
G α i2 , G α i3 and G α o are all Required for Normal Muscarinic Inhibition of the Cardiac...
11
J Mol Cell Cardiol 31, 1771–1781 (1999) Article No. jmcc.1999.1015, available online at http://www.idealibrary.com on Ga i2 ,Ga i3 and Ga o are all Required for Normal Muscarinic Inhibition of the Cardiac Calcium Channels in Nodal/ Atrial-like Cultured Cardiocytes Chianping Ye 1, *, Margaret O. Sowell 1, * , Peter M. Vassilev 1 , David S. Milstone 2 and Richard M. Mortensen 1 1 Endocrine-Hypertension Division, Department of Medicine and 2 Vascular Research Division, Department of Pathology, Brigham and Women’s Hospital and Harvard Medical School, Boston, MA 02115, USA (Received 18 December 1998, accepted in revised form 21 June 1999) C. Y , M. O. S, P. M. V , D. S. M R. M. M.Ga i2 ,Ga i3 and Ga o are all Required for Normal Muscarinic Inhibition of the Cardiac Calcium Channels in Nodal/Atrial-like Cultured Cardiocytes. Journal of Molecular and Cellular Cardiology (1999) 31, 1771–1781. The cardiac -type calcium current (I Ca,L ) is an important regulator of myocardial contractility. It is activated by sympathetic stimulation and inhibited by parasympathetic activity via muscarinic acetylcholine receptors. Muscarinic inhibition of I Ca,L occurs via activation of pertussis toxin (PTX)-sensitive heterotrimeric G-proteins. Although recent studies have shown that expression of G oa is important for this effect in adult mouse ventricular cells, two other PTX-sensitive G-proteins (G i2 and G i3 ) are also expressed in cardiocytes and are activated. Their role in the regulation of I Ca,L has not been examined. In addition, it is not known whether nodal/atrial cardiac cells use the same G-proteins. We show that gene inactivation of each of the three PTX-sensitive G a -proteins (a i2 , a i3 , and a o ) affects muscarinic inhibition of cardiac I Ca,L in embryonic stem (ES) cell-derived cardiocytes. Inactivation of either a i2 or a i3 markedly slows the time course of muscarinic inhibition of I Ca,L , and in cells where both a i2 and a i3 are inactivated the effects are not additive. We also establish an essential role for a o in this atrial/nodal-like cardiocyte system and show that a o acts proximal to NO generation. NO generation plays a critical role in I Ca,L regulation since the nitric oxide synthase (NOS) antagonist, -NMMA, blocked the inhibition of I Ca,L in WT and in a i2 /a i3 -null cells. In WT cells, the NO generating agent SIN-1 inhibited I Ca,L and the addition of carbachol resulted in faster inhibition, suggesting that pathways in addition to NO are also activated. This study shows that a i2 and a i3 play a critical role in the normal inhibition of cardiocyte I Ca,L . Thus, all muscarinic receptor activated G-proteins (G i2 ,G i3 and G o ) are necessary for normal inhibition and act through both NO and non-NO signaling pathways. 1999 Academic Press K W: G-proteins; Calcium channels; Isolated cardiocytes; Signal transduction; Muscarinic receptors; ES cells. cascade involving pertussis toxin (PTX)-sensitive Introduction G-proteins (Robishaw and Hansen, 1994). PTX- sensitive G-proteins also couple to b2-adrenergic Cardiac muscarinic receptors regulate the activity of a number of ion channels via a signaling receptors (Xiao et al., 1995; Daaka et al., 1997) Please address all correspondence to: Richard M. Mortensen, Endocrine-Hypertension Division, 221 Longwood Avenue, Boston, MA 02115, USA. * C. Ye and M. O. Sowell contributed equally to this work. Current address: Endocrine and Metabolism Section, Department of Medicine, Indiana University Medical Center, Indianapolis, IN. 0022–2828/99/091771+11 $30.00/0 1999 Academic Press
Transcript of G α i2 , G α i3 and G α o are all Required for Normal Muscarinic Inhibition of the Cardiac...

J Mol Cell Cardiol 31, 1771–1781 (1999)
Article No. jmcc.1999.1015, available online at http://www.idealibrary.com on
Gai2, Gai3 and Gao are all Required forNormal Muscarinic Inhibition of theCardiac Calcium Channels in Nodal/Atrial-like Cultured CardiocytesChianping Ye1,∗, Margaret O. Sowell1,∗†, Peter M. Vassilev1,David S. Milstone2 and Richard M. Mortensen1
1Endocrine-Hypertension Division, Department of Medicine and 2Vascular Research Division,Department of Pathology, Brigham and Women’s Hospital and Harvard Medical School, Boston,MA 02115, USA
(Received 18 December 1998, accepted in revised form 21 June 1999)
C. Y, M. O. S, P. M. V, D. S. M R. M. M. Gai2, Gai3 and Gao are all Requiredfor Normal Muscarinic Inhibition of the Cardiac Calcium Channels in Nodal/Atrial-like Cultured Cardiocytes.Journal of Molecular and Cellular Cardiology (1999) 31, 1771–1781. The cardiac -type calcium current (ICa,L) isan important regulator of myocardial contractility. It is activated by sympathetic stimulation and inhibited byparasympathetic activity via muscarinic acetylcholine receptors. Muscarinic inhibition of ICa,L occurs via activationof pertussis toxin (PTX)-sensitive heterotrimeric G-proteins. Although recent studies have shown that expressionof Goa is important for this effect in adult mouse ventricular cells, two other PTX-sensitive G-proteins (Gi2 andGi3) are also expressed in cardiocytes and are activated. Their role in the regulation of ICa,L has not been examined.In addition, it is not known whether nodal/atrial cardiac cells use the same G-proteins. We show that geneinactivation of each of the three PTX-sensitive Ga-proteins (ai2, ai3, and ao) affects muscarinic inhibition of cardiacICa,L in embryonic stem (ES) cell-derived cardiocytes. Inactivation of either ai2 or ai3 markedly slows the timecourse of muscarinic inhibition of ICa,L, and in cells where both ai2 and ai3 are inactivated the effects are notadditive. We also establish an essential role for ao in this atrial/nodal-like cardiocyte system and show that ao
acts proximal to NO generation. NO generation plays a critical role in ICa,L regulation since the nitric oxidesynthase (NOS) antagonist, -NMMA, blocked the inhibition of ICa,L in WT and in ai2/ai3-null cells. In WT cells,the NO generating agent SIN-1 inhibited ICa,L and the addition of carbachol resulted in faster inhibition, suggestingthat pathways in addition to NO are also activated. This study shows that ai2 and ai3 play a critical role in thenormal inhibition of cardiocyte ICa,L. Thus, all muscarinic receptor activated G-proteins (Gi2, Gi3 and Go) arenecessary for normal inhibition and act through both NO and non-NO signaling pathways.
1999 Academic Press
K W: G-proteins; Calcium channels; Isolated cardiocytes; Signal transduction; Muscarinic receptors;ES cells.
cascade involving pertussis toxin (PTX)-sensitiveIntroductionG-proteins (Robishaw and Hansen, 1994). PTX-sensitive G-proteins also couple to b2-adrenergicCardiac muscarinic receptors regulate the activity
of a number of ion channels via a signaling receptors (Xiao et al., 1995; Daaka et al., 1997)
Please address all correspondence to: Richard M. Mortensen, Endocrine-Hypertension Division, 221 Longwood Avenue, Boston, MA02115, USA.∗ C. Ye and M. O. Sowell contributed equally to this work.† Current address: Endocrine and Metabolism Section, Department of Medicine, Indiana University Medical Center, Indianapolis, IN.
0022–2828/99/091771+11 $30.00/0 1999 Academic Press

C. Ye et al.1772
and the cardiac suppressive effects may sig- taneously contracting nodal or atrial-like cellsexpress a number of cardiac specific ion channelsnificantly contribute to decreases in cardiac func-
tion. It is therefore important to define the including the -type calcium channel (Maltsev etal., 1993, 1994).signaling pathways through these G-proteins that
alter heart function. In the heart, three different In rabbit sinoatrial and atrioventricular nodalcells, muscarinic receptor regulation of ICa,L in-PTX-sensitive a subunits are expressed: ai2, ai3,
and ao. Muscarinic receptors have been shown volves a signaling cascade in which nitric oxide(NO) is generated (Han et al., 1996, 1997).to interact with all three a subtypes (Luetje et
al., 1987). Recent studies suggest that generation Inhibitors of nitric oxide synthase (NOS), such as-NMMA, block the inhibition of ICa,L by carbacholof specific responses in cardiocytes requires ex-
pression of specific G-protein a subtypes. In mouse while agents such as SIN-1, that spontaneouslygenerate NO, can inhibit ICa,L. We therefore ex-embryonic stem (ES) cell-derived cardiocytes, we
have shown that muscarinic receptor activation amined the effect of agents that modify the NOsignaling system on ICa,L responses in the ES cell-of the cardiac atrial type K+ channel, IKACh,
requires ai2 and ai3 but not ao (Sowell et al., derived cardiocytes from WT and a-null cells.Our data demonstrate important roles for ai21997). In adult ventricular cardiocytes isolated
from ao-null mice, an essential role for ao in and ai3 in determining the kinetics of inhibition.In addition, we show that expression of ao ismuscarinic receptor regulation of -type cardiac
calcium currents (ICa,L) but not IKACh was dem- essential for muscarinic receptor inhibition ofICa,L in nodal/atrial-like cells as it is in ventricularonstrated (Valenzuela et al., 1997).
Unlike modulation of IKACh activity where Gbc cells, and that the production of NO is importantas previously shown in rabbit nodal cardiocytes.protein interaction directly regulates channel ac-
tivity, G-protein modulation of -type calciumchannel activity is believed to occur primarily viasecond messengers (Mery et al., 1993; Campbelland Strauss, 1995). b-adrenergic receptors (bAR) Materials and Methodsstimulate ICa,L via a Gs initiated cascade thatinvolves activation of adenylyl cyclase, increases Cell lines and culturein cAMP, activation of PKA and phosphorylationof the channel. Muscarinic receptors use PTX- Wild type and mutant (ai2-, ai3- and ao-null) D3-ES
cells were maintained and differentiated as de-sensitive G-proteins (Gi or Go) to antagonize thebAR effect on ICa,L through activation of nitric scribed previously (Sowell et al., 1997). D3-ES cells
were targeted with constructs designed to inactivateoxide synthase and generation of nitric oxide(NO). Whether direct inhibition of adenylyl cyclase the endogenous ai2, ai3 and ao genes by inserting a
neomycin resistance (neo) gene in a required exon.via Gi/Go subunits or activation of phosphatasesby muscarinic receptors contribute substantially Heterozygous mutant lines were identified by South-
ern blot, expanded and selected in high con-to the anti-adrenergic effect on ICa,L remainsuncertain (Mery et al., 1993; Campbell and centration G418, which we have previously shown
enriches for homozygous diploid targeted linesStrauss, 1995).To fully characterize the G-protein requirements (Mortensen et al., 1992). Surviving clones that
were homozygous for the targeted allele werefor muscarinic receptor inhibition of the cardiacICa,L, we examined the ability of carbachol to identified by disappearance of the wild type band
in Southern analysis. Western analysis confirmedinhibit isoproterenol stimulated currents in EScell-derived cardiocytes lacking ai2, ai3, or ao. The inactivation of the corresponding genes.
To generate the cell line lacking both ai2 and ai3,panel of mutant ES cell lines was generated bytargeting specific a subunit genes. Muscarinic the previously produced ai3-null cell line (Sowell et
al., 1997) was transfected with the hygromycinreceptor signaling was then examined after dif-ferentiation of wild type and mutant cell lines to containing ai2 construct interrupting exon 6 (Mor-
tensen et al., 1991). Heterozygous ai2 recombinantcardiocytes. The ES cell-derived cardiocytes areparticularly useful for studies of receptor signaling clones were then further selected at high hy-
gromycin-B concentrations (1.0 mg/l) to isolatepathways as they recapitulate many characteristicsof native cardiocytes more closely than cardiac- homozygous null ai2/ai3 cells (Mortensen, 1993a,
b). After high concentration antibiotic selection,like cell lines (Kline et al., 1993). Extensiveelectrophysiological characterization of ES cell- clones were identified by Southern blotting to have
both ai2 alleles interrupted.derived cardiocytes has shown that these spon-
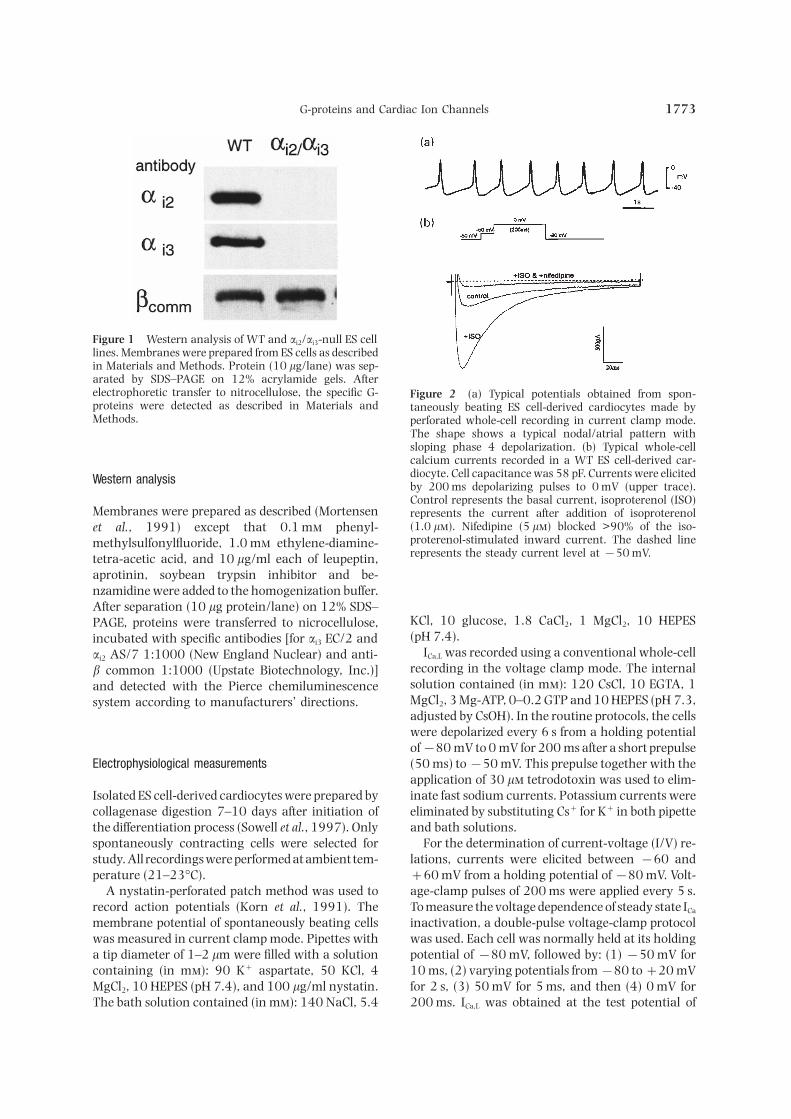
G-proteins and Cardiac Ion Channels 1773
Figure 1 Western analysis of WT and ai2/ai3-null ES celllines. Membranes were prepared from ES cells as describedin Materials and Methods. Protein (10 lg/lane) was sep-arated by SDS–PAGE on 12% acrylamide gels. Afterelectrophoretic transfer to nitrocellulose, the specific G- Figure 2 (a) Typical potentials obtained from spon-proteins were detected as described in Materials and taneously beating ES cell-derived cardiocytes made byMethods. perforated whole-cell recording in current clamp mode.
The shape shows a typical nodal/atrial pattern withsloping phase 4 depolarization. (b) Typical whole-cellcalcium currents recorded in a WT ES cell-derived car-diocyte. Cell capacitance was 58 pF. Currents were elicitedWestern analysis by 200 ms depolarizing pulses to 0 mV (upper trace).Control represents the basal current, isoproterenol (ISO)
Membranes were prepared as described (Mortensen represents the current after addition of isoproterenol(1.0 l). Nifedipine (5 l) blocked >90% of the iso-et al., 1991) except that 0.1 m phenyl-proterenol-stimulated inward current. The dashed linemethylsulfonylfluoride, 1.0 m ethylene-diamine-represents the steady current level at −50 mV.tetra-acetic acid, and 10 lg/ml each of leupeptin,
aprotinin, soybean trypsin inhibitor and be-nzamidine were added to the homogenization buffer.After separation (10 lg protein/lane) on 12% SDS–
KCl, 10 glucose, 1.8 CaCl2, 1 MgCl2, 10 HEPESPAGE, proteins were transferred to nicrocellulose,(pH 7.4).incubated with specific antibodies [for ai3 EC/2 and
ICa,L was recorded using a conventional whole-cellai2 AS/7 1:1000 (New England Nuclear) and anti-recording in the voltage clamp mode. The internalb common 1:1000 (Upstate Biotechnology, Inc.)]solution contained (in m): 120 CsCl, 10 EGTA, 1and detected with the Pierce chemiluminescenceMgCl2, 3 Mg-ATP, 0–0.2 GTP and 10 HEPES (pH 7.3,system according to manufacturers’ directions.adjusted by CsOH). In the routine protocols, the cellswere depolarized every 6 s from a holding potentialof−80 mV to 0 mV for 200 ms after a short prepulse(50 ms) to−50 mV. This prepulse together with theElectrophysiological measurementsapplication of 30 l tetrodotoxin was used to elim-inate fast sodium currents. Potassium currents wereIsolated ES cell-derived cardiocytes were prepared by
collagenase digestion 7–10 days after initiation of eliminated by substituting Cs+ for K+ in both pipetteand bath solutions.the differentiation process (Sowell et al., 1997). Only
spontaneously contracting cells were selected for For the determination of current-voltage (I/V) re-lations, currents were elicited between −60 andstudy. All recordings were performed at ambient tem-
perature (21–23°C). +60 mV from a holding potential of−80 mV. Volt-age-clamp pulses of 200 ms were applied every 5 s.A nystatin-perforated patch method was used to
record action potentials (Korn et al., 1991). The To measure the voltage dependence of steady state ICa
inactivation, a double-pulse voltage-clamp protocolmembrane potential of spontaneously beating cellswas measured in current clamp mode. Pipettes with was used. Each cell was normally held at its holding
potential of −80 mV, followed by: (1) −50 mV fora tip diameter of 1–2 lm were filled with a solutioncontaining (in m): 90 K+ aspartate, 50 KCl, 4 10 ms, (2) varying potentials from−80 to+20 mV
for 2 s, (3) 50 mV for 5 ms, and then (4) 0 mV forMgCl2, 10 HEPES (pH 7.4), and 100 lg/ml nystatin.The bath solution contained (in m): 140 NaCl, 5.4 200 ms. ICa,L was obtained at the test potential of

C. Ye et al.1774
Figure 3 Electrophysiological characterizations of WT, ao-null and ai2/ai3-null ES cell-derived cardiocytes. (a) Thecurrent density-voltage relationship (I/V curve) for each cell type is shown under basal conditions (Χ), after additionof isoproterenol (ISO, 1.0 l) (Β), and isoproterenol with carbachol (Carb, 10 l) (Φ). The currents were normalizedto the individual cell’s membrane capacitance, shown as pA/pF. Symbols and bars denote means±SEM (n=4–6). (b)Two-pulse experimental protocol is described in Materials and Methods. Steady-state inactivation curves of calciumcurrents were plotted as I/IMax (at 0 mV) against the voltages of the conditioning pulses. Data were fitted by Boltzmanndistribution equation (solid lines). Symbols and bars denote means±SEM (n=3–4).
0 mV by measuring the difference between peak in- preincubated with NGmonomethyl--arginine (-NMMA, 0.1 m) for 30 min prior to the experimentward current and the leak current at the end of the
2-s pulse. and -NMMA was continuously present throughoutthe recording period.The time dependent response to isoproterenol or
carbachol was determined using the routine voltage- Cell membrane capacitance (Cm) was measured bya voltage ramp (−80 to−60 mV) with a slope (dV/clamp protocol. Current amplitudes were obtained
by subtracting the steady current level at−50 mV dt) of 20 V/s. Cm was calculated as Cm=(Iramp−Iss)/(dV/dt) where Iramp is the current at the end of thefrom the peak current at 0 mV. After a stable whole-
cell configuration formed, the basal ICa,L was recorded ramp pulse and Iss is the steady state level of whole-cell current at−80 mV.for 4 min, isoproterenol (1.0 l) was added to the
bath solution and recordings continued for 5 min.Carbachol (10 l) or molsidomine (SIN-1, 0.1 m)was then added and the recordings continued for at Calculations and statisticsleast 6 min. The isoproterenol and carbachol werewashed out from the bath solution, ICa,L was recorded Data analysis used Graphpad Prism software on a
Macintosh G3 computer. The steady state in-for a few minutes, and isoproterenol was then re-added to assure that there was no significant current activation data were fitted with Boltzmann dis-
tribution equation. The potential eliciting half-rundown. For studying the role of NO, cells were
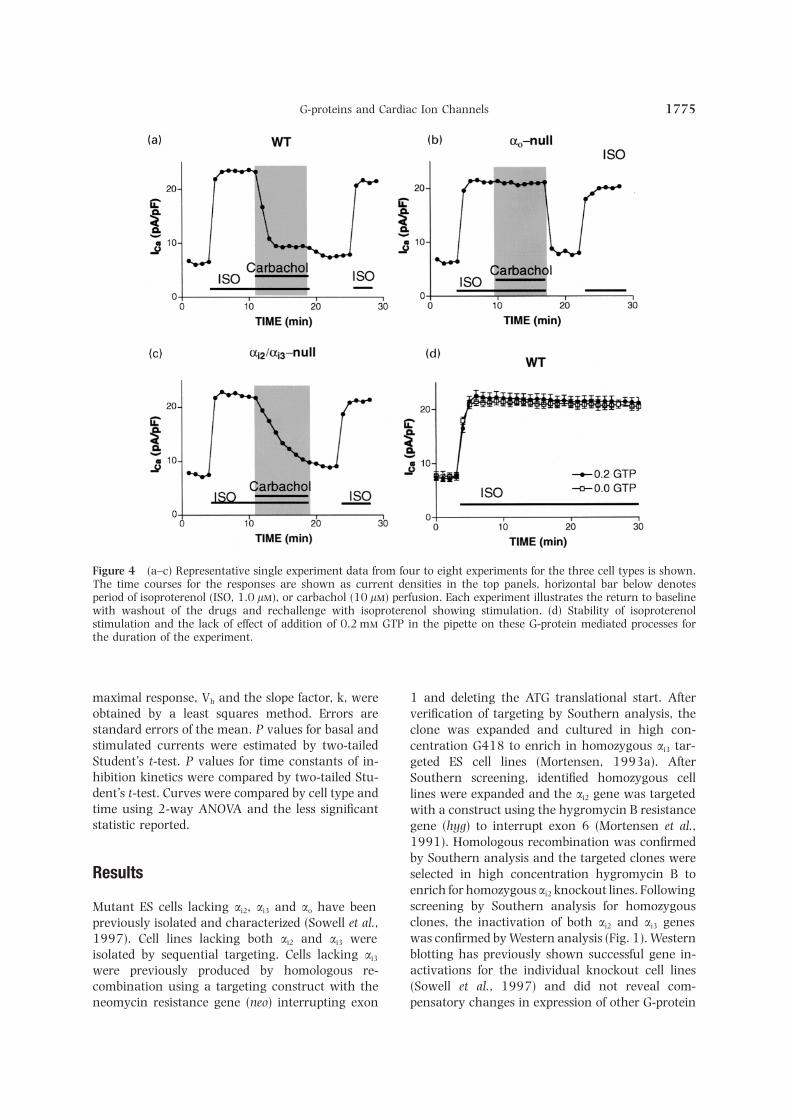
G-proteins and Cardiac Ion Channels 1775
Figure 4 (a–c) Representative single experiment data from four to eight experiments for the three cell types is shown.The time courses for the responses are shown as current densities in the top panels, horizontal bar below denotesperiod of isoproterenol (ISO, 1.0 l), or carbachol (10 l) perfusion. Each experiment illustrates the return to baselinewith washout of the drugs and rechallenge with isoproterenol showing stimulation. (d) Stability of isoproterenolstimulation and the lack of effect of addition of 0.2 m GTP in the pipette on these G-protein mediated processes forthe duration of the experiment.
maximal response, Vh and the slope factor, k, were 1 and deleting the ATG translational start. Afterobtained by a least squares method. Errors are verification of targeting by Southern analysis, thestandard errors of the mean. P values for basal and clone was expanded and cultured in high con-stimulated currents were estimated by two-tailed centration G418 to enrich in homozygous ai3 tar-Student’s t-test. P values for time constants of in- geted ES cell lines (Mortensen, 1993a). Afterhibition kinetics were compared by two-tailed Stu- Southern screening, identified homozygous celldent’s t-test. Curves were compared by cell type and lines were expanded and the ai2 gene was targetedtime using 2-way ANOVA and the less significant with a construct using the hygromycin B resistancestatistic reported. gene (hyg) to interrupt exon 6 (Mortensen et al.,
1991). Homologous recombination was confirmedby Southern analysis and the targeted clones wereselected in high concentration hygromycin B toResultsenrich for homozygous ai2 knockout lines. Followingscreening by Southern analysis for homozygousMutant ES cells lacking ai2, ai3 and ao have beenclones, the inactivation of both ai2 and ai3 genespreviously isolated and characterized (Sowell et al.,was confirmed by Western analysis (Fig. 1). Western1997). Cell lines lacking both ai2 and ai3 wereblotting has previously shown successful gene in-isolated by sequential targeting. Cells lacking ai3
activations for the individual knockout cell lineswere previously produced by homologous re-(Sowell et al., 1997) and did not reveal com-combination using a targeting construct with the
neomycin resistance gene (neo) interrupting exon pensatory changes in expression of other G-protein
C. Ye et al.1776
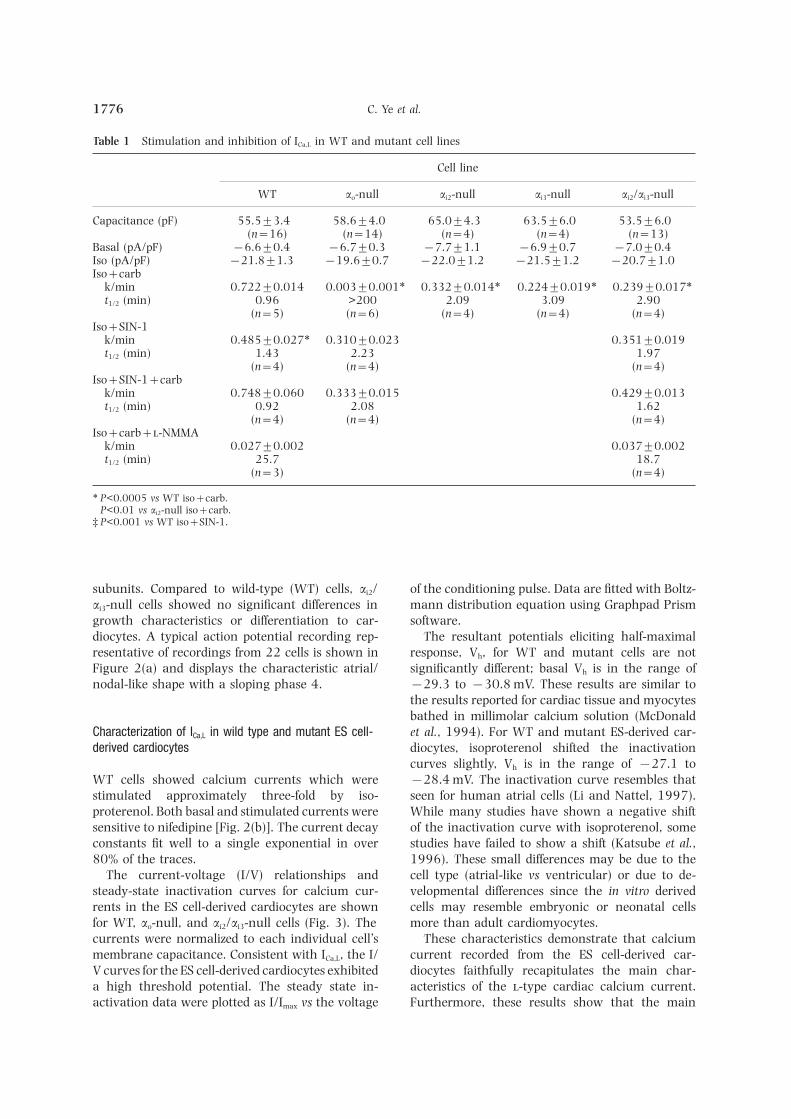
Table 1 Stimulation and inhibition of ICa,L in WT and mutant cell lines
Cell line
WT ao-null ai2-null ai3-null ai2/ai3-null
Capacitance (pF) 55.5±3.4 58.6±4.0 65.0±4.3 63.5±6.0 53.5±6.0(n=16) (n=14) (n=4) (n=4) (n=13)
Basal (pA/pF) −6.6±0.4 −6.7±0.3 −7.7±1.1 −6.9±0.7 −7.0±0.4Iso (pA/pF) −21.8±1.3 −19.6±0.7 −22.0±1.2 −21.5±1.2 −20.7±1.0Iso+carb
k/min 0.722±0.014 0.003±0.001∗ 0.332±0.014∗ 0.224±0.019∗† 0.239±0.017∗†t1/2 (min) 0.96 >200 2.09 3.09 2.90
(n=5) (n=6) (n=4) (n=4) (n=4)Iso+SIN-1
k/min 0.485±0.027∗ 0.310±0.023 0.351±0.019t1/2 (min) 1.43 2.23 1.97
(n=4) (n=4) (n=4)Iso+SIN-1+carb
k/min 0.748±0.060† 0.333±0.015 0.429±0.013t1/2 (min) 0.92 2.08 1.62
(n=4) (n=4) (n=4)Iso+carb+-NMMA
k/min 0.027±0.002 0.037±0.002t1/2 (min) 25.7 18.7
(n=3) (n=4)
∗ P<0.0005 vs WT iso+carb.† P<0.01 vs ai2-null iso+carb.‡ P<0.001 vs WT iso+SIN-1.
subunits. Compared to wild-type (WT) cells, ai2/ of the conditioning pulse. Data are fitted with Boltz-mann distribution equation using Graphpad Prismai3-null cells showed no significant differences in
growth characteristics or differentiation to car- software.The resultant potentials eliciting half-maximaldiocytes. A typical action potential recording rep-
resentative of recordings from 22 cells is shown in response, Vh, for WT and mutant cells are notsignificantly different; basal Vh is in the range ofFigure 2(a) and displays the characteristic atrial/
nodal-like shape with a sloping phase 4. −29.3 to −30.8 mV. These results are similar tothe results reported for cardiac tissue and myocytesbathed in millimolar calcium solution (McDonaldet al., 1994). For WT and mutant ES-derived car-Characterization of ICa,L in wild type and mutant ES cell-
derived cardiocytes diocytes, isoproterenol shifted the inactivationcurves slightly, Vh is in the range of −27.1 to
WT cells showed calcium currents which were −28.4 mV. The inactivation curve resembles thatseen for human atrial cells (Li and Nattel, 1997).stimulated approximately three-fold by iso-
proterenol. Both basal and stimulated currents were While many studies have shown a negative shiftof the inactivation curve with isoproterenol, somesensitive to nifedipine [Fig. 2(b)]. The current decay
constants fit well to a single exponential in over studies have failed to show a shift (Katsube et al.,1996). These small differences may be due to the80% of the traces.
The current-voltage (I/V) relationships and cell type (atrial-like vs ventricular) or due to de-velopmental differences since the in vitro derivedsteady-state inactivation curves for calcium cur-
rents in the ES cell-derived cardiocytes are shown cells may resemble embryonic or neonatal cellsmore than adult cardiomyocytes.for WT, ao-null, and ai2/ai3-null cells (Fig. 3). The
currents were normalized to each individual cell’s These characteristics demonstrate that calciumcurrent recorded from the ES cell-derived car-membrane capacitance. Consistent with ICa,L, the I/
V curves for the ES cell-derived cardiocytes exhibited diocytes faithfully recapitulates the main char-acteristics of the -type cardiac calcium current.a high threshold potential. The steady state in-
activation data were plotted as I/Imax vs the voltage Furthermore, these results show that the main
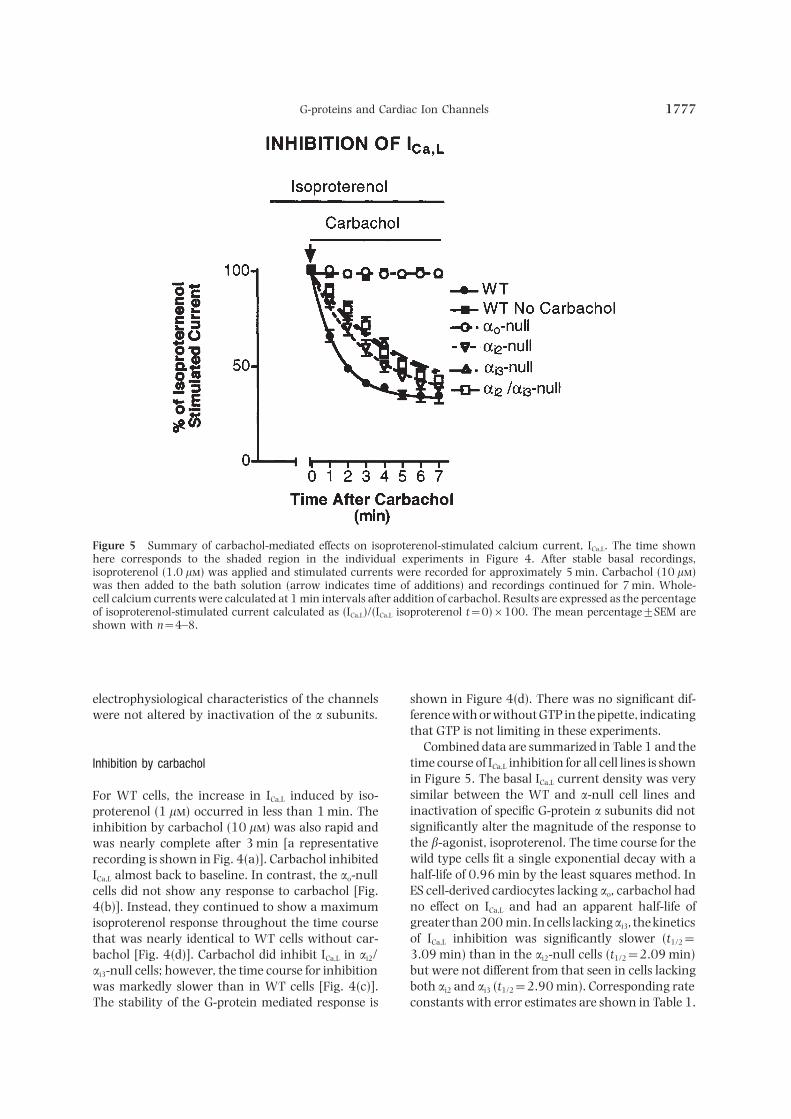
G-proteins and Cardiac Ion Channels 1777
Figure 5 Summary of carbachol-mediated effects on isoproterenol-stimulated calcium current, ICa,L. The time shownhere corresponds to the shaded region in the individual experiments in Figure 4. After stable basal recordings,isoproterenol (1.0 l) was applied and stimulated currents were recorded for approximately 5 min. Carbachol (10 l)was then added to the bath solution (arrow indicates time of additions) and recordings continued for 7 min. Whole-cell calcium currents were calculated at 1 min intervals after addition of carbachol. Results are expressed as the percentageof isoproterenol-stimulated current calculated as (ICa,L)/(ICa,L isoproterenol t=0)×100. The mean percentage±SEM areshown with n=4–8.
electrophysiological characteristics of the channels shown in Figure 4(d). There was no significant dif-ference with or without GTP in the pipette, indicatingwere not altered by inactivation of the a subunits.that GTP is not limiting in these experiments.
Combined data are summarized in Table 1 and thetime course of ICa,L inhibition for all cell lines is shownInhibition by carbacholin Figure 5. The basal ICa,L current density was verysimilar between the WT and a-null cell lines andFor WT cells, the increase in ICa,L induced by iso-inactivation of specific G-protein a subunits did notproterenol (1 l) occurred in less than 1 min. Thesignificantly alter the magnitude of the response toinhibition by carbachol (10 l) was also rapid andthe b-agonist, isoproterenol. The time course for thewas nearly complete after 3 min [a representativewild type cells fit a single exponential decay with arecording is shown in Fig. 4(a)]. Carbachol inhibitedhalf-life of 0.96 min by the least squares method. InICa,L almost back to baseline. In contrast, the ao-nullES cell-derived cardiocytes lacking ao, carbachol hadcells did not show any response to carbachol [Fig.no effect on ICa,L and had an apparent half-life of4(b)]. Instead, they continued to show a maximumgreater than 200 min. In cells lackingai3, the kineticsisoproterenol response throughout the time courseof ICa,L inhibition was significantly slower (t1/2=that was nearly identical to WT cells without car-3.09 min) than in the ai2-null cells (t1/2=2.09 min)bachol [Fig. 4(d)]. Carbachol did inhibit ICa,L in ai2/but were not different from that seen in cells lackingai3-null cells; however, the time course for inhibitionboth ai2 and ai3 (t1/2=2.90 min). Corresponding ratewas markedly slower than in WT cells [Fig. 4(c)].
The stability of the G-protein mediated response is constants with error estimates are shown in Table 1.

C. Ye et al.1778
Pretreatment of WT cells with 100 ng/ml PTX for18 h completely blocked the effect of carbachol onisoproterenol (1 l) stimulated ICa,L (n=3±SEMin nA/pF: WT=19.3±1.7, WT+carbachol=18.3±1.7). With inactivation of both ai2 and ai3 ina single cell (ai2/ai3-null cells), the inhibitory effectof carbachol on ICa,L was blocked by preincubationwith PTX, indicating that receptor signaling stillinvolved the remaining PTX-sensitive G-protein, Go
(ai2/ai3-null=18.7±2.2, ai2/ai3-null+carbachol=17.98±2.2).
Role of NO
Preincubation of WT or ai2/ai3-null cells with theNOS inhibitor -NMMA completely blocked in-hibition of ICa,L by carbachol. Addition of the NO-generator SIN-1 to isoproterenol-stimulated WTcells inhibited ICa,L (Fig. 6 and Table 1), in agreementwith results reported for nodal cardiocytes. In orderto test whether all effects of carbachol could be mim-icked by SIN-1, we compared the responses with car-bachol and SIN-1 in combination. In WT cells,carbachol and carbachol plus SIN-1 showed slightlyfaster kinetics of inhibition than SIN-1 alone, sug-gesting that at least some effects of carbachol do notoperate through NO. In the ao-null cells, SIN-1 wasable to inhibit ICa,L, indicating that NO generationwas distal to the requirement for ao.
Discussion
Muscarinic agonists regulate a number of cardiacion channels via signaling pathways initiated by re-ceptor activation of PTX-sensitive G-proteins (Ro-bishaw and Hansen, 1994). To determine if specificG-proteins are required for regulation of different ionchannels, we generated a panel of mouse ES cell lineswith targeted inactivation of specific PTX-sensitive asubunits. Using these mutant cells, we have pre-viously shown that expression of ai2 and ai3 but notao is required for muscarinic activation of the cardiac
Figure 6 Effect of -NMMA or SIN-1 on ICa,L in WT cells,inwardly rectifying K+ channel, IKACh (Sowell et al.,ao-null cells, and ai2/ai3-null cells. The arrow indicates1997). In this report, we present evidence that alltime of additions (carbachol, SIN-1, or SIN-1 plus car-three PTX-sensitive G-proteins expressed in heart, bachol) to the isoproterenol containing bath solution.
Gi2, Gi3 and Go, affect the anti-adrenergic action of The cells were preincubated with -NMMA for 30 minthe muscarinic agonist carbachol on cardiac calcium prior to the recordings and -NMMA was continuously
present during the experiment. Results are expressed ascurrents. Our results establish an essential role for aothe mean percent of isoproterenol-stimulated currentin muscarinic receptor regulation of the cardiac ICa,L ±SEM and calculated as described in Figure 5. Eachin these nodal/atrial-like cells and further show that point represents recordings of four to eight individual
both ai2 and ai3 have important roles in the normal cells. Results with carbachol alone are shown for com-regulatory process. parison.
A simple explanation for the inability of carbachol

G-proteins and Cardiac Ion Channels 1779
Figure 7 Working hypothesis for cardiac pathways controlling -type calcium channels by muscarinic stimulation.Three points in the pathway could be modified by ai G-proteins: (1) stimulation of NO, (2) inhibition of adenylyl cyclaseand (3) stimulation of a phosphatase decreasing the half-life of the stimulated phosphorylated channel. The NO-dependent pathway is thought to operate by increasing soluble guanylyl cyclase (GC) activity and increasing cGMPconcentration. The cGMP then activates a phosphodiesterase which decreases cAMP concentration thus opposing theeffects of bAR stimulation of adenylyl cyclase.
to inhibit ICa,L in the ao-null cells is that Go plays a muscarinic inhibition of ICa,L, consistent with the pro-posed NO-dependent pathways (Han et al., 1996).critical role in the pathway leading to activation of
NOS. Our finding that SIN-1 is able to inhibit ICa,L in However, expression of ao alone is not sufficientfor normal muscarinic responses. In the ai2/ai3-nullthe ao-null cells indicates that the regulatory steps
distal to NO generation are present and functional cells, ao is intact, yet the response of the cells to car-bachol is much slower. Inactivation of ai subunitsin the ao-null cells. Inactivation of ao does not disrupt
all muscarinic signaling since regulation of IKACh clearly has different effects on the regulatory processthan does inactivation of ao. Our results show that(Sowell et al., 1997; Valenzuela et al., 1997) and
inhibition of adenylyl cyclase is intact inao-null vent- ai2 and ai3 are not interchangeable since inactivationof these subunits slows channel inhibition by car-ricular cardiocytes (Jiang et al., 1998). The lack of
effect on inhibition of adenylyl cyclase in conjunction bachol to different degrees and the effect of ai2 andai3 inactivation were not additive. Therefore, therewith our results strongly argues that Go has an es-
sential role in regulating the cardiac calcium chan- may be common or related mechanisms that requireboth subunits. Addition of the NO generating agentnel via a NO-dependent mechanism independent of
any effects on adenylyl cyclase activity. These effects SIN-1 resulted in somewhat more rapid inhibition ofICa,L in the ai2/ai3-null cells than carbachol, so it isare likely dependent on cAMP via regulation of a
phosphodiesterase by NO. The role of phos- possible that absence of ai leads to inadequate gen-eration of NO.phodiesterases in regulating the cardiac calcium
channel has previously been investigated. Han et al. It is possible that these G-proteins are involved insignaling to other regulatory mechanisms. Possible(1995) showed that in sino-atrial nodes, 8-Br-cAMP
stimulated ICa,L and that activated ICa,L was then in- mechanisms include inhibition of adenylyl cyclaseby ai2 and ai3 or activation of phosphatases whichsensitive to muscarinic inhibition. Similar results
were obtained in atrial-ventricular nodes (Han et al., has been shown in some cells to be an effect of Gi
activation (Srikant and Shen, 1996; Caverzasio et1996). These results are consistent with the pro-posed NO-dependent stimulation of a phos- al., 1997). Lack of either of these mechanisms could
account for the altered kinetics.phodiesterase, since the cAMP analogue is not reg-ulated by phosphodiesterase. Phosphodiesterase in- The possible points of regulation are illustrated in
Figure 7. The Go heterotrimeric G-protein is shownhibitors have also been shown to reverse the

C. Ye et al.1780
tyrosine phosphorylation process. J Bone Miner Res 12:regulating the NO pathway but other pathways may1975–1983.also be involved. The three possibilities for the Gi2 D Y, L LM, L RJ, 1997. Switchingand Gi3 regulation discussed above are depicted as: of the coupling of the b2-adrenergic receptor to different
(1) contribution to regulation of NO, although they G proteins by protein kinase A. Nature 390: 88–91.are not required; (2) inhibition of adenylyl cyclase; H X, K L, B JL, K RA, S TW,
1996. Nitric oxide synthase (NOS3)-mediated cholin-and (3) regulation of a phosphatase which de-ergic modulation of Ca2+ current in adult rabbit atriov-phosphorylates the calcium channel. Further in-entricular nodal cells. Circ Res 78: 998–1008.vestigation will be required in a system where
H X, K L, Z YY, O DJ, L WD, K RA,enough cardiocytes can be obtained to perform bio- S TW, 1997. Nitric oxide regulation of atriov-chemical measurements. entricular node excitability. Can J Cardiol 13: 1191–
1201.The role of NOS in inhibiting ICa,L in the WT ESH X, S Y, G WR, 1995. A cellular mech-cell-derived cardiocytes is in agreement with results
anism for nitric oxide-mediated cholinergic control ofdescribed for native nodal heart cells (Mery et al.,mammalian heart rate. J Gen Physiol 106: 45–65.
1993; Han et al., 1996, 1997). The requirement for J M, G MS, B G, S K, P M,ao in ICa,L inhibition is the same as shown for vent- B P, S Y, R U, E G,ricular cells (Valenzuela et al., 1997). Therefore, the B L, 1998. Multiple neurological ab-
normalities in mice deficient in the G protein Go. ProcES cell-derived cardiocytes faithfully recapitulateNatl Acad Sci USA 95: 3269–3274.many features of native cardiocytes and are a valu-
K Y, Y H, N L, S N, 1996.able model for studies of cardiac receptor signalDifferences in isoproterenol stimulation of Ca2+ current
transduction. of rat ventricular myocytes in neonatal compared toThe importance of understanding the char- adult. Eur J Pharmacol 317: 391–400.
K RP, S S, D KP, S ME,acteristics of ES cell-derived cardiocytes is enhancedL N J, W AL, C WC, F LJ, 1993.by the recent production of human ES cells. TheseSpontaneous activity in transgenic mouse heart: com-cells are a possible source of human cardiocytes forparison of primary atrial tumor with cultured AT-1
tissue therapy (Shamblott et al., 1998; Thomson et atrial myocytes. J Cardiovasc Electrophysiol 4: 642–660.al., 1998). However, until the genetically ma- K SJ, C MA, H A, 1991. Perforated patchnipulated human ES cells are readily available, recording. Meth Neurosci 4: 264–373.
L GR, N S, 1997. Properties of human atrial ICacharacterization of the murine equivalent will serveat physiological temperatures and relevance to actionas a model for the behavior of in vitro derived car-potential. Am J Physiol 272: H227–H235.diocytes. In addition, murine ES cells can serve as a
L CW, G P, M G, U C, Smodel for testing tissue therapy paradigms in mice A, N NM, 1987. Tissue-specific regulationand for comparisons in murine disease models. of GTP-binding protein and muscarinic acetylcholine
receptor levels during cardiac development. Bio-In summary, multiple PTX-sensitive G-proteins af-chemistry 26: 4876–4884.fect muscarinic regulation of the cardiac calcium
M VA, R J, H J, W AM, 1993.current; expression of ao is essential while ai2 and ai3 Embryonic stem cells differentiate in vitro into car-appear to play an important role in regulating the diomyocytes representing sinusnodal, atrial and vent-kinetics of the inhibitory process. ricular cell types. Mech Dev 44: 41–50.
M VA, W AM, R J, B M, H- J, 1994. Cardiomyocytes differentiated in vitrofrom embryonic stem cells developmentally expressAcknowledgementscardiac-specific genes and ionic currents. Circ Res 75:233–244.
This work was supported by NIH CSDA (K08 MD TF, P S, T W, P DJ, 1994.HL03411-02) to MOS, NIH K01 AG00891-01 to Regulation and modulation of calcium channels in
cardiac, skeletal, and smooth muscle cells. Physiol RevCY, NIH R29 HL54095 to DSM, and an AHA74: 365–507.Established Investigator Award and the NIH R01
M PF, P C, B L, P F, FGM49122 and HL58606 to RMM.R, 1993. Nitric oxide regulates cardiac Ca2+ current.Involvement of cGMP-inhibited and cGMP-stimulatedphosphodiesterases through guanylyl cyclase ac-tivation. J Biol Chem 268: 26286–26295.References
M RM, 1993a. Double knockouts. Productionof mutant cell lines in cardiovascular research. Hyper-C DL, S HC, 1995. Regulation of calciumtension 22: 646–651.channels in the heart. Adv Second Messenger Phos-
M RM, 1993b. Homologous recombination. In:phoprotein Res 30: 25–88.Ausubel FBR, Kingston R, Moore D, Smith JA, SeidmanC J, P G, S A, B JP, 1997.J, Struhl K (eds). Current Protocols in Molecular Biology.Mechanism of the mitogenic effect of fluoride on os-
teoblast-like cells: evidences for a G protein-dependent Vol. Unit 9.15. John Wiley and Sons, Inc, 1–6.

G-proteins and Cardiac Ion Channels 1781
M RM, C DA, C S, G- cardiac muscarinic K+ channel, IK+Ach, in intactcells. Proc Natl Acad Sci USA 94: 7921–7926.L AAT, S JG, 1992. Production of
homozygous mutant ES cells with a single targeting S CB, S SH, 1996. Octapeptide somatostatinanalog SMS 201-995 induces translocation of in-construct. Mol Cell Biol 12: 2391–2395.
M RM, Z M, N EJ, S JG, tracellular PTP1C to membranes in MCF-7 humanbreast adenocarcinoma cells. Endocrinology 137:1991. Embryonic stem cells lacking a functional
inhibitory G-protein subunit (a i2) produced by gene 3461–3468.T JA, I-E J, S SS, Wtargeting of both alleles. Proc Natl Acad Sci USA 88:
7036–7040. MA, S JJ, M VS, J JM, 1998.Embryonic stem cell lines derived from human blast-R JD, H CA, 1994. Structure and function
of G proteins mediating signal transduction pathways ocysts. Science 282: 1145–1147.V D, H X, M U, F C, Min the heart. Alcohol Clin Exp Res 18: 115–120.
S MJ, A J, W S, B EM, L H, H P, P J, N EJ, F MC, 1997.G a(o) is necessary for muscarinic regulation of Ca2+JW, D PJ, B PD, H GR, G-
JD, 1998. Derivation of pluripotent stem cells channels in mouse heart. Proc Natl Acad Sci USA 94:1727–1732.from cultured human primordial germ cells [In Process
Citation]. Proc Natl Acad Sci USA 95: 13726–13731. X RP, J X, L EG, 1995. Functional couplingof the b2-adrenoceptor to a pertussis toxin-sensitiveS MO, Y C, R DA, H S, Q SJ,
V PM, M RM, 1997. Targeted in- G protein in cardiac myocytes. Mol Pharmacol 47:322–329.activation of ai2 or ai3 disrupts activation of the




















