilità tale riconoscimento
(Tab. 4.1).
Stadi Descrizione Range in anni
dalla morte
0 Niente fessure o esfoliazioni, forse presenza di pelle o legamenti, osso ancora umido. 0-1
1 Fratture parallele all’asse longitudinale dell’osso. 0-3
2 Esfoliazioni sulla parte esterna dell’osso, fessure presenti, bordo delle fratture è angolare. 2-6
3 Osso compatto alterato, ruvido omogeneamente, porta a una tessitura fibrosa; weathering
penetrato 1-1,5 mm massimo; bordi delle fratture arrotondate.
4-15
4 Fibrosità grossolana e superficie ruvida; schegge d’osso lungo la superficie, con weathering
penetrato all’interno delle cavità; fessure aperte.
6-15
5 Osso frammentato in situ, presenti grandi schegge, materiale osseo molto fragile. 6-15
Tab. 4.1 Stadi di weathering nei grandi mammiferi (Behrensmeyer 1978).
Questo tipo di alterazione deve essere considerato come uno dei processi biostratinomici
più importanti che può colpire lo scheletro degli animali.
4.3 Tracce di origine antropica
4.3.1 Le tracce di macellazione
Le tracce di origine antropica sono legate alle modalità di abbattimento dell’animale e al
suo sfruttamento. Per quanto riguarda l’abbattimento degli animali selvatici, è possibile
trovare le tracce dell’oggetto contundente che ha colpito l’animale, ad esempio la punta di
una freccia o una lancia. Per quanto riguarda le forme domestiche, per gli animali di grossa
taglia è possibile trovare un foro o una depressione nella regione frontale del cranio,
mentre per le forme di taglia piccola, si potranno trovare delle tracce nelle prime due
vertebre cervicali, atlante ed epistrofeo, il che può testimoniare un’avvenuta decapitazione
o sgozzamento (De Grossi Mazzorin, 2008). Per quanto riguarda lo sfruttamento, l’attività
umana produce sulle ossa una serie di tracce la cui localizzazione consente spesso di
ricostruire le tecniche di macellazione adottate (Binford 1981), come lo spellamento,
smembramento, disarticolazione, eviscerazione e la scarnificazione. A volte, lo
smembramento e la disarticolazione, possono essere omessi per gli animali di piccola
taglia, in quanto sono finalizzati a favorire il trasporto delle parti più ricche di carne (Kent
1993, Rosell 2001). I processi della macellazione sono (Càceres 2002):
Spellamento o scuoiatura: consiste nell’estrazione della pelle dell’animale. I tagli
sono situati in zone con un indice basso di carne e muscoli, ad esempio nel cranio
(mascella, mandibola, base di corna e palchi), metapodi e falangi (Binford 1981).

53
Smembramento: divisione dell’animale in parti più piccole. Normalmente consiste
nella separazione del cranio, del tronco e delle estremità. I segni, generalmente tagli e
incisioni, si localizzano nelle aree vicino le articolazioni del cranio con il collo e dei cinti
con le estremità.
Disarticolazione: separazione dei differenti elementi scheletrici di ogni membro. I
segni solitamente appaiono nelle epifisi e metafisi delle ossa lunghe. Si tratta di incisioni
profonde, corte e oblique.
Eviscerazione: estrazione delle viscere, sia all’interno della gabbia toracica che del
cranio. Le strie si possono trovare nella parte ventrale delle coste, nelle vertebre toraciche e
lombari.
Scarnificazione: estrazione della carne dall’osso. I segni di taglio si situano nelle
zone di inserzione muscolare o nelle diafisi delle ossa, con un orientamento parallelo e
obliquo all’asse longitudinale dell’osso.
Tutte queste fasi legate allo sfruttamento della carcassa, prevedono l’utilizzo di varie
tipologie di strumenti come lame, coltelli e asce, i quali possono essere di materiale diverso
come selce, rame, bronzo o ferro a seconda del periodo storico che si sta studiando.
Strumenti litici e metallici lasciano strie di tipo diverso, è importante quindi fare tale
discriminazione per capire che tipo di strumento è stato utilizzato (Binford 1981, Càceres
2002, Shipman y Rose, 1983). I tipi di tracce che si possono riscontrare sono (Càceres,
2002):
Incisioni: strie sottili di lunghezza, larghezza e profondità variabile. Si produce con
il contatto tra l’utensile e la superficie dell’osso mediante un movimento continuo del
taglio. La loro disposizione può essere isolata, raggruppata in uno o più gruppi,
l'orientamento può essere obliquo, trasversale e longitudinale rispetto all'asse principale
dell'osso.
Raschiatura: serie di incisioni poco profonde, ampie e parallele. Sono frutto di un
movimento che è distribuito lungo la superficie dell’osso in un direzione perpendicolare al
loro asse longitudinale. Questi segni si associano ad una azione di rimozione del periostio.
Taglio: incisione corta, ampia e profonda. Questo taglio è frutto dell’azione di un
colpo dello strumento sopra la superficie dell’osso diretto e perpendicolare, mirato a
tranciare l’osso e a sezionare le fibre muscolari.
Incisioni insistite: serie di incisioni corte e profonde, concentrate e sovrapposte.
Presentano un orientamento trasversale e obliquo frutto di un movimento ripetuto e
continuo dello strumento sopra la superficie dell’osso. Questo tipo di segni di taglio è

54
associato alla rimozione dell’inserzione muscolare o di altri elementi fortemente aderenti
all’osso.
Occorre precisare che le tracce di questo tipo, sono per lo più accidentali, infatti durante la
macellazione è opportuno evitare il contatto tra le superficie dell’osso e il filo dello
strumento, litico o metallico che sia, in quanto contatti ripetuti di questo tipo determinano
la perdita della capacità di taglio dello strumento (Giacobini, 1995).
Come detto pocanzi, strumenti di materie prime diverse (strumento litico e strumento
metallico), lasciano sulla superficie dell’osso strie di tipo diverso. L’accurata osservazione
delle tracce al microscopio a scansione elettronica, può in alcuni casi, permettere di risalire
al tipo di strumento che l’ha prodotta e addirittura al tipo di materia prima utilizzato
(Greenfield 2006, Mascaro, Perusin, Sarti 2009). L’azione di scorrimento del bordo
tagliente di uno strumento litico su una superficie ossea determina la formazione di un
solco che presenta schematicamente una sezione a V con pareti relativamente scoscese e
profondità almeno pari alla larghezza. Spesso in seguito ad una posizione più o meno
inclinata dello strumento rispetto alla superficie ossea le due pareti della V appaiono
disuguali e il solco presenta una parete più scoscesa dell’altra. Il profilo trasversale del
solco corrisponde a quello del margine attivo dello strumento che ha prodotto la stria.
Durante l’esecuzione del taglio, parti diverse del margine possono entrare in contatto con
la superficie ossea, il che porta ad una grande variabilità della superficie del solco; lo
stesso può accadere quando nel bordo dello strumento ci sono delle microfratturazioni
(Giacobini 1995, D’Errico 1989). La principale caratteristica micromorfologica delle
tracce lasciate da uno strumento litico, con valore diagnostico, consiste infatti nella
presenza di sottili striature secondarie all’interno del solco principale dovuta alla forma del
bordo attivo dello strumento litico (Shipman 1981, Olsen &Shipman 1988).
Per quanto riguarda gli strumenti in metallo, sono state fatte diverse sperimentazioni, per
analizzare le differenze fra tracce provocate da strumento litico e quelle provocate da
strumento in metallo. Quest’ultimi hanno dimostrato di essere più efficaci sotto diversi
punti di vista: sono più forti, più longevi, conservano il filo della lama per più tempo, sono
più affilati e richiedono meno energia per tagliare i tessuti molli (Walker, 1978). Al
contrario, gli strumenti in pietra hanno un potere tagliente più breve (Brose, 1975).
Le strie da strumento metallico sono più profonde e inclinate, inoltre possono avere una
terminazione aguzza/appuntita, oppure a base piatta. Anche questi tipi di tracce hanno una
sezione a V, ma con striature secondarie più fini e meno evidenti (De Grossi Mazzorin

55
2008). Alle estremità si assottigliano, dovuto al movimento dello strumento, in entrata e in
uscita. Il taglio può essere più profondo e stretto oppure profondo e largo, dipende dalla
natura della superficie tagliente dello strumento (Greenfield 1999).
Tutto questo è confermato da analisi in microscopia elettronica a scansione, finalizzate alla
discriminazione di tracce da strumento litico e quelle da strumento in metallo. Qui viene
riportata l’analisi effettuata nel sito archeologico di Via Bruschi, Sesto-Fiorentino, Firenze
(Mascaro, Perusin, Sarti 2009). Per quanto riguarda le tracce da strumento litico, nelle
immagini al SEM (Fig. 4.1) è molto evidente la sezione ampia e molto irregolare della
traccia, caratterizzata da una serie di strie secondarie dovute alle irregolarità del margine
tagliente.
Fig. 4.1 A: porzione medio-distale di tibia di Ovis vel Capra con serie insistita di strie derivate
dall’azione di uno strumento litico. B: ingrandimento allo stereomicroscopio. C: ingrandimento al
SEM; molto evidenti le strie secondarie prodotte dalle irregolarità del margine tagliente dello strumento (Mascaro, Perusin, Sarti 2009).
Per quanto riguarda invece le tracce da strumento metallico, l’immagine al SEM (Fig. 4.2)
raffigura la sezione delle incisioni mettendo in evidenza ancora una volta, alcuni elementi
caratteristici: l’accentuata profondità del segno, il profilo a base leggermente appiattita,
dovuto probabilmente ad una lama non perfettamente affilata, ed i margini lisci e uniformi
(Mascaro, Perusin, Sarti 2009).

56
Fig. 4.2 A: porzione di vertebra toracica di bovino con tracce imputabili all’azione di uno
strumento metallico. B: ingrandimento allo stereomicroscopio (il segmento in nero rappresenta il
punto di sezione di fig. 4.2 C). C: ingrandimento al SEM della sezione delle tracce (Mascaro, Perusin, Sarti 2009).
Esistono anche altri fattori esterni che possono influenzare la morfologia delle strie, come
la forza impiegata nell’azione del taglio e l’età dell’animale. Infatti, in un individuo di età
giovanile, i tagli presentano un lieve rialzamento dei bordi, dovuto al fatto che la parte
corticale non è del tutto compatta, risulta infatti molto porosa, offre quindi meno resistenza
al taglio (Caceres 2002).
A volte la discriminazione delle tracce può essere difficoltosa a causa dello stato di
conservazione del reperto osseo. Infatti le tracce di origine antropica, possono essere
occultate da vari fattori tafonomici pre o post deposizionali i quali determinano
l’asportazione di parti di reperti ossei originariamente recanti tracce di macellazione. In
linea generale questi fattori possono essere dati da azioni di rosicchiatura da parte di
carnivori e roditori, variazioni del grado di umidità che hanno portato a cambiamenti
dimensionali dell’osso e quindi aver favorito il distacco per esfoliazione di strati
superficiali della parte corticale, effetti dati da attività erosive, dal weathering e da effetti
abrasivi causati dal calpestio (Giacobini 1995).
In questo lavoro le tracce di macellazione sono state catalogate indicando:
La tipologia della traccia riconosciuta (incisione, raschiatura, taglio, incisioni
insistite);
Il numero delle tracce incontrate;

57
La loro disposizione, se sono isolate o aggruppate;
Il loro orientamento rispetto all’asse principale dell’osso, che può essere
trasversale, obliquo e parallelo;
La localizzazione, cioè in quale punto dell’osso sono state riscontrate.
4.3.2. Fratture
Un altro tipo di traccia, dovuto all’azione antropica è dato dalle fratture dell’elemento
osseo. Le ossa vengono frantumate per ottenerne il midollo osseo per l’alto valore nutritivo
che da esso si ricava o come materia prima per la lavorazione di strumenti. La morfologia
delle fratture dipende da tre fattori caratteristici:
Il tipo di forza che genera la frattura;
Lo stato dell’osso al momento della fratturazione;
Il tipo di tecnica impiegata per la fratturazione.
La frattura si viene a formare per un eccesso di tensione nella struttura ossea (Morlan
1983), determinata dall’azione di un colpo con un percussore sia di materiale duro che di
materiale tenero. Lo stato dell’osso, fresco o secco, al momento della frattura, è un
elemento fondamentale che ne determina la morfologia. Le fibre del collagene infatti,
forniscono una certa plasticità, in modo che le fratture in un osso fresco risultino allineate
in direzione della struttura dell’osso stesso, e la superficie del piano di frattura presenti una
tessitura “liscia-gradevole” (Johnson 1983). Le fratture a secco, essendoci una diminuzione
di elasticità dovuta alla perdita delle fibre di collagene, risulta essere con angoli retti o
misti e una superficie rugosa o a scanalature (Càceres 2002, Johnson 1983).
Queste caratteristiche sono state utilizzate da diversi autori per elaborare differenti metodi
di catalogazione e identificazione degli agenti responsabili delle fratture nei complessi
archeofaunistici (Villa y Mahieu, 1991; Capaldo, 1998; Alcántara et al., 2006). In questo
lavoro è stata realizzata l’analisi delle fratture di tutti i frammenti superiori a 20 mm di
lunghezza, prendendo in considerazione le seguenti caratteristiche:
Delineazione della frattura: contorno dell’osso dopo la frattura. Il piano di frattura
può essere longitudinale, trasversale o curvato, rispetto l’asse longitudinale dell’osso;
Angolo di frattura: si forma per l’intersezione del piano della superficie corticale e
la sezione midollare. Può essere obliquo, retto o misto.
Tessitura della frattura: può essere liscio o irregolare, questo aspetto è importante
perché si può capire se l’osso era fresco o secco al momento della fratturazione.

58
Per quanto riguarda le tecniche di percussione, ne esistono di due tipi, la percussione attiva
e quella passiva. La percussione attiva consiste nel percuotere l’osso direttamente contro
un altro oggetto; mentre nella percussione passiva, l’osso viene posto tra un incudine e
l’azione del percussore. Un altro modo per fratturare l’osso è quello per flessione o
peeling, viene riscontrato principalmente nelle coste in quanto la flessione può essere
esercitata con le mani. Il risultato sono estremi di frattura scheggiati (White, 1992). Questi
tipi di fratture, possono generare una serie di segni sulla superficie dell’osso, e una serie di
frammenti ossei con morfologie diagnostiche che indicano un intervento ti tipo antropico
(Turner, 1983; Pickering, 2006).
I segni principali dovuti ad una percussione sono:
Punto di impatto: depressione con strie microscopiche, concentriche o parallele alle
fibre di collagene. Sono causate per il collasso del corticale nella zona in cui l’osso ha
ricevuto la maggior forza del colpo (Peretto et al 1996).
Coni di percussione: schegge di tessuto compatto. Sono il frutto dell’azione della
forza massima esercitata per un colpo e la conseguente depressione del tessuto osseo.
4.3.3 Bollitura
All’interno dei campioni faunistici oggetto di analisi di questo lavoro, alcune ossa
sembrano presentare tracce di bollitura. Questa interpretazione rimane attualmente del tutto
ipotetica, alcune ossa sono state classificate come bollite poiché sembrano presentare le
caratteristiche di seguito descritte. L’esposizione indiretta delle ossa al fuoco a bassa
temperatura (≤ 100 C), legata ad attività culinarie, produce modifiche macroscopiche sulla
superficie delle ossa. In passato, tali modifiche non sono mai state descritte in maniera
sistematica (Solari et al 2013). Quando l’osso è esposto all’azione di bollitura, la superficie
diventa liscia, compatta, con un aspetto vetroso. Il colore diventa giallastro, inoltre quando
viene esposto ad una fonte di luce diretta, i bordi diventano traslucidi (Botella et al. 2000).
Un’altra particolarità è che quando viene battuto contro qualcosa, il suono emesso è
analogo a quello di un frammento di ceramica o di un pezzo di legno secco (Botella et al.
2000). A tal proposito sono stati fatti diversi studi sperimentali, per cercare di capire se un
osso è stato bollito o meno. Dalle sperimentazioni di Bosch et al (2011), è stato appurato
che le ossa trattate a bassa temperatura presentano delle caratteristiche macroscopiche
peculiari, si presentano più lisce, leggere e trasparenti; queste modifiche macroscopiche
corrispondo a delle modifiche a livello microscopico tessiturale. Bosch et al (2011) hanno
analizzato ossa umane contemporanee, bollite a diversi intervalli di tempo, poi analizzate

59
attraverso microscopia elettronica a scansione (SEM), spettroscopia a raggi X a dispersione
di energia (EDS), diffrazione di raggi X (XRD) e assorbimento di azoto (BET) e
confrontate poi con il materiale, del sito archeologico di età neolitica, della grotta di
Malalmuerzo, situato nella zona di Granada (Jiménez Brobeil, 1990; Ramos Muñoz, 1988-
1989). È stato constatato che l’ebollizione porta ad una stabilizzazione del collagene,
attraverso la matrice minerale, e alla modificazione della componente organica dell’osso,
rendendo la superficie più morbida e liscia con una bollitura di 4 h. Nel campione
archeologico le ossa neolitiche sono state divise in due gruppi, uno che presenta una
superficie disomogenea, mentre l’altro una superficie liscia ed omogenea. Questi gruppi
corrispondono rispettivamente alle ossa non bollite e bollite. Le ossa non bollite risultano
essere più danneggiate, in seguito all’alterazione provocata dalle radici e dai funghi; quelle
bollite appaiono meno degradate. Questo è dovuto al fatto che grazie alla stabilizzazione
del collagene, avviene la chiusura dei micropori che rallenta notevolmente i processi
diagenetici, rendendo l’osso più resistente a danneggiamenti esterni. Grazie a questa
sperimentazione è stato dimostrato che i criteri macroscopici per distinguere un osso
bollito da uno non bollito, coincidono con evidenze di tipo microscopiche (Bosch et al
2011).
4.3.4 Combustione
In entrambi i siti oggetto di analisi in questo lavoro, sono state trovate molte ossa
combuste. Le alterazioni che si verificano nei resti faunistici, quando sono esposti ad una
fonte di calore, sono molto diversificate, infatti possono portare ad un cambiamento della
colorazione, fessurazione e deformazione, e ad un alto grado di frammentazione che può
portare anche alla completa distruzione (Caceres 2002).
Cambio di colorazione. Il colore dell’elemento osseo cambia a seconda della temperatura e
del tempo di esposizione alla fonte di calore (Nicholson, 1993; Cáceres, 2002). I cambi di
colorazione si devono alla diversa risposta che offrono i componenti organici e inorganici
dell’osso alle alte temperature. Così il colore marrone si relazione con l’emoglobina
(Gejvall 1969), il colore nero testimonia la perdita di ossigeno, il che porta alla
carbonizzazione, (Mayne 1997), il bianco, che testimonia un avvenuta calcinazione, è
determinato dalla decomposizione dei componenti organici, che avviene a temperature
altissime (Shipman et al 1984).
In questo lavoro le ossa combuste sono state catalogate a seconda del loro grado di
colorazione:

60
Grado 1: la superficie ossea presenta macchie disperse di colore marrone;
Grado 2: il colore marrone è uniforme su tutta la superficie dell’osso;
Grado 3: l’intera superficie ossea è carbonizzata, presenta un colore nero uniforme;
Grado 4: il colore della superficie è grigio;
Grado 5: osso calcinato, colore bianco su tutta la superficie dell’osso.
Bisogna precisare che i processi di combustione, non sono le uniche alterazioni che
possono portare ad un cambiamento della colorazione, infatti anche l’ossido di manganese
porta ad una colorazione nera più o meno estesa sulla superficie dell’osso. È quind i
importante riuscire a fare questo tipo di discriminazione, per ricreare le modalità di
formazione del campione faunistico in esame (Shahak-Gross, 1997).
Fessurazione e deformazione. Sulla superficie degli elementi scheletrici, quando sono
sottoposti ad alte temperature, si vengono a formare delle fessure. Queste fessure tendono
ad avere un orientamento longitudinale lungo la diafisi, mentre lungo le epifisi tendono ad
avere un orientamento disordinato, quasi ramificato. Quest’ultime possono essere confuse
con quelle determinate dall’esposizione subaerea (Behresmeyer 1978). Alcuni autori hanno
cercato di descrivere queste fessure per facilitarne la discriminazione, affermando che le
fessure determinate da alte temperature sono più profonde rispetto a quelle determinate dal
weathering. In particolare le fessure sono più profonde quando l’osso, al momento della
cremazione, era fresco, inoltre l’orientamento oltre a essere longitudinale può essere anche
trasversale (Buikstra & Swegle 1989).
Le ossa sottoposte all’azione del fuoco, possono subire anche delle deformazioni, in
particolare la riduzione delle dimensioni. La riduzione può variare tra il 5% e il 32% della
dimensione originale, colpisce principalmente le ossa piatte e la diafisi delle ossa lunghe
(Shipman et al 1984, Buikstra & Swegle 1989). Questo tipo di deformazione deve essere
valutato con precisione quando bisogna prendere le misure osteometriche, in quanto è
facile incappare in errori di valutazione (Caceres 2002).
Frammentazione. Le proprietà fisiche dell’osso, sono basate su un mix di compattezza ed
elasticità, che garantisce la resistenza agli stress e l’assorbimento degli shock meccanici
(De Grossi Mazzorin 2008). Quando l’osso è combusto, perde queste proprietà e ne
favorisce la frammentazione e, quindi, ha una minor resistenza alla rottura (Stiner et al.
1995). La frammentazione può avvenire semplicemente per l’esposizione ad alte
temperature, partendo da un alto grado di fessurazione (Nicholson 1993), oppure per cause
esterne come per esempio il trampling. Quando l’osso raggiunge il grado 5 di
combustione, quindi raggiunge uno stato di calcinazione, va incontro alla frammentazione

61
o addirittura ad una completa distruzione, semplicemente per la pressione del sedimento
dopo essere stato interrato (Stiner et al. 1995).

62

63
Capitolo 5 “Registrazione dei dati”
5.1 Introduzione
Nel seguente capitolo verrà descritto il database che è stato utilizzato per la registrazione e
la catalogazione dei reperti analizzati. Verrà inoltre intrapresa una breve riflessione
sull’importanza dell’utilizzo di un database e i vantaggi che ne derivano.
5.2 Utilizzo di un database
L’utilizzo di un database è diventato ormai di fondamentale importanza nella ricerca
archeologica e archeozoologica moderna. I vantaggi nell’utilizzo dell’archivio
informatizzato sono dati dalla velocità di archiviazione dei dati con minime possibilità di
errore nella fase di immissione, il controllo continuo attraverso liste di facile costruzione,
riuscire ad effettuare in maniera agevole la combinazione tra campi con relativa
elaborazione (conteggi, creazione di tabelle e grafici) (Boscato et alii, 2001). Un altro
fattore importante, il quale è stato essenziale per la realizzazione di questo lavoro, è la
possibilità di un immediato confronto con dati archiviati provenienti da siti diversi. Inoltre
possono rappresentare una risorsa comune, in quanto possono essere accessibili a
molteplici utenti.
5.3 Registrazione dei dati
La gestione dei dati per la realizzazione del presente lavoro, è stata effettuata tramite
l’utilizzo di un database realizzato tramite il programma FileMaker Pro Advanced,
adoperato dal gruppo di ricerca dell’IPHES/Università Rovina i Virgili di Tarragona
(Spagna). Per meglio analizzare i dati in modo incrociato tale descrizione è stata resa
alfanumerica sulla base della creazione di varie categorie di classificazione (per esempio
taxon, elemento anatomico, porzione).
Il database è suddiviso in diverse schede, le quali rappresentano diverse tipologie di dati.
La prima scheda viene utilizzata per ogni tipologia di materiale (ossa, ceramica, litica,
etc.), nella quale vengono riportati i dati che sono stati registrati nella scheda di scavo, cioè
le informazioni generali del reperto (nome del sito, strato, anno, quadrato, numero,
materiale, categoria, taxon, coordinate, orientamento, quote e dimensioni) (Fig. 5.1).

64
Fig. 5.1 Database FileMaker pro Advanced
All’interno del database, vi sono delle schede dedicate all’analisi e alla registrazione dei
dati per ogni tipo di materiale rinvenuto. Di seguito vengono elencate quelle relative al
materiale faunistico:
Scheda generale: vengono inseriti i dati relativi al riconoscimento tassonomico e
anatomico, riportando anche le categorie di taglia, la porzione, la parte conservata, la
posizione e l’età (Fig. 5.2).
Fig. 5.2 Scheda generale FileMaker Pro Advanced

65
Scheda fratturazione: in questa scheda vengono catalogate il numero di fratture che
sono presenti nel reperto, e per ogni frattura vengono descritte le caratteristiche
(delineazione, angolo e tessitura) (Fig. 5.3).
Fig. 5.3 Scheda fratturazione FileMaker Pro Advanced
Scheda tracce di macellazione e combustione: vengono indicati il numero di tracce
riconosciute, la tipologia (incisioni, raschiature, tagli e incisioni insistite), la disposizione,
l’orientazione e la localizzazione. Inoltre è possibile registrare le finalità delle tracce
(scarnificazione, disarticolazione, spellamento, eviscerazione). In questa scheda viene
inserito anche il grado di combustione (1, 2, 3, 4, 5) e nel riquadro relativo alla descrizione
viene riportato se l’osso ha subito i processi di bollitura (Fig. 5.4)
Fig. 5.4 Scheda tracce di macellazione e combustione FileMaker Pro Advanced

66
Scheda tracce di morso: vengono indicate le varie tipologie di morsicatura (solchi,
depressioni, impronte), la loro localizzazione (corticale, bordo di frattura), la disposizione
(aggruppate, disperse), e se sono state determinate da carnivori o roditori (Fig 5.5).
Fig. 5.5 Scheda tracce di morso FileMaker Pro Advanced
Altre modifiche: in questa scheda vengono inserite altre tracce di origine naturale,
come le tracce da radici, trampling, ossido di manganese e concrezioni. Per ognuna di
queste tracce viene catalogata la loro disposizione sulla superficie dell’osso (generalizzata,
concentrata, isolata, dispersa) (Fig. 5.6).
Fig. 5.6 Altre modifiche FileMaker Pro Advanced

67
Scheda rimontaggio, localizzazione, osservazione: viene utilizzata in caso di
restauro del reperto e per inserire osservazioni di carattere generale (Fig. 5.7).
Fig. 5.7 Scheda rimontaggio, localizzazione, osservazione FileMaker Pro Advanced

68

69
Capitolo 6
“Il campione faunistico di Vallone Inferno”
6.1 Introduzione
Nel seguente capitolo vengono illustrate le analisi archeozoologiche effettuate sui reperti di
grandi mammiferi recuperati durante le campagne di scavo 2012, 2013 e parte del 2014 del
sito di Vallone Inferno. La fauna analizzata in questo lavoro proviene dallo strato 3.4.A,
scavato nel 2012, e dallo strato 3.4.B, scavato nel 2013 e terminato nel 2014. Questi strati
presentano una cronologia relativa che comprende la fase finale del Bronzo Antico e
l’inizio del Bronzo Medio. È stata recuperata anche un’età al radiocarbonio effettuata su un
seme, proveniente dallo strato 3.4.B, che ha restituito una datazione di 1616-1433 cal. BC.
6.2 La fauna analizzata
Il campione faunistico analizzato a Vallone Inferno è costituito da un NR pari a 1261, di
questi, 407 sono stati riconosciuti totalmente (NRDt), cioè sia a livello anatomico che
tassonomico, ossia il 32% del totale, 390 sono stati riconosciuti a livello anatomico
(NRDa), pari al 31%del totale. E’ stato possibile inserire 283 frammenti nelle categorie
osso lungo, osso piatto e osso articolare, pari al 22%, mentre le restanti 181 ossa sono state
classificate come indeterminate, ossia circa il 14% (Fig. 6.1.) (Tab. 6.1).
Fig. 6.1 NRDt, NRDa, categorie osso lungo, piatto e articolare, Indet, del campione faunistico di Vallone
Inferno proveniente dagli stati 3.4.A e 3.4.B.
0%
5%
10%
15%
20%
25%
30%
35%
NRDt NRDa Osso lungo,piatto e
articolare
Indet

70
Vallone Inferno 3.4.A 3.4.B TOT %
NRDt 204 203 407 32%
NRDa 202 188 390 31%
Osso lungo, piatto e
irregolare
166 117 283 22%
Indet 122 59 181 14%
Tot 694 567 1261 100%
Tab. 6.1 NR di Vallone Inferno
In figura 6.1 possiamo vedere che per la maggior parte del campione faunistico è stato
possibile effettuare il riconoscimento a livello tassonomico ed anatomico. Questo grazie sia
alla presenza di molti denti ritrovati, per un totale di 165, pari al 13% del totale, i quali
nella maggior parte dei casi hanno consentito il riconoscimento a livello specifico; il
riconoscimento è stato effettuato anche grazie alla presenza di molte parti diagnostiche
delle ossa lunghe, sia epifisi che metafisi.
Di seguito viene riportata la tabella che riassume gli elementi scheletrici riconosciuti a
livello anatomico (NRDa) (Tab. 6.2).
Sub-livelli 3.4.A 3.4.B NRDa %
ELEMENTI
ANATOMICI
Cranio 32 77 109 28%
Vertebra 41 39 80 21%
Mascella / 1 1 1%
Emimandibola 1 4 5 1%
Molare/premolare / 1 1 1%
Denti 1 / 1 1%
Coste 103 41 144 37%
Scapola / 2 2 1%
Omeri 6 3 9 2%
Radio 6 5 11 3%
Ulna / 1 1 1%
Metapodiali 4 2 6 2%
Falange 2 1 3 1%
Sacro 2 / 2 1%
Bacino 1 5 6 2%
Femore / 4 4 1%
Tibia 2 2 4 1%
Carpale/tarsale 1 / 1 1%
TOT 202 188 390 100%
Tab. 6.2 NRDa nei sub-livelli 3.4.A e 3.4.B.
In tabella 6.2 è possibile notare come gli elementi anatomici dello scheletro assile siano più
rappresentati rispetto allo scheletro appendicolare. Le ossa più rappresentate sono le coste
con il 37%, questa percentuale così alta è dovuta ad un alto grado di frammentazione,

71
dovuto sia a fattori antropici sia naturali, troviamo poi il cranio (28%) e le vertebre (21%).
Tutti gli altri elementi presentano delle percentuali bassissime, ascrivibili tra l’1 e il 3%.
È stato effettuato anche il conteggio delle ossa che sono state inserite all’interno delle
categorie osso lungo, piatto e articolare, di seguito viene riportato il grafico (Fig. 6.2).
Fig. 6.2 Categorie osso lungo, osso
piatto e osso articolare degli strati 3.4.A
e 3.4.B.
Dal grafico di figura 6.2 possiamo vedere come le ossa lunghe siano quelle più
rappresentate, raggiungono infatti il 69%, la categoria osso piatto è rappresentata per il
31%, mentre le ossa articolari sono in pratica rappresentate da un unico elemento.
Per le ossa che sono state riconosciute a livello anatomico e per quelle inserite nelle
categorie osso lungo, piano e articolare, è stato possibile attribuire una categoria di taglia
come si evince dal grafico di figura 6.3.
Fig. 6.3 Categoria di taglia, sub-livelli 3.4.A e 3.4.B.
Le ossa di taglia piccola siano le più rappresentate con circa il 65%. Dalla tabella presente
nel Capitolo 3, paragrafo 3.2.2, è possibile notare come gli animali di taglia piccola siano
compresi tra i 100 e i 10 kg, questa categoria include animali domestici come ovicaprini e
suini di età giovanile, animali selvatici come cervidi di età giovanile. Seguono animali di
0%
10%
20%
30%
40%
50%
60%
70%
Osso lungo Osso piatto Osso articolare
0%
10%
20%
30%
40%
50%
60%
70%
Taglia grande Taglia media Taglia piccola Taglia moltopiccola

72
taglia media con il 16%, percentuali ancora più basse le troviamo per animali di taglia
grande (5%), e animali di taglia piccola (12%).
Nella seguente tabella vengono riassunte le specie a grandi mammiferi riconosciute nei
sub-livelli 3.4.A e 3.4.B (Tab. 6.3). Di ogni specie viene riportato il NRDt, e la relativa
percentuale di abbondanza. Le percentuali vengo poi riportate nel grafico di figura 6.4, per
fornire con maggior chiarezza il dato sulle specie rappresentate.
Sub-livelli 3.4.A 3.4.B NRDt %
Taxa
Canide 10 4 14 3%
Felis sp. 1 1 2 1%
Sus scrofa 1 1 1%
Cervus elaphus 15 20 36 9%
Sus domesticus 27 13 40 10%
Bos taurus 7 16 23 6%
Capra hircus 1 1 1%
Ovis aries 1 5 6 1%
Ovicaprini 140 138 278 68%
Artiodattili 3 4 7 2%
Tot 204 203 407 100%
Tab 6.3. Grandi mammiferi recuperati nei livelli
3.4.A e 3.4.B.
Fig. 6.4 Percentuali di abbondanza dei mammiferi dei sub-livelli 3.4.A e 3.4.B.
Dalla tabella 6.3 e in figura 6.4 si evince come gli ovicaprini siano i taxa più rappresentati
in entrambi i sub-livelli analizzati, con il 68% del NRDt, rappresentano infatti, la base
fondamentale della pratica di allevamento. In alcuni casi è stato possibile riconoscere Ovis
aries e Capra hircus, grazie ai quarti premolari decidui e agli astragali. Seguono
abbastanza distaccati i maiali (10%) i cui resti sono stati trovati principalmente nello strato
3.4.A. Tuttavia, anche se subordinati rispetto agli ovicaprini, anch’essi dovevano aver
rappresentato una specie da allevamento per gli uomini del riparo. Una percentuale più
0%
10%
20%
30%
40%
50%
60%
70%

73
bassa la troviamo per Bos taurus (6%), questo lascia presupporre che si dovesse trattare di
un animale allevato solo raramente probabilmente a causa della sua grande taglia che
necessita di uno spazio di allevamento maggiore di quello offerto dal riparo. Tra gli
animali selvatici quello più rappresentato è Cervus elaphus (9%); in un solo caso è stato
possibile riconoscere il cinghiale, Sus scrofa, in particolare grazie a un canino che date le
dimensioni è stato attribuito alla forma selvatica. Questo testimonia una pratica secondaria,
ma pur sempre attiva della caccia. Infine sono stati trovati anche dei resti appartenenti a
carnivori, in particolare canidi (3%) e un solo resto attribuito ad un felide (1%), ma dato
l’esiguo numero dei resti, non è stato possibile effettuare la discriminazione tra forma
selvatica e domestica.
Di seguito vengono descritti i taxa riconosciuti, prima verranno trattate le forme
domestiche secondo un ordine di abbondanza, in seguito le forme selvatiche e infine i
carnivori.
6.3 Ovicaprini
Nel campione faunistico di Vallone Inferno analizzato in questo lavoro, gli ovicaprini sono
gli animali più rappresentati con il 68%, costituiscono perciò la base economica più
importante. Il NRDt equivale a 277, mentre il NMI è uguale a 13. Nella seguente tabella
vengono riportati gli elementi ossei riconosciuti, nei due sub-livelli, 3.4.A e 3.4.B (Tab.
6.4).
Sub-livelli 3.4.A 3.4.B NRDt % NME %
ELEMENTI
ANATOMICI
OVICAPRINI
Cranio 2 5 7 4%
Mascella 1 1 1% 1 2%
Emimandibola 8 10 18 10% 6 12%
Ioide 2 2 1%
Vertebra 1 1 1%
Costa 1 1 1%
Scapola 1 3 4 2% 2 4%
Omero 11 13 24 13% 12 4%
Radio 8 9 17 9% 6 12%
Ulna 2 2 4 2% 3 6%
Metacarpo 5 5 10 6%
Scafoide 1 1 1%
Semilunare 3 3 2%
Piramidale 1 1 2 1%
Uncinato 1 1 1%
Capitato-
trapezoide 3 3 2%
Femore 4 5 9 5% 4 8%
Tibia 6 7 13 7% 7 13%
Calcagno 1 5 6 3% 4 8%
Astragalo 2 5 7 4% 7 13%

74
Scafocuboide 1 1 1%
Metatarso 2 5 7 4%
Metapode 7 10 17 10%
Falange 13 8 21 12%
TOT 74 104 180 100% 52 100%
Tab. 6.4 Elementi anatomici degli ovicaprini di Vallone Inferno.
Dalla tabella 6.4 possiamo notare come lo scheletro appendicolare sia particolarmente
rappresentato, infatti troviamo alte percentuali per le ossa lunghe, spiccano in particolare
gli omeri con una percentuale pari al 13%. Di questo elemento anatomico sono state
ritrovate soprattutto zone della diafisi e delle metafisi. Anche le falangi sono ben
rappresentate (12%), segue il radio (9%), e la tibia (7%). Una buona rappresentazione è
data anche dai metapodi (10%), tanto che in alcuni casi è stato possibile distinguere il
metacarpo (6%), e il metatarso (4%). Inoltre sono state trovate varie ossa del carpo e del
tarso, soprattutto gli astragali rappresentati per il 4%. In tre casi è stato possibile effettuare
la misurazione dell’altezza al garrese, proprio grazie alla misura GLl degli astragali; queste
misurazioni però possono presentare un certo margine di errore, in quanto gli astragali
possono essere riferibili ad animali che non hanno ancora raggiunto l’età adulta (De Grossi
Mazzorin, 2008). Queste misure sono state tutte moltiplicate per il coefficiente di Teichert
(1975). L’altezza stimata per i tre astragali è di: 63,5 cm per il primo, 58,9 cm per il
secondo e 54,4 cm per il terzo. Data l’esiguità del campione il dato non ha valenza
statistica. Per quanto riguarda lo scheletro assile, troviamo percentuali molto basse di
presenza di neurocranio, vertebre e coste, mentre sono ben rappresentate le mandibole con
una percentuale pari al 10%.
Come detto prima, sono state trovate molte parti diafisarie e metafisarie delle ossa lunghe,
questo denota un certo grado di frammentazione. Di seguito viene riportato il grafico con
l’indice di frammentazione scheletrica, che è stato calcolato mettendo in rapporto il NME
con il NRDt; il valore vicino a 1 indica l’elemento poco frammentato, l’indice vicino a 0
indica l’elemento molto frammentato. Inoltre, è stato calcolato l’indice di sopravvivenza
scheletrica, mettendo in rapporto il NME con il NEE, basato sul NMI; il valore vicino a 1
indica lo scheletro completo, il valore vicino a 0 indica lo scheletro incompleto (Fig. 6.5).

75
Fig. 6.5 Indice di frammentazione degli elementi scheletrici e indice di sopravvivenza scheletrica degli
ovicaprini, del sito Vallone Inferno.
Dal grafico di figura 6.5 possiamo vedere come le ossa lunghe risultino essere molto
frammentate, con valori pari a 0,5, ad eccezione dell’ulna che invece presenta un valore
pari a 0,75. Per le ossa con valori pari a 1 troviamo un mascellare e gli astragali; in
particolare quest’ultimi grazie alla loro morfologia, durezza e densità, sono molto più
resistenti alla fratturazione. I motivi della frammentazione possono essere dovuti a cause
naturali o antropiche, ma questo tema verrà trattato nei capitoli successivi. Per quanto
riguarda l’indice di sopravvivenza scheletrica, vediamo che l’omero è l’elemento
anatomico più presente, con un valore pari a 0,5. Le altre ossa lunghe sono meno presenti
con un valore di circa 0,2. Infine, elementi mascellari e scapolari sono fortemente
sottorappresentati.
Per quanto riguarda il calcolo dell’età di morte, i resti dentari hanno restituito dati più
attendibili rispetto ai resti ossei; con quest’ultimi sono state semplicemente create tre classi
d’età (neonato, giovane, adulto), basandosi sulla fusione dell’epifisi e sul grado di porosità
dell’osso corticale (Fig. 6.6).
Fig. 6.6 Calcolo dell’età di morte da resti ossei di ovicaprini di Vallone Inferno.
Dal grafico di figura 6.6 possiamo vedere che le classi d’età giovane e neonato sono molto
più rappresentate rispetto alla classe d’età adulto. Per quanto riguarda i resti dentari, sono
00,20,40,60,8
11,2
NME/NRDt NME/NEE
0
10
20
30
40
50
Neonato Giovane Adulto

76
stati analizzati 97 denti, sia decidui che permanenti, in due casi sono stati trovati inclusi
nell’emimandibola. Di seguito viene riportata la tabella con le percentuali di
rappresentazione dei denti ritrovati (Tab. 6.5).
Sub-livelli 3.4.A 3.4.B NRDt % NME %
ELEMENTI
DENTARI
OVICAPRINI
dI inf 1 1 1%
dI3 dx 1 1 1% 1 1%
I inf sx 1 2 3 3% 3 4%
I inf 2 1 3 3%
I inf dx 1 1 1% 1 1%
I1 dx 3 1 4 4% 4 6%
I1 sx 1 1 1% 1 1%
I3 sx 2 1 3 3% 3 4%
I2/3 sx 1 1 2 2%
I 1 1 2 2%
dP2 sx 1 1 1% 1 1%
dP2 dx 3 3 3% 3 4%
dP2 sx 1 1 2 2% 2 3%
dP2 1 1 1% 1 1%
dP3 dx 5 5 5% 5 7%
dP3 sx 3 1 4 4% 4 6%
dP3 1 1 1%
dP4 dx 1 1 1% 1 1%
dP4 sx 4 3 7 7% 7 10%
P2 dx 1 1 1% 1 1%
P2 sx 1 1 1% 1 1%
P2 dx 1 1 1% 1 1%
P2 sx 1 1 1% 1 1%
P2 1 1 1%
P3 dx 3 3 3% 3 4%
P3 sx 1 1 1% 1 1%
P4 dx 1 1 1% 1 1%
P4 sx 1 1 1% 1 1%
M1 dx 4 4 4% 4 6%
M1 sx 3 3 3% 3 4%
M1 1 1 1%
M1 dx 1 1 1% 1 1%
M1 sx 3 3 3% 1 1%
M1/2 dx 2 2 2%
M1/2 sx 1 1 1%
M1/2 sx 1 1 1%
M1/2 1 1 1%
M2 dx 3 4 7 7% 7 10%
M2 sx 1 1 1% 1 1%
M2 dx 3 3 3% 3 4%
M3 dx 1 1 1% 1 1%
M3 dx 1 1 1% 1 1%

77
M3 sx 1 1 1% 1 1%
M3 sx 1 1 1% 1 1%
M 1 1 2 2%
M sup 2 1 3 3%
M/P 1 2 3 3%
TOT 58 39 97 100% 71 100%
Tab. 6.5 Resti dentari degli ovicaprini di Vallone Inferno.
Dalla tabella 6.5 si evince l’alto grado di varietà di elementi dentari ritrovati, infatti sono
stati riconosciuti incisivi, premolari, dal secondo al quarto, decidui e permanenti. Anche i
molari sono ben rappresentati, infatti troviamo sia il primo, il secondo e il terzo molare.
Alcuni denti erano particolarmente frammentati e non è stato possibile determinare il lato
e/o se erano inferiori o superiori. Il campione dentario è costituito dal 72% di denti
permanenti, mentre il restante 28% è costituito da decidui. Grazie ai denti è stato possibile
calcolare l’età di morte in maniera più attendibile, grazie alle tabelle di Habermehl (1961),
Barone (1980) e Silver (1969), che ne indicano lo stato di eruzione, mentre il metodo
Wilkens (2003) è stato usato per misurare lo stato d’usura. Di seguito viene riportata il
grafico con gli stadi di usura riconosciuti (Fig. 6.7).
Fig. 6.7 Stadi di usura Wilkens (2003) degli
ovicaprini di Vallone Inferno.
Dal grafico di figura 6.7 possiamo vedere come lo Stadio 1 e lo Stadio VII siano quelli più
rappresentati, corrispondo rispettivamente a 1-3 mesi d’età e più di 24 mesi d’età, quindi
alle classi neonato e adulto. Ben presenti sono i denti che appartengo allo Stadio 3 che
corrisponde a un’età di morte pari a 5-8 mesi, appartenente quindi alla classe d’età
infantile. Infine per i denti che presentano lo stadio di usura tra il V e il I, quindi tra 9-18
mesi, possono essere inseriti nella classe d’età giovane. Di seguito viene riportato il grafico
con le classi d’età riconosciute unendo i dati relativi ai denti e ai resti ossei (Fig. 6.8).
Stadio 1
Stadio 2
Stadio 3
Stadio 4Stadio 5Stadio I
Stadio II
Stadio VI
Stadio VII

78
Fig. 6.8 Calcolo dell’età di morte degli ovicaprini di Vallone Inferno.
Dal grafico di figura 6.8 si evince che le classi più rappresentate sono quella neonatale e
quella giovanile, con circa 50 reperti rinvenuti per ogni classe. Anche quella relativa all’età
adulta è ben rappresentata, seguono poi le classi sub-adulto e infantile. Sulla base dei
calcoli dell’età di morte si può ipotizzare che la base economica del riparo era basata su un
allevamento di tipo misto, mirato cioè sia alla produzione di carne, testimoniato dalla
presenza di individui giovani, sub-adulti e neonati, sia alla produzione di latte, testimoniato
dalla grande presenza della classe di età adulta. La classe degli adulti inoltre, oltre a
testimoniare la produzione di latte, probabilmente veniva impiegata anche per la
produzione di lana.
Per quanto riguarda il NMI, è stato calcolato in base al NME sia degli elementi ossei che
dentari, e alla classe d’età. Per i neonati è stato calcolato un NMI pari a 4, grazie ai terzi
premolari decidui destri, la classe infantile presenta un NMI pari a 2, grazie ai primi molari
destri, la classe giovani un NMI uguale a 2, grazie agli omeri destri, la classe sub-adulta è
rappresentata da 3 individui grazie ai molari sia inferiori che superiori sinistri, infine la
classe adulta è rappresentata da un NMI pari a 2, grazie agli astragali sinistri. Quindi il
NMI degli ovicaprini è pari a 13.
6.4 Ovis aries Linnaeus, 1758 & Capra hircus Linnaeus, 1758
In pochissimi casi è stato possibile distinguere le due specie osteologicamente affini,
pecora e capra, tramite tre quarti premolari inferiori, attraverso il metodo Payne (1985),
due di questi appartengono alla pecora (Ovis aries), mentre l’altro appartiene alla capra
(Capra hircus). Utili per tale discriminazione sono stati anche gli astragali, in particolare
ne sono stati trovati tre, tutti appartenenti alla pecora, riconosciuti tramite il criterio 1 del
metodo Zeder & Lapham (2010).
0
10
20
30
40
50
60

79
6.5 Sus domesticus Erxleben, 1777
Il maiale è la seconda specie più rappresentata, con circa il 10% del totale. Il NRDt è 38,
mentre il NMI è pari a 8. Nella seguente tabella vengono riportati gli elementi anatomici
riconosciuti (Tab. 6.6).
Sub-livelli 3.4.A 3.4.B NRDt % NME %
ELEMENTI
ANATOMICI
Sus domesticus
Cranio 1 1 3% Mascella 1 1 3% 1 5%
Emimandibola 3 3 8% 3 14%
Omero 1 1 3% 1 5%
Pelvi 1 1 3% 1 5%
III Metacarpo 1 1 3%
I Falange 1 1 3%
II Falange 1 1 3%
III Falange 1 1 3%
M1 sx 1 1 3% 1 5%
dP4 1 1 3%
dP4 dx 4 4 11% 4 19%
M2 dx 1 1 3% 1 5%
I inf 2 1 3 8%
I inf sx 1 1 3% 1 5%
I inf dx 2 2 5% 2 10%
dC 2 2 5%
P inf 1 1 3%
M1 1 1 3%
P4 1 1 3%
P4 dx 1 1 3% 1 5%
I3 sx 2 2 5% 2 10%
M 1 1 3%
dP3 sx 1 1 2 5% 2 10%
P2 1 1 3%
C inf dx 1 1 3% 1 5%
P sup sx 1 1 3%
TOT 26 12 38 100% 21 100%
Tab. 6.6 Elementi anatomici relativi ai suini di Vallone Inferno.
Dalla tabella 6.6 è possibile notare che gli elementi ossei sono meno numerosi rispetto a
quelli dentari, mentre i primi costituiscono solo il 29% del totale, i resti dentari
costituiscono il restante 71%. Tra gli elementi ossei spiccano le emimandibole, ne sono
state rinvenute tre e in una di queste sono stati trovati i denti ancora all’interno degli
alveoli, si tratta di un individuo di 2-4 settimane con dP3 e dP4. I quarti premolari decidui
sono quelli più numerosi con una percentuale pari al 11%. Questi resti sono stati
considerati appartenenti alla forma domestica, tranne un canino inferiore, che date le
dimensioni è stato attribuito alla forma selvatica. Il calcolo dell’età di morte è stato
effettuato soltanto sui denti, in quanto più numerosi. Anche per i denti di maiale sono state

80
utilizzate le tabelle di Habermehl (1961), Barone (1980) e Silver (1969), per lo stato di
usura è stato utilizzato il metodo Wilkens (2003). Gli stadi di usura sono stati riconosciuti
soltanto in quattro denti, due sono quarti premolari decidui inferiori i quali presentavano un
alto grado di usura, il secondo molare doveva essere in eruzione, quindi è stato attribuito lo
Stadio 4 (età compresa tra gli 8-12 mesi), gli altri due denti invece sono stati inseriti nello
Stadio 3, presentavano perciò un’età compresa tra i 6 e gli 8 mesi. Anche per i suini sono
state riconosciute cinque classi d’età, neonato, infantile, giovane, sub-adulto e adulto (Fig.
6.9).
Fig. 6.9 Calcolo dell’età di morte dei suini di Vallone Inferno.
Dal grafico di figura 6.9 possiamo vedere che la classe d’età dominante è quella infantile,
che comprende individui di circa 6-12 mesi, anche i neonati sono ben rappresentati,
soprattutto grazie ai canini decidui, questa classe comprende individui morti entro il primo
mese di vita. I sub-adulti, rappresentati da cinque reperti, comprendono individui di circa
14-20 mesi. Un unico reperto è stato attribuito alla classe di età adulta. Questo risultato è in
accordo con il profilo di mortalità del maiale che, essendo un animale strettamente rivolto
alla produzione di carne, l’età ideale di macellazione è a circa un anno. Invece, gli animali
adulti, utilizzati per scopi riproduttivi, sono in numero minore. Questo dato è confermato
anche dal NMI, il quale è stato calcolato dai resti dentari prendendo in considerazione la
lateralità e le classi d’età; infatti troviamo più rappresentati individui appartenenti alle
classi neonati e giovani, rispettivamente con 2 e 3 individui, mentre le altre classi sono
tutte rappresentate da un individuo. In totale quindi il NMI dei suini è pari a 8.
6.6 Bos taurus Linnaeus, 1758
Il bue è la specie domestica meno rappresentata di tutto il campione faunistico, infatti
presenta il NRDt pari a 23 corrispondente al 6% del totale (Tab. 6.7).
0
1
2
3
4
5
6
7
8
9
10
Neonati Infantili Giovani Sub-adulti Adulti

81
Sub-livelli 3.4.A 3.4.B NRDt % NME %
ELEMENTI
ANATOMICI
Bos taurus
Mandibola 3 3 13% 1 13%
Costa 1 1 4%
Vertebra 1 1 4%
Omero 2 2 9%
Radio 1 1 4%
Ulna 1 1 4% 1 13%
Metacarpo 2 2 9% 1 13%
Femore 1 1 4%
Astragalo 1 1 4% 1 13%
Semilunare 2 2 9%
Scafoide 1 1 4%
Scafocuboide 1 1 4% 1 13%
I falange 2 2 9%
III falange 1 1 4%
Incisivo inf. 1 1 4% 1 13%
I1 dx 1 1 4% 1 13%
Dp4 sx 1 1 4% 1 13%
TOT 7 16 23 100% 8 100%
Tab. 6.7 Elementi anatomici Bos taurus.
Da questa tabella si evince che sia lo scheletro assile che quello appendicolare sono poco
rappresentati, infatti troviamo soltanto una costa, una vertebra e tre frammenti di
mandibola, quest’ultime costituiscono l’elemento più rappresentato con il 13%, tali reperti
stati trovati esclusivamente nello strato 3.4.B. Per quanto riguarda lo scheletro
appendicolare, gli elementi più rappresentati sono l’omero, il metacarpo, il semilunare e la
prima falange, presentando una percentuale pari al 9%. Gli altri elementi anatomici
rinvenuti, presentano invece una percentuale pari al 4%.
Nonostante il numero esiguo degli elementi anatomici rinvenuti, in alcuni casi è stato
possibile il calcolo dell’età di morte, in particolare tramite gli elementi ossei, prendendo in
considerazione lo stato di fusione delle epifisi e il grado di porosità dell’osso corticale (Fig.
6.10).
Fig. 6.10 Calcolo dell’età di morte di Bos taurus di
Vallone Inferno.
0
2
4
6
8
10
Giovane Adulto

82
Dal garfico di figura 6.10, possiamo notare che sono state riconosciute soltanto due classi
d’età (giovane e adulta), dove quella più rappresentata è sicuramente quella adulta. Inoltre
è stato possibile calcolare il NMI, grazie all’ausilio del NME e prendendo in
considerazione l’età di morte; sono stati calcolati 2 individui, uno per ogni classe.
6.7 Cervus elaphus Linnaeus, 1758
Il cervo rosso rappresenta l’animale selvatico più rappresentato con il 9% del totale. Di
seguito viene riportata la tabella con gli elementi anatomici ritrovati (Tab. 6.8).
Sub-livelli 3.4.A 3.4.B NRDt % NME %
ELEMENTI
ANATOMICI
Cervus
elaphus
Palco 1 1 3%
I Falange 2 3 5 14%
II Falange 1 4 5 14%
III Falange 1 1 3%
Ulna 1 1 3%
Femore 1 1 3% 1 8%
Metacarpo 1 1 3%
Metatarso 3 3 8% 1 8%
Metapode 2 2 6%
Calcagno 2 2 6% 2 17%
Emimandibola 1 1 2 6% 1 8% Tibia 1 1 3% 1 8% Piramidale 1 1 3%
Scafo-cuboide 1 1 3%
I inf 2 2 6%
C sup 1 1 3%
C inf 1 1 3% 8% dP4 dx 1 1 3% 1 8% P3 sx 1 1 3% 1 8% P4 dx 1 1 3% 1 8% M1 sx 1 1 3% 1 8% M1 dx 1 1 3% 1 8% TOT 15 21 36 100% 12 100%
Tab. 6.8 Elementi anatomici Cervus elaphus.
Da questa tabella possiamo vedere che lo scheletro assile è particolarmente
sottorappresentato, infatti oltre a un frammento di palco, sono state trovate soltanto due
emimandibole, per le quali solo per una è stato possibile riconoscere il lato. Lo scheletro
appendicolare è rappresentato soprattutto dalle falangi, per un totale di 10, dai metapodi e
da altre ossa lunghe come un femore, una tibia e un’ulna. Per quanto riguarda i denti, ne
sono stati trovati 9, 8 permanenti e solo uno deciduo. Grazie a tre denti è stato possibile
identificare l’età di morte attraverso il metodo Brown & Chapman (1991). Il primo è un

83
quarto premolare deciduo inferiore destro, il quale è stato inserito nello stadio c (il primo e
il secondo molare sono già erotti, il terzo molare è ancora una gemma, sta per iniziare la
mineralizzazione del quarto premolare permanente), con un’età di circa 13 mesi. Gli altri
due denti, un quarto premolare superiore e un primo molare superiore, sono stati inseriti
nello stadio g (molariformi completamente formati), il quale corrisponde ad un età di circa
50 mesi. Il calcolo dell’età di morte è stato effettuato anche sulla base degli elementi ossei,
basandosi sullo stato di fusione delle epifisi, per cui è stato possibile determinare soltanto
due classi d’età, giovane e adulto (Fig. 6.11)
Fig. 6.11 Calcolo dell’età di morte di Cervus elaphus di Vallone Inferno.
Dal grafico di figura 6.11 possiamo vedere come gli individui adulti siano molto più
rappresentati, con 22 elementi anatomici riconosciuti, mentre la classe d’età giovane è
rappresentata da 5 elementi anatomici. Da questi dati, anche se poco numerosi, possiamo
ipotizzare la pratica di caccia selettiva, mirata principalmente agli individui adulti, in modo
da assicurare una maggiore quantità di carne. L’attività di caccia, da parte del gruppo
umano del riparo, rappresenta tuttavia una pratica economica secondaria. Infine il NMI è
stato calcolato in base al NME, sono stati riconosciuti soltanto 2 individui, uno appartenete
alla classe giovane e l’altro alla classe adulto.
6.8 Carnivori
I carnivori sono gli animali meno rappresentati di tutto il campione faunistico analizzato,
sono stati trovati infatti soltanto 16 reperti, 14 appartenenti a canidi, mentre due
appartenenti a Felis sp. Di quest’ultimo è stato trovato un frammento di un mascellare
destro, con presente il canino e il forame infraorbitario e un’ulna destra appartenente ad un
individuo adulto. Per quanto riguarda i canidi sono stati ritrovati 10 elementi ossei, quelli
più rappresentati sono le falangi, tre prime e una terza, e delle vertebre caudali. Per quanto
0
5
10
15
20
25
Giovane Adulto

84
riguarda gli elementi dentari ne sono stati trovati quattro, due canini, un premolare
superiore e un incisivo inferiore. A causa del cattivo stato di conservazione non è stato
possibile distinguere la forma selvatica dalla forma domestica.

85
Capitolo 7 “Analisi tafonomiche di Vallone Inferno”
7.1 Introduzione
Nel seguente capitolo verranno esposti i risultati ottenuti dalle analisi tafonomiche dal sito
di Vallone Inferno.
7.2 Tracce di origine naturale
Nel sito di VI sono stati trovati 607 resti ossei che presentano tracce di origine naturale ed
equivalgono al 48% del totale (Tab. 7.1). Il tipo di alterazione maggiormente riscontrata è
quella relativa all’ossido di manganese, rappresentata con il 62%, seguono le tracce da
radici con il 31%, mentre percentuali ancora più basse le troviamo sia per le rosicature di
carnivori (14%), sia per le alterazioni lasciate dai roditori e calpestio, entrambi
rappresentati con il 3%. Per quanto riguarda il weathering è stato riscontrato solo su due
frammenti (Fig 7.1).
Fig. 7.1 Tracce di origine naturale Vallone Inferno.
0
50
100
150
200
250
3.4.A 3.4.B
Radici Manganese Morsi di carnivori Morsi di roditori Calpestio Weathering

86
Vallone Inferno 3.4.A 3.4.B TOT %
NRDt 204 203 407 32%
NRDa 202 188 390 31%
Osso lungo, piatto e
irregolare
166 117 283 22%
Indet 122 59 181 14%
Tot 694 567 1261 100%
Ossa senza tracce
naturali
392 262 654 52%
Ossa con tracce
naturali
302 305 607 48%
Tot 694 567 1261 100%
Tab. 7.1 NR tracce di origina naturale.
7.2.1 Tracce di radici
Nel riparo sotto roccia di Vallone Inferno sono stati trovati 187 reperti ossei (31%), sui
quali sono state riconosciute tracce da radici. Il 52% sono state trovate nello strato 3.4.A,
mentre il restante 48% nello strato 3.4.B. Come esposto nei capitoli precedenti, le tracce da
radici sono state catalogate a seconda della loro disposizione sulla superficie dell’osso
(Gen: generalizzato, Conc: concentrato, Isol: isolato, Disp: disperso) (Tab. 7.2).
Tracce da radici VI 3.4.A 3.4.B Tot %
Gen. 34 27 61 33%
Conc. 37 37 74 40%
Isol. 4 6 10 5%
Disp. 22 20 42 22%
TOT 97 90 187 100%
Tab. 7.2 Tracce da radici di Vallone Inferno.
Dalla tabella 7.1 si evince che le tracce lasciate dalle radici sono molto abbondanti
all’interno del campione faunistico in esame, alterando profondamente la superficie
dell’osso. Le percentuali più alte sono quelle relative alle tracce che si dispongono in
maniera concentrata (attorno ad un unico punto) e generalizzata (su tutta la superficie),
rispettivamente con il 40% e il 33%. Percentuali più basse le riscontriamo nelle categorie
dispersa (22%) e isolata (5%). Le alterazioni che sono state riscontrate sono disposte in
modo orizzontale, cioè tendono a espandersi orizzontalmente lungo la superficie, con un
andamento ramificato. Solo in due casi sono state trovate disposte in modo verticale, che
porta invece alla perforazione della superficie. Questa alterazione potrebbe essere uno dei
fattori principali che ha portato all’obliterazione delle tracce di origine antropica.

87
7.2.2 Tracce di ossido di manganese
I resti osteologici che presentano questo genere di alterazione sono in totale 382 (62%),
tanto da costituire la traccia di origine naturale più abbondante; il 47% sono state trovate
nello strato 3.4.A, mentre il 53% nello strato 3.4.B. Anche le tracce da manganese sono
state catalogate a seconda della loro disposizione sulla superficie dell’osso (Gen., Conc.,
Isol., Disp.) (Tab. 7.3).
Tracce
manganese
VI
3.4.A 3.4.B TOT %
Gen. 77 77 154 40%
Conc. 6 4 10 3%
Isol. 2 4 6 2%
Disp. 96 116 212 55%
TOT 181 201 382 100%
Tab. 7.3 Tracce da ossido di manganese VI.
L’alta percentuale delle ossa che presentano questo tipo di tracce è ben spiegato dal fatto
che il sito di VI è un riparo sotto roccia, infatti, questo tipo di contesto, insieme a quello di
grotta, è il luogo ideale per la formazione delle alterazioni da ossido di manganese. Dalla
tabella 8.3 si evince che il 40% dei reperti scheletrici presenta una disposizione di tipo
generalizzata e il 55% una disposizione di tipo dispersa. Da questo è evidente che la
maggior parte delle ossa appaiono interamente coperte da manganese, anche se con
gradazioni differenti, restituendo il caratteristico colore scuro. Meno rappresentate sono le
categorie concentrata e isolata, rispettivamente 3% e 2%.
7.2.3 Tracce di rosicature di carnivori e roditori
Le rosicature di carnivori sono state riconosciute in 85 reperti ossei, pari al 12%; sono stati
individuati solchi, depressioni, impronte e alcuni casi di pitting (Fig. 7.2).
Fig. 7.2 Tracce di rosicature di carnivori a VI.
0%
10%
20%
30%
40%
3.4.A 3.4.B
Solchi Depressioni Impronta Pitting

88
La tipologia di alterazione maggiormente riscontrata è la depressione, originatasi attraverso
la pressione dei molari sulla superficie corticale dell’osso; nello strato 3.4.A la troviamo
con il 33%, mentre nello strato 3.4.B è pari al 24%. Seguono i solchi (strie prodotte dallo
sfregamento dei canini e dei molari) che al contrario delle depressioni sono maggiormente
rappresentati nello strato 3.4.B, con una percentuale pari al 15%, mentre nel 3.4.A è
rappresentata al 12%. Percentuali nettamente inferiori sono state riscontrate per le impronte
(tracce lasciate da più cuspidi dello stesso dente) pari all’1%, mentre il pitting lo troviamo
più rappresentato nello strato 3.4.B con il 4%. Inoltre sono state trovate alcune ossa che
presentano più tipologie di tracce contemporaneamente (Tab. 7.3).
Sub-livelli 3.4.A 3.4.B TOT %
Solchi +
depressioni
3 7 10 77%
Depressioni
+ impronte
1 1 2 15%
Solchi +
impronte
1 1 8%
TOT 4 9 13 100%
Tab. 7.3 Tracce di rosicature di carnivori, più tipi di traccia sullo stesso osso, VI.
La maggior parte (77%) sono quelle che presentano sia solchi che depressioni, disposti sia
nella parte corticale che nel bordo della frattura.
Infine, per quanto riguarda le tracce di rosicchiatura di denti di roditori, sono state
riscontrate su 21 reperti ossei (3%), questi appaiono con i caratteristici solchi paralleli
determinati dalla morfologia degli incisivi; 9 sono stati trovati nello strato 3.4.A e 12 nello
strato 3.4.B.
7.2.4 Calpestio
A VI sono state trovate 22 ossa che presentano tracce da calpestio, costituendo il 3%. Sono
state catalogate anch’esse a seconda della loro disposizione sulla superficie dell’osso
(Gen., Conc., Isol., Disp.) (Tab. 8.5).

89
Sub-
livelli
3.4.A 3.4.B TOT %
Gen. 3 / 3 14%
Conc. 5 7 12 52%
Isol. 2 / 2 10%
Disp. 2 3 5 24%
TOT 11 10 22 100%
Tab. 7.4 Tracce di calpestio, VI.
Dalla tabella 8.5 possiamo vedere che la categoria più rappresentata è quella relativa al
calpestio concentrato con il 52%, presente principalmente nello strato 3.4.B, segue la
disposizione dispersa con il 24%, presente in egual misura in entrambi gli strati, mentre
sono state riscontrate percentuali più basse nelle categorie generalizzato (14%) e isolato
(10%).
7.2.5 Tracce di weathering
Per catalogare le alterazioni da weathering, le quali si formano grazie alle variazioni di
temperatura e umidità, sono stati utilizzati i sei stadi indicati da Behrensmeyer (1978). A
VI sono state trovate solamente due ossa con questo tipo di alterazione, in particolare una
vertebra di taglia grande e un osso piatto di taglia media, entrambe inserite nello Stadio 1
(sono state riscontrate delle lievi fratture parallele all’asse longitudinale dell’osso). Questo
dato è compatibile con il contesto relativamente protetto di VI, probabilmente meno
soggetto a cambiamenti repentini di temperatura.
7.3 Tracce di origine antropica
Nel campione faunistico di Vallone Inferno sono stati trovati 355 reperti ossei che
presentano tracce di origine antropica, costituiscono il 28% del totale (Tab. 7.5). Tra queste
il 43% sono state trovate in uno stato di combustione, il 27% sono state classificate come
bollite, il 16% presentano fratture di origine antropica (coni di percussione, punti
d’impatto, peeling), infine le tracce di macellazione sono rappresentate soltanto dal 14%
(Fig. 7.3).

90
Fig. 7.3 Numero delle tracce antropiche rilevate, suddivise per tipologia e unità stratigrafica, (VI).
Vallone Inferno 3.4.A 3.4.B TOT %
NRDt 204 203 407 32%
NRDa 202 188 390 31%
Osso lungo, piatto e
irregolare
166 117 283 22%
Indet 122 59 181 14%
Tot 694 567 1261 100%
Ossa senza tracce
antropiche
475 431 906 72%
Ossa con tracce
antropiche
219 136 355 28%
Tot 694 567 1261 100%
Tab. 7.5 NR tracce di origine antropica
7.3.1 Tracce di combustione
Nel riparo sotto roccia di Vallone Inferno (VI) sono stati trovati 152 reperti scheletrici che
presentano tracce di combustione (43%); sono stati trovati diversi gradi di colorazione, a
seconda della temperatura e del tempo di esposizione dell’osso alla fonte di calore (Grado
1: marrone disperso; Grado 2: marrone uniforme; Grado 3: nero uniforme; Grado 4: grigio
uniforme; Grado 5: calcinato, bianco uniforme) (Tab. 7.5).
Tracce da
combustione
VI
3.4.A 3.4.B TOT %
Grado 1 8 3 11 7%
Grado 2 41 9 50 33%
Grado 3 55 6 61 40%
Grado 4 5 1 6 4%
Grado 5 23 1 24 16%
TOT 131 20 152 100%
Tab. 7.5 Tracce da combustione VI.
0%
5%
10%
15%
20%
25%
30%
35%
40%
3.4.A 3.4.B
Ossa combuste Ossa bollite Fratture antropiche Tracce di macellazione

91
Dalla tabella 7.5 possiamo notare che i gradi più frequenti sono il Grado 2 e il Grado 3
(rispettivamente 33% e 40%). Anche le ossa calcinate, Grado 5, sono ben rappresentate
con una percentuale pari al 16%, mostrando il caratteristico colore bianco. Gli altri gradi di
combustione sono invece rappresentati da percentuali più basse (Grado 1: 7%, Grado 4:
4%). Dalla tabella si evince anche che la maggior parte delle ossa combuste sono state
trovate nello strato 3.4.A, con una percentuale pari all’87%, mentre il restante 13% lo
troviamo nello strato 3.4.B. Tra le ossa appena descritte ne sono state trovate 34 (23%),
che presentano più gradazioni di colore nello stesso reperto (Tab. 7.6).
Tracce da
combustione
VI
3.4.A 3.4.B TOT %
Gradi 1-2 1 / 1 3%
Gradi 2-3 26 3 29 85%
Gradi 3-4 1 / 1 3%
Gradi 4-5 1 / 1 3%
Gradi 2-3-4 1 1 3%
Gradi 3-4-5 1 / 1 3%
TOT 30 4 34 100%
Tab. 7.6 Tracce da combustione VI, più gradazioni nello stesso reperto.
Dalla tabella 9.2 si nota che l’85% è rappresentato dai Gradi 2-3, restituendo quindi una
doppia colorazione, marrone e nera. Solamente in due casi è stata riconosciuta una tripla
colorazione, in un caso marrone, nera e grigia, nell’altro nera, marrone e bianca,
probabilmente dovuto a esposizioni differenti alla fonte di calore, delle varie parti
dell’osso.
7.3.2 Tracce di bollitura
Le ossa interpretate come bollite sono in totale 97 (27%), in questi casi la superficie
dell’osso si presenta liscia, compatta, con un aspetto vetroso, ed il colore diventa giallastro,
inoltre quando viene esposto ad una fonte di luce diretta, i bordi diventano traslucidi
(Botella et al. 2000). Nello strato 3.4.A sono stati trovati 20 reperti soggetti a bollitura, il
che corrisponde al 21% mentre i restanti 77 (79%) sono stati trovati nello strato 3.4.B.

92
7.3.3 Fratture antropiche
Le ossa che presentano fratture di origine antropica, sono in totale 56, ossia il 16% rispetto
al NR che presenta tracce di origine antropica. Sono stati riconosciute fratture per
percussione diretta e per flessione, si annoverano infatti coni di percussione, punti
d’impatto e peeling (Tab. 7.7).
Sub-livelli 3.4.A 3.4.B TOT %
Con. Perc. 11 2 13 23%
Impatto perc. 15 15 30 54%
Peeling 8 5 13 23%
TOT 34 22 56 100%
Tab. 7.7 Fratture antropiche VI.
Il tipo di frattura più rappresentata è l’impatto dovuto a percussione, con una percentuale
pari al 54%, mentre coni di percussione e peeling sono rappresentati dal 23%. Questi tipi di
fratture venivano effettuate per l’asportazione del midollo e per la preparazione della
materia prima per il confezionamento di strumenti. La frattura per flessione è stata
riscontrata esclusivamente sulle coste.
7.3.4 Tracce di macellazione
Le ossa che presentano tracce di macellazione hanno la percentuale più bassa, infatti ne
sono state trovate in totale 47, il che corrisponde al 14%. Tra queste il 68% è stato trovato
nel sub-livello 3.4.A, mentre il restante 32% nel sub-livello 3.4.B. Dal punto di vista
morfologico queste sembrano essere determinate da strumento litico in quanto troviamo un
solco che presenta schematicamente una sezione a V e le sottili striature secondarie
all’interno dello stesso. Dal punto di vista tipologico sono stati trovati incisioni, raschiature
e tagli (Fig. 7.4).

93
Fig. 7.4 Tracce di macellazione VI.
La tipologia di traccia più rappresentata è l’incisione, che è stata riscontrata con una
percentuale pari al 62% nello strato 3.4.A e il 22% nello strato 3.4.B. Con percentuali
nettamente più basse troviamo i tagli rappresentati con il 4% nello strato 3.4.A e con il 6%
nello strato 3.4.B; infine le raschiature si presentano con il 2% in entrambi i sub-livelli
(Tab. 7.8–7.9).
Taxon Ovicaprini Bos
taurus
Cervus
elaphus
Taglia
grande
Taglia
media
Taglia
piccola
TOT %
Cranio / / / / / 1 1 3%
Mandibola 1 / 1 / / 1 3 12%
Costa / / / / 2 12 13 41%
Omero 2 / / / / / 2 6%
Radio 1 / / / / / 1 3%
Bacino / / / / / 2 2 6%
Tibia 2 / / / / / 2 6%
Calcagno / / 1 / / / 1 3%
Metapode / 1 / / / / 1 3%
Osso lungo / / / / / 4 4 12%
Osso piatto / / / / 1 1 2 6%
TOT 7 1 2 / 3 21 32 100%
Tab. 7.8 NR che conservano tracce di macellazione suddivisi per elemento anatomico e taxon nel sub-
complesso 3.4.A dal sito di Vallone Inferno.
0%
20%
40%
60%
80%
3.4.A 3.4.B
Incisioni Raschiature Tagli

94
Taxon Ovicaprini Ovis
aries
Bos
taurus
Cervus
elaphus
Taglia
grande
Taglia
piccola
TOT %
Cranio 1 / / / / / 1 6%
Costa / / 1 / / 6 7 44%
Femore 1 / / / / / 1 6%
Tibia 1 / / / / / 1 6%
Falange / / / 1 / / 1 6%
Astragalo 1 2 / / / / 3 19%
Osso lungo / / / / 1 1 3%
TOT 4 2 1 1 1 7 15 100%
Tab. 7.9 NR che conservano tracce di macellazione suddivisi per elemento anatomico e taxon nel sub-
complesso 3.4.B dal sito di Vallone Inferno.
Il 40% delle tracce di macellazione sono state trovate sulle coste; occorre notare che la
maggior parte appartenevano ad individui di taglia piccola; si tratta principalmente di
incisioni, individuate sia sulla parte ventrale che sulla parte dorsale, mirate ad una
probabile eviscerazione. Di particolare interesse sono le tracce rinvenute sugli astragali, di
cui tre provenienti dal sub-livello 3.4.B, due dei quali appartengono a Ovis aries; su di essi
sono stati trovati soprattutto incisioni con un orientamento trasversale e parallelo tra loro,
finalizzate alla disarticolazione degli arti posteriori. Le incisioni sono state trovate anche su
ossa lunghe di ovicaprini, in particolare su due omeri, tre tibie e un femore,
prevalentemente situate nelle zone delle epifisi e metafisi; esse sono finalizzate
probabilmente all’azione di scarnificazione e rimozione delle masse muscolari. Per quanto
riguarda i tagli, si riporta il caso particolare di una falange di Cervus elaphus, sulla quale è
stato trovato un taglio che presenta un orientamento trasversale, situato nella parte
anteriore (craniale), finalizzato alla disarticolazione oppure allo spellamento. Le
raschiature, costituiscono la tipologia di traccia meno rappresentata, sono state individuate
con chiarezza in unico osso, una costa di un animale di piccola taglia, caratterizzato da
numerose strie, parallele tra di loro ma con un andamento a tratti discontinuo, determinato
probabilmente da uno strumento litico ritoccato.

95
Capitolo 8
“Il campione faunistico di Case Bastione”
8.1 Introduzione
Nel seguente capitolo vengono illustrate le analisi archeozoologiche effettuate sui reperti di
grandi mammiferi, recuperati durante la campagna di scavo 2013 del villaggio di Case
Bastione. La fauna analizzata in questo lavoro proviene dalle UU.SS. 1057, 2001, 2002,
2003, 2079, 2083, 2087 e 2089. Questi strati hanno una cronologia relativa risalente al
Bronzo antico, facies Castelluccio, mentre per la cronologia assoluta, è stata ottenuta da
due datazioni al radiocarbonio provenienti dallo strato 1057, la prima, grazie a un
campione di carbone che ha restituito una datazione di 2205-1951 cal. BC, la seconda, da
un campione faunistico il quale ha restituito una datazione di 2135-1921 cal. BC.
8.2 La fauna analizzata
Il campione faunistico analizzato a Case Bastione è costituito da un NR pari a 983, di
questi, 230 sono stati riconosciuti totalmente (NRDt), corrisponde al 23% del totale, 146
sono state riconosciute a livello anatomico (NRDa), pari al 15%; anche in questo caso sono
state riconosciute le classi osso lungo, piatto e articolare, a queste appartengono 370
reperti, pari al 38% del totale. Infine 237 ossa, sono state catalogate come indeterminate,
corrispondono al 24% del totale (Fig. 8.1) (Tab. 8.1).
Fig. 8.1 NRDt, NRDa, categorie osso lungo, piatto e articolare, Indet, del campione faunistico di Case
Bastione
0%5%
10%15%20%25%30%35%40%
NRDt NRDa Osso lungo,piatto e
articolare
Indet

96
Case Bastione Tot %
NR 983 100%
NRDt 230 23%
NRDa 146 15%
Osso lungo, piatto e articolare 370 38%
Indet 237 24%
Tot 983 100%
Tab. 8.1 NR Case Bastione
Dal grafico di figura 8.1 possiamo vedere come le categorie più rappresentate siano quelle
relative alle classi osso lungo, piatto e articolare e quelle considerate indeterminate. Questo
è dovuto ad un alto grado di frammentazione, dovuto probabilmente sia a processi
tafonomici, sia a fratture moderne, la questione verrà approfondita nei capitoli successivi.
Di seguito viene riportato il grafico con le percentuali relative alle categorie osso lungo,
piatto e articolare (Fig. 8.2).
Fig. 8.2 Categorie osso lungo, osso piatto e osso articolare di Case Bastione.
Dal grafico di figura 8.2 si evince come la categoria più rappresentata sia quella osso lungo
con quasi l’80%, si tratta soprattutto di elementi diafisari, dei quali è stata trovata soltanto
una faccia corticale che non presentava nessun elemento diagnostico. Molto più distaccate
sono le ossa piatte, rappresentate per il 24%, mentre una percentuale bassissima (2%), è
stata riconosciuta per la categoria osso articolare.
Per quanto riguarda gli elementi riconosciuti a livello anatomico (NRDa), di seguito viene
riportata la tabella che li riassume (Tab 8.2).
0%
10%
20%
30%
40%
50%
60%
70%
80%
Osso lungo Osso piatto Osso articolare

97
UU.SS. 1057 2001 2002 2003 2079 2083 2087 2089 NRDa %
ELEMENTI
ANATOMICI
Cranio 9 1 / / 4 8 2 / 24 16%
Atlante 1 / / / / / / / 1 1%
Emimandibola 3 / / / / 1 / / 4 3%
Vertebra 11 4 / / 6 2 / 5 28 19%
Coste 28 8 / / 9 4 2 / 51 35%
Scapola 1 2 / / 2 / 1 / 6 4%
Omeri 4 1 / / 5 1 1 1 13 9%
Radio 2 1 / / 1 1 / / 5 3%
Ulna 1 / / / / / / / 1 1%
Metapode 1 1 / 1 1 / / / 4 3%
Falange 1 / / / / / / / 1 1%
Bacino / / / / / 1 / / 1 1%
Femore 1 2 / / / / / / 3 2%
Tibia 1 1 / / 1 / / / 3 2%
Molare/premolare 1 / / / / / / / 1 1%
TOT 65 21 / 1 29 18 6 6 146 100%
Tab. 8.2 NRDa del campione faunistico di Case Bastione
Dalla tabella 8.2 possiamo notare che gli elementi più abbondanti sono le coste con il 35%
e le vertebre con il 19%, queste sono più rappresentate nell’US 1057, che in generale è lo
strato che ha restituito la maggior parte del materiale. Tra le vertebre è stato possibile
riconoscere una prima vertebra cervicale, l’atlante. Anche il cranio è ben documentato,
infatti corrisponde al 16%, mentre ad una percentuale più bassa troviamo le emimandibole
(3%). In questa tabella si evince anche come lo scheletro appendicolare risulti essere meno
rappresentato. Una situazione simile è stata riscontrata anche per Vallone Inferno, questo è
dovuto ad un alto grado di frammentazione che ha eliminato le parti diagnostiche. Tra
quelli più rappresentati abbiamo l’omero (9%), mentre gli altri elementi scheletrici
presentano delle percentuali comprese tra l’1 e il 4%.
Per le categorie osso lungo, piatto, articolare e per le ossa determinate anatomicamente,
sono state riconosciute anche le taglie; in particolare sono state trovate quattro categorie di
taglia: taglia grande, taglia media, taglia piccola e taglia molto piccola (Fig. 8.3).

98
Fig. 8.3 Categoria di taglia di Case Bastione.
Dal grafico di figura 8.3 si evince che le ossa di taglia piccola siano quelle più
rappresentate con il 62%; come già sostenuto nei capitoli precedenti a questa categoria di
taglia appartengono animali domestici come ovicaprini e suini di età giovanile, animali
selvatici come cervidi di età giovanile. Seguono poi gli animali di taglia media con una
percentuale pari al 23%, seguono con valori inferiori al 10% le categorie taglia grande e
taglia molto piccola.
Nella seguente tabella (Tab. 8.3) vengono riportate le specie a grandi mammiferi
riconosciute nelle UU.SS. oggetto di analisi di questo lavoro. Come per Vallone Inferno, di
ogni specie viene riportato il NRDt, e la relativa percentuale di abbondanza. Le percentuali
vengo poi riportate nel grafico di figura 8.4, per fornire con maggior chiarezza il dato sulle
specie rappresentate.
UU.SS. 1057 2001 2002 2003 2079 2083 2087 2089 NRDt %
Taxa
Cervus elaphus 14 / / / 4 / / / 18 8%
Canis familiaris / / 1 / 13 2 3 / 19 8%
Sus domesticus 13 1 / / 3 2 / / 19 8%
Bos taurus 4 2 / / 5 7 1 / 19 8%
Ovis aries 1 / / / 1 / / 2 2%
Ovicaprini 65 7 / 3 34 15 18 7 149 64%
Artiodattili 2 1 / / / / 1 / 4 2%
TOT 98 12 1 3 59 27 23 7 230 100%
Tab. 8.3 Specie riconosciute di Case Bastione.
0%
10%
20%
30%
40%
50%
60%
70%
Tagliagrande
Tagliamedia
Tagliapiccola
Taglia moltopiccola

99
Fig. 8.4 Percentuali di abbondanza mammiferi Case Bastione.
Dalla tabella 8.3 e dal grafico di figura 8.4 si evince con chiarezza che gli ovicaprini sono
gli animali più rappresentati, anche in questo caso costituiscono la base economica del sito.
Grazie a due reperti, in particolare quarti premolari decidui, è stato possibile effettuare il
riconoscimento a livello specifico, infatti è stata identificata la pecora (Ovis aries). Per
quanto riguarda gli altri animali domestici, presentano tutti la stessa abbondanza, pari
all’8%, indicando un allevamento di tipo misto che comprende in particolare il maiale ed il
bue, anche se con percentuali nettamente inferiori rispetto agli ovicaprini. Per quanto
riguarda gli animali selvatici, è stata riconosciuta un'unica specie, il cervo rosso, questo
presenta una percentuale pari all’8%, il che confrontata con quella degli altri animali
domestici, testimonia un’attività di caccia costante, ma pur sempre secondaria. Per quanto
riguarda i carnivori, sono stati ritrovati 19 reperti, corrispondente all’8%, attribuiti a canidi,
i quali hanno consentito la distinzione tra la forma domestica e quella selvatica, infatti è
stato riconosciuto il cane (Canis familiaris).
Anche in questo caso i taxa riconosciuti, verranno presentati seguendo un ordine di
abbondanza, prima verranno trattate le forme domestiche, in seguito le forme selvatiche.
8.3 Ovicaprini
Anche per Case Bastione gli ovicaprini sono gli animali più rappresentati, costituendo la
base economica del villaggio. Presentano infatti, una percentuale d’abbondanza pari al
64% del totale. Di seguito viene riportata la tabella con gli elementi ossei riconosciuti nel
campione faunistico analizzato (Tab. 8.4).
0%
10%
20%
30%
40%
50%
60%
70%

100
Tab. 8.4 Elementi anatomici degli ovicaprini di Case Bastione.
Dalla tabella 8.4 si evince che lo scheletro appendicolare è rappresentato soprattutto dalle
emimandibole, ne sono state trovate 18, le quali costituiscono il 17% del totale, alcune di
queste sono state trovate con i denti inclusi negli alveoli, molto utili per il calcolo dell’età
di morte. È stato trovato anche un frammento di mascellare destro che presenta ancora il
terzo molare superiore. Il resto degli elementi ossei rinvenuti appartengono soprattutto alle
ossa lunghe, quello più rappresentato è l’omero (22%), poi abbiamo il radio (14%), segue
la tibia con circa l’11%. Di questi sono state trovate moltissime epifisi e metafisi che hanno
consentito non solo il riconoscimento a livello specifico, ma anche la lateralità, infatti sono
stati molto utili per il conteggio del NME. Ben rappresentate sono le falangi (9%), mentre
le altre ossa lunghe sono rappresentate da percentuali più basse tra l’1 e il 4 %. Sono state
trovate anche alcune ossa del carpo e del tarso, di particolare importanza risultano essere
gli astragali, in quanto è stato possibile, anche se in un solo caso, effettuare il calcolo
dell’altezza al garrese, tramite i coefficienti di Teichert (1975). L’altezza stimata è 58,9
cm, perfettamente in media con quelle calcolate a Vallone Inferno.
Di seguito viene riportato il grafico con l’indice di frammentazione scheletrica, che è stato
calcolato mettendo in rapporto il NME con il NRDt, il valore vicino a 1 indica l’elemento
UU.SS. 105
7
200
1
200
2
200
3
207
9
208
3
208
7
208
9
NRDt % NM
E
%
ELEMENTI
ANATOMICI
OVICAPRINI
Mascella 1 / / / / / / / 1 1% / /
Emimandibola 9 / / / 5 / 3 1 18 17% 5 11%
Scapola 2 / / / / / 1 1 4 4% 2 4%
Omero 13 2 / / 4 3 / 1 23 22% 12 26%
Radio 6 1 / / 2 3 2 1 15 14% 9 19%
Ulna 2 1 / / 1 / / / 4 4% 4 9%
Bacino 1 / / / 1 1 / / 3 3% / /
Femore 1 / / / / / / / 1 1% / /
Tibia 3 2 / / 4 1 1 1 12 11% 10 21%
Patella 1 / / / / / / / 1 1% / /
Metacarpo / / / / / 2 / / 2 2% 1 2%
Metatarso 2 / / 1 1 / / / 4 4% 2 4%
Metapode 1 / / / 1 / / 1 3 3% / /
Falange 3 1 / 1 1 / 4 / 10 9% / /
Astragalo 1 / / / 1 / / / 2 2% 2 4%
Semilunare 1 / / / / / / / 1 1% / /
Capitato-
trapezoide
2 / / / / / / / 2 2% / /
TOT 49 7 / 2 21 10 11 6 106 100
%
47 100%

101
poco frammentato, l’indice vicino a 0 indica l’elemento molto frammentato; è stato
riportato anche l’indice di sopravvivenza scheletrica, mettendo in rapporto il NME con il
NEE, basato sul NMI, il valore vicino a 1 indica lo scheletro completo, il valore vicino a 0
indica lo scheletro incompleto (Fig. 8.5).
Fig. 8.5 Indice di frammentazione degli elementi scheletrici e indice di sopravvivenza scheletrica degli
ovicaprini, del sito Case Bastione.
Per quanto riguarda l’indice di frammentazione possiamo vedere che ossa come omero,
radio, metacarpo e metatarso, sono piuttosto frammentate, presentando un valore pari a 0,5.
Risultano essere meno frammentate l’ulna, la tibia e l’astragalo, presentando valori vicino
a 1. Il valore più basso è dato invece dalle emimandibole con un valore vicino a 0,3,
essendo queste ossa piatte sono più soggette a frammentazione. Per quanto riguarda
l’indice di frequenza scheletrica, le ossa più presenti all’interno del campione faunistico,
sono l’omero con un valore pari a 0,6, seguito da radio e tibia con un valore pari a 0,5. Tra
le altre ossa lunghe, sono poco presenti l’ulna e i metapodi. Lo scheletro assile è
rappresentato unicamente dalle emimandibole, anche se presentano un valore molto basso,
pari a 0,27. Per quanto riguarda il calcolo dell’età di morte, anche per Case Bastione, dalle
ossa lunghe sono state riconosciute tre classi d’età, neonato, giovane e adulto, a seconda
dello stato di fusione delle epifisi e del grado di porosità della parte corticale (Fig. 8.6).
Fig. 8.6 Calcolo dell’età di morte da resti ossei di ovicaprini di Case Bastione
00,20,40,60,8
11,2
NME/NRDt NME/NEE
0
10
20
30
40
Neonato Giovane Adulto

102
Dalla figura 8.6 è evidente che la classe degli individui adulti è quella più rappresentata
con quasi 40 reperti, da questo dato si può ipotizzare che l’allevamento era finalizzato
soprattutto alla produzione di carne e lana.
Di seguito viene riportata la tabella che riassume i resti dentari rinvenuti; fra questi è stato
possibile analizzarne 35, in quanto si presentano in un buono stato di conservazione, per
cinque denti invece non è stato possibile effettuare nessun tipo di analisi, questi non sono
stati inseriti nella tabella (Tab. 8.5).
UU.SS. 1057 2001 2002 2003 2079 2083 2087 2089 NRDt % NME %
ELEMENTI
DENTARI
OVICAPRINI
dP2 dx 1 1 2% 1 3%
dP3 sx 1 1 2% 1 3%
I inf 1 1 2%
P 1 1 2%
M/P 2 1 1 1 5 12%
P2 dx 1 1 2 5% 2 6%
P4 sx 1 1 2% 1 3%
P4 sx 1 1 2% 1 3%
P4 dx 1 1 1 3 7% 3 9%
M1 dx 1 1 2% 1 3%
M1 sx 1 1 2% 1 3%
M1 dx 1 1 2 5% 2 6%
M1 sx 2 1 3 7% 3 9%
M1/2 1 1 2%
M2 dx 1 1 2% 1 3%
M2 sx 1 1 1 3 7% 3 9%
M2 sx 2 2 4 10% 4 13%
M3 dx 1 1 2 5% 2 6%
M3 dx 1 1 2 5% 2 6%
M3 sx 1 1 1 1 4 10% 4 13%
M 1 1 2 5%
TOT 15 / / 1 13 5 7 1 42 100% 32 100%
Tab. 8.5 Resti dentari degli ovicaprini di Case Bastione.
Dalla tabella 8.5 possiamo vedere che la maggior parte sono denti permanenti, attribuiti
quindi ad animali adulti, di questi sono presenti i secondi e quarti premolari, mancano i
terzi, mentre i molari, sia superiori che inferiori, sono tutti rappresentati. Quelli più
numerosi sono i terzi molari superiori sinistri e i secondi molari superiori sinistri, con il
10%. I denti decidui sono rappresentati soltanto da due reperti, entrambi ritrovati nell’US
2087. Alcuni denti, che costituiscono il 12%, sono stati inseriti nella categoria
molare/premolare (M/P), a causa dell’alto stato di frammentazione.
Di seguito viene riportato il grafico (Fig. 8.7) con gli stadi di usura della Wilkens (2003); è
possibile notare come gli stadi più rappresentati sono quelli relativi ad animali adulti, dal II
al VII stadio, quindi con più di 24 mesi d’età; tra questi quelli più riscontrati sono lo stadio

103
IV e lo stadio V, dove abbiamo la perdita degli infundibuli del primo molare, mentre la
corona del terzo molare presenta una forma tendente al quadrato.
Fig. 8.7 Stadi di usura Wilkens (2003)
degli ovicaprini di Case Bastione.
Anche per Case Bastione, sono stati uniti i dati ricavati dal calcolo dell’età di morte, sia dai
resti ossei che da quelli dentari (Fig. 8.8).
Fig. 8.8 Calcolo dell’età di morte degli ovicaprini di Case Bastione
Dal grafico di figura 8.8 è possibile notare come la classe d’età più rappresentata sia quella
adulta, con 70 reperti riconosciuti. Grazie a questo dato è possibile ipotizzare una pratica di
allevamento mirata principalmente ad ottenere prodotti derivati come latte e lana. È
testimoniata anche la produzione di carne, soprattutto dalle classi neonato e giovane.
Per quanto riguarda il NMI, è stato calcolato in base al NME prendendo in considerazione
la classe d’età e la lateralità. Anche in questo conteggio la classe adulta risulta quella più
rappresentata, infatti sono stati calcolati 5 individui, in base alle ossa lunghe, in particolare
grazie a cinque radio destri e cinque omeri sinistri. Tutte le altre classi sono rappresentate
da un solo individuo. In totale quindi, nel campione faunistico di Case Bastione analizzato
in questo lavoro, il NMI degli ovicaprini è pari a 9.
Stadio 4
Stadio 5
Stadio II
Stadio III
Stadio IV
Stadio V
Stadio VI
Stadio VII
0
10
20
30
40
50
60
70
Neonato Infatile Giovane Sub-adulto Adulto

104
8.4 Sus domesticus Erxleben, 1777
Il maiale è tra le forme domestiche che sono state riconosciute all’interno del campione
faunistico di Case Bastione, con una percentuale pari all’8% e un NRDt pari a 19, il NMI è
uguale a 4. Di seguito viene riportata la tabella con gli elementi scheletrici riconosciuti
(Tab. 8.6).
UU.SS 1057 2001 2002 2003 2079 2083 2087 2089 NRDt % NME %
ELEMENTI
ANATOMICI
Sus
domesticus
Mascella 2 / / / / / / / 2 11% 2 29%
Mandibola / 1 / / 1 1 / / 3 16% 2 29%
Omero 1 / / / / / / / 1 5% 1 14%
II Metatarso / / / / / 1 / / 1 5% / /
Astragalo 1 / / / / / / / 1 5% / /
Falange 5 / / / 2 / / / 7 37% / /
I inf 1 / / / / / / / 1 5% / /
dI2 dx 1 / / / / / / / 1 5% 1 14%
C sup 1 / / / / / / / 1 5% / /
P3 sx 1 / / / / / / / 1 5% 1 14%
TOT 13 1 / / 3 2 / / 19 100% 7 100%
Tab. 8.6 Elementi anatomici relativi ai suini di Case Bastione.
Dalla tabella 8.6 possiamo vedere che gli elementi scheletrici relativi ai suini sono stati
trovati soltanto nelle UU.SS 1057, 2001, 2079 e 2083, nelle altre risultano totalmente
assenti. L’US che ha restituito più reperti è la 1057. In generale tra gli elementi ossei
spiccano le falangi, infatti ne sono state trovate 7, rappresentando così il 37%, fra queste
quelle più numerose sono le seconde falangi. Tra lo scheletro assile abbiamo due
frammenti di mascellare, il primo è un mascellare destro con P2 e P3 ancora infissi negli
alveoli, mentre il secondo è un mascellare sinistro con P4 ancora presente; questi due
frammenti appartengono ad individui diversi, in quanto hanno età differenti. Per quanto
riguarda le mandibole, soltanto una presenta ancora dei denti, in particolare M2 e M3, per
questa è stato possibile riconoscere lo stadio di usura I della Wilkens (2003), il terzo
molare si presenta infatti senza usura e con radici ancora aperte.
Per quanto riguarda il calcolo dell’età di morte è stato effettuato sia dagli elementi ossei,
prendendo in considerazione lo stato di fusione delle epifisi e la porosità della parte
corticale, sia attraverso i resti dentari tramite le tabelle di Habermehl (1961), Barone
(1980) e Silver (1969) (Fig. 8.9).

105
Fig. 8.9 Calcolo dell’età di morte dei suini di Case Bastione.
Dalla figura 8.9 possiamo vedere che sono state riconosciute quattro classi di età, neonato,
giovane, sub-adulto e adulto, manca la classe infantile. Quelle più rappresentate sono
giovane e adulto con 5 reperti per classe. Anche se il numero dei reperti è esiguo, possiamo
ipotizzare una pratica di allevamento mirata prevalentemente alla produzione di carne.
Per quanto riguarda il NMI, è stato calcolato in base al NME, prendendo in considerazione
lateralità e classi d’età. Gli individui calcolati sono in totale 4, uno per ogni classe di età.
8.5 Bos taurus Linnaeus, 1758
Tra le forme domestiche riconosciute a Case Bastione c’è anche il bue, questo è
rappresentato da 19 reperti i quali costituiscono l’8% del totale, il NMI è pari a 3. Di
seguito viene riportata la tabella che riassume gli elementi anatomici riconosciuti (Tab.
8.7).
UU.SS 1057 2001 2002 2003 2079 2083 2087 2089 NRDt % NME %
ELEMENTI
ANATOMICI
Bos taurus
Mandibola / / / / / 5 / / 5 26% 3 50%
Omero / / / / 1 / / / 1 5% / /
Femore 2 / / / / / / / 2 11% / /
Tibia / / / / / 1 / / 1 5% 1 17%
Calcagno / / / / / 1 / / 1 5% 1 17%
Metacarpo / / / / 1 / / / 1 5% 1 17%
Metapode / / / / 1 / / / 1 5% / /
Capitato-
trapezoide / / / / 1 / / / 1 5% / /
I Falange / / / / / / 1 / 1 5% / /
II Falange 1 1 / / / / / / 2 11% / /
M/P 1 / / / / / / 1 5% / /
M / 1 / / 1 / / / 2 11% / /
TOT 4 2 / / 5 7 1 / 19 100% 6 100%
Tab. 8.7 Elementi anatomici relativi ai bovini di Case Bastione.
0
1
2
3
4
5
Neonato Giovane Sub-adulto Adulto

106
Dalla tabella 8.7 si evince che sono stati riconosciuti pochi elementi anatomici, infatti lo
scheletro assile è rappresentato solo dalle mandibole, di queste sono stati trovati cinque
frammenti, costituiscono così il 26% del totale, soprattutto processi coronoidi e condilei.
Lo scheletro appendicolare è rappresentato da poche elementi anatomici: tra le ossa lunghe
degli arti anteriori troviamo soltanto un omero e un metacarpo, mentre tra le ossa lunghe
degli arti posteriori troviamo due femori e una tibia. Gli elementi dentari sono rappresentati
soltanto da tre reperti, ma dato l’alto stato di frammentazione non è stato possibile
riconoscere la lateralità e se si trattava di denti superiori o inferiori. Per quanto riguarda
l’età di morte è stato possibile riconoscere soltanto la classe adulta, questo può fare
ipotizzare, nonostante il numero esiguo dei reperti rinvenuti, che questi animali venivano
sfruttati principalmente per la produzione di prodotti secondari come il latte e la pelle,
secondariamente per la produzione di carne. Il NMI è stato possibile calcolarlo grazie alle
mandibole, in tutto ne sono state trovate cinque, tre sinistre, una destra, mentre per la
quinta non è stato possibile riconoscere il lato, quindi il NMI è pari a 3.
8.6 Canis familiaris Linnaeus, 1758
Tra i carnivori è stato riconosciuto il cane domestico, Canis familiaris, il quale è presente
con 19 reperti, costituendo anch’esso l’8% del totale. Di seguito viene riportata la tabella
che riassume gli elementi ossei rinvenuti (Tab. 8.8).
UU.SS 1057 2001 2002 2003 2079 2083 2089 2087 NRDt % NME %
ELEMENTI
ANATOMICI
Canis
familiaris
Cranio / / / / / 1 / / 1 5% 1 11%
Mandibola / / / / 5 / / 1 6 32% 4 44%
Calcagno / / / / 1 / / / 1 5% / /
P / / / / 1 / / / 1 5% / /
I1 / / / / 1 / / / 1 5% / /
I / / / / 3 / / / 3 16% / /
C inf sx / / / / 1 / / / 1 5% 1 11%
C inf dx / / / / 1 / / / 1 5% 1 11%
C sup dx / / / / / 1 / / 1 5% 1 11%
C / / 1 / / / / 1 2 11% / /
P2 dx / / / / / / 1 1 5% 1 11%
TOT / / 1 / 13 2 / 3 19 100% 9 100%
Tab. 8.8 Elementi anatomici di Canis familiaris di Case Bastione.

107
Dalla tabella 8.8 possiamo vedere che l’US che ha restituito il maggior numero di materiali
è la 2079, infatti sono stati trovati cinque frammenti di mandibola, quattro di questi con
molta probabilità appartengono allo stesso individuo, e diversi elementi dentari. Il pezzo
più sorprendente è tuttavia il cranio che è stato trovato nell’US 2083, in quanto presenta
l’osso incisivo, il nasale, mascellare sinistro, con quasi tutti i denti infissi negli alveoli,
troviamo infatti due incisivi, tre premolari e i due molari, anche parte dell’osso frontale,
zigomatico e lacrimale. Per il resto sono stati trovati soprattutto resti dentari, tra questi
quelli più numerosi risultano essere i canini. Per quanto riguarda il calcolo dell’età di
morte, è stata riconosciuta esclusivamente la classe adulta. Il NMI è stato calcolato in base
alle mandibole, per un totale di 2.
8.7 Cervus elaphus Linnaeus, 1758
Il cervo rosso è l’unica specie selvatica riconosciuta all’interno del campione faunistico di
Case Bastione. Anche in questo caso è rappresentato con l’8%, sono stati trovati infatti 18
reperti. Di seguito viene riportata la tabella con gli elementi anatomici riconosciuti (Tab.
8.9).
UU.SS. 1057 2079 NRDt % NME %
ELEMENTI
ANATOMICI
Cervus elaphus
Palco 5 2 7 39% / /
Radio 1 / 1 6% / /
Metapode 1 / 1 6% / /
Femore 1 / 1 6% / /
Tibia 2 2 4 22% 4 67%
Grande cuneiforme 1 / 1 6% / /
Piramidale 1 / 1 6% / /
dP4 dx 2 / 2 11% 2 33%
TOT 14 4 18 100% 6 100%
Tab. 8.9 Elementi anatomici relativi al Cervus elaphus di Case Bastione.
Dalla tabella 8.9 possiamo vedere che palco e tibia sono gli elementi più rappresentati, il
primo con il 39% e il secondo con il 22%. Per quanto riguarda i palchi, sono stati trovati
diversi frammenti, lunghi mediamente 2 cm, con uno stato di combustione pari a livello 3,
cioè interamente di colore nero. Questi ritrovamenti possono testimoniare la lavorazione
del palco, finalizzato alla creazione di strumenti, all’interno del villaggio. Per quanto
riguarda le tibie e le altre ossa lunghe ritrovate, si sono conservate principalmente metafisi
ed epifisi. Sono stati trovati anche due resti dentari, in particolare due quarti premolari

108
decidui destri inferiori, grazie ai quali è stato possibile calcolare l’età di morte attraverso il
metodo Brown & Chapman (1991). In uno è stato individuato lo stadio c (il primo e il
secondo molare già erotti, mentre il terzo è ancora una gemma), presenta un’età di 13 mesi.
Per il secondo è stato possibile stabilire lo stadio e (le corone del terzo molare e dei
premolari sono formate del tutto, mentre le radici del quarto premolare deciduo sono
completamente riassorbite), il quale indica un’età pari a 18 mesi. Anche gli altri elementi
scheletrici hanno fornito delle informazioni utili al calcolo dell’età di morte, infatti
troviamo un’epifisi prossimale di un femore, non saldata, e l’epifisi distale di una tibia non
ancora del tutto saldata alla metafisi. Tutte le altre ossa appartengono ad individui adulti.
Grazie agli elementi dentari e ossei è stato possibile quindi determinare tre classi d’età:
infantile, giovane e adulto (Fig. 8.10)
Fig. 8.10 Calcolo dell’età di morte di Cervus elaphus di Case Bastione
Anche per Case Bastione la classe d’età più rappresentata è quella adulta, nonostante il
numero esiguo dei reperti rinvenuti, possiamo comunque ipotizzare la pratica di caccia
selettiva. Per quanto riguarda il NMI è pari a 4, calcolato in base all’età e alla lateralità,
due individui appartengono alle classi infantile e giovane, mentre i restanti due alla classe
adulta.
0
1
2
3
4
5
6
Infantile Giovane Adulto

109
Capitolo 9 “Analisi tafonomiche di Case Bastione”
9.1 Introduzione
In questo capitolo verranno esposti i risultati delle analisi tafonomiche, sia di origine
naturale che antropiche dal sito di Case Bastione (CBS).
9.2 Tracce di origine naturale
Nel sito di CBS sono stati rinvenuti 401 reperti osteologici nei quali sono state riscontrate
tracce di origine naturale, queste equivalgono al 41% del totale (Tab 9.1). L’alterazione più
diffusa è quella da radici, infatti presenta una percentuale pari al 75%, segue quella relativa
alle rosicature di carnivori con il 16%. Percentuali ancora più basse sono state riscontrate
per le tracce da ossido di manganese, weathering, calpestio e rosicchiature ad opera di
roditori, rispettivamente 10%, 8%, 3% e 1% (Fig. 9.1).
Fig. 9.1 Tracce di origine naturale di CBS.
0
20
40
60
80
100
120
140
160
US 1057 US 2001 US 2002 US 2003 US 2079 US 2083 US2087 US 2089
Radici Manganese Rosicature di carnivori Rosicature di roditori Calpestio Weathering

110
Case Bastione Tot %
NR 983 100%
NRDt 230 23%
NRDa 146 15%
Osso lungo, piatto e articolare 370 38%
Indet 237 24%
Tot 983 100%
Ossa senza tracce naturali 582 59%
Ossa con tracce naturali 401 41%
Tot 983 100%
Tab. 9.1 NR tracce di origine naturale Case Bastione.
9.2.1 Tracce di radici
Nel villaggio di Case Bastione sono stati trovati 302 reperti ossei che presentano tracce di
radici. Si tratta dell’alterazione più abbondante, infatti corrisponde al 66%. Le tracce di
radici sono state catalogate a seconda della loro disposizione sulla superficie dell’osso
(Gen., Conc., Isol., Disp (Tab. 9.2).
Tracce da
radici CBS
1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Gen. 76 10 3 / 26 13 12 6 146 48%
Conc. 34 6 / / 15 10 5 4 74 25%
Isol. 1 / / / / / / / 1 1%
Disp. 30 6 / 1 25 7 7 5 81 27%
TOT 141 22 3 1 66 30 24 15 302 100%
Tab. 9.2 Tracce di radici Case Bastione.
Dalla tabella 9.2 possiamo vedere come la categoria generalizzata sulla superficie
dell’osso, sia quella predominante con il 48%, in alcuni casi è stata riscontrata
un’alterazione uniforme, molto profonda che coinvolge tutta la superficie dell’osso, questo
fattore può aver portato alla totale obliterazione di eventuali tracce di origine antropica.
Seguono le categorie dispersa e concentrata, rispettivamente con il 27% e il 25%, mentre in
un solo caso è stata riscontrata quella isolata. Solo in due casi l’alterazione presenta un
orientamento verticale, che porta quindi alla perforazione della superficie, mentre tutto il
resto dei reperti presentano un orientamento orizzontale, producendo dei solchi larghi, con
la sezione a U e con un andamento ramificato.

111
9.2.2 Tracce da ossido di manganese
Le tracce di ossido di manganese sono state trovate solamente in 42 campioni,
corrispondente al 9%. Anche in questo caso le tracce sono stata catalogate a seconda della
loro disposizione sulla superficie dell’osso (Gen., Conc., Isol., Disp) (Tab. 9.3).
Tracce da
manganese
CBS
1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Gen. 2 1 / / / / / / 3 7%
Conc. 4 / / / 2 / / / 6 14%
Isol. / 2 / / 3 / 1 / 6 14%
Disp 17 / / 2 7 / / 1 27 64%
TOT 23 3 / 2 12 / 1 1 42 100%
Tab. 9.3 Tracce da ossido di manganese da CBS.
La percentuale bassissima relativa a questo tipo di alterazione è probabilmente spiegato dal
fatto che il sito di CBS è un villaggio e quindi un contesto all’ aperto, il quale non
favorisce la formazione di questo genere di alterazioni. Dalla tabella 8.6 si evince che la
maggior parte delle ossa (64%) presentano una disposizione di manganese dispersa, si
trovano quindi in piccole unità distanziate tra loro.
9.2.3 Tracce di rosicature di carnivori e roditori
In totale sono stati trovati 64 reperti ossei che presentano tracce di rosicature di carnivori,
questi equivalgono al 14%, in particolare sono state riconosciuti solchi, depressioni,
impronte e pitting (Fig. 9.2).
Fig. 9.2 Tracce di rosicature di carnivori da CBS
0%
2%
4%
6%
8%
10%
12%
14%
16%
18%
20%
US 1057 US 2001 US 2002 US 2003 US 2079 US 2083 US 2087 US 2089
Solchi Depressioni Impronte Pitting

112
Da questo grafico possiamo vedere che non in tutte le UU.SS. sono state trovate questo
tipo di alterazioni, infatti sono totalmente assenti nelle UU.SS. 2002 e 2003. Il tipo di
tracce più rappresentato sono le depressioni, queste le troviamo maggiormente nello strato
1057, con una percentuale pari al 20% e nello strato 2079 (9%). Percentuali comprese tra il
3% e il 2% le troviamo nelle UU.SS. 2001, 2087 e 2089. Queste presentano una
disposizione varia, sono state trovate infatti sia sulla parte corticale dell’osso sia sul bordo
della frattura. Molto rappresentati sono anche i solchi, come per le depressioni li troviamo
principalmente nelle UU.SS. 1057 (11%) e 2079 (6%). Nelle altre UU.SS. (2001, 2087 e
2089) sono meno rappresentati, infatti sono state riscontrate percentuali pari al 2%. Infine
le impronte sono state trovate esclusivamente nell’US 1057, anche in questo caso con una
percentuale molto bassa, pari al 5%. Oltre alle ossa illustrate nel grafico di figura 8.4, ne
sono state trovate alcune (33%) che presentano più tipi di tracce contemporaneamente, in
particolare 16 (25%) che presentano sia solchi che depressioni, principalmente provenienti
dall’US 1057, in 4 casi (6%) sono state trovate sia depressioni che impronte, queste
provengono dall’US 2079, infine un solo reperto dove sono state trovate sia solchi che
impronte, rinvenuto nell’US 1057 (Tab. 9.4).
Tracce di
morso di
carnivori
1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Solchi +
depressioni
11 1 / / / 2 2 / 16 25%
Solchi +
impronte
1 / / / / / / / 1 1%
Depressioni
+ impronte
/ / / / 4 / / / 4 6%
TOT 12 1 / / 4 2 2 / 21 33%
Tab. 9.4 Tracce di rosicature di carnivori, più tipi di traccia sullo stesso osso, CBS.
Un altro tipo di alterazione è il pitting, determinato da piccole e abbondanti depressioni
presenti prevalentemente lungo le estremità dell’osso; come per le altre alterazioni è più
presente nell’US 1057 con l’11% e nell’US 2079 con il 5%, mentre nelle altre è
rappresentato da percentuali ancora più basse tra il 3% e il 2%.
Infine, per quanto riguarda le tracce di rosicchiature di roditori, ne sono state trovate
soltanto 6, pari all’1% del totale. Quattro sono state rinvenute nell’US 1057, mentre le altre
due sono state rinvenute nelle UU.SS. 2079 e 2087.

113
9.2.4 Calpestio
L’alterazione da calpestio è stata riscontrata in pochissimi casi, infatti è stata rinvenuta
soltanto in 12 reperti ossei, pari al 3%. Sono state catalogate a seconda della loro
disposizione sulla superficie dell’osso (Gen., Conc., Isol., Disp) (Tab. 9.5).
Tracce
da
calpestio
1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Gen / / / / 1 1 / / 2 17%
Conc 2 / / / 2 / / 1 5 42%
Isol / / / / 2 / / / 2 17%
Disp 1 1 / / 1 / / / 3 25%
TOT 3 1 / / 6 1 / 1 12 100%
Tab. 9.5 Tracce da calpestio di CBS.
Questo tipo di alterazione è stato rinvenuto principalmente nell’US 2079, con 6 reperti,
segue l’US 1057 con 3. La modalità di disposizione maggiormente riscontrata è quella
concentrata (Conc: numerose ma sempre nello stesso punto) con il 42%, segue quella
dispersa (Disp: più di una stria ma distanziate tra di loro) con il 25%, mentre le altre
presentano la stessa percentuale pari al 17%.
9.2.5 Tracce di weathering
I reperti ossei che hanno restituito questo tipo di alterazione sono in totale 33, il che
equivale al 3% del totale. Sono state catalogate secondo gli stadi di Behrensmeyer (1978).
Di seguito viene riportata la tabella che li riassume (Tab. 9.6).
Tracce da
weathering
CBS
1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Stadio 0 / / / / / / / / / /
Stadio 1 8 1 / / 7 4 3 / 23 74%
Stadio 2 2 / / / 4 1 2 1 10 26%
Stadio 3 / / / / / / / / / /
Stadio 4 / / / / / / / / / /
Stadio 5 / / / / / / / / / /
TOT 10 1 / / 11 5 5 1 33 100%
Tab. 9.6 Tracce da weathering di CBS, stadi di Behrensmeyer (1978).

114
Dalla tabella 9.6 possiamo notare che sono stati riscontrati soltanto due stadi di
Behrensmeyer (1978), lo Stadio 1 (74%) e lo Stadio 2 (26%), i quali presentano sia leggere
fessure lungo l’asse longitudinale dell’osso che esfoliazione nella parte esterna della
superficie. Le ossa che presentano questi tipi di tracce sono state riscontrate principalmente
nell’US 1057 e nell’US 2079, rispettivamente 10 e 11 reperti. CBS essendo un sito aperto,
potrebbe essere maggiormente soggetto ai cambiamenti di temperatura e quindi a questo
tipo di alterazione, rispetto a VI.
9.3 Tracce di origine antropica
Nel sito di Case Bastione sono stati trovati 235 reperti ossei che presentano tracce di
origine antropica, costituiscono il 24% del totale (Tab. 9.7). La maggior parte di questi
reperti sono stati rinvenuti in uno stato di combustione, con una percentuale pari al 78%;
sono state rinvenute anche ossa che hanno subito un trattamento di bollitura, questi
equivalgono all’11%, la stessa percentuale è data dalle ossa che presentano fratture
antropiche (peeling, coni di percussione e punti d’impatto) (11%), infine le ossa che hanno
restituito tracce da macellazione come incisioni, tagli e raschiature sono solamente 12 il
che equivale al 5%. (Fig. 9.3).
Fig. 9.3 Numero delle tracce antropiche rilevate, suddivise per tipologia e unità stratigrafica, dal sito di
Case Bastione.
0
10
20
30
40
50
60
70
80
90
US 1057 US 2001 US 2002 US 2003 US 2079 US 2083 US 2087 US 2089
Cobustione Bollitura Fratture antropiche Tracce di macellazione

115
Case Bastione Tot %
NR 983 100%
NRDt 230 23%
NRDa 146 15%
Osso lungo, piatto e articolare 370 38%
Indet 237 24%
Tot 983 100%
Ossa senza tracce antropiche 748 76%
Ossa con tracce antropiche 235 24%
Tot 983 100%
Tab. 9.7 NR tracce di origine antropica
9.3.1 Tracce di combustione
Nel villaggio di CBS sono state trovate 183 ossa con tracce di combustione, corrispondente
al 78%. Anche in questo caso sono state classificate a seconda della loro gradazione di
colore (Grado 1: marrone disperso; Grado 2: marrone uniforme; Grado 3: nero uniforme;
Grado 4: grigio uniforme; Grado 5: calcinato, bianco uniforme) (Tab. 9.7).
Tracce da
combustione
CBS
1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Grado 1 12 / / / 1 / / / 13 7%
Grado 2 26 8 / / 20 6 2 3 65 36%
Grado 3 39 10 1 / 30 7 2 2 91 50%
Grado 4 5 2 / / 5 / / / 12 7%
Grado 5 / 2 / / / / / / 2 1%
TOT 82 22 1 / 56 13 4 5 183 100%
Tab. 9.7 Tracce di combustione dal sito di Case Bastione.
Dalla tabella 9.6 possiamo vedere che tutte le UU.SS. analizzate hanno restituito ossa
combuste a varie gradazioni, tranne l’US 2003. La colorazione più frequente (50%) è
quella che corrisponde al Grado 3, quindi un colore nero uniforme, trovata principalmente
nelle UU.SS. 1057 e 2079. Le ossa che presentano un colore marrone uniforme (Grado 2),
sono in totale 65, corrispondenti quindi al 36%, per le altre gradazioni troviamo percentuali
più basse comprese tra l’1% e il 7%. Anche a CBS sono state trovate ossa che presentano
duplici o triplici colorazioni nello stesso reperto, probabilmente dovuto anche in questo
caso a differenti modalità di esposizione alla fonte di calore (Tab. 9.8).

116
Tracce da
combustione
CBS
1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Grado 1-2 / / / / 1 / / / 1 2%
Grado 2-3 15 4 / / 15 6 1 2 43 78%
Grado 3-4 3 / / / 4 / / / 7 13%
Grado 4-5 / / / / / / / / / /
Grado 1-2-3 3 / / / / / / / 3 5%
Grado 2-3-4 / 1 / / / / / / 1 2%
TOT 21 5 / / 20 6 1 2 55 100%
Tab. 9.8 Tracce di combustione dal sito di Case Bastione, con più gradazioni nello stesso reperto.
Dalla tabella 9.8 possiamo notare che tra i reperti ossei che presentano una duplice
colorazione, le più numerose sono quelle che hanno i colori marrone e nero, corrispondente
quindi al Grado 2-3 (78%), le troviamo soprattutto nelle UU.SS. 1057 e 2079, mentre sono
totalmente assenti nell’UU. SS. 2002 e 2003. Segue il Grado 3-4 con il 13%, determinato
da una colorazione nera e grigia. In pochi casi sono state trovate colorazioni triple, il 5%
corrispondente al il Grado 1-2-3 e un solo reperto per il grado 2-3-4. Bisogna tuttavia
sottolineare che le ossa combuste potrebbero non essere esclusivamente dovute a cause
antropiche legate all’alimentazione o allo smaltimento dei rifiuti, infatti dall’analisi
stratigrafica del sito è emerso che questo villaggio ha subito un incendio che ha portato
all’abbandono. Le varie gradazioni di colore si potrebbero spiegare quindi con la distanza
più o meno variabile dal nucleo principale dell’incendio. Questa rimane tuttavia un
interpretazione del tutto ipotetica.
9.3.2 Tracce di bollitura
Nel sito di Case Bastione sono state trovate pochissime ossa interpretate come bollite, in
totale sono 27 il che corrisponde all’ 11%. Queste ossa sono state rinvenute principalmente
nella parte centrale della Capanna 1, nelle UU.SS. 1057, 2079, 2083, 2087 e 2089,
sembrano essere invece totalmente assenti nelle UU. SS. 2001, 2002 e 2003.
9.3.3 Fratture antropiche
Per quanto riguarda le fratture di origine antropica, queste sono state trovate su 21 reperti
ossei ossia il 9%. Il numero è abbastanza esiguo, meno della metà rispetto a VI. Anche in
questo caso sono state riconosciute fratture per percussione diretta e per flessione (coni di
percussione, punti d’impatto e peeling) (Tab. 9.9).

117
UU.SS 1057 2001 2002 2003 2079 2083 2087 2089 TOT %
Con. perc. 2 / / / 3 2 1 / 8 38%
Impatto
perc.
2 1 / / 1 4 / / 8 38%
Peeling 2 1 / / / 2 / / 5 24%
TOT 6 2 / / 4 8 1 / 21 100%
Tab. 9.9 Fratture antropiche dal sito di Case Bastione.
Dalla tabella possiamo vedere che coni di percussione e punti d’impatto sono rappresentati
in egual misura, entrambi con il 38%, mentre la fratturazione per flessione è stata
riscontrata solamente in 5 coste (24%). Sono state rinvenute principalmente nelle UU.SS
2083, 2079 e 1057.
9.3.4 Tracce di macellazione
Nel campione faunistico di CBS sono state trovate pochissime ossa con tracce di
macellazione, in totale sono 12, le quali corrispondono al 5%. Dal punto di vista
morfologico queste sembrano essere determinate da strumento litico. Dal punto di vista
tipologico, anche in questo caso sono stati riconosciuti incisioni, tagli e raschiature (Tab.
9.10).
Taxon Ovicaprino Cervus
elaphus
Taglia
piccola
Indet. TOT %
Costa / / 1 / 1 8%
Omero 3 / / / 3 25%
Radio 1 / / / 1 8%
Ulna 1 / / / 1 8%
Metatarso / 1 / / 1 8%
Pisiforme / / 1 / 1 8%
Osso lungo / / 2 / 2 17%
Osso piatto / / 1 / 1 8%
Indet. / / / 1 1 8%
TOT 5 1 5 1 12 100%
Tab. 9.10 NR che conservano tracce di macellazione suddivisi per elemento anatomico e taxon (CBS).
Sono state trovate principalmente su ossa di ovicaprini (3 omeri, 1 radio e 1 ulna). Su due
omeri sono state riconosciute tracce di raschiature, entrambe all’altezza della metafisi
distale, in uno sono stati riconosciuti nella parte anteriore (craniale), nell’altro nella parte
laterale, mirati probabilmente all’asportazione del periostio o al recupero della massa
muscolare. Nell’ultimo omero, del quale si è conservata soltanto la metafisi distale, sono
state trovate 6 strie, localizzate nella parte posteriore (caudale), con un orientamento
trasversale. Alcune di queste appaiono molto sottili, determinate da un movimento ripetuto

118
dello strumento, mentre altre appaiono più larghe e parallele tra di loro. Per quanto
riguarda il radio, si sono conservate soltanto l’epifisi e la metafisi prossimale, nella parte
mediale sono state riconosciuti tre tagli con un orientamento trasversale, mirati
probabilmente a tranciare l’osso o sezionare le fibre muscolari. Particolare è il caso di un
pisiforme, dove sono state trovate tre incisioni molto evidenti, anche ad occhio nudo,
larghe, profonde e parallele tra di loro, finalizzate alla disarticolazione o spellamento.

119
Capitolo 10 “Analisi archeozoologiche, Vallone Inferno e Case
Bastione a confronto”
10.1 Introduzione
Nel seguente capitolo verranno confrontati i dati ottenuti dalle analisi archeozoologiche
effettuate su entrambi i campioni faunistici, discussi nei capitoli 6 e 8. Il fine è quello di
evidenziare le analogie e le differenze dei due siti siciliani, cronologicamente uguali ma
tipologicamente diversi. Ricordiamo che Vallone Inferno (VI) è un riparo sotto roccia,
mentre Case Bastione (CBS) è un villaggio. Le analisi archeozoologiche sono state
effettuate per i due campioni faunistici tramite le metodologie esposte nel capitolo 3.
10.2 I campioni faunistici a confronto
La fauna analizzata a VI presenta un NR pari a 1261, mentre a CBS il NR è pari 983. Per
entrambi i siti il NR è stato suddiviso in NRDt (numero dei resti determinati totalmente),
NRDa (numero dei resti determinati anatomicamente), Indet (numero dei resti
indeterminati), inoltre sono stati conteggiati anche le ossa inserite nelle categorie osso
lungo, piatto e articolare (Fig. 10.1).
Fig. 10.1 NRDt, NRDa, categorie osso lungo, piatto e articolare, Indet, di VI e CBS.
In figura 10.1 possiamo vedere che le percentuali dei NRDt e NRDa sono più alte per VI
(rispettivamente il 32% e il 31%), mentre per CBS troviamo percentuali comprese tra il
20% e il 15%. La situazione è totalmente opposta per quanto riguarda le categorie osso
lungo, piatto e articolare e le ossa considerate indeterminate, infatti a VI abbiamo per le
prime il 22%, per le seconde il 14%; mentre a CBS il 38%, sono state inserite nelle
0%
5%
10%
15%
20%
25%
30%
35%
40%
NRDt NRDa Osso lungo,piano,
articolare
Indet
VI CBS

120
categorie osso lungo, piatto e articolare, il 24% sono state considerate indeterminate.
Questo probabilmente è dovuto al fatto che a CBS sono state trovate più porzioni
diafisarie, le quali non sempre presentano degli elementi diagnostici utili al loro
riconoscimento. Bisogna aggiungere anche che molte ossa di CBS presentano delle fratture
moderne, rendendo ancora più arduo il riconoscimento a livello specifico. Inoltre a VI sono
stati trovati molti resti dentari (13%) dai quali nella maggior parte dei casi, è stato possibile
effettuare il riconoscimento a livello tassonomico.
Per quanto riguarda il NRDa di seguito viene riportato il grafico che ne riassume le
percentuali riconosciute per ciascun elemento anatomico (Fig. 10.2).
Fig. 10.2 NRDa di VI e CBS.
Da questo grafico si evince che le percentuali degli elementi anatomici riconosciuti sono
molto simili per ambedue i siti. Troviamo ben rappresentato lo scheletro assile, infatti il
cranio presenta percentuali comprese tra il 15% e il 30%, mentre le vertebre presentano
percentuali simili (21% per VI e 19% per CBS). Tuttavia, l’elemento più rappresentato
sono le coste, troviamo il 37% per VI e il 35% per CBS. Lo scheletro appendicolare è
sottorappresentato, infatti per entrambi i siti le percentuali sono comprese tra il 9% e l’1%.
Questa discrepanza tra i due distretti scheletrici si deve probabilmente al fatto che lo
scheletro assile essendo costituito da ossa piatte, data la loro particolare morfologia
tendono ad andare incontro più facilmente alla frammentazione, con il rischio di essere
sovra-rappresentate. Inoltre questa discrepanza può essere dovuta al fatto che le coste sono
ossa sovrannumerarie rispetto alle ossa degli arti.
Sono state confrontate anche le categorie osso lungo, piatto e articolare, i risultati vengono
mostrati nel grafico successivo (Fig. 10.3).
0%5%
10%15%20%25%30%35%40%
VI CBS

121
Fig. 10.3 Categorie osso lungo, piatto e articolare dei siti di VI e CBS.
Da questo grafico possiamo vedere che i dati relativi a queste categorie sono molto simili
in entrambi i siti. I frammenti più rappresentati appartengono a ossa lunghe infatti troviamo
il 69% per VI e il 74% per CBS, sono costituiti principalmente da frammenti dove è
riconoscibile il canale midollare e nella maggior parte dei casi troviamo una singola faccia
corticale; le percentuali dei frammenti di osso piatto sono più basse, infatti abbiamo il 31%
per VI e il 24% per CBS; infine per quanto riguarda le ossa articolari, troviamo percentuali
bassissime per entrambi i siti (VI 1%, CBS 2%).
Dai frammenti ossei inseriti nei NRDa e nelle categorie osso lungo, piatto e articolare, è
stato possibile riconoscerne la taglia (Fig. 10.4).
Fig. 10.4 Categorie di taglia dei siti di VI e CBS.
0%
10%
20%
30%
40%
50%
60%
70%
80%
Osso lungo Osso piano Osso articolare
VI CBS
0%
10%
20%
30%
40%
50%
60%
70%
Taglia grande Taglia media Taglia piccola Taglia moltopiccola
VI CBS

122
Dal grafico di figura 10.4 è possibile notare che la categoria di taglia più rappresentata è la
taglia piccola, la quale comprende maiali e cervi di età giovanile, ovicaprini e carnivori
come il cane e il lupo. Anche in questo caso i due siti presentano percentuali molto
similari, 64% per VI e 62% CBS. La categoria di taglia media risulta essere più
rappresentata a CBS con il 23%, mentre VI presenta una percentuale più bassa pari al 16%,
questa categoria comprende maiali e cervi di età adulta e bovini di età giovanile.
Rappresentate in modo minoritario sono le altre due categorie di taglia grande e molto
piccola, con percentuali comprese tra il 5% e il 10% circa.
10.2.1 Taxa riconosciuti e strategie economiche a confronto
Per quanto riguarda i taxa riconosciuti, di seguito viene riportato il grafico e la tabella che
ne riassumono le percentuali di abbondanza (Fig. 10.5; Tab. 10.1)
Fig. 10.5 Specie riconosciute nei campioni faunistici di VI e CBS.
0%
10%
20%
30%
40%
50%
60%
70%
VI CBS

123
NRDt
%
NMI NME
%
Neonato
%
Infantile
%
Giovane % Sub-adulto % Adulto
%
Taxa VI CBS V I CBS V I CBS VI CBS VI CBS VI CBS VI CBS VI CBS
Ovicaprini 68 % 64% 13 9 44% 53% 19% 3% 2% 1% 18% 7% 7% 1% 15% 47%
Ovis aries 1% 2% / / / / / / / / / / / / / /
Capra hircus 1% / / / / / / / / / / / / / / /
Sus scrofa d. 10% 8% 8 4 56% 37% 8 % 5% 26% / 8% 26% 13% 11% 3% 26%
Sus scrofa 1% / / / / / / / / / / / / / / /
Bos taurus 6 % 8% 2 3 35% 32% / / / / 13% / / / 43% 58%
Cervus elaphus 9% 8% 2 4 33% 33% / / / 11% 14% 16% / / 61% 32%
Canide 3% / / / / / / / / / / / / / / /
Canis familiaris / 8% / 2 / 47% / / / / / / / / / 74%
Felis sp. 1% / 1 / / / / / / / / / / / / /
Artiodattili 2% 2% / / / / / / / / / / / / / /
Tab 10.1 NRDt, NMI, NME e classi d’età dei campioni faunistici di VI e CBS.

124
I due campioni faunistici esaminati sono in linea generale molto similari tra loro, sia dal
punto di vista qualitativo sia dal punto di vista quantitativo. È possibile notare che gli
animali più rappresentati sono gli ovicaprini (68% per VI, 64% per CBS), costituendo così
la base fondamentale della pratica di allevamento. Per i ambedue i campioni è stato
possibile discriminare in pochissimi casi le due specie osteologicamente affini pecora-
capra (Ovis aries-Capra hircus). Sulla base dei calcoli dell’età di morte si può ipotizzare
che la base economica del riparo di VI, era basata su un allevamento di tipo misto, mirato
cioè sia alla produzione di carne, testimoniato dalla presenza di individui giovani, sub-
adulti e neonati, sia alla produzione di latte, testimoniato dalla classe di età adulta,
quest’ultima probabilmente veniva impiegata anche per la produzione di lana. Invece per
quanto riguarda CBS, è stata riscontrata prevalentemente la classe adulta, quindi è
possibile ipotizzare una pratica di allevamento mirata principalmente ad ottenere prodotti
derivati come latte e lana. È testimoniata anche la produzione di carne, soprattutto dalle
classi neonato e giovane. Esclusivamente per gli ovicaprini è stato possibile calcolare
l’altezza al garrese, grazie al ritrovamento degli astragali. L’altezza media è compresa tra
54,4 cm e 63,5 cm. La pratica dell’allevamento sembra essere basata in forma minore
anche sui maiali, infatti sono state riscontrate percentuali piuttosto basse (10% per VI, 8%
per CBS), con molta probabilità finalizzata alla produzione di carne. Con percentuali
altrettanto basse troviamo anche il bue, che sembra più rappresentato a CBS con l’8%,
mentre VI presenta una percentuale di abbondanza pari al 6%; in entrambi i siti è stata
riscontrata maggiormente la classe adulta, possiamo supporre quindi che questi animali
venivano sfruttati principalmente per la produzione di prodotti derivati come il latte,
secondariamente per la produzione di carne. L’attività di sussistenza dei due siti doveva
essere fondata anche sulla pratica della caccia, tuttavia, l’unica specie selvatica
riconosciuta è Cervus elaphus, rappresentato anch’esso da percentuali alquanto basse (9%
per VI, 8% per CBS); in particolare a CBS, nonostante i pochi reperti rinvenuti sono state
riconosciute diverse classi d’età, questo potrebbe testimoniare la pratica di caccia selettiva.
Infine, sono stati riconosciuti anche reperti ossei e dentari appartenenti a carnivori, a VI il
3% dei reperti appartiene a canidi, un solo reperto è stato attribuito al gatto, ma per
entrambi non è stato possibile distinguere la forma domestica da quella selvatica. Per CBS,
i carnivori sono rappresentati esclusivamente da canidi, però a differenza di VI è stato
possibile riconoscere il cane domestico (Canis familiaris), rappresentato con una
percentuale di abbondanza pari all’8%.

125
Capitolo 11 “Analisi tafonomiche a confronto”
11.1 Introduzione
Nel seguente capitolo verranno discusse le differenze e le analogie che sono state
riscontrate durante lo studio tafonomico dei due campioni faunistici. Inizialmente verranno
effettuate le considerazioni sulle tracce di origine naturale, subito dopo quelle sulle tracce
di origini antropiche.
11.2 Tracce di origine naturale, confronto tra VI e CBS.
I reperti con tracce di origine naturale presentano una percentuale d’abbondanza simile nei
due campioni faunistici, infatti troviamo il 48% per Vallone Inferno e il 41% per Case
Bastione. Nel seguente grafico vengono indicate le percentuali di abbondanza per ciascuna
tipologia di traccia (Fig. 11.1).
Fig. 11.1 Percentuali di abbondanza delle tracce di origine naturale riscontrate a VI e CBS.
Dal grafico (figura 11.1) possiamo vedere che ci sono molte differenze fra i due siti. Una
delle più evidenti riguarda le tracce di radici, a CBS è stata riscontrata una percentuale più
alta (75%) probabilmente dovuta alla tipologia di sito, infatti, trattandosi di un abitato,
siamo difronte ad un contesto di sito all’aperto, il che può aver favorito la crescita della
vegetazione rispetto a VI (31%), nel quale, trattandosi di un riparo sotto roccia la crescita
0%
10%
20%
30%
40%
50%
60%
70%
80%
Vallone Inferno Case Bastione
Radici Manganese Carnivori Roditori Calpestio Weathering

126
della vegetazione è meno accentuata. Per quanto riguarda l’ossido di manganese troviamo
una situazione totalmente opposta, infatti è molto più presente a VI con il 62%, mentre
CBS presenta una percentuale nettamente inferiore pari al 10%; anche questo fattore è
determinato dalla diversa tipologia di sito, infatti un contesto come il riparo sotto roccia,
favorisce la mobilitazione dei composti solubili di manganese in modo più accentuato
rispetto a un contesto di sito all’aperto. Un’altra differenza grossolana riguarda le tracce da
weathering, le quali sono più rappresentate a CBS (8%) probabilmente più soggetto a
continui cambiamenti di temperature rispetto a VI, il quale presenta soltanto l’1%. Per
quanto riguarda le altre alterazioni sono state registrate leggere differenze, infatti le tracce
di rosicature di carnivori presentano il 14% a VI e il 16% a CBS, le rosicature di roditori
sono poco più rappresentati a VI con il 3% mentre CBS presenta solo l’1%, infine il
calpestio è stato riscontrato con la stessa percentuale (3%) in entrambi i siti.
11.3 Tracce di origine antropica, confronto tra VI e CBS.
In entrambi i siti sono state trovate diverse tipologie di tracce antropiche, in particolare
tracce da combustione, trattamento da bollitura, tracce di macellazione e fratture
antropiche. Di seguito viene riportato il grafico che indica le frequenze per ciascuna
tipologia di traccia (Fig. 11.2).
Fig. 11.2 Percentuali di abbondanza delle tracce di origine antropica riscontrate a VI e CBS.
0%
10%
20%
30%
40%
50%
60%
70%
80%
Vallone Inferno Case Bastione
Ossa combuste Ossa bollite Fratture Tracce di macellazione

127
In figura 11.2 possiamo notare che anche per le tracce di origine antropica insistono
numerose differenze. Tra queste abbiamo quelle che presentano tracce da combustione,
molto più rappresentate a CBS con il 78%, mentre a VI presentano il 43%. Tuttavia, in
entrambi i casi, si tratta del processo tafonomico più abbondante. A giudicare dai dati
desunti dall’analisi stratigrafica dei due siti, è possibile ipotizzare che la causa dello stato
di combustione dei reperti ossei è differente. VI è un riparo sotto roccia utilizzato per la
stabulazione degli animali, ove sono stati rinvenuti in corso di scavo dei livelli di stallatico
(Forgia et alii, 2012), cioè veniva effettuata periodicamente una pulizia della superficie
accumulando e bruciando rifiuti e sterco. A CBS doveva essere praticata un’azione
differente di smaltimento dei rifiuti, le ossa combuste possono essere quindi dovute alle
pratiche di cottura e all’incendio traumatico che ha portato all’abbandono del villaggio
(Giannitrapani et alii, in cds). Per quanto riguarda le ossa che sono state interpretate come
bollite, sono maggiormente testimoniate a VI con il 27%, mentre CBS presenta una
percentuale più bassa pari all’11%. La distribuzione di queste ossa sulla superficie del
riparo di VI sembra abbastanza omogenea, mentre a CBS sembrano concentrarsi nella
parte centrale della Capanna 1, in particolare nelle UU.SS. 1057, 2079, 2083, 2087 e 2089,
questo potrebbe essere in connessione con il focolare che era situato proprio al centro della
capanna. Per quanto riguarda le fratture di origine antropica, queste presentano una
percentuale simile, il 16% a VI e l’11% a CBS, in entrambi sono stati riconosciuti punti
d’impatto, coni di percussione e la frattura per flessione (peeling), è quindi testimoniata la
lavorazione dell’osso per il recupero del midollo e per il confezionamento di strumenti.
Infine, per quanto riguarda le tracce di macellazione, sono quelle meno rappresentate, il
14% a VI e il 5% a CBS. La bassa frequenza può essere dovuta ai diversi processi
tafonomici, sia di origine naturale sia di origine antropica, pre e post-deposizionali, che
possono aver portato all’obliterazione di questi tipi di tracce attraverso la frammentazione
e l’asportazione della superficie corticale dell’osso. Dal punto di vista morfologico sono
state riscontrate tracce da strumento litico, questo conferma il fatto che durante il Bronzo
antico l’utilizzo degli strumenti in metallo era ancora abbastanza limitato (Greenfield,
1999). Dal punto di vista tipologico sono stati riconosciuti in entrambi i siti incisioni,
raschiature e tagli, rinvenuti principalmente sulle coste, sulle ossa del carpo e del tarso e
sulle zone metafisarie delle ossa lunghe. Sono state quindi riscontrate pratiche di
macellazione mirate principalmente all’eviscerazione, alla disarticolazione e al distacco
delle masse muscolari. Queste tracce sono state rinvenute soprattutto su ossa di ovicaprini,
in pochissimi casi sul bue e sul cervo.

128

129
CONCLUSIONI
L’analisi presentata in questo lavoro riguarda la gestione e lo sfruttamento delle risorse
faunistiche provenienti dai siti di Vallone Inferno (PA) e Case Bastione (EN). In
particolare sono stati analizzati i reperti faunistici provenienti dagli strati che testimoniano
una frequentazione durante l’antica età del Bronzo. Nel sito di Vallone Inferno sono stati
analizzati i sub-livelli 3.4.A e 3.4.B, scavati nel 2012, 2013 e 2014, mentre a Case Bastione
sono state analizzate le ossa rinvenute, durante la campagna di scavo 2013, nell’ Area β
all’interno della Capanna 1 (UU.SS. 1057, 2001, 2002, 2003, 2079, 2083, 2087, 2089);
l’obiettivo di questo lavoro è quello di confrontare i due campioni faunistici provenienti da
due contesti cronologicamente simili ma tipologicamente e funzionalmente differenti,
infatti Vallone Inferno si può considerare un riparo sotto roccia dedito alla stabulazione
degli animali, mentre Case Bastione è un contesto di abitato.
Il lavoro di analisi dei reperti archeo-faunistici è stato eseguito in due fasi, la prima
riguarda l’analisi archeozoologica, effettuata nel Dipartimento di Scienze della Terra e del
Mare della Facoltà di Scienze Naturali dell’Università di Palermo, seguito dalla Dott.ssa
Daria Petruso, nonché correlatrice di questo lavoro, la seconda riguarda l’analisi
tafonomica, eseguita nel Laboratorio di Archeozoologia e Tafonomia del Dipartimento di
Biologia ed Evoluzione dell’Università degli Studi di Ferrara, assist ito dalla Dott.ssa
Ursula Thun Hohenstein.
Dai dati desunti dalle analisi archeozoologiche si è evinto che la composizione degli
insiemi faunistici mostra molte analogie, infatti in entrambi i casi è presente una netta
prevalenza delle faune domestiche. La pratica dell’allevamento era sicuramente basata
soprattutto sugli ovicaprini; dal calcolo dell’età di morte è possibile ipotizzare un
allevamento di tipo misto, cioè mirato sia alla produzione di prodotti primari (carne) sia a
quelli secondari (latte, pelle, lana). Esclusivamente per gli ovicaprini inoltre, sono state
calcolate le altezze al garrese grazie a quattro astragali, tre provenienti da Vallone Inferno
e uno da Case Bastione, l’altezza media è compresa tra 54,4 cm e 63,5 cm.
L’allevamento era basato secondariamente anche sui suini, di cui sono stati riconosciuti
individui neonati fino ad una classe di età di circa 2 anni, venivano utilizzati quindi per la
produzione di carne. I due campioni faunistici sono costituiti anche da pochi resti di bue, in
entrambi i casi riguardano individui adulti, utilizzati principalmente per la produzione di
prodotti secondari.

130
La pratica di sussistenza era basata anche sulla caccia, le specie selvatiche rinvenute sono
Sus scrofa e Cervus elaphus, tuttavia in entrambi i siti, doveva trattarsi di una pratica
secondaria e alquanto sporadica. Questo vale soprattutto per Vallone Inferno in quanto
sono stati trovati pochissimi resti di cervo e uno solo relativo al cinghiale. Per Case
Bastione è stata attestata una caccia di tipo selettiva al cervo, in quanto sono state
riscontrate diverse classi d’età anche se il numero dei resti è piuttosto esiguo. Infine per
quanto riguarda i carnivori, sono attestati con basse percentuali i canidi, per Case Bastione
è stato possibile riconoscere il cane (Canis familiaris) rinvenuti in entrambi i siti, e i felidi
rinvenuti solo a Vallone Inferno (sono stati trovati due reperti attribuiti al gatto, ma non è
stato possibile discriminare la forma domestica da quella selvatica).
Per quanto concerne le analisi tafonomiche abbiamo visto che le tracce di origine naturale
rispecchiano il contesto in cui insistono i due siti in esame. I dati più significativi in questo
senso sono le percentuali relative alle tracce di ossido di manganese e radici, la prima
molto più presente nel riparo sotto roccia di Vallone Inferno, si formano infatti più
facilmente in riparo e grotte, mentre la seconda è nettamente più rappresentata nel villaggio
di Case Bastione. Per quanto riguarda le tracce di origine antropica, in particolare le tracce
di macellazione, sono poco rappresentate in entrambi i siti, a Case Bastione ne sono state
ritrovate solamente 12, dovuto probabilmente sia all’alterazione dovuta all’azione delle
radici che in molti casi ha portato all’asportazione della parte corticale dell’osso, sia alla
combustione che ha portato ad un’ulteriore frammentazione. Dai pochi dati desunti dalle
tracce di macellazione di Case Bastione è possibile ipotizzare l’azione di
spellamento/disarticolazione in quanto sono state trovate strie su alcune ossa del carpo e
del tarso e sulle epifisi di ossa lunghe, in particolare omeri di ovicaprini. A Vallone Inferno
è stata testimoniata l’eviscerazione, grazie alle numerose strie rinvenute sulle coste sia
sulla parte ventrale che dorsale, la disarticolazione grazie soprattutto al rinvenimento di
incisioni sugli astragali di ovicaprino e infine la scarnificazione grazie alle incisioni sulle
metafisi e alle poche raschiature che sono state rinvenute.
Questi dati sono stati confrontati con le analisi preliminari dei reperti faunistici provenienti
dal Riparo di San Tommaso (EN) (Mannino, 2006). I primi dati sembrano confermare una
strategia di allevamento in linea con i siti oggetto di studio di questo lavoro, infatti
troviamo tra le faune domestiche maggiormente rappresentati gli ovicaprini, in forma
minore il maiale e il bue, mentre tra le specie selvatiche è attestato solo il cervo (Mannino,
2006).

131
È stato preso in considerazione anche lo scarico del villaggio di Castelluccio (SR) (Orsi,
1983) dove sono stati trovati ampi resti di rifiuti; qui oltre a numerosi resti ceramici e
strumenti in osso, sono stati trovati anche resti ossei sia di animali selvatici che domestici
infatti sono stati rinvenuti il cane, bue, maiale, pecora, capra, cavallo e cervo.
Altri campioni faunistici protostorici siciliani provengono principalmente da abitati come
quelli di Messina (ME) (Mangano, 2000) e da luoghi di culto come Monte Grande (AG)
(Bedini, 1998), dove sono state attestate prevalentemente faune domestiche.
In conclusione i dati desunti dalle analisi archeozoologiche e tafonomiche sembrano in
linea con i confronti suddetti, illustrano per tanto un allevamento rivolto principalmente
agli ovicaprini, in secondo luogo al maiale, più raramente al bue; la pratica della caccia
risulta essere decisamente sporadica a Vallone Inferno, più presente a Case Bastione, ma
rimane comunque un’attività di sussistenza secondaria.

132

133
BIBLIOGRAFIA
AGENBROAD L. D. (1989): Spiral Fractured mammoth Bone from Nonhuman
Taphonomic Processes at Hot Spings Mammoth Site. In Bonnichsen, R. and Sorg, M. H.
(ed.) Bone Modification. Orono: University of Maine Center for the Study of the First
Americans. Pp: 139-148.
ALCÁNTARA, V. et al. (2006): “Determinación de procesos de fractura sobre
huesos frescos: un sistema de análisis de los ángulos de los planos de fracturación como
discriminador de agentes bióticos”. Trabajos de Prehistoria, 63 (1): 37-45.
ANDREWS, P. & Cook, J. (1985): Natural Modifications to Bones in a Temperate
Setting. Man. 20 pp: 675 -691.
BALDINI G., CASSATARO U., LONGO E., RECANI A., 1976. Recenti scoperte
sul paleolitico siciliano. Natura 67, 125–127.
BARONE R., Anatomie compare des mammifères domestiques, Paris: Vigot Freres,
Editeurs, 1976.
BARONE R., 1980, Anatomia comparata dei mammiferi domestici, vol. I,
Osteologia, Bologna.
BEDINI E. 1998. I reperti faunistici del deposito votivo del Bronzo Antico di Monte
Grande. In G. Castellana (a cura di), Il santuario castellucciano di Monte Grande e
l’approvvigionamento dello zolfo nel Mediterraneo nell’età del Bronzo, pp. 431-454.
BEHRENSMEYER A. K. (1978): Taphonomic and Ecologic Information from
Bone Weathering. Paleobiology. 4(2) Pp:150-62.
BEHRENSMEYER A. K., GORDON K. D., & YANAGI G. T., 1986. Trampling
as a cause of bone surface damage and pseudo-cut marks. Nature 319: pp. 768-771.
BEHRENSMEYER A. K., GORDON K. D., & YANAGI G. T., 1989. Nonhuman
bone modification in Miocene fossils from Pakistan. In (R. Bonnichsen and M. H. Sorg,
eds.) Bone modification, pp. 99-120. Orono: University of Maine Center for the Study of
the First Americans.
BINFORD, L. R. (1981): Bones. Ancient Men and Modern Myths. Ac. Press. New
York.
BLASCO, R. (2006): Estrategias de subsistencia de los homínidos del nivel XII de
la Cova del Bolomor (La Valldigna, Valencia). Tesis de Licenciatura. Tarragona.
Departament d’Història i Geografia. Universitat Rovira i Virgili.
BLUMENSCHINE, R. J. & SELVAGGIO, M. M. (1988): Percussion marks on
bone surfaces as a new diagnostic of hominid behaviour. Nature. 333 (6175) Pp: 763-5.

134
BOSCATO P., FRONZA V., SALVADORI F., 2000, Un archivio informatizzato
per la gestione dei reperti archeozoologici, in Brogiolo G.P. (a cura di): 2° Congresso
Nazionale di Archeologia Medievale, Brescia, settembre 2000, All’Insegna del Giglio,
Firenze: 46-52.
BOSCH P., et al., Boiled versus unboiled: a study on Neolithic and contemporary
human bones, Journal of Archaeological Science (2011), doi:10.1016/j.jas.2011.04.019.
BOTELLA, M.C., ALEMÁN, I, y JIMÉNEZ, S.A. (2000): Los huesos humanos.
Manipulación y alteraciones. Ediciones Bellaterra: Barcelona.
BOVIO MARCONI J., 1979. La grotta del Vecchiuzzo. Roma, 108 p.
BRAIN, C. K. (1981): The Hunters or the Hunted? An Introduction to African
Cave Taphonomy. University of Chicago Press. Chicago.
BROSE, D. S. (1975). Functional analysis of stone tools: a cautionary note on the
role of animal fats. American Antiquity 48, 86–103.
BROWN W. A. B., CHAPMAN N. G., Age assessment of red deer (Cervus
elaphus): from a scoring scheme based on radiographs of developing permanent
molariform teeth, J. Zoo/., Land. (1991) 225, 85-97.
BUIKSTRA J. E. & SWEGLE M. (1989): Bone modification due to burning:
experimental evidence. In Bonnichsen, R. and Sorg, M. H. (ed.) Bone Modification.
Orono, University of Maine Center for the Study of the First Americans. Pp: 247-258.
BUNN H. T. (1981): Archaeological Evidence for meat-eating by Plio-Pleistocene
hominids from Koobi Fora and Olduvai Gorge. Nature. 291 (5816) Pp: 574-576.
BUNN H. T. & KROLL E. M. (1986): Systematic Butchery by Plio/Pleistocene
Hominids at Olduvai Gorge, Tanzania. Current Anthropology. 27(5) Pp:431-452.
BURRIEL F., LUCENA F., ARRIBAS S., HERNÁNDEZ J., Química Analítica
Cualitativa, 13a ed., Paraninfo, Madrid, 1989.
CACERES I., Tafonomìa de yacimientos antròpicos en Karts. Complejo Galerìa
(Sierra de Atapuerca, Burgos), Vanguard Cave (Gibraltar) y Abric Romanì (Capellades,
Barcelona). Tarragona 2002.
CÁCERES, I., LOZANO, M. y SALADIÉ, P. (en prensa): “Evidence for Bronze
Age Cannibalism in El Mirador Cave (Sierra de Atapuerca, Burgos, Spain)”. American
Journal of Physical Anthropology.
CAPALDO, S.D. (1998): “Methods, marks and models for inferring hominid and
carnivore behavior”. Journal of Human Evolution, 35: 323-326.
COTTON F.A., WILKINSON G., Química Inorgánica avanzada, 4a ed., Limusa,
México, 1993.

135
D’ERRICO F., 1989, L’art gravé azilien. Analyse microscopique, reconstitution
gestuelle, signification, Thèse de Doctorat, Institut de Palèontologie Humaine, Paris.
DAVIS S. J. M., 1987, The Archaeology of Animals, Batsford, London.
DE GROSSI MAZZORIN J., Archeozoologia. Lo studio dei resti animali in
archeologia. Bari: Editori Laterza, 2008.
EFREMOV J. A., 1940, Taphonomy: New Branch of Paleontology, in «Pan-
American Geologist», 74, pp. 81-93.
EPM (Ente Parco delle Madonie), 2004. Guida geologica del Parco delle Madonie.
Ente Parco delle Madonie & Università degli Studi di Palermo, Palermo.
FIORILLO A. R. 1989. An experimental study of trampling: implications for the
fossil record. In (R. Bonnichsen and M. H. Sorg, eds.) Bone modification, pp. 61-71.
Orono: University of Maine Center for the Study of the First Americans.
FORGIA V., et al., New data on Sicilian prehistoric and historic evolution in a
mountain context, Vallone Inferno (Scillato, Italy). C. R. Palevol (2012).
GEJVALL N. G. (1969): Cremations. In Brothwell, D. and Higgs, E. (ed.) Science
in Archaeology. New York, Praeger. Pp: 468-479.
GIACOBINI G. 1995, Identificazione delle tracce di macellazione con strumenti
litici. Analisi di microscopia elettronica a scansione, Padusa, Atti 1° Convegno Nazionale
di Archeozoologia, Rovigo, pp. 29-38.
GIANNITRAPANI E., IANNÌ F., et al., Case Bastione: a prehistoric settlement in
the Erei uplands (Central Sicily), Origini, in corso di stampa.
GIFFORD-GONZALEZ, D. (1989a): Overview-Modern Analogues: Developing an
Interpretative Framework. In Bonnichsen, R. and Sorg, M. H. (ed.) Bone Modification.
Orono: University of Maine Center for the Study of the First Americans. pp: 43-52.
GIFFORD-GONZALEZ, D. (1989b): Ethnographic Analogues for Interpreting
Modified Bones: Some Cases from East Africa. In Bonnichsen, R. and Sorg, M. H. (ed.)
Bone Modification. Orono: University of Maine Center for the Study of the First
Americans. pp: 179-246.
GIFFORD-GONZALEZ D. P., DAMROSCH D. B., DAMROSCH D. R., PRYOR
J. & THUNEN R. L. (1985): The Third Dimension in Site Structure: An Experiment in
Trampling and Vertical dispersal. American Antiquity. 50(4) pp. 803-818.
GIUFFRIDA RUGGERI V., 1901–1902. Materiale paletnologico di una caverna
naturale di Isnello presso Cefalù in Sicilia. Atti della Società Romana di Antropologia
VIII, 337–68.
GIUFFRIDA RUGGERI V., 1905. Terzo contributo all’antropologia fisica dei
Siculi eneolitici. Atti della Società Romana di Antropologia XI, 56–103.

136
GRAYSON, D. K. (1988): Danger Cave, Last Supper Cave, and Hanging Rock
Shelter: the Faunas. American Museum of Natural History Anthropological Papers. 66(1)
Pp:1-130.
GREENFIELD H. J. 1999, The Origins of Metallurgy: Distinguishing Stone from
Metal Cut-marks on Bones from Archaeological Sites, in Journal of Archaeological
Science, 26, 797–808.
GREENFIELD, H. J., 2006, Slicing Cut Marks on Animal Bones: Diagnostics for
Identifying Stone Tool Type and Raw Material, Journal of Field Archaeology, 31 (2): 1 47-
163.
HABERMEHL K. H., 1961, Die Altersbestimmung bei Haustieren, Pelztieren und
beim jagdbaren Wild, Paul Parey Verlag, Berlin.
HAYNES, G. (1980): Prey bones and predators: potential ecologic information
from analysis of bone sites. Ossa. 7 Pp: 75-97.
HAYNES, G. (1983a): Frequencies of spiral and green-bone fractures on ungulate
limb bones in modern surface assemblages. American Antiquity. 48(1) Pp: 102-14.
HAYNES, G. (1983b): A guide for differentiating mammalian carnivore taxa
responsible for gnaw damage to herbivore limb bones. Paleobiology. 9 Pp: 164-72.
HILL C.A., Origin of black deposits in caves, National Speleological Society
Bulletin 44 (1982) 15e19.
HILSON S., Mammal Bones and teeth. An introductory guide to methods of
identification, Cambridge, 1986.
HOLLOWAY R. R., LUKESH S.S., MANISCALCO L., MCCONNEL B.E., La
Muculufa: un centro sul fiume Salso e il suo raggio di contatti, in L’antica età del Bronzo
in Italia, Atti del convegno, Viareggio 1995, pp. 297-303.
IANNÌ F., La Muculufa Santuario: considerazione tecnologiche, morfologiche e
stilistiche sulle classi vascolari e lo Stile di Muculufa, Riviste di Scienze Preistoriche LIX,
2009, pp 243-264.
IANNÌ F., L’insediamento di età preistorica di Case Bastione (Villarosa, Enna), da
Scheda siti di Tornambè e Case Bastione, www.academia.edu, 2013.
IRVING W. N., JOPLING A. V. & KRITSCH-AMSTRONG I. (1989): Studies of
Bone Technology and Taphonomy, Old Crow Basin, Yukon Territory. In Bonnichsen, R.
and Sorg, M. H. (ed.) Bone Modification. Orono: University of Maine Center for the Study
of the First Americans. Pp: 347 -379.
JIMÉNEZ BROBEIL S.A., 1990. Rituales funerarios neolíticos en la alta
Andalucía. Estado actual de la cuestión. Zephyrus XLIII, 125e130.

137
JOHNSON, E. (1983): “A framework for interpretation in bone technology” en
LEMOINE, G.M. y MACEACHEM, A.S. (Eds.): Carnivores, Human Scavengers and
Predators: A Question of Bone Technology. Calgary: 55-95.
KEAY S. J., Late Roman Amphorae in the Western Mediterranean. A typology and
economic study: the Catalan evidence. BAR International Series (i), Oxford,1984, p. 306
n. 1.
KENT, S. (1993): “Variability in Faunal Assemblages: The Influence of Hunting
Skill, Sharing, Dogs and Mode of Cooking on Faunal Remains at a Sedentary Kalahari
Community”. Journal of Anthropological Archaeology, 12: 323-385.
LAUDET, F. & FOSSE, P. (2001): Un Assemblage d'Os Grignoté par les Rongeurs
au Paléogène (Oligocène Supérieur, Phosphorites du Quercy). C.R. Acad. Sc. Paris. 333
Pp: 195-200.
LÓPEZ-GONZÁLEZ F., GRANDAL-D’ANGLADE A., VIDAL-ROMANÍ J. R.,
Deciphering bone depositional sequences in caves through the study of manganese
coatings. In Journal of Archaeological Science 33 (2006) pp. 707-717.
LYMAN R. L., 1994, Vertebrate Taphonomy, Cambridge University Press,
Cambridge.
MAGUIRE, J. M., PEMBERTON, D. & COLLETT, M. H. (1980): The
Makapansgat Limeworks grey breccia: Hominids, hyaenas, hystricids or hillwash.
Paleontologia Africana. 23 Pp: 75-98.
MANGANO G. 2000. Resti faunistici da insediamenti dell’antica e media età del
Bronzo a Messina. In Ati del 2° Convegno Nazionale di Archeozoologia. Asti, 14-16
novembre 1997, ABACO, Forli, pp. 257-261.
MANNINO G., 1989. L’abisso del Vento. Sicilia Archeologica 69–70, 49–52.
MANNINO M.A., PLUCIENNIK M. & GIANNITRAPANI E. (November 2006).
Risultati preliminari dello studio archeozoologico dei reperti osteologici dal Riparo San
Tommaso (Enna). V Convegno Nazionale di Archeozoologia [5th Italian National
Archaeozoology Conference], Museo di Rovereto, Rovereto (Trento), Italy.
MASCARO I., PERUSIN S., SARTI L., La “quotidianità” del metallo: analisi al
SEM delle tracce di macellazione dei reperti faunistici del sito campaniforme di Via
Bruschi a Sesto Fiorentino. In Atti del 6° Convegno Nazionale di Archeozoologia 2009,
pp. 7-9.
MAYNE CORREIA P. M. (1997): Fire Modification of Bone: A Review of the
Literature. In Haglund, W. D. and Sorg, M. H. (ed.) Forensic Taphonomy: The
Postmortem Fate of Human Remains, CRC Press, Inc. Pp: 275 -293.
MILLER, G. J. (1975): A Study of Cuts, Grooves, and Other Marks on Recent and
fossil Bone: II. Weathering, Cracks, Fractures, Splinters, and other Similar Natural
Phenomena. In Swanson, E. (ed.) Lithic Technology. The Hague, Mouton.

138
MINÀ PALUMBO F., 1869. Paletnologia sicula delle armi in pietra raccolte in
Sicilia, Palermo.
MORLAN R.E. (1980): Taphonomy and archaeology in the Upper Pleistocene of
the northern Yukon Territory: a glimpse of the peopling of the New World. 94 Mercury
Series. Ottawa.
MORLAN R.E. (1983): « Spiral fractures on limb bones: which ones are artificial?
», in LEMOINE, G.M. y MACEACHEM, A.S. (Ed): Carnivores, Human Scavengers and
Predators: A Question of Bone Technology. Calgary: 241-271.
NICHOLSON R. (1993): A Morphological Investigation of Burnt Animal Bone and
an Evaluation of its Utility in Archaeology. Journal of Archaeological Science. 20 Pp:411-
28.
OLIVER J. S. (1989): Analogues and Site Context: Bone Damages from Shield
Trap Cave (24 CB91), Carbon County, Montana, U.S.A. In Bonnichsen, R. and Sorg, M.
H. (ed.) Bone Modification. Orono, University of Maine Center for the Study of the First
Americans. pp: 73 -98.
OLSEN S., SHIPMAN P. 1988, Surface modification of bone: trampling versus
butchery, in “Journal of Archaeology Science”, 40, pp. 535-553.
ORLANDINI P., Il villaggio preistorico di Manfria, presso Gela, Palermo, 1962,
pp. 40-45.
ORSI P., Scarichi del villaggio di Castelluccio (Sicilia), Bullettino della
Paletnologia Italiana, XIX, 1893.
PALES L., LAMBERT C., Atlas Ostéologique pour server à l’identification des
Mammifères du Quaternaire, I Les membres Herbivores, Edition du centre national de la
recherche scientifique, 15, quai Anatole-France – Paris-VII, 1971.
PAYNE S., Morphological Distinctions between the Mandibular Teeth of Young
Sheep, Ok and Goats, Capra, in Journal of Archaeological Science, 1985, 12, 139-147.
PERETTO C.; ANCONETANI P.; CROVETTO C.; EVANGELISTA L.;
FERRARI M.; GIUSBERTI G.; THUN HOHENSTEIN U. & VIANELLO F. (1996):
Approccio Sperimentale alla Compresione delle Attività di Sussistenza Condotte nel Sito di
Isernia La Pineta (Molise-Italia). La Fratturazione Intenzionale. In Peretto, C. (ed.) I
Reperti Paleontologici del Giacimento Paleolitico di Isernia La Pineta. Isernia, Cosmo
Iannone Editore. pp: 187-452.
PICKERING, T. y EGELAND, C. (2006): “Experimental patterns of hammer
stone percussion damage on bones: implications for inferences of carcass processing by
humans”. Journal of Archaeological Science, 33: 459-469.
PROCELLI E., 2013, Popoli migranti: catalogo della mostra : Ramacca, Museo
archeologico civico.

139
RAMOS MUÑOZ, J, 1988-1989. Las industrias líticas del neolítico en Andalucía,
sus implicaciones espaciales y económicas. Zephyrus. Revista de prehistoria y arqueología
41e42, 113e148.
ROSELL, J. (2001): Patrons d’aprofitament de les biomasses animals Durant el
Pleistocè Inferior I Mig (Sierra de Atapuerca, Burgos) i Superior (Abric
Romaní,Barcelona). Tesis Doctoral Tarragona. Departament d’Historia I Geografia.
Universitat Rovira I Virgili.
SALADIÉ, P. (1998): Estudi zooarqueológic dels nivells del conjunt II de L’Abric
Romaní (Capellades, Barcelona). Tesis de Licenciatura. Tarragona. Departament
d’Historia i Geografia. Universitat Rovira i Virgili.
SCHMID E., Atlas of animal bones for Prehistorians, Archaelogists, and
Quaternary Geologists, Amsterdam, London, New York: Elsevier Publishing Company,
1972.
SHAHACK-GROSS, R., BAR-YOSEF, O. y WEINER, S. (1997): “Black-
Coloured
Bones in Hayonim Cave, Israel: Differentiation Between Burning and Oxide Staining”.
Journal of Archaeological Science, 24: 439-446.
SHIPMAN P. 1981, Applications of scanning electron microscopy to taphonomic
problems, in “Annals of the New York Academy of Science”, 376, pp. 357-385.
SHIPMAN, P. y ROSE, J. (1983): “Early Hominid Hunting, Butchering, and
Carcass- Processing Behaviors: Approaches to the Fossil Record”. Journal of
Anthropological Archaeology, 2: 57-98.
SHIPMAN, P. et al. (1984): “Burnt Bones and Teeth: an Experimental Study of
Color,
Morphology, Crystal Structure and Shrinkage”. Journal of Archaeological Science, 11:
303-325.
SILVER I. E., 1969, The ageing of domestic animals, D. Brothwell & E. S. Higgs
(Eds), Science in Archaeology, Thames & Hudson, London.
STINER, M. C. (1994): Honour Among Thieves. A Zooarchaeological Study of
Neandertal Ecology. Princeton University Press. Princeton.
STINER M. C.; WEINER S.; BAR-YOSEF O. & KUHN S. L. (1995): Differential
Burning, Recrystallization and Fragmentation of Archaeological Bone. Journal of
Archaeological Science. 22 Pp:223-37.
SOLARI A., OLIVERA D., GORDILLO I., BOSCH P., FETTER G., LARA V. H.,
NOVELO O. (2013), Cooked bones? Method and practice for identifying bones treated at
low temperature, in International Journale of Osteoarchaeology.

140
SUTCLIFFE, A. J. & COLLINGS, H. D. (1972): Gnawed Bones from the Crag and
Forest Red Deposits of East Anglia. Suffolk Natural History. 15(6) Pp: 497 -498.
TURNER, C.G. (1983): “Taphonomic reconstruction of human violence and
cannibalism based on mass burials in the American Southwest” en LEMOINE, G.M. y
MACEACHEM, A.S.: Carnivores, Human Scavengers and Predator: A Question of Bone
Technology. Calgary: 219-240.
TUSA S., La Sicilia nella preistoria, Sellerio, Palermo, 1999.
TUSA S., Sicilia preistorica, Dario Flaccovio Editore, 1994, p. 126.
VILLA, P. y MAHIEU, E. (1991): “Breakage patterns of human long bones”.
Journal of Human Evolution, 21: 27-48.
VON DEN DRIESCH A., BOESSNECK J., 1974. “Kritische Anmerkunger zur
Widerristhöenberechnung aus Längemassen vor- und frühgeschitlicher Tierknochen,
Säugetierkundliche Mitteilungen”, Munchen, pp. 325-348.
VON DER DRIESCH A., 1976. A Guide to Measrement of the Animal Bones from
Archaeological Sites. In “Peabody Museum Bullettins”, l, pp. 1-138.
WALKER, P. L. (1978). Butchering and stone tool function. American Antiquity
43(4), 710–715.
WHITE, T. (1992): Prehistoric Cannibalism at Mancos 5MTUMR-2346. Princeton:
Princeton University Press.
WILKENS B., 2003, Archeozoologia. Manuale per lo studio dei resti faunistici
dell’area mediterranea. CD rom, Schio.
WILSON R. J., Brick and tiles in Roman Sicily, in A. Mc Whirr (Ed), Roman brick
and tile, London 1979, pp. 11-37.
YELLEN J. E. (1991): Small Mammals: Post-discard Pattering of Kung San
Faunal Remains. Journal of Anthropological Archaeology. 10 Pp: 152-192.
ZEDER M. A., LAPHAM H. A., Assessing the reliability of criteria used to
identify postcranial bones in sheep, Ovis, and goats, Capra, Journal of Archaeological
Science, 37, (2010), 2887-2905.

141
APPENDICE

142

143
TABELLE OSTEOMETRICHE
Legenda:
- Str.: strato
- q.: quadrato
- n.: numero
- Dx: destro
- Sx: sinistro
Misure espresse in mm
Vallone Inferno
Ovicaprini
Prima falange
Str. q. n. Bd SD
3.4.A G19 40 11 9
Seconda falange
Str. q. n. GLpe GL Bp SD Bd
3.4.A G19 15 25 25 11 6 7
3.4.A F23 15 21 23 11 8 9
3.4.B G22 45 21 22 11 7 8
3.4.B H18 8 / 23 11 8 8
Terza falange
Str. q. n. Ld MBS DLS
3.4.A G21 27 24 7 24
Radio
Str. q. n. Lato Bp
3.4.A F20 85 dx 33
Omero
Str. q. n. Lato Bd
3.4.A G19 28 dx 30

144
Tibia
Str. q. n. Lato Bd SD Dd
3.4.A F19 57 sx 27 14 21
Scafoide
Str. q. n. GB
3.4.A H18 24 21
Scafocuboide
Str q. n. GB
3.4.B F23 22 22
Capitato-trapezoide
Str q. n. GB
3.4.B I23 57 16
3.4.B G21 67 18
3.4.B H18 14 17
Semilunare
Str q. n. GB
3.4.B I23 58 12
3.4.B F20 31 14
Piramidale
Str q. n. GB
3.4.B I23 59 16
Uncinato
Str q. n. GB
3.4.B I23 60 13
Astragalo
Str. q. n. Lato GLl GLm Dl Dm Bd
3.4.B G20 30 Dx 28 26 16 16 17

145
Ovis aries
Astragalo
Str. q. n. Lato GLl GLm Dl Dm Bd
3.4.B F19 3 Dx / 26 15 14 17
3.4.B F20 15 Sx 26 25 15 15 17
3.4.B H18 19 Sx 24 23 14 14 15
Bos taurus
Ulna
Str. q. n. Lato LO DPA SDO BPC
3.4.B E21 12 Dx 54 36 31 23
Metacarpo
Str. q. n. Lato Bd
3.4.A G18 10 Dx 58
Terza falange
Str. q. n. DLS Ld MBS
3.4.A G19 46 71 53 22
Scafoide
Str. q. n. GB
3.4.B G19 2 31
Semilunare
Str. q. n. GB
3.4.A H19 14 51
Cervus elaphus
Femore
Str. q. n. Lato Bp
3.4.A I21 12 Sx 63

146
Metatarso
Str. q. n. Lato Bp Dp
3.4.A G19 52 Dx 30 33
Prima falange
Str. q. n. Dp Bp
3.4.A G22 48 22 17
Str. q. n. GLpe GL Bp SD Bd
3.4.B G23 16 52 55 20 15 19
Seconda falange
Str. q. n. GL SD Bd
3.4.A G19 9 35 14 12
Str. q. n. GL Bp SD Bd
3.4.B G21 65 33 15 13 13
Calcagno
Str. q. n. Lato GL Gb
3.4.A G19 24 Dx 91 28
Canide
Prima falange
Str. q. n. GL Bp SD Bd
3.4.A E20 31 23 6 3 5
3.4.A H18 21 19 5 5 7
3.4.A H18 27 18 5 4 7
Felis sp.
Ulna
Str. q. n. Lato DPA SDO
3.4.B G21 54 Dx 13 11

147
Case Bastione
Ovicaprini
Omero
Str. q. n. Lato Bd
1057 A 101 II NW 7 Dx 24
Radio
Str. q. n. Lato Bp BFp
2083 A 102 IV SE 35 Sx 26 24
Tibia
Str. q. n. Lato Bd
1057 A 101 I SE 37 Dx 23
Metatarso
Str. q. n. Lato Bp
1057 A 101 I SE 33 Sx 17
Prima falange
Str. q. n. SD Bd
1057 A 101 III NE 10 11 12
2087 A 102 I NW 10 7 9
Seconda falange
Str. q. n. GLpe GL Bp SD Bd
2001 A 100 IV NE 8 18 20 10 7 9
2087 A 102 I NE 25 17 19 10 8 7,5
Capitato trapezoide
Str. q. n. GB
1057 A 101 III SW 19 17
Astragalo
Str. q. n. Lato GLl GLm Dl Dm Bd
1057 A 101 I SE 19 Sx 26 25 14 13 12

148
Sus domesticus
Secondo metatarso
Str. q. n. GL
2083 A 102 IV SE 26 66
Prima falange
Str. q. n. Bd
2079 A 102 II SW 1 16
Seconda falange
Str. q. n. GL Bp SD Bd
1057 A 101 II NW 13 23 15 11 12
1057 A 101 III NE 9 21 15 13 13
Bos taurus
Tibia
Str. q. n. Bd
2083 A 102 IV SW 20 54
Metacarpo
Str. q. n. Lato Bp
2079 A 102 III NW 54 Sx 33
Capitato-trapezoide
Str. q. n. GB
2079 A 102 IV SE 19 29
Prima falange
Str. q. n. SD Bd
2087 A 102 I NE 38 22 27
Seconda falange
Str. q. n. GLpe GL Bp SD Bd
1057 A 100 II SW 9 44 51 30 24 26

149
2001 A 100 IV NE 3 / 39 26 21 22
Cervus elaphus
Piramidale
Str. q. n. GB
1057 A 100 II SE 4 26
Metatarso
Str. q. n. Bd
1057 A 101 I SE 9 37

150
TAVOLA N. 1
Vallone Inferno str. 3.4.A q. G18 n. 10; Epifisi e metafisi distale di un metacarpo destro di Bos taurus con
strie di macellazione. E-F presenti nel condilo nel lato plantare, G-H situate nella metafisi nella parte laterale.
A B
C D
E F
G-H
G H
E-F

151
TAVOLA N. 2
Vallone Inferno str. 3.4.A q. F19 n.69; Costa taglia media con strie da strumento litico, situate nella parte interna
(C-D-E-F).
A
B
C D
C-D-E-F
E F

152
TAVOLA N. 3
Vallone Inferno str. 3.4.B q. F22 n. 30; costa taglia piccola, raschiature da strumento litico rinvenute nella
parte interna (C-D).
Immagine al SEM delle raschiature, ove
sono visibili i solchi a sezione quadrangolare e le striature secondarie.
TekneHub (E).
A
B
C D
C-D
E

153
TAVOLA N. 4
Vallone Inferno str. 3.4.B q. F20 n. 30; astragalo destro di ovicaprino (A-B-C-D, tagli
presenti nella parte mediale (E-F-G).
A B
C D
E F
G
E-F-G

154
TAVOLA N. 5
Vallone Inferno str. 3.4.B q. F20 n. 31; semilunare di ovicaprino, nell’immagine B sono presenti
tracce di morsi di roditori.
Vallone Inferno str. 3.4.B q. F20 n. 7; Osso
lungo taglia media; pitting nella figura D.
A B
C D
E
D
B

155
TAVOLA N. 6
Case Bastione, Area β, Capanna 1, str. 1057 q. A 100 II NE n. 7; radio sinistro di ovicaprino, con
tre tagli con orientamento trasversale e paralleli tra di loro, presenti nella parte mediale. L’osso è
parzialmente combusto.
A B
C D
E F
E-F
E-F
E-F

156
TAVOLA N. 7
Case Bastione, Area β, Capanna 1, str. 1057 q. A 101 III SW n. 33; pisiforme con strie da
strumento litico.
Dettaglio al SEM del solco delle strie a fondo quadrangolare, ove sono visibili le striature
secondarie prodotte dallo strumento litico. TekneHub.
D
E F
A
E
F
B
C
C-D

157
TAVOLA N. 8
Vallone Inferno str. 3.4.A q. G19 n. 45; emimandibola destra di Ovis aries.
Vallone Inferno str. 3.4.B q. H19 n. 1; canino Sus scrofa.

158
TAVOLA N. 9
Vallone Inferno str. 3.4.A q. G19 n. 46; terza falange di Bos taurus con tracce di ossido di
manganese.
Vallone Inferno str. 3.4.A q. G21 n. 25; emimandibola sinistra di cervide con il quarto premolare
deciduo.

159
TAVOLA N. 10
Case Bastione Area β Capanna 1 str. 1057 q. A 100 II NE n. 2; terzo molare destro di ovicaprino
con alcune tracce da combustione.
Case Bastione Area β Capanna 1; omero sinistro di Sus domesticus.

160
TAVOLA N. 11
Case Bastione Area β Capanna 1; diverse gradazioni di colore a seconda della diversa esposizione alla fonte di calore: A: Grado 2; B: Grado 3; C: Grado: 4; D: Grado 5. Il Grado 1 corrisponde a un
colore marrone non uniforme.
Case Bastione Area β Capanna 1; frammento di un cranio attribuito a Canis familiaris, sono
presenti P1, P2, P3 e M1 ed M2.
A
B C D

161
TAVOLA N. 12
Vallone Inferno 3.4.B F20 n. 15; astragalo sinistro di pecora (Ovis aries), sono presenti strie da strumento
litico nella parte mediale (B-C).
Dettagli al SEM dei solchi delle strie di macellazione con sezione a V, in cui sono visibili le
striature secondarie prodotte dallo scorrimento di uno strumento litico. TekneHub (D-E).
A
B C
B-C
D E
D-E

162

163
RINGRAZIAMENTI
I primi ringraziamenti vanno sicuramente alla dott.ssa Daria Petruso la quale è stata una
delle protagoniste del mio percorso di crescita professionale, sin dalla laurea triennale. La
ringrazio per il sostegno datomi nei momenti più difficili. Un grosso ringraziamento va alla
dott.ssa Vincenza Forgia, che mi ha dato la possibilità concreta di approcciarmi al mondo
dell’archeologia e dell’archeozoologia, se sono arrivato a questo punto molto lo devo a lei.
Ringrazio tantissimo la mia relatrice, la dott.ssa Ursula Thun Hohenstein, per tutto quello
che ho imparato da lei e per il tempo dedicatomi. Ringrazio il professor Oscar Belvedere,
la dott.ssa Rosa Maria Cucco e il dott. Stefano Vassallo, che hanno reso possibile la
realizzazione dello scavo di Vallone Inferno il quale sta dando a giovani archeologici la
possibilità di crescere professionalmente. Ringrazio anche la dott.ssa Patricia Martín
Rodríguez, dal punto di vista professionale la considero come un punto di riferimento
molto difficile da raggiungere. Ringrazio Marco Bertolini, per i suoi preziosi consigli,
anche lui mi ha aiutato moltissimo. Un grosso ringraziamento va anche alla dott.ssa Lisa
Volpe che ha permesso di realizzare le analisi al SEM. Ringrazio il dott. Enrico
Giannitrapani e il dott. Filippo Iannì, non solo per avermi dato la possibilità di analizzare il
campione faunistico di Case Bastione, anche per la loro disponibilità e per la fiducia
datami. Ringrazio tantissimo la mia compagna, per il suo appoggio e per i suoi
incoraggiamenti a non mollare mai, sono sicuro che senza di lei non ce l’avrei mai fatta.
Naturalmente ringrazio anche i miei genitori, tutta la mia famiglia, i miei amici e miei
colleghi che mi sono stati sempre vicini e con i quali ho condiviso questo lungo percorso.
Grazie a tutti.


![Lettura di "Inferno XXVIII" [2013]](https://static.fdokumen.com/doc/165x107/6320a8fcc5de3ed8a70dd3e8/lettura-di-inferno-xxviii-2013.jpg)

















