
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania
46
Journal of World Prehistory, Vol. 13, No. 4, 1999 Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania Richard Cosgrove1,2 Over the past 18 years the results of archaeological field research in many parts of Australia have challenged the view of late Pleistocene archaeological unity and homogeneity. Debates are increasingly focused on issues of identi- fying regional behavioral variability and away from a 'normalization' of Australia's prehistory. Prior notions of cultural transformations from 'simple' (late Pleistocene) to 'complex' (late Holocene) social behaviors, technological homogeneity, and the timing of colonization are being increasingly scruti- nized. One such area that has become a spearhead of this challenge is Tasmania, which continues to demonstrate its ability to confront preconcep- tions about the nature of past human behaviors. INTRODUCTION Locked in the jaws of the roaring forties, the island of Tasmania is some 64,000 km2 in area and lies at the foot of the Australian continent (Fig. 1). The western region is remote, mountainous, wet, and covered by temperate rainforest and infertile button grass plain. Its coast is wild and highly productive. Sea mammals, birds, fish, and shellfish are nourished by the rich nutrients upwelling from the Southern Ocean. The Central Plateau 1School of Archaeological and Historical Studies, La Trobe University, Melbourne, Austra- lia, 3083. 2Correspondence should be addressed, to the author at School of Archaeological and Historical Studies, Department of Archaeology, La Trobe University, Melbourne, Australia 3083; e-mail: [email protected]. KEY WORDS: Australian prehistory; late Pleistocene Tasmania; hunter-gatherers; human colonization; palaeoecology; behavioral variability. 357 0892-7537/99/1200-0357$16.00/0© 1999 Plenum Publishing Corporation
Transcript of Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania

Journal of World Prehistory, Vol. 13, No. 4, 1999
Forty-Two Degrees South: The Archaeology ofLate Pleistocene Tasmania
Richard Cosgrove1,2
Over the past 18 years the results of archaeological field research in manyparts of Australia have challenged the view of late Pleistocene archaeologicalunity and homogeneity. Debates are increasingly focused on issues of identi-fying regional behavioral variability and away from a 'normalization' ofAustralia's prehistory. Prior notions of cultural transformations from 'simple'(late Pleistocene) to 'complex' (late Holocene) social behaviors, technologicalhomogeneity, and the timing of colonization are being increasingly scruti-nized. One such area that has become a spearhead of this challenge isTasmania, which continues to demonstrate its ability to confront preconcep-tions about the nature of past human behaviors.
INTRODUCTION
Locked in the jaws of the roaring forties, the island of Tasmania issome 64,000 km2 in area and lies at the foot of the Australian continent(Fig. 1). The western region is remote, mountainous, wet, and covered bytemperate rainforest and infertile button grass plain. Its coast is wild andhighly productive. Sea mammals, birds, fish, and shellfish are nourished bythe rich nutrients upwelling from the Southern Ocean. The Central Plateau
1School of Archaeological and Historical Studies, La Trobe University, Melbourne, Austra-lia, 3083.
2Correspondence should be addressed, to the author at School of Archaeological and HistoricalStudies, Department of Archaeology, La Trobe University, Melbourne, Australia 3083;e-mail: [email protected].
KEY WORDS: Australian prehistory; late Pleistocene Tasmania; hunter-gatherers; humancolonization; palaeoecology; behavioral variability.
357
0892-7537/99/1200-0357$16.00/0© 1999 Plenum Publishing Corporation

Fig. 1. Distribution of late Pleistocene Tasmanian sites with off-shore bathometry indicated.
area is cold, mountainous, and alpine, the source of many of Tasmania'slargest rivers. To the east is the drier, warmer, more open Midlands valleybounded by the wooded coastal ranges and the calmer inlets and bays ofthe Tasman Sea. This picturesque and rugged landscape evolved over thelast 10,000 years. Throughout this period Tasmania remained separated
Cosgrove358

from the Australian mainland by the waters of Bass Strait, and the Aborigi-nes endured the longest period of physical isolation of any human groupon the globe (Jones, 1978).
During the 18th and 19th centuries the Europeans' view of the indige-nous Tasmanians fluctuated between "noble savage" and "most primitivepeople on Earth" (Taylor, 1893; Jones, 1992). These perceptions also fo-cused attention on the nature and makeup of earliest human societies. TheTasmanians were seen to represent the lowest rung on the Paleolithic socialladder, their level of technology akin to ecofacts frozen in primordial time(Sollas, 1911). The interpretation of Tasmania's Holocene archaeologicalrecord has followed this path, where the remains were seen as the productof 10,000 years of isolation leading to cultural conservatism. It was perceivedas an example of a social system in decline, becoming more simplifiedover time (Jones, 1971a, 1977a). The rather minimal repertoire of materialculture items described at contact and the subsequent archaeological discov-eries of the cessation of fish-eating and the loss of bone-point technologyfrom the culture about 3500 B.P. reinforced this view. The loss, rather thanreplacement, was emphasized (Jones, 1971a, 1977b, 1984, 1995), althoughalternative explanations that highlighted economic adaptations were putforward (Allen, 1979; Bowdler, 1980; Thomas, 1981). The early impressionwas that Tasmanian Aborigines were representative of the Paleolithic age,populated by a society that provided, along with Terra del Fuegians andHottentots, the intellectual justification for the belief that societies evolvedfrom simple to complex.
MODERN HUMANS ON THE MOVE
The magnitude of the effects of the lengthiest human isolation in theworld is still debated among Australian archaeologists. One thing is clear,however: the people who first entered Tasmania at least 35,000 radiocarbonyears ago were fully modern humans. Over this period human behaviorchanged in response to varied social and ecological conditions, the charac-ter, frequency, and magnitude of which are the focus of this paper.
Remarkably, these colonists had moved far from their tropical entrypoint in northern Australia into the most southerly latitudes of GreaterAustralia. This feat is even more phenomenal as it was the final act ofsettlement of the western Pacific Rim that began perhaps 70,000 yearsearlier in Africa. The Aboriginal Tasmanians were part of this global spreadof anatomically modern humans, their ancestors using watercraft technolog-ically advanced enough to cross Wallacea, the biogeographic barrier sepa-rating the placental faunas of Eurasia and the unique marsupial faunas of
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 359

Greater Australia. The only other mammals to make this journey wererats and bats. They entered a landscape of giant kangaroos, wombats andemus, large carnivorous reptiles, marsupial lions, and egg-laying mammals.At their entry point in the tropics, plants and marine resources would havebeen familiar, to these travelers but many others would be totally unknownto them. Nevertheless, by 35,000 radiocarbon years ago humans had colo-nized all of the major environmental zones in Australia (Allen, 1989; Jones,1990; Smith, 1989; Cosgrove, 1989; Allen et al., 1989; O'Connor, 1990).
It is in this sense that the Aboriginal Tasmanians remain central toquestions about the timing and tempo of the final phase of continental andglobal settlement (Allen and Holdaway, 1995; Roberts et al., 1994; Cos-grove, 1995b; O'Connell and Allen, 1998). They are also central to generaldebates about the interaction of human biology, behavior, and technology(Holdaway and Cosgrove, 1997). For example, Tasmanians' stone technol-ogy has been equated with the early middle Paleolithic of the NorthernHemisphere, providing a contrast to the technological developments onthe Australian mainland during the mid-Holocene (Jones, 1977a). Theparadox is that these early colonists were anatomically modern, implyingno necessary link between morphology, behavior, and technology.
Equally, the perceptions of the relatively conservative character of latePleistocene Australian human behavior (White and O'Connell, 1982, p.72) and the nature of the archaeological database that has emerged fromsites in Tasmania over the last 18 years strengthen this situation. Over 40cave and open sites of late Pleistocene age demonstrate a level of richness,distribution, and variability not seen in other Australian archaeologicalsites (Kiernan et al., 1983; Cosgrove, 1989,1995b, 1997; Cosgrove and Jones,1989, Cosgrove et al., 1990; Brown et al., 1991; Porch and Allen, 1995; Allen,1996; Sim, 1998). These sites range, geographically, from near sea level toupland periglacial zones on the edge of the Tasmanian Central Plateau andspan the time period between 750 B.P. and 35,000 B.P. (Figs. 1 and 2).Intriguingly, the rich deposits in many of the caves compare favorably withthe densities found in the upper Paleolithic French, Spanish, and Russiansites. This is an aspect they do not share with sites of similar age onmainland Australia.
DYNAMIC LANDSCAPES
While the palaeoecology has been discussed in detail elsewhere (Cos-grove et al, 1990; Porch and Allen, 1995), a brief review of the importanttrends will be given. The changing nature of the environment between
Cosgrove360

361Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania

35,000 and 10,000 B.P. and human responses to them can be usefully viewedat a regional scale.
In the west before 45,000 B.P. the vegetation consisted mainly of alpineand subalpine taxa and the larger tree species were confined to loweraltitudes than today. In fact, between 21,000 and 18,000 B.P. the tree linewas depressed by at least 230 m in the west coast region (Gibson et al.,1987). Pollen core data analyzed from various lowland west coast locationssuggest that between 44,000 and 25,000 B.P. annual average temperatureswere 5°C colder than today, with a wetter climate (Colhoun et al., 1994).Lake levels are known to have fluctuated during this time, but contractedafter about 21,000 B.P. with the onset of glacial conditions (Colhoun andvan de Geer 1986; Porch and Allen, 1995). At 18,000 B.P. temperatureswere depressed by at least 6.5° C. Between 24,000 and 20,000 B.P. wetheath and herb communities, with increasing grass pollen after 22,000 B.P.,appear, peaking during the Late Glacial Maximum. Grass produces largequantities of pollen and is overrepresented at the expense of other species(Kirkpatrick, 1986). Grass was more likely restricted to more fertile sub-strates and limited in its distribution to patches of alluvial soils and limestonegeology (Kirkpatrick, 1986, p. 235; Cosgrove et al., 1990).
Further afield in the east, and north onto the Bassian Plain, the environ-ment was dominated by grasses and herbfields with limited occurrences ofheath and scrub (Hope, 1978; Colhoun et al., 1982; Colhoun, 1978). Muchwindier conditions, up to 8 km/hr faster, were also experienced aboutthis time, particularly in northeast Tasmania (Bowden, 1983). Sand sheetdevelopment and linear dunes dated to about 15,000 B.P. suggest similarconditions in southeast Tasmania (Sigleo and Colhoun, 1982). The east wasdry, drought-prone, and sparsely vegetated. Cold winters and short, coolsummers with strong seasonal variation were probably experienced duringthis time (Sigleo, 1978).
Amelioration of the glacial climate is detected in pollen data about14,000 B.P. Although glaciers reached their maximum extent just before19,000 B.P., deglaciation had probably occurred before 13,000 B.P. in mostlocalities (Colhoun et al., 1994). At this time sea levels were beginning torise over the exposed Bass Plain that had earlier allowed people to enterTasmania during the late Pleistocene. The link was finally severed about11,000 B.P. (Sim, 1998). The flow of outside genes, technology, and socialinfluences was cut for the next 10,000 years.
SETTING THE SCENE: FIRST ARRIVALS
People got to Tasmania by crossing exposed portions of the Bassianland bridge that joined Tasmania to Australia. The initial line of travel
362 Cosgrove

would have been via a narrow neck of land connecting Wilson's Promon-tory, Flinders Island, and northeast Tasmania (Fig. 1). This is the shallowestpart of Bass Strait, requiring a drop of only 55 m to expose, first, anarchipelago of islands and, shortly after, continuous dry land 300-400 kmwide (Sim, 1998, p. 291). Determining how and when they arrived is depen-dent upon an understanding of the timing and configuration of the landbridge and associated archaeological evidence.
The submarine topography along the 55-m sill depth is close to criticalsea level oscillation between 50,000 and 30,000 B.P. Calculated estimatesof variation are ±10 m and ±500 years (Sim, 1998, pp. 290-291). Therehas been considerable debate as to when people were able to move across.It is still not precisely known when and for how long the sill was exposedat any one time prior to 30,000 B.P. (Bloom, 1988; Cann et al., 1993; Jones,1995; Cosgrove, 1995; Sim, 1998; Jackson, 1999, p. 88).
Figure 2 displays the radiocarbon dates of cave deposits correspondingto sea-level changes over the last 55,000 radiocarbon years (Blom, 1988;Cann et al., 1993; Chappell, 1994; Chappell et al., 1996). Using estimatesbased on oxygen isotopes from deep sea cores and the Huon Peninsuladata from New Guinea (Chappell, 1994,1996), one can postulate continuousland link between the Australia and Tasmania about 50,000-10,000 B.P.(Cosgrove, 1995a; Sim, 1998, pp. 289-93). In this scenario people could haveentered Tasmania at any time during this period, although all archaeologicalevidence points to a date of around 35,000 radiocarbon years B.P. (Cos-grove, 1995a).
Blom (1988), on the other hand, has estimated at least three majorperiods of connection at 55,000-50,000, 37,000-31,000 and 29,000-12,000radiocarbon years ago (Fig. 2). Studies from south Australia show a marineinundation of Gulf St Vincent between 45,000 and 30,000 radiocarbon yearsago (Cann et al, 1988, 1993). Extrapolating these data to Bass Strait, anextravagant claim by Jackson (1999, p. 67) asserts that occupation occurredc. 150,000 B.P. since there was little probability of a land bridge between150,000 and 23,000 B.P. Earlier human presence prior to 45,000 B.P. hasbeen argued from C-14-dated vegetation changes noted in pollen cores andthe presence of noncultural burnt plant material in geological sediments(Jackson, 1999, p. 74). The implications of this contradictory evidence arediscussed below.
The two oldest sites are Warreen Cave and Parmerpar Meethanerrockshelter (Fig. 2). They are in limestone and quartzite geology, respec-tively. The basal date for Warreen is currently 34,790 ± 510 B.P. (Allen,1996, p. 154), while two AMS dates of 33,850 ± 450 and 33,260 ± 420 B.P.have been recorded for Parmerpar Meethaner (Cosgrove, 1995a). Thesetwo dates are underlain by a radiocarbon age of 39,970 ± 950 B.P., 10 cm
363Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania

down in a sterile layer. It has been suggested that these layers represent alandscape before human colonization. In Bone Cave the lower occupationlevels have been dated to 29,000 ± 520 and 28,330 ± 630 B.P. respectively(Allen, 1996, p. 113). In the basal layers of the Acheron Cave ages of29,800 ± 720 B.P. have been recorded (Allen 1996, p. 184). Slightly olderdates have been assayed for Nunamira Cave and ORS 7 rockshelter,30,840 ± 480 and 30,750 ± 1340 B.P., respectively.
Much younger dates from sites on three Bass Strait islands have beenrecorded. Cave Bay Cave has a basal age of about 23,000 B.P. (Bowdler,1984), while sites on Prime Seal Island and Badger Island have radiocarbondates of 23,015 ± 210 and 23,180 ± 1280 B.P., respectively (Sim, 1998,p. 258). These are ca. 12,000 radiocarbon years younger than the oldestTasmanian mainland site of Warreen at 35,000 radiocarbon years.
Thirty-five thousand years is argued to be too young by some workersbecause contamination by younger carbonates will underestimate the realage of initial human occupation, which is suggested to be as early as 150,000B.P. (Jackson, 1999, pp. 68, 88). Apart from the startling fact that thiswould require the Australian continent to be colonized more than 100,000years earlier than is generally accepted (O'Connell and Allen, 1998, p. 140;Roberts el al., 1990,1994; David et al, 1997), all radiocarbon dates shouldshow similar 'suppressed' ages. Importantly, the dates obtained from pollenand geological and marine cores do not reflect this equally from eitherTasmania or continental Australia.
In Tasmania dates of >45,000 radiocarbon years B.P. on charcoal fromnoncultural deposits in the state's north (Cosgrove and Murray, 1993) andradiocarbon ages from noncultural sediments on King Island (D'Costaet al., 1993) of between 39,400 and >55,000 B.P. have been obtained.Radiocarbon dates on wood of 38,800 ± 1300 B.P. (Jordan et al., 1991)and 48,700 + 2900/-2100 B.P. (Fitzsimons and Colhoun, 1991) have beenassayed, while the sterile deposits in Parmerpar Meethaner have an AMSdate on charcoal of 39,970 ± 950 B.P. There are other examples of dateson various organic materials in excess of 35,000 years from geologicalsamples cited by Colhoun (1985). European studies have also shown thatdates in excess of 35,000 radiocarbon years are possible using both conven-tional and AMS radiocarbon techniques. In an analysis of Chatelperronianand Aurignacian sites of southwest Europe, for example, Zilha and d'Errico(1999) tabulate a series of AMS dates, 20 of which are older than 35,000 B.P.
This strongly implies that irrespective of what the radiocarbon datesactually represent in 'real' years or their level of contamination, there isno apparent problem in obtaining radiocarbon ages in excess of 35,000 B.P.There are now over 140 radiocarbon dates from late Pleistocene archaeolog-ical sites that show internal chronological consistency between 13,000 and
364 Cosgrove

35,000 B.P. (Cosgrove, 1995a; Holdaway and Porch, 1996; Allen, 1996).Leaving aside the contradictory sea level estimates (Blom, 1988; Cann etal, 1993; Cosgrove, 1995a), if people had entered Tasmania before 35,000B.P., this should be reflected in the radiocarbon chronology.
O'Connell and Allen (1998) have recently examined claims of conti-nental human occupation in excess of 40,000 B.P. They explore the correla-tion between luminescence and radiocarbon dates, concluding that "C14dates ought not to be more than about 4,000 to 5,000 years short of theirtrue ages between 30,000 and 45,000 years, and might well be essentiallyidentical between 45,000 and 50,000. Both of these predictions are partlyconfirmed by recent experimental results" (p. 140).
They also examined the frequency of >40,000 B.P. ages from noncul-tural and cultural deposits from all over Australia. They found that radiocar-bon dates from noncultural deposits in excess of 45,000 B.P. were consis-tently reported, while dates from unequivocal cultural deposits were noolder than 40,000 B.P. Again they conclude that if Pleistocene Australiawas inhabited earlier than this, some radiocarbon dates of 45,000-50,000should be expected (O'Connell and Allen, 1998, p. 142). The same logicapplies to Tasmania. All the available archaeological evidence suggests thatpeople colonized western and northern Tasmania about 35,000 radiocarbonyears ago. There is no direct archaeological evidence for initial Aboriginaloccupation of Tasmania before 35,000 years.
PHYSICAL ANTHROPOLOGY
Discoveries of late Pleistocene human skeletal material from NanwoonCave in the Florentine River valley and at a cave site on King Islandprovide some limited insights into the physical characteristics of these earlycolonists. A partial occipital bone was discovered at the base of a rubblechute in Nanwoon Cave in 1987 and dated by stratigraphic association tobe more than 16,000 years old (Jones et al., 1988). The bone was stainedby manganese, but was very well preserved, the presence of unfused suturessuggesting that the individual was quite young (Webb, 1988). It was thin,about 1.0-1.5 mm in thickness, rounded with few muscular markings (Webb,1988). Overall its morphology was very gracile and modern in appearance.The remains are similar in form to late Pleistocene gracile skeletal materialfrom Lake Mungo and Keilor in New South Wales and Victoria, respec-tively.
A skeleton of a male found on King Island and indirectly dated toabout 14,000 B.P. suggests that people at about this time were gracile,reflecting modern morphological traits (Thorne and Sim, 1994). There is
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 365

no evidence of the robust forms like those found at Kow Swamp, Victoria,dated between 13,000 and 9,000 B.P., nor is there any similarity with thevery robust specimen WLH 50 from the Walandra Lakes (Webb, 1989, p.67). It is assumed on the basis of the limited evidence that the people whofirst entered Tasmania were anatomically modern humans.
SETTLEMENT PATTERN
The discoveries and subsequent analyses of late Pleistocene materialculture have reinforced the idea of widespread regional behavioral variabil-ity. These facts challenge many earlier notions, particularly that the latePleistocene was a time of archaeological unity across the continent andthat there was little regional variability (White and O'Connell, 1982, p. 72).The quality and quantity of the data from various parts of Tasmania attestto the richness and complexity of this archaeological record (Kiernan etal, 1983; Jones, 1990; Cosgrove, 1995b; Sim, 1998; McWilliams et al, 1999).
The preservation conditions in all southwest limestone cave depositsare exceptional. It is not uncommon to excavate 200,000 pieces of boneand 20,000 stone artifacts from less than 1 m3 of deposit (Kiernan et al,1983; Cosgrove, 1995b; Allen, 1996). Importantly, there are few majordisturbances to the deposits and most are sealed with a thin layer of lime-stone capping. To the east and north, sites spanning this period are verydifferent, with lower densities of cultural remains, but possessing similarpreservation conditions (Bowdler, 1984; Brown, 1993; Cosgrove, 1995a,b;Sim, 1998). All the cave deposits have escaped the modifications by agricul-tural practices found in various Eurasian and African sites after the lastIce Age.
The wide distribution and tempo of occupation of late Pleistocenesites indicate adaptations to changing ecological and social circumstances,particularly to increasing aridity after 25,000 B.P. Low-level occupation orabandonment has been suggested for some sites like Bone Cave at theheight of glacial conditions at 18,000 B.P. because of depressed moistureregimes identified in lake basins (Holdaway and Porch, 1995). However,the presence of charcoal hearths at Warreen, Pallawa Trountra, and PinigaNairana Caves about 18,000-19,000 B.P. indicate that these climatic condi-tions did not act equally on all human groups. These sites lie within thelarge Gordon-Franklin River catchment, one that probably provided reli-able water throughout the period. There is no evidence in western Tasmaniaof permafrost or semiarid conditions at the height of the last glacial maxi-mum that would have restricted resource availability or human movement(Fitzsimons and Colhoun, 1989). Glaciers at 19,000 B.P were at their great-
366 Cosgrove

est extent and affected the Central Plateau and isolated high-altitude valleys(Colhoun et al., 1996). Spring thaw and summer melt would have allowedat least seasonal flows through these river systems. However this does notexplain the lack of archaeological evidence in southeast Tasmania at theLast Glacial Maximum (LGM) given the presence of the large DerwentRiver. More likely, it was the relatively more stable ecosystem of southwestTasmania that attracted people and their resources to the area. This isdiscussed further below.
Increased movement of peoples and occupation of the Bass Plain afterca. 22,000 has been argued by Porch and Allen (1995, p. 723), who use rawartifact counts to track these changes. The increases only occur in relativeterms and when corrected for excavated sediment weight, artifact numbersare exceedingly small (Brown, 1993, Tables 4, 8, and 10). Similarly, thesites of Cave Bay Cave, Beaton Shelter, and Mannalargenna Cave havefar fewer artifacts and bone remains than any of the southwest sites occupiedat this time. Increases in artifact discard were noted at Beaton Shelter andMannalargenna Cave between ca. 15,000 B.P. and ca. 18,000 B.P. (Sim,1998, p. 275). However, the highest number is 12 flakes per 100 kg ofsediment in spit 23. The other 32 spits each contain less than 2 artifactsper 100 kg of sediment. In fact, Cave Bay Cave is effectively abandonedafter ca. 18,000 B.P. and not reoccupied until ca. 6600 B.P. (Bowdler, 1984,p. 51). The evidence indicates extremely low levels of occupation on theBass Plain, which contrasts markedly with increases in activity identifiedin the valleys in western Tasmania. It is the lower elevation sites like thosein the protected Franklin River valley in the west, such as Kutikina Cave,Deena Reena Cave, and the Flying Fox site, that show occupation for thefirst time and large increases in use after 19,000 B.P. (Jones, 1990; Allenand Cosgrove, 1996, pp. 6-7). Densities of over 200,000 marsupial bonesweighing over 30 kg and ca. 35,000 stone artifacts occur in ca. 0.65 m3 ofexcavated deposit in Kutikina Cave. These become particularly dense ca.17,000 B.P.
It has also been argued that sites at higher elevations appear to beoccupied earlier and abandoned later than those at lower elevations, al-though the evidence it is not straightforward (Porch and Allen, 1995, p.721). For example, Kutikina Cave is dated to between 15,000 and 19,770B.P. It lies on the Franklin River at an elevation of about 40 m above sealevel. Conversely, Warreen Cave is in the adjacent valley, only ca. 100 mhigher and 11 km distant, yet it was occupied much earlier, at 35,000 B.P.,showing increased levels of activity between 25,000 and 20,000 B.P. It ceasedto be occupied after 18,000 B.P. due to roof collapse. Sites such as Boneand Nunamira Caves at about 400 m fit the prediction and have occupationbetween about 30,000 and 13,000 B.P. Conversely, Mackintosh, Piniga Nai-
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 367

rana, and Maneena (Nelson River) Caves at 250, 200, and 240 m abovesea level, respectively, have evidence of occupation no earlier than ca.20,000 B.P. and go out of sustained use before ca. 11,000 B.P. They alsohave relatively lower levels of cultural materials.
Porch and Allen (1995) recognize that all these sites would have beenwithin easy reach of humans at most times of the year. Temperature varia-tions would have been on the order of a few degrees Centigrade andoverall there appears to be only a weak correlation with occupation periods,altitude, and temperature. Rather, the sites more likely reflect the varioushuman responses to changes in valley ecology, and their differential usewas probably linked to seasonal adjustments in resource availability andsocial obligations at a local scale. The presence of artifacts made on Darwinglass in many of the sites suggests movement of up to 100 km or more overthe region. In this way the sites probably operated as node points ona landscape from which people could take advantage of sedentary andpredictable resources (Cosgrove et al, 1990).
SITE TYPES
All the detailed analyses of assemblages are from cave sites. The focuson cave sites has remained the basis of the late Pleistocene prehistory ofthe region. Some workers have lamented this fact, but so far only one opensite, the Flying Fox in the Franklin River valley, has been dated to17,100 ± 1350 B.P. (Jones, 1990; Cosgrove et al., 1990; Freslov, 1993;Thomas, 1993; Cosgrove et al, 1994; Allen and Cosgrove, 1996, p. 7).This situation is partly a product of poor visibility within dense temperaterainforest, although even where ground visibility is good, few open siteshave been found. Surveys of the button grass plains and bare alkaline pans(Harris et al, 1988) surrounding Warreen Cave and within the Maxwellvalley have failed to find any evidence of open air sites despite minimalvegetation cover and excellent visibility.
Undated sites have been recorded outside the region, especially in theKing River valley and on the bare hills surrounding Queenstown (Corbett,1980; Cosgrove, 1985; Freslov, 1993). Artifact raw material of the typefound in late Pleistocene caves has been recognized on open sites. Wheredates have been obtained, however, they only range between ca. 300 and1500 B.P. (Kiernan et al., 1983; Freslov, 1993, p. 233). Open sites dated toca. 3,500 B.P. (Hartzell, 1993) in northwest Tasmania show the utilizationof wet eucalypt/rainforest vegetation after the mid-Holocene. In ParmerparMeethaner continued human occupation occurs through the forest phaseinto modern times, but at diminished levels compared to the late Pleistocene
368 Cosgrove

(Cosgrove, 1995a). Humans were able to exploit forests in a way that hasnot been detected in the southwest rainforests. At present there is no cleararchaeological evidence of continued occupation within the southwest zone.
Other site types include two stone sources used in artifact manufacture.The first, Darwin Crater, is a meteorite impact site formed about 700,000years ago (Fudali and Ford, 1979) and is the source of Darwin glass (Jonesand Allen, 1984, pp. 94,96). This material remains confined to the southwestregion. It is most abundant in cave deposits close to the crater, in particularMackintosh Cave, and at more than 100 km distant, Bone Cave has a singlepiece. On inspection of the crater in 1984, Jones found most of the naturallyoccurring pieces to be small, the largest 30-40 mm long (Jones and Allen,1984). Artifacts of this material have been consistently made using bipolartechniques. The strategy reflects the overall small original size of Darwinglass pieces collected by Aboriginal people visiting the crater. It appearsthat no quarrying was needed since most of the Darwin glass can be obtainedfrom upturned tree roots (Jones and Allen, 1984).
The second stone source more than likely occurs within the RaggedRange, although its precise location is unknown (Sheppard, 1997). At pres-ent it is the sole location for the cryptocrystalline chert common in thesites on the eastern boundary of the southwest zone, particularly Bone andNunamira Caves (Allen et al., 1988; Cosgrove, 1995b). The chert has alimited distribution, found most commonly in Nunamira (26.8%) and Bone(20.6%) Caves close to the range, but drops off in Pallawa Trounta (1.3%)and Warreen (1.6%) deposits (Sheppard, 1997). It suggests that raw materi-als were exploited locally and did not extend beyond sites in close proximityto sources in any large quantities, Darwin glass follows this trend. It isfound in small quantities in cave deposits and is most numerous in sites closeto the crater, for example, the Mackintosh Cave (Stern and Marshall, 1993).
Systematic ground surveys through the mid-1980s and early 1990sidentified three Aboriginal hand stencil art sites located deep inside lime-stone caves (Harris et al., 1988; Cosgrove and Jones, 1989). These red-ochre stencils of human hands are associated with sites that have no obviousoccupation deposit. Preliminary dating of the art indicates that they maybe on the order of 11,000 B.P. (Loy et al., 1990). At the site of WargataMina in the Cracroft River valley, they are located about 60 m from daylight,while a further seven complete arms in red ochre have been stenciled ontocave walls 1 km inside the cave (A. McGowan, personal communication).A number of children's hands have been placed very low down on thesouthern wall, indicating visits by family groups. At Ballawinne Cave 16red hand stencils have been placed along three walls about 20 m from theentrance in complete darkness. All appear to be adult in size. The last siteis Keyhole Cavern, containing three hands again about 40-50 m inside the
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 369

cave (Allen and Cosgrove, 1996, p. 35). No figurative art is known. It isassumed that this art is Pleistocene in age since it is stylistically similar,and placement of art is consistently deep inside caves.
TEMPO OF OCCUPATION
Occupation in pre-25,000 B.P. layers in a number of sites suggests aninitial burst of human activity. This is particularly evident in ParmerparMeethaner, Bone Cave, and Nunamira Cave, where bone and stone artifactdiscard rates are relatively high compared to the period bracketing 25,000-18,000 B.P. Conversely, in Warren Cave occupation is quite intense around22,000 B.P., suggesting local rather than regional influences on occupation.
Increased levels of occupation and wider regional connections occurafter 16,000 B.P. (Cosgrove, 1995a; Porch and Allen, 1995). This is reflectedin the overall stone artifact and bone discard rates rising after 16,000 B.P.In Bone, Stone, and Nunamira Caves there is also good evidence for agreater use of internal floor space after 16,000 B.P. Strong correlationsexist between sites during particular time cycles, with increases in discardrates occurring between >30,000 and 27,000 B.P. and between 18,000 and13,000 B.P. The trend is detected in the chronologically longer deposits ofNunamira, Bone, Pallawa Trounta, and Parmerpar Meethaner rockshelters.These are separated by periods of relatively lower discard rates.
Holdaway and Porch (1995) suggested that the occupation of sitesoccurred on a cyclical basis. They based their argument on the fluctuatingarray of radiocarbon determinations and strong correlation they presentwith past global climatic changes based on pollen and water regimes. Themagnitude and frequency of these fluctuations varied among sites, whereless frequent use or abandonment was linked to periods of relatively coolerand drier climatic conditions.
Similar observations have been made about the long-term influencesof climatic change, particularly the correlation with periods of sea levelrise and fall, during flooding sequences of the Flinders and Bass landbridgesand Tasmanian coastlines (Cosgrove, 1995a). The pulses of occupation,particularly the increases in discard between 35,000 and 25,000 B.P. andbetween 16,000 and 13,000 B.P., have been argued to originate at times ofsevere climatic instability (Cosgrove 1995a, p. 99).
Very fine scale climatic 'flickers' have been identified from various icecores in both the northern and southern hemispheres (Dansgaard et al.,1989; Taylor et al, 1993; White et al, 1994). They are short, representingyearly temperature differences of up to 7°C. They are especially noticeableduring the transitions between glacial and interglacial periods. It is apparent
370 Cosgrove

that these fluctuations are very intense between ca. 15,000 and ca. 13,000B.P. and between 40,000 and 27,000 B.P. Coincidentally, these are timeswhen occupation in many of the cave sites is most intense, as seen in large-scale changes of fauna and stone artifact discard. The sites that show thisphenomenon most clearly are Nunamira, possibly Kutikina, Bone, PallawaTrounta, and Parmerpar Meethaner rockshelters (Cosgrove, 1995a). Theincrease in activity in these southwest sites may reflect the climatic instabilityin eastern Tasmania compared to the relatively more stable southwest. Itcould indicate attempts by humans to mitigate the effects of unpredictableresources and lower risk and uncertainty, although these explanations needfurther testing with archaeological and ecological data.
AN ABANDONED LANDSCAPE?
The initial discovery of Kutikina Cave and its excavation provided thefirst evidence for repeated occupation of the southwest region during thelast glacial period (Kiernan et al, 1983). A cessation of occupation occurredafter ca. 15,000 B.P. Further archaeological work has reinforced this trend,with all cave sites going out of use by at least 13,000 B.P. (Allen, 1996).Figure 2 shows this strong chronological pattern. Those sites that lie outsidethe southwest, such as Parmerpar Meethaner and ORS 7, continue to beoccupied into the Holocene period, although at much lower intensities(Cosgrove, 1995a).
This has led many to argue that the southwest was abandoned afterthe end of the LGM due to increasing rainforest cover, moister conditions,and, importantly, a shrinking terrestrial resource base (Jones, 1984, 1995;Cosgrove et al, 1994; Cosgrove, 1995b; Porch and Allen, 1995). With in-creasing rainforest, the prey animals so commonly represented in all cavedeposits were displaced from their grassland/herbfield habitats, and sohumans moved away from the region. The economic strategy that hadsustained people for over 1000 generations came to an end with encroachingtemperate rainforest.
While the archaeological evidence from the southwest provides over-whelming support for abandonment of this late Pleistocene economic strat-egy, some workers have questioned this interpretation (Thomas, 1993,1995;Jackson, 1999). In particular, some have asserted that human groups contin-ued to regularly use the western region through the Holocene period. Ifthis were the case, continued occupation of southwest shelters and cavesshould be expected in a similar manner to that documented at ParmerparMeethaner and ORS 7 (Fig. 2).
The evidence in support of continued use is circumstantial and based
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 371

only on changing vegetation regimes, especially the appearance of fire-adapted species and their survival into the Holocene period detected inpollen cores. The argument is based on the fact that pyrogenic plant commu-nities have been identified in sediments dated to 38,000 B.P. and thatrainforest has been excluded from extensive areas of the southwest for atleast the last 12,000 years. The dominant species is the sedge Gymnoschoe-nus spp. (buttongrass) that grows extensively on infertile substrates and isfire-adapted. It is assumed that since there is evidence for the long-termpresence of fire-adapted sedges, people must have been present and werethe major ignition source for the fires. A further assumption is that thesefires must have been regularly lit to sustain these vegetation types through-out this period (Thomas, 1995, p. 299).
However, rainforest is very fire-sensitive and can be effectively ex-cluded from its range by regular burning. Fires lit by people in the recentpast have destroyed large tracts of rainforest vegetation and even underlow-intensity burning its distribution is severely restricted (Jackson, 1978,pp. 98-101; Marsden-Smedley, 1998). The abrupt abandonment of all cavesites associated with rainforest suggests that either the use of fire sticks wasineffectual against encroaching rainforest or that people did not deliberatelykeep areas around them open. Aboriginal people probably also recognized,as modern ecologists have shown, that regular firing decreases the fertilityof soils (Kirkpatrick and Dickson, 1984). In fact it has been observed that"The nutrient content of the biomass in the ecosystems occupying thesiliceous soils in areas of high rainfall in Tasmania is not such as to behighly attractive to large numbers of marsupial herbivores, even after fire(Bowman and Jackson, 1981), when compared with grassy ecosystems onmore fertile soils" (Kirkpatrick and Duncan, 1987, p. 77).
Field data showing that at least 70% of Tasmania's native fauna livein the infertile regions of southwest Tasmania, but at very low densities,support this observation. Hocking and Guiler (1982), for example, recordedonly five Bennett's wallabies after 2400 trap nights in a range of vegetationtypes. Conversely, in the grassy woodlands of eastern Tasmania 1,586,306brushtail possums, 8,110,410 ring-tail possums, and 1,802,600 Bennett'swallaby skins were presented for royalties between 1923 and 1970 (Guiler,1973). Driessen recorded over 2,092,071 Bennett's wallaby shot between1981 and 1986 (1988, pp. 22). This supports the contention of a directrelationship between high soil fertility, grass-herbfield growth, and in-creased densities of grazing herbivores.
As argued above, all the caves within the southwest associated withrainforest today were abandoned about 13,000 B.P. with the exception ofWarreen, vacated about 17,000 B.P. after the roof collapsed. The only siteto have postglacial evidence of human presence is Nunamira Cave. It has
372 Cosgrove

a few flakes and a core lying atop a thick layer of limestone capping formedafter 11,000 B.P. Only one open site of Holocene age has been identifiedat the Denison River mouth containing a few flakes and a quartzite coredated to 300 ± 150 B.P. (Kiernan et al., 1983). The five dated sites in theKing River valley have late Holocene ages of between ca. 300 and ca. 1200B.P. (J. Freslov, personal communication). As discussed previously, wheregood visibility is present and associated with sedgeland, no open sites havebeen detected within the southwest zone (Cosgrove et al., 1994).
There is therefore a gap of at least 12,000 years between late Pleisto-cene cave and late Holocene open site occupation. There is no convincingarchaeological data to suggest that people continued to occupy any of thesites associated with rainforest after 13,000 B.P. Whether or not peoplewere in the area during the Holocene associated with infertile sedgelandis a debatable point and one that could be tested further. The most parsimo-nious explanation for the pattern at present is that people abandoned theirsuccessful 20,000-year economic strategy at the end of the Pleistocene. Withfewer prey animals and plants to sustain a human presence, the cave siteswere abandoned (Jones, 1984; Cosgrove et al., 1990).
TECHNOLOGICAL CHANGE
Two issues in particular have dominated the discussion of stone tooltechnology and raw material use in late Pleistocene Tasmanian archaeology.The first is the timing for the appearance of a small rounded tool, the"thumbnail scraper," and associated Darwin glass (Jones, 1989,1990,1995),the second is the idea of increasing mobility after 16,000 B.P, with a concom-itant rise in the intensity of tool curation (McNiven, 1994). The debateshave focused on these because they are the most typologically distinctiveand were seen as concomitant with technological variation in the Europeanupper Paleolithic. The newer evidence suggests that these notions are pre-mature because until now only a small proportion of the entire stoneassemblage had been studied. With the completion of the Southern ForestArchaeological Project database (McWilliams et al., 1999), its analysis isleading to a fuller understanding of technological variability. Many earlyclaims made for the direction and tempo of technological change need tobe reexamined in light of this more recent data (e.g., McNiven, 1994; Jones,1990, 1995).
Thumbnail Scrapers
These small, disc-shaped scrapers have been a focus for study eversince the first Ice Age sequences in Kutikina Cave came to light in 1983
373Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania
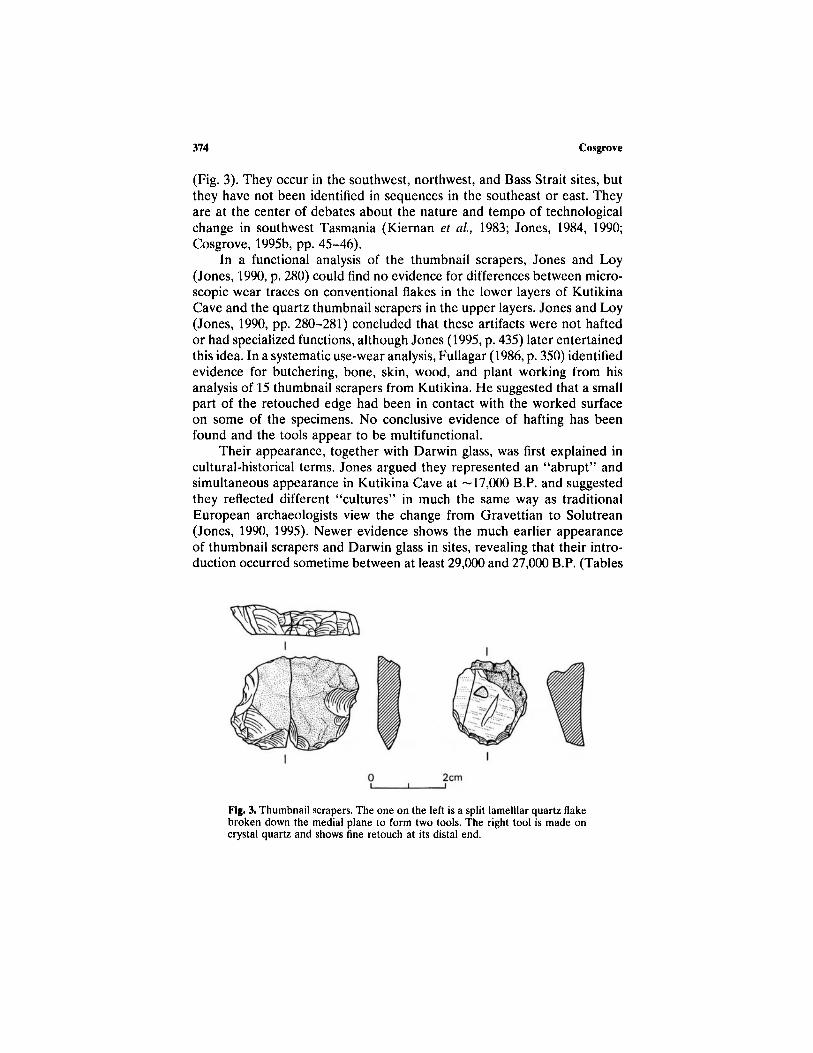
(Fig. 3). They occur in the southwest, northwest, and Bass Strait sites, butthey have not been identified in sequences in the southeast or east. Theyare at the center of debates about the nature and tempo of technologicalchange in southwest Tasmania (Kiernan et al, 1983; Jones, 1984, 1990;Cosgrove, 1995b, pp. 45-46).
In a functional analysis of the thumbnail scrapers, Jones and Loy(Jones, 1990, p. 280) could find no evidence for differences between micro-scopic wear traces on conventional flakes in the lower layers of KutikinaCave and the quartz thumbnail scrapers in the upper layers. Jones and Loy(Jones, 1990, pp. 280-281) concluded that these artifacts were not haftedor had specialized functions, although Jones (1995, p. 435) later entertainedthis idea. In a systematic use-wear analysis, Fullagar (1986, p. 350) identifiedevidence for butchering, bone, skin, wood, and plant working from hisanalysis of 15 thumbnail scrapers from Kutikina. He suggested that a smallpart of the retouched edge had been in contact with the worked surfaceon some of the specimens. No conclusive evidence of hafting has beenfound and the tools appear to be multifunctional.
Their appearance, together with Darwin glass, was first explained incultural-historical terms. Jones argued they represented an "abrupt" andsimultaneous appearance in Kutikina Cave at —17,000 B.P. and suggestedthey reflected different "cultures" in much the same way as traditionalEuropean archaeologists view the change from Gravettian to Solutrean(Jones, 1990, 1995). Newer evidence shows the much earlier appearanceof thumbnail scrapers and Darwin glass in sites, revealing that their intro-duction occurred sometime between at least 29,000 and 27,000 B.P. (Tables
Fig. 3. Thumbnail scrapers. The one on the left is a split lamelllar quartz flakebroken down the medial plane to form two tools. The right tool is made oncrystal quartz and shows fine retouch at its distal end.
374 Cosgrove
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania
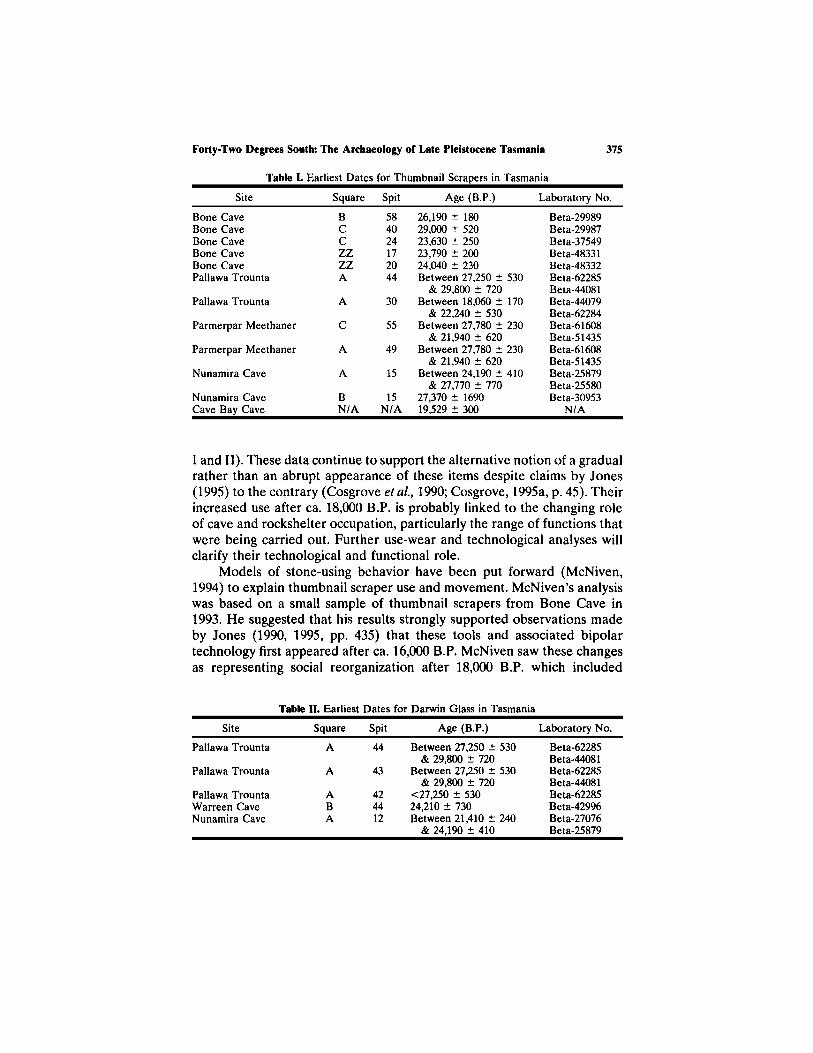
Table I. Earliest Dates for Thumbnail Scrapers in Tasmania
Site
Bone CaveBone CaveBone CaveBone CaveBone CavePallawa Trounta
Pallawa Trounta
Parmerpar Meethaner
Parmerpar Meethaner
Nunamira Cave
Nunamira CaveCave Bay Cave
Square
BCCZZZZA
A
C
A
A
BN/A
Spit
584024172044
30
55
49
15
15N/A
Age (B.P.)
26,190 ± 18029,000 ± 52023,630 ± 25023,790 ± 20024,040 ± 230Between 27,250 ± 530
& 29,800 ± 720Between 18,060 ± 170
& 22,240 ± 530Between 27,780 ± 230
& 21,940 ± 620Between 27,780 ± 230
& 21,940 ± 620Between 24,190 ± 410
& 27,770 ± 77027,370 ± 169019,529 ± 300
Laboratory No.
Beta-29989Beta-29987Beta-37549Beta-48331Beta-48332Beta-62285Beta-44081Beta-44079Beta-62284Beta-61608Beta-51435Beta-61608Beta-51435Beta-25879Beta-25580Beta-30953
N/A
I and II). These data continue to support the alternative notion of a gradualrather than an abrupt appearance of these items despite claims by Jones(1995) to the contrary (Cosgrove et al., 1990; Cosgrove, 1995a, p. 45). Theirincreased use after ca. 18,000 B.P. is probably linked to the changing roleof cave and rockshelter occupation, particularly the range of functions thatwere being carried out. Further use-wear and technological analyses willclarify their technological and functional role.
Models of stone-using behavior have been put forward (McNiven,1994) to explain thumbnail scraper use and movement. McNiven's analysiswas based on a small sample of thumbnail scrapers from Bone Cave in1993. He suggested that his results strongly supported observations madeby Jones (1990, 1995, pp. 435) that these tools and associated bipolartechnology first appeared after ca. 16,000 B.P. McNiven saw these changesas representing social reorganization after 18,000 B.P. which included
Table II. Earliest Dates for Darwin Glass in Tasmania
Site
Pallawa Trounta
Pallawa Trounta
Pallawa TrountaWarreen CaveNunamira Cave
Square
A
A
ABA
Spit
44
43
424412
Age (B.P.)
Between 27,250 ± 530& 29,800 ± 720
Between 27,250 ± 530& 29,800 ± 720
<27,250 ± 53024,210 ± 730Between 21,410 ± 240
& 24,190 ± 410
Laboratory No.
Beta-62285Beta-44081Beta-62285Beta-44081Beta-62285Beta-42996Beta-27076Beta-25879
375
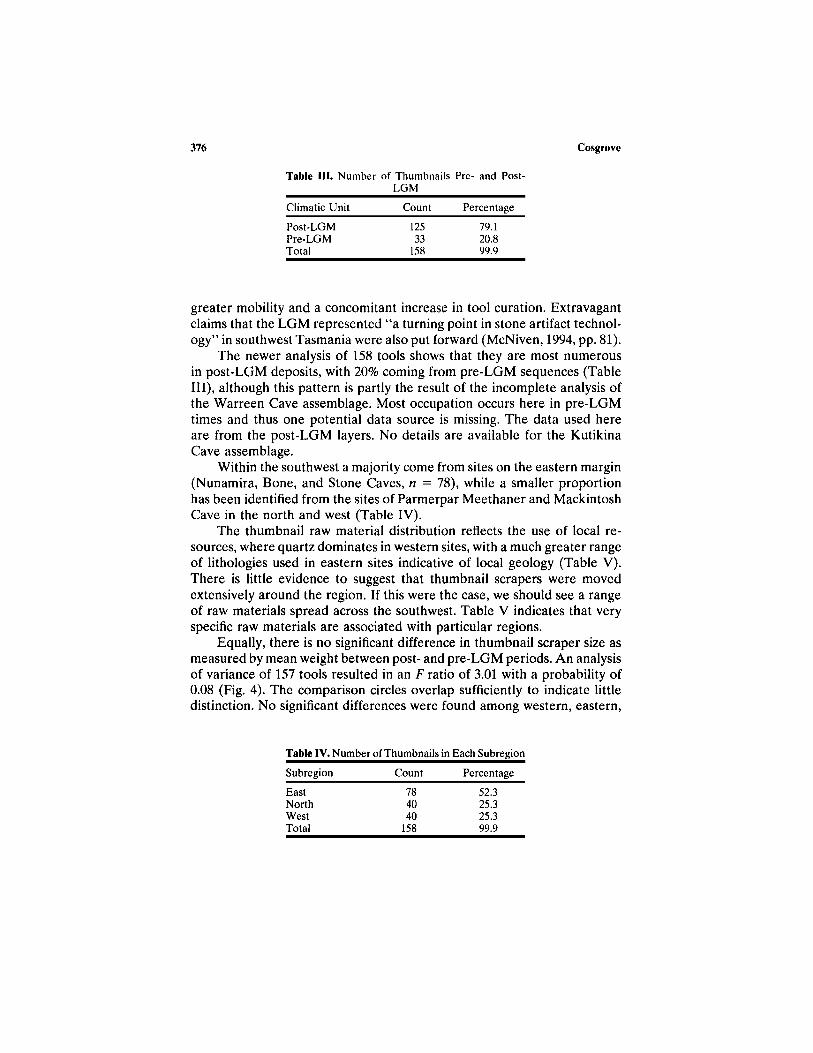
Table III. Number of Thumbnails Pre- and Post-LGM
Climatic Unit
Post-LGMPre-LGMTotal
Count
12533
158
Percentage
79.120.899.9
greater mobility and a concomitant increase in tool curation. Extravagantclaims that the LGM represented "a turning point in stone artifact technol-ogy" in southwest Tasmania were also put forward (McNiven, 1994, pp. 81).
The newer analysis of 158 tools shows that they are most numerousin post-LGM deposits, with 20% coming from pre-LGM sequences (TableIII), although this pattern is partly the result of the incomplete analysis ofthe Warreen Cave assemblage. Most occupation occurs here in pre-LGMtimes and thus one potential data source is missing. The data used hereare from the post-LGM layers. No details are available for the KutikinaCave assemblage.
Within the southwest a majority come from sites on the eastern margin(Nunamira, Bone, and Stone Caves, n = 78), while a smaller proportionhas been identified from the sites of Parmerpar Meethaner and MackintoshCave in the north and west (Table IV).
The thumbnail raw material distribution reflects the use of local re-sources, where quartz dominates in western sites, with a much greater rangeof lithologies used in eastern sites indicative of local geology (Table V).There is little evidence to suggest that thumbnail scrapers were movedextensively around the region. If this were the case, we should see a rangeof raw materials spread across the southwest. Table V indicates that veryspecific raw materials are associated with particular regions.
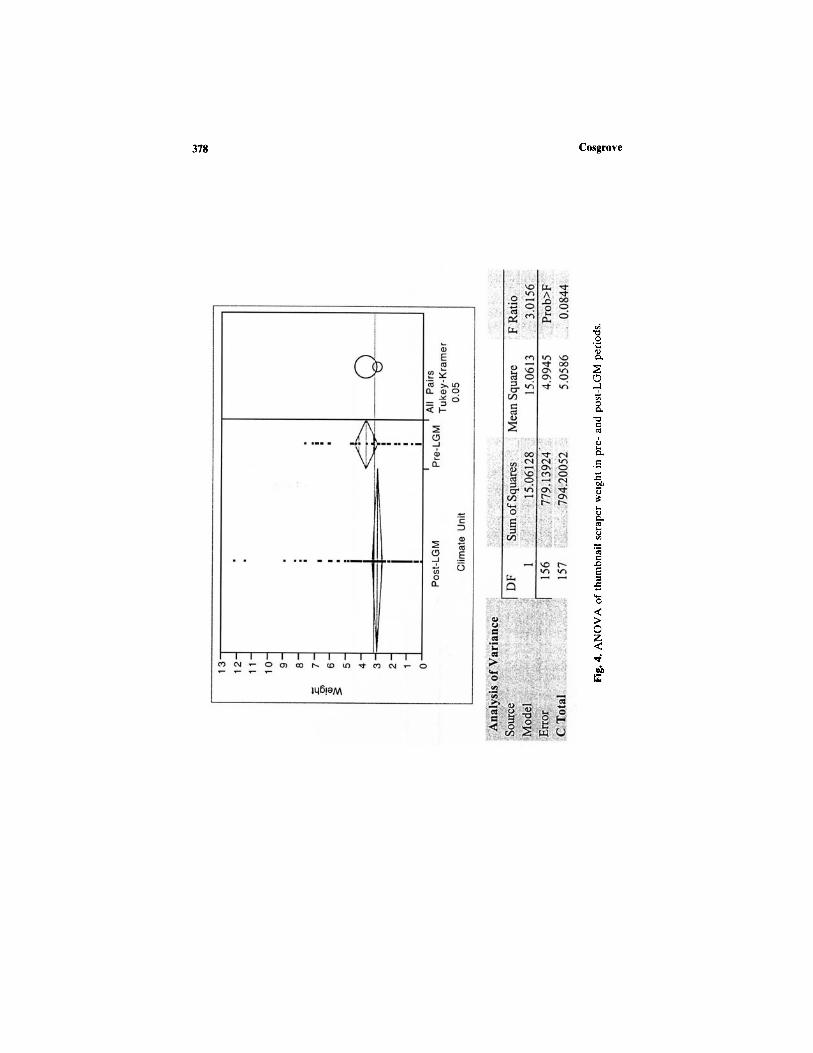
Equally, there is no significant difference in thumbnail scraper size asmeasured by mean weight between post- and pre-LGM periods. An analysisof variance of 157 tools resulted in an F ratio of 3.01 with a probability of0.08 (Fig. 4). The comparison circles overlap sufficiently to indicate littledistinction. No significant differences were found among western, eastern,
376 Cosgrove
Table IV. Number of Thumbnails in Each Subregion
Subregion
EastNorthWestTotal
Count
784040
158
Percentage
52.325.325.399.9

Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 377
Cosgrove378

and northern sites either (Fig. 5). Tools made on local quartz in the westare slightly smaller (mean = 2.4 g) than those found in the east (mean =3.4 g) and north (mean = 3.4 g). However, the comparison circles againshow no significant difference among subregions. A different pattern isreflected generally in individual sites (Fig. 6), where, for example, largerquartz thumbnails are found at Parmerpar Meethaner. The comparisoncircles indicate a significant difference between the small tools found atthe Mackintosh site and larger ones at Nunamira Cave, Warreen Cave,and Parmerpar Meethaner rockshelter. The data show that differences areat a site rather than a regional level as previously proposed by McNiven(1994).
The results from a much larger sample from six sites do not supportearlier interpretations. First, there is no evidence for an abrupt introductionof thumbnail technology into sites after 18,000 B.P. accompanied by Darwinglass. Although there are increased discard rates at sites after 16,000 B.P.,there is no evidence for size difference to suggest increased curation awayfrom raw material sources. In any case there is no a priori reason to believethat this represents any greater mobility of people either before or afterLGM times. The high percentage of local use of thumbnail scraper rawmaterials, the lack of evidence that they became significantly smaller withdistance, their gradual introduction, and the incidental but limited interre-gional use of stone resources (Sheppard 1997) refute the earlier notionsby McNiven (1994) and Jones (1995).
The regular movement of significant quantities of raw material of over150 km is a pattern described in the European Paleolithic and argued toreflect social interaction (Kozlowski, 1991; Rensink et al., 1991; Feblot-Augustins 1993; Thacker, 1997, p. 83). In Tasmania the acquisition of stoneand its transport over long distances is relatively limited. There is littleevidence for lithic exchange goods, which may, on the face of it, suggestlack of social interaction (Gamble, 1993). Organic exchange goods mayhave been used, but there is no evidence of these. The fact that this limitedAboriginal stone raw material distribution reflected in regional geology(Sutherland, 1973) continues into the Holocene is interesting because thereis no evidence of social or genetic isolation because of low population levelswithin the Aboriginal populations (Pardoe, 1986,1991). In fact it has beensuggested that Holocene population estimates by Jones (1971b) are toolow and that late Pleistocene numbers were high (Pardoe 1986). In addition,there are no strong morphological groupings that could be expected inclosed social networks like those argued for Row Swamp robust groupfound in the Murray Valley (Pardoe, 1988).
The archaeological distribution of Tasmanian Aboriginal stone rawmaterials may not be a very useful attribute for tracking interaction in the
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 379

Cosgrove380

Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 381

same way as it is used in the Eurasian upper Paleolithic or Americananthropological models of stone-using behavior. Certainly the wide regionaldistribution of small amounts of Darwin glass of up to 100 km from itssource provides us with a very important insight into the relationshipsbetween cave sites. However, there is no evidence that it left the southwestregion and was moved over extensive distances. It may also be the casethat the time scale over which these processes operated is too large, spatiallyand temporally, to identify the subtle features in artifact patterning at anyone moment in time.
Bone Technology
A number of bone points made from the fibulae of the red-neckedwallaby were also identified (Cosgrove, 1997) and were inferred to representclothing manufacture, although other, alternative uses have been suggestedthrough use-wear analysis (Webb and Allen, 1990). The earliest evidencefor such activities appears at Bone and Warreen Caves at 23,420 ± 340B.P. and before 27,160 ± 250 B.P., respectively (Table VI). Suggestionshave been made that these tools also functioned as spear points, but thereis no conclusive evidence of hafting on the shafts of these tools. No clearcorrelations exist between prey animals and the number of bone pointsexcavated. In Nunamira Cave, for example, similar amounts of depositwere excavated as in other sites, yet no bone points were found. Someexplanations for this present themselves, such as functional differencewithin and between caves. Nevertheless, until late Pleistocene researchmoves beyond the limited sampling of cave sites, we are constrained bythe nature of the data and explanations of its variability.
FAUNAL REMAINS
On average about 70% of the bone identified to species from all south-western sites are Bennett's wallaby (Macropus rufogriseus) and about 5%
Table VI. Earliest Dates for Bone Tools in Tasmania
Site
Bone CaveWarreen Cave
Warreen Cave
Square
BB
A
Spit
4257
17
Age (B.P.)
23,420 ± 340Between 31,610 + 370
& 27,160 ± 25023,370 ± 470
Laboratory No.
Beta-48330Beta-46873/ETH-8509Beta-46872/ETH-8508Beta-26962
382 Cosgrove

are attributed to the wombat (Vombatus ursinus). Sites located outside thezone have a generally similar array of terrestrial human prey, but they areat much lower densities (Bowdler, 1984; Cosgrove, 1995a; Sim, 1998). Othermedium- and small-sized animals are present, although many of these havearrived at the sites by natural processes. Table VII lists the NISP and weightfor all species found in five southwest cave sites. Due to high fragmentationlevels in some sites, a large proportion remain unidentified.
Species like the eastern grey kangaroo (Macropus giganteus) and emu(Dromaius novaehollandi), known to exist in some of the valleys as fossildeposits (Murray, 1978; Murray et al., 1980), and the eastern barred banicoot(Perameles gunnii) are very rare. They prefer open, grassy woodland habi-tats. In the Bass Plain sites, species like the eastern hare wallaby (Lagorch-estes leporides), eastern grey kangaroo (Macropus giganteus), and rufousrat kangaroo (Aepyrymnus rufescens) reflect the dry, open grassy ecologicalsettings of these sites during the glacial period (Brown, 1993, p. 262).Marine shell is also found throughout the deposits of Mannerlargenna Cavecollected and utilized as tools by Aborigines from the nearby coast.
The species from the southwest late Pleistocene cave sites, on the otherhand, show an animal community adapted to moist, LGM glacial conditionssupporting low, shrubby vegetation communities like those on the highwestern mountain regions today (Kirkpatrick, 1986). The almost total ab-sence of species preferring open environments reflects the lack of extensivegrasslands, particularly in sites to the west. Of these sites, only NunamiraCave, located in the broad, open Florentine River valley, contains evidenceof emu eggshell (Cosgrove et al., 1990). It occurs in layers dated to ca.28,000 B.P. and in increased quantities after 16,000 B.P., suggesting the sitewas occupied at least during late winter to early spring, the harshest time forhunters and gatherers and their prey (Cosgrove et al., 1990). The existence ofemu leg bones and vertebrae at Nunamira also supports interpretationsmade by Murray et al. (1980) for Beginners Luck Cave that open valleyconditions prevailed at this time. The limited evidence for this flightlessbird shows low-intensity hunting within the valley. It is difficult to judgewhether this was because of its low numbers or because emu butchery waspracticed away from caves, or whether it was nonpreferred prey.
In Tasmania there were no large predators such hyenas, leopards, orwolves that make significant contributions of human prey-sized animals tocultural deposits. These can pose interpretative challenges, particularly inidentifying the source of the bone and whether the prey were scavengedor hunted by humans (Lyman, 1994, pp. 205-216; Stiner, 1994, pp. 95-138).In the Tasmanian sites owls and marsupial carnivores are the main nonhu-man accumulators of the rat- and mouse-sized fauna (Marshall and Cos-grove, 1990; Northwood, 1990; Gale, 1994; Garvey, 1999). It explains the
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 383

384 Cosgrove
Table VII. Species in Southwest Tasmanian Sites
MammalAntechinus (marsupial mouse)Antechinus swainsonii (dusky marsupial mouse)Cercartetus (pygmy possum)Cercartetus lepidus (little pygmy possum)Dasyurus (quoll)Dasyurus maculatus (tiger cat)Dasyurus viverrinus (native cat)Hydromys chrosogaster (water rat)Isoodon obesulus (southern brown bandicoot)Large macropodLarge mammalMacropus giganteus (eastern grey kangaroo)Macropus rufogriseus (Bennett's wallaby)Mastacomys fuscus (broad-tooth rat)Medium mammalOrnithorhynchus anatinus (platypus)Perameles gunnii (eastern barred bandicoot)Petaurus breviceps (sugar glider)Potorous tridactylus (potoroo)Pseudocheirus peregrinus (ring-tail possum)Pseudomys (native mouse)Pseudomys higginsi (long-tailed mouse)Rattus lutreolus (swamp rat)Sarcophilus harrisii (Tasmanian devil)Small macropodSmall mammalSmall marsupialSminthopsis leucopus (white-footed dunnart)Tachyglossus aculeatus (echidna)Thylogale billardierii (rufous allaby)Trichosurus vulpecula (brush-tail possum)Unidentified mammalUnidentified rodentVombatus ursinus (wombat)ReptileUnidentified reptileBirdTyto spp. (owl)Dromaius novaehollandi (Tasmanian emu)Emu egg shellGallinula mortierii (Tasmanian native hen)Unidentified birdUnidentifiedUnidentified taxon
Total
NISP
25313286325
9304
115
417025436
49262953
2357123
11
24137
469
11313
2147173278
23
8643
56734172724
49
1124327
162
206470
264711
Weight (g)
16.910.491.258.845.67.64
73.850.091.31
1677.479135.08
31.917210.26
154.65227.1759.320.60.054.19
17.960.43.137.93
39.8889.21
385.3428.30.062
134.919.24
86.65242.75
4287.65
1.6
0.726.675.3
22.3847
14823.12
48883.48

high proportion of these animals listed in Table VII. They may also beresponsible for the much lower numbers of medium-sized prey between 1and 5 kg such as the two possum species, ring-tail (Pseudocheirus peregrinus)and brushtail (Trichosurus vulpecula), the brown banicoot (Isoodon obesu-lus), and the potoroo (Potorous tridactylus). Some species, like possums,only appear in the upper layers of the deposits at the end of the glacialand herald a change from nonforested to forested conditions.
It is clear, however, that humans have hunted the two larger marsupialspecies, Bennett's wallaby (Macropus rufogriseus) and wombat (Vombatusursinsus) (Kiernan et al., 1983; Cosgrove, 1995b; Cosgrove and Allen, 2000).The platypus (Ornithorhynchus anatinus) is also a minor, but significanthuman prey animal found in the sites (Marshall, 1992). This animal is shyand elusive and could only have found its way there through human hunting.
The lack of the two large carnivore species Sarcophilus harrisii(Tasmanian devil) and Thylacinus cynocephalus (Tasmanian tiger) is nota-ble. Importantly, there is no evidence of megafauna in any of the sites,thought to have become extinct in this region by 35,000 B.P. (Kiernan etal., 1983; Bowdler, 1984; Cosgrove, 1989,1995b; Jones, 1990; Brown, 1993;Allen, 1995; Porch and Allen 1995; Sim, 1998). This probably reflects thesubtle ecological differences between regions and the different ecologicalrequirements of megafaunal species that allowed them to survive in otherparts of Australia alongside human communities (Furby, 1995; Fullagarand Furby, 1997).
The highest site densities of bone occur in levels dated to the post-LGM, after ca. 17,000 B.P. (Fig. 7). Although the volumes of excavateddeposit differ slightly between sites, the trend is strong in the six sitesexamined (Allen, 1996). Warreen Cave is the exception, with much higherbone discard during preglacial times. The deposit dated to between 23,000and 19,000 B.P. contains a rich bed of burnt and smashed wallaby andwombat bone. The dominance of Bennett's wallaby is overwhelming, whilevariability in the minor species, for example, more wombat remains in thewestern sites and the presence of emu and their eggs in the east, reflectsthe exploitation of local animal communities (Cosgrove, 1995b).
The other notable finding is the very low abundance of medium-sizedmammals in all sites. These have a high probability of being incorporatedinto the deposits whole, since they are easily transported and do not needto be cut up into body parts for removal to sites for processing and consump-tion. This lends support to the contention that these species were beingoverlooked in favor of the larger Bennett's wallaby meat package. Thishas some important implications, and at least two alternative argumentscould be made for this pattern. The first has been longstanding, that theseanimals were deliberately targeted by human predators (Kiernan et al,
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 385

386 Cosgrove
Fig. 7. Pre- and post-LGM NISP for Macropus rufogriseus at each cave site.

1983) and that the medium-sized mammals, although present, were largelyignored. The second argument assumes that these smaller animals weresimply not as abundant as the wallaby and that people were encounteringwallabies much more often than any other mammals. It is difficult to choosebetween explanations because of the low resolution of most deposits. How-ever, an understanding of hunting strategies is crucial in illuminating preda-tor-prey relationships in sub-Antarctic environments (Cosgrove and Allen,2000). This has implications for detecting forward planning, preparation,and prey search patterns. Models of 'encounter' and 'intercept' huntingtechniques have been developed elsewhere to explain the predominanceof one or two human prey animals in Northern Hemisphere sites (Boyle,1990, pp. 96-101; Burke and Pike-Tay, 1991, pp. 69-79; Thacker 1997,pp. 88-92). Alternative explanations have been put forward for the latePleistocene Tasmanian sites (Cosgrove et al, 1990). This is discussed fur-ther below.
The other important feature of the bone assemblage is the very highnumber of small mammals, particularly of muridae, correlated with theterminal Pleistocene and the onset of wetter conditions after 12,000 B.P.in most sites. Where earlier sequences of occupation are preserved, suchas at Warreen Cave, much of the small fauna occur in the lower depositsbefore 25,000 B.P. (Cockbill, 1999). Environmental evidence from pollencores obtained from swamp sites in western Tasmania shows that moistconditions prevailed with rainforest and eucalypt vegetation present aboutthis time (Colhoun and van de Geer, 1986). Between 25,000 and 13,000B.P. there are relatively fewer small species, but variations do occur betweensites. Overall the pattern reflects the presence of owls, the most likelycontributor of small fauna to these sites (Garvey 1999), and their preferredhabitat, wooded landscapes.
Both wallaby and wombat are important components of late Pleisto-cene economic base in all the southwest sites examined. Their distributionthrough time and among sites, however, does vary. Tables VIII and IXshow the NISP, MNI, and mean weight for three sites. For Bone andNunamira Caves most discard of heavier bone pieces occurs after 18,000B.P. The relatively higher number of fragments between 25,000 and 30,000B.P. reflects more intense burning and increased fragmentation. At WarreenCave the highest number and average weights occur between 19,000 and24,000 B.P. Although the difference in altitude between Warreen Cave andthe other sites is only ca. 200 m, quite different chronological patterns ofwallaby and wombat exploitation occur.
Outside the region, especially at Parmerpar Meethaner and ORS 7,very few bones of wallaby have been identified. Preservation is not as goodat Parmerpar Meethaner, although many burnt bones have been identified,
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 387

388 Cosgrove

Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 389

with almost no wombat or wallaby teeth recorded. There appears to be asubstantial geographical and temporal difference among regions in resourcedistribution and acquisition.
Butchery Pattern
It has been observed on a number of occasions that there is an under-representation of specific body portions for Bennett's wallaby, reflectingdifferent aspects of the processing of skeletal elements (Cosgrove et al.1990; Cosgrove, 1995b; Cosgrove and Allen, 2000). Figure 8 displays theNISP and weight of Macropus rufogriseus body portions from a combineddatabase of six sites. Only complete elements, proximal, distal ends, andvertebrae centrum were used in the analysis. The results indicate highnumber and weight of lower limb, foot, and forequarter bones with lowrepresentation of other body portions. The patterns are relatively similarbetween post- and pre-LGM periods. This supports previous findings fromthe analysis of faunal remains at individual sites that the hindquarters arethe most common body part returned to the sites. The most commonelements are the tibia, femur, metatarsals 4/5, phalanges, humerus, radius,ulna, and ribs, while the other elements are underrepresented. The lowrepresentation of axial and girdle parts, particularly the vertebrae, parallelsprevious findings where it was suggested the pattern reflected crushing andgrinding to extract within-bone nutrients (Cosgrove, 1995b).
It is tempting to compare these results with those of O'Connell andMarshall (1989, p. 401), who found distinct macropod body partitioningamong the Alyawara. Their study indicated that the lumbar and sacralportions were the most highly ranked, followed by forequarters and hind-limbs. The lowest ranked were the head/cervical and feet, respectively.They suggested that these along with the tail were the most likely potionsto be left in the field rather than taken back to base camp. Two of thehighest ranked portions are found in the combined data, although thehighest ranked, lumbar and sacral portions, are not strongly represented.Feet, on the other hand, are well represented. At face value the combineddata reflect bones mostly discarded at field butchery stations, althoughmuch caution is needed in the interpretation.
In particular, linking ethnographic observation and archaeological datahas been a confounding problem long associated with the inappropriateuse of different temporal scales. Solutions for linking the two have beenelusive because the mechanisms for doing so are theoretically underdevel-oped (Bailey, 1983; Fletcher, 1989; Wobst, 1990). Thus, the results shouldbe tempered in light of the large time periods amalgamated in the present
390 Cosgrove

Fig. 8. Pre- and post-LGM NISP and weight (g) for Macropus rufogriseus body portions.

analysis. They are general trends that represent an accumulation of boneprocessing episodes over long time spans, produced by an amalgam ofbutchering behaviors of unknown duration in an array of sites. It is thereforeunlikely that we are seeing individual butchering events.
In a practical sense meaningful archaeological comparisons can onlybe attempted when a full understanding of the differential element fragmen-tation is made. For example, strong, dense metatarsal proximal and distalends are more likely to survive and be identified than less dense crushedand processed vertebrae. Examination of these important taphonomic vari-ables is currently under way (Cosgrove and Allen, 2000).
MODELS OF LAND USE
The variability of the archaeological record of late Pleistocene Tasma-nia has often been explained in frameworks of global climatic change(Bowdler, 1984; Jones, 1990; Cosgrove, 1995a; Porch and Allen, 1995; Hol-daway and Porch, 1995). While these fluctuations are extremely importantin understanding larger scale alterations to environmental structure, it isat the local and regional level that changes become important for humansoperating at yearly or seasonal scales. Unfortunately, many of the sites donot allow any finer time resolution than a few thousand years (Holdawayand Porch, 1995; Cosgrove and Allen, 1995, p. 23). Although this is thecase, attempts have been made to understand the archaeological variabilityacross regional Tasmania during the late Pleistocene period. It is clear thatthere are strong contrasts in the patterning of archaeological deposits foundin late Pleistocene southwestern caves and sites outside this zone (Bowdler,1984; Cosgrove, 1995a; Sim, 1998).
Initial suggestions by Kiernan et al (1983) proposed that there waswidespread steppe grassland in southwest Tasmania that supported largenumbers of macropods and that people had targeted these in a way similarto Northern Hemisphere reindeer hunters. If this was the case, then weneeded to explain why the high densities of bony remains and stone artifactswere not found in sites elsewhere, particularly in the equally well grassedareas to the north and east.
Rather than focusing on environmental changes at the large scale,investigations were made into the contemporary ecological requirementsof both plants and animals at the local scale. Studies showed that it waslikely that Poa spp. grasses would only grow on the small restricted alluvialand limestone soils during the late Pleistocene (Kirkpatrick, 1986; Kirkpat-rick and Duncan, 1987). Because of the widespread distribution of siliceoussoils in southwest Tasmania, areas of shrub and buttongrass sedge, which
392 Cosgrove

grow in wet, infertile areas, would have also been extensive. No grassesgrow in these conditions today, and where they do, they only colonize thebest soils and well-drained limestone outcrops as refugia. Therefore, duringthe late Pleistocene the southwest vegetation structure was probably charac-terized by limited fertile grassland patches surrounded by extensive lowshrubs and heath vegetation growing on siliceous soils. These 'coarse-grained' distributions (Wiens, 1976,1977,1985) were attractive habitats forthe Bennett's wallaby to gather (Cosgrove et al, 1990).
Bennett's wallaby are intermediate feeders of browse and graze, whilestudies have shown that unlike their much larger, gregarious relatives,Macropus giganteus, they are solitary. Where patches of grass occur theycongregate in large numbers, this being their principal food. They are alsovery sedentary, with a small home range of 15-20 hectares, and move theircenters of feeding activity less than 30 m every 2-3 years (Johnson, 1987,p. 131). These animals would have been tethered to the patches in muchthe same manner as marsupial 'lawns' are used today (Cosgrove et al., 1990).This provided hunters with a predictable macropod resource correlated withfertile soils, limestone geology, and cave sites. This patchy ecology had arelatively more reliable and stable moisture supply than the east.
The east was drought-prone with unpredictable moisture input andscattered resources. A reduction in vegetation would have caused soil desta-bilization and erosion. Large dune fields and lunettes systems attest to thisperiod of low precipitation and evaporation (Bowden, 1983). Thus theecological data suggest that western and eastern Tasmanian ecosystemsdisplayed wide variability. This dictated to a large extent the structure anddistribution of human prey animals and plant resources prior to, during,and subsequent to the Last Glacial Maximum. It has been argued that thisis one reason for the extremely rich archaeological record in the southwestcaves. A model was put forward to illustrate the differences between theselimestone cave deposits and those elsewhere (Cosgrove et al, 1990). Withinthis ecological structure it was argued that a range of social, economic, andtechnological relationships were played out.
The model only represents conditions at any one point in time and itis not intended to represent the ecological structure of the entire latePleistocene. The palaeoecology was unlikely to be stable and probablyvaried at small and medium scales over the long term. However, it is usefulbecause it allows the archaeological record to be investigated within anecological framework. Figure 9 is a modified version of the original modelpublished by Cosgrove et al. (1990). The dynamics of the southeast couldbe expanded to reflect the palaeoecology of most of eastern Tasmania.This model is intended to highlight the possible human resource relation-ships and links with zones outside the southwest that were not depicted
393Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania

394 Cosgrove
Fig. 9. Palaeoecological model indicating the possible human-landscape-resource relation-ships during the Tasmanian late Pleistocene. Based on Stanner (1965) and Peterson andLong (1986).
previously. It is based on ethnographically observed Aboriginal land usepatterns by Stanner (1965) and Peterson and Long (1986).
The model predicts that where resources were clumped and predictablehuman groups could be expected to move between them within their estateranges. Here wallabies were captured and processed at nearby cave sites.When the harvest from hunting diminished, people moved onto the nextpatch to secure necessary animal and plant products. Thus, people movedfrom patch to patch hunting ecologically tethered wallabies, possibly usinga mixture of 'encounter' and 'intercept' subsistence strategies (Cosgroveand Allen, 2000). These movements were probably not continuous andperhaps involved seasonal contact with the coast and areas outside thesouthwest. Evidence for this contact, however, is yet to be identified.
Where resources were dispersed relative to human estate ranges, move-ments between them could be expected to be wider and more frequent.

People in the drier east were probably more mobile and moved to resourceswhich were relatively more scattered. In this way the model predicts an'encounter' subsistence strategy in the east (Cosgrove et al, 1990, p. 74).Although there are pitfalls in using the ethnographic record in this way(Wobst, 1978), it is heuristic for understanding possible human-landscapeinteraction during the late Pleistocene in Tasmania.
Although criticisms have been leveled at the model (Thomas, 1993;Holdaway and Porch, 1995; Dunnett 1993), no substantive arguments havebeen mounted to refute it. At present there is no evidence of coastalconnections between the caves like that identified at Mannalargenna Cave(Brown, 1993; Sim, 1998), although this is not to suggest that there werenone. Some evidence in the form of coastal silcrete has been found atNanwoon Cave in the Florentine River valley (Cosgrove, 1996, p. 51). Noris there sufficient data at present to understand how the eastern and westernzones were socially or economically articulated. It is clear, however, thatthe humans inhabiting the southwest abandoned an economic strategy thathad been highly successful for at least 20,000 years at the end of thelate Pleistocene.
CONCLUSION
The late Pleistocene archaeology of Tasmania provides an opportunityto investigate human responses to a period of intense global change in theSouthern Hemisphere. The evidence shows that people entered Tasmaniaat least 35,000 radiocarbon years ago and occupied the periglacial uplandsand the Bass Plain. Inland they were far from the coast, hunting a rangeof terrestrial animals using a technology sufficiently sophisticated to sustainthis economy for 20,000 years. It is not known what role plants or marineresources played in the overall subsistence base since there is no evidenceof these in the cave sites. Presumably some plants were used, although inthis sub-Antarctic environment these may have been limited. This wouldhave had consequences for the availability of carbohydrates. In this casethe wallaby probably played an important role providing a marrow substi-tute for carbohydrates, which would be one reason for the high densitiesof marrow-bearing long bones in the sites (Cosgrove, 1995b, p. 118). Theymay be considered the terrestrial LGM equivalent of low-key dependableshellfish resources of the Holocene coasts (Meehan, 1984). It is clear thatalthough there are many features within the southwest cave sites that linkthem, like Darwin glass, thumbnail scrapers, bone points, hand stencil art,and raw materials, equally there is much variability within and betweenregions.
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 395

Understanding this variability is a major goal for Tasmanian archaeo-logical research despite its having been stifled in recent years for politicalends (Murray and Allen, 1995). Future archaeological work should comeabout through genuine collaborative efforts between professional archaeol-ogists, heritage managers, and indigenous Tasmanian communities seekingto comprehend this fascinating record of human colonization and set-tlement.
ACKNOWLEDGMENTS
The initial fieldwork upon which this paper is partly based was carriedout while I was a doctoral student at La Trobe University and in receiptof a Commonwealth Postgraduate Research Award from the AustralianGovernment. The Australian Institute of Aboriginal and Torres Strait Is-lander Studies and La Trobe University funded the work. I thank mycolleague, Prof. Jim Allen, for his valuable support through the good andbad years and Prof. Tim Murray for encouragement to take a differentview of the archaeological record. The Southern Forests ArchaeologicalResearch Project and The Management of Archaeological Resources inForested Areas Project were funded by The Australian Research Counciland the Tasmanian Forest Research Council Inc., respectively. The fieldcrews and laboratory volunteers, too many to mention individually, helpedin numerous ways and are thanked for their untiring energy and support.I particularly thank Darrell West, Eric Stadler, and Barry Blain for theircontinued support of the Tasmanian research.
REFERENCES
Allen, H. (1979). Left out in the cold: why the Tasmanians stopped eating fish. Artefact 4:1-10.Allen, J. (1996). Report of the Southern Forests Archaeological Project, Vol. 1, School of
Archaeology, La Trobe University, Melbourne.Allen, J., and Cosgrove, R. (1996a). Background history of the Southern Forests Archaeologi-
cal Project. In Allen, J. (ed.), Report of the Southern Forests Archaeological Project,Volume 1, Site descriptions, stratigraphies and Chronologies, La Trobe University, Mel-bourne, pp. 3-19.
Allen, J., and Cosgrove, R. (1996b). Fieldwork methods: excavations, surveys and laboratoryprocedures. In Allen, J. (ed.), Report of the Southern Forests Archaeological Project,Volume 1, Site descriptions, stratigraphies and chronologies, La Trobe University, Mel-bourne, pp. 31-30.
Allen, J., and Holdaway, S. (1995). The question of the contamination of older radiocarbondeterminations in Australia. Antiquity 69:101-112.
Allen, J., Cosgrove, R., and Brown, S. (1988). New archaeological data from the southernforests region, Tasmania: a preliminary statement. Australian Archaeology 27:75-88.
Cosgrove396

Bailey, G. (1983). Concepts in Quaternary prehistory. Annual Review of Anthropology12:165-192.
Bowden, A. R. (1983). Relict terrestrial dunes: legacies of a former climate in coastal northeast-ern Tasmania. Zeitschrift fur Geomorphologie, Supplement 45:153-174.
Bowdler, S. (1984). Hunter Hill, Hunter Island, Terra Australis 8, Department of Prehistory,Research School of Pacific Studies, Australian National University, Canberra, Australia.
Bowman, D. M. J. S., and Jackson, W. D. (1981). Vegetation succession in southwest Tasmania.Search 12(10):358-362.
Boyle, K. V. (1990). Upper Palaeolithic faunas from South-West France: a zooarchaeologicalperspective. BAR International Series 557.
Brown, S. (1993). Mannalargenna Cave: a Pleistocene site in Bass Strait. In Smith, M., Spriggs,M., and Fankhauser, B. (eds.), Sahul in review, Department of Prehistory, ResearchSchool of Pacific Studies, Australian National University, Canberra, pp. 258-271.
Burke, A., and Pike-Tay, A. (1997). Reconstructing "L'Age Du Renne." In Jackson, J. J.,and Thacker, P. T. (eds.), Caribou and reindeer hunters of the Northern Hemisphere,Avebury, pp. 69-79.
Cann, J., Belperio, A. P., Gostin, V. A., and Murray-Wallace, C. V. (1988). Sea-level history,45,000 to 30,000 B.P., inferred from benthic foraminifera, Gulf St. Vincent, South Austra-lia. Quaternary Research 29:153-175.
Cann, J., Belperio, A. P., Gostin, V. A., and Rice, R. L. (1993). Contemporary benthicforaminifera in Gulf St. Vincent, South Australia and a refined sea-level history. AustralianJournal of Earth Sciences 40:197-211.
Chappell, J. (1994). Upper Quaternary sea levels, coral terraces, oxygen isotopes and deep-sea temperatures. Journal of Geography 103(7):828-840.
Chappell, J., Omura, A., Esat, T., McCulloch, M., Pandolfi, J., Ota, Y., and Pillans, B. (1996).Reconciliation of late Quaternary sea levels derived from coral terraces at Huon Peninsulawith deep sea oxygen isotope records. Earth and Planetary Letters 141:227-236.
Cockbill, J. (1999). Bones of contention, Unpublished B.A. Honours Thesis, Department ofArchaeology, La Trobe University, Melbourne.
Colhoun, E. A. (1978). The late Quaternary environment of Tasmania as a backdrop to man'soccupance. Records of the Queen Victoria Museum 61:1-12.
Colhoun, E. A. (1985). Radiocarbon dates for Tasmania, 1956-1984. Papers and Proceedingsof the Royal Society of Tasmania 119:39-54.
Colhoun, E., and van de Geer, G. (1986). Holocene to middle last glaciation vegetation historyat Tullabardine Dam, western Tasmania. Proceedings of the Royal Society of LondonB 229:177-207.
Colhoun, E., van de Geer, G., and Mook, W. G. (1982). Stratigraphy, pollen analysis andpalaeoclimatic interpretation of Pulbeena Swamp, northwestern Tasmania. QuaternaryResearch 18:108-126.
Colhoun, E., van de Geer, G., Fitzsimons, S. J., and Heusser, L. E. (1994). Terrestrial andmarine records of the last glaciation from western Tasmania: do they agree? QuaternaryScience Review 13:293-300.
Colhoun, E. A., Hannan, D., and Kiernan, K. (1996). Late Wisconsin glaciation of Tasmania.Papers and Proceedings of the Royal Society of Tasmania 130:33-45.
Corbett, K. D. (1980). A record of Aboriginal implement sites in the Queenstown area,Tasmania. Papers and Proceedings of the Royal Society of Tasmania 114:35-39.
Cosgrove, R. (1985). King River power development environmental plan: archaeological recon-naissance survey, A report to the Hydro-Electric Commission, Tasmania.
Cosgrove, R. (1989). Thirty thousand years of human colonisation in Tasmania: new Pleisto-cene dates. Science 243:1703-1705.
Cosgrove, R. (1995a). Late Pleistocene behavioural variation and time trends: the case fromTasmania. Archaeology in Oceania 21:19-36.
Cosgrove, R. (1995b). The illusion of riches: scale, resolution and explanation in TasmanianPleistocene human behaviour, Tempus Reparatum, BAR International Series 608, Oxford.
Cosgrove, R. (1996). Nunamira Cave. In Allen, J. (ed.), Report of the Southern Forests
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 397

Archaeological Project, Volume 1, Site descriptions, stratigraphies and chronologies, LaTrobe University, Melbourne, pp. 43-68.
Cosgrove, R. (1997). The Tasmanian Aborigines through the Ice Age. In Webb, E. (ed),Windows on meteorology: Australian perspective, CSIRO, Melbourne, pp. 46-58.
Cosgrove, R., and Allen, J. (1996). Research strategies and theoretical perspectives. In Allen,J. (ed.), Report of the Southern Forests Archaeological Project, Volume 1, Site descriptions,stratigraphies and chronologies, La Trobe University, Melbourne, pp. 21-30.
Cosgrove, R., and Allen, J. (2000). Species targeting as a hunting strategy in the late Pleistocene:evidence from southwest Tasmania, the restaurant at the end of the Pleistocene universe.In Ambrose, W. (ed.), Rhys Jones' festschrift volume of essays, ANH publications, Depart-ment of Archaeology and Natural History Research School of Pacific and Asian Studies,ANU, Canberra (in press).
Cosgrove, R., and Murray, T. (1993). The management of archaeological resources in forestedareas, Tasmanian Forest Research Council Report, 1992-1993.
Cosgrove, R., Allen, J., and Marshall, B. (1990). Palaeoecology and Pleistocene human occupa-tion in south central Tasmania. Antiquity 64(242) :59-78.
Cosgrove, R., Allen, J., and Marshall, B. (1994). Late Pleistocene human occupation inTasmania: a reply to Thomas. Australian Archaeology 38:28-35.
Dansgaard, W., White, J. W. C., and Johnsen, S. J. (1989). The abrupt termination of theYounger Dryas climate event. Nature 339:532-533.
David, B., Roberts, R., Tuniz, C., Jones, R., and Head, J. (1997). New optical and radiocarbondates from Ngarrabullgan Cave, a Pleistocene archaeological site in Australia: implicationsfor the comparability of time clocks and for the human colonisation of Australia. Antiq-uity 71:183-188.
D'Costa, D. M., Grindrod, J., and Ogden, R. (1993). Preliminary environmental reconstructionsfrom late Quaternary pollen and mollusc assemblages at Egg Lagoon, King Island, BassStrait. Australian Journal of Ecology 18:351-366.
Driessen, M. M. (1988). Age structure and reproduction of Bennett's and Rufous wallabiesin Tasmania. Unpublished technical report to Australian National Parks and WildlifeService, Canberra.
Dunnett, G. (1993). Diving for dinner: some implications from Holocene middens for therole of coasts in the late Pleistocene of Tasmania. In Smith, M. A., Spriggs, M., andFankhauser, B. (eds.), Sahul in review, Department of Prehistory, Research School ofPacific Studies, Australian National University, Canberra, pp. 247-257.
Feblot-Augustins, J. (1993). Mobility strategies in the late middle Palaeolithic of central Europeand Western Europe: elements of stability and variability. Journal of AnthropologicalArchaeology 12(3):211-265.
Fletcher, R. (1989). Social theory and archaeology: diversity, paradox and potential. Man-kind 19(l):65-75.
Fitzsimons, S. J., and Colhoun, E. A. (1989). Pleistocene clastic dykes in the King Valley,western Tasmania. Australian Journal of Earth Sciences 36:351-363.
Fitzsimons, S. J., and Colhoun, E. A. (1991). Pleistocene glaciation of the King Valley, westernTasmania, Australia. Quaternary Research 36:135-156.
Freslov, J. (1993). The role of open sites in the investigation of Pleistocene phenomena inthe inland southwest of Tasmania. In Smith, M. A., Spriggs, M., and Fankhauser, B.(eds.), Sahul in review, Department of Prehistory, Research School of Pacific Studies,Australian National University, Canberra, pp. 233-239.
Fudali, R. F., and Ford, R. J. (1979). Darwin glass and Darwin Crater: a progress report.Meteoritics 14:283-296.
Fullagar, R. (1986). Use-wear and residues on stone tools: functional analysis and its applicationto two southeastern Australian archaeological assemblages. Unpublished Ph.D. Thesis,Department of Archaeology, La Trobe University, Melbourne.
Fullagar, R. L. K., and Furby, J. H. (1997). Pleistocene seed-grinding implements from thearid zone. Antiquity 71:300-307.
Furby, J. H. (1995). Megafauna under the microscope: archaeology and palaeoenvironment atCuddle Springs. Ph.D. Thesis, University of New South Wales, Sydney.
398 Cosgrove

Gale, K. (1994). When size really does count: an occupational history and comparative analysisof Stone Cave and Bone Cave from southwest Tasmania. Unpublished B. A. (Hons) Thesis,School of Archaeology, La Trobe University, Melbourne.
Gamble, C. (1993). Exchange, foraging and local hominid networks. In Scarre, C., and Healy,F. (eds.), Trade and exchange in prehistoric Europe, Oxbow Books, Oxford, pp. 35-44.
Garvey, J. (1999). Taphonomic analysis of the small vertebrate fauna from the archaeologicalsite Derwent River Shelter (DRS 7), Tasmania. Unpublished B.A. Honours Thesis, Depart-ments of Zoology and Archaeology, La Trobe University, Melbourne.
Gibson, N., Keirnan, K. W., and Macphail, M. K. (1987). A fossil bolster plant from the KingRiver, Tasmania. Papers and Proceedings of the Royal Society of Tasmania 121:35-42.
Guiler, E. R. (1973). Some thoughts on man and mammals in the Central Plateau. In Banks,M. (ed.), The Lake Country, Royal Society of Tasmania, pp. 125-133.
Harris, S., Ranson, D., and Brown, S. (1988). Maxwell River Archaeological Survey 1986.Australian Archaeology 27:89-97.
Hartzell, L. (1993). Preliminary report on phase 3 archaeological investigations in the SurreyHills, northwest Tasmania: test excavations at the Brecciated Chert Quarry site TASI5354, (SQ N9/W13.5). Report to Department of Archaeology, La Trobe University,and Department of Parks, Wildlife and Heritage, Tasmanian Forest Research Council,Tasmanian Aboriginal Lands Council and the Tasmanian Forestry Commission, Hobart.
Hocking, G. J., and Guiler, E. R. (1983). The mammals of the lower Gordon River regionsouth-west Tasmania. Australian Wildlife Research 10:1-23.
Holdaway, S., and Cosgrove, R. (1997). The archaeological attributes of behaviour: differenceor variability. Endeavour 21(2):66-71.
Holdaway, S., and Porch, N. (1995). Cyclical patterns in the Pleistocene human occupationof southwest Tasmania. Archaeology in Oceania 30:74-82.
Holdaway, S., and Porch, N. (1996). Dates as data: an alternative approach to the constructionof chronologies for Pleistocene sites in southwest Tasmania. In Allen, J. (ed.), Report ofthe Southern Forests Archaeological Project: Volume 1, School of Archaeology, La TrobeUniversity, Melbourne.
Hope, G. S. (1978). The late Pleistocene and Holocene vegetational history of Hunter Island,north-western Tasmania. Australian Journal of Botany 26:493-514.
Jackson, W. (1978). 'Ecological Drift': an argument against the continued practise of hazardreduction burning. In Gee, H., and Fenton, J. (eds.), The southwest book: a Tasmanianwilderness, Australian Conservation Foundation, Melbourne, pp. 98-101.
Jackson, W. (1999). Palaeohistory of vegetation change: the last 2 million years. In Reid,J. B. (ed.), Vegetation of Tasmania, Australian Biological Resources Study, Canberra,pp. 64-88.
Jones, R. (1971a). Rocky Cape and the problem of the Tasmanians. Unpublished Ph.D. Thesis,University of Sydney.
Jones, R. (1971b). The demography of hunters and farmers in Tasmania. In Mulvaney, D. J.,and Golson, J. (eds.), Aboriginal man and environment in Australia. Australian NationalUniversity Press, Canberra, pp. 271-287.
Jones, R. (1977a). The Tasmanian paradox. In Wright, R. V. S. (ed.), Stone tools as culturalmarkers: change, evolution and complexity, Australian Institute of Aboriginal Studies,Canberra, pp. 189-204.
Jones, R. (1977b). Man as an element of a continental fauna: the case of the sundering ofthe Bassian Bridge. In Allen, J., Golson, J., and Jones, R. (eds.), Sunda and Sahul:prehistoric studies in Southeast Asia, Melanesia and Australia, Australian National Univer-sity, Canberra, pp. 317-386.
Jones, R. (1978). Why did the Tasmanians stop eating fish? In Gould, R. A. (ed.). Explorationsin ethnoarchaeology, University of New Mexico Press, Albuquerque, pp. 11-47.
Jones, R. (1984). Hunters and history: a case study from western Tasmania. In Schrire, C.(ed.), Past and present in hunter gatherer studies, Academic Press, London, pp. 27-65.
Jones, R. (1990). From Kukadu to Kutikina: the southern continent at 18,000 years ago. InGamble, C., and Soffer, O. (eds.), The world at 18,000 B.P. Volume 2: Low latitudes,Unwin Hyman, London, pp. 264-295.
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 399

Jones, R. (1992). Philosophical time travelers. Antiquity 66:744-757.Jones, R. (1995). Tasmanian archaeology: establishing the sequences. Annual Review of An-
thropology 24:423-446.Jones, R., and Allen, J. (1984). Archeological investigations in the Andrew River valley,
Acheron River valley and at Precipitous Bluff—southwest Tasmania—February 1984.Australian Archaeology 19:86-101.
Jones, R., Cosgrove, R., Allen, J., Cane, S., and Kiernan, K. (1988). An archaeological recon-naissance of karst caves within the Southern Forests region of Tasmania, September1987. Australian Archaeology 26:1-23.
Jordan, G. L., Carpenter, R. J., and Hill, R. S. (1991). Late Pleistocene vegetation and climatenear Melaleuca Inlet, southwestern Tasmania. Australian Journal of Botany 39:315-333.
Kiernan, K., Jones R., and Ranson, D. (1983). New evidence from Fraser Cave for glacialage man in southwest Tasmania. Nature 301:28-32.
Kirkpatrick, J. (1986). Tasmanian alpine biogeography and ecology and interpretations of thepast. In Barlow, B. A. (ed.), Flora and fauna of alpine Australasia. CSIRO, Melbourne,pp. 229-242.
Kirkpatrick, J., and Dickson, K. J. M. (1984). The impact of fire on Tasmanian alpine vegetationand soils. Australian Journal of Botany 32:613-629.
Kirkpatrick, J., and Duncan, F. (1987). Tasmanian high altitude grassy vegetation: its distribu-tion, community composition and conservation status. Australian Journal of Ecology12:73-86.
Kozlowski, J. K. (1991). Raw material procurement in the upper Palaeolithic of central Europe.In Monet-White, A., and Holen, S. (eds.), Raw material economies among prehistorichunter-gatherers, University of Kansas, Lawrence, Kansas, pp. 187-196.
Loy, T., Jones, R., Nelson, D. E., Meehan, B., Vogel, J., Southern, J., and Cosgrove, R. (1990).Accelerator radiocarbon dating of human blood proteins in pigments from late Pleistoceneart sites in Australia. Antiquity 64(242):110-116.
Lyman, L. (1994). Vertebrate taphonomy. Cambridge University Press, Cambridge.Marsden-Smedely, J. B. (1998). Changes in southwestern Tasmanian fire regimes since the
early 1800s. Papers and Proceedings of the Royal Society of Tasmania 132:15-29.Marshall, B. (1992). Late Pleistocene human exploitation of the platypus in southern Tasmania.
In Augee, M. L. (ed.), Platypus and echidnas, Royal Zoological Society of NSW, Sydney,pp. 268-276.
Marshall, B., and Cosgrove, R. (1990). Tasmanian Devil (Sarcophilus harrisii) scat bone:signature criteria and archaeological implications. Archaeology in Oceania 25:102-113.
McNiven, I. (1994). Technological organisation and settlement in southwest Tasmania afterthe glacial maximum. Antiquity 68:75-82.
McWilliams, R., Allen, J., Cosgrove, R., and Holdaway, S. (1999). Welcome to the SouthernForests Archaeological Project Data Base, Volume 3 of the SFAP. In Cosgrove, R. (ed.),Hands of time [CD-ROM]. School of Archaeological and Historical Studies, La TrobeUniversity, Melbourne.
Meehan, B. (1984). Shell bed to shell midden. Australian Institute of Aboriginal Studies. Can-berra.
Murray, (1978). Life during the late Pleistocene. In Gee, H., and Fenton, J. (eds.), The southwestbook: a Tasmanian wilderness, Australian Conservation Foundation, Melbourne, pp.98-101.
Murray, P. F., Goede, A., and Bada, J. L. (1980). Pleistocene human occupation at BeginnersLuck Cave, Florentine Valley, Tasmania. Archaeology and Physical Anthropology inOceania 15:142-152.
Murray, T., and Allen, J. (1995). The forced repatriation of cultural properties to Tasmania,Antiquity 69:871-874.
Northwood, C. (1990). A taphonomic analysis of Sarcophilus harrisii in Australian archaeologi-cal sites. Unpublished B.A. Honours Thesis.
O'Connell, J., and Allen, J. (1998). When did humans first arrive in greater Australia andwhy is it important to know? Evolutionary Anthropology 6(4):132-146.
Cosgrove400

O'Connell, J., and Marshall, B. (1989). Analysis of kangaroo body part transport among theAlyawara of central Australia. Journal of Archaeological Science 16:393-405.
O'Connor, S. (1990). Thirty thousand years in the Kimberley. Unpublished Ph.D. Thesis,Centre for Prehistory, University of Western Australia.
Pardoe, C. (1986). Population genetics and population size in prehistoric Tasmania. AustralianArchaeology 22:1-6.
Pardoe, C. (1988). Cemetery as symbol: the distribution of prehistoric Aboriginal burialgrounds in south-eastern Australia. Archaeology in Oceania 23:1-16.
Pardoe, C. (1991). Isolation and evolution in Tasmania. Current Anthropology 32(1):1-21.Peterson, N., and Long, J. (1986). Australian Territorial Organisation, University of Sydney.Porch, N., and Allen, J. (1995). Tasmania: archaeological and palaeo-ecological perspectives.
Antiquity 69(265):714-732.Rensink, E., Kolen, J., and Spieksma, A. (1991). Patterns of raw material distribution in the
upper Pleistocene of northwest and central Europe. In Monet-White, A., and Holen, S.(eds.), Raw material economies among prehistoric hunter-gatherers, University of Kansas,Lawrence, Kansas, pp. 141-160.
Roberts, R. G., Jones, R., and Smith, M. (1990). Thermoluminescence dating of a 50,000-year old human occupation site in northern Australia. Nature 345:153-156.
Roberts, R. G., Jones, R., Spooner, N. A., Head, M. J., and Murray, A. S. (1994). The humancolonisation of Australia: optical dates of 53,000 and 60,000 years bracket human arrivalat Deaf Adder Gorge Northern Territory. Quaternary Science Review 13:575-583.
Sheppard, P. J. (1997). Characterisation of cherts from sites in southwest Tasmania. Archaeol-ogy in Oceania 32:47-53.
Sigleo, W. R. (1978). A study of late Quaternary environment and man from four sites insoutheastern Tasmania. Unpublished Ph.D. thesis, Department of Geography, Universityof Tasmania, Hobart.
Sigleo, W. R., and Colhoun, E. A. (1982). Terrestrial dunes, man and the late Quaternaryenvironment in southern Tasmania. Palaeogeography, Palaeoclimatology, Palaeoecol-ogy 39:87-121.
Sim, R. (1998). The archaeology of isolation? Prehistoric occupation in the Furneaux Groupof Islands, Bass Strait, Tasmania. Unpublished Ph.D. Thesis, Department of Archaeologyand Natural History, Australian National University, Canberra.
Sim, R., and Thorne, A. (1990). Pleistocene human remains from King Island, southeasternAustralia. Australian Archaeology 31:44-51.
Stanner, W. E. H. (1965). Aboriginal territorial organisation: estate, range, domain and regime.Oceania 36:1-26.
Stern, N., and Marshall, B. (1993). Excavations at Mackintoah 90/1: a discussion of stratigraphy,chronology and site formation. Archaeology in Oceania 28:183-192.
Stiner, M. (1994). Honor among thieves: a zooarchaeological study of Neanderthal ecology,Princeton University Press, Princeton, New Jersey.
Sutherland, L. (1972). The classification, distribution, analysis and sources of material in flakedstone implements of Tasmanian Aborigines. Records of the Queen Victoria Museum42:1-46.
Taylor, E. B. (1893). On the Tasmanians as representatives of Palaeolithic man. Journal ofthe Anthropological Institute 23:141-152.
Taylor, K. C., Lamorey, G. W., Doyle, G. A., Alley, R. B., Grootes, P. M., Mayewski, P. A.,White, J. W. C., and Barlow, L. K. (1993). The flickering switch of late Pleistocene climatechange. Nature 361:432-436.
Thacker, P. (1997). The significance of Rangifer as a human prey species during the centralEuropean upper Palaeolithic. In Jackson, J. J., and Thacker, P. T. (eds.), Caribou andreindeer hunters of the Northern Hemisphere, Avebury, pp. 82-104.
Thomas, I. (1993). Late Pleistocene environments and aboriginal settlement patterns in Tasma-nia. Australian Archaeology 36:1-11.
Thomas, I. (1995). Where have all the forests gone? New pollen evidence from MelaleucaInlet in Southwestern Tasmania. In Dixon, G., and Aitken, D. (eds.), Institute of Australian
Forty-Two Degrees South: The Archaeology of Late Pleistocene Tasmania 401

Geographers: Conference Proceedings, 1993, Department of Geography and Environmen-tal Science, Monash University, Melbourne, pp. 295-301.
Thomas, N. (1981). Social theory, ecology and epistemology: theoretical issues in Australianprehistory. Mankind 13(2):165-177.
Thorne, A., and Sim, R. (1994). The gracile male skeleton from late Pleistocene King Island,Australia. Australian Archaeology 38:8-10.
Webb, C., and Allen, J. (1990). A functional analysis of Pleistocene bone tools from two sitesin south west Tasmania. Archaeology in Oceania 25:75-78.
Webb, S. (1988). Human skeletal remains from the Nanwoon Cave, southwestern Tasmania.Report to the Tasmanian Aboriginal Community.
Webb, S. (1989). The Willandra Lakes hominids, Department of Prehistory, Research Schoolof Pacific Studies, Australian National University, Canberra.
White, J. W. C., Ciais, P., Figge, R. A., Kenny, R., and Markgraf, V. (1994). A high-resolutionrecord of atmospheric CO2 content from carbon isotopes in peat. Nature 367:153-156.
Wiens, J. A. (1976). Population responses to patchy environments. Annual Review of EcologicalSystems 7:81-120.
Wiens, J. A. (1977). On competition and variable environments. American Scientist 65:590-597.Wiens, J. A. (1985). Vertebrate responses to environmental patchiness in arid and semiarid
ecosystems. In Pickett, S. T. A., and White, P. S. (eds.), The ecology of natural disturbanceand patch dynamics, Academic Press, San Diego, pp. 169-193.
Wobst, H. M. (1990). Afterword minitime and megaspace in the Palaeolithic at 18 K andotherwise. In Soffer, O., and Gamble, C. (eds.), The world at 18,000 B.P. Volume 2, Lowlatitudes. Unwin Hyman, London, pp. 322-334.
Wobst, H. M. (1978). The archaeo-ethnology of hunter-gatherers and the tyranny of theethnographic record in archaeology. American Antiquity 43(2):303-309.
Zilhao, J., and d'Errico, F. (1999). The chronology and taphonomy of the earliest Aurignacianand its implications for the understanding of Neanderthal extinction. Journal of WorldPrehistory 13(1):1-40.
402 Cosgrove




















