
Floral size variation causes differentiation of pollinators and genetic parameters in Alpinia...
9
1 23 Plant Systematics and Evolution ISSN 0378-2697 Volume 299 Number 5 Plant Syst Evol (2013) 299:865-871 DOI 10.1007/s00606-013-0768-z Floral size variation causes differentiation of pollinators and genetic parameters in Alpinia nieuwenhuizii, a flexistylous ginger (Zingiberaceae) Atsuko Takano, Johnny Gisil & Monica Suleiman
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Floral size variation causes differentiation of pollinators and genetic parameters in Alpinia...

1 23
Plant Systematics and Evolution ISSN 0378-2697Volume 299Number 5 Plant Syst Evol (2013) 299:865-871DOI 10.1007/s00606-013-0768-z
Floral size variation causes differentiationof pollinators and genetic parameters inAlpinia nieuwenhuizii, a flexistylous ginger(Zingiberaceae)
Atsuko Takano, Johnny Gisil & MonicaSuleiman

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Wien. This e-offprint is for personal
use only and shall not be self-archived
in electronic repositories. If you wish to
self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL ARTICLE
Floral size variation causes differentiation of pollinatorsand genetic parameters in Alpinia nieuwenhuizii, a flexistylousginger (Zingiberaceae)
Atsuko Takano • Johnny Gisil • Monica Suleiman
Received: 30 September 2012 / Accepted: 2 February 2013 / Published online: 24 February 2013
� Springer-Verlag Wien 2013
Abstract Floral size dimorphism, pollination, and
genetic variation of Alpinia nieuwenhuizii (Zingiberaceae),
a flexitylous ginger, were studied. This study revealed that
floral size differed among habitats (i.e., roadsides/river-
sides vs. forest floors). The effective pollinators of small-
flowered populations of the species on a forest floor were
different from those of large-flowered populations along
roadsides/riversides. Using inter-simple sequence repeat
(ISSR) PCR, considerable genetic differentiation was
detected between small- and large-flowered populations.
These results indicate that reproductive isolation in A.
nieuwenhuizii owing to the differentiation of pollen vectors
between two floral size morphs may lead to genetic dif-
ferentiation between the two morphs.
Keywords Alpinia � Flexistyly � ISSR � Genetic analysis �Pollinator ecotype � Zingiberaceae
Introduction
Speciation in plants can result from the formation of eco-
types adapted to the environment of a local pollinator (Grant
and Grant 1962; Stebbins 1970), and several pollination
ecotypes have been reported, e.g., in Gilia splendens
Douglas ex H. Mason and A.D. Grant (Grant and Grant
1965), Campanula punctata L. (Inoue and Amano 1986),
Platanthera ciliaris Lindl. (Robertson and Wyatt 1990),
Isodon umbrosus (Maxim.) H. Hara and Isodon shikokianus
(Makino) H. Hara (Suzuki 1992), and Satyrium hallackii
Bolus (Johnson 1997). These studies have provided evidence
that floral differentiation (e.g., size or length of spur) occurs
among populations and is induced by pollinator differences
between floral types. Pollinator differences may lead to
genetic differentiation between floral types because the gene
flow between types is reduced. There are no published
studies that provide data regarding genetic variations among
pollinator ecotypes, and it remains to be established whether
there is genetic differentiation between ecotypes.
Alpinia nieuwenhuizii Val. is endemic to Borneo and is
found in various habitats, including riversides, roadsides,
and montane forest floors (Smith 1985). This species is
easily distinguished from other Bornean Alpinia species by
its strongly paniculate inflorescence and small ligule
(maximum of 1.5 cm; Smith 1985). It is also known to
exhibit flexistyly, a form of unique stylar polymorphism
combining reciprocal herkogamy and dichogamy, which
enhances outcrossing (Cui et al. 1995, 1996; Li et al. 2001,
2002; Zhang et al. 2003; Takano et al. 2005; Sun et al.
2007). Takano et al. (2005) observed visitors to flowers of
A. nieuwenhuizii growing in a freshwater swamp forest
(altitude, 15 m) near the mouth of the Segama River,
Sabah, Malaysia and concluded that two species of large
carpenter bees, Xylocopa (Koptorthosoma) latipes and
Xylocopa (Zonohirsuta) collaris alboxantha, were the
effective pollinators. However, floral size differentiation
has been reported in A. nieuwenhuizii, with flowers
appearing to be smaller in highland populations than in
lowland populations (Poulsen 2006). This suggests that
pollinators may differ between highland and lowland
populations of A. nieuwenhuizii.
A. Takano (&)
Museum of Nature and Human Activities, Hyogo, Sanda,
Hyogo, Japan
e-mail: [email protected]
J. Gisil � M. Suleiman
Institute for Tropical Biology and Conservation, University
Malaysia Sabah, Locked Bag 2073, 88999 Kota Kinabalu,
Sabah, Malaysia
123
Plant Syst Evol (2013) 299:865–871
DOI 10.1007/s00606-013-0768-z
Author's personal copy

This study aimed to: (1) confirm floral size differentia-
tion among populations of A. nieuwenhuizii, (2) identify
the effective pollinators of montane populations of A.
nieuwenhuizii, (3) clarify genetic variations within and
among A. nieuwenhuizii populations using inter-simple
sequence repeat (ISSR) PCR, and (4) evaluate genetic
differences between populations with different floral sizes.
To our knowledge, this is the first study of genetic variation
in a flexistylous ginger and of genetic differentiation
between pollination ecotypes.
Materials and methods
All five study sites (Kimanis, Trail4, Mahua, Ranau, and
Segama) were located in Sabah, Eastern Malaysia
(Table 1). The first three populations were located on the
forest floor of lower montane forests in Crocker Range
Park (CRP), western Sabah. The remaining two popula-
tions were located in very different habitats: the Ranau
population was situated along the side of a trunk road from
Kota Kinabalu through Ranau, and the Segama population
was located on the banks of the Segama River, which runs
through a lowland area (Takano et al. 2005).
Measurement of floral morphology
To describe the floral size, five floral characteristics were
selected and measured: floral tube length, labellum length
and width, filament length, and anther length (Fig. 1). The
number of flowers examined is shown in Table 2. Data
collected from the measurements of each floral part were
analyzed by analysis of variance (ANOVA). When differ-
ences were significant, a multiple comparison was per-
formed using the Tukey–Kramer HSP multiple comparison
test. All statistical tests were performed using Statview J.
ver. 5 (SAS Institute Inc., Cary, NC USA).
Observations of flower visitors in the lower montane
forest
Visitors to flowers of A. nieuwenhuizii in the montane
forest were observed directly at Kimanis during the periods
of 13–16 November 2008 and 15–18 September 2009. We
found two flowering individuals (one cataflexistylous and
one that started to release pollen in the morning, but no
stigma movement occurred in the afternoon) in 2008 and
three (one analflexistylous and two cataflexistylous) in
2009. All observations of flower visitors were made
between 8:00 and 17:00 (a total of 12.5 h in 2008 and
37.9 h in 2009). Flower visitors were only recorded when
they touched the anther or stigma. Some visitors were
captured for identification. Vouchers of flower visitors
were deposited at the Institute for Tropical Biology and
Conservation (ITBC), University Malaysia Sabah (BORH).
ISSR-PCR analysis
Background
Inter simple sequence repeat (ISSR) markers are generated
from single-primer PCRs where the primer is designed
from di- or trinucleotide repeat motifs with a 50 or 30
anchoring sequence of one to three nucleotides (Gupta
Table 1 Populations of Alpinia nieuwenhuizii studying floral morphology, ISSR variation, and flower visitors
Population Locality Latitude and Longitude alt. (m) Habitat
Kimanis Ulu Kimanis, Papar 5� 300 16.80 0 N, 116� 470 000 0 E 555 below montane forest
Trail4 In Crocker Range, Papar 5� 290 19.40 0 N, 116� 010 07.80 0 E 700 below montane forest
Mahua Crocker Range, Tambunan 5� 470 50.00 0 N, 116� 260 000 0 E 1,200 below montane forest
Ranau Along route A4, Ranau 5� 560 35.80 0 N, 116� 390 00.10 0 E 737 road side
Segama Tidong village, kinabatangan 5� 240 48.80 0 N, 118� 430 35.60 0 E 15 below freshwater swamp forest
All sites are located in Sabah E Malaysia, Borneo
Fig. 1 Measured parts of an Alpinia nieuwenhuizii flower: floral tube
length, labellum length and width, filament length, and anther length
866 A. Takano et al.
123
Author's personal copy
et al. 1994; Zietkiewicz et al. 1994). Recent ISSR studies
of natural populations have demonstrated the hyper-vari-
able nature of these markers and their potential for popu-
lation-level studies (Wolfe et al. 1998; Camacho and
Liston 2001; Ge et al. 2005; Gupta et al. 2008).One limi-
tation of the ISSR technique, which is the same as for
Random Amplification of Polymorphic DNA (RAPD;
Williams et al. 1990), is that bands are scored as dominant
markers and genetic diversity estimates are based on
diallelic characters (Ge et al. 2005).
ISSR-PCR analysis
Leaves from 92 individuals of A. nieuwenhuizii were col-
lected from the five populations (Table 3). To minimize the
possibility of sampling from the same genets, shoots that
were at least 5–6 m apart from each other were selected.
Plant material was stored in silica gel immediately after
collection and kept at -20 �C prior to DNA extraction.
Total DNA was isolated from 0.5 to 1.0 g of silica gel-
dried leaves, using a modified version of the 19 cetyltri-
methylammonium bromide extraction protocol from Doyle
and Doyle (1987). We screened 17 ISSR primers for the
814.1 markers (50 ? 30): [(CT)8-TG], M1 [CAA-(GA)5],
ISSR-2 [(AC)8-TA], ISSR-4 [(AG)8-TA], ISSR-5 [(CA)8-
G], ISSR-6 [(CT)8-GC], UBC-808 [(AG)8-C], UBC-820
[(GT)8-TC], UBC-827 [(AC)8-CG], UBC-830 [(TG)8-GG],
UBC-831 [(AT)8-CA], UBC-843 [(CT)8-TGA], UBC-852
[(TC)8-CGA], UBC-864 [(ATG)6], UBC-868 [(GAA)6],
UBC-870 [(TGC)6], and UBC-874 [(CCCT)4]. We asses-
sed the reproducibility of the bands by running duplicate
reactions on different days. Nine of the primers (ISSR-2,
ISSR-4, ISSR-6, UBC-808, UBC-820, UBC-827, UBC-
852, UBC-864, and UBC-868) produced 67 strong, clear,
and reproducible polymorphic bands, which were used for
further analysis. PCRs were carried out in a total volume of
25 ll containing: 2.5 ll of 109 PCR buffer, 2.5 mM
MgCl2, 2.5 mM dNTPs, 0.4 lM of primer, 1.25 U of Taq
polymerase (TaKaRa Biotechnology, Otsu Japan), and
50 ng of genomic DNA. The ISSR-PCR amplifications
were conducted with a PCR Thermal Cycler (MP) (Takara
Bio Inc., Japan) under the following conditions: initial
denaturation for 1 min at 94 �C followed by 36 cycles of
40 s at 94 �C, 45 s at the annealing temperature of each
primer (46–52 �C), and 90 s at 72 �C, with a final exten-
sion for 5 min at 72 �C. The amplification products were
separated by electrophoresis in 2 % (w/v) agarose gels in
19 TAE buffer at 100 V for 2 h. A 100 bp ladder was used
as a size reference. The gel was stained with ethidium
bromide and photographed using a gel imaging system
(ImageSaver AE-6905C; ATTO, Tokyo, Japan). The
banding patterns were scored manually.
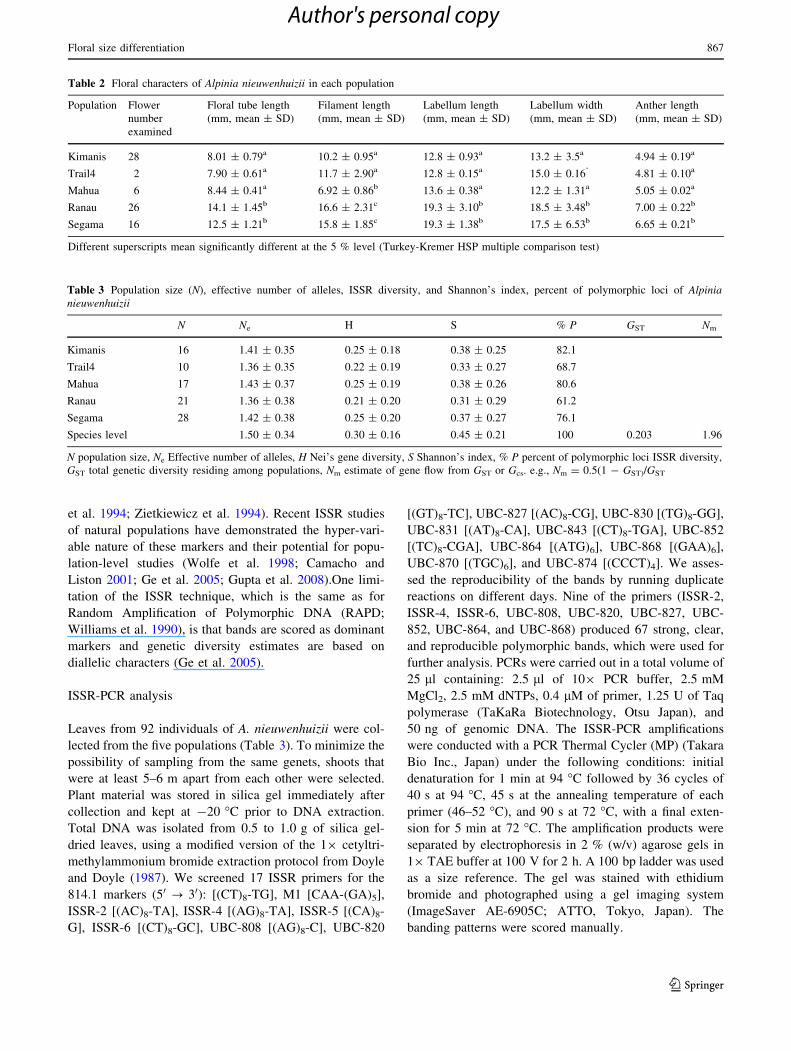
Table 2 Floral characters of Alpinia nieuwenhuizii in each population
Population Flower
number
examined
Floral tube length
(mm, mean ± SD)
Filament length
(mm, mean ± SD)
Labellum length
(mm, mean ± SD)
Labellum width
(mm, mean ± SD)
Anther length
(mm, mean ± SD)
Kimanis 28 8.01 ± 0.79a 10.2 ± 0.95a 12.8 ± 0.93a 13.2 ± 3.5a 4.94 ± 0.19a
Trail4 2 7.90 ± 0.61a 11.7 ± 2.90a 12.8 ± 0.15a 15.0 ± 0.16‘ 4.81 ± 0.10a
Mahua 6 8.44 ± 0.41a 6.92 ± 0.86b 13.6 ± 0.38a 12.2 ± 1.31a 5.05 ± 0.02a
Ranau 26 14.1 ± 1.45b 16.6 ± 2.31c 19.3 ± 3.10b 18.5 ± 3.48b 7.00 ± 0.22b
Segama 16 12.5 ± 1.21b 15.8 ± 1.85c 19.3 ± 1.38b 17.5 ± 6.53b 6.65 ± 0.21b
Different superscripts mean significantly different at the 5 % level (Turkey-Kremer HSP multiple comparison test)
Table 3 Population size (N), effective number of alleles, ISSR diversity, and Shannon’s index, percent of polymorphic loci of Alpinianieuwenhuizii
N Ne H S % P GST Nm
Kimanis 16 1.41 ± 0.35 0.25 ± 0.18 0.38 ± 0.25 82.1
Trail4 10 1.36 ± 0.35 0.22 ± 0.19 0.33 ± 0.27 68.7
Mahua 17 1.43 ± 0.37 0.25 ± 0.19 0.38 ± 0.26 80.6
Ranau 21 1.36 ± 0.38 0.21 ± 0.20 0.31 ± 0.29 61.2
Segama 28 1.42 ± 0.38 0.25 ± 0.20 0.37 ± 0.27 76.1
Species level 1.50 ± 0.34 0.30 ± 0.16 0.45 ± 0.21 100 0.203 1.96
N population size, Ne Effective number of alleles, H Nei’s gene diversity, S Shannon’s index, % P percent of polymorphic loci ISSR diversity,
GST total genetic diversity residing among populations, Nm estimate of gene flow from GST or Gcs. e.g., Nm = 0.5(1 - GST)/GST
Floral size differentiation 867
123
Author's personal copy

ISSR data analysis
Amplified fragments were scored as ‘‘1’’ for the presence
and ‘‘0’’ for the absence of homologous bands. A binary
data matrix of different ISSR phenotypes was established
and analysed using the program POPGENE version 1.31
(Yeh et al. 1999). Hardy–Weinberg equilibrium was
assumed, and the following genetic diversity parameters
were estimated at the population and species levels: the
percentage of polymorphic bands (% P), the effective
number of alleles per locus (Ne), Nei’s (1973) gene
diversity (H), and Shannon’s index (S). To examine the
population genetic structure, gene differentiation (GST)
among populations was estimated. To visualize the genetic
relationships among populations, a dendrogram was con-
structed based on Nei’s genetic distance (D) and using an
unweighted pair-group cluster analysis method with arith-
metic averages (UPGMA).
Results
Measurement of floral morphology
Table 2 shows the measurements of floral morphology.
The measured size of each floral organ was consistent
within each population studied. No significant differences
were found among the three populations in the lower
montane forests, except for the filament length of the
Mahua population. The same trend was observed in both
the roadside and riverside populations. Flowers of the three
forest populations were significantly smaller than those in
Ranau and Segama (P [ 0.05, Tukey–Kramer HSP mul-
tiple comparison test), suggesting two flower size classes in
A. nieuwenhuzii.
Observations of flower visitors
Flower visitors to the small-flowered form at Kimanis
in 2008
The most frequent visitor to the small-flowered form
(hereafter SF form) of A. nieuwenhuizii at Kimanis was
Amegilla calceifera (Fig. 2a; 62 visits). The second most
common visitor was the stingless bee Tetragonilla collina
(Fig. 2b; 35 visits), followed by Trigona melanocephala (4
visits), and finally a carpenter bee, Xylocopa caerulea
(Fig. 2c; 2 visits). The most frequent visiting time for
A. calceifera and T. collina was during the early morning
(6:00–8:00), followed by a period after midday (12:00–
14:00). Amegilla calceifera descended onto the labellum,
foraged for nectar, and typically collected pollen by
scratching the anther with their forelegs. We visually
confirmed pollen attachment on the back of the bees while
they were moving on the labellum. Tetragonilla collina
descended directly onto the opened thecae and collected only
pollen. Xylocopa caerulea foraged for nectar and did not
collect pollen, although the back of the insects did touch the
thecae during visits to the flower (Fig. 2c).
Flower visitors at Kimanis in 2009
Amegilla calceifera was observed visiting the flowers of A.
nieuwenhuizii 181 times during this observation period.
The next most common visitors were another stingless-bee
(unidentified, but clearly different from T. collina and T.
melanocephala), which visited the flowers 139 times, and
X. caerulea, which visited 68 times. Visits by A. calceifera,
to both cataflexistylous and anaflexistylous flowers,
occurred most frequently between 10:00–12:00 and then
12:00–14:00. In contrast, visits by the stingless bee were
concentrated in the morning and occurred only on flowers
in the male stage. Xylocopa caerulea visited the flower
most frequently between 16:00–17:00 and then early in the
morning (6:00–8:00), with visits to both cataflexistylous
and anaflexistylous flowers.
ISSR-PCR analysis
The nine selected ISSR primers generated 67 bands (loci)
that ranged from 300 to 3,000 base pairs (bp) across all 92
individuals of the five populations of A. nieuwenhuizii. The
primers yielded 6–9 bands each, with an average of 7.4
bands per primer. All 67 bands were polymorphic. The
mean percentage of polymorphic bands (% P) at the pop-
ulation level was 73.7 %, with a range of 61.2 % (Ranau)
to 82.1 % (Kimanis) (Table 3). The mean effective number
of alleles per locus (Ne) was 1.36 at the population level
and 1.50 at the species level. By assuming Hardy–Wein-
berg equilibrium, the mean gene diversity was estimated to
be 0.24 (H) within populations and 0.30 at the species level
(Table 3). Shannon’s index had a mean of 0.36 within
populations and 0.45 at the species level.
Population genetic structure
Statistically significant genetic differentiation among the
populations of A. nieuwenhuizii was observed as follows.
The GST was estimated to be 0.203 (Table 3), which means
that 20.3 % of the genetic variability was distributed
among populations. The overall level of inferred gene flow
(Nm) was estimated to be 1.96 individuals per generation
among populations (Table 3), indicating a moderate gene
flow via pollen and seed dispersal between populations.
Nei’s genetic distance (D) among populations varied from
0.044 to 0.148 (Table 4). Genetic distances among the
868 A. Takano et al.
123
Author's personal copy

Kimanis, Trail4, and Mahua populations for the SF form
ranged between 0.044 and 0.066. The genetic distance
between the Mahua and Ranau populations, with the large-
flowered form (hereafter LF form), was 0.120, which is two
to three times the distances between SF form populations in
CRP, despite their closer geographical distance (29.1 km)
compared with the distances between Mahua-Kimanis
(56.4 km) and Mahua-Trail4 (57 km; Table 4). The three
SF form populations formed a cluster and the two LF form
populations formed another cluster on a phenogram based
on Nei’s genetic distances (Fig. 3).
Discussion
Differences in the effective pollinator
among populations of the SF and LF forms
This study confirmed the existence of floral size dimor-
phism in A. nieuwenhuizii. All three populations with the
SF form were located under a montane forest canopy at
high altitudes, ranging from 555 to 1,200 m, in CRP. In
contrast, the LF form was established in open areas along a
trunk road (Ranau, ca. 700 m altitude) and along a river
Fig. 2 Flower visitors foraging pollen and/or nectar of Alpinianieuwenhuizii. a, d Amegilla calceifera collected and became dusted
with pollen, in addition to sucking nectar, when visiting the small
flower type (a, at Kimanis), but only nectar consumption, with no
pollen attachment on the body, was observed on the large flower type
(d, at Ranau). b Tetragonilla collina landed directly on the thecae and
foraged for pollen; no pollen attachment was observed. c Xylocopacaerulea landed on the labellum, sucked nectar, and was dusted with
pollen on its back whenever it came to a flower in the male stage
Table 4 Geographic distance (km, above diagonal) and Nei’s genetic
distance (below diagonal) of Alpinia nieuwenhuizii
Kimanis Trail4 Mahua Ranau Lsegama
Kimanis 1.9 56.4 85.6 300.9
Trail4 0.044 57 86.1 300.2
Mahua 0.066 0.059 29.1 258.2
Ranau 0.139 0.147 0.12 237.7
Lsegama 0.092 0.117 0.099 0.148
Floral size differentiation 869
123
Author's personal copy

bank (Segama, ca. 15 m altitude). Therefore, floral size
difference did not correspond to differences in altitude as
suggested by Poulsen (2006). Instead, we suggest that floral
dimorphism may be induced by environmental differences,
because floral, as well as vegetative, traits can exhibit
phenotypic plasticity in response to spatially variable
environments (Herrera et al. 2006). Individuals of A. nie-
uwenhuizii in the CRP populations were scattered on the
forest floor of montane rainforest where limited sunlight is
available. Many species of ground herbs, grasses and
sedges grow together, and A. nieuwenhuizii never becomes
dominant there. In contrast, populations are found along
riverside and riversides/roadsides that are sunny, and where
there is no upper layer of tall trees. Species composition is
very simple, and A. nieuwenhuizii is often dominant.
Examinations of herbarium sheets at the University
Malaysia Sabah (BORH), the Museum of Nature and
Human Activities, Hyogo (HYO), the Forest Research
Centre (SAN), and Sabah Parks (SNP) also supported our
idea: the specimens collected from montane forest floors
had smaller flowers, and those collected from roadsides and
riversides had larger flowers.
It is generally considered that flower size is strongly
associated with pollinator size (e.g., Inoue and Amano
1986; Inoue and Amano 1986), and the results obtained in
this study indicate that a difference in flower size is asso-
ciated with the differentiation of the effective pollinator.
The frequent flower visitors observed at Kimanis were
A. calceifera, stingless bees (T. collina and an unidentified
species), and a carpenter bee (X. caerulea). Among these
species, stingless bees cannot serve as pollinators for A.
nieuwenhuizii because they collect only pollen (at least for
this species) and visit flowers in the male stage only. In
flexistylous flowers, the stigma is behind the thecae when
the flower is in the male stage; thus, pollen-collecting bees
do not touch the stigma. In contrast, A. calcelfera and X.
caerulea are both honey suckers, and pollen was confirmed
to attach to their backs. By visiting the flowers during both
the male and female stages, they are effective pollinators
for the SF form (Fig. 2a, c). Takano et al. (2005) studied
flower visitors to the LF form of A. nieuwenhuizii at Se-
gama and reported that two large carpenter bees (X. latipes
and X. alboxantha) were the effective pollinators. Visits of
X. latipes to the flowers of A. nieuwenhuizii were also
observed at Ranau in 2004 (Takano, personal observation).
However, large carpenter bees were not observed visiting
the SF form at Kimanis, despite the wide distribution of
both bee species in Southeast Asia. Although there were
frequent observations of A. caleifera (=37 visits during
11.8 h observation, Takano unpublished data) and the
unidentified stingless bee on the LF form at Ranau,
A. calceifera did not touch the anther because the flower
was too large (Fig. 2d). This suggests that even
though A. calceifera visits both forms of the flower, it can
be an effective pollinator for only the SF form of
A. nieuwenhuizii.
Genetic differentiation among populations of the SF
and LF forms
Flexistyly is believed to be a mechanism for promoting
outcrossing (e.g., Li et al. 2001, 2002; Zhang et al. 2003).
The present study provides evidence that A. nieuwenhuizii
maintains high genetic diversity as well as genetic differ-
entiation among populations similar to that of other
outcrossing species. The observed mean gene diversity
within populations of A. nieuwenhuizii (H = 0.296) was
higher than the average reported for outcrossing species
(H = 0.260) (Nybom and Bartish 2000; Nybom 2004), and
the value of genetic differentiation among populations of
A. nieuwenhuizii (GST = 0.20) was slightly lower than the
values for other species (GST = 0.23, Nybom and Bartish
2000).
However, there may be a barrier preventing gene flow
between populations of the LF and SF forms. The genetic
distances between the Ranau and CRP populations
(GD = 0.119–0.147) were much larger than those among
the three CRP populations (GD = 0.044–0.066). The UP-
GMA phenogram indicated that the Ranau population
formed a cluster with the Segama population, which both
consisted of individuals of the LF form, but not with the
Fig. 3 An unrooted UPGMA phenogram based on Nei’s (1973)
genetic distance among five populations of Alpinia nieuwenhuizii.Population names follow those in Table 1
870 A. Takano et al.
123
Author's personal copy

CRP population. This indicates that gene flow has fre-
quently occurred among the CRP populations, but has been
limited between Ranau and the CRP populations. The
barrier to gene flow could be the effective pollinator, which
is a consequence of the size of the flowers. The amegilla
bee (A. calceifera) is an effective pollinator of CRP pop-
ulations, but not the Ranau population. Although it is still
unknown which genetic or plastic variations induce the
difference in flower size, this change may result in the
differentiation of effective pollinators, leading to genetic
differentiation among different floral size forms.
Acknowledgments We sincerely thank the Economic Planning
Unit, Prime Minister’s Department, for giving us permission to
undertake research in Malaysia (UPE: 40/200/19/1523). Sabah Parks
allowed us to use their facilities in the CRP. We are grateful to the
curators of BORH, HYO, SAN, and SNP for allowing us to examine
their facilities and collections. Prof. Dr. Makoto Kato (Kyoto Univ.)
kindly identified flower visitors. Drs. H. Okada and T. Denda pro-
vided critical comments on an earlier version of the manuscript. We
also thank two anonymous reviewers for their constructive comments.
This study was partly supported by a Grant-in-Aid for young scien-
tists (B) (no. 18770075 to A.T.) from the Japanese Ministry of
Education, Culture, Sports, Science, and Technology.
References
Camacho FJ, Liston A (2001) Population structure and genetic
diversity of Botrychium pumicola (Ophioglossaceae) based on
inter-simple sequence repeats (ISSR). Am J Bot 88:1065–1070
Cui XL, Wei RC, Huang RF (1995) A preliminary study on the
genetic system of Amomum tsao-ko. J Yunnan Univ Nat Sci
17:290–297
Cui XL, Wei RC, Huang RF (1996) A study on the breeding system
of Amomum tsao-ko. In: Wu TL, Wu QG, Chen ZY (eds.) Proc.
2nd Symp. Fam. Zingiberaceae, Guangzhou, China, pp 288–296
Doyle JJ, Doyle JD (1987) A rapid DNA isolation procedure for small
quantities of fresh leaf tissue. Phytochem Bull 19:11–15
Ge XJ, Yu Y, Yuan YM, Huang HW, Yan C (2005) Genetic diversity
and geographic differentiation in endangered Ammopiptanthus(Leguminosae) populations in desert regions of northwest China
as revealed by ISSR analysis. Ann Bot 95:843–851
Grant KA, Grant V (1962) Mechanical isolation of Salvia apiana and
Salvia mellifera (Labiatae). Evolution 18:196–212
Grant V, Grant KA (1965) Flower pollination in the phlox family.
New York. pp 69–73
Gupta M, Chyi YS, Romero-Severson J, Owen JL (1994) Amplifi-
cation of DNA markers from evolutionarily diverse genomes
using single primers of simple-sequence repeats. Theor Appl
Genet 89:998–1006
Gupta S, Srivastava M, Mishra GP, Naik PK, Chauhan RS, Tiwari
SK, Kumar M, Singh R (2008) Analogy of ISSR and RAPD
markers for comparative analysis of genetic diversity among
different Jatropha curcas genotypes. African J Biotech 7:4230–
4243
Herrera CM, Castellanos MC, Medrano M (2006) Geographical
context of floral evolution: towards and improved research
programme in floral diversification. In: Harder LD, Barrett SCH
(eds) Ecology and evolution of flowers. Oxford University Press,
New York, pp 278–294
Inoue K, Amano M (1986) Evolution in Campanula punctata Lam. In
the Izu Islands: changes of pollinators and evolution of breeding
systems. Plant Species Biol 1:89–97
Johnson SD (1997) Pollination ecotypes of Satyrium hallackii(Orchidaceae) in South Africa. Bot J Linne Soc 123:225–235
Li QJ, Xu ZF, Kress WJ, Xia YM, Zhang L, Deng XB, Gao JY, Bai
ZL (2001) Flexible style that encourages outcrossing. Nature
410:432
Li QJ, Kress WJ, Xu ZF, Xia YM, Zhang L, Deng XB, Gao JY (2002)
Mating system and stigmatic behaviour during flowering of
Alpinia kwangsiensis (Zingiberaceae). Pl Syst Evol 232:123–132
Nei M (1973) Analysis of gene diversity in subdivided populations.
Proc Natl Acad Sci USA 70:332–3323
Nybom H (2004) Comparison of different nuclear DNA markers for
estimating intraspecific genetic diversity in plants. Mol Ecol
13:1143–1155
Nybom H, Bartish IV (2000) Effects of life history traits and sampling
strategies on genetic diversity estimates obtained with RAPD
markers in plants. Perspective Plant Ecol Evol Syst 3:93–114
Poulsen AD (2006) A pocket guide; ginger of Sarawak. Natural
History Publications, Borneo, p 16
Robertson JL, Wyatt R (1990) Evidence for pollination ecotypes in
the yellow fringed orchid, Platanthera ciliaris. Evolution
44:121–133
Smith RM (1985) A review of Bornean Zingiberaceae: 1 (Aplineae
p.p.) Notes RBG Edinb 42:261–314
Stebbins GL (1970) Adaptive radiation of reproductive characteristics
in angiosperms, 1: pollination mechanisms. Annual Rev Ecol
Syst 1:307–326
Sun S, Gao JY, Liao WJ, Li QJ, Zhang DY (2007) Adaptive
significance of flexistyly in Alpinia blepharocalyx (Zingibera-
ceae): a hand pollination experiment. Ann Bot 99:661–666
Suzuki K (1992) Bumble bee pollinators and pollination ecotypes of
Isodon umbrosus and I. shikokianus (Lamiaceae). Plant Species
Biol 7:37–48
Takano A, Gisil J, Yusoff M, Tachi T (2005) Floral and pollinator
behaviour of flexistylous Bornean ginger, Alpinia nieuwenhuizii(Zingiberaceae). Pl Syst Evol 252:167–173
Williams JGK, Kubelik AR, Livak KJ, Rafalski AJA, Tingey SV
(1990) DNA polymorphism amplified by arbitrary primers are
useful as genetic markers. Nucleic Acids Res 22:6531–6535
Wolfe AD, Xiang QY, Kephart S (1998) Assessing hybridization in
natural populations of Penstemon (Scrophulariaceae) using
hypervariable intersimple sequence repeat (ISSR) bands. Mol
Ecol 7:1107–1125
Yeh FC, Yang RC, Boyle T (1999) POPGENE. Microsoft Windows-
based freeware for population genetic analysis. Release 1.31.
University of Alberta, Edomonton
Zhang L, Li QJ, Deng XB, Ren PY, Gao JY (2003) Reproductive
biology of Alpinia blepharocalyx (Zingiberaceae): another
example of flexistyly. Pl Syst Evol 241:67–76
Zietkiewicz E, Rafalski A, Labuda D (1994) Genome fingerprinting
by simple sequence repeat (SSR)- anchored polymerase chain
reaction amplification. Genomics 20:176–183
Floral size differentiation 871
123
Author's personal copy




















