
Forebrain HCN1 Channels Contribute to Hypnotic Actions of Ketamine

European Journal ofPain (1999) 3: 7-12
Functional anatomy of hypnotic analgesia: a PET study of patients with fibromyalgia
Gustav Wika,b, H5kan Fischer c,d, Bjijrn Brag6eb, Basil Finer-b and Mats Fredriksonc
aDepartment of Clinical Neurosciences, Karolinska Institute and Hospital, Stockholm, Sweden; bKronan Pain Clinic, Stockholm, Sweden: =Department of Clinical Psychology, Uppsala University, Uppsala, Sweden; and dUppsala University PET-centre, Uppsala University, Uppsala, Sweden
Hypnosis is a powerful tool in pain therapy. Attempting to elucidate cerebral mechanisms behind hypnotic analgesia, we measured regional cerebral blood flow with positron emission tomography in patients with fibromyalgia, during hypnotically-induced analgesia and resting wakefulness. The patients experienced less pain during hypnosis than at rest. The cerebral blood-flow was bilaterally increased in the orbitofrontal and subcallosial cingulate cortices, the right thalamus, and the left inferior parietal cortex, and was decreased bilat- erally in the cingulate cortex. The observed blood-flow pattern supports notions of a multifactorial nature of hypnotic analgesia, with an interplay between cortical and subcortical brain dynamics.
INTRODUCTION
Although it is widely accepted that hypnosis effectively reduces the perception of clinical and experimental pain (Hilgard, 1986), the cerebral mechanisms mediating hypnotic anal- gesia are only in part understood. Reflecting that pain itself is multidimensional, involving attentional, sensory, and affective components (Melzack & Casey, 1968), hypnotic analgesia may engage multiple pain-reducing mecha- nisms. Using 133Xenon imaging, Crawford et al. (1993) studied central neural effects of hypnotic analgesia in males with high and low hypnotic susceptibility. They found bilaterally-increased regional cerebral blood flow (rCBF) in the orbito-frontal cortex among highly hypnotiza- ble men, and suggested that hypnotic analgesia involves supervisory attentional control systems that co-operate in the regulation of thalamo- cortical activities.
Paper received 30 May 1997 and accepted in revised form 10 October 1998. Correspondence to: Gustav Wik, Kronan Pain Clinic, Mariebergsg. 5, S- 112 8 1 Stockholm, Sweden.
One mechanism of action in hypnotic anal- gesia may relate to descending thalamo-spinal pathways for nociceptive control (Jesse1 & Kely, 1991), mediated by endogenous opiates (Frenk et al., 1986) or other neurochemical systems (Sternbach, 1982). Another mechanism for hyp- notic analgesia is indicated by the neo-dissocia- tion theory (Hilgard & Hilgard, 1983), proposing that hypnotic analgesia reduces the awareness of pain, once nociception has reached a higher processing level. Studying a specific nociceptive reflex, Kiernan et al. (1995) found that hypnotic analgesia produced signifi- cant reductions, not only in ratings of pain and unpleasantness, but also in a spinally-mediated nociceptive reflex: the R-III reflex. As a conse- quence, they suggested that hypnotic analgesia involves the activation of a hierarchical pain control system engaging the two mechanisms described above: (a) antinociceptive activity related to spinal cord mechanisms; and (b) processes serving to prevent awareness of pain once the nociceptive signal has reached higher centres. Additionally, they proposed a third mechanism: (c) changes in the meanings related to the painful sensations.
1090.3801/99/010007 + 06 $12.00/O 0 1999 European Federation of Chapters of the International Association for the Study of Pain

8 G. WIK ETAL.
The fibromyalgia syndrome (FS) is a chronic disabling condition of unknown aetiology (Smythe, 1972). Widespread pain from relatively normal musculosceletal tissue is a major FS com- plaint, with low threshold for nociceptive stimuli and super-sensitive tender-points as a diagnostic hallmark (Wolfe et al., 1990). In the present study, we used positron emission tomography (PET)-based measures of rCBF to investigate FS patients during resting wakefulness and hypnotic analgesia. The study aimed to elucidate and extend the 13”Xenon rCBF correlates of hypnotic analgesia of Crawford et al. (1993) and to search for neuroanatomical underpinnings for the notion of an hierarchical pain control system being activated during hypnosis, as suggested by Kiernan et al. (1995).
MATERIALS AND METHODS
Eight right-handed women, mean age (SD) 47 (+ 4.6) years (range 42-56 years), with fibromyal- gia were included in the study. They were recruited from patients that had been referred to the Kronan Pain Clinic, Stockholm, for treat- ment of chronic disabling pain, and fulfilled the FS criteria put forward by the American College of Rheumatology (1990). Hypnotizability was tested before inclusion, and patients partici- pating in the study had scores of more than 9 according to the Harvard Group Scale of Hypnotic Susceptibility Form A (Shor & Orne, 1963). The study was approved by the local Ethics Committee and the Radiation Safety Committee of the Karolinska Hospital. Informed consent was obtained from all participants.
To control for the visual aspect of resting dif- ferences, the subjects were visually stimulated during scanning, using videotapes with scenes of individuals walking in a park (Wik et al., 1996). The conditions of resting wakefulness and hyp- notic analgesia were counterbalanced between subjects. During resting wakefulness, patients were told to be comfortable and watch the video- tapes. Hypnosis was induced by gently talking to the subjects, instructing them to be relaxed and to go into a deep trance, to watch the videotapes, and not to feel any pain whatsoever. Directly
after each scan, pain ratings were made on a visual analogue scale that spanned from 0 (no pain) to 10 (unbearable pain).
All scans were performed in a darkened room, with sounds reduced to a minimum. For attenua- tion corrections, a transmission scan was first performed. Immediately before each emission scan, 25 mCi (925MBq) of [i50]butanol (Berridge et al, 1990) was administrated to the subjects. A functional blood flow image was determined for each individual by summing uptake data from 10 images covering 100 s. An eight-ring PET scan- ner (PC 2048-l 5B) was used (Litton et al., 1990). The transaxial image resolution is 5 mm full width at half maximum (FWHM), while the axial resolution is 6 mm FWHM. Anatomical standardization of all individual CBF images into a standard brain shape was performed using the Greitz computerized brain atlas (Greitz et al., 1991). To make cross-study comparisons possi- ble, coordinates from the Talairach and Tournoux (1988) brain atlas have been imported into the Greitz atlas software, and can be produced for each data-point. Talairach coordinates for peak activation are given for brain regions being acti- vated. Anatomically-normalized and -filtered images of 20 mm at FWHM were used to gener- ate standardized Z-maps, based on an estimate of the standard deviation pooled over all intrac- erebral voxels (Worsley et al., 1992). When assessing the significance of rCBF changes, Z-values corresponding to a Bonferroni cor- rected p-value of 0.05 were estimated and dis- played (Worsley et al., 1992). The extent of activated regions was estimated by including all voxels with a Z-value corresponding to a non- Bonferroni corrected p-value of 0.001, directly or indirectly connected (using a six-connectivity criterion) to a significant peak value.
RESULTS
Pain ratings were lower after the hypnotic state than after the resting (mean 4.3 + 2.5 vs 3.2 +2,7, p=O.O66).
As compared to the resting state, hypnotic analgesia increased rCBF bilaterally in the subcallosial Jcingulate gyrus (BA 25) the right
European Journal of Pain (1999), 3

HYPNOTIC ANALGESIA AND PET 9
TABLE 1. Significant rCBF peak-differences and locations between hypnotic analgesia and rest in fibromyalgic patients, using the Worsley statistical method and a pvalue cut at 0.05.
Brain areas Brodmann areas
Talairach coordinates
X Y 2 Z-value
Increases R Subcallosal L Subcallosal L Inferior Parietal R Thalamus R Orbitofrontal (r/=4) L Orbitofrontal (n=4)
Decreases R Anterior cingulate L Anterior cingulate R Posterior cingulate L Posterior cingulate
25 25
40139
11 11
24133 24133 23131 23131
2 9 -2 4.24 -1 7 -2 4.57
-43 -30 22 5.15 9 -18 17 6.32 1 42 -22 4.18
-5 46 -22 4.39
-1 1
-1
-10 26 3.71 -9 26 3.71
-21 26 4.42 -28 26 4.47
R, right; I, left. Talairach coordinates are expressed in mm and and refer to: (x) the medial- lateral position relative to midline (R=+); (y) anterior-posterior position relative to the anterior commissure (anterior=+); and (4 superior-inferior position relative to the commisural line (superior=+).
thalamus, and the left inferior parietal cortex (BA 39 and 40) (Table 1 and Fig. 1). Because of its inferior and frontal location, the orbitofrontal cortex was imaged only in four patients. A sepa- rate analysis of these patients, however, revealed increased orbitofrontal cortex activation during hypnotic analgesia (Table 1). Decreases in rCBF during hypnotic analgesia were seen bilaterally in cluster, including the posterior cingulate gyrus (BA 23/3 I), and the posterior part of the anterior cingulate gyrus (BA 24) (Table 1 and Fig. 2).
DISCUSSION
The rCBF pattern contrasting hypnotic analgesia and resting wakefulness found in this study, underpin and extend the previous 133Xenon find- ings by Crawford et al. (1993). It is also consis- tent with a functional hierarchical neural network for pain control (Kiernan et al., 1995), activating an interplay between cortical and sub- cortical brain dynamics.
With the exeption of increased rCBF in the left inferior parietal cortex, rCBF changes were all related to limbic system structures. The orbitofrqntal cortex, as well as being part of the limbic system, has been shown in animal studies to have widespread connections to the subcal- losial area, the thalamus, and the cingulate gyrus
(Hirose et al., 1992; Morecraft et al., 1992; Ray & Price 1993), i.e. those brain regions in which we observed a changed rCBF during hypnotic anal- gesia. The increased rCBF in this region is in accordance with Crawford et al. (1993), and may be essential in fronal-limbic aspects of pain pro- cessing. The subcallosial gyrus cingulus area (BA 25/33) was bilaterally more activated during hyp- notic analgesia than resting wakefulness. Besides being closely, functionally and anatomically, related to the orbitofrontal cortex, animal studies show that the subcallosial area projects to multi- ple limbic and paralimbic regions, including pain-related structures as mid-line thalamic nuclei and the central grey (Takagishi & Chiba, 1991). Areas 25 and 33 are the least differentiated in the anterior cingulate cortex (Devinsky et al., 1995), and are engaged during internal states associated with emotions, processing of painful stimuli, vocalization and attention to action. In addition, the thalamus was more activated dur- ing hypnotically-induced analgesia than resting wakefulness. Besides limbic-system aspects, the thalamus also has a clear-cut role in pain defense, possibly related to analgesic processes. A prereq- uisite for pain control by descending pathways in hypnotic analgesia, however, is that mental activ- ities can pace thalamic and spinal anti-nocicep- tive mechanisms. Some support for this comes from animal studies showing cortical influences
European Journal of fain (1999), 3
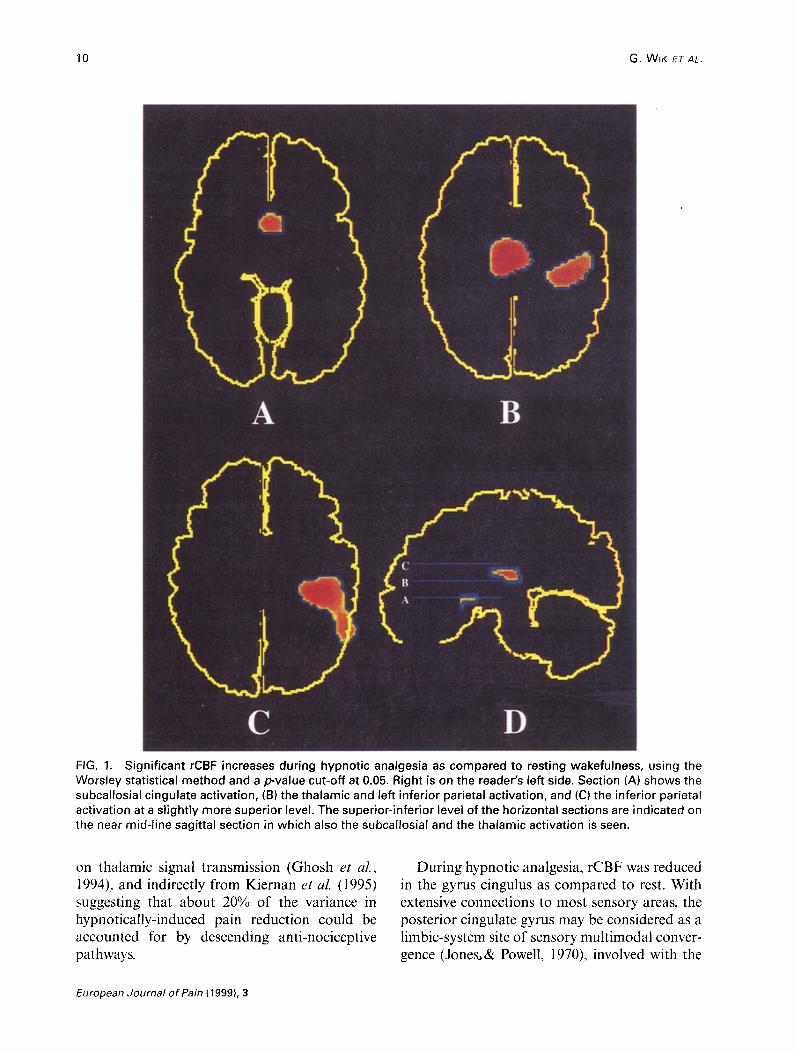
10 G. WIK ETAL.
FIG. 1. Significant rCBF increases during hypnotic analgesia as compared to resting wakefulness, using the Worsley statistical method and a pvalue cut-off at 0.05. Right is on the reader’s left side. Section (A) shows the subcallosial cingulate activation, (B) the thalamic and left inferior parietal activation, and (Cl the inferior parietal activation at a slightly more superior level. The superior-inferior level of the horizontal sections are indicated on the near mid-line sagittal section in which also the subcallosial and the thalamic activation is seen.
on thalamic signal transmission (Ghosh et al., During hypnotic analgesia, rCBF was reduced 1994) and indirectly from Kiernan et al, (1995) in the gyrus cingulus as compared to rest. With suggesting that about 20% of the variance in extensive connections to most sensory areas, the hypnotically-induced pain reduction could be posterior cingulate gyrus may be considered as a accounted for by descending anti-nociceptive limbic-system site of sensory multimodal conver- pathways. gence (Jones,& Powell, 1970), involved with the
European Journal of Pain (1999), 3

HYPNOTIC ANALGESIA AND PET 11
components disappeared. Hsieh et al. (1995) found more cingulate activation in chronic ongo- ing neuropathic pain than in a pain-alleviated state, and suggested this activation to reflect sen- sorial and affectional aspects of pain. Thus, the cingulate deactivation may reflect that FS patients pay less emotional attention to pain sig- nalling (reduced secondary hyperalgesia) during hypnotic analgesia than resting wakefulness.
A pain processing role for the inferior parietal cortex is supported by PET studies showing pari- eta1 activation following experimental pain (Coghill et al., 1994; Derbyshire et al., 1994), and that infarcts of the parietal lobe may be accom- panied by pain (Masson et al., 199 1; Schahmann & Leifer, 1992), sometimes induced by intellec- tual concentration (Michel et al., 1990). In accor- dance with the neo-dissociation theory of hypnotic analgesia (Hilgard & Hilgard 1983), the left inferior parietal activation may then reflect psychological mechanisms serving to prevent awareness of pain, once signalling has reached higher centres.
FIG. 2. Near mid-line sagittal section showing rCBF decreases during hypnotic analgesia as compared to resting wakefulness using the Worsley statistical method and a pvalue cut-off at 0.05. The cluster of deactivation contains mainly the posterior cingulate, but extends to the rostal ‘nociceptive’ portion (Devinsky et al., 1995) of the anterior cingulate cortex.
encoding of the associative significance of stim- uli (Baleydier & Mauguiere, 1980). During rest- ing wakefulness, posterior cingulate activity may thus reflect the attention to, and encoding of, multiple sensory events, including Iibromyalgic pain signalling. During hypnotic analgesia, how- ever, the state of consciousness is altered, and external suggestions take precedence over the normal perception (Ludwig, 1966; Smyth & Lowy, 1983). Thus, being absorbed by the hyp- notically-induced message subjects may become less attentive to details of sensory afference (Smyth & Lowy, 1983). Moreover, the cluster of cingulate deactivation during hypnotic analgesia extended bilaterally into the deep rostal anterior gyrus cingulus. In fact, the anterior part of the deactivation that we now observe corresponds to what Devinsky et al. (1995) suggested to be the cingulate ‘nociceptive cortex’ (cf. their Fig. 1, p. 281). Foltz and White (1962) indicated that this brain region may have a function related to emo- tional and attentive processing of pain. They found that anxious patients who ‘augmented’ their pain were those most likely to benefit from cingulotomy, and that their patients continued to feel the pain after the cingulotomy, without dis- tress. Thus, in spite of pain perception not being altered, the adverse emotional and the attentional
ACKNOWLEDGMENTS
We wish to thank Britt-Marie Berggren, Walter Pulka and Peter Soderholm for excellent techni- cal assistance, and Morten Juell for valuable comments on the manuscript. The project was supported by the Swedish Medical Research Council (10594), the Swedish Council for Research in Humanities and Social Sciences and the Siiderstriim-Kiinigska Foundation.
REFERENCES
Baleydier C, Mauguiere F. The duality of the cingulate gyrus in monkey: neuroanatomical study and functional hypothesis. Bruin 1980; 103: 525-554.
Berridge M, Cassidy M, Miraldy F. A routine, automated synthesis of 150-labeled butanol for positron tomogra- phy. J Nucl Med 1990; 31: 112771731.
Coghill RC, Talbot JD, Evans AC, Meyer E, Cijedde A, Bushnell MC, Duncan GH. Distributet processing of pain and vibration by the human brain. J Neurosci 1994; 14: 409554108.
Crawford HJ, Gur RC, Skolnick B, Gur RE, Benson DM. Effects of hypnosis on regional cerebral blood flow dur- ing ischemic pain with and without suggested hypnotic analgesia. Znt J P.~~rchoph~siol 1993; 15: 18 lP 195.
European Journal of Pain (1999), 3

12 G. WIK ET AL.
Derbyshire SWG, Jones AKP, Devani P, Friston KJ, Feinmann C, Harris M, Pearce S, Watson JDG, Frackowiak RSJ. Cerebral responses to pain in patients with atypical facial pain measured by positron emission tomography. J Neurol Neurosurg Psychiatry. 1994; 57: 1166-l 172.
Devinsky 0, Morrell MJ, Vogt BA. Contributions of ante- rior cingulate cortex to behavior. Brain 199.5; 118: 279-306.
Foltz EL, White LE. Pain ‘relief by frontal cingulumtomy. J Neurosurg 1962; 19: 89-100.
Frenk H, Cannon JT, Lewis JW, Liebeskind JC. Neural and neurochemical mechanisms of pain inhibition. In: Sternbach RA, editor. The Psychology of Pain. New York: Raven Press, 1986: 2.548.
Ghosh S, Murray GM, Turman AB, Rowe MJ. Corticothalamic influences on transmission of tactile information in the ventroposterolateral thalamus of the cat: effect of reversible inactivation of somatosensory cortical areas I and II. Exp Brain Res 1994; 100: 276-286.
Greitz T, Bohm C, Holte S, Eriksson L. A computerized brain atlas: construction, anatomical content, and some applications. J Comput Assist Tomogr 1991; 15: 26-38.
Hilgard ER. Hypnosis and Pain. In: Sternbach R, editor. Psychology of Pain. New York: Raven Press, 1986: 1977221.
Hilgard ER, Hilgard JR. Hypnosis in the Relief of Pain. Los Altos: William Kaufmann Inc, 1983.
Hirose S, Ino T, Takada M, Kimura J, Akiguchi I, Mizuno N. Topographic projections from the subiculum to the limbic regions of the medial frontal cortex in the cat. Neurosci Lett 1992; 139: 61-64.
Hsieh J-C, Belfrage M, Stone-Elander S, Hansson P, Ingvar M. Central representation of chronic ongoing neuro- pathic pain studied by positron emission tomography. Pain 1995; 63: 225-236.
Jesse1 TM, Kely DD. Pain and analgesia. In: Kandel ER, Schwartz JH, Jesse1 TM, editors. Principles of Neural Science. Norwalk: Appleton and Lange, 1991: 385-399.
Jones EG, Powell TPS. An anatomical study of converging sensory pathways within the cerebral cortex of the mon- key. Brain 1970; 93: 7933820.
Kiernan D, Dane JR, Phillips LH, Price DD. Hypnotic analgesia reduces R-III nociceptive reflex: further evi- dence concerning the multifactorial nature of hypnotic analgesia. Pain 1995; 60: 39-47.
Litton JE, Holte S, Eriksson L. Evaluation of the Karolinska new positron camera system: the Scanditronix PC2048-15B. IEEE Trans Nucl Sci 1990; 37:743S748.
Ludwig AM. Altered states of consciousness. Arch Gen Psychiatry 1966; 15: 225-234.
Masson C, Koskas P, Cambier J, Masson M. Syndrome cor- tical pseudothalamique gauche et asymbolie a la douleur. Rev Nemo1 (Paris) 1991; 147: 668-670.
Melzack R, Casey KL. Sensory, motivational, and central control determinants of pain. In: Kenshalo D, editor. The Skin Senses. Springfield: Thomas, 1968; 423443. ,
Michel D, Laurent B, Convers P, Garcia-Larrea L, Le-Bas JF, Mauguiere F, Schott B. Cortical pain. Clinical, elec- trophysiologic and topographic study of 12 cases. Rev Neurol (Paris) 1990; 146: 405414.
Morecraft RJ, Geula C, Mesulam MM. Cytoarchitecture and neural afferents of orbitofrontal cortex in the brain of the monkey. J Comp Nemo1 1992; 323: 341-358.
Ray JP, Price JL. The organization of projections from the mediodorsal nucleus of the thalamus to orbital and medial prefrontal cortex in macaque monkeys. J Camp Neuroll993; 337: l-3 1.
Schahmann JD, Leifer D. Parietal pseudothalamic pain syn- drome. Arch Nemo1 1992; 49: 1032-1037.
Shor RE, Orne EC. Norms on the Harvard group scale of hypnotic susceptibility, Form A. Int J Clin Exp Hypn 1963; 11: 3947.
Smyth LD, Lowy D. Auditory vigilance during hypnosis: a brief communication. Znt J Clin Exp Hypnosis 1983; 31: 67-71.
Smythe HA. Non-articular rheumatism and the fibrositis syndromes. In: Hollander E, McCarty DJ, editors. Arthritis and Allied Conditions. Philadelphia, PA: Lea and Febiger, 1972: 808-816.
Sternbach RA. On strategies for identifying neurochemical correlates of hypnotic analgesia. Int J Clin E,xp Hypn 1982;30:251-256.
Takagishi M, Chiba T. Efferent projections of the infralim- bit (area 25) region of the medial prefrontal cortex in the rat: an anterograde tracer PHA-L study. Brain Res 199 1; 566:26-39.
Talairach J, Tournoux P. Co-Planar Stereotactic Atlas of’ the Human Brain. Stuttgart: Georg Thieme Verlag, 1988.
Wik G, Fredrikson M, Fischer H. Cerebral correlates of anticipated fear: a PET study of specific phobia. Intern J Neuroscience 1996; 87: 267-276.
Wolfe F, Smythe HA, Yunus MB, Bennett RM, Bombardier C, Goldenberg DL, Tugwell P, Campbell SM, Abeles M, Clark P The American College of Reumatology 1990 criteria for the classification of libromyalgia. Arthritis Rheum 1990; 33: 160-173.
Worsley KJ, Evans AC, Marrett S, Neelin I? A three-dimen- sional statistical analysis for CBF activation studies in human brain. J Cereh Blood FIOW Metab 1992; 12: 900-9 18.
European Journal of Pain (1999). 3
Copyright © 2022 FDOKUMEN




















