
Flores-Nunes et al., 2014
13
MOLECULAR AND CELLULAR EFFECTS OF CONTAMINATION IN AQUATIC ECOSYSTEMS Changes in protein expression of pacific oyster Crassostrea gigas exposed in situ to urban sewage Fabrício Flores-Nunes & Tânia Gomes & Rui Company & Roberta R. M. Moraes & Silvio T. Sasaki & Satie Taniguchi & Márcia C. Bicego & Cláudio M. R. Melo & Afonso C. D. Bainy & Maria J. Bebianno Received: 15 August 2014 /Accepted: 4 November 2014 # Springer-Verlag Berlin Heidelberg 2014 Abstract The composition and concentration of substances in urban effluents are complex and difficult to measure. These contaminants elicit biological responses in the exposed organ- isms. Proteomic analysis is a powerful tool in environmental toxicology by evidencing alterations in protein expression due to exposure to contaminants and by providing a useful frame- work for the development of new potential biomarkers. The aim of this study was to determine changes in protein expres- sion signatures (PES) in the digestive gland of oysters Crassostrea gigas transplanted to two farming areas (LIS and RIB) and to one area contaminated by sanitary sewage (BUC) after 14 days of exposure. This species is one of the most cultivated molluscs in the world. The identified proteins are related to the cytoskeleton (CKAP5 and ACT2), ubiquitination pathway conjugation (UBE3C), G protein- coupled receptor and signal transduction (SVEP1), and cell cycle/division (CCNB3). CKAP5 showed higher expression in oysters kept at BUC in comparison with those kept at the farming areas, while ACT2, UBE3C, SVEP1, and CCNB3 were suppressed. The results suggest that these changes might lead to DNA damage, apoptosis, and interference with the immune system in oyster C. gigas exposed to sewage and give initial information on PES of C. gigas exposed to sanitary sewage, which can subsequently be useful in the development of more sensitive tools for biomonitoring coastal areas, par- ticularly those devoted mainly to oyster farming activities. Keywords Urban sewage . Crassostrea gigas . Proteomic analysis . Two-dimensional gel electrophoresis Introduction Urban sewage is one of the major sources of contaminants into the marine and estuarine ecosystems, causing adverse effects in target tissues of exposed organisms, which may reflect changes in the population and community structures (Abessa et al. 2005; Kennish 1992; Martins et al. 2008). Periodically, complex mixtures of solids, nutrients, hydrocarbons, metals, pesticides, pharmaceuticals, and personal care products from sanitary, hospital, gas stations, small industries, and runoffs are discharged in these areas (Abessa et al. 2005; Bolong et al. 2009; USEPA 2009). Many ecotoxicological studies in marine and estuarine environments use bivalve molluscs as sentinel organisms since they are sessile, euryhaline, show wide geographical distribution, and possess filter feeding habits with great ca- pacity to accumulate contaminants (Cajaraville et al. 2000; Beeby 2001 ). Among bivalve molluscs, the oyster Crassostrea gigas holds a prominent position for being the most cultivated, consumed, and studied marine mollusc in the world (Mao et al. 2006; Saavedra and Bachère 2006). In southern Brazil, as well as in other countries around the world, Responsible editor: Philippe Garrigues Electronic supplementary material The online version of this article (doi:10.1007/s11356-014-3821-8) contains supplementary material, which is available to authorized users. F. Flores-Nunes : R. R. M. Moraes : A. C. D. Bainy Laboratory for Biomarkers of Aquatic Contamination and Immunochemistry, Federal University Santa Catarina, Florianópolis, Brazil T. Gomes : R. Company : M. J. Bebianno (*) CIMA, Faculty of Science and Technology, University of Algarve, Campus de Gambelas, 8005-139 Faro, Portugal e-mail: [email protected] S. T. Sasaki : S. Taniguchi : M. C. Bicego Laboratory of Marine Organic Chemistry, Oceanographic Institute, University of São Paulo, São Paulo, Brazil C. M. R. Melo Laboratory of Marine Molluscs, Federal University of Santa Catarina, Florianópolis, Brazil Environ Sci Pollut Res DOI 10.1007/s11356-014-3821-8
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Flores-Nunes et al., 2014

MOLECULAR AND CELLULAR EFFECTS OF CONTAMINATION IN AQUATIC ECOSYSTEMS
Changes in protein expression of pacific oyster Crassostreagigas exposed in situ to urban sewage
Fabrício Flores-Nunes & Tânia Gomes & Rui Company &
Roberta R. M. Moraes & Silvio T. Sasaki & Satie Taniguchi &Márcia C. Bicego & Cláudio M. R. Melo & Afonso C. D. Bainy & Maria J. Bebianno
Received: 15 August 2014 /Accepted: 4 November 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract The composition and concentration of substancesin urban effluents are complex and difficult to measure. Thesecontaminants elicit biological responses in the exposed organ-isms. Proteomic analysis is a powerful tool in environmentaltoxicology by evidencing alterations in protein expression dueto exposure to contaminants and by providing a useful frame-work for the development of new potential biomarkers. Theaim of this study was to determine changes in protein expres-sion signatures (PES) in the digestive gland of oystersCrassostrea gigas transplanted to two farming areas (LISand RIB) and to one area contaminated by sanitary sewage(BUC) after 14 days of exposure. This species is one of themost cultivated molluscs in the world. The identified proteinsare related to the cytoskeleton (CKAP5 and ACT2),ubiquitination pathway conjugation (UBE3C), G protein-coupled receptor and signal transduction (SVEP1), and cellcycle/division (CCNB3). CKAP5 showed higher expression
in oysters kept at BUC in comparison with those kept at thefarming areas, while ACT2, UBE3C, SVEP1, and CCNB3were suppressed. The results suggest that these changes mightlead to DNA damage, apoptosis, and interference with theimmune system in oysterC. gigas exposed to sewage and giveinitial information on PES of C. gigas exposed to sanitarysewage, which can subsequently be useful in the developmentof more sensitive tools for biomonitoring coastal areas, par-ticularly those devoted mainly to oyster farming activities.
Keywords Urban sewage .Crassostrea gigas . Proteomicanalysis . Two-dimensional gel electrophoresis
Introduction
Urban sewage is one of the major sources of contaminants intothe marine and estuarine ecosystems, causing adverse effectsin target tissues of exposed organisms, which may reflectchanges in the population and community structures (Abessaet al. 2005; Kennish 1992; Martins et al. 2008). Periodically,complex mixtures of solids, nutrients, hydrocarbons, metals,pesticides, pharmaceuticals, and personal care products fromsanitary, hospital, gas stations, small industries, and runoffsare discharged in these areas (Abessa et al. 2005; Bolong et al.2009; USEPA 2009).
Many ecotoxicological studies in marine and estuarineenvironments use bivalve molluscs as sentinel organismssince they are sessile, euryhaline, show wide geographicaldistribution, and possess filter feeding habits with great ca-pacity to accumulate contaminants (Cajaraville et al. 2000;Beeby 2001). Among bivalve molluscs, the oysterCrassostrea gigas holds a prominent position for being themost cultivated, consumed, and studied marine mollusc in theworld (Mao et al. 2006; Saavedra and Bachère 2006). Insouthern Brazil, as well as in other countries around the world,
Responsible editor: Philippe Garrigues
Electronic supplementary material The online version of this article(doi:10.1007/s11356-014-3821-8) contains supplementary material,which is available to authorized users.
F. Flores-Nunes :R. R. M. Moraes :A. C. D. BainyLaboratory for Biomarkers of Aquatic Contamination andImmunochemistry, Federal University Santa Catarina, Florianópolis,Brazil
T. Gomes : R. Company :M. J. Bebianno (*)CIMA, Faculty of Science and Technology, University of Algarve,Campus de Gambelas, 8005-139 Faro, Portugale-mail: [email protected]
S. T. Sasaki : S. Taniguchi :M. C. BicegoLaboratory of Marine Organic Chemistry, Oceanographic Institute,University of São Paulo, São Paulo, Brazil
C. M. R. MeloLaboratory of Marine Molluscs, Federal University of SantaCatarina, Florianópolis, Brazil
Environ Sci Pollut ResDOI 10.1007/s11356-014-3821-8

this species was introduced and has been cultivated intensive-ly (EPAGRI 2013), which points to the need of developingtools for monitoring the levels and effects of contaminants inoysters kept in farming zones or other more impacted areas.
Contaminants may activate effector cell receptors, which,in turn, interact with specific DNA sequences promoting thetranscription of multiple genes. Multiple messenger RNAs(mRNAs) are processed and sent to the cytoplasm for proteinsynthesis, thus constituting the cellular response (Piña et al.2007). Therefore, genomic and proteomic techniques are im-portant tools for assessing the global profile of molecular andbiological responses in exposed animals and can help in boththe development of oyster farming industries (Saavedra andBachère 2006) and in programs of ecological risk assessment(Kling et al. 2008). Although mRNA sequences could be usedto predict the biological processes, proteins are performers ofthe actual cell response (Rudert et al. 2000). In this context,proteomic analysis reflects the change in protein expressionby environmental adaptations induced by pollutants and/orpathological processes (López 2005; Moore 2002) and there-fore enables the development of new tools for biomonitoring(Thompson et al. 2012a) to complement transcriptomic stud-ies (López 2005; Pandey and Mann 2000).
Several methodologies have been used for proteomic anal-ysis, but the two-dimensional electrophoresis (2DE) on poly-acrylamide gels is the most established and used tool (Lemoset al. 2010). The identification of induced and/or inhibitedproteins is one way to identify exposure-effect relationshipsassociated to certain stressors in order to propose molecularmechanisms of response (Nesatyy and Suter 2007).“Proteomic maps,” also known as protein expression profiles(PEPs), are groups of specific proteins differentiallyexpressed, reflecting a cellular response elicited by a chal-lenge (Bradley et al. 2002) that can be related to a stressor, aswell as recognize potential protein-protein interactions(Rudert et al. 2000) without the need for the identification ofproteins (Apraiz et al. 2009; Shepard et al. 2000).
Advances in techniques of protein separation and identifi-cation have expanded the use of proteomics for application toa large number of sentinel organisms (Veldhoen et al. 2012),providing a better relationship between environmental stressand pollutant toxicity (Campos et al. 2012). This is speciallyimportant in the case of bivalve molluscs used as edibleresources as oysters C. gigas. The use of proteins as bio-markers has been applied to mollusc farming (López et al.2002) and for evaluating different environmental impacts suchas exposure to toxic algae (Ronzitti et al. 2008), to protozoalinfections (Cao et al. 2009), to organic contaminants (Apraizet al. 2009; Monsinjon et al. 2006; Olsson et al. 2004), tourban effluents (Amelina et al. 2007), to trace metals (Choraet al. 2008; Chora et al. 2009b; Shepard and Bradley 2000;Thompson et al. 2012a), and to nanoparticles (Gomes et al.2013).
Considering the social and economic importance of bivalveaquaculture in coastal zones near the urban areas, the under-standing of the impact of sanitary sewage in such areas is stilllimited. The aim of this study was to evaluate the PEPs in thedigestive gland of C. gigas exposed in the field to urbansanitary sewage discharges.
Materials and methods
Experimental design
Oysters C. gigas (6–8 cm) were collected in the farming areaof the Laboratory of Marine Molluscs (LMM) of the FederalUniversity of Santa Catarina (UFSC) at the Sambaqui beach(SAM; Florianópolis, SC) and transplanted to two oysterfarming areas (LIS: Santo Antonio de Lisboa beach, NorthBay; RIB: Ribeirão da Ilha beach, South Bay) and to an areaimpacted by discharges of urban sanitary sewage used as acontaminated site in this study (BUC: Bücheler river mouth)(Fig. 1).
After 14 days of exposure, oysters were collected at eachsite (n=10), digestive glands were dissected, immediatelyfrozen in liquid nitrogen, and stored separately at −80 °C.
Analysis of fecal coliforms, detergents, organic matter, oil,and grease in water samples
Water samples were collected at 10 cm depth at the beginning(0 h), 24 h, and 14 days of exposure in all sites. Water analysiswas performed in the same sampling day by counting coliformbacteria (fecal coliforms, CF), detergents (DTG), organic mat-ter (OM), and oil and grease (O/G) according to standardprotocols (Clesceri et al. 1998).
Analysis of organic contaminants in sediments and oysters
Oysters and sediment samples (one pool of whole oystertissues, n=8, and 250 g of sediments) were lyophilized for72 h (Thermo Savant, module D), macerated, and homoge-nized in mortar with pestle, and stored in glass bottles previ-ously cleaned with solvent.
The sediment and tissue samples were freeze-dried andhomogenized. An amount of, respectively, 20 and 1 g wasSoxhlet-extracted with a 50 % mixture of residue grade n-hexane and dichloromethane for 8 h in accordancewith UNEP(1992) and McLeod et al. (1986), with some minor modifica-tion. Before extraction, PCB 103, PCB 198, n-hexadecene, n-eicosene, naphthalene-d8, acenaphthene-d10, phenanthrene-d10, chrysene-d12, perylene-d12, and dodecyl 1-benzene(1C12LAB) were added to all the samples, blanks, and refer-ence material from the National Institute of Standards andTechnology (NIST) (SRM 1944 for sediment and SRM
Environ Sci Pollut Res

2974 for oyster) as surrogates. The organochlorine extract waspurified in an alumina chromatographic column. The hydro-carbon extracts were fractionated into F1 (aliphatics and linearalkylbenzenes), F2 (polycyclic aromatic hydrocarbons), andF3 (only for sediment, steroids) by silica gel-alumina columnchromatography. Fraction 3 was evaporated to dryness, andsteroids were derivatized to form trimethylsilyl ethers usingbis(trimethylsilyl) trifluoroacetamide (BSTFA) with 1 %trimethylchlorosilane (TMCS) for 90 min at 65 °C.Organochlorine pesticides were analyzed in an AgilentTechnologies gas chromatograph in splitless mode using anelectron capture detector (GC-ECD). Aliphatic hydrocarbonsand steroids were determined on a gas chromatography withflame ionization detector (GC-FID). PAHs, PCBs, and LABs
were analyzed by anAgilent 6890 gas chromatograph coupledto a 5973N mass spectrometer (GC/MS) in a selected ionmode (SIM). Certified standards at five different concentra-tions were used for calculations. PAH, PCBs, and LABsidentification was based onmass/charge ratio of the individualquantitation ion (m/z) and retention times of certifiedstandards.
Proteomic analysis
The sample preparation as well as the protocol used for two-dimensional electrophoresis is described by Chora et al.(2009a). For each study site (LIS, RIB, and BUC), five poolswere prepared using two digestive glands each (n=5)
Fig. 1 Sites used for oysterstransplantation in the region ofFlorianópolis. 1 SAM: Sambaquibeach, LMM/UFSC (oysterssupplier); 2 LIS: Santo Antoniode Lisboa beach, North bay oysterfarming area, North bay; 3 RIB:Ribeirão da Ilha beach, South bayoyster farming area, South bay; 4BUC: Bücheler river mouth,contaminated site
Environ Sci Pollut Res

suspended in a specific buffer (10 mM HEPES and 250 mMsucrose solution) containing 1 mMDTT, 1 mM EDTA, 1 mMPMSF, 10 % of protease inhibitor (protease inhibitor CocktailSigma P8340), and homogenized using an Ultra-Turrax ho-mogenizer IKA-Werke on an ice bath at 4 ° C in a 3:1 (buffer/tissue). The resulting homogenate was centrifuged at15,000×g for 2 h (4 °C), and the supernatant was stored at−80 °C for later use. The quantitation of total proteins wascarried out according to the Bradford method using bovineserum albumin (BSA) as standard (Bradford, 1976). Aliquotsof 150 μg of proteins were suspended in protein precipitatingsolution (1:9; 10 % trichloroacetic acid in cold acetone con-taining 20 mM DTT) for 2 h at −20 °C, centrifuged(10,000×g; 30 min; 4 ° C), the supernatant discarded, andthe pellet washed three times with cold acetone. The residualacetone was removed by air-drying.
Two-dimensional electrophoresis
Proteins were first separated by isoelectric focusing (IEF; firstdimension), followed by separation by molecular weight bytwo-dimensional electrophoresis (2-DE). Each sample con-taining 150 μg of protein was incubated for 30 min in300 μl of lysis buffer (7 M urea, 2 M thiourea, 4 % CHAPS,0.8 % Pharmalyte, 65 mM DTT, and traces of bromophenolblue), centrifuged (14,000×g, 10 min, 4 °C), and applied overa GE Immobiline DryStrip, pH 4–7, 18 cm. After 6 h ofpassive rehydration, followed by 6 h of active rehydration(50 V), the IEF was performed (20 °C and 50 A per strip) in aProtean® IEF Cell (Bio-Rad, CA, USA) following a five stepprogram: 1000 V for 1 h, 4000 V for 1 h, 8000 V, and 8000 Vfor 1 h for 5 h to reach a total of 50,000 Vh.
Before 2-DE, strips were equilibrated in buffer (6 M urea,75 mM Tris, pH 8.8, 4 % SDS, and 29.3 % glycerol) contain-ing 2 % DTT for 15 min, followed by further immersion for15 min in buffer containing 2.5 % iodoacetamide. The 2-DEwas performed in 10 % polyacrylamide gels using the verticalsystem of Protean®XLCell Format Cell (Bio-Rad, CA, USA)at 20 °C in two steps: 90 V for 30 min for a gradual entry ofthe sample into the top of the gel and 300 V until the samplewent through the whole gel (∼5 h).
The gels were silver-stained in compliance with a protocolcompatible with mass spectrometry analysis (Blum et al.1987). To ensure reproducibility, four gels were used forimage analysis.
Acquisition and image analysis
After silver-staining, the gels run with samples from each sitewere scanned using a densitometer (GS-800, Bio-Rad, CA,USA). Data analysis and statistical tests were performed usingPDQuest 8.0 software (Bio-Rad, CA, USA). A master gel wasconstructed by combining the 2-DE maps (n=4 gels), where
the 2-DE maps from the oyster farming areas (LIS and RIB),used as reference sites, were overlaid with the 2-DEmap fromthe contaminated site (BUC).
The spot intensity normalization was carried out inorder to enable an accurate comparison among the gels,where the volume of each protein spot was divided by thetotal volume of all detected spots in each image. Thenormalized volumes were compared with the correspond-ing values of the reference gels (calibration). The numberof protein spots was determined for each valid gel, andthe qualitative and quantitative differences in protein pat-terns between the farming and contaminated sites weredetermined. After image analysis, the protein spots with ahigher or lower expression than those from the referencegroup were selected for further sequencing.
Digestion of spots and protein identification by massspectrometry
Proteins of oysters exposed in situ to urban sanitary sewagewere selected and removed manually from silver-stained gels,digested with trypsin (Shevchenko et al. 2006), and submittedto “peptide mass fingerprint” (PMF) for identification of pep-tides and acquisition of mass spectra using the Ultraflex IIMALDI-TOF-TOF (Bruker Daltonics) operating at positivepolarity mode. The reflection spectra were acquired in therange m/z 900–3500. A total number of 3000 spectra wereobtained at each dot position with a frequency of 50 Hz laser.For the experiments, MS/MS ion peptides with S/N higherthan 25 and peak intensity higher than 800 were selected forMS/MS. Laser shots (300 and 1000) were used to acquire theMS and MS/MS, respectively. The laser power was 2 to 5 %above the threshold of ionization.
Data acquisition and processing were performed using thesoftware FlexAnalysis 3.0 (Bruker Daltonics) with the peakdetection algorithm SNAP. The obtained peptide mass list wassent to the softwareMASCOTusing NCBI database. Searcheswere conducted using the following parameters: taxonomy,other metazoans; proteolytic enzyme, trypsin peptides toler-ance, 100 ppm; fixed modifications, carbamidomethyl (C),variable modification, oxidation (M); peptide charge state, +1, missed cleavages allowed until 1. The limit of significancewas set at 95 %. MS BLAST searches (NCBI / Blastp) wereperformed on a database of non-redundant protein sequencesavailable for C. gigas (taxid: 29159) using protein-proteinBLAST algorithm (default parameters).
Statistical analysis
Differences among the levels of protein expression in diges-tive gland of oysters from different sites were compared using
Environ Sci Pollut Res
nonparametric tests, namelyMann-WhitneyU rank. The levelof significance was set at 0.05. The results were analyzedusing GraphPad Prism ® 5.01 and XLSTAT (ExcelWindows) software.
Results
Water quality
The levels of fecal coliforms, detergents, organic matter, andoil and grease were detected in the water samples of BUC siteat the beginning of the exposure and after 24 h and 14 days ofexposure evidence that this area is contaminated by sanitarysewage. SAM, RIB, and LIS sites showed undetected or verylow levels of these contaminants during the exposure period(Suppl. Fig. 1).
Organic contaminants in sediments
Sediment analysis showed that the level of ΣDDT {sumof the p,p′- and o,p-isomers of DDT [1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane], DDE [1,1-dichloro-2,2-bis(p-chlorophenyl)ethylene], and DDD [1,1-dichloro-2,2-b i s ( p - ch l o r opheny l ) e t h ane ] } c augh t a t SAM(=11.9 ng g−1) was 3–7 times higher when comparedto other sites (RIB, 1.6; LIS, 3.8; BUC, 2.9 ng g−1)(Suppl. Fig. 2). In contrast, the content of total aliphatichydrocarbons (ΣAH) in sediments sampled at BUC siteshowed higher levels (52.6 ng g−1) than in the othersites (LIS, 18.4; SAM, 10.0; RIB, 3.67 ng g−1). Total
linear alkylbenzenes (ΣLAB) were also significantlyhigher in BUC than in the other sites (BUC, 108.0;RIB, 12.1; SAM, 1.56 ng g−1; LIS: below detectionlimit). Lower levels of total PAHs were observed insediments from RIB (14.5 ng g−1) and higher levels inBUC (112.1 ng g−1), while LIS and SAM showedsimilar values (96.9 and 88.2 ng.g−1, respectively).
Coprostanol, a biomarker for the presence of humanfecal matter in the environment, was detected in sedimentsof BUC, with levels 125-fold higher (2.55 μg g−1) than inthe sediments caught at the other sites (SAM, 0.02; RIB,0.08; LIS, 0.05 μg g−1) (Suppl. Fig. 3a). The ratio betweenfecal sterols used as indicative of contamination by sanitarysewage (Grimalt et al. 1990; Mudge and Norris, 1997)observed in sediment from BUC was higher than the othersites (Suppl. Fig. 3b).
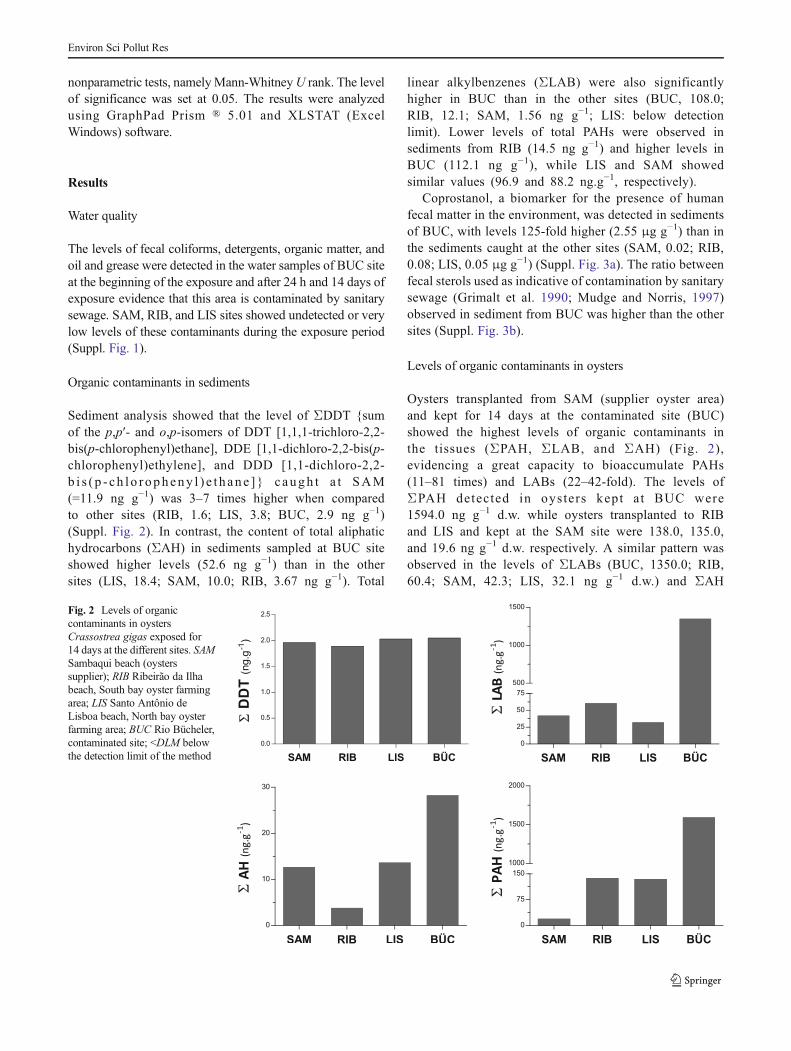
Levels of organic contaminants in oysters
Oysters transplanted from SAM (supplier oyster area)and kept for 14 days at the contaminated site (BUC)showed the highest levels of organic contaminants inthe tissues (ΣPAH, ΣLAB, and ΣAH) (Fig. 2),evidencing a great capacity to bioaccumulate PAHs(11–81 times) and LABs (22–42-fold). The levels ofΣPAH detected in oysters kept a t BUC were1594.0 ng g−1 d.w. while oysters transplanted to RIBand LIS and kept at the SAM site were 138.0, 135.0,and 19.6 ng g−1 d.w. respectively. A similar pattern wasobserved in the levels of ΣLABs (BUC, 1350.0; RIB,60.4; SAM, 42.3; LIS, 32.1 ng g−1 d.w.) and ΣAH
Fig. 2 Levels of organiccontaminants in oystersCrassostrea gigas exposed for14 days at the different sites. SAMSambaqui beach (oysterssupplier); RIB Ribeirão da Ilhabeach, South bay oyster farmingarea; LIS Santo Antônio deLisboa beach, North bay oysterfarming area; BUC Rio Bücheler,contaminated site; <DLM belowthe detection limit of the method
Environ Sci Pollut Res

(BUC, 28.3; LIS, 13.7; SAM, 12.7; RIB, 3.84 ng g−1).In terms of ΣDDT concentrations, the levels in oysters fromBUC (2.05 ng g−1) and LIS (2.03 ng g−1) were similar tolevels detected in animals from the initial of the experiment(SAM, 1.96 ng g−1), while oysters kept in RIB showed unde-tected levels (Fig. 2).
Protein expression profiles
PEPs were obtained by comparing samples of digestivegland from oysters C. gigas kept at sites containingsanitary sewage effluents (BUC) with samples fromoysters from mollusc farming areas (BUC×LIS; BUC×RIB) (Figs. 3 and 4). Data analysis of the PEPs obtain-ed for all sites showed a mean of 525±24 protein spots(matched to three and four gels) in a total of approxi-mately 1000 proteins. Among the detected spots, 118±18 is unique to the oysters kept at BUC. Comparing thePEPs, 257 and 280 protein spots statistically differ,respectively, between BUC×LIS and BUC×RIB and109 spots were common between all sites (p<0.05)(Table 1).
PEPs from the oyster digestive gland from the con-taminated site were compared to those from the twofarming areas to identify protein expression signatures(PESs) (BUC×LIS and BUC×RIB). Among the 131protein spots detected in LIS and the 106 spots in
RIB, 57 spots were also present in BUC, indicatingpossible PESs common among farmed and contaminatedorganisms (Fig. 5a). Among the differential proteinspots detected in LIS and RIB compared to BUC, 30spots are common to the two comparisons, where 13proteins were 2-fold downregulated (−2× spots) and 17proteins 2-fold upregulated (+2× spots) (Fig. 5b, c).
Protein identification
Of the 30 spots selected for identification by massspectrometry, six showed homology to proteins identi-fied and available at the NCBI database (http://blast.ncbi.nlm.nih.gov), one of which was actin, selected asa positive control for bivalve sequencing. Theidentif ied proteins are related to the differentbiological functions, namely cytoskeleton (CKAP5 andACT2), ubiquitination/conjugation pathway (UBE3C),G protein-coupled receptor/signal transduction pathway(SVEP1), and cell cycle/cell division (CCNB3)(Table 2; Figs. 3 and 4).
Discussion
A proteomic approach is an important tool for providingprospects of unknown responses to environmental stress, such
Mw kDa
250
20
pH 4 7
25
37
50
75
100
150
Fig 3 Master gel representativeof PESs up-regulated commonbetween BUC×RIB and BUC×LIS. Proteins identified byMALDI–TOF–TOF arehighlighted. One hundred fiftymicrograms of protein contentwas separated on 18 cm IPG stripsin pH 4–7 gradients. The seconddimension was performed in 10%SDS–PAGE gels. Molecularweight and pH are indicated in thefigure
Environ Sci Pollut Res

as the impact of urban wastewater at the molecular phenotypelevel, by quantifying changes in protein expression that re-flects changes in protein synthesis, post-translation, or degra-dation (Tomanek 2011). Up to the present, there are no reportsof protein changes in response to in situ exposure to sanitarysewage effluents in bivalves. Therefore, the present studyaims to analyze proteomic changes in the digestive gland ofoysters C. gigas exposed to urban sanitary sewage dischargesand to compare with those kept at farming areas.
The levels of FC, DTG, O/G, and OM in water and PAHs,AHs, LABs, and fecal sterols in sediments confirmed thatBUC are highly contaminated, especially by sanitary sewagedischarges (Suppl. Fig. 1, Fig. 2, and Fig. 3). This was also
confirmed by the levels of PAHs (11–81-fold) and LABs(22–42-fold) accumulated in the oysters C. gigas comparedwith those from the two farming areas (LIS and RIB) (Fig. 2).
Previous studies carried out with C. gigas kept in this samecontaminated site showed that sanitary sewage effluentscaused changes in the activity of antioxidant enzymes(Souza et al. 2012; Zanette et al. 2008) and gene transcription(Medeiros et al. 2008). However, no information is availableabout the effects of these mixtures on protein expression inthis species.
Five differentially expressed proteins were identified in theoysters exposed in situ to sanitary sewage discharges. Theidentified proteins are related to the cytoskeleton (CKAP5 and
pH 4 7
Mw kDa
250
20
25
37
50
75
100
150
Fig. 4 Master gel representativeof PES down-regulated commonbetween BUC×RIB and BUC×LIS. Proteins identified byMALDI–TOF–TOF arehighlighted. One hundred fiftymicrograms of protein contentwas separated on 18 cm IPG stripsin pH 4–7 gradients. The seconddimension was performed in 10%SDS–PAGE gels. Molecularweight and pH are indicated in thefigure
Table 1 Number of protein spots in oysters kept in the contaminated site versus farming areas
Local/experiment Spot gels average Prot only (n) Prot stat dif (p<0,05) 2-fold common spots (n)
↓ ↑
BUC vs. LIS 539 131 257 51 74489 70
Common betweenexperiments
109 13 17
BUC vs. RIB 539 534 280 74 49106 101
BUC Bücheler river mouth (contaminated site), LIS Santo Antonio de Lisboa beach (North bay oyster farming area), RIB Ribeirão da Ilha Beach (Southbay oyster farming area), ↓ expression levels downregulated less than 2-fold, ↑ expression levels upregulated higher than 2-fold
Environ Sci Pollut Res

ACT2), ubiquitination and conjugation pathway (UBE3C), Gprotein-coupled receptor and signal transduction (SVEP1),and cell cycle/cell division (CCNB3). Only the proteinCKAP5 showed higher expression in oysters from the con-
taminated site, while ACT2, UBE3C, SVEP1, and CCNB3had lower expression in this group (Table 2).
The cytoskeleton proteins showed the highest and thelowest change in expression in the contaminated oysters(Table 2). CKAP5 (spot 0209; Table 2; Fig. 3) showed a 13/2-fold up-regulated, and ACT2 (spot 2403; Table 2; Fig. 4)showed 2/3-fold down-regulation in oysters exposed to sani-tary sewage effluents at BUC site when compared to thefarming areas (LIS and RIB), respectively. As xenobioticsenter the cells through the cell membrane, cytoskeleton-related proteins might serve as biomarkers of exposure and/or susceptibility to environmental contaminants (Gómez-Mendikute et al. 2002). The inhibition of proteins related tothe cytoskeleton has been previously reported in bivalvesexposed to different contaminants, such as metals andpolychlorinated biphenyls (Apraiz et al. 2006; Rodríguez-Ortega et al. 2003). Although this response indicates damageto proteins, the unique identification of cytoskeletal proteinsmay reflect their abundance/prevalence in databases or eventheir role as major targets of oxidative stress (Amelina et al.2007; Dalle-Donne et al. 2001; Rodríguez-Ortega et al. 2003;Thompson et al. 2012b; Tomanek et al. 2011). However, thereis no information concerning the role of the cytoskeletalprotein CKAP5 in bivalves, but its function may be associatedwith cellular challenges endured by oysters against the expo-sure to contaminants associated with urban effluent disposal.These proteins are presented on the microtubules that form thecytoskeleton and have interactions with cyclin B, an essentialprotein for the cell cycle regulation, especially at the mitoticphase (Basto et al. 2007; Charrasse et al. 2000; Chen et al.2009; Kinoshita et al. 2002). In terrestrial vertebrates, CKAP5
Fig. 5 Venn diagram with specific and common proteins spots withstatistical difference (p<0.05) between contaminated×farming (a),Venn diagram with number of proteins of interest down-regulated (b)and up-regulated (c) common between BUC×RIB and BUC×LIS. BUCBücheler river mouth, contaminated site; LIS Santo Antonio de Lisboabeach, North bay oyster farming area; RIB Ribeirão da Ilha beach, Southbay oyster farming area
Table 2 Identification of differentially expressed proteins by MALDI-TOF-TOF mass spectrometry in Crassostrea gigas exposed to sanitary sewage
Spot no.a name PDQuest Accessionnumber
TheoricMr (Da)/pI
Protein identification Function
Av. ratiob MALDI-TOF-TOF
Scorec % Coverage Matchedpeptides
209 Cytoskeleton-associated protein 5
13/2 ↑ gi|405971153 199.7/8.73 1041 99 14 Cytoskeleton
2403 Actin 2 2/3 ↓ gi|18565105 41.7/5.3 702 100 10 Cytoskeleton
5104 Ubiquitin-proteinligase E3C
4/8 ↓ gi|405973722 122.8/8.46 807 94 5 Ubiquitinationconjugation pathway
5506 Sushi, von Willebrandfactor type A, EGF andpentraxin domain-containing protein 1
2/5 ↓ gi|405974932 48.6/ 6.86 318 51 5 Cell cycle/cell divison
– Actin – gi|3182893 41.7/5.3 759 100 10 Cytoskeleton
a Spot number on 2DE master gel (Figs. 3 and 4)b Fold change increase (↑) or decrease (↓) in terms of intensity between the oysters exposed to BUC×LIS and BUC×RIB. Average ratio calculated byPDQuest using four replicates in each group. For all comparisons, the p value is <0.05c Scores of thematches usingMASCOT (http://www.matrixscience.com) andNCBI/BLAST (http://blast.ncbi.nlm.nih.gov), percentage of coverage, andnumber of matched peptides in the identified proteins
Environ Sci Pollut Res

are microtubule-associated proteins (MAPs) responsible forencoding proteins in human tumor-expressed gene (TOG),while in frog, Xenopus spp. are XMAP215 homologues(Cassimeris and Morabito 2004; Francone et al. 2007).These proteins are related to cell dysfunction in tumor cells(Bogoch and Linial 2008; Conte et al. 2003; Kinoshita et al.2002; Paulsen et al. 2009; Røe et al. 2010).
Actin is a ubiquitous cytosolic protein with vital functionsin the cytoskeleton and is one of the initial antioxidant pro-tection mechanisms against glutathionylation and carbonyla-tion of proteins (Dalle-Donne et al. 2003; McDonagh et al.2005). Glutathionylation comes from reversible disulfidebridge formation between GSH and protein-SH groups as astrategy for protecting proteins during oxidative stress (e.g.,protection of thiol group of cysteine) (Dalle-Donne et al.2003; McDonagh and Sheehan 2006). Moreover, thiol groups(sulphenic acid, nitrosothiols, disulphides, and mixeddisulphides) in many proteins are redox-sensitive and theirredox state often determines protein activities and function(Wang et al. 2012). However, in the digestive gland of mus-sels, the glutathionylation of actin is unlikely to occur (Dalle-Donne et al. 2003), as this tissue presents a high ratio of theredox system GSH/GSSG (McDonagh and Sheehan 2006).
Under high ROS production levels and/or strong jeopar-dizing of antioxidant defenses, the oxidation of reactive/accessible thiol groups can alter protein function reversiblyor irreversibly. Reversible oxidation protects the protein fromirreversible oxidation and/or acts as a molecular switch forregulation of protein function, whereas irreversible proteinoxidation often leads to permanent loss of function (Wanget al. 2012). Protein modifications are usually associated withirreversible permanent loss of function and can lead tothe elimination of damaged proteins by their accumulationin proteasome system or in the form of insoluble aggregates(Dalle-Donne et al. 2007).
The actin is arranged in such a way that some of its aminoacid targets are very sensitive to oxidation, making it one ofthe most susceptible proteins to carbonylation under oxidativestress conditions (Castro et al. 2013), particularly in the case ofdigestive gland of bivalves, where the protein carbonylation iselevated (Dowling et al. 2006). Carbonylation changes thefunction of this protein, promoting its aggregation which maylead to proteosomal degradation preventing its accumulation(Castro et al. 2013). Thus, the down-regulation of ACT2 in thedigestive gland of oysters exposed to sanitary sewage may berelated to the carbonylation of this protein that could havebeen denatured or degraded with loss of function andidentification.
The protein proteosomal degradation is a non-lysosomalmechanism mainly for recycling essential amino acids in cells(Jung et al. 2009). This complex has a catalytic function withtwo predominant forms: the catalytic core 20S (marking ATPand ubiquitin independent) and 26S form (ATP-dependent
and ubiquitination) (Jung et al. 2009). The 26S proteasomegenerally requires the previous action of ubiquitin(polyubiquitination) that consists of three types of enzymes:E1 ubiquitin-activating enzyme, E2 ubiquitin-conjugating en-zyme, and E3 ubiquitin ligases (Castro et al. 2013; Pickart2004). The E3 ubiquitin ligases are enzymes that modifyproteins post-translationally by binding to a single ubiquitin(monoubiqu i t ina t ion) or a cha in of ub iqu i t ins(polyubiquitination) (Glickman and Ciechanover, 2002). Themajority of polyubiquitinated proteins are targeted for degra-dation by the 26S proteasome, whereas monoubiquitinationregulates the function and location of modified proteins ordirect them to lysosomes (Kudryashova et al. 2005).
Previous studies have indicated carbonylation andubiquitination of proteins in the digestive gland of marinemolluscs under oxidative stress (Chora et al. 2008; Dowlinget al. 2006). In addition, actin carbonylated in excess canundergo aggregation and inhibit the proteolytic activity ofthe proteasome (Castro et al. 2012). It is possible that changesin ACT2 expression associated with carbonyl aggregatescould inhibit the proteasome degradation pathway and, con-sequently, decrease the recycling of ubiquitin by lowering theproteosomal activity. The down-regulation of UBE3C (spot5104; Table 2; Fig. 4) detected in this study (4- and 8-fold forBUC×LIS and BUC×RIB, respectively) suggests a relation-ship with its low availability in the cytosol due to a possiblelow amount of ubiquitinated proteins and the recycling of theubiquitin through proteosomal degradation. This is furthersupported byMcDonagh and Sheehan (2006), which suggeststhat protein carbonylation in mussels is an independent pro-cess of the ubiquitination pathway, where the accumulation ofubiquitinated proteins could impair the ubiquitination path-way under oxidative stress in bivalves. The analysis of proteincarbonylation and redox proteomic should be further consid-ered in oysters exposed to sanitary sewage, as well as theidentification of ubiquitination pathways and degradation.
On the other hand, it was recently observed that CKAP5interacts with USP19, a specific protease which edits andrecycles ubiquitin, and it was demonstrated that USP19 inter-acts with ubiquitinated proteins having the ubiquitin ligase E3as the main target (Velasco et al. 2013). Accordingly, thelargest quantities of CKAP5 in oysters exposed to urbansewage could be related to a need for maintenance of availableubiquitin in the cytosol, as an indirect response to the loweramount of UBE3C.
Among the cellular proteins, cyclins are the regulators ofthe cell cycle (Glickman and Ciechanover 2002), being essen-tial for DNA repair and the completion of mitosis(Shackelford et al. 2000). Under oxidative stress conditions,permanent damage or loss of coordination of the cell cyclepromotes abnormal chromosome segregation and induces ap-optosis and cell death (Langley 2007; Shackelford et al. 2000;Tomanek 2011). In this study, down-regulation of the CCNB3
Environ Sci Pollut Res

protein was observed (2- and 5-fold for BUC×LIS and BUC×RIB, respectively; spot 7218; Table 2; Fig. 4). This is ahomologue, a protein to cyclin B3, related to cell cycle/celldivision.
The mitotic process is highly dependent on activity of cyclinB3 (Barzilai and Yamamoto 2004). The destruction of cyclin Bis required for the end of mitosis and is specifically mediated bythe ubiquitin E3 ligase by 26S proteasome (Glickman andCiechanover 2002; Min and Lindon 2012; Yu et al. 1996).Thus, the down-regulation of CCNB3 (2/5-fold) might interferewith the coordination of the cell cycle, causing DNA damage inoysters exposed to urban sewage. Furthermore, changes in thisprotein can also be related to the down-regulation of UBE3C(4- and 8-fold for BUC×LIS and BUC×RIB, respectively) andconsequently the ubiquitination of ACT2 (as previously men-tioned), given its specificity as a target protein for degradationvia the 26S proteasome.
The decrease in the expression of CCNB3 in oysters couldalso be related to changes in CKAP5. This cytoskeletal proteinis related to regulation of mitosis, as described above, and it issuggested as a possible interaction of CKAP5 with the p53tumor suppressor proteins, known as sensors of DNA damage,that inhibit the G1 phase of the cell cycle and regulate apo-ptosis (Chen et al. 2009; Langley 2007; Piekorz et al. 2002).CKAP5 up-regulation could be an early defense against cel-lular damage caused by the smaller CCNB3 (cyclin G2phase→M) and to the need for signaling cell cycle arrest inthe early stages of mitotic regulation (G0→G1) possibly toavoid cell damage (Suppl. Fig. 4).
Oysters exposed to sanitary effluents also showed a down-regulation (2/5-fold for BUC×LIS and BUC×RIB, respec-tively) of a protein Sushi, von Willebrand factor type A, EGF,and pentraxin domain-containing protein 1 (SVEP1; spot5506; Table 2; Fig. 4), whose function is unknown in bivalvemolluscs. The SVEP1 is a multi-domain protein of the extra-cellular matrix (ECM) comprising multiple domains charac-teristic of this structure (e.g., vonWillebrand factor A domain;pentraxin domain, EGF-like domains, and “sushi” domains)(Sato-Nishiuchi et al. 2012). The association of these domainsin the same molecule has been associated with immune func-tions (e.g., complement system, acute phase response of in-fection, and tissue injury) and/or cell adhesion (Bouchut et al.2006; Gilgès et al. 2000; Iwanaga et al. 1992; Schwarz et al.2008).
The domain von Willebrand factor type A domain (VWA)participates in a variety of functions, including homeostasis andinteractions between extracellular matrix (ECM) molecules,playing an important role in protein-protein interactions andthe assembly of protein subunits (e.g., proteins multimeric,microtubule, and cytoskeleton proteins) (Bouchut et al. 2006;Fitzgerald et al. 2002;Whittaker and Hynes 2002). In terrestrialmolluscs, the VWAs are present in the hemolymph as part ofthe coagulant immune system, playing a key role in the
processes of susceptibility/resistance in host-parasite interac-tions (Bouchut et al. 2006). Sushi domains/EGF-like are relatedto the modulation of immune system activation cascade andcoagulation of the hemolymph of crustaceans and related toendotoxin (LPS), a major component of the surface of Gram-negative bacteria (Iwanaga et al. 1992). Thus, the down-regulation of SVEP1 expression observed could be related toa reduction of the immune system in oysters exposed to sanitarysewage. In a parallel study, Souza et al. (2012) observed thatoysters transplanted to BUC site showed Salmonella (Gram-negative proteobacteria) after 14 days of exposure, which couldbe related to the above response.
Conclusion
This study is based on PEP changes in oysters C. gigasexposed to sanitary effluents as an impact of the overallresponse of organisms to urbanized coastal environments. Ingeneral, this study demonstrates the applicability of proteo-mics in oysters exposed to sanitary sewage effluents, althoughonly five of 30 selected proteins were identified.Most proteinsfrom invertebrate species used as biomarkers in pollutioncontrol programs, as in the case of molluscs, which are avail-able in databases, are usually related to cytoskeletal proteins(Rodríguez-Ortega et al. 2003). In accordance, two of theidentified proteins are associated with the cytoskeleton andcell structure (ACT2 and CKAP5). However, other identifiedproteins showed that exposure to urban sanitary sewage maycause changes in other cellular processes of digestive glandcells of oyster C. gigas, namely ubiquitination, cell cycle, andsignal transduction (UBE3C, CCNB3, and SVEP1). The re-sults suggest a possible relationship between the proteinsCKAP5, ACT2, UBE3C, and CCNB3, whose changes mightinterfere in the cellular apoptosis and in the immune system ofoysters exposed to sanitary sewage.
Acknowledgments FFN was a fellow doctoral PDEE (CAPES, Pro-cess 1452/10-0). The research leading to these results has receivedfunding from the EU project GENERA within the framework of theMarie Curie IRSES Actions (FP7-PEOPLE-2009-IRSES, Proposal no.247559) and CNPq grant number 483028/2012-6. Analysis of organiccontaminants was conducted in partnership with the Laboratory of Or-ganic Chemistry of the Oceanographic Institute of the University of SãoPaulo (LQO-IO-USP). ACDB, MCB, and CMRM are recipients ofCNPq productivity fellowship. MJB is recipient of PVE-CNPqfellowship.
References
Abessa DMS, Carr RS, Rachid BRF et al (2005) Influence of a Braziliansewage outfall on the toxicity and contamination of adjacent sedi-ments. Mar Pollut Bull 50:875–885
Environ Sci Pollut Res

Amelina H, Apraiz I, Sun W, Cristobal S (2007) Proteomics-basedmethod for the assessment of marine pollution using liquid chroma-tography coupled with two-dimensional electrophoresis. J ProteomeRes 6:2094–2104
Apraiz I, Mi J, Cristobal S (2006) Identification of proteomic signaturesof exposure to marine pollutants in mussels (Mytilus edulis). MolCell Proteomics 5:1274–1285
Apraiz I, Cajaraville MP, Cristobal S (2009) Peroxisomal proteomics:biomonitoring in mussels after the Prestige’s oil spill. Mar PollutBull 58:1815–1826
Barzilai A, Yamamoto K-I (2004) DNA damage responses to oxidativestress. DNA Repair (Amst) 3:1109–1115
Basto R, Gergely F, Draviam VM et al (2007) Hsp90 is required tolocalise cyclin B and Msps/ch-TOG to the mitotic spindle inDrosophila and humans. J Cell Sci 120:1278–1287
Beeby A (2001) What do sentinels stand for ? Environ Pollut 112:285–298
Blum H, Beier H, Gross HJ (1987) Improved silver staining of plantproteins, RNA and DNA in polyacrylamide gels. Electrophoresis 8:93–99
Bogoch Y, Linial M (2008) Coordinated expression of cytoskeletonregulating genes in the accelerated neurite outgrowth of P19 embry-onic carcinoma cells. Exp Cell Res 314:677–690
Bolong N, Ismail AF, Salim MR, Matsuura T (2009) A review of theeffects of emerging contaminants in wastewater and options for theirremoval. Desalination 239:229–246
Bouchut A, Roger E, Coustau C et al (2006) Compatibility in theBiomphalaria glabrata/Echinostoma caproni model: potential in-volvement of adhesion genes. Int J Parasitol 36:175–184
Bradford MB (1976) A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72:248–254
Bradley BP, Shrader EA, Kimmel DG, Meiller JC (2002) Protein expres-sion signatures: an application of proteomics. Mar Environ Res 54:373–377
CajaravilleMP, BebiannoMJ, Blasco J et al (2000) The use of biomarkersto assess the impact of pollution in coastal environments of theIberian Peninsula : a practical approach. Sci Total Environ 247:295–311
Campos A, Tedesco S, Vasconcelos V, Cristobal S (2012) Proteomicresearch in bivalves: towards the identification of molecular markersof aquatic pollution. J Proteomic 75:4346–4359
Cao A, Fuentes J, Comesaña P et al (2009) A proteomic approachenvisaged to analyse the bases of oyster tolerance/resistance tobonamiosis. Aquaculture 295:149–156
Cassimeris L, Morabito J (2004) TOGp, the human homolog ofXMAP215 / Dis1, is required for centrosome integrity, spindle poleorganization, and bipolar spindle assembly. Mol Biol Cell 15:1580–1590
Castro JP, Ott C, Jung T et al (2012) Free radical biology and medicinecarbonylation of the cytoskeletal protein actin leads to aggregateformation. Free Radic Biol Med 53:916–925
Castro JP, Jung T, Grune T, Almeida H (2013) Actin carbonylation : fromcell dysfunction to organism disorder. J Proteomics 92:171–180
Charrasse S, Lorca T, Dorée M, Larroque C (2000) The XenopusXMAP215 and its human homologue TOG proteins interact withcyclin B1 to target p34cdc2 tomicrotubules duringmitosis. Exp CellRes 254:249–256
Chen J, Zhou J, Sanders CK et al (2009) A surface display yeast two-hybrid screening system for high-throughput protein interactomemapping. Anal Biochem 390:29–37
Chora S, McDonagh B, Sheehan D et al (2008) Ubiquitination andcarbonylation as markers of oxidative-stress in Ruditapesdecussatus. Mar Environ Res 66:95–97
Chora S, Bebianno M, Roméo M (2009a) Two-dimensional electropho-resis protocols. In: Tyther R, Sheehan D (eds) Methods Mol. Biol.
Two Dimens. Electrophor. Protoc. vol. 519. Humana Press, Totowa,NJ, pp 1–8
Chora S, Starita-Geribaldi M, Guigonis J-M et al (2009b) Effect ofcadmium in the clam Ruditapes decussatus assessed by proteomicanalysis. Aquat Toxicol 94:300–308
Clesceri LS, Greenberg AE, Eaton AD (1998) Standard methods for theexamination of water and wastewater, 20th edn. American PublicHealth Association, the American Water Works Association and theWater Environment Federation, Washington
Conte N, Delaval B, Ginestier C et al (2003) TACC1-chTOG-aurora Aprotein complex in breast cancer. Oncogene 22:8102–8116
Dalle-Donne I, Rossi R, Milzani A et al (2001) The actin cytoskeletonresponse to oxidants: from small heat shock protein phosphorylationto changes in the redox state of actin itself. Free Radic Biol Med 31:1624–1632
Dalle-Donne I, Rossi R, Giustarini D et al (2003) Actin S-glutathionylation: evidence against a thiol-disulphide exchangemechanism. Free Radic Biol Med 35:1185–1193
Dalle-Donne I, Rossi R, Giustarini D et al (2007) S-glutathionylation inprotein redox regulation. Free Radic Biol Med 43:883–898
Dowling V, Hoarau PC, RomeoM et al (2006) Protein carbonylation andheat shock response in Ruditapes decussatus following p, p’-dichlorodiphenyldichloroethylene (DDE) exposure: a proteomic ap-proach reveals that DDE causes oxidative stress. Aquat Toxicol 77:11–18
EPAGRI - Agricultural Research and Rural Extension of Santa Catarina(2013) Fact summary of mariculture, http://migre.me/fmlh8,accessed 11 March 2013 (in Portuguese)
Fitzgerald J, Tay Ting S, Bateman JF (2002) WARP is a new member ofthe von Willebrand factor A-domain superfamily of extracellularmatrix proteins. FEBS Lett 517:61–66
Francone VP, Maggipinto MJ, Kosturko LD, Barbarese E (2007)The microtubule-associated protein tumor overexpressedgene/cytoskeleton-associated protein 5 is necessary for mye-lin basic protein expression in oligodendrocytes. J Neurosci27:7654–7662
Gilgès D, Vinit MA, Callebaut I, Coulombel L et al (2000) Polydom: asecreted protein with pentraxin, complement control protein, epider-mal growth factor and vonWillebrand factor A domains. Biochem J352:49–59
Glickman MH, Ciechanover A (2002) The ubiquitin-proteasome proteo-lytic pathway : destruction for the sake of construction. Physiol Rev82:373–428
Gomes T, Pereira CG, Cardoso C, Bebianno MJ (2013) Differentialprotein expression in mussels Mytilus galloprovincialis exposed tonano and ionic Ag. Aquat Toxicol 136–137:79–90
Gómez-Mendikute A, Etxeberria A, Olabarrieta I, Cajaraville MP (2002)Oxygen radicals production and actin filament disruption in bivalvehaemocytes treated with benzo (a) pyrene. Mar Environ Res 54:431–436
Grimalt JO, Ferninder P, Bayona JM, Albaiges J (1990) Assessment offecal sterols and ketones as indicators of urban sewage inputs tocoastal waters. Environ Sci Technol 24:357–363
Iwanaga S, Miyata T, Tokunaga F, Muta T (1992) Molecular mechanismof hemolymph clotting system in Limulus. Thromb Res 68:1–32
Jung T, Catalgol B, Grune T (2009) The proteasomal system. Mol AspecMed 30:191–296
Kennish MJ (1992) Ecology of estuaries: anthropogenic effects. CRCPress, London, p 494
Kinoshita K, Habermann B, Hyman AA (2002) XMAP215 : a keycomponent of the dynamic microtubule cytoskeleton. Trends CellBiol 12:267–273
Kling P, Norman A, Andersson PL et al (2008) Gender-specific proteo-mic responses in zebrafish liver following exposure to a selectedmixture of brominated flame retardants. Ecotoxicol Environ Saf 71:319–327
Environ Sci Pollut Res

Kudryashova E, Kudryashov D, Kramerova I, SpencerMJ (2005) Trim32is a ubiquitin ligase mutated in limb girdle muscular dystrophy type2H that binds to skeletal muscle myosin and ubiquitinates actin. JMol Biol 354:413–424
Langley BC (2007) The cell cycle and oxidative neuronal cell death. In:Qureshi GA, Parvez SH (eds) Oxidative Stress Neurodegener.Disord., 1a. Elsevier B.V, Oxford, pp 165–182
LemosMFL, Soares AMVM, Correia AC, Esteves AC (2010) Proteins inecotoxicology—how, why and why not? Proteomics 10:873–887
López JL (2005) Role of proteomics in taxonomy: the Mytilus complexas a model of study. J Chromatogr B 815:261–274
López JL, Marina A, Vázquez J, Alvarez G (2002) A proteomic approachto the study of the marine mussels Mytilus edulis and M.galloprovincialis. Mar Biol 141:217–223
Mao Y, Zhou Y, Yang H, Wang R (2006) Seasonal variation in metabo-lism of cultured pacific oyster, Crassostrea gigas, in Sanggou Bay,China. Aquaculture 253:322–333
Martins CC, Ferreira JA, Taniguchi S et al (2008) Spatial distribution ofsedimentary linear alkylbenzenes and faecal steroids of Santos Bayand adjoining continental shelf, SWAtlantic, Brazil: origin and fateof sewage contamination in the shallow coastal environment. MarPollut Bull 56:1359–1363
McDonagh B, Sheehan D (2006) Redox proteomics in the blue musselMytilus edulis: carbonylation is not a pre-requisite for ubiquitinationin acute free radical-mediated oxidative stress. Aquat Toxicol 79:325–333
McDonagh B, Tyther R, Sheehan D (2005) Carbonylation andglutathionylation of proteins in the blue mussel Mytilusedulis detected by proteomic analysis and Western blotting:actin as a target for oxidative stress. Aquat Toxicol 73:315–326
McLeod WD, Brown DW, Friedman AJ, Burrows DG et al (1986)Standard Analytical Procedures of the NOAA National AnalyticalFacility, 1985–1986. Extractable Toxic Organic Components, 2nded. U.S. Department of Commerce, NOAA Technical Memo,NMFS-F/ NWC-9. NOAA Northwest Fisheries Science Center,Seattle, p 121
Medeiros ID, Siebert MN, Toledo-Silva G et al (2008) Induced geneexpression in oyster Crassostrea gigas exposed to sewage. EnvironToxicol Pharmacol 26:362–365
Min M, Lindon C (2012) Substrate targeting by the ubiquitin-proteasomesystem in mitosis. Semin Cell Dev Biol 23:482–491
Monsinjon T, Andersen OK, Leboulenger F, Knigge T (2006) Dataprocessing and classification analysis of proteomic changes: a casestudy of oil pollution in the mussel, Mytilus edulis. Proteome Sci 4:17
Moore MN (2002) Biocomplexity: the post-genome challenge in ecotox-icology. Aquat Toxicol 59:1–15
Mudge SM, Norris CE (1997) Lipid biomarkers in the Conwy Estuary(North Wales, U.K.): a comparison between fatty alcohols andsterols. Mar Chem 57:61–84
Nesatyy VJ, Suter MJ (2007) Proteomics for the analysis of environmen-tal stress responses in organisms. Environ Sci Technol 41:6891–6900
Olsson B, Bradley BP, Gilek M et al (2004) Physiological and proteomicresponses in Mytilus edulis exposed to PCBs and PAHs extractedfrom Baltic Sea sediments. Hydrobiologia 514:15–27
Pandey A, Mann M (2000) Proteomics to study genes and genomes.Nature 405:837–846
Paulsen RD, Soni DV, Wollman R et al (2009) A genome-wide siRNAscreen reveals diverse cellular processes and pathways that mediategenome stability. Mol Cell 35:228–239
Pickart CM (2004) Back to the future with ubiquitin review. Cell 116:181–190
Piekorz RP, Hoffmeyer A, Duntsch CD et al (2002) The centrosomalprotein TACC3 is essential for hematopoietic stem cell function and
genetically interfaces with p53-regulated apoptosis. EMBO J 21:653–664
Piña B, CasadoM, Quirós L (2007) Analysis of gene expression as a newtool in ecotoxicology and environmental monitoring. Trends AnalChem 26:1145–1154
Rodríguez-Ortega MJ, Grøsvik BE, Rodríguez-Ariza A et al (2003)Changes in protein expression profiles in bivalve molluscs(Chamaelea gallina) exposed to four model environmental pollut-ants. Proteomics 3:1535–1543
Røe OD, Anderssen E, Sandeck H et al (2010) Malignant pleural meso-thelioma: genome-wide expression patterns reflecting general resis-tance mechanisms and a proposal of novel targets. Lung Cancer 67:57–68
Ronzitti G,Milandri A, Scortichini G et al (2008) Protein markers of algaltoxin contamination in shellfish. Toxicon 52:705–713
Rudert F, Ge L, Ilag L (2000) Functional genomics with protein-proteininteractions. Biotechnol; Ann Rev 5:45–86
Saavedra C, Bachère E (2006) Bivalve genomics. Aquaculture 256:1–14Sato-Nishiuchi R, Nakano I, Ozawa A et al (2012) Polydom/SVEP1 is a
ligand for integrin α9β1. J Biol Chem 287:25615–25630Schwarz RS, Bosch TCG, Cadavid LF (2008) Evolution of polydom-like
molecules: identification and characterization of cnidarian polydom(Cnpolydom) in the basal metazoan Hydractinia. Dev CompImmunol 32:1192–1210
Shackelford RE, Kaufmann WK, Paules RS (2000) Oxidative stressand cell cycle checkpoint function. Free Radic Biol Med 28:1387–1404
Shepard JL, Bradley BP (2000) Protein expression signatures and lyso-somal stability in Mytilus edulis exposed to graded copper concen-trations. Mar Environ Res 50:457–463
Shepard JL, Olsson B, Tedengren M, Bradley BP (2000) Protein expres-sion signatures identified inMytilus edulis exposed to PCBs, copperand salinity stress. Mar Environ Res 50:337–340
Shevchenko A, Tomas H, Havlis J et al (2006) In-gel digestion for massspectrometric characterization of proteins and proteomes. NatProtoc 1:2856–2860
Souza DSM, Ramos APD, Flores-Nunes F et al (2012) Evaluation oftropical water sources and mollusks in southern Brazil using micro-biological, biochemical, and chemical parameters. EcotoxicolEnviron Saf 76:153–161
Thompson EL, Taylor DA, Nair SVet al (2012a) Proteomic discovery ofbiomarkers of metal contamination in Sydney rock oysters(Saccostrea glomerata). Aquat Toxicol 109:202–212
Thompson EL, Taylor DA, Nair SV et al (2012b) Proteomicanalysis of Sydney rock oysters (Saccostrea glomerata) ex-posed to metal contamination in the field. Environ Pollut170:102–112
Tomanek L (2011) Environmental proteomics: changes in the proteomeof marine organisms in response to environmental stress, pollutants,infection, symbiosis, and development. Ann Rev Mar Sci 3:373–399
Tomanek L, Zuzow MJ, Ivanina AV et al (2011) Proteomic response toelevated PCO2 level in eastern oysters, Crassostrea virginica: evi-dence for oxidative stress. J Exp Biol 214:1836–1844
UNEP (1992). Determinations of petroleum hydrocarbons in sediments,Reference Methods for Marine Pollution studies 20, 77p.
USEPA (2009) Occurrence of contaminants of emerging concern inwastewater from nine publicly owned treatment works. 85p.
Velasco K, Zhao B, Callegari S et al (2013) An N-terminal SIAH-interacting motif regulates the stability of the ubiquitin spe-cific protease (USP)-19. Biochem Biophys Res Commun 433:390–395
Veldhoen N, Ikonomou MG, Helbing CC (2012) Molecular pro-filing of marine fauna: integration of omics with environmen-tal assessment of the world’s oceans. Ecotoxicol Environ Saf76:23–38
Environ Sci Pollut Res

Wang H, Wang S, Lu Y et al (2012) Proteomic analysis of early-responsive redox-sensitive proteins in Arabidopsis. J ProteomeRes 11:412–424
Whittaker CA, Hynes RO (2002) Distribution and evolution of vonWillebrand / integrin a domains : widely dispersed domainswith roles in cell adhesion and elsewhere. Mol Biol Cell 13:3369–3387
Yu H, King RW, Peters JM, Kirschner MW (1996) Identification of anovel ubiquitin-conjugating enzyme involved in mitotic cyclin deg-radation. Curr Biol 6:455–466
Zanette J, Flores-Nunes F, Medeiros ID et al (2008) Comparison of theantioxidant defense system in Crassostrea rhizophorae andCrassostrea gigas exposed to domestic sewage discharges. MarEnviron Res 66:196–198
Environ Sci Pollut Res




















