
Functional analysis of the essential bifunctional tobacco enzyme 3-dehydroquinate...
15
Journal of Experimental Botany, Vol. 58, No. 8, pp. 2053–2067, 2007 doi:10.1093/jxb/erm059 Advance Access publication 26 April, 2007 RESEARCH PAPER Functional analysis of the essential bifunctional tobacco enzyme 3-dehydroquinate dehydratase/shikimate dehydrogenase in transgenic tobacco plants Li Ding 1, *, Daniel Hofius 1,† , Mohammad-Reza Hajirezaei 1 , Alisdair R. Fernie 2 , Frederik Bo ¨ rnke 1,‡ and Uwe Sonnewald 1,‡,§ 1 Institut fu¨r Pflanzengenetik und Kulturpflanzenforschung (IPK), Corrensstrasse 3, 06466 Gatersleben, Germany 2 Max-Planck-Institute for Molecular Plant Physiology, Department of Lothar Willmitzer, 14476 Golm-Potsdam, Germany Received 17 January 2007; Revised 27 February 2007; Accepted 28 February 2007 Abstract In plants, the shikimate pathway occurs in the plastid and leads to the biosynthesis of aromatic amino acids. The bifunctional 3-dehydroquinate dehydratase/shiki- mate dehydrogenase (DHD/SHD) catalyses the conver- sion of dehydroquinate into shikimate. Expression of NtDHD/SHD was suppressed by RNAi in transgenic tobacco plants. Transgenic lines with <40% of wild- type activity displayed severe growth retardation and reduced content of aromatic amino acids and down- stream products such as cholorogenic acid and lignin. Dehydroquinate, the substrate of the enzyme, accu- mulated. However, unexpectedly, so did the pro- duct, shikimate. To exclude that this finding is due to developmental differences between wild-type and transgenic plants, the RNAi approach was additionally carried out using a chemically inducible promoter. This approach revealed that the accumulation of shikimate was a direct effect of the reduced activity of NtDHD/SHD with a gradual accumulation of both dehydroquinate and shikimate following induction of gene silencing. As an explanation for these findings the existence of a parallel extra-plastidic shikimate pathway into which dehydroquinate is diverted is proposed. Consistent with this notion was the identifi- cation of a second DHD/SHD gene in tobacco (NtDHD/ SHD-2) that lacked a plastidic targeting sequence. Expression of an NtDHD/SHD-2–GFP fusion revealed that the NtDHD/SHD-2 protein is exclusively cytosolic and is capable of shikimate biosynthesis. However, given the fact that this cytosolic shikimate synthesis cannot complement loss of the plastidial pathway it appears likely that the role of the cytosolic DHD/SHD in vivo is different from that of the plastidial enzyme. These data are discussed in the context of current models of plant intermediary metabolism. Key words: 3-Dehydroquinate dehydratase/shikimate dehydro- genase, Nicotiana tabacum, RNA interference, secondary metabolism, shikimate pathway. Introduction The shikimate pathway is central for the biosynthesis of aromatic amino acids, folates, vitamins, and a number of aromatic compounds and secondary metabolites in bacte- ria, plants, fungi, and apicomplexa parasites but is absent from humans and other higher animals (Bentley, 1990; Herrmann, 1995; Herrmann and Weaver, 1999). In plants, aromatic amino acids are used not only as protein building blocks, but also as precursors for a large number of secondary metabolites (i.e. pigments, flavonoids, auxins, * Present address: Max-Planck-Institute of Molecular Cell Biology and Genetics, Pfotenhauerstr. 108, 01307 Dresden, Germany y Present address: Department of Molecular Biology, Copenhagen Biocenter, University of Copenhagen, Ole Maaloes Vej 5, 2200 Copenhagen N, Denmark z Present address: Friedrich-Alexander-Universita ¨ t, Lehrstuhl fu ¨ r Biochemie, Staudtstr. 5, 91058 Erlangen, Germany § To whom correspondence should be addressed. E-mail: [email protected] Abbreviations: DAHP, 3-deoxy-D-arabino-heptulosonate-7-phosphate; DHD/SHD, 3-dehydroquinate dehydratase/shikimate dehydrogenase; EPSP, enolpyruvylshikimate-3-phosphate; EPSPS, enolpyruvylshikimate-3-phosphate synthase; PEP, phosphoenolpyruvate; NADP, nicotinamide adenine dinucleotide phosphate; TCA, trichloracetic acid. ª The Author [2007]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved. For Permissions, please e-mail: [email protected] by guest on July 30, 2014 http://jxb.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Functional analysis of the essential bifunctional tobacco enzyme 3-dehydroquinate...

Journal of Experimental Botany, Vol. 58, No. 8, pp. 2053–2067, 2007
doi:10.1093/jxb/erm059 Advance Access publication 26 April, 2007
RESEARCH PAPER
Functional analysis of the essential bifunctional tobaccoenzyme 3-dehydroquinate dehydratase/shikimatedehydrogenase in transgenic tobacco plants
Li Ding1,*, Daniel Hofius1,†, Mohammad-Reza Hajirezaei1, Alisdair R. Fernie2, Frederik Bornke1,‡ and
Uwe Sonnewald1,‡,§
1 Institut fur Pflanzengenetik und Kulturpflanzenforschung (IPK), Corrensstrasse 3, 06466 Gatersleben, Germany2 Max-Planck-Institute for Molecular Plant Physiology, Department of Lothar Willmitzer, 14476 Golm-Potsdam,Germany
Received 17 January 2007; Revised 27 February 2007; Accepted 28 February 2007
Abstract
In plants, the shikimate pathway occurs in the plastid
and leads to the biosynthesis of aromatic amino acids.
The bifunctional 3-dehydroquinate dehydratase/shiki-
mate dehydrogenase (DHD/SHD) catalyses the conver-
sion of dehydroquinate into shikimate. Expression of
NtDHD/SHD was suppressed by RNAi in transgenic
tobacco plants. Transgenic lines with <40% of wild-
type activity displayed severe growth retardation and
reduced content of aromatic amino acids and down-
stream products such as cholorogenic acid and lignin.
Dehydroquinate, the substrate of the enzyme, accu-
mulated. However, unexpectedly, so did the pro-
duct, shikimate. To exclude that this finding is due
to developmental differences between wild-type and
transgenic plants, the RNAi approach was additionally
carried out using a chemically inducible promoter.
This approach revealed that the accumulation of
shikimate was a direct effect of the reduced activity
of NtDHD/SHD with a gradual accumulation of both
dehydroquinate and shikimate following induction of
gene silencing. As an explanation for these findings
the existence of a parallel extra-plastidic shikimate
pathway into which dehydroquinate is diverted is
proposed. Consistent with this notion was the identifi-
cation of a second DHD/SHD gene in tobacco (NtDHD/
SHD-2) that lacked a plastidic targeting sequence.
Expression of an NtDHD/SHD-2–GFP fusion revealed
that the NtDHD/SHD-2 protein is exclusively cytosolic
and is capable of shikimate biosynthesis. However,
given the fact that this cytosolic shikimate synthesis
cannot complement loss of the plastidial pathway it
appears likely that the role of the cytosolic DHD/SHD in
vivo is different from that of the plastidial enzyme.
These data are discussed in the context of current
models of plant intermediary metabolism.
Key words: 3-Dehydroquinate dehydratase/shikimate dehydro-
genase, Nicotiana tabacum, RNA interference, secondary
metabolism, shikimate pathway.
Introduction
The shikimate pathway is central for the biosynthesis ofaromatic amino acids, folates, vitamins, and a number ofaromatic compounds and secondary metabolites in bacte-ria, plants, fungi, and apicomplexa parasites but is absentfrom humans and other higher animals (Bentley, 1990;Herrmann, 1995; Herrmann and Weaver, 1999). In plants,aromatic amino acids are used not only as protein buildingblocks, but also as precursors for a large number ofsecondary metabolites (i.e. pigments, flavonoids, auxins,
* Present address: Max-Planck-Institute of Molecular Cell Biology and Genetics, Pfotenhauerstr. 108, 01307 Dresden, Germanyy Present address: Department of Molecular Biology, Copenhagen Biocenter, University of Copenhagen, Ole Maaloes Vej 5, 2200 Copenhagen N, Denmarkz Present address: Friedrich-Alexander-Universitat, Lehrstuhl fur Biochemie, Staudtstr. 5, 91058 Erlangen, Germany§ To whom correspondence should be addressed. E-mail: [email protected]: DAHP, 3-deoxy-D-arabino-heptulosonate-7-phosphate; DHD/SHD, 3-dehydroquinate dehydratase/shikimate dehydrogenase; EPSP,enolpyruvylshikimate-3-phosphate; EPSPS, enolpyruvylshikimate-3-phosphate synthase; PEP, phosphoenolpyruvate; NADP, nicotinamide adeninedinucleotide phosphate; TCA, trichloracetic acid.
ª The Author [2007]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
phytoalexins, lignin, and tannins) (Herrmann, 1995). Inaddition, pathway intermediates may serve as substratesfor a number of other metabolic pathways includingbiosynthesis of quinate and derived products such aschlorogenate (Herrmann and Weaver, 1999). It has beenestimated that, under normal growth conditions, 20% ofthe carbon fixed by plants can be directed towards theshikimate pathway (Haslam, 1993).The shikimate pathway consists of seven metabolic steps
beginning with the condensation of phosphoenolpyruvate(PEP) and erythrose 4-phosphate to form 3-deoxy-D-arabino-heptulosonate-7-phosphate (DAHP) and ending withthe synthesis of chorismate from 5-enolpyruvylshikimate3-phosphate (EPSP). The organization of the sevenenzymes is markedly different between the three king-doms. While the seven enzymatic steps in most prokar-oytes are catalysed by monofunctional enzymes; in fungi,reactions 2–6 are catalysed by the multifunctional AROMcomplex.In plants, the shikimate pathway is believed to be con-
fined to plastids (Herrmann, 1995; Schmid and Amrhein,1995; Mustafa and Verpoorte, 2005; Weber et al., 2005).This is supported by biochemical and molecular data.Most shikimate pathway enzymes are synthesized as pre-cursors with an N-terminal extension, allowing the post-translational uptake of the protein into chloroplasts(Della-Cioppa et al., 1986; Schmid et al., 1992). Thus,the shikimate pathway appears to be firmly established inplastids. However, it has been proposed that the chloroplast-localized biosynthetic activity does not account for 100%of the aromatic amino acid biosynthesis observed.Hence, a spatially separated pathway in the cytosol hasbeen proposed (Buchholz et al., 1979). Evidence fora cytosolic shikimate pathway comes from the isolation ofDAHP synthase (Ganson et al., 1986), EPSP synthase(Mousdale and Coggins, 1985), and chorismate mutase(d’Amato, 1984) isoforms lacking N-terminal transitpeptides. Moreover, since plastid-targeted shikimate ki-nase (Schmid et al., 1992) and EPSPS synthase (Della-Cioppa et al., 1986) are active when they retain theirtransit peptides, these enzymes could also potentially beconstituents of a cytosolic shikimate pathway. However,to date, operation of a complete extra-plastidial pathwayhas not been established.In plants, steps three and four of the shikimate pathway
are catalysed by the bifunctional enzyme 3-dehydroquinatedehydratase (EC 4.2.1.10)/shikimate dehydrogenase (EC1.1.1.25) (DHD/SHD), which catalyses the dehydration ofdehydroquinate to dehydroshikimate and the reversiblereduction of dehydroshikimate to shikimate, respectively(Fig. 1A). Three different classes of SHDs have been iden-tified across kingdoms on the basis of phylogenetic andbiochemical analysis: shikimate dehydrogenase (AroE),shikimate/quinate dehydrogenase (YdiB), and shikimatedehydrogenase-like (SHD-L) (Michel et al., 2003; Singh
et al., 2005). All three classes of SHDs can catalyse theoxidation of shikimate in the presence of nicotinamideadenine dinucleotide phosphate (NADP+) in vitro, albeitwith varying efficiencies. The natural substrate of theSHD-L, however, has not yet been determined (Singhet al., 2005). The plant SHD has been shown to be theAroE type (Singh and Christendat, 2006).
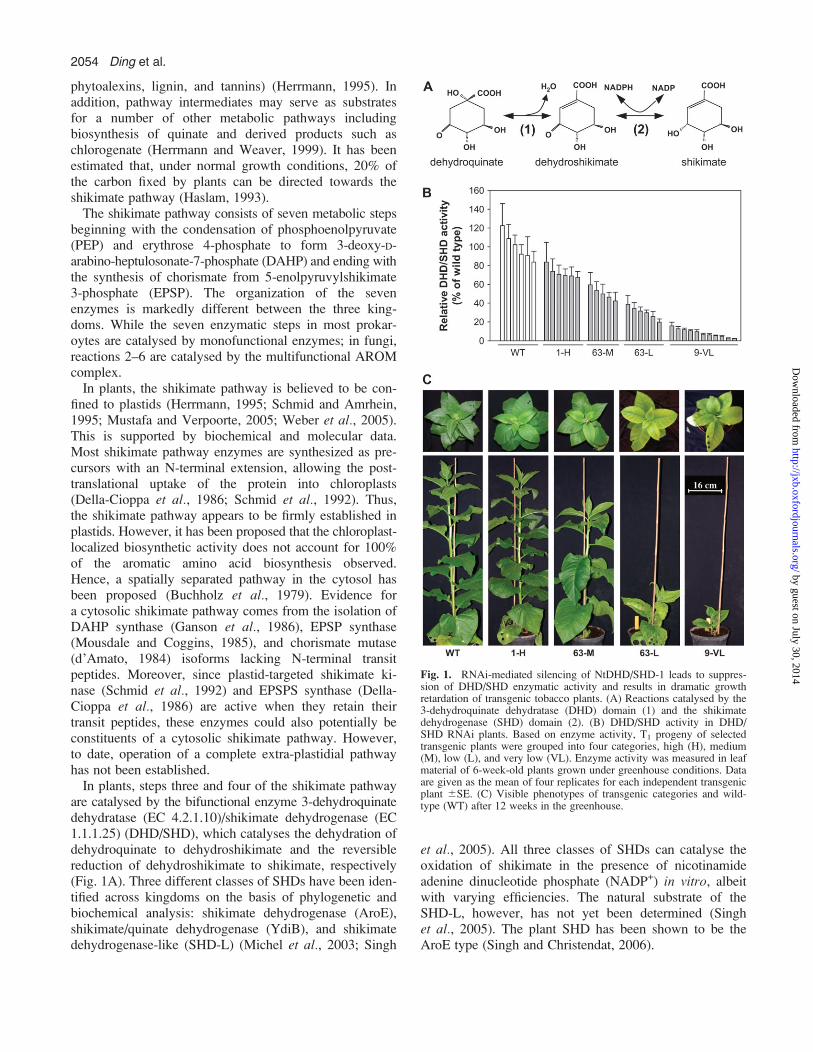
Fig. 1. RNAi-mediated silencing of NtDHD/SHD-1 leads to suppres-sion of DHD/SHD enzymatic activity and results in dramatic growthretardation of transgenic tobacco plants. (A) Reactions catalysed by the3-dehydroquinate dehydratase (DHD) domain (1) and the shikimatedehydrogenase (SHD) domain (2). (B) DHD/SHD activity in DHD/SHD RNAi plants. Based on enzyme activity, T1 progeny of selectedtransgenic plants were grouped into four categories, high (H), medium(M), low (L), and very low (VL). Enzyme activity was measured in leafmaterial of 6-week-old plants grown under greenhouse conditions. Dataare given as the mean of four replicates for each independent transgenicplant 6SE. (C) Visible phenotypes of transgenic categories and wild-type (WT) after 12 weeks in the greenhouse.
2054 Ding et al.
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

The ratio of SHD to DHD activity has been reportedto be 9:1; thus dehydroquinate is readily converted toshikimate without accumulation of dehydroshikimate(Fiedler and Schultz, 1985). Bifunctional enzymes playimportant roles in regulating the partitioning of metabolitesat branch points in metabolic pathways to ensure sufficientproduction of a desired compound. DHD/SHD is found atthe branch point for quinate synthesis. Dehydroquinate anddehydroshikimate, the substrates for DHD and SHD,respectively, can be partitioned towards quinate metabo-lism (Bentley, 1990). Substrate channelling from the DHDto the SHD site will ensure a constant flux through theshikimate pathway for chorismate biosynthesis.Partial or full-length cDNA clones encoding DHD/SHD
of pea (Deka et al., 1994), tobacco (Bonner and Jensen,1994), and tomato (Bischoff et al., 2001) have beenisolated. In 1994, Bonner and Jensen isolated a partialcDNA encoding tobacco DHD/SHD by screening a Nico-tiana tabacum cDNA expression library with a DHD/SHD-specific polyclonal antibody. This cDNA comprisedthe entire coding region of the mature enzyme and anadditional 69 upstream nucleotides, postulated to encodepart of the transit peptide for plastidic targeting. Themature sequence translates into a protein with both enzy-matic activities, the dehydratase at the amino-terminal andthe dehydrogenase at the carboxyl-terminal part. The puta-tive transit peptide is rich in hydroxylated amino acid re-sidues but has a net negative charge, a feature not seen forother chloroplast transit peptides (Gavel and von Heijne,1990). In 2001, Bischoff and co-workers cloned a full-length cDNA encoding tomato DHD/SHD (Bischoff et al.,2001). The amino acid sequence deduced revealed thatDHD/SHD is most probably synthesized as a precursor witha very short (13 amino acids) plastid-specific transit peptide,which was highly similar to the tobacco counterpart.To elucidate the physiological role of DHD/SHD in
plants, its activity was decreased by RNAi in transgenictobacco plants. Resulting plants were thoroughly investi-gated for growth and biochemical and molecular parameters.Silencing of DHD/SHD resulted in a dramatic reduction ofaromatic compounds and a strongly reduced growth oftransgenic plants. Unexpectedly, DHD/SHD-deficient plantsaccumulated shikimate, the product of DHD/SHD activity.Stimulated by this rather unexpected result a second DHD/SHD isoform, capable of synthesizing shikimate within thecytosol, was isolated and biochemically characterized. Thisnewly isolated cytosolic DHD/SHD isoform was designatedNtDHD/SHD-2 and seems likely to be involved in theshikimate synthesis observed in the cytosol.
Materials and methods
Plant material and growth conditions
Tobacco plants (Nicotiana tabacum cv. Samsun NN) were grown intissue culture under a 16 h light/8 h dark regime (150 lM quanta
m�2 s�1 light, 21 �C) at 50% humidity on Murashige–Skoogmedium (Sigma, St Louis, MO, USA) containing 2% (w/v) Suc. Forbasic molecular and biochemical characterization, kanamycin-selected plants of the T1 generation were cultivated in soil ina greenhouse with 16 h supplemental light (;250 lM m�2 s�1)followed by 8 h darkness. The temperature regime followed theday/night cycle with 25 �C/18 �C. Detailed investigation of growthparameters was performed with plants grown on sand undergreenhouse conditions and watered daily with a nutrient solution asdescribed (Geiger et al., 1999).
Cloning of full-length NtDHD/SHD-1 from tobacco and
expression in Escherichia coli
To obtain a full-length DHD/SHD clone from Nicotiana tabacum,the lacking 5# end of a published DHD/SHD sequence (Bonner andJensen, 1994; GenBank accession no. L32794) was amplified byRACE using the SMART RACE cDNA amplification kit (Clontech,Palo Alto, CA, USA) and a gene-specific primer (5#-TTCATCAC-CAGCATACTGACCCCCTTCCC-3#). The resulting PCR product(;500 bp) was ligated into the pCR-blunt vector (Invitrogen,Carlsbad, CA, USA) and sequenced. Sequence information allowedassembly of a putative full-length cDNA using the InforMaxVectorNTI software (InforMax, Inc., USA). Finally, the entirecDNA clone, designated NtDHD/SHD-1, was amplified by RT-PCRfrom tobacco leaves using M-MLV[H-] reverse transcriptase(Promega, Madison, WI, USA) and oligo(dT)[30]V primer forreverse transcription, as well as Pfu polymerase (Stratagene) andgene-specific primers (5#-GGGGACTTTTTCAAGCTGGCTGCT-3# and 5#-TTGATACAAATCATACAATAATGTG-3#) for PCRamplification. Identity of the sequence was verified and submittedto GenBank/DDBJ/EMBL (accession no. AY578144).For expression in E. coli, the coding region of mature NtDHD/
SHD-1 protein lacking the apparent transit peptide [nucleotides(nts) 118–1785] was PCR amplified with gene-specific primers5#-GGATCCGGGGAGGCAATGACGAGGAAC-3# and 5#-CTGCAGTTAATTCCTCCGAAGCACAAATGG-3#. After sub-cloning into pCR-blunt (Promega), the PCR fragment was ligatedinto the BamHI and PstI sites of pQE9 and transferred into E. coliM15(pREP4) cells for protein expression (Qiagen, Hilden, Germany).Recombinant E. coli cells were harvested 3–4 h after induction with0.05 mM isopropylthio-b-galactoside, and soluble 63 His-taggedNtDHD/SHD-1 protein was purified under native conditions usingnickel–nitrilotriacetic acid agarose according to the manufacturer’sinstruction.
RNAi plasmid construction and tobacco transformation
To generate the NtDHD/SHD-1 RNAi construct, a partial frag-ment of tobacco NtDHD/SHD-1 (GenBank, AY578144) comprisingnts 816–1368 was PCR amplified using the primers5#-GGATCCGTTGGGACCAGATACCAGAATA-3# and 5#-GTCGACGCTAAGCTCGTCAAGAGACAAAG-3# (BamHI andSalI restriction sites are underlined). The resulting PCR productwas subcloned into a pCR-blunt vector and subsequently inserted asa BamHI/SalI fragment in sense orientation downstream of theGA20oxidase intron in the pUC-RNAi vector as described by Chenet al. (2003). The same fragment was ligated in antisenseorientation into the BglII/XhoI sites of pUC-RNAi already carryingthe sense fragment. Finally, the resulting RNAi fragment wasexcised from pUC-RNAi using the flanking PstI sites and insertedinto the SbfI site of pBinAR (Hofgen and Willmitzer, 1990)between the CaMV 35S promoter and ocs terminator, yielding theconstruct pBin-NtDHD/SHD-1-RNAi. The construct Alc-NtDHD/SHD-1-RNAi for inducible silencing was prepared using thestrategy detailed in Chen et al. (2003), employing the same genefragment as above.
Molecular analysis of shikimate synthesis in tobacco 2055
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Transformation of tobacco plants by Agrobacterium-mediatedgene transfer using A. tumefaciens C58,C1,pGV2260 was carriedout as described by Rosahl et al. (1987).
Cloning of NtDHD/SHD-2, binary plasmid construction, and
Agro-infiltration
A partial 270 bp EST clone homologous to tobacco NtDHD/SHD-1was identified from a sequenced tobacco cDNA library andamplified by RT-PCR from tobacco leaf material using gene-specific primers (5#-TCGAAGAGCAGGAGAGCTGAAATT-3#and 5#-GCGTCTTTCACGCAGAGTAGTCTTT-3#). The resultingPCR fragment was ligated into pCR-blunt and sequenced to verifythe identity of the NtDHD/SHD isoform. 5# and 3# fragments of 1.2and 0.7 kb, respectively, were obtained by RACE using theSMART RACE cDNA amplification kit (Clontech, Palo Alto, CA,USA) and gene-specific primers (5#-RACE, 5#-GCGTCTTTCACG-CAGAGTAGTCTTTCTA-3#; 3#-RACE, 5#-TCGAAGAGCAG-GAGAGCTGAAATTGTGA-3#). PCR products were cloned intopCR-blunt, and sequences were assembled by the softwareVectorNTI (InforMax, Inc.). Finally, the full-length cDNA clonewas amplified by RT-PCR from tobacco leaves using RACEfragment-deduced oligonucleotides, and the entire sequence, desig-nated NtDHD/SHD-2, was determined (GenBank accession no.AY578143).To create a binary construct for transient expression in plants,
the entire coding region of NtDHD/SHD-2 was amplified by PCRusing the primers 5#-GTCGACATGGGTAGTGTTGGATTGTTG-3#and 5#-GTCGACTTAGAATTTTGCCATAACGATGT-3#, therebyintroducing SalI restriction sites (underlined) at the 5# end and 3#end, respectively. After subcloning, the PCR fragment was insertedinto the SalI site of pBinAR downstream of the CaMV 35S promoterto yield pBin-NtDHD/SHD-2.For infiltration of N. tabacum leaves, the A. tumfaciens strain
C58C1 was infiltrated into the abaxial air space of 4-week-oldplants as described (Voinnet et al., 2003). The p19 protein oftomato bushy stunt virus was used to suppress gene silencing. Co-infiltration of Agrobacterium strains containing pBin-NtDHD/SHD-2and the p19 construct was carried out at OD600 of 1.0.
Construction of fusion proteins with GFP variants and
microprojectile bombardment
To obtain reporter gene fusion constructs for transient expression inleaf tissue, the coding regions of the respective NtDHD/SHDisoform were either cloned by RT-PCR from leaf tissue (NtDHD/SHD-1) or PCR amplified from the plasmid pBin-NtDHD/SHD-2,using primers 5#-CCCGGGATGGAGTTGGTAGTGGATTC-AGG-3# and 5#-TCTAGAATTCCTCCGAAGCACAAATGGTA-3#for NtDHD/SHD-1, and primers 5#-CCCGGGATGGGTAGTG-TTGGATTGTTGAA-3# and 5#-TCTAGAGAATTTTGCCAT-AACGATGTCGC-3# for NtDHD/SHD-2, respectively. Theoligonucleotides were designed to delete the stop codon and tointroduce SmaI sites at the 5# end and XbaI sites at the 3# end,respectively. PCR fragments were subcloned and inserted into theSmaI/XbaI sites of a pFF19-based vector (Timmermans et al., 1990)containing either mGFP5, EYFP, or ECFP instead of the GUS gene(pFF19–GFP, pFF19–YFP, pFF19–CFP, kindly provided by AWachter, Heidelberg, Germany), to yield plasmids pFF19–NtDHD/SHD-1,YFP, pFF19–NtDHD/SHD-2,GFP, or pFF19-NtDHD/SHD-2,CFP. DNA-plasmids were coated onto 1 lm gold particles, and theabaxial epidermis of tobacco was bombarded either with a singleconstruct (pFF19–GFP vector, pFF19–NtDHD/SHD-2,GFP) or simul-taneously with two constructs (pFF19–NtDHD/SHD-1,YFP andpFF19–NtDHD/SHD-2,GFP) using a Helios Gene Gun accordingto the manufacturer’s protocol (Bio-Rad Laboratories, Hercules,
CA, USA). Images were obtained with a confocal microscope LSM510 META (Carl Zeiss, Gottingen, Germany). Excitation light of458, 488, and 514 nm produced by an argon laser and differentemission filters of 475–510, 510–525, and 520–545 nm alloweddetection of CFP, GFP, or YFP, respectively. Images weresuperimposed with scans of chlorophyll-derived red fluorescenceand transmission images by means of the Zeiss LSM Version 3.0software.
Measurement of DHD/SHD enzyme activity
DHD/SHD enzyme activity of recombinant NtDHD/SHD-1 proteinand of tobacco plants was determined with shikimate as substrateby quantifying the production of NADPH spectrophotometrically at334 nm. An aliquot (0.5 lg) of purified 63 His-NtDHD/SHD-1protein (see above) was incubated in 800 ll of reaction buffer(200 lM NADP in 100 mM glycine–NaOH buffer, pH 9.0) at roomtemperature for 3 min and the reaction was started by adding 2 ll of100 mM shikimate. For determination of DHD/SHD activity instable transformants or agro-infiltrated leaves, 500 mg of leafmaterial was homogenized in liquid nitrogen and extracted with500 ll of protein extraction buffer [50 mM TRIS–HCl, pH 6.8,5 mM MgCl2, 5 mM mercaptoethanol, 15% (v/v) glycerol, 1 mMEDTA, 1 mM EGTA, and 0.1 mM Pefabloc protease inhibitor].Twenty microlitres of leaf extracts were incubated in 780 ll ofreaction buffer (1 mM NADP in 100 mM glycine–NaOH buffer,pH 9.0) for 5 min at room temperature, and the reaction was startedin the spectrophotometer (Uvikon 922, Kontron Instruments) afterreaching a stable baseline by addition of shikimate as described.
Enzymatic determination of shikimate and
dehydroshikimate/dehydroquinate
Shikimate was determined enzymatically in plant extracts bymeasuring the turnover of NADP to NADPH following the additionof recombinant DHD/SHD protein (see above). Five hundredmilligrams of leaf tissue were extracted with trichloroacetic acid(TCA) as described previously (Jelitto et al., 1992), and 50 ll of theneutralized extract [16% (v/v) TCA] were incubated in 550 ll ofreaction buffer 1 (100 lM NADP in 100 mM glycine–NaOHbuffer, pH 9.0). The production of NADPH was determinedspectrophotometrically at 334 nm with 203 multiplication usinga Sigma ZWS-II photometer (Sigma, Darmstadt, Germany), and thereaction was started by adding 0.5 lg of purified DHD/SHDenzyme. Calculation was based on calibration curves prepared with1–20 nmol shikimate ml�1 giving rise to the linear equationy¼1.0504x+0.1974 (R2¼0.9993). Recovery experiments were un-dertaken to confirm the accuracy of the measurements. A definedportion of shikimate (1–20 nmol ml�1) was added to the leafmaterial before extraction and determined exactly as described. Therecovery rates were between 92% and 101% of the amount added.Since DHD/SHD catalyses a reversible reaction, dehydroquinate/
dehydroshikimate was determined in the same extract when thereaction was carried out in the reverse direction. Ten micrilitres ofTCA extract was incubated in 590 ll of reaction buffer 2 (100 lMNADPH in 100 mM HEPES–KOH buffer, pH 7.2) and the turnoverof NADPH to NADP was followed at 334 nm with 10 timesmultiplication after addition of recombinant DHD/SHD protein asdescribed.
Carbohydrate and amino acid determination
Soluble sugars and starch were determined in leaf samples extractedwith 80% (v/v) ethanol/20 mM HEPES–KOH pH 7.5 as described(Stitt et al., 1989). Amino acid contents were measured from thesame extracts with HPLC as described (Rolletschek et al., 2002).
2056 Ding et al.
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Single amino acids were identified by co-chromatography withauthentic standards and quantified by comparison with internalstandards.
Chlorophyll determination
Chlorophyll was measured in ethanol extracts and concentrationswere determined as described in Lichtenthaler (1987).
Chlorogenic acid determination
Leaf samples (100 mg) were homogenized in liquid nitrogen andsuccessively extracted with 500, 300, and 200 ll of methanol(gradient grade; Sigma-Aldrich, Germany). Supernatants of eachextraction step were collected after centrifugation (16 000 g, 5 min)and the pooled samples were used for further analysis. Prior to themeasurements, the methanol concentration of the sample wasbrought up to 20 % using buffer A (9.5% acetonitrile and 1% aceticacid in purest H2O). A Waters HPLC system (Waters, Eschborn,Germany) consisting of a gradient pump (Waters no. 600), a degas-ing module, an autosampler (no. 717), and a UV detector (no. 417)was used for determination of chlorogenic acid. The gradient wasaccomplished with buffer A and buffer B (100 % acetonitrile). Forseparation of chlorogenic acid, a reversed phase column [Luna 5lC18(2), 25034.6 mm; Phenomenex, Aschaffenburg, Germany] wasused. The column was equilibrated with buffer A at a flow rate of1 ml min�1 and chlorogenic acid was recorded by the UV detectorat 280 nm.
Lignin determination
Lignin content was determined spectrophotometrically (280 nm)using thioglycolic acid following the protocol as described byCampbell and Ellis (1992). Calculation was done according toa calibration curve prepared with pure lignin (5–25 mg). Ligninstaining of transverse sections of petioles was performed by meansof phloroglycinol–HCl reagent essentially as described by Biemeltet al. (2004). Tobacco petioles (6 weeks old) were decolorized in70% (v/v) ethanol for 6–8 h, washed with distilled water, and slicedby scalpel. The sections were stained in 1% (w/v) phloroglucinol(Sigma-Aldrich) for 1–2 h and analysed using a light microscope(Axioscop, Zeitz, Germany).
Shikimate feeding experiment
Excised leaf discs from mature tobacco leaves (about 40 mg) werefloated on 50 mM HEPES (pH 7.0) buffer solution containing 20 mMshikimate at room temperature for 16 h. As a control, leaf discs wereexposed to the buffer solution without shikimate. Afterwards, the leafdiscs were washed three times with distilled water, dried on filterpaper, and frozen in liquid nitrogen for further analysis.
RNA isolation and northern blot analysis
Total RNA was extracted from tobacco leaf material as described byLogemann et al. (1987). Thirty micrograms of RNA per sample wereseparated on a 1.5% (w/v) formaldehyde–agarose gel, transferred toa nitrocellulose membrane (Genescreen; NEN Lifescience Products,Boston, MA, USA), and fixed by UV-crosslinking. Radioactivelabelling of cDNA fragments was performed using the High Prime-Kit (Roche, Mannheim, Germany) and [a-32P]CTP. Hybridizationwas carried out as described previously (Chen et al., 2003) andsignals were detected and quantified by means of a phosphoimaginganalyser (FLA-3000, Fujifilm, Tokyo, Japan). Shikimate pathwaygene-specific cDNAs used for hybridization were as follows:DAHPS (3-deoxy-D-heptulosonate-7-phosphate synthase; GenBankaccession no. M64261), DHQS (dehydroquinate synthase; accession
no. L46847), DHD/SHD (3-dehydroquinate dehydratase/shikimatedehydrogenase; accession no. L32794), SK (shikimate kinase;accession no. X63560), EPSPS (enolpyruvylshikimate-3-phosphatesynthase; accession no. M61904), and CS (chorismate synthase;accession no. Z21791).
Western blot analysis
Protein extraction and western blot analysis followed the protocoldescribed in Hajirezaei et al. (2002). Soluble protein amounts oftobacco leaf extracts were determined according to Bradford (1976)and adjusted to 1 lg protein ll�1 final concentration. Twentymicrograms of total protein were separated by 12.5 % SDS–PAGEand electro-transferred to a nitrocellulose membrane (Porablot;Macherey-Nagel, Duren, Germany). Immunoreaction was carriedout with a polyclonal anti-NtDHD/SHD-1 generated againstaffinity-purified 63 His-tagged NtDHD/SHD-1 protein (see above)in rabbits using custom service from Eurogentec (Seraing, Belgium).Development of the immunoblots was performed with the Super-signal West Pico Chemiluminescent Substrate system (Pierce,Rockford, IL, USA), after incubation with an anti-rabbit horseradishperoxidase-conjugated secondary antibody (Pierce).
Results
Biochemical characterization of recombinantNtDHD/SHD-1
A cDNA fragment encoding the predicted matureNtDHD/SHD-1 was PCR amplified from tobacco cDNAand heterologously expressed in E. coli. The recombinantprotein was purified to apparent homogeneity using metal-affinity purification (data not shown), and this enzymepreparation was used to determine a number of bio-chemical parameters. Enzyme pH preference was de-termined in both the forward and reverse reaction indifferent buffer systems. The pH optimum for the anabolicreaction (from dehydroquinate to shikimate) was between6.8 and 7.2. For the catabolic reaction, a higher pHoptimum was determined. The reaction velocity reacheda maximum at pH 9.0 and pH 9.4. The effect oftemperature on NtDHD/SHD-1 activity was determinedfor both the forward and reverse direction. The velocity ofthe NtDHD/SHD-1 catalysed reaction rose linearly from5 �C to 30 �C, reaching a maximum value at 30 �C, whichremained constant until 45 �C. Thereafter, the reactionvelocity dropped rapidly as a result of the deactivation ofthe protein (data not shown). Thus the optimum temper-ature for NtDHD/SHD-1 was determined to be between30 �C and 45 �C. DHD/SHD catalyses two neighbouringreversible reactions. Limited by the detection of dehydro-shikimate, only the shikimate–NADP and dehydroquinate–NADPH pairs could be employed to determine the kineticproperties of NtDHD/SHD-1. The SHD domain, whichcatalysed the oxidation of shikimate to dehydroshikimate,exhibited typical Michaelis–Menten kinetics with Km(NADP)
of 3167 lM and Km(shikimate) of 130615 lM (data notshown).
Molecular analysis of shikimate synthesis in tobacco 2057
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

RNA interference of NtDHD/SHD-1 strongly inhibitsDHD/SHD activity in transgenic tobacco plants
To analyse functionally the in planta role of NtDHD/SHD-1, the corresponding transcript was targeted throughan RNA interference (RNAi) strategy. To this end, afragment of the tobacco NtDHD/SHD-1, ranging from nt816 to nt 1368 of the respective cDNA clone (GenBank,AY578144), was amplified by PCR and inserted first inantisense orientation downstream of the CaMV 35Spromoter, followed by a short intron, and by the sameNtDHD/SHD fragment, but in sense orientation. This re-sulting plasmid pBin-DHD/SHD-RNAi was transformedinto tobacco plants by Agrobacterium-mediated genetransfer, and 80 kanamycin-resistant primary transform-ants were screened by RNA blot hybridization aftertransfer into the greenhouse. Fourteen transgenic plantsshowed decreased transcript levels (data not shown), anddepletion of DHD/SHD was further confirmed by enzymeactivity measurement. Six independent transgenic lineswith 16–87% remaining DHD/SHD activities were finallyselected for detailed analysis in the T1 generation. Basedon enzyme activities, DHD/SHD RNAi plants from the T1
generation were classified into four groups: high (H,DHD/SHD activities 85–60%); middle (M, DHD/SHD ac-tivities 60–40%); low (L, DHD/SHD activities 40–20%);and very low (VL, DHD/SHD activities 20–5%) (Fig. 1B).
Silencing of NtDHD/SHD-1 severely inhibits growth oftransgenic tobacco plants
Silencing of NtDHD/SHD-1 led to dramatic phenotypicalchanges of the transgenic tobacco plants. A 40–60% re-duction of DHD/SHD activity (group M plants) led tochlorotic leaves. Bleached areas developed in intercostalregions of mature leaves. A further reduction (60–80%)resulted in a dwarfen phenotype. When DHD/SHD ac-tivity dropped below 20% of wild type, transgenic plantsstopped growing and frequently died in 1–2 weeks aftertransfer from tissue culture to the greenhouse (Fig. 1C).To correlate the reduction in DHD/SHD activity with
biomass production, NtDHD/SHD-1-RNAi plants (T1
generation) were germinated and cultivated in sand cultureas described in Materials and methods. Growth parameterswere acquired by measuring fresh weight (FW), dryweight (DW), and plant height (Table 1). An up to 40%reduction of DHD/SHD did not result in growth re-tardation. Whereas 40% to 60% reduction of enzymeactivity led to a 33% reduction in fresh weight, a 42%reduction in dry weight, and a 37% reduction in plantheight, respectively. In plants with 20% or lower DHD/SHD activity, biomass production severely decreased to7% (fresh weight and dry weight) and 36% (plant height)as compared with wild-type plants. These results indicatethat the loss of DHD/SHD activity leads to a concomitantreduction in biomass production of transgenic plants.
The shikimate pathway links primary and secondarymetabolism. Under normal growth conditions, it was esti-mated that about 20% of fixed carbon is directed towardsthe shikimate pathway (Haslam, 1993). To determinewhether reduced flux towards the shikimate pathwaywould influence primary metabolism, carbohydrate con-tents in DHD/SHD-silenced plants were determined.Similar amounts of carbohydrates were detected in allDHD/SHD RNAi plants when compared with wild type(data not shown). This result suggests that the inhibitionof DHD/SHD had no effect on primary metabolism
Silencing of NtDHD/SHD-1 leads to accumulation ofdehydroquinate and shikimate
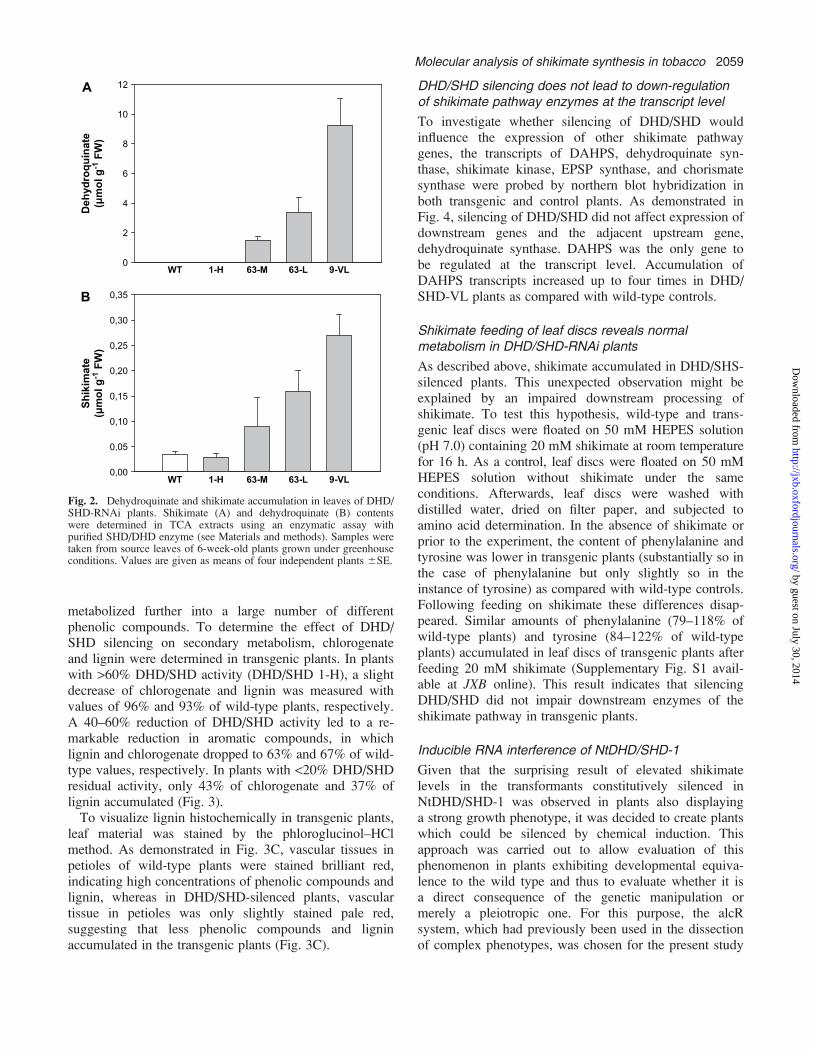
Inhibition of DHD/SHD activity was expected to lead toan accumulation of the substrate (dehydroquinate) and adepletion of the product (shikimate) of the enzymereaction. Therefore, dehydroquinate and shikimate levelswere determined in leaf extracts of wild-type and trans-genic plants using the enzymatic assay mentioned above.In wild-type plants and NtDHD/SHD-1 RNAi plants with>60% DHD/SHD activity, no dehydroquinate could bedetected (Fig. 2A), whereas in plants with <60% residualDHD/SHD activity, 1.7–9.3 lmol of dehydroquinate weremeasured per gram leaf material (fresh weight). Accumu-lation of dehydroquinate in these plants correlated withthe reduction in DHD/SHD activity (see Fig. 1B). Un-expectedly, shikimate, the product of DHD/SHD, alsoaccumulated in DHD/SHD RNAi plants; 3.5- to 8-foldhigher levels of shikimate were detected in plants with<60% residual DHD/SHD activity as compared with wild-type controls (Fig. 2B), whereas in plants with >60%DHD/SHD activity, no shikimate accumulation wasobserved.
Inhibition of DHD/SHD leads to reduced chlorogenateand lignin content
The shikimate pathway converts erythrose 4-phosphateand PEP into aromatic amino acids that could be
Table 1. Growth parameters of tobacco plants with decreasedDHD/SHD activity
Tobacco seeds were germinated in soil under greenhouse conditions andtransferred to small pots containing sand. Plants were fertilized dailywith 100 ml of nutrient solution (Geiger et al., 1999) and allowed togrow for 6 weeks. Results are mean values of six wild-type plants or T1
progeny of each independent transgenic line 6SE.
Genotype Group Plantheight (cm)
ShootFW (g)
ShootDW (g)
FW/DW
Wild-type – 13.562.6 7.1361.24 0.4160.18 17.39DHD/SHD-1 H 12.962.3 6.3261.46 0.3460.10 18.59DHD/SHD-63 M 8.662.1 4.8360.67 0.2460.07 20.13DHD/SHD-63 L 7.261.7 1.7960.19 0.1060.05 17.9DHD/SHD-9 VL 4.961.2 0.5060.083 0.0360.01 16.67
2058 Ding et al.
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
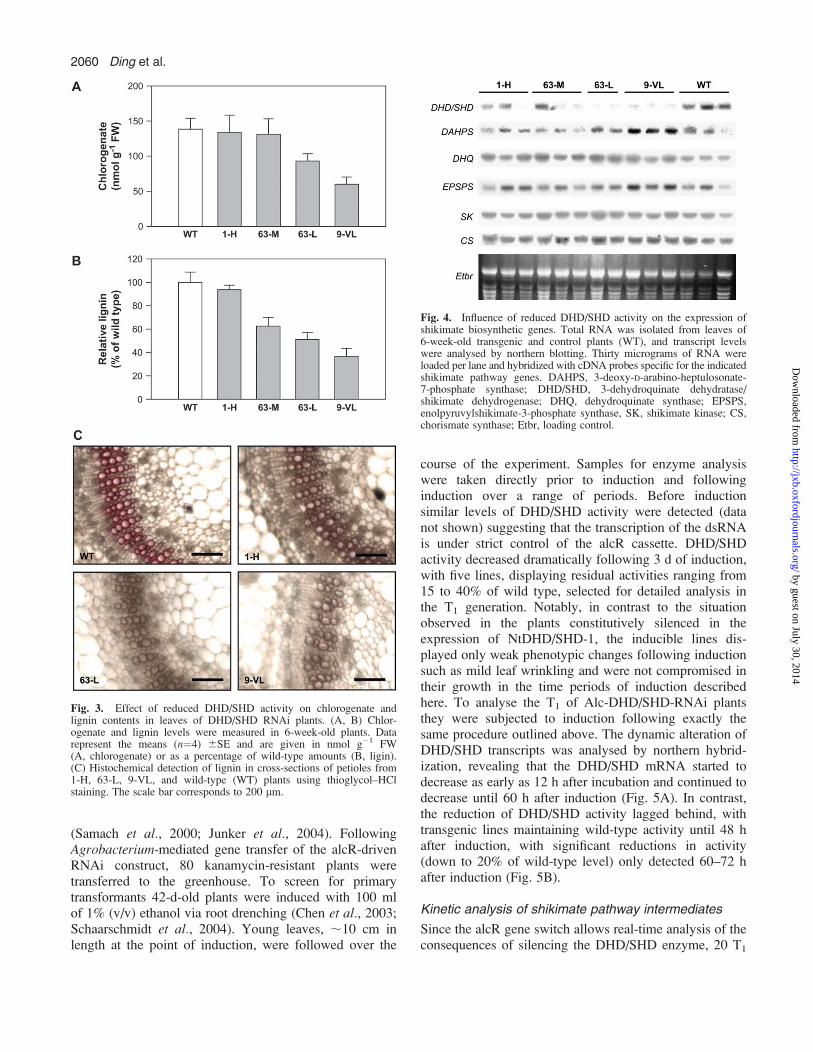
metabolized further into a large number of differentphenolic compounds. To determine the effect of DHD/SHD silencing on secondary metabolism, chlorogenateand lignin were determined in transgenic plants. In plantswith >60% DHD/SHD activity (DHD/SHD 1-H), a slightdecrease of chlorogenate and lignin was measured withvalues of 96% and 93% of wild-type plants, respectively.A 40–60% reduction of DHD/SHD activity led to a re-markable reduction in aromatic compounds, in whichlignin and chlorogenate dropped to 63% and 67% of wild-type values, respectively. In plants with <20% DHD/SHDresidual activity, only 43% of chlorogenate and 37% oflignin accumulated (Fig. 3).To visualize lignin histochemically in transgenic plants,
leaf material was stained by the phloroglucinol–HClmethod. As demonstrated in Fig. 3C, vascular tissues inpetioles of wild-type plants were stained brilliant red,indicating high concentrations of phenolic compounds andlignin, whereas in DHD/SHD-silenced plants, vasculartissue in petioles was only slightly stained pale red,suggesting that less phenolic compounds and ligninaccumulated in the transgenic plants (Fig. 3C).
DHD/SHD silencing does not lead to down-regulationof shikimate pathway enzymes at the transcript level
To investigate whether silencing of DHD/SHD wouldinfluence the expression of other shikimate pathwaygenes, the transcripts of DAHPS, dehydroquinate syn-thase, shikimate kinase, EPSP synthase, and chorismatesynthase were probed by northern blot hybridization inboth transgenic and control plants. As demonstrated inFig. 4, silencing of DHD/SHD did not affect expression ofdownstream genes and the adjacent upstream gene,dehydroquinate synthase. DAHPS was the only gene tobe regulated at the transcript level. Accumulation ofDAHPS transcripts increased up to four times in DHD/SHD-VL plants as compared with wild-type controls.
Shikimate feeding of leaf discs reveals normalmetabolism in DHD/SHD-RNAi plants
As described above, shikimate accumulated in DHD/SHS-silenced plants. This unexpected observation might beexplained by an impaired downstream processing ofshikimate. To test this hypothesis, wild-type and trans-genic leaf discs were floated on 50 mM HEPES solution(pH 7.0) containing 20 mM shikimate at room temperaturefor 16 h. As a control, leaf discs were floated on 50 mMHEPES solution without shikimate under the sameconditions. Afterwards, leaf discs were washed withdistilled water, dried on filter paper, and subjected toamino acid determination. In the absence of shikimate orprior to the experiment, the content of phenylalanine andtyrosine was lower in transgenic plants (substantially so inthe case of phenylalanine but only slightly so in theinstance of tyrosine) as compared with wild-type controls.Following feeding on shikimate these differences disap-peared. Similar amounts of phenylalanine (79–118% ofwild-type plants) and tyrosine (84–122% of wild-typeplants) accumulated in leaf discs of transgenic plants afterfeeding 20 mM shikimate (Supplementary Fig. S1 avail-able at JXB online). This result indicates that silencingDHD/SHD did not impair downstream enzymes of theshikimate pathway in transgenic plants.
Inducible RNA interference of NtDHD/SHD-1
Given that the surprising result of elevated shikimatelevels in the transformants constitutively silenced inNtDHD/SHD-1 was observed in plants also displayinga strong growth phenotype, it was decided to create plantswhich could be silenced by chemical induction. Thisapproach was carried out to allow evaluation of thisphenomenon in plants exhibiting developmental equiva-lence to the wild type and thus to evaluate whether it isa direct consequence of the genetic manipulation ormerely a pleiotropic one. For this purpose, the alcRsystem, which had previously been used in the dissectionof complex phenotypes, was chosen for the present study
Sh
ikim
ate
(µ
mo
l g
-1 F
W)
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
Deh
yd
ro
qu
in
ate
(µ
mo
l g
-1 F
W)
0
2
4
6
8
10
12A
B
1-HWT 63-M 63-L 9-VL
1-HWT 63-M 63-L 9-VL
Fig. 2. Dehydroquinate and shikimate accumulation in leaves of DHD/SHD-RNAi plants. Shikimate (A) and dehydroquinate (B) contentswere determined in TCA extracts using an enzymatic assay withpurified SHD/DHD enzyme (see Materials and methods). Samples weretaken from source leaves of 6-week-old plants grown under greenhouseconditions. Values are given as means of four independent plants 6SE.
Molecular analysis of shikimate synthesis in tobacco 2059
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
(Samach et al., 2000; Junker et al., 2004). FollowingAgrobacterium-mediated gene transfer of the alcR-drivenRNAi construct, 80 kanamycin-resistant plants weretransferred to the greenhouse. To screen for primarytransformants 42-d-old plants were induced with 100 mlof 1% (v/v) ethanol via root drenching (Chen et al., 2003;Schaarschmidt et al., 2004). Young leaves, ;10 cm inlength at the point of induction, were followed over the
course of the experiment. Samples for enzyme analysiswere taken directly prior to induction and followinginduction over a range of periods. Before inductionsimilar levels of DHD/SHD activity were detected (datanot shown) suggesting that the transcription of the dsRNAis under strict control of the alcR cassette. DHD/SHDactivity decreased dramatically following 3 d of induction,with five lines, displaying residual activities ranging from15 to 40% of wild type, selected for detailed analysis inthe T1 generation. Notably, in contrast to the situationobserved in the plants constitutively silenced in theexpression of NtDHD/SHD-1, the inducible lines dis-played only weak phenotypic changes following inductionsuch as mild leaf wrinkling and were not compromised intheir growth in the time periods of induction describedhere. To analyse the T1 of Alc-DHD/SHD-RNAi plantsthey were subjected to induction following exactly thesame procedure outlined above. The dynamic alteration ofDHD/SHD transcripts was analysed by northern hybrid-ization, revealing that the DHD/SHD mRNA started todecrease as early as 12 h after incubation and continued todecrease until 60 h after induction (Fig. 5A). In contrast,the reduction of DHD/SHD activity lagged behind, withtransgenic lines maintaining wild-type activity until 48 hafter induction, with significant reductions in activity(down to 20% of wild-type level) only detected 60–72 hafter induction (Fig. 5B).
Kinetic analysis of shikimate pathway intermediates
Since the alcR gene switch allows real-time analysis of theconsequences of silencing the DHD/SHD enzyme, 20 T1
Fig. 3. Effect of reduced DHD/SHD activity on chlorogenate andlignin contents in leaves of DHD/SHD RNAi plants. (A, B) Chlor-ogenate and lignin levels were measured in 6-week-old plants. Datarepresent the means (n¼4) 6SE and are given in nmol g�1 FW(A, chlorogenate) or as a percentage of wild-type amounts (B, ligin).(C) Histochemical detection of lignin in cross-sections of petioles from1-H, 63-L, 9-VL, and wild-type (WT) plants using thioglycol–HClstaining. The scale bar corresponds to 200 lm.
Fig. 4. Influence of reduced DHD/SHD activity on the expression ofshikimate biosynthetic genes. Total RNA was isolated from leaves of6-week-old transgenic and control plants (WT), and transcript levelswere analysed by northern blotting. Thirty micrograms of RNA wereloaded per lane and hybridized with cDNA probes specific for the indicatedshikimate pathway genes. DAHPS, 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase; DHD/SHD, 3-dehydroquinate dehydratase/shikimate dehydrogenase; DHQ, dehydroquinate synthase; EPSPS,enolpyruvylshikimate-3-phosphate synthase, SK, shikimate kinase; CS,chorismate synthase; Etbr, loading control.
2060 Ding et al.
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
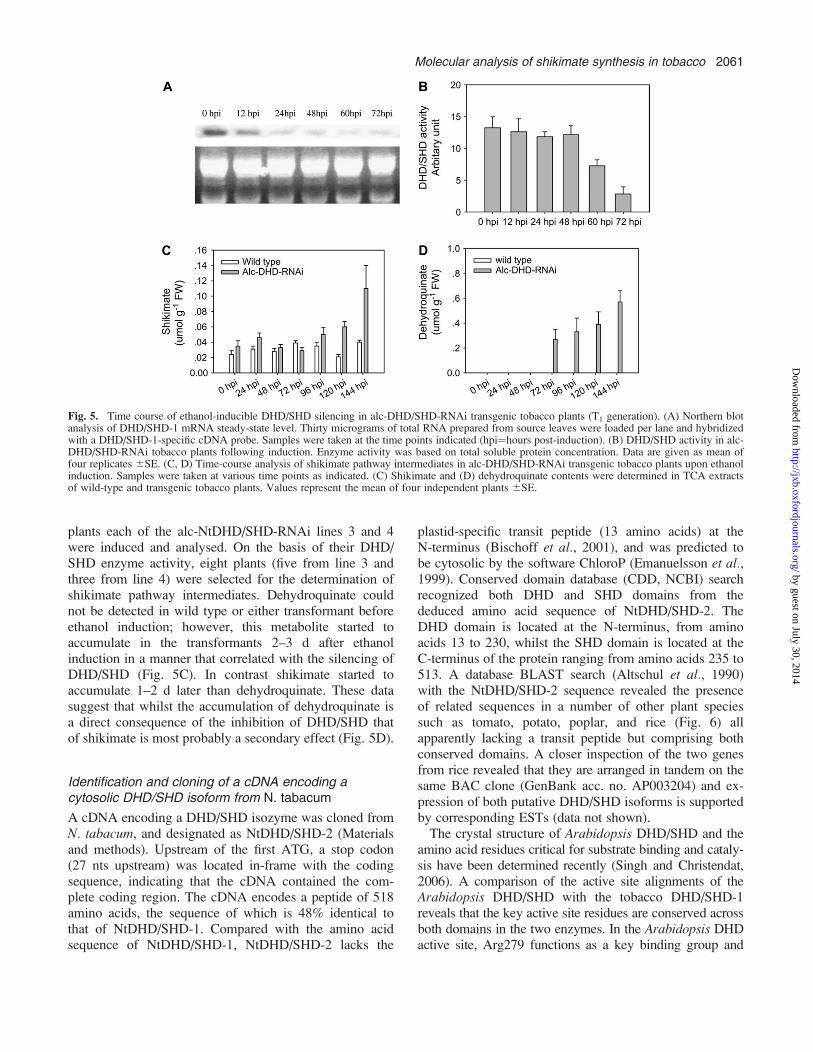
plants each of the alc-NtDHD/SHD-RNAi lines 3 and 4were induced and analysed. On the basis of their DHD/SHD enzyme activity, eight plants (five from line 3 andthree from line 4) were selected for the determination ofshikimate pathway intermediates. Dehydroquinate couldnot be detected in wild type or either transformant beforeethanol induction; however, this metabolite started toaccumulate in the transformants 2–3 d after ethanolinduction in a manner that correlated with the silencing ofDHD/SHD (Fig. 5C). In contrast shikimate started toaccumulate 1–2 d later than dehydroquinate. These datasuggest that whilst the accumulation of dehydroquinate isa direct consequence of the inhibition of DHD/SHD thatof shikimate is most probably a secondary effect (Fig. 5D).
Identification and cloning of a cDNA encoding acytosolic DHD/SHD isoform from N. tabacum
A cDNA encoding a DHD/SHD isozyme was cloned fromN. tabacum, and designated as NtDHD/SHD-2 (Materialsand methods). Upstream of the first ATG, a stop codon(27 nts upstream) was located in-frame with the codingsequence, indicating that the cDNA contained the com-plete coding region. The cDNA encodes a peptide of 518amino acids, the sequence of which is 48% identical tothat of NtDHD/SHD-1. Compared with the amino acidsequence of NtDHD/SHD-1, NtDHD/SHD-2 lacks the
plastid-specific transit peptide (13 amino acids) at theN-terminus (Bischoff et al., 2001), and was predicted tobe cytosolic by the software ChloroP (Emanuelsson et al.,1999). Conserved domain database (CDD, NCBI) searchrecognized both DHD and SHD domains from thededuced amino acid sequence of NtDHD/SHD-2. TheDHD domain is located at the N-terminus, from aminoacids 13 to 230, whilst the SHD domain is located at theC-terminus of the protein ranging from amino acids 235 to513. A database BLAST search (Altschul et al., 1990)with the NtDHD/SHD-2 sequence revealed the presenceof related sequences in a number of other plant speciessuch as tomato, potato, poplar, and rice (Fig. 6) allapparently lacking a transit peptide but comprising bothconserved domains. A closer inspection of the two genesfrom rice revealed that they are arranged in tandem on thesame BAC clone (GenBank acc. no. AP003204) and ex-pression of both putative DHD/SHD isoforms is supportedby corresponding ESTs (data not shown).The crystal structure of Arabidopsis DHD/SHD and the
amino acid residues critical for substrate binding and cataly-sis have been determined recently (Singh and Christendat,2006). A comparison of the active site alignments of theArabidopsis DHD/SHD with the tobacco DHD/SHD-1reveals that the key active site residues are conserved acrossboth domains in the two enzymes. In the Arabidopsis DHDactive site, Arg279 functions as a key binding group and
Fig. 5. Time course of ethanol-inducible DHD/SHD silencing in alc-DHD/SHD-RNAi transgenic tobacco plants (T1 generation). (A) Northern blotanalysis of DHD/SHD-1 mRNA steady-state level. Thirty micrograms of total RNA prepared from source leaves were loaded per lane and hybridizedwith a DHD/SHD-1-specific cDNA probe. Samples were taken at the time points indicated (hpi¼hours post-induction). (B) DHD/SHD activity in alc-DHD/SHD-RNAi tobacco plants following induction. Enzyme activity was based on total soluble protein concentration. Data are given as mean offour replicates 6SE. (C, D) Time-course analysis of shikimate pathway intermediates in alc-DHD/SHD-RNAi transgenic tobacco plants upon ethanolinduction. Samples were taken at various time points as indicated. (C) Shikimate and (D) dehydroquinate contents were determined in TCA extractsof wild-type and transgenic tobacco plants. Values represent the mean of four independent plants 6SE.
Molecular analysis of shikimate synthesis in tobacco 2061
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
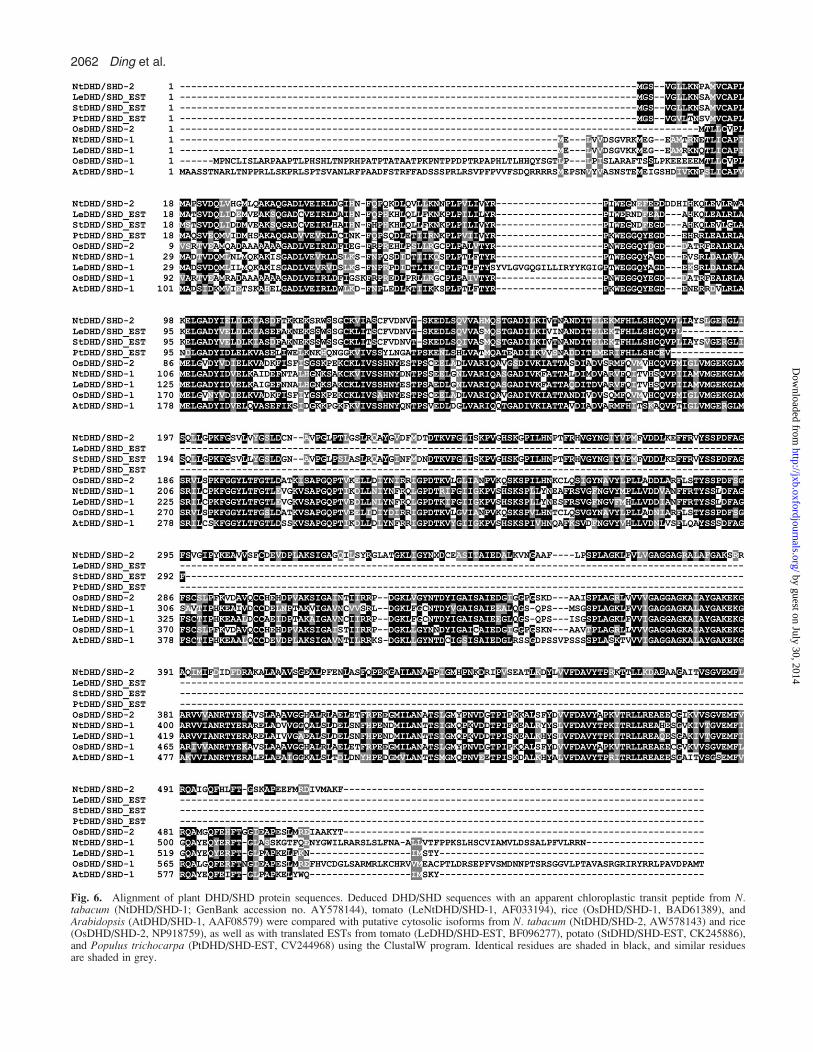
Fig. 6. Alignment of plant DHD/SHD protein sequences. Deduced DHD/SHD sequences with an apparent chloroplastic transit peptide from N.tabacum (NtDHD/SHD-1; GenBank accession no. AY578144), tomato (LeNtDHD/SHD-1, AF033194), rice (OsDHD/SHD-1, BAD61389), andArabidopsis (AtDHD/SHD-1, AAF08579) were compared with putative cytosolic isoforms from N. tabacum (NtDHD/SHD-2, AW578143) and rice(OsDHD/SHD-2, NP918759), as well as with translated ESTs from tomato (LeDHD/SHD-EST, BF096277), potato (StDHD/SHD-EST, CK245886),and Populus trichocarpa (PtDHD/SHD-EST, CV244968) using the ClustalW program. Identical residues are shaded in black, and similar residuesare shaded in grey.
2062 Ding et al.
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
Lys241 and His214 are catalytic groups (Singh andChristendat, 2006). These residues correspond to Arg207,Lys169, and His142, respectively, in NtDHD/SHD1 (Fig.6). In the NtSHD/DHD-2 polypeptide the Lys residue(Lys160), which is involved in the formation of a Schiffbase intermediate (Singh and Christendat, 2006), is alsoconserved; however, the His residue is substituted by Phe(Phe134) and the Arg is replaced by Gln (Gln198). Incontrast, Lys385 and Asp423 (numbered with respect tothe Arabidopsis protein), located in the SDH active siteare conserved in all three DHD/SHD enzymes and havebeen proposed to be involved in proton transfer duringcatalysis (Singh and Christendat, 2006). The crystalstructure of the Arabidopsis SDH domain indicated thatSer336, Ser338, and Tyr550 are important for bindingshikimate (Singh and Christendat, 2006). All threeresidues are conserved in NtDHD/SHD-1 (Ser264,Ser266, and Tyr473, respectively). In NtDHD/SHD-2only the Ser336 (Ser254 in NtSHD/DHD-2) and theTyr550 (Tyr467) are conserved, with the Ser338 sub-stituted by Gly (Gly256).
Enzymatic activity of NtDHD/SHD-2 and in vivolocalization of NtDHD/SHD-1 and 2
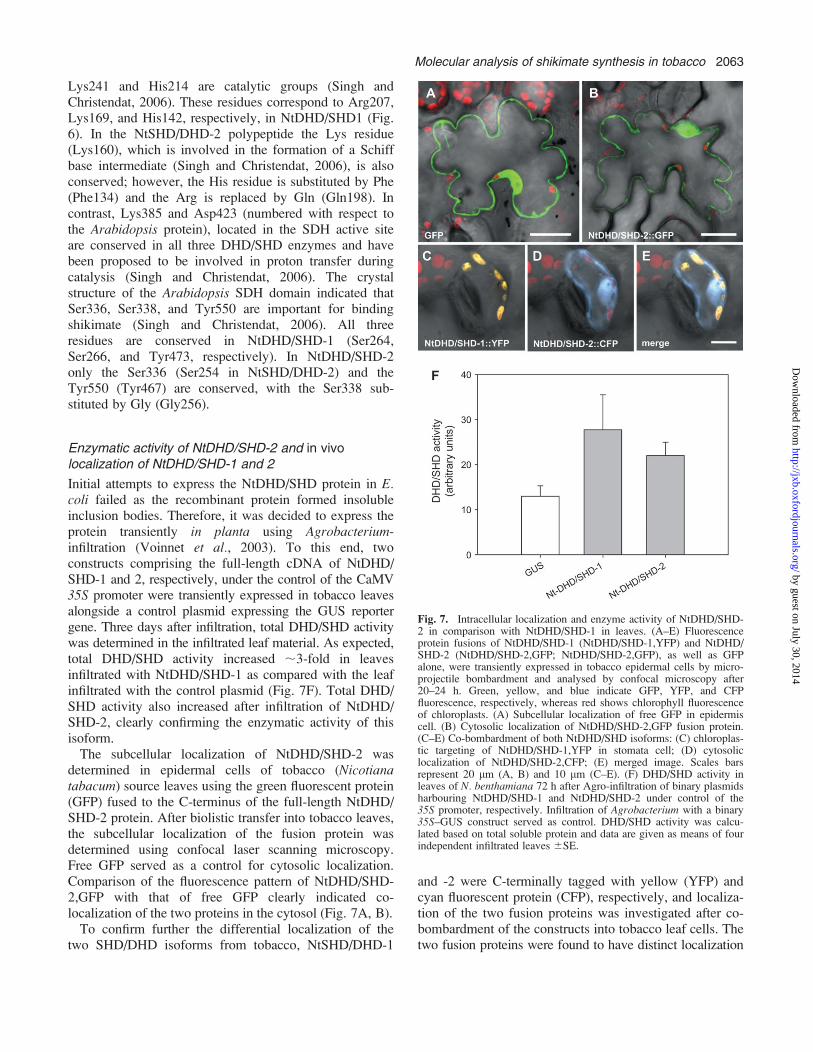
Initial attempts to express the NtDHD/SHD protein in E.coli failed as the recombinant protein formed insolubleinclusion bodies. Therefore, it was decided to express theprotein transiently in planta using Agrobacterium-infiltration (Voinnet et al., 2003). To this end, twoconstructs comprising the full-length cDNA of NtDHD/SHD-1 and 2, respectively, under the control of the CaMV35S promoter were transiently expressed in tobacco leavesalongside a control plasmid expressing the GUS reportergene. Three days after infiltration, total DHD/SHD activitywas determined in the infiltrated leaf material. As expected,total DHD/SHD activity increased ;3-fold in leavesinfiltrated with NtDHD/SHD-1 as compared with the leafinfiltrated with the control plasmid (Fig. 7F). Total DHD/SHD activity also increased after infiltration of NtDHD/SHD-2, clearly confirming the enzymatic activity of thisisoform.The subcellular localization of NtDHD/SHD-2 was
determined in epidermal cells of tobacco (Nicotianatabacum) source leaves using the green fluorescent protein(GFP) fused to the C-terminus of the full-length NtDHD/SHD-2 protein. After biolistic transfer into tobacco leaves,the subcellular localization of the fusion protein wasdetermined using confocal laser scanning microscopy.Free GFP served as a control for cytosolic localization.Comparison of the fluorescence pattern of NtDHD/SHD-2,GFP with that of free GFP clearly indicated co-localization of the two proteins in the cytosol (Fig. 7A, B).To confirm further the differential localization of the
two SHD/DHD isoforms from tobacco, NtSHD/DHD-1
and -2 were C-terminally tagged with yellow (YFP) andcyan fluorescent protein (CFP), respectively, and localiza-tion of the two fusion proteins was investigated after co-bombardment of the constructs into tobacco leaf cells. Thetwo fusion proteins were found to have distinct localization
Fig. 7. Intracellular localization and enzyme activity of NtDHD/SHD-2 in comparison with NtDHD/SHD-1 in leaves. (A–E) Fluorescenceprotein fusions of NtDHD/SHD-1 (NtDHD/SHD-1,YFP) and NtDHD/SHD-2 (NtDHD/SHD-2,GFP; NtDHD/SHD-2,GFP), as well as GFPalone, were transiently expressed in tobacco epidermal cells by micro-projectile bombardment and analysed by confocal microscopy after20–24 h. Green, yellow, and blue indicate GFP, YFP, and CFPfluorescence, respectively, whereas red shows chlorophyll fluorescenceof chloroplasts. (A) Subcellular localization of free GFP in epidermiscell. (B) Cytosolic localization of NtDHD/SHD-2,GFP fusion protein.(C–E) Co-bombardment of both NtDHD/SHD isoforms: (C) chloroplas-tic targeting of NtDHD/SHD-1,YFP in stomata cell; (D) cytosoliclocalization of NtDHD/SHD-2,CFP; (E) merged image. Scales barsrepresent 20 lm (A, B) and 10 lm (C–E). (F) DHD/SHD activity inleaves of N. benthamiana 72 h after Agro-infiltration of binary plasmidsharbouring NtDHD/SHD-1 and NtDHD/SHD-2 under control of the35S promoter, respectively. Infiltration of Agrobacterium with a binary35S–GUS construct served as control. DHD/SHD activity was calcu-lated based on total soluble protein and data are given as means of fourindependent infiltrated leaves 6SE.
Molecular analysis of shikimate synthesis in tobacco 2063
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

patterns (Fig. 7C, E). As observed before for the GFPfusion protein, the NtDHD/SHD-2,CFP signal clearlylocalized to the cytosol, whereas the NtDHD/SHD-1,YFPfluorescence was confined to the plastids. When takentogether these data clearly indicate that NtDHD/SHD-2encodes a cytosolic isoform of tobacco DHD/SHD.
Discussion
It has been demonstrated previously that compromisingplastidial shikimate biosynthesis, by the antisense inhibitionof DAHP synthase, leads to potato plants exhibiting astunted growth phenotype characterized by reduced stemlength, width, and lignin content (Jones et al., 1995). Earlystudies also documented the importance of this pathway inthe synthesis of plant secondary compounds includingindole acetic acid, pigments, such as anthrocyanins, andantimicrobial compounds such as phytoalexins and liginwhich have been demonstrated to play vital roles in plantdefence (Vance et al., 1980; Moerschbacher et al., 1990;Nicholson and Hammerschmidt, 1992), wound healing(Bostock and Stermer, 1989), and maintenance of struc-tural integrity and water transport capacity (Lewis andYakamoto, 1990). More recently, the mutant cue1 wasidentified in a genetic screen for chlorophyll a/b-bindingprotein underexpressors (Li et al., 1995); the mutationmapped to the PEP/phosphate translocator of the plastidinner envelope membrane which has been proposed toimport PEP into the stroma, where it serves as one of thesubstrates of the shikimate pathway (Fischer et al., 1997).Phenotypically cue1 is quite distinctive; it exhibits a re-ticulate leaf phenotype with dark green paraveinal andlight green interveinal regions as well as exhibitingdeficiencies in chloroplast and mesophyll development(Li et al., 1995; Streatfield et al., 1999). However,a detailed genetic and biochemical analysis of thisphenotype revealed that it was not simply caused bya general restriction of the shikimate pathway (Voll et al.,2003). In the present study it is demonstrated that RNAi-mediated reduction of the expression of NtDHD/SHD-1 intobacco also leads to severe phenotypic and metabolicalterations. A 50% reduction of DHD/SHD activity (DHD/SHD-M plants) resulted in strongly bleached leaves, anda further reduction of DHD/SHD activity to 40% of thewild-type value led to strong decreases in growth param-eters, highlighting the importance of DHD/SHD for plantdevelopment. Further reduction of endogenous DHD/SHDactivity (DHD/SHD-VL plants) led to a lethal phenotype,in which case transgenic plants were stunted, and died2–3 weeks after being transferred to the greenhouse. Thesephenotypes are largely consistent with what would beexpected following a severe restriction in shikimate bio-synthesis. Moreover, the approximately linear correlationbetween DHD/SHD residual activity and the level of
phenolic compounds that the transgenic lines containhighlights the crucial role of this plastid-localized enzymein controlling the flux of carbon towards secondarymetabolism.Given the marked reduction in the expression of the
bifunctional DHD/SHD and the clear growth phenotypeand lignin deficiency exhibited by the transgenic lines itwas somewhat surprising that these plants displayedincreased, as opposed to decreased, levels of dehydroqui-nate and shikimate. Expression analysis of genes encodingall seven enzymes of the shikimate pathway in the DHD/SHD RNAi plants revealed that DAHPS was the only onefound to be regulated at the transcriptional level. Activa-tion of DAHPS at the transcriptional level has beendescribed previously following abiotic stresses such asmechanical wounding (Dyer et al., 1989) or glyphosatetreatment (Pinto et al., 1988), indicating its potentialimportance in controlling carbon flow through the shiki-mate pathway. However, the fact that the up-regulation ofDAHPS was not accompanied by a co-ordinate change inother shikimate pathway enzymes suggests that this isunlikely to be the major mechanism underlying theincreased dehydroquinate and shikimate levels of thetransformants. Another possible reason for the accumula-tion of dehydroquinate and shikimate is a restriction in thein vivo activity of downstream enzymes of the shikimatepathway. In keeping with this hypothesis is the fact thatthe transformants generated in this study displayed re-duced levels of lignin and phenylpropanoids. However,leaves of transgenic and wild-type plants were able tosynthesize a similar level of phenylalanine and tyrosinefollowing incubation in shikimate, suggesting that re-duction in lignin and phenylpropanoids was merely due toreduced substrate supply to downstream enzymes ofchorismate synthesis rather than an active inhibition ofthe enzyme activities per se.Silencing of a pathway enzyme typically disturbs the
conversion of substrate to product, leading to an accumu-lation of the substrate and a depletion of the product(Rolleston, 1972). In Nt-DHD/SHD-RNAi plants, how-ever, the reduction of the DHD/SHD enzyme activitiesresulted in a build-up of both dehydroquinate andshikimate, which are substrate and product of this enzyme,respectively. The accumulation of dehydroquinate andshikimate in transgenic plants was, furthermore, stronglynegatively correlated with DHD/SHD residual activity.The accumulation of a shikimate observed in transgenicplants is unexpected for two reasons. On one hand, as theproduct of DHD/SHD, the silencing of DHD/SHD intransgenic plants would be anticipated to result in a re-duced biosynthesis of shikimate in planta. On the otherhand, the silencing of DHD/SHD did not impair themetabolism of shikimate by the downstream enzymes.Alternatively, shikimate could be synthesized from dehy-droquinate via quinate, thus bypassing the DHD/SHD
2064 Ding et al.
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

reaction. In this scenario, quinate is synthesized fromdehydroquinate by an NADP-dependent quinate oxidore-ductase and subsequently converted to shikimate bya quinate hydrolase (Leuschner et al., 1995). However,given the proposed plastidial localization of the quinatehydrolase, this sequence of reactions is unlikely to beresponsible for the shikimate accumulation as one wouldexpect that any shikimate produced in plastids would beavailable to support aromatic amino acid and ligninsynthesis.When the considerations above are taken together it is
clear that shikimate content should decrease in DHD/SHD-silenced plants. To interpret the accumulation ofshikimate in DHD/SHD-silenced plants a metabolite shut-tle model is proposed. According to this model, thesilencing of NtDHD/SHD-1 leads to a high-level accumu-lation of dehydroquinate in chloroplasts. A proportion ofthis metabolite, which notably accumulates to very highlevels in the transgenics, could be transported fromchloroplasts to the cytosol and be converted to shikimateby a postulated cytosolic DHD/SHD isozyme. The extra-plastidial shikimate produced by this enzyme is, however,most probably neither efficiently re-imported into chlor-oplasts, nor metabolized by downstream shikimate path-way enzymes in the cytosol, resulting in an elevatedcellular shikimate level in the transformants. The cloningof a cytosolic DHD/SHD-like enzyme (NtDHD/SHD-2)gives strong support to this model, although the actualexport of dehydroquinate from the plastid into the cytosolhas thus far not been experimentally demonstrated.However, the enzymatic activity of NtDHD/SHD-2 hasbeen confirmed by Agrobacterium-infiltration, and sub-cellular localization using GFP-fusion proteins clearlyrevealed that this enzyme is a cytosolic protein.Active site alignment between the Arabidopsis DHD/
SHD and NtDHD/SHD-2 revealed that many of the aminoacid residues critical for substrate binding and catalysisare conserved between the two proteins, suggesting thatthese enzymes support similar catalytic functions. How-ever, there appears to be deviation in amino acid type atsome other key positions. The role of the active siteresidues for binding and catalysis in the Arabidopsis SHDdomain has been assessed by site-directed mutagenesis(Singh and Christendat, 2006). All critical residues ex-cept Ser338 (changed to Gly256 in NtDHD/SHD-2)are conserved between the Arabidopsis DHD/SHD andNtDHD/SHD-2. Mutagenesis of Ser338 to Ala in theArabidopsis enzyme resulted in a 10-fold reduction in theturnover rate, and this residue has been implicated to beinvolved in binding and also in positioning of thesubstrate for catalysis (Singh and Christendat, 2006).Given the replacement of this critical residue by Gly inNtSHD/DHD-2 it appears likely that the cytosolic enzymeretains its ability to synthesize shikimate, albeit witha lower turnover rate. Interestingly, NtSHD/DHD-2 con-
tains an additional Ser (Ser247) which is supposed to bepositioned deeper in the shikimate binding pocket. Takentogether, the conservation of some of the critical residuesfor catalysis is in close agreement with the DHD/SHDactivity of NtDHD/SHD-2 which it is possible todemonstrate. However, on the basis of comparisonto SDH-L forms from other organisms, it is tempting tospeculate that the preferred substrate of the enzyme in vivois in fact a derivative of shikimate exhibiting a largerfunctional group at the C1 position ring (Singh andChristendat, 2006). Thus, NtDHD/SHD-2 might involvedifferent, albeit chemically related, substrates thanNtDHD/SHD-1 and thus lead to a different product thanthe reaction catalysed by plastidial enzyme. Hence, itappears unlikely that the biological function of NtDHD/SHD-2 in vivo is the synthesis of shikimate in the cytosol.Circumstantial evidence in support of such a hypothesis isprovided by the fact that overexpression of DHD/SHD-2did not result in as great an increase in activity as thatobserved on overexpression of DHD/SHD-1. Furthermore,several examples in the literature indicate that genes ofplant secondary metabolism have been recruited fromancestors of primary metabolism (Pichersky and Gang,2000; Ober, 2005). New genes almost always arise bygene duplication followed by divergence. If the originalgene had an essential function, as is the case for DHD/SHD, gene duplication is a prerequisite (Pichersky andGang, 2000). The genomic organization of NtDHD/SHD-1 and 2 is unknown; however, a closer inspection of thetwo rice orthologues revealed their tandem location withinthe genome, strongly implicating that they arose through agene duplication event. It is a common feature of enzymeevolution that the substrate of a new enzyme often closelyresembles that of the enzyme from which it is derived.Thus relatively few amino acid substitutions can allow analtered enzyme to recognize a novel substrate whilstretaining the ability to utilize the original substrate albeitat lower efficiency (Pichersky and Gang, 2000). Futureexperiments will focus on the biochemical characteriza-tion and on a structure–function analysis of DHD/SHD-2to elucidate its biological function further. Furthermore,non-aqueous fractionation will have to elucidate thesubcellular distribution of shikimate in DHD/SHD-silenced plants.In conclusion, in the current experiments, it was
possible to provide direct evidence for the essential roleof a plastidial isoform of DHD/SHD in the synthesis ofphenylalanine and its secondary metabolite derivativesincluding chlorogenic acid and lignin. These plants werealso characterized by dramatic growth deficiencies and anelevated content of shikimate. After cloning of anadditional gene for a DHD/SHD-like enzyme and con-firming a cytosolic localization for its product, it is proposedthat this enzyme is responsible for the shikimate accumu-lation observed in DHD/SHD-silenced tobacco plants.
Molecular analysis of shikimate synthesis in tobacco 2065
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Intriguingly, despite this capability, this pathway is notable to complement that localized in the plastid, mostprobably due to a spatial restriction in the availability ofthe shikimate to the chloroplast. The fact that theshikimate produced by DHD/SHD does not enter thearomatic amino acid or lignin biosynthesis pathways ishighly interesting and suggests that the cytosolic isoformmay, in vivo, catalyse an alternative reaction.As such, the future elucidation of the functional role of
cytosolic DHD/SHD is likely to provide important insightinto the evolution of both shikimate and intermediarymetabolism in plants.
Supplementary material
Supplementary material can be found at JXB online.Supplementary figure 1. Tyrosine (Tyr) and phenylal-
anine (Phe) content in leaf discs of DHD/SHD-RNAiplants prior to and following exogenous application ofshikimate in comparison to the wild type controls.
References
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990.Basic local alignment search tool. Journal Molecular Biology215, 403–410.
Bentley R. 1990. The shikimate pathway: metabolic tree with manybranches. Critical Reviews in Biochemistry and Molecular Bi-ology 25, 307–384.
Biemelt S, Tschiersch H, Sonnewald U. 2004. Impact of alteredgibberellin metabolism on biomass accumulation, lignin bio-synthesis, and photosynthesis in transgenic tobacco plants. PlantPhysiology 135, 254–265.
Bischoff M, Schaller A, Bieri F, Kessler F, Amrhein N,Schmid J. 2001. Molecular characterization of tomato 3-dehydroquinate dehydratase-shikimate:NADP oxidoreductase.Plant Physiology 125, 1891–1900.
Bonner CA, Jensen RA. 1994. Cloning of cDNA encoding thebifunctional dehydroquinase shikimate dehydrogenase of aromatic-amino-acid biosynthesis in Nicotiana tabacum. BiochemicalJournal 302, 11–14.
Bostock RM, Stermer BA. 1989. Perspectives on wound healingin resistance to pathogens. Annual Review of Phytophathology 27,343–371.
Bradford MM. 1976. A rapid and sensitive method for quantifica-tion of microgram quantities of proteins utilizing the principle ofprotein-dye coupling. Analytical Biochemistry 72, 248–254.
Buchholz B, Reupke B, Bickel H, Schultz G. 1979. Reconstruc-tion of amino acid biosynthesis by combining spinach chloro-plasts with other leaf organelles. Phytochemistry 18, 1109–1111.
Campbell MM, Ellis BE. 1992. Fungal elicitor-mediated responsesin pine cell cultures. III. Purification and characterization ofphenylalanine ammonia-lyase. Plant Physiology 98, 62–70.
Chen S, Hofius D, Sonnewald U, Bornke F. 2003. Temporal andspatial control of gene silencing in transgenic plants by inducibleexpression of double-stranded RNA. The Plant Journal 36, 731–740.
d’Amato TA. 1984. Subcellular localization of chorismate-mutaseisoenzymes in protoplasts from mesophyll and suspension-cultured cells of Nicotiana sylvestris. Planta 162, 104–108.
Deka RK, Anton IA, Dunbar B, Coggins JR. 1994. Thecharacterisation of the shikimate pathway enzyme dehydroqui-nase from Pisum sativum. FEBS Letters 349, 397–402.
Della-Cioppa G, Bauer SC, Klein BK, Shah DM, Fraley RT,Kishore GM. 1986. Translocation of the precursor of 5-enolpyruvylshikimate-3-phosphate synthase into chloroplasts ofhigher plants in vitro. Proceedings of the National Academy ofSciences, USA 83, 6873–6877.
Dyer WE, Henstrand JM, Handa AK, Herrmann KM. 1989.Wounding induces the first enzyme of the shikimate pathway inSolanaceae. Proceedings of the National Academy of Sciences,USA 86, 7370–7373.
Emanuelsson O, Nielsen H, von Heijne G. 1999. ChloroP,a neural network-based method for predicting chloroplast transitpeptides and their cleavage sites. Protein Science 8, 978–984.
Fiedler E, Schultz G. 1985. Localization, purification andcharacterization of shikimate oxidoreductase-dehydroquinate hy-drolase from stroma of spinach chloroplasts. Plant Physiology 79,212–218.
Fischer K, Kammerer B, Gutensohn M, Arbinger B, Weber A,Hausler RE, Flugge UI. 1997. A new class of plastidicphosphate translocators: a putative link between primary andsecondary metabolism by the phosphoenolpyruvate/phosphateantiporter. The Plant Cell 9, 453–462.
Ganson RJ, D’Amato TA, Jensen RA. 1986. The two-isozymesystem of 3-deoxy-D-arabino-heptulosonate 7-phosphate synthasein Nicotiana silvestris and other higher plants. Plant Physiology82, 203–210.
Gavel Y, von Heijne G. 1990. A conserved cleavage-site motif inchloroplast transit peptides. FEBS Letters 261, 455–458.
Geiger M, Haake V, Ludewig F, Sonnewald U, Stitt M. 1999.The nitrate and ammonium nitrate supply have a major influenceon the response of photosynthesis, carbon metabolism, nitrogenmetabolism and growth to elevated carbon dioxide in tobacco.Plant, Cell and Environment 22, 1177–1199.
Hajirezaei MR, Peisker M, Tschiersch H, Palatnik JF,Valle EM, Carrillo N, Sonnewald U. 2002. Small changes inthe activity of chloroplastic NADP+-dependent ferredoxin oxido-reductase lead to impaired plant growth and restrict photosyn-thetic activity of transgenic tobacco plants. The Plant Journal 29,281–293.
Haslam E. 1993. Shikimic acid, metabolism and metabolites.Chichester: John Wiley and Sons..
Herrmann KM. 1995. The shikimate pathway: early steps inthe biosynthesis of aromatic compounds. The Plant Cell 7,907–919.
Herrmann KM, Weaver LM. 1999. The shikimate pathway.Annual Review of Plant Physiology and Plant Molecular Biology50, 473–503.
Hofgen R, Willmitzer L. 1990. Biochemical and genetic analysisof different patatin isoforms expressed in various organs of potato(Solanum tuberosum). Plant Science 66, 221–230.
Jelitto T, Sonnewald U, Willmitzer L, Hajirezaei MR, Stitt M.1992. Inorganic pyrophosphate content and metabolites in leavesand tubers of potato and tobacco plants expressing E. colipyrophosphatase in their cytosol: biochemical evidence thatsucrose metabolism has been manipulated. Planta 188, 238–244.
Jones JD, Henstrand JM, Handa AK, Herrmann KM,Weller SC. 1995. Impaired wound induction of 3-deoxy-D-arabino-heptulosonate-7-phosphate (DAHP) synthase and alteredstem development in transgenic potato plants expressing a DAHPsynthase antisense construct. Plant Physiology 108, 1413–1421.
Junker BH, Wuttke R, Tiessen A, Geigenberger P,Sonnewald U, Willmitzer L, Fernie AR. 2004. Temporallyregulated expression of a yeast invertase in potato tubers allows
2066 Ding et al.
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

dissection of the complex metabolic phenotype obtained follow-ing its constitutive expression. Plant Molecular Biology 56,91–110.
Leuschner C, Herrmann KM, Schultz G. 1995. The metabolismof quinate in pea roots (purification and partial characterization ofa quinate hydrolyase). Plant Physiology 108, 319–325.
Lewis NG, Yakamoto E. 1990. Lignin, occurrence, biogenesis, anddegradation. Annual Review of Plant Physiology and PlantMolecular Biology 41, 455–496.
Li H, Culligan K, Dixon RA, Chory J. 1995. Cue1: A mesophyllcell-specific positive regulator of light-controlled gene expressionin Arabidopsis. The Plant Cell 7, 1599–1610.
Lichtenthaler HK. 1987. Chlorophylls and carotenoids, pigmentsof photosynthetic biomembranes. Methods in Enzymology 148,350–382.
Logemann J, Schell J, Willmitzer L. 1987. Improved method forthe isolation of RNA from plant tissues. Analytical Biochemistry163, 16–20.
Michel G, Roszak AW, Sauve V, Maclean J, Matte A,Coggins JR, Cygler M, Lapthorn AJ. 2003. Structures ofshikimate dehydrogenase aroe and its paralog ydib: a commonstructural framework for different activities. Journal of BiologicalChemistry 278, 19463–19472.
Moerschbacher BM, Noll U, Gorrichon L, Reisener H-J. 1990.Specific inhibition of lignification breaks hypersensitive resis-tance of wheat to stem rust. Plant Physiology 93, 465–470.
Mousdale DM, Coggins JR. 1985. Subcelluar localization of thecommon shikimate-pathway enzymes in Pisum sativum l. Planta163, 241–249.
Mustafa NR, Verpoorte R. 2005. Chorismate derived c6c1compounds in plants. Planta 222, 1–5.
Nicholson RL, Hammerschmidt R. 1992. Phenolic compoundsand their role in disease resistance. Annual Review of Phytopa-thology 30, 369–389.
Ober D. 2005. Seeing double: gene duplication and diversificationin plant secondary metabolism. Trends in Plant Science 10, 444–449.
Pichersky E, Gang DR. 2000. Genetics and biochemistry ofsecondary metabolites in plants: an evolutionary perspective.Trends in Plant Science 5, 439–445.
Pinto JE, Dyer WE, Weller SC, Herrmann KM. 1988.Glyphosate induces 3-deoxy-D-arabino-heptulosonate 7-phosphatesynthase in potato (Solanum tuberosum L.) cells grown insuspension culture. Plant Physiology 87, 891–893.
Rolleston FS. 1972. A theoretical background to the use ofmeasured intermediates in the study of the control of intermediarymetabolism. Current Topics in Cellular Regulation 5, 47–75.
Rolletschek H, Hajirezaei MR, Wobus U, Weber H. 2002.Antisense-inhibition of ADP-glucose pyrophosphorylase in Vicianarbonensis seeds increases soluble sugars and leads to higherwater and nitrogen uptake. Planta 214, 954–964.
Rosahl S, Schell J, Willmitzer L. 1987. Expression of a tuber-specific storage protein in transgenic tobacco plants—demonstra-tion of an esterase-activity. EMBO Journal 6, 1155–1159.
Samach A, Onouchi H, Gold SE, Ditta GS, Schwarz-Sommer Z, Yanofsky MF, Coupland G. 2000. Distinct roles ofconstans target genes in reproductive development of Arabidop-sis. Science 288, 1613–1616.
Schaarschmidt S, Qu N, Strack D, Sonnewald U, Hause B. 2004.Local induction of the alc gene switch in transgenic tobacco plantsby acetaldehyde. Plant Cell Physiology 45, 1566–1577.
Schmid J, Amrhein N. 1995. Molecular organization of theshikimate pathway in higher plants. Phytochemistry 39, 737–749.
Schmid J, Schaller A, Leibinger U, Boll W, Amrhein N. 1992.The in vitro synthesized tomato shikimate kinase precursor isenzymatically active and is imported and processed to the matureenzyme by chloroplasts. The Plant Journal 2, 375–383.
Singh SA, Christendat D. 2006. Structure of Arabidopsisdehydroquinate dehydratase-shikimate dehydrogenase and impli-cations for metabolic channeling in the shikimate pathway.Biochemistry 45, 7787–7796.
Singh S, Korolev S, Koroleva O, Zarembinski T, Collart F,Joachimiak A, Christendat D. 2005. Crystal structure of a novelshikimate dehydrogenase from Haemophilus influenzae. Journalof Biological Chemistry 280, 17101–17108.
Stitt M, Lilley RMC, Gerhardt R, Heldt HW. 1989. Determina-tion of metabolite levels in specific cells and subcellular compart-ments of plant leaves. Methods in Enzymology 174, 518–552.
Streatfield SJ, Weber A, Kinsman EA, Hausler RE, Li J, Post-Beittenmiller D, Kaiser WM, Pyke KA, Flugge UI, Chory J.1999. The phosphoenolpyruvate/phosphate translocator is re-quired for phenolic metabolism, palisade cell development, andplastid-dependent nuclear gene expression. The Plant Cell 11,1609–1622.
Timmermans MC, Maliga P, Vieira J, Messing J. 1990. The Pffplasmids: cassettes utilising camv sequences for expression offoreign genes in plants. Journal of Biotechnology 14, 333–344.
Vance CP, Kirk TK, Sherwood RT. 1980. Lignification as amechanism of disease resistance. Annual Review of Phytopha-thology 18, 259–288.
Voinnet O, Rivas S, Mestre P, Baulcombe D. 2003. An enhancedtransient expression system in plants based on suppression ofgene silencing by the p19 protein of tomato bushy stunt virus.The Plant Journal 33, 949–956.
Voll L, Hausler RE, Hecker R, Weber A, Weissenbock G,Fiene G, Waffenschmidt S, Flugge UI. 2003. The phenotype ofthe Arabidopsis cue1 mutant is not simply caused by a generalrestriction of the shikimate pathway. The Plant Journal 36, 301–317.
Weber APM, Schwacke R, Flugge UI. 2005. Solute transportersof the plastid envelope membrane. Annual Review of PlantBiology 56, 133–164.
Molecular analysis of shikimate synthesis in tobacco 2067
by guest on July 30, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from




















