
Gametogenesis and maturity stages scale of Raja asterias Delaroche, 1809 (Chondrichthyes, Raijdae)...
10
BIODIVERSITY IN ENCLOSED SEAS Gametogenesis and maturity stages scale of Raja asterias Delaroche, 1809 (Chondrichthyes, Raijdae) from the South Ligurian Sea M. Barone S. De Ranieri O. Fabiani A. Pirone F. Serena Ó Springer Science+Buisness Media B.V. 2007 Abstract This stu\dy was aimed at the acquisition of basic life-history information on Raja asterias through an objective approach in the assignment of maturity stage, using both histological techniques and a multivariate analysis aimed at macroscopic evaluations of the reproductive system. The sam- ples examined were collected from landings of commercial trawl vessels from the port of Viareg- gio. From July 2001 to August 2002 a total of 351 specimens, 166 females and 155 males, were purchased. A description of the main stages of oogenesis and spermatogenesis is presented here. Moreover, on the basis of the maturity stage definition, maturity ogives were constructed. Length at 50% maturity was estimated to be 51.7 cm and 56.1 cm total length for males and females respectively. Finally an informal table for the maturity stage assignment, which can be useful for a quick and easier definition both on board and in the laboratory, is formulated. Keywords Shark fisheries Sexual maturity Oogenesis Spermatogenesis Introduction The starry ray Raja asterias Delaroche, 1809 is common in the whole Mediterranean excluding probably the Black Sea (Fischer et al., 1987; Relini et al., 2000). Although along the Atlantic coasts of northern Morocco and in southern Portugal it has been recorded (Whitehead et al., 1984), R. asterias may be considered in general terms a Mediterranean endemic species. It is a benthic species generally found in shallow waters on muddy or sandy soft bottoms, mostly concen- trated at depths up to 150 m (Serena et al., 1988). In the southern Ligurian Sea port of Viareggio, R. asterias is among the most important compo- nents of the fish assemblages caught all year round with beam-trawling. Elasmobranchs are particu- larly vulnerable to fishing pressure, because of their biological characteristics: low growth rates, late sexual maturation, low fecundity, and a long reproductive cycle. The decline of many species as a consequence of removals that went beyond the Guest editors: G. Relini & J. Ryland Biodiversity in Enclosed Seas and Artificial Marine Habitats M. Barone F. Serena (&) Agenzia Regionale per la Protezione Ambientale della Toscana, Via Marradi, 114, 56100 Livorno, Italy e-mail: [email protected] S. De Ranieri Centro Interuniversitario di Biologia Marina, Livorno, Italy O. Fabiani A. Pirone Sezione di anatomia – Dipartimento di produzioni animali, Universita ` di Pisa, Pisa, Italy 123 Hydrobiologia (2007) 580:245–254 DOI 10.1007/s10750-006-0448-x
Transcript of Gametogenesis and maturity stages scale of Raja asterias Delaroche, 1809 (Chondrichthyes, Raijdae)...

BIODIVERSITY IN ENCLOSED SEAS
Gametogenesis and maturity stages scale of Raja asteriasDelaroche, 1809 (Chondrichthyes, Raijdae) from the SouthLigurian Sea
M. Barone Æ S. De Ranieri Æ O. Fabiani ÆA. Pirone Æ F. Serena
� Springer Science+Buisness Media B.V. 2007
Abstract This stu\dy was aimed at the acquisition
of basic life-history information on Raja asterias
through an objective approach in the assignment of
maturity stage, using both histological techniques
and a multivariate analysis aimed at macroscopic
evaluations of the reproductive system. The sam-
ples examined were collected from landings of
commercial trawl vessels from the port of Viareg-
gio. From July 2001 to August 2002 a total of 351
specimens, 166 females and 155 males, were
purchased. A description of the main stages of
oogenesis and spermatogenesis is presented here.
Moreover, on the basis of the maturity stage
definition, maturity ogives were constructed.
Length at 50% maturity was estimated to be
51.7 cm and 56.1 cm total length for males and
females respectively. Finally an informal table for
the maturity stage assignment, which can be useful
for a quick and easier definition both on board and
in the laboratory, is formulated.
Keywords Shark fisheries � Sexual maturity �Oogenesis � Spermatogenesis
Introduction
The starry ray Raja asterias Delaroche, 1809 is
common in the whole Mediterranean excluding
probably the Black Sea (Fischer et al., 1987;
Relini et al., 2000). Although along the Atlantic
coasts of northern Morocco and in southern
Portugal it has been recorded (Whitehead et al.,
1984), R. asterias may be considered in general
terms a Mediterranean endemic species. It is a
benthic species generally found in shallow waters
on muddy or sandy soft bottoms, mostly concen-
trated at depths up to 150 m (Serena et al., 1988).
In the southern Ligurian Sea port of Viareggio,
R. asterias is among the most important compo-
nents of the fish assemblages caught all year round
with beam-trawling. Elasmobranchs are particu-
larly vulnerable to fishing pressure, because of
their biological characteristics: low growth rates,
late sexual maturation, low fecundity, and a long
reproductive cycle. The decline of many species as
a consequence of removals that went beyond the
Guest editors: G. Relini & J. RylandBiodiversity in Enclosed Seas and Artificial MarineHabitats
M. Barone � F. Serena (&)Agenzia Regionale per la Protezione Ambientaledella Toscana, Via Marradi, 114, 56100 Livorno, Italye-mail: [email protected]
S. De RanieriCentro Interuniversitario di Biologia Marina,Livorno, Italy
O. Fabiani � A. PironeSezione di anatomia – Dipartimento di produzionianimali, Universita di Pisa, Pisa, Italy
123
Hydrobiologia (2007) 580:245–254
DOI 10.1007/s10750-006-0448-x

sustainable levels has been stressed by many
authors (Musick, 1999; Stevens et al., 2000).
However, it seems that this did not occur for R.
asterias. On the grounds where the fishery oper-
ates, data on catch trends for the last 15 years
suggest that the biomass of the stock is in a steady
state (Serena et al., 1988; Abella & Serena, 2005).
Almost nothing is known about the reproduc-
tive biology of the starry ray in the South Ligurian
Sea. Lo Bianco (1908) reported observation of
specimens from the Golfo di Napoli, Tortonese
(1956) of specimens from markets of Livorno,
Napoli and the Adriatic Sea, Capape (1977) also
studied the biology of R. asterias in the Tunisian
waters. This species is oviparous, mature females
being present throughout the year (Capape, 1977)
and the egg-cases are usually deposited at a depth
of about 30–40 m. The only available data on
rates of egg laying and incubation were obtained
under experimental conditions. The rate of egg
lying estimated by Lo Bianco (1908) was one egg
capsule every 4 days; the incubation period lasted
about 5–6 months (Capape, 1977). As newborn
juveniles of about 8 cm TL have been found
distributed primarily in the northern Tyrrhenian
Sea at depth of 8–12 m, this area is very likely to
be a nursery (Abella et al., 1997).
This study was aimed at the acquisition of basic
life-history information on R. asterias through an
objective approach in the assignment of maturity
stage, using both histological techniques and a
multivariate analysis aimed at macroscopic evalu-
ations of the reproductive system. A description of
the main stages of oogenesis and spermatogenesis,
the definition of a maturity scale and an estimate of
length at first maturity are showed here.
Materials and methods
The samples examined were collected from land-
ings of commercial vessels utilizing a variant of
the beam-trawl called rapido and operating from
the estuary of the Arno River to the Isle of Tino.
From July 2001 to August 2002, monthly pur-
chases amounted to a total of 351 specimens, 166
females and 155 males. At the laboratory, total
length (nearest 0.5 cm below), from tip of snout
to terminal point of the caudal fin, disc width and
sex were recorded for each specimen. In addition,
in females the maximum nidamental gland width
(nearest 0.1 mm) was measured.
Methodology for definition of maturation
stages
In order to determine maturity stages, for
females, the size of the ovarian eggs, the condi-
tion of the oviducts, the measurements of the
nidamental gland, and the presence of egg-cases
were considered; for males the measurements and
the consistency of the claspers, the development
of the sperm ducts and the gonads (testes) were
observed. Each characteristic was associated with
a score. As a result, each specimen was allocated
a code made of four scores. In order to group
together specimens with the same macroscopic
features, data of reproductive apparatus were
analyzed with multivariate techniques carried out
through the option ‘‘agglomerative hierarchical
clustering’’ of S-Plus program (MathSoft, 1999).
Determination of the length at maturity
The length at 50% maturity (L50) for both sexes
was estimated. Females were classified as mature
when they were in stages up to III (vitellogenic
follicles, well differentiated oviducts, nidamental
gland of large size, including specimens with egg
capsules), conferred by the previous method;
males when they were in stage IV (claspers
longer than tips of posterior pelvic fin lobes and
with skeleton hardened, sperm ducts filled,
gonads wide, rose-coloured, with seminiferous
follicles filling the whole volume).
The proportion of mature females and males at
length was described through a logistic function
as follows:
Y ¼ 1=1þ eaþbX
where Y is the estimated mature proportion, a
and b are the estimated coefficients of the logistic
equation and X is the total length. The length at
maturity can be estimated as the ratio of the
coefficients (a/b) by substituting Y = 0.5 in equa-
tion. The curve fitting was done using the specific
tool included in Microsoft Excel.
246 Hydrobiologia (2007) 580:245–254
123
Histological techniques
The ovaries of 34 females and the testes of 7
males, representing all stages of morphological
maturity, were extracted in their entirely and
fixed in 10% formaldehyde 0.1 M phosphate
buffer, pH 7.4. Small fragments of each gonad
were extensively washed in fresh water for one
day, dehydrated in a graded ethanol series
(30 min at 30%, 50%, 70%; 1 h at 80%; 2 h at
95%), then embedded in glycol methacrylate
resin (JB-4, Polysciences Inc.). Microtomed sec-
tions of 5 lm were collected on glass slides coated
with 0.5% gelatine containing 0.05% chrome
alum. Sections were subsequently stained with
toluidine blue, methylene blue and PAS reaction,
cleared in xylene, mounted with DPX and exam-
ined with a light microscope (·10–·1000 magni-
fication).
Methodology for ovarian egg counts
One of the ovaries of 166 female specimens was
fixed in modified Gilson’s solution (Simpson,
1951), which composition was: 100 ml ethanol
60%, 15 ml nitric acid 80%, 18 ml acetic acid,
20 g mercury chloride, 800 ml distilled water. The
total number of eggs in each ovary was counted
and their diameter was measured with the eye-
piece micrometer of a stereoscope.
Results
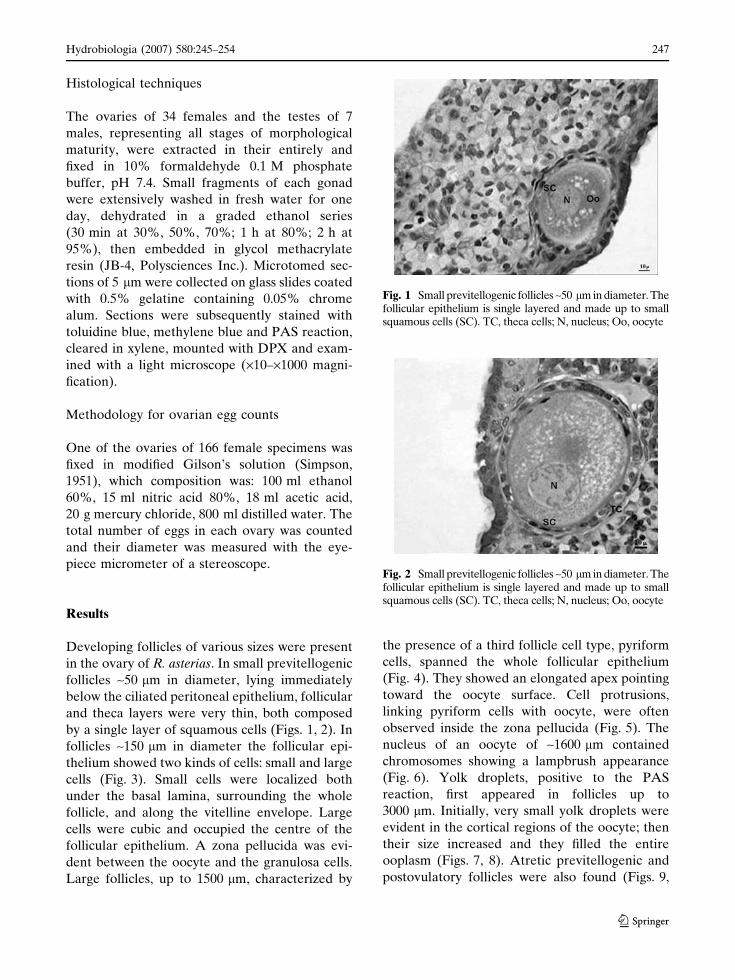
Developing follicles of various sizes were present
in the ovary of R. asterias. In small previtellogenic
follicles ~50 lm in diameter, lying immediately
below the ciliated peritoneal epithelium, follicular
and theca layers were very thin, both composed
by a single layer of squamous cells (Figs. 1, 2). In
follicles ~150 lm in diameter the follicular epi-
thelium showed two kinds of cells: small and large
cells (Fig. 3). Small cells were localized both
under the basal lamina, surrounding the whole
follicle, and along the vitelline envelope. Large
cells were cubic and occupied the centre of the
follicular epithelium. A zona pellucida was evi-
dent between the oocyte and the granulosa cells.
Large follicles, up to 1500 lm, characterized by
the presence of a third follicle cell type, pyriform
cells, spanned the whole follicular epithelium
(Fig. 4). They showed an elongated apex pointing
toward the oocyte surface. Cell protrusions,
linking pyriform cells with oocyte, were often
observed inside the zona pellucida (Fig. 5). The
nucleus of an oocyte of ~1600 lm contained
chromosomes showing a lampbrush appearance
(Fig. 6). Yolk droplets, positive to the PAS
reaction, first appeared in follicles up to
3000 lm. Initially, very small yolk droplets were
evident in the cortical regions of the oocyte; then
their size increased and they filled the entire
ooplasm (Figs. 7, 8). Atretic previtellogenic and
postovulatory follicles were also found (Figs. 9,
Fig. 1 Small previtellogenic follicles ~50 lm in diameter. Thefollicular epithelium is single layered and made up to smallsquamous cells (SC). TC, theca cells; N, nucleus; Oo, oocyte
Fig. 2 Small previtellogenic follicles ~50 lm in diameter. Thefollicular epithelium is single layered and made up to smallsquamous cells (SC). TC, theca cells; N, nucleus; Oo, oocyte
Hydrobiologia (2007) 580:245–254 247
123

10). The former were characterized by follicle and
theca cells that later become hypertrophic. In
postovulatory follicles the basal lamina, positive
to the PAS reaction, appeared collapsed and
invaded the central lumen.
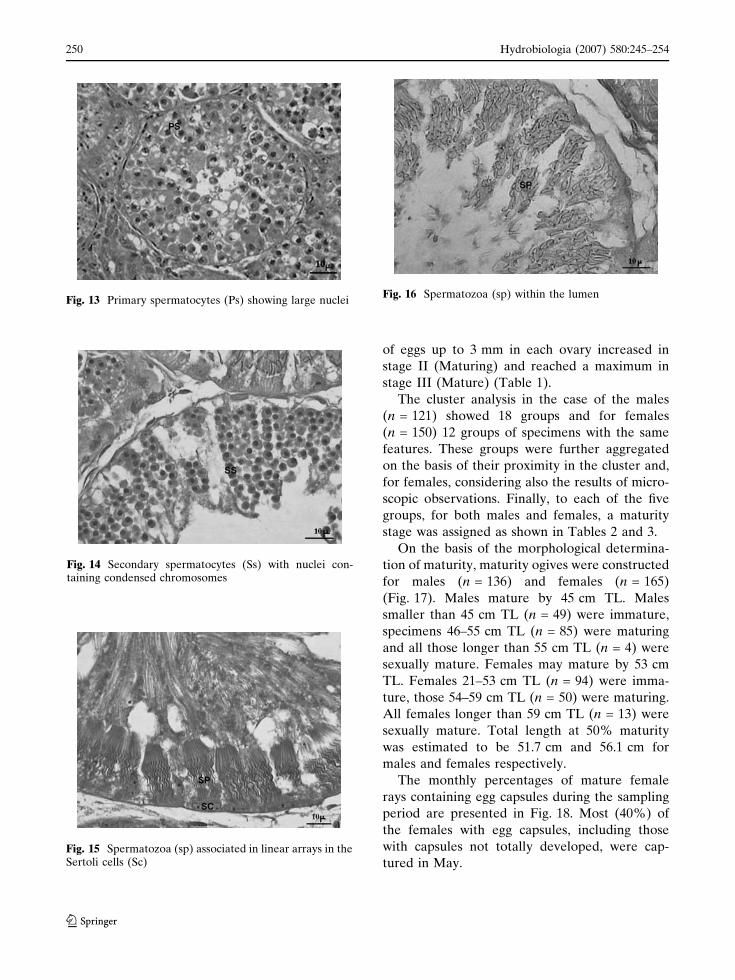
In the testes, six stages of spermatocyst devel-
opment were observed. At the beginning Sertoli
cells lined the lumen and a lesser number of
spermatogonia were in a peripherical position in
the spermatocyst (Fig. 11). In the second stage
the Sertoli cells were also found in peripherical
position just inside the basement membrane
(Fig. 12). Successively, the primary spermatocytes
showed large nuclei and in the fourth stage
secondary spermatocytes appeared with nuclei
Fig. 4 Large previtellogenic follicles, up to 1500 lm indiameter. The follicular epithelium is multilayered andmade up to small (SC), large (LC) and pyriform (PC) cells.Oo, oocyte; ZP, zona pellucida; BL, basal lamina
Fig. 3 Previtellogenic follicle ~100 lm in diameter. Thefollicular epithelium is double layered and made up tosmall (SC) and large (LC) cells. ZP, zona pellucida
Fig. 6 Nucleus (N) of oocyte 1600 lm in diametercontained lampbrush chromosomes (LC)
Fig. 5 Intercellular bridges (IB) link pyriform (PC) cellswith oocyte (Oo)
Fig. 7 Follicles up to 3000 lm filled with yolk platelets (Y)
248 Hydrobiologia (2007) 580:245–254
123

containing condensed chromosomes (Figs. 13,
14). The fifth stage was characterized by sper-
matids with ellipsoidal nuclei and emerging
flagella. In this stage, spermatozoa began to
aggregate around the periphery of the spermato-
cyst to form tight bundles (Fig. 15). In the
terminal stage of spermatogenesis, spermatozoa
were present in the lumen (Fig. 16).
The diameter of oocytes contained in the
ovaries of 118 specimens were measured and
distributed in classes of 150 lm. The diameters
were correlated to the maturity stage previously
assigned. Table 1 shows the presence of oocytes
up to 3 mm in diameter, in which it was supposed
that vitellogenesis had begun. The mean number
Fig. 8 Follicles up to 3000 lm filled with yolk platelets (Y)
Fig. 9 Atretic previtellogenic follicle. Granulosa cells (C)and theca cells (TC) are hypertrophic
Fig. 10 Postovulatory follicle. The basal lamina (BL)appears collapsed and invades the central lumen (L)
Fig. 11 Sertoli cell nuclei (Sc) migrating toward theperiphery
Fig. 12 Sertoli cells (Sc) in peripherical position justinside the basement membrane
Hydrobiologia (2007) 580:245–254 249
123
of eggs up to 3 mm in each ovary increased in
stage II (Maturing) and reached a maximum in
stage III (Mature) (Table 1).
The cluster analysis in the case of the males
(n = 121) showed 18 groups and for females
(n = 150) 12 groups of specimens with the same
features. These groups were further aggregated
on the basis of their proximity in the cluster and,
for females, considering also the results of micro-
scopic observations. Finally, to each of the five
groups, for both males and females, a maturity
stage was assigned as shown in Tables 2 and 3.
On the basis of the morphological determina-
tion of maturity, maturity ogives were constructed
for males (n = 136) and females (n = 165)
(Fig. 17). Males mature by 45 cm TL. Males
smaller than 45 cm TL (n = 49) were immature,
specimens 46–55 cm TL (n = 85) were maturing
and all those longer than 55 cm TL (n = 4) were
sexually mature. Females may mature by 53 cm
TL. Females 21–53 cm TL (n = 94) were imma-
ture, those 54–59 cm TL (n = 50) were maturing.
All females longer than 59 cm TL (n = 13) were
sexually mature. Total length at 50% maturity
was estimated to be 51.7 cm and 56.1 cm for
males and females respectively.
The monthly percentages of mature female
rays containing egg capsules during the sampling
period are presented in Fig. 18. Most (40%) of
the females with egg capsules, including those
with capsules not totally developed, were cap-
tured in May.Fig. 15 Spermatozoa (sp) associated in linear arrays in theSertoli cells (Sc)
Fig. 14 Secondary spermatocytes (Ss) with nuclei con-taining condensed chromosomes
Fig. 13 Primary spermatocytes (Ps) showing large nucleiFig. 16 Spermatozoa (sp) within the lumen
250 Hydrobiologia (2007) 580:245–254
123

Table 1 Number ofoocytes in females ofR. asterias at differentmaturity stages
Stage No. offemales
No. ofoocytes
Mean numberoocytes <3 mm/female
Mean numberoocytes >3 mm/female
I 71 6371 86 6II 19 1794 74 21III 11 1852 135 33IV 11 1599 117 27V 6 637 84 16
Table 2 Maturity scale for females of R. asterias
Stage State Description
Macroscopic Microscopic
I Immature Oocytes uniformly small and white. Ovary verysmall. Oviducts thread-like and nidamental glandabsent or little differentiated.
The anterior region of the ovary containingnumerous follicles in first phase ofprevitellogenesis, up to 150 lm in diameter.Atretic follicles present.
II Maturing Ovary small, oocytes differentiatedto various small sizes but all white. Oviducts andnidamental gland well developed but small(<2 mm maximum width).
Sections appear filled with previtellogenic folliclesof various sizes.
III Mature Ovary walls more transparent. Ovary filled withyellow Oocytes, some large. Oviducts welldeveloped and nidamental gland large(>2 mm maximum width).
Sections of oocytes of 3–7 mm show yolk droplets.
IV Extruding Ovary filled with large yellow oocytes. Egg capsulesmore or less formed in one or both oviducts.
Only the follicular walls were embedded because oflarge dimension of oocytes.Yolk droplets evident in the cortical region.
V Resting Ovary walls transparent. Oocytes ofdifferent sizes, white or yellow. Oviducts emptybut much enlarged and vascularized.Nidamental gland small (<2 mm).
Postovulatory follicles observed.
Table 3 Maturity stagesfor males of R. asterias
Stage State Description
I Immature Clasper shorter or as long as the extreme tips of posterior pelvic finlobes. Ducti deferentes not differentiated or narrow. Testes small andwhite occupying up to the half of abdominal cavity.
II Virgin Clasper longer than tips of posterior pelvic fin lobes but skeleton stillflexible. Ducti deferentes well developed. Testes occupying over thehalf of abdominal cavity with walls transparent but seminiferousfollicles not fill the whole testes.
III Maturing Clasper longer than tips of posterior pelvic fin lobes, skeleton hardenedbut axial cartilages still soft. Ducti deferentes whitish. Testes enlarged,not entirely filled with seminiferous follicles.
IV Mature Clasper longer than tips of posterior pelvic fin lobes, skeleton hardenedwith axial cartilages hardened and pointed. Ducti deferentes full.Testes wide, rose-coloured, with seminiferous follicles filling wholevolume.
V Resting Clasper longer than tips of posterior pelvic fin lobes, skeleton hardenedwith axial cartilages hardened and pointed. Testes scarcely occupyinghalf of abdominal cavity.
Hydrobiologia (2007) 580:245–254 251
123

The monthly abundances of females of R.
asterias at different stages of maturity is presented
in Fig. 19. The greatest number (80%) of females
at stage I (Immature) was observed in August.
This percentage diminished from September to
December, when the specimens at stage II increased
until the 50%. Stage III (Mature) stays almost
constant throughout the year, but stage IV (Extrud-
ing) were mainly observed in May and June, when it
reached the 20 %, decreasing in autumn.
Discussion
The analysis of histological maturity in female of
R. asterias indicates that both ovaries contain
developing follicles of various sizes. In all the
ovaries examined, the great majority of follicles
was in the phase of previtellogenesis, character-
ized by the progressive growth of the oocytes and
the increase in complexity of the follicular
epithelium. The change observed in the follicular
epithelium, during the early previtellogenic stage,
corresponds to that described by Andreuccetti
et al. (1999) in the same species. In addition, an
evidence of the phase of previtellogenesis is the
presence of chromosome with lampbrush appear-
ance in the nucleus of oocytes 50–1500 lm in
diameter. References to lampbrush chromosomes
in elasmobranch oocytes have not been found,
although the lampbrush chromosomes were de-
scribed by Guraya (1986) in the ovary of several
teleosts, in which they appear during the first
meiotic prophase. Furthermore, vitellogenis starts
when the follicle measures about 3000 lm in
diameter and is indicated by the appearance of
minute platelets of yolk in the cortical region. We
have been unable to describe the successive
phases of oogenesis because of the large size of
the oocytes. Instead, the count of oocytes allowed
us to observe that only few follicles continue with
vitellogenesis; therefore, the ovary of R. asterias
seems to contain a reserve of oocytes, present
both at every maturity stage and throughout the
year.
A brief analysis of the histology of testes
indicates that spermatogenesis of R. asterias is
similar to that described by Hamlett (1999) in
Urolophus jamaicensis. In the testes spermato-
cysts at different stages of development were
present, in accordance with the statement that the
gonads of rajids continuously produce reproduc-
tive gametes (Dodd, 1983).
Fig. 17 Maturity ogives of R. asterias
Fig. 19 Running average (2ra3) of maturity stages inR. asterias
Fig.18 Percentage and running average (2ra3) of egg-capsules in females of R. asterias
252 Hydrobiologia (2007) 580:245–254
123

Gross morphology of the reproductive tracts of
females at any time, supported by the histology,
facilitate the assignment of maturity stages, and
indicate that the process of maturation is both
progressive and gradual. In males only qualitative
analysis of the gonads were done; however, the
assignment of maturity stages was based on
accurate observations not only of claspers, as
was done by Capape (1977), but also of gonads
and sperm ducts.
The maturity stages both for females and males
resulting from the cluster analysis were numer-
ous. However, this method was used to eliminate
the subjectivity of the current maturity scales
(Stehmann, 2002) postponing the classification
into different stages until separate examination of
each feature of the reproductive tracts had been
carried out. The first three groups resulting from
the clustering were joined together in stage I
(Immature) both in males and females. Between
females classified as immature, the differences
consisted in the appearance of oviducts and
nidamental gland during growth; in all cases the
oocytes are small and white. In males, the
appearance of sperm ducts and the start of the
development of the gonads were the reasons
determining the different groups in the cluster. In
addition, in females, stage III (Mature) is char-
acterized by two phases in which the oocytes grow
and become yellow, contemporary the nidamen-
tal gland reaches a large size. Another finding,
resulting from the application of cluster analysis,
was the stage of resting hypothesized for both
males and females and rarely described for
elasmobranchs.
In females, at the onset of maturity, the ovaries
contain vitellogenic oocytes and the nidamental
gland is of medium dimension; in males claspers
are longer than the tips of pelvic fins, axial
cartilages are pointed, and gonads contain sem-
iniferous ampullae filling the whole volume. The
estimation of size at first sexual maturity (Lm) for
males (51.7 cm) is somewhat discordant from that
estimated by Capape (1977) (54 cm); also in the
case of females the estimated value of Lm
(56.1 cm) is lower than that calculated by Capape
(1977) and Tortonese (1956) (60 cm). These
differences could be related to environmental
and/or geographical aspects (Frisk et al., 2001).
In evaluating the reproductive cycle, the low
number of specimens reaching lengths at 50%
maturity, 36 females and 46 males, was insuffi-
cient to estimate the gonado-somatic indexes for
each month. From the occurrence of mature
females and females with egg-capsules during the
sampling period, it is concluded that the main
period of spawning activity occurs from March to
July. On the basis of these results, the tables of
maturity stages provide a more reliable tool for
describing rajid life history data.
Acknowledgements We wish to express our thanks toProf. Giorgio Mancino (Sezione di Biologia cellulare edello sviluppo del Dipartimento di Fisiologia eBiochimica—Universita di Pisa) for his advice onhistological investigations. We are very grateful toAlvaro Abella and Romano Baino (Agenzia Regionaleper la Protezione Ambientale della Toscana) for theiruseful support in processing the data. Many thanks also toCecilia Mancusi for her help and encouragement. We owespecial thanks to Caroline Bennett (Fao-AdriaMed) forher assistance with the translation of this paper.
References
Abella, A. J. & F. Serena, 2005. Comparison of elasmo-branch catches from Research Trawl Surveys andcommercial landings at port of Viareggio, Italy, in thelast decade. e-Journal of Northwest Atlantic FisheryScience V35: art. 23.
Abella, A., R. Auteri, R. Baino, A. Lazzeretti, P. Righini,F. Serena, R. Silvestri, A. Voliani & A. Zucchi, 1997.Reclutamento di forme giovanili nella fascia costieratoscana. Biologia Marina del Mediterraneo 4(1): 172–181.
Andreuccetti, P., M. Iodice, M. Prisco & R. Gualtieri,1999. Intercellular bridges between granulosa cellsand the oocyte in the elasmobranch R. asterias.Anatomical Record 255(2): 180–187.
Capape, C., 1977. Contribution a la biologie des Rajidaedes cotes tunisiennes. 4. Raja asterias Delaroche, 1809:repartition geographique et bathymetrique, sexualite,reprodution et fecondite. Bulletin du Museum d’His-toire Naturelle, Paris, 3e ser., 435, Zool., 305: 305–326.
Dodd, J. M., 1983. Reproduction in cartilagineus fishes(Chondrichthyes). In Hoar, W. S., D. J. Randall &E/M/ Donaldson (eds), Fish Physiology, Vol. 9.Academic Press, San Diego, pt A, 31–95.
Fischer, W., M. L. Bouchot & M. Schneider, (redacteurs)1987. Fishes FAO d’identification des especes pour lesbesoins de la peche. (Revision 1). Mediterranee etMer Noire. Zone de peche 37, Vol. II. Rome, FAO,Vertebres, 847–876.
Frisk, G. M., J. T. Miller & M. J. Fogarty, 2001. Estimationand analysis of biological parameters in elasmobranch
Hydrobiologia (2007) 580:245–254 253
123

fishes: a comparative life history study. CanadianJournal of Fisheries and Aquatic Science 58(5): 969–981.
Guraya, S. S., 1986. The Cell and Molecular Biology ofFish Oogenesis. Monographs in Developmental Biol-ogy, Vol. 18. Karger, Basel, 223 p.
Hamlett, W. C., 1999. Sharks, Skates and Rays: TheBiology of the Elasmobranchs Fishes. John HopkinsUniversity Press, Maryland, 515 pp.
Lo Bianco, S., 1908. Notizie biologiche riguardantispecialmente il periodo di maturita sessuale deglianimali del golfo di Napoli. Pubblicazione dellaStazione Zoologica di Napoli 19: 513–761.
MathSoft, 1999. S-PLUS User’s Guide. Data AnalysisProducts Division, MathSoft, Seattle, WA, 620 pp.
Musick, A. J., 1999. Criteria to define extinction risk inmarine fishes. The American Fisheries Society initia-tive. Fisheries 24(12): 6–14.
Relini, G., F. Biagi, F. Serena, A. Belluscio, M. T. Spedicato,P. Rinelli, M. C. Follesa, C. Piccinetti, N. Ungaro,L. Sion & D. Levi, 2000. I selaci pescati con lostrascico nei mari italiani. Biologia Marina delMediterranaeo 7(1): 347–384.
Serena, F., R. Baino & P. Righini, 1988. Geographical anddepth distribution of Rays in Northern Tyrrenian Sea.Commission Internationale pour l’Exploration Scien-tifique de la Mer Mediterranee 31(II): 277.
Simpson, A. C., 1951. The fecundity of the plaice. FisheriesInvestigation, Ministry of Agriculture, Fisheries andFood (G.B.), Ser. 11 17(5): 1–27.
Stehmann, M. F. W., 2002. Proposal of a maturity stagesscale for oviparous and viviparous cartilaginous fishes(Pisces, Chondrichthyes). Archive of Fishery andMarine Research 50(1): 23–48.
Stevens, J. D., R. Bonfil, N. K. Dulvy & P. A. Walker,2000. The effects of fishing on shark, rays andchimaeras (chondrichthyans), and implications formarine ecosystem. ICES Journal of Marine Science57: 476–494.
Tortonese, E., 1956. Leptocardia, Cyclostomata, Selachii.Fauna d’Italia, Vol. II. Calderini (ed.), Bologna,545 pp.
Whitehead, P. J. P., M. L. Bauchot, J. C. Hureau,J. Nielsen & E. Tortonese, 1984. Fishes of theNorth-Eastern Atlantic and Mediterranean (FNAM),Vol. I. UNESCO, Paris, 510 p.
254 Hydrobiologia (2007) 580:245–254
123




















