
A Flexible Bioreactor System for Constructing In Vitro Tissue and Organ Models

Eng. Life Sci. 2012, 12, No. 5, 1–10 1
Vasil Georgiev1
Ivan Ivanov1
Strahil Berkov2
Mladenka Ilieva1
Milen Georgiev1
Tatiana Gocheva1
Atanas Pavlov1,3
1Laboratory of AppliedBiotechnologies, The StephanAngeloff Institute ofMicrobiology, BulgarianAcademy of Sciences, Plovdiv,Bulgaria
2Department of Photochemistry,AgroBioInstitute, Sofia, Bulgaria
3Department of OrganicChemistry and Microbiology,University of Food Technologies,Plovdiv, Bulgaria
Research Article
Galanthamine production by Leucojumaestivum L. shoot culture in a modified bubblecolumn bioreactor with internal sections
Shoot culture of summer snowflake (Leucojum aestivum L.) was successfully cul-tivated in an advanced modified glass-column bioreactor with internal sectionsfor production of Amaryllidaceae alkaloids. The highest amounts of dry biomass(20.8 g/L) and galanthamine (1.7 mg/L) were achieved when shoots were cultured at22◦C and 18 L/(L·h) flow rate of inlet air. At these conditions, the L. aestivum shootculture possessed mixotrophic-type nutrition, synthesizing the highest amounts ofchlorophyll (0.24 mg/g DW (dry weight) chlorophyll A and 0.13 mg/g DW chloro-phyll B). The alkaloids extract of shoot biomass showed high acetylcholinesteraseinhibitory activity (IC50 = 4.6 mg). The gas chromatography–mass spectrome-try (GC/MS) profiling of biosynthesized alkaloids revealed that galanthamine andrelated compounds were presented in higher extracellular proportions while ly-corine and hemanthamine-type compounds had higher intracellular proportions.The developed modified bubble-column bioreactor with internal sections providedconditions ensuring the growth and galanthamine production by L. aestivum shootculture.
Keywords: Acetylcholinesterase inhibitors / Alkaloid profiling / Amaryllidaceae alkaloids /Column bioreactor / Cultivation conditions
Received: September 26, 2011; revised: February 3, 2012; accepted: March 21, 2012
DOI: 10.1002/elsc.201100177
1 Introduction
Amaryllidaceae alkaloids are known to exhibit a wide range ofbiological activities [1]. The best investigated galanthamine (anisoquinoline alkaloid), is long acting, selective, reversible, andcompetitive acetylcholinesterase inhibitor used for symptomatictreatment of senile dementia of the Alzheimer’s type [2], po-liomyelitis, and other neurological diseases [3, 4]. Another valu-able member of the Amaryllidaceae alkaloids family is lycorine (apyrrolophenanthridine alkaloid) that possesses a strong antivi-ral effect against poliovirus, measles, and Herpes simplex type1 viruses [5], as well as high antiretroviral [6], strong antimi-totic [7–10], and cytotoxic activities [11]. Nowadays, the galan-thamine is marketed as a hydrobromide salt under the name
Correspondence: Dr. Vasil Georgiev ([email protected]),Laboratory of Applied Biotechnologies, The Stephan Angeloff In-stitute of Microbiology, Bulgarian Academy of Sciences, 139 RuskiBlvd, 4000 Plovdiv, Bulgaria.
Abbreviations: DW, dry weight; GC/MS, gas chromatography–massspectrometry; IC50, half maximal inhibitory concentration; TIC, totalion current
Razadyne R© (formerly Reminyl R©) and Nivalin R© for the treat-ment of Alzheimer’s disease, poliomyelitis, and other neurolog-ical diseases [12]. Although chemical synthesis of galanthaminehas been successfully performed [13], the main commercialsources for the production of the galanthamine are both daf-fodils (Narcissus) and snowflake (Leucojum aestivum L.) [4, 14].Since 1989, prescription regime of the utilization of L. aestivumL. has been imposed and, from these perspectives, galanthamineproduction from in vitro cultures is considered as an attractivealternative. Nevertheless, the galanthamine has been found inmany Amaryllidaceae plants, only two plant species have beenstudied for their in vitro galanthamine production – Narcissusconfusus Pugsley and L. aestivum L. [12]. It was established thatthe cell differentiation is a prerequisite for galanthamine biosyn-thesis as the lowest levels of galanthamine have been found incalli [14–16]. Diop and coworkers [17] had success to obtainhairy roots from L. aestivum L. after Agrobacterium rhizogenestransformation using ultrasound methodology. However, thetransformed root culture does not express galanthamine. Shoot-clump cultures had a higher ability to synthesize this alkaloidas their biosynthetic capacity was strongly influenced from il-lumination [15], concentrations of the macronutrients [14, 18],growth regulators, and elicitation as well [19, 20]. On the basis
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

2 V. Georgiev et al. Eng. Life Sci. 2012, 12, No. 5, 1–10
of these studies, the optimal content of the nutrient medium forbiosynthesis of galanthamine by L. aestivum L. shoots cultivatedin liquid medium was proposed [18]. At the moment, primaryexperiments for developing biotechnological process for galan-thamine production based on temporary immersion technologyhas been performed [21]. However, for the correct estimation ofthe potential of in vitro systems for galanthamine production,different bioreactor-based processes should be investigated.
In the present study, for the first time we describe the pro-cess of galanthamine biosynthesis by L. aestivum L. shoot cul-ture in an illuminated bubble-column bioreactor with internalsections.
2 Materials and methods
2.1 Leucojum aestivum L. shoot culture
The shoot cultures were established by planting previously ob-tained calli [15] on Murashige and Skoog nutrient medium (MS)nutrient medium supplemented with 30 g/L sucrose, 5.5 g/L‘‘Plant agar’’, 1.15 mg/L 1-naphthylacetic acid (NAA, Duchefa,The Netherlands) and 2.0 mg/L 6-benzylaminopurine (BAP,Duchefa, The Netherlands). Single shoots, differentiated fromcallus tissue were propagated to give a raise of homogenous lines.The cultivation of the established shoot clumps was carried outat 26◦C under illumination 16:8 h (light:dark) at a light inten-sity of 110 μmol/(m2.s) - SYLVANIA Gro-Lux fluorescent lamp(F18W/GRO-LUX). The best growing and glanthamine produc-ing shoot line was L. aestivum 80. This line was selected for thenext experiments. It was subcultivated every 28 days under thesame conditions as described before.
2.2 Conditions of the bioreactor cultivation
For bioreactor cultivation of L. aestivum shoots, a new systembased on bubble-column bioreactor was developed (Fig. 1). Thereactor consisted of a glass column (length = 68 cm, internaldiameter = 5.5 cm), covered by glass jacket (length = 55 cm,diameter = 7 cm). The top of the column was designed withconical ground-in glass socket (50/55 mm) and was closed byconically tapered ground glass joint, equipped with a gas-outport. The bottom of the column was equipped with ceramicsparger and sampling port. For creating internal sections, sixpolypropylene copolymer (PPCO) rings (diameter = 5.3 cm,thickness = 2 mm) were attached to a glass rod (length =66 cm, diameter = 6 mm) forming five equal cultivation sec-tions (length = 8 cm).
The L. aestivum line 80 shoot culture was cultivated for 35days in modified bubble-column bioreactor with internal sec-tions (Fig. 1) filled with 1 L optimized Murashige and Skoognutrient medium (MS) medium [18] under illumination 16:8 h(light:dark) with light intensity of 110 μmol/(m2.s) - SYLVANIAGro-Lux fluorescent lamp (F18W/GRO-LUX). The optimizationof cultivation temperature was performed by using constant flowrate of inlet air of 18 L/(L·h) and different temperatures (18◦C,22◦C, and 26◦C). The experiments for optimization of flow rateof inlet air were performed at optimal temperature (22◦C). For
experiments, different cultivations at flow rate of inlet air of 9,18, and 27 L/(L·h) were performed in agreement to the experi-mental design. In all experiments, the bioreactor was inoculatedwith 60 g (fresh weight) of shoot culture grown for 28 days onsolid medium as described above.
2.3 Extraction of chlorophyll
After bioreactor cultivation, the biomass was collected andlyophilized. For determination of chlorophyll contents, about200 mg of this biomass were extracted three times with 5 mL80% (v/v) acetone for 10 min. Combined extracts were filteredusing Whatman 1 paper and used for analyses.
2.4 Extraction of alkaloids
2.4.1 Intracellular alkaloidsTwo hundred milligrams of lyophilized biomass were measuredand extracted directly with 5 mL methanol in an ultrasonic bathfor 15 min (three times). The combined extracts were concen-trated under vacuum and dissolved in 2 × 2 mL of 3% sulfuricacid. The neutral compounds were removed by extraction (threetimes) with diethyl ether. The alkaloids were fractionated afterbasification of the extracts with 1 mL of 25% ammonia and ex-traction with chloroform (3 × 3 mL). The chloroform extractswere then dried over anhydrous sodium sulfate and evaporated todryness.
2.4.2 Extracellular alkaloidsFifty milliliters of liquid medium were evaporated to dryness anddissolved in 10 mL methanol. After centrifugation and separationof pellets, 8 mL of supernatants were evaporated to dryness andresiduals were dissolved in 2 × 2 mL of 3% sulfuric acid andthen processed as described above.
2.5 Analyses
2.5.1 Dry biomassThe growth of the shoots was calculated by measurement of drybiomass, accumulated dry biomass (ADB), and the growth index(GI) [15].
2.5.2 Chlorophyll contentChlorophyll A and chlorophyll B were determined spectrophoto-metrically at 663 nm and 645 nm following the method describedby [22].
2.5.3 Galanthamine contentThe galanthamine quantification was performed on WatersHPLC system, equipped with Waters 1525 binary pump (Mil-ford, USA), a Dual λ Waters 2487 absorbance detector (Milford,USA) and Breeze 3.0 Breeze 3.30 SPA software by the methoddescribed previously [21].
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

Eng. Life Sci. 2012, 12, No. 5, 1–10 Galanthamine production by Leucojum aestivum L. shoot culture 3
Figure 1. Construction (A) and schematic diagram (B) of modified bubble-column bioreactor with internal sections: 1, bubble column withinternal sections; 2, fluorescent lamps; 3, filters; 4, sampling. Twenty-eight day of cultivation of L. aestivum line 80 shoot culture in modifiedbubble-column bioreactor with internal sections at 22◦C and flow rate of inlet air of 18 L/(L·h) (C).
2.5.4 gas chromatography–mass spectrometry(GC/MS) analysis
The GC-MS analyses were performed with a Hewlett Packard6890+/MSD 5975 (Hewlett Packard, Palo Alto, CA, USA) onHP-5 MS column (30 m × 0.25 mm × 0.25 μm) as describedpreviously [23].
The spectra of coeluting chromatographic peaks were exam-ined and deconvoluted by using AMDIS 2.6 (NIST, Gaithersburg,MD, USA) software before area integration. The contribution ofeach compound in the extracts is shown in Table 2 as a percent-age of the total ion current (TIC). The alkaloids were identifiedby comparing their mass spectral fragmentations with those of
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

4 V. Georgiev et al. Eng. Life Sci. 2012, 12, No. 5, 1–10
Table 1. Chlorophyll content at the end of cultivation of L. aes-tivum 80 shoot culture in bubble-column bioreactor with internalsections. The presented values are means ± SD (n = 4).
Cultivation Chlorophyll A Chlorophyll Bconditions (mg/g DW) (mg/g DW)
18◦C 18 L/(L·h) 0.19 ± 0.01 0.10 ± 0.0122◦C 9 L/(L·h) 0.21 ± 0.02 0.12 ± 0.0122◦C 18 L/(L·h) 0.24 ± 0.01 0.13 ± 0.0122◦C 27 L/(L·h) 0.19 ± 0.01 0.09 ± 0.0126◦C 18 L/(L·h) 0.21 ± 0.01 0.11 ± 0.01
DW, dry weight.
standard references from NIST 08 database (NIST Mass SpectralDatabase, PC-Version 5.0 (2008), National Institute of Stan-dardization and Technology, Gaithersburg, MD, USA) and/or asdescribed literature data [24, 25].
The area of the GC-MS peaks depends not only on the con-centration of the corresponding compounds but also on theintensity of their mass spectral fragmentation. The presenteddata do not express absolute values (do not represent a truequantification) but can be used for comparison of the samples,obtained during different cultivation regimes.
2.5.5 Inhibition of the acetylcholinesteraseThe acetylcholinesterase inhibition assays were performed spec-trophotometrically at 405 nm according to the method describedby [26], modified as described previously [23]. The results werepresented as inhibition rate (%).
2.5.6 Sugars assaySucrose, glucose, and fructose in the culture medium were de-termined by means of enzyme test combination (Megazyme,Ireland, Cat. No.: K-SUFRG).
2.5.7 Inorganic ions assayNitrate, ammonium, and phosphate ions were determinedby chemical test combinations (MERCK, Germany, Cat. No.1.14773.0001, 1.14752.0001, 1.14842.0001)
2.6 Statistical analyses
The software package MINITAB 14 was used for the regressionanalysis of the process, assessment of the obtained experimentalresults, and for model’s development using the response surfacemethodology. The MINITAB 14 software also was used for theoptimization of the regression model for galanthamine biosyn-thesis.
The results presented in the study have been summarizedfrom two independent experiments, repeated twice.
3 Results
3.1 Bioreactor cultivation for galanthamineproduction by L. aestivum L. shoot culture.Effects of temperature and flow rate of inlet air
For the experiment, modified version of bubble-column biore-actor was used (Fig. 1). This bioreactor system was developedto cover the specific requirements, raised by the morpholog-ical characteristics of L. aestivum L. shoots. The allocation ofthe initial inoculum into the five internal sections completelyprevents the shoots flotation and improves the mass transfer be-tween biomass and culture medium. During cultivation in thisbioreactor system, the L. aestivum L. shoots showed balancedgrowth at all tested regimes. They visibly increased their lineardimensions and meristem cells differentiated in new shoots aswell.
Both the temperature of cultivation and the flow rate of in-let air are the most important variables that could influence onbiomass accumulation and galanthamine biosynthesis by L. aes-tivum L. shoot culture during its cultivation in bubble-columnbioreactor with internal sections. The results are presented onFigs. 2 and 3.
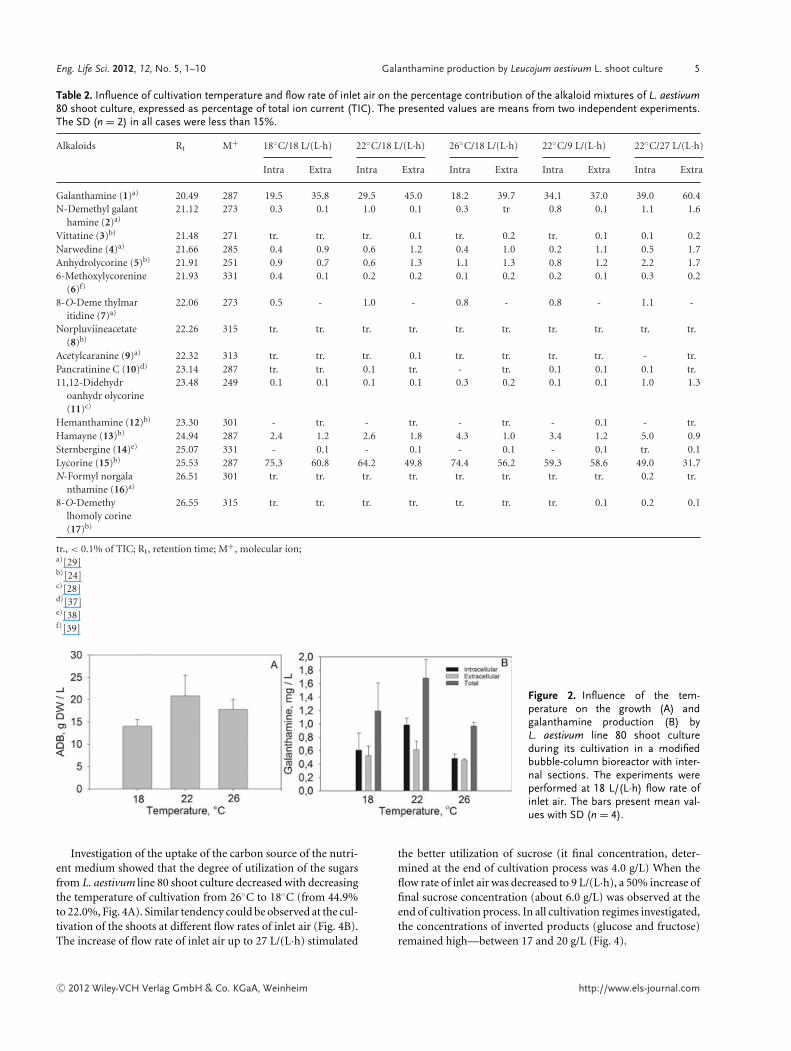
The highest amount of dry biomass (20.8 g/L) L. aestivumline 80 shoot culture accumulated during its cultivation at 22◦C.At the same temperature, the highest GI (4.8) was also achieved.Increased and decreased temperatures of cultivation led to re-duction of accumulated shoot dry biomass (14.0 g/L at 18◦C and17.8 g/L at 26◦C, respectively) (Fig. 2A). The maximal galan-thamine production by L. aestivum line 80 shoots cultivatedin modified bubble-column bioreactor (1.7 mg/L, consist of 1.0mg/L intracellular and 0.7 mg/L extracellular galanthamine) wasachieved at the optimal temperature for biomass accumulation(Fig. 2B). When the cultivation temperature was increased to26◦C, the galanthamine production decreased with 43% (0.96mg/L) and the amounts of both intracellular and extracellulargalanthamine were equal (0.48 mg/L). The biomass and galan-thamine productions were also found to be affected by the valuesof the flow rate of inlet air (Fig. 3). The achieved results showedthat the flow rate of inlet air had comparable effect to this of thetemperature. The best results for the growth of L. aestivum line 80shoot culture during its cultivation in modified bubble-columnbioreactor with internal sections was achieved at 22◦C and 18L/(L·h) flow rate of inlet air. At this regime of cultivation, themaximal production of galanthamine (1.7 mg/L) was achievedas well, whereas the decrease or increase of inlet air flow ratesreduced the galanthamine production by 32% and 63%, respec-tively (Fig. 3B). It should be noted, that simultaneously withthe maximal galanthamine production, the L. aestivum line 80shoot culture biosynthesized the highest levels of other majoralkaloid—lycorine (8.3 mg/L, consists of 6.3 mg/L intracellularand 2.0 mg/L extracellular lycorine). When cultured at opti-mal conditions [22◦C and 18 L/(L·h) air flow rate] the highestamounts of chlorophyll (0.24 mg/g DW (dry weight) chlorophyllA and 0.13 mg/g DW chlorophyll B, respectively—Table 1) wasregistered in shoots biomass as well. This could be considered asindirect confirmation for the balanced growth of the investigatedplant in vitro system.
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com
Eng. Life Sci. 2012, 12, No. 5, 1–10 Galanthamine production by Leucojum aestivum L. shoot culture 5
Table 2. Influence of cultivation temperature and flow rate of inlet air on the percentage contribution of the alkaloid mixtures of L. aestivum80 shoot culture, expressed as percentage of total ion current (TIC). The presented values are means from two independent experiments.The SD (n = 2) in all cases were less than 15%.
Alkaloids Rt M+ 18◦C/18 L/(L·h) 22◦C/18 L/(L·h) 26◦C/18 L/(L·h) 22◦C/9 L/(L·h) 22◦C/27 L/(L·h)
Intra Extra Intra Extra Intra Extra Intra Extra Intra Extra
Galanthamine (1)a) 20.49 287 19.5 35.8 29.5 45.0 18.2 39.7 34.1 37.0 39.0 60.4N-Demethyl galant
hamine (2)a)21.12 273 0.3 0.1 1.0 0.1 0.3 tr 0.8 0.1 1.1 1.6
Vittatine (3)b) 21.48 271 tr. tr. tr. 0.1 tr. 0.2 tr. 0.1 0.1 0.2Narwedine (4)a) 21.66 285 0.4 0.9 0.6 1.2 0.4 1.0 0.2 1.1 0.5 1.7Anhydrolycorine (5)b) 21.91 251 0.9 0.7 0.6 1.3 1.1 1.3 0.8 1.2 2.2 1.76-Methoxylycorenine
(6)f)21.93 331 0.4 0.1 0.2 0.2 0.1 0.2 0.2 0.1 0.3 0.2
8-O-Deme thylmaritidine (7)a)
22.06 273 0.5 - 1.0 - 0.8 - 0.8 - 1.1 -
Norpluviineacetate(8)b)
22.26 315 tr. tr. tr. tr. tr. tr. tr. tr. tr. tr.
Acetylcaranine (9)a) 22.32 313 tr. tr. tr. 0.1 tr. tr. tr. tr. - tr.Pancratinine C (10)d) 23.14 287 tr. tr. 0.1 tr. - tr. 0.1 0.1 0.1 tr.11,12-Didehydr
oanhydr olycorine(11)c)
23.48 249 0.1 0.1 0.1 0.1 0.3 0.2 0.1 0.1 1.0 1.3
Hemanthamine (12)b) 23.30 301 - tr. - tr. - tr. - 0.1 - tr.Hamayne (13)b) 24.94 287 2.4 1.2 2.6 1.8 4.3 1.0 3.4 1.2 5.0 0.9Sternbergine (14)e) 25.07 331 - 0.1 - 0.1 - 0.1 - 0.1 tr. 0.1Lycorine (15)b) 25.53 287 75.3 60.8 64.2 49.8 74.4 56.2 59.3 58.6 49.0 31.7N-Formyl norgala
nthamine (16)a)26.51 301 tr. tr. tr. tr. tr. tr. tr. tr. 0.2 tr.
8-O-Demethylhomoly corine(17)b)
26.55 315 tr. tr. tr. tr. tr. tr. tr. 0.1 0.2 0.1
tr., < 0.1% of TIC; Rt, retention time; M+, molecular ion;a)[29]b)[24]c)[28]d)[37]e)[38]f)[39]
Figure 2. Influence of the tem-perature on the growth (A) andgalanthamine production (B) byL. aestivum line 80 shoot cultureduring its cultivation in a modifiedbubble-column bioreactor with inter-nal sections. The experiments wereperformed at 18 L/(L·h) flow rate ofinlet air. The bars present mean val-ues with SD (n = 4).
Investigation of the uptake of the carbon source of the nutri-ent medium showed that the degree of utilization of the sugarsfrom L. aestivum line 80 shoot culture decreased with decreasingthe temperature of cultivation from 26◦C to 18◦C (from 44.9%to 22.0%, Fig. 4A). Similar tendency could be observed at the cul-tivation of the shoots at different flow rates of inlet air (Fig. 4B).The increase of flow rate of inlet air up to 27 L/(L·h) stimulated
the better utilization of sucrose (it final concentration, deter-mined at the end of cultivation process was 4.0 g/L) When theflow rate of inlet air was decreased to 9 L/(L·h), a 50% increase offinal sucrose concentration (about 6.0 g/L) was observed at theend of cultivation process. In all cultivation regimes investigated,the concentrations of inverted products (glucose and fructose)remained high—between 17 and 20 g/L (Fig. 4).
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

6 V. Georgiev et al. Eng. Life Sci. 2012, 12, No. 5, 1–10
Figure 3. Influence of the flow rateof inlet air on the growth (A)and galanthamine production (B) byL. aestivum line 80 shoot cultureduring its cultivation in a modifiedbubble-column bioreactor with inter-nal sections. The experiments wereperformed at 22◦C. The bars presentmean values with SD (n = 4).
Figure 4. Final sugar concentrationsand degree of utilization (*) fromL. aestivum line 80 shoot culture cul-tivated in modified bubble-columnbioreactor with internal sections atdifferent temperatures (A) and flowrates of inlet air (B). The bars presentmean values with SD (n = 4).
The temperature of cultivation did not influence the uti-lization of nitrate ions from L. aestivum line 80 shoot culture(Fig. 5A). At all tested temperature levels, about 60% of ni-trate ions were utilized from the culture medium. However,it influenced on the utilization of phosphate and ammoniumions. Maximal degrees of utilization (99% for phosphate ionsand 95% for ammonium ions) were achieved at the cultivationat a temperature of 22◦C and 18 L/(L·h) flow rate of inlet air(Fig. 5). At lower temperature of cultivation (18◦C) L. aestivumline 80 shoot culture utilized 62% and 70% of supplied phos-phate and ammonium ions, respectively. At higher temperatureof cultivation (26◦C) 86% of phosphate and 93% of ammoniumions were exhausted from the culture medium (Fig. 5A).
The flow rate of inlet air had stronger effect on the uptakeof main inorganic ions from the culture medium, comparedto the effect of temperature (Fig. 5B). The maximal degrees ofutilization were achieved at 18 L/(L·h) inlet air. Both increasedand decreased flow rate of inlet air leaded to lower activity ofcatabolism of the investigated ions.
3.2 gas chromatography–mass spectrometry(GC/MS) profiling
Seventeen alkaloids of galanthamine, lycorine, homolycorine,and hemanthamine types were detected in the studied sam-ples (Tables 2 and 3). Intra and extracellular alkaloid patternswere similar. The only differences have been observed in com-positions of minor compounds. For example, compound 7 wasfound in the intracellular alkaloid fractions while compounds12 and 14 were found mainly in the extracellular ones. Thealkaloid patterns were dominated by lycorine type, followed bygalanthamine-type compounds with principal alkaloids lycorine(15) and galanthamine (1), respectively.
With respect to the intracellular alkaloid patterns, the highestgalanthamine proportion, 39% of total alkaloids, was measuredat 22◦C with 27 L/(L·h) flow rate of inlet air, which is higher thanthose measured at 18◦C or 26◦C (20% and 18%, respectively).The extracellular galanthamine proportions followed similartrend. In contrast to the galanthamine content, which highest
Figure 5. Degree of utilization ofammonium, nitrate, and phos-phate ions from L. aestivum line 80shoot culture cultivated in a modi-fied bubble-column bioreactor withinternal sections at different tem-peratures (A) and flow rates of inletair (B). The bars present mean val-ues with SD (n = 4).
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

Eng. Life Sci. 2012, 12, No. 5, 1–10 Galanthamine production by Leucojum aestivum L. shoot culture 7
Table 3. Influence of the cultivation temperature and the flow rate of inlet air on the distribution of the alkaloid types in alkaloid mixturesof L. aestivum 80 shoot culture, expressed as percentage of total ion current (TIC). The presented values are means from two independentexperiments. The SD (n = 2) in all cases were less than 15%.
Types alkaloids 18◦C/18 L/ 22◦C/18 L/ 26◦C/18 L/ 22 ◦C/9 L/ 22◦C/27 L/(L·h) (L·h) (L·h) (L·h) (L·h)
Intracellular Extracellular Intracellular Extracellular Intracellular Extracellular Intracellular Extracellular Intracellular Extracellular
1. Galanthamine 20.2 36.8 31.1 46.3 18.9 40.7 35.1 38.2 40.8 63.72. Lycorine 76.3 61.7 64.9 51.4 75.8 58.8 60.2 60.0 52.2 34.83. Homolycorine 0.4 0.1 0.2 0.3 0.1 0.2 0.2 0.2 0.5 0.34. Hemanthamine 2.9 1.2 3.6 1.9 5.1 1.2 4.2 1.4 6.2 1.1
amounts (1.7 mg/L, 29.5% of total alkaloids) were achieved atoptimal for the growth and photosynthesis flow rate of inlet air[18 L/(L·h)], the higher galanthamine proportions were foundat flow rates of inlet air of 27 L/(L·h) (39% of total alkaloids)and 9 L/(L·h) (34.1% of total alkaloids), (Table 2). In general, thegalanthamine proportion in the alkaloid fractions from medium(extracellular galanthamine) was higher than those found inthe plant tissues. The opposite trend was observed for lycorine(Tables 2 and 3). The alternations of the alkaloid types due tothe culture conditions followed those of the principal alkaloids,lycorine, and galanthamine (Table 3).
3.3 Acetylcholinesterase inhibitory action of alkaloidextracts
The activities of intracellular and extracellular alkaloid extractsto inhibit the activity of acetylcholinesterase were investigatedduring the cultivation of L. aestivum line 80 shoot culture inmodified bubble-column bioreactor with internal sections atdifferent temperatures and flow rates of inlet air. The analy-ses showed that the shoot biomass extracts have higher acetyl-cholinesterase inhibitory activities than the alkaloid fractionsfrom the culture media (Table 4). The biomass extracts fromL. aestivum line 80 shoot culture grown at 18◦C and 26◦C, bothwith flow rate of 18 L/(L·h) showed low acetylcholinesterase in-hibitory activities (IC50 = 19.5 and IC50 = 10.4 mg dry biomass,respectively). At these conditions, the extracts were with thelower galanthamine proportions (19.5 and 18.2 of total alkaloids,respectively), but the lycorine proportions were higher (75.3%
Table 4. Acetylcholinesterase inhibitory activity of alkaloid mix-tures of L. aestivum 80 shoot culture, cultivated in glass-bubblecolumn bioreactor. The presented values are means ± SD (n =4).
Cultivation IC50, IC50,conditions mg dry biomass mL culture medium
18◦C 18 L/(L·h) 19.5 ± 1.8 64.3 ± 5.422◦C 9 L/(L·h) 6.5 ± 0.1 58.9 ± 6.122◦C 18 L/(L·h) 4.6 ± 0.9 49.9 ± 1.022◦C 27 L/(L·h) 9.2 ± 0.3 45.1 ± 4.726◦C 18 L/(L·h) 10.4 ± 0.7 37.9 ± 4.7Galanthamine 0.31 ± 0.05 -Lycorine Not active -
and 74.4% of total alkaloids, respectively) (Table 2). When theshoot culture was cultivated at 22◦C with different flow rates [9,18, and 27 L/(L·h)] and galanthamine proportions in them wereraised (34.1%, 29.5%, and 39% of total alkaloids, respectively),the acetylcholinesterase inhibitory activities of alkaloid extractswere increased (IC50 = 6.5, IC50 = 4.6, and IC50 = 9.2 mg drybiomass, respectively) (Tables 2 and 4).
3.4 Statistical optimization of galanthamineproduction
Based on the achieved results concerning influence of the tem-perature of cultivation and the flow rate of inlet air (investigatedindependent variables) on the galanthamine biosinthesis byL. aestivum line 80 shoot culture, the statistical regression modelwas obtained, taking into account the influence of the tempera-ture (X1) and flow rate of inlet air (X2) on the amount of totalproduced galanthamine (Y, equation 1).
Y = −18.5 + 1.63X 1 + 0.31X 2 − 0.04X 21 − 0.01X 2
2 (1)
The received coefficient of determination (R2 = 73.3) wasgood enough taking into account the degree of differentiation ofthe investigated plant in vitro systems.
The optimization procedures carried out using “Responseoptimizer” of MINITAB 14 software gave the following values ofvariable X1 and X2 for maximum biosynthesis of galanthamine(Y) by L. aestivum line 80 shoot culture:
X ∗1 = 21.6; X ∗
2 = 16.5; Y∗max = 1.71mg/L
The deviation between theoretically studied maximalamounts of galanthamine and experimentally obtained (at tem-perature of cultivation 22◦C and flow rate of inlet air 18 L/(L·h)was only 0.03 mg/L (1.7%). On this basis, we propose temper-ature of cultivation 22◦C and flow rate of inlet air 18 L/(L·h)as optimal for galanthamine production by L. aestivum line 80shoot culture in modified bubble-column bioreactor with inter-nal sections.
4 Discussion
Nowadays, the production of plant-derived substances is lim-ited by environmental, ecological, and climatic conditions. Thus,
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

8 V. Georgiev et al. Eng. Life Sci. 2012, 12, No. 5, 1–10
there are likely to be shortages of some products due to combinedpressures of limitations of supplies [27]. The galanthamine is amolecule which is biosynthesized by L. aestivum L., a speciesunder protection in Bulgaria [28] and in several European coun-tries [4]. The problem, associated with industrial production ofthis important medicine could be overcome using plant in vitrotechnologies.
Leucojum aestivum line 80 shoot culture was obtained viacallus 5 years ago [15]. Further, the nutrient medium for max-imal galanthamine yields has been optimized [18] and the pro-files of the both extra and intracellular alkaloid fractions of theL. aestivum L. shoots have been investigated as well [28, 29].
As far as the critical point for the successful industrial ex-ploitation of plant cells growing in vitro is the up scaling, a specialattention should be paid on the choice of the most appropriatebioreactor system for each production process individually. Ourprevious investigations [21] showed that the temporary immer-sion is a prospective technology for galanthamine productionby L. aestivum L. shoots; however, there are few but very im-portant limitations: large-scale temporary immersion apparatusis not available; the labor cost during the exploitation of suchcultivation is too high. It is well known that the improvement ofexisted bioreactor designs or development of new constructionsis an essential step in development of effective biotechnologi-cal processes for secondary metabolite production [30]. Becauseof these circumstances, we estimated the potential of bubble-column bioreactor with regard to galanthamine production. Thecritical point for the success of the cultivations was developmentof the system with internal sections (Fig. 1). With this system,the problems with floatation of the shoots were overcome anda disturbance of the internal stream ways of the fluids (mediumand air) was not observed. The rings of internal sections weredesigned with five “large” and five “small” holes (Fig. 1), whichallowed good mixing in the bioreactor and at the same timeprevented floatation of the plant material. The design of thismodified bioreactor system is very simple and is susceptible forfurther scale up.
The growth of the L. aestivum shoot line 80 was fast. Theaccumulated amount of dry biomass (20.8 g/L) at the optimalconditions of cultivation was 1.7-fold higher than the maximalamounts achieved at the cultivation in temporary immersionRITA system [21]. The achieved doubling time (175 h) and GI(4.8) were also higher than these, achieved at temporary immer-sion conditions of cultivation (Td = 282 h and GI = 3.0).
Comparing the galanthamine biosynthesis by L. aestivumline 80 shoot culture cultivated in a modified bubble-columnbioreactor with internal sections and in a temporary immer-sion system [21], it should be noted that at the optimal condi-tions of both cultivation systems, the shoots synthesized 1.3-foldhigher amounts of galanthamine in the modified bubble column(1.7 mg/L vs. 1.3 mg/L, respectively). This is very good result ascompared to previously reported data concerning galanthaminecontents in collectable areal parts of wild growing L. aestivum L.plants (0.120 mg/g fresh weight and 0.078 mg/g fresh weight forleaves and stems, respectively [28]).
All discussed advantages for galanthamine production byL. aestivum line 80 shoots in modified bubble column with inter-nal sections were probably due to better hydrodynamic environ-ment in the cultivation vessel, which leaded to better availability
of the main nutrients of the medium at the optimal conditions ofcultivation [22◦C and 18 L/(L·h) flow rate of inlet air]. At theseconditions L. aestivum line 80 shoots completely utilized phos-phate, 95% of ammonium and 60% of nitrate ions (Fig. 4). At thesame time, only 33.5% of carbon source was used, which clearlyunderlined that L. aestivum line 80 shoot culture possessed amixotrophic-type nutrition during its cultivation in modifiedbubble-column bioreactor with internal sections and thereforesynthesized amounts of chlorophyll pigments were higher thanthese synthesized at temporary immersion conditions. More-over, our previous investigations showed that the biosynthesis ofgalanthamine is related to photosynthesis [28].
The results from the conducted experiments in modifiedbubble-column bioreactor clearly showed that the alkaloid pat-terns, biomass accumulation, and chlorophyll synthesis, wereinfluenced by both temperature of cultivation and flow rate ofinlet air. While the highest galanthamine yields coincided withthe optimal growth conditions for biomass and chlorophyll accu-mulation [22◦C, 18 L/(L·h)], higher both intra and extracellulargalanthamine proportions were found at the optimal tempera-ture but at the lower and higher flow rates.
The principal alkaloids, lycorine, and galanthamine show dif-ferent ability to be released into the medium by the tissue cul-tures. Galanthamine and related compounds showed higher ex-tracellular proportions while lycorine and hemanthamine-typecompounds, on the contrary, had higher intracellular propor-tions.
It is worth of remark that beside pharmaceutically valuablegalanthamine, L. aestivum line 80 shoot culture synthesized manyrelated alkaloids that could be of pharmacological importance.Moreover, the analyses of acetylcholinesterase inhibitory activ-ities of intracellular and extracellular alkaloid fractions showedthat there were synergistic effects of galanthamine and the otherconcomitant alkaloids, especially lycorine (Tables 2 and 4). Theseobservations support the conclusion of Orchan et al. [31] thatlycorine might contribute to anti-acetylcholinesterase effect ofthe mixed alkaloid extracts, but it is not directly responsiblefor the mentioned activity. Lycorine (15) is of huge interestbecause of its strong biological and pharmacological activities[5, 6, 8–10, 32–35]. Hamayne (13) possesses cytostatic and anti-cancer activities [36]. Other alkaloids identified in minor per-centages of TIC: 8-O-demethylmaritidine (7), pancratinine C(10); hemanthamine (12) and sternbergine (14), recently werereported to possess strong cytostatic and anticancer activities[9,10,36]. Vittatine (3) and anhydrolycorine (5) possess antibac-terial activity [33] and improve the analgesic effect of morphine[35], and anhydrolycorine (5) possesses anticancer activity [9].
5 Concluding remarks
In conclusion, as a result of our investigations, we propose thebubble column bioreactor with internal sections as suitable cul-tivation system for galanthamine production by L. aestivumshoot cultures. This bioreactor system provided good nutri-ent and mass transfer conditions and ensures the maximal cul-ture growth and galanthamine production by L. aestivum line80 shoot culture. The bioreactor design is suitable for devel-opment of large volume vessels for further upscaling studies.
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

Eng. Life Sci. 2012, 12, No. 5, 1–10 Galanthamine production by Leucojum aestivum L. shoot culture 9
The higher galanthamine proportions in extracellular fractionsrevealed the possibilities for its recovery from the culturemedium which will additionally increase the productivity of thesystem. Beside galanthamine production, the presented resultsclearly underlined that the L. aestivum line 80 shoot culture, cul-tivated in this type of bioreactor in optimized nutrient mediumis a good biological matrix for producing related Amaryllidaceaealkaloids, possessing valuable biological and pharmacologicalactivities. Some of these alkaloids have strong synergistic effectson acetylcholinesterase inhibitory activity of galanthamine andtheir action should be investigated in more details.
Practical Application
In this manuscript, we describe for the first time the biosyn-thesis of galanthamine by Leucojum aestivum L. shoot cul-ture in a newly developed glass-column bioreactor with in-ternal sections. Galanthamine is an isoquinoline alkaloid,which has been widely used for symptomatic treatmentof senile dementia of the Alzheimer’s type, poliomyeli-tis, and other neurological diseases. Optimization of cul-tivation conditions including the influence of temperatureand the flow rate of inlet air was performed. Multimetabo-lite analyses of biosynthesized alkaloids as well as acetyl-cholinesterase inhibitory activity of produced alkaloid mix-tures were done for each cultivation regime. The analy-ses showed that galanthamine and related compoundswere secreted into culture medium, whereas lycorine andhemanthamine-type compounds were accumulated mainlyin shoot biomass. The presented results enable new per-spectives in bioreactor designs, as well as allow more ef-fective cultivation of shoot cultures, which could be usedfor development of large-scale galanthamine productionprocesses.
This research has been supported by National Science Fund ofBulgaria under contract number DO-02/105 - 2009.
The authors have declared no conflict of interest.
6 References
[1] Jin, Z., Amaryllidaceae and Sceletium alkaloids. Nat. Prod. Rep.2011, 28, 1126–1142.
[2] Erdogan Orhan, I., Orhan, G., Gurkas, E., An overview onnatural cholinesterase inhibitors - a multi-targeted drug class- and their mass production. Mini Rev. Med. Chem. 2011, 11,836–842.
[3] Ptak, A., El Tahchy, A., Dupire, F., Boisbrun, M. et al., LCMSand GCMS for the screening of alkaloids in natural and in vitroextracts of Leucojum aestivum. J. Nat. Prod. 2009, 72, 142–147.
[4] Parolo, G., Abeli, T., Rossi, G., Dowgiallo, G. et al., Biologicalflora of Central Europe: Leucojum aestivum L. Perspect. PlantEcol. Evol. Syst., 2011, 13, 319–330.
[5] Li, S. Y., Chen, C., Zhang, H. Q., Guo, H. Y. et al., Identificationof natural compounds with antiviral activities against SARS-associated coronavirus. Antiviral Res. 2005, 67, 18–23.
[6] Szlavik, L., Gyuris, A., Minarovits, J., Forgo, P. et al., Al-kaloids from Leucojum vernum and antiretroviral activityof Amaryllidaceae alkaloids. Planta Med. 2004, 70, 871–873.
[7] Liu, J., Hu, W. X., He, L. F., Ye, M. et al., Effects of lycorineon HL-60 cells via arresting cell cycle and inducing apoptosis.FEBS Lett. 2004, 578, 245–250.
[8] Liu, X. S., Jiang, J., Jiao, X. Y., Wu, Y. E. et al., Lycorine inducesapoptosis and down-regulation of Mcl-1 in human leukemiacells. Cancer Lett. 2009, 274, 16–24.
[9] Lamoral-Theys, D., Andolfi, A., Van Goietsenoven, G.,Cimmino, A. et al., Lycorine, the main phenanthridine Amaryl-lidaceae alkaloid, exhibits significant antitumor activity in can-cer cells that display resistance to proapoptotic stimuli: an in-vestigation of structure-activity relationship and mechanisticinsight. J. Med. Chem. 2009, 52, 6244–6256.
[10] Lamoral-Theys, D., Decaestecker, C., Mathieu, V., Dubois, J.et al., Lycorine and its derivatives for anticancer drug design.Mini Rev. Med. Chem. 2010, 10, 41–50.
[11] Weniger, B., Italiano, L., Beck, J. P., Bastida, J. et al., Cytotoxicactivity of Amaryllidaceae alkaloids. Planta Med. 1995, 61, 77–79.
[12] Berkov, S., Georgieva, L., Kondakova, V., Atanasov, A. et al.,Plant sources of galanthamine: phytochemical and biotechno-logical aspects. Biotechnol. & Biotechnol. Eq. 2009, 23, 1170–1176.
[13] Magnus, P., Sane, N., Fauber, B. P., Lynch, V., Concise synthe-ses of (−)-galanthamine and (±)-codeine via intramolecularalkylation of a phenol derivative. J. Am. Chem. Soc. 2009, 131,16045–16047.
[14] Selles, M., Bergonon, S., Viladomat, F., Bastida, J. et al., Effectof sucrose on growth and galanthamine production in shoot-clump cultures of Narcissus confusus in liquid-shake medium.Plant Cell Tissue Organ Cult. 1997, 49, 129–136.
[15] Pavlov, A., Berkov, S., Courot, E., Gocheva, T. et al., Galan-thamine production by Leucojum aestivum in vitro systems.Process Biochem. 2007, 42, 734–739.
[16] Ptak, A., Tahchy, A., Wyzgolik, G., Henry, M., Laurain-Mattar,D., Effects of ethylene on somatic embryogenesis and galan-thamine content in Leucojum aestivum L. cultures. Plant CellTissue Organ Cult. 2010, 102, 61–67.
[17] Diop, M. F., Hehn, A., Ptak, A., Chretien, F. et al., Hairyroot and tissue cultures of Leucojum aestivum L. - relation-ships to galanthamine content. Phytochem. Rev. 2007, 6, 137–141.
[18] Georgiev, V., Berkov, S., Georgiev, M., Burrus, M. et al., Opti-mized nutrient medium for galanthamine production in Leu-cojum aestivum L. in vitro shoot system. Z. Naturforsch. C. 2009,64, 219–224.
[19] Colque, R., Viladomat, F., Bastida, J., Codina, C., Improvedproduction of galanthamine and related alkaloids by methyljasmonate in Narcissus confusus shoot-clumps. Planta Med.2004, 70, 1180–1188.
[20] El Tahchy, A., Bordage, S., Ptak, A., Dupire, F. et al., Effectsof sucrose and plant growth regulators on acetylcholinesteraseinhibitory activity of alkaloids accumulated in shoot cultures
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com

10 V. Georgiev et al. Eng. Life Sci. 2012, 12, No. 5, 1–10
of Amaryllidaceae. Plant Cell Tissue Organ Cult. 2011, 106,381–390.
[21] Ivanov, I., Georgiev, V., Georgiev, M., Ilieva, M. et al.,Galanthamine and related alkaloids production by Leucojumaestivum L. shoot culture using a temporary immersion tech-nology. Appl. Biochem. Biotechnol. 2011, 163, 268–277.
[22] Bajracharya, D., Experiments in Plant Physiology: A LaboratoryManual, Narosa Publishing House, New Delhi 2003, pp. 51–52.
[23] Georgiev, V., Ivanov, I., Berkov, S., Pavlov, A., Alkaloids biosyn-thesis by Pancratium maritimum L. shoots in liquid culture.Acta Physiol. Plant. 2011, 33, 927–933.
[24] Berkov, S., Bastida, J., Viladomat, F., Codina, C., Analysis ofgalanthamine-type alkaloids by capillary gas chromatography–mass spectrometry in plants. Phytochem. Anal. 2008, 19, 285–293.
[25] Torras-Claveria, L., Berkov, S., Jauregui, O., Caujape, J. et al.,Metabolic profiling of bioactive Pancratium canariense extractsby GC-MS. Phytochem. Anal. 2010, 21, 80–88.
[26] Perry, N. S. L., Noughton, P. J., Theobald, A. E., Jenner, P. et al.,In-vitro inhibition of human erythrocyte acetylcholine esteraseby Salvia lavandulaefolia essential oil and constituent terpenes.J. Pharm. Pharmacol. 2000, 52, 895–902.
[27] Pavlov, A., Plant cells and algae in bioreactors. Eng. Life Sci.2009, 9, 154–155.
[28] Berkov, S., Pavlov, A., Georgiev, V., Bastida, J. et al., Alka-loid synthesis and accumulation in Leucojum aestivum in vitrocultures. Nat. Prod. Commun. 2009, 4, 359–364.
[29] Berkov, S., Pavlov, A., Ilieva, M., Burrus, M. et al., CGC-MSof alkaloids in Leucojum aestivum plants and their in vitrocultures. Phytochem. Anal. 2005, 16, 98–103.
[30] Bley, T., Editorial: Bioprocess-oriented plant design – turning
basic research into practical applications. Eng. Life Sci. 2011,11, 333–334.
[31] Orhan, I. E., Yilmaz, B. S., Altun, M. L., Saltan, G. et al., Anti-acetylcholinesterase and antioxidant appraisal of the bulb ex-tracts of five Sternbergia species. Rec. Nat. Prod. 2011, 5, 193–201.
[32] Sener, B., Orhan, I., Satayavivad, J., Antimalarial activityscreening of some alkaloids and the plant extracts fromAmaryllidaceae. Phytother. Res. 2003, 17, 1220–1223.
[33] Evidente, A., Andolfi, A., Abou-Donia, A. H., Touema, S. M.et al., (-)-Amarbellisine, a lycorine-type alkaloid from Amaryl-lis belladonna L. growing in Egypt. Phytochemistry 2004, 65,2113–2118.
[34] Li, Y., Liu, J., Tang, L. J., Shi, Y. W. et al., Apoptosis inducedby lycorine in KM3 cells is associated with the G0/G1 cell cyclearrest. Oncol. Rep. 2007, 17, 377–384.
[35] Kornienko, A., Evidente, A., Chemistry, biology, and medicinalpotential of narciclasine and its congeners. Chem. Rev. 2008,108, 1982–2014.
[36] Campbell, W. E., Nair, J. J., Gammon, D. W., Codina, C. et al.,Bioactive alkaloids from Brunsvigia radulosa. Phytochemistry2000, 53, 587–591.
[37] Cedron, J., Oberti, J., Estevez-Braun, A., Ravelo, A. et al., Pan-cratium canariense as an important source of Amaryllidaceaealkaloids. J. Nat. Prod. 2009, 72, 112–116.
[38] Evidente, A., Iasiello, I., Randazzo, G., Isolation of sternbergine,a new alkaloid from bulbs of Sternbergia lutea. J. Nat. Prod.1984, 47, 1003–1008.
[39] Kreh, M., Matusch, R., Witte, L., Capillary gaschromatography-mass spectrometry of Amaryllidaceaealkaloids. Phytochemistry 1995, 38, 773–776.
C© 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim http://www.els-journal.com
Copyright © 2022 FDOKUMEN




















