
Générer la forme en biologie: une caractéristique du processus développemental ?
21
Générer la forme en biologie, une caractéristique du processus développemental ? Antonine Nicoglou Labex “Who am I?”, CRPMS, “Evolution and development of Metazoans” IJM & IHPST Paris. La question qui consiste à se demander comment la forme est générée repose sur l’étude de la morphogenèse – du grec : morphê (μορφή) pour « forme » et genesis (γένεσις) pour « naissance, formation, production ». Le terme sert, depuis le XIX e siècle, à caractériser le processus qui, en biologie, permet d’expliquer le développement des structures caractéristiques des espèces vivantes. Pourtant, dès l’Antiquité (Aristote, De la génération des animaux), l’examen des structures visibles des êtres vivants constitue un objet d’étude privilégié pour le premier « philosophe- biologiste 1 » qui s’attache à décrire et à théoriser l’ontogenèse et l’organisation progressive de ces êtres vivants. Si Platon, quant à lui, entendait par le terme grec eidos (εἶδος) la forme d’une chose dans l’esprit et, considérant les idées comme des substances éternelles, situait la forme au-dessus de la matière, Aristote récuse une telle conception et place sur le même plan forme et matière. Plus précisément, il considère que la forme (μορφή : qui constitue, selon lui, l’aspect extérieur d’un objet) s’oppose à la matière. Dans un même temps, il considère que l’idée ou le mouvement – l’ eidos donc –, porté par le sperme du mâle, « in-forme » la matière inerte. Mais en distinguant deux modes de génération, le déploiement de structures déjà préexistantes du développement de novo, et en prenant partie pour la seconde thèse, il suggère alors la possibilité d’une structure (ou d’une forme) émergente à partir de la matière. En argumentant de la sorte, il est le premier philosophe à adopter un présupposé d’ordre « rationnel » quant à la forme : il ne se réfère ni à l’activité d’un démiurge, comme dans le Timée de Platon, ni à un quelconque récit mythique, comme chez les présocratiques, pour rendre compte de la genèse de la forme. Une telle démarche rationnelle quant à la genèse de la forme des êtres vivants ne réapparaîtra qu’à la fin du XVI e siècle à travers les travaux sur la génération du médecin et physiologiste William Harvey, Exercitationes de generatione animalium (1651). 1 Nous appliquons ici cette expression qui, bien qu’étant un anachronisme, permet de rendre compte d’une certaine filiation entre les problématiques anciennes et celles plus contemporaines
Transcript of Générer la forme en biologie: une caractéristique du processus développemental ?

Générer la forme en biologie, une caractéristique du processus développemental ?
Antonine Nicoglou Labex “Who am I?”, CRPMS, “Evolution and development of
Metazoans” IJM & IHPST Paris.
La question qui consiste à se demander comment la forme est générée repose sur
l’étude de la morphogenèse – du grec : morphê (µορφή) pour « forme » et genesis
(γένεσις) pour « naissance, formation, production ». Le terme sert, depuis le XIXe
siècle, à caractériser le processus qui, en biologie, permet d’expliquer le
développement des structures caractéristiques des espèces vivantes. Pourtant, dès
l’Antiquité (Aristote, De la génération des animaux), l’examen des structures visibles
des êtres vivants constitue un objet d’étude privilégié pour le premier « philosophe-
biologiste1 » qui s’attache à décrire et à théoriser l’ontogenèse et l’organisation
progressive de ces êtres vivants. Si Platon, quant à lui, entendait par le terme grec
eidos (εἶδος) la forme d’une chose dans l’esprit et, considérant les idées comme des
substances éternelles, situait la forme au-dessus de la matière, Aristote récuse une
telle conception et place sur le même plan forme et matière. Plus précisément, il
considère que la forme (µορφή : qui constitue, selon lui, l’aspect extérieur d’un objet)
s’oppose à la matière. Dans un même temps, il considère que l’idée ou le mouvement
– l’ eidos donc –, porté par le sperme du mâle, « in-forme » la matière inerte. Mais en
distinguant deux modes de génération, le déploiement de structures déjà
préexistantes du développement de novo, et en prenant partie pour la seconde
thèse, il suggère alors la possibilité d’une structure (ou d’une forme) émergente à
partir de la matière. En argumentant de la sorte, il est le premier philosophe à
adopter un présupposé d’ordre « rationnel » quant à la forme : il ne se réfère ni à
l’activité d’un démiurge, comme dans le Timée de Platon, ni à un quelconque
récit mythique, comme chez les présocratiques, pour rendre compte de la genèse
de la forme. Une telle démarche rationnelle quant à la genèse de la forme des êtres
vivants ne réapparaîtra qu’à la fin du XVIe siècle à travers les travaux sur la génération
du médecin et physiologiste William Harvey, Exercitationes de generatione
animalium (1651).
1 Nous appliquons ici cette expression qui, bien qu’étant un anachronisme, permet de rendre
compte d’une certaine filiation entre les problématiques anciennes et celles plus contemporaines

Ce n’est qu’à partir de la fin du XVIIIe siècle que les réflexions autour de la genèse
de la forme s’inscriront dans le cadre de l’embryologie, la discipline des sciences du
vivant qui jusqu’au XXe siècle s’attachera à décrire les transformations progressives
de l’œuf fécondé en organisme adulte. L’embryologie prend naissance dans un
contexte où l’affrontement entre les partisans d’une conception épigénétique de la
génération (le développement de la forme étant considéré comme se faisant
progressivement) et les partisans d’une conception préformationniste de la génération
(le développement de la forme étant considéré comme dépendant de structures d’ores
et déjà préétablies dès l’origine) est saillant. Ainsi, les questions philosophiques
associées au développement de la forme et qui avaient déjà été soulevées par Aristote
restent très présentes dans les premiers moments de la discipline scientifique de
l’embryologie : la forme doit-elle être considérée comme étant préétablie ou bien doit-
elle être conçue comme apparaissant progressivement ? Comment expliquer
l’apparition de formes particulières ? Comment expliquer que certaines formes soient
très conservées et récurrentes ?
À partir des années 1950, la discipline de l’embryologie se transforme
progressivement. Avec l’identification de la structure d’ADN, elle se rapproche de la
biologie moléculaire pour devenir « la biologie du développement ». Par ailleurs, le
début du XXe siècle ayant marqué non seulement l’avènement de la génétique mais
aussi la formulation par Charles Darwin de la théorie de l’évolution par le biais de la
sélection naturelle, de nouvelles questions viennent enrichir le champ des réflexions
quant à l’origine de la forme en biologie : L'information génétique renouvelle-t-elle la
forme aristotélicienne ? (Canguilhem, 1983) Quelle place adoptent les gènes dans la
détermination de la forme ? La forme est-elle principalement déterminée par
l’information génétique ? Quels autres facteurs peuvent entrer en jeu dans
l’établissement de la forme : l’environnement, le processus développemental lui-
même, etc. ? Quels impacts ces nouveaux questionnements peuvent-ils avoir quant à
notre compréhension des mécanismes de l’évolution ?
L’objet de ce chapitre est d’examiner les questions philosophiques liées à la
morphogenèse dans la biologie contemporaine. Ces questions ont été réélaborées par
la biologie évolutionnaire du développement (l’évo-dévo) – le champ disciplinaire
apparu dans les années 1970 qui entend rassembler les données fournies par la
biologie du développement et celles de l’évolution afin de proposer une nouvelle
synthèse de l’évolution (Pigliucci, 2007). Si, à partir des années 1940, la biologie

évolutionnaire s’était focalisée principalement sur les explications apportées par la
génétique des populations et par la génétique mendélienne, à partir des années 1970
certains biologistes du développement ont voulu déplacer la focale depuis les gènes
vers le développement. Toutefois, ce chapitre ne s'en tient pas là, et plutôt que
d’examiner les seules questions philosophiques relatives à l’articulation entre
évolution et développement, entre explication lointaine et explication prochaine
(Mayr, 1961) – comme c’est souvent le cas dans les manuels anglo-saxons de
philosophie de la biologie –, j’ai voulu montrer que certaines questions
contemporaines associées spécifiquement à la génération de la forme en biologie (et
parfois liées aux recherches en évo-dévo) s’articulaient avec des réflexions
philosophiques anciennes, voire immuables, relatives à l’ontogenèse.
Ces questions philosophiques seront abordées en se référant à des notions aujourd’hui
ou jadis familières des biologistes et que la philosophie de la biologie cherche, encore
et toujours, à éclairer davantage. Toutes ces notions permettent aux scientifiques de
caractériser les phénomènes qu’ils observent lorsqu’ils s’interrogent sur la genèse de
la forme : qu’il s’agisse de la « préformation », du « programme », du
« déterminisme », du « type »/ou de l’ « archétype », de la « robustesse », de la
« plasticité phénotypique »/ou de la « plasticité développementale », du
« développement » ou encore de la « croissance ».
Cinq questions principales constitueront les cinq parties de ce chapitre et me
permettront de définir et d’articuler ces différentes notions entre elles. Ces questions
sont : Qu’est ce que le « préformationnisme 2 » ? Y-a-t-il un ou plusieurs
« programme(s) » à l’origine de la forme en biologie ? La génération de la forme :
entre « robustesse » et « plasticité » ? Quel « développement » pour éclairer notre
compréhension de l’évolution de la forme ? Comment distinguer la génération de la
forme en biologie de la génération de la forme en physique ? Si les trois premières
parties porteront plus spécifiquement sur le problème du développement de la forme
en biologie, il en sera autrement des deux dernières. La quatrième partie me permettra
de rendre compte de la manière dont s’articulent les problèmes relatifs au
développement de la forme avec ceux relatifs à son évolution. La cinquième partie
soulèvera, quant à elle, un problème que l’on pourrait qualifier de « limitrophe », car
souvent traité de manière périphérique par les biologistes du développement depuis
2 Expression qui fait référence à l’ensemble des théories de la préformation depuis l’avènement de l’embryologie au XVIIIe siècle.

l’avènement de la génétique, mais qui reste néanmoins essentiel du point de vue de la
philosophie des sciences, puisqu’il s’agit du rapport entre les différentes sciences, et
plus particulièrement du rapport entre la physique et la biologie en ce qui concerne la
question de la forme. Après avoir présenté ces cinq questions relatives à la forme en
biologie, je reviendrai, en conclusion, sur la manière dont il faut penser le rapport qui
associe le processus développemental à la génération de la forme.
I. Qu’est ce que le « préformationnisme » ?
La notion de « préformationnisme » est couramment employée et connue des
embryologistes dès le XVIIe siècle. Le dévoilement de structures embryonnaires,
jusqu’alors invisibles à l’œil nu, dans un contexte où le microscope fait
progressivement son apparition, nécessite une réinterprétation des observations de
l’épigénéticien William Harvey (1578-1657) dans la ligne d’un proto-
préformationnisme dont Pierre Gassendi (1592-1655) se fait l’instigateur. Ce dernier
fournira le cadre épistémologique nécessaire à une telle conception en soutenant que
si toutes les parties essentielles de l’embryon se forment simultanément, elles
n’apparaissent que successivement dans le développement du fœtus. La caution
empirique de l’argumentation préformationniste ne surviendra, quant à elle, qu’à
partir de la publication de De formatione pulli in ovo de Malpighi (1628-1694) en
1672, lequel soutenait la thèse de la préformation d’une structure complexe
directement issue de la fécondation et sujette à déploiement et à agrandissement
progressif lors de l’incubation. Déjà avant la fin du XVIIe siècle et l’apparition des
premières conceptions proto-préformationnistes, certains savants pensaient que
l’embryon était déjà d’une certaine manière préformé dans le corps du parent avant la
fécondation. L’idée selon laquelle tous les embryons existaient d’ores et déjà, dès
l’origine du monde, apparaissait ainsi dans les écrits de Malebranche, Swammerdam,
Perrault et d’autres encore. Ces théories de la préexistence, fondées pour la plupart sur
le concept d’ « emboîtement », étaient apparues dans un contexte où les théories
épigénétiques du développement, qui avaient été proposées par Harvey et aussi par
Descartes, soulevaient des difficultés – comme celle, par exemple, de savoir comment
l’embryon parvenait à atteindre un tel niveau de complexité dans son organisation au
cours de son seul développement. Les théories de la préexistence offraient une
résolution de ce problème en argumentant que l’organisation était déjà « incorporée »,
« préprogrammée » par le Créateur dans l’œuf.

L’embryologie prend donc naissance dans un contexte théorique, où s’affrontent
encore deux conceptions divergentes de la génération. Mais à partir du XVIIIe siècle,
ce ne seront plus uniquement des présupposés métaphysiques différents qui
s’affronteront, mais également des observations divergentes du développement
exposées par des savants aux épistémologies différentes (Roe, 1981). D’un côté,
Albrecht von Haller (1708-1777), converti tardivement au préformationnisme,
observe un liquide visqueux dans l’œuf qui cache selon lui au regard l’existence de
structures déjà formées dans l’œuf non fécondé. Ces structures et ces formes se
déploieraient et s’offriraient ensuite progressivement au regard au cours du
développement. Haller propose une explication a posteriori de la génération de la
forme qui suit son observation. En effet, il avait été confronté à une insatisfaction
empirique avec la théorie épigénétique du développement, car elle nécessitait de faire
appel à des forces extérieures et « constructrices » pour expliquer le développement.
En se tournant vers le préformationnisme, il lui est possible de considérer que toutes
les forces à l’œuvre dans le processus développemental sont simples et mécaniques en
nature ; leur utilisation et orientation ne dépendant que du Créateur qui, à l’origine, a
construit tous les organes préformés de tous les embryons à venir et a, en plus,
organisé dans ces structures toutes les forces qui seront ensuite activées au cours du
développement. Haller explique donc les phénomènes qu’il observe en s’appuyant sur
des prérogatives empiriques, mécanistes3 et sur la religion. De l’autre côté, Caspar
Friedrich Wolff (1735-1794) considère que les processus vitaux doivent être
expliqués en s’appuyant principalement sur une approche rationaliste4. À l’inverse de
Haller, l’épigénéticien5 Wolff observe l’apparition et la modification progressive des
structures embryonnaires pendant le développement. Se référant à la fois à des
preuves observationnelles et à des forces mécaniques physiques pour appuyer ses
explications, celles-ci se situant toujours dans un schéma de raisonnement déductif
basé sur le principe de raison suffisante, il adopte davantage une approche a priori
dans son raisonnement explicatif. En considérant que la matière est conçue comme
3 L’approche mécaniste repose sur une conception matérialiste qui perçoit la plupart des
phénomènes suivant le modèle des liens de cause à effet. 4 Aux yeux du rationalisme, l’expérience sensible a elle seule ne saurait donner de
connaissance véritable. Il n’est possible d’accéder à la connaissance du réel qu'en vertu d'une explication par la raison déterminante, suffisante et nécessaire.
5 Il est également considéré comme le « père de l’embryologie », voir J. Needham dans son ouvrage, A History of Embryology, New York, Abelard-Schuman, 1959, p. 43. Voir également J. M. Cooper, “Metaphysics in Aristotle's Embryology,” in Proceedings of the Cambridge Philological Society, 214, 1988, p. 14–41.

quelque chose possédant des formes, des qualités, des modes et des attributs, il rejette
néanmoins les critiques de Haller, puisqu’il ne lui est pas nécessaire de se référer à
des forces constructrices pour expliquer le développement de la forme par épigenèse.
Jusqu’en 1760, la plupart des embryologistes adopteront la théorie de la préformation,
car il s’agit de la seule explication mécanique du développement qui soit en accord
avec la conception religieuse dominante du monde. Par ailleurs, chez la plupart des
penseurs préformationnistes du XVIIIe siècle, tout comme chez Haller, la matière est
conçue comme une entité passive et activée uniquement sous l’influence de forces
mécaniques simples imposées par Dieu. Le développement ne peut être que
programmé dans des structures préexistantes, car des forces mécaniques aveugles,
agissant sur une simple matière passive, ne peuvent, à elles seules, être responsables
de la production des organismes vivants.
Avec Johann Friedrich Blumenbach (1752-1840) et les autres embryologistes
allemands, la source de l’organisation embryonnaire ne fait plus partie des problèmes
à expliquer. L’organisation devient plutôt la seule chose que l’on considère comme
acquise. Influencés par la pensée kantienne, les embryologistes allemands embrassent
une vision téléologique du développement (l’organisation téléologique est comprise
comme concept régulateur). S’accordant avec Haller, Blumenbach et ses successeurs
rejettent l’idée selon laquelle l’organisation pourrait être expliquée en ne se référant
qu’à des forces mécaniques (par épigenèse). En acceptant l’organisation comme un
fait téléologique, il devient possible non seulement d’expliquer de manière mécanique
comment fonctionne et est maintenue cette organisation, mais aussi d’envisager des
investigations empiriques des différents mécanismes à l’origine des phénomènes
vitaux. Le courant de l’épigenèse germanique (qui diffère quelque peu de la
conception épigénétique de Wolff) prend alors naissance. Dès lors, il n’est plus
nécessaire de rendre compte du processus développemental dans son ensemble (de
répondre à la question « pourquoi y a-t-il développement ? »), pour comprendre la
génération de la forme (pour répondre à la question « comment se déroule le
développement ? »). La formation de l’individu devient une partie du processus
développemental qui inclut la totalité du domaine organique (Canguilhem, Lapassade,
Piquemal, Ulmann, 1962).
Cette brève généalogie explique pourquoi on fait encore référence dans les
discussions contemporaines sur la génération et le développement, à une potentielle
opposition entre une conception épigénétique et une conception préformationniste de

la génération (Maienschein, 1992). Cette question réapparaît dès lors que l’on
s’interroge sur la spécificité du processus développemental. Ainsi, dans la biologie du
développement, il existe des explications qui se concentrent sur une interprétation
précise du déroulement des processus morphologiques (dans la lignée des travaux de
Wolff) et des explications qui s’attachent à spécifier un contenu informationnel ou
bien des dispositions précises présentes durant le développement (dans la droite ligne
d’une tradition davantage préformationniste). Ce dernier type d’explication se
focalise, aujourd’hui, principalement sur l’analyse génétique alors que le premier
prend également en compte les processus épigénétiques (régulation génomique et/ou
influence environnementale sur le développement). D’aucuns ont vu une résolution
possible du débat entre épigenèse et préformation en considérant que les gènes et les
processus épigénétiques contribuaient les uns et les autres au développement. À
l’inverse, la vision qui attribue aux seuls gènes un rôle déterminant dans la génération
de la forme revient à dire qu’il existe dès le zygote un « programme » nécessaire à la
réalisation du développement. Le biologiste Richard Dawkins a même parlé de
« recette » pour le développement, laquelle serait déterminée par les gènes (1982) : la
génération de la forme serait déterminée par l’information génétique. Cette
conception a fait l’objet d’âpres discussions suite aux découvertes en épigénétique
montrant que les gènes seuls ne suffisaient pas et que d’autres éléments que les gènes
pouvaient entrer dans la « recette ». On peut voir, à travers la métaphore de Dawkins,
l’expression moderne d’une théorie de la préformation qui a eu et continue encore
d’avoir une influence sur les recherches en biologie du développement.
II. Un ou bien des « programme(s) » à l’origine de la forme en biologie ?
La notion de « programme », plus encore que celle de préformation qui reste
davantage associée à l’idée de préexistence de la forme, a acquis une place importante
dans la biologie contemporaine. François Jacob affirme dans La logique du vivant :
« Chaque œuf contient, […] dans les chromosomes reçus de ses parents, tout son
propre avenir, les étapes de son développement, la forme et les propriétés de l’être qui
en émergera [; l’] organisme [devenant] ainsi la réalisation d’un programme prescrit
par l’hérédité ». À son instar, de nombreux biologistes décrivent les processus par
lesquels la cellule œuf se transforme en organisme adulte en termes d’exécution d’un
« programme développemental ». L’usage de la notion de « programme » en biologie
correspond à la description des relations qui associent spécifiquement les gènes aux

différents structures et processus pour lesquels ils jouent un rôle causal (Atlan 1986 ;
Kupiec 2008). En majorité, les biologistes considèrent que les gènes jouent un rôle
causal primordial quant à une structure ou une forme donnée en tant qu’ils sont
« porteurs de l’information » relativement à la formation de ces différents produits.
Autrement dit, même si les gènes peuvent ne pas constituer la seule cause ou la cause
directe de la formation de ces produits et que l’information dont ils sont porteurs peut
requérir la coopération de différents facteurs environnementaux pour qu’ils puissent
être formés, la réalisation du programme dépendra néanmoins de leur présence.
Jacob souligne avec raison que dans l’idée de programme se trouvent deux notions
que l’intuition associe aux êtres vivants : la mémoire et le projet. Dès lors, la notion
de programme semble, d’une certaine façon, réconcilier l’opposition entre
préformation et épigenèse : la « mémoire » se situant du côté d’une conception de la
préformation, alors que le « projet » se situe, quant à lui, davantage du côté de la
conception épigénétique. Forts de ce nouveau concept, les biologistes se sont
consacrés ces dernières années à l’analyse de ce programme. En dépassant l’idée
initiale de « programme génétique » et en la remplaçant progressivement par celle de
« programme développemental », ils ont cherché à montrer que l’information du
programme pouvait reposer non pas uniquement sur des gènes mais sur des « réseaux
d’expression génique ». Ces réseaux prennent en compte toutes les interactions
régulatrices qui peuvent exister entre les gènes et qui sont susceptibles de moduler
l’information dont ils sont porteurs. La mise en évidence de ces réseaux – dont les
interactions viennent non seulement moduler mais aussi enrichir l’information portée
par les gènes – constitue une analyse fine de la mémoire du programme. Qu’en est-il
du projet du programme ?
Le biologiste peut s’intéresser au programme de deux manières différentes. D’une
part, il peut se concentrer sur la structure du programme, son fonctionnement, sa
logique ; c’est ce qu’il fait lorsqu’il analyse non seulement l’information portée par
les gènes mais aussi celle portée par les réseaux géniques. D’autre part, il peut
enquêter sur l’histoire des programmes, leur dérive et éventuellement les lois qui
permettent d’expliquer leurs modifications à travers les générations en fonction, par
exemple, des systèmes écologiques auxquels ils appartiennent. Cette seconde
approche fait moins souvent l’objet de discussions que la première approche parmi les
philosophes de la biologie, et ce, bien qu’elle fasse tout autant l’objet de recherches et
d’analyses. Analyser l’histoire des programmes implique d’examiner comment

l’information génétique ou bien comment les réseaux d’expression géniques (ce qui
peut apparaître plus simple à modéliser) ont pu évoluer dans le temps (Lynch 2008).
Le débat reste vif quand à la capacité qu’auraient les biologistes à déterminer le
caractère adaptatif (ayant évolué par sélection naturelle) ou non-adaptatif de cette
évolution (Lynch 2007). Ces deux approches renvoient au deux manières les plus
courantes pour le biologiste d’appréhender l’explication de la genèse de la forme par
le biais d’un « programme ». Dans un cas comme dans l’autre, il s’agit d’une
conception simple du « programme », au sens où l’information est portée par les
gènes ou les réseaux d’expression génique. Dès lors, le mode d’ « in-formation » ne
varie pas au cours du temps, seules les structures de cette information (l’agencement)
en sont modifiées.
Pourtant, il est possible d’adopter un autre usage de la notion de « programme
développemental », moins répandu que les deux usages précédents, lorsqu’il est
question de la genèse de la forme en développement. Depuis les années 2000, certains
biologistes du développement ont réussi à montrer que le processus de morphogenèse
reposait sur un ensemble de molécules relativement limité (Forgacs and Newman
2005, Newman & Bhat 2008, 2009; Newman & Müller, 2000 ; Gilbert et al. 1996). Le
biologiste Stuart A. Newman a, par exemple, identifié des molécules-types
(Cadhérine, Notch, Wnt, etc.) impliquées dans des épisodes cellulaires
développementaux spécifiques (comme l’adhésion, l’inhibition latérale, l’anisotropie
cellulaire de surface, etc.) et responsables de certaines formes types (comme
l’invagination cellulaire, la polarisation, la tubularisation, etc.) présentes lors du
développement chez de très nombreux clades6. Il nomme ces épisodes, qu’il associe
aux molécules-types, des « Blocs de Construction Développementaux7 » (BCD) et
montre que ces BCD se produisent dans les différents clades indépendamment des
gènes recrutés. En s’appuyant sur ces observations, Newman suggère que le processus
de morphogenèse peut lui-même être considéré comme étant devenu programmatique
au cours du temps (Newman 2014). Autrement dit, au cours de l’histoire évolutive, les
processus morphogénétiques seraient devenus de plus en plus déterminés par
l’information génétique. Dans la conception de Newman, l’idée de « programme
développemental » fait référence à l’association historique de certains gènes dans des
6 Un clade est un groupe d’organismes présentant des similarités morphologique. Le clade
correspond à l’unité de base de la classification phylogénétique des êtres vivants. 7 [Developmental Pattern Modules]. Voir Newman & Bhat 2008, 2009.

BCD. Il oppose un temps évolutif révolu, où la morphogenèse était plastique (où les
gènes n’étaient pas spécifiquement associés à une forme donnée), à un temps évolutif
avancé, où le processus développemental est devenu programmatique (la genèse
d’une forme donnée étant désormais associée de manière plus spécifique à des
facteurs génétiques donnés). Dans cet usage de la notion de « programme
développemental », la génération de la forme est dissociée de la réalisation du
programme génétique, puisqu’on considère que le rapport entre les deux ne
correspond plus qu’à une étape spécifique dans l’histoire évolutive. Dès lors, la notion
de « programme développemental » implique de rendre compte davantage du
« projet » du programme (des moyens de sa réalisation) que de sa « mémoire ».
Si une dissociation est possible entre la génération de la forme et la réalisation
du programme développemental, il en est autrement de la question du « type »
(molécules types, BCD) qui semble nécessairement associé à une forme donnée.
Comment expliquer la stabilité ou la robustesse de la forme-type ? C’est là l’objet de
la prochaine section.
III. La génération de la forme : entre « robustesse » et « plasticité » ?
Si certains concepts, comme celui d’espèce, peuvent être considérés comme des
émanations de la théorie de l’évolution telle que l’a formulée Darwin à la fin du XIXe
siècle8, ce n’est aucunement le cas du concept de forme, dont la compréhension
repose sur ses usages métaphysiques antiques. Depuis Aristote, et surtout depuis le
XVIIe siècle, on oppose le concept de forme à celui de matière : la matière est ce qui
reste indéterminé, la forme étant nécessairement déterminée. Comme nous l’avons
indiqué précédemment, à partir de l’épigenèse germanique, à la fin du XVIIIe siècle, il
n’est plus nécessaire d’expliquer les raisons du processus développemental :
l’organisation du vivant dans son ensemble est expliquée téléologiquement. C’est
dans ce contexte que prend naissance l’embryologie expérimentale. Une conséquence
de ce nouveau paradigme développemental est l’importance des données
embryologiques pour l’histoire naturelle (Roe, 1981). C’est également à cette époque
là que la notion d’archétype – l’idée selon laquelle toutes les formes animales sont des
variations d’un nombre limité de types idéaux – est élaborée (Buffon (1750), puis
Owen (1848)). Or c’est cette notion qui contribue en premier à l’émergence de l’idée
8 cf. chapitre A. Barberousse.

de « programme », avant même que le support génétique n’existe. L’usage que fait le
biologiste Stuart Newman de la notion de « programme développemental » semble
étroitement lié à cette notion, même s’il n’examine plus uniquement les formes à
l’échelle organique mais aussi à l’échelle cellulaire, voire moléculaire.
Depuis le début des années 2000, la notion de « robustesse développementale » est
venue s’adjoindre, voire se substituer, à celle de « programme développemental » et
permet, en un sens, d’éviter l’ambiguïté soulevée par la notion de « programme ».
L’idée de « robustesse » (Keller, 2002) renseigne davantage sur la structure du
processus développemental que sur sa finalité (le projet) ou son origine (la mémoire).
À la robustesse des mécanismes développementaux sont associées des formes
considérées, elles-aussi, comme robustes (c’est-à-dire stables et constantes au sein des
espèces et tout au long de leur histoire évolutive). Par exemple, certains stades
développementaux se caractérisent par la robustesse des formes développementales
chez différentes espèces, reposant sur des processus morphologiques similaires. C’est
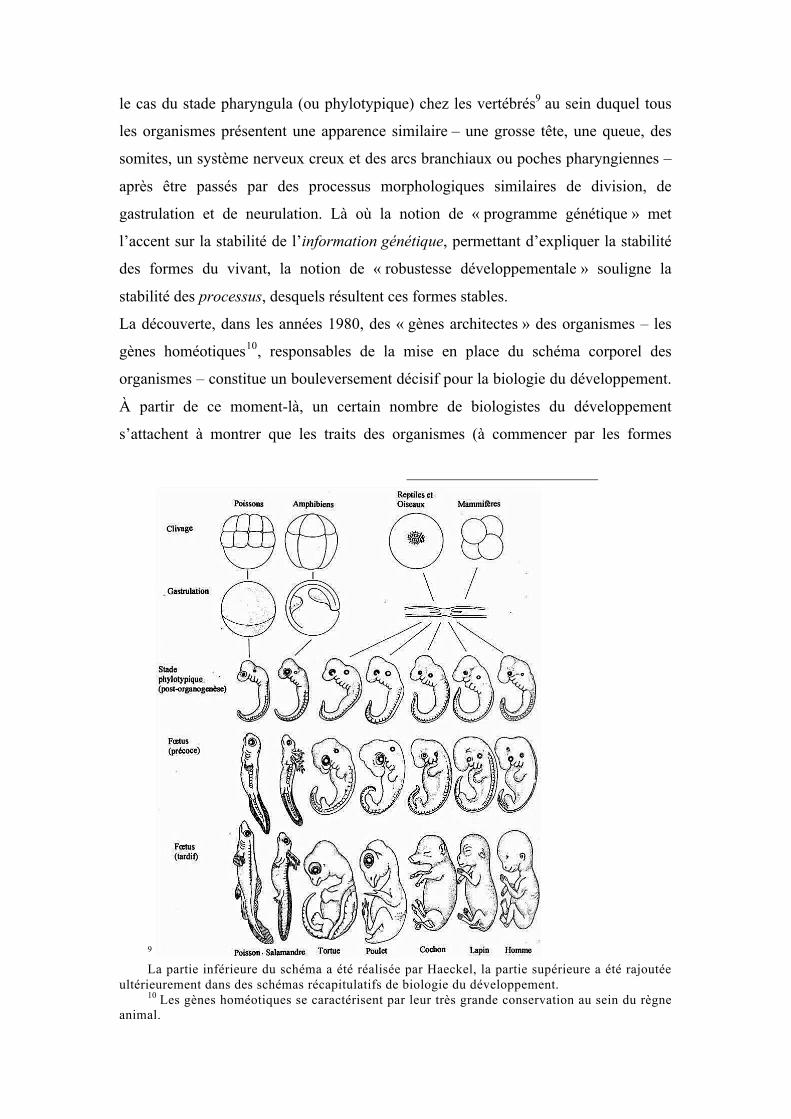
le cas du stade pharyngula (ou phylotypique) chez les vertébrés9 au sein duquel tous
les organismes présentent une apparence similaire – une grosse tête, une queue, des
somites, un système nerveux creux et des arcs branchiaux ou poches pharyngiennes –
après être passés par des processus morphologiques similaires de division, de
gastrulation et de neurulation. Là où la notion de « programme génétique » met
l’accent sur la stabilité de l’information génétique, permettant d’expliquer la stabilité
des formes du vivant, la notion de « robustesse développementale » souligne la
stabilité des processus, desquels résultent ces formes stables.
La découverte, dans les années 1980, des « gènes architectes » des organismes – les
gènes homéotiques10, responsables de la mise en place du schéma corporel des
organismes – constitue un bouleversement décisif pour la biologie du développement.
À partir de ce moment-là, un certain nombre de biologistes du développement
s’attachent à montrer que les traits des organismes (à commencer par les formes
9 La partie inférieure du schéma a été réalisée par Haeckel, la partie supérieure a été rajoutée
ultérieurement dans des schémas récapitulatifs de biologie du développement. 10 Les gènes homéotiques se caractérisent par leur très grande conservation au sein du règne
animal.

complexes comme les organes) ne peuvent pas être expliqués en se référant aux seules
variations de fréquence génétique – comme cela avait été principalement le cas depuis
l’avènement de la génétique des populations. La mise en évidence des gènes
homéotiques permet aussi d’illustrer l’importance d’une nouvelle forme
d’homologie11 (distincte de l’homologie de structure, mais qui permet d’expliquer la
similarité d’organes complexes chez des espèces différentes) : l’homologie de
processus (Gilbert 1996). Dès lors, des changements affectant les complexes des
gènes Hox (une famille de gènes homéotiques), présents chez la drosophile, peuvent
contribuer à la perturbation de son schéma corporel (par exemple, les segments
thoracique-ailes et abdominaux-pattes seront inversés). La redondance de ces
complexes de gènes au sein du règne animal (et végétal) permet d’expliquer que des
fonctions similaires soient retrouvées dans des contextes différents (Kitano, 2004).
Ainsi, l’expression d’un même gène homéotique peut conduire à la formation de deux
cœurs différents, comme le cœur des insectes et le cœur des vertébrés, dont la
formation a reposé sur des processus identiques et qui à l’arrivée possèdent la même
fonction – la capacité à pomper du sang –, et ce bien que leur forme précise diffère
(Manak & Scott 1994). Plus encore que l’idée de stabilité de la forme, c’est l’idée de
stabilité de processus développemental conduisant à une stabilité de la fonction qui est
traduite à travers l’usage de la notion de « robustesse développementale ».
Depuis une dizaine d’années, les biologistes du développement ont fait resurgir un
autre concept, apparu pour la première fois dans le milieu des années 1960 en
génétique (Bradshaw 1965), pour rendre compte de la pluralité des formes qui
peuvent être générées dans le vivant en dépit d’une information génétique stable. Il
s’agit du concept de « plasticité phénotypique ». Ce dernier, qui servait
principalement à expliquer les raisons pour lesquelles une même espèce (possédant la
même information génétique) pouvait adopter des formes différentes selon les
environnements dans lesquels elle se développait, est devenu un concept essentiel
pour la formulation d’une nouvelle synthèse évolutionnaire incluant le
développement. Ainsi, le concept de « plasticité développementale » (West-Eberhard
2003) ne permet plus tant de rendre compte de la stabilité de la forme que des moyens
par lesquels la forme peut être générée.
11 En biologie, l’homologie désigne généralement le lien évolutif qui existe entre deux traits
(en général anatomiques) observés chez deux espèces différentes et qui est dû au fait que toutes deux l’ont hérité d’un ancêtre commun.

IV. Quel « développement » pour éclairer notre compréhension de
l’évolution de la forme ?
Dans le schéma traditionnel permettant de décrire l’évolution des formes du vivant, et
adopté depuis l’avènement de la Synthèse Moderne de l’évolution (Huxley 1942) – le
courant théorique qui a contribué au rapprochement disciplinaire de la biologie
évolutionnaire et de la génétique mendélienne par le biais de la génétique des
populations –, le développement est laissé pour compte. Comme cela sera montré par
la suite (Arthur 1997, Hall 2003), il est tout au plus considéré comme une « boîte
noire », puisque seuls les signaux entrants (l’information génétique) et les signaux
sortants (les traits phénotypiques) comptent. La compréhension des processus et des
mécanismes qui caractérisent le développement n’est pas nécessaire pour comprendre
la variation, qui est ce sur quoi la sélection va agir. L’évolution de la variation est
expliquée par le fait que des mutations, survenues aléatoirement, vont ou non se
répandre dans la population, conduisant potentiellement à des modifications stables de
la forme tout au long de l’échelle de temps évolutive, en fonction du caractère
adaptatif de la forme en question. Depuis les années 1980, certaines approches ont
cherché à redonner une nouvelle importance au développement dans les explications
évolutionnistes en montrant, entre autres choses, l’importance et la spécificité des
gènes développementaux (comme les gènes homéotiques ou, plus récemment, les
gènes régulateurs) pour expliquer l’évolution des formes du vivant (Carroll 1995).
En 2003, la biologiste Mary Jane West-Eberhard propose dans un ouvrage de
synthèse, intitulé Developmental Plasticity and Evolution, un schéma qui repose en
grande partie sur le concept de « plasticité développementale ». De l’étude poursuivie
sur une période de vingt ans du comportement des insectes (principalement les
abeilles) corrélé à leur développement, l’auteure a retiré la conviction qu’une nouvelle
synthèse de la théorie de l’évolution, qui prenne mieux en compte le développement,
était devenue nécessaire. Si le livre appartient bien au champ disciplinaire de la
biologie évolutionnaire du développement (l’évo-dévo), West-Eberhard ne se
contente pas de rassembler des données issues de la biologie de l’évolution et de la
biologie du développement, elle entend en sus proposer une nouvelle théorie
synthétique de l’évolution qui inclurait le développement dans sa définition. Lorsque
le livre paraît, il crée un certain émoi dans la communauté des biologistes, car il
propose un modèle radicalement nouveau pour rendre compte de l’évolution des

formes. Au sein de ce nouveau schéma conceptuel, les gènes ne sont plus considérés
comme étant des initiateurs, mais seulement comme ayant un rôle secondaire dans
l’évolution (la formule anglaise est plus parlante : “followers, not leaders in
evolution”). Autrement dit, les changements dans les fréquences génétiques sont
considérés comme intervenant dans un second temps plutôt que comme expliquant
initialement l’apparition d’une nouvelle forme phénotypique. Si la plupart des
théories qui expliquent la diversité des formes du vivant continuent de s’appuyer
principalement sur des études en génétique quantitative, West-Eberhard renverse les
rapports de priorité en faisant passer l’évolution du génome à l’arrière-plan de
l’analyse pour se concentrer principalement sur les formes du vivant observées.
Le processus général qu’elle décrit est le suivant : un changement dans
l’environnement dans lequel se trouve l’organisme en développement conduit à une
« réponse développementale plastique » et à une accommodation phénotypique –
« l’ajustement immédiat aux changements résultant de la flexibilité adaptative
multidimensionnelle du phénotype, sans changement génétique » (West-Eberhard,
2003, 140) – ce qui va permettre, dans un second temps, l’amélioration de
l’adaptation individuelle au nouvel environnement. Les nouvelles formes, résultantes
de cette plasticité développementale, sont ensuite sélectionnées. Enfin, un changement
dans les fréquences alléliques (l’accommodation génétique) permet d’améliorer et
d’intégrer ce changement de forme durablement. West-Eberhard rapporte le célèbre
exemple d’une telle accommodation fourni par le vétérinaire hollandais Slijper. En
1942, ce dernier avait observé une chèvre qui, à sa naissance, ne possédait que deux
pattes, mais dont l’organisme avait réagi à ce lourd handicap par des spécialisations
morphologiques et comportementales qui avaient finalement permis à la chèvre de
survivre et même de se déplacer. West-Eberhard s’appuie, tout au long de son livre,
sur cet exemple surprenant qui illustre la portée générale et les enjeux lointains de son
investigation. Le point important, qui fait la force de cet exemple particulier, est qu’en
dépit des malformations congénitales majeures dont souffre la chèvre, la fonction de
locomotion parvient à être préservée au cours de l’ontogenèse. Selon West-Eberhard,
les accommodations adaptatives qui favorisent le développement de la fonction (la
locomotion, par exemple) ont une probabilité élevée de contribuer à l’apparition d’une
nouveauté fonctionnelle (un nouveau mode de locomotion), elle-même liée à une
nouvelle organisation morphologique – une nouvelle forme. Pour peu que cette
nouvelle organisation morphologique soit viable et compatible avec la genèse de

l’individu, la nouveauté fonctionnelle qui en résulte peut conduire à une augmentation
de la fréquence d’apparition de la forme nouvelle (deux pattes au lieu de quatre) dans
la population. Ceci peut conduire éventuellement, sur de très nombreuses générations,
à la production d’une sous-population d’individus présentant le trait en question. Le
processus peut alors être suivi d’une « accommodation génétique » qui se traduit par
un changement dans la fréquence génétique qui affecte la régulation, l’expression ou
les effets secondaires de la nouvelle forme, sous l’effet du processus de
sélection/variation décrit par Darwin. Au sein de ce processus original,
l’environnement devient un participant clé dans la génération et la sélection des
adaptations, mais aussi dans la capacité adaptative des organismes eux-mêmes. Ainsi,
le génome est considéré comme une composante « physique », au même titre que
l’environnement (l’un et l’autre « modelant » l’organisme en devenir). La forme
générée est, quant à elle, conçue comme la conséquence de l’expression de ces
facteurs physiques, et sa réalisation dépend en grande partie du processus
développemental.
Venons-en maintenant à la question de la spécificité de la génération de la forme en
biologie.
V. Comment distinguer la génération de la forme en biologie de la génération de la
forme en physique ?
Plus que l’idée de « vivant », c’est l’idée de « complexification » qui est centrale dès
lors que l’on traite de la question du développement de la forme en biologie. Cette
idée est non seulement spécifique à la biologie, mais elle est surtout propre au
processus développemental puisque, contrairement au processus évolutif qui, comme
l’avait souligné Darwin, ne va pas nécessairement vers une complexité plus grande,
l’ontogenèse se caractérise essentiellement par une complexification progressive.
Ainsi, l’organisme à l’issue de son développement sera, par définition, plus complexe
que la cellule œuf originelle dont il provient. L’apparition de formes diverses (au fur
et à mesure de l’organogenèse, par exemple) participe de cette complexification. Au
cours de ce processus développemental, la mise en place de la forme repose à la fois
sur des mécanismes cellulaires mais aussi, à une échelle plus fine, sur des mécanismes
qui peuvent être appréhendés par le biais de lois physico-chimiques. Le
rapprochement entre certains mécanismes morphologiques et des mécanismes d’ordre
physico-chimiques a conduit certains biologistes à adopter une vision unifiée de la

morphologie et de la biochimie. Le biochimiste, Joseph Needham est, à ce titre, une
figure importante. Ce dernier a même été jusqu’à considérer que les formes issues des
développements embryologiques étaient déterminées par des substances chimiques,
lesquelles reposaient d’ores et déjà sur une organisation élaborée qu’il a qualifié de
« morphologie moléculaire » (Needham, 1968). La frontière entre forme physique et
forme biologique semblait rompue.
Dans la lignée de ces travaux, le biologiste Stuart Newman montre, quant à lui, quels
sont les liens mais aussi les différences entre les mécanismes physico-chimiques à
l’origine de la forme et les morphologies cellulaires (Forgacs & Newman 2005).
Ainsi, en s’appuyant sur deux caractéristiques qui se manifestent à la fois dans les
tissus d’origine animale et dans les matériaux non-vivants – « la matière molle12 » (de
Gennes 1992) et les « média excitables13 » (Beloussov 1998) –, il argumente en
faveur d’une distinction de la génération de la forme en physique et en biologie en
montrant que si les deux caractéristiques physico-chimiques existent bien hors de la
biologie, il est néanmoins rare de les trouver de manière simultanée dans des
matériaux non vivants. Le matériau biologique à l’origine de la forme vivante serait
donc, en un sens, là encore plus complexe que le matériau à l’origine de la forme en
physique. Plus précisément, le processus développemental, duquel résulte la forme, se
caractériserait par sa nature « hybride » (Newman, 2014) à la différence de n’importe
quel autre processus physico-chimique complexe. Au sens où le processus
développemental ferait appel à différents types de média physico-chimiques de
manière cumulés là où ils sont généralement isolés ou « purs » dans les matériaux non
vivants. Par ailleurs, tout au long du processus développemental (et à la différence du
processus de croissance), le rôle et l’expression des facteurs physico-chimiques et
génétiques ne cessent de se modifier, générant une situation de constant déséquilibre
qui sera à l’origine de la différenciation progressive de la forme. Le caractère hybride
du processus développemental permet dans ce contexte d’expliquer la génération de
formes intermédiaires successives jusqu’à l’établissement d’une forme complexe
définie (un organe par exemple) qui, bien qu’ayant été contrainte par des facteurs et
des processus constants, sera toujours le résultat de la réalisation d’un programme
original.
12 « matière molle » [Soft Matter] matériaux viscoélastiques condensés tels que des colloïdes, des cristaux liquides, des mousses, ou des gels.
13 « média excitables » [excitable media] systèmes chimiques, électriques, mécanistiques (réactions d’oscillation chimique).

Conclusion
L’étude de la génération de la forme chez les êtres vivants d’un point de vue
philosophique est un sujet certes ancien mais qui requiert une nouvelle interprétation
dans la biologie contemporaine. En effet, et tout d’abord, s’il y a bien un domaine
pour lequel la philosophie de la biologie ne peut se passer d’une analyse historique,
c’est bien celui de la biologie du développement. Car même si les outils théoriques et
empiriques d’observation de l’organisme en développement ont radicalement changé
depuis l’Antiquité et même depuis le XVIIIe siècle, les problèmes philosophiques
relatifs à la genèse de la forme ne sont pas nouveaux. Ensuite, la compréhension des
problèmes contemporains relatifs à la génération de la forme a eu tendance à être
négligée au profit de questions relatives à l’interaction des explications
développementales avec les explications évolutionnaires. Dès lors, les biologistes ont
cherché à expliquer la génération de la forme en se référant principalement aux
mécanismes de l’évolution. Or, et comme j’ai cherché à le montrer à travers ce
chapitre, le problème de la génération de la forme en biologie soulève des questions
philosophiques qui lui sont propres et dont la formulation remonte bien avant la
théorie de la sélection naturelle par Darwin. C’est donc en examinant la spécificité du
processus développemental en tant qu’il est une morphogenèse biologique et non
uniquement physico-chimique que l’on peut appréhender intégralement la question de
la génération de la forme en biologie et donc aussi la manière dont ce problème peut
être repensé au sein de la théorie de l’évolution.
Bibliographie
Arthur, W. (1997). The Origin of Animal Body Plans: A Study in Evolutionary
Developmental Biology. Cambridge, New York : Cambridge University Press.
Atlan, H. (1986). [1979]. Entre le cristal et la fumée : essai sur l’organisation du
vivant. Paris : Éditions du Seuil.

Beloussov, L. (1998). The Dynamic Architecture of a Developing Organism.
Dordrecht : Kluwer Academic Publishers.
Bradshaw, A.D. (1965). « Evolutionary Significance of Phenotypic Plasticity in
Plants », Advances in Genetics, 13, p. 115–155.
Buffon, (1830) [1750]. Œuvres complètes, Histoire des Animaux. Paris : F.D. Pillot.
Canguilhem, G. (1983). « La nouvelle connaissance de la vie. Le concept et la vie »,
in Études d’histoire et de philosophie des Sciences, 5ème édition augmentée, Paris,
Vrin, pp 335-364.
Canguilhem, G., Lapassade, G., Piquemal, J., Ulmann J. (2003) [1962]. Du
développement à l’évolution au XIXe siècle. 1ère édition « Quadrige », Paris, Presses
Universitaires de France.
Carroll, S.B. (1995). « Homeotic Genes and the Evolution of Arthropods and
Chordates », in Gee, H. (ed.) Shaking the Tree: Readings from Nature in the History
of Life. University of Chicago Press.
Cooper, J.M. (1988). « Metaphysics in Aristotle’s Embryology », Proceedings of the
Cambridge Philological Society, 214, p. 14–41.
Dawkins, R. (1982). The Extended Phenotype: The Long Reach of the Gene. Oxford :
Oxford University Press.
De Gennes, P.G. (1992). « Soft matter », Science, 256, p. 495–497.
Forgacs, G. & Newman, S.A. (2005). Biological physics of the developing embryo.
Cambridge UK, New York : Cambridge University Press.
Gilbert, S.F. (1996). « Cellular dialogues during development », in Neri, G., Martini-
Neri, E. & Opitz, J.M. (Eds.), Gene Regulation and Fetal Development: Procedings
of the Third International Congress on Fetal Genetic Pathology. New York : Wiley–
Liss.
Gilbert, S.F., Opitz, J.M. & Raff, R.A. (1996). « Resynthesizing Evolutionary and
Developmental Biology », Developmental Biology, 173, 2, p. 357–372.
Hall, B.K. (2003). « Unlocking the Black Box Between Genotype and Phenotype:
Cell Condensations as Morphogenetic (modular) Units », Biology and
Philosophy, 18, 2, p. 219–247.
Huxley, J. (1942). Evolution. The Modern Synthesis.
Jacob F. (1970). La logique du vivant : une histoire de l’hérédité. Paris : Éditions
Gallimard.

Keller, E.F. (2002). « Developmental Robustness », Annals of the New York Academy
of Sciences, 981, 1, p.189–201.
Kitano, H. (2004). « Biological Robustness » Nature Reviews Genetics 5, no. 11
(2004): 826–37.
Kupiec, J.-J. (2008). L’origine des individus. Paris : Fayard.
Lynch, M. (2007). « The Evolution of Genetic Networks by Non-Adaptive
Processes », Nature Reviews Genetics, 8, 10, p. 803–813.
Lynch, M. (2008). The Origins of Genome Architecture. 1st ed. Sinauer Associates
Inc.
Mayr, E. (1961). « Cause and Effect in Biology : kinds of causes, predictability, and
teleology are viewed by a practicing biologists », Science, 134, 3489, p.
1501–1506.
Maienschein J., “Epigenesis and Preformationism” In The Stanford Encyclopedia
of Philosophy, edited by Edward N. Zalta, Spring 2012,
http://plato.stanford.edu/archives/spr2012/entries/epigenesis/.
Manak, J.R. & Scott, M.P. (1994) « A Class Act: Conservation of Homeodomain
Protein Functions », Development, p. 61–77.
Needham, J. (1959). A History of Embryology. New York : Abelard-Schuman.
Needham, J. (1968). Order and Life. Cambridge Mass. & London : the M.I.T. Press.
Newman, S.A. (2014), « Physico-genetics of morphogenesis: the hybrid nature of
developmental mechanisms », in Minelli, A. & Pradeu T. (eds.), Towards a
Theory of Development. Oxford : Oxford University Press.
Newman, S.A. & Bhat, R. (2008). « Dynamical Patterning Modules: Physico-Genetic
Determinants of Morphological Development and Evolution », Physical
Biology, 5, p. 1–14.
Newman, S.A. & Bhat, R. (2009). « Dynamical Patterning Modules: A ‘pattern
Language’ for Development and Evolution of Multicellular Form »,
International Journal of Developmental Biology, 53, p. 693–705.
Newman, S.A., & G. B. Müller, “Epigenetic mechanisms of character
origination”. Journal of Experimental Zoology Part B (Molecular and
Developmental Evolution) 288, 304–17. (2000).

Owen, R. (1848). On the Archetype and Homologies of the Vertebrate Skeleton.
London: John van Voorst, Paternoster Row.
http://archive.org/details/onarchetypeandh01owengoog
Pigliucci, M. “Do we need and extended evolutionary synthesis? Evolution,
61(12), 2743 –2749. (2007).
Roe, S. (1981). Matter, Life, and Generation : Eighteenth-Century Embryology
and the Haller-Wolff Debate. Cambridge : Cambridge University Press.
West-Eberhard, M.J. (2003). Developmental Plasticity and Evolution. Oxford :
Oxford University Press.




















