
Scattering of Electromagnetic Waves from Two-Dimensional Randomly Rough Penetrable Surfaces

Genetic characterization of populations of the
ectoparasitic caligid, Lepeophtheirus salmonis
(Krøyer 1837) using randomly amplified
polymorphic DNA
BryonyA Dixon, Andrew P Shinn & Christina SommervilleInstitute of Aquaculture, University of Stirling, Stirling, Scotland, UK
Correspondence: A P Shinn, Parasitology, Institute of Aquaculture, University of Stirling, Stirling, FK9 4LA, Scotland, UK. E-mail:
Abstract
Genetic variability within salmon louse, Le-peophtheirus salmonis (Caligidae: Copepoda), popula-tions parasitizing farmed and wild Scottish Atlanticsalmon (Salmo salar) was investigated using analysisof randomly ampli¢ed polymorphic DNA (RAPD)fragments. Seven individual decamer primers wereused to analyse samples of salmon lice collected from15 di¡erent locations in Scotland. The polymerasechain reaction products were separated usingagarose gel electrophoresis and the resulting bandpatterns were analysed using a semi-automated ana-lytical scoring system. Dendrograms were producedusing the unweighted pair-group average (UPGMA)method using Dice similarity values. The summarydendrogram of the analysis of all RAPD bandsshowed two separate clusters of salmon lice, the lar-ger being sub-divided into a further two sections.Thecollections of lice occupying each of these sub-divi-sions, however, were a mix of sites, which did not ex-hibit a structured geographical pattern.
Keywords: Lepeophtheirus salmonis, sea lice, cope-pod parasite, RAPDs, Salmo salar
Introduction
Sea lice (Lepeophtheirus salmonis (Kr�yer 1837) andCaligus elongatus v. Nordmann 1832) are responsiblefor serious mortalities and the loss of production inAtlantic salmon, Salmo salar L., farming in Scotland.These parasites also a¡ect wild salmonids and a100% prevalence of lice have been documented forwild Atlantic salmon caught in marine coastal
waters o¡ Scotland (Todd,Walker, Hoyle, Northcott,Walker & Ritchie 2000). Previous studies have inves-tigated the allelic frequencies of polymorphic en-zymes (allozymes) of populations of the salmonlouse, L. salmonis, and have shown varying degreesof polymorphism. Isdal, Nylund and N�vdal (1997)demonstrated two distinct populations of salmon licein Norway based on allozyme data determined fromspecimens collected from six di¡erent localities onthe Norwegian coast. Todd,Walker,Wol¡, Northcott,Walker, Ritchie, Hoskins, Abbott and Hazon (1997)used both allozymes and randomly ampli¢ed poly-morphic DNA (RAPD) analysis to investigate salmonlice variation at the population level from both wildand farmed Atlantic salmon and from sea trout(Salmo trutta L.) in Scotland. The allozyme resultsshowed an overall pattern of genetic homogeneity,although data were obtained for only two poly-morphic loci. In addition, they found genetic homo-geneity between samples fromwild salmon and fromfarmed salmon although the two sets of data used inthe comparison were from the east and west coastsrespectively. However, Todd et al. (1997) found that,unlike their allozyme results, which implied genetichomogeneity, the RAPD analysis showed signi¢-cantly high levels of genetic di¡erentiation. Of thesites examined, the authors found four farmed sal-monid sites to be genetically distinct from one an-other. Furthermore, these were also di¡erent fromthose samples of L. salmonis taken fromwild salmonand sea trout suggesting ‘putative’ farm markers foreach farm site examined. Lepeophtheirus salmonissampled from the wild salmon and sea trout showeda pattern of genetic homogeneity, although the
Aquaculture Research, 2004, 35, 730^741 doi:10.1111/j.1365-2109.2004.01095.x
730 r 2004 Blackwell Publishing Ltd

authors suggested that the use of one of the primersmight permit the possibility of further elucidationbetween the two groups. The hypotheses of Toddet al. (1997) were that a combination of founder e¡ectand strong selection pressures, such as chemothera-peutant use, produced the RAPD phenotypes ob-served in the farmed salmon samples. The currentpaper attempts to further evaluate the usefulness ofthe primers described byTodd et al. (1997) by expand-ing the study to encompass salmon louse samplesfrom a much greater geographical area from bothwild and farmed salmon in Scotland.
Materials and methods
Two experiments are described both using the sameprimers as published byTodd et al. (1997). Experiment1 used starved and unstarved L. salmonis collectedfrom farmed S. salar at a single site (Loch Linnhe)(Fig. 1). The purpose of this was to evaluate the need
for starving salmon lice before processing, thus elim-inating host products from the gut that may produceadditional RAPD-polymerase chain reaction (RAPD-PCR) bands. Experiment 2 was a larger study en-compassing L. salmonis samples collected from 15di¡erent locations (Fig.1) from both farmed and wildsalmon from the north, east and west coasts of Scot-land. In Experiment 2, all salmon lice were starvedfor 48 h prior to DNA extraction and PCR analysis toeliminate host products from the gut.
Sample collection and processing
Salmon lice (usually 50^300) were collected fromap-proximately100 to 200 farmed S. salar at commercialharvests from 12 di¡erent multipen sites (see Fig. 1)on the west coast of Scotland and transported tothe laboratory in fresh aerated seawater. Salmon lice(usually 20^60) were also collected from multiplewild salmon using seine net at three sites (LochDuich, River North Esk and the River South Esk)(Fig. 1). The collection dates and conditions of
Loch Eriboll
Loch Laxford
Skye
Uist
Loch Duich Wd
Loch Carron
North Esk WdSouth Esk Wd
Arran
Loch Fyne
Loch MelfortShuna
MullLoch Linnhe
Loch Sunart
Figure 1 Map of Lepeophtheirus sal-monis collection sites in Scotland forrandomly ampli¢ed polymorphic DNAanalysis (Experiments 1 and 2). Sam-ples with the su⁄x ‘Wd’ and shownas triangles were collected from wildAtlantic salmon, whilst all other sitesshown as circles represent salmonlouse collections from farmed Atlanticsalmon.
Aquaculture Research, 2004, 35, 730^741 RAPD analysis of L. salmonis populations BA Dixon et al.
r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741 731

sampling for all samples are given inTable1. Salmonlice were maintained in fresh sea water (33%, 10 1C)with aeration prior to use.In Experiment 1, ¢ve individual, randomly se-
lected, adult female L. salmonis were processed im-mediately upon return to the laboratory, as detailedbelow. These females had red guts suggesting thatthey had taken a recent blood meal and, therefore,were assumed to be unstarved. The remaining speci-mens of adult female L. salmonis were starved for48 h by maintaining them in aerated fresh sea water(33%, 10 1C) before a further ¢ve, randomly selected,adult females were removed for processing. Female L.salmonis were chosen because their larger size en-abled complete homogenization and a more e⁄cientDNA extraction. In Experiment 2, all adult female L.salmonis from all sites were kept for a minimum of48-h starvationbefore processing to permit themeta-bolism, assimilation and excretion of host productsfrom the gut (O. Andrade-Salas, pers. comm.). Indivi-dual salmon lice were then processed by maceratingin approximately 30 mL of homogenization bu¡er(1-mM Tris-HCl, pH 7.0, 1-mM EDTA, 0.1-mM 2-mer-captoethanol) using a pellet pestle (Anachem, Luton,UK). Samples were then frozen in liquid nitrogen andstored at �70 1C until required.
DNA extraction
Genomic DNA was extracted from frozen, homoge-nized salmon louse tissue (circa 20mg) using thePuregeneTM DNA extraction kit (Gentra Systems,
Minneapolis, MN, USA) according to the manufac-turer’s instructions. The puri¢ed DNA pellets wereair dried at room temperature overnight and re-sus-pended in 50-mL TE bu¡er (0.1-mM EDTA, 10-mMTris-HCl, pH 8.0). The DNA concentration, purityand quality (A260/A280) were determined using spec-trophotometry and samples were stored at 4 1C untilneeded.
Polymerase chain reaction
Each PCR included one of the following sevenprimers (Operon, Alameda, CA, USA): OPH-01 5 0-GGTCGGAGAA-3 0, OPH-02 5 0-TCGGACGTGA-3 0,OPH-11 5 0-CTTCCGCAGT-3 0, OPH-14 5 0-ACCAGGT-TGG-3 0, OPH-15 5 0-AATGGCGCAG-3 0, OPH-18 5 0-GAATCGGCCA-3 0, OPA-02 5 0-TGCCGAGCTG-3 0. Allindividuals for all samples within each primer setwere ampli¢ed in the same PCR run. Although thisrestricted the number of individuals available foreach sample site, it ensured that all comparisons be-tween sample sites for each primer were under mini-mal in£uence fromvariation between PCR assays.Each PCR tube contained a Ready-To-GoTM RAPD
analysis bead (Amersham Pharmacia Biotech UK,Little Chalfont, UK) containing thermostable poly-merases AmpliTAQR and Sto¡el fragment, 0.4mM ofeach dNTP, 2.5-mg bovine serum albumin (BSA) , 3-mM MgCl2, 30-mM KCl and10-mM Tris, pH 8.3. ThePCR reaction also contained 1 mL of genomic DNA(50^100 ng), 1 mL of primer (25 pmol) and 23 mL ofsterile distilled water (SDW). Ampli¢cation was
Table 1 Details of the Lepeophtheirus salmonis samples collected and analysed within Experiment 2
Date sampled Nameof site Conditions No. collected Treatment history prior to L. salmonis collection
March 1995 Loch Carron Farm harvest 175 Never treated
April 1995 Loch Fyne Farm harvest 288 Aquagard June 1994; H2O2 September 1994
April 1995 Isle of Shuna Farm harvest 220 H2O2 July, September, November 1994
April 1995 Loch Linnhe Farm harvest 205 Never treated (high freshwater influx)
May 1995 Loch Laxford Farm harvest 180 No details available
May 1995 Loch Eriboll Farm harvest 120 H2O2 April 1995
May 1995 Loch Melfort Farm harvest 110 No details available
May 1995 Isle of Arran Farm harvest 175 Aquagard April 1994
June 1995 Isle of Mull Farm harvest 125 No details available
June 1995 Loch Duich Wild, net caught 55 –
July 1995 River North Esk Wild, net caught 82 –
July 1995 Isle of Skye Farm harvest 130 Three dichlorvos treatments after 1993
July 1995 Loch Sunart Farm harvest 60 No details available
July 1995 River South Esk Wild, net caught 20 –
August 1995 Isle of Uist Farm harvest 44 No details available
For each population, four starved, adult female lice were selected randomly from a population of between 20 and 300 individuals andprocessed for analysis (H2O2, hydrogen peroxide).
RAPD analysis of L. salmonis populations BA Dixon et al. Aquaculture Research, 2004, 35, 730^741
732 r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741

carried out using Amersham Pharmacia Biotech’sReady-To-GoTM RAPD analysis beads and followingthe recommended protocol of an initial denaturationfor 5min at 95 1C followed by 45 cycles of 1min at94 1C, 1min at 36 1C and 2min at 72 1C, followed bya ¢nal hold at 25 1C.
Band visualization
Ampli¢ed RAPD products were separated on 1.0%agarose gels (1 � TBE) containing ethidium bromide(0.5 mgmL�1) and photographed on an ultraviolettransilluminator. One microlitre of a low-molecular-weight DNA marker (1-kb DNA ladder molecularweight marker#G5711, Promega UK, Southampton,UK) was loaded as a means of assessing the size ofampli¢ed products. Samples from ¢ve populationswere loaded per gel; each population sample consist-ing of four randomly selected individuals. Negativecontrols (DNA absent) were run at random through-out the study. The reproducibility of RAPD band pro-¢les was tested by re-analysing 25 individuals (from¢ve di¡erent sampling sites) and comparing both pro-¢les as scored using the GelDoc ID advanced soft-ware. Only clearly resolved RAPD-PCR pro¢les wereconsidered.
Statistical analysis
All band fragments were analysed using an auto-mated system (GelDoc 1D Advanced, 1997) (PhoretixInternational, Newcastle upon Tyne, UK) scoredusing a presence/absence system (P/A5 1/0). Simi-larity matrices were constructed using the Dice simi-larity coe⁄cient (Dice 1945) calculated as thenumber of shared bands divided by the total numberof bands. This coe⁄cient is identical to the similaritycoe⁄cient of Nei and Li (1979). Dendrograms for eachprimer were produced based on band sharing usingthe unweighted pair-group average (UPGMA) matrixas the amalgamation (linkage) rule using the soft-ware package StatisticaTM (ver.5.5, StatSoft1999;Tul-sa, OK, USA). A summary dendrogramwas producedfor Experiment 2 using StatisticaTM by taking anaverage of band-sharing values over the seven pri-mers using the same methods as for the individualprimer dendrograms.The potential e¡ect of an indivi-dual primer set having a disproportionately large biason the ¢nal result was eliminated by investigating thesystematic removal of each primer set from the sum-mary table of coe⁄cients. The data for Experiment 2,
for each individual primer set and data for all primersets combined, were then used in Arlequin ver 2.000(Schneider, Roessli & Exco⁄er 2000) to runananaly-sis of molecular variance (AMOVA) (Exco⁄er, Smouse& Quattro1992).
Results
Experiment1
The dendrograms from Experiment 1 did not show aclear division between starved and unstarved sam-ples when the band fragments were analysed. Highvariability in the RAPD band patterns between indi-vidual lice meant a ¢rm conclusion could not bedrawn regarding PCR ampli¢cation of host-derivedDNA fragments but the recommendation for starvingsalmon lice prior to analysis was upheld.
Experiment 2
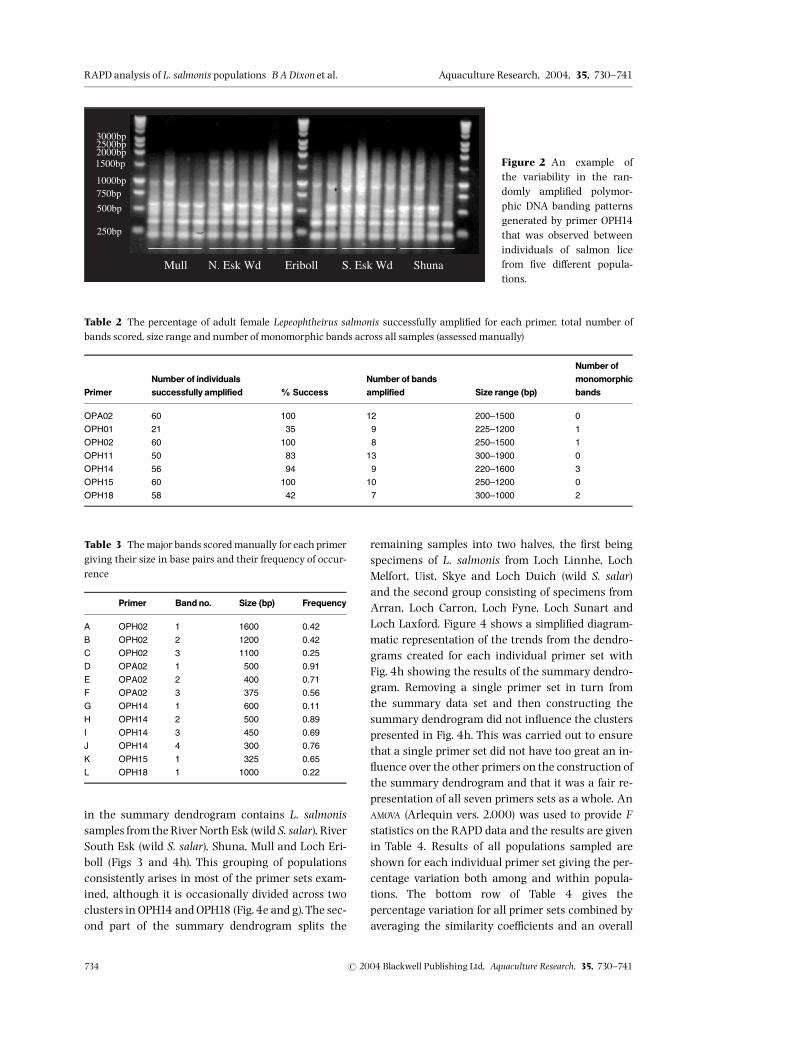
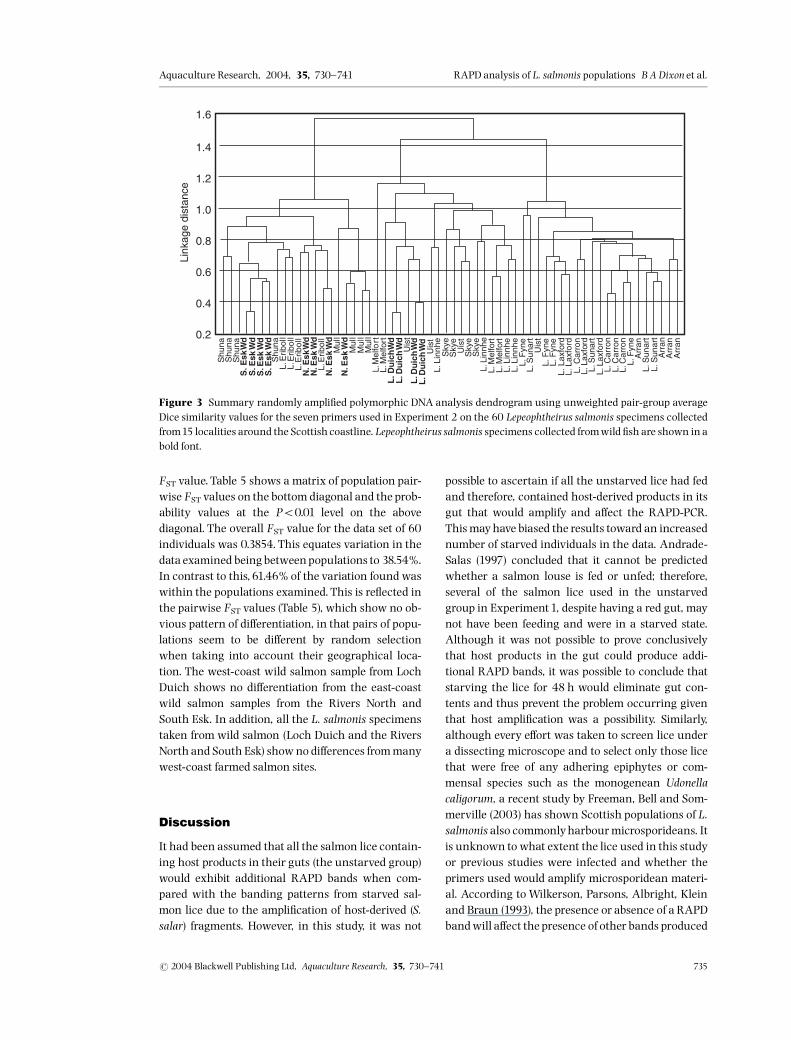
An example of the variability observed in the RAPDbanding patterns between the di¡erent populationsof lice is shown for one primer, primer OPH14, in Fig.2. Details of sample ampli¢cation including percent-age success and number and size range of band frag-ments per primer, estimated manually, are given inTable 2 along with band size and allele frequenciesof 12 polymorphic bands scored by eye in Table 3.Cluster analysis using dendrograms of these 12manually scored polymorphic bands showed thesame patterns as created when scoring all bandsthrough the automated system and is thus coveredin the discussion for this analysis. Assuming thateach RAPD band represents a single diallelic locusin Hardy^Weinberg (H^W) equilibrium, the pres-ence/absence data set can be converted into allelefrequencies (e.g. Apostol, Black, Reiter & Miller 1996;Lu & Rank1996). However, because RAPDs behave asdominantmarkers (i.e.15AAorAa and 05 aa), allelefrequency estimates are less accurate than with co-dominant markers (Lynch & Milligan 1994) and dueto the small sample sizes considered in this study, es-timates of allelic frequencies per population samplewere not calculated.Figure 3 shows the summary dendrogram of all
seven primer sets combined for Experiment 2, whileFig.4a^g summarizes the distribution of lice for eachprimer used. Figure 4h provides a summary of thedistribution of lice consistently observed across themajority of primer sets. The ¢rst group to segregate
Aquaculture Research, 2004, 35, 730^741 RAPD analysis of L. salmonis populations BA Dixon et al.
r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741 733
in the summary dendrogram contains L. salmonissamples from the River North Esk (wild S. salar), RiverSouth Esk (wild S. salar), Shuna, Mull and Loch Eri-boll (Figs 3 and 4h). This grouping of populationsconsistently arises in most of the primer sets exam-ined, although it is occasionally divided across twoclusters in OPH14 and OPH18 (Fig.4e and g).The sec-ond part of the summary dendrogram splits the
remaining samples into two halves, the ¢rst beingspecimens of L. salmonis from Loch Linnhe, LochMelfort, Uist, Skye and Loch Duich (wild S. salar)and the second group consisting of specimens fromArran, Loch Carron, Loch Fyne, Loch Sunart andLoch Laxford. Figure 4 shows a simpli¢ed diagram-matic representation of the trends from the dendro-grams created for each individual primer set withFig. 4h showing the results of the summary dendro-gram. Removing a single primer set in turn fromthe summary data set and then constructing thesummary dendrogram did not in£uence the clusterspresented in Fig. 4h. This was carried out to ensurethat a single primer set did not have too great an in-£uence over the other primers on the construction ofthe summary dendrogram and that it was a fair re-presentation of all seven primers sets as a whole. AnAMOVA (Arlequin vers. 2.000) was used to provide Fstatistics on the RAPD data and the results are givenin Table 4. Results of all populations sampled areshown for each individual primer set giving the per-centage variation both among and within popula-tions. The bottom row of Table 4 gives thepercentage variation for all primer sets combined byaveraging the similarity coe⁄cients and an overall
250bp
500bp
750bp1000bp
3000bp2500bp2000bp1500bp
Mull N. Esk Wd Eriboll S. Esk Wd Shuna
Figure 2 An example ofthe variability in the ran-domly ampli¢ed polymor-phic DNA banding patternsgenerated by primer OPH14that was observed betweenindividuals of salmon licefrom ¢ve di¡erent popula-tions.
Table 2 The percentage of adult female Lepeophtheirus salmonis successfully ampli¢ed for each primer, total number ofbands scored, size range and number of monomorphic bands across all samples (assessed manually)
PrimerNumber of individualssuccessfully amplified % Success
Number of bandsamplified Size range (bp)
Number ofmonomorphicbands
OPA02 60 100 12 200–1500 0
OPH01 21 35 9 225–1200 1
OPH02 60 100 8 250–1500 1
OPH11 50 83 13 300–1900 0
OPH14 56 94 9 220–1600 3
OPH15 60 100 10 250–1200 0
OPH18 58 42 7 300–1000 2
Table 3 The major bands scored manually for each primergiving their size in base pairs and their frequency of occur-rence
Primer Bandno. Size (bp) Frequency
A OPH02 1 1600 0.42
B OPH02 2 1200 0.42
C OPH02 3 1100 0.25
D OPA02 1 500 0.91
E OPA02 2 400 0.71
F OPA02 3 375 0.56
G OPH14 1 600 0.11
H OPH14 2 500 0.89
I OPH14 3 450 0.69
J OPH14 4 300 0.76
K OPH15 1 325 0.65
L OPH18 1 1000 0.22
RAPD analysis of L. salmonis populations BA Dixon et al. Aquaculture Research, 2004, 35, 730^741
734 r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741
FST value. Table 5 shows a matrix of population pair-wise FST values on the bottom diagonal and the prob-ability values at the Po0.01 level on the abovediagonal. The overall FST value for the data set of 60individuals was 0.3854. This equates variation in thedata examined being betweenpopulations to 38.54%.In contrast to this,61.46% of the variation found waswithin the populations examined. This is re£ected inthe pairwise FST values (Table 5), which show no ob-vious pattern of di¡erentiation, in that pairs of popu-lations seem to be di¡erent by random selectionwhen taking into account their geographical loca-tion. The west-coast wild salmon sample from LochDuich shows no di¡erentiation from the east-coastwild salmon samples from the Rivers North andSouth Esk. In addition, all the L. salmonis specimenstaken from wild salmon (Loch Duich and the RiversNorth and South Esk) showno di¡erences frommanywest-coast farmed salmon sites.
Discussion
It had been assumed that all the salmon lice contain-ing host products in their guts (the unstarved group)would exhibit additional RAPD bands when com-pared with the banding patterns from starved sal-mon lice due to the ampli¢cation of host-derived (S.salar) fragments. However, in this study, it was not
possible to ascertain if all the unstarved lice had fedand therefore, contained host-derived products in itsgut that would amplify and a¡ect the RAPD-PCR.This mayhave biased the results toward an increasednumber of starved individuals in the data. Andrade-Salas (1997) concluded that it cannot be predictedwhether a salmon louse is fed or unfed; therefore,several of the salmon lice used in the unstarvedgroup in Experiment1, despite having a red gut, maynot have been feeding and were in a starved state.Although it was not possible to prove conclusivelythat host products in the gut could produce addi-tional RAPD bands, it was possible to conclude thatstarving the lice for 48 h would eliminate gut con-tents and thus prevent the problem occurring giventhat host ampli¢cation was a possibility. Similarly,although every e¡ort was taken to screen lice undera dissecting microscope and to select only those licethat were free of any adhering epiphytes or com-mensal species such as the monogenean Udonellacaligorum, a recent study by Freeman, Bell and Som-merville (2003) has shown Scottish populations of L.salmonis also commonlyharbourmicrosporideans. Itis unknown to what extent the lice used in this studyor previous studies were infected and whether theprimers used would amplify microsporidean materi-al. According to Wilkerson, Parsons, Albright, Kleinand Braun (1993), the presence or absence of a RAPDbandwill a¡ect the presence of other bands produced
Shu
na
Shu
na
Shu
na
S. E
skW
dS
.Esk
Wd
S.E
skW
dS
.Esk
Wd
Shu
na
L. E
ribol
lL.
Erib
oll
L.E
ribol
lN
. Esk
Wd
N.E
skW
dL.
Erib
oll
N.E
skW
dM
ull
N.E
skW
dM
ull
Mul
lM
ull
L. M
elfo
r tL.
Mel
fort
L. D
uic
hWd
L.D
uic
hW
dU
ist
L.D
uic
hW
dL
.Du
ich
Wd
Uis
tL.
Lin
nhe
Sky
eS
kye
Uis
tS
kye
Sky
eL.
Linn
heL.
Mel
fort
L.M
elfo
rtL.
Linn
heL.
Linn
heL.
Fyn
eL.
Sun
art
Uis
tL.
Fyn
eL.
Fyn
eL.
Lax
ford
L. L
axfo
rdL.
Car
ron
L. L
axfo
rdL.
Sun
art
L.La
xfor
dL.
Car
ron
L.C
arro
nL.
Car
ron
L. F
yne
Arr
anL.
Sun
art
L. S
unar
tA
rran
Arr
anA
rran
Link
age
dist
ance
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Figure 3 Summary randomly ampli¢ed polymorphic DNA analysis dendrogram using unweighted pair-group averageDice similarity values for the seven primers used in Experiment 2 on the 60 Lepeophtheirus salmonis specimens collectedfrom15 localities around the Scottish coastline. Lepeophtheirus salmonis specimens collected fromwild ¢sh are shown inabold font.
Aquaculture Research, 2004, 35, 730^741 RAPD analysis of L. salmonis populations BA Dixon et al.
r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741 735

using RAPD-PCR because of competition for primerbinding sites. Thus, if gut host products or ectocom-mensals were present in or on L. salmonis and wereampli¢ed using RAPD-PCR, this may a¡ect thepresence of bands derived from salmon lice DNA. Tobe certain of the e¡ects of salmonid DNA on theRAPD-PCR, a further set of RAPD analyses wouldbe useful including salmonid DNA alone and alsosalmonid DNA mixed with louse DNA at varyingconcentrations. This could further evaluate the po-
tential of host DNA to amplify in RAPD-PCR. Thus,due to the competitive nature of RAPD-PCR, the si-tuation is far more complicated than simply lookingfor additional ‘host’-derived bands in the analysis.Given the uncertainty in concluding whether non-louse-derived products were present in RAPD-PCR,it was decided to starve all salmon lice for 48 hbefore processing to minimize the possibility of mis-interpreting RAPD bands through non-louse DNAinterference.
Loch Duich WdShunaSkye
Loch LaxfordLoch FyneLoch CarronArranLoch Linnhe
UistLoch MelfortNorth Esk Wd
Loch EribollSouth Esk Wd
Loch Sunart
Mull
Loch Linnhe
Loch Eriboll
UistSkye
ShunaMullS. Esk Wd
Loch FyneArranLoch Laxford
Loch Melfort
N. Esk Wd
Loch Duich WdSkye
Loch SunartLoch Duich Wd
Loch MelfortUist
South Esk Wd(Loch Eriboll)
Shuna
Arran(Loch Laxford)Loch Carron
Mull
Loch LinnheLoch Fyne
(Loch Laxford) N. Esk Wd(Loch Eriboll)
Loch Melfort
Loch Fyne (Loch Carron) (Loch Linnhe)
Loch Eriboll North Esk Wd
(South Esk Wd)
Arran
(d) OPH 11
(a) OPA 02 (b) OPH 01
(c) OPH 02
Figure 4 A summary of the populations of salmon lice that fall into each cluster within each unweighted pair-groupaverage dendrogram produced for each primer set (a^g) and from all pooled data (h). Only those sites that showed a ma-jority of individuals within a cluster are shown.
RAPD analysis of L. salmonis populations BA Dixon et al. Aquaculture Research, 2004, 35, 730^741
736 r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741

The individual dendrograms, summarized as dia-grams in Fig. 4a^g, produced for each primer set inExperiment 2 mirrored the broad categories de¢nedin the summary dendrogram (Fig. 3). To avoid repeti-tion, the general trends observed from the analysiswill be discussedwith reference to the summary den-drogram only. The summary dendrogram of the ana-lysis of all RAPD bands (Fig. 3) showed two separateclusters of salmon lice, the larger group itself beingsub-divided into two sections. The smaller groupconsisted of the east-coast populations of L. salmonisfrom the Rivers North and South Esk (wild S. salar)
along with salmon lice from Shuna, Mull and LochEriboll. It is clear, referring to the map of samplingsites (Fig.1), that there is no geographical connectionbetween all of these sites. However, it may be ex-pected that the north-coast samples from Loch Eri-boll show some similarity to the east-coast samplesgiven that a clockwise circulation pattern of currentsexists around Scotland (McManus 1992). A similartrend is also shown by Todd et al. (1997) from resultsof an initial RAPD experiment (‘RAPD markers, Ex-periment 1’). The authors found a north-/east-coastversus west-coast split between the populations but
MullShuna
(Loch Eriboll)S. Esk Wd
Loch SunartN. Esk Wd
(Loch Eriboll)(Loch Fyne)Uist
Loch Duich WdLoch Melfort Arran
Loch LinnheLoch LaxfordLoch Carron
(Skye)
Skye
ShunaLoch EribollN. Esk WdS. Esk WdMull
Uist
ArranLoch LinnheLoch LaxfordLoch FyneLoch Melfort
Loch Duich Wd
Loch Linnhe
Mull(N. Esk Wd)
Loch CarronArran
Loch Fyne
Loch Laxford
Loch Duich Wd
ShunaLoch EribollS. Esk Wd
UistSkye
Loch SunartLoch Melfort
Mull
ShunaS. Esk WdL. Eriboll
ArranLoch Fyne
Loch LaxfordL. Sunart
UistLoch Melfort Loch Linnhe
Skye
N. Esk Wd
Loch Duich Wd
(g) OPH 18
(e) OPH 14 (f) OPH 15
(h) All primers
Figure 4 (Continued).
Aquaculture Research, 2004, 35, 730^741 RAPD analysis of L. salmonis populations BA Dixon et al.
r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741 737

concluded that this e¡ect could be due to admixtureof thewild and farmed populations included. It seemsinexplicable why the samples from Mull and Shunapopulations should cluster together with thesenorth- and east-coast samples. Mull is a west-coastisland site and Shuna, although also an island, liesvery close to the mainland just south of Mull. Even ifa clockwise circulation pattern is assumed, there areseveral other sampling sites, for example Loch Su-nart, Skye, Uist and Loch Laxford, that lie betweenthe Mull/Shuna and the Loch Eriboll/River Eskgroup.The larger grouping of salmon lice populations
from the summary dendrogram (Fig. 3) containedthe majority of the west-coast farmed salmon sites
examined. This larger group was sub-divided intotwo.The ¢rst group being Loch Linnhe, LochMelfort,Uist, Skye and Loch DuichWild and the second groupconsisting of Arran, Loch Carron, Loch Fyne, LochSunart and Loch Laxford. Initially, these two clustersseem to have no apparent geographical basis. How-ever, it is interesting to note that Loch Duich (wild S.salar) clusters together with Uist and Skye, which arethe two most westerly island sites.Likelycandidates to produce genetic di¡erentiation
are low gene £ow between localities; di¡ering selec-tion pressure at each locality; local mutations andgenetic drift. All of these factors, along with localmigration of the host, may in£uence genetic struc-ture of the populations to varying degrees. Due tothe lack of any clear geographical pattern to the sal-mon lice di¡erentiation observed in this study, theresults seem to be a result of di¡ering selectionpressures on certain populations of L. salmonis. Thiscould be a result of environmental conditions such assalinity variation but could also be from farmedsalmon sites that areheavily treatedwith chemother-apeutants selecting for resistant populations of sal-mon lice. Di¡erences may also result from the levelof inbreeding between lice within a population, thesize of that population and the age of the populationat the time the samples were taken.For most marine invertebrates, the general as-
sumption is that having a planktonic larval stageconfers dispersal ability and thus high rates of gene£ow (Johnson & Black 1982; Hunt & Ayre 1989). An
Table 4 AMOVA of the RAPD data produced from Experi-ment 2 analysing15 populations of Lepeophtheirus salmoniscollected in Scotland
Primer
%Variationamongpopulations
% Variationwithin populations FST
OPA02 37.60 62.40 0.376
OPH01 53.12 46.88 0.531
OPH02 30.12 69.88 0.301
OPH11 33.19 66.81 0.332
OPH14 41.87 58.13 0.419
OPH15 45.71 54.29 0.457
OPH18 45.14 54.86 0.451
Total 38.54 61.46 0.385
Table 5 Matrix showing population pairwise FST (below diagonal) of RAPD phenotypes and the probability that theobserved di¡erence is di¡erent from a random distance (above diagonal) for all seven RAPD primer datasets combined
Population 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
1. Arran – 0.207 0.090 0.018 0.045 0.018 o0.01 o0.01 0.036 0.036 0.036 o0.01 o0.01 0.027 0.036
2. Loch Carron 0.035 – 0.018 o0.01 0.036 o0.01 o0.01 o0.01 o0.01 0.018 0.027 0.036 0.018 0.054 0.045
3. Loch Laxford 0.169 0.152 – 0.162 o0.01 0.027 o0.01 0.018 0.018 o0.01 0.027 0.036 0.054 o0.01 0.045
4. Loch Fyne 0.184 0.121 0.094 – o0.01 o0.01 0.045 0.018 o0.01 0.036 0.027 0.045 o0.01 0.036 0.027
5. Loch Linnhe 0.095 0.197 0.232 0.215 – o0.01 o0.01 0.027 0.018 o0.01 0.036 0.027 0.027 o0.01 0.027
6. Mull 0.436 0.451 0.530 0.425 0.519 – 0.054 0.018 o0.01 0.027 0.027 0.063 0.018 o0.01 0.036
7. North Esk wild 0.424 0.421 0.511 0.418 0.490 0.201 – 0.108 0.036 o0.01 0.054 o0.01 0.018 0.027 0.018
8. Loch Eriboll 0.437 0.415 0.479 0.385 0.508 0.338 0.072 – 0.036 0.036 o0.01 o0.01 0.036 0.045 0.018
9. South Esk wild 0.519 0.541 0.613 0.536 0.606 0.464 0.358 0.294 – 0.045 o0.01 0.027 0.045 0.027 0.036
10. Shuna 0.438 0.450 0.502 0.416 0.516 0.419 0.373 0.233 0.262 – 0.027 0.063 0.018 o0.01 o0.01
11. Loch Sunart 0.251 0.253 0.345 0.219 0.352 0.346 0.287 0.306 0.437 0.352 – 0.0252 o0.01 0.586 0.054
12. Skye 0.250 0.271 0.371 0.292 0.341 0.427 0.410 0.391 0.505 0.372 0.057 – o0.01 0.279 0.027
13. Loch Duich wild 0.546 0.578 0.639 0.561 0.614 0.600 0.580 0.605 0.680 0.594 0.404 0.345 – 0.036 o0.01
14. Uist 0.292 0.279 0.360 0.286 0.367 0.427 0.388 0.377 0.503 0.411 � 0.004 0.044 0.350 – 0.117
15. Loch Melfort 0.325 0.357 0.439 0.347 0.402 0.382 0.335 0.369 0.460 0.347 0.107 0.180 0.382 0.142 –
RAPD, randomly ampli¢ed polymorphic DNA.The ¢gures in bold relate to signi¢cant di¡erences.
RAPD analysis of L. salmonis populations BA Dixon et al. Aquaculture Research, 2004, 35, 730^741
738 r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741

analysis of genetic heterogeneity of local populationsis thought to give an indication of the extent of larvaldispersal and also an indirect measure of gene £ow(Todd, Lambert & Thorpe1998).The planktonic larvalstages of L. salmonis persist onaverage for10.4 days at10 1C (emergent nauplius I to newly moulted chali-mus I) (Johnson & Albright1991), but a small numberof individuals may remain infective for up to 30 daysat lower temperatures (Boxaspen, Asplin, Nilsen &Holst 2003). The situation with L. salmonis is furthercomplicated by a high host mobility. Sea trout andwild salmon have very di¡erent behavioural patternsupon entering the marine environment. Sea trouttend to remain in the coastal waters; thus, there isthe potential for cross transfer between wild andfarmed ¢sh in coastal regions (Whelan 1993). Wildsalmon are reported to move o¡shore and becomeoceanic quite quickly and do not remain in coastalareas where levels of salmon lice larvae might behigh.The host mobility and larval dispersal factors tend
to point towards a well-dispersed population. How-ever, the results from the present RAPD study showsome degree of population di¡erentiation over a largegeographic scale. The fact that the di¡erentiationdoes not seem to conform to any geographical pat-tern suggests that there may be selectionafter disper-sal has occurred. Selection pressure could a¡ectpopulation heterogeneity by at least two routes:¢rstly, by preferential recruitment to some popula-tions (for example, tolerance to low salinity in areaswith high freshwater land runo¡) or secondly,through the use of chemotherapeutants on farmedsites which build up resistant populations of salmonlice. Allele frequencies would be expected to changeif this gave a selective advantage for resistance tochemotherapeutants (Jones, Sommerville & Wootten1992). Continued treatment would preferentially se-lect for those salmon lice with resistance and with-out an external source of new recruits, new alleleswould quickly become established in the population.There are several published reports documenting thebuildup of resistance to several chemotherapeutants,including hydrogen peroxide, used in the salmonfarming industry (Jones et al. 1992; Treasurer,Wadsworth & Grant 2000).If this hypothesis held true, then the small cluster
represented in the dendrogram could potentially re-present unselected populations, i.e. Rivers Northand South Esk (wild S. salar), Loch Eriboll, Mull andShuna. The larger cluster of salmon lice from thesummary dendrogram could represent farmed sites
where a high selection pressure, such as the use ofchemotherapeutants at Loch Fyne or tolerance tofresh water at Loch Linnhe, was exerting itself. If so,the farms in questionwould be at Loch Carron, LochLaxford, Arran, Loch Fyne, Loch Melfort, Loch Su-nart, Loch Linnhe, Uist and Skye. It could be arguedthat this theory is invalidated by the sample LochDuich (wild S. salar) present in this latter group. How-ever, the samples from the Loch Duich (wild S. salar)population were collected from an area less than1km from a farmed salmon site in Loch Duich; if thissite were also under heavy selection pressures, thenthe wild salmon sample may be picking up selectedsalmon lice from the loch system. Several studies onmarine invertebrates (Hedgecock 1986; Todd et al.1998) show large-scale genetic homogeneity butsigni¢cant microgeographic heterogeneity. This hasbeen concluded to be due to selection post settlementrather than a genetic heterogeneity of recruits(Hedgecock 1986) in populations with high gene£ow and a similar pattern seems to be the case forL. salmonis.The use of AMOVA (Arlequin) in the current study
revealed an overall FST value of 0.385 indicating that38.5% of the overall variation occurs between popu-lations. All FST values for individual primer setsshowed very similar values ranging from 0.301(OPH01) to 0.531 (OPH02). The overall result of 0.385is comparable with the ¢gure of 0.362 found byTodd et al. (1997) and to that found for other marineinvertebrates, e.g. 0.462 for the asteroid Patiriella exi-gua (Lamarck 1815) (Hunt 1993) and 0.396 for theprosobranch gastropod Bembicium vittatum Philippi,1846 (Parsons 1996). This serves to re-iterate thefact that there is evident di¡erentiation observedin the current study despite a larval dispersal stage,high host movement and proposed high gene£ow but no observed correlation with geographicallocalities.There appears to be evidence in the present study
of some segregation in the salmon louse populationsthat occur in Scotland although it cannot bedescribed as a ‘farmed versus wild’di¡erentiation. Anumber of studies have been carried out usingRAPDs and allozyme analysis to assess di¡erentia-tion in populations of L. salmonis (Isdal et al. 1997;Todd et al.1997) and all ¢nd varying levels of di¡eren-tiation. It may be that the RAPD technique is simplytoo insensitive to detect delicate £uctuations in thepopulations under study and it should be kept inmind that there are a number of limitations in theRAPD technique.
Aquaculture Research, 2004, 35, 730^741 RAPD analysis of L. salmonis populations BA Dixon et al.
r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741 739

Randomly ampli¢ed polymorphic DNA band frag-ments are notoriously di⁄cult to reproduce exactlybetween laboratories (Penner, Bush,Wise, Kim, Dom-ier & Kasha 1993) due to the highly competitive na-ture of the RAPD-PCR brought about by the use ofshort primers and low annealing temperatures. Adi¡erence of just 1 1C in the annealing temperaturecan produce qualitatively di¡erent results in a RAPDanalysis. The thermocycle programme used by Toddet al. (1997), whichwas unavailable at the time of thepresent study, has notable di¡erences from the oneused in this study, the most important being a 4 1Cdi¡erence in annealing temperature. The thermo-cycle used in the present study was chosen becauseit was recommended byAmersham Pharmacia Bio-tech as the optimal one for use with Ready-To-GoTM
RAPDanalysis beads. Penner et al. (1993) suggest thata higher annealing temperature suchas the one usedby Todd et al. (1997) could result in a decrease in thetotal number of fragments which are ampli¢ed to be-gin with, leading to an increase in the competitive-ness of larger fragments. Alternatively, accessoryhost-ampli¢ed bands could be present if the salmonlice are not starved before processing. If completestarvation of salmon lice did not occur, it is feasiblethat contamination of the sample occurred from thepresence of host/¢sh DNAcarried over in the gut of L.salmonis.The current study used a smaller number ofindividuals per sample site but covered a much great-er sampling area, to ensure that every individualused in the study could be ampli¢ed by each RAPDprimer in the same PCR assay. It was felt that theadvantages of eliminating PCR run variation out-weighed the limitations of reducing the numbers ofindividuals per sampling site particularly as theprimers used were already thought to show di¡eren-tiation between sea lice from farmed salmonids sitesin Scotland. Sea lice were selected at random fromeach sample and the presence of small-scale hetero-geneity such as between individual pens was not in-vestigated. In addition, samples were collected over ashort time period (6 months) to try to limit seasonale¡ects. A further explanation for the lack of compar-able di¡erentiation found within this study with thatof Todd et al. (1997) may result from the di¡erent po-pulations of salmon lice that were sampled in eachstudy. Todd et al. (1997) analysed lice collected fromeight salmon localities, four wild (Rivers Dionard,Tay, South and North Esk) and four farm sites (Argyll,Sutherland, Uist andWester-ross). The current studyalso included lice collected fromwild salmon caughtin the Rivers South and North Esk and from farms in
the Argyll (Melfort, Shuna, Fyne and Arran),Wester-ross (Carron), Uist and Sutherland (Laxford and Eri-boll) districts, although it is not possible to ascertainif the same farms were sampled. Even if it is assumedthat the same sites were sampled, the current studypresents new data for a further nine salmon sites. Inaddition, the current study only used female L. sal-monis whilst the study of Todd et al. (1997) includedboth male and female L. salmonis in their study.Whilst restricting the current study to female sal-mon lice may have biased the RAPD fragments ob-tained, it was felt by the authors that reducingvariability among the samples used would providegreater discrimination on awider geographical basis.It is possible that if the salmon lice in the study by
Todd et al. (1997) were not starved before processingthat some of the more unique bands that presentedthemselves could be due to host product ampli¢ca-tion. In addition to the higher annealing temperatureand the use of both male and female L. salmonis, thismay explain why Todd et al. (1997) found speci¢cRAPDmarkers in their studywhere nonewere foundin the present study.
Acknowledgments
This work was supported by a BBSRC studentshipaward to Professor Sommerville and an industrialCASE award by Scottish Quality Salmon.The authorswould like to thank all the ¢sh farm companies vis-ited throughout the course of this study for their kindassistance in the collection of samples. In particular,thanks are due to Paul and Michael McCaskill forassisting with the collection of L. salmonis from wildsalmon.
References
Andrade-Salas O. (1997) Feeding and digestion in the ectopara-sitic copepod Lepeophtheirus salmonis (Kr�yer,1837). PhDthesis, University of Stirling, Stirling, UK,311pp.
Apostol B.L., BlackW.C., Reiter P. & Miller B.R. (1996) Popu-lation genetics with RAPD-PCR markers: the breedingstructure of Aedes aegypti in Puerto Rico. Heredity 76,325^334.
Boxaspen K., Asplin L., Nilsen F. & Holst J.C. (2003) Biolo-gical factors a¡ecting dispersal of salmon lice and regio-nal size of farm areas. Sealice 2003: Sixth InternationalConference on the Biology and Control of Sealice, St An-drews, Canada,1^4 July 2003. p.6.
Dice L.R. (1945) Measures of the amount of ecologic associa-tion between species. Ecology 26, 297^302.
RAPD analysis of L. salmonis populations BA Dixon et al. Aquaculture Research, 2004, 35, 730^741
740 r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741

Exco⁄er L., Smouse P.E. & Quattro J.M. (1992) Analysisof molecular variance inferred from metric distancesamong DNA haplotypes^application to human mito-chondrial DNA restriction data. Genetics131, 479^491.
Freeman M.A., Bell A.S. & Sommerville C. (2003) A hyper-parasitic microsporidean infecting the salmon louse,Lepeophtheirus salmonis: an rDNA-basedmolecular phylo-genetic study. Journal of Fish Diseases 26,667^676.
Hedgecock D. (1986) Is gene £ow from pelagic larval disper-sal important in the adaption and evolution of marine in-vertebrates? Bulletin of Marine Science 39,550^564.
Hunt A. (1993) E¡ects of contrasting patterns of larval dis-persal on the genetic connectedness of local populationsof two intertidal star¢sh, Patiriella calcar and P. exigua.Marine Ecology Progress Series 92,179^186.
Hunt A. & Ayre D.J. (1989) Population structure in the sexu-ally reproducing sea anemone Oulactis muscosa. MarineBiology102,537^544.
Isdal E., Nylund A. & N�vdal G. (1997) Genetic di¡erencesamong salmon lice from six Norwegian coastal sites: evid-ence from allozymes. Bulletin of the European Associationof Fish Pathology17,17^22.
Johnson M.S. & Black R. (1982) Chaotic genetic patchinessin an intertidal limpet, Siphonaria sp. Marine Biology 70,157^164.
Johnson S.C. & Albright L.J. (1991) Development, growth,and survival of Lepeophtheirus salmonis (Copepoda: Cali-gidae) under laboratory conditions. Journal of the MarineBiological Association of the United Kingdom 71, 425^436.
Jones M.W., Sommerville C. & Wootten R. (1992) Reducedsensitivity of the salmon louse, Lepeophtheirus salmonis,to the organophosphate dichlorvos. Journal of Fish Dis-eases15,197^202.
Lu R. & Rank G.H. (1996) Use of RAPD analyses to estimatepopulation genetic parameters in the alfalfa leaf-cuttingbee,Megachile rotundata. Genome 39,655^663.
Lynch M. & Milligan B.G. (1994) Analysis of population gen-etic structure with RAPD markers. Molecular Ecology 3,91^99.
McManus J. (1992) A hydrographic framework for marineconservation in Scotland. Proceedings of the Royal Societyof Edinburgh, Section B ^ Biological Sciences100,3^26.
Nei M. & Li W.H. (1979) Mathematical models for studyinggenetic variation in terms of restriction endonucleases.Proceedings of the National Academy of Sciences USA 76,5269^5273.
Parsons K.E. (1996) The genetic e¡ects of larval dispersaldepend upon spatial scale and habitat characteristics.Marine Biology126, 403^414.
Penner G., Bush A.,Wise R., KimW., Domier L. & Kasha K.(1993) Reproducibility of random ampli¢ed polymorph-ic DNA (RAPD). PCR Methods and Applications 2,341^345.
Schneider S., Roessli D. & Exco⁄er L. (2000) Arlequin ver.2.000: A Software for Population Genetic Data Analysis.Genetics and Biometry Laboratory, University of Geneva,Switzerland.
StatSoft (1999) STATISTICA forWindows [Computer ProgramManual]. StatSoft,Tulsa, OK, USA.
Todd C.D., Lambert W.J. & Thorpe J.P. (1998) The geneticstructure of intertidal populations of two species of nudi-branch molluscs with planktonic and pelagic lecitho-trophic larval stages: are pelagic larvae ‘for’ dispersal?Journal of Experimental Marine Biology and Ecology 228,1^28.
Todd C.D.,Walker A.M.,Wol¡ K., Northcott S.J.,Walker A.F.,Ritchie M.G., Hoskins R., Abbott R.J. & Hazon N. (1997)Genetic di¡erentiation of populations of the copepod sealouse Lepeophtheirus salmonis (Kr�yer) ectoparasitic onwild and farmed salmonids around the coasts of Scot-land: evidence from RAPDmarkers. Journal of Experimen-tal Marine Biology and Ecology 210, 251^274.
Todd C.D.,WalkerA.M., Hoyle J.E., Northcott S.J.,WalkerA.F.& Ritchie M.G. (2000) Infestations of wild adult Atlanticsalmon (Salmo salar L.) by the ectoparasitic copepod sealouse Lepeophtheirus salmonis Kr�yer: prevalence, inten-sity and the spatial distribution of males and females onthe host ¢sh. Hydrobiologia 429,118^196.
Treasurer J.W.,Wadsworth S. & Grant A. (2000) Resistanceof sea lice, Lepeophtheirus salmonis (Kr�yer), to hydrogenperoxide on farmed Atlantic salmon, Salmo salar L. Aqua-culture Research 31,855^860.
Whelan K.F. (1993) Decline of sea trout in the west ofIreland: an indication of forthcoming marine problemsfor salmon? In: Salmon in the Sea and New EnhancementStrategies (ed. by D. Mills), pp. 171^183. Fishing NewsBooks, Oxford, UK.
Wilkerson R.C., Parsons T.J., Albright D.G., Klein T.A. &Braun M.J. (1993) Random ampli¢ed polymorphic DNA(RAPD) markers readily distinguish cryptic mosquitospecies (Diptera; Culicidae: Anopheles). Insect MolecularBiology1, 205^211.
Aquaculture Research, 2004, 35, 730^741 RAPD analysis of L. salmonis populations BA Dixon et al.
r 2004 Blackwell Publishing Ltd, Aquaculture Research, 35, 730^741 741
Copyright © 2022 FDOKUMEN













![(DISSERTATION) THE NINETEENTH-CENTURY BRITISH PLANTATION SETTLEMENT AT LAMANAI, BELIZE (1837–1868) [pgs 101-111]](https://static.fdokumen.com/doc/165x107/6344dc63df19c083b107cac7/dissertation-the-nineteenth-century-british-plantation-settlement-at-lamanai.jpg)






