
Weighted and Ergodic Theorems for Series of Differences of Convolutions

DevelopmentalBrain Research, 45 (1989) 303-308 303 Elsevier
BRD60299
Formation and growth of the cerebral convolutions. II. Cell death in the gyrus suprasylvius and
adjoining sulci in the cat*
I. Ferrer 1, M. Hernandez-Martf 2, E. Bernet 2 and M. Calopa 1 I Unidad de Neuropatologia, Departarnento Anatomfa Patol6gica, Hospital Principes de Espatia, Hospitalet de Llobregat and
2Departamento de Anatomla Patoldgica, C.S. La Fe, Valencia (Spain)
(Accepted 25 October 1988)
Key words: Cell death; Cerebral convolution; Cerebral cortex
Cell death, calculated by counting pyknotic nuclei to assess the number of dying cells, in the gyrus suprasylvius (GS-Syl) and adjoin- ing sulci sulcus lateralis (SL) and sulcus suprasylvius (SS-Syl) was studied in cats aged 5, 15, 25 days and 6 months. Three patterns of cell death were characterized: (1) an ascending gradient from the inner to the upper cortical layers; (2) a lateromedial gradient from the SS-Syl towards the SL; and (3) a predominance of cell death in the sulcal zones. These patterns are in accordance with the sequence of cortical neurogenesis, the lateromedial pattern of the whole formation and growth of the GS-Syl and adjoining sulci, and the differ- ences in the cortical thickness between the sulci and the gyral crown.
Naturally occurring cell death and neuron loss dur- ing development is a common phenomenon in sever- al regions of the nervous system 2'15'1s'19. In the cere-
bral cortex degenerating cells, characterized by their extremely pyknotic nuclei and virtual absence of cy-
toplasm, have been observed in hamsters and mice 9'2l'22, as well as in the olfactory cortex of the
rat 5. In the cat's cerebral cortex many of these cells
are neurons according to their morphology as re-
vealed with the Golgi method, ultrastructure and im- munocytochemical characteristics4'33; furthermore,
they form a heterogeneous group with different neu- ropeptide and neurotransmitter expression 1.4,34. Ex-
cessive production of cortical neurons during devel- opment has also been demonstrated by quantitative
studies showing a decrease of 25% layer VI neurons in the mouse during the first ten days of postnatal life ~1'12, and a decrease in the number of neurons of
about 16% in the cerebral cortex of the cat from day 15 to 25 (ref. 8). A temporospatial ascending gra-
dient of cell death has been observed in the lyssence- phalic brain of the hamster 9, but little is known about
the patterns of cell death in gyrencephalic species.
For this reason in the present study we have at-
tempted to examine the sequence and possible inter-
laminar and regional gradients of cell death during the formation and growth of the gyrus suprasylvius
and adjoining sulci in the cat. Cats aged 5, 15, 25 days
and 6 months were killed under phenobarbitone
anaesthesia. The brains were perfused with saline and immediately afterwards fixed by perfusion with
1% glutaraldehyde-2.5% paraformaldehyde (v/v) in
phosphate buffer. The brains were removed from the skull and samples of the left gyrus suprasylvius (GS-
Syl) were obtained for electron microscopical exami- nation while the rest of the brain was immersed in the same fixative for several weeks. Three animals of each age were used.
For electron microscopical study the samples were postfixed in 2% osmium tetroxide for 2 h, dehy-
* This study is dedicated to A. Ibafiez Cacho. Correspondence: I. Ferret, Unidad Neuropatologfa, Departamento Anatomfa Patol6gica, Hospital Principes de Espafia, 08907 Hos- pitalet del Llobregat, Spain.
0165-3806/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

304
drated in ethanol and proprileneoxide and embedded
in epon: semi-thin sections were stained with Tolui- dine blue and selected ultrathin sections with uranyl
acetate and lead citrate. Morphological studies on paraffin sections were focused on the GS-Syl and ad-
joining sulci, sulcus lateralis (SL) and sulcus supra-
sylvius (SS-Syl) of the right cerebral hemisphere,
contained between two perpendicular lines that fol-
lowed and extended from the course of the ascending
branch of the sulcus suprasylvius and the ascending branch of the sulcus suprasylvius anterior according to the nomenclature of Sanides and Hoffman 25. (La-
get ~4 named these latter sulci by postsylvian sulcus
and anterior ectosylvian sulcus, respectively). The
selected cortical region was embedded in paraffin
and sections 10/~m thick were obtained in the coronal
plane every 100 ~m by means of a sliding microtome. The sections were numbered consecutively and
stained with Cresyl violet. The 3 sections located halfway along the gyrus (n/2 +_ 1) were used for the
present quantitative study. A photomontage of the
complete gyrus and adjacent sulci, obtained at a magnification × 400, was constructed for each of the
sections. For all the photomontages a line was traced
at the midlevel of the cellular cerebral cortex. This line was later divided into 20 equal segments from the
depth of SL to the depth of SS-Syl, and a series of
lines radial to the cortical surface were subsequently drawn at these points following the curvature of the
cortical nerve cell columns8; these lines divided the
whole of the coronal section into fields labelled 1-20
from SS-Syl to SL (Fig, 1).
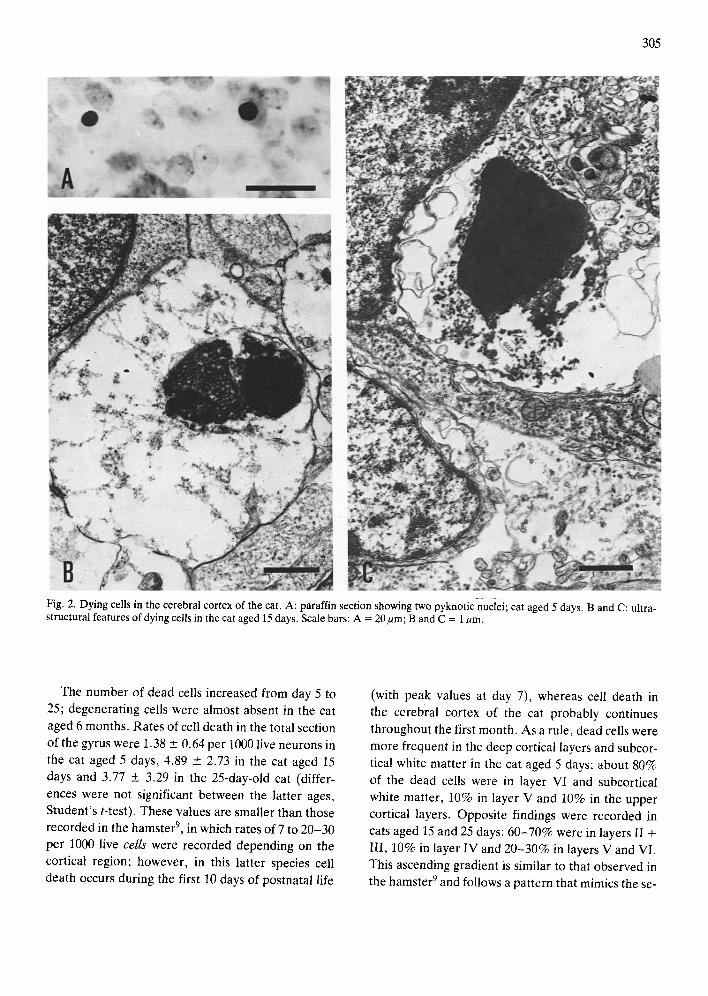
Dead cells were recognized in paraffin and semi- thin sections by their darkly and homogeneously stained, shrunken and sometimes fragmented nuclei, and their pale or absent cytoplasm 3'26'2729. Electron
microscopical examination of these cells confirmed
the degeneration of the nucleus and the virtual lack of organelles in the clear cytoplasm, characteristics which conform to similar findings currently used to identify dead cells in different regions of the nervous system 3'6'23'32 (Fig. 2). Because this method does not
permit us to discriminate the cell type, no attempt was made to recognize glial cells and neurons among
dead cells. The following parameters were measured at every
age: (1) number of dead cells in the total section of the gyrus and in each one of the 20 consecutive fields;
50AYS S.L.
~ ~ l 5 D Y
C 5mm
Fig. 1. A: lateral view of the brain of the cat aged 6 months. The vertical lines mark the limits of the cortical region, containing the gyrus suprasylvius (GS-Syl), sulcus lateralis (SL) and sulcus suprasylvius (SS-Syl), selected for quantitative studies. SE- SylA, sulcus suprasylvius anterior. B: division of the cerebral cortex into 20 consecutive fields, labelled 1-20, from SS-Syl to SL in the cat aged 5 days. C: camera lucida drawings of coronal sections of the brain of cats at different postnatal ages in which it may be observed the displacement of the gyral ridge that oc- curs during the growth of the SL. V: ventricle (see ref. 8 for fur- ther details).
(2) percentage of dead cells in the different cortical
layers in the total section of the gyrus. Since the number of neurons and dead cells varied
from one field to the next consecutive field, and from
one section to another, the quantity of cell death was expressed as the mean values +__ S.D. of dead cells per
1000 live neurons. On the other hand, since the upper cortical layers are not cleary defined in cats aged 5 days, mainly because significant neuronal migration is still in progress 17, layers I I - I I I were considered
conjointly. The methods and correcting factors to know the number of nerve cells have been described in the first part of this seriesS; here again the number of dead cells was corrected according to the average length of the nuclei at every age and the section thick-
ness.
305
Fig. 2. Dying cells in the cerebral cortex of the cat. A: paraffin section showing two pyknotic nuclei; cat aged 5 days. B and C: ultra- structural features of dying cells in the cat aged 15 days. Scale bars: A = 20gm; B and C = 1/~m.
The number of dead cells increased from day 5 to 25; degenerating cells were almost absent in the cat
aged 6 months. Rates of cell death in the total section of the gyrus were 1.38 _+ 0.64 per 1000 live neurons in the cat aged 5 days, 4.89 + 2.73 in the cat aged 15
days and 3.77 +_ 3.29 in the 25-day-old cat (differ- ences were not significant between the latter ages, Student's t-test). These values are smaller than those recorded in the hamster 9, in which rates of 7 to 20-30 per 1000 live cells were recorded depending on the cortical region; however, in this latter species cell death occurs during the first 10 days of postnatal life
(with peak values at day 7), whereas cell death in the cerebral cortex of the cat probably continues throughout the first month. As a rule, dead cells were
more frequent in the deep cortical layers and subcor- tical white matter in the cat aged 5 days: about 80%
of the dead cells were in layer VI and subcortical white matter, 10% in layer V and 10% in the upper
cortical layers. Opposite findings were recorded in cats aged 15 and 25 days: 60-70% were in layers II + III, 10% in layer IV and 20-30% in layers V and VI.
This ascending gradient is similar to that observed in the hamster 9 and follows a pattern that mimics the se-
3t)6
quence of neuronal generation and maturation in the cerebral cortex from the inner to the upper cortical
layers. The greater amount of cell death observed in the
inner cortical layers during the first days of postnatal life agrees with similar findings observed in the cat's cerebral cortex using other methods; thus, Valverde and Facal-Valverde 33 using combined Golgi and elec- tron microscopical methods observed degenerating neurons in the inner cortical layers and subcortical plate of the temporal cortex during the first 10 days of life; similarly, Chun et al. 4 described the presence of transitory populations of neurons (according to their ultrastructural characteristics and reactivity to MAP,_) in the cortical subplate; these neurons estab- lish ipsilateral and contralateral connections and are constituted by different subpopulations with distinct
expressions of somatostatin (SRIF), neuropeptide Y (NPY) and cholecystokinin (CCK); finally, Whale and Meyer 34 observed 3 subpopulations of NPY-con- raining neurons in the visual cortex of the cat; sub- population 2 and early members of subpopulation 3 are located in the inner cortical layers and white mat- ter and become eliminated by day 48. In contrast, knowledge is scarce about cell death in the upper cor- tical layers of the cat during development. Subpop- ulation 1 of NPY-containing neurons is distributed throughout the cortical thickness and is eliminated by day 12 (ref. 34). Nevertheless, naturally occurring cell death in the upper cortical layers is well docu- mented in hamsters and mice 9"2z'22 during the first
days of life, and transitory populations of SR1F-con- taining neurons have been observed in the cerebral cortex of the rat 2°.
In a previous quantitative study we have observed in the same cortical region an increase in the number of cortical neurons from birth to day 15 that was fol- lowed by a decrease of about 16% in the cat aged 25 days and a cumulative loss of 21% in the cat aged six months s. That increase was in accordance with re- suits from tritiated thymidine studies 17'2~ indicating that neurons generated at prenatal days 50-57 do not reach their definite sites in the upper cortical layers (layers II + III) until about two weeks later. Hence, considering all these data, it may be inferred that neuronal recruitment and cell loss overlap one anoth- er during cortical histogenesis in the cat. A positive balance is found during the first two weeks and this is
z,,a.i
Z
N
5 DAYS
15 DAYS
10 ~,T,,lr Tr 25 DAYS 5
T . . . . . . . . . . . . . . . . . . .
S.L. S.S-SYL.
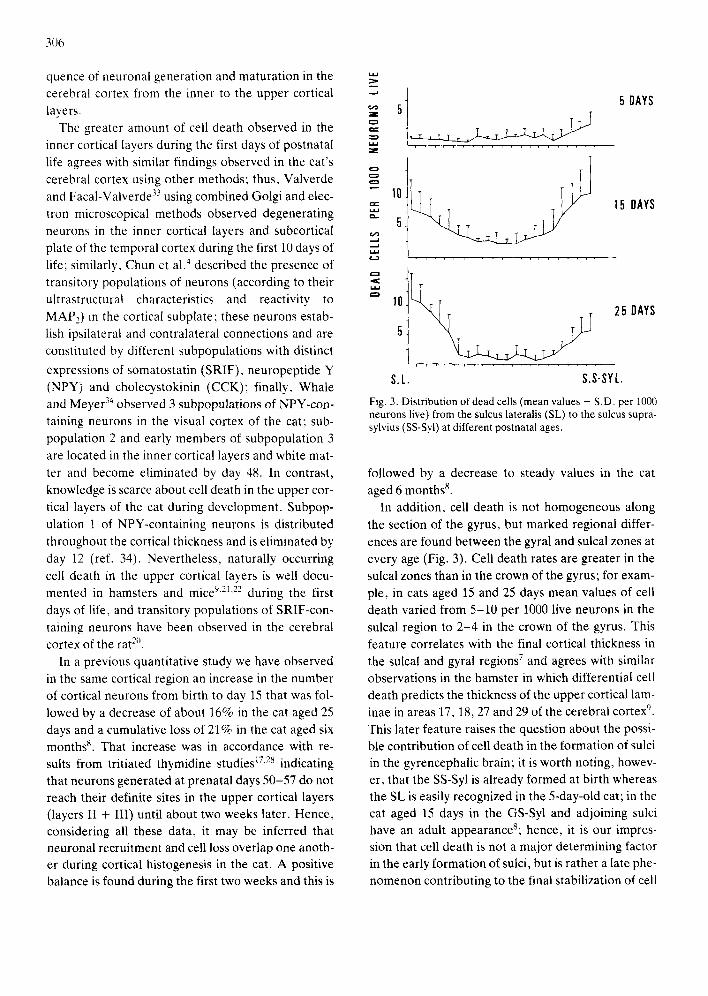
Fig. 3. Distribution of dead cells (mean values + S.D. per 1000 neurons live) from the sulcus lateralis (SL) to the sulcus supra- sylvius (SS-Syl) at different postnatal ages.
followed by a decrease to steady values in the cat aged 6 months 8.
In addition, cell death is not homogeneous along the section of the gyrus, but marked regional differ- ences are found between the gyral and sulcal zones at every age (Fig. 3). Cell death rates are greater in the sulcal zones than in the crown of the gyrus; for exam- ple, in cats aged 15 and 25 days mean values of cell death varied from 5-10 per 1000 live neurons in the sulcal region to 2 -4 in the crown of the gyrus. This feature correlates with the final cortical thickness in the sulcal and gyral regions 7 and agrees with similar observations in the hamster in which differential cell death predicts the thickness of the upper cortical lam- inae in areas 17, 18, 27 and 29 of the cerebral cortex '~. This later feature raises the question about the possi- ble contribution of cell death in the formation of sulci in the gyrencephalic brain; it is worth noting, howev- er, that the SS-Syl is already formed at birth whereas the SL is easily recognized in the 5-day-old cat; in the cat aged 15 days in the GS-Syl and adjoining sulci have an adult appearanceS; hence, it is our impres- sion that cell death is not a major determining factor in the early formation of sulci, but is rather a late phe- nomenon contributing to the final stabilization of cell

numbers in the sulcal and gyral regions, Finally, cell death rates are not synchronous in the
SS-Syl and SL; the rate of cell death is greater in the SS-Syl than in the SL in cats aged 5 and 15 days, but the contrary occurs in the cat aged 25 days (Fig. 3). These results are in accordance with the whole for- mation and growth of this gyrus and adjoining sulci; this gyrus emerges from the region adjacent to the al- most fully formed, at birth, SS-Syl and grows toward the developing SL 8. Morphological studies of the cor- tical infolding process in the ferret have also stressed a tangential spreading of the more mature tissue at the gyral crown while at the site of the future sulci growth is retarded 3°,31. Thus, the lateromedial gra-
dient of cell death agrees with the general latero-me-
dial gradient of formation and growth of the GS-Syl and with the lateromedial displacement of neurons toward the developing sulcus (SL) previously de- scribed in the cat between days 1 and 25 (ref. 8).
The significance of naturally occurring cell death in the cerebral cortex has been the subject of several studies focused on the relation between transitory populations of neurons and extracortical afferencies, although other factors may also be involved 2'15'1s'19.
On the one hand, reduction and elimination of neu- rons in the cortical subplate is simultaneous with the displacement of thalamic afferencies (waiting in the white mat ter 5,24'2s) toward the middle layers of the
cortex; neurons in the subplate are, therefore, con- sidered putative transitory targets for thalamic affe- rencies during the 'waiting per iod ' until definite con- nections can be established with neurons in the upper cortical layers 4,28,33. On the other hand, loss of neu-
rons in the cortical plate is closely related to the arri- val of thalamic afferencies; thus, in the visual cortex
307
of the mouse there is a loss of layer VI neurons during the first ten days of life which is simultaneous with the formation of specific thalamo-cortical connec- tionsll, lz.
In the visual cortex of the cat the final position of neurons in layers II + III is not reached until about the third week and it is not until this age that segrega- tion of ocular dominance columns occurs in layer IV 16,28. It may be significant that during the same pe-
riod dying cells are mainly found in the upper cortical layers in the GS-Syl.
It is far from our intention to theorize, on the basis of the present morphological data, about the role of deafferentation or competi t ion on the death of corti- cal neurons; there is not, as yet, an explanation for
the region differences between the gyral and sulcal zones as regards the different cell death rates, nor is there an explanation for the lateromedial gradient of cell death during development. Studies focusing on regional differences and differential temporal gra- dients in the arrival of thalamic afferencies to the GS- Syl and adjoining sulci would probably be illuminat- ing. Finally, it is worth emphasizing that little is known about glial cell death during development. In addition to nerve cell recruitment in the upper corti- cal layers 17,28, there is a major increase in the number
of glial cells in the cerebral cortex of the cat during the same period13; this feature, together with the lim-
itations of the method here employed (counting of pyknotic nuclei), does not permit us to rule out the presence of indeterminate numbers of glial cells among dying cells.
We wish to thank Mr. L. Barnet t for his assistance in the stylistic revision of the manuscript.
1 Bear, M.F., Carnes, K.N. and Ebner, F.F., Postnatal ch~knges in the distribution of acetylcholinesterase in the kitten striate cortex, J. Comp. Neurol., 237 (1985) 519-532.
2 Cowan, W.M., Neuronal death as a regulative mechanism in the control of cell number in the nervous system, In M. Rockstein (Ed.), Development and Aging in the Nervous System, Academic, New York, 1973, pp. 19-41.
3 Chu-Wang, I.W. and Oppenheim, R.W., Cell death ofmo- toneurons in the chick embryo spinal cord. I. Light and electron microscopical study of naturally-occurring and in- duced cell loss during development, J. Comp. Neurol., 177 (1978) 33-58.
4 Chun, J.J.M., Nakamura, N.J. and Shatz, C.J., Transient
cells of the developing mammalian telencephalon are pep- tide immuno-reactive neurons, Nature (Lond.), 325 (1987) 617-620.
5 Crandall, J.E. and Caviness, V.S., Thalamo-cortical con- nections in newborn mice, J. Comp. Neurol., 228 (1984) 542-556.
6 Cunningham, T.J., Naturally occurring neuron death and its regulation by developing natural pathways, Int. Rev. Cy- toL, 74 (1982) 163-187.
7 Ferrer, I., F~bregues, I. and Condom, E., A Golgi study of the sixth layer of the cerebral cortex. III. Neuronal changes during normal and abnormal cortical folding, J. Anat., 152 (1987) 71-82.
8 Ferrer, I., Hernandez-Martf, M., Bernet, E. and Galofr6,

308
E., Formation and growth of the cerebral convolutions. I. Postnatal development of the median-suprasylvian gyrus and adjoining sulci in the cat, J, Anat., in press.
9 Finlay, B.L. and Slattery, M., Local differences in the amount of early cell death in neocortex predict adult local specializations, Science, 219 (1983) 1349-1351.
10 Friedman, B. and Price, J.L., Age-dependent cell death in the olfactory cortex: lack of transneuronal degeneration in neonates, J. Comp. Neurol., 246 (1986) 20-31.
11 Heumann, D. and Leuba, G., Neuronal death in the devel- opment and aging of the cerebral cortex of the mouse, Neu- ropathol. Appl. Neurobiol., 9 (1983)297-311.
12 Heumann, D. and Rabinowitz, T., Postnatal development of the dorsal lateral geniculate nucleus in the normal and enucleated albino mouse, Exp. Brain Res., 38 (1980) 75-85.
13 Hillebrand. H., Quantitative Untersuchungen fiber Postna- tale Verfinderungen der Glia im Corpus Callosum der Katze, Z. Zellforsch., 73 (1966) 303-312.
14 Lager, P., ElOments de Neuro-Anatomie Fonctionelle: TOlencOphale, Masson, Paris, 1976, p. 55.
15 Kfillen, B., Degeneration and regeneration in the verte- brate central nervous system during embryogenesis, Prog. Brain Res., 14 (1965) 77-96.
16 LeVay, S., Stryker, M.P. and Shatz, C.J., Ocular domi- nance columns and the development in layer IV of the cat's visual cortex: a quantitative study, J. Comp. Neurol., 179 (1978) 223-244.
17 Luskin. M.B. and Shatz. C.J., Neurogenesis of the cat's pri- mary visual cortex, J. Comp. Neurol.. 242 (1985) 611-631.
18 Oppenheim, R.W.. Neuronal cell death and some related progressive phenomena during neurogenesis: a selective historical review and progress report. In W.M. Cowan (Ed.), Studies in Developmental Neurobiology, Oxford University Press, New York, 1981, pp. 74-133.
19 Oppenheim, R.W., Naturally occurring cell death during normal development, Trends Neurosci., 8 (1985) 487-493.
20 Parnavelas, J.G. and Cavanagh, M.E., Transient expres- sion of neurotransmitters in the developing neocortex, Trends Neurosci., 11 (1988) 92-93.
21 Pearlman. A.L., The visual cortex of the normal mouse and the reeler mutant. In A. Peters and E.G. Jones (Eds.), Ce- rebral Cortex, Vol. 3, Visual Cortex, Plenum, New York and London, 1985, pp. 1-18.
22 Pearlman, A.L., Bautista, J.O. and Cohen, J.P., Cell death in the neocortex of the normal mouse and the reeler mu- tant, Soc. Neurosci. Abstr., 9 (1983) 322.
23 Pilaf, G. and Landmesser, L., Ultrastructural differences during embryonic cell death in normal and peripherally de- prived ciliary ganglia, J. Cell Biol., 68 (1976) 339-356.
24 Rakic, P., Prenatal development of the visual system in the Rhesus monkey, Phil. Trans. R. Soc. Lond. Ser. B, 278 (1977) 245-260.
25 Sanides, F. and Hoffman, J., Cyto- and Myelo-architecture of the visual cortex of the cat and the surrounding integra- tion cortices, J. Hirnforsch., 11 (1969) 79-104.
26 Sengelaub, D.R. and Finlay, B.L., Cell death in the mam- malian visual system during normal development. I. Reti- nal ganglion cells, J. Comp. Neurol., 204 (1982) 311-317.
27 Sengelaub, D.R., Jacobs, L.F. and Finlay, B.L,, Regional differences in normally occurring cell death in the develop- ing hamster lateral geniculate nuclei, Neurosci. Lett., 55 (1985) 103-108.
28 Shatz, C.J. and Luskin, M.B., The relationship between the geniculocortical afferents and their cortical target cells during development of the cat's primary visual cortex, J. Neurosci., 6 (1986) 3655- 3668.
29 Silver, J. and Hughes, A.F.W., The role of cell death dur- ing morphogenesis of the mammalian eye, J. Morphol., 140 (1973) 159-170.
30 Smart, I.H.M. and McSherry, G.M., Gyms formation in the cerebral cortex in the ferret. I. Description of the exter- nal changes, J. Anat., 146 (1986a) 141-152.
31 Smart, I.H.M. and McSherry, G.M., Gyrus formation in the cerebral cortex in the ferret. II. Description of the inter- nal histological changes, J. Anat., 147 (1986b) 27-43.
32 Trujillo, C.Ma., Yanes, C.M., Marrero, A., Perez, M.A. and Martin, J.M., Cell death in the embryonic brain of Gal- Iota galloti (reptilia; Lacertidae): a structural and ultra- structural study, J. Anat., 150 (1987) 11-21.
33 Valverde, F. and Facal-Valverde, M.V., Transitory pop- ulation of cells in the temporal cortex of kittens, Dev. Brain Res., 32 (1987) 283-288.
34 Whale, P. and Meyer, G., Morphology and quantitative changes of transient NPY-ir neuronal populations during early postnatal development of the cat visual cortex, J. Comp. Neurol., 261 (1987) 165-192.
Copyright © 2022 FDOKUMEN




















