
Genetic effects of a maximal harvest schedule
12
Zoo Biology 5:203-214 (1986) Genetic Effects of a Maximal Harvest Schedule Bennett Dyke, Timothy B. Gage, and Paul M. Mamelka Genetics Department, Southwest Foundation for Biomedical Research, San Antonio, Texas A mathematical method of maximizing the reproductive efficiency of captive populations is evaluated for its efficacy in very small breeding groups and for its effect on the loss of genetic variability. The evaluation consists of comparing reproductive performance of simulated populations having demographic charac- teristics similar to a small troop of rhesus kept at the Vilas Park Zoo in Madison, WI, under two age-specific removal strategies. It was found that a “maximal harvest schedule” (in which a portion of the animals reaching age 1, and all animals age 19, were removed) resulted in a higher rate of reproduction than did a “random harvest schedule” that removed a fixed proportion of all age classes. Little or no difference between the two removal strategies was found in between- locus variance calculated for 40 simulated genetic loci. From these results we conclude that a maximal harvest strategy may be an effective way to increase reproductive efficiency in small captive populations, and that such a strategy per se may not accelerate the loss of genetic variability. Key words: captive breeding, reproductive efficiency, genetic variability, rhesus monkeys INTRODUCTION There are a number of circumstances in which it may be advantageous to maximize the rate of growth or productivity of a captive animal population [Foose et al, 19861. This is often true for domesticated animals but may also be important in efforts to minimize the threat of extinction of small populations and in programs of planned colonization or reintroduction of wild populations. In many instances, popu- lation growth may be stimulated simply by increasing available nutrients or territory, but in closely managed populations, it may also be possible, where resources are limited, to improve productivity by manipulating the demographic structure. A potential danger in accelerating growth rates, particularly in closed popula- tions, is a concomitant increase in the rate of loss of genetic variability due to genetic drift. We report here on a computer simulation experiment designed to test the genetic Received for publication September 17, 1985; accepted October 22, 1985. Address reprint requests to Dr. Bennett Dyke, Genetics Department, Southwest Foundation for Biorned- ical Research, PO Box 28147, San Antonio, TX 78284. 0 1986 Alan R. Liss, Inc.
Transcript of Genetic effects of a maximal harvest schedule

Zoo Biology 5:203-214 (1986)
Genetic Effects of a Maximal Harvest Schedule Bennett Dyke, Timothy B. Gage, and Paul M. Mamelka
Genetics Department, Southwest Foundation for Biomedical Research, San Antonio, Texas
A mathematical method of maximizing the reproductive efficiency of captive populations is evaluated for its efficacy in very small breeding groups and for its effect on the loss of genetic variability. The evaluation consists of comparing reproductive performance of simulated populations having demographic charac- teristics similar to a small troop of rhesus kept at the Vilas Park Zoo in Madison, WI, under two age-specific removal strategies. It was found that a “maximal harvest schedule” (in which a portion of the animals reaching age 1, and all animals age 19, were removed) resulted in a higher rate of reproduction than did a “random harvest schedule” that removed a fixed proportion of all age classes. Little or no difference between the two removal strategies was found in between- locus variance calculated for 40 simulated genetic loci. From these results we conclude that a maximal harvest strategy may be an effective way to increase reproductive efficiency in small captive populations, and that such a strategy per se may not accelerate the loss of genetic variability.
Key words: captive breeding, reproductive efficiency, genetic variability, rhesus monkeys
INTRODUCTION
There are a number of circumstances in which it may be advantageous to maximize the rate of growth or productivity of a captive animal population [Foose et al, 19861. This is often true for domesticated animals but may also be important in efforts to minimize the threat of extinction of small populations and in programs of planned colonization or reintroduction of wild populations. In many instances, popu- lation growth may be stimulated simply by increasing available nutrients or territory, but in closely managed populations, it may also be possible, where resources are limited, to improve productivity by manipulating the demographic structure.
A potential danger in accelerating growth rates, particularly in closed popula- tions, is a concomitant increase in the rate of loss of genetic variability due to genetic drift. We report here on a computer simulation experiment designed to test the genetic
Received for publication September 17, 1985; accepted October 22, 1985.
Address reprint requests to Dr. Bennett Dyke, Genetics Department, Southwest Foundation for Biorned- ical Research, PO Box 28147, San Antonio, TX 78284.
0 1986 Alan R. Liss, Inc.

204 Dyke, Gage, and Mamelka
effects of an optimized production schedule based upon demographic manipulation. The simulated population is based upon "ZOO troop 4," a small troop of rhesus monkeys belonging to the Wisconsin Regional Primate Research Center (WRPRC) but kept at the Vilas Park Zoo in Madison, Wisconsin.
Although rhesus monkeys are common in their natural environments, and in little danger of extinction, they represent a good model for the study of captive populations, including those of rarer species whose continued survival depends at the moment on captive breeding in zoos. Like more exotic species, rhesus monkeys are relatively undomesticated and of tropical origin, characteristics that require special attention to diet, health care, and behavioral monitoring. Unlike most zoo animals, however, demographic and genetic data are available from large colonies where animals are raised for biomedical research, a fact that makes this species particularly attractive for the study of population processes in captive breeding groups.
Because of the reduced availability of wild animals, captive colonies in zoos and other installations are faced increasingly with the necessity of maintaining self- sustaining breeding populations for an indefinite period of time. Under these circum- stances, issues of long-term colony survival become critical. A fundamental conflict arises between the practical requirements of colony management and the goal of preventing serious reduction in fertility and viability that accompanies the loss of genetic variability in the population (inbreeding depression).
Unfortunately, several characteristics of captive breeding programs make the maintenance of genetic variability a difficult task.
SMALL SIZES OF BREEDING COLONIES
A major source of loss of genetic variability is random genetic drift, or the failure of genes to be passed to the next generation simply by chance. Loss from this source is inversely proportional to population size, and because most breeding colo- nies (particularly in zoos) are very small, drift is an important concern. Economic constraints make it unlikely that colony size can be increased sufficiently to reduce the effects of drift in a substantial way.
ISOLATION OF COLONIES
Breeding colonies tend to be geographically and reproductively isolated from one another-quite a different situation from the wild, where genes lost from one group can be reintroduced from another simply because mating can occur across population boundaries. Intercolony transfer of animals has been proposed as a way to remedy the problem of isolation. However, the effectiveness of this practice depends upon the extent to which animals will tolerate the disruption of established social and breeding hierarchies and upon expensive scientific and logistic planning at the national or international level.
IMBALANCE OF SEX RATIO AND VARIABLE NUMBERS OF OFFSPRING
Like small population size, an imbalanced sex ratio of breeders and a nonbinomi- a1 distribution of offspring per parent affects the magnitude of loss of genes through genetic drift. Equalizing the sex ratio of breeding animals can sometimes be accom-

Genetic Effects of Maximal Harvest 205
plished, but there are cases (such as harem groups) where it is inconsistent with the species’ social and breeding structure. Controlling the distribution of numbers of offspring is likely to be a difficult task, particularly in the case of animals that are subfertile due to biological causes.
SHORT GENERATION LENGTH
Colony managers often retain breeding animals only during their peak reproduc- tive years. This practice increases efficiency of production, but it can reduce the average time between the birthdates of parents and their offspring, thereby increasing the rate at which drift occurs. Choice of last-born offspring to replace parental breeding stock may be a way to slow the rate of loss of genetic variability, although the effectiveness of this policy is not clear at present.
METHODS
Developing a management strategy that takes into account both short-term financial constraints and longer-term genetic requirements is a formidable task, particularly since there is little experience to guide us, and the risks of relying upon trial and error are so great. In order to overcome these difficulties, we have turned to computer simulation as a means to experiment with the interacting effects of compet- ing management goals: we create artificial populations that have the demographic and genetic characteristics of real breeding colonies, subject them to various management strategies, and observe the outcome.
The strategy of interest here is a method of increasing reproductive efficiency by adjusting the age-sex structure of the population in such a way as to maximize the number of births for a given colony size. Use of this method is not intended to solve or circumvent many of the serious problems described above, but it is attractive in that it appears to be relatively inexpensive, easy to implement, and minimally disrup- tive to colony structure. The approach is based on a result due to Beddington and Taylor [1973] which shows that any age-specific property of the Leslie matrix model can be maximized by partial removal of individuals in a single age category (usually early in life) and by complete removal of a later age group (usually toward the end of the life span). The method is of potential economic value in captive breeding programs and has been cited by Goodman El9801 as an appropriate tool for wildlife manage- ment, although to our knowledge it has never been applied for either purpose.
Parameters required for the determination of a maximum harvest model are age-specific birth and death rates. We have modified the basic single-sex model so that it will accommodate both sexes by adjusting female fertility rates according to the sex ratio at birth and by incorporating sex-specific death rates. Vital rates have been taken from an analysis of the Wisconsin Regional Primate Research Center rhesus colony, where records have been kept for over 20 yr on more than 4,700 animals.
The solid line in Figure 1 represents the stable age-sex structure based upon age-specific rates of fertility and mortality calculated from data collected from the WRPRC rhesus colony over the 20-yr period, and a removal schedule that takes from the colony a fixed proportion of animals without regard to age. This “random harvest schedule” is in fact quite similar to the pattern of removal that actually applied to the

206 Dyke, Gage, and Mamelka
Proportion
MALES FEMALES
Fig. 1. (solid line) and maximal harvest schedule (broken line).
Stable age-sex structures based on WRPRC birth and death rates for random harvest schedule
WRPRC colony, where a detailed demographic analysis of the population [Dyke et al, 19861 revealed that age-specific removal rates were remarkably uniform across all ages. More than twice the number of animals were removed for sale or research than died during this time.
The pyramid formed by the broken line represents the population structure as it would appear with the same fertility and mortality rates, but with age-specific removal rates adjusted for maximum production efficiency according to the Beddington and Taylor model-that is, so that the colony would produce the maximum number of births from a given total colony size. We refer to this strategy as “maximal harvest.” Differences between the two strategies can be seen by comparing the two pyramids: the maximal scheme specifies harvesting about 43% of the females and 80% of the males in their first year and culling all adults at age 19. This minimizes maintenance in the colony of those animals that would otherwise die before being removed and of the older breeders that are no longer reproducing at a high rate.
Theoretically, the random strategy should permit an annual harvest of about 6 % of the living population, while the maximal strategy should yield about 9%, a 50% improvement. This result assumes that birth, death, and removal rates are constant over time, and that the population being analyzed is large enough that random fluctuation in numbers of vital events is negligible. These conditions seldom hold in captive animal colonies, and we know little about the accuracy and reliability of deterministic projection techniques when they are applied to more realistic situations. Computer simulation can be used very effectively here to evaluate the importance of these limitations.
Perhaps more important than uncertainties about the efficacy of a maximal harvest strategy as applied to small populations are serious questions about its genetic

Genetic Effects of Maximal Harvest 207
consequences. On the one hand, comparison of the two pyramids suggests that removal of all animals at age 19 might well reduce the generation length, and if the scheme is successful, its adoption could lead to fewer animals being retained as breeders. Either result would increase the magnitude of genetic drift. On the other hand, systematic removal at a single early age, and truncation at the end of the reproductive period, might reduce the variance of offspring number, thereby reducing the effects of drift. Real-life evaluation of maximal harvest would be costly, requiring experimentation with many colonies, and it would take many years before results would be available. Loss of genetic material, should it occur as a result of experimen- tation, would be very difficult to replace. Here again, simulation can provide answers that are impractical to get otherwise.
The computer program we have developed for simulation of primate breeding colonies (ColonySim) is a stochastic discrete event microsimulation model written in FORTRAN (see Appendix). Input consists of age-specific probabilities of birth, death, and harvest, sex ratio at birth, and a starting population of known demographic and genetic composition for which vital events are generated on a yearly basis. Output consists of a record of genotypes and vital events for each animal in the initial population, and their simulated descendents. These records are then summarized and analyzed by using standard demographic and genetic analysis programs.
In the experiments described here, each simulation replicate was begun with the same initial population. To produce this population, the zoo troop 4 colony (as it existed in August 1982) was projected forward 10 yr with the simulation program, using birth and death probabilities derived from vital statistics of the entire WRPRC rhesus colony. This was done to increase size somewhat (from 76 to 94 animals) and to allow us to start each replicate with a population whose pedigree had a realistic structure and depth and whose underlying age-sex structure was as consistent as possible with the vital rates used throughout the experiments. Under the influence of these birth and death rates alone, simulated populations would be expected to grow at a rate of 6.3% per year.
Artificial genotypes for each of 40 unlinked genetic loci were assigned to individuals without parents in the founding group by sampling from Hardy-Weinberg distributions with gene frequencies of 0.5. The purpose of assigning this somewhat unlikely array of genotypes was to provide a convenient vehicle for the simulation of Mendelian segregation rather than to create a realistic genotypic configuration. This random assignment of genotypes resulted in a mean frequency across all loci of 0.48. Subsequently, genotypes were assigned to simulated offspring according to the geno- types of their parents and according to the laws of segregation.
The control for our experiment was established by running the simulation with birth and death probabilities as described and with a random harvest schedule. In order to reproduce the overall sex ratio of zoo troop 4 in the simulated populations, twice the number of males as females were removed. Because population size differences can themselves affect our experimental results, we also prevented large departures from zero growth by adjusting harvest rates up or down on the basis of a yearly census. This is probably a fairly realistic form of population control.
The simulation was run for 100 simulated years, which, although ambitious from a practical point of view, has the advantage of increasing the generality of the results by ensuring that any inconsistencies between the initial population structure and the input probabilities based on the entire Wisconsin rhesus colony would have
208 Dyke, Gage, and Mamelka
10
4 0 t
6
m o a lI.xIIl 10
0 10 20 30 40 50 60 70 80 90 100
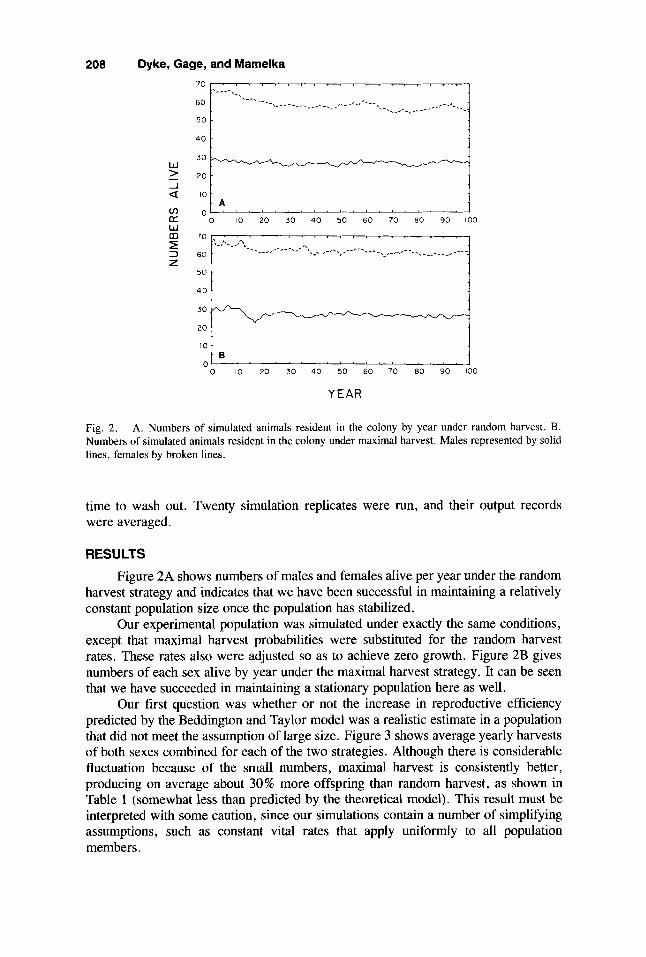
Fig. 2. A. Numbers of simulated animals resident in the colony by year under random harvest. B. Numbers of simulated animals resident in the colony under maximal harvest. Males represented by solid lines, females by broken lines.
time to wash out. Twenty simulation replicates were run, and their output records were averaged.
RESULTS
Figure 2A shows numbers of males and females alive per year under the random harvest strategy and indicates that we have been successful in maintaining a relatively constant population size once the population has stabilized.
Our experimental population was simulated under exactly the same conditions, except that maximal harvest probabilities were substituted for the random harvest rates. These rates also were adjusted so as to achieve zero growth. Figure 2B gives numbers of each sex alive by year under the maximal harvest strategy. It can be seen that we have succeeded in maintaining a stationary population here as well.
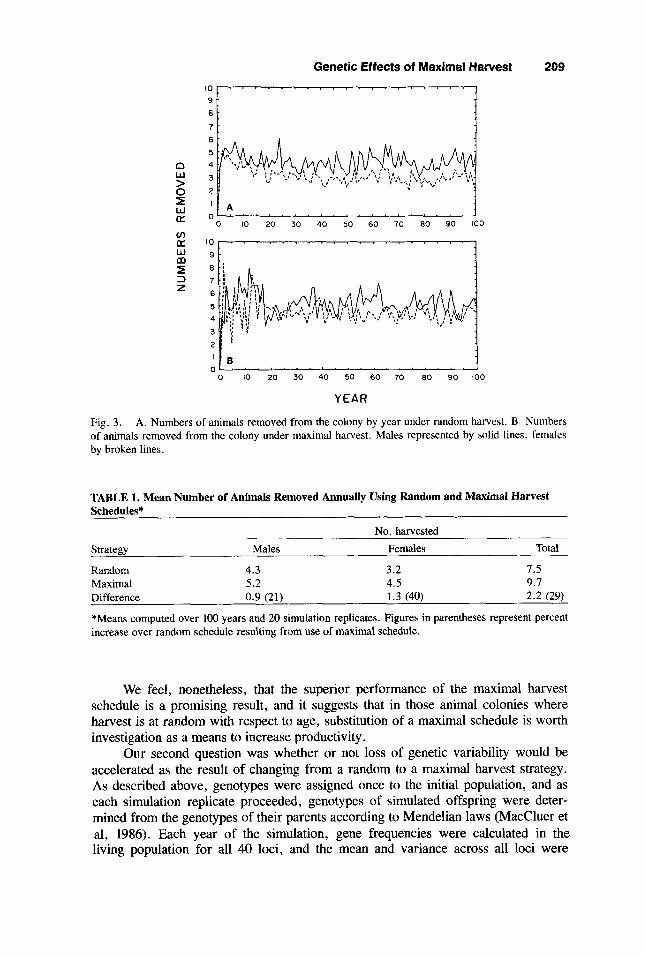
Our first question was whether or not the increase in reproductive efficiency predicted by the Beddington and Taylor model was a realistic estimate in a population that did not meet the assumption of large size. Figure 3 shows average yearly harvests of both sexes combined for each of the two strategies. Although there is considerable fluctuation because of the small numbers, maximal harvest is consistently better, producing on average about 30% more offspring than random harvest, as shown in Table 1 (somewhat less than predicted by the theoretical model). This result must be interpreted with some caution, since our simulations contain a number of simplifying assumptions, such as constant vital rates that apply uniformly to all population members.
Genetic Effects of Maximal Harvest 209
7
LL '0 10 20 30 40 50 60 70 80 90 100
' f B 0 0 10 20 30 40 50 60 70 80 90 100
YEAR
Fig. 3. A. Numbers of animals removed from the colony by year under random harvest. B. Numbers of animals removed from the colony under maximal harvest. Males represented by solid lines, females by broken lines.
TABLE 1. Mean Number of Animals Removed Annually Using Random and Maximal Harvest Schedules*
No. harvested Strategy Males Females Total
Random 4.3 3.2 1.5 Maximal 5.2 4.5 9.1 Difference 0.9 (21) 1.3 (40) 2.2 (29)
*Means computed over 100 years and 20 simulation replicates. Figures in parentheses represent percent increase over random schedule resulting from use of maximal schedule.
We feel, nonetheless, that the superior performance of the maximal harvest schedule is a promising result, and it suggests that in those animal colonies where harvest is at random with respect to age, substitution of a maximal schedule is worth investigation as a means to increase productivity.
Our second question was whether or not loss of genetic variability would be accelerated as the result of changing from a random to a maximal harvest strategy. As described above, genotypes were assigned once to the initial population, and as each simulation replicate proceeded, genotypes of simulated offspring were deter- mined from the genotypes of their parents according to Mendelian laws (MacCluer et al, 1986). Each year of the simulation, gene frequencies were calculated in the living population for all 40 loci, and the mean and variance across all loci were

210 Dyke, Gage, and Mamelka
351 . . . I . , . . . . . , . , , , , , . , 30
25
20
W u z [L 5 3 cn 0 10 20 30 40 50 60 70 80 90 100 3
0 -I 30
25
W 3 2o I- w l5
10
05
" 0 10 20 30 40 50 60 70 80 90 100
Y E A R
A. Means of gene frequencies for 40 loci by year for each of 20 simulation replicates under Fig. 4. random harvest. B. Means of gene frequencies under maximal harvest.
tabulated. We expect that frequencies in individual replicates will drift away from their initial values as the generations pass, although the average across all replicates should not. The greater the tendency to dispersal, the greater the likelihood of loss of genetic variability. Loss of variability can be seen most dramatically in those loci where the frequency drifts to 0 or 1, meaning that an allele has been Iost altogether. The question of the relative risk of loss of variability due to our two harvest strategies can be answered then by determining which strategy exhibits the greatest dispersal of gene frequencies. The results of this test can be seen in Figure 4A and 4B, where the mean of 40 gene frequencies for each of ten replicates is plotted for 100 years of simulation. Figure 4A shows results for random harvest and Figure 4B results for maximal harvest. The gradual drift away from the initial mean gene frequency of 0.48 over time is apparent in each set of curves. It is somewhat difficult to compare the two strategies visually with respect to the dispersal of gene frequencies, but the major difference between the two sets of curves appears to occur at about year 20, in which the spread of gene frequencies is somewhat greater in the populations under the maximal harvest regime. This increase is due for the most part to a temporary decline in numbers, particularly of males, during the time that the population is in transition between its initial composition and the ultimate structure that results from the age- and sex-specific pattern of removals according to the maximal schedule (see Fig. 1). As time passes, however, the differences between the two sets of curves become less obvious.
A more appropriate measure of dispersal is the between-locus variance of gene frequencies measured from the initial mean of 0.48 which is directly proportional to the risk of loss of genetic variability. Between-locus variance is shown by year for
Genetic Effects of Maximal Harvest 211
1 !$! 6 10 20 30 40 50 60 70 80 90 100
z a 56
I . . . . . . , , I . , . , . , . , > / I 0 10 20 30 40 50 60 70 80 90 100
YEAR
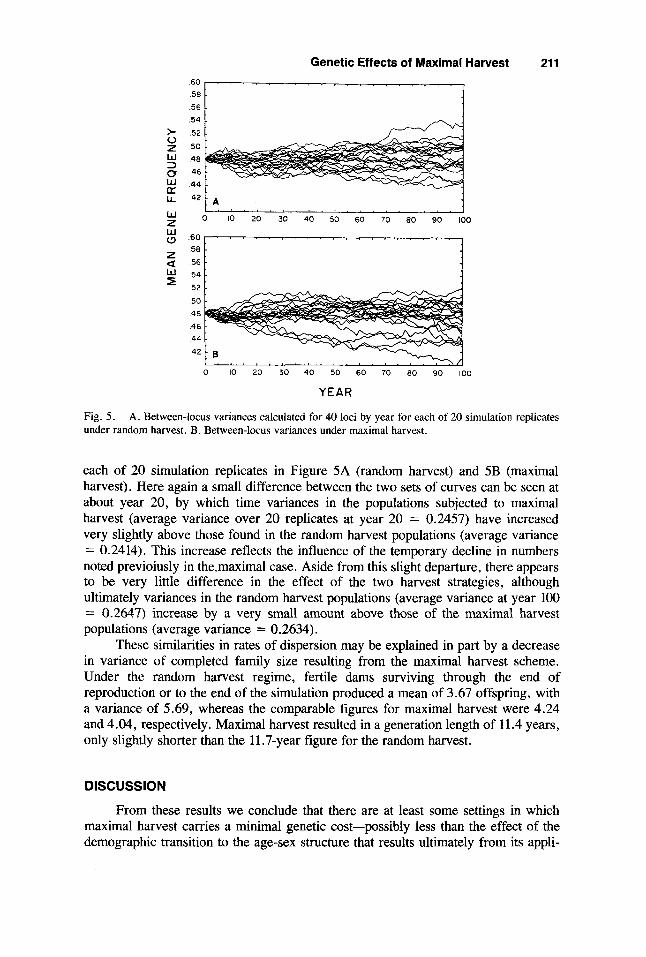
Fig. 5 . under random harvest. B. Between-locus variances under maximal harvest.
A. Between-locus variances catculated for 40 loci by year for each of 20 simulation replicates
each of 20 simulation replicates in Figure 5A (random harvest) and 5B (maximal harvest). Here again a small difference between the two sets of curves can be seen at about year 20, by which time variances in the populations subjected to maximal harvest (average variance over 20 replicates at year 20 = 0.2457) have increased very slightly above those found in the random harvest populations (average variance = 0.2414). This increase reflects the influence of the temporary decline in numbers noted previoiusly in the.maximal case. Aside from this slight departure, there appears to be very little difference in the effect of the two harvest strategies, although ultimately variances in the random harvest populations (average variance at year 100 = 0.2647) increase by a very small amount above those of the maximal harvest populations (average variance = 0.2634).
These similarities in rates of dispersion may be explained in part by a decrease in variance of completed fardy size resulting from the maximal harvest scheme. Under the random harvest regime, fertile dams surviving through the end of reproduction or to the end of the simulation produced a mean of 3.67 offspring, with a variance of 5.69, whereas the comparable figures for maximal harvest were 4.24 and 4.04, respectively. Maximal harvest resulted in a generation length of 11.4 years, only slightly shorter than the 11.7-year figure for the random harvest.
DISCUSSION
From these results we conclude that there are at least some settings in which maximal harvest carries a minimal genetic cost-possibly less than the effect of the demographic transition to the age-sex structure that results ultimately from its appli-

212 Dyke, Gage, and Mamelka
cation. Presumably, initiation of a maximal harvest schedule could be adjusted in such a way as to reduce some of the effects of this transition as well.
Ryman et a1 [1981] used simulation in a study of potential loss of genetic variability resulting from various age- and sex-dependent strategies for hunting moose; they found substantial differences between strategies. The contrast between their results and ours is probably due to the broad range of their hunting patterns, which (aside from a random harvest scheme) produced age structures quite unlike that resulting from our maximal harvest. In particular, their harvest strategies resulted in marked differences in generation length, whereas ours did not. There is some uncertainty in the results of the Ryman study, however, owing to their simulation methods and the way they have measured genetic variability. By simulating the reproduction of very large numbers of females alone and then performing their computations on a fixed number of dams, they have ignored any effects of mating structure or temporal fluctuations in population size on genetic drift. Since the magnitude of these effects is not known for any of the hunting strategies (in either simulated or real populations), evaluation of their results is somewhat difficult. The estimate of effective population number which they used is an approximation largely dependent upon demographic structure and tends to underestimate the effects of mating structure and temporal fluctuations of population size. We prefer our method of incorporating genetic transmission in our simulations because it permits a more direct estimate of genetic variability (the between-locus variance) that avoids these difficulties.
It should be pointed out that even if maximal harvest has little or no genetic effect, the problem of loss of genetic variability may remain a serious one. We have not addressed here the problems of small size, isolation, skewed sex ratio, or inbreeding, all of which affect genetic structure regardless of harvest strategy. None- theless, it is encouraging to have evidence that increased reproductive efficiency need not add significantly to the problems of genetic management. Although adoption of a maximal harvest schedule requires a careful prior analysis, there is no reason to reject it out of hand on genetic grounds.
CONCLUSIONS
1. A maximal harvest strategy based upon adjustment of demographc structure may be an effective way to increase the overall reproductive efficiency of small captive breeding colonies.
2. Under demographic conditions approximating those described here, maximal harvest per se appears to have little effect on the rate of loss of genetic variability.
ACKNOWLEDGMENTS
This work is a continuation of a project begun in 1980 by the late Dr. Martin Curie-Cohen at the University of Wisconsin. Demographic and pedigree data from the WRPRC rhesus colony were organized and verified by Dr. Curie-Cohen and his associates, Charles E. Kahn, Jr., Lesleigh Luttrell, and Gail Beyer, without whose efforts this analysis would not have been possible. This work was supported by NIH Grant RR 01216. Computer facilities were supplied by a grant from the Robert J. Kleberg, Jr. and Helen C. Kleberg Foundation.

Genetic Effects of Maximal Harvest 213
APPENDIX ColonySim
CoZonySim is a stochastic discrete-time microsimulation designed for experi- mentation with the demographic and genetic structures of captive primate colonies. The simulation consists of five FORTRAN 77 program modules, making it conve- niently exportable to any general-purpose computer having a minimum of 1 megabyte of high-speed memory available to the user. The five programs are:
1. Program GetSpecs sets input parameters User and experiment identifiers Initial seed for random number generator Number of breeding colonies (1-8) Initial and maximum sizes of breeding groups (by sex) Transfer (migration) between colonies Simulation cycle length (“years, ” “months, ” etc) Duration of simulation (in cycles) Sex ratio at birth Mating rules (including social and dominance hierarchies) Initial gene frequencies in each colony for up to 40 loci with specifiable linkage relationships
2. Program LifeTubZ derives age-specific fertility, mortality, harvest, and trans- fer rates for each colony by
a. calculating them from an existing animal records file (such as the
b. reading predefined rates from a file designated by the user. 3 . Program GetCoZony assembles a starting population for each colony by
a. specifying a group of animals from an existing animal records file, maintaining actual kinship, demographic, and genotypic charcteristics,
b. randomly selecting an arbitrary number of animals from an animal records file, maintaining demographic characteristics only, or
c. creating an entirely artificial group, based on arbitrary kinship, demo- graphic, and genotypic parameters.
4. Program AgeCoZony projects the initial colony or colonies forward in time, subjecting each individual to probabilities of death, harvest, transfer, and giving birth as specified in LifeTubZ. Genotypes of newborn individuals are assigned according to genotypes of their parents and the Mendelian laws. The process can be halted at predefined points for evaluation of colony status and then restarted at a later time.
5. Program ListCoZony generates and displays demographic and genetic statistics in graphic and tabular form at specified intervals.
WRPRC rhesus data base), or
REFERENCES
Beddington, J.R.; Taylor, D.B. Optimal age spe- NAL OF PRIMATOLOGY 10:257-269, 1986. cific harvesting of a population. BIOMETRICS Foose, T.J.; Lande, R.; Flesness, N.R.; Rabb, G.; 29~801-809, 1973. Read, B. Propagation plans. ZOO BIOLOGY
R.W.; Stone; W.H. A demographic analysis of Goodman, D. Demographic intervention for the Wisconsin Regional Primate Research Center closely managed populations, pp. 171-195 in Rhesus colony, 1963-1982. AMERICAN JOUR- CONSERVATION BIOLOGY. M. SoulC; B.A.
Dyke, B.; Gage, T.B.; Mamelka, P.M.; Goy, 5:139-146, 1986.

214 Dyke, Gage, and Mamelka
Wilcox, eds. Sunderland, Massachusetts, Sinauer Associates, 1980.
MacCluer, J.M.; VandeBerg, J.L.; Read B.; Ry- der, O.A.; Pedigree analysis by computer simu-
Ryman, N.; Baccus, R.; Reuterwall, C.; Smith, M.H. Effective population size, generation inter- Val, and potential loss of genetic variability in game species under different hunting regimes.
lation. ZOO BIOLOGY 5:147-160, 1986. OIKOS 36:257-266, 1981.




















