
Outbreaks of Edwardsiella ictaluri Infection in Ayu ... - J-Stage

© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society
Agricultural and Forest Entomology (2007), 9, 141–158 DOI:10.1111/j.1461-9563.2007.00327.x
Introduction
Subsequent to its discovery in Macedonia in 1984 ( Deschka & Dimic, 1986 ), the horse-chestnut leaf miner Cameraria ohridella Deschka & Dimic has spread very rapidly all over Europe and was first observed in Switzerland in 1998 ( Kenis & Forster, 1998 ). In all invaded regions, the leaf miner reaches outbreak densities, causing severe damage to horse-
chestnut Aesculus hippocastanum L. an important ornamen-tal tree throughout most of Europe. Despite yearly fluctuations in population densities, no outbreak has ever collapsed, a phenomenon rarely encountered in insects ( Berryman, 1987 ). To understand the mechanisms that allow this insect to build and maintain outbreak densities, and potentially to develop sustainable control methods, it is es-sential to assess all mortality factors and other population traits acting on the host through all of its developmental stages. Surprisingly, very little is known about the different mortality factors affecting C. ohridella , with the notable exception of parasitism. The latter has been the focus of
Factors favouring the development and maintenance of outbreaks in an invasive leaf miner Cameraria ohridella (Lepidoptera: Gracillariidae): a life table study
Sandrine Girardoz * † , Donald L. J . Quicke † ‡ § and Marc Kenis * * CABI Switzerland Centre, 1, rue des Grillons, 2800 Delémont, Switzerland , † Division of Biology, Imperial College London, Silwood Park Campus, Ascot, Berks SL5 7PY, U.K. , ‡ Centre for Population Biology, Silwood Park, Ascot, Berks SL5 7PY, U.K. and § Department of Entomology, Natural History Museum, London SW7 5BD, U.K.
Abstract 1 Life tables were constructed for seven generations of the invasive horse-chestnut leaf miner Cameraria ohridella in Switzerland, to assess the factors allowing the moth to build and maintain outbreak densities and to identify ‘gaps’ among their mortality factors that could be targeted by new control methods. The fecundity of females was calculated and the mortality factors affecting all developmental stages determined.
2 Females contained 106.6 eggs. In captivity, the mean number of eggs laid per fe-male varied between 34 and 82 eggs, with a maximum of 184 eggs. Egg mortal-ity was 18 – 75% and was mostly due to unknown factors.
3 The main mortality factors affecting larvae and pupae in the leaves were preda-tion by birds and arthropods in all generation and leaf senescence in the autumn generation. Parasitoids killed less than 5% of the population.
4 Nearly 90% of overwintering pupae died in dead leaves, the majority of them be-ing killed by earthworms and other leaf decomposers. As a result, the overwinter-ing generations were the only ones showing a net rate of increase less than one. Non-overwintering generations had net rates of increase between four and 11, al-lowing populations to build up from spring to autumn.
5 The persistence of high outbreak densities in Europe is explained by a combina-tion of at least three factors: (i) high fecundity; (ii) multivoltinism; (iii) the unu-sually low impact of traditional natural enemies of leaf miners and particularly parasitoids.
6 Yearly variations in population densities could be at least partly explained by dif-ferences in larval mortality due to leaf senescence and intraspecific competition in the last generation in autumn.
Keywords Cameraria ohridella , fecundity , invasive species , life tables , mortality factors , parasitoids , predators
Correspondence: Marc Kenis, CABI Switzerland Centre, 1, Rue des Grillons, 2800 Delémont, Switzerland. Tel.: +41 32 421 48 84; fax: +41 32 421 48 71; e-mail: [email protected].

142 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
several studies both in Central Europe ( Grabenweger & Lethmayer, 1999; Hellrigl, 2001; Grabenweger, 2003; Girardoz et al. , 2006; Volter & Kenis, 2006 ) and in the Balkans where the host tree originates and where the moth was first discovered ( Freise et al. , 2002; Grabenweger et al. , 2005a ). Approximately 30 native generalist parasitoid spe-cies have been recorded as attacking this alien species. However, parasitism rates are low and, although reliable data on population dynamics are lacking, there is common agreement that the role of parasitoids in the natural regula-tion of C. ohridella populations is negligible.
Mortality factors other than parasitism have been poorly studied. Predation was investigated only recently in a single study ( Grabenweger et al. , 2005b ) and there are hardly any reliable and quantifiable data on other mortality factors and population traits such as winter, egg and adult mortality, fe-cundity, host resistance, intraspecific competition and insect-host plant synchronization.
In the present study, we present quantitative data on mor-tality factors and other traits affecting the population dynam-ics of C. ohridella in Switzerland, and integrate them into life tables ( Bellows et al. , 1992; Van Driesche & Bellows, 1996 ). The main aim was to understand the roles of these factors in the natural control and population dynamics of the moth, and to identify the causes of permanent outbreaks. Ideally, parallel studies should be carried out in the region of origin of the moth, or in regions where it occurs at low den-sity, to allow comparison between outbreaking and non-out-breaking populations with the aim of identifying differences leading to outbreaks. Such a comparative study would also allow us to spot ‘gaps’ among the mortality factors in the in-vaded region that could be targeted by new control methods, such as the importation of a key natural enemy from the re-gion of origin. However, the origin of C. ohridella is not yet known, and nor is there any region where the moth is known to occur at low density ( Kenis et al. , 2005 ). Consequently, such comparisons are not currently possible. In the mean-time, data on population traits of other leaf mining moths producing no, or temporary, outbreaks ( Kirizoglu, 1976; Faeth & Simberloff, 1981; Van Driesche & Taub, 1983; Hespenheide, 1991; Connor & Beck, 1993 ) can be used to assess the specificity of the permanent outbreak behaviour of C. ohridella .
Materials and methods
Field sites
The study was carried out at a site in north-eastern Switzerland, in Delémont (47°21 ¢ 59 ¢ ¢ N, 7°19 ¢ 59 ¢ ¢ E), in the Canton Jura. It consisted of an alley of 54 horse-chestnut trees situated along a road leading to a forest where the first mines of C. ohridella were observed in 2000.
Population size
Population size was measured at the end of each generation, when the majority of mines contained pupae. Two genera-
tions per year were observed in Delémont in 2001, 2002, 2004 and 2005, and three in 2003. Fifteen horse-chestnut leaves were chosen on 15 trees, at random, and the mine den-sities were estimated using the damage key of Gilbert & Grégoire (2003) , based on a visual assessment of the relative mined area per leaf. Each leaf was assigned to one of the eight scores defined by the authors: 0% damage, 2, 5, 10, 25, 50, 75 and 100% damage. The median value was taken as an indicator of the population size of one generation. Gilbert & Grégoire (2003) showed that their damage scoring system provides a good tool to estimate and compare population densities and it is now increasingly used in population stud-ies on C. ohridella ( Gilbert et al. , 2005; Grabenweger et al. , 2005b; Kehrli et al. , 2005 ). The sample size of 15 leaves was chosen after an analysis showing that the median value did not change with larger sample sizes (S. Girardoz and M. Kenis, unpublished data).
Fecundity
Potential fecundity was studied in 2001 (first generation, i.e. females emerging in early summer from the first generation of mines), 2003 (first and second generations) and 2004 (first generation). Pupae were separated by sex according to Freise & Heitland (1999) and stored in glass vials. Females were frozen within the day of emergence and kept until they were dissected under saline water (0.9%), and the number of oocytes in the ovaries was counted. Between 37 and 125 females were dissected per generation.
Realized fecundity was studied in 2001 (first and second generations), 2002 and 2003 (overwintering generation, i.e. females emerging from overwintering pupae from the second generation of the preceeding year, and first and second gen-erations). Freshly emerged (less than 8 h old) females and males were first maintained for a few hours in laboratory conditions in a cylinder (1.3 L) for mating (maximum 30 in-dividuals per cylinder). Females were then transferred indi-vidually to muslin cages (20 × 20 × 30 cm) with one male and one horse-chestnut leaf with its petiole immersed in wa-ter to increase the chances of the female being mated. These cages were placed outdoors in a shaded tunnel, and females were provided daily with water. Food was not provided be-cause it had been shown previously that females fed with honey neither lived longer nor laid more eggs than unfed ones ( Girardoz, 2001 ). The eggs laid on the leaves were counted after the female’s death and kept until hatching to assess viability. Only fertile eggs were included in the calcu-lation of fecundity, and females that had laid only sterile eggs were suspected to have been unmated and were discarded. Between seven and 28 fertile females per generation were in-cluded in the analyses. It cannot be excluded that some de-gree of unfertility also occurs under natural conditions. However, the proportion of sterile females occurring in na-ture is not known and is undoubtedly much lower than that observed in our laboratory experiments, where mating was not easily obtained and up to half of the females were found to be sterile. Therefore, it was decided to discard them from the analyses.

Life tables of C. ohridella 143
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Egg mortality
In 2002, 2003 and 2004, horse-chestnut seedlings (70 – 100 cm high) or cut branches placed in water were exposed to C. ohridella females in cages in the laboratory. The females were left on the leaves for three days to allow them to lay eggs, which were then counted and their spatial distribution noted. Tree seedlings or cut branches in water were then ex-posed in the canopies of mature trees in the last phase of the oviposition periods, to minimize additional natural oviposi-tion on the exposed leaves. After the majority of the eggs had hatched, the branches or seedlings were taken back to the laboratory and the eggs were scored as flat/empty, disap-peared, opened or hatched. The number of eggs exposed per generation was very variable (20 – 1173), but only generations for which at least 80 eggs could be exposed were included in the analyses.
To determine whether egg mortality was due to predators and, if so, which kind of predators, an experiment was car-ried out in 2004. Five seedlings per generation were exposed to C. ohridella females as described above. For each seed-ling, the leaves were separated into three batches with a simi-lar number of eggs on them. One batch was enclosed in a gauze bag, to exclude flying and crawling predators. The sec-ond batch of leaves had their petioles smeared with Tangle-coating (The Tanglefoot Company, Grand Rapids, Michigan) to exclude crawling predators only. A third batch was left as a control. The seedlings were then placed amongst the foli-age of mature trees, in an attempt to place their stems/trunk in contact with branches but keeping their leaves away from any contact. The seedlings were brought back to the labora-tory after the majority of the eggs had hatched. The eggs and first-instar larvae were counted and classified as in previous years.
Larval and pupal mortality during the summer and autumn
Summer larval and pupal mortality were studied in Delémont from 2002 to 2004, which corresponds to seven generations of the moth (two in 2002 and 2004, three in 2003). For each generation, 400 first instar larvae were chosen at random on the lower branches (max 2 m above ground level) of a mini-mum of 30 different trees. Leaves with selected mines were tagged, the position of the mines on the leaf was noted and the cohort of mines was checked every week to monitor larva and parasitoid development and fate. When mines with feed-ing stages of the larvae stopped growing, they were brought to the laboratory and examined. The developmental stage was noted and the mortality factor of dead or dying larvae classi-fied into the categories: (1) Parasitism (an egg or larva of a parasitoid was found in the mine); (2) Host feeding by para-sitoids (the integument of the host larva was stuck to the up-per inner surface of the mine as defined in Askew & Shaw, 1979 ); (3) Predation by birds (i.e. the upper surface of the mine was ripped open); (4) Predation by arthropods; (5) Plant defence reaction (i.e. necrosis around a young mine); (6) Competition with the leaf fungus Guignardia aesculi (Peck) Stewart (Ascomycetes); (7) Intra-specific competition at high
densities; (8) Senescence of the host leaf; (9) Unknown cause (i.e. no obvious sign of any mortality factor mentioned above). Categories 6, 7 and 9 were combined as ‘other’ for the analysis because the difference between these three fac-tors was not always obvious, and the numbers dying from each of these factors were low.
For all generations but the last one of the season, pupae were left on the trees until the adults emerged. If no adult had emerged one month after pupation, two thirds of the mines were brought back to the laboratory and examined. The remaining third was left on the trees to assess mortality affecting summer diapausing pupae.
The proportions of larvae and pupae killed by the different mortality factors were assessed for each stage and each gen-eration investigated.
Overwintering mortality
To assess winter mortality of C. ohridella pupae in the leaf litter, leaves were collected at leaf fall in Delémont on 30 October 2002 and 27 October 2003. For each leaf, the number of pupae was noted on a small plastic tag attached to the main vein of the leaf. One thousand pupae were selected per year, and separated in ten groups of 100 mines. Five groups were placed in an envelope of chicken wire, five groups were placed between a plastic mesh on the bottom and the chicken wire on top. The chicken wire was used to prevent the loss of leaves. Earthworms, and possibly other leaf decomposers, were partly (2002 – 03) or entirely (2003 – 04) excluded by separating the leaves from the ground by means of plastic mesh, 3 mm in diameter in the first year, 0.2 mm in the sec-ond. The batches of leaves were placed amongst the leaf lit-ter underneath mature horse-chestnuts at the study site. The leaves were removed in the spring, as soon as the first adult C. ohridella was caught in a pheromone trap at the site ( Kalinova et al. , 2003 ) (i.e. on 17 April 2003 and 18 April 2004). The remaining pupae were counted and the mortality was assessed. In winter 2003 – 04, one control grid and one with mesh were checked monthly from November to April to assess when pupae disappear. Only two grids were inspected monthly, to minimize disturbance.
Construction of life tables
The columns of the life tables were defined in accordance with the structure of Van Driesche & Bellows (1996) as fol-lows: (i) l x , the number of individuals entering stage x ; (ii) d x , the number of individuals dying in stage x ; (iii) factor d x , the number of individuals dying from a given factor; (iv) marginal attack rates ( m x ); (v) apparent mortality; (vi) real mortality; and (viii) k -values . Further details can be found in Southwood (1978) and Bellows et al. (1992) . Marginal at-tack rates were calculated after Elkinton et al. (1992) . In the case of two contemporaneous mortality factors, of which one is always decided to be the ‘winner’ in case of competition, we used Eq. (6) from Elkinton et al. (1992) . In the case of more than two contemporaneously acting mortality factors, we used Eq. (9).

144 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
The number of eggs entering the next generation ( F 1 prog-eny) was estimated as the number of adults × sex ratio × fe-male realized fecundity. The sex ratio used in the life tables was of 0.54, as calculated by Freise (2001) . The net repro-ductive rate ( R 0 ), was taken as the ratio of the F 1 progeny to the initial number of individuals entering the life table.
It was not possible to assess the fecundity of C. ohridella females and the mortality affecting certain stages in all years and all generations. Potential fecundity could not be obtained in 2002 and in the second generation of 2004. For the miss-ing data, we used those of the corresponding generation in 2003. Data on the realized fecundity of females could not be obtained in 2004, and thus the corresponding data of 2003 were used. In 2003, egg mortality could not be assessed for the first generation, so we used the mean of the first genera-tions of the years 2002 and 2004. Winter mortality was not assessed for the winter 2004 – 05. Instead, we used the aver-age of the two previous years.
For certain mortality factors, some assumptions had to be made. For mortality through leaf senescence, we assumed that larvae that were still in the feeding stages and spinning stages in the last sample when leaves were senescing would die, whereas pupae were assumed to survive (S. Girardoz and M. Kenis, unpublished data). The few pupae of the first gen-erations that were entering into an early diapause were con-sidered as dead because we observed that, in general, these could not survive until leaf senescence (S. Girardoz and M. Kenis, unpublished data). By contrast, we considered that diapausing pupae of the second generation were able to survive until the winter, even when a third generation was observed.
Results
Population size
Population densities in Delémont increased over the years 2001 – 03 ( Table 1) but the 2004 populations were lower in comparison with the previous year, with an estimated dam-age of 5% and 25% for the first and second generations, re-spectively. The population increased again in 2005, with a total defoliation observed in the second generation.
Fecundity
Freshly emerged C. ohridella females contained a mean of 106.63, mostly immature, eggs (SE = 0.99, n = 314). A maxi-mum of 180 eggs per female was found. Females of the first
generation of 2001 were found to have 124.59 eggs on aver-age (SE = 2.25, n = 37). In 2003, females had 102.74 eggs on average (SE = 1.58, n = 105) in the first generation and 100.25 (SE = 1.07, n = 125) in the second generation. In 2004, females of the first generation had 118.17 eggs on av-erage (SE = 3.08, n = 47). The difference between the years was highly significant [one-way analysis of variance (anova): F = 10.01, d.f = 2, P < 0.001] with fecundity in 2003 being significantly lower than in the two other years (Scheffe test, P < 0.001).
The realized fecundity for all generations is shown in Table 2. The mean number of eggs laid per female varied be-tween 34 and 82 eggs, with a maximum of 184 eggs. The number of eggs strongly decreased with years and the differ-ence was significant (two-way anova, F = 4.93, d.f. = 2, P = 0.009; fecundity lower in 2001 than in 2002 and 2003, Scheffe test, P < 0.01). There was no significant difference between generations ( F = 0.49, d.f. = 2; P = 0.611).
Egg mortality
Results for egg mortality are given in Table 3. Survival rates were found to vary little between the first and second genera-tion with 61% to 82% of the eggs hatching. The one third generation observed showed a much lower survival rate, per-haps because the eggs were laid on severely attacked leaves, including on mines, in which case hatching larvae were una-ble to survive. The majority of the eggs that did not hatch were simply not recovered. However, in some generations, a sizeable number of eggs were found to be unhatched, flat and dry ( Table 3 ). Very few eggs were found destroyed with any obvious signs of predation.
Results obtained in the experiment set up in 2004 to assess the role of predators are shown in Fig. 1. There were no sig-nificant differences in mortality between the different treat-ments neither in the first generation (Friedman: n = 15, c 2 = 3.00, d.f. = 1, P = 0.083) nor in the second ( n = 15, c 2 = 3.00, d.f. = 1, P = 0.093) and approximately 70% of eggs hatched in both generations.
Winter mortality
The pupal mortality observed in the winters 2002 – 03 and 2003 – 04 is shown in Fig. 2. In both years, less than 15% of the pupae survived, as shown in the control samples. Most pupae had disappeared, eaten by predators or saprophages. Leaf veins were often found half buried in the soil, suggesting that they had been moved there by earthworms. In 2002 – 03, the number of pupae that disappeared was not significantly greater in the
Table 1 Estimated damage from Cameraria ohridella at the end of each generation in Delémont over the years 2001 – 05
2001 2002 2003 2004 2005
First generation 2% 10% 25% 5% 50% Second generation 25% 50% 50% 25% 100% Third generation – – 75% – –
Percentages indicate the leaf surface damaged by the mines.

Life tables of C. ohridella 145
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
control than in the earthworm- excluded batches (Mann – Whitney U -test: Z = – 1.984, P = 0.056).
By contrast, in 2003 – 04, the number of pupae disappeared was significantly greater in the control than in the earth-worm-excluded batches, in which more than 50% of the pu-pae were still alive in spring ( Z = – 2.611, P = 0.008). These results suggest that earthworms play an important role in the overwintering mortality of C. ohridella and that the mesh used in 2002 – 03 may have not been fine enough to exclude earthworms effectively. The monthly monitoring of two grids suggested that earthworms are active mainly in early spring ( Fig. 3) as numbers started to differ in March
Larval and pupal mortality and construction of life tables
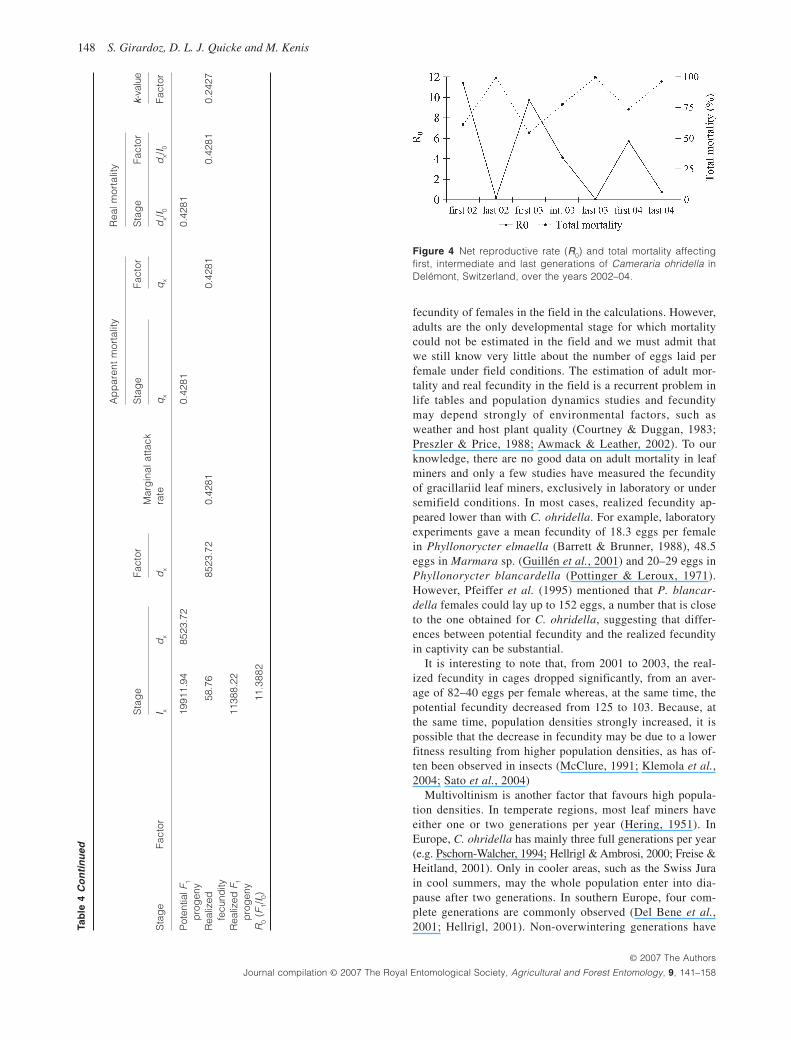
Life tables are presented in Tables 4 – 10. All generations ex-cept for the last ones of the season had a very high net repro-ductive rate: they increased from 4.1-fold to 11.4-fold. Only the last, overwintering generations had a net increase rate be-low 1, due to high mortality rates on overwintering pupae ( Fig. 4).
Total mortality ranged from 55.3% of the total population to 77.6% in the first and intermediate generations and from 96.0% to 99.6% for the last generation of the season ( Fig. 4 ). Egg mortality generally accounted for the death of 18 – 36% of the population of one generation, the only third generation examined had the highest egg mortality rate observed, reach-ing 75% of the total population. Real mortality rates in the first- and second-instar larvae were usually below 10%. First- (L1) and second- (L2) instar larvae were subject to only a few mortality factors, mostly plant defence reaction and un-
known mortality, whereas parasitism and predation were rare. Third-instar larvae (L3) were subject to more mortality fac-tors than the younger larvae, but a maximum of 5% of the population died in this stage in the first and second genera-tions. Fourth-instar larvae (L4) suffered the highest mortality amongst feeding stages: 2 – 11% of the total population died at this stage. In spring and summer, the major mortality fac-tor affecting L4 was predation either by birds or by arthro-pods (with marginal attack rates of 1 – 19%). Birds feeding on C. ohridella were mostly great tits Parus major L. and blue tits Parus caeruleus L. whereas other predators were not clearly identified, although a bush-cricket, Meconema merid-ionale (Orthoptera, Tettigoniidae) was often seen on leaves and has been described by Grabenweger et al. (2005b) as a predator of C. ohridella. In the last generations, leaf senes-cence was a major cause of mortality in 2002 and 2003 (e.g. with marginal attack rates of 10% and 20%). Parasitism re-mained low, with marginal attack rates of less than 1% in most cases. The main parasitoids attacking C. ohridella were Minotetrastichus frontalis (Nees) and Pnigalio agraules (Walker) (Hymenoptera: Eulophidae). Patterns for spinning stages were similar to those observed for L4. Indeed, for non-overwintering generations, the major mortality was often bird predation, with marginal attack rates of 1 – 11%, but real mortality remaining under 7%. For overwintering genera-tions, the major mortality factor was leaf senescence, with marginal attack rates of 5 – 44%. Pupae of all generations were attacked less than L4 and spinning stages, with only 0 – 9% of the population dying in that stage. Only one genera-tion, the intermediate generation in 2003, had higher values, with 26% of the total population dying in the pupal stage, which was mostly due to a higher impact of birds.
Table 2 Realized fecundity of Cameraria ohridella females in Delémont, Switzerland in 2001, 2002 and 2003
2001 2002 2003
Overwintering generation a – 41.62 ± 6.60 (16, 85) 38.35 ± 4.68 (17, 81) 1st generation 81.69 ± 13.74 (13, 184) 58.76 ± 11.31 (17, 141) 40.26 ± 4.56 (27, 95) 2nd generation b 82.41 ± 7.88 (28, 161) 58.57 ± 5.40 (7, 87) 34.00 ± 5.50 (5, 52) Mean 82.08 ± 6.83 (41, 184) 51.87 ± 5.62 (40, 141) 39.96 ± 3.01 (49, 95)
a Overwintering generation, females emerging from overwintering pupae (i.e. from the second generation of the preceeding year). b 2nd generation, females emerged before winter, even when, as in 2001 and 2002, the majority of the pupae of the second generation went into diapause. Numbers indicate the mean ± SE number of eggs per female (no. of females, maximum number of eggs).
Table 3 Mortality of Cameraria ohridella eggs over the years 2002 – 04 in Delémont
2002 2003 2004
First generation
Second generation
Second generation
Third generation
First generation
Second generation
Total 1173 88 112 246 292 331 Eggs dry 4 4 89 76 2 Eggs disappeared 372 32 15 94 28 97 Eggs opened 1 1 1 Eggs hatched 801 56 92 62 187 232 Survival rate 0.68 0.64 0.82 0.25 0.64 0.70
Numbers are given only for generations for which a minimum of 80 eggs could be exposed.

146 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
‘Other mortality’ was observed in all stages, but remained low. Marginal attack rates were, in most cases, below 10%.
Discussion
Subsequent to the first discovery of C. ohridella in Macedonia in 1984 and its subsequent spread to most of Europe, out-breaks have continued unabated. Despite yearly fluctuations in population levels, no outbreak has ever collapsed. Our life tables confirm these observations. For all non-overwintering generations, high net rates of increase were found, which were consistent with the increases in populations observed at the study sites. A combination of at least three factors can explain the permanence of outbreak densities in central Europe: (i) high fecundity; (ii) multivoltinism; and (ii) an unusually low impact of the traditional natural enemies of leaf miners, in particular parasitoids.
The fecundity of C. ohridella was previously thought to be rather low. Skuhravy (1998) had suggested a fecundity of 20 – 30 eggs whereas Freise (2001) had measured an average realized fecundity of 23 and a potential fecundity of 42, with a maximum of 80 eggs per female. We found fecundities that were substantially higher. Both dissections of ovarioles
(potential fecundity) and oviposition tests in cages (realized fecundity) showed that a female can lay up to 180 eggs. On average, potential fecundity was much higher than realized fecundity, probably because conditions in cages were not op-timal for oviposition tests, and many tested females laid few eggs. Because realized fecundity was used in the calculation of the life tables, it is likely that we underestimated the real
Figure 1 Percentage of eggs hatched (white bars), unhatched (striped bars) and disappeared (grey bars) after the three treatments (exclusion of all predators, exclusion of crawling predators and control), in the first (A) and second (B) generation of Cameraria ohridella in 2004 in Delémont, Switzerland.
Figure 2 Mortality of pupae of Cameraria ohridella during the winter 2002 – 03 (A) and the winter 2003 – 04 (B) in Delémont, Switzerland.
Figure 3 Numbers of pupae of Cameraria ohridella present in two grids during the winter 2003 – 04 in Delémont, Switzerland. The grids were not examined in February because they were covered by snow.

Life tables of C. ohridella 147
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Tab
le 4
Life
tab
le f
or t
he f
irst
gen
erat
ion
of C
amer
aria
ohr
idel
la in
Del
émon
t, S
witz
erla
nd,
2002
Ap
par
ent
mor
talit
y
Rea
l mor
talit
y
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Eg
g
10
00
319.
46
0.
3195
0.31
95
Eg
gs
dis
app
ear
off l
eaf
31
6.05
78
0.31
61
0.
3161
0.31
61
0.16
50
E
gg
s fl a
t and
em
pty
3.39
85
0.00
50
0.
0034
0.00
34
0.00
22
L1
68
0.54
6.
81
0.
0100
0.00
68
Leaf
sen
esce
nce
6.
8054
0.
0100
0.01
00
0.
0068
0.
0044
L2
673.
74
69.7
6
0.10
35
0.
0698
P
lant
def
ense
rea
ctio
n
34.0
272
0.05
19
0.
0505
0.03
40
0.02
32
O
ther
15.3
122
0.02
37
0.
0227
0.01
53
0.01
04
P
aras
itism
8.50
68
0.01
32
0.
0126
0.00
85
0.00
58
H
ost f
eed
ing
6.80
54
0.01
06
0.
0101
0.00
68
0.00
46
Le
af s
enes
cenc
e
3.40
27
0.00
51
0.
0051
0.00
34
0.00
22
P
red
atio
n (a
rthr
opod
s)
1.
7014
0.
0027
0.00
25
0.
0017
0.
0012
L3
603.
98
32.3
3
0.05
35
0.
0323
O
ther
13.6
109
0.02
29
0.
0225
0.01
36
0.01
01
Le
af s
enes
cenc
e
8.50
68
0.01
41
0.
0141
0.00
85
0.00
62
P
aras
itism
6.80
54
0.01
15
0.
0113
0.00
68
0.00
50
P
red
atio
n (b
irds)
1.70
14
0.00
29
0.
0028
0.00
17
0.00
13
P
red
atio
n (a
rthr
opod
s)
1.
7014
0.
0029
0.00
28
0.
0017
0.
0013
L4
571.
66
74.8
6
0.13
10
0.
0749
P
red
atio
n (b
irds)
20.4
163
0.03
76
0.
0357
0.02
04
0.01
66
P
red
atio
n (a
rthr
opod
s)
17
.013
6 0.
0314
0.02
98
0.
0170
0.
0139
Par
asiti
sm
13
.610
9 0.
0252
0.02
38
0.
0136
0.
0111
Oth
er
10
.208
2 0.
0190
0.01
79
0.
0102
0.
0083
Leaf
sen
esce
nce
8.
5068
0.
0149
0.01
49
0.
0085
0.
0065
Hos
t fee
din
g
5.
1041
0.
0095
0.00
89
0.
0051
0.
0042
S
pin
496.
80
28.9
2
0.05
82
0.
0289
P
aras
itism
15.3
122
0.03
13
0.
0308
0.01
53
0.01
38
P
red
atio
n (b
irds)
8.50
68
0.01
71
0.
0171
0.00
85
0.00
75
O
ther
3.40
27
0.00
70
0.
0068
0.00
34
0.00
31
H
ost f
eed
ing
1.70
14
0.00
35
0.
0034
0.00
17
0.00
15
Pup
ae
46
7.87
80
.26
0.
171
5
0.08
03
Pre
dat
ion
(bird
s)
35
.858
9 0.
0766
0.07
66
0.
0359
0.
0346
Par
asiti
sm
23
.906
0 0.
0545
0.05
11
0.
0239
0.
0243
Oth
er
15
.368
1 0.
0354
0.03
28
0.
0154
0.
0156
Pre
dat
ion
(art
hrop
ods)
1.70
76
0.00
40
0.
0036
0.00
17
0.00
17
H
ost f
eed
ing
1.70
76
0.00
40
0.
0036
0.00
17
0.00
17
P
upae
in d
iap
ause
1.70
76
0.00
40
0.
0036
0.00
17
0.00
17
Ad
ults
387.
62
S
ex r
atio
0.5
P
oten
tial
fe
cund
ity
10
2.74
Con
tinue
d
148 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
fecundity of females in the field in the calculations. However, adults are the only developmental stage for which mortality could not be estimated in the field and we must admit that we still know very little about the number of eggs laid per female under field conditions. The estimation of adult mor-tality and real fecundity in the field is a recurrent problem in life tables and population dynamics studies and fecundity may depend strongly of environmental factors, such as weather and host plant quality ( Courtney & Duggan, 1983; Preszler & Price, 1988; Awmack & Leather, 2002 ). To our knowledge, there are no good data on adult mortality in leaf miners and only a few studies have measured the fecundity of gracillariid leaf miners, exclusively in laboratory or under semifield conditions. In most cases, realized fecundity ap-peared lower than with C. ohridella . For example, laboratory experiments gave a mean fecundity of 18.3 eggs per female in Phyllonorycter elmaella ( Barrett & Brunner, 1988 ), 48.5 eggs in Marmara sp. ( Guillén et al. , 2001 ) and 20 – 29 eggs in Phyllonorycter blancardella ( Pottinger & Leroux, 1971 ). However, Pfeiffer et al. (1995) mentioned that P. blancar-della females could lay up to 152 eggs, a number that is close to the one obtained for C. ohridella , suggesting that differ-ences between potential fecundity and the realized fecundity in captivity can be substantial.
It is interesting to note that, from 2001 to 2003, the real-ized fecundity in cages dropped significantly, from an aver-age of 82 – 40 eggs per female whereas, at the same time, the potential fecundity decreased from 125 to 103. Because, at the same time, population densities strongly increased, it is possible that the decrease in fecundity may be due to a lower fitness resulting from higher population densities, as has of-ten been observed in insects ( McClure, 1991; Klemola et al. , 2004; Sato et al. , 2004 )
Multivoltinism is another factor that favours high popula-tion densities. In temperate regions, most leaf miners have either one or two generations per year ( Hering, 1951 ). In Europe, C. ohridella has mainly three full generations per year (e.g. Pschorn-Walcher, 1994; Hellrigl & Ambrosi, 2000; Freise & Heitland, 2001 ). Only in cooler areas, such as the Swiss Jura in cool summers, may the whole population enter into dia-pause after two generations. In southern Europe, four com-plete generations are commonly observed ( Del Bene et al. , 2001; Hellrigl, 2001 ). Non-overwintering generations have
Figure 4 Net reproductive rate ( R 0 ) and total mortality affecting first, intermediate and last generations of Cameraria ohridella in Delémont, Switzerland, over the years 2002 – 04.
Ap
par
ent
mor
talit
y
Rea
l mor
talit
y
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Pot
entia
l F 1
p
rog
eny
19
911.
94
8523
.72
0.
4281
0.42
81
Rea
lized
fecu
ndity
58.7
6
8523
.72
0.42
81
0.
4281
0.42
81
0.24
27
Rea
lized
F 1
p
rog
eny
11
388.
22
R 0
( F 1 /
l 0 )
11
.388
2
Tab
le 4
Co
nti
nu
ed

Life tables of C. ohridella 149
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Tab
le 5
Life
tab
le f
or t
he s
econ
d,
over
win
terin
g g
ener
atio
n of
Cam
erar
ia o
hrid
ella
in D
elém
ont,
Sw
itzer
land
, 20
02
Ap
par
ent
mor
talit
y
Rea
l mor
talit
y
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Eg
g
10
00.0
0 36
3.64
0.36
36
0.
3636
E
gg
s d
isap
pea
r of
f lea
f
363.
6364
0.
3636
0.36
36
0.
3636
0.
1963
L1
636.
36
7.95
0.01
25
0.
0080
Le
af s
enes
cenc
e
7.95
45
0.01
25
0.
0125
0.00
80
0.00
55
L2
62
8.41
60
.45
0.
0962
0.06
05
Pla
nt d
efen
se r
eact
ion
25
.454
5 0.
0417
0.04
05
0.
0255
0.
0185
Leaf
sen
esce
nce
15
.909
1 0.
0253
0.02
53
0.
0159
0.
0111
Oth
er
15
.909
1 0.
0263
0.02
53
0.
0159
0.
0116
Hos
t fee
din
g
1.
5909
0.
0027
0.00
25
0.
0016
0.
0012
Pre
dat
ion
(art
hrop
ods)
1.59
09
0.00
27
0.
0025
0.00
16
0.00
12
L3
56
7.95
39
.77
0.
0700
0.03
98
Oth
er
28
.636
4 0.
0509
0.05
04
0.
0286
0.
0227
Leaf
sen
esce
nce
9.
5455
0.
0168
0.01
68
0.
0095
0.
0074
Pre
dat
ion
(art
hrop
ods)
1.59
09
0.00
29
0.
0028
0.00
16
0.00
13
L4-5
528.
18
105.
00
0.
1988
0.10
50
Leaf
sen
esce
nce
52
.500
0 0.
0994
0.09
94
0.
0525
0.
0455
Oth
er
42
.954
5 0.
0867
0.08
13
0.
0430
0.
0394
Pre
dat
ion
(bird
s)
4.
7727
0.
0100
0.00
90
0.
0048
0.
0044
Pre
dat
ion
(art
hrop
ods)
3.18
18
0.00
67
0.
0060
0.00
32
0.00
29
H
ost f
eed
ing
1.59
09
0.00
34
0.
0030
0.00
16
0.00
15
Sp
in
42
3.18
22
0.85
0.52
19
0.
2208
Le
af s
enes
cenc
e
187.
4367
0.
4429
0.44
29
0.
1874
0.
2541
Pre
dat
ion
(bird
s)
17
.500
0 0.
0568
0.04
14
0.
0175
0.
0254
Oth
er
9.
5455
0.
0314
0.02
26
0.
0095
0.
0139
Par
asiti
sm
4.
7727
0.
0158
0.01
13
0.
0048
0.
0069
Pre
dat
ion
(art
hrop
ods)
1.59
09
0.00
53
0.
0038
0.00
16
0.00
23
Pup
ae
20
2.34
11
.09
0.
0548
0.01
11
Pre
dat
ion
(bird
s)
5.
5435
0.
0274
0.02
74
0.
0055
0.
0121
Oth
er
4.
6195
0.
0232
0.02
28
0.
0046
0.
0102
Hos
t fee
din
g
0.
9239
0.
0047
0.00
46
0.
0009
0.
0020
P
upae
in d
iap
ause
19
1.25
18
2.17
0.95
25
0.
1822
E
arth
wor
ms
17
0.82
83
0.94
26
0.
8932
0.17
08
1.24
10
O
ther
9.42
50
0.14
59
0.
0493
0.00
94
0.06
85
P
aras
itism
1.91
25
0.03
15
0.
0100
0.00
19
0.01
39
Ad
ults
9.08
Sex
rat
io
0.
5
Pot
entia
l fec
und
ity
10
2.74
Pot
entia
l F 1
pro
gen
y
466.
61
292.
43
0.
6267
0.62
67
R
ealiz
ed fe
cund
ity
38
.35
29
2.43
0.
6267
0.62
67
0.
6267
0.
4279
R
ealiz
ed F
1 p
rog
eny
17
4.19
R 0
( F 1 /
l 0 )
0.
1742
150 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Tab
le 6
Life
tab
le f
or t
he f
irst
gen
erat
ion
of C
amer
aria
ohr
idel
la in
Del
émon
t, S
witz
erla
nd,
2003
Ap
par
ent
mor
talit
y
Rea
l mor
talit
y
Sta
ge
Fa
ctor
Mar
gin
al a
ttack
ra
te
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Eg
g
10
00
178.
57
0.
178
6
0.17
86
Eg
gs
dis
app
ear
off l
eaf
13
3.92
86
0.13
39
0.
1339
0.13
39
0.06
24
E
gg
s fl a
t and
em
pty
35.7
143
0.03
86
0.
0357
0.03
57
0.01
71
E
gg
s op
ened
8.92
86
0.00
98
0.
0089
0.00
89
0.00
43
L1
82
1.43
12
.32
0.
0150
0.01
23
Pla
nt d
efen
se r
eact
ion
10
.267
9 0.
0125
0.01
25
0.
0103
0.
0055
Oth
er
2.
0536
0.
0025
0.00
25
0.
0021
0.
0011
L2
809.
11
197.
14
0.
2437
0.19
71
Pla
nt d
efen
se r
eact
ion
10
6.78
57
0.14
04
0.
1320
0.10
68
0.06
57
P
aras
itism
39.0
179
0.05
38
0.
0482
0.03
90
0.02
40
O
ther
24.6
429
0.03
43
0.
0305
0.02
46
0.01
52
H
ost f
eed
ing
14.3
750
0.02
02
0.
0178
0.01
44
0.00
88
Le
af s
enes
cenc
e
8.21
43
0.01
02
0.
0102
0.00
82
0.00
44
P
red
atio
n (a
rthr
opod
s)
4.
1071
0.
0058
0.00
51
0.
0041
0.
0025
L3
611.
96
49.2
9
0.08
05
0.
0493
Le
af s
enes
cenc
e
16.4
286
0.02
68
0.
0268
0.01
64
0.01
18
O
ther
10.2
679
0.01
73
0.
0168
0.01
03
0.00
76
P
aras
itism
10.2
679
0.01
73
0.
0168
0.01
03
0.00
76
P
red
atio
n (a
rthr
opod
s)
6.
1607
0.
0104
0.01
01
0.
0062
0.
0046
Hos
t fee
din
g
4.
1071
0.
0070
0.00
67
0.
0041
0.
0030
Pre
dat
ion
(bird
s)
2.
0536
0.
0035
0.00
34
0.
0021
0.
0015
L4
562.
68
20.5
4
0.03
65
0.
0205
P
red
atio
n (a
rthr
opod
s)
12
.321
4 0.
0221
0.02
19
0.
0123
0.
0097
Pre
dat
ion
(bird
s)
6.
1607
0.
0109
0.01
09
0.
0062
0.
0048
Par
asiti
sm
2.
0536
0.
0037
0.00
36
0.
0021
0.
0016
S
pin
542.
14
26.1
0
0.04
81
0.
0261
P
red
atio
n (b
irds)
10.0
397
0.01
85
0.
0185
0.01
00
0.00
81
P
aras
itism
10.0
397
0.01
88
0.
0185
0.01
00
0.00
82
P
red
atio
n (a
rthr
opod
s)
4.
0159
0.
0076
0.00
74
0.
0040
0.
0033
Oth
er
2.
0079
0.
0038
0.00
37
0.
0020
0.
0016
P
upae
516.
04
68.3
9
0.13
25
0.
0684
P
upae
in d
iap
ause
26.9
418
0.05
45
0.
0522
0.02
69
0.02
43
P
red
atio
n (b
irds)
16.5
796
0.03
21
0.
0321
0.01
66
0.01
42
P
red
atio
n (a
rthr
opod
s)
14
.507
1 0.
0297
0.02
81
0.
0145
0.
0131
Par
asiti
sm
10
.362
2 0.
0213
0.02
01
0.
0104
0.
0094
A
dul
ts
44
7.65
Sex
rat
io
0.
54
P
oten
tial f
ecun
dity
102.
74
P
oten
tial F
1 p
rog
eny
24
881.
31
1513
1.70
0.60
82
0.
6082
Rea
lized
fecu
ndity
40.2
593
15
131.
70
0.60
82
0.
6082
0.60
82
0.40
69
Rea
lized
F 1
pro
gen
y
9749
.61
R
0 ( F
1 / l 0 )
9.74
96
Life tables of C. ohridella 151
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Tab
le 7
Life
tab
le f
or t
he s
econ
d g
ener
atio
n of
Cam
erar
ia o
hrid
ella
in D
elém
ont,
Sw
itzer
land
, 20
03
Ap
par
ent
mor
talit
y R
eal m
orta
lity
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Eg
g
10
00
178.
57
0.
1786
0.17
86
Eg
gs
dis
app
ear
off l
eaf
13
3.92
86
0.13
39
0.
1339
0.13
39
0.06
24
E
gg
s fl a
t and
em
pty
35.7
143
0.03
86
0.
0357
0.03
57
0.01
71
E
gg
s op
ened
8.92
86
0.00
98
0.
0089
0.00
89
0.00
43
L1
82
1.43
41
.07
0.
0500
0.04
11
Pla
nt d
efen
se r
eact
ion
28
.750
0 0.
0353
0.03
50
0.
0288
0.
0156
Oth
er
10
.267
9 0.
0127
0.01
25
0.
0103
0.
0056
Leaf
sen
esce
nce
2.
0536
0.
0025
0.00
25
0.
0021
0.
0011
L2
780.
36
84.2
0
0.10
79
0.
0842
P
lant
def
ense
rea
ctio
n
47.2
321
0.06
20
0.
0605
0.04
72
0.02
78
O
ther
32.8
571
0.04
36
0.
0421
0.03
29
0.01
93
P
red
atio
n (b
irds)
2.05
36
0.00
28
0.
0026
0.00
21
0.00
12
Le
af s
enes
cenc
e
2.05
36
0.00
26
0.
0026
0.00
21
0.00
11
L3
69
6.16
51
.34
0.
0737
0.05
13
Pre
dat
ion
(art
hrop
ods)
14.3
750
0.02
12
0.
0206
0.01
44
0.00
93
P
red
atio
n (b
irds)
12.3
214
0.01
82
0.
0177
0.01
23
0.00
80
O
ther
12.3
214
0.01
82
0.
0177
0.01
23
0.00
80
Le
af s
enes
cenc
e
4.10
71
0.00
59
0.
0059
0.00
41
0.00
26
P
lant
def
ense
rea
ctio
n
4.10
71
0.00
61
0.
0059
0.00
41
0.00
27
P
aras
itism
4.10
71
0.00
61
0.
0059
0.00
41
0.00
27
L4
64
4.82
80
.09
0.
1242
0.08
01
Pre
dat
ion
(bird
s)
45
.178
6 0.
0721
0.07
01
0.
0452
0.
0325
Pre
dat
ion
(art
hrop
ods)
12.3
214
0.02
02
0.
0191
0.01
23
0.00
89
O
ther
12.3
214
0.02
02
0.
0191
0.01
23
0.00
89
Le
af s
enes
cenc
e
6.16
07
0.00
96
0.
0096
0.00
62
0.00
42
P
aras
itism
4.10
71
0.00
68
0.
0064
0.00
41
0.00
30
Sp
in
56
4.73
76
.75
0.
1359
0.07
68
Pre
dat
ion
(bird
s)
61
.607
1 0.
1106
0.10
91
0.
0616
0.
0509
Pre
dat
ion
(art
hrop
ods)
8.21
43
0.01
55
0.
0145
0.00
82
0.00
68
P
aras
itism
4.10
71
0.00
78
0.
0073
0.00
41
0.00
34
Le
af s
enes
cenc
e
2.82
37
0.00
50
0.
0050
0.00
28
0.00
22
Pup
ae
48
7.98
26
3.55
0.54
01
0.
2636
P
red
atio
n (b
irds)
181.
1909
0.
4137
0.37
13
0.
1812
0.
2319
Leaf
sen
esce
nce
70
.005
6 0.
1435
0.14
35
0.
0700
0.
0673
Oth
er
10
.294
9 0.
0299
0.02
11
0.
0103
0.
0132
Par
asiti
sm
2.
0590
0.
0060
0.00
42
0.
0021
0.
0026
A
dul
ts
22
4.43
Sex
rat
io
0.
54
P
oten
tial f
ecun
dity
100.
248
P
oten
tial F
1 p
rog
eny
12
171.
38
8043
.35
0.
6608
0.66
08
R
ealiz
ed fe
cund
ity
34
8043
.35
0.66
08
0.
6608
0.66
08
0.46
96
Rea
lized
F 1
pro
gen
y
4128
.03
R
0 ( F
1 / l 0 )
4.12
80
152 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Tab
le 8
Life
tab
le f
or t
he t
hird
, ov
ewin
terin
g g
ener
atio
n of
Cam
erar
ia o
hrid
ella
in D
elém
ont,
Sw
itzer
land
, 20
03
Ap
par
ent
mor
talit
y R
eal m
orta
lity
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Eg
g
10
00
756.
10
0.
7561
0.75
61
Eg
gs
dis
app
ear
off l
eaf
38
2.11
38
0.38
21
0.
3821
0.38
21
0.20
91
E
gg
s fl a
t and
em
pty
361.
7886
0.
4909
0.36
18
0.
3618
0.
2932
Eg
gs
open
ed
12
. 195
1 0.
0225
0.01
22
0.
0122
0.
0099
L1
243.
90
17.6
8
0.07
25
0.
0177
O
ther
9.14
63
0.03
82
0.
0375
0.00
91
0.01
69
P
lant
def
ense
rea
ctio
n
8.53
66
0.03
57
0.
0350
0.00
85
0.01
58
L2
22
6.22
25
.61
0.
1132
0.02
56
Leaf
sen
esce
nce
10
.365
9 0.
0458
0.04
58
0.
0104
0.
0204
Oth
er
7.
9268
0.
0365
0.03
50
0.
0079
0.
0162
Pla
nt d
efen
se r
eact
ion
7.
3171
0.
0337
0.03
23
0.
0073
0.
0149
L3
200.
61
27.4
4
0.13
68
0.
0274
Le
af s
enes
cenc
e
10.9
756
0.05
47
0.
0547
0.01
10
0.02
44
O
ther
8.53
66
0.04
47
0.
0426
0.00
85
0.01
99
P
aras
itism
3.65
85
0.01
94
0.
0182
0.00
37
0.00
85
H
ost f
eed
ing
2.43
90
0.01
30
0.
0122
0.00
24
0.00
57
P
red
atio
n (a
rthr
opod
s)
1.
2195
0.
0065
0.00
61
0.
0012
0.
0028
Pre
dat
ion
(bird
s)
0.
6098
0.
0033
0.00
30
0.
0006
0.
0014
L4
173.
17
68.9
0
0.39
79
0.
0689
Le
af s
enes
cenc
e
34.7
561
0. 2
007
0.
200
7
0.03
48
0.09
73
O
ther
14.6
341
0.10
21
0.
0845
0.01
46
0.04
68
P
red
atio
n (b
irds)
8.53
66
0.06
09
0.
0493
0.00
85
0.02
73
P
aras
itism
7.92
68
0.05
67
0.
0458
0.00
79
0.02
53
P
red
atio
n (a
rthr
opod
s)
2.
4390
0.
0178
0.01
41
0.
0024
0.
0078
Hos
t fee
din
g
0.
6098
0.
0045
0.00
35
0.
0006
0.
0019
S
pin
104.
27
73.7
8
0.70
76
0.
0738
Le
af s
enes
cenc
e
39.0
244
0.37
43
0.
3743
0.03
90
0.20
36
P
red
atio
n (b
irds)
32.3
171
0.41
64
0.
3099
0.03
23
0.23
39
P
aras
itism
2.43
90
0.03
98
0.
0234
0.00
24
0.01
77
Pup
ae
30
.49
4.27
0.14
00
0.
0043
P
red
atio
n (b
irds)
4.26
83
0.14
00
0.
1400
0.00
43
0.06
55
Pup
ae in
dia
pau
se
26.2
2 22
.64
0.
8636
0.02
26
Ear
thw
orm
s
22.3
0 0.
8595
0.85
06
0.
0223
0.
8523
Par
asiti
sm
0.
17
0.01
49
0.
0065
0.00
02
0.00
65
O
ther
0.17
0.
0149
0.00
65
0.
0002
0.
0065
A
dul
ts
3.
58
S
ex r
atio
0.54
Pot
entia
l fec
und
ity
10
0.24
8
Pot
entia
l F 1
pro
gen
y
193.
90
128.
14
0.
6608
0.66
08
R
ealiz
ed fe
cund
ity
34
128.
14
0.66
08
0.
6608
0.66
08
0.46
96
Rea
lized
F 1
pro
gen
y
65.7
6
R 0
( F 1 /
l 0 )
0.
0658

Life tables of C. ohridella 153
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Tab
le 9
Life
tab
le f
or t
he f
irst
gen
erat
ion
of C
amer
aria
ohr
idel
la in
Del
émon
t, S
witz
erla
nd,
2004
Ap
par
ent
mor
talit
y R
eal m
orta
lity
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Eg
g
10
00
316.
42
0.
3164
0.31
64
Eg
gs
dis
app
ear
off l
eaf
23
9.72
38
0.23
97
0.
2397
0.23
97
0.11
90
E
gg
s fl a
t and
em
pty
70.6
389
0.08
14
0.
0706
0.07
06
0.03
69
E
gg
s op
ened
6.06
06
0.00
73
0.
0061
0.00
61
0.00
32
L1
68
3.58
51
.27
0.
0750
0.05
13
Pla
nt d
efen
se r
eact
ion
41
.014
6 0.
0605
0.06
00
0.
0410
0.
0271
Leaf
sen
esce
nce
6.
8358
0.
0100
0.01
00
0.
0068
0.
0044
Oth
er
3.
4179
0.
0052
0.00
50
0.
0034
0.
0023
L2
632.
31
93.9
9
0.14
86
0.
0940
P
lant
def
ense
rea
ctio
n
49.5
593
0.08
14
0.
0784
0.04
96
0.03
69
O
ther
18.7
984
0.03
17
0.
0297
0.01
88
0.01
40
P
red
atio
n (a
rthr
opod
s)
10
.253
7 0.
0174
0.01
62
0.
0103
0.
0076
Leaf
sen
esce
nce
8.
5447
0.
0135
0.01
35
0.
0085
0.
0059
Hos
t fee
din
g
5.
1268
0.
0087
0.00
81
0.
0051
0.
0038
Par
asiti
sm
1.
7089
0.
0029
0.00
27
0.
0017
0.
0013
L3
538.
32
30.7
6
0.05
71
0.
0308
P
red
atio
n (a
rthr
opod
s)
18
.798
4 0.
0349
0.03
49
0.
0188
0.
0154
Par
asiti
sm
5.
1268
0.
0098
0.00
95
0.
0051
0.
0043
Oth
er
3.
4179
0.
0065
0.00
63
0.
0034
0.
0028
Pla
nt d
efen
se r
eact
ion
3.
4179
0.
0065
0.00
63
0.
0034
0.
0028
L4
507.
56
109.
37
0.
2155
0.10
94
Pre
dat
ion
(art
hrop
ods)
97.4
097
0.19
44
0.
1919
0.09
74
0.09
39
P
red
atio
n (b
irds)
3.41
79
0.00
76
0.
0067
0.00
34
0.00
33
Le
af s
enes
cenc
e
3.41
79
0.00
67
0.
0067
0.00
34
0.00
29
P
aras
itism
3.41
79
0.00
76
0.
0067
0.00
34
0.00
33
O
ther
1.70
89
0.00
38
0.
0034
0.00
17
0.00
16
Sp
in
39
8.18
44
.43
0.
1116
0.04
44
Pre
dat
ion
(art
hrop
ods)
27.3
431
0.07
02
0.
0687
0.02
73
0.03
16
P
red
atio
n (b
irds)
6.83
58
0.01
80
0.
0172
0.00
68
0.00
79
Le
af s
enes
cenc
e
5.12
68
0.01
29
0.
0129
0.00
51
0.00
56
P
aras
itism
3.41
79
0.00
91
0.
0086
0.00
34
0.00
40
O
ther
1.70
89
0.00
45
0.
0043
0.00
17
0.00
20
Pup
ae
35
3.75
90
.57
0.
2560
0.09
06
Pre
dat
ion
(art
hrop
ods)
35.8
878
0.11
06
0.
1014
0.03
59
0.05
09
Le
af s
enes
cenc
e
23.9
252
0.06
76
0.
0676
0.02
39
0.03
04
P
red
atio
n (b
irds)
11.9
626
0.03
83
0.
0338
0.01
20
0.01
70
O
ther
8.54
47
0.02
75
0.
0242
0.00
85
0.01
21
P
aras
itism
6.83
58
0.02
21
0.
0193
0.00
68
0.00
97
Con
tinue
d

154 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
high net rates of increase, as shown in this study, which allows C. ohridella to build outbreak populations year after year.
Parasitoids are known as the most important natural ene-mies of leaf miners ( Askew & Shaw, 1979; Faeth & Simberloff, 1981; Van Driesche & Taub, 1983; Hespenheide, 1991; Hawkins et al. , 1997 ) and parasitism rates of 40% and higher are commonly cited. Faeth & Simberloff (1981) men-tioned parasitism rates of 40 – 50% on a Cameraria sp. and noted that exclusion of parasitoids considerably reduced the mortality of larvae. Askew & Shaw (1979) studied Phyllonorycter spp. on oak and birch and found out that pop-ulations were regulated and kept at low levels almost entirely by the action of parasitoids. With C. ohridella being an ex-otic insect in central Europe, it is expected to be less heavily parasitized than native leaf miners. However, leaf miners are notorious among insects for being parasitized, mainly by polyphagous parasitoids attacking various groups of leaf miners on different host plants. Consequently, it is common for exotic leaf miners to become heavily parasitized by na-tive parasitoids. Examples include Phyllonorycter robiniella introduced in Europe in the 1980s, which exhibits parasitism rates of 30 – 70% ( Stojanovic & Markovic, 2005 ), Phyllocnistis citrella , introduced to Spain in 1993 and parasitized up to 70% by native parasitoids 4 years later ( Urbaneja et al. , 2000 ), and Phyllonorycter leucographella , which was al-ready parasitized at 56% 16 years after its introduction to England ( Godfray et al. , 1995 ). In Europe, C. ohridella has indeed been adopted by over 30 native, polyphagous parasi-toids. However, parasitism rates remain low (Grabenweger et al. , 2005a; Girardoz et al. , 2006; Volter & Kenis, 2006) . Several reasons have been proposed to explain the low para-sitism rates observed in C. ohridella . One of them is a lack of synchronization between parasitoid emergence and the first generation of C. ohridella in spring ( Grabenweger, 2004; Girardoz et al. , 2006 ). Indeed, the bulk of the parasitoids emerge at least 6 weeks before the first suitable C. ohridella larvae are available, suggesting that alternate hosts are needed. However, Girardoz et al. (2006) showed that parasit-ism is not greater in forests, where alternate hosts are sup-posed to be more available, than in cities. Furthermore, parasitism rates do not increase in the subsequent generations in summer and autumn, when generations overlap and syn-chronization is not problematical. Other possible reasons for low observed parasitism rates could be that parasitoids are poorly attracted or even repelled by A. hippocastanum , which does not host any other leaf miner, or the low physiological suitability of larvae and pupae of the non-European genus Cameraria for European parasitoids. Laboratory rearing tests and olfactometry observations are needed to test these hypotheses.
Predation rates were very variable but were often greater than parasitism rates. Predation by birds was often the main mortality factor on mature larvae of non-overwintering gen-erations. Predation has been poorly studied in C. ohridella . Zegula et al. (2002) and Radeghieri (2004) made observa-tions on bird and ant predation, respectively, but it is the re-cent study by Grabenweger et al. (2005b) that provides the most complete information on predation on C. ohridella . They determined and quantified invertebrate and bird
A
pp
aren
t m
orta
lity
Rea
l mor
talit
y
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
P
upae
in d
iap
ause
3.41
79
0.01
11
0.
0097
0.00
34
0.00
48
Ad
ults
263.
18
S
ex r
atio
0.54
Pot
entia
l fec
und
ity
10
2.74
Pot
entia
l F 1
pro
gen
y
1462
7.96
88
96.0
7
0.60
82
0.
6082
Rea
lized
fecu
ndity
40.2
593
88
96.0
7 0.
6082
0.60
82
0.
6082
0.
4069
R
ealiz
ed F
1 p
rog
eny
57
31.8
9
R 0
( F 1 /
l 0 )
5.
7319
Tab
le 9
Co
nti
nu
ed

Life tables of C. ohridella 155
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Tab
le 1
0 Li
fe t
able
for
the
sec
ond
, ov
erw
inte
ring
gen
erat
ion
of C
amer
aria
ohr
idel
la in
Del
émon
t, S
witz
erla
nd,
2004
Ap
par
ent
mor
talit
y
Rea
l mor
talit
y
Sta
ge
Fa
ctor
M
arg
inal
atta
ck
rate
Sta
ge
Fact
or
Sta
ge
Fact
or
k -va
lue
Sta
ge
Fact
or
l x d
x d
x q
x q
x d
x / l 0
d x /
l 0 Fa
ctor
Eg
g
10
00
311.
02
0.
3110
0.31
10
Eg
gs
dis
app
ear
off l
eaf
30
6.47
81
0.30
65
0.
3065
0.30
65
0.15
89
E
gg
s fl a
t and
em
pty
4.54
55
0.00
66
0.
0045
0.00
45
0.00
29
L1
68
8.98
29
.28
0.
0425
0.02
93
Pla
nt d
efen
se r
eact
ion
17
.224
4 0.
0252
0.02
50
0.
0172
0.
0111
Oth
er
8.
6122
0.
0127
0.01
25
0.
0086
0.
0055
Leaf
sen
esce
nce
3.
4449
0.
0050
0.00
50
0.
0034
0.
0022
L2
659.
69
60.2
9
0.09
14
0.
0603
P
lant
def
ense
rea
ctio
n
34.4
488
0.05
33
0.
0522
0.03
44
0.02
38
O
ther
17.2
244
0.02
70
0.
0261
0.01
72
0.01
19
Le
af s
enes
cenc
e
5.16
73
0.00
78
0.
0078
0.00
52
0.00
34
P
red
atio
n (a
rthr
opod
s)
3.
4449
0.
0055
0.00
52
0.
0034
0.
0024
L3
599.
41
29.2
8
0.04
89
0.
0293
P
red
atio
n (a
rthr
opod
s)
17
.224
4 0.
0290
0.02
87
0.
0172
0.
0128
Leaf
sen
esce
nce
8.
6122
0.
0144
0.01
44
0.
0086
0.
0063
Oth
er
3.
4449
0.
0059
0.00
57
0.
0034
0.
0026
L4
570.
13
55.1
2
0.09
67
0.
0551
P
red
atio
n (b
irds)
15.5
020
0.02
82
0.
0272
0.01
55
0.01
24
P
red
atio
n (a
rthr
opod
s)
13
.779
5 0.
0251
0.02
42
0.
0138
0.
0110
Oth
er
12
.057
1 0.
0220
0.02
11
0.
0121
0.
0097
Leaf
sen
esce
nce
10
.334
6 0.
0181
0.01
81
0.
0103
0.
0079
Par
asiti
sm
1.
7224
0.
0032
0.00
30
0.
0017
0.
0014
Hos
t fee
din
g
1.
7224
0.
0032
0.00
30
0.
0017
0.
0014
S
pin
515.
01
51.6
7
0.10
03
0.
0517
Le
af s
enes
cenc
e
29.2
815
0.05
69
0.
0569
0.02
93
0.02
54
P
red
atio
n (b
irds)
18.9
469
0.03
68
0.
0368
0.01
89
0.01
63
P
red
atio
n (a
rthr
opod
s)
3.
4449
0.
0069
0.00
67
0.
0034
0.
0030
P
upae
463.
34
36.1
7
0.07
81
0.
0362
P
red
atio
n (b
irds)
29.2
815
0.06
32
0.
0632
0.02
93
0.02
84
P
red
atio
n (a
rthr
opod
s)
3.
4449
0.
0077
0.00
74
0.
0034
0.
0034
Oth
er
3.
4449
0.
0077
0.00
74
0.
0034
0.
0034
P
upae
in d
iap
ause
42
7.17
38
7.15
0.90
63
0.
3871
E
arth
wor
ms
37
2.46
12
0.89
75
0.
8719
0.37
25
0.98
93
O
ther
11.9
125
0.07
03
0.
0279
0.01
19
0.03
16
P
aras
itism
2.77
38
0.01
68
0.
0065
0.00
28
0.00
74
Ad
ults
40.0
2
Sex
rat
io
0.
54
P
oten
tial f
ecun
dity
100.
25
P
oten
tial F
1 p
rog
eny
21
70.2
7 14
34.2
1
0.66
08
0.
6608
Rea
lized
fecu
ndity
34.0
0
1434
.21
0.66
08
0.
6608
0.66
08
0.46
96
Rea
lized
F 1
pro
gen
y
736.
07
R
0 ( F
1 / l 0 )
0.73
61

156 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
predation in Austria and Switzerland and identified tits and the bush-cricket Meconema meridionale as the main preda-tors, which confirms our observations in Switzerland. Both Grabenweger et al. (2005b) and our study found that bird predation was mainly on spinning larvae and pupae whereas invertebrate predation occurred nearly exclusively on the last feeding stages. Grabenweger et al. (2005b) found M. meridi-onale to have a measurable negative impact on C. ohridella populations. Bird predation was estimated at 2 – 4% (i.e. of similar magnitude as parasitism). Although the comparison of predation rates between the two studies is difficult because of the different methods of measurement used, it appears that predation by birds was sometimes much greater in our study. For example, in the second and third generations of 2003, birds ate 7 – 41% of each of the stages L4, spinning larvae and pupae. In the second moth generation, birds killed 29% of the total generation, as shown by the real mortality rates in Table 7 . The fact that predation by birds was much higher in 2003, when moth populations were highest, compared with the two other years, suggests that predation may be density dependent, although Grabenweger et al. (2005b) could not draw any clear conclusions regarding density-dependence. Avian predation on C. ohridella requires further research, particularly its density-dependence and its evolution over time. Considering the great learning capacity of tits, it cannot be ruled out that they will adapt their behaviour to this new prey at a faster rate than arthropod predators or parasitoids ( Grabenweger et al. , 2005b ).
Predation on leaf-mining Lepidoptera by birds and inverte-brates has often been documented, but few studies provide a precise account of mortality due to all predators. Kirizoglu (1976) measured predation rates on larvae of Caloptilia syring-ella due to invertebrates of 33 and 50% in a spring and autumn generation, respectively. Predators were also reported to kill 24.7% of the leaf-mining stages of Cameraria hamadryadella ( Connor & Beck, 1993 ), 10 – 30% of those of a Cameraria sp. ( Faeth & Simberloff, 1981 ) and up to 60% of those of the inva-sive Phyllocnistis citrella ( Urbaneja et al. , 2000 ).
The main mortality factors of C. ohridella occur on the last, overwintering generation, which is the only one having a net rate of increase usually lower than one. Indeed, it is well known that, in most cases, population densities in spring are lower than in the previous generation in autumn. Our ob-servations showed that nearly 90% of the pupae in dead leaves died during winter, a proportion that corresponds to what other authors have found on Phyllonorycter spp. Pottinger & Leroux (1971) found that 84% of populations of P. blancardella died during the winter and for Phyllonorycter spp. on oak, this percentage was of 50% ( Connor, 1984 ). Laing et al. (1986) determined that the majority of this mor-tality is due to earthworms, which bury and eat the leaves and therefore destroy the pupae or prevent the hatching adults from reaching the surface. Our study was made at one loca-tion only and it remains to be determined whether earth-worms and other leaf decomposers have a similar impact in other localities and on other soils. The observations were made in the countryside, at the edge of a meadow and a for-est. In urban areas, where most horse-chestnut trees are found in Central Europe, most dead leaves are removed or blown
away, which probably has a similar effect on moth popula-tions as leaf decomposers in rural areas. Leaf removal in au-tumn has an important impact on populations of C. ohridella in the next spring ( Gilbert et al. , 2003; Pavan et al. , 2003 ), suggesting again that winter mortality has a key role in the population dynamics of the moth.
Other major causes of mortality in the last generation are leaf senescence and intraspecific competition. Climatic con-ditions during the summer largely determine the synchroni-zation between the development of C. ohridella and leaf senescence in autumn. In Delémont, during the relatively cool summers in 2002 and 2004, the development of C. ohridella was rather slow and only two generations devel-oped. Nearly all pupae of the second generation entered into diapause and, consequently, populations in the next year in-creased compared with the previous year. By contrast, in 2003, the unusually warm summer induced a third genera-tion. Only a part of this third generation reached the pupal stage due to high larval mortality, which resulted in a low level of attack in spring 2004.
When horse-chestnuts are heavily attacked by C. ohridella , larvae may die from intraspecific competition, and leaf se-nescence occurs earlier than normal ( Pschorn-Walcher, 2001 ), a phenomenon that is often seen in leaf-miners when they occur at high densities ( Quiring & McNeil, 1984; Faeth, 1992; Connor & Beck, 1993 ). Connor & Beck (1993) even described epidemic years of Cameraria hamadryadella (Clemens) where 53% of the population died as a result of intraspecific competition and only 0.6% of the population survived. Intra-specific competition did not appear as an im-portant mortality factor in our life tables. In 2002 and 2004, populations were too low to suffer from competition. In 2003, mortality through intraspecific competition was re-corded, in the ‘other mortality’ category, but was probably largely underestimated, especially in the last generations. First, leaf samples were always taken from the trees, even late in the season when the heaviest attacked leaves (i.e. those in which intraspecific competition was most likely to occur) had already dropped. Second, leaf senescence and in-traspecific competition are clearly overlapping mortality fac-tors. High mine densities (i.e. intraspecific competition) cause early leaf senescence and, most probably, delays in lar-val development due to lower food quality. Thus, at least a part of the mortality attributed to leaf senescence could have been attributed to intraspecific competition.
Acknowledgements
We thank Giselher Grabenweger (BOKU, Vienna) and Hannes Bauer (Natural History Museum, Bern) for their help in the identification of parasitoids, Natalia Guazzone, Sébastien Marcombe, Ghislaine Cortat and Audrey Favereau for their help in field and laboratory work, Hefin Jones and Jon Knight for fruitful discussion and comments on the manuscript, and Werner Heitland for his support during the whole study. The project was funded by the Swiss Federal Office for Education and Science as part of the European Union FWP5 project CONTROCAM (QLK5-CT-2000-01684), and by the EU FWP6 project ALARM (GOCE-CT-2003-506675).

Life tables of C. ohridella 157
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
References
Askew , R.R. & Shaw , M.R . ( 1979 ) Mortality factors affecting the leaf-mining stages of Phyllonorycter (Lepidoptera: Gracillariidae) on oak and birch. 1. Analysis of the mortality factors . Zoological Journal of the Linnean Society , 67 , 31 – 49 .
Awmack , C.S. & Leather , S.R . ( 2002 ) Host plant quality and fecun-dity in herbivorous insects . Annual Review of Entomology , 47 , 817 – 844 .
Barrett , B.A. & Brunner , J.F . ( 1988 ) Fecundity, longevity, and devel-opmental rates of Phyllonorycter elmaella (Lepidoptera: Gracil-lariidae) and its major parasitoid, Pnigalio flavipes (Hymenoptera: Eulophidae) . Melanderia , 46 , 29 – 35 .
Bellows , T.S. Jr , Van Driesche , R.G. & Elkinton , J.S . ( 1992 ) Life- table construction and analysis in the evaluation of natural enemies . Annual Review of Entomology , 37 , 587 – 614 .
Berryman , A.A . ( 1987 ) The theory and classification of outbreaks . Insect Pest Outbreaks: Ecological and Evolutionary Perspectives ( ed. by P. Barbosa and J. C. Schultz .), pp . 3 – 30 . Academic Press , New York, New York .
Connor , E.F . ( 1984 ) The causes of overwintering mortality of Phyl-lonorycter on Quercus robur . Ecological Entomology , 9 , 23 – 28 .
Connor , E.F. & Beck , M.W . ( 1993 ) Density -related mortality in Cameraria hamadryadella (Lepidoptera: Gracillariidae) at epi-demic and endemic densities . Oikos , 66 , 515 – 525 .
Courtney , S.P. & Duggan , A.E . ( 1983 ) The population dynamics of the orange tip butterfly, Anthocharis cardamines , in Britain . Ecological Entomology , 8 , 271 – 281 .
Del Bene , G. , Gargani , E. , Landi , S. & Bonifacio , A . ( 2001 ) Camer-aria ohridella e malattie fogliari dell’ippocastano in Toscana . Italus Horus , 8 , 41 – 49 .
Deschka , G. & Dimic , N . ( 1986 ) Cameraria ohridella sp. n. (Lep., Lithocolletidae) aus Mazedonien, Jugoslawien . Acta Entomolog-ica Jugoslavica , 22 , 11 – 23 .
Elkinton , S. , Buonaccorsi , J.P. , Bellows , T.S. Jr & Van Driesche , R.G . ( 1992 ) Marginal attack rate, k-values and density depen-dence in the analysis of contemporaneous mortality factors . Research on Population Ecology , 34 , 29 – 44 .
Faeth , S.H . ( 1992 ) Interspecific and intraspecific interactions via plant responses to folivory: an experimental field test . Ecology , 73 , 1802 – 1813 .
Faeth , S.H. & Simberloff , D . ( 1981 ) Population regulation of a leaf-mining insect, Cameraria sp. Nov at increased field densities . Ecology , 62 , 620 – 624 .
Freise , J.F . ( 2001 ) Untersuchungen zur Biologie und Ökologie der Roßkastanien-Miniermotte ( Cameraria Ohridella DESCH. & DIM 1986) (Lepidoptera: Gracillariidae) . PhD Thesis, Technische Uni-versität München . Munich, Germany .
Freise , J.F. & Heitland , W . ( 1999 ) A brief note on sexual differences in pupae of the horse-chestnut leaf miner, Cameraria ohridella Deschka & Dimic (1986) (Lep., Gracillariidae), a new pest in Central Europe on Aesculus hippocastanum . Journal of Applied Entomology , 123 , 191 – 192 .
Freise , J. & Heitland , W . ( 2001 ) Neue Aspekte zur Biologie und Ökologie der Roßkastanien-Miniermotte, Cameraria ohridella Deschka & Dimic (1986) (Lep., Gracillariidae), einem neuartigen Schädling an Aesculus hippocastanum . Mitteilungen der Deutschen Gesellschaft für Allgemeine und Angewandte Entomol-ogie , 13 , 135 – 139 .
Freise , J.F. , Heitland , W. & Toshevski , I . ( 2002 ) Parasitism of the horse-chestnut leaf miner, Cameraria ohridella , Deschka and Dimic (Lep., Gracillariidae), in Serbia and Macedonia . Anzeiger für Schädlingskunde , 75 , 152 – 157 .
Gilbert , M. & Grégoire , J.-C. ( 2003 ) Visual, semi-quantitative as-sessments allow accurate estimates of leafminer population densi-
ties: an example comparing image processing and visual evaluation of damage by the horse chestnut leafminer Cameraria ohridella (Lep., Gracillariidae) . Journal of Applied Entomology , 127 , 354 – 359 .
Gilbert , M. , Guichard , S. , Freise , J. et al . ( 2005 ) Forecasting Cameraria ohridella invasion dynamics in recently invaded countries: from validation to prediction . Journal of Applied Ecology , 42 , 805 – 813 .
Gilbert , M. , Svato š , A. , Lehmann , M. & Bacher , S . ( 2003 ) Spatial patterns and infestation processes in the horse chestnut leafminer Cameraria ohridella Deschka & Dimic (Lepidoptera: Gracillari-idae): a tale of two cities . Entomologia Experimentalis et Appli-cata , 107 , 25 – 37 .
Girardoz , S . ( 2001 ) A preliminary evaluation of life table parameters for the horse-chestnut leaf-miner , Cameraria ohridella (Deschka and Dimic) ( Lep., Gracillariidae) in Switzerland . International Diploma Thesis , Imperial College London .
Girardoz , S. , Kenis , M. & Quicke , D.L.J . ( 2006 ) Recruitment of na-tive parasitoids by an exotic leaf miner, Cameraria ohridella : host-parasitoid synchronization and influence of the environment . Agricultural and Forest Entomology , 8 , 48 – 56 .
Godfray , H.C.J. , Agassiz , D.L.J. , Nash , D.R. & Lawton , J.H . ( 1995 ) The recruitment of parasitoid species to two invading herbivores . Journal of Animal Ecology , 64 , 393 – 402 .
Grabenweger , G . ( 2003 ) Parasitism of different larval stages of Cam-eraria ohridella . Biocontrol , 48 , 671 – 684 .
Grabenweger , G . ( 2004 ) Poor control of the horse chestnut leafminer, Cameraria ohridella (Lepidoptera: Gracillariidae), by native Eu-ropean parasitoids: a synchronisation problem . European Journal of Entomology , 101 , 189 – 192 .
Grabenweger , G. , Avtzis , N. , Girardoz , S. , Hrasovec , B. , Tomov , R. & Kenis , M . ( 2005a ) Parasitism of Cameraria ohridella (Lepi-doptera, Gracillariidae) in natural and artificial horse-chestnut stands in the Balkans . Agricultural and Forest Entomology , 7 , 291 – 296 .
Grabenweger , G. , Kehrli , P. , Schlick-Steiner , B. , Steiner , F. , Stolz , M. & Bacher , S . ( 2005b ) Predator complex of the horse chestnut leafminer Cameraria ohridella : identification and impact assess-ment . Journal of Applied Entomology , 129 , 353 – 362 .
Grabenweger , G. & Lethmayer , C . ( 1999 ) Occurrence and phenol-ogy of parasitic Chalcidoidea on the horse chestnut leafminer, Cameraria ohridella Deschka & Dimic (Lep., Gracillariidae) . Journal of Applied Entomology , 123 , 257 – 260 .
Guillén , M. , Davis , D.R. & Heraty , J.M . ( 2001 ) Systematics and bi-ology of a new, polyphagous species of Marmara (Lepidoptera: Gracillariidae) infesting grapefruit in the Southwestern United States . Proceedings of the Entomological Society of Washington , 103 , 636 – 654 .
Hawkins , B.A. , Cornell , H.V. & Hochberg , M.E . ( 1997 ) Predators, parasitoids, and pathogens as mortality agents in phytophagous insect populations . Ecology , 78 , 2145 – 2152 .
Hellrigl , K . ( 2001 ) Neue Erkenntnisse und Untersuchungen über die Roßkastanien-Miniermotte . Gredleriana , 1 , 9 – 91 .
Hellrigl , K. & Ambrosi , P . ( 2000 ) Die Verbreitung der Rosskastanien-Miniermotte Cameraria ohridella Desch. & Dimic (Lepid., Gracillariidae) in der Region Südtirol-Trentino . Anzeiger für Schädlingskunde , 73 , 25 .
Hering , E.M . ( 1951 ) Biology of the Leaf Miners . Junk’s Gravenhage , The Netherlands .
Hespenheide , H.A . ( 1991 ) Bionomics of leaf-mining insects . Annual Review of Entomology , 36 , 535 – 560 .
Kalinova , B. , Svatos , A. , Kindl , J. , Hovorka , O. , Hrdy , I. , Kuldova , J. & Hoskovec , M . ( 2003 ) Sex pheromone of horse-chestnut leafminer Cameraria ohridella and its use in a pheromone-based monitoring system . Journal of Chemical Ecology , 29 , 387 – 404 .

158 S. Girardoz, D. L. J. Quicke and M. Kenis
© 2007 The Authors
Journal compilation © 2007 The Royal Entomological Society, Agricultural and Forest Entomology, 9, 141–158
Kehrli , P. , Lehmann , M. & Bacher , S . ( 2005 ) Mass-emergence de-vices: a biocontrol technique for conservation and augmentation of parasitoids . Biological Control , 32 , 191 – 199 .
Kenis , M. & Forster , B . ( 1998 ) Die Rosskastanien-Miniermotte: neu in der Schweiz . Der Gartenbau , 39 , 16 – 17 .
Kenis , M. , Tomov , R. , Svatos , A. et al . (2005) The horse-chestnut leaf miner in Europe – prospects and constraints for biological control . Proceedings of the Second International Symposium on Biological Control of Arthropods , September 12 – 16 , 2005 , Davos, Switzerland ( ed . by M.S. Hoddle ), pp . 77 – 90 . USDA Forest Ser-vice , Morgantown, West Virginia .
Kirizoglu , I . ( 1976 ) Zur Kenntnis der Fliedermotte, Gracilaria syrin-gella (F.) (Lepid., Gracillariidae. 2. Biologie; 3. Oekologie, Grad-ologie und Bekämpfung . Zeischrift für Angewandte Entomologie , 81 , 163 – 187 .
Klemola , T. , Ruohomaki , K. , Andersson , T. & Neuvonen , S . ( 2004 ) Reduction in size and fecundity of the autumnal moth, Epirrita autumnata , in the increase phase of a population cycle . Oecolo-gia , 141 , 47 – 56 .
Laing , J.E. , Heraty , J.M. & Corrigan , J.E . ( 1986 ) Leaf burial by the earthworm, Lumbricus terrestris (Oligochaeta: Lumbricidae), as a major factor in the population dynamics of Phyllonorycter blan-cardella (Lepidoptera: Gracillariidae) and its parasites . Environ-mental Entomology , 15 , 321 – 326 .
McClure , M.S . ( 1991 ) Density-dependant feedback and population cycles in Adelges tsugae (Homoptera, Adelgidae) on Tsuga ca-nadensis . Environmental Entomology , 20 , 258 – 264 .
Pavan , F. , Barro , P. , Bernardinelli , I. , Gambon , N. & Zandigiacomo , P . ( 2003 ) Cultural control of Cameraria ohridella on horse chest-nut in urban areas by removing fallen leaves in autumn . Journal of Arboriculture , 29 , 253 – 258 .
Pfeiffer , D.G. , Hull , L.A. , Biddinger , D.J. & Killian , J.C . ( 1995 ) Apple-Indirect pests . Mid-Atlantic Orchard Monitoring Guide ( ed . by H. W. Hogmire ), pp . 18 – 43 . Northeast Regional Agricultural Engineering Service , Ithaca, New York .
Pottinger , R.P. & Leroux , E.J . ( 1971 ) The biology and dynamics of Lithocolletis blancardella (Lepidoptera: Gracillariidae) on apple in Quebec. H. Mortality factors . Memoirs of the Entomological Society of Canada , 77 , 76 – 91 .
Preszler , R.W. & Price , P.W . ( 1988 ) Host quality and sawfly popula-tions: a new approach to life table analysis . Ecology , 69 , 2012 – 20 .
Pschorn-Walcher , H . ( 1994 ) Freiland-Biologie des eingeschleppten Rosskastanien- Miniermotte Cameraria ohridella Deschka et Dimic (Lep., Gracillariidae) im Wienerwald . Linzer Biologische Beiträge , 26 , 633 – 642 .
Pschorn-Walcher , H . ( 2001 ) Zehn Jahre Rosskastanien-Miniermotte ( Cameraria ohridella Deschka &Dimic; Lep., Gracillariidae) im Wienerwald . Linzer Biologische-Beitraege , 33 , 941 – 947 .
Quiring , D.T. & McNeil , J.N . ( 1984 ) Exploitation and interference intraspecific larval competition in the Dipteran leaf miner, Agro-myza frontella (Rondani) . Canadian Journal of Zoology , 62 , 421 – 427 .
Radeghieri , P . ( 2004 ) Cameraria ohridella (Lepidoptera, Gracillari-idae) predation by Crematogaster scutellaris (Hymenoptera, For-micidae) in Northern Italy (Preliminary note) . Bulletin of Insectology , 57 , 63 – 64 .
Sato , T. , Shinkaji , N. & Amano , H . ( 2004 ) Effects of larval density on larval survivorship and imaginal fecundity of Dacne picta (Co-leoptera: Erotylidae) . Applied Entomology and Zoology , 39 , 591 – 596 .
Skuhravy , V . ( 1998 ) Zur Kenntnis der Blattminen-Motte Cameraria ohridella Desch. & Dim. (Lep., Lithocolletidae) an Aesculus hip-pocsatanum L. in der Tschechischen Republik . Anzeiger für Schädlingskunde Pflanzenschutz Umweltschutz , 71 , 82 – 84 .
Southwood , T.R.E . ( 1978 ) Ecological Methods: With Particular Reference to the Study of Insect Populations . Chapman & Hall , U.K.
Stojanovic , A. & Markovic , C . ( 2005 ) Parasitoid complex of Phyllo-norycter robiniella (Clemens, 1859) (Lepidoptera, Gracillariidae) in Serbia . Journal of Pest Science , 78 , 109 – 114 .
Urbaneja , A. , Llácer , E. , Tomás , O. , Garrido , A. & Jacas , J.-A. ( 2000 ) Indigenous natural enemies associated with Phyllocnistis citrella (Lepidoptera: Gracillariidae) in eastern Spain . Biological Control , 18 , 199 – 207 .
Van Driesche , R.G. & Bellows , T.S. Jr ( 1996 ) Biological Control . Chapman & Hall , New York, New York .
Van Driesche , R.G. & Taub , G . ( 1983 ) Impact of parasitoids on Phyl-lonorycter leafminers infesting apple in Massachusetts, U.S.A . Protection Ecology , 5 , 303 – 317 .
Volter , L. & Kenis , M . ( 2006 ) Parasitoid complex and parasitism rates of the horse chestnut leafminer, Cameraria ohridella (Lepi-doptera: Gracillariidae) in the Czech Republic, Slovakia and Slo-venia . European Journal of Entomology , 103 , 367 – 370 .
Zegula , T. , Kranz , J. , Arnold , C. & Buchheim , A . ( 2002 ) Die Ross-kastanien-Miniermotte Cameraria ohridella (Deschka and Dimic) als potenzielle Nahrungsquelle für einheimische Singvögel . Char-adrius , 38 , 232 – 238 .
Accepted 2 January 2007 First published online 2 March 2007
Copyright © 2022 FDOKUMEN




















