
Фофанова Е Г и др
11
БИОЛОГИЯ МОРЯ, 2014, том 40, № 1, с. 46–56 46 УДК 595.141+591.481 ЗООЛОГИЯ БЕСПОЗВОНОЧНЫХ реСниЧнЫе и нерВнЫе ЭЛеМенТЫ У юВениЛьнЫХ СаМоК аннеЛидЫ dInOPhIlUS GyrOcIlIAtUS (o. sChMIDt, 1848) (AnneLIDA: PoLyChAetA) 1 © 2014 г. е. Г. фофанова, Л. П. незлин, е. е. Воронежская Институт биологии развития им. Н.К. Кольцова РАН, Москва 119334 e-mail: [email protected] Статья принята к печати 6.06.2013 г. У ювенильных самок Dinophilus gyrociliatus (O. Schmidt, 1848) исследовали ресничные и нервные элементы после иммуномечения антителами к тубулину, серотонину и FMRFамиду. С помощью антител к тубулину на по- верхности тела животных выявлены два головных и семь туловищных ресничных шнуров, головные ресничные поля и вентральная ресничная полоска. Внутри тела обнаружены реснички пищеварительного канала и пять пар протонефридиев. Установлено, что нервная система представлена головным нейропилем с отходящими тяжами, семью продольными туловищными стволами, комиссурами и кольцевыми нервами. С помощью антител к серото- нину выявлены головной нейропиль, многочисленные тела нейронов в основании вентральной ресничной поло- ски, окологлоточное нервное кольцо и семь продольных вентральных стволов. Антитела к FMRFамиду позволили обнаружить около десяти нейронов в головном отделе, пять продольных стволов, окологлоточное и каудальное нервные кольца. Представленные данные демонстрируют черты упрощения в строении нервной системы динофи- люса, возможно, связанные с педоморфозом. Ключевые слова: аннелиды, Dinophilus, тубулин, серотонин (5-НТ), FMRFамид, нервная система. Ciliary and nervous structures in juvenile females of the annelid dinophilus gyrociliatus (o. schmidt, 1848) (Annelida: Polychaeta). E. G. Fofanova, L. P. Nezlin, E. E. Voronezhskaya (Koltsov Institute of Developmental Biology, Russian Academy of Sciences, Moscow 119334) Ciliary and nerve structures were described in juvenile female Dinophilus gyrociliatus (O. Schmidt, 1848) after immunochemical staining with tubulin, serotonin, and FMRFamide antibodies. Anti-tubulin antibodies visualized the fol- lowing external structures: two head and seven trunk ciliary bands, a ventral ciliary band, and head ciliary fields. Gut cilia and five pairs of protonephridia were detected inside the body. The nervous system consists of an oval head neuropile with anterior and posterior nerves extending from it, seven longitudinal nerve cords, commissures, and circular nerves. Anti- serotonin antibodies visualized the head neuropile, neurons at the base of the ventral ciliary band, an oesophageal ring, and seven longitudinal ventral cords. Anti-FMRFamide antibodies visualized about ten neurons in the cerebral ganglion, five longitudinal cords, and oesophageal and caudal nerve rings. The presented data suggest the simplification of the ner- vous system structure in D. gyrociliatus, which probably reflects pedomorphosis. (Biologiya Morya, 2014, vol. 40, no. 1, pp. 46–56). Keywords: annelids, Dinophilus, tubulin, serotonin (5-HT), FMRFamide, nervous system. 1 Исследования поддержаны грантами РФФИ № 12-04-01510, № 12-04-10119 и № 13-04-10164, а также частично проводились в рамках реали- зации Федеральной целевой программы "Научные и научно-педагогические кадры инновационной России" на 2009–2013 годы при финансовой поддержке Министерства образования и науки РФ (Государственный контракт № 12.741.12.0155). ФОФАНОВА И ДР. РеСНИЧНые Последние данные о морфологии беспозвоночных, полученные с применением методов иммуногистохимии и лазерной сканирующей микроскопии, позволили ина- че взглянуть на раннее развитие представителей группы Lophotrochozoa (Воронежская, Ивашкин, 2010; Незлин, 2010). В настоящее время проводится ревизия имею- щихся данных. Наиболее перспективными представля- ются два направления: 1) изучение последовательных стадий развития и 2) сравнительный анализ морфологии редких малоизученных групп животных. Одной из таких групп являются представители семейства Dinophilidae (Annelida: Polychaeta). Эти животные привлекают вни- мание зоологов с момента открытия, так как взрослые особи сочетают в себе признаки нескольких типов и не- сут черты неотении (Иванов и др., 1941; Беклемишев, 1964; Struck, 2006). Многие зоологи относили динофилид к эуницид- ным полихетам, придавая особое значение простоте в структуре глоточного бульбуса и морфологии личинок (Беклемишев, 1964; Свешников, 1967; Hermans, 1969; Akesson, 1977), но данные молекулярно-генетического анализа не подтвердили эту точку зрения (Struck et al., 2005; Struck, 2006). Вагин (2007) относил динофилид (вместе с мизостомидами и моллюсками) к надтипу Trochozoa Oligomera, противопоставляя их Trochozoa Heteronomia (аннелиды и архианнелиды). Старобогатов (1983) объединял динофилид и мизостомид в тип Myzostomozoes. Идея о том, что динофилиды – самосто-
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Фофанова Е Г и др

БИОЛОГИЯ МОРЯ, 2014, том 40, № 1, с. 46–56
46
УДК 595.141+591.481 ЗООЛОГИЯ БЕСПОЗВОНОЧНЫХ
реСниЧнЫе и нерВнЫе ЭЛеМенТЫ У юВениЛьнЫХ СаМоК аннеЛидЫ dInOPhIlUS GyrOcIlIAtUS
(o. sChMIDt, 1848) (AnneLIDA: PoLyChAetA)1
© 2014 г. е. Г. фофанова, Л. П. незлин, е. е. ВоронежскаяИнститут биологии развития им. Н.К. Кольцова РАН, Москва 119334
e-mail: [email protected]
Статья принята к печати 6.06.2013 г.
У ювенильных самок Dinophilus gyrociliatus (O. Schmidt, 1848) исследовали ресничные и нервные элементы после иммуномечения антителами к тубулину, серотонину и FMRFамиду. С помощью антител к тубулину на по-верхности тела животных выявлены два головных и семь туловищных ресничных шнуров, головные ресничные поля и вентральная ресничная полоска. Внутри тела обнаружены реснички пищеварительного канала и пять пар протонефридиев. Установлено, что нервная система представлена головным нейропилем с отходящими тяжами, семью продольными туловищными стволами, комиссурами и кольцевыми нервами. С помощью антител к серото-нину выявлены головной нейропиль, многочисленные тела нейронов в основании вентральной ресничной поло-ски, окологлоточное нервное кольцо и семь продольных вентральных стволов. Антитела к FMRFамиду позволили обнаружить около десяти нейронов в головном отделе, пять продольных стволов, окологлоточное и каудальное нервные кольца. Представленные данные демонстрируют черты упрощения в строении нервной системы динофи-люса, возможно, связанные с педоморфозом.
Ключевые слова: аннелиды, Dinophilus, тубулин, серотонин (5-НТ), FMRFамид, нервная система.
Ciliary and nervous structures in juvenile females of the annelid dinophilus gyrociliatus (o. schmidt, 1848) (Annelida: Polychaeta). E. G. Fofanova, L. P. Nezlin, E. E. Voronezhskaya (Koltsov Institute of Developmental Biology, Russian Academy of Sciences, Moscow 119334)
Ciliary and nerve structures were described in juvenile female Dinophilus gyrociliatus (O. Schmidt, 1848) after immunochemical staining with tubulin, serotonin, and FMRFamide antibodies. Anti-tubulin antibodies visualized the fol-lowing external structures: two head and seven trunk ciliary bands, a ventral ciliary band, and head ciliary fields. Gut cilia and five pairs of protonephridia were detected inside the body. The nervous system consists of an oval head neuropile with anterior and posterior nerves extending from it, seven longitudinal nerve cords, commissures, and circular nerves. Anti-serotonin antibodies visualized the head neuropile, neurons at the base of the ventral ciliary band, an oesophageal ring, and seven longitudinal ventral cords. Anti-FMRFamide antibodies visualized about ten neurons in the cerebral ganglion, five longitudinal cords, and oesophageal and caudal nerve rings. The presented data suggest the simplification of the ner-vous system structure in D. gyrociliatus, which probably reflects pedomorphosis. (Biologiya Morya, 2014, vol. 40, no. 1, pp. 46–56).
Keywords: annelids, Dinophilus, tubulin, serotonin (5-HT), FMRFamide, nervous system.
1 Исследования поддержаны грантами РФФИ № 12-04-01510, № 12-04-10119 и № 13-04-10164, а также частично проводились в рамках реали-зации Федеральной целевой программы "Научные и научно-педагогические кадры инновационной России" на 2009–2013 годы при финансовой поддержке Министерства образования и науки РФ (Государственный контракт № 12.741.12.0155).
ФОФАНОВА И ДР.РеСНИЧНые
Последние данные о морфологии беспозвоночных, полученные с применением методов иммуногистохимии и лазерной сканирующей микроскопии, позволили ина-че взглянуть на раннее развитие представителей группы Lophotrochozoa (Воронежская, Ивашкин, 2010; Незлин, 2010). В настоящее время проводится ревизия имею-щихся данных. Наиболее перспективными представля-ются два направления: 1) изучение последовательных стадий развития и 2) сравнительный анализ морфологии редких малоизученных групп животных. Одной из таких групп являются представители семейства Dinophilidae (Annelida: Polychaeta). Эти животные привлекают вни-мание зоологов с момента открытия, так как взрослые особи сочетают в себе признаки нескольких типов и не-
сут черты неотении (Иванов и др., 1941; Беклемишев, 1964; Struck, 2006).
Многие зоологи относили динофилид к эуницид-ным полихетам, придавая особое значение простоте в структуре глоточного бульбуса и морфологии личинок (Беклемишев, 1964; Свешников, 1967; Hermans, 1969; Akesson, 1977), но данные молекулярно-генетического анализа не подтвердили эту точку зрения (Struck et al., 2005; Struck, 2006). Вагин (2007) относил динофилид (вместе с мизостомидами и моллюсками) к надтипу Trochozoa Oligomera, противопоставляя их Trochozoa Heteronomia (аннелиды и архианнелиды). Старобогатов (1983) объединял динофилид и мизостомид в тип Myzostomozoes. Идея о том, что динофилиды – самосто-

РеСНИЧНые 47
БИОЛОГИЯ МОРЯ том 40 № 1 2014
ятельный тип, до сих пор находит сочувствие в отече-ственной литературе (Буруковский, 2010).
Первые упоминания о Dinophilus появились во вто-рой половине XIX века (Schmidt, 1848, цит. по: Hermans, 1969; Harmer, 1889). С этого момента и до начала XX века опубликовано множество работ о находках самых разных видов, выделяемых по количеству сегментов и ресничных шнуров и относящихся к роду Dinophilus (Harmer, 1889; Nelson, 1907; Shearer, 1912). Описано око-ло 10 видов, которые пытались разделить на две группы. Основные диагностические критерии для такого раз-деления – количество сегментов, строение реснично-го аппарата, окраска тела, а главное, жизненный цикл. В результате в первую группу вошли виды без полового диморфизма, во вторую – с крайней степенью полового диморфизма (Nelson, 1907; Shearer, 1912).
В середине XX века были опубликованы работы по биологии Dinophilus, однако их авторы часто или не определяли видовую принадлежность (Jägersten, 1944), либо относили описываемое животное к одному из ранее выделенных видов (Котикова, 1973; Мамкаев, Селиванова, 1979). Вслед за Нельсоном (Nelson, 1907) Хартман (Hartman, 1959) в своем определителе к роду Dinophilus отнес 5 видов, из которых 4 вида (все окра-шенные, без выраженного полового диморфизма) вхо-дили в первую группу и один вид D. gyrociliatus – во вторую. Позже было показано, что представители всех видов первой группы рода Dinophilus являются одним видом D. taeniatus (Harmer, 1889; Donworth, 1986).
Таким образом, в настоящее время к роду Dinophilus принято относить два вида: D. gyrociliatus и D. taeniatus. Изучение деталей строения ресничных элементов с использованием современных методов ак-туально для определения систематической принадлеж-ности видов данного рода. Строение нервной системы исследовали в основном на взрослых особях D. taeniatus и D. gyrociliatus, имеются сведения о морфологии нерв-ной системы карликового самца D. gyrociliatus. Причем информация, приводимая разными авторами, весьма противоречива (см. "Обсуждение"). В данной работе проведено детальное исследование ресничной и нерв-ной систем D. gyrociliatus с использованием высоко-чувствительных методов иммунохимического марки-рования, совмещенного с конфокальной сканирующей микроскопией.
МАТеРИАЛ И МеТОДИКА
Культивирование животных
Работа выполнена в лаборатории сравнительной физио-логии Института биологии развития (ИБР) им. Н.К. Кольцова РАН. Лабораторная культура динофилид была создана на основе кладок и ювенильных особей, обнаруженных нами в пробах морской воды из Средиземного моря (Неаполитанская зоологическая станция). Взрослых животных содержали со-гласно методике, описанной ранее (Фофанова, Воронежская, 2008).
Иммуноцитохимическое маркирование
Для маркировки ресничек и нервных элементов исполь-зовали антитела против ацетилированного тубулина (моно-клональные, выработанные в мыши, T-6793, Sigma, USA); для маркировки специфичных нервных элементов – антитела про-тив серотонина (5-гидрокситриптамин, 5-HT) и нейропептида FMRFамида (все поликлональные, выработаны в кролике, Cat. Nrs. 428002, 410002, Immunostar, USA). Первичные антитела выявляли при помощи вторичных антител против иммуногло-булинов кролика и мыши (goat-anti-rabbit IgG, goat-anti-mouse IgG), меченных флуоресцентными маркерами Alexa-488 и Alexa-546 (все Molecular Probes, USA).
Иммунореакцию проводили на тотальных препаратах ювенильных червей (1–3 дня после вылупления). Животных предварительно расслабляли в 7% растворе MgCl2 на морской воде в течение 2–3 мин. Затем фиксировали 4% параформаль-дегидом (PFA) на 0.01 M фосфатном буфере (PBS, рН 7.4) в те-чение ночи при температуре 10°С. Последующее иммунохи-мическое маркирование проводили по стандартной методике (Voronezhskaya et al., 2003; Fofanova, Voronezhskaya, 2012). Для двойного окрашивания в одном препарате использовали первичные антитела, выработанные в разных животных, в сле-дующих разведениях: для серотонина и FMRFамида – 1 : 2000, для тубулина – 1 : 5000. Вторичные антитела разводили в про-порции 1 : 800. Ядра маркировали DAPI или Hoechst-33342 (все Molecular Probes, USA) в концентрации 2–3 мкг/мл. Препараты заключали в 70% глицерин в PBS.
Предварительный анализ препаратов производили с по-мощью эпифлуоресцентного микроскопа DMRXA2 (Leica, Germany) и микроскопа Zeiss Axioskop (Zeiss, Germany). Для детального изучения препаратов использовали лазерные кон-фокальные сканирующие микроскопы TCS-SPE и TCS-SP5 (Leica, Germany), оборудованные соответствующим набором лазеров, фильтров и детекторов. Двухмерные изображения и трехмерные реконструкции создавали на основе серий опти-ческих срезов с шагом 0.5–0.7 мкм с помощью программ Leica LAS AF и Image J. Чтобы получить иллюстрации, серию опти-ческих срезов проецировали в одно изображение и импорти-ровали в программу Adobe Photoshop CS 3, изменяли только яркость и контрастность изображения. Рисунки 1–3 и 5 пред-ставлены в виде монохромных негативных изображений.
РеЗУЛЬТАТы
Особенности морфологии
Тело червей (самки) практически не окрашено, прозрачно, длина ювенильных особей составляет 0.4–0.6 мм, половозрелых – 0.8–1.5 мм. В теле выделяются го-ловной и туловищный отделы, а также каудальный отро-сток, отделенный от туловищного отдела бороздой (рис. 1А, Б). В туловищном отделе видны 6 латеральных бо-розд, которые более четко выражены у ювенильных осо-бей (рис. 1А), однако различимы и у взрослых (рис. 1Б).
При содержании животных в пластиковых кон-тейнерах кладки появляются регулярно, в каждой кладке четко различаются яйца двух типов: большие (90–120 мкм) – будущие самки, и маленькие (50 мкм) – будущие самцы (рис. 1В, Г). На пятый день после отклад-ки яиц ювенильные самки покидают яйцевую оболочку.

48 ФОФАНОВА И ДР.
БИОЛОГИЯ МОРЯ том 40 № 1 2014
После выхода ювенильных самок из кокона под общей оболочкой кладки видны оставшиеся мелкие эмбрионы, это карликовые самцы (рис. 1Д), которые не покидают кладку. При дальнейшем наблюдении отмечено помут-нение и последующий распад содержимого кладки.
При окрашивании ДНК у всех изученных юве-нильных особей (n = 60) сразу после вылупления в ту-ловищном отделе между пятой и шестой латеральными бороздами обнаружены две симметрично расположен-ные сферические области диаметром 8–13 мкм, отлича-ющиеся наличием многочисленных мелких клеточных ядер с высокой плотностью хроматина (рис. 2Б, В, Г), характерных для сперматозоидов (Franzen, 1977). При окрашивании эмбрионов непосредственно перед вылу-плением (n = 60) области с повышенной концентрацией хроматина отсутствовали (рис. 2А).
Внешние и внутренние ресничные структуры
Поверхностные ресничные структуры представ-лены двумя головными шнурами, семью туловищными шнурами и вентральной ресничной полоской (рис. 3А, Б, В, Д, е). Головные шнуры охватывают голову полу-кольцом, не замкнутым с дорсальной стороны и рас-ширяющимся на вентральной стороне (рис. 3А, Б, В). В передней части головы, ограниченной первым голов-ным шнуром, находится область с несколькими пуч-ками длинных (15–18 мкм) ресничек (рис. 3А, Б, Д). Дорсолатерально на границе головного и туловищного отделов симметрично по обеим сторонам тела выявля-ются два ресничных поля овальной формы (рис. 3Б, В, Г, Г'). Реснички, составляющие эти поля, существенно ко-роче (6–8 мкм) ресничек основных шнуров (15–17 мкм), а их основания расположены плотнее друг к другу
рис. 1. Кладки и внешняя морфология самок Dinophilus gyrociliatus. А – ювенильная самка непосредственно после вылупления; Б – взрос-лая самка. Четко видно деление тела на головной (hr), туловищный (br) и каудальный (cr) отделы; в головном отделе просматриваются пара глаз (e) и пара мощных ресничек (small arrows); на границе головного и туловищного отделов расположен рот (m), в туловищном отделе четко видны 6 латеральных борозд (arrows); у взрослой самки латеральные борозды менее выражены; в середине туловищного от-дела просматривается темный кишечник. В – кладка, отложенная молодым червем в возрасте 7 дней и содержащая два крупных женских яйца и одно мелкое мужское яйцо (показано стрелкой); Г – кладка, отложенная взрослым червем в возрасте 18 дней и содержащая пять крупных женских яиц и два мелких мужских яйца (стрелки); Д – кладка после вылупления ювенильных самок, видны оставшиеся мелкие мужские яйца (стрелки). Масштаб: А – 60; Б – 120; В, Г, Д – 90 мкм.

РеСНИЧНые 49
БИОЛОГИЯ МОРЯ том 40 № 1 2014
(рис. 3Г, Г'). Мы не смогли обнаружить нервные волок-на непосредственно под ресничным полем, хотя один из крупных нервных тяжей (вентролатеральный, см. далее) проходит в непосредственной близости под основанием ресничек (рис. 3Г').
В туловищном отделе выделяется вентральная рес-ничная полоска, к ней прилегают концы семи кольцевых шнуров (рис. 3А, Б), которые располагаются на наиболее выпуклых участках тела между латеральными борозда-ми. Первый туловищный шнур на дорсальной стороне не замкнут (рис. 3В). Длина ресничек во всех шнурах одинакова и составляет 15–17 мкм. Реснички вентраль-ной полоски заметно короче (7–9 мкм).
Среди внутренних ресничных элементов можно выделить реснички пищеварительного канала и пять пар протонефридиев (рис. 3Д, Ж). Длина ресничек в перед-ней части пищевода, средней и задней кишке составляет 9–10 мкм, а в глотке и остальном пищеводе – 5–6 мкм. Протонефридии расположены в области латеральных борозд (рис. 3Ж). Расстояние между первыми двумя па-рами протонефридиев меньше, чем между остальными, т.е. они сближены (рис. 3Д, Ж).
Элементы нервной системы, выявляемые при иммуномечении антителами к тубулину
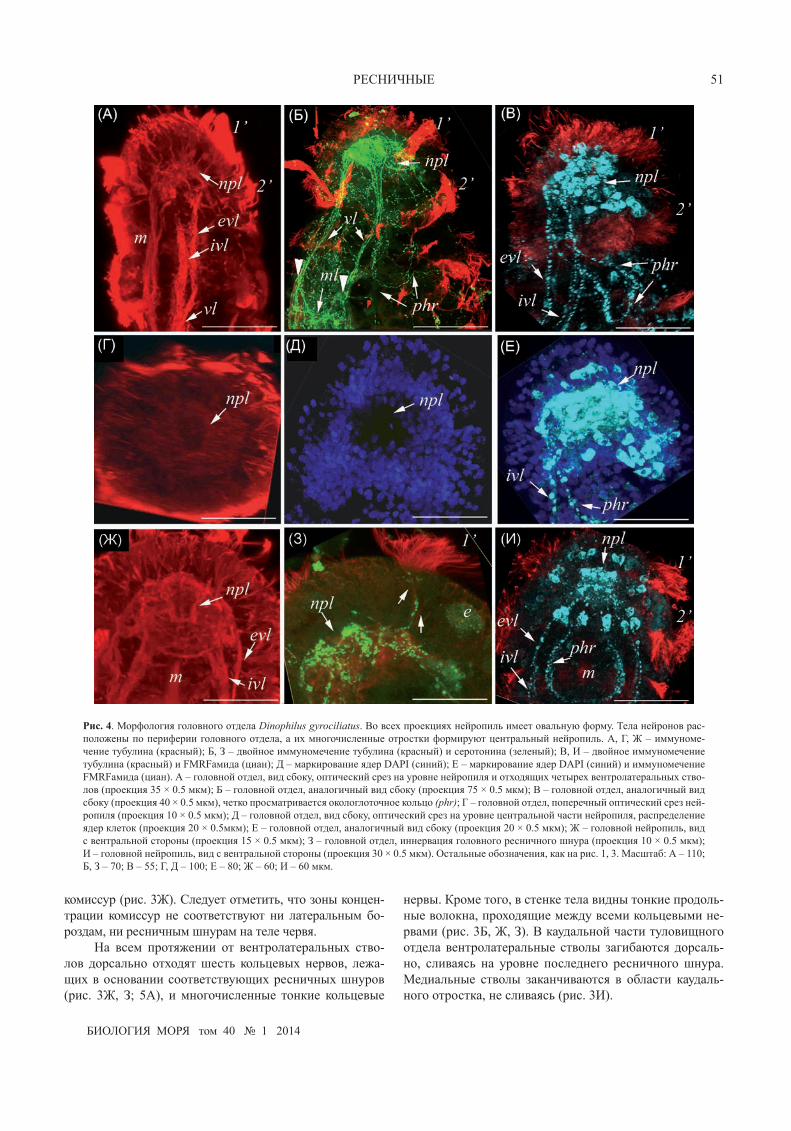
Кроме ресничных структур с помощью антител к тубулину выявлены элементы нервной системы. В го-ловном отделе расположен мощный нейропиль, во всех
проекциях (спереди, сбоку, на поперечном оптическом срезе) он имеет овальную форму (рис. 3Д–Ж, 4А–И, 5А–В). Область в центральной части нейропиля харак-теризуется пониженным содержанием волокон (рис. 4Г, Д, Ж). От дорсальной части нейропиля отходят много-численные волокна к основаниям ресничных шнуров и длинных ресничек, расположенных в передней части го-ловы (рис. 3Д, е, Ж; 4А, Г, Ж, З; 5А).
Справа и слева от нейропиля отходят по два тол-стых тяжа: дорсальные и вентральные, которые слива-ются, формируя два мощных вентролатеральных ствола (рис. 3е, Ж; 4А, Ж). От места слияния дорсального и вентрального стволов отходят медиальные отростки, ко-торые сливаясь, образуют медиальный тяж (рис. 3е, Ж), а также тонкие промежуточные продольные тяжи (рис. 3Ж, 5Г).
В туловищном отделе четко выделяется семь про-дольных стволов: пара наиболее мощных вентролатераль-ных, более тонкий медиальный и по паре промежуточных, расположенных с каждой стороны между медиальным и вентролатеральным стволами (рис. 3Ж, 5Г). На всем протяжении вентролатеральные стволы соединены между собой комиссурами (рис. 3е, Ж; 5Ж). В зависимости от размера червя число комиссур варьирует от 12 до 15; они располагаются по телу животного неравномерно, однако распределение комиссур позволяет выделить пять зон концентрации. На уровне первых двух сближенных пар протонефридиев наблюдается максимальное количество
рис. 2. Окрашивание ядер DAPI на разных стадиях развития Dinophilus gyrociliatus. Между 5-й и 6-й латеральными бороздами только у ювенильной особи видны две симметрично расположенные области с повышенной ядерной плотностью. А – эмбрион на 4-й день раз-вития, общий вид (проекция 40 × 0.5 мкм); Б–Г – ювенильная особь непосредственно после вылупления: Б – общий вид (проекция 60 × 0.5 мкм), В – вид сбоку на уровне 5–6-й латеральных борозд (проекция 60 × 0.5 мкм), Г – вид спереди на том же уровне (проекция 60 × 0.5 мкм). Масштаб: А – 80, Б – 150, В – 60, Г – 65 мкм.

50 ФОФАНОВА И ДР.
БИОЛОГИЯ МОРЯ том 40 № 1 2014
рис. 3. Иммуномечение тубулина у ювенильной особи Dinophilus gyrociliatus непосредственно после вылупления. А–Г – внешние рес-ничные структуры: А – вид с вентральной стороны (проекция 25 × 0.5 мкм), Б – вид с латеральной стороны (проекция 20 × 0.5 мкм), В – вид с дорсальной стороны (проекция 15 × 0.5 мкм). В головном отделе видны два ресничных шнура (1' и 2'), в туловищном – 7 ресничных шнуров (1–7) и вентральная ресничная полоска (vcf); оба ресничных шнура головного отдела, а также первый ресничный шнур туловищного отдела не замкнуты на дорсальной стороне; непосредственно над первым туловищным шнуром симметрично по обе-им сторонам тела дорсолатерально расположены два ресничных поля (показано стрелкой). Г – ресничное поле (стрелка) при большем увеличении, вид с латеральной стороны (проекция 15 × 0.5 мкм); Г' – ресничное поле (стрелка) и подлежащие внутренние тубулин-иммунореактивные структуры (проекция 25 × 0.5 мкм); Д–И – внутренние тубулин-иммунореактивные структуры (ресничные и нерв-ные): Д – вид на уровне кишечника, фронтальная проекция 15 × 0.5 мкм; е – вид сбоку, оптический срез на уровне вентролатеральных стволов (проекция 25 × 0.5 мкм); Ж – вид с вентральной стороны, оптический срез на уровне вентральной нервной цепочки (проекция 20 × 0.5 мкм); З – иннервация туловищных ресничных шнуров (проекция 15 × 0.5 мкм); И – каудальная часть туловищного отдела, вид сбоку (проекция 20 × 0.5 мкм). В туловищном отделе просматриваются реснички в кишечнике (стрелки на Д). В головном отделе (Д–Ж) выявляется мощный нейропиль (npl), от которого отходят наружный (evl) и внутренний (ivl) стволы, сливающиеся в вентролатеральный ствол (vl). Непосредственно под вентральной ресничной полоской (vcf) расположены медиальный ствол (ml), промежуточные (imc) и медиовентральные (mvc) стволы. В туловищном отделе от вентролатеральных стволов (vl) отходят латеральные нервы (ln). Помимо нервной системы выявляется пять пар протонефридиев (pn). Остальные обозначения, как на рис. 1. Масштаб: А, е – 90; Б, В, Ж – 120; Г – 25; Г' – 20; Д – 100; З – 30; И – 100 мкм.
РеСНИЧНые 51
БИОЛОГИЯ МОРЯ том 40 № 1 2014
комиссур (рис. 3Ж). Следует отметить, что зоны концен-трации комиссур не соответствуют ни латеральным бо-роздам, ни ресничным шнурам на теле червя.
На всем протяжении от вентролатеральных ство-лов дорсально отходят шесть кольцевых нервов, лежа-щих в основании соответствующих ресничных шнуров (рис. 3Ж, З; 5А), и многочисленные тонкие кольцевые
нервы. Кроме того, в стенке тела видны тонкие продоль-ные волокна, проходящие между всеми кольцевыми не-рвами (рис. 3Б, Ж, З). В каудальной части туловищного отдела вентролатеральные стволы загибаются дорсаль-но, сливаясь на уровне последнего ресничного шнура. Медиальные стволы заканчиваются в области каудаль-ного отростка, не сливаясь (рис. 3И).
рис. 4. Морфология головного отдела Dinophilus gyrociliatus. Во всех проекциях нейропиль имеет овальную форму. Тела нейронов рас-положены по периферии головного отдела, а их многочисленные отростки формируют центральный нейропиль. А, Г, Ж – иммуноме-чение тубулина (красный); Б, З – двойное иммуномечение тубулина (красный) и серотонина (зеленый); В, И – двойное иммуномечение тубулина (красный) и FMRFамида (циан); Д – маркирование ядер DAPI (синий); е – маркирование ядер DAPI (синий) и иммуномечение FMRFамида (циан). А – головной отдел, вид сбоку, оптический срез на уровне нейропиля и отходящих четырех вентролатеральных ство-лов (проекция 35 × 0.5 мкм); Б – головной отдел, аналогичный вид сбоку (проекция 75 × 0.5 мкм); В – головной отдел, аналогичный вид сбоку (проекция 40 × 0.5 мкм), четко просматривается окологлоточное кольцо (phr); Г – головной отдел, поперечный оптический срез ней-ропиля (проекция 10 × 0.5 мкм); Д – головной отдел, вид сбоку, оптический срез на уровне центральной части нейропиля, распределение ядер клеток (проекция 20 × 0.5мкм); е – головной отдел, аналогичный вид сбоку (проекция 20 × 0.5 мкм); Ж – головной нейропиль, вид с вентральной стороны (проекция 15 × 0.5 мкм); З – головной отдел, иннервация головного ресничного шнура (проекция 10 × 0.5 мкм); И – головной нейропиль, вид с вентральной стороны (проекция 30 × 0.5 мкм). Остальные обозначения, как на рис. 1, 3. Масштаб: А – 110; Б, З – 70; В – 55; Г, Д – 100; е – 80; Ж – 60; И – 60 мкм.

52 ФОФАНОВА И ДР.
БИОЛОГИЯ МОРЯ том 40 № 1 2014
Элементы нервной системы, выявляемые при иммуномечении антителами к 5-НТ
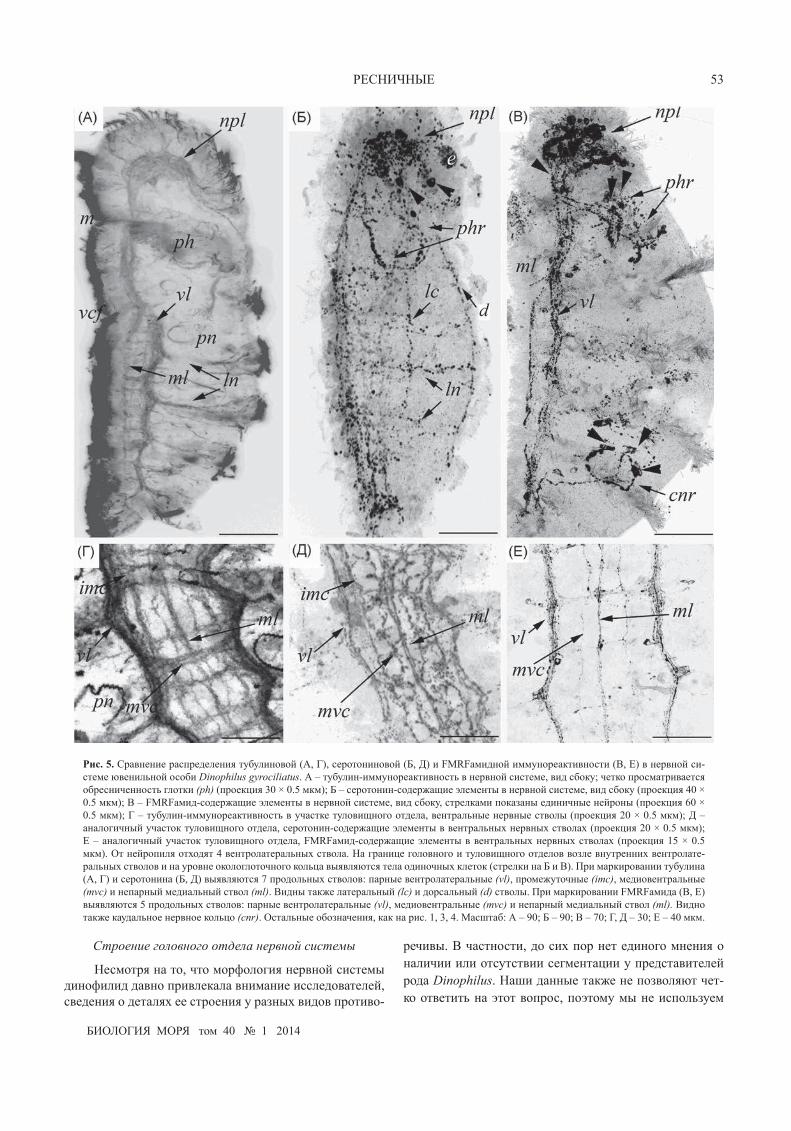
Общий план расположения 5-НТ-подобной им-мунореактивности сходен с выявляемым при имму-номечении антителами к тубулину. В головном отделе расположен нейропиль, в туловищном отделе видны вентральные нервные стволы с отходящими от них коль-цевыми нервами (рис. 4Б, З; 5Б, Д).
В головном отделе находятся единичные тела нейронов, посылающие свои отростки в нейропиль (рис. 5Б). От нейропиля отходят отдельные волокна к основаниям ресничек (рис. 4З, 5Б). От каудальной части нейропиля симметрично с боковых сторон отходит пара вентролатеральных стволов (рис. 4Б), которые продол-жаются в туловищном отделе (рис. 4Б; 5Б, Д). На грани-це головного и туловищного отделов от боковых стволов отходит пара более тонких тяжей, которые, сливаясь в виде подковы, охватывают глотку (рис. 4Б, 5Б).
В туловищном отделе на уровне первой латераль-ной борозды вентролатеральные стволы попарно раз-деляются на две части: наружную (более дорсальную) и внутреннюю (более вентральную) (рис. 4Б). Наружные стволы продолжаются в каудальном направлении не-посредственно под вентральной ресничной полоской. Внутренние стволы загибаются дугой навстречу друг другу и формируют медиальный нервный ствол. В свою очередь, от медиального ствола с обеих сторон отделяет-ся по одному тонкому стволу (рис. 4Б). Таким образом, под вентральной ресничной полоской насчитывается семь продольных стволов (рис. 4Б, 5Д). Ближе к по-верхности, непосредственно под вентральной реснич-ной полоской, залегают многочисленные тела нейронов (рис. 5З).
На всем протяжении туловищного отдела от вен-тролатеральных стволов отходят тонкие кольцевые не-рвы, расположенные непосредственно под ресничны-ми шнурами и охватывающие тело червя полукольцом (рис. 5Б).
Элементы нервной системы, выявляемые при иммуномечении антителами к FMRFамиду
Общий план строения FMRFaмид-иммун ореак тив-ной нервной системы сходен с выявляемым при иммуно-мечении антителами к тубулину.
В головном отделе находится около 30 FMRFaмид-иммунореактивных клеток. Их тела расположены в по-верхностном слое дорсальной части головного отдела нервной системы, а отростки следуют в центральный нейропиль (рис. 4В, е, И; 5В). С обеих сторон нейропиля отходят продольные дорсальный и вентральный вентро-латеральные стволы, которые переходят в туловищный отдел, образуя парные вентролатеральные и медиовен-тральные стволы (рис. 5В, е).
На границе головного и туловищного отделов меж-ду вентральными продольными стволами располагается комиссура. От комиссуры дорсально отходят по два тол-
стых волокна, следующих по правой и левой сторонам глотки (рис. 4В, е, И). Эти волокна ветвятся в средней и дистальной части глотки, сливаясь, они формируют сеть, охватывающую глотку с вентральной стороны (рис. 4В, е, И; 5В). По ходу волокон выявляются тела 4–6 нейро-нов (рис. 5В).
Каудальнее первой латеральной борозды от вну-тренних вентролатеральных стволов отходят отростки, которые сливаются и образуют медиальный ствол (рис. 5В). Всего в туловищном отделе можно выделить пять продольных стволов: парные вентролатеральные и ме-диовентральные, а также медиальный (рис. 5е). На всем протяжении продольные стволы соединены между со-бой нерегулярно расположенными комиссурами (рис. 5е). Кроме того, в средней части тела на уровне третьего и четвертого кольцевых ресничных шнуров от наружных вентролатеральных стволов отходят волокна, ветвящие-ся в толще тела (рис. 5В).
На уровне пятой латеральной борозды от медиаль-ного тяжа отходит единичное нервное волокно, охваты-вающее кольцом с дорсальной стороны заднюю кишку на уровне пятого кольцевого ресничного шнура, каудаль-ное нервное кольцо (рис. 5В). На прямолинейном участ-ке этого волокна видны два нейрона, а на кольцевом рас-положены три единичных нейрона (рис. 5В).
ОБСУЖДеНИе
Видовая принадлежность
По совокупности выявленных особенностей био-логии, внешнего и внутреннего строения наш объект относится к виду Dinophilus gyrociliatus. Тело червя не окрашено, имеет характерное распределение ресничных шнуров и протонефридиев, что согласуется с диагности-ческими признаками для D. gyrociliatus. Убедительным доказательством видовой принадлежности является на-личие карликового самца в жизненном цикле: мелкие яйца развиваются в самцов, которые не вылупляются, а умирают, не покидая кладки (Shearer, 1912).
Вопрос о времени оплодотворения у D. gyrociliatus до сих пор остается дискуссионным. В литературе встречаются две гипотезы: либо оплодотворение про-исходит в кладке непосредственно перед вылуплени-ем и самцы гибнут, не выходя из кладки, либо опло-дотворение происходит после вылупления, самцы еще в кладке прикрепляются к самкам и некоторое время живут в общей популяции. Согласно нашим данным, парные области с многочисленными плотно лежащи-ми мелкими ядрами с высокой плотностью хроматина, характерными для сперматозоидов (Franzen, 1977), вы-являются только у ювенильных и взрослых особей и не встречаются у эмбрионов. В оболочках кладок после вылупления самок всегда присутствуют остатки мелких яиц. Полученные результаты свидетельствуют в пользу второй гипотезы: оплодотворение происходит внутри кладки, и самка получает запас спермы, который рас-ходует в течение жизни.
РеСНИЧНые 53
БИОЛОГИЯ МОРЯ том 40 № 1 2014
Строение головного отдела нервной системы
Несмотря на то, что морфология нервной системы динофилид давно привлекала внимание исследователей, сведения о деталях ее строения у разных видов противо-
речивы. В частности, до сих пор нет единого мнения о наличии или отсутствии сегментации у представителей рода Dinophilus. Наши данные также не позволяют чет-ко ответить на этот вопрос, поэтому мы не используем
рис. 5. Сравнение распределения тубулиновой (А, Г), серотониновой (Б, Д) и FMRFамидной иммунореактивности (В, е) в нервной си-стеме ювенильной особи Dinophilus gyrociliatus. А – тубулин-иммунореактивность в нервной системе, вид сбоку; четко просматривается обресниченность глотки (ph) (проекция 30 × 0.5 мкм); Б – серотонин-содержащие элементы в нервной системе, вид сбоку (проекция 40 × 0.5 мкм); В – FMRFамид-содержащие элементы в нервной системе, вид сбоку, стрелками показаны единичные нейроны (проекция 60 × 0.5 мкм); Г – тубулин-иммунореактивность в участке туловищного отдела, вентральные нервные стволы (проекция 20 × 0.5 мкм); Д – аналогичный участок туловищного отдела, серотонин-содержащие элементы в вентральных нервных стволах (проекция 20 × 0.5 мкм); е – аналогичный участок туловищного отдела, FMRFамид-содержащие элементы в вентральных нервных стволах (проекция 15 × 0.5 мкм). От нейропиля отходят 4 вентролатеральных ствола. На границе головного и туловищного отделов возле внутренних вентролате-ральных стволов и на уровне окологлоточного кольца выявляются тела одиночных клеток (стрелки на Б и В). При маркировании тубулина (А, Г) и серотонина (Б, Д) выявляются 7 продольных стволов: парные вентролатеральные (vl), промежуточные (imc), медиовентральные (mvc) и непарный медиальный ствол (ml). Видны также латеральный (lc) и дорсальный (d) стволы. При маркировании FMRFамида (В, е) выявляются 5 продольных стволов: парные вентролатеральные (vl), медиовентральные (mvc) и непарный медиальный ствол (ml). Видно также каудальное нервное кольцо (cnr). Остальные обозначения, как на рис. 1, 3, 4. Масштаб: А – 90; Б – 90; В – 70; Г, Д – 30; е – 40 мкм.

54 ФОФАНОВА И ДР.
БИОЛОГИЯ МОРЯ том 40 № 1 2014
термин "сегмент", а отмечаем лишь относительное рас-положение внешних и внутренних структур.
В ранних работах (Harmer, 1889; Schimkewitsch, 1895; Nelson, 1907; Shearer, 1912) у взрослых особей Dinophilus описывается крупный гантелевидный го-ловной мозг, занимающий практически всё простран-ство головного отдела. Авторы выделяют в нем два ганглия, между которыми находится нейропиль, вклю-чающий одну или две комиссуры. Аналогичная органи-зация головного ганглия была показана для карликово-го самца (Windoffer, Westheide, 1988a, b). Трехмерная электронно-микроскопическая реконструкция по серий-ным ультратонким срезам позволила выявить два го-ловных ганглия, соединенных толстой и тонкой комис-сурами (Windoffer, Westheide, 1988a). В более поздних исследованиях видов без полового диморфизма опи-сывается единый головной нейропиль округлой фор-мы (Беньяш и др., 1992; Müller, Westheide, 2002). Нам не удалось выделить подразделения головного отдела нервной системы на парные образования ни при вы-явлении ядер клеток, ни при маркировании нейропиля антителами к тубулину, ни при иммунохимическом мар-кировании 5-НТ-иммунореактивных или FMRFaмид-иммунореактивных нервных элементов. Нейропиль головного отдела имеет овальную форму во всех трех проекциях. При этом распределение отростков в нейро-пиле равномерное, они не формируют комиссуры. Тела клеток расположены вокруг области нейропиля также достаточно равномерно.
Парные ресничные поля, расположенные в голов-ном отделе латеродорсально перед первым туловищ-ным шнуром, были обнаружены у взрослых особей D. gyrociliatus (Müller, Westheide, 2002) и по аналогии с по-лихетами названы авторами нухальным органом, получа-ющим иннервацию от дорсокаудальной части головного ганглия. Нам не удалось выявить подобной иннервации аналогичных ресничных структур ни от каких участков нервной системы, что, возможно, связано с более ран-ним возрастом исследуемых животных.
Строение туловищного отдела нервной системы
В самых ранних исследованиях в туловищном от-деле динофилид описаны пять или шесть пар гангли-ев, располагавшихся вдоль двух мощных продольных нервных стволов, причем каждая пара ганглиев соеди-нялась двумя или тремя комиссурами (Harmer, 1889; Schimkewitsch, 1895; Nelson, 1907). Такое представле-ние вошло во все отечественные учебники. Однако ни в одной из более поздних работ при всем многообразии использованных методов не упоминается о наличии пар-ных туловищных ганглиев. Нами они также не обнару-жены.
При окрашивании на ацетилхолинестеразу опи-саны пять продольных стволов (Jägersten, 1944; Котикова, 1973), соединенных между собой неравно-
мерно расположенными комиссурами, число которых по данным разных авторов варьирует от четырех до семи. Кроме того, выявлена окологлоточная иннерва-ция (Donworth, 1986; Беньяш и др., 1992). В туловищ-ном отделе нервной системы карликового самца при электронно-микроскопических исследованиях серийных срезов обнаружены два крупных продольных ствола, соединенных семью комиссурами (Windoffer, Westheide, 1988a). При иммунохимическом окрашивании анти-телами против тубулина авторы насчитали пять про-дольных стволов, соединенных 18 комиссурами (Müller, Westheide, 2002).
Результаты наших исследований по обнаружению нервных элементов с помощью антител против тубу-лина и отдельных нервных клеток при мечении 5-НТ и FMRFамида больше всего сходны с данными Мюллер и Вестхайде (Müller, Westheide, 2002). При иммунохи-мическом выявлении серотонина в туловищном отде-ле видны многочисленные тела нейронов. Однако они расположены отдаленно от вентральных продольных стволов, непосредственно под ресничным эпителием вентральной полоски. К тому же тела клеток не обра-зуют выраженных скоплений. Тел нейронов, иммуно-реактивных к FMRFамиду, существенно меньше, чем 5-НТ-иммунореактивных, при этом в вентральной части тела они концентрируются около вентральных стволов в месте перехода головного отдела в туловищный. Таким образом, мы не можем выделить места концентрации нервных клеток, которые были бы организованы туло-вищными ганглиями.
Отмеченные у ювенильных особей семь продоль-ных стволов по форме и расположению сходны со ство-лами, обнаруженными у взрослых животных. Меньшее количество стволов у взрослых особей, скорее все-го, можно объяснить слиянием вентролатеральных и промежуточных стволов, которые в работе Мюллер и Вестхайде (Müller, Westheide, 2002) описаны как дуги, прилегающие к наиболее мощному латеральному ство-лу. В этом же стволе наиболее интенсивно представле-ны 5-НТ- и FMRFамид-иммунореактивные волокна. Во всех остальных стволах 5-НТ-иммунореактивные волокна представлены более или менее равномерно, а FMRFамид-иммунореактивные волокна выявляются только в пяти стволах (во всех, кроме промежуточных), при этом наиболее интенсивно они выражены в меди-альном продольном стволе.
Периферическая иннервация
Кроме латеральных стволов 5-НТ-иммунореактив-ные волокна находятся во всех кольцевых нервах, а так-же в обнаруженном нами дорсальном продольном ство-ле. Эти волокна расположены в поверхностных слоях эпителия, в непосредственной близости от ресничных шнуров. Такая организация 5-НТ-иммунореактивных волокон, а также выявленная концентрация тел 5-НТ-

РеСНИЧНые 55
БИОЛОГИЯ МОРЯ том 40 № 1 2014
иммунореактивных нейронов и интенсивная сеть во-локон с варикозными расширениями под вентральной ресничной полоской позволяют предположить, что у D. gyrociliatus серотонин вовлечен в иннервацию локо-моторных структур.
Обнаруженные нами FMRFамид-иммунореактив-ные волокна иннервируют в основном внутренние ор-ганы. Интересным представляется наличие каудального кольцевого волокна на уровне пятой латеральной бо-розды. Подобная структура была описана у карликово-го самца D. gyrociliatus (Windoffer, Westheide, 1988a, b) и у D. vorticoides (Беньяш и др., 1992). Логично, что у самца D. gyrociliatus данному волокну авторы припи-сали функцию иннервации пениса. Однако в случае D. vorticoides эта структура была выявлена у представи-телей обоих полов. В нашем исследовании каудальное кольцевое волокно обнаружено у самок D. gyrociliatus. Расположение волокна вокруг задней кишки указыва-ет, скорее, на его участие в иннервации соответствую-щего отдела пищеварительного тракта. Наличие тел FMRFамид-иммунореактивных нейронов и соответству-ющей иннервации глотки и пищеварительного канала позволяет предположить участие пептида FMRFамида в регуляции функционирования пищеварительной систе-мы у D. gyrociliatus.
Таким образом, выявленные нами черты строения нервной системы ювенильных самок D. gyrociliatus: еди-ный головной отдел округлой формы, множественные продольные вентральные стволы, соединенные комиссу-рами, рассредоточенные по периферии нервные клетки – больше напоминают строение нервной системы личинок аннелид, чем взрослых особей данной группы. Можно предположить, что такие черты упрощения связаны с пе-доморфозом, характерным для данного вида динофилид.
Авторы выражают глубокую благодарность ано-нимному рецензенту за ценные замечания, позволившие исправить недостатки рукописи. Мы также благодар-ны Ю.Б. Шмуклеру за привезенные с Неаполитанской зоологической станции пробы морской воды, в кото-рых были обнаружены динофилюсы, послужившие основой для создания лабораторной культуры. Работа была выполнена с использованием оборудования ЦКП Научно-образовательного центра ИБР РАН.
СПИСОК ЛИТеРАТУРы
Беклемишев В.Н. Основы сравнительной анатомии беспозво-ночных. М.: Советская наука. 1964. T. 1. 432 с.
Беньяш И.А., Ерлыкова Н.Н., Евдонин Л.А. К анатомии нерв-ной системы Dinophilus vorticoides // Многощетинковые черви и их экологическое значение. СПб.: Изд-во СПбГУ. 1992. C. 5–9.
Буруковский Р.Н. Зоология беспозвоночных. СПб.: Проспект науки. 2010. 960 с.
Вагин В.Л. Об аннелидной теории происхождения мизостомид, их положении в системе Trochozoa и выделение нового типа Myzostomida // Уч. зап. Казан. ун-та. 2007. Т. 149. С. 31–41.
Воронежская Е.Е., Ивашкин Е.Г. Пионерные нейроны: осно-ва или ограничивающий фактор разнообразия нервных систем Lophotrochozoa // Онтогенез. 2010. Т. 41, № 6. С. 403–413.
Иванов А.В., Петрушевский Г.К., Полянский Ю.И., Стрелков А.А. Большой практикум по зоологии беспозвоночных. Ч. I. Л.: Учпедгиз. 1941. 436 с.
Котикова Е.А. Новые данные о нервной системе Archiannelida // Зоол. журн. 1973. Т. 52, № 11. С. 1611–1615.
Мамкаев Ю.В., Селиванова Р.В. Исследования организации Dinophilus vorticoides // Тр. ЗИН АН СССР. 1979. Т. 84. С. 73–83.
Незлин Л.П. Золотой век сравнительной морфологии: лазерная сканирующая микроскопия и нейрогенез трохофорных животных // Онтогенез. 2010. Т. 41, № 5. С. 378–380.
Свешников В.А. Личинки архианнелид и полихет залива Посьет Японского моря // Исслед. фауны морей. 1967. Т. 5, № 13. С. 125–159.
Старобогатов Я.И. Брахицельная (гидроцельная) гипотеза происхождения целома // Сб. тр. ЗИН АН СССР. 1983. Т. 109. С. 111–122.
Фофанова Е., Воронежская Е. Содержание в культуре и вы-явление ресничных, нервных и мышечных элементов архианнелиды Dinophilus // XV Международ. конф. Ломоносов-2008. 2008. С. 16–17.
Akesson B. Parasite-host relationships and phylogenetic system-atics. The taxonomic position of dinophilids // Mikrofauna Meeresboden. 1977. Vol. 61. P. 19–28.
Donworth P.J. A reappraisal and validation of the species Dinophilus taeniatus Harmer, 1889 and of taxonomically significant fea-tures in monomorphic dinophilids (Annelida: Polychaeta) // Zool. Anz. 1986. Vol. 216. P. 32–38.
Fofanova E., Voronezhskaya E. The structure of archiannelid Dinophilus gyrociliatus ventral nerve cords // Acta. Biol. Hung. 2012. P. 88–90.
Franzen A. Ultrastructure of spermatids and spermatozoa in Archiannelida // Zoon. 1977. Vol. 5. P. 97–105.
Harmer S. Notes on the anatomy of Dinophilus // J. Mar. Biol. Assoc. U.K. 1889. Vol. 1, no. 2. P. 119–143.
Hartman O. Catalogue of the polychaetous annelids of the world // Allan Hancock Found. Publ. Occ. Pap. 1959. Vol. 23. P. 613–614.
Hermans C. The systematic position of the Archiannelida // Syst. Zool. 1969. Vol. 18. P. 85–102.
Jägersten G. Zur Kenntnis der Morphologie: Enzystierung und Taxonomie von Dinophilus // K. Sven. Vet. Akad. Handl. 1944. Bd. 21. S. 1–90.
Müller M.C.M., Westheide W. Comparative analysis of the nervous systems in presumptive progenetic dinophilid polychaetes (Annelida) by immunohistochemistry and cLSM // Acta Zool. 2002. Vol. 83. P. 33–48.
Nelson J.A. The morphology of Dinophilus conklini n. sp. // Proc. Acad. Natl. Sci. Phila. 1907. Vol. 59. P. 82–143.
Schimkewitsch W. Zur Kenntniss des Baues und der Entwicklung des Dinophilus vom Weissem Meere // Z. Wiss. Zool. 1895. Bd. 59. S. 46–79.
Shearer G. The problem of sex determination in Dinophilus gy-rociliatus: Part. 1 – The sexual cycle // Quart. J. Micr. Sci. 1912. Vol. 57, no. 3. P. 329–369.
Struck T. Progenetic species in polychaetes (Annelida) and prob-lems assessing their phylogenetic affiliation // Integr. Comp. Biol. 2006. Vol. 46, no. 4. P. 558–568.

56 ФОФАНОВА И ДР.
БИОЛОГИЯ МОРЯ том 40 № 1 2014
Struck T., Halanych K., Purschke G. Dinophilidae (Annelida) is most likely not a progenetic Eunicida: evidence from 18S and 28S rDNA // Mol. Phyl. Evol. 2005. Vol. 37. P. 619–623.
Voronezhskaya E.E., Tsitrin E.B., Nezlin L.P. Neuronal development in larval polychaete Phyllodoce maculata (Phyllodocidae) // J. Comp. Neurol. 2003. Vol. 455. P. 299–309.
Windoffer R., Westheide W. The nervous system of the male Dinophilus gyrociliatus (Polychaeta, Dinophilidae): electron microscopical reconstruction of the nervous anatomy and ef-fector cells // J. Comp. Neurol. 1988a. Vol. 272. P. 475–488.
Windoffer R., Westheide W. The nervous system of the male Dinophilus gyrociliatus (Polychaeta, Dinophilidae). I. Number, types and distribution pattern of sensory cells // Acta Zool. 1988b. Vol. 69, no. 1. P. 55–64.




















